Overview of the Division of Intramural Clinical and Biological Research National Institute on Alcohol Abuse and Alcoholism 2011-2012

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Overview of the Division of Intramural Clinical
and Biological Research
National Institute on Alcohol Abuse and Alcoholism
2011-2012

About the covers: The covers show, in fluorescent green, the striatal direct pathway (front) and the the striatal indirect pathway (back) in a section of mouse brain. Activation of both pathways is critical for control of movement. From work performed by members of the NIAAA Laboratory for Integrative Neuroscience and colleagues. For details, Cui G, Jun SB, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature in press. Managing and Scientific Editor: Peter B. Silverman, PhD, JD, Deputy Scientific Director Editing Assistant: Emily K. Surette, BS For further information contact: George Kunos, MD, PhD Scientific Director National Institute on Alcohol Abuse and Alcoholism 5625 Fishers Lane, Room 2S-24 Bethesda, MD 20892 Phone: 301 443 2069 Fax: 301 480 0257 e-mail: [email protected]

National Institute on Alcohol Abuse and Alcoholism
Overview of the
Division of Intramural Clinical and Biological Research
2011-2012
U.S. Department of Health and Human Services Public Health Service National Institutes of Health


i
Table of Contents Foreword by George Kunos, MD, PhD, Scientific Director, NIAAA .................................. iii Division of Intramural Clinical and Biological Research Organizational Chart ............... vi Board of Scientific Counselors ................................................................................................ vii Laboratory of Clinical and Translational Studies ...................................................................1
Section on Molecular Pathophysiology ..............................................................................2
Section on Brain Electrophysiology and Imaging .............................................................13
Section on Clinical Assessment and Treatment Evaluation ..............................................20
Section on Human Psychopharmacology ..........................................................................28
Section on Clinical Psychoneuroendocrinology & Neuropsychopharmacology ...............45
Laboratory of Membrane Biochemistry and Biophysics ......................................................50
Section on Nuclear Magnetic Resonance ..........................................................................54
Section of Molecular Pharmacology and Toxicology .......................................................63
Section on Nutritional Neuroscience .................................................................................74
Laboratory of Neurogenetics ...................................................................................................91
Section on Human Neurogenetics ......................................................................................97
Section of Comparative Behavioral Genomics ................................................................125
Laboratory for Integrative Neuroscience .............................................................................136
Section on Synaptic Pharmacology .................................................................................139
Section on Neuronal Structure .........................................................................................145

ii
Table of Contents
Laboratory of Physiologic Studies .........................................................................................153
Section on Neuroendocrinology .....................................................................................154
Section on Oxidative Stress – Tissue Injury ..................................................................169
Laboratory of Molecular Physiology ....................................................................................182
Section on Transmitter Signaling ...................................................................................184
Section on Model Synaptic Systems ..............................................................................187
Section on Cellular Biophotonics ...................................................................................191
Laboratory of Metabolic Control ..........................................................................................193
Laboratory of Neuroimaging .................................................................................................198
Laboratory of Epidemiology and Biometry .........................................................................212
Laboratory of Behavioral and Genomic Neuroscience .......................................................227
Laboratory of Molecular Signaling .......................................................................................231
Laboratory of Liver Diseases .................................................................................................245
Chemical Biology Research Branch, Drug Design and Synthesis Section .........................251
DICBR Publications 2011-2012 .............................................................................................267

iii
Foreword The Division of Intramural Clinical and Biological Research (DICBR) of the National Institute on Alcohol Abuse and Alcoholism (NIAAA) has as its overall goals understanding the biological basis of alcohol use disorders and alcohol-induced morbidities, and developing novel strategies and tools for the prevention and treatment of these disorders. Research is conducted at multiple levels: cellular/molecular, animal studies in rodents and non-human primates, human studies of the genetics and epidemiology of alcoholism and co-morbidities, and validation of novel molecular targets for alcohol use disorders. Collaborations among the various units and between these units and investigators from other NIH institutes or extramural institutions reflect the integrative, multidisciplinary nature of ongoing research. Research into the causes and consequences of alcoholism is not a discipline in its own right, rather it relies on and employs the tools of a wide range of biological disciplines. The outstanding scientists heading the various research units of our intramural program are recognized for their contributions to their chosen research fields, be it molecular biology, neuroscience, biochemistry, physical chemistry, physiology, genetics, epidemiology or psychiatry, and apply their expertise to answer questions related to alcohol use disorders. Currently, there are twelve intramural NIAAA Laboratories, several of which are divided further into a number of Sections, each headed by a principal investigator, each of whom provides a description of their ongoing research in this volume. In addition to these units, Dr. Kenner Rice heads the Chemical Biology Research Branch (CBRB), jointly sponsored by NIDA and NIAAA. The Laboratory of Clinical and Translational Studies (LCTS), headed by Dr. Markus Heilig, is the only clinical laboratory in our intramural program. It has recently become the core of the joint NIAAA/NIDA clinical program under Dr. Heilig’s leadership, with a focus on mechanisms and treatment of addictive disorders, including alcoholism, through a combination of preclinical studies aimed to discover and validate new molecular targets, and clinical proof-of-concept studies that test these novel targets through the use of therapeutic agents directed against such targets. This work also takes advantage of state-of-the-art functional brain imaging techniques. The Laboratory of Membrane Biochemistry and Biophysics (LMBB), headed by Dr. Klaus Gawrisch, explores the relationship between alcohol-induced changes in membrane structure and function, and investigates the biological functions of polyunsaturated fatty acids, with special emphasis on docosahexaenoic acid (DHA), and analyzes the mechanisms of oxidative stress-induced organ damage. The Laboratory of Neuroimaging (LNI), headed by Dr. Nora Volkow, uses in vivo brain imaging to gain insight into the neurochemical basis of addictive disorders, including drug-seeking behavior, alcoholism and alcohol dependence. The Laboratory of Neurogenetics (LNG), headed by Dr. David Goldman, aims to identify genes that predispose to or protect from alcoholism and comorbid disorders. Their approach combines functional genomics in in vitro systems, large scale analyses of gene expression, and linkage analyses including both candidate gene and genome-wide approaches. With support from the NIDA IRB, plans have been developed to incorporate LNG into another joint NIAAA/NIDA research program focusing on the genetics of addictive disorders. The Laboratory of Epidemiology and Biometry (LEB), headed by Dr. Bridget Grant, designs, conducts and analyzes the data collected from national epidemiologic surveys on alcohol use disorders and related conditions, in order to gain insight into their prevalence, co-morbidities, prevention, treatment needs and societal costs. The unifying theme of research in the Laboratory of Integrative Neuroscience (LIN), headed by Dr. David Lovinger, is forebrain mechanisms of cognition and behavioral control that play a role in addiction, which are investigated using a combination of electrophysiological, cell and

iv
molecular biological techniques and paradigms. The newly created Laboratory of Behavioral and Genomic Neuroscience (LBGN) headed by the recently tenured Dr. Andrew Holmes focuses on the molecular underpinnings of addictive behaviors and their modulation by stress. The Laboratory of Molecular Signaling (LMS), headed by Dr. Hee-Yong Kim, explores the signaling pathways involved in the effects of polyunsaturated fatty acids on neuronal survival and their modulation by ethanol, using a combination of cell biological and mass spectrometry-based proteomic approaches. Research in the Laboratory of Physiologic Studies (LPS), headed by Dr. George Kunos, focuses on neuroendocrine mechanisms that regulate appetitive functions, including alcohol drinking behavior, on liver biology. Mechanisms of oxidative and nitrosative stress and their modulation by the endocannabinoid system are studied in the similarly named section, headed by the recently tenured Dr. Pal Pacher. The Laboratory of Liver Diseases (LLD), headed by Dr. Bin Gao, investigates the molecular pathology of alcoholic and nonalcoholic fatty liver diseases and seeks targets for their treatment. The Laboratory of Metabolic Control (LMC), headed by Dr. Richard Veech, studies how the flux through various metabolic pathways is involved in controlling cellular energy status, with particular emphasis on the metabolism of ketone bodies and its therapeutic implications. Finally, the Laboratory of Molecular Physiology (LMP), headed by Dr. Stephen Ikeda, investigates cellular, subcellular and molecular mechanisms underlying synaptic transmission in the nervous system, using electrophysiological and advanced cellular imaging techniques, as well as zebrafish genetics. Despite an essentially flat budget during the last 5 years, research productivity has reached new highs in our intramural program. Significant scientific advances published in 2011/2012 include:
• Concurrent activation of striatal direct and indirect pathways during action initiation. (LIN and LMP, Nature doi:10.1038/nature11846 2012).
• Enhanced endocannabinoids promote fear plasticity. (LBGN and LPS, Molecular Psychiatry doi: 10.1038/mp.2012.72; Faculty 1000 2x ‘must read’ http://f1000.com/717948500).
• Chronic alcohol remodels prefrontal neurons and disrupts NMDA receptor-mediated fear extinction encoding (LBGN, Nature Neuroscience 15:1359-1361, 2012; Faculty 1000 ‘must read’ http://f1000.com/717956468).
• Cocaine supersensitivity and enhanced motivation for reward in mice lacking D2 autoreceptors (LIN, Nature Neuroscience 14:1033-38, 2011).
• Pharmacogenetic approaches to the treatment of alcohol addiction (LCTS and LNG, Nature Reviews Neuroscience 12:670-84, 2011).
• Stress-related neuropeptides and addictive behaviors: beyond the usual suspects. (LCTS, Neuron 76:192-208, 2012.)
• A human serotonin transporter polyadenylation polymorphism selectively modulates the retention of fear extinction memory (LBGN, Proc Natl Acad Sci USA 109:5493-5498, 2012).
• Paradoxical reversal learning enhancement by stress or prefrontal cortical damage: rescue with BDNF (LBGN, Nature Neuroscience 14:1507-1509; Faculty 1000 ‘recommended’ paper http://f1000.com/13405009).
• Identification of adult renal progenitor cells capable of nephron formation and regeneration in zebrafish (LMP, Nature 470: 95-100, 2011).
• Phosphatidylserine as a critical modulator of Akt activation (LMS, Journal of Cell Biology 192:979-992, 2011).

v
• Interleukin-22 induces hepatic stellate cell senescence and restricts liver fibrosis (LLD, Hepatology 56:1150-1159, 2012; Faculty of 1000 Medicine, http://f1000.com/714297813).
• Hyperactivation of anandamide synthesis and regulation of cell cycle progression via CB1 receptors in the regenerating liver (LPS and LNG, Proc Natl Acad Sci USA 108:6323-8, 2011; editorial in PNAS, 108:6339-6340, 2011).
• AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin resistant mice (LLD, Cell Metabolism 13:376-88, 2011).
• Peripheral CB1 receptor inverse agonism reduces obesity by reversing leptin resistance (LPS, Cell Metabolism 16:167-179, 2012; highlighted in Nat Rev Drug Discovery, Nat Rev Endocrinol, 2012).
• Reduced cannabinoid CB1 receptor binding in alcohol dependence measured with positron emission tomography (LCTS, Molecular Psychiatry doi:10.1038/mp.2012.100).
• Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol (LNG, Proc Natl Acad Sci USA 108:6626-31, 2011).
• Hepatic cannabinoid receptor-1 mediates diet-induced insulin resistance via inhibition of insulin signaling and clearance in mice (LPS, Gastroenterology 142:1218-28, 2012; editorial on page 1063, ‘Covering the Cover’, p. 1049; Faculty of 1000: Stieger B: 2012. F1000.com/716247804).
Research excellence in the DICBR has been recognized in the last two years in ways additional to an exceptional publication record. The Society for Neuroscience presented Dr. Xin Jin, a postdoctoral fellow in the Laboratory for Integrative Neuroscience, with the 2011 Gruber International Research Award in Neuroscience, recognized Dr. Rui Costa, a former member and currently guest researcher in LIN and an investigator at the Champalimaud Center for the Unknown in Lisbon, Portugal, with a 2012 Young Investigator Award, and presented the prestigious Jacob W. Waletzky Award for 2012 to Dr. Andrew Holmes, Laboratory of Behavioral and Genomic Neuroscience, for research in the area of substance abuse and brain and nervous system. Despite the challenging budgetary environment, our investigators have been able to maintain the viability of our research program by taking advantage of funding opportunities from outside of the NIH and entering into cooperative research and development agreements (CRADAs) with outside entities. DICBR researchers have also submitted several invention reports in 2011-2012. Even more important than adequate resources, however, is the creativity and uncompromising dedication of our intramural investigators.
George Kunos, MD, PhD, Scientific Director National Institute on Alcohol Abuse and Alcoholism
vi

vii
NIAAA Board of Scientific Counselors 2011-2012
The Board of Scientific Counselors is comprised of eminent scientists with expertise in alcohol research and related areas of scientific endeavor. Employed outside of NIH, they, along with ad hoc reviewers, periodically convene to review the quality of NIAAA intramural science and scientists and advise the Scientific Director. NIAAA is most appreciative of their efforts. Tatiana Foroud, PhD, Chair Chancellor's Professor P. Michael Conneally Professor of Medical and Molecular Genetics Director of Hereditary Genomics Division Department of Medical & Molecular Genetics Indiana University School of Medicine Rexford S. Ahima, MD, PhD Associate Professor of Medicine Division of Endocrinology, Diabetes & Metabolism University of Pennsylvania Medical School Yusuf A. Hannun, MD Director, Stony Brook Cancer Center Vice Dean for Cancer Medicine Joel Kenny Professor of Medicine Stony Brook University Health Science Center Kenneth S. Kendler, MD Rachel Brown Banks Distinguished Professor of Psychiatry and Professor of Human Genetics Director, Psychiatric Genetics Research Program Director of the Virginia Institute for Psychiatric and Behavioral Genetics Department of Psychiatry Medical College of Virginia of Virginia Commonwealth University Virginia Institute for Psychiatric and Behavioral Genetics Scott M. Thompson, PhD Professor, Department of Physiology University of Maryland School of Medicine Kathleen T. Brady, MD, PhD Professor, Department of Psychiatry and Behavioral Sciences Director, Clinical Neuroscience Division Associate Dean, Clinical and Translational Research Director, South Carolina Clinical and Translational Research Institute Medical University of South Carolina Susan G. Amara, PhD Detre Professor Chair, Department of Neurobiology University of Pittsburgh

1
_____________________________________________________________________
Laboratory of Clinical and Translational Studies ______________________________________________________________________ Markus A. Heilig, MD, PhD, Chief Suzanne Bell, Administrative Officer Dena Stringer, Administrative Assistant Catherine Little, IRB Coordinator Melanie Schwandt, PhD, Database and Biostatistics Coordinator Juan Rivas, IT-support Karen Smith, Biomedical Informationist Medications development: Target discovery and validation
l to r: C Little, D Stringer, K Smith, M Heilig, M Schwandt
Our laboratory as a whole applies a translational approach to developing novel pharmacological treatments for alcohol dependence. In order to accomplish our mission, the Laboratory has five highly interactive components with diverse focus and expertise. The Section on Molecular Pathophysiology provides a resource for target discovery and validation. A particular focus is on the neurobiology of stress and negative affect, which are related to alcohol dependence and relapse processes in important ways, and of high relevance for several other psychiatric conditions. This section utilizes rodent models, molecular studies and pharmacological approaches to identifying novel mechanisms and treatment targets, and validating them to a level that can provide a rationale for human development. The Section on Clinical and Treatment Evaluation represents the other end of the drug evaluation spectrum within the intramural program, providing the platform for clinical evaluation of pharmacological therapies for alcohol use disorders. Between the two are the Section on Brain Electrophysiology and Imaging, the Section on Human Psychopharmacology and the Section on Clinical Psychoneuroendocrinology and Neuropsychopharmacology (joint with NIDA). The Section on Brain Electrophysiology and Imaging focuses on the study of the effect of brain structure and function on the development of alcoholism, and the effect of alcoholism on brain structure and function. The Section on Human Psychopharmacology studies alcohol pharmacokinetics and pharmacodynamics as a function of gender, family history and alcohol use history, and also provides tools for producing pharmacokinetically controlled brain alcohol exposure that can be used to assess the actions of experimental therapeutics. The Section on Clinical Psychoneuroendocrinology and Neuropsychopharmacology focuses on neuroendocrinology of appetitive behaviors, as potential targets for new medications. The research conducted in each of these components is presented in greater detail in the pages that follow. Dr. Heilig: [email protected]

2
______________________________________________________________________
Section on Molecular Pathophysiology ______________________________________________________________________ Markus A. Heilig, MD, PhD, Chief Jenica Tapocik, PhD, Staff Scientist Melanie Schwandt, PhD, Staff Scientist Robert Eskay, PhD, Senior Biologist Hui Sun, MD, Biologist Ruslan Damadzic, MD, Biologist Erick Singley, BS, Chemist Jesse Schank, PhD, Postdoctoral IRTA Estelle Barbier, PhD, Visiting Fellow Eric Augier, PhD, Visiting Fellow Meghan Flanigan, Postbac IRTA Nate Juergens, Postbac IRTA Courtney King, Postbac IRTA Matt Solomon, Technical IRTA Faaz Rehman, Postbac IRTA Ali Pincus, Technical IRTA Camilla Karlsson, Doctoral student Medications development: Target discovery and validation The Section on Molecular Pathophysiology uses behavioral, molecular and pharmacological approaches in animal models for discovery and validation of novel mechanisms that can be targeted for treatment of alcoholism. A particular focus is on the neurobiology of stress and negative affect, processes that are central to alcohol dependence and relapse, and also relevant for other psychiatric disorders such as depression and anxiety. The section collaborates closely with the imaging and clinical sections on human early proof-of-concept and mechanistic studies. Animal models for target discovery Although addiction is a complex disorder that cannot be fully modeled in experimental animals, rat and mouse models can provide important discovery and validation tools. These models must be applied in a thoughtful manner, because laboratory rodents ordinarily do not spontaneously consume amounts of EtOH that are sufficient to produce pharmacological effects. Consumption in non-dependent, genetically heterogeneous animals can therefore be for taste, caloric content or other reasons unrelated to the addictive properties of alcohol. Against that background, we use two main categories of models: Pre-existing genetic susceptibility factors Genetic selection for high alcohol preference enriches alleles that contribute to high voluntary alcohol intake during the initiation phase of alcohol use. This approach is well established in the field. Our group has collaborated over the years with several laboratories that have maintained selected preferring lines, more recently including Dr. Ciccocioppo (Camerino, Italy: msP rat), and the Indiana Alcohol Center (P-rat). Many different alleles contribute to the phenotype of

3
high alcohol preference, and animals selected for this trait can be phenotypically very different. For instance, we have previously found that the Finnish AA line is impulsive, while both mSP and P rats show high measures of anxiety-like behavior, suggesting a role for negatively reinforced alcohol drinking. An emerging interest in the laboratory, and one related to genetic susceptibility factors, is pharmacogenetics. Prior work by us and others has suggested that genetic variation at the locus encoding the mu-opioid receptor (OPRM1) modulates both alcohol reward, and therapeutic response to naltrexone, so far the best established pharmacotherapy for alcoholism. While human studies have been somewhat conflicting, prior work in non-human primates allowed us to obtain robust evidence that carriers of a rhesus variant functionally equivalent to the minor human OPRM1 118G allele, 77G, show a robust response to naltrexone, while alcohol preference is not at all influenced by naltrexone in individuals homozygous for the major allele at this locus. To establish the mechanistic role of OPRM1 variation, and to isolate the influence of A118G variation from other markers in the same linkage disequilibrium block, we embarked on two complementary approaches. Collaborating with other components of LCTS, we studied the effects of this SNP on central dopamine (DA) release in response to a pharmacokinetically controlled alcohol challenge, using positron emission tomography (PET) and displacement of the DA-D2 ligand 11C-raclopride. In parallel, we created two lines of humanized mice, where exon 1 of the OPRM1 gene was replaced with the human 118A, or the 118G allele, respectively (1). The human PET component of this study indicated a more vigorous DA response to alcohol in carriers of the OPRM1 118G allele than in subjects homozygous for the major 118A allele. Using the humanized mouse lines, we were able to isolate this effect specifically to the A118G SNP, and directly demonstrate, using brain microdialysis, enhanced DA release in 118GG animals. Further work with the humanized mouse lines revealed elevated alcohol consumption in 118GG mice, and experiments are underway to investigate whether 118GG mice will be preferentially or selectively sensitive to NTX (Thorsell et al., in preparation). In collaboration with a laboratory at University of North Carolina, we are also investigating the consequences of this receptor variation for brain reward as measured by intracranial self-stimulation. Preliminary data from that work indicate that, among several ligands, morphine distinguishes between the receptor variants, indicating a loss-of-function mutation for morphine effects (Robinson et al, in preparation). Accordingly, in 118G mice, morphine is less potent both for Ca2+ flux effects in dispersed ganglionic neurons, as well as analgesia (2). Long-term neuroadaptations Over the course of developing alcohol dependence, long term neuroadaptations occur in numerous brain systems. These include, importantly, those that mediate behavioral stress responses (reviewed in (3)). Figure 1 shows the conceptual framework for this view.

4
use heavy use earlydependence
late dependence Šneuroadapted state
Impulsive stage
abstinencerelapse
abstinencerelapse
abstinencerelapse
Compulsive stage
Bingeintoxication
AbstinenceNeutral affect
Protracted abstinenceNegative affect
Prolonged intoxication
Rewardcraving
Pleasurableeffects
Relief craving
Relief
Figure 1. Conceptual framework for targeting negatively reinforced, neuroadaptive drinking.
In prior work, we had demonstrated that prolonged exposure of the rat brain to repeated cycles of intoxication and mild withdrawal results in a long-term, possibly life-long up-regulation of voluntary alcohol intake, presumably encoded by long-term changes in gene expression patterns. The up-regulation, shown in Figure 2, is to at least twice the levels seen in animals without a history of dependence, and is selectively sensitive to the clinically effective drug acamprosate. This model has offered a valuable tool for evaluating the activity of candidate medications.
)y 7
a ***
d/ 6
gk/g 5
( e 4
katn 3
i lo 2
nah 1
Et
0Control Post-dependent
Ethanol vapor Recovery Voluntary exposure to eliminate ethanol consumption 3 week
acute withdrawal ”detox”
Figure 2. Excessive voluntary alcohol consumption following a history of dependence.

5
Target validation In addition to up-regulated voluntary alcohol intake, the post-dependent state is also characterized by an increased sensitivity to stress, a major factor triggering relapse in alcohol dependent patients, and reinstatement of alcohol-seeking in experimental animals. Recent findings from our and other laboratories point to glutamatergic signaling, as well as several stress-related neuropeptide systems as a promising category of candidate targets, preferentially sensitive in the post-dependent state. Metabotropic Glutamatergic Receptor 2 (mGluR2) We have collaborated with Dr. Wolfgang Sommer to complete an ambitious project he initiated while still a post-doctoral fellow in the laboratory. Using laser capture microscopy, Sommer et al found that following a history of alcohol dependence, mGluR2 expression is almost entirely lost in infralimbic medial prefrontal cortex (mPFC). When this expression was restored using a viral vector, the phenotype of escalated alcohol self-administration was rescued (Sommer et al, in press, J Neurosci). This indicates that alcohol dependence and the escalation of voluntary alcohol intake that are associated with it are related to a loss of the break on glutamatergic neuronal activity in the mPFC, in agreement with the hypothesis that a hyperglutamatergic state emerges following a history of dependence. Additional work in collaboration with the Goldman and Lovinger laboratories (Zhou et al., in review) supports a critical role for mGluR2. Together, these data provide validation for mGluR2 as a candidate treatment target in alcohol use disorders. To obtain a screening model for candidate medications that would not require long term dependence induction, we attempted to emulate the hyperglutamatergic state using mice with a loss of the glial glutamate transporter GLAST, and predicted that the GLAST deletion would result in an escalation of alcohol intake. Contrary to our hypothesis, we found that GLAST null-mutants consumed very low amounts of alcohol, and did not show place preference for alcohol. In collaboration with the Lovinger laboratory, we found that this is presumably related to a loss of endocannabinoid-mediated long term depression, a developmental effect (4). Corticotropin-Releasing Hormone 1 Receptor (CRH1R), and Glucocorticoid Receptors (GR) We have previously found that both the elevated self-administration of alcohol and the increased behavioral sensitivity to stress in the post-dependent state is in a large part mediated by an up-regulation of CRH1R within the amygdala. This converges with our findings of a stress-sensitive / anxious behavioral phenotype in the msP rat, accompanied by an innate up-regulation of CRH1R in the amygdala and several other brain regions. The up-regulated CRH1 gene expression in msP rats is driven by a polymorphism in the promoter region of the receptor gene in this line. Using the selective CRH1R antagonist antalarmin, and the novel, orally available and brain penetrant CRH1 antagonist MTIP, we have shown that this contributes both to the elevated self-administration of alcohol and the anxious phenotype of msP rats (reviewed in (3)). These findings provided a validation of the CRH1R as a treatment target in alcoholism, and led to clinical studies that are currently ongoing in our program. An outstanding question in the field has been whether the effects of CRH1 antagonists are related to the HPA-axis, central stress-reactive mechanisms, or both. A collaborative project with

6
the laboratory of Dr. Spanagel showed that a forebrain selective deletion of the Crh1r gene using a nestin-promoter resulted in suppression of drinking that had escalated due to a history of dependence, or stress. In contrast, baseline self-administration was unaffected (5). This supports our hypothesis that a recruitment of extrahypothalamic, rather than HPA-axis related Crh1r systems is involved in escalation following a history of dependence. However, the activity of the HPA-axis is involved in the neuroadaptive processes that lead to escalation, as the GR antagonist mifepristone prevents post-dependent escalation (6). We also examined the role of HPA axis activity for neuronal death that occurs following binge alcohol intake, and found that neuronal death in the entorhinal cortex can be prevented using mifepristone. These results suggest that elevated circulating corticosterone in part mediates alcohol-induced neurotoxicity in the brain through activation of GR (Cippitelli et al., in press). Neurokinin 1 Receptors (NK1Rs) Substance P (SP) mediates stress responses through actions in the amygdala, while NK1R antagonists have anti-stress / anti-anxiety properties. In 2008, we reported that genetic inactivation of NK1Rs suppresses alcohol intake in mice, while NK1R antagonism alleviates alcohol craving in humans. We subsequently went on to show that NK1R gene deletion blocks alcohol reward, as well as escalation of voluntary alcohol intake over repeated cycles of access and deprivations. Divergence between human and rodent NK1Rs posed a challenge to obtaining a fuller pharmacological validation of the SP-NK1 system as an alcoholism treatment target. To address this, we first showed that in wild type C57BL/6 mice, the NK1R antagonist L-703606 dose dependently suppressed alcohol intake, but was ineffective in NK1R knock outs, demonstrating the receptor specificity of the drug effect. We then used an NK1R antagonist specifically engineered for high affinity to the rat receptor, L822429, and assessed its effects on alcohol-related behaviors in rats. At the doses used, L822429 did not suppress alcohol self-administration or cue-induced reinstatement in non-dependent Wistar rats, but potently and dose-dependently suppressed stress-induced reinstatement of alcohol-seeking, with a complete suppression at the highest dose. This effect was highly specific behaviorally (6). While L822429 is not effective in suppressing the modest rates of alcohol self-administration in non-dependent Wistars, it potently and dose-dependently suppresses escalated self-administration rates in genetically selected, alcohol preferring P-rats (Figure 3A). Within the central amygdala (CeA), elevated TacR1 expression was accompanied by elevated NK1R binding and an infusion of L822429 directly within the CeA replicated the systemic antagonist effects on alcohol self-administration in P rats. All P-rats but only 18% of their founder Wistar population were CC homozygous for a 1372G/C single nucleotide polymorphism. In silico analysis indicated that the TacR1 -1372 genotype could modulate binding of the transcription factors GATA-2 and E2F-1. Electromobility shift and luciferase reporter assays suggested that the -1372C allele confers increased transcription factor binding and transcription (Figure 3B) (Schank et al, in press Biol Psychiat). Genetic deletion of NK1R has also been shown to decrease the reinforcing properties of opioids, but it was unknown whether pharmacological NK1R blockade has the same effect. We determined that L822429 reduced heroin self-administration and the motivation to consume heroin in short (ShA) and long (LgA) access groups. L822429 also decreased anxiety-like behavior in both groups. Expression of TacR1 was decreased in reward- and stress-related brain areas both in ShA and LgA rats compared with heroin-naïve rats, but did not differ between the two heroin-experienced groups. In contrast, passive exposure to heroin produced increases in
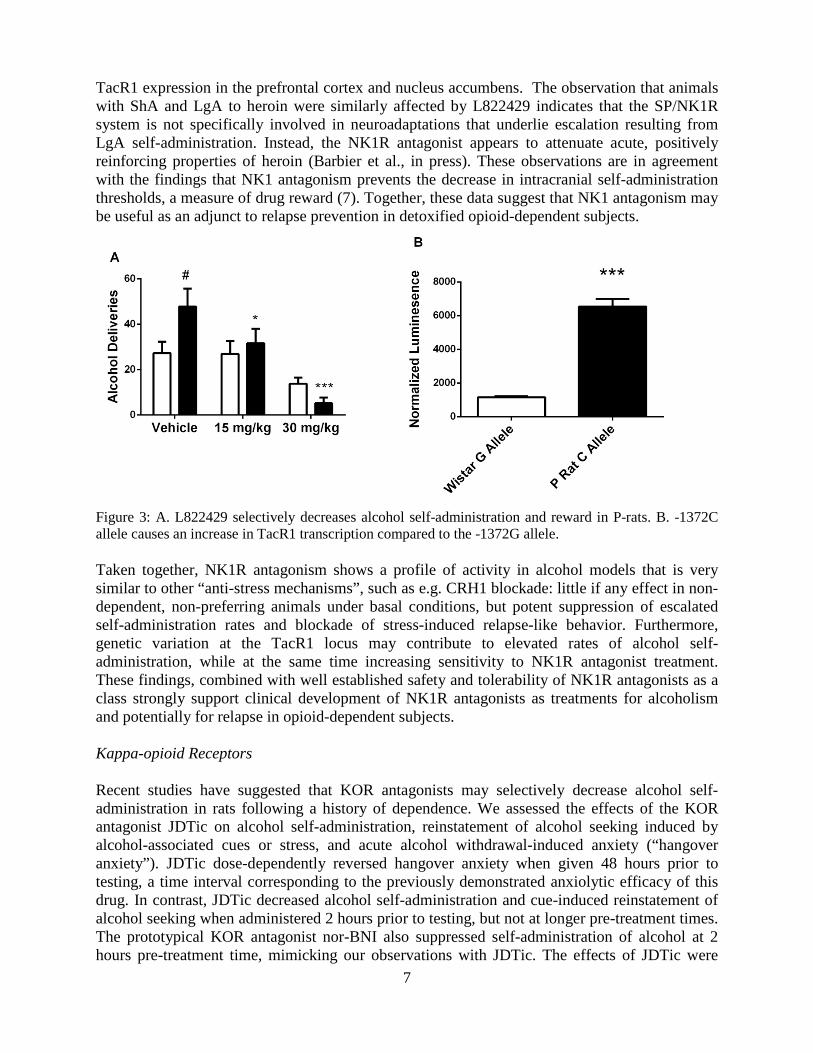
7
TacR1 expression in the prefrontal cortex and nucleus accumbens. The observation that animals with ShA and LgA to heroin were similarly affected by L822429 indicates that the SP/NK1R system is not specifically involved in neuroadaptations that underlie escalation resulting from LgA self-administration. Instead, the NK1R antagonist appears to attenuate acute, positively reinforcing properties of heroin (Barbier et al., in press). These observations are in agreement with the findings that NK1 antagonism prevents the decrease in intracranial self-administration thresholds, a measure of drug reward (7). Together, these data suggest that NK1 antagonism may be useful as an adjunct to relapse prevention in detoxified opioid-dependent subjects.
Figure 3: A. L822429 selectively decreases alcohol self-administration and reward in P-rats. B. -1372C allele causes an increase in TacR1 transcription compared to the -1372G allele. Taken together, NK1R antagonism shows a profile of activity in alcohol models that is very similar to other “anti-stress mechanisms”, such as e.g. CRH1 blockade: little if any effect in non-dependent, non-preferring animals under basal conditions, but potent suppression of escalated self-administration rates and blockade of stress-induced relapse-like behavior. Furthermore, genetic variation at the TacR1 locus may contribute to elevated rates of alcohol self-administration, while at the same time increasing sensitivity to NK1R antagonist treatment. These findings, combined with well established safety and tolerability of NK1R antagonists as a class strongly support clinical development of NK1R antagonists as treatments for alcoholism and potentially for relapse in opioid-dependent subjects. Kappa-opioid Receptors Recent studies have suggested that KOR antagonists may selectively decrease alcohol self-administration in rats following a history of dependence. We assessed the effects of the KOR antagonist JDTic on alcohol self-administration, reinstatement of alcohol seeking induced by alcohol-associated cues or stress, and acute alcohol withdrawal-induced anxiety (“hangover anxiety”). JDTic dose-dependently reversed hangover anxiety when given 48 hours prior to testing, a time interval corresponding to the previously demonstrated anxiolytic efficacy of this drug. In contrast, JDTic decreased alcohol self-administration and cue-induced reinstatement of alcohol seeking when administered 2 hours prior to testing, but not at longer pre-treatment times. The prototypical KOR antagonist nor-BNI also suppressed self-administration of alcohol at 2 hours pre-treatment time, mimicking our observations with JDTic. The effects of JDTic were

8
behaviorally specific, as it had no effect on stress-induced reinstatement of alcohol seeking, self-administration of sucrose, or locomotor activity. Our results support an involvement of KOR in regulation of alcohol-related behaviors and provide support for KOR antagonists, including JDTic, to be evaluated as medications for alcoholism. Exploratory targets Studies by our group and others have over many years indicated that NPY is a potent endogenous anti-stress system that counteracts behavioral stress responses mediated by CRH. We and others have also validated the role of NPY in modulating stress responses in humans, using functional genetic variation, post-mortem brain tissue expression, and psychiatric analyses in a longitudinal cohort. As predicted by those findings, central NPY administration potently blocks stress-induced relapse to alcohol seeking. Based on these observations, it has long been a hypothesis that potentiation of NPY signaling would potentially be beneficial in alcohol use disorders. An NPY Y1 agonist with drug-like properties, has, however, not emerged. As an alternative approach, we have examined whether endogenous NPY drive could be potentiated through the use of blockade of presynaptic Y2 receptors. Under a co-operative research and development agreement (CRADA) with Johnson & Johnson, we recently completed an evaluation of JNJ-31020028, a novel, non-peptide, brain penetrant Y2 antagonist, in a range of alcohol-related models (8). Although showing some activity, the effects of JNJ-31020028 followed an inverted-U-shaped dose-response curve, possibly reflecting activity at Y2 heteroceptors on glutamatergic terminals at higher doses. Other ways of modulating NPY function may therefore be required. It has been reported that blockade of type 1 receptors for melanin concentrating hormone (MCH1) produces anti-anxiety and anti-depressant-like effects. In an initial evaluation of this mechanism in alcohol-related behaviors, we found that the MCH1 antagonist GW803430 potently suppressed operant alcohol self-administration and cue-induced reinstatement of alcohol-seeking. Unexpectedly, stress-induced reinstatement was unaffected. Furthermore, GW803430 decreased sucrose self-administration, cue-induced reinstatement and progressive ratio. However, saccharin self-administration was unaffected (9). Recently we identified that GW803430 decreased consumption in high consuming rats within the intermittent access paradigm (Wistar and P-rats) (Karlsson et al., in preparation). These data provide the first evidence for MCH1 receptors as a candidate target for pharmacotherapy in alcoholism. Proinflammatory activity has a detrimental effect in behavioral responses to stress and addictive processes. Interleukin -1 beta (IL-1β) is a key proinflammatory cytokine that is induced by stress as well as alcohol exposure, and has been suggested to modulate stress- and alcohol-related behaviors. We investigated a potential role of the receptor for IL-1β (IL-1R) in regulation of alcohol consumption and stress related drinking using IL-1R knockout mice (IL-1R KO). Access to increasing concentrations of alcohol revealed that IL-1R KO mice drink significantly less than wildtypes (WT) at pharmacologically active concentrations. This decreased consumption was associated with enhanced sensitivity to the sedative effects of alcohol in the IL-1R KO mice. Repeated cycles of deprivation induced escalation in WT mice but at a lesser degree in the IL-1R KO mice at each cycle. Although the IL1R is involved in stress responses, the IL-1R KO mice do not show differences in stress induced alcohol consumption (Karlsson et al, in preparation). A potential explanation is that other cytokines, such as TNF-α and its receptor, TNF-1R, may act in parallel to the IL-1R signaling pathways, and also contribute to stress- and alcohol-related behaviors. Recently, we found that mice lacking both the IL-1R and the TNF-1R consume

9
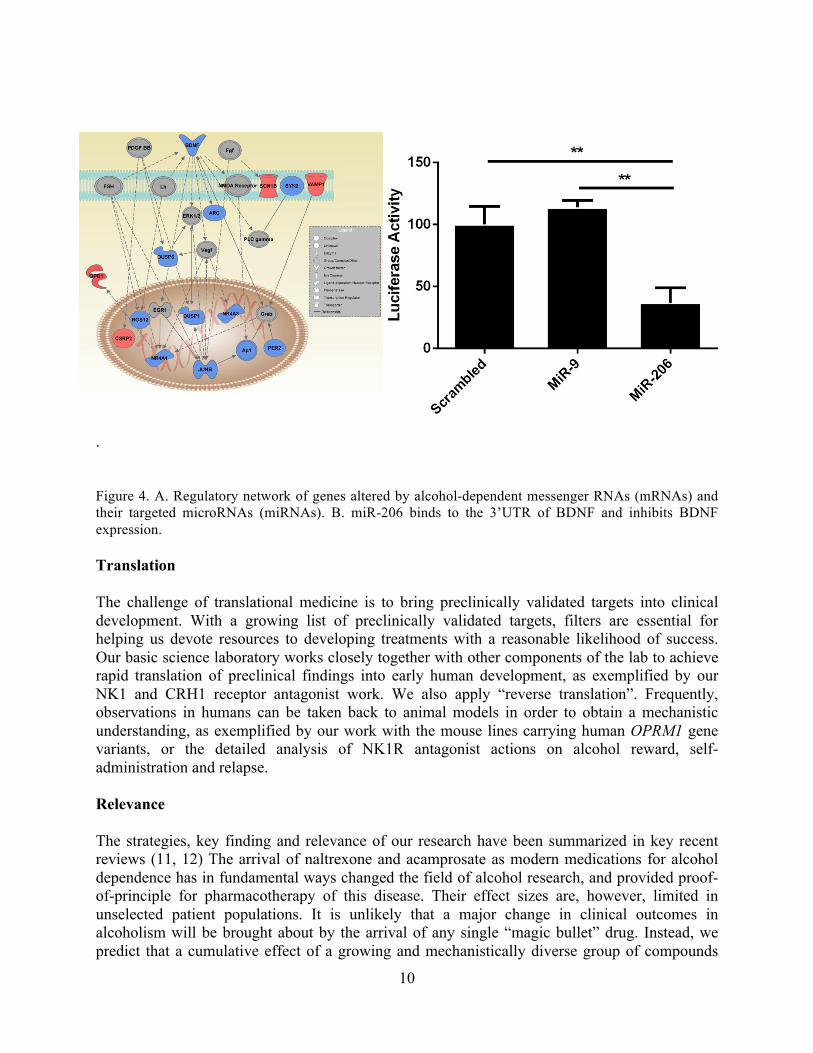
significantly less alcohol than their WT controls. WT mice escalate their intake over time, while no escalation was observed in IL-1R-TNF-1R double knockout mice. We are currently evaluating a TNF-1R inhibitor to assess the specific role of TNF-1R on escalated alcohol consumption (Karlsson et al., in preparation). Finally, we have collaborated with the laboratory of Dr. Roberto Ciccocioppo on exploring the potential of the neuropeptide S (NPS) receptor as a treatment target. In the course of this collaboration, we developed a screenable assay for the NPS receptor, and in collaboration with the NIH Chemical Genomics Center (NCGC; now part of NCATS) identified a lead molecule, NCG001865684, that was determined to be brain penetrant upon peripheral administration in rats. In vitro, NCGC00185684 shows biased antagonist properties, and preferentially blocks ERK-phosphorylation over intracellular cAMP- or calcium-responses to NPS. In vivo, pretreatment with NCG001865684 was found to block alcohol-induced ERK-phosphorylation in the rat central amygdala, a region involved in regulation of alcohol intake. NCGC00185684 also decreases operant alcohol self-administration, and lowers alcohol reward as measured using progressive ratio responding. These effects are behaviorally specific, in that they are observed at doses that do not influence locomotor activity or reinstatement responding following extinction. Taken together, these data provide an initial validation of the NPSR as a therapeutic target in alcoholism (Thorsell et al., submitted). Novel Target Discovery: Gene expression profiling and epigenetic modifications In an effort to identify previously unknown candidate drug targets, a series of projects are being carried out looking at changes in gene expression and epigenetic modifications (microRNA and methylation) in the post-dependent rat model. Bioinformatic analysis of the initial runs applying RNAseq, CHIPseq and microRNAseq (miR) has identified several interesting, differentially expressed transcripts as well as several cases of miR-mediated changes in gene expression (10). Figure 4A represents a gene regulatory network altered by alcohol dependence in the mPFC, where brain derived neurotrophic factor (BDNF) is functioning as the node gene. In silico analysis determined that BDNF is targeted by an alcohol dependent deregulated microRNA, miR-206. A follow up study determined the functional role of miR-206 in alcohol dependence. Inhibition of miR-206 in the mPFC decreased alcohol consumption and preference in post-dependent rats but not control rats. In addition, miR-206 was able to bind and inhibit the expression of BDNF (Figure 4B) (Tapocik et al., in preparation). In collaboration with Dr. Paul Kenny, we have determined that miR-206 knockout mice consume and prefer alcohol significantly less than WT mice at pharmacologically active doses of alcohol (Tapocik et al., in preparation). We have also found that DNA methylation in the mPFC is regulated by a history of alcohol dependence, and contributes to the persistent escalation in alcohol consumption observed in post-dependent rats. ICV infusion of a DNA methylation inhibitor normalized both mPFC DNA methylation and alcohol consumption in post-dependent rats. Conversely, systemic injections and direct injections of a methyl donor induced DNA hypermethylation in the mPFC of non-dependent rats and escalated alcohol consumption. The expression of genes encoding synaptic vesicle proteins is altered following a history of alcohol dependence. This was prevented by DNA methylation inhibition, suggesting that DNA methylation regulates behavioral adaptations to chronic alcohol exposure (Barbier et al., submitted). Currently we are inhibiting two synaptic vesicle proteins (Syt1 & Syt2) in the mPFC that may be methylated after alcohol dependence to see if it causes an escalation in alcohol self-administration.
10
. Figure 4. A. Regulatory network of genes altered by alcohol-dependent messenger RNAs (mRNAs) and their targeted microRNAs (miRNAs). B. miR-206 binds to the 3’UTR of BDNF and inhibits BDNF expression. Translation The challenge of translational medicine is to bring preclinically validated targets into clinical development. With a growing list of preclinically validated targets, filters are essential for helping us devote resources to developing treatments with a reasonable likelihood of success. Our basic science laboratory works closely together with other components of the lab to achieve rapid translation of preclinical findings into early human development, as exemplified by our NK1 and CRH1 receptor antagonist work. We also apply “reverse translation”. Frequently, observations in humans can be taken back to animal models in order to obtain a mechanistic understanding, as exemplified by our work with the mouse lines carrying human OPRM1 gene variants, or the detailed analysis of NK1R antagonist actions on alcohol reward, self-administration and relapse. Relevance The strategies, key finding and relevance of our research have been summarized in key recent reviews (11, 12) The arrival of naltrexone and acamprosate as modern medications for alcohol dependence has in fundamental ways changed the field of alcohol research, and provided proof-of-principle for pharmacotherapy of this disease. Their effect sizes are, however, limited in unselected patient populations. It is unlikely that a major change in clinical outcomes in alcoholism will be brought about by the arrival of any single “magic bullet” drug. Instead, we predict that a cumulative effect of a growing and mechanistically diverse group of compounds

11
genetic as well as experiential characteristics of the patient. Our program has the potential to contribute in major ways to this development. Key Prior Reference
Heilig M, Koob GF. 2007. A key role for corticotropin-releasing factor in alcohol dependence. Trends Neurosci 30: 399-406
Publications 2011 – 2012
1. Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, Damadzic R, Eskay R, Schoor M, Thorsell A, Schwandt ML, Sommer WH, George DT, Parsons LH, Herscovitch P, Hommer D, Heilig M. 2011. A genetic determinant of the striatal dopamine response to alcohol in men. Mol Psychiatry 16: 809-17
2. Mahmoud S, Thorsell A, Sommer WH, Heilig M, Holgate JK, Bartlett SE, Ruiz-Velasco V. 2011. Pharmacological consequence of the A118G mu opioid receptor polymorphism on morphine- and fentanyl-mediated modulation of Ca(2)(+) channels in humanized mouse sensory neurons. Anesthesiology 115: 1054-62
3. Karlsson RM, Adermark L, Molander A, Perreau-Lenz S, Singley E, Solomon M, Holmes A, Tanaka K, Lovinger DM, Spanagel R, Heilig M. 2012. Reduced alcohol intake and reward associated with impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology 63: 181-9
4. Molander A, Vengeliene V, Heilig M, Wurst W, Deussing JM, Spanagel R. 2012. Brain-Specific Inactivation of the Crhr1 Gene Inhibits Post-Dependent and Stress-Induced Alcohol Intake, but Does Not Affect Relapse-Like Drinking. Neuropsychopharmacology 37: 1047-56
5. Vendruscolo LF, Barbier E, Schlosburg JE, Misra KK, Whitfield TW, Jr., Logrip ML, Rivier C, Repunte-Canonigo V, Zorrilla EP, Sanna PP, Heilig M, Koob GF. 2012. Corticosteroid-dependent plasticity mediates compulsive alcohol drinking in rats. J Neurosci 32: 7563-71
6. Schank JR, Pickens CL, Rowe KE, Cheng K, Thorsell A, Rice KC, Shaham Y, Heilig M. 2011. Stress-induced reinstatement of alcohol-seeking in rats is selectively suppressed by the neurokinin 1 (NK1) antagonist L822429. Psychopharmacology (Berl) 218: 111-9
7. Robinson JE, Fish EW, Krouse MC, Thorsell A, Heilig M, Malanga CJ. 2012. Potentiation of brain stimulation reward by morphine: effects of neurokinin-1 receptor antagonism. Psychopharmacology (Berl) 220: 215-24
8. Cippitelli A, Rezvani AH, Robinson JE, Eisenberg L, Levin ED, Bonaventure P, Motley ST, Lovenberg TW, Heilig M, Thorsell A. 2011. The novel, selective, brain-penetrant neuropeptide Y Y2 receptor antagonist, JNJ-31020028, tested in animal models of alcohol consumption, relapse, and anxiety. Alcohol 45: 567-76
9. Karlsson C, Zook M, Ciccocioppo R, Gehlert DR, Thorsell A, Heilig M, Cippitelli A. 2012. Melanin-concentrating hormone receptor 1 (MCH1-R) antagonism: reduced

12
appetite for calories and suppression of addictive-like behaviors. Pharmacol Biochem Behav 102: 400-6
10. Tapocik JD, Solomon M, Flanigan M, Meinhardt M, Barbier E, Schank JR, Schwandt M, Sommer WH, Heilig M. 2012. Coordinated dysregulation of mRNAs and microRNAs in the rat medial prefrontal cortex following a history of alcohol dependence. Pharmacogenomics J
11. Heilig M, Goldman D, Berrettini W, O'Brien CP. 2011. Pharmacogenetic approaches to the treatment of alcohol addiction. Nat Rev Neurosci 12: 670-84
12. Schank JR, Ryabinin AE, Giardino WJ, Ciccocioppo R, Heilig M. 2012. Stress-Related Neuropeptides and Addictive Behaviors: Beyond the Usual Suspects. Neuron 76: 192-208
Dr. Heilig: [email protected]

13
______________________________________________________________________
Section on Brain Electrophysiology and Imaging ______________________________________________________________________
Daniel Hommer*, MD, Chief Reza Momenan, PhD, Acting Chief, Staff Scientist Daniel Rio, PhD, Physicist Mike Kerich, BS, Program Analyst Betsy Davis, Psychology Technician Vladimir Senatorov, MD, PhD, Postdoctoral IRTA Xi Zhu, PhD, Postdoctoral IRTA Caitlin Durkee, Postbac IRTA Claire Mann, Postbac IRTA Jennifer Liu, Postbac IRTA Rabina Joshi, Postbac Technical IRTA Structural Imaging This research is designed to determine in more detail the association of brain structure with alcoholism and the risk for alcoholism. During the past 15 years we have collected full, volumetric T-1 weighted MR images of the brain using 1.5 T and 3 T scanners to image the brain in nearly 400 alcoholics (70% male) and 210 healthy, non-alcoholic comparison subjects (60% male). Past work in the BEI section has demonstrated that alcoholics show greater brain degeneration than non-alcoholics. Alcoholic women are more affected than alcoholic men and significant differences in brain shrinkage between alcoholics and controls can be detected by the age of 25 to 30 years. Based on an automated segmentation program developed in our section, the intracranial contents are divided into CSF, gray and white matter. Our measurement techniques allow us to independently estimate overall brain growth and brain degeneration. Growth determines maximum brain size achieved during life and can be accurately estimated by intracranial volume (ICV) since ICV remains constant throughout adult life. Brain growth drives expansion of ICV, and once brain growth ceases during mid-adolescence, ICV does not change. As the brain shrinks with aging or pathological processes, degeneration can be measured by the ratio of cerebral volume (or gray matter or white matter volume) to ICV. Using the extensions of the same methods we have investigated the effects of substance-dependence on the volume of medial and orbital frontal cortex, as well as the entire striatum. These regions are known to be involved in motivation, social behavior and decision making. The structural findings may relate to functional pathology, since we have previously found reduced posterior medial frontal cortex activation by risky rewards in alcohol-dependent patients. Over the past two years we have made use of several methodological advances in the automated measurement and analysis of brain volumes. An automated method for dividing the brain into right and left hemispheres was developed and validated. We have also utilized emerging methods in the field to study other aspects of structural images such as cortical thickness analysis, voxel based morphometry (VBM), and diffusion tensor imaging (DTI) tractography, a measure of structural connectivity.
*Dr. Hommer tragically passed away January 2, 2013. Dan is enormously missed by his colleagues at NIAAA.
14
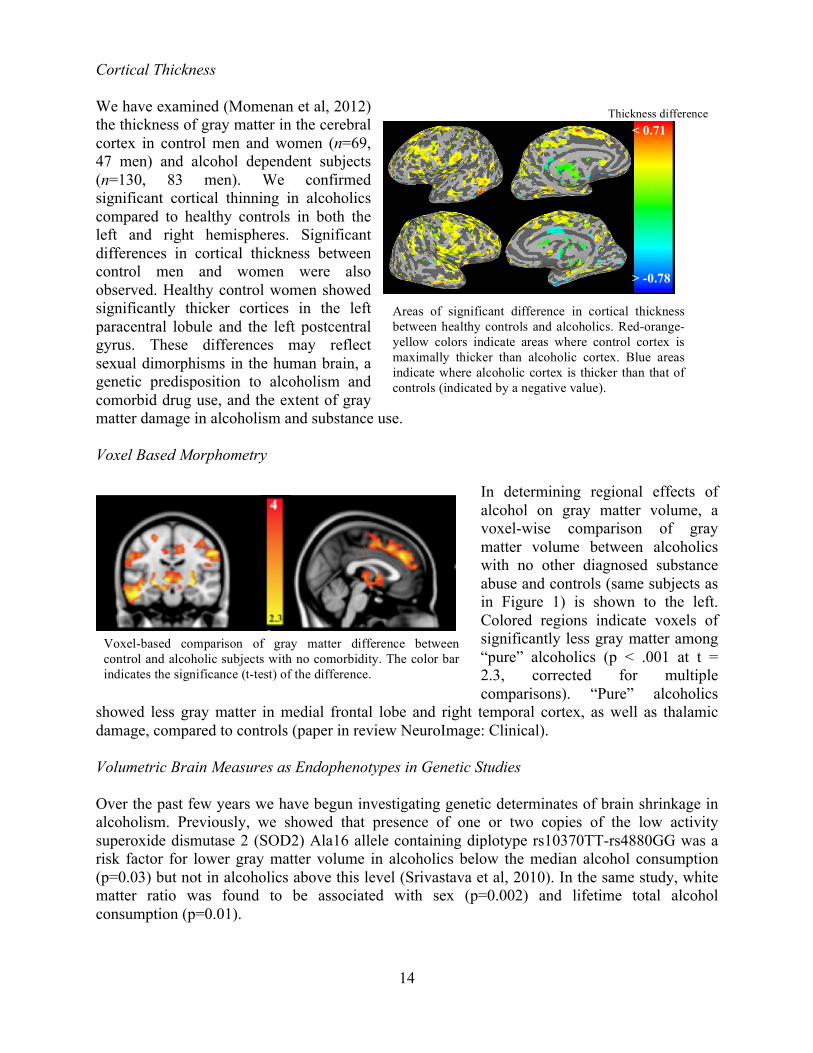
Cortical Thickness We have examined (Momenan et al, 2012) the thickness of gray matter in the cerebral cortex in control men and women (n=69, 47 men) and alcohol dependent subjects (n=130, 83 men). We confirmed significant cortical thinning in alcoholics compared to healthy controls in both the left and right hemispheres. Significant differences in cortical thickness between control men and women were also observed. Healthy control women showed significantly thicker cortices in the left paracentral lobule and the left postcentral gyrus. These differences may reflect sexual dimorphisms in the human brain, a genetic predisposition to alcoholism and comorbid drug use, and the extent of gray matter damage in alcoholism and substance use. Voxel Based Morphometry
In determining regional effects of alcohol on gray matter volume, a voxel-wise comparison of gray matter volume between alcoholics with no other diagnosed substance abuse and controls (same subjects as in Figure 1) is shown to the left. Colored regions indicate voxels of significantly less gray matter among “pure” alcoholics (p < .001 at t = 2.3, corrected for multiple comparisons). “Pure” alcoholics
showed less gray matter in medial frontal lobe and right temporal cortex, as well as thalamic damage, compared to controls (paper in review NeuroImage: Clinical). Volumetric Brain Measures as Endophenotypes in Genetic Studies Over the past few years we have begun investigating genetic determinates of brain shrinkage in alcoholism. Previously, we showed that presence of one or two copies of the low activity superoxide dismutase 2 (SOD2) Ala16 allele containing diplotype rs10370TT-rs4880GG was a risk factor for lower gray matter volume in alcoholics below the median alcohol consumption (p=0.03) but not in alcoholics above this level (Srivastava et al, 2010). In the same study, white matter ratio was found to be associated with sex (p=0.002) and lifetime total alcohol consumption (p=0.01).
Thickness difference < 0.71
> -0.78
Areas of significant difference in cortical thickness between healthy controls and alcoholics. Red-orange-yellow colors indicate areas where control cortex is maximally thicker than alcoholic cortex. Blue areas indicate where alcoholic cortex is thicker than that of
controls (indicated by a negative value).
Voxel-based comparison of gray matter difference between control and alcoholic subjects with no comorbidity. The color bar indicates the significance (t-test) of the difference.

15
We are currently studying the influence of catechol-O-methyltransferase (COMT) Val158 Met polymorphism on volumetric measures of different brain structures in alcohol dependent patients. Preliminary findings indicate that compared to the Val/Val or Met/Met genotype, female alcoholics with the Val/Met genotype have significantly smaller gray matter volume, particularly in both right and left putamen and right nucleus accumbens (right). In agreement with this observation, female alcoholics with Val/Met showed significantly larger CSF volume. Although volumetric measures are a powerful category of endophenotypes that may allow detection of genetic effects in small samples, these findings await replication.
More recently, preliminary results show that AA homozygotes (rs6198) and G allele carriers had similar hippocampal volumes, but among those that had experienced emotional mistreatment, G allele carriers had larger hippocampal volumes than AA homozygotes (left).
Functional Imaging Our research is focused on two aspects of brain function that are critically important for addictive disorders: motivation and emotion. Alcoholism is characterized by changes in a person’s motivation such that previously motivating social or personal goals are replaced by the desire to obtain and consume alcohol. Alcoholics also suffer from higher levels of negative emotionality than non-alcoholics, and aversive stimuli that provoke negative emotionality are established triggers of relapse. Motivation We have used event-related functional magnetic resonance imaging (fMRI) to examine how the brain responds during the anticipation of working to gain reward or to avoid punishment. This has been done using a modified version of risk-taking task by (Matthews et al, 2004). (3). Thirty-two healthy adult volunteers (27.0 ± 4.1 years old, 15 males) were scanned while playing this task. The risk-taking task required participants to choose between a safe choice where they were guaranteed to win $0.25, and a risky choice which presented the possibility of winning $1 or $5, but also the risk of losing those amounts. Each trial of the task consisted of four events: 1) cue presentation, 2) choice, 3) a delay, and 4) notification of the outcome. Our analyses in this report focus exclusively on the notification of outcome.
Met/Met Val/Met Val/Val Rs4680
Mean (Gray/Total) vs. rs4680
[TG
ray/
Tota
l
Male (n = 163) Female (n = 88)
16
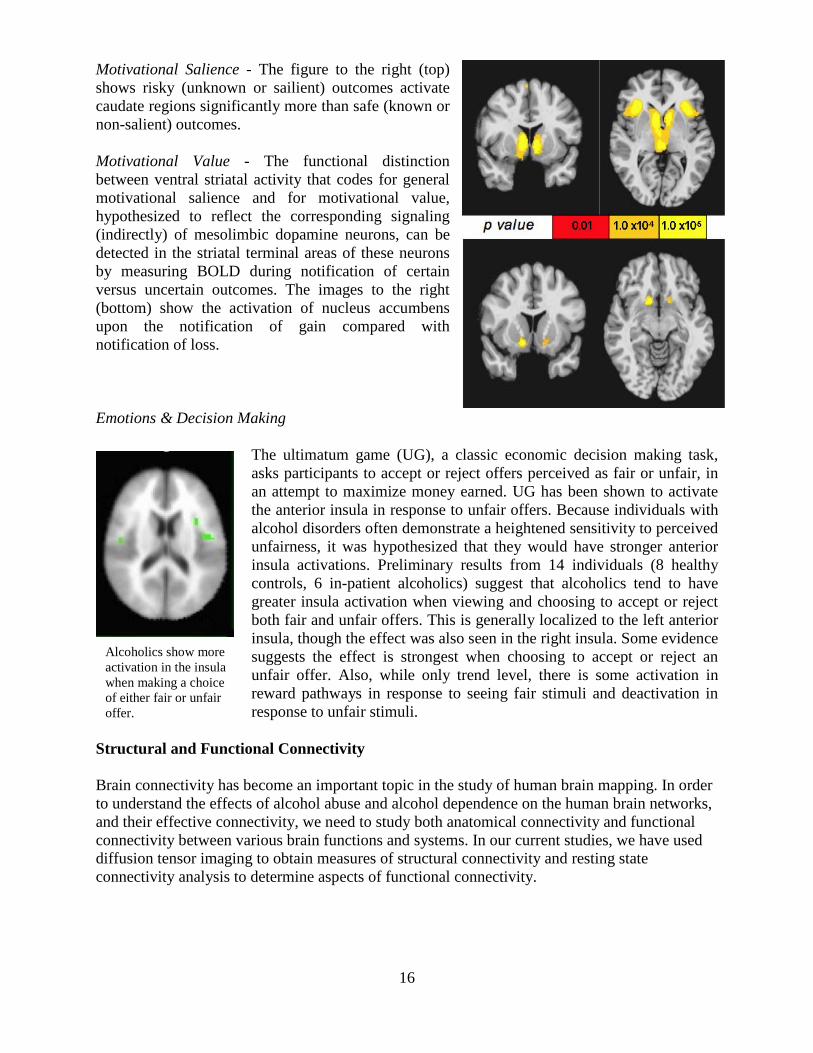
Motivational Salience - The figure to the right (top) shows risky (unknown or sailient) outcomes activate caudate regions significantly more than safe (known or non-salient) outcomes. Motivational Value - The functional distinction between ventral striatal activity that codes for general motivational salience and for motivational value, hypothesized to reflect the corresponding signaling (indirectly) of mesolimbic dopamine neurons, can be detected in the striatal terminal areas of these neurons by measuring BOLD during notification of certain versus uncertain outcomes. The images to the right (bottom) show the activation of nucleus accumbens upon the notification of gain compared with notification of loss. Emotions & Decision Making
The ultimatum game (UG), a classic economic decision making task, asks participants to accept or reject offers perceived as fair or unfair, in an attempt to maximize money earned. UG has been shown to activate the anterior insula in response to unfair offers. Because individuals with alcohol disorders often demonstrate a heightened sensitivity to perceived unfairness, it was hypothesized that they would have stronger anterior insula activations. Preliminary results from 14 individuals (8 healthy controls, 6 in-patient alcoholics) suggest that alcoholics tend to have greater insula activation when viewing and choosing to accept or reject both fair and unfair offers. This is generally localized to the left anterior insula, though the effect was also seen in the right insula. Some evidence suggests the effect is strongest when choosing to accept or reject an unfair offer. Also, while only trend level, there is some activation in reward pathways in response to seeing fair stimuli and deactivation in response to unfair stimuli.
Structural and Functional Connectivity Brain connectivity has become an important topic in the study of human brain mapping. In order to understand the effects of alcohol abuse and alcohol dependence on the human brain networks, and their effective connectivity, we need to study both anatomical connectivity and functional connectivity between various brain functions and systems. In our current studies, we have used diffusion tensor imaging to obtain measures of structural connectivity and resting state connectivity analysis to determine aspects of functional connectivity.
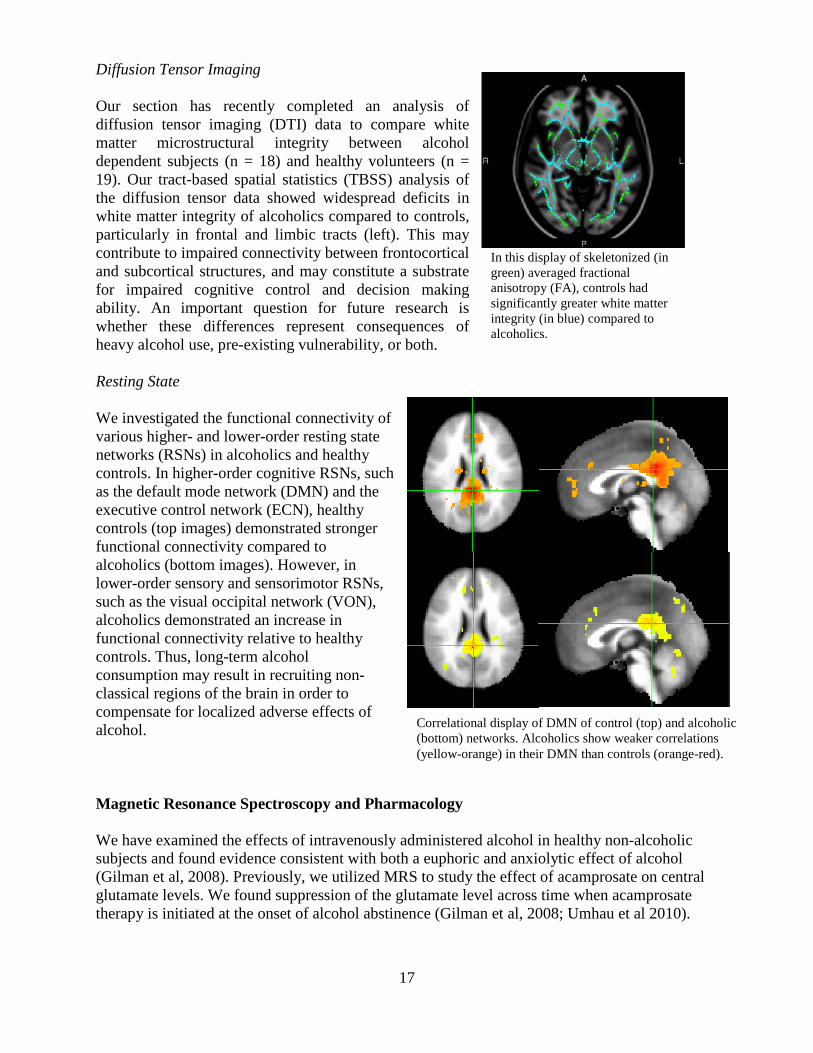
Alcoholics show more activation in the insula when making a choice of either fair or unfair offer.
17
Diffusion Tensor Imaging Our section has recently completed an analysis of diffusion tensor imaging (DTI) data to compare white matter microstructural integrity between alcohol dependent subjects (n = 18) and healthy volunteers (n = 19). Our tract-based spatial statistics (TBSS) analysis of the diffusion tensor data showed widespread deficits in white matter integrity of alcoholics compared to controls, particularly in frontal and limbic tracts (left). This may contribute to impaired connectivity between frontocortical and subcortical structures, and may constitute a substrate for impaired cognitive control and decision making ability. An important question for future research is whether these differences represent consequences of heavy alcohol use, pre-existing vulnerability, or both. Resting State We investigated the functional connectivity of various higher- and lower-order resting state networks (RSNs) in alcoholics and healthy controls. In higher-order cognitive RSNs, such as the default mode network (DMN) and the executive control network (ECN), healthy controls (top images) demonstrated stronger functional connectivity compared to alcoholics (bottom images). However, in lower-order sensory and sensorimotor RSNs, such as the visual occipital network (VON), alcoholics demonstrated an increase in functional connectivity relative to healthy controls. Thus, long-term alcohol consumption may result in recruiting non-classical regions of the brain in order to compensate for localized adverse effects of alcohol. Magnetic Resonance Spectroscopy and Pharmacology We have examined the effects of intravenously administered alcohol in healthy non-alcoholic subjects and found evidence consistent with both a euphoric and anxiolytic effect of alcohol (Gilman et al, 2008). Previously, we utilized MRS to study the effect of acamprosate on central glutamate levels. We found suppression of the glutamate level across time when acamprosate therapy is initiated at the onset of alcohol abstinence (Gilman et al, 2008; Umhau et al 2010).
In this display of skeletonized (in green) averaged fractional anisotropy (FA), controls had significantly greater white matter integrity (in blue) compared to alcoholics.
Correlational display of DMN of control (top) and alcoholic (bottom) networks. Alcoholics show weaker correlations (yellow-orange) in their DMN than controls (orange-red).

18
Ethanol Peak
We now are in the process of determining the effect of acute alcohol infusion on brain metabolite levels as well as determining the rate of absorption of alcohol in light social drinkers in contrast to heavy drinkers. The above figures are sample MRS spectra of a 25x25x25 mm3 single voxel in ACC before (left) and after (right) ethanol infusion. References Gilman JM, Hommer DW. 2008. Modulation of brain response to emotional images by alcohol cues in alcohol-dependent patients. Addiction Biology 13: 423-34 Matthews SC, Simmons AN, Lane SD, Paulus MP. 2004. Selective activation of the nucleus accumbens during risk-taking decision making. Neuroreport 15: 2123-7 Momenan R, Steckler L.E., Saad Z., van Rafelghem S., Kerich, M., and Hommer, D.W. 2012. Effects of alcohol dependence on cortical thickness. Psychiatry Research: Neuroimaging 204: 101-11 Srivastava V, Buzas B, Momenan R, Oroszi G, Pulay AJ, Enoch MA, Hommer DW, Goldman D. 2010. Association of SOD2, a mitochondrial antioxidant enzyme, with gray matter volume shrinkage in alcoholics. Neuropsychopharmacology 35: 1120-8 Umhau JC, Momenan R, Schwandt ML, Singley E, Lifshitz M, Doty L, Adams LJ, Vengeliene V, Spanagel R, Zhang Y, Shen J, George DT, Hommer D, Heilig M. 2010. Effect of acamprosate on magnetic resonance spectroscopy measures of central glutamate in detoxified alcohol-dependent individuals: a randomized controlled experimental medicine study. Archives of General Psychiatry 67: 1069-77 Publications 2011-2012 1. Bjork JM, Chen G, Hommer DW. 2012. Psychopathic tendencies and mesolimbic
recruitment by cues for instrumental and passively obtained rewards. Biol Psychol 89: 408-15
2. Bjork JM, Smith AR, Chen G, Hommer DW. 2012. Mesolimbic recruitment by nondrug rewards in detoxified alcoholics: Effort anticipation, reward anticipation, and reward delivery. Human Brain Mapping 33: 2174-88

19
3. Chen CH, Walker J, Momenan R, Rawlings R, Heilig M, Hommer DW. 2012. Relationship Between Liver Function and Brain Shrinkage in Patients with Alcohol Dependence. Alcoholism-Clinical and Experimental Research 36: 625-32
4. Gilman JM, Ramchandani VA, Crouss T, Hommer DW. 2012. Subjective and Neural Responses to Intravenous Alcohol in Young Adults with Light and Heavy Drinking Patterns. Neuropsychopharmacology 37: 467-77
5. Gilman JM, Smith AR, Ramchandani VA, Momenan R, Hommer DW. 2012. The effect of intravenous alcohol on the neural correlates of risky decision making in healthy social drinkers. Addiction Biology 17: 465-78
6. Momenan R, Steckler LE, Saad ZS, van Rafelghem S, Kerich MJ, Hommer DW. 2012. Effects of alcohol dependence on cortical thickness as determined by magnetic resonance imaging. Psychiatry Research-Neuroimaging 204: 101-11
7. Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, Damadzic R, Eskay R, Schoor M, Thorsell A, Schwandt ML, Sommer WH, George DT, Parsons LH, Herscovitch P, Hommer D, Heilig M. 2011. A genetic determinant of the striatal dopamine response to alcohol in men. Molecular Psychiatry 16: 809-17
8. Vatsalya V, Issa JE, Hommer DW, Ramchandani VA. 2012. Pharmacodynamic Effects of Intravenous Alcohol on Hepatic and Gonadal Hormones: Influence of Age and Sex. Alcoholism-Clinical and Experimental Research 36: 207-13
9. Zhang L, Kerich M, Schwandt ML, Rawlings RR, McKellar JD, Momenan R, Hommer DW, George DT. 2011. Smaller right amygdala in Caucasian alcohol-dependent male patients with a history of intimate partner violence: a volumetric imaging study. Addiction Biology DOI: 10.1111/j.1369-1600.2011.00381.x
Dr. Momenan: [email protected]

20
______________________________________________________________________
Section on Clinical Assessment and Treatment Evaluation ______________________________________________________________________ David Ted George, MD, Chief Victoria Brown, BA, Postbac IRTA Kristie Diamond, MSN, CRNP Debra Hill, MSW, Social Worker Cheryl Jones, BA, Psychologist Laura Kwako, PhD Postdoctoral IRTA Vidda Moussavi, BS, Postbac IRTA Monte Phillips, BS, Biologist Vera Spagnola, MD Postdoctoral IRTA John Umhau, MD, MPH, Clinical Fellow Jennifer Warmingham, BA, Postbac IRTA Mission The Section of Clinical Assessment and Treatment Evaluation (SCATE) is the clinical component of the LCTS. It combines the provision of state-of-the art assessment and treatment of patients with alcohol use disorders, with the execution of experimental medicine studies. The section has developed a unique approach in which experimental therapeutics are evaluated in the short term under closely controlled inpatient conditions. Behavioral, neuroendocrine and imaging based readouts are used to obtain surrogate markers of potential clinical efficacy, or to further the understanding of pathophysiological mechanisms in addiction and related disorders. Experimental medicine studies of current therapeutic mechanisms One goal of the section has been to study the effects of pharmacological interventions on alcohol craving and other surrogate markers under carefully controlled conditions. These studies allow mechanisms of action to be examined. Currently approved medications for the treatment of alcoholism, acamprosate and naltrexone, have documented albeit modest efficacy overall, but their mechanisms of action in alcohol dependent patients is not clear. Effect of acamprosate on alcohol craving induced by pharmacological stressors: yohimbine and meta-chlorophenylpiperazine (mCPP) A potentially attractive approach to inducing craving responses is through administration of the α2-adrenergic antagonist yohimbine. In rats, yohimbine acts as pharmacological stressor that reinstates alcohol seeking and leads to escalation of voluntary alcohol intake. We therefore evaluated whether yohimbine administration would increase alcohol craving in alcoholics. In an attempt to assess the predictive validity of this approach, the approved alcoholism medication acamprosate was also evaluated for its ability to modulate yohimbine-induced cravings. For comparison, mCPP was also used as a pharmacological challenge.
Treatment-seeking alcohol dependent inpatients in early abstinence were randomized to parallel arms that received placebo or acamprosate (2997 mg daily). Following two weeks of medication, subjects within both arms underwent three challenge sessions with yohimbine, mCPP or saline
21
infusion under double-blind conditions, carried out in counterbalanced order, and separated by at least 5 days. Ratings of craving and anxiety, as well as biochemical measures were obtained.
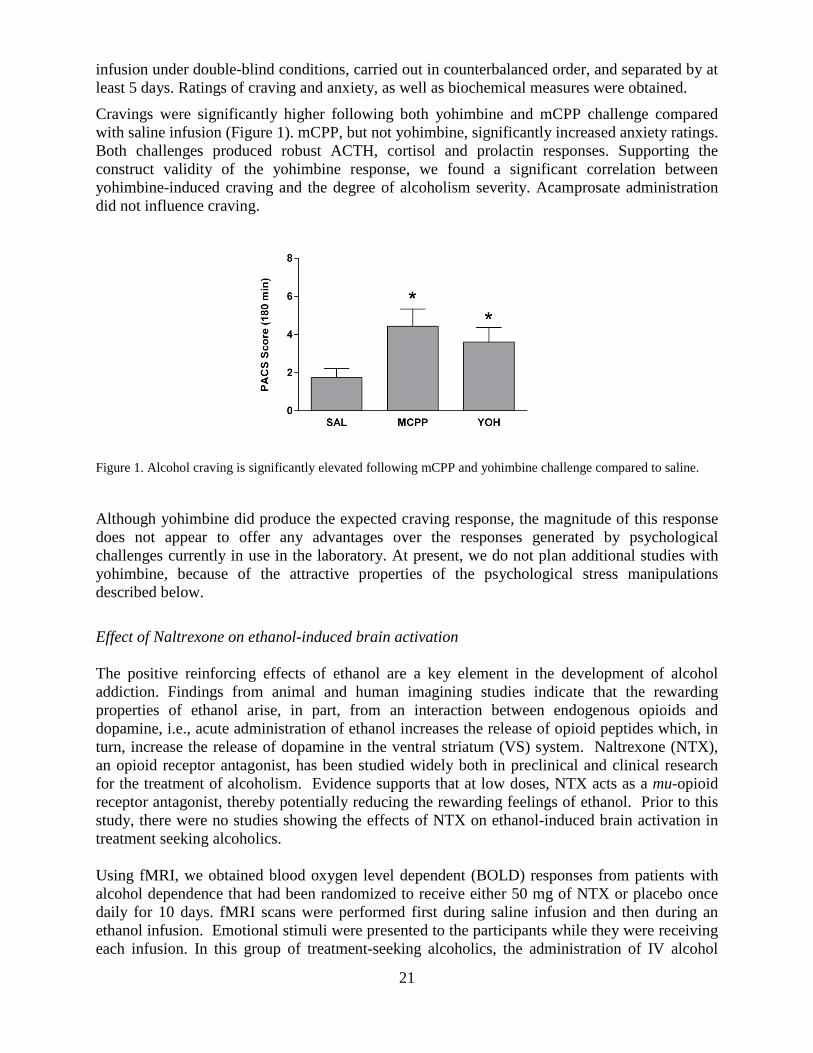
Cravings were significantly higher following both yohimbine and mCPP challenge compared with saline infusion (Figure 1). mCPP, but not yohimbine, significantly increased anxiety ratings. Both challenges produced robust ACTH, cortisol and prolactin responses. Supporting the construct validity of the yohimbine response, we found a significant correlation between yohimbine-induced craving and the degree of alcoholism severity. Acamprosate administration did not influence craving.
Figure 1. Alcohol craving is significantly elevated following mCPP and yohimbine challenge compared to saline.
Although yohimbine did produce the expected craving response, the magnitude of this response does not appear to offer any advantages over the responses generated by psychologicalchallenges currently in use in the laboratory. At present, we do not plan additional studies with yohimbine, because of the attractive properties of the psychological stress manipulationsdescribed below.
Effect of Naltrexone on ethanol-induced brain activation The positive reinforcing effects of ethanol are a key element in the development of alcohol addiction. Findings from animal and human imagining studies indicate that the rewarding properties of ethanol arise, in part, from an interaction between endogenous opioids and dopamine, i.e., acute administration of ethanol increases the release of opioid peptides which, in turn, increase the release of dopamine in the ventral striatum (VS) system. Naltrexone (NTX), an opioid receptor antagonist, has been studied widely both in preclinical and clinical research for the treatment of alcoholism. Evidence supports that at low doses, NTX acts as a mu-opioid receptor antagonist, thereby potentially reducing the rewarding feelings of ethanol. Prior to this study, there were no studies showing the effects of NTX on ethanol-induced brain activation in treatment seeking alcoholics. Using fMRI, we obtained blood oxygen level dependent (BOLD) responses from patients with alcohol dependence that had been randomized to receive either 50 mg of NTX or placebo once daily for 10 days. fMRI scans were performed first during saline infusion and then during an ethanol infusion. Emotional stimuli were presented to the participants while they were receiving each infusion. In this group of treatment-seeking alcoholics, the administration of IV alcohol
22
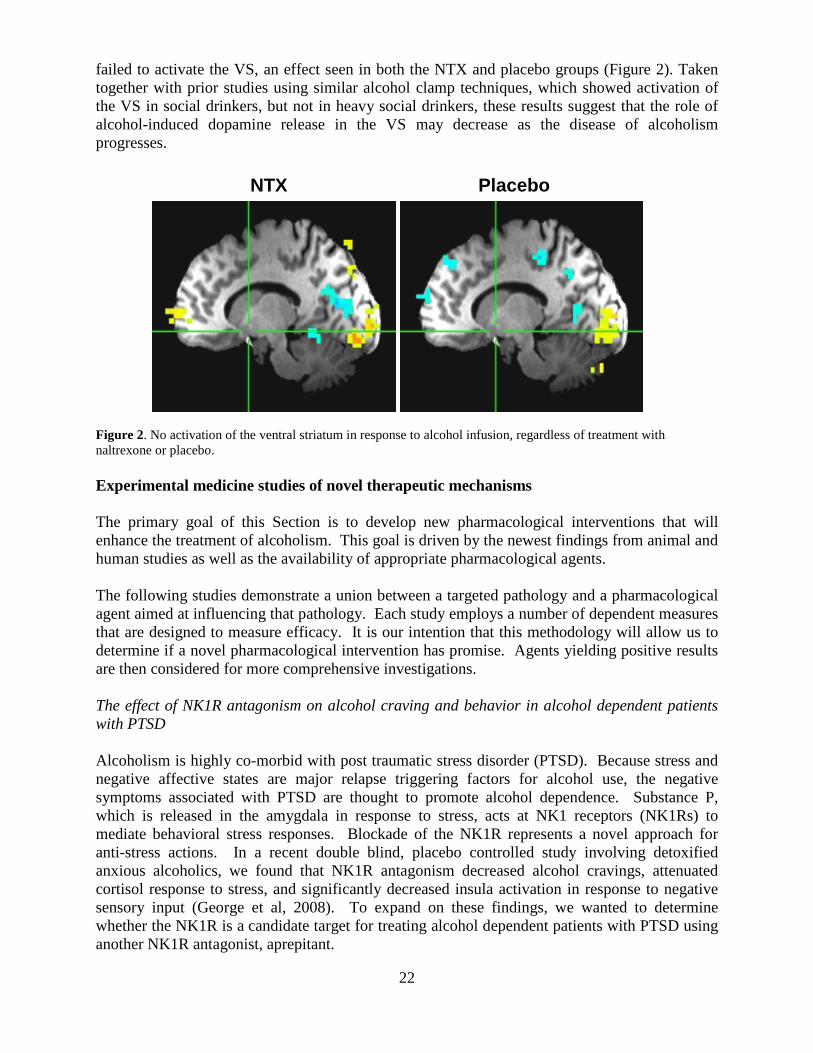
failed to activate the VS, an effect seen in both the NTX and placebo groups (Figure 2). Taken together with prior studies using similar alcohol clamp techniques, which showed activation of the VS in social drinkers, but not in heavy social drinkers, these results suggest that the role of alcohol-induced dopamine release in the VS may decrease as the disease of alcoholism progresses. Figure 2. No activation of the ventral striatum in response to alcohol infusion, regardless of treatment with naltrexone or placebo. Experimental medicine studies of novel therapeutic mechanisms The primary goal of this Section is to develop new pharmacological interventions that will enhance the treatment of alcoholism. This goal is driven by the newest findings from animal and human studies as well as the availability of appropriate pharmacological agents. The following studies demonstrate a union between a targeted pathology and a pharmacological agent aimed at influencing that pathology. Each study employs a number of dependent measures that are designed to measure efficacy. It is our intention that this methodology will allow us to determine if a novel pharmacological intervention has promise. Agents yielding positive results are then considered for more comprehensive investigations. The effect of NK1R antagonism on alcohol craving and behavior in alcohol dependent patients with PTSD Alcoholism is highly co-morbid with post traumatic stress disorder (PTSD). Because stress and negative affective states are major relapse triggering factors for alcohol use, the negative symptoms associated with PTSD are thought to promote alcohol dependence. Substance P, which is released in the amygdala in response to stress, acts at NK1 receptors (NK1Rs) to mediate behavioral stress responses. Blockade of the NK1R represents a novel approach for anti-stress actions. In a recent double blind, placebo controlled study involving detoxified anxious alcoholics, we found that NK1R antagonism decreased alcohol cravings, attenuated cortisol response to stress, and significantly decreased insula activation in response to negative sensory input (George et al, 2008). To expand on these findings, we wanted to determine whether the NK1R is a candidate target for treating alcohol dependent patients with PTSD using another NK1R antagonist, aprepitant.
NTX Placebo
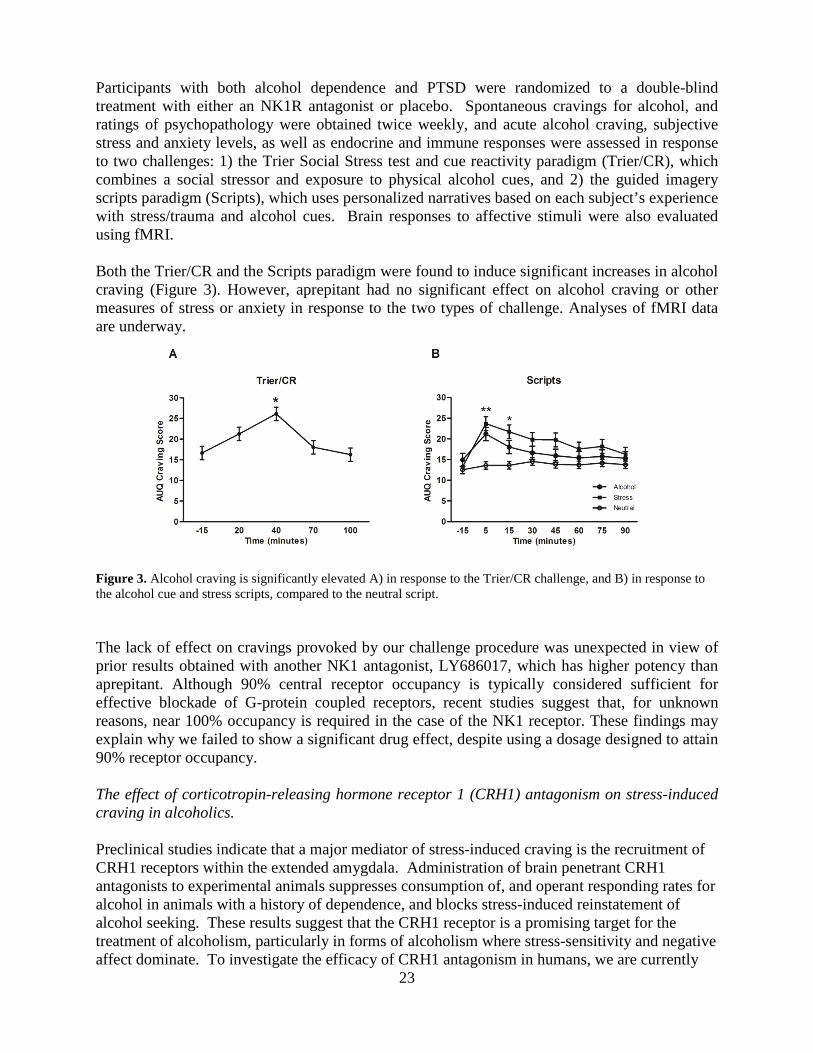
23
Participants with both alcohol dependence and PTSD were randomized to a double-blind treatment with either an NK1R antagonist or placebo. Spontaneous cravings for alcohol, and ratings of psychopathology were obtained twice weekly, and acute alcohol craving, subjective stress and anxiety levels, as well as endocrine and immune responses were assessed in response to two challenges: 1) the Trier Social Stress test and cue reactivity paradigm (Trier/CR), which combines a social stressor and exposure to physical alcohol cues, and 2) the guided imagery scripts paradigm (Scripts), which uses personalized narratives based on each subject’s experience with stress/trauma and alcohol cues. Brain responses to affective stimuli were also evaluated using fMRI. Both the Trier/CR and the Scripts paradigm were found to induce significant increases in alcohol craving (Figure 3). However, aprepitant had no significant effect on alcohol craving or other measures of stress or anxiety in response to the two types of challenge. Analyses of fMRI data are underway. Figure 3. Alcohol craving is significantly elevated A) in response to the Trier/CR challenge, and B) in response to the alcohol cue and stress scripts, compared to the neutral script. The lack of effect on cravings provoked by our challenge procedure was unexpected in view of prior results obtained with another NK1 antagonist, LY686017, which has higher potency than aprepitant. Although 90% central receptor occupancy is typically considered sufficient for effective blockade of G-protein coupled receptors, recent studies suggest that, for unknown reasons, near 100% occupancy is required in the case of the NK1 receptor. These findings may explain why we failed to show a significant drug effect, despite using a dosage designed to attain 90% receptor occupancy. The effect of corticotropin-releasing hormone receptor 1 (CRH1) antagonism on stress-induced craving in alcoholics. Preclinical studies indicate that a major mediator of stress-induced craving is the recruitment of CRH1 receptors within the extended amygdala. Administration of brain penetrant CRH1 antagonists to experimental animals suppresses consumption of, and operant responding rates for alcohol in animals with a history of dependence, and blocks stress-induced reinstatement of alcohol seeking. These results suggest that the CRH1 receptor is a promising target for the treatment of alcoholism, particularly in forms of alcoholism where stress-sensitivity and negative affect dominate. To investigate the efficacy of CRH1 antagonism in humans, we are currently
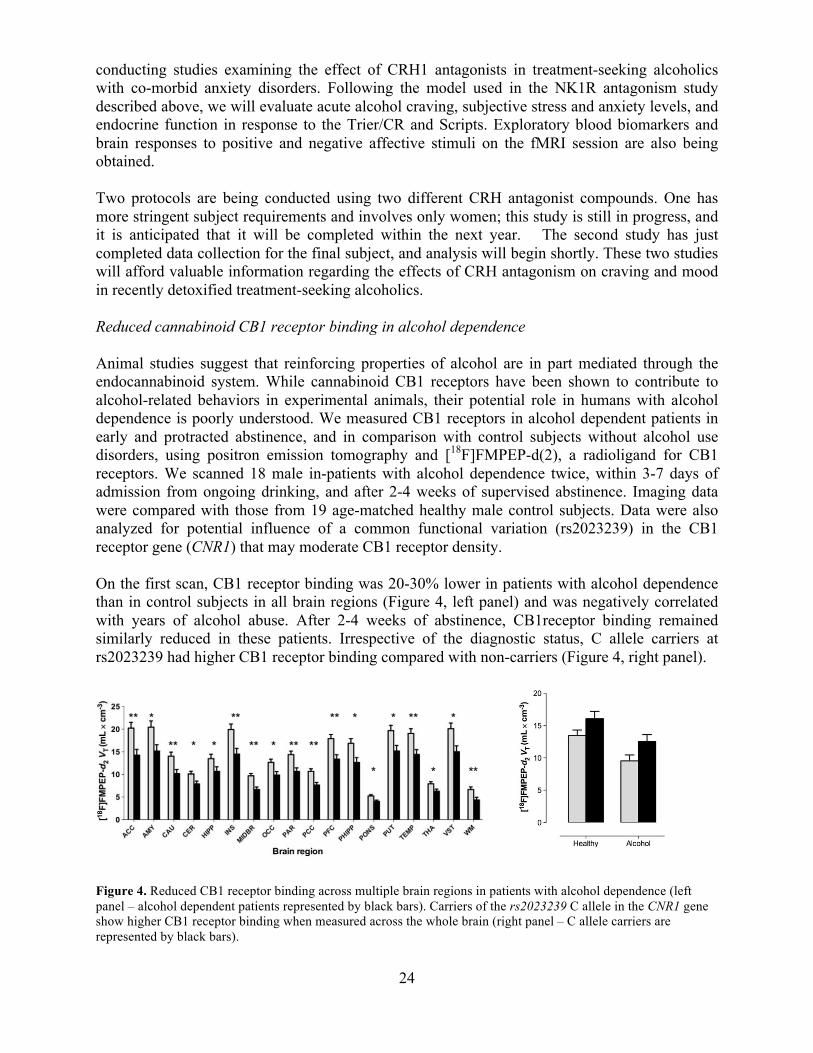
24
conducting studies examining the effect of CRH1 antagonists in treatment-seeking alcoholics with co-morbid anxiety disorders. Following the model used in the NK1R antagonism study described above, we will evaluate acute alcohol craving, subjective stress and anxiety levels, and endocrine function in response to the Trier/CR and Scripts. Exploratory blood biomarkers and brain responses to positive and negative affective stimuli on the fMRI session are also being obtained. Two protocols are being conducted using two different CRH antagonist compounds. One has more stringent subject requirements and involves only women; this study is still in progress, and it is anticipated that it will be completed within the next year. The second study has just completed data collection for the final subject, and analysis will begin shortly. These two studies will afford valuable information regarding the effects of CRH antagonism on craving and mood in recently detoxified treatment-seeking alcoholics. Reduced cannabinoid CB1 receptor binding in alcohol dependence Animal studies suggest that reinforcing properties of alcohol are in part mediated through the endocannabinoid system. While cannabinoid CB1 receptors have been shown to contribute to alcohol-related behaviors in experimental animals, their potential role in humans with alcohol dependence is poorly understood. We measured CB1 receptors in alcohol dependent patients in early and protracted abstinence, and in comparison with control subjects without alcohol use disorders, using positron emission tomography and [18F]FMPEP-d(2), a radioligand for CB1 receptors. We scanned 18 male in-patients with alcohol dependence twice, within 3-7 days of admission from ongoing drinking, and after 2-4 weeks of supervised abstinence. Imaging data were compared with those from 19 age-matched healthy male control subjects. Data were also analyzed for potential influence of a common functional variation (rs2023239) in the CB1 receptor gene (CNR1) that may moderate CB1 receptor density. On the first scan, CB1 receptor binding was 20-30% lower in patients with alcohol dependence than in control subjects in all brain regions (Figure 4, left panel) and was negatively correlated with years of alcohol abuse. After 2-4 weeks of abstinence, CB1receptor binding remained similarly reduced in these patients. Irrespective of the diagnostic status, C allele carriers at rs2023239 had higher CB1 receptor binding compared with non-carriers (Figure 4, right panel).
Figure 4. Reduced CB1 receptor binding across multiple brain regions in patients with alcohol dependence (left panel – alcohol dependent patients represented by black bars). Carriers of the rs2023239 C allele in the CNR1 gene show higher CB1 receptor binding when measured across the whole brain (right panel – C allele carriers are represented by black bars).

25
In a previous study, we had shown that the CB1 receptor inverse agonist rimonabant was not effective when evaluated in chronic alcohol dependence. The current findings suggest that CB1 receptors may have different roles in early vs late phases of alcohol dependence, suggesting that enhanced, rather than blocked CB1 signaling may be beneficial in late stage, treatment seeking alcohol dependent patients. Role of Proinflammatory Signaling in Alcohol Craving Activation of the innate immune system via alcohol consumption is thought to have a role in the pathogenesis of alcoholism. Proinflammatory cytokines, such as IL1β, IL6 and TNF α are produced by activated peripheral macrophages and Kupffer cells. Once in the central nervous system, these cytokines give rise to local production of proinflammatory mediators by microglia as well as activation of the hypothalamic pituitary adrenal axis. Activation of the transcription factor NFκB is an important part of the cascade, since it is induced by IL1β and in turn leads to increased transcription for IL1β itself. NFκB transcription can be opposed by activation of gamma peroxisome proliferator-activated receptors (PPARγ). PPARγ are activated as part of the neuroprotective responses to excitotoxic processes and inflammatory damage. For this reason,the PPARγ has emerged as a candidate target for addiction pharmacotherapy. To determine the role of proinflammatory signaling in alcohol craving, the PPARγ agonist pioglitazone, which modulates glial activity, is evaluated as an experimental treatment. Guided imagery auditory scripts are used as an established set of stimuli to induce craving. Low dose lipopolysaccharide (LPS) administration which activates proinflammatory signaling is used as a novel challenge, and is evaluated for its ability to provoke alcohol craving. If LPS in fact induces alcohol craving, the present design will allow evaluation of whether pioglitazone can inhibit this response. A lumbar puncture is performed and cerebrospinal fluid obtained to determine the effect of pioglitazone on levels of proinflammatory cytokines. Neuroendocrine, psychological and physiological measures are collected for exploratory purposes. An fMRI scan is obtained to evaluate the effect of pioglitazone on BOLD signal in response to emotionally salient visual cues. Pilot study of deep brain stimulation for refractory alcoholism Current treatments for alcoholism utilizing psychological, behavioral and pharmacological approaches often have limited success, with estimated relapse rates ranging from 40% to 70% depending on the study population. More effective interventions are needed for patients with alcoholism that are resistant to these traditional forms of treatment. Preclinical studies indicate that deep brain stimulation (DBS) may have efficacy for the treatment of substance abuse disorders, and preliminary data from human case studies support the efficacy of DBS to treat patients with alcoholism. We are conducting a pilot clinical study to test the safety and efficacy of deep brain stimulation (DBS) of the nucleus accumbens, ventral striatum, and central capsule (VC/CS) in patients with treatment-resistant alcoholism. Following neurosurgical implantation of the DBS in the VC/VS, but prior to initiation of the titration phase, baseline cognitive and behavioral testing and two positron emission tomography scans (i.e. FDG and raclopride) will be performed. Approximately four weeks following placement of the electrodes, a randomized, sham-controlled trial with an open label extension will be instituted whereby participants will be randomized and blinded to undergo either titration for two weeks followed by 24 weeks with DBS system ON, or titration followed by 24 weeks with DBS system OFF.

26
After 24 weeks with DBS ON or OFF, the label extension phase will occur. All participants will be readmitted to the hospital for the second two-week titration period. The cognitive and behavioral assessments performed at baseline and the PET scans will be repeated. Following the inpatient titration phase, they will be discharged from the hospital and followed in the outpatient clinic for 24 weeks with the DBS ON. Cognitive and behavioral assessments which were performed at baseline will be repeated at the end of this 24 week period. Participants will be followed monthly after this time for 9 months, for a total of 24 months of active enrollment.
Reference
George DT, Gilman J, Hersh J, Thorsell A, Herion D, Geyer C, Peng X, Kielbasa W, Rawlings R, Brandt JE, Gehlert DR, Tauscher JT, Hunt SP, Hommer D, Heilig M. 2008. Neurokinin 1 receptor antagonism as a possible therapy for alcoholism. Science 319: 1536-1539.
Publications 2011-2012 1. Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, Damadzic R,
Eskay R, Schoor M, Thorsell A, Schwandt ML, Sommer WH, George DT, Parsons LH, Herscovitch P, Hommer D, Heilig M. 2011. A genetic determinant of the striatal dopamine response to alcohol. Molecular Psychiatry, 16:809-817.
2. George DT, Phillips MJ, Lifshitz M, Lionetti TA, Spero DE, Ghassemzedeh N, Doty L,
Umhau JC, Rawlings RR. 2011. Fluoxetine treatment of alcoholic perpetrators of domestic violence: a 12-week, double-blind, randomized, placebo-controlled intervention study. J Clin Psychiatry, 72:60-5.
3. Umhau JC, Schwandt ML, Usala J, Geyer C, Singley E, George DT, Heilig M. 2011.
Pharmacologically induced alcohol craving in treatment seeking alcoholics correlates with alcoholism severity, but is insensitive to acamprosate. Neuropsychopharmacology, 36:1178-1186.
4. Snell LD, Ramchandani VA, Herion D, Heilig M, George DT, Pridzun L, Helander A,
Saba L, Schwandt ML, Phillips M, Tabakoff B. 2012. The Biometric Measurement of Alcohol Consumption. Alcoholism: Clinical and Experimental Research, 36:332-34.
5. Huang M-C, Schwandt M, Ramchandani V, George D, Heilig M. 2012. Impact of
multiple types of childhood trauma exposure on risk of psychiatric comorbidity among alcoholic inpatients. Alcoholism: Clinical and Experimental Research, 36:598-606.
6. Zhang L, Kerich M, Schwandt M, Rawlings R, McKellar J, Momenan R, Hommer D,
George DT. 2012. Smaller right amygdala in alcoholic patients with a history of intimate partner violence: a volumetric imaging study. Addiction Biology (online doi: 10.1111/j.1369-1600.2011.00381.x).

27
7. Hirvonen J, Zanotti-Fregonara P, Umhau JC, George DT, Rallis-Frutos D, Lyoo CH, Li CT, Hines CS, Sun H, Terry GE, Morse C, Zoghbi SS, Pike VW, Innis RB, Heilig M. 2012. Reduced cannabinoid CB(1) receptor binding in alcohol dependence measured with positron emission tomography. Mol Psychiatry (online doi: 10.1038/mp.2012.100).
8. Schwandt ML, Heilig MA, Hommer DW, George DT, Ramchandani VA. 2012.
Childhood trauma exposure and alcohol dependence severity in adulthood: mediation by emotional abuse severity and neuroticism. Alcoholism: Clinical and Experimental Research (online doi: 10.1111/acer.12053).
9. Umhau JC, George DT, Heaney RP, Lewis MD, Ursano RJ, Heilig M, Hibbeln JR,
Schwandt ML. 2012. Low vitamin D status and suicide: a case-control study of active duty military service members. PLOS One (online doi: 10.1371/journal.pone.0051543).
Dr. George: [email protected]

28
______________________________________________________________________
Section on Human Psychopharmacology ______________________________________________________________________ Vijay A. Ramchandani, PhD, Acting Chief Vatsalya Vatsalya, MD, MS, Postdoctoral Visiting Fellow Bethany Stangl, PhD, Postdoctoral IRTA Jia Yan, PhD, Postdoctoral IRTA Marion Coe, BA, Postbac IRTA Molly Zametkin, BA, Postbac IRTA Veronica Schmidt, Research Contractor
L to R: M Zametkin, V Schmidt, J Yan, M Coe, V Ramchandani, B Stangl, V Vatsalya.
Alcoholism is a serious and chronic disease that has a tremendously negative impact on the health, work, family and relationships for the affected individual, and poses a substantial burden on society at large. Thus, there is a great and urgent need to understand the risk factors for development of alcohol problems as well as to develop treatments for this debilitating disorder, and research on the clinical pharmacology of alcohol, is critical to achieving this. Clinical pharmacological investigations include examination of the genetic and environmental determinants of the pharmacokinetics (PK) and pharmacodynamics (PD) of the drug (in this case, alcohol). While the PK and PD of alcohol have been widely studied, there is still a need to understand the sources of variability that may contribute to the risk of developing alcoholism. Moreover, an improved understanding of the brain systems underlying the pharmacodynamic effects of alcohol can lead to the development of novel pharmacotherapeutic agents for the treatment of alcoholism. Goals The Section on Human Psychopharmacology conducts studies characterizing the pharmacokinetics and CNS pharmacodynamics of alcohol in humans using behavioral, neuroendocrine, electrophysiological and functional imaging measures. These studies, conducted
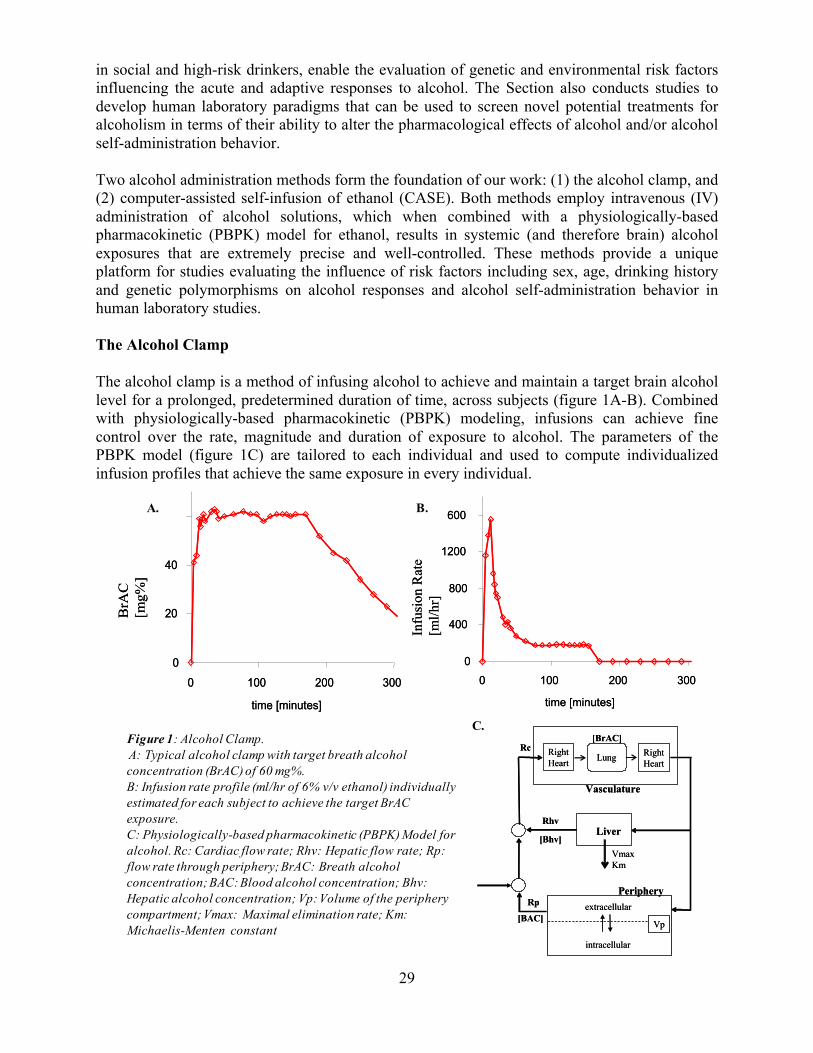
29
in social and high-risk drinkers, enable the evaluation of genetic and environmental risk factors influencing the acute and adaptive responses to alcohol. The Section also conducts studies to develop human laboratory paradigms that can be used to screen novel potential treatments for alcoholism in terms of their ability to alter the pharmacological effects of alcohol and/or alcohol self-administration behavior. Two alcohol administration methods form the foundation of our work: (1) the alcohol clamp, and (2) computer-assisted self-infusion of ethanol (CASE). Both methods employ intravenous (IV) administration of alcohol solutions, which when combined with a physiologically-based pharmacokinetic (PBPK) model for ethanol, results in systemic (and therefore brain) alcohol exposures that are extremely precise and well-controlled. These methods provide a unique platform for studies evaluating the influence of risk factors including sex, age, drinking history and genetic polymorphisms on alcohol responses and alcohol self-administration behavior in human laboratory studies. The Alcohol Clamp The alcohol clamp is a method of infusing alcohol to achieve and maintain a target brain alcohol level for a prolonged, predetermined duration of time, across subjects (figure 1A-B). Combined with physiologically-based pharmacokinetic (PBPK) modeling, infusions can achieve fine control over the rate, magnitude and duration of exposure to alcohol. The parameters of the PBPK model (figure 1C) are tailored to each individual and used to compute individualized infusion profiles that achieve the same exposure in every individual.
0
20
40
60
0 100 200 300
time [minutes]
BrA
C
[mg%
]
0
20
40
60
0 100 200 300
time [minutes]
BrA
C
[mg%
]
0
400
800
1200
1600
0 100 200 300
time [minutes]
Infu
sion
Rat
e [m
l/hr]
0
400
800
1200
1600
0 100 200 300
time [minutes]
Infu
sion
Rat
e [m
l/hr]
Figure 1: Alcohol Clamp.A: Typical alcohol clamp with target breath alcohol concentration (BrAC) of 60 mg%. B: Infusion rate profile (ml/hr of 6% v/v ethanol) individually estimated for each subject to achieve the target BrAC exposure. C: Physiologically-based pharmacokinetic (PBPK) Model for alcohol. Rc: Cardiac flow rate; Rhv: Hepatic flow rate; Rp: flow rate through periphery; BrAC: Breath alcohol concentration; BAC: Blood alcohol concentration; Bhv: Hepatic alcohol concentration; Vp: Volume of the periphery compartment; Vmax: Maximal elimination rate; Km: Michaelis-Menten constant
Liver
extracellular
intracellular
RightHeart Lung
[BrAC]RightHeart
Vasculature
Periphery
[BAC]
[Bhv]
Rc
Rhv
Rp
VmaxKm
Vp
Liver
extracellular
intracellular
RightHeart Lung
[BrAC]RightHeart
Vasculature
Periphery
[BAC]
[Bhv]
Rc
Rhv
Rp
VmaxKm
Vp
A. B.
C.

30
The alcohol clamp was developed by Ramchandani and O’Connor (2006) to minimize the substantial inter-individual variation in the PK of alcohol, including alcohol absorption and metabolic rates. This variation, which can be attributed to factors such as alcohol intake, food consumption and composition, use of other drugs, sex, age, body weight and composition, and genetic polymorphisms of the alcohol metabolizing enzymes, alcohol dehydrogenase and aldehyde dehydrogenase, confounds the examination of the acute PD effects of alcohol and its determinants. It is therefore critical to control the variability in the absorption and metabolism of alcohol and the resulting blood (and therefore brain) alcohol exposure that drives the pharmacological effects of alcohol. The alcohol clamp provides a unique platform to evaluate the effects of any number of determinants on the response to alcohol, including the magnitude, route and rate of exposure to alcohol, sex, age, family history of alcoholism, and recent drinking history. Alcohol response phenotypes obtained under the alcohol clamp can be used to examine candidate genes that may be associated with the response to alcohol and might underlie the risk for alcoholism. These response measures can also be used to evaluate the effects of medications being developed for the treatment of alcohol use disorders, particularly those that act by modifying the rewarding effects of alcohol. Another major application of the clamp method follows from the ability to obtain a direct measure of the alcohol elimination rate (AER). During the alcohol clamp, when the breath alcohol concentration (BrAC) and the infusion rate are both at steady-state, the infusion rate becomes a direct measure of the elimination rate of alcohol in grams per hour. This direct measurement of the AER allows the evaluation of various determinants of alcohol metabolism, including sex, age, ethnicity, and polymorphisms of the alcohol metabolizing enzymes, as well as the role of various factors such as food intake, lean body mass, liver blood flow and menopause in explaining the variability in alcohol.
Computer-assisted self-infusion of ethanol (CASE) in humans A common feature of almost all addictive drugs is that they are self-administered by both experimental animals and humans. Human alcohol self-administration is generally assessed by measuring the ingestion of alcoholic beverages, typically in a laboratory bar setting. However, these methods tend to be imprecise and may be unreliable due to substantial variability in alcohol exposures, either through differences in the definition of standardized drinks and drinking schedules or through high inter-individual PK variability, or both. In addition, these methods are subject to various non-pharmacological influences such as alcohol expectancy, beverage preference, choice and monetary value of incentives or disincentives for drinking.
A computer-assisted method of alcohol self-administration (CASE), using the PBPK-model based infusion algorithm, provides subjects with the flexibility to choose when to push a button to receive alcohol, while providing the investigator with flexibility in controlling the subsequent time course of breath (and therefore brain) alcohol exposure. This method assures the same increments across all participants and prevents the BrAC from exceeding any pre-set upper limit (figure 2). Thus, the CASE method minimizes PK variability, and focuses on assessment of behavior driven by the pharmacological effects of the drug. This is advantageous as it avoids non-pharmacological sources of variation in self-administration. However, the CASE method does not readily measure the component of alcohol reward that is driven by conditioned effects such as taste, smell and expectancy.

31
Figure 2. Schematic for computer-assisted self-infusion of ethanol (CASE) method.
The first paradigm developed using the CASE method was the so-called “open-bar” paradigm (Zimmermann et al., 2013). In this paradigm, participants have free access to alcohol (requiring a single button-press to receive infusions) for a period of 2 hours. This paradigm maintains the ad lib nature of an open bar, where drinks are provided for free, but the highly variable response to oral dosing is replaced by a standardized exposure to alcohol that is called a “slopelet”. A slopelet is a fixed linear increase in BrAC (for example an increase in BrAC at the rate of 4 mg% per min for 2 min resulting in an increase of 8 mg% per button-press) followed by a fixed linear decrease in BrAC (for example at -0.5 mg% per min) until the subject pushes the button for an additional exposure. This is achieved by administering an infusion rate profile, individually computed based on the PBPK model for alcohol, to achieve the desired BrAC exposure. The primary measures of self-administration include the number of button- presses, average BrAC, and peak BrAC. Preliminary studies indicated that the CASE system is a practical method to investigate human alcohol self-administration behavior. More recently, CASE was used to examine the influence of family history of alcoholism on alcohol self-administration, and as hypothesized, family history positive individuals showed higher levels of alcohol self-administration compared to family history negative individuals. The CASE paradigm is a valuable tool for evaluating determinants that may underlie self-administration behavior in humans, in particular the influence of factors such as drinking history and sex, since the infusion profile is individually tailored based on a subject’s sex and morphometric measures. The fact that CASE is sensitive to drinking history makes this method an important tool for evaluating the effect of pharmacological agents on the rate, magnitude and pattern of exposure to alcohol achieved in individuals. Reduction in alcohol self-administration, assessed as a change from baseline in CASE measures following treatment with these agents, may serve as a biomarker of the clinical effectiveness of these agents in the treatment of alcohol-dependence. Genetic Influences on Alcohol Responses There has been extensive research on the genetic underpinnings of alcoholism and studies have identified a number of genetic variations that have been associated with alcohol dependence and associated phenotypes. While many of these studies have been replicated, others have not been as successful. Moreover, the clinical phenotypes examined in most of these studies demonstrate

32
substantial heterogeneity and variability, resulting in smaller effect sizes for genetic influences and thus necessitating larger subject samples. Consequently, there has been an increasing interest in examining quantitative endophenotypes for genetic association studies for alcohol.
One group of quantitative endophenotypes includes those associated with the acute response to alcohol. It is widely accepted that the pharmacological effects of alcohol are linked to risk for alcohol dependence; however, there have been very few studies that have actually examined this relationship. Longitudinal studies by Schuckit and colleagues have identified relationships between subjective responses to alcohol and alcohol-related outcomes; however these studies need to be replicated. There have also been IV alcohol studies that have demonstrated family-history differences in the acute response to alcohol, suggesting genetic influences on the pharmacodynamics of alcohol. In addition, studies have identified genetic factors, such as the OPRM1 A118G and GABRA2 polymorphisms, that explain some of the variation in the pharmacodynamic response to acute alcohol administration. Role of OPRM1 A118G Polymorphism on alcohol-induced striatal dopamine release Mesolimbic dopamine release is a key signal for the reinforcing effects of drugs of abuse, including alcohol, and endogenous opioids are thought to exert their effects by modulating this release. A functional polymorphism (A118G) in the mu-opioid receptor (OPRM1) gene alters the affinity of the mu-opioid receptor for its endogenous ligand, and has been associated with greater subjective responses to alcohol, increased risk for alcoholism, and the response to naltrexone treatment. Additionally, alcohol preference and alcohol-induced stimulation has been observed in rhesus macaques carrying the functional equivalent of the OPRM1 A118G variant. This differential response to alcohol observed as a function of the OPRM1 A118G polymorphism may reflect differential activation of striatal dopamine release following acute alcohol administration. Therefore, we examined the influence of the OPRM1 A118G polymorphism on subjective and neuroendocrine responses and on striatal dopamine release, measured by 11C-raclopride displacement using positron emission tomography (PET), following intravenous (IV) alcohol administration. Healthy male social drinkers were screened to obtain two groups: 1) subjects homozygous for the major 118A allele (118AA); and 2) subjects carrying 1 or 2 copies of the variant 118G allele (118GX). Subjects underwent two PET scanning sessions with 11C-raclopride while they received IV infusions of either alcohol (6% v/v) or saline, according to an infusion-rate profile based on a physiologically-based pharmacokinetic model for alcohol, to achieve target breath alcohol levels of 0.08 g% or 0 g%, in counter-balanced order. During the infusions, serial blood samples were collected for cortisol and adrenocorticotropic hormone (ACTH) levels and self-ratings of subjective effects were obtained. The 118GX group, but not the 118AA group, showed significant decreases in 11C-raclopride binding potential in striatal regions, indicating greater dopamine release, following alcohol compared to placebo. Subject groups did not differ in peak scores for “high” and “intoxication”, however the 118GX subjects showed higher scores for “liking effects” and “wanting more” than the 118AA subjects. The 118GX group showed significantly higher cortisol and ACTH levels at baseline and greater alcohol-induced decreases in cortisol and ACTH levels compared to the 118AA group (Ramchandani et al., 2011).
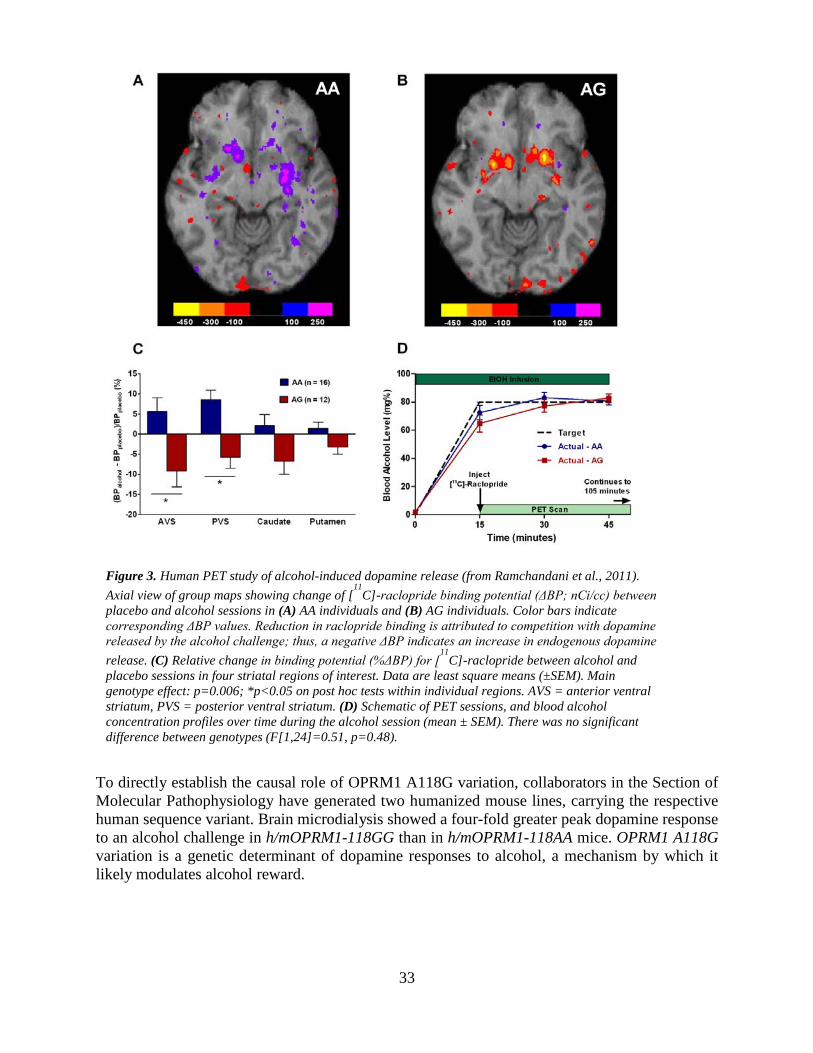
33
Figure 3. Human PET study of alcohol-induced dopamine release (from Ramchandani et al., 2011). Axial view of group maps showing change of [
11C]-raclopride binding potential (ΔBP; nCi/cc) between
placebo and alcohol sessions in (A) AA individuals and (B) AG individuals. Color bars indicate corresponding ΔBP values. Reduction in raclopride binding is attributed to competition with dopamine released by the alcohol challenge; thus, a negative ΔBP indicates an increase in endogenous dopamine release. (C) Relative change in binding potential (%ΔBP) for [
11C]-raclopride between alcohol and
placebo sessions in four striatal regions of interest. Data are least square means (±SEM). Main genotype effect: p=0.006; *p<0.05 on post hoc tests within individual regions. AVS = anterior ventral striatum, PVS = posterior ventral striatum. (D) Schematic of PET sessions, and blood alcohol concentration profiles over time during the alcohol session (mean ± SEM). There was no significant difference between genotypes (F[1,24]=0.51, p=0.48).
To directly establish the causal role of OPRM1 A118G variation, collaborators in the Section of Molecular Pathophysiology have generated two humanized mouse lines, carrying the respective human sequence variant. Brain microdialysis showed a four-fold greater peak dopamine response to an alcohol challenge in h/mOPRM1-118GG than in h/mOPRM1-118AA mice. OPRM1 A118G variation is a genetic determinant of dopamine responses to alcohol, a mechanism by which it likely modulates alcohol reward.

34
Ongoing Studies and Future Plans Currently the Section’s research in this area is moving in two directions: continuing the focused examination of the role of the OPRM1 locus in the acute response to alcohol, and using a broader approach of association studies to examine the genetics of the acute response to alcohol. Given our previous study demonstrating that alcohol-induced dopamine release is influenced by the OPRM1 A118G polymorphism in human, and pre-clinical studies that have also demonstrated that alcohol administration results in increased central release of endogenous opioid peptides, it is postulated that these opioid peptides (particularly beta-endorphin) activate mu-opioid receptors on GABA neurons in the ventral tegmental area (VTA), resulting in disinhibition of DA neurons and consequently DA release in the striatum (Heilig et al., 2009). The Section plans to examine the effect of IV alcohol, and the role of the OPRM1 A118G polymorphism, on endogenous opioid release following acute alcohol administration in healthy male and female social drinkers using PET employing [11C] carfentanil, a selective mu-opioid receptor agonist that provides a measure of brain mu-receptor occupancy, and can be used to measure changes in endogenous opioid release following acute alcohol administration in humans. Comparison of alcohol-induced endogenous opioid release between subjects who carry the common and variant forms of the OPRM1 A118G SNP would provide important information about the role of this system in the acute effects of alcohol, and specifically establish whether differential opioid activation to alcohol is upstream of the detected differential alcohol-induced release of DA.
A complementary approach to examining the effect of candidate gene loci on the pharmacological effects of alcohol is to use genome-wide association approaches. This approach is based on the assumption that common genetic variation contributes to disease risk, and allows a comprehensive test of association across the genome, rather than testing only one gene at a time. Genome-wide association studies have identified several loci with significant associations with alcohol dependence. These analyses are typically limited by low statistical power, thus necessitating large sample sizes. However, recent studies using well characterized quantitative endophenotypes in relatively small, carefully ascertained samples have successfully demonstrated whole genome-based associations with alcohol-related phenotypes, such as EEG. More recently approaches such as pathway-based analysis can enable the examination of neurobiology-based gene networks for their association with these quantitative endophenotypes.
Studies evaluating the response to alcohol have used varying definitions of risk and of the measures or phenotypes that characterize the response to alcohol. Studies by Schuckit have used the level of response (LR) phenotype, which posits that a low level of response is associated with increased risk, and is based predominantly on subjective measures mostly associated with the sedative and ataxic effects of alcohol. These studies have demonstrated that family history positive (FHP) subjects have a lower level of response; however it is unclear if this lower level is the result of a lower sensitivity or a greater tolerance development in the high risk FHP group. In contrast, our previous work and those of others have shown both higher initial responses and greater acute tolerance development in FHP subjects. This lends support to the differentiator model, proposed by Newlin and Thompson, that posits that a higher initial response on the ascending limb of the BrAC-time curve and a lower response on the descending limb of the BrAC-time curve are associated with increased risk. Thus there is a need to study both the initial

35
effect as well as the adaptive or acute tolerance to the effects of alcohol in studies evaluating the genetics of alcohol responses in humans.
In studies of oral alcohol administration, measures of acute tolerance are obtained by comparing responses at the same BAC level on the ascending and descending limbs of the BrAC-time curve. A lower response on the descending limb, compared to the ascending limb, is indicative of acute functional tolerance and the difference between measures can be used as a metric of acute tolerance. In these studies, however, even though the BrAC level is the same, the rate of change is different in magnitude and direction, and there is a variable interval of time during which the BrACs peak and decline between measurements. Studies using the alcohol clamp method define acute tolerance as the difference between responses measured at the start and at the end of a pre-determined duration of constant clamped exposure to the same BAC level. In this method, the level of exposure is constant, rate of change of exposure is zero and the only variable factor is time. It is unclear however, which metric of acute tolerance would provide greater sensitivity to genetic influences in genetic studies of the acute response to alcohol.
The Section is currently conducting a study to identify genes that are associated with the response to acutely administered alcohol in non-dependent drinkers. The study will be conducted in two phases. In phase I, subjects with positive family history of alcoholism (FHP) and with no family history of alcoholism (FHN) will undergo two ethanol infusion sessions to compare different breath alcohol concentration exposures. In one session (Clamp session), participants will be infused with 6% ethanol in saline using an individualized infusion profile to achieve and clamp breath alcohol concentrations (BrAC) at 60 mg% for 2 hours. In the other session (Oral-mimic session), participants will be infused with 6% ethanol and saline using another individualized infusion profile to achieve a BrAC-time profile that would be typically obtained after oral administration (ascending limb to peak BrAC followed by descending limb). During each session, serial BrACs will be obtained, and a battery of subjective (self-rating questionnaires) and objective measures (psychomotor performance, behavioral disinhibition tasks) will be obtained at baseline and two points during the infusion to assess the initial response and adaptive response to alcohol. The measures of initial response and adaptive response will be compared between sessions to determine which provides greater sensitivity (higher effect size) for detecting family history differences. In phase II, participants will undergo a one session study, and will receive an ethanol infusion to achieve the BrAC-time profile (Clamp or Oral-mimic) that provides the greater sensitivity to family history of alcoholism in phase I of the study. The same battery of subjective and objective measures will be obtained to assess the initial response and adaptive response to alcohol. These response measures will be used as endophenotypes for examination of genetic association with a set of candidate genes, based on previous clinical and pre-clinical studies. Identification of genes that are associated with these endophenotypes is important in several contexts, including the identification of susceptibility to alcoholism, understanding the biological pathways involved in the acute pharmacological actions of alcohol, and ultimately, in informing the development of better pharmacological treatments to help those who wish to reduce their alcohol consumption. Other Determinants of Alcohol Responses Influence of sex and age on alcohol metabolism and responses Previous research indicates that alcohol metabolism differs between men and women, and may also be influenced by age. There appears to be a complex interaction between sex, age and

36
alcohol metabolism, and differences in sex steroidal hormones, estrogen and testosterone, may underlie this interaction. Studies have also shown sex- and age-related differences in alcohol pharmacodynamics, although the underlying determinants of these differences are unclear. The objective of this study was to evaluate the influence of sex and age on the metabolism and acute response to alcohol in social drinkers. This was a randomized, two-session crossover study in 48 male and female - 24 young (21-25 years) and 24 older (55-65 years) - social drinkers. Participants underwent the alcohol clamp at target BrACs of 0 and 50 mg% for 3 hrs. The alcohol elimination rate (AER) and behavioral, hormonal and physiological responses to alcohol were evaluated. Analysis of the pharmacokinetic data indicated that AERs were significantly higher in males than in females, as expected. AER per unit lean body mass (AER/LBM) was similar between males and females, indicating that sex differences in lean body mass may contribute to the observed sex differences in AER. Age did not appear to influence AER, although AER/LBM was higher in older compared to younger subjects. LBM was significantly associated with AER (r2=0.42) across subjects. Liver volume, measured using magnetic resonance imaging, was also significantly associated with AER (r2=0.23) across subjects. Analysis of the pharmacodynamic data showed a significant effect of alcohol on peak subjective perceptions of high, intoxication and drug effects. There were no sex differences in subjective responses to clamped BrACs. Older subjects reported similar peak ratings of “high” and “intoxicated” as younger subjects. However, they showed significantly lower peak ratings for “liking drug effects” and “wanting more” than younger subjects. Analysis of the effect of alcohol on growth hormone (GH), insulin-like growth factor-1 (IGF-1) and sex steroid (estrogen, testosterone) levels revealed a complex pattern of responses that appeared to differ by sex and age (Vatsalya et al., 2012). Change in free testosterone levels showed a significant treatment-by-baseline interaction, suggesting that the alcohol-induced decrease in testosterone levels was seen primarily in males. Change in estradiol levels showed a main effect of treatment with alcohol and a significant treatment-by-baseline interaction as well as a significant treatment-by-sex interaction, indicating that alcohol resulted in an increase in estradiol in females and a decrease in estradiol in males, compared to placebo. Change in IGF1 levels did not show a main effect of treatment; however it demonstrated a main effect of baseline and a trend for treatment-by-age interaction. Change in GH showed a main effect of baseline and main effect of sex, and a trend for treatment-by-baseline interaction, suggesting that males showed greater increases in GH levels across treatment compared to females. Analysis of the cardiovascular responses indicated a significant decrease in heart rate and an increase in heart rate variability measures during the ascending limb of the IV alcohol infusion, with no significant differences observed during the clamped interval. This suggests an initial dominant parasympathetic cardiac response, followed by a progressively increasing sympathetic response to alcohol. Further, the change in HR was significantly associated with changes in subjective responses measured during the ascending limb. These results are currently being written up for publication. Additional analyses, including PK-PD modeling, are being conducted to evaluate the influence of age and sex on subjective responses and heart rate changes during the alcohol clamp. Findings from these studies will provide a better understanding of age- and sex-related differences in

37
alcohol metabolism, which may underlie medically important differences in individual responses to alcohol. Functional MR Imaging Studies of Alcohol Response The Section has collaborated with the Section on Brain Electrophysiology and Imaging (SBEI) to conduct studies examining the effect of acute alcohol infusion on BOLD signal changes using functional magnetic resonance imaging (fMRI). The first study examined the effect of IV alcohol, administered to a target BrAC of 0.08%, in social drinkers (Gilman et al., 2008). The brain response to facial images portraying neutral and fearful emotions was measured using fMRI. Alcohol robustly activated ventral striatal reward areas in response to neutral faces, and attenuated the response to fearful stimuli in limbic and visual brain areas. Self-ratings of intoxication were significantly correlated with striatal activation, indicating that striatal activation may contribute to the subjective experience of pleasure and reward during intoxication. In the second study (Gilman et al., 2012a), the brain response to intravenous ethanol (target BrAC of 0.08%) and facial images portraying neutral and fearful emotions was compared between groups of heavy drinkers (HDs) and social drinkers (SDs). HDs reported lower subjective alcohol effects than SDs. Alcohol significantly activated the nucleus accumbens in SDs, but not in HDs. Self-reported ratings of intoxication correlated with striatal activation, suggesting that activation may reflect subjective experience of intoxication. Fearful faces significantly activated the amygdala in the SDs only, and this activation was attenuated by alcohol. This study shows that HDs not only experience reduced subjective effects of alcohol, but also demonstrate a blunted response to alcohol in structures that are a part of the brain’s reward circuitry. These findings indicate that reduced subjective and neural response to alcohol in heavy drinkers may be a result of development of tolerance to alcohol, or of pre-existing decreased sensitivity to alcohol‘s effects. More recently, in collaboration with SBEI, we have completed a study examining the neural correlates of risky decision making and the influence of alcohol in social drinkers (Gilman et al., 2012b). In this study, participants were given intravenous alcohol or placebo while undergoing fMRI and playing a risk-taking game. The game allowed us to examine the neural response to choosing a safe or risky option, anticipating outcome, and receiving feedback. We found that alcohol increased risk-taking behavior, particularly among participants who experienced more stimulating effects of alcohol. fMRI scans demonstrated that alcohol increased activation in the striatum to risky compared to safe choices, and dampened the neural response to notification of both winning and losing throughout the caudate, thalamus, and insula. This study suggests that alcohol may increase risk-taking behavior by both activating brain regions involved in reward when a decision is made, and by dampening the response to negative and positive feedback.
These studies have been critical in identifying the neural correlates of the rewarding and anxiolytic effects of alcohol, as well as the effect of drinking history on these effects. The effect of alcohol on decision making has improved our understanding of increased risk-taking behavior seen after alcohol consumption. Ongoing Studies and Future Plans
The Section has several ongoing or planned collaborative studies to examine neural correlates of the acute effects of alcohol. We continue to collaborate with SBEI to conduct MRI studies of the acute effects of alcohol. SBEI has developed a version of the monetary incentive delay (MID)

38
task (called Alcohol-MID) that directly measures motivation for alcohol. As described in the SBEI report, this task involves subjects being scanned while they perform a MID task during which they can button press to obtain points that determine how much intravenous alcohol or snack food they will receive at the conclusion of the scan. This task is being used in an experimental medicine study of varenicline (see below). Another current study is being conducted in collaboration with SBEI to examine the effect of acute alcohol administration on brain levels of glutamate and other metabolites using magnetic resonance spectroscopy (MRS) in social and heavy drinkers. In a collaborative study with Drs. Mark Hallett and Dietrich Haubenberger, NINDS Intramural Program, the alcohol clamp is being used to examine electrophysiological correlates of the effect of alcohol to suppress tremor in patients with essential tremor. This study will use TMS-induced intracortical inhibition (a mechanism known to be mediated by GABA) to examine the tremor response to alcohol, and to examine correlations between objective and subjective measures of alcohol response in these patients. Determinants of Intravenous Alcohol Self-administration Drinking history, Initial Sensitivity and Personality Measures
Initial studies conducted by the Section using the Computer-Assisted Self-infusion of Ethanol (CASE) method were aimed at characterizing the open-bar paradigm in healthy social drinkers, who underwent two identical CASE sessions. During each CASE session, subjects first underwent a directed priming phase, where they were prompted to push a button to receive standardized alcohol infusions. This was followed by an ad-lib phase, where they have free access to the standardized alcohol infusions. BrAC was measured serially, and physiological and subjective measures of alcohol effects and urges were assessed. Primary self-administration measures include number of button presses, average BrAC and peak BrAC.
Analysis of data generated thus far indicates a high degree of test-retest reliability between self-administration measures across sessions (all correlation coefficients > 0.7). Correlations between measures within each session demonstrated a high level of internal consistency (all correlation coefficients > 0.7). There was a significant association between recent drinking history measures and self-administration measures. Regression analyses of the influence of recent drinking measures revealed the number of drinks per drinking day to be the strongest predictor of self-administration measures (p < 0.01). There did not appear to be any sex differences in self-administration measures. A retrospective measure of initial sensitivity to alcohol (Self-Report of the Effects of Alcohol Questionnaire) was positively associated with ad lib alcohol self-administration. There was also a significant positive association between measures of impulsivity, specifically motor impulsivity (measured using the BIS) and sensation-seeking (measured using the UPPS+P scale) and measures of self-administration. Measures of “liking drug effects” (measured using DEQ) and urges (measured using AUQ) following the priming phase predicted the number of button presses and average BrAC during the ad lib phase. Subjective measures of craving following the priming phase were significantly associated with self-administration measures. There was a strong association between peak alcohol effects of feeling drug effects, liking drug effects, intoxication, stimulation and self-administration measures. In summary, heavier drinkers, individuals with low responsiveness to alcohol (higher SRE scores), and individuals with greater reward sensitivity demonstrated higher alcohol self-administration in the laboratory. Self-administration was associated with perceptions of

39
intoxication and stimulation, and supports the role of rewarding alcohol effects underlying alcohol intake behavior. Additional analyses of these data are ongoing to characterize changes in objective response measures including heart rate variability and skin blood flow, and their relationship to the subjective response to alcohol during IV self-administration. The Section plans to conduct quantitative PK-PD modeling to characterize the exposure-response relationship between self-administration measures and response to alcohol. In addition, the Section plans to continue to collect data on IV alcohol self-administration to have an adequate sample size to examine genetic underpinnings of this IV self-administration behavior. Ongoing Work and Future Plans Recent efforts have been made by the Section to develop more operant paradigms of human alcohol self-administration, including a progressive ratio (PR) schedule paradigm. In contrast to the open-bar paradigm, this paradigm requires participants to press the button an increasing number of times for each subsequent alcohol exposure (e.g., 10, 20, 40, 80, etc., button presses for the 1st, 2nd, 3rd, 4th, etc. infusions). Outcome measures of this paradigm include the “breakpoint”, i.e., the point at which the individual stops working for more alcohol, as well as the average and peak BrACs achieved during the study. This type of operant paradigm is expected to assess motivation for reward (in this case, alcohol), and is based on the principle that people will work harder for greater rewards. There is extensive animal work to support the utility of operant paradigms in characterizing motivation for reward, both for the evaluation of genetic determinants of ethanol drinking as well as for screening molecular targets for medication development for alcoholism. These operant paradigms have been used in animal models and in a few human studies to evaluate self-administration behavior for heroin and cocaine. However, very little research has been done using alcohol in humans. Since the reinforcing value of drugs is a function not only of their pharmacodynamic effects, but also the rate at which these effects are produced, as well as the slope of the brain exposure curve, it is important to robustly demonstrate that humans will respond on a progressive ratio schedule for IV alcohol. To our knowledge, the only published studies to date using a PR schedule paradigm for alcohol self-administration used ingestion of 6 gm oral drinks of ethanol in return for geometric increases in schedule of incremental work required. Thus, the PR schedule paradigm represents an interesting and novel extension of IV alcohol self-administration methods. Development and evaluation of this operant (PR schedule) method would complement free-choice (open-bar) measures of IV alcohol self-administration measures in improving the understanding of both genetic and environmental determinants of alcohol self-administration behavior. It would also offer an attractive translational biomarker in evaluation of experimental therapeutics.
Given the paucity of data on operant alcohol self-administration in humans, initial studies are focusing on evaluating the test-retest characteristics of this paradigm and an examination of factors such as recent drinking history and sex on alcohol self-administration behavior. Preliminary results from these studies have demonstrated excellent test-retest reliability (correlation coefficients for self-administration measures between 2 sessions > 0.8). These studies have also indicated that self-administration measures obtained from the progressive-ratio paradigm are sensitive to drinking history and expectancy of alcohol effects. This demonstration of the repeatability of the PR schedule paradigm and sensitivity to drinking history provides the basis for using this paradigm to evaluate effects of medications on motivation for alcohol and self-administration behavior in humans, as discussed below.

40
Early Life Stress and Alcohol-related Outcomes Studies examining genetic influences on alcoholism have indicated a heritability of approximately 50%, thereby indicating a substantial contribution of environmental factors, both shared and non-shared, on alcoholism and other alcohol-related outcomes. One of these environmental factors is exposure to early life stress. There is growing evidence that exposure to stressful and adverse childhood experiences are associated with a range of adverse outcomes in adults. Individuals who have adverse childhood experiences have a greater vulnerability and/or prevalence of substance abuse, including alcoholism, and psychiatric diagnoses, including depression. Individuals who have adverse childhood experiences tend to show higher levels of neuroticism and lower levels of conscientiousness than controls. Individuals with early life stress or trauma show alterations in electrophysiological measures as well as volumetric measures of brain function and structure. In addition, studies have identified a role for genetic factors and their interaction with this environmental determinant in the relationship between early life stress factors and outcomes including anxiety disorders, post-traumatic stress disorder, depression, suicidality, and alcoholism. Studies have also indicated a role of early life negative events on earlier onset of drinking, and other hazardous patterns of drinking such as binge drinking and drinking to cope in non-dependent individuals. In collaboration with the Section on Clinical Assessment and Treatment Evaluation (SCATE), the Section has launched an effort to examine the prevalence of early life stress in alcoholics as well as non-dependent drinkers, and evaluate the influence of early-life stress, including adverse childhood experiences such as trauma and abuse, on a range of alcohol-related outcomes. Early life stress measures have included the childhood trauma questionnaire (CTQ) and the early life stress questionnaire (ELSQ). Alcohol-related outcomes include measures of alcohol use (drinking quantity, frequency, pattern) as well as IV self-administration in non-dependent drinkers, and alcohol dependence severity, withdrawal symptoms, anxiety and depression symptomatology, suicidality, comorbid psychiatric diagnoses (including PTSD, other anxiety disorders and substance use disorders) in treatment-seeking alcoholics. We are also examining the mediating role of personality measures, including neuroticism, impulsivity and aggression on the relationship between early life stress and alcohol-related outcomes. Analysis of a sample of 376 treatment-seeking alcoholics indicated a high prevalence of childhood trauma, including emotional abuse (46%), physical abuse (46%) and sexual abuse (24%). Rates in a complementary group of 209 non-dependent drinkers were significantly lower: emotional abuse (11%), physical abuse (15%) and sexual abuse (6%). In addition, childhood trauma was found to influence alcohol dependence severity, an effect that was mediated by neuroticism. When individual trauma types were examined, emotional abuse was found to be the primary predictor of alcohol dependence severity. Physical abuse also had a moderate direct effect on alcohol dependence severity. Results of mediation analyses, conducted to model associations between childhood trauma measures and alcohol-related outcomes, indicated that the relationship between emotional abuse and dependence severity was mediated through the impulsivity sub-facet of neuroticism. These findings suggest that childhood trauma may play a significant role in the development and severity of alcohol dependence through an internalizing pathway involving negative affect (Schwandt et al., 2013).
Another analysis of this dataset focused on the relationship between various types of childhood trauma and lifetime psychiatric comorbidities and suicide attempts in treatment-seeking alcoholics. Results indicated that the prevalence of at least 2 types of childhood trauma was 32%,

41
and the prevalence of at least 3 types of childhood trauma was 19% in this sample. A history of emotional abuse increased the risk of mood disorder, in particular major depressive disorder, as well as PTSD. Physical abuse contributed to the prediction of suicide attempts, while sexual abuse was associated with a diagnosis of anxiety disorder, PTSD, and multiple comorbidities (e.g., anxiety and mood disorder).The number of reported trauma types or the total scores on the CTQ predicted an increased risk of having single or multiple psychiatric comorbidities as well as suicide attempts. These results indicate a childhood trauma exposure-specific enhancement of risk for various psychiatric outcomes among alcoholic inpatients, and point to the need to consider this important factor in determining severity of psychiatric illness and developing prevention and treatment strategies for alcoholic individuals (Huang et al., 2012). Examination of the effect of early life stress in non-dependent drinkers has focused on drinking measures, both recent drinking history (as measured by the timeline followback) as well as IV alcohol self-administration (as measured by CASE). In a dataset of 203 non-dependent drinkers, the total score on the CTQ and the number of trauma types experienced was positively associated with recent drinking history measures including total drinks and drinks per drinking day (p<0.001). Those that reported childhood emotional abuse, emotional and physical neglect had heavier drinking (drinks per drinking day). In a subset of participants that underwent the CASE paradigm, the presence of a history of childhood trauma as well as the number of trauma types were associated with the total amount of ethanol administered and the average BrAC achieved during the session. These findings are being written up for publication. Current and future plans include examination of the effect of early life stress on neuroanatomical measures in alcoholics and controls. Previous studies have shown associations between early life stress and reductions in hippocampal volume, and pre-clinical studies have shown increased amygdala volume among animals exposed to prenatal stress. Chronic alcohol exposure has also been associated with reductions in total brain volume, as well as hippocampal volume; however there are no studies examining the interaction between early life stress exposure and alcohol exposure on structural brain measures. The Section, in collaboration with SCATE and SBEI, plans to measure brain volume as well as sub-cortical volumes in alcoholics and controls with and without early life stress to examine the effects of alcohol and early life stress, and their interaction, on brain volumetric measures. Future plans also include the examination of genetic influences on the relationship between early life stress and brain volumetric measures in alcoholics. These studies will provide valuable information on gene-by-environment interactions on the effects of alcohol on brain structure and function.
Human Laboratory Paradigms in Medication Development for Alcoholism
There has been much research on the pathophysiological mechanisms underlying the development of alcohol dependence, and this has led to the discovery of several promising targets for pharmacotherapy of alcohol dependence. While there are well-established animal models of alcoholism that characterize various aspects of the addictive process, there is a need for continued development of human laboratory models that can help translate the pre-clinical findings to clinical populations. A recent review has proposed a so-called “Rosetta Stone approach” to medications development, wherein pre-clinical screening is combined dynamically with human laboratory studies to help identify treatments that are likely to succeed in clinical trials and to facilitate further drug development.

42
Medication development research for alcohol has focused on two broadly defined targets: (1) the mesocorticolimbic system, which is involved in the positive reinforcing effects of alcohol, and alcohol intake/reinforcement behaviors; and (2) the stress response and glutamate systems, which become dysregulated during the withdrawal-negative affect stage and anticipation/craving stage of addiction. Human laboratory studies that model alcohol intake and acute intoxication effects include challenge studies with standardized exposure to alcohol as well as self-administration studies. Studies that model drug anticipation and craving effects include studies of cue-reactivity, cue-induced craving, stress-induced craving, and brain imaging studies of craving behavior. The Section is also invested in developing and utilizing human alcohol self-administration paradigms, using the CASE method, to examine the effect of pharmacological agents being developed for the treatment of alcoholism. Human self-administration studies directly examine drug-taking behavior and thus play a critical role in informing decisions regarding the potential usefulness of pharmacotherapeutics for alcohol. Previous studies have established that oral alcohol self-administration is a useful tool to study the effect of pharmacological agents on self-administration behavior, and this effect can serve as a biomarker of the clinical effectiveness of these agents in the potential treatment of alcohol dependence (Zimmermann et al., 2013). IV alcohol self-administration, using the CASE method as described above, holds promise to complement these paradigms by adding the following new features: (i) the IV method, while non-naturalistic, avoids the influence of olfactory, gustatory and other cues associated with oral alcohol consumption, which can be both a limitation and an advantage. (ii) The CASE method yields peak BrAC levels under baseline conditions that are significantly associated with drinking patterns, and therefore may reflect hazardous (binge and/or heavy) drinking more closely than oral self-administration paradigms. (iii) The CASE method demonstrates good test-retest reliability, suggesting that within-subjects designs can be reliably used for medication testing. Thus, the CASE method may serve as a complementary translational tool to screen drugs that have demonstrated the ability to modify free choice or operant self-administration of alcohol in animal models of alcoholism.
The first experimental medicine initiated by the Section was designed to examine the effect of varenicline, an α4β2 (nicotinic) acetylcholine receptor (nAchR) partial agonist, on alcohol self-administration using the CASE method in non-treatment seeking heavy drinkers. Varenicline, an oral medication approved by the FDA for smoking cessation, has demonstrated effectiveness in reducing alcohol consumption in both a rodent model of alcohol dependence and in heavy drinking smokers. Our study, which is currently ongoing, is a randomized, placebo-controlled study that investigates the ability of varenicline to decrease IV alcohol self-administration in non-treatment-seeking heavy drinkers. The study includes 21-60 year-old non-treatment-seeking heavy drinkers in good physical and psychiatric health. Subjects will undergo a baseline CASE session, after which they will be randomized to 3 weeks of treatment with varenicline or placebo in this parallel-design study. Following approximately 2 weeks of treatment, subjects will undergo an fMRI scan to examine the effect of varenicline on incentive motivation for alcohol and the underlying brain reward system activation, as well as on activation of brain reward systems in response to intravenously administered alcohol. At the end of 3 weeks of treatment, subjects will repeat the CASE session to measure changes in IV self-administration. Following the medication phase, all subjects will receive brief intervention for their alcohol use. We anticipate this study to demonstrate the utility of the CASE paradigm in evaluating pharmacologic agents being developed for the treatment of alcohol dependence.

43
Ongoing Work and Future Plans The Section plans to continue the development of self-administration paradigms as models for studying potential medications for alcohol dependence. Given the current efforts of the Molecular Pathophysiology and CATE sections as well as others in studying novel pharmacological agents targeting the negative affect phase of the addiction cycles, the Section plans to examine the relationship between stress and IV alcohol self-administration in humans. Previous studies that have examined the effect of acute stress exposure have yielded mixed results, probably related to the type of subjects and methods used. Recent studies have demonstrated that acute stress exposure induced alcohol cravings, and the magnitude of this craving was associated with relapse, in treatment-seeking alcoholics. The Section plans to use guided imagery of stressful situations, individualized for each subject, to acutely elicit a stress response and evaluate the craving and self-administration response to this stress exposure in non-treatment-seeking heavy drinkers. In addition to providing important information on the modulation of alcohol self-administration behavior by acute stress, this paradigm could serve as a model to examine the effect of pharmacological agents, such as CRH-antagonists and NK1-antagonists, on stress-induced self-administration. These experimental studies would help further establish the use of these human laboratory paradigms in medications development for alcohol addiction. References
Gilman JM, Ramchandani VA, Davis MB, Bjork JM, Hommer DW. 2008. Why we like to drink: a functional magnetic resonance imaging study of the rewarding and anxiolytic effects of alcohol. J Neurosci 28:4583-91.
Kwo PY, Ramchandani VA, O'Connor S, Amann D, Carr LG, et al. 1998. Gender differences in alcohol metabolism: relationship to liver volume and effect of adjusting for body mass. Gastroenterology 115:1552-7.
Ramchandani VA, Bolane J, Li TK, O'Connor S. 1999. A physiologically-based pharmacokinetic (PBPK) model for alcohol facilitates rapid BrAC clamping. Alcohol Clin Exp Res 23:617-23.
Ramchandani VA, Bosron WF, Li TK. 2001. Research advances in ethanol metabolism. Pathol Biol (Paris) 49:676-82.
Ramchandani VA, Flury L, Morzorati SL, Kareken D, Blekher T, et al. 2002. Recent drinking history: association with family history of alcoholism and the acute response to alcohol during a 60 mg% clamp. J Stud Alcohol 63:734-44.
Ramchandani VA, O'Connor S. 2006. Studying alcohol elimination using the alcohol clamp method. Alcohol Res Health 29:286-90.
Ramchandani VA, Plawecki M, Li T-K, O’Connor S. 2009. Intravenous ethanol infusions can mimic the time course of breath alcohol concentrations following oral alcohol administration in healthy volunteers. Alcohol Clin Exp Res 33:938-44.

44
Publications 2011-2012 1. Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, Damadzic R,
Eskay R, Schoor M, Thorsell A, Schwandt ML, Sommer WH, George DT, Parsons LH, Herscovitch P, Hommer D, Heilig M. 2011. A genetic determinant of the striatal dopamine response to alcohol in men. Molecular Psychiatry 16: 809-17
2. Roh S, Matsushita S, Hara S, Maesato H, Matsui T, Suzuki G, Miyakawa T, Ramchandani VA, Li TK, Higuchi S. 2011. Role of GABRA2 in Moderating Subjective Responses to Alcohol. Alcohol Clin Exper Res 35: 400-7
3. Gilman JM, Ramchandani VA, Crouss T, Hommer DW. 2012. Subjective and Neural Responses to Intravenous Alcohol in Young Adults with Light and Heavy Drinking Patterns. Neuropsychopharmacology 37: 467-77
4. Gilman JM, Smith AR, Ramchandani VA, Momenan R, Hommer DW. 2012. The effect of intravenous alcohol on the neural correlates of risky decision making in healthy social drinkers. Addiction Biology 17: 465-78
5. Huang MC, Schwandt ML, Ramchandani VA, George DT, Heilig M. 2012. Impact of Multiple Types of Childhood Trauma Exposure on Risk of Psychiatric Comorbidity Among Alcoholic Inpatients. Alcohol Clin Exper Res 36: 598-606
6. Kalu N, Ramchandani VA, Marshall V, Scott D, Ferguson C, Cain G, Taylor R. 2012. Heritability of Level of Response and Association with Recent Drinking History in Nonalcohol-Dependent Drinkers. Alcohol Clin Exper Res 36: 522-9
7. Schwandt ML, Heilig M, Hommer DW, George DT, Ramchandani VA. 2012. Childhood Trauma Exposure and Alcohol Dependence Severity in Adulthood: Mediation by Emotional Abuse Severity and Neuroticism. Alcohol Clin Exp Res
8. Snell LD, Ramchandani VA, Saba L, Herion D, Heilig M, George DT, Pridzun L, Helander A, Schwandt ML, Phillips MJ, Hoffman PL, Tabakoff B, Trait WISS. 2012. The Biometric Measurement of Alcohol Consumption. Alcohol Clin Exper Res36: 332-41
9. Vatsalya V, Issa JE, Hommer DW, Ramchandani VA. 2012. Pharmacodynamic Effects of Intravenous Alcohol on Hepatic and Gonadal Hormones: Influence of Age and Sex. Alcohol Clin Exper Res 36: 207-13
10. Hendler RA, Ramchandani VA, Gilman J, Hommer DW. 2013. Stimulant and sedative effects of alcohol. Curr Top Behav Neurosci 13: 489-509
11. Ramchandani VA. 2012. Genetic aspects of alcohol metabolism. In Alcohol, Nutrition and Health Consequences (Watson RR and Preedy VR, ed). New York, Humana Press.
12. Zimmermann US, O’Connor S, Ramchandani VA. 2013. Modeling alcohol self-administration in the human laboratory. Curr Top Behav Neurosci 13:315-53 (In Press)
Dr. Ramchandani: [email protected]

45
______________________________________________________________________
Section on Clinical Psychoneuroendocrinology and Neuropsychopharmacology (a joint NIAAA-NIDA section)
______________________________________________________________________ Lorenzo Leggio, MD, PhD, MSc, Acting Chief Mary R. Lee, MD, Staff Clinician Allison A. Feduccia, PhD, Postdoctoral IRTA Petra Suchankova Karlsson, PhD, Postdoctoral Guest Researcher Emily Oot, Postbac Technical IRTA Jared Bollinger, Postbac IRTA Lexi Dias, Postbac IRTA
E Oot, M. Lee, J. Bollinger, A. Feduccia, L. Leggio, L. Dias, P. Suchankova Karlsson
Preface The Section on Clinical Psychoneuroendocrinology and Neuropsychopharmacology (CPN) has recently formed (summer 2012) as a joint effort between NIAAA and NIDA after the recent recruitment of Dr. Lorenzo Leggio, as a tenure-track clinical investigator. Before joining NIH, Dr. Leggio was a faculty member at the Brown University Medical School and Center for Alcohol and Addiction Studies (CAAS). Dr. Leggio’s clinical research has primarily focused on the treatment of alcoholism, with a special emphasis on the role of feeding-related as well as GABAergic pathways; and on the medical consequences of alcoholism, with a special emphasis on alcoholic liver diseases. As a Principal Investigator, Dr. Leggio received extramural research funding from NIAAA and NIDA. Additionally, he received research funding from the European Foundation for Alcohol Research (ERAB), Brown University CAAS, ABMRF/The Foundation for Alcohol Research, and the Brain & Behavior Research Foundation (formerly NARSAD).

46
Dr. Leggio’s current research at the NIH Intramural Research Program is jointly supported by NIAAA and NIDA. Additional support is also provided by Dr. Leggio’s NARSAD Young Investigator Grant awarded by the Brain & Behavior Research Foundation. Mission The mission of the CPN Section is to contribute to a deeper understanding of possible pharmacological treatments for alcohol and drug use disorders. In order to identify possible novel medications for addiction, we conduct outpatient and inpatient clinical studies using a combination of state-of-the-art and novel biobehavioral and pharmacological procedures performed under well-controlled human laboratory conditions. The CPN section is particularly interested in: 1) investigating feeding-related pathways (e.g. ghrelin, GLP-1, insulin, hypothalamus-pituitary-thyroid axis) as novel neuropharmacological targets for alcoholism; and 2) identifying biobehavioral mechanisms and markers that predict treatment outcomes of promising medications (e.g. GABAB agonists) for the treatment of alcoholism. Clinical validation of novel targets: focus on feeding-related pathways As part of our inaugural studies, we are focusing on the role of the ghrelin pathway in alcoholism. Ghrelin is a 28-amino acid peptide acting as the endogenous ligand for the growth hormone secretagogue receptor (GHS-R). Ghrelin stimulates appetite by acting on the hypothalamic arcuate nucleus (ARC), a region that controls the intake of food and other substances, including alcohol. In addition to the ARC, GHS-Rs are also highly expressed in the caudal brain stem, the ventral tegmental area (VTA), hippocampus, substantia nigra, and dorsal and medial raphe nuclei. The expression of the GHS-R in the mesolimbic dopamine (DA) pathway suggests that ghrelin could play a role in reward processing. The role of ghrelin in DA reward processing and the role of the DA reward system in alcoholism suggest a role of ghrelin in alcoholism. Consistent with this hypothesis, preclinical studies demonstrate that both ghrelin and ethanol activate the cholinergic-dopaminergic reward link, implying neurochemical analogies between ghrelin and ethanol. This supports the hypothesis that ghrelin is involved in mediating the rewarding properties of ethanol. Additional animal experiments demonstrate that central ghrelin action not only stimulates reward processing but is also required for alcohol stimulation of that system. Human studies show reduced ghrelin levels in actively drinking alcoholics; increased ghrelin levels during alcohol abstinence; and a positive correlation between ghrelin level and alcohol craving scores. The CPN section aims to further investigate the ghrelin pathway in alcoholism in order to demonstrate its putative role in the biological and behavioural mechanisms that lead to alcohol craving and drinking. The ultimate goal of this research is to demonstrate that the ghrelin system may represent a novel pharmacological target for the treatment of alcoholism. Phenotypes for promising medications: focus on the GABAB pathway The selective GABAB receptor agonist baclofen has been identified as a possible medication to reduce alcohol craving and intake in alcoholic individuals. In keeping with several preclinical studies, most clinical studies have demonstrated baclofen’s effects in reducing alcohol craving and intake and promoting alcohol abstinence. However, one trial with heavy drinking individuals with a low severity of dependence reported a robust treatment effect, but no differences between baclofen and placebo were found. The inconsistency of baclofen’s effects on alcohol drinking among previous treatment trials suggests that different alcoholic individuals may respond

47
differently to baclofen. Baclofen consistently reduces anxiety in alcoholic patients, and analyses of positive vs. null findings with baclofen suggest that alcoholic patients with higher levels of anxiety at baseline may represent a sub-population particularly responsive to baclofen treatment. One of our goals is to assess under well-controlled laboratory conditions the effects of baclofen in alcoholics with high anxiety levels. The overall goal of this research is to identify phenotypes that respond best to a pharmacotherapy (baclofen) in order to develop more personalized treatments. Ecological assessment of alcohol craving and drinking We conduct clinical studies that utilize a number of techniques including computer-assisted self-administration of ethanol (in collaboration with the HP Section at LCTS) and fMRI (in collaboration with the BEI Section at LCTS). Furthermore, we conduct studies that investigate the effects of putatively effective medications under well-controlled conditions, but in a setting that mirrors a ‘real world’ scenario where individuals usually drink. In order to operationalize this, we have recently developed a simulated bar-like environment that acts as a human laboratory room (Figure 1). In this mock ‘real-world’ setting, we test the effect of novel medications and collect accurate behavioral and biological markers of medication response.
Figure 1. Human laboratory testing room at CPN. Key Prior References
1. Addolorato G, Leggio L, Abenavoli L, Agabio R, Caputo F, et al. 2006. Baclofen in the treatment of alcohol withdrawal syndrome: a comparative study vs diazepam. American Journal of Medicine 119:276.e13-8.

48
2. Addolorato G, Leggio L, Ferrulli A, Cardone S, Vonghia L, et al. 2007. Effectiveness and safety of baclofen for maintenance of alcohol abstinence in alcohol-dependent patients with liver cirrhosis: randomised, double-blind controlled study. Lancet 370:1915-22.
3. Leggio L, Ferrulli A, Malandrino N, Miceli A, Capristo E, et al. 2008. Insulin but not insulin growth factor-1 correlates with craving in currently drinking alcohol-dependent patients. Alcoholism: Clinical and Experimental Research 32:450-8.
4. Leggio L, Ferrulli A, Cardone S, Malandrino N, Mirijello A, et al.2008. Relationship between the hypothalamic-pituitary-thyroid axis and alcohol craving in alcohol-dependent patients: a longitudinal study. Alcoholism: Clinical and Experimental Research 32:2047-53.
5. Leggio L, Ray LA, Kenna GA, Swift RM. 2009. Blood glucose level, alcohol heavy drinking, and alcohol craving during treatment for alcohol dependence: results from the Combined Pharmacotherapies and Behavioral Interventions for Alcohol Dependence (COMBINE) Study. Alcoholism: Clinical and Experimental Research 33:1539-44.
6. Ray LA, Krull JL, Leggio L. 2010. The Effects of Naltrexone Among Alcohol Non-Abstainers: Results from the COMBINE Study. Frontiers in Psychiatry 1:26.
7. Leggio L, Garbutt JC, Addolorato G. 2010. Effectiveness and safety of baclofen in the treatment of alcohol dependent patients. CNS and Neurological Disorders Drug Targets 9:33-44.
8. Leggio L. 2010. Role of the ghrelin system in alcoholism: Acting on the growth hormone secretagogue receptor to treat alcohol-related diseases. Drug News & Perspectives 23:157-66.
Publications 2011 – 2012
1. Montalto M, Gallo A, Ferrulli A, Visca D, Campobasso E, et al. 2011. Fecal calprotectin concentrations in alcoholic patients: a longitudinal study. European Journal of Gastroenterology and Hepatology 23:76-80.
2. Leggio L, Addolorato G, Cippitelli A, Jerlhag E, Kampov-Polevoy AB, Swift RM. 2011. Role of feeding-related pathways in alcohol dependence: A focus on sweet preference, NPY, and ghrelin. Alcoholism: Clinical and Experimental Research 35:194-202.
3. Caputo F, Francini S, Brambilla R, Vigna-Taglianti F, Stoppo M, et al. 2011. Sodium oxybate in maintaining alcohol abstinence in alcoholic patients with and without psychiatric comorbidity. European Neuropsychopharmacology 21:450-6.
4. Addolorato G, Leggio L, Ferrulli A, Cardone S, Bedogni G, et al. 2011. Dose-response effect of baclofen in reducing daily alcohol intake in alcohol dependence: secondary analysis of a randomized, double-blind, placebo-controlled trial. Alcohol and Alcoholism 46:312-7.

49
5. Edwards S, Kenna GA, Swift RM, Leggio L. 2011. Current and promising pharmacotherapies, and novel research target areas in the treatment of alcohol dependence: a review. Current Pharmaceutical Design 17:1323-32.
6. Leggio L, Kenna GA, Ferrulli A, Zywiak WH, Caputo F, et al. 2011. Preliminary findings on the use of metadoxine for the treatment of alcohol dependence and alcoholic liver disease. Human Psychopharmacology 26:554-9.
7. Leggio L, Ferrulli A, Cardone S, Nesci A, Miceli A, et al. 2012. Ghrelin system in alcohol-dependent subjects: role of plasma ghrelin levels in alcohol drinking and craving. Addiction Biology 17:452-64.
8. Kenna GA, Swift RM, Hillemacher T, Leggio L. 2012. The relationship of appetitive, reproductive and posterior pituitary hormones to alcoholism and craving in humans. Neuropsychology Review 22:211-28.
9. Tang YL, Hao W, Leggio L. 2012. Treatments for alcohol-related disorders in China: a developing story. Alcoholism and Alcoholism 47:563-70.
10. Caputo F, Vignoli T, Leggio L, Addolorato G, Zoli G, et al. 2012. Alcohol use disorders in the elderly: a brief overview from epidemiology to treatment options. Experimental Gerontology 47:411-6.
11. Leggio L, Promrat K, Kenna GA. 2012. A Call to Action: Alcohol Interventions in HIV-Infected Patients. Frontiers in Psychiatry 3:35.
12. Leggio L, Ferrulli A, Zambon A, Caputo F, Kenna GA, et al. 2012. Baclofen promotes alcohol abstinence in alcohol dependent cirrhotic patients with hepatitis C virus (HCV) infection. Addictive Behaviors 37:561-4.
13. Addolorato G, Leggio L, Hopf FW, Diana M, Bonci A. 2012. Novel therapeutic strategies for alcohol and drug addiction: focus on GABA, ion channels and transcranial magnetic stimulation. Neuropsychopharmacology 37:163-77.
14. Heberlein A, Leggio L, Stichtenoth D, Hillemacher T. 2012. The treatment of alcohol and opioid dependence in pregnant women. Current Opinion in Psychiatry 25:559-64.
15. Kenna GA, Roder-Hanna N, Leggio L, Zywiak WH, Clifford J, et al. 2012. Association of the 5-HTT gene-linked promoter region (5-HTTLPR) polymorphism with psychiatric disorders: review of psychopathology and pharmacotherapy. Pharmacogenomics and personalized medicine 5:19-35.
16. Leggio L, Zywiak WH, McGeary JE, Edwards S, Fricchione SR, et al. 2012. A human laboratory pilot study with baclofen in alcoholic individuals. Pharmacology, Biochemistry and Behavior 103:784-791.
17. Leggio L, Kenna GA. 2012. Doxasozin for Alcoholism. Alcoholism: Clinical and Experimental Research Dec 27. doi: 10.1111/acer.12064.
Dr. Leggio: [email protected]

50
______________________________________________________________________
Laboratory of Membrane Biochemistry and Biophysics ______________________________________________________________________ Klaus Gawrisch, PhD, Chief The mission of the Laboratory of Membrane Biochemistry & Biophysics (LMBB), established in 1992, is to study the alterations in cell membrane structure and function caused by alcohol abuse with a focus upon polyunsaturated lipids. The lab has a particular emphasis on the most highly unsaturated essential fatty acid found in the mammalian tissues, docosahexaenoic acid (DHA). This fatty acid typically occurs as a phospholipid in brain and retinal membranes where it is highly enriched and so these systems are of great interest to those within the laboratory. A nutritional approach to membrane structure and composition is often taken both with respect to essential fatty acid profiles and alcohol content since these are powerful modulators of lipid content and membrane properties. Methodology The LMBB is truly multi-disciplinary in its methodological approaches which include psychiatry, nutritional neuroscience, pharmacology, toxicology, biochemistry, biophysics, and spectroscopy. Clinical studies are performed with the cooperation of the clinical laboratory within DICBR. This currently involves a randomized, placebo controlled trial of the effects of dietary fatty acid intake on the spectrum of fatty acids in the human body, on craving, impulsivity and relapse in aggressive alcoholics. Recent metabolic trials have included the use of orally administered stable isotopically labeled essential fatty acids in adult alcoholics and controls as well as in newborn infants. Biochemical studies include molecular biological techniques as well as studies of essential fatty acid metabolism using sophisticated tracer techniques and negative chemical ionization (NCI) GC/MS as the detection tool, methods developed within our laboratory. Artificial membranes are formed using traditional liposomes where proteins like rhodopsin and the cannabinoid type II receptor, CB2, are incorporated and also newer technology developed within the NMR Section using flow-through lipid nanotube arrays made up of anodic aluminum oxide. Receptor activation can then be studied using ligand binding and G protein activation assays as well as various spectroscopic techniques. A range of biophysical studies are conducted using solid state NMR, including magic angle spinning in combination with multidimensional NMR techniques, NMR diffusion measurements with application of pulsed magnetic field gradients. Our principal equipment includes 500 and 800 MHz nuclear magnetic resonance spectrometers. The laboratory is also equipped with differential scanning and isothermal titration calorimetric equipment, time resolved and steady state fluorimeters and circular dichroism spectrometers. The LMBB is also pioneering the application of robotic technology for chemical derivatization as part of the development of high throughput methods for fatty acid analysis. It can be seen then that a full spectrum of approaches is taken to study the functions of polyunsaturated lipids ranging from the clinical to NMR studies at the molecular level.

51
LMBB consists of three sections: the Section on Nutritional Neuroscience which is led by CAPT Joseph R. Hibbeln, MD; the Section on Nuclear Magnetic Resonance, led by Klaus Gawrisch, PhD; and the Section on Molecular Pharmacology and Toxicology, led by B. J. Song, PhD. Laboratory administrative personnel are within the Office of the Chief. Accomplishments The activities of the individual sections are described separately in greater detail in the chapters that follow. Major contributions of the LMBB include: 1. Suicides rates among active duty US Military have increased to record numbers. Deficiencies of highly unsaturated omega-3 polyunsaturated fatty acids (n-3 HUFAs) may be a significant contributing risk factor for psychiatric pathology, including suicide. Among n=1,600 active duty US military, risk of suicide death was 62% greater with low serum DHA status. Subsequently, a $10 million grant has been awarded to conduct a randomized controlled trial (RCT) to assess if 4 gm/ day of n-3 HUFAs are effective in reducing risk of suicidal behaviors among US Military veterans. CAPT Joseph R. Hibbeln, M.D. SNN, LMBB and Prof. Bernadette Marriott, Ph.D. the Medical University of South Carolina are co- principle investigators on the award. 2. Advice to substitute polyunsaturated fats for saturated fats is a key component of worldwide dietary guidelines for coronary heart disease risk reduction. However, clinical benefits of the most abundant polyunsaturated fatty acid, omega-6 linoleic acid, have not been established. We recovered data from the Sydney Diet Heart Study, a single blinded, parallel group, randomized controlled trial conducted in 1966-73. Interventions were replacement of dietary saturated fats with omega 6 linoleic acid sources. The intervention group (n=221) had higher rates of death than controls (n=237) (all cause %, hazard ratio 1.62 (95% confidence interval 1.00 to 2.64), P=0.05; cardiovascular disease 1.70 (1.03 to 2.80), P=0.04; coronary heart disease 1.74 (1.04 to 2.92), P=0.04). These findings could have important implications for worldwide dietary advice to substitute omega-6 linoleic acid, or polyunsaturated fats without specificity, for saturated fats. 3. Suppressing hyperactive endocannabinoid tone is a critical target for reducing obesity. The backbone of both endocannabinoids 2-arachidonoylglycerol (2-AG) and anandamide (AEA) is the omega-6 fatty acid arachidonic acid (AA). We posited that excessive dietary intake of omega-6 linoleic acid, (LA) the precursor of AA, would induce endocannabinoid hyperactivity and promote obesity in rodents. Increasing dietary LA from 1 percent of energy (en%) to 8 en% elevated AA-phospholipids (PL) in liver and erythrocytes, tripled 2-AG + 1-AG and AEA associated with increased food intake, feed efficiency, and adiposity in mice. These animal diets modeled 20th century increases of human LA consumption which closely correlates with increasing prevalence rates of obesity. 4. Genetic variants may interact with dietary intakes of essential fatty acids to increase, or decrease the risk of early developmental disorders including low IQ conduct disorder and delinquent behaviors. This cluster of behaviors in children substantially increases the likelihood of developing substance abuse and criminally deviant behaviors later in life. Here we examined interactions of variants in the FADS-2 gene with breast vs. formula feeding 5934 children aged 8

52
years. Breast milk contains preformed products so the FADS-2 gene while formula contained only precursors. Among formula fed children IQ was 4.3 points higher for the GG genotype (that could produce HUFAs) compared to CC+CG genotypes (suboptimal HUFA production). These data indicate a specific link between gene variants, metabolism in that pathway and a meaningful functional outcome. 5. The Dietary Guidance for Americans 2010 was updated to contain advice for pregnant women to consume at least 8 to 12 oz of seafood per week directly as a result of our 2007 Lancet publication. In that study we evaluated the relative risks and benefits of fish consumption during pregnancy in a large epidemiological study. Women whose seafood consumption during pregnancy were in concordance with the limits advised in 2004 by the FDA and EPA, create nutritional inadequacies for their children during early development including low verbal IQ and deviant prosocial and peer behaviors. 6. A key mechanism for the allosteric modulation of rhodopsin function has been established – the enhancement of rhodopsin activation by negative curvature elastic stress in the lipid matrix. Curvature stress is caused by an asymmetry in the shape of lipid molecules. Lipids with small headgroups like phosphatidylethanolamines and wide hydrocarbon chains like the polyunsaturated DHA form monolayers with negative curvature. In a flat lipid matrix, these lipids experience negative curvature elastic stress that, upon photoactivation, favors formation of metarhodopsin-II (MII), the photointermediate that activates G protein. Functional studies with reconstitution of rhodopsin into a large set of different lipids yielded excellent quantitative agreement between curvature elastic properties of lipid monolayers and MII formation. Curvature elastic properties were determined independently in inverse hexagonal lipid phases. Moreover, since all G protein-coupled receptors (GPCR) of class A undergo a similar conformational change upon activation, such allosteric modulation by negative curvature elastic stress is likely to occur in many other GPCR as well. 7. Cannabinoid receptors are one of the most ubiquitous GPCR and have attracted much attention because of their role in health and disease including drugs of abuse and alcoholism. At our laboratory, the human cannabinoid type II receptor, CB2, is recombinantly expressed in E. coli at the level of milligrams, purified, and successfully reconstituted in the functional form into lipid bilayers. The receptor was sufficiently stabilized in the micellar phase by a proper mixture of detergents, lipid additives, and strongly binding ligands to enable efficient purification and reconstitution into membranes of controlled composition. Upon reconstitution, the receptor regains long-term stability that enables a variety of functional and structural studies. The recombinant CB2 fully retained its ligand binding properties and is capable of G protein activation. 8. Structural studies on the GPCR rhodopsin revealed that the lipid matrix adjusts its hydrophobic thickness to the length of hydrophobic transmembrane helices of GPCR. For rhodopsin, the optimal hydrophobic thickness of the lipid bilayer is about 27±1 Å. Any mismatch severely alters rhodopsin function due to additional curvature elastic stress in the lipid matrix and as a result of GPCR aggregation. Furthermore, GPCR adjust their structure to the properties of the lipid matrix far more nimbly than generally assumed. For rhodopsin, with increasing hydrophobic thickness of bilayers an increase of helical content was observed.

53
9. An efficient redox proteomics method to identify and characterize oxidatively-modified proteins was developed. This simple method has been used to identify such proteins in animal models of alcoholic and non-alcoholic fatty liver disease caused by high fat or ischemia-reperfusion injury. We also used this redox proteomics method in translational research by demonstrating the beneficial effects of polyunsaturated fatty acids including DHA on reducing alcohol-mediated oxidative stress, mitochondrial dysfunction and fatty liver. 10. We used peroxisomal proliferator activated receptor α (PPARα)-null mice and CYP2E1-null mice, respectively, to study the molecular mechanisms of alcohol, high fat, and drug-induced nitroxidative stress, mitochondrial dysfunction, fat accumulation, inflammation, and subsequent organ damage. We recently showed that CYP2E1 is important in the development of insulin resistance and nonalcoholic steatohepatitis caused by high fat diets. 11. We recently reported that CYP2E1 is important in promoting protein nitration, although the identities and functions of nitrated proteins are unknown. Therefore, we developed an antibody-based purification method to identify nitrated proteins in acetaminophen-exposed mice. Mass-spectral results of the purified proteins confirmed that many mitochondrial proteins were nitrated and all five enzymes we examined were inactivated. Time-dependent experiments with or without antioxidant N-acetylcysteine showed that nitration of many proteins at early time points contributed to mitochondrial dysfunction and liver injury at later time points, indicating a causal role of protein nitration in promoting liver injury. This method is being applied to further studies of characterizing nitrated proteins in alcohol-exposed animals and human pathological specimens. 12. We have reported that JNK phosphorylates Bax to promote mitochondrial-dependent apoptosis and acute liver injury by toxic substrates of CYP2E1. However, identities and functional roles of many other JNK-target proteins are relatively unknown. Therefore, we also developed a purification method to identify JNK-target phospho-proteins in mitochondria from mouse models of acute liver injury by carbon tetrachloride or acetaminophen. Our data showed that JNK was rapidly activated, then translocated to mitochondria and phosphorylated mitochondrial proteins. Mass-spectral data of the affinity-purified phospho-proteins showed that more than 100 mitochondrial proteins were phosphorylated in carbon tetrachloride-exposed mice. Time-dependent experiments with or without specific JNK inhibitors further revealed that JNK-mediated phosphorylation of many mitochondrial proteins at early time points is critical to mitochondrial dysfunction and liver injury at later time points, indicating a causal role of protein phosphorylation in liver injury. This method is being used to study functional characteristics of phospho-proteins in animals exposed to binge alcohol and human pathological specimens. Dr. Gawrisch: [email protected]

54
______________________________________________________________________
Section on Nuclear Magnetic Resonance
______________________________________________________________________ Klaus Gawrisch, PhD, Chief Alexei A. Yeliseev, PhD, Staff Scientist Olivier Soubias, PhD, Staff Scientist John K. Northup, Guest Researcher Tomohiro Kimura, PhD, Research Fellow Silvia Locatelli-Hoops, PhD, Contractor Walter E. Teague, Jr., PhD, Physical Chemist Kirk G. Hines, BS, Biologist Jeffrey Dwulit-Smith, BS, Postbac IRTA Lioudmila Zoubak, MS, Contractor
We conduct structural and functional studies on reconstituted G protein-coupled membrane receptors (GPCR). The goal is to investigate GPCR at functional conditions in a fluid lipid matrix with a biologically relevant composition of lipids. In particular the neuronal GPCR are located in membranes containing high concentrations of polyunsaturated fatty acids (PUFA). The mechanisms by which polyunsaturated lipid species may influence biological function at the molecular level have attracted considerable attention. Two mechanisms of PUFA action on proteins are considered: on one hand, PUFA may influence function by altering biophysical properties of the lipid matrix as well as by PUFA-specific interactions at the lipid/protein interface. On the other hand, PUFA and their derivatives may act as ligands that interact with membrane proteins specifically.
l to r, top: KG Hines, S Locatelli-Hoops, T Kimura, J Dwulit-Smith, K Gawrisch bottom: O Soubias, WE Teague Jr., AA Yeliseev, JK Northup, L Zoubak

55
Bacterial expression, purification, and functional reconstitution of the cannabinoid receptor type II, CB2, for functional and structural studies Yeliseev, Kimura, Locatelli-Hoops, Dwulit-Smith, Zoubak, Hines, Northup Cannabinoid receptors are one of the most ubiquitous GPCR and have attracted much attention because of their role in health and disease including drugs of abuse and alcoholism. They belong to the large family of rhodopsin-like class A GPCR that has several hundred protein members. Cannabinoid receptors are also remarkable because their endogenous ligands, 2-arachidonylglycerol (2-AG) and N-arachidonylethanolamine (anandamide), are synthesized from lipid-bound, arachidonic acid hydrocarbon chains (AA, 20:4n6). It had been suggested that availability of endogenous ligands in tissues may be influenced by the nutritional balance of ω-3 to ω-6 fatty acids, a hypothesis which was confirmed recently for 2-AG in studies of the Section of Nutritional Neurosciences of LMBB. We are interested both type I and type II cannabinoid receptors, CB1 and CB2. However, the higher inherent instability of recombinantly expressed CB1 led to the decision to tackle CB2 first.
Fig. 1. Snake plot of the human cannabinoid type 2 (CB2) receptor. Amino-acid residues in the wild-type receptor are shown with gray background. The recombinant CB2 receptor that was cleaved from the CB2-130 fusion protein contains additional amino acids in N-, and C-terminal ends (white background) (Yeliseev, et al., 2005; Yeliseev, et al., 2007). CB2 is primarily localized in the peripheral tissues of the immune system. It has been found at very high concentrations in monocytes, macrophages, B-cells, and T-cells where it is involved in immune regulation. The class of cannabinoid receptors, like other GPCR have become the object of high interest, in part because of their role as therapeutically important targets of drugs. One of the major difficulties in rational design and synthesis of potential modulators of GPCR activity is the lack of information on GPCR structure and function at the molecular level. The first and most important step for any structural study at high resolution is to obtain milligram quantities of pure and functional receptor at high concentration. A variety of methods for structural and functional characterization will be employed but main emphasis is on nuclear magnetic resonance (NMR). NMR studies may provide valuable insights into structure and function of CB2 at near physiological conditions.
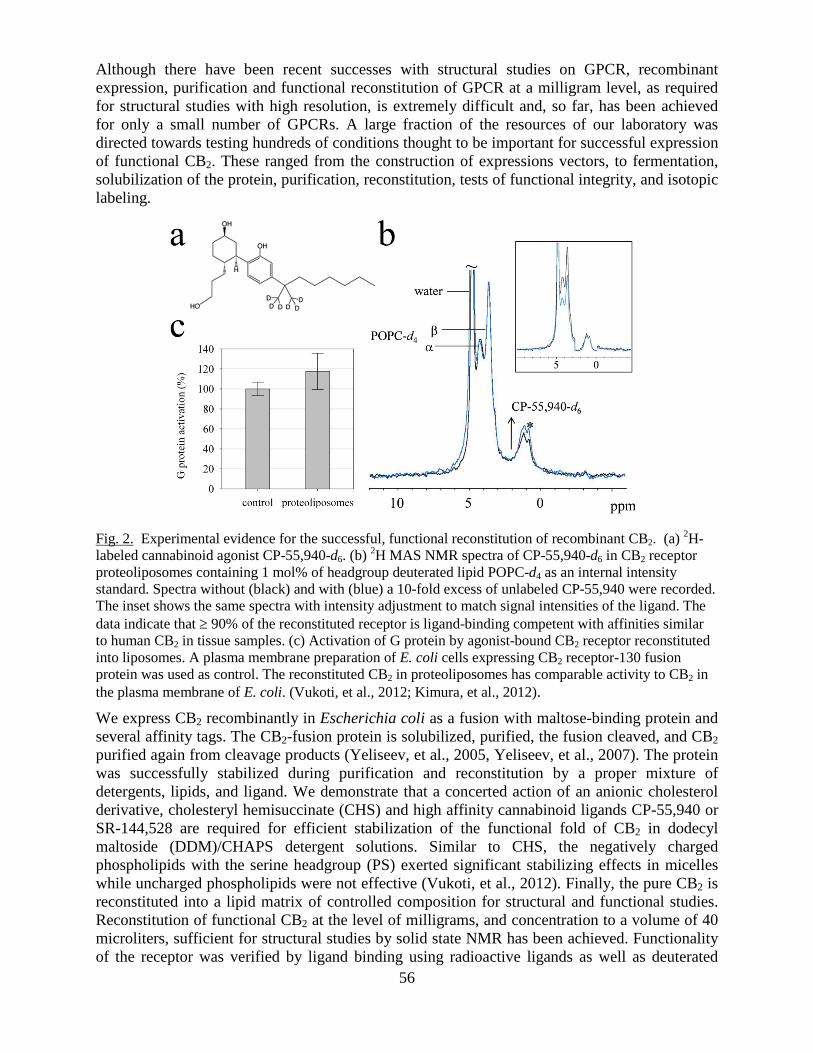
56
Although there have been recent successes with structural studies on GPCR, recombinant expression, purification and functional reconstitution of GPCR at a milligram level, as required for structural studies with high resolution, is extremely difficult and, so far, has been achieved for only a small number of GPCRs. A large fraction of the resources of our laboratory was directed towards testing hundreds of conditions thought to be important for successful expression of functional CB2. These ranged from the construction of expressions vectors, to fermentation, solubilization of the protein, purification, reconstitution, tests of functional integrity, and isotopic labeling.
Fig. 2. Experimental evidence for the successful, functional reconstitution of recombinant CB2. (a) 2H-labeled cannabinoid agonist CP-55,940-d6. (b) 2H MAS NMR spectra of CP-55,940-d6 in CB2 receptor proteoliposomes containing 1 mol% of headgroup deuterated lipid POPC-d4 as an internal intensity standard. Spectra without (black) and with (blue) a 10-fold excess of unlabeled CP-55,940 were recorded. The inset shows the same spectra with intensity adjustment to match signal intensities of the ligand. The data indicate that ≥ 90% of the reconstituted receptor is ligand-binding competent with affinities similar to human CB2 in tissue samples. (c) Activation of G protein by agonist-bound CB2 receptor reconstituted into liposomes. A plasma membrane preparation of E. coli cells expressing CB2 receptor-130 fusion protein was used as control. The reconstituted CB2 in proteoliposomes has comparable activity to CB2 in the plasma membrane of E. coli. (Vukoti, et al., 2012; Kimura, et al., 2012).
We express CB2 recombinantly in Escherichia coli as a fusion with maltose-binding protein and several affinity tags. The CB2-fusion protein is solubilized, purified, the fusion cleaved, and CB2 purified again from cleavage products (Yeliseev, et al., 2005, Yeliseev, et al., 2007). The protein was successfully stabilized during purification and reconstitution by a proper mixture of detergents, lipids, and ligand. We demonstrate that a concerted action of an anionic cholesterol derivative, cholesteryl hemisuccinate (CHS) and high affinity cannabinoid ligands CP-55,940 or SR-144,528 are required for efficient stabilization of the functional fold of CB2 in dodecyl maltoside (DDM)/CHAPS detergent solutions. Similar to CHS, the negatively charged phospholipids with the serine headgroup (PS) exerted significant stabilizing effects in micelles while uncharged phospholipids were not effective (Vukoti, et al., 2012). Finally, the pure CB2 is reconstituted into a lipid matrix of controlled composition for structural and functional studies. Reconstitution of functional CB2 at the level of milligrams, and concentration to a volume of 40 microliters, sufficient for structural studies by solid state NMR has been achieved. Functionality of the receptor was verified by ligand binding using radioactive ligands as well as deuterated

57
ligands in combination with 2H-MAS NMR and by G protein activation studies using recombinantly produced G protein in a [35S]GTP-γ-S assay. Composition, size, and homogeneity of proteoliposomes were investigated by analytical NMR, fluorescence spectroscopy using labeled lipid and CB2, dynamic light scattering, and sucrose gradient centrifugation (Kimura, et al., 2012).
While the receptor proved to be vulnerable to degradation in micellar solution, the reconstituted CB2 in a lipid matrix at sufficiently low temperatures has long-term stability that enables functional and structural studies (Vukoti, et al., 2012). Exploratory NMR experiments conducted on a 2-mg sample of homogeneously 13C- and -15N labeled CB2 (Berger, et al., 2010) and comparison of experimental results with simulated spectra obtained from the atomic coordinates of a CB2 model (Hurst, et al., 2010) have demonstrated feasibility of the experimental concept. Specific isotopic labeling schemes have been developed to achieve the desired spectral resolution for a structural analysis.
The lipid matrix as allosteric modulator of rhodopsin function Soubias, Teague, Hines, Dwulit-Smith, Northup Retinal membranes of mammals, like synaptosomal membranes in brain, contain up to 50 mol% of lipid with the ω-3 hydrocarbon chain docosahexaenoic acid (DHA, 22:6n3). Rhodopsin, the mammalian dim-light photoreceptor, is ideally suited to study the importance of a polyunsaturated lipid matrix for GPCR function. Membrane composition strongly modulates the early steps of the visual process in rod cells. Photoactivation of rhodopsin in lipid bilayers results within milliseconds in a metarhodopsin I (MI) metarhodopsin II (MII) equilibrium that is very sensitive to the lipid composition. MII is the rhodopsin photointermediate that is capable of activating G protein. The strong similarities in structure and function of class A, rhodopsin-like GPCR suggests that results on lipid-rhodopsin interaction could be generalized to function of other GPCR.
Phosphoethanolamines (PE) comprise about 40% of retinal and synaptosomal membranes and, within these membranes, about 50% of hydrocarbon chains in PE are the six-fold unsaturated docosahexaenoic acid (22:6n-3, DHA). Increased levels of PE significantly boost formation of the photointermediate metarhodopsin-II (MII) that is capable of activating the G protein transducin. PE lipids have the tendency to form inverse hexagonal lipid phases (HII) composed of highly curved monolayers with small area per lipid at the PE headgroup and a larger area near the terminal methyl groups of hydrocarbon chains. When forced into a lamellar arrangement, those monolayers are under considerable negative curvature elastic stress.
Fig. 3: The lipids 1-perdeuterio-stearyl-2-docosahexaenoyl-sn-glycero-3-phosphoethanolamine (18:0d35-22:6n-3PE) and 1-perdeuterio-stearyl-2-docosapentaenoyl-sn-glycero-3-phosphoethanolamine (18:0d35-22:5n-6PE).
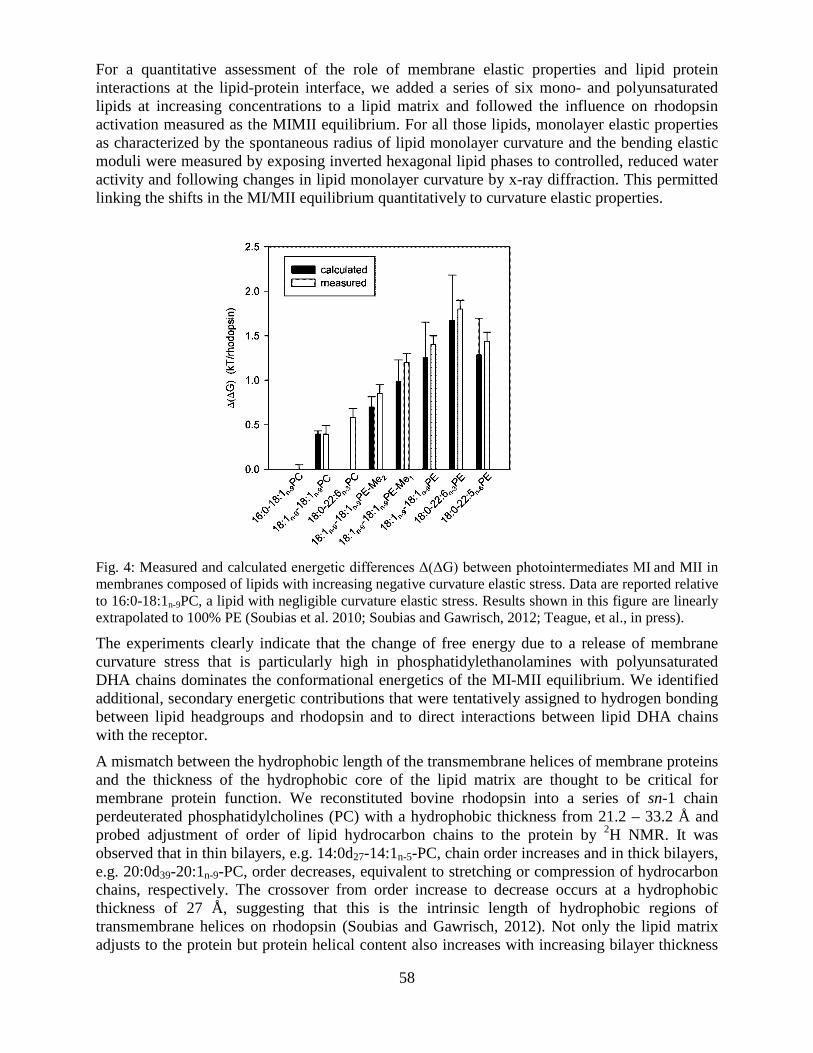
58
For a quantitative assessment of the role of membrane elastic properties and lipid protein interactions at the lipid-protein interface, we added a series of six mono- and polyunsaturated lipids at increasing concentrations to a lipid matrix and followed the influence on rhodopsin activation measured as the MIMII equilibrium. For all those lipids, monolayer elastic properties as characterized by the spontaneous radius of lipid monolayer curvature and the bending elastic moduli were measured by exposing inverted hexagonal lipid phases to controlled, reduced water activity and following changes in lipid monolayer curvature by x-ray diffraction. This permitted linking the shifts in the MI/MII equilibrium quantitatively to curvature elastic properties.
Fig. 4: Measured and calculated energetic differences Δ(ΔG) between photointermediates MI and MII in membranes composed of lipids with increasing negative curvature elastic stress. Data are reported relative to 16:0-18:1n-9PC, a lipid with negligible curvature elastic stress. Results shown in this figure are linearly extrapolated to 100% PE (Soubias et al. 2010; Soubias and Gawrisch, 2012; Teague, et al., in press).
The experiments clearly indicate that the change of free energy due to a release of membrane curvature stress that is particularly high in phosphatidylethanolamines with polyunsaturated DHA chains dominates the conformational energetics of the MI-MII equilibrium. We identified additional, secondary energetic contributions that were tentatively assigned to hydrogen bonding between lipid headgroups and rhodopsin and to direct interactions between lipid DHA chains with the receptor.
A mismatch between the hydrophobic length of the transmembrane helices of membrane proteins and the thickness of the hydrophobic core of the lipid matrix are thought to be critical for membrane protein function. We reconstituted bovine rhodopsin into a series of sn-1 chain perdeuterated phosphatidylcholines (PC) with a hydrophobic thickness from 21.2 – 33.2 Å and probed adjustment of order of lipid hydrocarbon chains to the protein by 2H NMR. It was observed that in thin bilayers, e.g. 14:0d27-14:1n-5-PC, chain order increases and in thick bilayers, e.g. 20:0d39-20:1n-9-PC, order decreases, equivalent to stretching or compression of hydrocarbon chains, respectively. The crossover from order increase to decrease occurs at a hydrophobic thickness of 27 Å, suggesting that this is the intrinsic length of hydrophobic regions of transmembrane helices on rhodopsin (Soubias and Gawrisch, 2012). Not only the lipid matrix adjusts to the protein but protein helical content also increases with increasing bilayer thickness

59
increasing hydrophobic thickness. A hydrophobic mismatch between rhodopsin and the surrounding lipid matrix promotes rhodopsin oligomerization the extent of which is protein concentration dependent. Oligomerization shifts the MI/MII equilibrium towards MI and reduces the amplitude of protein-induced adjustments of lipid order. The observation is likely to be applicable to the entire class of rhodopsin-like G protein-coupled membrane receptor.
MII
MI
Fig. 5: Cartoon highlighting the structural differences between the rhodopsin photointermediates MI and MII. Structural data on MIand MII suggest that MII tends to be more hourglass-shaped. This generates negative curvature in lipid monolayers near the protein. Please note that structural differences are exaggerated for clarity.
GPCR reconstitution into solid-supported layers
Soubias, Locatelli-Hoops, Gorshkova, Vaish, Silin, Teague, Vanderah, Yeliseev Immobilization of GPCR at solid interfaces offers unique opportunities for functional and structural studies at high sensitivity like surface plasmon resonance (SPR) and neutron scattering. If combined with the use of porous substrates like anodic aluminum oxide filters (AAO), the surface area per unit of volume is sufficiently large for application of NMR. We followed two strategies for protein immobilization. (i) Expression of CB2 as a fusion with Rho-tag (peptide composed of the last nine amino acids of rhodopsin) in E. coli was evaluated in terms of protein levels, accessibility of the tag, and activity of the receptor. Highly pure and functional CB2 was obtained and its affinity to the monoclonal 1D4 antibody that binds the Rho-tag characterized by SPR. Either the purified receptor or fusion CB2 from the crude cell extract was captured onto a 1D4 -coated CM4 chip (Biacore) in a uniform orientation, in a quantitative fashion as demonstrated by the SPR signal. The accessibility of the extracellular surface of immobilized CB2 for specific interaction with the novel monoclonal antibody raised against reconstituted, recombinant CB2 was demonstrated. The method is ready for use in functional studies on CB2. Furthermore, (ii) in collaboration with the National Institute of Standards and Technology we fabricated chips for SPR using novel, water soluble nitrilotriacetic acid (NTA)-terminated oligo(ethylene glycol) thiols that were synthesized at NIST to capture histidine-tagged GPCR in a defined orientation with negligible nonspecific binding. Experiments were conducted with His-tagged, recombinant CB2. It was demonstrated that the membrane protein surface density can be controlled by tuning the surface density of the capture molecule in mixed, self-assembled monolayers. The advantage of the method is that GPCR are captured without the

60
use of a tethered large biomolecule like an antibody which is particularly useful for structural studies, e.g. by neutron diffraction (Vaish, et al., in press).
Last but not least, we developed reagents and methods for formation of tubular, single lipid bilayers containing high concentrations of GPCR that cover the pore walls of porous anodic aluminum oxide (AAO) filters. One square centimeter of the filter material with a thickness of 60 micrometers yields up to 500 cm2 of oriented membranes, which is sufficient for multinuclear solid state NMR studies. To obtain meaningful information on the influence of membrane spanning proteins on the surrounding lipid matrix, it is of critical importance to study reconstituted membranes of controlled curvature, free of detergents, and fully hydrated. The method fulfills all the requirements and has the additional advantage of yielding single bilayers that are freely accessible to the water phase from one side which allows unimpeded delivery of ligands, and oriented bilayers that are packed in the substrate at very high density. The latter results in a significant gain in sensitivity and resolution of NMR resonances, such that experiments on single, supported lipid bilayers are feasible. The majority of our recent NMR experiments on rhodopsin have been aided by the use of AAO filters.
References
Berger C, Ho JTC, Kimura T, Hess S, Gawrisch K, Yeliseev A. 2010. Preparation of stable isotope-labeled peripheral cannabinoid receptor CB2 by bacterial fermentation. Protein Expression Purification 70: 236-47
Figure 6. Schematic representation of single lipid bilayers adsorbed to the inner surface of an AAO pore. The reconstituted GPCR is shown as red rectangles. A single bilayer forms a good seal with the AAO surface by the interaction of a small percentage of the lipids. The lipids adsorb as wavy tubules. These tubules possess undulation with a radius of curvature of 100-400 nm. Trapped between the tubules and the AAO surface are pockets of water with an average thickness of 3 nm. To the right from the enlarge pore an illustration of shape of the lipid bilayer tubules inside the AAO pore is shown (Soubias and Gawrisch, in press).

61
Hurst DP, Grossfield A, Lynch DL, Feller S, Romo TD, Gawrisch K, Pitman MC, Reggio PH. 2010. A lipid pathway for ligand binding is necessary for a cannabinoid G protein-coupled receptor. J. Biol. Chem. 285: 17954-64
Kimura T, Yeliseev AA, Vukoti K, Rhodes SD, Cheng K, Rice KC, Gawrisch K. 2012. Recombinant Cannabinoid Type 2 Receptor in Liposome Model Activates G Protein in Response to Anionic Lipid Constituents. J. Biol. Chem. 287: 4076-87
Soubias O, Gawrisch K. 2012. The role of the lipid matrix for structure and function of the GPCR rhodopsin. Biochim. Biophys. Acta 1818: 234-40
Soubias O, Niu SL, Mitchell DC, Gawrisch K. 2008. Lipid-rhodopsin hydrophobic mismatch alters rhodopsin helical content. J. Am. Chem. Soc. 130: 12465-71 Soubias O, Teague WE, Hines KG, Mitchell DC, Gawrisch K. 2010. Contribution of Membrane elastic energy to rhodopsin function. Biophys. J. 99: 817-824
Vukoti K, Kimura T, Macke L, Gawrisch K, Yeliseev A. 2012. Stabilization of Functional Recombinant Cannabinoid Receptor CB2 in Detergent Micelles and Lipid Bilayers. Plos One 7: e46290
Yeliseev A, Zoubak L, Gawrisch K. 2007. Use of dual affinity tags for expression and purification of functional peripheral cannabinoid receptor. Protein Expression Purification 53: 153-63 Yeliseev AA, Wong KK, Soubias O, Gawrisch K. 2005. Expression of human peripheral cannabinoid receptor for structural studies. Protein Sci. 14: 2638-53
Publications 2011-2012
1. Mihailescu M, Soubias O, Worcester D, White SH, Gawrisch K. 2011. Structure and Dynamics of Cholesterol-Containing Polyunsaturated Lipid Membranes Studied by Neutron Diffraction and NMR. J. Membrane Biol. 239: 63-71
2. Vukoti K, Kimura T, Macke L, Gawrisch K, Yeliseev A. 2012. Stabilization of Functional Recombinant Cannabinoid Receptor CB2 in Detergent Micelles and Lipid Bilayers. Plos One 7: e46290
3. Kimura T, Yeliseev AA, Vukoti K, Rhodes SD, Cheng K, Rice KC, Gawrisch K. 2012. Recombinant Cannabinoid Type 2 Receptor in Liposome Model Activates G Protein in Response to Anionic Lipid Constituents. J. Biol. Chem. 287: 4076-87
4. Soubias O, Gawrisch K. 2012. The role of the lipid matrix for structure and function of the GPCR rhodopsin. Biochim. Biophys. Acta 1818: 234-40
5. Krepkiy D, Gawrisch K, Swartz KJ. 2012. Structural Interactions between Lipids, Water and S1-S4 Voltage-Sensing Domains. J. Mol. Biol. 423: 632-47
6. Gawrisch K. 2012. Tafazzin senses curvature. Nature Chem. Biol. 8: 811-2 Colloborators
Christian Berger, PhD, Inst. Med. Phys. Biophys., Univ. Leipzig, Germany Kejun Cheng, PhD, DDS, NIDA, NIH, Bethesda, MD Scott E. Feller, PhD, Dept. Chemistry, Wabash College, Crawfordsville, IN Inna I. Gorshkova, PhD, NIBIB, NIH, Bethesda, MD

62
Alan M. Grossfield, PhD, Dept. Biochem. Biophys., Univ. Rochester Med. Ct. Rochester, NY Daniel Huster, PhD, Inst. Med. Phys. Biophys., Univ. Leipzig, Germany Dmitriy V. Krepkiy, PhD, MPB, NINDS, NIH, Bethesda, MD Susan Krueger, PhD, NIST, Gaithersburg, MD Mihaela Mihailescu, PhD, Inst. Biosci. Biotech. Research, Univ. MD, Gaithersburg, MD Drake C. Mitchell, PhD, Dept. Physics Lib. Arts Sciences, Portland State Univ., Portland, OR Richard W. Pastor, PhD, BBC, NHLBI, NIH, Bethesda, MD Michael C. Pitman, PhD, IBM TJ Watson Research Center, Yorktown Heights, NY Patricia H. Reggio, PhD, Dept. Chem. Biochem., Univ North Carolina, Greensboro, NY Vitalii Silin, PhD, IBBR, NIST, Gaithersburg, MD Kenton J. Swartz, PhD, MPB, NINDS, NIH, Bethesda, MD Kenner C. Rice, PhD, CBRB, NIDA/NIAAA, NIH, Bethesda, MD Amit Vaish, PhD, IBBR, NIST, Gaithersburg, MD David J. Vanderah, PhD, IBBR, NIST, Gaithersburg, MD Sarah L. Veatch, PhD, Dept. Chem., Univ. Michigan, Ann Arbor, MI Stephen H. White, PhD, Dept. Physiol./Biophys., Univ. of California, Irvine, CA Joshua Zimmerberg, MD/PhD, LCMB, NICHD, NIH, Bethesda, MD
Dr. Gawrisch: [email protected]

63
______________________________________________________________________
Section of Molecular Pharmacology and Toxicology ______________________________________________________________________ Byoung-Joon (B.J.) Song, PhD, Chief Mohamed Abdelmegeed, MD, PhD, Research Fellow Sehwan Jang, PhD, Visiting Fellow Atrayee Banerjee, PhD, Postdoctoral IRTA Lauren E. Henderson, BS, Postbac IRTA Alcohol/drug metabolism, functional consequence, and signaling mechanisms in alcoholic and nonalcoholic fatty liver and tissue injury In my laboratory we study the functional role of two enzymes in promoting oxidative/nitrative tissue injury. The two enzymes are ethanol-inducible cytochrome P450 2E1 (CYP2E1) and mitochondrial aldehyde dehydrogenase (ALDH2). We also investigate the signaling mechanisms toward cell death upon exposure to potentially toxic compounds including ethanol, acetaminophen (APAP, a major ingredient of Tylenol), carbon tetrachloride (CCl4), nitrosamines and various abused drugs. In addition, we study the effect of high fat diets and other substances on causing nonalcoholic fatty liver (NAFLD) or tissue injury. CYP2E1, localized in both endoplasmic reticulum (ER) and mitochondria, is involved in the metabolism of more than 80 potentially toxic substrates. ALDH2 is involved in the metabolism of toxic acetaldehyde (AcAH), a product of ethanol metabolism, as well as lipid peroxides such as 4-hydroxynonenal (HNE) and malondialdehyde (MDA).
Figure 1. Major pathways of oxidative ethanol metabolism. Under chronic alcohol consumption or binge alcohol drinking, CYP2E1 is activated while ALDH2 activity is suppressed. The activity changes in the two enzymes CYP2E1 and ALDH2 are likely to increase oxidative/nitrative stress.
In the past, we have cloned the cDNAs for rat and human CYP2E1 and showed at least seven distinct means of regulation of CYP2E1 protein. CYP2E1 can be induced by many small molecules including ethanol and in pathophysiological conditions such as fasting, high fat diet,

64
and diabetes. Since year 2000, we have re-focused our research by studying the functional implications of activated CYP2E1 with the suppressed ALDH2 activity in alcohol- or drug-mediated oxidative/nitrative stress, mitochondrial dysfunction, and ER stress, contributing to cell/organ injury. Figure 1 shows the major pathways of ethanol metabolism. Under normal conditions, ethanol is sequentially oxidized to acetaldehyde and acetate mainly through cytosolic alcohol dehydrogenase (ADH) and mitochondrial ALDH2, respectively. However, after binge alcohol drinking or long-term chronic alcohol consumption, CYP2E1 plays a major role in ethanol metabolism. Under these conditions, CYP2E1 is induced (activated) while ALDH2 activity is suppressed. Because of reactive oxygen species (ROS) produced by CYP2E1-mediated reactions, we observe increased oxidative stress with elevated levels of toxic acetaldehyde and lipid peroxides. These conditions likely lead to mitochondrial dysfunction, ER stress and tissue injury, especially in the presence of another risk factor such as APAP (one example of many drugs), high fat, hepatitis virus and smoking. 1. Characterization of covalently-modified proteins in alcohol- or drug-exposed tissues Song, Abdelmegeed, Jang, Banerjee in collaboration with Salem, Pacher, Gonzalez, Veenstra, Yu 1-A. Protein oxidation Increased oxidative/nitrative stress is a major contributing factor in alcohol- or drug-mediated mitochondrial dysfunction, ER stress and cell/tissue damage. The majority of ROS and reactive nitrogen species (RNS) in alcohol-exposed cells/tissues is produced through direct inhibition of the mitochondrial respiratory chain, induction or activation of CYP2E1, nitric oxide synthase, NADPH-oxidase, myeloperoxidase, xanthine oxidase, etc (Purohit et al., 2009). Despite the well-established roles of ROS/RNS in alcohol-induced cellular dysfunction and injury, it is poorly understood which proteins are oxidatively-modified and whether their functions are altered in proportion to the severity of cell/tissue injury. We hypothesized that oxidative modifications (e.g., Cys oxidation to sulfenic, sulfinic, sulfonic acid, or disulfide, S-nitrosylation, S-glutathionylation, etc) of cellular proteins promote inactivation of many target proteins in the cytoplasm and mitochondria, contributing to ER stress and mitochondrial dysfunction, respectively, prior to pathological tissue injury observed at later time points.
To test this hypothesis and further characterize the oxidatively-modified proteins, we developed a simple, effective method by using biotin-N-maleimide (biotin-NM) as a specific probe to positively identify oxidized Cys residues of many proteins in alcohol-exposed mouse tissues

65
(reviewed in Song et al., 2010). Cys residues in various proteins can be oxidatively-modified under increased oxidative/nitrative states. By using the Cys-targeted redox proteomics method as illustrated in Figure 2, we can identify oxidatively-modified proteins in different sub-cellular fractions including mitochondria and study their activity changes. As shown in the right panel of Figure 2, the number/intensity of oxidized proteins detected by this method positively correlated with the levels of increased oxidative/nitrative stress, as demonstrated in the alcohol-exposed hepatoma cells and animal tissues. We have applied this targeted proteomics method to identify oxidized mitochondrial proteins in experimental models of alcoholic fatty liver (AFLD) and NAFLD. Our results showed that many mitochondrial proteins involved in mitochondrial electron transfer, energy production, β-oxidation of fatty acids, antioxidant defense, and chaperone activities were also oxidatively-modified in alcohol-exposed rat livers (summarized in Figure 3). The activities of all four oxidized proteins, shown in blue color in Figure 3, were significantly decreased in rats or mice exposed to alcohol (Moon et al., 2006). The activity of the oxidized ALDH2 proteins correlated with the presence of S-nitrosylated Cys band and the suppressed ALDH2 activity was completely restored by pre-incubation with a reducing agent DTT, suggesting a reversible oxidative-modification of Cys residues including the active site Cys. These results also implicate oxidative stress as a signaling mechanism to modify protein function and activity. Based on these results, we believe that oxidative modifications and subsequent inactivation of these enzymes and other proteins are likely to contribute to mitochondrial dysfunction, fat accumulation, and eventually tissue injury.
Figure 3. Summary of oxidatively-modified mitochondrial proteins in alcohol-exposed rat liver. Oxidized mitochondrial proteins were purified from alcohol-exposed rats, identified by mass spectral analysis, and then grouped under different functions. The enzymes in blue color were oxidized and inactivated.
Similar results were observed in rats exposed to the abused substance MDMA (ecstasy, 3,4-methylenedioxymethamphetamine) (Moon et al, 2008b). We also observed similar patterns of oxidized mitochondrial proteins in mouse models of hepatic ischemia reperfusion (I/R) injury (conducted in collaboration with Dr. Pal Pacher, LPS) (Moon et al., 2008a). Mass spectral analyses and activity measurements verified that many key mitochondrial enzymes involved in cellular defense [i.e., ALDH2 and superoxide dismutase (SOD2)], fat metabolism (i.e., mitochondrial 3-ketoacyl-CoA thiolases), and energy supply (i.e., ATP synthase) were
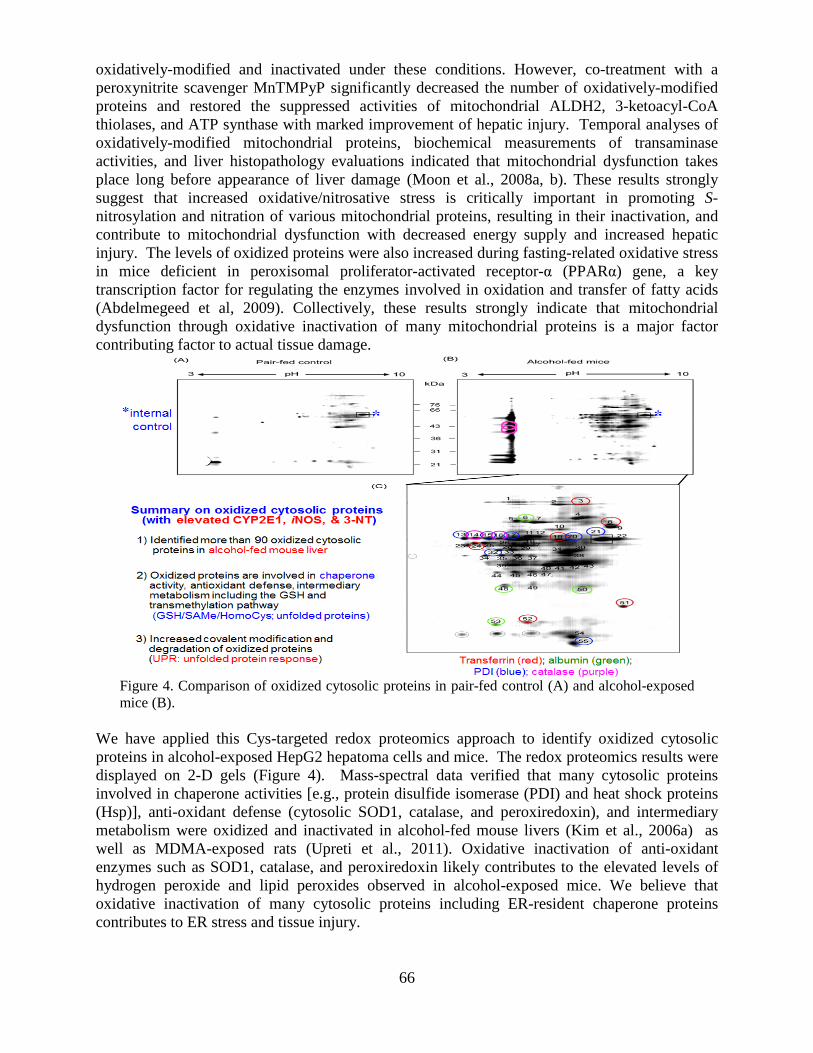
66
oxidatively-modified and inactivated under these conditions. However, co-treatment with a peroxynitrite scavenger MnTMPyP significantly decreased the number of oxidatively-modified proteins and restored the suppressed activities of mitochondrial ALDH2, 3-ketoacyl-CoA thiolases, and ATP synthase with marked improvement of hepatic injury. Temporal analyses of oxidatively-modified mitochondrial proteins, biochemical measurements of transaminase activities, and liver histopathology evaluations indicated that mitochondrial dysfunction takes place long before appearance of liver damage (Moon et al., 2008a, b). These results strongly suggest that increased oxidative/nitrosative stress is critically important in promoting S-nitrosylation and nitration of various mitochondrial proteins, resulting in their inactivation, and contribute to mitochondrial dysfunction with decreased energy supply and increased hepatic injury. The levels of oxidized proteins were also increased during fasting-related oxidative stress in mice deficient in peroxisomal proliferator-activated receptor-α (PPARα) gene, a key transcription factor for regulating the enzymes involved in oxidation and transfer of fatty acids (Abdelmegeed et al, 2009). Collectively, these results strongly indicate that mitochondrial dysfunction through oxidative inactivation of many mitochondrial proteins is a major factor contributing factor to actual tissue damage.
Figure 4. Comparison of oxidized cytosolic proteins in pair-fed control (A) and alcohol-exposed mice (B).
We have applied this Cys-targeted redox proteomics approach to identify oxidized cytosolic proteins in alcohol-exposed HepG2 hepatoma cells and mice. The redox proteomics results were displayed on 2-D gels (Figure 4). Mass-spectral data verified that many cytosolic proteins involved in chaperone activities [e.g., protein disulfide isomerase (PDI) and heat shock proteins (Hsp)], anti-oxidant defense (cytosolic SOD1, catalase, and peroxiredoxin), and intermediary metabolism were oxidized and inactivated in alcohol-fed mouse livers (Kim et al., 2006a) as well as MDMA-exposed rats (Upreti et al., 2011). Oxidative inactivation of anti-oxidant enzymes such as SOD1, catalase, and peroxiredoxin likely contributes to the elevated levels of hydrogen peroxide and lipid peroxides observed in alcohol-exposed mice. We believe that oxidative inactivation of many cytosolic proteins including ER-resident chaperone proteins contributes to ER stress and tissue injury.

67
As recently reviewed (Song et al., 2010), the Cys-targeted redox proteomics method using biotin-NM has significant advantages over other redox proteomics methods. In addition, we can predict the functional changes of the detected proteins by searching the literature for active site Cys residues in the proteins of interest. In contrast, many global proteomics methods for detecting changes in protein expression, despite their own advantages and merits, may not necessarily provide valuable information about functional changes of the target proteins since the activities of many (mitochondrial) proteins can be suppressed (or increased) without significant quantitative changes in their expressed levels. Despite our characterizations of oxidatively-modified proteins in our experimental models, we believe that the actual number of oxidized proteins could be much higher since some mitochondrial or cytosolic proteins, expressed in low quantities, might not be detected by the redox proteomics analysis because of detection limits of mass spectrometry. In addition, some oxidized proteins could be rapidly degraded through ubiquitin-dependent and -independent proteolysis as we discussed (Song et al., 2010; Abdelmegeed et al., 2010) and thus could not be detected by the current method. 1-B. Protein nitration Many laboratories reported important roles of protein nitration in promoting various types of cell/tissue injury caused by exposure to alcohol, APAP, MDMA, or lipopolysaccharide (LPS) and pathological conditions such as I/R injury and diabetes. Despite extensive reports on the critical role of nitrated proteins in acute liver injury (mostly via necrosis), it is poorly understood which cellular (including mitochondrial) proteins are nitrated and how their functions are altered to promote mitochondrial dysfunction and hepatotoxicity. We and other laboratories reported that only a few proteins such as cytosolic superoxide dismutase (Cu-Zn-SOD, SOD1), catalase, and mitochondrial SOD2 (Mn-SOD, SOD2) are nitrated in APAP-exposed animals. Further, we recently demonstrated the important role of CYP2E1 in promoting protein nitration and ubiquitin-dependent degradation of many proteins (Abdelmegeed et al., 2010).
Figure 5. Summary of nitrated mitochondrial proteins in APAP-exposed mouse liver. Nitrated mitochondrial proteins were affinity-purified from APAP-exposed mice and identified by mass spectral analysis and grouped under different functions. The enzymes in blue color were nitrated and inactivated.
Based on the numerous spots of nitrated proteins displayed on 2-D gel (Abdelmegeed et al., 2010), we hypothesized that many more proteins could be nitrated and that nitrated (mitochondrial) proteins could contribute to acute hepatotoxicity. Thus, we aimed to

68
systematically identify nitrated proteins from mice exposed to APAP (a single ip injection of 350 mg/kg) for 2 h, where liver injury and ALT levels were very low. The main reason we chose this time point was to investigate the causal relationship between protein nitration and APAP-induced mitochondrial dysfunction and liver injury. Our data showed that many cytosolic and mitochondrial proteins were rapidly nitrated at 1 or 2 h while liver damage, determined by serum ALT levels and histology, was prominent at 24 h following APAP exposure. We therefore purified nitrated proteins by using the antibody-affinity columns from APAP-exposed mouse livers only for 2 h. Mass spectral analysis of the affinity-purified proteins showed that protein nitration following APAP exposure was extensive and far greater than what other laboratories have reported. In fact, our data verified that more than 30 cytosolic and 65 mitochondrial proteins involved in anti-oxidant defense, energy supply, amino acid and fat metabolic pathways were nitrated by APAP exposure. Figure 5 summarizes some of the mitochondrial proteins that were nitrated at 2 h following APAP exposure. The enzyme activities of a few selected nitrated proteins such as ALDH2, glutathione peroxidase, SOD2, ATP synthase, and 3-ketoacyl-CoA thiolase (thiolase) were suppressed in APAP-exposed mice compared to those in vehicle-control mice. The suppressed activities of these proteins were restored by co-treatment with an antioxidant N-acetylcysteine (NAC). Immunoprecipitation followed by immunoblot with anti-3-NT antibody confirmed that these proteins were nitrated in APAP-exposed mice but not in mice co-treated with APAP and NAC, which also prevented APAP-induced liver injury. These results not only establish the causal role of protein nitration in APAP-induced liver injury but also provide a new mechanism by which many nitrated proteins promote mitochondrial dysfunction and hepatotoxicity (Abdelmegeed et al, 2012, under revision). It is possible that many other critical proteins, expressed in small amounts, could be also nitrated and their functions could be altered under nitrative stress. Our method can be applied to study the identities and functional roles of nitrated proteins in different tissues and/or various disease states such as diabetes. 1-C. Protein phosphorylation. It is well-established that mitogen-activated protein kinases (MAPKs) are activated under increased oxidative and nitrative stress. We previously reported the important role of the activated c-Jun N-terminal protein kinase (JNK) in promoting cell death by phosphorylating critical proteins including pro-apoptotic Bax (Kim et al., 2006b) and mitochondrial ALDH2 (Moon et al., 2010, Song et al., 2011). To better understand the roles of JNK and its target proteins in regulating mitochondrial function and cell/tissue damage, we also tried to identify and characterize JNK-mediated phosphorylation of many mitochondrial proteins. For specific activation of JNK without activating other MAPKs such as p38K, we chose a model of acute liver injury caused by a single dose (50 mg/kg, ip) of CCl4, another substrate of CYP2E1. JNK was rapidly activated within 30 min after CCl4 exposure and translocated to mitochondria to phosphorylate many mitochondrial proteins between 1 and 8 h where liver injury was minimal. However, these events were not observed in the corresponding Cyp2e1-null mice. To further characterize the role of protein phosphorylation, we purified phosphorylated mitochondrial proteins from vehicle-treated control mice and CCl4-exposed mice, respectively, for 2 h by using metal-affinity columns. Mass-spectral analysis of the purified phospho-proteins and differential proteomics analysis with a ratio of control versus CCl4-exposed tissues greater than 1.5 revealed that more than 100 mitochondrial proteins were phosphorylated by activated JNK. These proteins include ATP synthase, α-ketoglutarate dehydrogenase, ALDH2, etc involved in energy supply and cellular defense, respectively. The activities of these phospho-proteins were markedly suppressed in CCl4-exposed mice but significantly restored in CCl4-exposed mice pretreated with a selective JNK inhibitor, which blocked the mitochondrial translocation of activated p-JNK, JNK-mediated protein phosphorylation, mitochondrial swelling, and prevented liver
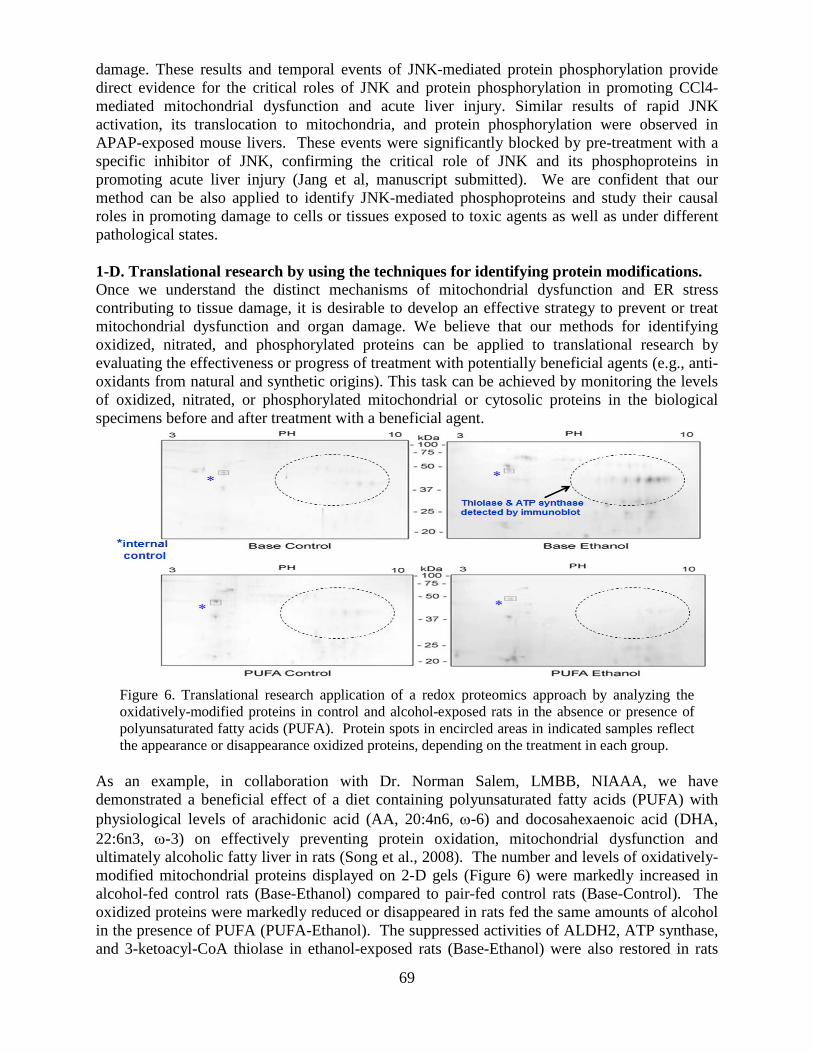
69
damage. These results and temporal events of JNK-mediated protein phosphorylation provide direct evidence for the critical roles of JNK and protein phosphorylation in promoting CCl4-mediated mitochondrial dysfunction and acute liver injury. Similar results of rapid JNK activation, its translocation to mitochondria, and protein phosphorylation were observed in APAP-exposed mouse livers. These events were significantly blocked by pre-treatment with a specific inhibitor of JNK, confirming the critical role of JNK and its phosphoproteins in promoting acute liver injury (Jang et al, manuscript submitted). We are confident that our method can be also applied to identify JNK-mediated phosphoproteins and study their causal roles in promoting damage to cells or tissues exposed to toxic agents as well as under different pathological states. 1-D. Translational research by using the techniques for identifying protein modifications. Once we understand the distinct mechanisms of mitochondrial dysfunction and ER stress contributing to tissue damage, it is desirable to develop an effective strategy to prevent or treat mitochondrial dysfunction and organ damage. We believe that our methods for identifying oxidized, nitrated, and phosphorylated proteins can be applied to translational research by evaluating the effectiveness or progress of treatment with potentially beneficial agents (e.g., anti-oxidants from natural and synthetic origins). This task can be achieved by monitoring the levels of oxidized, nitrated, or phosphorylated mitochondrial or cytosolic proteins in the biological specimens before and after treatment with a beneficial agent.
Figure 6. Translational research application of a redox proteomics approach by analyzing the oxidatively-modified proteins in control and alcohol-exposed rats in the absence or presence of polyunsaturated fatty acids (PUFA). Protein spots in encircled areas in indicated samples reflect the appearance or disappearance oxidized proteins, depending on the treatment in each group.
As an example, in collaboration with Dr. Norman Salem, LMBB, NIAAA, we have demonstrated a beneficial effect of a diet containing polyunsaturated fatty acids (PUFA) with physiological levels of arachidonic acid (AA, 20:4n6, ω-6) and docosahexaenoic acid (DHA, 22:6n3, ω-3) on effectively preventing protein oxidation, mitochondrial dysfunction and ultimately alcoholic fatty liver in rats (Song et al., 2008). The number and levels of oxidatively-modified mitochondrial proteins displayed on 2-D gels (Figure 6) were markedly increased in alcohol-fed control rats (Base-Ethanol) compared to pair-fed control rats (Base-Control). The oxidized proteins were markedly reduced or disappeared in rats fed the same amounts of alcohol in the presence of PUFA (PUFA-Ethanol). The suppressed activities of ALDH2, ATP synthase, and 3-ketoacyl-CoA thiolase in ethanol-exposed rats (Base-Ethanol) were also restored in rats

70
fed the ethanol-DHA/AA-supplemented diet (PUFA-Ethanol). These results suggest that PUFA diet containing physiologically relevant levels of DHA/AA can protect against alcohol-mediated mitochondrial dysfunction and AFLD. By the similar approaches to Cys-redox proteomics, the levels of nitrated or phosphorylated proteins can be studied in the absence and presence of many natural anti-oxidants or synthetic compounds. These newly-developed proteomics approaches may also be used to find potential biomarkers of disease states in other tissues such as brain, heart, and lung. Finally, in collaboration with Dr. Frank J. Gonzalez, NCI, NIH, we have started analyzing different mouse strains to study the mechanisms of mitochondrial dysfunction, fatty liver diseases and inflammation by monitoring different post-translational modifications (PTMs) of mitochondrial proteins. Our data showed that Ppara-null mice are far more sensitive to oxidative stress, mitochondrial dysfunction and fatty liver diseases than are wild-type mice, while Cyp2e1-null mice are quite resistant in the mouse models of AFLD and NAFLD. We recently showed that CYP2E1 is critically important in the development of high fat-induced nonalcoholic steatohepatitis (NASH) through increasing insulin resistance and various types of protein modifications (Abdelmegeed et al., 2012). These results and other results with different mouse strains have been recently reported from this laboratory (see publication list below). 2. Signaling mechanisms during cell death caused by ethanol and other toxic compounds or pathological conditions Song, Abdelmegeed, Jang, Banerjee, Henderson We have previously reported that activated JNK and/or p38K promote cell death. In our models, JNK and p38K were activated by toxic substrates of CYP2E1 such as APAP, 4-hydroxynonenal, CCl4, and long chain fatty acids as well as non-CYP2E1 substrates such as troglitazone, hydrogen peroxide, etoposide, and staurosporine. We also showed that all these compounds activate JNK and/or p38 kinase, which phosphorylate Bax, contributing to translocation of Bax to mitochondria to initiate mitochondria-dependent apoptosis (Kim et al., 2006b). Furthermore, our unpublished results showed that ethanol alone, APAP, and hepatic I/R procedure activated both JNK and p38 kinase, promoting phosphorylation of Bax prior to its translocation to mitochondria to initiate mitochondria-dependent apoptosis/necrosis. We also studied the role of JNK in promoting tissue injury through phosphorylation of other mitochondrial proteins (see above Section 1-C). Our results showed that JNK activated at 1 or 2 h after CCl4 exposure translocated to mitochondria and phosphorylated many mitochondrial proteins including ALDH2, leading to its inactivation, elevation of lipid peroxides and tissue damage (Moon et al., 2010). This represents a novel, regulatory mechanism of ALDH2. Based on this new information and other modifications reported from other laboratories including small molecule chemical activators (Figure 7), we have reviewed various types of ALDH2 regulation and their functional implications (Song et al, 2011).

71
Figure 7. Multiple types of regulation of ALDH2 and its functional implications. The negative and positive signs represent suppression and activation of ALDH2 activity, respectively.
References Abdelmegeed MA, Moon KH, Chen C, Gonzalez FJ, Song BJ. 2010. Role of cytochrome P450 2E1 in protein nitration and ubiquitin-mediated degradation during acetaminophen toxicity. Biochemical Pharmacology 79:57-66
Abdelmegeed MA, Jang SH, Banerjee A, Hardwick JP, Song BJ. 2012. Robust protein nitration contributes to acetaminophen-induced mitochondrial dysfunction and acute liver injury. Free Radical Biology and Medicine under revision.
Abdelmegeed MA, Moon KH, Hardwick JP, Gonzalez FJ, Song BJ. 2009. Role of peroxisome proliferator-activated receptor-alpha in fasting-mediated oxidative stress. Free Radical Biololgy and Medicine 47:767-78. Kim BJ, Hood BL, Aragon RA, Hardwick JP, Conrads TP, et al. 2006a. Increased oxidation and degradation of cytosolic proteins in alcohol-exposed mouse liver and hepatoma cells. Proteomics 6:1250-60
Kim BJ, Ryu SW, Song BJ. 2006b. JNK- and p38 kinase-mediated phosphorylation of Bax leads to its activation and mitochondrial translocation and to apoptosis of human hepatoma HepG2 cells. Journal of Biological Chemistry 281:21256-65
Moon KH, Hood BL, Kim BJ, Hardwick JP, Conrads TP, et al. 2006. Inactivation of oxidized and S-nitrosylated mitochondrial proteins in alcoholic fatty liver of rats. Hepatology 44:1218-30
Moon KH, Hood BL, Mukhopadhyay P, Rajesh M, Abdelmegeed MA, et al. 2008a. Oxidative inactivation of key mitochondrial proteins leads to dysfunction and injury in hepatic ischemia reperfusion. Gastroenterology 135:1344-57
Moon KH, Upreti VV, Yu LR, Lee IJ, Ye X, et al. 2008b. Mechanism of 3,4-methylenedioxymethamphetamine (MDMA, ecstasy)-mediated mitochondrial dysfunction in rat liver. Proteomics 8:3906-18

72
Moon KH, Lee YM, Song BJ. 2010. Inhibition of hepatic mitochondrial aldehyde dehydrogenase by carbon tetrachloride through JNK-mediated phosphorylation. Free Radical Biology and Medicine 48:391-8
Purohit V, Gao B, Song BJ. 2009. Molecular Mechanisms of Alcoholic Fatty Liver. Alcoholism-Clinical and Experimental Research 33:191-205
Song BJ, Abdelmegeed MA, Yoo SH, Kim BJ, Jo SA, et al. 2011. Post- translational modifications of mitochondrial aldehyde dehydrogenase and biomedical implications. Journal of Proteomics 74:2691-702.
Song BJ, Moon KH, Olsson NU, Salem N, Jr. 2008. Prevention of alcoholic fatty liver and mitochondrial dysfunction in the rat by long-chain polyunsaturated fatty acids. Journal of Hepatology 49:262-73
Song BJ, Suh SK, Moon KH. 2010. A simple method to systematically study oxidatively modified proteins in biological samples and its applications. In Methods in Enzymology, Vol 473: Thiol Redox Transitions in Cell Signaling, Pt A: Chemistry and Biochemistry of Low Molecular Weight and Protein Thiols 473:251-64
Upreti VV, Moon KH, Yu LR, Lee IJ, Eddington ND, et al. 2011. Increased oxidative-modifications of cytosolic proteins in 3,4-methylenedioxymeth amphetamine (MDMA, ecstasy)-exposed rat liver. Proteomics 11:202-11.
Publications 2011-2012 1. Song BJ, Abdelmegeed MA, Yoo SH, Kim BJ, Jo SA, et al. 2011. Post-
translational modifications of mitochondrial aldehyde dehydrogenase and biomedical implications. Journal of Proteomics 74:2691-702.
2. Upreti VV, Moon KH, Yu LR, Lee IJ, Eddington ND, et al. 2011. Increased oxidative-modifications of cytosolic proteins in 3,4-methylenedioxymeth amphetamine (MDMA, ecstasy)-exposed rat liver. Proteomics 11:202-11.
3. Yoo SH, Park O, Henderson LE, Abdelmegeed MA, Moon KH, Song BJ. 2011. Lack of
PPARα exacerbates lipopolysaccharide-induced liver toxicity through STAT1 inflammatory signaling and increased oxidative/nitrosative stress. Toxicology Letter 202, 23-9.
4. Abdelmegeed MA, Yoo SH, Henderson LE, Gonzalez FJ, Woodcroft KJ, Song BJ. 2011.
PPAR protects male mice from high fat-induced non-alcoholic fatty liver. Journal of Nutrition 141:603-10.
5. Abdelmegeed MA, Banerjee A, Yoo SH, Jang SH, Gonzalez FJ, Song BJ. 2012. Critical role
of cytochrome P450 2E1 (CYP2E1) in the development of high-fat induced non-alcoholic steatohepatitits. Journal of Hepatology 57:860-6.

73
6. Byun K, Bayarsaikhan E, Kim D, Kim CY, Mook-Jung I, et al. 2012. Induction of neuronal death by microglial AGE-albumin: implications for Alzheimer’s disease. PLoS One 7:e37917.
7. Purohit V, Rapaka R, Kwon OS, Song BJ. 2012. Role of alcohol and smoking in the
development of hepatocellular carcinoma. Life Sciences [Epub 2012 Oct 30].
8. Hardwick JP, Eckman K, Lee YK, Abdelmegeed MA, Esterle A, et al. 2012. Eicosanoids in metabolic syndrome. Advances in Pharmacology in press.
9. Abdelmegeed MA, Jang SH, Banerjee A, Hardwick JP, Song BJ. 2012. Robust protein nitration contributes to acetaminophen-induced mitochondrial dysfunction and acute liver injury. Free Radical Biology and Medicine under revision.
Collaborators Norman Salem, Jr. PhD, LMBB, NIAAA, Bethesda, MD Joe Hibbeln, MD, LMBB, NIAAA, Bethesda, MD Pal Pacher, MD, LPS, NIAAA, Bethesda, MD Frank G. Gonzalez, PhD, LM, NCI, Bethesda, MD James P. Hardwick, PhD, Northeastern Ohio University College of Medicine, OH Timothy D. Veenstra, PhD, LAC, SAIC-Frederick, NCI-Frederick, MD Li-Rong Yu, PhD, National Center for Toxicology Research, FDA, Jefferson, AR Ali Keshavarzian, MD, Rush University Medical Center, Chicago, IL Dr. Song: [email protected]

74
______________________________________________________________________
Section on Nutritional Neuroscience ______________________________________________________________________ CAPT Joseph R. Hibbeln, MD, Acting Chief LCDR Christopher Ramsden, MD Yu Hong Lin, PhD, Staff Scientist Jim Loewke, Chemist Sharon Majchrzak, MS, Chemist Rachel Gow, Guest Researcher Katherine Ness, Postbac IRTA Amit Ringel, Postbac IRTA William E. Lands, PhD, Special Volunteer Norman Salem, Jr., PhD, Special Volunteer
Overview The central focus of the Section on Nutritional Neuroscience is to examine the impact of deficiencies in omega-3 highly unsaturated fatty acids (n-3 HUFAs) on adverse neurodevelopment and psychiatric outcomes. Alcohol and illicit substance use are linked to n-3 HUFA status because 1) deficiencies in pregnancy and early childhood appear to increase risk for childhood disruptive behaviors, and an adverse developmental trajectory 2) substance misuse causes deficiencies in tissue n-3 HUFAs both directly by increasing degradation and indirectly via decreased dietary intakes and 3) resulting deficiencies of n-3 HUFAs may accelerate addictive processes via dopamine depletion, endocannabinoid excesses and increased risks for depressive and aggressive phenotypes. This section translates basic biochemical and biophysical findings regarding these nutrients into clinical applications. The portfolio of projects include genetic epidemiology, large-scale longitudinal cohort studies, studies of basic metabolism, and randomized placebo-controlled clinical intervention trials among populations potentially affected by insufficient intakes. Active clinical intervention protocols increasing n-3 HUFAs intake include US Veterans at risk for suicide attempts and three pediatric populations with disruptive behaviors. Clinical intervention protocols that also reduce n-6 polyunsaturated fatty acids (PUFAs) include patients suffering from chronic daily headaches, overweight women and a military age population consuming US military diets. The assessment of the safety of increasing consumption of seafood, rich in n-3 HUFAs, and decreasing consumption of seed oils rich in n-6 PUFAs has yielded findings that have changed dietary advice for pregnancy women and has the potential to change current worldwide dietary advice to consume PUFA to specificity n-3 or n-6 fatty acids. Projects cover 7 topic areas 1. n-3 HUFA deficiencies in early neurodevelopment. 2. n-3 HUFA deficiencies in alcohol and substance misuse. 3. n-3 HUFA deficiencies in depressive disorders 4. Excessive n-6 PUFA intakes as a cause of n-3 HUFA tissue deficiencies. 5. Excessive n-6 PUFA intake increased risk of cardiovascular disorder death.

75
6. Excessive n-6 PUFA intakes as a cause of endocannabinoid hyperactivity. 7. Reduction of suicide risk among US Military: diets to eliminate n-3 HUFA deficiencies. 1. n-3 HUFA deficiency in early neurodevelopment: contributions to international policies The 2011 USDA Dietary Guidelines for fish and seafood consumption in pregnancy were substantially modified based upon our 2007 paper in the Lancet 1. The new recommendations to promote seafood consumption during pregnancy are directly derived from our finding that the benefits of seafood consumption outweighed the risks from trace levels of heavy metals. The Food and Drug Administration (FDA) has issued (19 Jan 2009) a draft revision of scientific background to update the 2004 FDA/ the Environmental Protection Agency (EPA) advice to pregnant and fertile women regarding seafood consumption 2. We participated in the drafting of this document and our data from the ALSPAC study is central to the new risk benefit modeling. We anticipate a forthcoming revision of the FDA/EPA 2004 advisory due to these contributions. In 2008, a Joint UK Parliamentary Inquiry of the House of Lords and House of Commons reviewed the data linking n-3 deficiencies to aggression, depression behavioral disorders and suboptimal development during pregnancy 3. They recommended that on a precautionary basis, the Food Standards Agency should reconsider its advice to pregnant women about fish consumption, with a view to encouraging them to eat two portions of oily fish, or the equivalent in omega-3 PUFAs, a week (rather than that people should eat two portions of fish a week, of which one should be oily). They also recommended that the Scientific Advisory Committee on Nutrition (SACN) should be asked to define further the optimum intake of omega-3 polyunsaturated fatty acids (n-3 PUFAs) in different stages of life, especially for pregnant women and children. The studies of the Section of Nutritional Neurosciences were cited as key data in the formulation of these recommendations. 1.1 Risk of harm from seafood in methyl-mercury vs. nutritional benefits of seafood We are extending this work by beginning to conduct tissue based risk benefit analyses. We have achieved quantification of fatty acid compositions in a large number of subjects (n=6,500 maternal RBCs, n=4,090 umbilical cord blood samples, n=5,500 serum of the children at 7 years old). Our collaborator in the CDC, Robert Jones, PhD, has completed quantification of trace metals (Hg, Se, Fe and Pb) in n= 5,000 samples from early pregnancy. In collaboration with the FDA, NOAA, CDC and the University of Bristol, we are developing quantitative risk benefit models for multiple neurodevelopmental outcomes including IQ and adverse behavioral problems. These models will be able to further inform public policy advice for fish consumption. 1.2 Dietary predictors of maternal prenatal blood mercury levels: ALSPAC study Background: Very high levels of prenatal maternal mercury have adverse effects on the developing brain of the fetus. There is concern as to whether all possible sources of mercury should be avoided. However, although seafood is known to be contaminated with mercury, little is known as to what other dietary components are sources of mercury or the contribution of diet to the overall levels of blood mercury. Here we sought to quantify the contribution of components of the mother’s diet to her prenatal blood mercury level. Methods: Blood was collected from women (n=4484) in the Avon Longitudinal Study of Parents and Children (ALSPAC). Information on diet and sociodemographic factors were collected in pregnancy using self-completion questionnaires. The whole blood was assayed for total mercury (Hg) using ICP-DRC-MS. Contributions of dietary and sociodemographic measures to the mercury level used
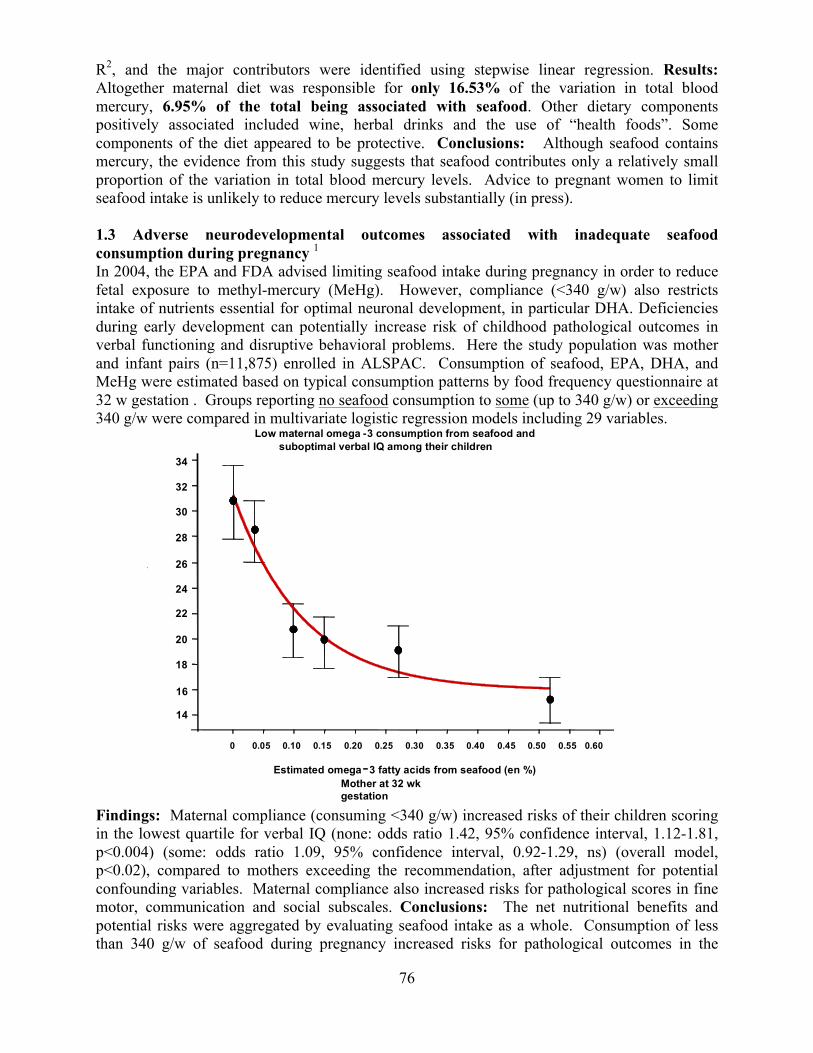
76
R2, and the major contributors were identified using stepwise linear regression. Results: Altogether maternal diet was responsible for only 16.53% of the variation in total blood mercury, 6.95% of the total being associated with seafood. Other dietary components positively associated included wine, herbal drinks and the use of “health foods”. Some components of the diet appeared to be protective. Conclusions: Although seafood contains mercury, the evidence from this study suggests that seafood contributes only a relatively small proportion of the variation in total blood mercury levels. Advice to pregnant women to limit seafood intake is unlikely to reduce mercury levels substantially (in press). 1.3 Adverse neurodevelopmental outcomes associated with inadequate seafood consumption during pregnancy 1 In 2004, the EPA and FDA advised limiting seafood intake during pregnancy in order to reduce fetal exposure to methyl-mercury (MeHg). However, compliance (<340 g/w) also restricts intake of nutrients essential for optimal neuronal development, in particular DHA. Deficiencies during early development can potentially increase risk of childhood pathological outcomes in verbal functioning and disruptive behavioral problems. Here the study population was mother and infant pairs (n=11,875) enrolled in ALSPAC. Consumption of seafood, EPA, DHA, and MeHg were estimated based on typical consumption patterns by food frequency questionnaire at 32 w gestation . Groups reporting no seafood consumption to some (up to 340 g/w) or exceeding 340 g/w were compared in multivariate logistic regression models including 29 variables.
III
Low maternal omega - 3 consumption from seafood and suboptimal verbal IQ among their children
Estimated omega - 3 fatty acids from seafood (en %) Mother at 32 wk gestation
Findings: Maternal compliance (consuming <340 g/w) increased risks of their children scoring in the lowest quartile for verbal IQ (none: odds ratio 1.42, 95% confidence interval, 1.12-1.81, p<0.004) (some: odds ratio 1.09, 95% confidence interval, 0.92-1.29, ns) (overall model, p<0.02), compared to mothers exceeding the recommendation, after adjustment for potential confounding variables. Maternal compliance also increased risks for pathological scores in fine motor, communication and social subscales. Conclusions: The net nutritional benefits and potential risks were aggregated by evaluating seafood intake as a whole. Consumption of less than 340 g/w of seafood during pregnancy increased risks for pathological outcomes in the
0 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40 0.45 0.50 0.55 0.60
34 32 30 28
26 24 22 20 18
16 14
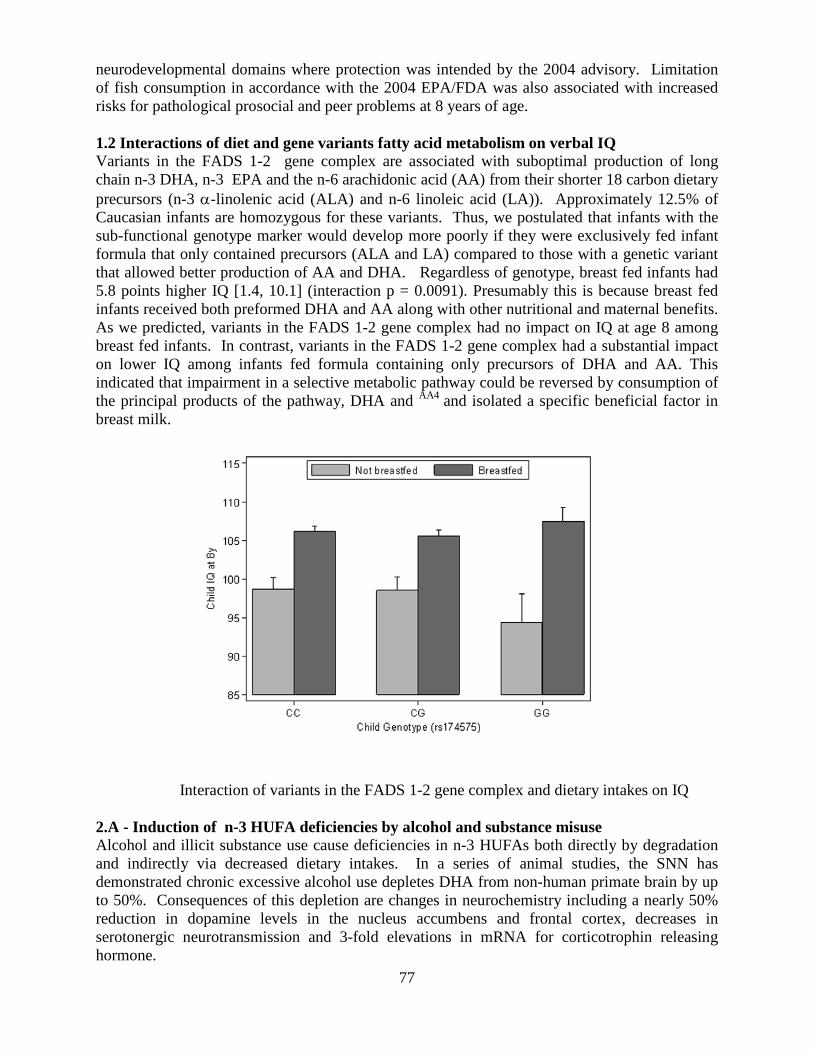
77
neurodevelopmental domains where protection was intended by the 2004 advisory. Limitation of fish consumption in accordance with the 2004 EPA/FDA was also associated with increased risks for pathological prosocial and peer problems at 8 years of age. 1.2 Interactions of diet and gene variants fatty acid metabolism on verbal IQ Variants in the FADS 1-2 gene complex are associated with suboptimal production of long chain n-3 DHA, n-3 EPA and the n-6 arachidonic acid (AA) from their shorter 18 carbon dietary precursors (n-3 α-linolenic acid (ALA) and n-6 linoleic acid (LA)). Approximately 12.5% of Caucasian infants are homozygous for these variants. Thus, we postulated that infants with the sub-functional genotype marker would develop more poorly if they were exclusively fed infant formula that only contained precursors (ALA and LA) compared to those with a genetic variant that allowed better production of AA and DHA. Regardless of genotype, breast fed infants had 5.8 points higher IQ [1.4, 10.1] (interaction p = 0.0091). Presumably this is because breast fed infants received both preformed DHA and AA along with other nutritional and maternal benefits. As we predicted, variants in the FADS 1-2 gene complex had no impact on IQ at age 8 among breast fed infants. In contrast, variants in the FADS 1-2 gene complex had a substantial impact on lower IQ among infants fed formula containing only precursors of DHA and AA. This indicated that impairment in a selective metabolic pathway could be reversed by consumption of the principal products of the pathway, DHA and AA4 and isolated a specific beneficial factor in breast milk.
Interaction of variants in the FADS 1-2 gene complex and dietary intakes on IQ 2.A - Induction of n-3 HUFA deficiencies by alcohol and substance misuse Alcohol and illicit substance use cause deficiencies in n-3 HUFAs both directly by degradation and indirectly via decreased dietary intakes. In a series of animal studies, the SNN has demonstrated chronic excessive alcohol use depletes DHA from non-human primate brain by up to 50%. Consequences of this depletion are changes in neurochemistry including a nearly 50% reduction in dopamine levels in the nucleus accumbens and frontal cortex, decreases in serotonergic neurotransmission and 3-fold elevations in mRNA for corticotrophin releasing hormone.
78
2.1 Essential fatty acid metabolism in alcoholism The first in vivo studies of the essential fatty acid (EFA) metabolism in alcoholics and smokers have been performed within the Section of Nutritional Neuroscience. In addition, our research with infants and adults report some of the first basic studies of normative physiology. Highly sensitive and selective methodology has been developed within our research group allowing for the safe and non-invasive assessment of EFA metabolism in vivo. This approach takes advantage of the stable isotope labeled EFAs, deuterated linoleic and linolenic acids, along with negative chemical ionization gas chromatography /mass spectrometry to simultaneously measure the precursor and product fatty acids in the blood stream after metabolism and export in the liver. This methodology has advanced with new capabilities to simultaneously measure different components of the EFA pathway and to model the resulting data so as to obtain rate related parameters. Thus, arguably the worldwide lead in the metabolomics of essential fatty acids has been gained.
Docosahexaenoic Acid (C22:6n-3, DHA)
C
O
OH
Our metabolic studies were among the first clinical studies performed in human adults and showed that a diet low in n-3 fats leads to an increase in formation and transport of long chain polyunsaturates to the plasma compartment5. Smokers also have somewhat increased activity in this regard. Smoking alcoholics have a further marked increase in the deuterium enrichment of plasma DHA when deuterated α-linolenate is orally ingested. Both smoking and alcohol are peroxidative challenges that are hypothesized to lead to increased EFA degradation. The apparently greater level of formation/transport of the fatty acids may, in part, compensate for these losses. When the intensity of the alcohol challenge is too great with respect to its frequency and dosage, metabolism cannot keep pace with the increased catabolism and tissue levels fall, consistent with our findings in many species. This fall in long chain polyunsaturate levels then has adverse consequences for organ function and may lead, in particular, to liver and brain pathology. Dietary impact of these fats is variable in the alcoholic population and may be protective for alcohol-induced organ pathology; this may help to explain why some alcoholics develop liver disease or organic brain syndrome and others do not. 2.2 Docosahexaenoic acid metabolism assessed by Positron Emission Tomography Docosahexaenoic acid (DHA, 22:6n-3) is a critical constituent of brain, but its metabolism has not been measured in the human brain in vivo. In monkeys, using positron emission tomography (PET), we first showed that intravenously injected [1- 11C]DHA mostly entered non-brain organs, with approximately 0.5% entering brain. Then, using PET and intravenous [1-11C]DHA in 14 healthy adult humans, we quantitatively imaged regional rates of incorporation (K*) of DHA. We also imaged cerebral blood flow (rCBF) using PET and intravenous [15O]water. Values of K* for DHA were higher in gray than white matter regions and correlated significantly with values of rCBF in 12 of 14 subjects despite evidence that rCBF does not directly influence K*. For the entire human brain, the net DHA incorporation rate Jin, the product of K* and the unesterified plasma DHA concentration, equaled 3.8 ± 1.7 mg/day. This net rate is equivalent to the net rate of DHA consumption by brain and, considering the reported amount of DHA in

79
brain, indicates that the half-life of DHA in the human brain approximates two and a half years. Thus, PET with [1-11C] DHA can be used to quantify regional and global human brain DHA metabolism in relation to health and disease 6. This methodology has been applied to compare chronic alcoholics to healthy controls to determine if brain uptake of DHA is increased, decreased or unchanged. 2.B - Consequences of n-3 HUFA deficiencies for alcohol and substance misuse 2.3 Low n-3 status and early developmental phenotypes of alcohol misuse In collaboration with Marc Schukit, M.D., Ph.D. we prospectively characterized alcoholic phenotypes, especially responses to first exposures to alcohol among nearly n=8,000 children in ALSPAC. A low level of response (LR), or low sensitivity, to alcohol as established by alcohol challenges has been shown to predict future heavier drinking, alcohol-related problems and alcohol use disorders. Previously only one study evaluated the predictive validity of a second measure of LR as determined by the Self-Report of the Effects of Alcohol (SRE) Questionnaire. We found that the SRE scores appear to have value in predicting future heavier drinking and alcohol problems in 12-year olds that go beyond the information offered by the earlier drinking pattern alone. We will evaluate if deficient dietary intakes or low serum levels of n-3 HUFAs at age 7 y or from umbilical cord blood, increase risk of the LR phenotype or predict future heavy drinking 7. 2.4 Low plasma levels of DHA are associated with increased relapse vulnerability. We posited that low levels of n-3 HUFAs would influence behaviors leading to the abuse of substances through their actions on serotonergic and dopaminergic and other mechanisms. Substance abusers tend to have poor dietary habits, deficient intake of n-3 PUFAs, which are available from dietary sources only, and subsequent low n-3 plasma levels. We explored whether n-3 plasma levels would predict relapse rates. Thirty-five patients admitted to substance abuse clinics were enrolled and followed for one year. Six patients dropped out shortly after study entry, 11 relapsed in the course of the study and dropped out, 7 relapsed but completed the study, and 11 did not relapse and completed the study. Non-relapsers were found to have significantly higher levels of DHA calculated as μg/ml and % total fatty acids, when compared to relapsers (p = .031 and p = .010, respectively) and to relapsers and non-completers combined (p = .014 and p = .009, respectively). These pilot data suggest the existence of a relationship between low levels of DHA and relapse vulnerability in some individuals who abuse substances 8. 2.5 Randomized placebo controlled trial of n-3 HUFAs to reduce relapse These observational studies were supported by a randomized placebo controlled trial of 2 gm EPA+ DHA among (n=96) alcoholics in recovery. Among those with documented compliance, the number of days of drinking were substantially fewer (3.2d/90d) compared to those with unchanged EPA + DHA in cerebrospinal fluid (17.5d/90d) 9.
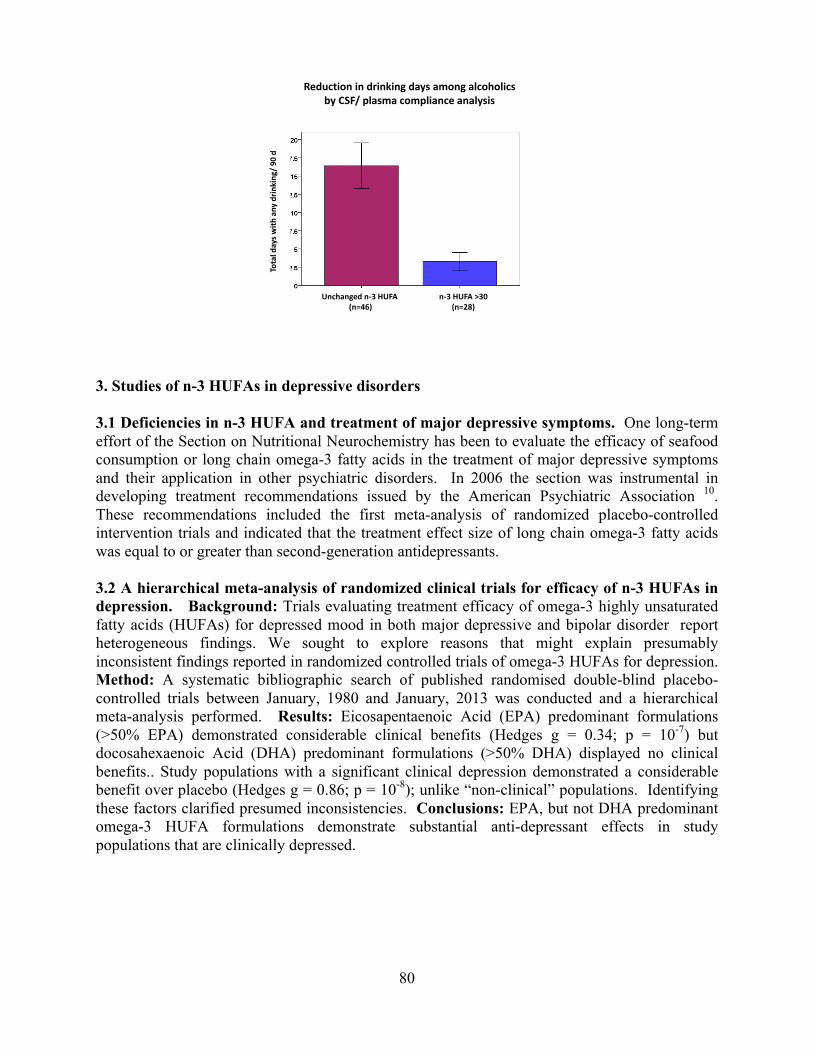
80
3. Studies of n-3 HUFAs in depressive disorders 3.1 Deficiencies in n-3 HUFA and treatment of major depressive symptoms. One long-term effort of the Section on Nutritional Neurochemistry has been to evaluate the efficacy of seafood consumption or long chain omega-3 fatty acids in the treatment of major depressive symptoms and their application in other psychiatric disorders. In 2006 the section was instrumental in developing treatment recommendations issued by the American Psychiatric Association 10. These recommendations included the first meta-analysis of randomized placebo-controlled intervention trials and indicated that the treatment effect size of long chain omega-3 fatty acids was equal to or greater than second-generation antidepressants. 3.2 A hierarchical meta-analysis of randomized clinical trials for efficacy of n-3 HUFAs in depression. Background: Trials evaluating treatment efficacy of omega-3 highly unsaturated fatty acids (HUFAs) for depressed mood in both major depressive and bipolar disorder report heterogeneous findings. We sought to explore reasons that might explain presumably inconsistent findings reported in randomized controlled trials of omega-3 HUFAs for depression. Method: A systematic bibliographic search of published randomised double-blind placebo-controlled trials between January, 1980 and January, 2013 was conducted and a hierarchical meta-analysis performed. Results: Eicosapentaenoic Acid (EPA) predominant formulations (>50% EPA) demonstrated considerable clinical benefits (Hedges g = 0.34; p = 10-7) but docosahexaenoic Acid (DHA) predominant formulations (>50% DHA) displayed no clinical benefits.. Study populations with a significant clinical depression demonstrated a considerable benefit over placebo (Hedges g = 0.86; p = 10-8); unlike “non-clinical” populations. Identifying these factors clarified presumed inconsistencies. Conclusions: EPA, but not DHA predominant omega-3 HUFA formulations demonstrate substantial anti-depressant effects in study populations that are clinically depressed.
Reduction in drinking days among alcoholics by CSF/ plasma compliance analysis
Total days w
ith any drin
king
/ 90
d
Unchanged n-‐3 HUFA (n=46)
n-‐3 HUFA >30(n=28)
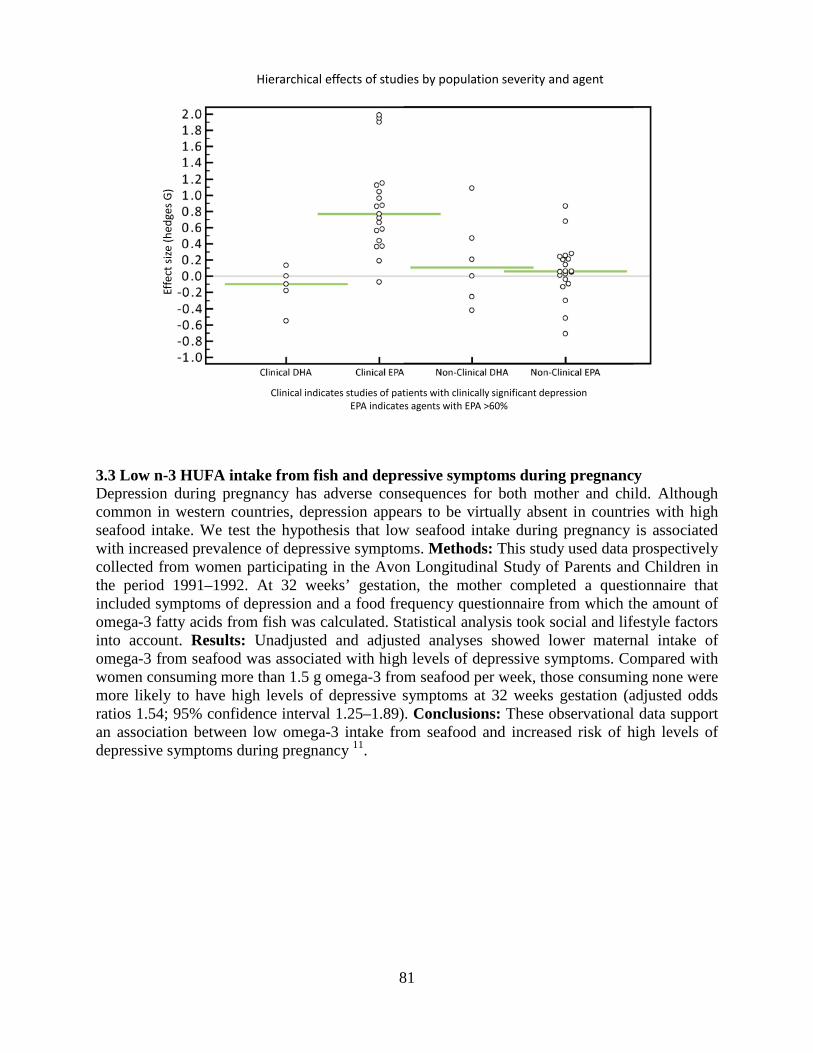
81
Effe
ct si
ze (h
edge
s G)
Hierarchical effects of studies by population severity and agent
Clinical indicates studies of patients with clinically significant depressionEPA indicates agents with EPA >60%
3.3 Low n-3 HUFA intake from fish and depressive symptoms during pregnancy Depression during pregnancy has adverse consequences for both mother and child. Although common in western countries, depression appears to be virtually absent in countries with high seafood intake. We test the hypothesis that low seafood intake during pregnancy is associated with increased prevalence of depressive symptoms. Methods: This study used data prospectively collected from women participating in the Avon Longitudinal Study of Parents and Children in the period 1991–1992. At 32 weeks’ gestation, the mother completed a questionnaire that included symptoms of depression and a food frequency questionnaire from which the amount of omega-3 fatty acids from fish was calculated. Statistical analysis took social and lifestyle factors into account. Results: Unadjusted and adjusted analyses showed lower maternal intake of omega-3 from seafood was associated with high levels of depressive symptoms. Compared with women consuming more than 1.5 g omega-3 from seafood per week, those consuming none were more likely to have high levels of depressive symptoms at 32 weeks gestation (adjusted odds ratios 1.54; 95% confidence interval 1.25–1.89). Conclusions: These observational data support an association between low omega-3 intake from seafood and increased risk of high levels of depressive symptoms during pregnancy 11.
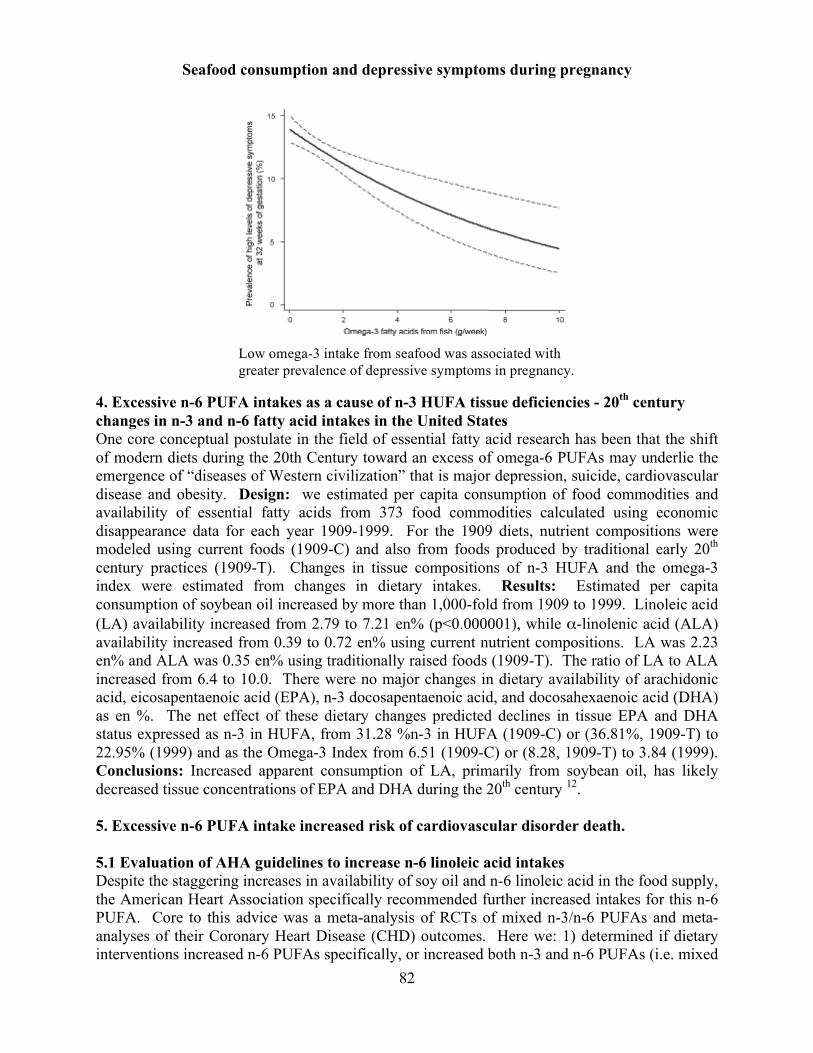
82
Seafood consumption and depressive symptoms during pregnancy
Low omega-3 intake from seafood was associated with greater prevalence of depressive symptoms in pregnancy.
4. Excessive n-6 PUFA intakes as a cause of n-3 HUFA tissue deficiencies - 20th century changes in n-3 and n-6 fatty acid intakes in the United States One core conceptual postulate in the field of essential fatty acid research has been that the shift of modern diets during the 20th Century toward an excess of omega-6 PUFAs may underlie the emergence of “diseases of Western civilization” that is major depression, suicide, cardiovascular disease and obesity. Design: we estimated per capita consumption of food commodities and availability of essential fatty acids from 373 food commodities calculated using economic disappearance data for each year 1909-1999. For the 1909 diets, nutrient compositions were modeled using current foods (1909-C) and also from foods produced by traditional early 20th century practices (1909-T). Changes in tissue compositions of n-3 HUFA and the omega-3 index were estimated from changes in dietary intakes. Results: Estimated per capita consumption of soybean oil increased by more than 1,000-fold from 1909 to 1999. Linoleic acid (LA) availability increased from 2.79 to 7.21 en% (p<0.000001), while α-linolenic acid (ALA) availability increased from 0.39 to 0.72 en% using current nutrient compositions. LA was 2.23 en% and ALA was 0.35 en% using traditionally raised foods (1909-T). The ratio of LA to ALA increased from 6.4 to 10.0. There were no major changes in dietary availability of arachidonic acid, eicosapentaenoic acid (EPA), n-3 docosapentaenoic acid, and docosahexaenoic acid (DHA) as en %. The net effect of these dietary changes predicted declines in tissue EPA and DHA status expressed as n-3 in HUFA, from 31.28 %n-3 in HUFA (1909-C) or (36.81%, 1909-T) to 22.95% (1999) and as the Omega-3 Index from 6.51 (1909-C) or (8.28, 1909-T) to 3.84 (1999). Conclusions: Increased apparent consumption of LA, primarily from soybean oil, has likely decreased tissue concentrations of EPA and DHA during the 20th century 12. 5. Excessive n-6 PUFA intake increased risk of cardiovascular disorder death. 5.1 Evaluation of AHA guidelines to increase n-6 linoleic acid intakes Despite the staggering increases in availability of soy oil and n-6 linoleic acid in the food supply, the American Heart Association specifically recommended further increased intakes for this n-6 PUFA. Core to this advice was a meta-analysis of RCTs of mixed n-3/n-6 PUFAs and meta-analyses of their Coronary Heart Disease (CHD) outcomes. Here we: 1) determined if dietary interventions increased n-6 PUFAs specifically, or increased both n-3 and n-6 PUFAs (i.e. mixed
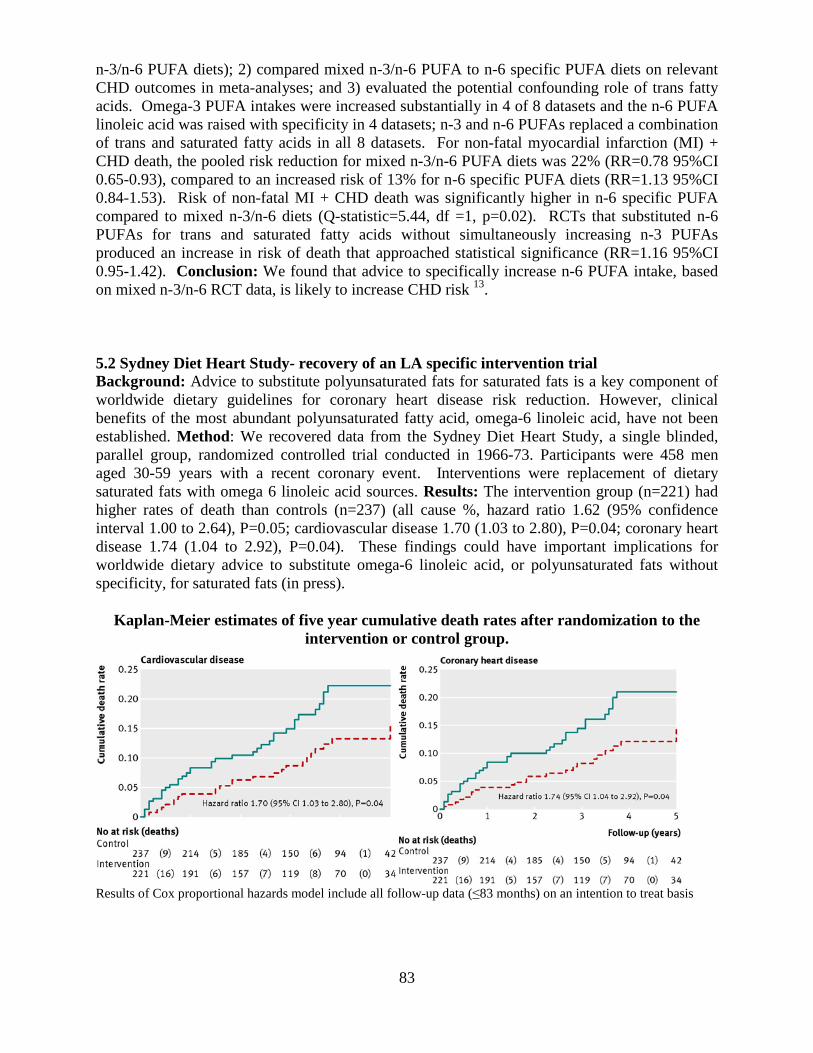
83
n-3/n-6 PUFA diets); 2) compared mixed n-3/n-6 PUFA to n-6 specific PUFA diets on relevant CHD outcomes in meta-analyses; and 3) evaluated the potential confounding role of trans fatty acids. Omega-3 PUFA intakes were increased substantially in 4 of 8 datasets and the n-6 PUFA linoleic acid was raised with specificity in 4 datasets; n-3 and n-6 PUFAs replaced a combination of trans and saturated fatty acids in all 8 datasets. For non-fatal myocardial infarction (MI) + CHD death, the pooled risk reduction for mixed n-3/n-6 PUFA diets was 22% (RR=0.78 95%CI 0.65-0.93), compared to an increased risk of 13% for n-6 specific PUFA diets (RR=1.13 95%CI 0.84-1.53). Risk of non-fatal MI + CHD death was significantly higher in n-6 specific PUFA compared to mixed n-3/n-6 diets (Q-statistic=5.44, df =1, p=0.02). RCTs that substituted n-6 PUFAs for trans and saturated fatty acids without simultaneously increasing n-3 PUFAs produced an increase in risk of death that approached statistical significance (RR=1.16 95%CI 0.95-1.42). Conclusion: We found that advice to specifically increase n-6 PUFA intake, based on mixed n-3/n-6 RCT data, is likely to increase CHD risk 13. 5.2 Sydney Diet Heart Study- recovery of an LA specific intervention trial Background: Advice to substitute polyunsaturated fats for saturated fats is a key component of worldwide dietary guidelines for coronary heart disease risk reduction. However, clinical benefits of the most abundant polyunsaturated fatty acid, omega-6 linoleic acid, have not been established. Method: We recovered data from the Sydney Diet Heart Study, a single blinded, parallel group, randomized controlled trial conducted in 1966-73. Participants were 458 men aged 30-59 years with a recent coronary event. Interventions were replacement of dietary saturated fats with omega 6 linoleic acid sources. Results: The intervention group (n=221) had higher rates of death than controls (n=237) (all cause %, hazard ratio 1.62 (95% confidence interval 1.00 to 2.64), P=0.05; cardiovascular disease 1.70 (1.03 to 2.80), P=0.04; coronary heart disease 1.74 (1.04 to 2.92), P=0.04). These findings could have important implications for worldwide dietary advice to substitute omega-6 linoleic acid, or polyunsaturated fats without specificity, for saturated fats (in press).
Kaplan-Meier estimates of five year cumulative death rates after randomization to the intervention or control group.
Results of Cox proportional hazards model include all follow-up data (≤83 months) on an intention to treat basis

84
6. Excessive n-6 PUFA intakes as a cause of endocannabinoid hyperactivity Suppression of hyperactive endocannabinoid tone is a critical target for reducing obesity. The backbone of both 2-arachidonoylglycerol (2-AG) and anandamide (AEA) is arachidonic acid (AA), an omega-6 essential fat available only from diet. Here we posited that excessive dietary intake of linoleic acid (LA), a precursor of AA, would induce endocannabinoid hyperactivity and promote obesity. Results: Increasing LA from 1% of energy (en%) to 8 en% elevated AA-phospholipids in liver and erythrocytes, nearly doubled hepatic 2-AG causing increased food intake, feed efficiency and adiposity in mice. Reducing AA-phospholipids by adding long chain omega-3 fats to 8 en% LA diets resulted in metabolic patterns resembling 1 en% LA diets. Selectively reducing LA to 1 en% reversed the obesogenic properties of a 60 en% fat diet. This finding is especially significant as 60% fat diets, containing 8 en% LA, are the diets most commonly used to induce obesity in experimental animals. The animal diets we used to induce obesity here modeled 20th century increases of human LA consumption, changes that closely correlate with increasing prevalence rates of obesity. Conclusion: Reducing excessive AA-phospholipids in humans may reduce endocannabinoid hyperactivity and obesity 14. Implications for substance use disorders. Food is the most commonly used and abused substance in the United States. These findings raise the prospect that satiety for addictive substances, in addition to food, could be up or down regulated by selecting specific dietary intakes of n-3 and n-6 fatty acids. These form the precursor pool of the n-6 derived endocannabinoids 2-AG and AEA which in turn may regulate satiety for rewarding substances.
Dietary fatty acids as precursors of endocannabinoids Shift in omega-3 HUFAs in the US diet over the last 100 years.
AA (20:4 n-‐6)EPA (20:5 n-‐3)DHA (22:6 n-‐3)
25 M years ago 20th Century
omega-‐3 rich diets omega-‐6 rich dietsseed oils
LA (18:2 n-‐6)ALA (18:3 n-‐3)Δ6 desatΔ5 desatΔ6 desatFADS 1-‐2
Flux
PLC PLA2 NAT
% n-‐3 in HUFA rich membranes % n-‐6 in HUFA rich membranes
AA-‐PL
EPA-‐PL
DHA-‐PL
EPA-‐PL
DHA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
DHA-‐PL
EPA-‐PL
DHA-‐PL
EPA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
EPA-‐PL
DHA -‐PL
EPA-‐PL
DHA-‐PL
EPA-‐PL
AA-‐PL
EPA-‐PL
EPA-‐PL
DHA -‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
DHA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
AA-‐PL
2-‐AG Anandamide (AEA)
AAOHOH
H
HONAA
Impaired satiety ↑ obesity↑ fat storage ↑ depression?
CB-‐1CB-‐2
O
EPAOH
OH
n-3 endocannabinoids
↑ Synaptic proliferation and plasticity↑ TBI resilience?
?H
HONDHA
O

85
6. The US Military diets as a model system to eliminate n-3 HUFA deficiencies The US military may provide an important model system to evaluate the health and economic benefits of lowering n-6 PUFA and raising n-3 PUFA. In order to evaluate the relevance of existing data to military objectives of reducing prevalence rates of mental illnesses, enhancing wound and surgical recoveries and increasing wellness, The Section of Nutritional Neurosciences and the Samueli Institute co-organized the following conference at NIAAA: Nutritional Armor for the Warfighter: Can Omega-3 Fatty Acids Enhance Stress Resilience, Wellness and Military Performance? October 13-14, 2009. Two expert panels recommended that sufficient data exist to increase the n-3 content of the US Military diets and that an mDRI for the intakes of EPA and DHA should be established. This conference was followed by an Institute of Medicine conference evaluating strategies to prevent and treat traumatic brain injury with a substantial focus on n-3 HUFAs. 6.1 US Military suicides and inadequate long chain omega-3 fatty acid status We sought to determine if deficiencies of neuroactive n-3 HUFA, in particular DHA, were associated with increased risk of suicide death among a large random sample of active duty US military. Methods: Serum fatty acids were quantified as % of total fatty acids, among US military suicide deaths (n= 800) and controls (n=800) matched for age, date of collection, sex, rank and year of incident. Participants were Active Duty US Military personnel (2002-2008). Outcome measures included death by suicide, post deployment health assessment questionnaire and ICD-9 mental health diagnosis data. Results: Risk of suicide death was 14% higher, per standard deviation [SD] lower DHA % (OR =1.14, 95% CI; 1.02-1.27, p<0.03), in adjusted logistic regressions. Among men risk of suicide death was 62% greater with low serum DHA status (adjusted Odds Ratio [OR] =1.62, 95% CI 1.12-2.34, p<0.01, comparing DHA below 1.75% [n=1,389] to above [n=141]). Risk of suicide death was 54% greater in those who reported having seen wounded, dead or killed coalition personnel (OR = 1.54, 95% CI; 1.12-2.12, p< 0.007.) Conclusion: This US military population had a very low and narrow range of n-3 LC PUFA status. Although these data suggest that low serum DHA may be a risk factor for suicides, well designed intervention trials are needed to evaluate causality.
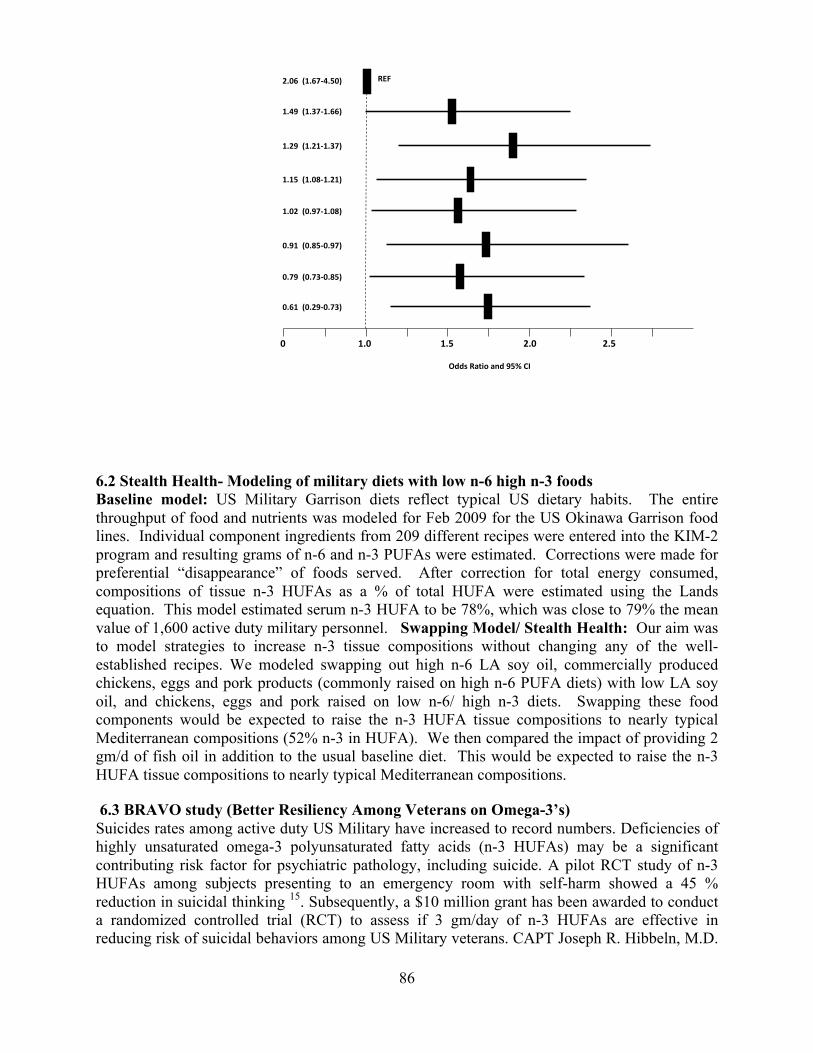
86
6.2 Stealth Health- Modeling of military diets with low n-6 high n-3 foods Baseline model: US Military Garrison diets reflect typical US dietary habits. The entire throughput of food and nutrients was modeled for Feb 2009 for the US Okinawa Garrison food lines. Individual component ingredients from 209 different recipes were entered into the KIM-2 program and resulting grams of n-6 and n-3 PUFAs were estimated. Corrections were made for preferential “disappearance” of foods served. After correction for total energy consumed, compositions of tissue n-3 HUFAs as a % of total HUFA were estimated using the Lands equation. This model estimated serum n-3 HUFA to be 78%, which was close to 79% the mean value of 1,600 active duty military personnel. Swapping Model/ Stealth Health: Our aim was to model strategies to increase n-3 tissue compositions without changing any of the well-established recipes. We modeled swapping out high n-6 LA soy oil, commercially produced chickens, eggs and pork products (commonly raised on high n-6 PUFA diets) with low LA soy oil, and chickens, eggs and pork raised on low n-6/ high n-3 diets. Swapping these food components would be expected to raise the n-3 HUFA tissue compositions to nearly typical Mediterranean compositions (52% n-3 in HUFA). We then compared the impact of providing 2 gm/d of fish oil in addition to the usual baseline diet. This would be expected to raise the n-3 HUFA tissue compositions to nearly typical Mediterranean compositions. 6.3 BRAVO study (Better Resiliency Among Veterans on Omega-3’s) Suicides rates among active duty US Military have increased to record numbers. Deficiencies of highly unsaturated omega-3 polyunsaturated fatty acids (n-3 HUFAs) may be a significant contributing risk factor for psychiatric pathology, including suicide. A pilot RCT study of n-3 HUFAs among subjects presenting to an emergency room with self-harm showed a 45 % reduction in suicidal thinking 15. Subsequently, a $10 million grant has been awarded to conduct a randomized controlled trial (RCT) to assess if 3 gm/day of n-3 HUFAs are effective in reducing risk of suicidal behaviors among US Military veterans. CAPT Joseph R. Hibbeln, M.D.
Odds Ratios of MaleSuicide Death and Octiles of DHA Status
2.01.0 2.51.50
Odds Ratio and 95% CI
REF
0.61 (0.29-‐0.73)
0.79 (0.73-‐0.85)
0.91 (0.85-‐0.97)
1.02 (0.97-‐1.08)
1.15 (1.08-‐1.21)
1.29 (1.21-‐1.37)
2.06 (1.67-‐4.50)
1.49 (1.37-‐1.66)
DHA %, mean (range)

87
SNN, LMBB and Prof. Bernadette Marriott, Ph.D. the Medical University of South Carolina are co- principal investigators on the award. References
1. Hibbeln JR, Davis JM, Steer C, Emmett P, Rogers I, Williams C, Golding J. 2007. Maternal seafood consumption in pregnancy and neurodevelopmental outcomes in childhood (ALSPAC study): an observational cohort study. Lancet 369: 578-85
2. U.S. Food and Drug Administration. January 15, 2009. Draft Risk & Benefit Assessment Report, Draft Summary of Published Research, Peer Review Report
3. Associate Parliamentary Food and Health Forum. 2008. The Links Between Diet and Behaviour. The influence of nutrition on mental health.
4. Steer CD, Davey Smith G, Emmett PM, Hibbeln JR, Golding J. FADS2 polymorphisms modify the effect of breastfeeding on child IQ. PLoS One 5: e11570
5. Pawlosky RJ, Hibbeln JR, Herion D, Kleiner DE, Salem N, Jr. 2009. Compartmental analysis of plasma and liver n-3 essential fatty acids in alcohol-dependent men during withdrawal. J Lipid Res 50: 154-61
6. Umhau JC, Zhou W, Carson RE, Rapoport SI, Polozova A, Demar J, Hussein N, Bhattacharjee AK, Ma K, Esposito G, Majchrzak S, Herscovitch P, Eckelman WC, Kurdziel KA, Salem N, Jr. 2009. Imaging incorporation of circulating docosahexaenoic acid into the human brain using positron emission tomography. J Lipid Res 50: 1259-68
7. Schuckit MA, Smith TL, Heron J, Hickman M, Macleod J, Lewis G, Davis JM, Hibbeln JR, Brown S, Zuccolo L, Miller LL, Davey-Smith G. 2011. Testing a level of response to alcohol-based model of heavy drinking and alcohol problems in 1,905 17-year-olds. Alcohol Clin Exp Res 35: 1897-904
8. Buydens-Branchey L, Branchey M, Hibbeln JR. 2009. Low plasma levels of docosahexaenoic acid are associated with an increased relapse vulnerability in substance abusers. Am J Addict 18: 73-80
9. Hibbeln JR. June 2010. Reduction of Relapse Among Chronic Alcoholics with n-3 HUFAs: A Randomized Placebo Controlled Trial. ISSFAL International Congress Maastricht Netherlands
10. Freeman MP, Hibbeln JR, Wisner KL, Davis JM, Mischoulon D, Peet M, Keck PE, Jr., Marangell LB, Richardson AJ, Lake J, Stoll AL. 2006. Omega-3 fatty acids: evidence basis for treatment and future research in psychiatry. J Clin Psychiatry 67: 1954-67
11. Golding J, Steer C, Emmett P, Davis JM, Hibbeln JR. 2009. High levels of depressive symptoms in pregnancy with low omega-3 fatty acid intake from fish. Epidemiology 20: 598-603
12. Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. 2011. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr 93: 950-62

88
13. Ramsden CE, Hibbeln JR, Majchrzak SF, Davis JM. 2010. n-6 fatty acid-specific and mixed polyunsaturate dietary interventions have different effects on CHD risk: a meta-analysis of randomised controlled trials. Br J Nutr 104: 1586-600
14. Avelheim A, Federova I, Osei-Hyiaman D, Pawlosky R, Hibbeln JR. 2010. Isolation of dietary linoleic acid, an endocannabinoid precursor, as a proximal cause of obesity. ISSFAL International Congress Maastricht Netherlands, Abstract
15. Hallahan B, Hibbeln JR, Davis JM, Garland MR. 2007. Omega-3 fatty acid supplementation in patients with recurrent self-harm. Single-centre double-blind randomised controlled trial. Br J Psychiatry 190: 118-22
Publications 2011-2012 1. Freeman MP, Hibbeln JR, Silver M, Hirschberg AM, Wang B, Yule AM, Petrillo LF,
Pascuillo E, Economou NI, Joffe H, Cohen LS. 2011. Omega-3 fatty acids for major depressive disorder associated with the menopausal transition: a preliminary open trial. Menopause-the Journal of the North American Menopause Society 18: 279-84
2. Ramsden CE, Hibbeln JR, Majchrzak SF, Davis JM. 2010. n-6 fatty acid-specific and
mixed polyunsaturate dietary interventions have different effects on CHD risk: a meta-analysis of randomised controlled trials. Br J Nutr 104: 1586-600
3. Wells EM, Jarrett JM, Lin YH, Caldwell KL, Hibbeln JR, Apelberg BJ, Herbstman J,
Halden RU, Witter FR, Goldman LR. 2011. Body burdens of mercury, lead, selenium and copper among Baltimore newborns. Environmental Research 111: 411-7
4. Harper KN, Hibbeln JR, Deckelbaum R, Quesenberry CP, Schaefer CA, Brown AS.
2011. Maternal serum docosahexaenoic acid and schizophrenia spectrum disorders in adult offspring. Schizophrenia Research 128: 30-6
5. Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. 2011. Changes in
consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. American Journal of Clinical Nutrition 93: 950-62
6. Lewis MD, Hibbeln JR, Johnson JE, Lin YH, Hyun DY, Loewke JD. 2011. Suicide
deaths of active-duty US military and omega-3 fatty-acid status: a case-control comparison. J Clin Psychiatry 72: 1585-90
7. Ramsden CE, Mann JD, Faurot KR, Lynch C, Imam ST, MacIntosh BA, Hibbeln JR, Loewke J,
Smith S, Coble R, Suchindran C, Gaylord SA. 2011. Low omega-6 vs. low omega-6 plus high omega-3 dietary intervention for Chronic Daily Headache: Protocol for a randomized clinical trial. Trials 12
8. Buydens-Branchey L, Branchey M, Hibbeln JR. 2011. Higher n-3 fatty acids are associated with more intense fenfluramine-induced ACTH and cortisol responses among cocaine-abusing men. Psychiatry Research 188: 422-7
9. Ramsden CE, Hibbeln JR, Majchrzak-Hong SF, Davis JM. 2011. Don't disregard the
essential distinction between PUFA species. British Journal of Nutrition 106: 953-7

89
10. Ramsden CE, Hibbeln JR, Majchrzak-Hong SF, Davis JM. 2011. Don't disregard the
essential distinction between PUFA species Comment on Ramsden et al Response. British Journal of Nutrition 106: 959-60
11. Schuckit MA, Smith TL, Heron J, Hickman M, Macleod J, Lewis G, Davis JM, Hibbeln
JR, Brown S, Zuccolo L, Miller LL, Davey-Smith G. 2011. Testing a level of response to alcohol-based model of heavy drinking and alcohol problems in 1,905 17-year-olds. Alcohol Clin Exp Res 35: 1897-904
12. Ramsden CE, Hibbeln JR, Majchrzak-Hong SF. 2011. All PUFAs are not created equal:
absence of CHD benefit specific to linoleic acid in randomized controlled trials and prospective observational cohorts. World Rev Nutr Diet 102: 30-43
13. Steer CD, Hibbeln JR, Golding J, Smith GD. 2012. Polyunsaturated fatty acid levels in
blood during pregnancy, at birth and at 7 years: their associations with two common FADS2 polymorphisms. Human Molecular Genetics 21: 1504-12
14. Alvheim AR, Malde MK, Osei-Hyiaman D, Lin YH, Pawlosky RJ, Madsen L, Kristiansen K, Froyland L, Hibbeln JR. 2012. Dietary Linoleic Acid Elevates Endogenous 2-AG and Anandamide and Induces Obesity. Obesity 20: 1984-94
15. Lin YH, Salem N, Wells EM, Zhou WY, Loewke JD, Brown JA, Lands WEM, Goldman LR, Hibbeln JR. 2012. Automated High-Throughput Fatty Acid Analysis of Umbilical Cord Serum and Application to an Epidemiological Study. Lipids 47: 527-39
16. Lin PY, Mischoulon D, Freeman MP, Matsuoka Y, Hibbeln J, Belmaker RH, Su KP. 2012. Are omega-3 fatty acids antidepressants or just mood-improving agents? The effect depends upon diagnosis, supplement preparation, and severity of depression. Mol Psychiatry 17: 1161-3; author reply 3-7
17. Lattka E, Koletzko B, Zeilinger S, Hibbeln JR, Klopp N, Ring SM, Steer CD. 2012.
Umbilical cord PUFA are determined by maternal and child fatty acid desaturase (FADS) genetic variants in the Avon Longitudinal Study of Parents and Children (ALSPAC). Br J Nutr: 1-15
18. Alvheim AR, Torstensen BE, Lin YH, Lillefosse HH, Lock EJ, Madsen L, Hibbeln JR, Malde MK. 2012. Dietary linoleic acid elevates endogenous 2-arachidonoylglycerol and anandamide in Atlantic salmon (Salmo salar L.) and mice, and induces weight gain and inflammation in mice. Br J Nutr: 1-10
19. Golding J, Northstone K, Emmett P, Steer C, Hibbeln JR. 2012. Do omega-3 or other
fatty acids influence the development of 'growing pains'? A prebirth cohort study. BMJ Open 2
20. Ramsden CE, Ringel A, Feldstein AE, Taha AY, MacIntosh BA, Hibbeln JR, Majchrzak-Hong SF, Faurot KR, Rapoport SI, Cheon Y, Chung YM, Berk M, Mann JD. 2012.

90
Lowering dietary linoleic acid reduces bioactive oxidized linoleic acid metabolites in humans. Prostaglandins Leukotrienes and Essential Fatty Acids 87: 135-41
21. Lin YH, Loewke JD, Hyun DY, Leazer J, Hibbeln JR. 2012. Fast Transmethylation of Serum Lipids Using Microwave Irradiation. Lipids 47: 1109-17
Collaborators Prof. Jean Golding, PhD Prof. John M. Davis, MD Prof. Bernadette Marriott, PhD Hee-Yong Kim, PhD Robert Pawlosky, PhD Marc Schuckit MD, Elizabeth Sublette, PhD John C. Umhau, MD Marlene Freeman, MD Robert Jones, PhD, CDC COL Michael Lewis, MD Andrew Young, PhD Laure Buydens-Branchey, MD DARPA NOAA Dr. Hibbeln: [email protected]

91
______________________________________________________________________
Laboratory of Neurogenetics ______________________________________________________________________
David Goldman, MD, Chief
The Laboratory of Neurogenetics (LNG), established 1992, studies the influence of genetic factors on neural mechanisms of alcoholism. Alcoholism is one of the most heritable of the addictions, which are common, complex diseases influenced by both substance-specific and shared genetic mechanisms (Goldman et al, 2005). The major paradigm of LNG is the study of inter-individual variation and its relation to behavior. The aim is to relate genotype to complex behavioral phenotypes, with the goal of identifying vulnerability and protective alleles responsible for the substantial heritability of alcoholism. To accomplish this, LNG studies intermediate phenotypes which assay functions of neurobiological systems integral to alcoholism and other addictions. These mechanisms include anxiety/stress response, executive cognitive function and behavioral control, and reward. Identification of alleles influencing these mechanisms will lead to better understanding of vulnerability, gene-environment interactions, diagnostic markers to individualize treatment, and molecular targets for intervention.
Methodology: Although moderately to highly heritable, alcoholism is multi-etiologic, and to some extent it is an end-stage diagnosis, meaning that the clinical phenotype does not well-reflect the genes or neural functional differences that influenced initial vulnerability and resilience. This conception is validated, at least in a crude way, by genetic studies reporting

92
association of alcoholism with diverse traits that are in some cases not biologically related but in other cases are actually mutually exclusive. For example extroversion (and impulsivity), introversion, alcohol response, alcohol metabolism and EEG differences including attentional responses have all been tied to alcoholism. None of these represents the full spectrum of vulnerability, but they represent a diversity of vulnerability mechanisms, and at least two (introversion/extroversion) appear to act in opposition. Mechanistic diversity of alcoholism has also been validated through the restricted, and still far from sufficient, collection of loci that have so far been identified to influence risk of alcoholism, in that the loci identified work on disparate processes such as alcohol metabolism (e.g. ALDH2 His487Lys), impulsivity (e.g. HTR2B Q20*) and cognition and anxiety (e.g. COMT Val158Met). To penetrate the etiologic heterogeneity of alcoholism and thereby detect the functional loci responsible for genetic risk and understand their actions, LNG utilizes sampling frameworks and phenotypes identified as relevant for alcoholism from neuroscience. We have studied founder populations in which genetic and environmental heterogeneity are reduced. We have identified alleles that modulate risk through specific mechanisms that include stress resiliency and impulsivity, and some of these also to be weaker predictors of the disease of alcoholism. LNG uses an integrative approach meshing multiple genetic and neurobehavioral paradigms. Genomic approaches include family linkage, genome wide association, exome sequencing, whole genome sequencing, whole genome evaluation of RNA expression by RNA-Seq and RNA microarrays, and whole-genome evaluation of epigenetic modifications to DNA and chromatin. To follow up and enhance genomic studies, functional genomics is conducted within in vitro systems and in animal models including the rhesus macaque, rat and mouse. We study the role of genes and environmental factors in powerful contexts including large population-based and family based datasets. These include population isolates such as Finns and American Indians, populations of individuals who have been exposed to risk factors such as stress, and case/control samples in which cases are extreme for the phenotype of interest and controls are otherwise well-matched. We collect and collaboratively study intermediate phenotypes including variations in electrophysiology and brain imaging on which gene actions are stronger than on the complex trait. We develop and extensively use linkage and genome informatics tools. Many of these studies are inherently complex, expensive and time-consuming, necessitating intramural and extramural collaborations enabling access to clinical and technical resources. Via The Center for Addictions Genomics (TCAG, Director, David Goldman, MD), we have partnered in the genomics of addiction with the National Institute of Drug Abuse (NIDA).
Organization: LNG has two sections: Human Neurogenetics led by David Goldman, also Chief of the Lab, and Comparative Behavioral Genomics led by Christina Barr, a Tenure-Track investigator. Within the Section of Human Neurogenetics is the DNA/Data Core led by Colin Hodgkinson. The Core manages large datasets and resources in genome informatics, linkage informatics, DNA sequencing, high throughput genotyping and expression analysis. The Center for Addictions Genomics (TCAG) is a partnership with NIDA in which LNG is the lead laboratory. Accomplishments: As a center for genomic studies on alcoholism, LNG is engaged in diverse projects. Highlighted discoveries include a functional NPY promoter locus whose effects on anxiety and stress response are stronger on molecules (RNA and the neuropeptide) and neuroimaging responses than on clinically observable behavior (Zhou et al, 2008), and we have continued to pursue effects of NPY, including on placebo response. We found an HTR2B stop codon with a role in impulsivity and alcoholism (Bevilacqua et al, Nature, 2010) and we and our collaborators have continued to follow through with functional studies on HTR2B, showing that

93
this receptor regulates phasic dopamine release crucial for addiction and other behavior. We found that a Grm2 stop codon is the origin of an alcohol preference QTL in the Alcohol-Preferring rat, a widely used model organism for alcoholism, causing uncompensated differences in glutamate function in these selected animals. The Grm2 stop codon is common in outbred Wistar rats, and thus of broad interest to neuroscientists who use this and similar model organisms (Zhou et al, submitted). In an imaging genetic study we showed that the CHRNA5 Asp398Asn polymorphism implicated in nicotine addiction strongly predicts the strength of this addiction-related circuit (Hong et al, PNAS, 2010) and an EEG GWAS detected three genome wide significant regions (Hodgkinson et al, PNAS, 2010). More broadly, LNG’s contributions are:
1) Analysis of alcoholism, a complex disorder, in isolates and founder populations that are
genetically and environmentally more homogeneous, and in animal models including strains selected for alcoholism-related phenotypes. We defined and characterized the impact of alcoholism in American Indian communities, which on a worldwide basis are among the most afflicted by alcoholism, but neglected by research. The studies extend from analysis of phenotype and familial transmission to two whole genome linkage scans identifying putative locations of vulnerability genes, a genome wide association study, and a collaborative pharmacogenetic study (with Stephanie O’Malley, Yale) on treatment response in Alaskan Natives.
2) A leading role in identification of functional variants in neurogenetic candidate genes.
Common functional variants LNG discovered or demonstrated to have in vitro functionality include serotonin, dopamine, opioid receptor variants, neuropeptide Y, the serotonin transporter, and the common stop codon in HTR2B that is restricted to the Finnish population (Bevilacqua et al, 2010). These are major targets in behavior and neuropharmacology.
3) LNG is a groundbreaking genetics laboratory for studies relating alleles to in vivo
functional measures. We led or participated as the genetic component in seminal “imaging genetics” studies on genes such as COMT and HTTLPR. In other genetic studies with neuroimaging components, we discovered multilevel association of a haplotype determining low expression of neuropeptide Y (an anxiolytic peptide) to NPY mRNA and peptide levels, brain imaging responses, and anxiety and anxiety disorders (Zhou et al, 2008). A functional variant of MnSOD2 alters response to oxidative stress and predicts brain shrinkage in alcoholics (Srivastava et al, 2010). A functional polymorphism of the human HTR2C receptor gene alters striatal dopamine release (Mickey et al, 2012). We discovered that CHRNA5 Asp398Asn, a functional nicotinic receptor missense variant linked to nicotine addiction by GWAS, may alter risk by altering connectivity of a circuit linking anterior cingulate to ventral striatum (Hong et al, 2010). Our discovery of the role of the HTR2B stop codon in impulsivity led us to further studies of this receptor in collaboration with other labs, showing that in mice and people this receptor is located on dopamine neurons and regulates phasic dopamine release. Our discovery that a Grm2 stop codon is an allele leading to increased alcohol preference in the P (Alcohol Preferring) rat was translated into an understanding that this leads to many of the global differences in behavior and glutamate function observed in these animals. Recently, we discovered brain expression and functional variation in the human vesicular

94
monoamine transporter (VMAT1) that modulates emotional processing (Lohoff et al, 2013). The functional studies are now extending to studies on stem cells and neurons isolated from living patients, and using induced pluripotent stem cells from blood (for example in a close partnership with Peter Schmidt, NIMH, on the cellular basis of premenstrual dysphoric disorder).
4) LNG is a collaborative center enhancing efforts of intramural and extramural labs. We
developed the Addictions Array, a 130 gene, 1536 SNP array and used it to assist more than a dozen external labs (>20,000 samples), performing the genotyping for most (Hodgkinson et al, 2008). Applications of this array include pharmacogenetic studies. In the COMBINE study we replicated association of the functional Asp40 allele to favorable clinical outcome in alcoholics treated with naltrexone (Anton et al, 2008). A functionally deficient variant of MnSOD2, an antioxidant enzyme, increased gray matter loss in moderate drinkers (Srivastava et al, 2010). We work collaboratively with other laboratories using our capabilities in massively parallel DNA sequencing and genotyping, for example performing genome wide association in suicide (Columbia Suicide Consortium) and alcoholism (with the Laboratory of Clinical and Translational Studies, NIAAA).
5) LNG is an exponent of gene/environment interaction, including the use of intermediate phenotypes and cross-population studies. We found that the common, stable low voltage alpha EEG trait [LVA] identifies a subtype of alcoholism associated with anxiety disorders, and in two American Indian studies has shown the crucial role of early life stress exposures in increasing the risk of alcoholism and other psychiatric disorders. A genome-wide association study (GWAS) revealed several genome-wide significant regions that regulate the resting EEG. In the Memphis Longitudinal Study (extramural PI, David Olds) we are investigating gene by environment interactions in high risk children.
6) With P.J. Brooks, a former LNG scientist and now at NIAAA extramural and the Office
of Rare Diseases and with Akira Yokoyama (Kurihama institute), we performed whole genome sequencing for mutations associated with upper GI cancer in ALDH2 deficient individuals who consume alcohol – acetaldehyde being a potent mutagen.
7) Christina Barr, VMD, PhD, a Tenure Track investigator, makes powerful cross-species
comparisons using polymorphisms in non-human primates that are orthologous to human polymorphisms. These studies led to a better understanding of the roles of human polymorphisms in genes such as the serotonin transporter, MAOA, NPY and OPRM1. For example there are functionally similar CRH polymorphisms in both species that have strong effects on HPA function and behavior in the rhesus macaque (Barr et al, 2008). Deep sequencing also enabled us to take a genome-level view of variation in nonhuman primates, as compared to humans (Yuan et al, 2012), finding that the macaque, a widely used model organism, is more variable but less variable than would be expected in functional variation, a signature of selection.

95
References
Anton RF, Oroszi G, O'Malley S, Couper D, Swift R, Pettinati H, Goldman D. 2008. An evaluation of mu-opioid receptor (OPRM1) as a predictor of naltrexone response in the treatment of alcohol dependence: results from the Combined Pharmacotherapies and Behavioral Interventions for Alcohol Dependence (COMBINE) study. Arch Gen Psychiat 65: 135-44 Barr C, Dvoskin RL, Yuan, Q, Lipsky, R, Gupte, M, Hu X, Zhou Z, Schwandt M, Lindell, SG, McKee M, Becker ML, Kling MA, Gold P, Higley D, Heilig M, Suomi SJ, Goldman, D. 2008. CRH Haplotype as a Factor Influencing Cerebrospinal Fluid Levels of Corticotropin-Releasing Hormone, Hypothalamic-Pituitary-Adrenal Axis Activity, Temperament, and Alcohol Consumption in Rhesus Macaques. Arch Gen Psychiat 65: 934-944 Bevilacqua L, Doly S, Kaprio J, Yuan Q, Tikkanen R, Paunio T, Zhou Z, Wedenoja J, Maroteaux L, Diaz S, Belmer A, Hodgkinson CA, Dell'osso L, Suvisaari J, Coccaro E, Rose RJ, Peltonen L, Virkkunen M, Goldman D. 2010. A population-specific HTR2B stop codon predisposes to severe impulsivity. Nature 468: 1061-1066
Goldman D, Oroszi G, Ducci F. The genetics of addictions: uncovering the genes. 2005.
Nat Genet Rev 6: 521-32
Hodgkinson CA, Enoch MA, Srivastava V, Cummins-Oman JS, Ferrier C, Iarikova P, Sankararaman S, Yamini G, Yuan Q, Zhou Z, Albaugh B, White KV, Shen PH, Goldman D. 2010. Genome-wide association identifies candidate genes that influence the human electroencephalogram. Proc Natl Acad Sci U S A 107: 8695-700
Hong LE, Hodgkinson CA, Yang Y, Sampath H, Ross TJ, Buchholz B, Salmeron BJ, Srivastava V, Thaker GK, Goldman D, Stein EA. 2010. A genetically modulated, intrinsic cingulate circuit supports human nicotine addiction. Proc Natl Acad Sci U S A 107: 13509-14
Lohoff FW, Hodge R, Narasimhan S, Nall A, Ferraro TN, Mickey BJ, Heitzeg MM, Langenecker SA, Zubieta JK, Bogdan R, Nikolova YS, Drabant E, Hariri AR, Bevilacqua L, Goldman D, Doyle GA. 2013. Functional genetic variants in the vesicular monoamine transporter 1 modulate emotion processing. Mol Psychiatry [Epub ahead of print]
Mickey BJ, Sanford BJ, Love TM, Shen PH, Hodgkinson CA, Stohler CS, Goldman D, Zubieta JK. 2012. Striatal dopamine release and genetic variation of the serotonin 2C receptor in humans. J Neurosci 32: 9344-50
Srivastava V, Buzas B, Momenan R, Oroszi G, Pulay AJ, Enoch MA, Hommer DW, Goldman D. 2010. Association of SOD2, a mitochondrial antioxidant enzyme, with gray matter volume shrinkage in alcoholics. Neuropsychopharmacology 35: 1120-8
Yuan Q, Zhou Z, Lindell SG, Higley JD, Ferguson B, Thompson RC, Lopez JF, Suomi SJ, Baghal B, Baker M, Mash DC, Barr CS, Goldman D. 2012 The rhesus macaque is three times as diverse but more closely equivalent in damaging coding variation as compared to the human. BMC Genet 13: 52

96
Zhou Z, Zhu G, Hariri AR, Enoch MA, Scott D, Sinha R, Virkkunen M, Mash DC, Lipsky RH, Hu XZ, Hodgkinson CA, Xu K, Buzas B, Yuan Q, Shen PH, Ferrell RE, Manuck SB, Brown SM, Hauger RL, Stohler CS, Zubieta JK, Goldman D. 2008. Genetic variation in human NPY expression affects stress response and emotion. Nature 452: 997-1001
Zhou Z, Yuan Q, Mash DC, Goldman D. 2011. Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol. Proc Natl Acad Sci U S A 108: 6626-31
David Goldman: [email protected]

97
______________________________________________________________________
Section of Human Neurogenetics ____________________________________________________________
David Goldman, MD, Chief Mary-Anne Enoch, MD, Staff Scientist Colin Hodgkinson, PhD, Staff Scientist Kornel Schuebel, PhD, Staff Scientist Qiaoping Yuan, PhD, Computer Scientist Pei-hong Shen, Computer Scientist Elisa Moore, Computer Scientist Laura Bevilacqua, MD, Visiting Fellow1 Vibhuti Srivastava, PhD, Visiting Fellow2 Rony Panarsky, PhD, Visiting Fellow Seungeun Yeo, Visiting Fellow Zhifeng Zhou, PhD, Scientist 1 New York University Department of Psychiatry (Residency); 2 MD Anderson Hospital; 3 Tauro College of Osteopathic Medicine; 4 Kurihama Institute. Human Neurogenetics, directed by David Goldman, MD, identifies functional loci that modulate pathways to vulnerability to alcoholism, other addictions, and related psychiatric disorders. To accomplish this it generates clinical datasets and collaborates with multiple laboratories. Its activities encompass human research protocols, large scale DNA sequence variant detection using massively parallel sequencing, array and capillary electrophoresis based genotyping, in vitro and in vivo functional analyses of receptor variants, linkage studies of markers and candidate alleles, genome linkage scans, genome-wide associations, genome-wide epigenetic analyses, transcriptome analyses, and genome informatics. Brief biographical sketches of members of the Section of Human Neurogenetics:
Mary-Anne Enoch, MD, (Staff Scientist) joined LNG in 1995. She pursues two major interests: the use of intermediate phenotypes, including electrophysiological phenotypes and dimensional anxiety, for understanding the genetics and neurobiology of alcoholism and co-morbid disorders, and the influence of gene-environment interactions on addiction. The large psychiatric
and electrophysiological phenotype datasets Dr. Enoch developed, or for which she led electrophysiology studies, are US Caucasians, Plains Indians, and Southeastern American Indians. She has published papers on the interactions of sex with gene effects, and more recently on the interaction of childhood trauma and genetic variation to increase risk for addictive disorders and associated psychopathology. Recent findings include associations between GABRA2 and GABRG1 haplotypes and alcoholism, additive effects between functional variants in 5-HTTLPR and HTR3B on risk for alcohol and drug dependence, and the association of CRHBP, identified as a candidate gene through a whole genome linkage scan, with EEG power, alcoholism and anxiety. Dr. Enoch serves on the NIH Neuroscience IRB.
Longina Akhtar, Biologist Cheryl Marietta, Biologist Basel Baghal, Postbac IRTA 3 Isioma Mordi, Postbac IRTA Sasha Rosser, Postbac IRTA Zach Hommer, Postbac IRTA Bo-Jeong Kim, Postbac IRTA Mitsuru Kimura, MD, Visiting Scientist4 Elena Gorodetsky, PhD, Contractor Serguei Zoubak, PhD, Guest Researcher P.J. Brooks, PhD, Guest Researcher

98
Colin Hodgkinson, PhD, (Staff Scientist) joined LNG in 2003 and leads the DNA/Data core. He directs high throughput sequencing and genotyping, genetic linkage, and functional genetic analyses. Using the high-density haplotype approach in a large case-control dataset, Dr. Hodgkinson replicated and extended linkage of DISC1 to schizoaffective disorder, and is exploring the
relationship of DISC1 and genes to which it is functionally related in various phenotypes. He identified multiple genes influencing the electroencephalogram by genome-wide association.
Kornel Schuebel, PhD, (Staff Scientist) joined LNG in 2008. He is intimately involved with all aspects of genome-wide discovery and “functionation” of DNA methylation, chromatin, and RNA metabolism as it pertains to gene expression alterations in pathological brain disorders, with a focus on fetal alcohol syndrome. Dr. Schuebel directs analyses utilizing massively parallel
sequencing.
Laura Bevilacqua, MD, was a Fogarty fellow appointed 2007, and in 2012 became a Resident in Psychiatry at New York University. She received her M.D. in 2005 from the University of Pisa, Italy, where she also trained in Psychiatry. She is interested in functional genomics, from genome informatics to in vivo and in vitro functional assays. She is focusing on the role of serotonin
and dopamine domain genes in impulsivity and dyscontrol towards the identification of functional alleles in addictions and related behaviors. In this respect she performed next generation sequencing studies on dyscontrol behaviors and plans to extend it to whole genome transcriptomes and microRNAs. She discovered a stop codon that is common in the Finnish population and that influences impulsivity.
Rony Panarsky, PhD, is a Fogarty fellow from the Hebrew University of Jerusalem, Israel, appointed 2013. He is interested in understanding mechanisms of addiction using cell and whole animal models. He uses microarray and next generation sequencing technology for whole genome transcriptome and DNA methylation analyses together with induced pluripotent
stem cells and neurons derived from these cells.
Seungeun Yeo, PhD, is a Fogarty fellow from Yeungnam University, South Korea, appointed 2011. She is interested in understanding mechanisms of addiction and behavior using genome wide detection of genes that have cis-regulatory loci altering gene expression in neural tissue. She uses microarray and next generation sequencing technology for whole genome transcriptome
analyses with analysis of differential allele expression.
Zhifeng Zhou, PhD, who joined LNG in 2002, is a molecular biologist. He uses high-throughput sequencing approaches to study drug- and alcohol-related genome-wide changes in gene expression and epigenetic regulation. Dr. Zhou also uncovered multilevel effects of haplotype-based neuropeptide Y (NPY) gene expression variation on emotion and stress responses and linked
haplotype-based genetic association of the TPH2 gene with susceptibility to depression and suicidal behavior. Recently he has mapped gene expression and epigenetic change in the hippocampus of alcoholics and cocaine addicts and performed a comparative study of the P and

99
NP rats, identifying an mGluR2 stop codon responsible for changes in glutamate function in P rats, and partially responsible for alcohol preference in this strain.
Qiaoping Yuan, PhD, who joined LNG in 2005 is a bioinformatics and analytical IT tools resource for all members of LNG. He is in the DNA/Data Core where he leads genome informatics including creating analytic frameworks and pathways for the next-gen sequencer data. He is involved in all genomics projects of LNG, both at the front end of designing arrays and
planning analyses, and at the analytical phases, for example publishing a paper describing overall enhanced genetic diversity without proportional increase in functional variation in the rhesus macaque, an important model organism in biomedical research.
Pei-Hong Shen, MS, who joined LNG in 2004 is involved in all aspects of data analysis and data management to identify genetic variation, define molecular functionality, and link genetic variation to intermediate phenotypes and complex behavior. She is a bioinformatics and analytical IT tools resource for all members of LNG.
Cheryl Marietta, MS, with NIAAA since 1980, is a Senior Research Assistant. She is involved in the Induced Pluripotent Stem Cell (IPSC) project and will be generating neurons from the IPSC’s as well as assisting with various other projects. She is Chair of the NIAAA Safety Committee.
Longina Akhtar, MS, with LNG since 1983, manages the front-end of multiple collaborations requiring preparation of DNA and/or lymphoblastoid cell lines. She performs or supervises all the primary cell culture, DNA extractions from cells and blood, and cell storage – including off-site backup. She assists in the construction of DNA panels for linkage studies. Her position
is broad in scope and responsibility in that she is the direct, and frequent point of interaction with many collaborating investigators.
Elisa Moore, Sr. Systems Analysis/Specialist and Robotics, has been with LNG since 1993. She plans, designs, and implements information systems that meet LNG’s unique and demanding bioinformatics requirements. She designed network VLAN’s enabling terabyte data-flows between next-generation sequencers, cluster servers and mass storage arrays. She supports specialized
computers on DNA sequencers, analyzers and real-time PCR equipment. She also programs DNA liquid handling robotics and works on other “front-end” aspects of high throughput genetic analysis.
Elena Gorodetsky, MD, PhD, a part of LNG 2003, is a former Guest Researcher and now a Contractor performing and coordinating large scale sequencing and genotyping projects, and expanding on analyses of selected model organisms.

100
Mitsuru Kimura MD, PhD, a member of LNG 2009-2012, was a guest researcher on sabbatical from the Kurihama Alcoholism Center in Tokyo. He received his Ph.D. from Keio University in 2009. He studies the clinical and molecular effects of alcohol-metabolizing enzyme variants. He is interested in
epigenetic change induced by chronic exposure to alcohol and acetaldehyde.
P.J. Brooks, PhD, who has been affiliated with LNG for the past decade, has been an Adjunct investigator since 2010. He is now with the NIAAA Division of Metabolism and Health Effects (DMHE), working on the relationship of alcohol to cancer. He also has an appointment with the NIH Office of Rare Diseases Research. He collaborates on studies of the interaction of alcohol and
alcohol metabolizing genotypes to produce mutation, and cancer.
Neelima Dubey, PhD, a part of LNG since 2011, is working on a sex-steroid related project with Dr. Peter J. Schmidt (NIMH). Dr Dubey performs whole transcriptome sequencing and DNA methylation studies and is also working on induced pluripotent stem cell (IPSCs) from these patients towards studies in neuronal cells, and to understand the neural basis of differences in behavioral
hormonal responses in women.
Serguei Zoubak, PhD, has collaborated with LNG as a Special Volunteer since 2010. CNRM (Center for Neuroscience and Regenerative Medicine) was established to address the current needs of the medical community to better diagnose and intervene for the prevention of the long term consequences resulting from traumatic brain injury (TBI), particularly in the context
experienced by service members in Operation Iraqi Freedom and Operation Enduring Freedom. Dr Zoubak works with the dataset of Vietnam War Veterans with different forms of traumatic brain injury to discover genetic predictors underlying impairment of cognitive and sensory motor learning skills that accompany many forms of TBI.

101
Trainees in the Section of Human Genetics:
Bojeong Kim, BS, 2010 Cornell University Biological Engineering Pre-Medical Zach Hommer, BA, 2008 University of Maryland History Pre-Medical Basel Baghal, BS, 2009 Rutgers University Physics Pre-Medical
Isioma Mordi, BS, 2008 InterAmerican University of Puerto Rico Biotechnology Pre-Medical Sasha Rosser, BS, 2011 George Mason University Neuroscience Pre-Graduate
Erica Busch, BS, 2012 University of Virginia Biology Pre-Graduate (Based in the NIAAA Lab of Behavioral and Genomic Neuroscience)
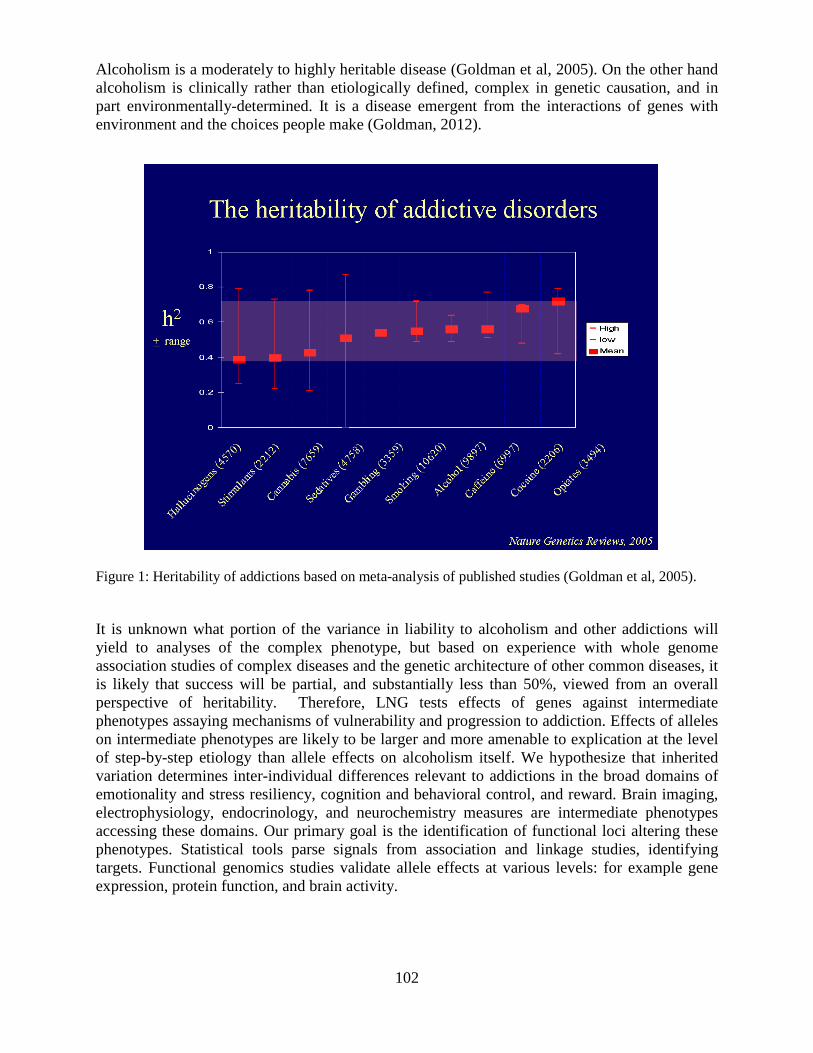
102
Alcoholism is a moderately to highly heritable disease (Goldman et al, 2005). On the other hand alcoholism is clinically rather than etiologically defined, complex in genetic causation, and in part environmentally-determined. It is a disease emergent from the interactions of genes with environment and the choices people make (Goldman, 2012).
Figure 1: Heritability of addictions based on meta-analysis of published studies (Goldman et al, 2005). It is unknown what portion of the variance in liability to alcoholism and other addictions will yield to analyses of the complex phenotype, but based on experience with whole genome association studies of complex diseases and the genetic architecture of other common diseases, it is likely that success will be partial, and substantially less than 50%, viewed from an overall perspective of heritability. Therefore, LNG tests effects of genes against intermediate phenotypes assaying mechanisms of vulnerability and progression to addiction. Effects of alleles on intermediate phenotypes are likely to be larger and more amenable to explication at the level of step-by-step etiology than allele effects on alcoholism itself. We hypothesize that inherited variation determines inter-individual differences relevant to addictions in the broad domains of emotionality and stress resiliency, cognition and behavioral control, and reward. Brain imaging, electrophysiology, endocrinology, and neurochemistry measures are intermediate phenotypes accessing these domains. Our primary goal is the identification of functional loci altering these phenotypes. Statistical tools parse signals from association and linkage studies, identifying targets. Functional genomics studies validate allele effects at various levels: for example gene expression, protein function, and brain activity.

103
Identifying targets for functional genomics The Primary Hypotheses of LNG and the Section of Human Neurogenetics are:
1. The genetic architecture of alcoholism is simplified in founder populations with reduced genetic heterogeneity, enabling detection of effects of alleles that are rare worldwide.
2. Allele effects are stronger on relevant intermediate phenotypes. Effects on intermediate phenotypes offer a path to validate allele effects and understand mechanism and might eventually contribute to the neuroscience-based redefinition of alcoholism.
3. Allele effects are stronger in contexts of exposure.
The Aims are:
Aim 1: In founder populations, using phenotypes based on the neuroscience of addiction, identify genomic regions harboring risk alleles.
Aim 2: Identify functional loci influencing alcoholism and trace allele effects through intermediate phenotypes and studies of allele effects in appropriate contexts of exposure. Aim 3: Validate allele effects in animal models.
Genome-wide scans for linkage and whole genome association The genome wide association study (GWAS) offers the ability to detect the action on a trait of alleles of moderate frequency (i.e. >0.05) and effect (i.e. odds ratio >1.5). As such, GWAS is highly complementary to family linkage. LNG recently completed a whole genome family linkage scan (Enoch et al, 2008) and GWAS (Illumina 550k panel) in its Plains Indian alcoholism/electrophysiology dataset. The resting EEG measures collected in this sample are intermediate phenotypes in arousal-related behaviors such as alcoholism and anxiety disorders. The family linkage yielded a genome wide significant signal in an interval which includes CRHBP (Enoch et al, 2008). The GWAS detected genome-wide significant signals at multiple locations (Figure 2) including a whole-genome significant peak on Chromosome 1 in a region previously implicated for Beta2 EEG power (Hodgkinson et al, 2010).
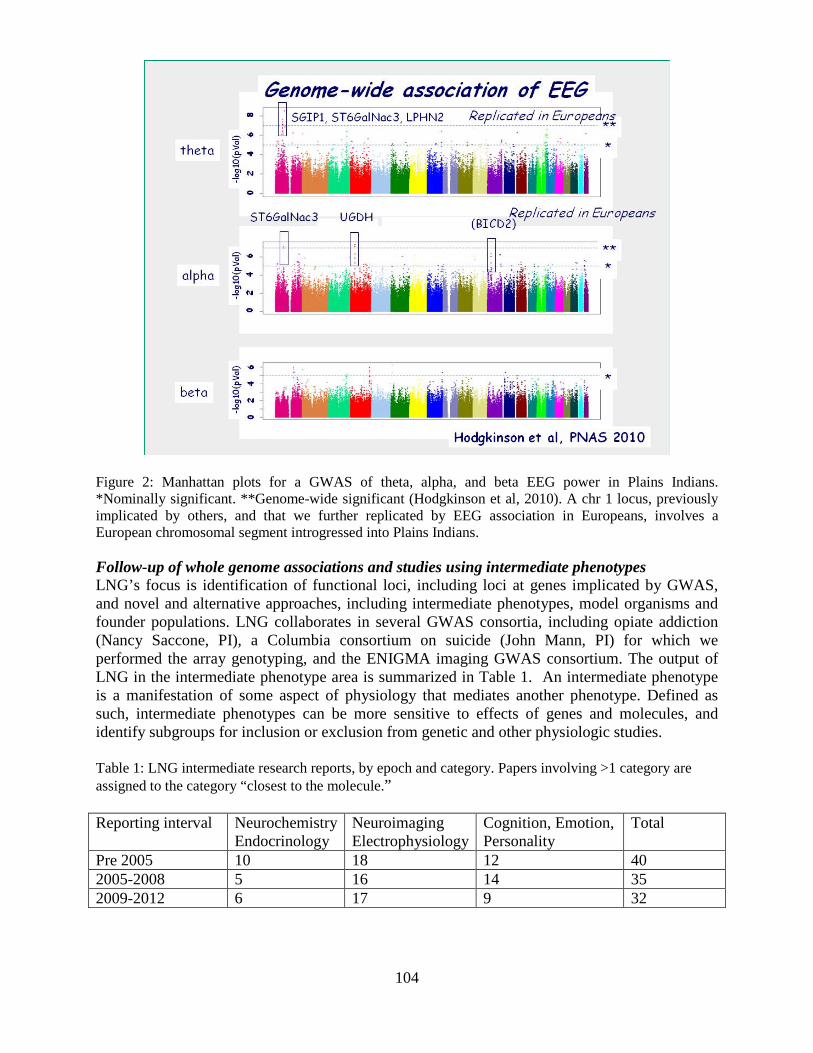
104
Figure 2: Manhattan plots for a GWAS of theta, alpha, and beta EEG power in Plains Indians. *Nominally significant. **Genome-wide significant (Hodgkinson et al, 2010). A chr 1 locus, previously implicated by others, and that we further replicated by EEG association in Europeans, involves a European chromosomal segment introgressed into Plains Indians. Follow-up of whole genome associations and studies using intermediate phenotypes LNG’s focus is identification of functional loci, including loci at genes implicated by GWAS, and novel and alternative approaches, including intermediate phenotypes, model organisms and founder populations. LNG collaborates in several GWAS consortia, including opiate addiction (Nancy Saccone, PI), a Columbia consortium on suicide (John Mann, PI) for which we performed the array genotyping, and the ENIGMA imaging GWAS consortium. The output of LNG in the intermediate phenotype area is summarized in Table 1. An intermediate phenotype is a manifestation of some aspect of physiology that mediates another phenotype. Defined as such, intermediate phenotypes can be more sensitive to effects of genes and molecules, and identify subgroups for inclusion or exclusion from genetic and other physiologic studies.
Table 1: LNG intermediate research reports, by epoch and category. Papers involving >1 category are assigned to the category “closest to the molecule.”
Reporting interval Neurochemistry
Endocrinology Neuroimaging Electrophysiology
Cognition, Emotion, Personality
Total
Pre 2005 10 18 12 40 2005-2008 5 16 14 35 2009-2012 6 17 9 32
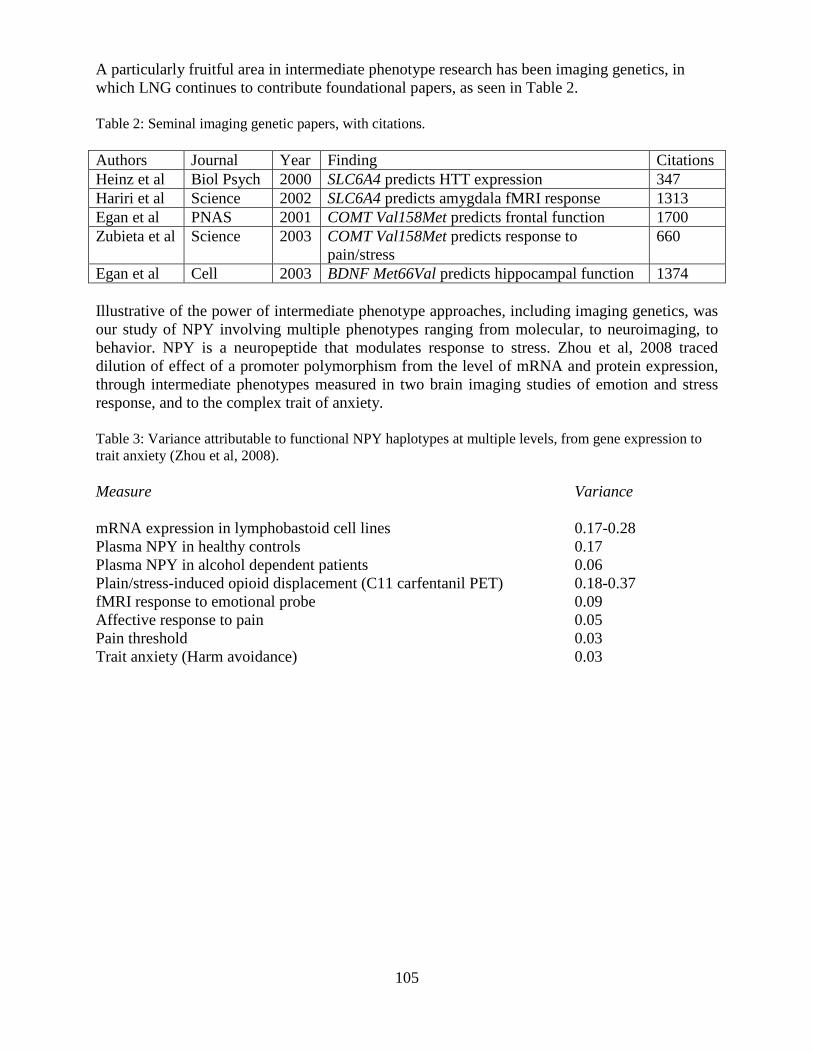
105
A particularly fruitful area in intermediate phenotype research has been imaging genetics, in which LNG continues to contribute foundational papers, as seen in Table 2. Table 2: Seminal imaging genetic papers, with citations.
Authors Journal Year Finding Citations Heinz et al Biol Psych 2000 SLC6A4 predicts HTT expression 347 Hariri et al Science 2002 SLC6A4 predicts amygdala fMRI response 1313 Egan et al PNAS 2001 COMT Val158Met predicts frontal function 1700 Zubieta et al Science 2003 COMT Val158Met predicts response to
pain/stress 660
Egan et al Cell 2003 BDNF Met66Val predicts hippocampal function 1374 Illustrative of the power of intermediate phenotype approaches, including imaging genetics, was our study of NPY involving multiple phenotypes ranging from molecular, to neuroimaging, to behavior. NPY is a neuropeptide that modulates response to stress. Zhou et al, 2008 traced dilution of effect of a promoter polymorphism from the level of mRNA and protein expression, through intermediate phenotypes measured in two brain imaging studies of emotion and stress response, and to the complex trait of anxiety. Table 3: Variance attributable to functional NPY haplotypes at multiple levels, from gene expression to trait anxiety (Zhou et al, 2008). Measure Variance mRNA expression in lymphobastoid cell lines 0.17-0.28 Plasma NPY in healthy controls 0.17 Plasma NPY in alcohol dependent patients 0.06 Plain/stress-induced opioid displacement (C11 carfentanil PET) 0.18-0.37 fMRI response to emotional probe 0.09 Affective response to pain 0.05 Pain threshold 0.03 Trait anxiety (Harm avoidance) 0.03
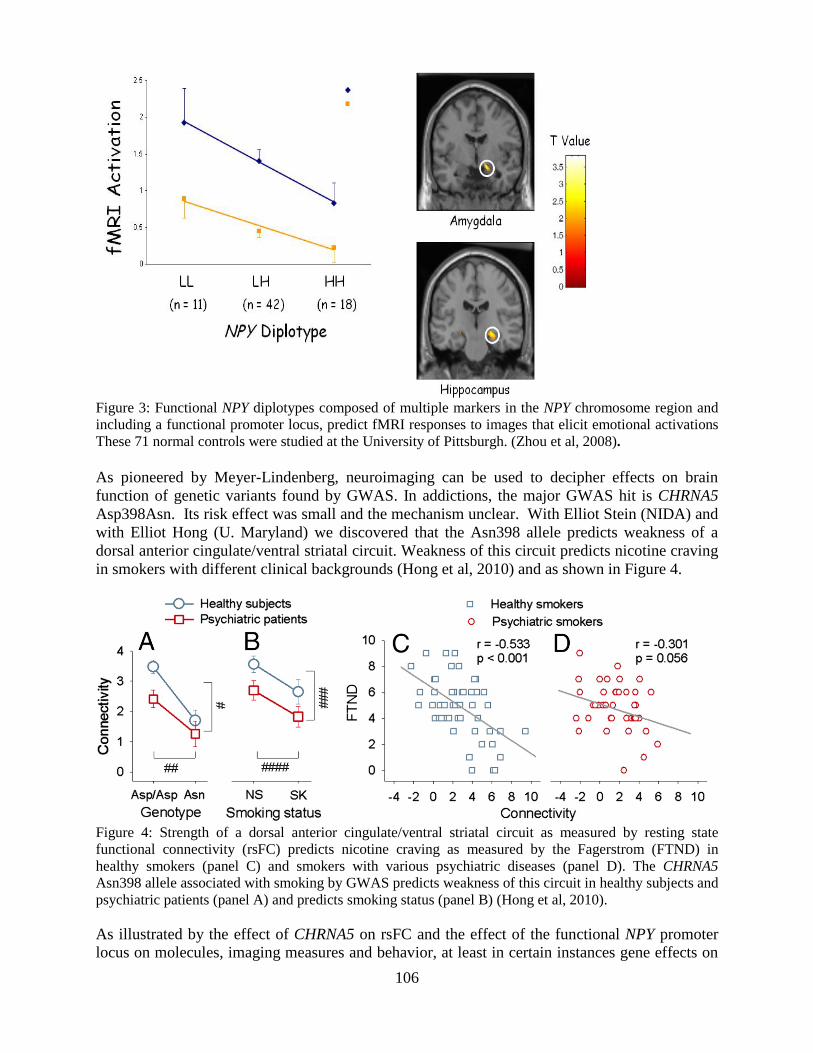
106
Figure 3: Functional NPY diplotypes composed of multiple markers in the NPY chromosome region and including a functional promoter locus, predict fMRI responses to images that elicit emotional activations These 71 normal controls were studied at the University of Pittsburgh. (Zhou et al, 2008). As pioneered by Meyer-Lindenberg, neuroimaging can be used to decipher effects on brain function of genetic variants found by GWAS. In addictions, the major GWAS hit is CHRNA5 Asp398Asn. Its risk effect was small and the mechanism unclear. With Elliot Stein (NIDA) and with Elliot Hong (U. Maryland) we discovered that the Asn398 allele predicts weakness of a dorsal anterior cingulate/ventral striatal circuit. Weakness of this circuit predicts nicotine craving in smokers with different clinical backgrounds (Hong et al, 2010) and as shown in Figure 4.
Figure 4: Strength of a dorsal anterior cingulate/ventral striatal circuit as measured by resting state functional connectivity (rsFC) predicts nicotine craving as measured by the Fagerstrom (FTND) in healthy smokers (panel C) and smokers with various psychiatric diseases (panel D). The CHRNA5 Asn398 allele associated with smoking by GWAS predicts weakness of this circuit in healthy subjects and psychiatric patients (panel A) and predicts smoking status (panel B) (Hong et al, 2010). As illustrated by the effect of CHRNA5 on rsFC and the effect of the functional NPY promoter locus on molecules, imaging measures and behavior, at least in certain instances gene effects on

107
intermediate phenotypes are larger than on emergent behavioral traits. The value of intermediate phenotypes is easily over-generalized, in particular because of their expense but also because of the need to directly relate the intermediate phenotype to the disease, and to understand mechanistic relevance. For both CHRNA5 and NPY we took some of those steps. In the case of NPY, mechanistic relevance of NPY level to stress response was shown by others, for example in studies of war veterans; amygdala fMRI response predicts emotionality, and C11 carfentanil displacement predicts pain threshold and affective response to pain following pain challenge. In each of these instances there is also strong evidence from neuroscience that these correlations represent causation, and not reverse causation, which would anyway not well explain the stronger effects of the genes on the intermediate phenotypes. As reviewed (Goldman and Ducci, 2007), effect sizes of alleles on intermediate phenotypes were larger (median odds ratio 3.9) as compared to allele effects on more heterogeneous complex traits (odds ratio 1.9 for 24 WGA loci identified for seven complex diseases in the WTCC study), those complex traits also having different measurement properties. Selection bias is present because larger intermediate phenotype datasets would yield additional associations (both true and false-positive) with alleles of smaller effect. In addition, several functional loci that alter neuroimaging phenotypes are imperfectly captured by large SNP arrays used for GWAS, and therefore the failure to detect GWAS signals at these genes is unremarkable. Overall, we conclude that genetic studies with intermediate phenotypes are an increasingly important paradigm for detecting, validating and understanding effects of genes in complex behavior. For several functional loci, multiple convergent results involving intermediate phenotypes are now available, with contributions from LNG. Other functional loci that appear to have validated effects in complex behavior include the MAOA VNTR (dyscontrol), HTTLPR (emotionality), COMT (emotionality and cognition), and BDNF (hippocampal structure and episodic memory). LNG has contributed to each of these discoveries as has been reported in scientific journals and described in more detail elsewhere (including previous editions of this publication). For HTTLPR, a functional serotonin transporter promoter locus, we found association to brain transporter density, prediction of amygdala metabolic response after emotional stimuli, gene x gene interaction of HTTLPR and COMT in processing emotional stimuli, and a new functional HTTLPR allele and gene x stress interaction predicting suicidality in substance dependent patients. For COMT, we participated in the first study linking Val158Met to cognition, replicated the association of Val158Met to cognition in various contexts, and discovered the role of COMT in emotion and the processing of aversive stimuli. Gene x environment and gene x gene interaction For each of the functional loci just discussed there are particular results in the literature that are likely to constitute false positive and false negative signals. Also, effect sizes (and thus predictive value) are difficult to estimate, and are in flux. It is important that functional loci are studied in different contexts and with more refined approaches accessing different domains of brain function, representing a maturation of the science in the form of more fine-grained analysis. Understanding allele effects on behavior also requires gene x gene as well as gene x environment interaction studies, because allele effects may not be additive. Several recent discoveries of gene by stress and gene by gene interaction are summarized: FKBP5 and CRHBP: 1. FKBP5 and CRHBP variation predispose, independently and additively, to suicidal behavior in substance dependent African Americans who have experienced significant childhood trauma (Roy et al, 2010, Roy et al, 2012).

108
2. FKBP5 interacts with childhood trauma, particularly physical abuse, to predict overt aggressive behavior in Italian prisoners (Bevilacqua et al, 2012). GABRA2 1. In a group of African American substance dependent individuals, one of the two uniquely African GABRA2 haplotypes predicted heroin addiction, whereas the other haplotype appeared to confer resilience to addiction after exposure to severe childhood trauma. There was also an independent GABRA2 SNP that interacted with childhood trauma to influence addiction vulnerability, particularly to cocaine (Enoch et al, 2010a). 2. GABRA2 and GABRG1 haplotypes were shown to be independent predictors of alcoholism in two samples: Finns and Plains Indians (Enoch et al, 2009). HTR3B 1. In a sample of African American substance dependent patients, an HTR3B gain-of-function allele and low 5-HTTLPR activity had an additive (non-interactive) effect on alcohol+drug dependence that accounted for 13% of the variance. Potentially, increased synaptic 5-HT coupled with increased 5-HT3 receptor responsiveness may result in enhanced dopamine transmission in the reward circuit. Our results have pharmacogenetic implications for 5-HT3 therapeutic antagonists such as ondansetron (Enoch et al, 2011). 2. Genetic variation within HTR3B predicts alcoholism with comorbid antisocial personality disorder in Finns and resting EEG power in Plains Indians and US Caucasians (Ducci et al, 2009a). Studies on gene targets from animal models: Beginning with gene targets identified in model organisms, several functional loci or haplotypes were identified and replicated by association in humans. In a rat experimental pain model involving sciatic neurotomy, our collaborator Clifford Wolf used mRNA expression arrays to identify that phantom limb pain in these animals was accompanied by large increases in expression of GTP Cyclohydrolase Gch1 and other biopterin-biosynthetic enzymes, in the dorsal root ganglion on the ipsilateral side of the nerve lesion. Inhibition of GCH1 prevented a large increase in biopterin levels in DRG occurring post-neurotomy, and prevented development of enhanced experimental pain. LNG has studied pain models because of the importance of affective response in determining pain response, and the robust effects of genes such as COMT and NPY that affect emotionality or pain response. The rat Gch1/pain finding was translated to humans at LNG, in collaboration with Guest Researchers Mitchell Max and Inna Belfer, via association studies in 162 post-diskectomy patients and 547 normal controls studied for experimental pain threshold, and by functional studies of GCH1. A GCH1 haplotype leading to lower GCH1 expression was associated with enhanced resilience to pain, consistent with the rat model (Tegeder et al, 2006). Gene targets on a 1536 SNP “Addictions Array: To study candidate genes primarily implicated via behavioral pharmacology studies in animal models LNG designed a genotyping array (Hodgkinson et al, 2008). The array also contains ancestry informative markers (AIMs) used to compute ethnic factor scores and closely approximating results previously observed with other AIMs panels applied to the same reference populations (for example Rosenberg et al, 2002), and as illustrated in Figure 5.

109
Figure 5: Ethnic factor scores for 1051 individuals in 51 CEPH reference populations using 186 AIMs from the Addictions Array selected from HapMap as described. 7-factor solution computed with Structure 2.0 (Pritchard) by Pei-Hong Shen. The Addictions Array has been genotyped across >20,000 unique DNAs, including the CEPH reference populations. LNG has provided genotypes, ancestry factor scores, and in some cases diplotypes determined in the context of larger reference populations, and contributing to several published research reports (e.g. Zhou et al, 2008; Bevilacqua et al, 2010) in which the ancestry scores were used to detect or control for ethnic stratification. Further, we were able to use the ancestry scores to show that high rates of heroin and cocaine dependence in African Americans are not due to African ancestry, but are secondary to socioeconomic factors (Ducci et al, 2009b). Identification of functional loci and validation in model organisms via deep sequencing GWAS has not accounted for most of the genetic variance of common complex diseases, encouraging deep sequencing, or more descriptively massively parallel sequencing, to detect rare and uncommon functional variants that would be missed by GWAS (reviewed by Biesecker, 2010; 2012). Deep sequencing had been successfully applied to rare Mendelian diseases for which families were available. We have applied several ideas that have been championed by Leal, including sequencing of extreme individuals (Liu and Leal, 2012) and aggregation of putatively functional alleles to generate gene-based signals Liu and Leal, 2010). In addition, we used model organisms and founder populations, as will be illustrated in the two studies that conclude this report. In human studies, we used founder populations, namely Finns and American Indians, in whom disease genetic heterogeneity is reduced. Reviewing Finnish disease heritage, Peltonen et al (1999) noted some 35 diseases in which Finns had one predominant allele, and in contrast disease origins in other European populations included multiple alleles at multiple genes. Peltonen (Kristiansson et al, 2008) and others, including us, have expanded the argument for the use of population isolates for complex disease identification, where the advantages are more salient. LNG adopted the isolate strategy at the inception of this lab in 1992. Impulsivity: Locus detection from MAOA to HTR2B Impulsivity, describing action without foresight, is a precursor and mediating factor in several psychiatric diseases including addictions and personality disorders. Impulsivity is a modifier of other psychiatric diseases, for example contributing to suicidality. Functional alleles modulate impulsivity, as shown by the heritability of impulsivity and associated disorders. Alleles mediating risk for impulsivity may be both rare and common, and found at the same gene as will
Africa Middle East
Asia Oceania Europe America

110
be discussed for MAOA, which in several ways represents the precursor to our discovery of an HTR2B stop codon, but which also allows important contrasts. MAOA has a functional VNTR locus that influences impulsivity. The effects appear context dependent and also provide context for other behavioral determinants. In this regard we found that MAOA low expression genotypes are permissive for aggression enhancing effects of testosterone (Sjoberg et al, 2008). Stronger effects of the MAOA VNTR on brain neurotransmitter receptor density (Mickey et al, 2008) were observed. Also in line with the more complex or weak effects of such polymorphisms on the behavior, the MAOA VNTR’s effects appear to be strongly altered by interaction with stress exposure (Ducci et al, 2007a, Enoch et al, 2010b). Discovery of the MAOA VNTR was preceded by the discovery of a stop codon that led to impulsivity in one Dutch family and produced neurochemical changes that would be rationally expected (Brunner et al, 1993). This MAOA stop codon was genotyped by us and others but not observed outside this one family. In contrast to the family-genetic analysis performed by Brunner, we initially sequenced extreme, unrelated cases and controls from a founder population and followed up results in both populations and families. The sequencing was performed on 14 serotonin and dopamine genes in pools representing equal numbers of extreme cases of antisocial behavior (ASPD) and controls. Sequencing of the 14 candidate genes in Finnish probands and controls led to detection of a large panel of missense variants amongst which were four probably damaging variants, and these were found in significant excess in cases (Bevilacqua et al, 2010). The excess was due to Q20*, a stop codon in HTR2B that on an individual basis was significant in case/control and familial co-transmission association with ASPD/Alcoholism, with moderate p values on the order of <10-2. Here we would note the convergence of impulsivity and ASPD with alcoholism and alcohol use. The probands we sequenced and analyzed for genetic association had been convicted of criminal offenses, many for impulsive crimes. Most had diagnoses of ASPD, and in almost all cases met criteria for DSM-III-R alcohol dependence. Thus in this population the impulsive ASPD individuals represent a phenotypically extreme type of alcoholic. Furthermore, the carriers of *20 who had committed violent crimes had almost always done so while inebriated, in response to some minor irritation, and seemingly for no gain. As shown in Figure 6, *20 led to variable nonsense-mediated decay of the HTR2B mRNA and complete blockade of receptor expression, these experiments taking advantage of HTR2B expression in transformed B lymphocytes. The stop codon was necessary for severe impulsivity displayed by some individuals in our sample of violent offenders, but not sufficient. Other important factors were male sex and inebriation, as mentioned. It did not lead to global cognitive deficits. *20 is apparently population-specific but common in Finns, with an allele frequency of 0.010 to 0.015 in the general population of Finland (as later confirmed by genotyping >20,000 Finns), and a frequency of 0.037 in the 228 Finnish cases we genotyped. A survey of other populations including the Human Diversity Panel of 51 populations (N =1064), and borderline personality disorder cases collected by Emil Coccarro (University of Chicago) yielded one additional carrier of *20. This was a female alcoholic of Finnish ancestry. Ancestry analyses confirmed that the stop codon was found in Finns of Finnish ancestry but that it was not associated with ASPD because of its association with Finnish ancestry
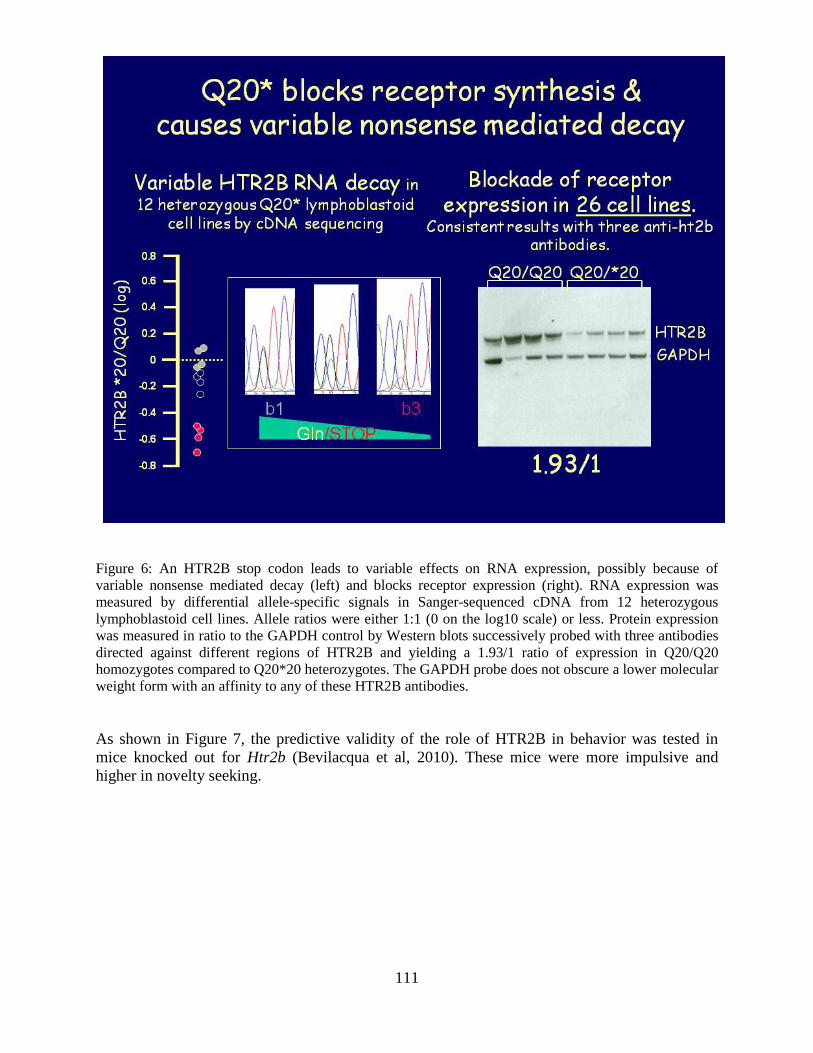
111
Figure 6: An HTR2B stop codon leads to variable effects on RNA expression, possibly because of variable nonsense mediated decay (left) and blocks receptor expression (right). RNA expression was measured by differential allele-specific signals in Sanger-sequenced cDNA from 12 heterozygous lymphoblastoid cell lines. Allele ratios were either 1:1 (0 on the log10 scale) or less. Protein expression was measured in ratio to the GAPDH control by Western blots successively probed with three antibodies directed against different regions of HTR2B and yielding a 1.93/1 ratio of expression in Q20/Q20 homozygotes compared to Q20*20 heterozygotes. The GAPDH probe does not obscure a lower molecular weight form with an affinity to any of these HTR2B antibodies. As shown in Figure 7, the predictive validity of the role of HTR2B in behavior was tested in mice knocked out for Htr2b (Bevilacqua et al, 2010). These mice were more impulsive and higher in novelty seeking.
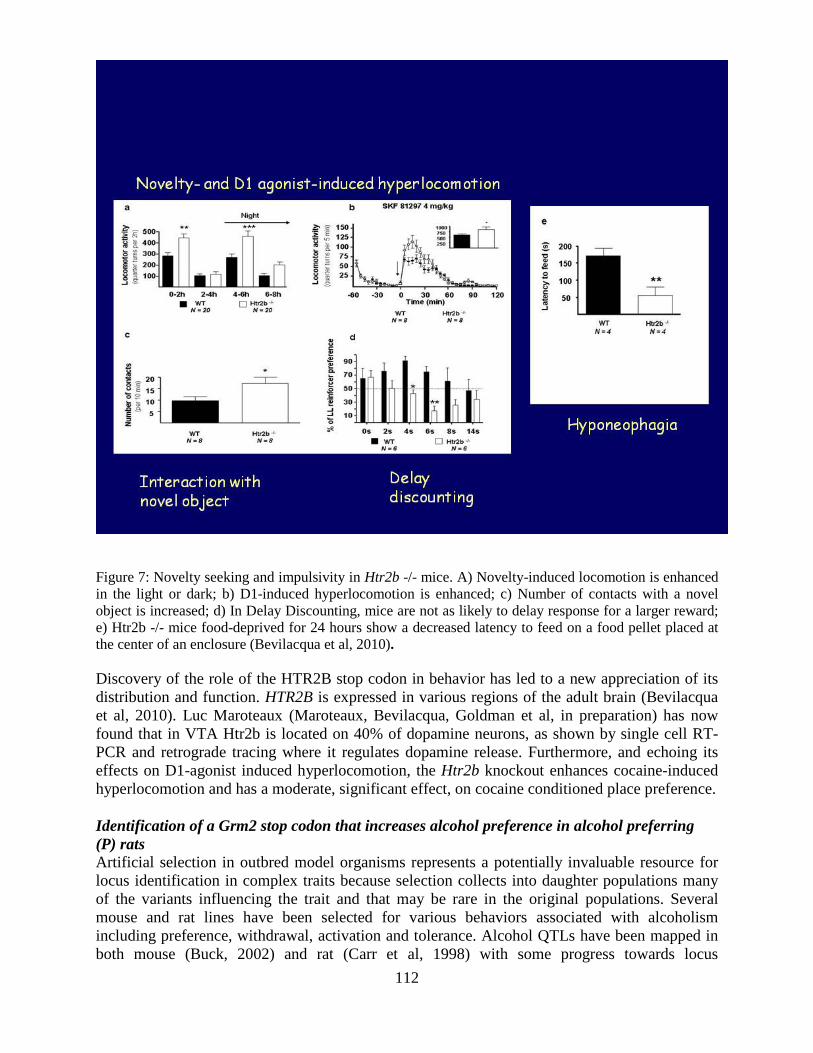
112
Figure 7: Novelty seeking and impulsivity in Htr2b -/- mice. A) Novelty-induced locomotion is enhanced in the light or dark; b) D1-induced hyperlocomotion is enhanced; c) Number of contacts with a novel object is increased; d) In Delay Discounting, mice are not as likely to delay response for a larger reward; e) Htr2b -/- mice food-deprived for 24 hours show a decreased latency to feed on a food pellet placed at the center of an enclosure (Bevilacqua et al, 2010). Discovery of the role of the HTR2B stop codon in behavior has led to a new appreciation of its distribution and function. HTR2B is expressed in various regions of the adult brain (Bevilacqua et al, 2010). Luc Maroteaux (Maroteaux, Bevilacqua, Goldman et al, in preparation) has now found that in VTA Htr2b is located on 40% of dopamine neurons, as shown by single cell RT-PCR and retrograde tracing where it regulates dopamine release. Furthermore, and echoing its effects on D1-agonist induced hyperlocomotion, the Htr2b knockout enhances cocaine-induced hyperlocomotion and has a moderate, significant effect, on cocaine conditioned place preference. Identification of a Grm2 stop codon that increases alcohol preference in alcohol preferring (P) rats Artificial selection in outbred model organisms represents a potentially invaluable resource for locus identification in complex traits because selection collects into daughter populations many of the variants influencing the trait and that may be rare in the original populations. Several mouse and rat lines have been selected for various behaviors associated with alcoholism including preference, withdrawal, activation and tolerance. Alcohol QTLs have been mapped in both mouse (Buck, 2002) and rat (Carr et al, 1998) with some progress towards locus

113
identification in both species, for example Mpdz, at a chromosome 4 location for alcohol withdrawal in the mouse (Fehr et al, 2002). The alcohol preferring P rat has been regarded as a model for human alcoholism because of its excessive voluntary consumption producing sustained high blood alcohol concentrations, drug consumption for CNS effects, signs of intoxication, and relapse-like behavior (Li et al, 1993). Expression and genetic studies implicating Npy and alpha synuclein in the P rat have led to cross-species studies in humans (Foroud et al, 2007). A series of mouse lines for alcohol-related traits has also been produced through selection. However, the identities of the functional variants accounting for the mouse and rat alcohol QTL’s have remained obscure despite the substantial progress in mapping chromosome locations and progress in the neurobiology of alcohol preference, including the role of glutamate. Following on from earlier RNA-Seq and ChIP-Seq studies in postmortem hippocampus from human alcoholics, cocaine addicts and controls (Zhou et al, 2011, Enoch et al, 2012) and the hippocampus of rhesus macaque (Yuan et al, 2012), we used genomic sequencing combined with functional genetic analyses including linkage analysis to identify loci modulating alcohol preference in the P/NP rat model (Zhou et al, submitted). The variant detection was accomplished from RNA-Seq and ChIP-Seq data obtained from total hippocampus of alcohol-naïve animals, subsequently followed by exome sequencing for greater completeness. Sequencing of 8 P and 8 NP rats was performed on genomic and gene-centric levels using an Illumina GA2X sequencer. Bioinformatic analysis predicted damaging variations genetically fixed in P and NP rats. Function was confirmed by measurement of RNA and protein (the latter via Western blots with an mGluR2-specific antibody), and by linkage in iP x iNP F2s made by cross-mating inbred P (iP) and NP (iNP) strains. We identified 19,129 SNPs that were homozygous and that segregated between P and NP rats. Subsequently we sequenced exomes of six unrelated P rats, six NP rats and four Wistar rats and constructed a genome map highlighting differences between P and NP rats (Figure 8). The map reveals regions that were randomly fixed by inbreeding to differentiate these two lines, as well as a clear excess of large fixed block regions in opposite allelic configuration between P and NP that are more likely to harbor loci responsive to the artificial selection and that therefore became more rapidly fixed.

114
-----------------------------------------------------------------------------------------------------------------------
Figure 8: A genomic map of blocks of segregating variants that differentiate the P and NP rat, based on exome sequencing of unrelated animals (6 P rats, 6 NP rats and 4 Wistar rats). Indicated are locations of severely damaging variants, 11/13 being located in large blocks. As shown in larger size at the bottom, the Grm2 and Lcn2 stop codons that were linked to alcohol preference in iP x iNP F2 mice are located in large segregating blocks on chromosomes 8 and 3, respectively. Among 235 segregating missense variants, 22 were predicted to be damaging. A stop codon at codon 407 of the metabotropic glutamate receptor 2 (Grm2) was homozygous in P rats. This stop codon was also found at lower frequency in outbred Wistar rats, the parental strain of P and NP. Loss of mGluR2 protein expression in P rats was complete (Figure 7). The Grm2 stop codon led

115
to a 32% increase in alcohol consumption and a 28% increase in preference in F2 progeny from an intercross of inbred P (iP) and iNP lines, indicating that it is unlikely that this stop codon represents a randomly fixed genetic difference, but that it or a locus nearby alters alcohol preference. P rats were also homozygous for a nonsense variant at codon 137 of lipocalin 2 (Lcn2). This stop codon was also linked to higher alcohol consumption. Both stop codons mapped to large blocks, on chr 8 and chr 3, fixed in opposite allelic configuration in P and NP (Figure 8). The size of the blocks harboring Lcn2 and Grm2 and the presence of nearby fixed blocks may indicate that extended haplotypes in these genomic regions were selected to high opposing frequency and subsequently large pieces within them were fixed relatively early by the artificial selection. We hypothesize that loci such as Grm2 and Lcn2 located in these blocks drove response to selection.
2601601108060
4050
30
20
15
10
NP1NP2 P1 P2
kDa
mGluR2
GAPDH
Figure 9: The Grm2 protein is expressed in brain of NP rats (Lanes 1 and 2) but undetectable in P rats (lanes 3 and 4), by Western blot. As mentioned, the Grm2 stop codon is functional, blocking expression of the mGluR2 receptor protein (Figure 9). To evaluate the global effects of the Grm2 stop codon, we analyzed the hippocampal RNA-Seq profiles from P and NP rats, subjecting these data to gene ontology analysis. Global evaluation of RNA expression revealed strong perturbations of glutamate function in P rats. We propose that amongst the alterations in gene expression between P and NP rats (Kimpel et al, 2007) the Grm2 stop codon is an important origin of glutamate related differences. In humans, alcohol preference and alcoholism are multifactorial. We targeted trait anxiety as a phenotype for GRM2 association studies because of increased anxiety observed in murine glutamate genetic models, the higher anxiety of P rats and the association of trait anxiety with alcoholism in different populations of alcoholics with and without ASPD (Ducci et al, 2007b). We found that GRM2 was associated with dimensional anxiety (Harm Avoidance) in two independent populations. To investigate the possible role of GRM2 in human alcohol consumption, alcoholism, and anxiety, 5300 Finns and 320 Plains Indians were genotyped on the Illumina 550k platform. In both Finns and Plains Indians, the same two GRM2 SNPs were associated with Harm Avoidance (p<0.002), but not alcohol consumption or alcoholism. Survey of 1000 Genomes data and several hundred individuals on whom we have performed targeted
NP P
Log2 Expression Level P = 0.0001
FDR = 0.0145
a
b
c
d
NP
P
SeqRe
ad Cou
nt
407 C/C 407 C/* 407 */*
Alcoho
l Con
sumption
(247) (87) (46)0
1
2
3
4
5
407 C/C 407 C/* 407 */*
Alcoho
l Preference
(247) (87) (46)0
0.10.20.30.40.50.60.7
e
f
c.1221C>Ap.Cys 407 *
Ligand Binding Trans-‐membrane
1 872
*111,837,086 111,850,133111,844,799Chr8
Grm2
mGluR2
Grm
2 *407A
llele Freq
WistarRats
P Rats(Gen 30)
P Rats(Gen 70)
0
0.2
0.4
0.6
0.8
1

116
resequencing at GRM2 did not yield stop codons, so presumably functional alleles at the human GRM2 gene are rare, or more subtle, in their action. References
Anton RF, Oroszi G, O'Malley S, Couper D, Swift R, Pettinati H, Goldman D. 2008. An evaluation of mu-opioid receptor (OPRM1) as a predictor of naltrexone response in the treatment of alcohol dependence: results from the Combined Pharmacotherapies and Behavioral Interventions for Alcohol Dependence (COMBINE) study. Arch Gen Psychiat 65:135-44.
Bevilacqua L, Doly S, Kaprio J, Yuan Q, Tikkanen R, Paunio T, Zhou Z, Wedenoja J, Maroteaux L, Diaz S, Belmer A, Hodgkinson CA, Dell'osso L, Suvisaari J, Coccaro E, Rose RJ, Peltonen L, Virkkunen M, Goldman D. 2010. A population-specific HTR2B stop codon predisposes to severe impulsivity. Nature 468(7327):1061-6.
Biesecker LG. 2010. Exome sequencing makes medical genomics a reality. Nat Genet 42:13-4.Biesecker LG. 2012. Opportunities and challenges for the integration of massively parallel genomic sequencing into clinical practice: lessons from the ClinSeq project. Genet Med 14:393-8.
Brooks PJ, Enoch MA, Goldman D, Li TK, Yokoyama A. 2009. The alcohol flushing response: an unrecognized risk factor for esophageal cancer from alcohol consumption. PLoS Med 6(3):e50.
Brooks PJ, Goldman D, Li TK. 2009. Alleles of alcohol and acetaldehyde metabolism genes modulate susceptibility to oesophageal cancer from alcohol consumption. Hum Genomics 3(2):103-5.
Brunner HG, Nelen M, Breakefield XO, Ropers HH, van Oost BA. 1993. Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science 262:578-80.
Buck KJ, Rademacher BS, Metten P, Crabbe JC. 2002. Mapping murine loci for physical dependence on ethanol. Psychopharmacology (Berl) 160:398-407.
Carr LG, Foroud T, Bice P, Gobbett T, Ivashina J, Edenberg H, Lumeng L, Li TK. 1998. A quantitative trait locus for alcohol consumption in selectively bred rat lines. Alcohol Clin Exp Res 22:884-7.
Ducci F, Enoch M-A, Hodgkinson C, Xu K, Catena M, Robin RW, Goldman D. 2007a. Interaction between a functional MAOA locus and childhood sexual abuse predicts alcoholism and antisocial personality disorder in adult women. Mol Psychiatry 13:334-7.
Ducci F, Enoch MA, Funt S, Virkkunen M, Albaugh B, Goldman D. 2007b. Increased anxiety and other similarities in temperament of alcoholics with and without antisocial personality disorder across three diverse populations. Alcohol 41:3-12.

117
Ducci F, Enoch MA, Yuan Q, Shen PH, White K, Hodgkinson C, Albaugh B, Virkkunen M, Goldman D. 2009a. Genetic variation within HTR3B predicts alcoholism with comorbid antisocial personality disorder and resting EEG power. Alcohol 43:73-84.
Ducci F, Roy A, Shen PH, Yuan Q, Yuan NP, Hodgkinson CA, Goldman LR, Goldman D. 2009b. Association of substance use disorders with childhood trauma but not African genetic heritage in an African American cohort. Am J Psychiatry 166(9):1031-40.
Enoch MA, Shen PH, Ducci F, Yuan Q, Liu J, White KV, Albaugh B, Hodgkinson CA, Goldman D. 2008. Common genetic origins for EEG, alcoholism and anxiety: the role of CRH-BP. PLoS ONE 3:e3620.
Enoch M-A, Hodgkinson CA, Yuan Q, Albaugh B, Virkkunen M, Goldman D. 2009. GABRG1 and GABRA2 haplotypes as independent predictors for alcoholism in two populations. Neuropsychopharmacology 34:1245-54.
Enoch M-A, Hodgkinson CA, Yuan Q, Shen P-H, Goldman D, Roy A. 2010a. The Influence of GABRA2, Childhood Trauma and their Interaction on Alcohol, Heroin and Cocaine Dependence. Biol Psychiatry 67(1):20-27.
Enoch M-A, Steer CD, Newman TK, Gibson N, Goldman D. 2010b. The Impact of Early Life Stress, MAOA, and Gene-Environment Interaction on Behavioral Disinhibition in Pre-Pubertal Children. Genes Brain Behav 9:65-74.
Fehr C, Shirley RL, Belknap JK, Crabbe JC, Buck KJ. 2002. Congenic mapping of alcohol and pentobarbital withdrawal liability loci to a <1 centimorgan interval of murine chromosome 4: identification of Mpdz as a candidate gene. J Neurosci 22:3730-8.
Foroud T, Wetherill LF, Liang T, Dick DM, Hesselbrock V, Kramer J, Nurnberger J, Schuckit M, Carr L, Porjesz B, Xuei X, Edenberg HJ. 2007. Association of alcohol craving with alpha-synuclein (SNCA). Alcohol Clin Exp Res 31:537-45.
Goldman D, Oroszi G, Ducci F. 2005. The genetics of addictions: uncovering the genes. Nat Genet Rev 6:521-32.
Goldman D, Ducci F. 2007. Deconstruction of vulnerability to complex diseases: enhanced effect sizes and power of intermediate phenotypes. ScientificWorldJournal. 7:124-30. Review.
Hodgkinson CA , Xu K, Yuan Q, Shen P-H, Heinz E, Lobos EA, Binder E, Cubells J, Ehlers CL, Gelernter J, Mann J, Riley B, Roy A, Tabakoff B, Todd RD, Zhou Z, Goldman D. 2008. Addictions biology: haplotype based analysis for 130 candidate genes on a single array.Alcohol Alcohol 43: 505-15.
Hodgkinson CA, Enoch MA, Srivastava V, Cummins-Oman JS, Ferrier C, Iarikova P, Sankararaman S, Yamini G, Yuan Q, Zhou Z, Albaugh B, White KV, Shen PH, Goldman D. 2010. Genome-wide association identifies candidate genes that influence the human electroencephalogram. Proc Natl Acad Sci U S A 107(19):8695-700.

118
Hong LE, Hodgkinson CA, Yang Y, Sampath H, Ross TJ, Buchholz B, Salmeron BJ, Srivastava V, Thaker GK, Goldman D, Stein EA. 2010. A genetically modulated, intrinsic cingulate circuit supports human nicotine addiction. Proc Natl Acad Sci U S A 107(30):13509-14.
Kristiansson K, Naukkarinen J, Peltonen L. 2008. Isolated populations and complex disease gene identification. Genome Biol 9(8):109.
Li TK, Lumeng L, Doolittle, DP. 1993. Selective breeding for alcohol preference and associated responses. Behav Genet 23:163-70.
Liu DJ, Leal SM. 2010. A novel adaptive method for the analysis of next-generation sequencing data to detect complex trait associations with rare variants due to gene main effects and interactions. PLoS Genet 6:e1001156.
Liu DJ, Leal SM. 2012. A Unified Framework for Detecting Rare Variant Quantitative Trait Associations in Pedigree and Unrelated Individuals via Sequence Data. Hum Hered 73:105-22.
Mickey BJ, Ducci F, Hodgkinson CA, Langenecker SA, Goldman D, Zubieta JK. 2008. Monoamine Oxidase A Genotype Predicts Human Serotonin 1A Receptor Availability in Vivo. J Neurosci 28:11354-9.
Peltonen L, Jalanko A, Varilo T. 1999. Molecular genetics of the Finnish disease heritage. Hum Mol Genet 8:1913–23.
Rosenberg NA, Pritchard JK, Weber JL, Cann HM, Kidd KK, Zhivotovsky LA, Feldman MW. 2002. Genetic structure of human populations. Science 298:2381-5.
Roy A, Gorodetsky E, Yuan Q, Goldman D, Enoch M-A. 2010. The Interaction of FKBP5, a Stress Related Gene, with Childhood Trauma Increases the Risk for Attempting Suicide. Neuropsychopharmacology 35:1674-83.
Sjöberg RL, Ducci F, Barr CS, Newman TK, Dell'osso L, Virkkunen M, Goldman D. 2008. A non-additive interaction of a functional MAO-A VNTR and testosterone predicts antisocial behavior. Neuropsychopharmacology 33:425-30.
Srivastava V, Buzas B, Momenan R, Oroszi G, Pulay AJ, Enoch MA, Hommer DW, Goldman D. 2010. Association of SOD2, a mitochondrial antioxidant enzyme, with gray matter volume shrinkage in alcoholics. Neuropsychopharmacology 35(5):1120-8.
Tegeder I, Costigan M, Griffin RS, Abele A, Belfer I, Schmidt H, Ehnert C, Nejim J, Marian C, Scholz J, Wu T, Allchorne A, Diatchenko L, Binshtok AM, Goldman D, Adolph J, Sama S, Atlas SJ, Carlezon WA, Parsegian A, Lötsch J, Fillingim RB, Maixner W, Geisslinger G, Max MB, Woolf CJ. 2006. GTP cyclohydrolase and tetrahydrobiopterin regulate pain sensitivity and persistence. Nature Medicine 12(11):1269-77.

119
Zhou Z, Zhu G, Hariri AR, Enoch MA, Scott D, Sinha R, Virkkunen M, Mash DC, Lipsky RH, Hu XZ, Hodgkinson CA, Xu K, Buzas B, Yuan Q, Shen PH, Ferrell RE, Manuck SB, Brown SM, Hauger RL, Stohler CS, Zubieta JK, Goldman D. 2008. Genetic variation in human NPY expression affects stress response and emotion. Nature 452:997-1001.
Publications 2011-2012
1. Abraham J, Brooks PJ. 2011. Divergent effects of oxidatively induced modification to the C8 of 2'-deoxyadenosine on transcription factor binding: 8,5'(S)-cyclo-2'-deoxyadenosine inhibits the binding of multiple sequence specific transcription factors, while 8-oxo-2'-deoxyadenosine increases binding of CREB and NF-kappa B to DNA. Environ Mol Mutagen 52:287-95.
2. Abraham J, Balbo S, Crabb D, Brooks PJ. 2011. Alcohol metabolism in human cells
causes DNA damage and activates the Fanconi anemia-breast cancer susceptibility (FA-BRCA) DNA damage response network. Alcohol Clin Exp Res 35:2113-20.
3. Barnett JH, Xu K, Heron J, Goldman D, Jones PB. 2011. Cognitive effects of genetic
variation in monoamine neurotransmitter systems: a population-based study of COMT, MAOA, and 5HTTLPR. Am J Med Genet B Neuropsychiatr Genet 156(2):158-67.
4. Bevilacqua L, Carli V, Sarchiapone M, George DK, Goldman D, Roy A, Enoch MA.
2012. Interaction between FKBP5 and childhood trauma and risk of aggressive behavior Arch Gen Psychiatry 69(1):62-70.
5. Bevilacqua L, Goldman D. 2011. Genetics of emotion. Trends Cogn Sci 15(9):401-8.
6. Goldman D. 2011. Molecular etiologies of schizophrenia: are we almost there yet? Am J
Psychiatry 2 168:879-81.
7. Bletzer KV, Yuan NP, Koss MP, Polacca M, Eaves ER, Goldman D. 2011.Taking humor seriously: talking about drinking in Native American focus groups. Med Anthropol 30(3):295-318.
8. Burghardt PR, Love TM, Stohler CS, Hodgkinson C, Shen PH, Enoch MA, Goldman D, Zubieta JK. 2012. Leptin regulates dopamine responses to sustained stress in humans. J Neurosci 32(44):15369-76.
9. Cirulli F, Reif A, Herterich S, Lesch KP, Berry A, Francia N, Aloe L, Barr CS, Suomi SJ, Alleva E. 2011. A novel BDNF polymorphism affects plasma protein levels in interaction with early adversity in rhesus macaques. Psychoneuroendocrinology 36(3):372-9.
10. Copeland WE, Sun H, Costello EJ, Angold A, Heilig MA, Barr CS. 2011. Child μ-opioid receptor gene variant influences parent-child relations. Neuropsychopharmacology 36(6):1165-70.

120
11. Deo AJ, Huang YY, Hodgkinson CA, Xin Y, Oquendo MA, Dwork AJ, Arango V, Brent
DA, Goldman D, Mann JJ, Haghighi F. 2012. A large-scale candidate gene analysis of mood disorders: evidence of neurotrophic tyrosine kinase receptor and opioid receptor signaling dysfunction. Psychiatr Genet 2012 Dec 30 [epub ahead of print].
12. Dlugos AM, Hamidovic A, Hodgkinson C, Shen PH, Goldman D, Palmer AA, de Wit H. 2011. OPRM1 gene variants modulate amphetamine-induced euphoria in humans. Genes Brain Behav 10:199-209.
13. Ducci F, Goldman D. 2012. The genetic basis of addictive disorders. Psychiatr Clin
North Am 35:495-519.
14. Enoch MA. 2011. The role of early life stress as a predictor for alcohol and drug dependence. Psychopharmacology (Berl) 214(1):17-31.
15. Enoch MA, Zhou Z, Kimura M, Mash DC, Yuan Q, Goldman D. 2012. GABAergic gene
expression in postmortem hippocampus from alcoholics and cocaine addicts; corresponding findings in alcohol-naïve P and NP rats. PLoS One 7(1):e29369.
16. Enoch MA. 2012. The influence of gene-environment interactions on the development of
alcoholism and drug dependence. Curr Psychiatry Rep 14(2):150-8.
17. Enoch MA, Gorodetsky E, Hodgkinson C, Roy A, Goldman D. 2011. Functional genetic variants that increase synaptic serotonin and 5-HT3 receptor sensitivity predict alcohol and drug dependence. Mol Psychiatry 16:1139-46.
18. Forbes CE, Poore JC, Barbey AK, Krueger F, Solomon J, Lipsky RH, Hodgkinson CA,
Goldman D, Grafman J. 2011. Cereb Cortex. BDNF Polymorphism-Dependent OFC and DLPFC Plasticity Differentially Moderates Implicit and Explicit Bias. Cereb Cortex 22(11):2602-9.
19. Goldman D. 2012. Our Genes, Our Choices: How Genotype and Gene Interactions Affect
Behavior. Elsevier, Academic Press.
20. Goldman D, Srivastava V. 2012. Genetics and Neuroimaging, in Adinoff and Stein (Eds) Neuroimaging in the Addictions, Wiley-Blackwell, Addiction Press.
21. Heilig M, Goldman D, Berrettini W, O'Brien CP. 2011. Pharmacogenetic approaches to
the treatment of alcohol addiction. Nat Rev Neurosci 12:670-84.
22. Higham JP, Barr CS, Hoffman CL, Mandalaywala TM, Parker KY, Maestrapieri D.
2011. Mu-opioid receptor (OPRM1) variation, oxytocin levels and maternal attachment in free-ranging rhesus macaques Macaca mulatta. Behav Neurosci 125(2):131-6.
23. Hong E, Yang X, Wonodi I, Hodgkinson CA, Goldman D, Stine OC, Stein EA, Thaker
G. 2011. A CHRNA5 allele related to nicotine addiction and schizophrenia. Genes Brain Behav 10: 530-535.

121
24. Horibata K, Saijo M, Bay MN, Lan L, Kuraoka I, Brooks PJ, Honma M, Nohmi T, Yasui
A, Tanaka K. 2011. Mutant Cockayne syndrome group B protein inhibits repair of DNA topoisomerase I-DNA covalent complex. Genes Cells 16:101-14.
25. Hsu DT, Mickey BJ, Langenecker SA, Heitzeg MM, Love TM, Wang H, Kennedy SE,
Peciña M, Shafir T, Hodgkinson CA, Enoch MA, Goldman D, Zubieta JK. 2012. Variation in the corticotropin-releasing hormone receptor 1 (CRHR1) gene influences fMRI signal responses during emotional stimulus processing. J Neurosci 32:3253-60.
26. Jonassaint CR, Szatkiewicz JP, Bulik CM, Thornton LM, Bloss C, Berrettini WH, Kaye
WH, Bergen AW, Magistretti P, Strober M, Keel PK, Brandt H, Crawford S, Crow S, Fichter MM, Goldman D, Halmi KA, Johnson C, Kaplan AS, Klump KL, La Via M, Mitchell JE, Rotondo A, Treasure J, Woodside DB. 2011. Absence of association between specific common variants of the obesity-related FTO gene and psychological and behavioral eating disorder phenotypes. Am J Med Genet B Neuropsychiatr Genet 156B(4):454-61.
27. Kano S, Colantuoni C, Han F, Zhou Z, Yuan Q, Wilson A, Takayanagi Y, Lee Y, Rapoport J, Eaton W, Cascella N, Ji H, Goldman D, Sawa A. Genome-wide profiling of multiple histone methylations in olfactory cells: further implications for cellular susceptibility to oxidative stress in schizophrenia. Mol Psychiatry 2012 Aug 28. [Epub ahead of print]
28. Krueger F, Pardini M, Huey E D, Raymont V, Solomon J, Lipsky R H, Hodgkinson CA, Goldman D, Grafman J. 2011. The Role of the Met66 Brain-Derived Neurotrophic Factor Allele in the Recovery of Executive Function after Combat-Related Traumatic Brain Injury. J Neuroscience 31:598-606.
29. Lee MR, Gallen CL, Zhang X, Hodgkinson CA, Goldman D, Stein EA, Barr CS. 2011.
Functional Polymorphism of the Mu-Opioid Receptor Gene (OPRM1) Influences Reinforcement Learning in Humans. PLoS One 6(9):e24203.
30. Lindell SG, Yuan Q, Zhou Z, Goldman D, Thompson RC, Lopez JF, Suomi SJ, Higley JD, Barr CS. 2012. The serotonin transporter gene is a substrate for age and stress dependent epigenetic regulation in rhesus macaque brain: potential roles in genetic selection and gene × environment interactions. Dev Psychopathol 24(4):1391-400.
31. Liu J, Zhou Z, Hodgkinson CA, Yuan Q, Shen PH, Mulligan CJ, Wang A, Gray RR, Roy A, Virkkunen M, Goldman D, Enoch MA. 2011. Haplotype-based study of the association of alcohol-metabolizing genes with alcohol dependence in four independent populations. Alcohol Clin Exp Res 35:304-16.
32. Love TM, Enoch MA, Hodgkinson CA, Pecina M, Mickey B, Koeppe RA, Stohler CS,
Goldman D, Zubieta JK. 2012. Oxytocin Gene Polymorphisms Influence Human Dopaminergic Function in a Sex-Dependent Manner. Biol Psychiatry 72:198-206.

122
33. Marsh AA, Crowe SL, Yu HH, Gorodetsky EK, Goldman D, Blair RJ. 2011. Serotonin transporter genotype (5-HTTLPR) predicts utilitarian moral judgments. PLoS One 6(10):e25148.
34. Marsh AA, Yu HH, Pine DS, Gorodetsky EK, Goldman D, Blair RJ. 2012. The influence of oxytocin administration on responses to infant faces and potential moderation by OXTR genotype.Psychopharmacology (Berl.) 224(4):469-76.
35. Mickey BJ, Zhou Z, Heitzeg MM, Heinz E, Hodgkinson CA, Hsu DT, Langenecker SA, Love TM, Pecina M, Shafir T, Stohler CS, Goldman D, Zubieta JK. 2011. Emotion processing, major depression, and functional genetic variation of neuropeptide Y. Arch Gen Psychiatry 68:158-66.
36. Mickey BJ, Sanford BJ, Love TM, Shen PH, Hodgkinson CA, Stohler CS, Goldman D, Zubieta JK. 2012. Striatal dopamine release and genetic variation of the serotonin 2C receptor in humans. J Neurosci 32(27):9344-50.
37. Morozova TV, Goldman D, Mackay TF, Anholt RR. 2012. The genetic basis of alcoholism: multiple phenotypes, many genes, complex networks. Genome Biol 13(2):239.
38. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, Godlewski G, Harvey-White J, Mordi I,
Cravatt BF, Lotersztajn S, Gao B, Yuan Q, Schuebel K, Goldman D, Kunos G. 2011. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proc Natl Acad Sci U S A 108:6323-8.
39. Pardini M, Krueger F, Hodgkinson C, Raymont V, Ferrier C, Goldman D, Strenziok M,
Guida S, Grafman J. 2011. Prefrontal cortex lesions and MAO-A modulate aggression in penetrating traumatic brain injury. Neurology 76:1038-45.
40. Pardini M, Krueger F, Koenigs M, Raymont V, Hodgkinson C, Zoubak S, Goldman D, Grafman J. 2012. Fatty-acid amide hydrolase polymorphisms and post-traumatic stress disorder after penetrating brain injury. Transl Psychiatry Jan 31;2:e75.
41. Peciña M, Mickey BJ, Love T, Wang H, Langenecker SA, Hodgkinson C, Shen PH, Villafuerte S, Hsu D, Weisenbach SL, Stohler CS, Goldman D, Zubieta JK. DRD2 polymorphisms modulate reward and emotion processing, dopamine neurotransmission and openness to experience. Cortex 2012 [Epub ahead of print].
42. Ray LA, Barr CS, Blendy JA, Oslin D, Goldman D, Anton RF. 2011. The Role of the
Asn40Asp Polymorphism of the Mu Opioid Receptor Gene (OPRM1) on Alcoholism Etiology and Treatment: A Critical Review. Alcohol Clin Exp Res 36:385-94.
43. Reimold M, Knobel A, Rapp MA, Batra A, Wiedemann K, Ströhle A, Zimmer A,
Schönknecht P, Smolka MN, Weinberger DR, Goldman D, Machulla HJ, Bares R, Heinz A. 2011. Central serotonin transporter levels are associated with stress hormone response and anxiety. Psychopharmacology (Berl) 213:563-72.

123
44. Roiser JP, Levy J, Fromm SJ, Goldman D, Hodgkinson CA, Hasler G, Sahakian BJ, Drevets WC. 2012. Serotonin transporter genotype differentially modulates neural responses to emotional words following tryptophan depletion in patients recovered from depression and healthy volunteers. J Psychopharmacol 26:1434-42.
45. Root TL, Szatkiewicz JP, Jonassaint CR, Thornton LM, Pinheiro AP, Strober M, Bloss C,
Berrettini W, Schork NJ, Kaye WH, Bergen AW, Magistretti P, Brandt H, Crawford S, Crow S, Fichter MM, Goldman D, Halmi KA, Johnson C, Kaplan AS, Keel PK, Klump KL, La Via M, Mitchell JE, Rotondo A, Treasure J, Woodside DB, Bulik CM. 2011. Association of Candidate Genes with Phenotypic Traits Relevant to Anorexia Nervosa. Eur Eat Disord Rev 19:487-93.
46. Rostami E, Krueger F, Zoubak S, Dal Monte O, Raymont V, Pardini M, Hodgkinson CA,
Goldman D, Risling M, Grafman J. 2011. BDNF polymorphism predicts general intelligence after penetrating traumatic brain injury. PLoS One 6(11):e27389.
47. Roy A, Hodgkinson CA, Deluca V, Goldman D, Enoch MA . 2012. Two HPA axis
genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. J Psychiatr Res 46:72-9.
48. Roy A, Roy M, Goldman D. 2011. Childhood trauma and depressive symptoms in type 1
diabetes. J Clin Psychiatry 72:1049-53.
49. Salehi B, Preuss N, van der Veen JW, Shen J, Neumeister A, Drevets WC, Hodgkinson C, Goldman D, Wendland JR, Singleton A, Gibbs JR, Cookson MR, Hasler G. Age-modulated association between prefrontal NAA and the BDNF gene. Int J Neuropsychopharmacol 2012 Dec 20:1-9. [Epub ahead of print]
50. Schneider ML, Moore CF, Barr CS, Larson JA, Kraemer GW. 2011. Moderate prenatal alcohol exposure and serotonin genotype interact to alter CNS serotonin function in rhesus monkey offspring. Alcohol Clin Exp Res 35(5):912-20.
51. Schwandt ML, Lindell SG, Higley JD, Heilig M, Barr CS. 2011. OPRM1 gene variation
influences hypothalamic-pituitary-adrenal axis function in response to a variety of stressors in rhesus macaques. Psychoneuroendocrinology 36(9):1303-11.
52. Spinelli S, Schwandt ML, Lindell SG, Heilig M, Suomi SJ, Higley JD, Goldman D, Barr
CS. 2012. The serotonin transporter gene linked polymorphic region is associated with the behavioral response to repeated stress exposure in infant rhesus macaques. Dev Psychopathol 24(1):157-65.
53. Tikkanen R, Auvinen-Lintunen L, Ducci F, Sjöberg RL, Goldman D, Tiihonen J,
Ojansuu I, Virkkunen M. 2011. Psychopathy, PCL-R, and MAOA genotype as predictors of violent reconvictions. Psychiatry Res 185:382-6.
54. Wang J, Cao H, You C, Yuan B, Bahde R, Gupta S, Nishigori C, Niedernhofer LJ,
Brooks PJ, Wang Y. 2012. Endogenous formation and repair of oxidatively induced G[8-5 m]T intrastrand cross-link lesion. Nucleic Acids Res 40:7368-74.

124
55. Xu K, Hong KA, Zhou Z, Hauger RL, Goldman D, Sinha R. 2012. Genetic modulation of plasma NPY stress response is suppressed in substance abuse: Association with clinical outcomes. Psychoneuroendocrinology 37:554-64.
56. Young-Wolff KC, Enoch MA, Prescott CA. 2011. The influence of gene-environment
interactions on alcohol consumption and alcohol use disorders: a comprehensive review. Clin Psychol Rev 31(5):800-16.
57. Yuan Q, Zhou Z, Lindell SG, Higley JD, Ferguson B, Thompson RC, Lopez JF, Suomi SJ, Baghal B, Baker M, Mash DC, Barr CS, Goldman D. 2012. The rhesus macaque is three times as diverse but more closely equivalent in damaging coding variation as compared to the human. BMC Genet 13:52.
58. Zhou Z, Yuan Q, Mash DC, Goldman D. 2011. Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol. Proc Natl Acad Sci U S A 108:6626-31.
David Goldman: [email protected]

125
______________________________________________________________________
Section of Comparative Behavioral Genomics
______________________________________________________________________ Christina S. Barr, VMD, PhD, Acting Chief Stephen Lindell, MS, Biologist Carlos Driscoll, PhD, Postdoctoral IRTA Maggie Baker, Postbac IRTA Jessica Clemente, Postbac IRTA Kevin Blackistone, Technical IRTA The focus of the Section of Comparative Behavioral Genomics is the examination of intra- and inter-specific variation in genetic and epigenetic effects as they relate to variation in behavioral traits that are known risk factors for alcohol use disorders. Our section partners with both intramural and extramural scientists to obtain samples from a variety of species in order to identify functional genetic variation that can be functionally analyzed and determined to be candidates for predicting individual differences in addiction-related behaviors. We then perform genetic or epigenetic studies in order to home in on the factors that are reliably predictive of functional and behavioral differences. Through the use of samples and behavior datasets that were previously collected through the intramural program, we are able also to perform more refined molecular studies and multi-tiered genotype-phenotype correlations in order to investigate the substrates through which G x E interactions might contribute to risk for alcohol use disorders.
Stephen Lindell, MS, has been with NIAAA since 1993. His graduate work involved identification and characterization of a functional NPY promoter variant that predicts individual differences in stress resilience and alcohol consumption in rhesus macaques. More recently, he has been using next generation sequencing technologies in order to identify genetic variants that predict individual differences in ethanol consumption in rhesus. In addition to continuing his work in primate behavioral genetics, he functions as manager for the Section of Comparative Behavioral Genomics.
Carlos Driscoll, D.Phil. Before joining NIAAA Carlos did work on genetic variation in three population groups of felids – lions, cheetahs and pumas- as they pertained to phylogenetic inference. Later work described the evolutionary relationships among Felis silvestris subspecies, including the domestic cat. Dr. Driscoll is now focused on detecting the causative genetics behind domestic behavior and is working to translate those findings to individual differences in temperament, reward sensitivity, and reactivity in human subjects.

126
The neurobiological systems that influence addiction vulnerability in humans may do so by acting on reward pathways, behavioral dyscontrol, and vulnerability to stress and anxiety. In certain instances, genetic variants that are functionally similar or orthologous to those that moderate risk for human psychiatric disorders are maintained across species, and some of our studies have suggested there to be convergent evolution or allelic variants being driven to or maintained by natural selection in multiple primate species. We have examined such variation in order to model how genetic variation moderates risk in certain environmental contexts (Barr et al, Arch Gen Psychiatry, 2004; Barr et al, PNAS, 2004; Barr et al, PNAS, 2008; Barr et al, PNAS, 2009; Barr et al, Arch Gen Psychiatry, 2008, Lindell et al, Arch Gen Psychiatry, 2010; Schwandt et al, Biol Psychiatry, 2010). We have also been searching for spontaneous genetic variation that occurs across species, including those species that have been subject to intense artificial selection, in order to find signatures for selection on behavior and to “replicate” our findings in primates. Of relevance to the NIAAA mission, these findings reinforce the potential for comparative behavioral genomics studies to demonstrate how the prevalence of human genetic variants that are linked to psychiatric disorders, including the addictions, may be rooted in the fact that such variants contribute to adaptive or favorable traits in the absence of environmental stressors or alcohol. Neurogenetics of Reward Sensitivity- Candidate Gene-Based Studies The reward systems are critical to survival because they are involved in driving ingestion of food, social interactions, and sexual activity. During times when food availability is scarce, these are particularly important because they relate to resource acquisition and control. Of interest, some of the behaviors that can increase likelihood of gaining access to limited resources (i.e., aggression) also involve the reward pathways. One system that is activated in response to both natural and artificial rewards is the endogenous opioid system. In multiple primate species, there are non-synonymous SNPs (Single Nucleotide Polymorphisms) in the first exon of the OPRM1 gene, which produce amino acid changes in the ligand-binding domain of the receptor. Among these are the A118G SNP in humans and the C77G SNP in rhesus macaques. Studies examining intermediate phenotypes likely to be under the control of this receptor (for example, HPA axis activity or alcohol response) suggest gain-of function roles for these polymorphisms (Barr et al, 2007; Schwandt et al, 2011), though opposite effects have been reported at the molecular level. The OPRM1 SNPs present in human and in rhesus macaques not only predict responses to artificial rewards, such as alcohol, but to natural rewards as well. In rhesus, the 77G allele predicts increased expression of attachment of an infant to its mother, and this is particularly true following repeated exposures to periods of maternal separation (Barr et al, 2008). This finding has also been replicated in a study performed using a sample of human children aged 9-13, in
Maggie Baker, BS, 2010 Humboldt State University Biology PhD candidate
Jessica Clemente, BS, 2011 Mercyhurst University Biochemistry Pre-Med
Kevin Blackistone, BA, 2000 University of Maryland- BC VPAV Bioinformatics

127
which various measures of attachment to the caregiver were increased as a function of parental inconsistency or unavailability (Copeland et al, 2011). These studies provide examples how the macaque can be used to both confirm findings derived from human neurogenetic or pharmacogenetic studies and, more importantly, provide a powerful model for informing analyses that should be performed in humans. Moreover, such studies provide a clear demonstration of how genetic factors that influence traits that would normally be “adaptive” can also drive “maladaptive” outcomes in certain environmental contexts (Figure 1).
Figure 1. Genetic Variation at the OPRM1 Gene Could Have Conferred Selective Advantage in Ancestral or Ancient Populations (Adaptive Response?), but Also Increases Risk for Alcohol Problems in Modern Humans (Maladaptive Outcomes?). Macaque infants carrying a functional allele at the OPRM1 gene, which influences reward sensitivity, exhibit increased maternal attachment, especially as a function of repeated maternal separation. They also exhibit increased alcohol preference and consumption, likely through the pathways of novelty seeking and alcohol response.
Broadening of our Search for Functional Variation through Exome and Whole Genome Sequencing We have recently been examining a larger number of species, both primate and non-primate, that have marked differences in behaviors relating to the addictions. In addition to focused sequencing and whole genome sequencing of various animal species (including dog, cat, horse, cow, and rat), we sequenced the exomes of rhesus macaque subjects whom we had chosen based on marked differences in reactivity and temperament. The advantages of sequencing exomes are that DNA can easily be obtained from peripheral tissues (such as blood, hair follicles or skin), that there is a high degree of efficiency, and that there is immense potential for identifying functional variants, since only the protein coding sequences are screened for variation. Moreover, we were able to use a human-based assay in order to perform this study because of the high degree of sequence similarity across species. We identified a number of non-synonymous SNPs (variants that predicted amino acid changes), some of which were predicted to be “damaging” by in silico analysis. Among these were SNPs in the GABARAA6 and CRHR2 genes, which encode the alpha-6 subunit of the GABA-A receptor and the corticotropin releasing hormone receptor (Figure 2). Variation at these genes have been shown to predict individual differences in alcohol and stress response. Other primate samples, including New World Monkey
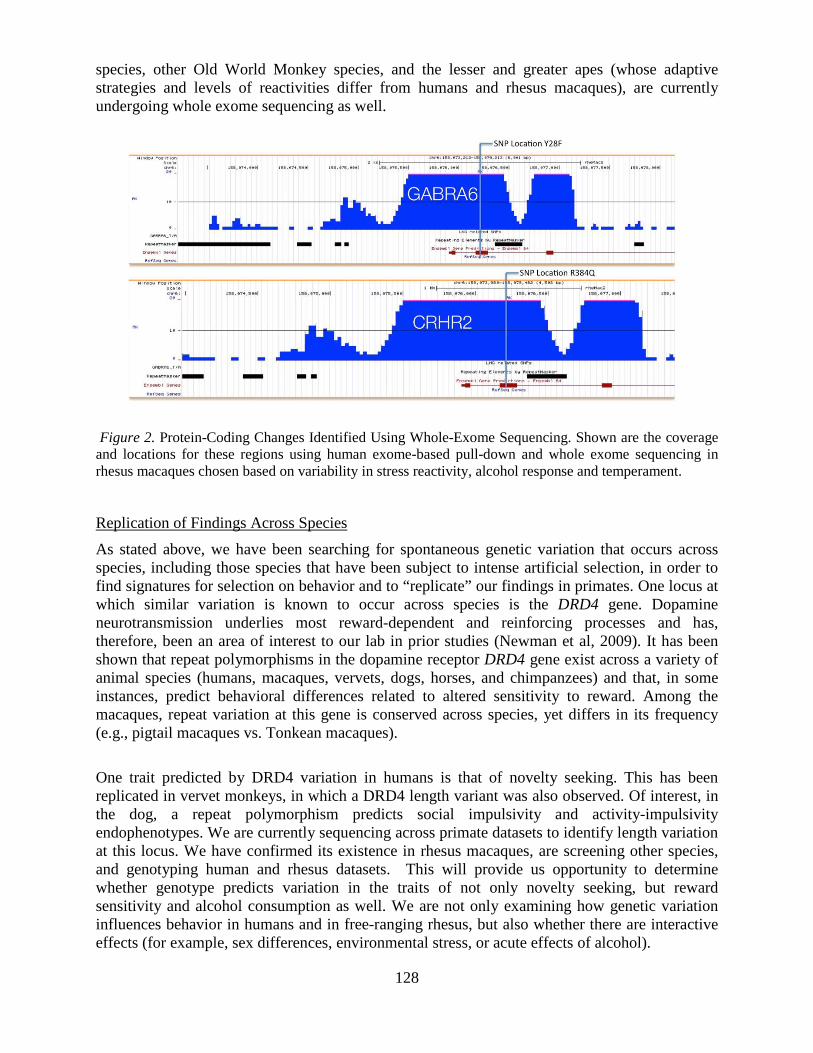
128
species, other Old World Monkey species, and the lesser and greater apes (whose adaptive strategies and levels of reactivities differ from humans and rhesus macaques), are currently undergoing whole exome sequencing as well.
Figure 2. Protein-Coding Changes Identified Using Whole-Exome Sequencing. Shown are the coverage and locations for these regions using human exome-based pull-down and whole exome sequencing in rhesus macaques chosen based on variability in stress reactivity, alcohol response and temperament.
Replication of Findings Across Species As stated above, we have been searching for spontaneous genetic variation that occurs across species, including those species that have been subject to intense artificial selection, in order to find signatures for selection on behavior and to “replicate” our findings in primates. One locus at which similar variation is known to occur across species is the DRD4 gene. Dopamine neurotransmission underlies most reward-dependent and reinforcing processes and has, therefore, been an area of interest to our lab in prior studies (Newman et al, 2009). It has been shown that repeat polymorphisms in the dopamine receptor DRD4 gene exist across a variety of animal species (humans, macaques, vervets, dogs, horses, and chimpanzees) and that, in some instances, predict behavioral differences related to altered sensitivity to reward. Among the macaques, repeat variation at this gene is conserved across species, yet differs in its frequency (e.g., pigtail macaques vs. Tonkean macaques).
One trait predicted by DRD4 variation in humans is that of novelty seeking. This has been replicated in vervet monkeys, in which a DRD4 length variant was also observed. Of interest, in the dog, a repeat polymorphism predicts social impulsivity and activity-impulsivity endophenotypes. We are currently sequencing across primate datasets to identify length variation at this locus. We have confirmed its existence in rhesus macaques, are screening other species, and genotyping human and rhesus datasets. This will provide us opportunity to determine whether genotype predicts variation in the traits of not only novelty seeking, but reward sensitivity and alcohol consumption as well. We are not only examining how genetic variation influences behavior in humans and in free-ranging rhesus, but also whether there are interactive effects (for example, sex differences, environmental stress, or acute effects of alcohol).

129
Neurogenetics of Stress Response and Molecular Mechanisms for G x E Interactions As noted above, rhesus macaques provide opportunity to examine gene x environment (G x E) interactions in a controlled, prospective manner. For a number of years, we have been studying rhesus macaques in order to model how spontaneous genetic variation that influences stress reactivity and/or reward sensitivity, traits that could be adaptive in certain contexts, could also increase risk for alcohol related problems in modern humans (Barr, in press). One of the first variants to be studied in our lab was the serotonin transporter linked polymorphism (HTTLPR), for which functional equivalents exist in humans and macaques. This variant is one that has been shown to interact with repeated, severe life stressors to predict depression, psychopathology and alcohol intake in both human and nonhuman primates (Barr et al, 2004a, b, and c; Schneider et al, 2011; Kraemer et al, 2008; Schneider et al, 2010). In addition to interactive effects with severe or long-term stressors, we recently showed in infant macaques that there are effects of HTTLPR genotype on responses to short-term maternal separation stress, with incremental increases in anxiety-like responses as a function of repeated separation. This suggests that HTTLPR genotype moderates acute responses to stress, and that by promoting sensitization to a stressor, could contribute to risk for behavioral pathology with repeated stress exposure. Our results show that this sensitization process is moderated by genotype from a very early developmental stage (Spinelli et al, in press). One mechanism by which stress could interact with genotype is via epigenetic modifications. The emergence of next generation sequencing technologies has broadened our potential for discovery of epigenetic effects. Using archived brain tissue, we have used these approaches and have been performing ChIP-Seq (Sequencing of chromatin immunopreciptated DNA) in order to examine effects of early stress on epigenetic regulation in tissue derived from stress-sensitive brain regions (hippocampus). One commonly studied epigenetic mark is the histone 3 protein which is tri-methylated at lysine 3 (modified histone, H3K4me3). This is a histone that marks “active” promoters and which would, therefore, be expected to facilitate gene expression. We found there to be interactive effects between a stressful life history and HTTLPR genotype on epigenetic regulation of the serotonin transporter gene in brain (Figure 3). Consistent with this, there were similar interactive effects on central serotonin system function (as assessed by measuring cerebrospinal fluid levels of 5-HIAA), sampled from another cohort of subjects. In addition to the effects of stress, we found that age of sampling is a factor that interacts with genotype and rearing history, both with regard to H3K4me3 binding and CSF 5-HIAA. These data indicate that effects of genotype may vary according to an individual’s life history and environmental exposures. The methodology employed in this study involved antibody-based isolation of H3K4me3, after which the levels of H3K4me3 binding were assessed by sequencing the histone-associated DNA, such that the number of reads for a region are the index for the relative degree of H3K4me3 binding and, therefore, epigenetic regulation via this mechanism. One by-product of using this approach is that genetic variation can also be identified. We identified 11 SNPs within the serotonin transporter gene and its regulatory region, 3 of which were present in the region of H3K4me3 binding, overlapping with a CpG island (a region that can act as a substrate for another, more commonly studied epigenetic regulation, that is, DNA methylation) and within 100 nucleotides of the transcription start site. Of interest, there are 3 SNPs present in the corresponding region in humans as well (Lindell et al, 2012). Though little is known about their
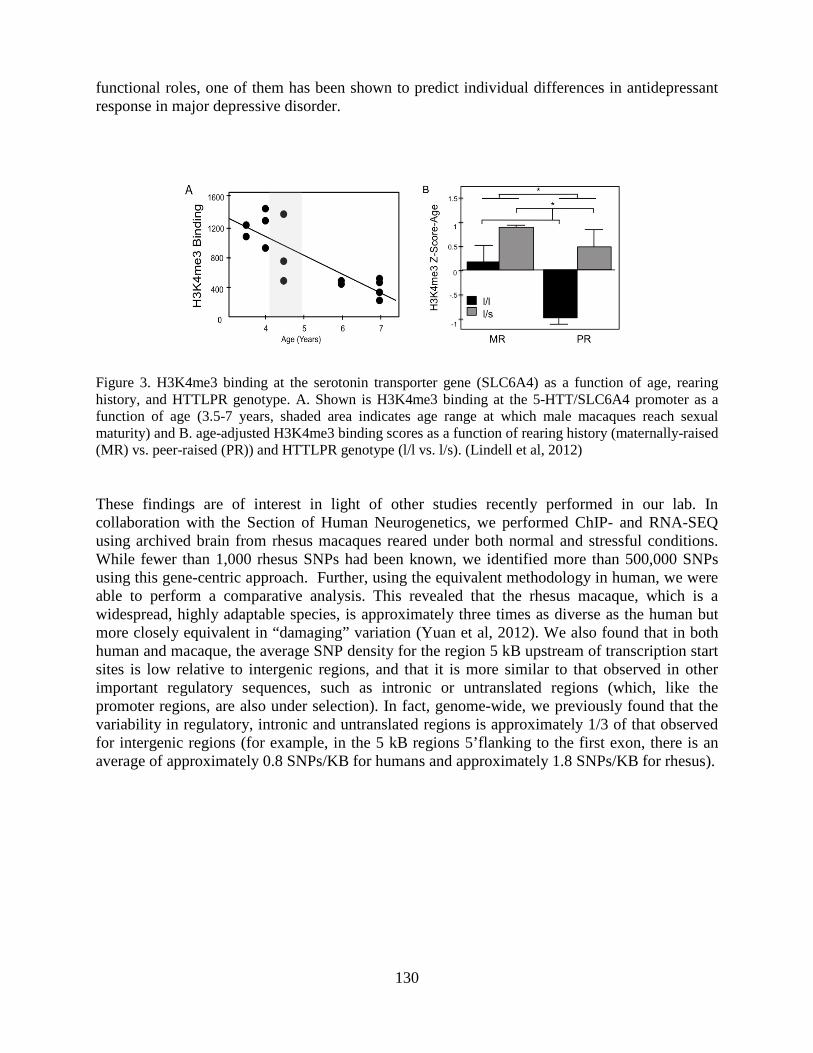
130
functional roles, one of them has been shown to predict individual differences in antidepressant response in major depressive disorder.
Figure 3. H3K4me3 binding at the serotonin transporter gene (SLC6A4) as a function of age, rearing history, and HTTLPR genotype. A. Shown is H3K4me3 binding at the 5-HTT/SLC6A4 promoter as a function of age (3.5-7 years, shaded area indicates age range at which male macaques reach sexual maturity) and B. age-adjusted H3K4me3 binding scores as a function of rearing history (maternally-raised (MR) vs. peer-raised (PR)) and HTTLPR genotype (l/l vs. l/s). (Lindell et al, 2012) These findings are of interest in light of other studies recently performed in our lab. In collaboration with the Section of Human Neurogenetics, we performed ChIP- and RNA-SEQ using archived brain from rhesus macaques reared under both normal and stressful conditions. While fewer than 1,000 rhesus SNPs had been known, we identified more than 500,000 SNPs using this gene-centric approach. Further, using the equivalent methodology in human, we were able to perform a comparative analysis. This revealed that the rhesus macaque, which is a widespread, highly adaptable species, is approximately three times as diverse as the human but more closely equivalent in “damaging” variation (Yuan et al, 2012). We also found that in both human and macaque, the average SNP density for the region 5 kB upstream of transcription start sites is low relative to intergenic regions, and that it is more similar to that observed in other important regulatory sequences, such as intronic or untranslated regions (which, like the promoter regions, are also under selection). In fact, genome-wide, we previously found that the variability in regulatory, intronic and untranslated regions is approximately 1/3 of that observed for intergenic regions (for example, in the 5 kB regions 5’flanking to the first exon, there is an average of approximately 0.8 SNPs/KB for humans and approximately 1.8 SNPs/KB for rhesus).
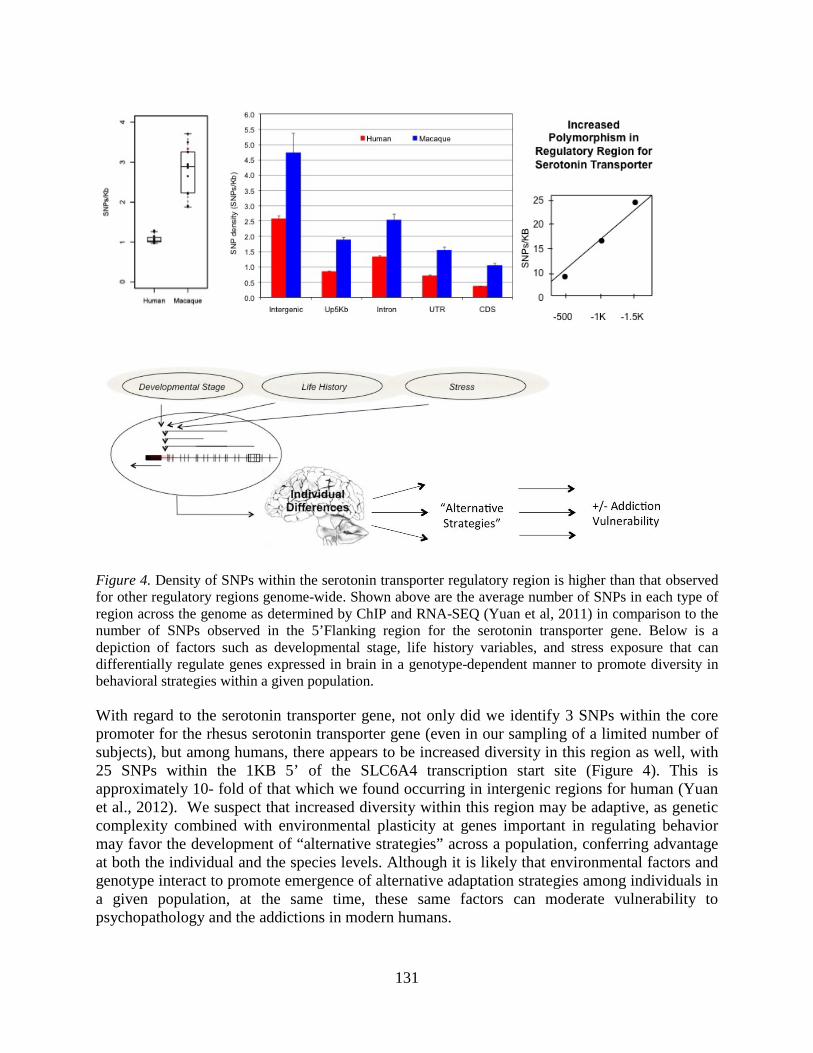
131
Figure 4. Density of SNPs within the serotonin transporter regulatory region is higher than that observed for other regulatory regions genome-wide. Shown above are the average number of SNPs in each type of region across the genome as determined by ChIP and RNA-SEQ (Yuan et al, 2011) in comparison to the number of SNPs observed in the 5’Flanking region for the serotonin transporter gene. Below is a depiction of factors such as developmental stage, life history variables, and stress exposure that can differentially regulate genes expressed in brain in a genotype-dependent manner to promote diversity in behavioral strategies within a given population. With regard to the serotonin transporter gene, not only did we identify 3 SNPs within the core promoter for the rhesus serotonin transporter gene (even in our sampling of a limited number of subjects), but among humans, there appears to be increased diversity in this region as well, with 25 SNPs within the 1KB 5’ of the SLC6A4 transcription start site (Figure 4). This is approximately 10- fold of that which we found occurring in intergenic regions for human (Yuan et al., 2012). We suspect that increased diversity within this region may be adaptive, as genetic complexity combined with environmental plasticity at genes important in regulating behavior may favor the development of “alternative strategies” across a population, conferring advantage at both the individual and the species levels. Although it is likely that environmental factors and genotype interact to promote emergence of alternative adaptation strategies among individuals in a given population, at the same time, these same factors can moderate vulnerability to psychopathology and the addictions in modern humans.
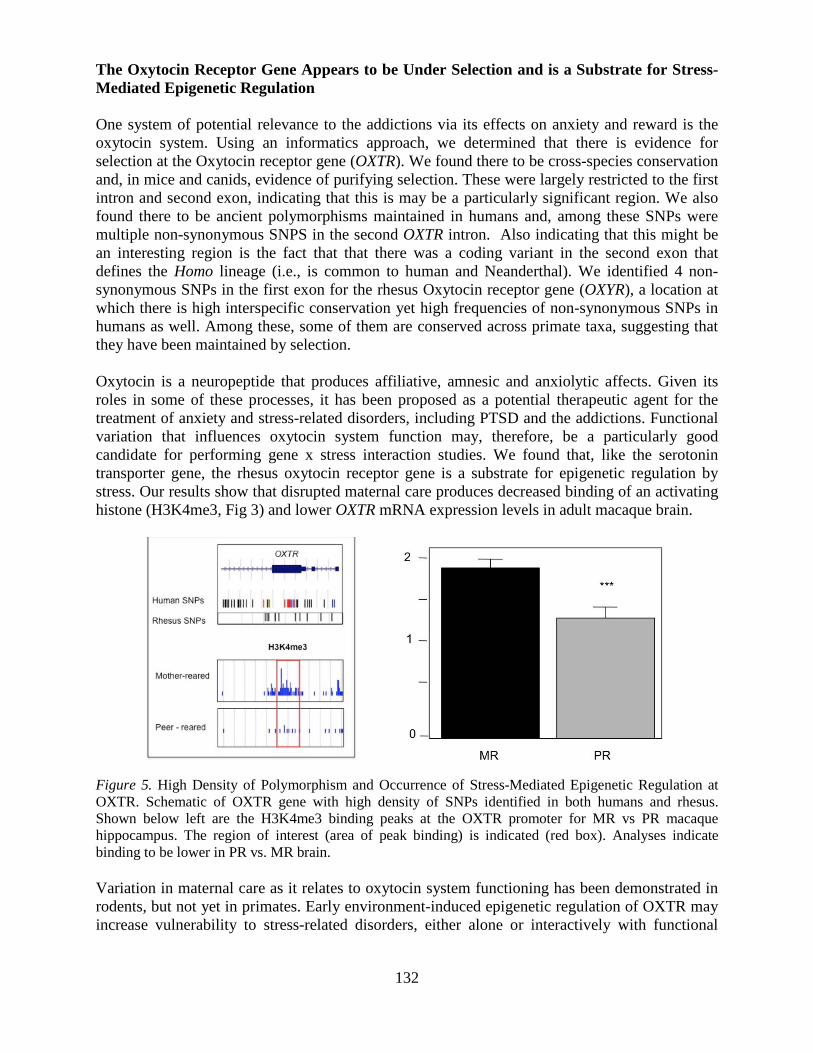
132
The Oxytocin Receptor Gene Appears to be Under Selection and is a Substrate for Stress-Mediated Epigenetic Regulation One system of potential relevance to the addictions via its effects on anxiety and reward is the oxytocin system. Using an informatics approach, we determined that there is evidence for selection at the Oxytocin receptor gene (OXTR). We found there to be cross-species conservation and, in mice and canids, evidence of purifying selection. These were largely restricted to the first intron and second exon, indicating that this is may be a particularly significant region. We also found there to be ancient polymorphisms maintained in humans and, among these SNPs were multiple non-synonymous SNPS in the second OXTR intron. Also indicating that this might be an interesting region is the fact that that there was a coding variant in the second exon that defines the Homo lineage (i.e., is common to human and Neanderthal). We identified 4 non-synonymous SNPs in the first exon for the rhesus Oxytocin receptor gene (OXYR), a location at which there is high interspecific conservation yet high frequencies of non-synonymous SNPs in humans as well. Among these, some of them are conserved across primate taxa, suggesting that they have been maintained by selection. Oxytocin is a neuropeptide that produces affiliative, amnesic and anxiolytic affects. Given its roles in some of these processes, it has been proposed as a potential therapeutic agent for the treatment of anxiety and stress-related disorders, including PTSD and the addictions. Functional variation that influences oxytocin system function may, therefore, be a particularly good candidate for performing gene x stress interaction studies. We found that, like the serotonin transporter gene, the rhesus oxytocin receptor gene is a substrate for epigenetic regulation by stress. Our results show that disrupted maternal care produces decreased binding of an activating histone (H3K4me3, Fig 3) and lower OXTR mRNA expression levels in adult macaque brain.
Figure 5. High Density of Polymorphism and Occurrence of Stress-Mediated Epigenetic Regulation at OXTR. Schematic of OXTR gene with high density of SNPs identified in both humans and rhesus. Shown below left are the H3K4me3 binding peaks at the OXTR promoter for MR vs PR macaque hippocampus. The region of interest (area of peak binding) is indicated (red box). Analyses indicate binding to be lower in PR vs. MR brain. Variation in maternal care as it relates to oxytocin system functioning has been demonstrated in rodents, but not yet in primates. Early environment-induced epigenetic regulation of OXTR may increase vulnerability to stress-related disorders, either alone or interactively with functional

133
genetic variation. This may also have implications for vulnerability to the addictions and other disorders in which disrupted social cognition or empathy are observed. References
Barr CS, Newman TK, Lindell S, Shannon C, Champoux M, Lesch KP, Suomi SJ, Goldman D, Higley JD. 2004. Interaction between serotonin transporter gene variation and rearing condition in alcohol preference and consumption in female primates. Arch Gen Psychiatry 61: 1146-52
Barr CS, Newman TK, Schwandt M, Shannon C, Dvoskin RL, Lindell SG, Taubman J, Thompson B, Champoux M, Lesch KP, Goldman D, Suomi SJ, Higley JD. 2004. Sexual dichotomy of an interaction between early adversity and the serotonin transporter gene promoter variant in rhesus macaques. Proc Natl Acad Sci U S A 101: 12358-63
Barr CS, Schwandt M, Lindell SG, Chen SA, Goldman D, Suomi SJ, Higley JD, Heilig M. 2007. Association of a functional polymorphism in the mu-opioid receptor gene with alcohol response and consumption in male rhesus macaques. Arch Gen Psychiatry 64: 369-76
Barr CS, Schwandt ML, Lindell SG, Higley JD, Maestripieri D, Goldman D, Suomi SJ, Heilig M. 2008. Variation at the mu-opioid receptor gene (OPRM1) influences attachment behavior in infant primates. Proc Natl Acad Sci U S A 105: 5277-81
Barr CS, Dvoskin RL, Yuan Q, Lipsky RH, Gupte M, Hu X, Zhou Z, Schwandt ML, Lindell SG, McKee M, Becker ML, Kling MA, Gold PW, Higley D, Heilig M, Suomi SJ, Goldman D. 2008. CRH haplotype as a factor influencing cerebrospinal fluid levels of corticotropin-releasing hormone, hypothalamic-pituitary-adrenal axis activity, temperament, and alcohol consumption in rhesus macaques. Arch Gen Psychiatry 65: 934-44
Copeland WE, Sun H, Costello EJ, Angold A, Heilig MA, Barr CS. 2011. Child mu-Opioid Receptor Gene Variant Influences Parent-Child Relations. Neuropsychopharmacology 36: 1165-70
Kraemer GW, Moore CF, Newman TK, Barr CS, Schneider ML. 2008. Moderate level fetal alcohol exposure and serotonin transporter gene promoter polymorphism affect neonatal temperament and limbic-hypothalamic-pituitary-adrenal axis regulation in monkeys. Biol Psychiatry 63: 317-24
Lindell SG, Schwandt ML, Sun H, Sparenborg JD, Bjork K, Kasckow JW, Sommer WH, Goldman D, Higley JD, Suomi SJ, Heilig M, Barr CS. 2010. Functional NPY variation as a factor in stress resilience and alcohol consumption in rhesus macaques. Arch Gen Psychiatry 67: 423-31
Lindell SG, Yuan QP, Zhou ZF, Goldman D, Thompson RC, Lopez JF, Suomi SJ, Higley JD, Barr CS. 2012. The serotonin transporter gene is a substrate for age and stress dependent epigenetic regulation in rhesus macaque brain: Potential roles in genetic selection and Gene x Environment interactions. Devel Psychopathol 24: 1391-400

134
Newman TK, Parker CC, Suomi SJ, Goldman D, Barr CS, Higley JD. 2009. DRD1 5'UTR variation, sex and early infant stress influence ethanol consumption in rhesus macaques. Genes Brain Behav 8: 626-30
Schneider ML, Moore CF, Larson JA, Barr CS, Dejesus OT, Roberts AD. 2009. Timing of moderate level prenatal alcohol exposure influences gene expression of sensory processing behavior in rhesus monkeys. Front Integr Neurosci 3: 30
Schwandt ML, Lindell SG, Sjoberg RL, Chisholm KL, Higley JD, Suomi SJ, Heilig M, Barr CS. 2010. Gene-environment interactions and response to social intrusion in male and female rhesus macaques. Biol Psychiatry 67: 323-30
Schwandt ML, Lindell SG, Higley JD, Suomi SJ, Heilig M, Barr CS. 2011. OPRM1 gene variation influences hypothalamic-pituitary-adrenal axis function in response to a variety of stressors in rhesus macaques. Psychoneuroendocrinology 36: 1303-11
Yuan QP, Zhou ZF, Lindell SG, Higley JD, Ferguson B, Thompson RC, Lopez JF, Suomi SJ, Baghal B, Baker M, Mash DC, Barr CS, Goldman D. 2012. The rhesus macaque is three times as diverse but more closely equivalent in damaging coding variation as compared to the human. BMC Genetics 13:52
Publications 2011-2012
1. Barr CS. 2012. Temperament in Animals, in Zentner and Shiner (Eds), Handbook of Temperament, Guilford Press, 251-272.
2. Cirulli F, Reif A, Herterich S, Lesch KP, Berry A, Francia N, Aloe L, Barr CS, Suomi SJ, Alleva E. 2011. A novel BDNF polymorphism affects plasma protein levels in interaction with early adversity in rhesus macaques. Psychoneuroendocrinology 36: 372-9
3. Copeland WE, Sun H, Costello EJ, Angold A, Heilig MA, Barr CS. 2011. Child mu-Opioid Receptor Gene Variant Influences Parent-Child Relations. Neuropsychopharmacology 36: 1165-70
4. Driscoll CA, Luo S, MacDonald D, Dinerstein E, Chestin I, Pereladova O, O'Brien SJ. 2011. Restoring tigers to the Caspian region. Science 333: 822-3
5. Driscoll C, Yamaguchi N, O'Brien SJ, Macdonald DW. 2011. A suite of genetic markers useful in assessing wildcat (Felis silvestris ssp.)-domestic cat (Felis silvestris catus) admixture. J Hered 102 Suppl 1: S87-90
6. Higham JP, Barr CS, Hoffman CL, Mandalaywala TM, Parker KJ, Maestripieri D. 2011. Mu-opioid receptor (OPRM1) variation, oxytocin levels and maternal attachment in free-ranging rhesus macaques Macaca mulatta. Behav Neurosci 125: 131-6
7. Lee MR, Gallen CL, Zhang X, Hodgkinson CA, Goldman D, Stein EA, Barr CS. 2011. Functional polymorphism of the mu-opioid receptor gene (OPRM1) influences reinforcement learning in humans. Plos One 6: e24203

135
8. Lindell SG, Yuan QP, Zhou ZF, Goldman D, Thompson RC, Lopez JF, Suomi SJ, Higley JD, Barr CS. 2012. The serotonin transporter gene is a substrate for age and stress dependent epigenetic regulation in rhesus macaque brain: Potential roles in genetic selection and Gene x Environment interactions. Devel Psychopathol 24: 1391-400
9. Ray LA, Barr CS, Blendy JA, Oslin D, Goldman D, Anton RF. 2012. The Role of the Asn40Asp Polymorphism of the Mu Opioid Receptor Gene (OPRM1) on Alcoholism Etiology and Treatment: A Critical Review. Alcohol Clin Exp Res 36: 385-94
10. Schneider ML, Moore CF, Barr CS, Larson JA, Kraemer GW. 2011. Moderate Prenatal Alcohol Exposure and Serotonin Genotype Interact to Alter CNS Serotonin Function in Rhesus Monkey Offspring. Alcohol Clin Exp Res 35: 912-20
11. Schwandt ML, Lindell SG, Higley JD, Suomi SJ, Heilig M, Barr CS. 2011. OPRM1 gene variation influences hypothalamic-pituitary-adrenal axis function in response to a variety of stressors in rhesus macaques. Psychoneuroendocrinology 36: 1303-11
12. Spinelli S, Schwandt ML, Lindell SG, Heilig M, Suomi SJ, Higley JD, Goldman D, Barr CS. 2012. The serotonin transporter gene linked polymorphic region is associated with the behavioral response to repeated stress exposure in infant rhesus macaques. Devel Psychopathol 24: 157-65
13. Yuan QP, Zhou ZF, Lindell SG, Higley JD, Ferguson B, Thompson RC, Lopez JF, Suomi SJ, Baghal B, Baker M, Mash DC, Barr CS, Goldman D. 2012. The rhesus macaque is three times as diverse but more closely equivalent in damaging coding variation as compared to the human. BMC Genetics 13:52
Dr. Barr: [email protected]

136
______________________________________________________________________
Laboratory of Integrative Neuroscience ______________________________________________________________________ David M. Lovinger, PhD, Chief Research in the Laboratory for Integrative Neuroscience (LIN) examines the role of particular molecules in acute alcohol intoxication, alcohol seeking behavior, addiction and habitual behavior. Another aim of research in LIN is to analyze the molecular basis of synaptic plasticity in striatum and other brain regions implicated in action and habit control. An important unifying theme of research within LIN is investigation of forebrain mechanisms involved in aspects of cognition and behavioral control that contribute to addiction. We seek to understand the neural basis of addiction-related brain functions such as decision-making, goal-directed behavior, habit formation and habitual behavior. We are also examining mechanisms underlying specific responses to addictive drugs, including acute intoxication and neural adaptations to chronic drug exposure, such as tolerance, dependence and drug seeking/taking behavior. Gaining an understanding of these functions will necessarily involve determining which neural circuits mediate these processes, what physiological and synaptic mechanisms underlie circuit function and plasticity, and what neural and synaptic molecules play key parts in synaptic, neuronal and circuit function. We believe that these goals can best be achieved by bringing together investigators with a shared interest in these subjects and with diverse technological expertise so that we can approach the problem at multiple levels of analysis. These aims are being pursued via the coordinated effort of investigators in two independent, interactive sections. Members of these sections have expertise in analyses ranging from molecular biology to physiological and behavioral analysis at the whole animal level. The Section on Synaptic Pharmacology (SSP) is headed by Dr. Lovinger. Members of the SSP have expertise in several areas of molecular and cellular neuroscience. These include molecular techniques for working with cDNA as well as analysis of protein expression, protein phosphorylation and structure-pharmacology relationships. Advanced cell culture techniques are also in use by section members. Members of this section also have considerable expertise in electrophysiological techniques for analysis of single neurons and small neural circuits in brain slices, as well as in vivo electrophysiology, photometry and neurochemistry. Expertise in cellular imaging also contributes to the present work and future aims of the SSP. Research in SSP has focused on three main areas. We have continued our long-standing efforts to understand the cellular and molecular mechanisms underlying plasticity at GABAergic and glutamatergic synapses in the striatum. This work led to an interest in the role of endogenous cannabinoids (endocannabinoids) and serotonin in synaptic modulation and plasticity in this brain region. Our interest in striatal synaptic plasticity has always been driven by a desire to better understand information storage in this brain region that plays crucial roles in action-outcome, habit and skill learning. Newly developed striatal-specific gene-targeted mouse

137
models are now making it possible for us to determine if striatal synaptic plasticity plays important roles in learning and memory. Our long-term goal in this project is to understand how the molecules and cells of the striatum contribute to the types of learning and memory involving that brain region, and to develop pharmacological and genetic manipulations that can alter striatal-based information storage. The second long-term area of emphasis in SSP is alcohol actions that contribute to intoxication and alcohol abuse/addiction. We continue to explore mechanisms of ethanol (EtOH) action on ligand-gated ion channels using a combination of molecular biological, electrophysiological and kinetic approaches. We are also characterizing presynaptic actions of EtOH that contribute to potentiation of GABAergic transmission. Our studies have established the presynaptic terminal as the likely site of this EtOH action. Recent studies spurred by ongoing collaborations have focused on comparison of electrophysiological and behavioral effects of EtOH in gene-targeted mice lacking molecules that are targets for acute alcohol actions. These studies are designed to examine the effects of acute ethanol as well as neuroadaptations brought about the chronic alcohol exposure. We are also working with members of the Section on Neuronal Structure and other collaborators to examine effects of chronic EtOH exposure and consumption on striatal neurophysiology and synaptic transmission. Our role in this part of the research effort is to examine EtOH sensitivity of receptors and alcohol effects on synaptic transmission using in vitro preparations. This section has also expanded its focus to include research on neural mechanisms of skill learning, goal-directed behavior, habit formation and addiction. One of the major areas of emphasis within the section is on the role of cortical-basal ganglia circuitry in these neural functions. Assessment of the role of particular molecules within these brain regions is also an area of emphasis. To this end, members of this section have implemented advanced methodology for in vivo electrophysiological recording and photometry in awake mice during different phases of task learning, and are developing in vivo voltammetric techniques as well. These techniques are used to analyze transgenic and gene-targeted mice. Members of this section are designing and using sophisticated behavioral paradigms to assess behavioral sequencing, as well as goal-directed and habit learning in mice. In addition, the outstanding molecular biology expertise in this section has allowed for creation of unique and powerful molecular tools and gene-targeted mice that can be used by this section, and by other LIN investigators. The Section on Neuronal Structure (SNS), headed by Dr. Veronica Alvarez is examining dynamic changes in neuronal morphology and dendritic spine function in relation to synaptic transmission, neurophysiology and behavior. In this section, advance microscopy methods, including multiphoton laser microscopy, are used to examine the fine structure of different parts of identified neurons. These microscopy techniques are also used to produce rapid and highly localized increases in neurotransmitter levels using laser uncaging, and to examine electrophysiological responses and intracellular signaling produced by these neurotransmitters. Research in this section also involves innovative techniques for gene expression and fluorescent labeling in individual cells. Behavioral models of cocaine and EtOH self-administration, ongoing within this section and in collaboration with other LIN sections, are used to examine how changes in neuronal morphology and physiology contribute to neuronal plasticity and neuroadaptions during addiction. We envision that the shared interests of LIN investigators and the interactive nature of the laboratory will allow investigators from different groups to combine their expertise in the

138
analysis of neural function at multiple levels. For example, changes in addiction-related behavior in a particular mouse model can be examined in conjunction with analysis of neuronal morphology, signaling in subcellular compartments, and neural circuit function at the micro and macro levels. Examination of molecular expression, location and function can also be applied in the same animals. Ultimately, this should allow us to gain a thorough understanding of the way in which particular neuronal molecules contribute to cellular and circuit functions that ultimately influence behavior. We also envision that interactions and collaborations with other NIAAA laboratories will enhance our ability to analyze these neural functions from the molecule to the whole mouse. One theme that brings together the different lines of research within LIN is an interest in mechanisms of goal directed actions, cue-controlled actions, addiction and habitual behavior. We predict that gaining a better understanding of synaptic plasticity in the striatum and related brain regions will help us to better appreciate how information relevant to habit formation becomes encoded. Our overarching hypothesis is that effects of addictive substances on synaptic structure and transmission will interface with natural plasticity mechanisms to focus habits on drugs of abuse as well as associated stimuli and responses. Thus, understanding the neural basis of habit formation and the molecular actions of intoxicating/addictive substances will allow us to gain a more thorough understanding of addiction. Ultimately, it is hoped that these lines of research can contribute to development of better therapies for neurological disorders and alcoholism. Dr. Lovinger: [email protected]

139
______________________________________________________________________
Section on Synaptic Pharmacology ______________________________________________________________________ David M. Lovinger, PhD, Chief Margaret I. Davis, PhD, Staff Scientist Li Zhang, MD, Research Medical Officer Yolanda Mateo, PhD, Staff Scientist Wei Xiong, PhD, Research Fellow Guohong Cui, PhD, Research Fellow Brian Mathur, PhD, Postdoc IRTA Verginia Cuzon Carlson, PhD, Postdoc IRTA Christina Gremel, PhD, Postdoc IRTA Brady Atwood, PhD, Postdoc IRTA David Kupferschmidt, Visiting Fellow Patrick Cody, Postbac IRTA Coordinated Activity of Striatal Direct and Indirect Pathway Neurons During Movement Sequencing (Cui et al. in press) Our research group has a long-standing interest in the contribution of striatal neuronal activity to performance of action sequences. The medium-sized spiny projection neurons (SPNs) of the striatum give rise to two output pathways, the direct pathway that acts through the Substantia Nigra to increase net cortical-spinal output, and the indirect pathway that acts through the external segment of the Globus pallidus to inhibit output. Activity in these two pathways controls the initiation and sequencing of actions, but the way in which the pathways work together has not been clear. A popular model suggests that direct pathway activity serves as a “go’ signal to initiate movement/sequences, while the indirect pathway halts actions via a “no-go” signal. This model suggests that the direct pathway should be active at the start of movement sequences, and the indirect pathway should be active when movement stops and even during periods when little movement is observed. Alternatively, the two pathways may work in a coordinated manner to promote the timing or sequencing of actions. For example, the direct pathway may facilitate a desired set of actions while the indirect pathway suppresses unwanted actions. In this scenario, the two pathways would be co-activated at the beginning, and perhaps during, action sequences. These models have been hard to disentangle mainly because it has been difficult to measure activity in the two pathways separately while an animal is performing an action sequence. To address this question, we developed a new fiber optic-based photometry system for measuring neuronal calcium fluctuations (as an indicator of neuronal activity) deep in the brain. The system combines laser-driven time-correlated single photon counting (TCSPC) spectroscopy with optical fibers implanted into mouse striatum. A genetically encoded calcium indicator (GECI) protein called GCaMP3 is expressed in direct or indirect pathway neurons via intrastriatal injection of an adeno associated virus (AAV) containing a double-inverted open reading frame (DIO) GCaMP3 construct in mice expressing cre recombinase under the control of either the D1 receptor (direct pathway) or A2A receptor (indirect pathway) promoters. In this
l to r, front: M Davis, A Luo, V Cuzon Carlson, C Gremel rear: G Cui, W Xiong, D Kupferschmidt, B Atwood, B Mathur, M Pava, Y Mateo, D Lovinger

140
way, the GCaMP3 protein is expressed only in the projection neurons of interest. We then implant the fiber into the striatum in one brain hemisphere, and measure calcium transients as the mouse performs a lever-pressing task in which it is required to perform 10 presses to obtain food. The TCSPC system allows us to measure fluorescence lifetime, providing a signature of GCaMP3 different from that of endogenous brain fluorescence signals. We can also separate fluorescence emission into different spectral components using a polychromator. With these measures we can clearly distinguish GCaMP3 signals reflective of intracellular calcium levels from other fluorescent events. We have observed fast, transient increase in GCaMP3 fluorescence in both sets of striatal projection neurons. These transients consist of increases in fluorescence intensity of a duration similar to transients observed with imaging in brain slices or cerebral cortex, and no change in fluorescence lifetime. The calcium transients are blocked by general anesthetics and a blocker of neuronal action potential firing, indicating that they are neuronal in origin. Transients in both direct and indirect pathway neurons are observable when mice are moving during task performance, but are few in number and very low in frequency when mice are sitting still. The latter observation indicates that the indirect pathway is not active when no overt movements occur, a first piece of evidence inconsistent with a simple no-go role for this pathway. When mice initiate sequences of action, such as moving from lever pressing to food retrieval, or vice versa, transients occur in a coincident fashion in both direct and indirect pathway projection neurons. This activity only occurs when the animal moves in a direction away from the hemisphere in which the transients are measured (e.g. a contraversive movement). The onset of increased transient frequency precedes the first evidence of movement by ~500 msec and continues for ~1 sec during movement. These findings indicate that direct and indirect pathway striatal projection neurons increase their activity in a coordinated manner just before and during the performance of action sequences. This finding is most consistent with models in which the two pathways work together to promote proper sequencing or timing of action sequences. These findings have implications for Parkinson’s Disease, in which improperly coordinated direct and indirect pathway activity likely contributes to akinesia. Understanding the activity of these pathways will also help us to gain a better understanding of changes in striatal output in response to drugs of abuse including alcohol. The technology used to carry out this study can be applied to measuring neuronal calcium signals in other brain regions, and should also be useful for measuring a variety of characteristics of fluorescent signals in brain (e.g. fluorescence lifetime changes), using different fluorescent reporter molecules. Differential, Postsynaptic Voltage-Dependent Endocannabinoid-Mediated Long-Term Synaptic Depression at Different GABAergic Striatal Synapses (Mathur et al., submitted) Endocannabinoids (eCBs) mediate short- and long-term depression of synaptic strength by retrograde transsynaptic signaling. In previous studies we characterized eCB-dependent long-term depression (LTD) at glutamatergic and GABAergic synapses onto striatal SPNs. At GABAergic synapses onto SPNs in the dorsolateral stiratum we found that eCB-LTD could be induced by stimulation with a 60s duration train of 1Hz stimulation (Adermark and Lovinger 2009). This stimulation has little effect on glutamatergic transmission in DLS. Physiological studies in vivo have indicated that the membrane potentials of SPNs often reside at or near the resting potential (the “down-state”) or at more depolarized potentials where action

141
potential firing is active (the “up-state”). Our original experiments on GABAergic LTD were conducted during recordings in which the postsynaptic SPN membrane potential was near the up-state, and thus we wanted to determine if such plasticity could be observed when neurons were in the down-state. Furthermore, the electrical stimulation used to measure GABAergic synaptic responses in our past studies could not be used to distinguish which GABAergic inputs were depressed after the LTD induction protocol. The SPNs receive GABAergic inputs from several sources, including major inputs from other SPNs and from the fast-spiking interneurons (FSIs). To specifically interrogate these pathways, we expressed the light-activated channel rhodopsin 2 (ChR2) protein in SPNs and FSIs using injection of a DIO ChR2 AAV construct into mice with Cre expressed in the different GABAergic striatal neuronal subtypes. In brain slices from these mice we then examined eCB-LTD induced using 1Hz stimulation and other protocols and measured as the change in amplitude of optically-evoked GABAergic inhibitory postsynaptic currents (oIPSCs) arising from SPN or FSI activation. Induction of LTD was attempted at a -80 mV down-state potential and a -60 mV up-state potential. Our findings revealed evidence for two distinct forms of eCB-LTD with different induction mechanisms, involving different eCBs, and involving different afferent inputs and postsynaptic SPN subtypes. To identify molecules involved LTD, we used a variety of pharmacological manipulations and mice in which key proteins were knocked out either constitutively or conditionally and cell specifically. These experiments revealed that up-state LTD depends on coordinated activation of glutamatergic and GABAergic inputs, with induction requiring activation of L-type voltage-gated calcium channels. This form of LTD is mediated by postsynaptic generation and release of the eCB 2-arachidonoyl glycerol (2-AG) and activation of cannabinoid type 1 (CB1) receptors. The “up-state” eCB-LTD appears to exclusively occur at synapses made by presynaptic SPNs onto postsynaptic SPNs of both the direct and indirect pathways. The “down-state” form of eCB-LTD does not require glutamatergic transmission or activation of L-type channels. The eCB that mediates this form of LTD appears to be arachidonoyl ethanolamide (AEA, also known as anandamide), acting on presynaptic CB1 receptors. The down-state eCB-LTD is less input-specific than up-state LTD, as it is expressed at both SPN-SPN and FSI-SPN synapses. Interestingly, down-state eCB-LTD is only observed at synapses onto postsynaptic direct pathway SPNs. We also uncovered evidence that calcium influx through L-type channels may suppress down-state eCB-LTD, favoring the up-state form. These findings indicate that the efficacy of transmission at different GABAergic synapses onto SPNs is controlled by a combination of afferent input and postsynaptic membrane potential. Inputs from SPNs have subtle effects on dendritic integration within other SPNs, while FSI inputs can strongly control the excitability and output of SPNs. Thus the differential plasticity at these GABAergic synapses allows for a wide range of information storage and circuit control in different brain and behavioral states. One interesting feature of the down-state LTD is that it would allow for stronger direct pathway output, perhaps broadening the repertoire within certain action sequences. Long-term Alcohol Consumption Alters Striatal Physiology associated with Highly Regulated Intake in Non-Human Primates (Cuzon-Carlson et al. 2011, Cuzon-Carlson et al., in preparation) The basal ganglia have been implicated in control of drug taking, drug abuse and addiction. Emerging evidence indicates that striatal subregions that are part of the “sensorimotor” circuitry

142
(e.g. dorsolateral striatum in rodents, putamen nucleus in primates) participate in control of habitual behaviors including habitual/compulsive drug use. We are thus interested in how prolonged and excessive use of a drug, in our case alcohol, alters the circuitry within these striatal subregions. One approach we have taken to this question is to examine synaptic transmission and neuronal excitability in striatal SPNs in a primate model of voluntary alcohol intake. Rhesus and cynomolgus macaque monkeys are trained to drink alcohol and then are allowed 22 hours of drinking access per day for 1-3 years, in some cases with intermittent forced abstinence periods. The animals are then brought to necropsy where we obtain striatal tissue and can cut and record from brain slices to examine GABAergic miniature inhibitory postsynaptic currents (mIPSCs), glutamatergic miniature excitatory postsynaptic currents (mEPSCs), and action potential firing in response to current injection. In our initial study of monkeys that drank alcohol for 3 years with 3 forced abstinence periods, we observed decreased frequency of mIPSCs in putamen SPNs, indicative of decreased GABA release or synapse number (Cuzon Carlson et al. 2011). The mIPSC frequency was strongly negatively correlated with average drinking and blood alcohol levels, suggesting that disinhibition of the putamen may contribute to heavy drinking. We also observed increased excitability of SPNs using action potential frequency with current injection as the measure. In addition, an increase in glutamatergic mEPSC frequency was observed, along with an increase in the average number of dendritic spines on SPNs. These last findings suggest that alcohol drinking is associated with an increase in the number of glutamatergic synapses on putamen SPNs. Overall, the findings from this study suggest that the projection neurons of the striatum are hyperexcitable following long-term alcohol drinking. This could contribute to habitual control of alcohol seeking and drinking behavior. Indeed, the animals showed invariant drinking patterns during the last year of alcohol intake. These studies have been extended to examine putamen slices from animals that have been drinking for one year with no forced abstinence periods. In addition, the age of onset of drinking varied in different monkey cohorts, ranging from late adolescence to full adulthood. Findings to date indicate that the decrease in mIPSC frequency is observed following one year of alcohol drinking with no abstinence, and that this effect is much larger in young adult and adolescent-onset drinkers than in animals that began drinking as adults. Interestingly, the heaviest drinking was observed in these younger-onset groups. Thus, decreases in GABAergic function in the striatum appear to accompany heavy drinking per se, and are not a consequence of abstinence. Age of drinking onset is one factor that may predispose animals to heavy drinking and GABAergic dysfunction. We hope that these findings will prove useful for predicting and controlling alcohol intake in laboratory animals. These studies may also help us to understand vulnerabilities of the human brain to alcohol drinking, and aid in the development of therapies to curtail alcohol abuse. References
Adermark L and Lovinger DM. 2009. Frequency-dependent inversion of net striatal output by endocannabinoid-dependent plasticity at different synaptic inputs. Journal of Neuroscience 29: 1375-80
Cui G, Jun SB, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM. Concurrent
activation of striatal direct and indirect pathways during action initiation. Nature in press.

143
Cuzon Carlson VC, Seabold GK, Helms CM, Garg N, Odagiri M, Rau AR, Daunais J, Alvarez VA, Lovinger DM, Grant KA. 2011. Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates. Neuropsychopharmacology 36: 2513-28
Publications 2011-2012 1. Bello EP, Mateo Y, Gelman DM, Noain D, Shin JH, Low MJ, Alvarez VA, Lovinger
DM, Rubinstein M. 2011. Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D-2 autoreceptors. Nature Neuroscience 14: 1033-U128
2. Borghese CM, Blednov YA, Quan Y, Iyer SV, Xiong W, Mihic SJ, Zhang L, Lovinger DM, Trudell JR, Homanics GE, Harris RA. 2012. Characterization of Two Mutations, M287L and Q266I, in the alpha 1 Glycine Receptor Subunit That Modify Sensitivity to Alcohols. Journal of Pharmacology and Experimental Therapeutics 340: 304-16
3. Cachope R, Mateo Y, Mathur BN, Irving J, Wang HL, Morales M, Lovinger DM, Cheer JF. 2012. Selective Activation of Cholinergic Interneurons Enhances Accumbal Phasic Dopamine Release: Setting the Tone for Reward Processing. Cell Reports 2: 33-41
4. Cuzon Carlson VC, Mathur BN, Davis MI, Lovinger DM. 2011. Subsets of Spiny Striosomal Striatal Neurons Revealed in the Gad1-GFP BAC Transgenic Mouse. Basal Ganglia 1: 201-11
5. Cuzon Carlson VC, Seabold GK, Helms CM, Garg N, Odagiri M, Rau AR, Daunais J, Alvarez VA, Lovinger DM, Grant KA. 2011. Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates. Neuropsychopharmacology 36: 2513-28
6. Dang MT, Yokoi F, Cheetham CC, Lu J, Vo V, Lovinger DM, Li YQ. 2012. An anticholinergic reverses motor control and corticostriatal LTD deficits in Dyt1 Delta GAG knock-in mice. Behavioural Brain Research 226: 465-72
7. Jun SB, Cuzon Carlson V, Ikeda S, Lovinger D. 2011. Vibrodissociation of neurons from rodent brain slices to study synaptic transmission and image presynaptic terminals. J Vis Exp
8. Karlsson RM, Adermark L, Molander A, Perreau-Lenz S, Singley E, Solomon M, Holmes A, Tanaka K, Lovinger DM, Spanagel R, Heilig M. 2012. Reduced alcohol intake and reward associated with impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology 63: 181-9
9. Kim HY, Moon HS, Cao DH, Lee J, Kevala K, Jun SB, Lovinger DM, Akbar M, Huang BX. 2011. N-Docosahexaenoylethanolamide promotes development of hippocampal neurons. Biochemical Journal 435: 327-36
10. Klug JR, Mathur BN, Kash TL, Wang HD, Matthews RT, Robison AJ, Anderson ME, Deutch AY, Lovinger DM, Colbran RJ, Winder DG. 2012. Genetic Inhibition of CaMKII in Dorsal Striatal Medium Spiny Neurons Reduces Functional Excitatory Synapses and Enhances Intrinsic Excitability. Plos One 7

144
11. Lin X, Parisiadou L, Sgobio C, Liu GX, Yu J, Sun LX, Shim H, Gu XL, Luo J, Long CX, Ding JH, Mateo Y, Sullivan PH, Wu LG, Goldstein DS, Lovinger D, Cai HB. 2012. Conditional Expression of Parkinson's Disease-Related Mutant alpha-Synuclein in the Midbrain Dopaminergic Neurons Causes Progressive Neurodegeneration and Degradation of Transcription Factor Nuclear Receptor Related 1. Journal of Neuroscience 32: 9248-64
12. Mathur BN, Capik NA, Alvarez VA, Lovinger DM. 2011. Serotonin Induces Long-Term Depression at Corticostriatal Synapses. Journal of Neuroscience 31: 7402-11
13. Morton RA, Luo GX, Davis MI, Hales TG, Lovinger DM. 2011. Fluorophore assisted light inactivation (FALI) of recombinant 5-HT(3)A receptor constitutive internalization and function. Molecular and Cellular Neuroscience 47: 79-92
14. Torres-Altoro MI, Mathur BN, Drerup JM, Thomas R, Lovinger DM, O'Callaghan JP, Bibb JA. 2011. Organophosphates dysregulate dopamine signaling, glutamatergic neurotransmission, and induce neuronal injury markers in striatum. Journal of Neurochemistry 119: 303-13
15. Xiong W, Wu XW, Lovinger DM, Zhang L. 2012. A Common Molecular Basis for Exogenous and Endogenous Cannabinoid Potentiation of Glycine Receptors. Journal of Neuroscience 32: 5200-8
Collaborators
Dr. Veronica Alvarez, Section on Neuronal Structure, LIN, NIAAA Dr. Andrew Holmes, Laboratory of Behavioral and Genomic Neuroscience, NIAAA Dr. Rui Costa, Champalimaud Neuroscience Institute, Lisbon, Portugal Dr. Stephen R. Ikeda, Laboratory of Molecular Physiology, NIAAA Dr. Steven S. Vogel, Laboratory of Molecular Physiology, NIAAA Dr. George Kunos, Laboratory of Physiologic Studies, NIAAA Dr. Hee-Yong Kim, Laboratory of Molecular Signaling, NIAAA Dr. Eric Delpire, Vanderbilt University School of Medicine Dr. Roger Colbran, Vanderbilt University School of Medicine Dr. Danny Winder, Vanderbilt University School of Medicine Dr. Kathleen Grant, Oregon Health Sciences University Dr. Marcelo Rubinstein, University of Buenos Aires, Argentina Dr. Timothy Hales, University of Dundee, Scotland Dr. Huaibin Cai, NIA intramural research program Dr. Yuqing Li, University of Alabama Birmingham Dr. Joseph Cheer, University of Maryland School of Medicine
Dr. Lovinger: [email protected]

145
______________________________________________________________________
Section on Neuronal Structure ______________________________________________________________________ Veronica Alvarez, PhD, Acting Chief Jung Hoon Shin, PhD, Postdoctoral IRTA Martin Adrover, PhD, Postdoctoral IRTA Alice Dobi, PhD, Postdoctoral IRTA Roland Bock, MSc, Technical IRTA Paul F. Kramer, BS, Postbac IRTA Nyssa Sherazee, BS, Postbac IRTA Mark Wilcox, BS, Postbac IRTA Robert Fuino, BS, Postbac IRTA
l to r: G Seabold, JH Shin, R Fuino, M Adrover, P Kramer, R Bock, V Alvarez, A Dobi, N Sherazee
Mission Research in the Section on Neuronal Structure is focused on understanding the effects of drugs of abuse on synapses and neuronal connectivity with the purpose of revealing the mechanisms that induce the long-term behavior changes that characterize individuals suffering from drug abuse and dependence disorders. The projects focus on cocaine and ethanol, two prominent drugs of abuse in the US and we are investigating the short and long term actions on synaptic transmission in brain regions that are implicated in reward motivated learning: the nucleus accumbens (NAc) and the ventral tegmental area (VTA). One salient characteristic of addiction is that not every person exposed to drugs of abuse becomes an addict. For example, only 20% of cocaine users become compulsive drug-takers. Similarly, only a fraction of research animals that self-administer cocaine show addiction-like behaviors (1-3). One important part of our research is focused on understanding what is different in the brain of animals that developed addictive behaviors. Understanding the changes in the brain that contribute to compulsive drug taking and the vulnerability factors of becoming an addict are crucial for improving treatment and prevention of this disease.

146
Research Aims The overarching hypothesis addressed by our research is that repeated cocaine or ethanol exposures cause long-term changes in neuronal connectivity among nuclei of the mesolimbic system that lead to life-long disruptions in reward-motivated behaviors and learning. Below is a description of the three main aims of our research and a few examples of the studies completed and underway. AIM 1: Effects of acute and chronic cocaine exposure on synaptic transmission in the nucleus accumbens and regions of the mesolimbic system The goal of these studies is to dissect the initial synaptic changes that occur as a consequence of a few repeated exposures. The studies use novel technical approaches such as electrophysiological recording combined with two-photon imaging, glutamate uncaging and optogenetic stimulation to study glutamatergic transmission in the nucleus accumbens. The role of protracted cocaine withdrawal in the development of functional and structural plasticity in the NAc (4) This study tested the hypothesis that prolonged cocaine withdrawal is required for the enhancement of glutamatergic transmission induced by cocaine in the NAc. Most adaptations induced by cocaine in the NAc are only apparent several weeks after the last cocaine injection (5-8). These and other observations led to the hypothesis that cocaine withdrawal plays an important role in the development of neuronal adaptations in the NAc, and raises the possibility that withdrawal itself might be the trigger of this plasticity. To test this, the study compared the changes in glutamatergic transmission induced by two different cocaine treatments: a short treatment followed by prolonged withdrawal, and a longer treatment in which cocaine exposure was continued and protracted withdrawal was avoided. Under these two treatments, mice received different total doses of cocaine (210 mg/kg in the short and 600 mg/kg in the long treatment). However, the time after the last and the first injections were matched in the treatments, such that we could evaluate and compare the effect of prolonged withdrawal with the effect of elapsed time after the first injection. The results showed that glutamatergic inputs onto D1-MSNs are particularly susceptible to the effects of cocaine and locomotor sensitization. Protracted cocaine withdrawal is not required for these synaptic adaptations and; while few repeated cocaine injections are sufficient to trigger the plasticity, its expression is time-dependent. We speculate that the changes take many days to develop because they require the activity of the whole basal ganglia circuit loop. Recurrent network activity was shown to drive striatal synaptogenesis during early postnatal development (9). Future studies will test the hypothesis that the cocaine-induced synaptogenesis is triggered by initial changes in striatal output that affect the activity of the network loop and increase spine density and functional glutamatergic inputs. Acute cocaine effects on glutamatergic and dopaminergic transmission in the nucleus accumbens (Adrover, Shin and Alvarez, in preparation)
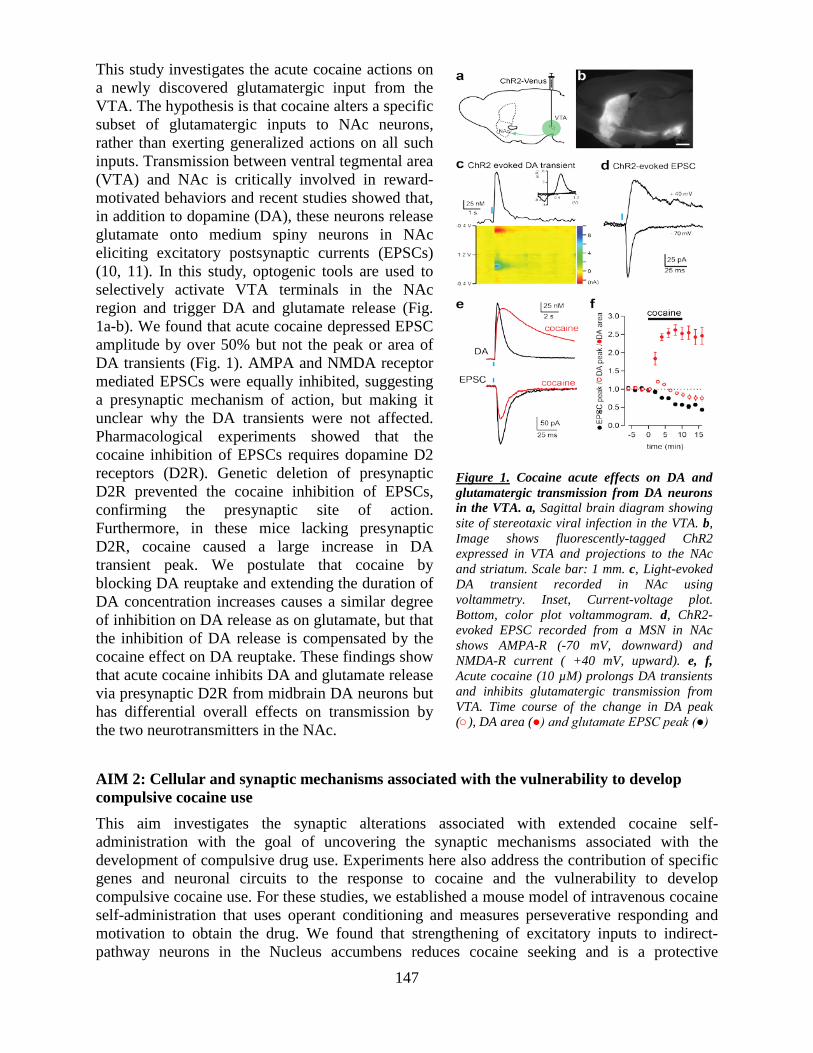
147
Figure 1. Cocaine acute effects on DA and glutamatergic transmission from DA neurons in the VTA. a, Sagittal brain diagram showing site of stereotaxic viral infection in the VTA. b, Image shows fluorescently-tagged ChR2 expressed in VTA and projections to the NAc and striatum. Scale bar: 1 mm. c, Light-evoked DA transient recorded in NAc using voltammetry. Inset, Current-voltage plot. Bottom, color plot voltammogram. d, ChR2-evoked EPSC recorded from a MSN in NAc shows AMPA-R (-70 mV, downward) and NMDA-R current ( +40 mV, upward). e, f, Acute cocaine (10 µM) prolongs DA transients and inhibits glutamatergic transmission from VTA. Time course of the change in DA peak (○), DA area (●) and glutamate EPSC peak (●)
This study investigates the acute cocaine actions on a newly discovered glutamatergic input from the VTA. The hypothesis is that cocaine alters a specific subset of glutamatergic inputs to NAc neurons, rather than exerting generalized actions on all such inputs. Transmission between ventral tegmental area (VTA) and NAc is critically involved in reward-motivated behaviors and recent studies showed that, in addition to dopamine (DA), these neurons release glutamate onto medium spiny neurons in NAc eliciting excitatory postsynaptic currents (EPSCs) (10, 11). In this study, optogenic tools are used to selectively activate VTA terminals in the NAc region and trigger DA and glutamate release (Fig. 1a-b). We found that acute cocaine depressed EPSC amplitude by over 50% but not the peak or area of DA transients (Fig. 1). AMPA and NMDA receptor mediated EPSCs were equally inhibited, suggesting a presynaptic mechanism of action, but making it unclear why the DA transients were not affected. Pharmacological experiments showed that the cocaine inhibition of EPSCs requires dopamine D2 receptors (D2R). Genetic deletion of presynaptic D2R prevented the cocaine inhibition of EPSCs, confirming the presynaptic site of action. Furthermore, in these mice lacking presynaptic D2R, cocaine caused a large increase in DA transient peak. We postulate that cocaine by blocking DA reuptake and extending the duration of DA concentration increases causes a similar degree of inhibition on DA release as on glutamate, but that the inhibition of DA release is compensated by the cocaine effect on DA reuptake. These findings show that acute cocaine inhibits DA and glutamate release via presynaptic D2R from midbrain DA neurons but has differential overall effects on transmission by the two neurotransmitters in the NAc. AIM 2: Cellular and synaptic mechanisms associated with the vulnerability to develop compulsive cocaine use This aim investigates the synaptic alterations associated with extended cocaine self-administration with the goal of uncovering the synaptic mechanisms associated with the development of compulsive drug use. Experiments here also address the contribution of specific genes and neuronal circuits to the response to cocaine and the vulnerability to develop compulsive cocaine use. For these studies, we established a mouse model of intravenous cocaine self-administration that uses operant conditioning and measures perseverative responding and motivation to obtain the drug. We found that strengthening of excitatory inputs to indirect-pathway neurons in the Nucleus accumbens reduces cocaine seeking and is a protective

148
mechanism against the expression of compulsive cocaine use. We are now incorporating optogenetic and chemicogenetic (Designer REceptors Activate by Designer Drugs) in vivo approaches to activate or inhibit specific neuronal circuits during the task. These studies, which have required large investment of time and resources to overcome technical difficulties, have now proven successful. Strengthening the accumbal indirect pathway promotes resilience to compulsive cocaine use (Bock. et al., Nature Neuroscience, in press) A hallmark of addiction is the loss of control over drug intake, which is seen only in a fraction of those exposed to stimulant drugs like cocaine. The cellular mechanisms underlying vulnerability or resistance to compulsive drug use are still unknown. The hypothesis behind this study is that the vulnerability to develop compulsive cocaine use arises in part from inherent differences across animals in the ability to recruit the D2-MSNs and engage the indirect accumbal-tegmental pathway. Evidence from previous studies suggests that activation of D2 receptors is involved in the locomotor response to cocaine and conditioned place preference (12,13). However, it is still unclear what the role of D2-MSNs is with regard to voluntary cocaine self-administration and the development of compulsive cocaine use. Experiments also test the hypothesis that inhibition of D2-MSNs impairs self-control over cocaine intake and renders individuals more susceptible to the rewarding effects of the drug. The results showed that individual variability in the development of highly motivated and perseverative behavior toward cocaine was associated with synaptic plasticity in medium spiny neurons expressing dopamine D2 receptors (D2-MSNs) in the nucleus accumbens. Potentiation of glutamatergic inputs onto indirect pathway D2-MSNs was associated with resilience towards compulsive cocaine seeking. Inhibition of D2-MSNs using a chemicogenetic approach enhanced the motivation to obtain cocaine while optogenetic activation of D2-MSNs suppressed cocaine self-administration. In conclusion, this study establishes that synaptic potentiation in D2-MSN inputs is a critical mechanism for controlling the expression of compulsive behaviors towards cocaine. We propose that this cell-specific synaptic potentiation facilitates the recruitment of indirect pathway neurons and protects against the development addictive behaviors. Role of D2 receptors on dopaminergic neurons (autoreceptors) in the regulation of reward-motivated and motor learning (14) D2 receptors expressed in DA neurons (D2 autoreceptors) had been proposed to exert a negative feedback regulation on DA neuron firing, DA synthesis and release. However, pharmacological and genetic approaches used so far have been unable to address directly the contribution of D2 autoreceptors in vivo because these receptors are also expressed in striatal MSNs at high levels (postsynaptic receptors). This study tested the hypothesis that selective elimination of D2 autoreceptors would enhance DA release and boost DA-mediated behaviors. Midbrain DA neurons from autoD2-KO mice lack DA-mediated somatodendritic synaptic response and were not inhibited by DA release. AutoD2-KO mice displayed elevated DA synthesis and release, hyperlocomotion and increased sensitivity to the psychomotor effects of cocaine compared to D2f/f littermates. AutoD2-KO mice also exhibited increased place preference for cocaine and enhanced motivation for food reward. The study highlights the importance of D2 autoreceptors in the regulation of DA neurotransmission and demonstrates that D2 autoreceptors are key players in normal motor function, food seeking behavior and sensitivity to the locomotor and rewarding properties of cocaine.

149
AIM 3: Chronic intermittent ethanol drinking and its effect on striatal synaptic function and morphology This aim studies the effects of chronic intermittent ethanol drinking on synaptic transmission and morphology in the striatum. We completed a collaborative study that employed a model of ethanol oral self-administration in non-human primates and showed increased dendritic spine density in the striatum of ethanol drinkers (15). In addition, we established in the laboratory a mouse model of intermittent ethanol binge-drinking and, using a lickometer, characterized the pattern of drinking with high temporal resolution during the acquisition of the binge-drinking behavior. Synaptic morphology in the NAc and the dorsal striatum was addressed using diolisitic labeling and confocal imaging (Wilcox et al., under review). Changes in striatal synaptic morphology associated with extended ethanol drinking in non-human primates (15) In this study performed in collaboration with the laboratories of Drs. Kathleen Grant and David Lovinger, we investigated the synaptic changes, both at the functional and structural level, that are associated with chronic ethanol self-administration in monkeys. Chronic intermittent ethanol drinking was established in macaque monkeys to explore neuronal adaptations in the striatum, a brain region controlling habitual behaviors. Prolonged drinking with repeated abstinence narrowed the variability in daily intake, increased the rate of drinking and led to consistently higher blood ethanol concentrations more than twice the legal intoxication limit. Following extensive drinking, we found increased dendritic spine density and enhanced glutamatergic transmission in the putamen, but not the caudate, striatal subregions. GABAergic transmission was suppressed in the putamen selectively in heavy drinkers. Intrinsic excitability of medium spiny neurons was also enhanced in putamen of alcohol-drinking monkeys in comparison to non-drinkers. These morphological and physiological changes indicate a shift in the balance of inhibitory/excitatory transmission that biases the circuit toward stronger synaptic activation of putamen output as a consequence of prolonged heavy drinking/relapse. The resultant potential for increased putamen activation may underlie a key feature of alcoholism, contributing to the evolved pattern of regulated drinking leading to sustained intoxication. Dissection of the drinking behavior and analysis of spine morphology in a mouse model of binge-like ethanol drinking (Wilcox and Carlson Cuzon et al., under review) Research into the neurobiology of heavy and binge-like ethanol drinking has been limited by the low-levels of voluntary ethanol consumption shown by most mouse strains. Recently, a model of intermittent access to ethanol has been shown to elicit binge-like drinking and pharmacologically relevant blood ethanol concentrations (BECs) in mice. Termed “Drinking in the Dark”, this model takes advantage of the circadian patterns of mice to achieve reliably high levels of consumption in a two hour drinking session. C57BL/6J mice reach BECs higher than 80 mg/dl, and show signs of intoxication such as motor impairment. DID is a robust paradigm that has been successfully used to investigate neuronal circuits and signals that modulate binge-like ethanol consumption. Until now, it has been thought to work by limiting the access to ethanol to just a few hours a day during active circadian phase but the exact behavioral mechanism responsible for inducing fast rate drinking remains unclear. In this study, we established DID in our laboratory and showed that it produces reliable escalation of voluntary ethanol intake and blood ethanol concentration (Fig. 2a-c). We used lickometers to record with high temporal resolution the drinking pattern of mice during
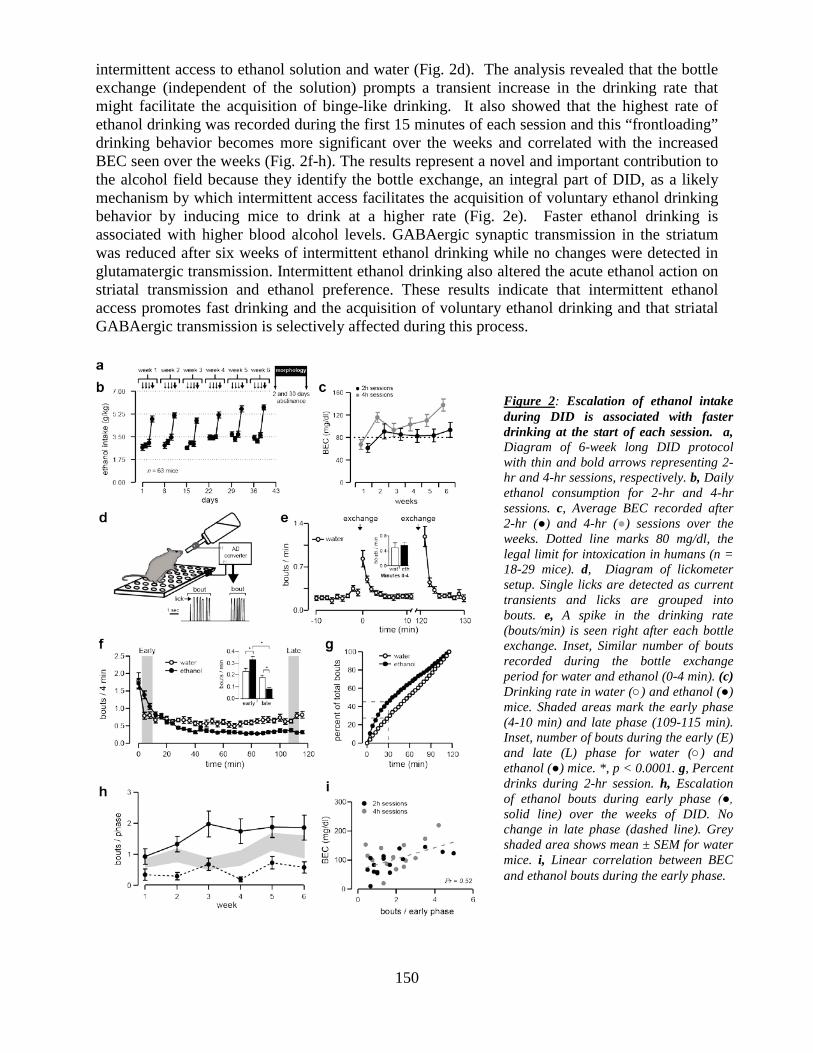
150
intermittent access to ethanol solution and water (Fig. 2d). The analysis revealed that the bottle exchange (independent of the solution) prompts a transient increase in the drinking rate that might facilitate the acquisition of binge-like drinking. It also showed that the highest rate of ethanol drinking was recorded during the first 15 minutes of each session and this “frontloading” drinking behavior becomes more significant over the weeks and correlated with the increased BEC seen over the weeks (Fig. 2f-h). The results represent a novel and important contribution to the alcohol field because they identify the bottle exchange, an integral part of DID, as a likely mechanism by which intermittent access facilitates the acquisition of voluntary ethanol drinking behavior by inducing mice to drink at a higher rate (Fig. 2e). Faster ethanol drinking is associated with higher blood alcohol levels. GABAergic synaptic transmission in the striatum was reduced after six weeks of intermittent ethanol drinking while no changes were detected in glutamatergic transmission. Intermittent ethanol drinking also altered the acute ethanol action on striatal transmission and ethanol preference. These results indicate that intermittent ethanol access promotes fast drinking and the acquisition of voluntary ethanol drinking and that striatal GABAergic transmission is selectively affected during this process.
Figure 2: Escalation of ethanol intake during DID is associated with faster drinking at the start of each session. a, Diagram of 6-week long DID protocol with thin and bold arrows representing 2-hr and 4-hr sessions, respectively. b, Daily ethanol consumption for 2-hr and 4-hr sessions. c, Average BEC recorded after 2-hr (●) and 4-hr (●) sessions over the weeks. Dotted line marks 80 mg/dl, the legal limit for intoxication in humans (n = 18-29 mice). d, Diagram of lickometer setup. Single licks are detected as current transients and licks are grouped into bouts. e, A spike in the drinking rate (bouts/min) is seen right after each bottle exchange. Inset, Similar number of bouts recorded during the bottle exchange period for water and ethanol (0-4 min). (c) Drinking rate in water (○) and ethanol (●) mice. Shaded areas mark the early phase (4-10 min) and late phase (109-115 min). Inset, number of bouts during the early (E) and late (L) phase for water (○) and ethanol (●) mice. *, p < 0.0001. g, Percent drinks during 2-hr session. h, Escalation of ethanol bouts during early phase (●, solid line) over the weeks of DID. No change in late phase (dashed line). Grey shaded area shows mean ± SEM for water mice. i, Linear correlation between BEC and ethanol bouts during the early phase.

151
References 1. Belin D, Mar AC, Dalley JW, Robbins TW, Everitt BJ. 2008. High impulsivity predicts
the switch to compulsive cocaine-taking. Science 320: 1352-5 2. Dalley JW, Fryer TD, Brichard L, Robinson ES, Theobald DE, Laane K, Pena Y,
Murphy ER, Shah Y, Probst K, Abakumova I, Aigbirhio FI, Richards HK, Hong Y, Baron JC, Everitt BJ, Robbins TW. 2007. Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement. Science 315: 1267-70
3. Deroche-Gamonet V, Belin D, Piazza PV. 2004. Evidence for addiction-like behavior in the rat. Science 305: 1014-7
4. Dobi A., Seabold G.K., Christensen C.H., Bock R. and Alvarez V.A. (2011) Cocaine-induced plasticity in the nucleus accumbens is cell specific and develops without withdrawal. Journal of Neuroscience 31(5):1895-904.
5. Conrad KL, Tseng KY, Uejima JL, Reimers JM, Heng LJ, Shaham Y, Marinelli M, Wolf ME. 2008. Formation of accumbens GluR2-lacking AMPA receptors mediates incubation of cocaine craving. Nature 454: 118-21
6. Kourrich S, Rothwell PE, Klug JR, Thomas MJ. 2007. Cocaine experience controls bidirectional synaptic plasticity in the nucleus accumbens. J Neurosci 27: 7921-8
7. Robinson TE, Gorny G, Mitton E, Kolb B. 2001. Cocaine self-administration alters the morphology of dendrites and dendritic spines in the nucleus accumbens and neocortex. Synapse 39: 257-66
8. Thomas MJ, Kalivas PW, Shaham Y. 2008. Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br J Pharmacol 154: 327-42
9. Kozorovitskiy Y, Saunders A, Johnson CA, Lowell BB, Sabatini BL. 2012. Recurrent network activity drives striatal synaptogenesis. Nature 485: 646-50
10. Stuber GD, Hnasko TS, Britt JP, Edwards RH, Bonci A. 2010. Dopaminergic terminals in the nucleus accumbens but not the dorsal striatum corelease glutamate. J Neurosci 30: 8229-33
11. Tecuapetla F, Patel JC, Xenias H, English D, Tadros I, Shah F, Berlin J, Deisseroth K, Rice ME, Tepper JM, Koos T. 2010. Glutamatergic signaling by mesolimbic dopamine neurons in the nucleus accumbens. J Neurosci 30: 7105-10
12. Bello Gay E., Mateo Y., Gelman D.M., Noaín D., Shin J.H., et. al. (2011) Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D2 autoreceptors. Nature Neuroscience 14: 1033-8.
13. Durieux PF, Bearzatto B, Guiducci S, Buch T, Waisman A, Zoli M, Schiffmann SN, de Kerchove d'Exaerde A. 2009. D2R striatopallidal neurons inhibit both locomotor and drug reward processes. Nat Neurosci 12: 393-5
14. Ferguson SM, Eskenazi D, Ishikawa M, Wanat MJ, Phillips PE, Dong Y, Roth BL, Neumaier JF. 2011. Transient neuronal inhibition reveals opposing roles of indirect and direct pathways in sensitization. Nat Neurosci 14: 22-4
15. Cuzon Carlson V, Seabold GK, Helms CM, Garg N, Odagiri M, et al., (2011) Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates. Neuropsychopharmacology 36: 2513-28.

152
Publications 2011-2012 1. Cui C., Noronha A, Morikawa H., Alvarez VA, Stuber GD, et al. (2012) New Insights on
Neurobiological Mechanisms underlying Alcohol Addiction. Neuropharmacology 67C: 223-232. Review.
2. Bello Gay E., Mateo Y., Gelman D.M., Noaín D., Shin J.H., et. al. (2011) Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D2 autoreceptors. Nature Neuroscience 14: 1033-8.
3. Dobi A., Seabold G.K., Christensen C.H., Bock R. and Alvarez V.A. (2011) Cocaine-induced plasticity in the nucleus accumbens is cell specific and develops without withdrawal. Journal of Neuroscience 31(5):1895-904.
4. Kramer P.F., Christensen C.H., Hazelwood L.A., Dobi A., Bock R., et al. (2011) Dopamine D2 receptor overexpression alters behavior and physiology in Drd2-EGFP mice. Journal of Neuroscience 31(1):126-32. Faculty of 1000 Article factor: FFa/6.
5. Cuzon Carlson V, Seabold GK, Helms CM, Garg N, Odagiri M, et al., (2011) Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates. Neuropsychopharmacology 36: 2513-28.
6. Mathur BN, Capik NA, Alvarez VA, Lovinger DM. (2011) Serotonin induces long-term depression at corticostriatal synapses. Journal of Neuroscience 31(20):7402-11.
7. Sherazee N. and Alvarez VA. (2012) Chapter 29: DiOlisitcs-delivery of fluorescent dyes into cells. In "Biolistic DNA Delivery: Methods and Protocols" Editors: S. Sudowe and AB Reske-Kunz. Publisher: Springer
Collaborators Dr. David Lovinger, Laboratory for Integrative Neuroscience, NIAAA Dr. Andrew Holmes, Laboratory for Integrative Neuroscience, NIAAA Dr. David Sibley, Molecular Neuropharmacology Section, NINDS. Dr. Marcelo Rubinstein, INGEBI, University of Buenos Aires, Argentina Dr. Kathleen A. Grant, Dept Behavioral Neuroscience, Oregon National Primate Research Center Dr. Todd Thiele, Dept. of Psychology, University of North Carolina Dr. Alvarez: [email protected]

153
______________________________________________________________________
Laboratory of Physiologic Studies ______________________________________________________________________ George Kunos, MD, PhD, Chief
The common theme in the Laboratory of Physiologic Studies is the use of in vivo animal models of human pathological states relevant to alcohol use disorders combined with the use of biochemical, cell- and molecular biological tools to explore underlying disease mechanisms and test novel therapeutic paradigms. The Section on Neuroendocrinology conducts studies on the biology of endogenous cannabinoids and their receptors. Work focuses on the role of the endocannabinoid system in the control of energy homeostasis, including energy intake and peripheral energy metabolism, and its role in cardiovascular and behavioral regulation. The work on energy intake aims to explore the role of endocannabinoids in the neuroendocrine control of appetite, including the appetitive aspects of ethanol drinking behavior. Endocannabinoids are also involved in the regulation of peripheral metabolism, and research in this section was first to identify the liver as a primary target of the effect of endocannabinoids on lipid metabolism and steatosis, induced either by high-fat diets or chronic alcohol intake. More recently, our laboratory was the first to document the therapeutic potential of peripherally restricted CB1 receptor antagonists in animal models of visceral obesity. These compounds retain the metabolic efficacy of globally acting (i.e. brain-penetrant) CB1 antagonists in reducing excess body weight and adiposity and reversing insulin resistance, diabetes and hepatic steatosis but, unlike the latter, are devoid of centrally mediated behavioral effects predictive of neuropsychiatric side effects in humans. Among peripheral CB1 antagonists, inverse agonists have greater metabolic efficacy than neutral antagonists, and the peripheral CB1 receptor inverse agonist we characterized is currently undergoing toxicology screening, supported by the NIH BrIDGs program, in preparation for its testing in proof-of- principle clinical trials. Another novel compound introduced by our Laboratory, which increases endocannabinoid tone by blocking the anandamide degrading enzyme fatty acid amide hydrolase (FAAH), was tested in collaboration with Dr. Andrew Holmes’ group (Laboratory of Behavioral and Genomic Neuroscience) and found effective in promoting fear extinction in a rodent model of the post-traumatic stress syndrome. The Section on Oxidative Stress Tissue Injury examines the role of oxidative-nitrosative stress and consequent poly(ADP-ribose) polymerase (PARP) activation in various forms of cardio- vascular pathologies, including those that are induced by chronic alcohol intake or develop as a consequence of aging. The preferred approach is to use a combination of state-of-the-art in vivo methodologies to analyze hemodynamic functions in combination with cell and molecular biological methods to study the expression and function of cellular proteins involved in tissue injury and inflammation, or acting as anti-inflammatory, protective and tissue repair mechanisms. As an example of the latter, the role of endocannabinoids as anti-inflammatory mediators acting via CB2 cannabinoid receptors in peripheral tissues is analyzed in ischemia-reperfusion models of tissue injury.
Dr. Kunos: [email protected]

______________________________________________________________________
Section on Neuroendocrinology ______________________________________________________________________ George Kunos, MD, PhD, Chief Jie Liu, MD, PhD, Medical Officer Judith Harvey-White, BS, Research Chemist Grzegorz Godlewski, PharmD, PhD, Staff Scientist Bani Mukhopadhyay, PhD, Staff Scientist Yossi Tam, DDS, PhD, Staff Scientist Resat Cinar, PhD, Visiting Fellow Tony Jourdan, PhD, Visiting Fellow Gergő Szanda, MD, PhD, Visiting Fellow
The Biology and Functions of the Endocannabinoid System and its Role in Metabolic Regulation
Endocannabinoids are lipid-like mediators discovered in the 1990s that bind to cannabinoid (CB) 1 and 2 receptors to trigger effects similar to those of marijuana1. Endocannabinoids and their CB1 receptors (CB1R) are expressed at high levels in the brain, but are also present in many peripheral tissues2. The Section on Neuroendocrinology, Laboratory of Physiologic Studies, has been focusing on exploring the role of the endocannabinoid system in the regulation of appetitive functions, including caloric intake3 and alcohol drinking behavior4, and in the control of lipid and carbohydrate metabolism with emphasis on their role in obesity and its metabolic complications such as insulin resistance/diabetes, fatty liver and dyslipidemias5,6. Of particular current interest is exploring the role of the peripheral endocannabinoid system in metabolic regulation and developing and testing novel, peripherally restricted CB1R antagonists for the treatment of obesity/metabolic syndrome as well as alcoholic fatty liver. Globally acting (i.e. brain-penetrant) CB1 antagonists showed promise in the treatment of the metabolic syndrome7, but their further therapeutic development was halted in 2008 due to neuropsychiatric side effects8. We and others have shown that CB1R are expressed at low yet functional levels in peripheral tissues, including liver, fat, muscle and endocrine pancreas, where their activation promotes de novo lipogenesis and induces insulin and leptin resistance5,6. Thus, peripherally restricted CB1 antagonists may retain some or most of the metabolic benefit of global CB1 blockade while causing less frequent or no neuropsychiatric side effects. In a proof-of-principle study published in 2010, we demonstrated that a novel, selective, potent, and non-brain-penetrant analog of rimonabant, AM6545, was effective in improving metabolic endpoints in a mouse model of diet-induced obesity/metabolic syndrome (DIO) without causing behavioral changes which result from blockade of CB R in the CNS9
1 . Unlike the inverse agonist rimonabant, AM6545 is a neutral CB1R antagonist. AM6545 was equieffective with rimonabant in reversing hepatic steatosis, but it was less effective in reducing body weight, insulin and leptin resistance, and had minimal to no effect on food intake. To determine if the greater efficacy of rimonabant was due to its additional central actions or to its inverse agonist properties, we tested the peripherally restricted CB1R inverse agonist JD-5037, developed by the small biotech company, Jenrin Discovery, Inc.
154
l to r: R Cinar, Y Tam, J Liu, G Godlewski G Szanda, B Mukhopadhyay, T Jourdan, G Kunos
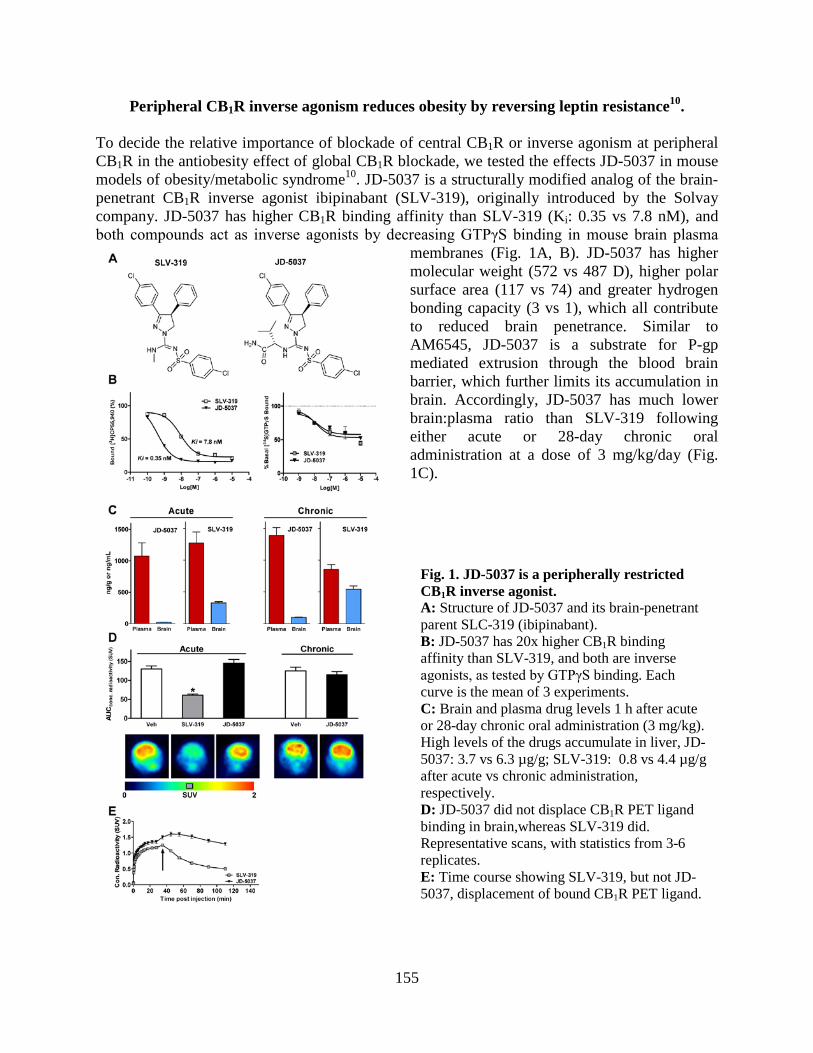
155
Peripheral CB1R inverse agonism reduces obesity by reversing leptin resistance10. To decide the relative importance of blockade of central CB1R or inverse agonism at peripheral CB1R in the antiobesity effect of global CB1R blockade, we tested the effects JD-5037 in mouse models of obesity/metabolic syndrome10. JD-5037 is a structurally modified analog of the brain-penetrant CB1R inverse agonist ibipinabant (SLV-319), originally introduced by the Solvay company. JD-5037 has higher CB1R binding affinity than SLV-319 (Ki: 0.35 vs 7.8 nM), and both compounds act as inverse agonists by decreasing GTPγS binding in mouse brain plasma
membranes (Fig. 1A, B). JD-5037 has higher molecular weight (572 vs 487 D), higher polar surface area (117 vs 74) and greater hydrogen bonding capacity (3 vs 1), which all contribute to reduced brain penetrance. Similar to AM6545, JD-5037 is a substrate for P-gp mediated extrusion through the blood brain barrier, which further limits its accumulation in brain. Accordingly, JD-5037 has much lower brain:plasma ratio than SLV-319 following either acute or 28-day chronic oral administration at a dose of 3 mg/kg/day (Fig. 1C).
Fig. 1. JD-5037 is a peripherally restricted CB1R inverse agonist. A: Structure of JD-5037 and its brain-penetrant parent SLC-319 (ibipinabant). B: JD-5037 has 20x higher CB1R binding affinity than SLV-319, and both are inverse agonists, as tested by GTPγS binding. Each curve is the mean of 3 experiments. C: Brain and plasma drug levels 1 h after acute or 28-day chronic oral administration (3 mg/kg). High levels of the drugs accumulate in liver, JD-5037: 3.7 vs 6.3 µg/g; SLV-319: 0.8 vs 4.4 µg/g after acute vs chronic administration, respectively. D: JD-5037 did not displace CB1R PET ligand binding in brain,whereas SLV-319 did. Representative scans, with statistics from 3-6 replicates. E: Time course showing SLV-319, but not JD-5037, displacement of bound CB1R PET ligand.

156
The free concentration of JD-5037 in brain is further reduced by its very high non-specific brain protein binding of 99.7%. Nevertheless, potential in vivo occupancy of central CB1R was analyzed by CB1 PET. Acute or 28-day chronic pretreatment of DIO mice with 3 mg/kg JD-5037 failed to displace the PET ligand, whereas acute pretreatment with the same dose of SLV-319 caused ~60% displacement (Fig. 1D). This confirmed the absence of functionally relevant levels of JD-5037 in the brain, which was further confirmed by JD-5037 being inactive in behavioral paradigms: it did not inhibit CB1R agonist-induced catalepsy, did not induce hyperambulatory activity, and did not elicit anxiety-like behavior in the elevated plus maze. As illustrated in Fig. 2, oral treatment of male DIO mice with 3 mg/kg/day JD-5037 or SLV-319 for 28 days was equieffective, completely reversing the diet-induced weight gain, reducing adiposity without affecting lean body mass, as assessed by MRI, and caused similar hypophagia that lasted for about two weeks. The two compounds also completely reversed the diet-induced hepatic steatosis and the associated hepatocellular damage, normalized blood glucose and insulin levels, reversed glucose intolerance and insulin resistance, reversed the hyperleptinemia and hypoadiponectinemia, and improved the plasma lipid profile. Indirect calorimetry revealed that CB1R blockade increased total energy expenditure by a selective increase in fat burning, as reflected by the decrease in respiratory quotient. Surprisingly, JD-5037 and SLV-319 caused equal hypophagia, indicating a peripheral mechanism for an effect thought to be centrally regulated. A clue about the potential mechanism came from experiments in leptin deficient ob/ob and leptin receptor defective db/db mice, in both of which JD-5037 treatment failed to affect food intake and body weight, while it still improved hepatic steatosis and glycemic control similar to its effects in DIO mice. This suggested that appetite and weight reduction by CB1R blockade in DIO mice may reflect reversal of the leptin resistance of these animals. This was further supported by the finding that pretreatment of DIO mice with a novel pegylated leptin antagonist greatly attenuated the hypophagic and weight reducing effects of JD-503710. Hyperleptinemia is thought to be responsible for the leptin resistance in DIO11. Normalization of the elevated plasma leptin by JD-5037 treatment was rapid and positively correlated with the parallel decrease in both food intake and body weight. Furthermore, the leptin-induced reduction in food intake, body weight and hypothalamic STAT3 phosphorylation, which are lost in leptin-resistant DIO mice, were restored to responses seen in control mice following chronic oral treatment of DIO mice with JD-5037, 3 mg/kg/day10. The rapid decrease in plasma leptin caused by JD-5037 treatment involves both decreased leptin secretion and increased clearance. CB1R blockade reversed the obesity-induced increase in ob mRNA gene expression in fat tissue, suggesting a direct effect on leptin gene expression. Furthermore, CB1R agonists increased leptin secretion by cultured, differentiated 3T3 L1 adipocytes, and the effects were blocked by 100 nM JD-5037. Blockade of CB1R on sympathetic nerve terminals may also contribute to decreased leptin secretion: norepinephrine acting via β3-adrenergic receptors on adipocytes is known to suppress leptin secretion12,13, and the high-fat diet-induced reduction in adipose tissue norepinephrine levels and β3-receptor expression was reversed by JD-5037 treatment10. .
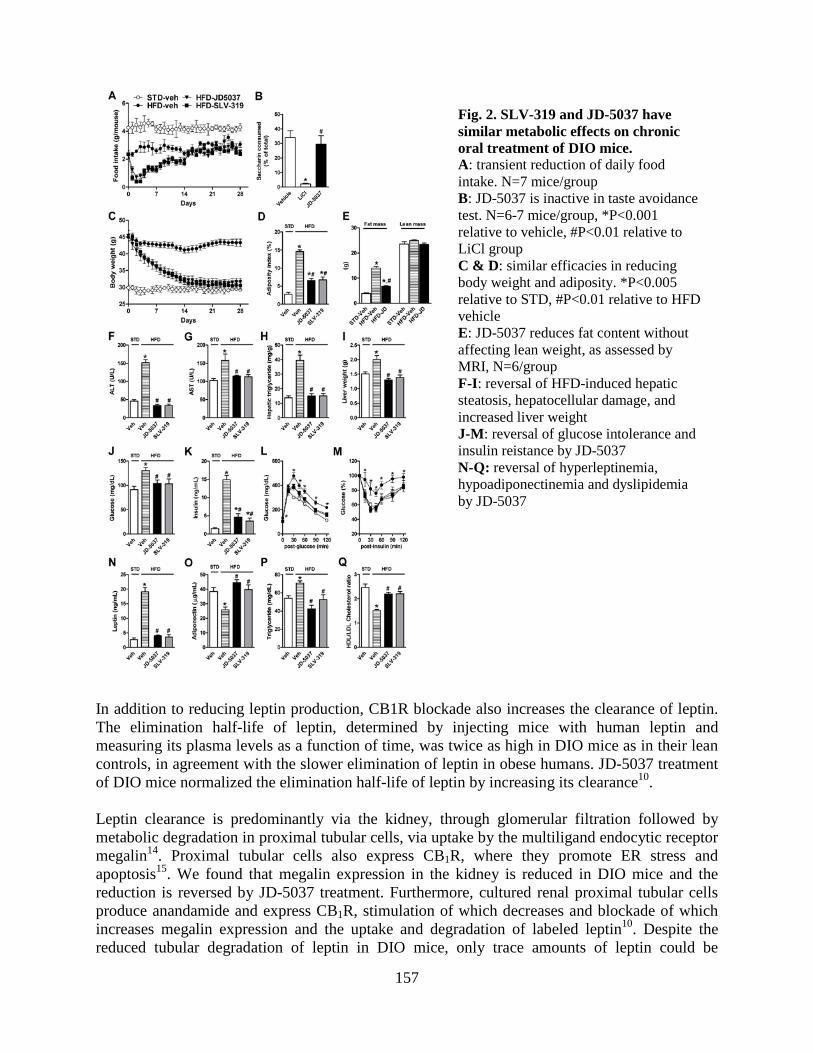
157
In addition to reducing leptin production, CB1R blockade also increases the clearance of leptin. The elimination half-life of leptin, determined by injecting mice with human leptin and measuring its plasma levels as a function of time, was twice as high in DIO mice as in their lean controls, in agreement with the slower elimination of leptin in obese humans. JD-5037 treatment of DIO mice normalized the elimination half-life of leptin by increasing its clearance10.
Leptin clearance is predominantly via the kidney, through glomerular filtration followed by metabolic degradation in proximal tubular cells, via uptake by the multiligand endocytic receptor megalin14. Proximal tubular cells also express CB1R, where they promote ER stress and apoptosis15. We found that megalin expression in the kidney is reduced in DIO mice and the reduction is reversed by JD-5037 treatment. Furthermore, cultured renal proximal tubular cells produce anandamide and express CB1R, stimulation of which decreases and blockade of which increases megalin expression and the uptake and degradation of labeled leptin10. Despite the reduced tubular degradation of leptin in DIO mice, only trace amounts of leptin could be
Fig. 2. SLV-319 and JD-5037 have similar metabolic effects on chronic oral treatment of DIO mice. A: transient reduction of daily food intake. N=7 mice/group B: JD-5037 is inactive in taste avoidance test. N=6-7 mice/group, *P<0.001 relative to vehicle, #P<0.01 relative to LiCl group C & D: similar efficacies in reducing body weight and adiposity. *P<0.005 relative to STD, #P<0.01 relative to HFD vehicle E: JD-5037 reduces fat content without affecting lean weight, as assessed by MRI, N=6/group F-I: reversal of HFD-induced hepatic steatosis, hepatocellular damage, and increased liver weight J-M: reversal of glucose intolerance and insulin reistance by JD-5037 N-Q: reversal of hyperleptinemia, hypoadiponectinemia and dyslipidemia by JD-5037

158
detected in their urine. This suggests that reduced levels of leptin reach the tubules by filtration, and/or unmetabolized leptin may be reabsorbed into the circulation. Glomerular cells have been shown to contain CB1R, stimulation of which causes apoptosis16 and reduced filtration17. We found that increased plasma creatinine levels in DIO mice were normalized by JD-5037 treatment, suggesting that CB1R blockade improves glomerular filtration. Thus, peripheral CB1R blockade promotes leptin clearance in DIO mice by an action at multiple sites in the kidney.
A key finding in this study was the greater efficacy of peripheral CB1R inverse agonism compared to neutral antagonism in reducing food intake and body weight. In agreement with reversal of hyperleptinemia being the cause of these effects, it was only partially reduced by AM6545, but completely reversed by JD-5037. Thus, in addition to increased EC activity in obesity18, the fraction of CB1R in the active conformation may be also increased in peripheral tissues involved in metabolic regulation, to account for the greater efficacy of inverse agonists over neutral antagonists.
In summary, peripherally restricted CB1R inverse agonists have therapeutic potential not only for the metabolic complications of visceral obesity, including diabetes, fatty liver and dyslipidemia, but also in reducing obesity itself. A key component in the mechanism of their antiobesity action is reversal of leptin resistance10.
Hepatic CB1R mediates diet-induced insulin resistance via inhibition of insulin signaling
and clearance19
We have earlier reported that hepatocyte-specific CB1R knockout (hCB1R-/-) mice become obese on HFD but remain insulin sensitive, suggesting the involvement of hepatic CB1R in whole body insulin resistance6. Here we have further explored the role of the liver in diet-induced insulin resistance using a hyperinsulinemic euglycemic clamp. The involvement of CB1R and the signaling pathways they engage were analyzed by comparing glycemic control in wild-type mice with that in mice with genetic deletion of CB1R globally (CB1R-/-) or in hepatocytes only (hCB1R-/-), or in a ‘rescue model’ of global CB1R-/- mice with transgenic re-expression of CB1R in heptocytes only (htgCB1R-/-). The tissue-specific expression profile of CB1R in these 4 strains is illustrated in Fig. 3C. Note that CB1R is overexpressed in the liver of htgCB1R-/- mice to the same extent as in the liver of wild-type mice on HFD6.
To explore the role of endocannabinoids in glucose homeostasis, we analyzed the effects of a single dose of anandamide (10 mg/kg ip) using the intraperitoneal glucose tolerance and insulin sensitivity tests in mice on standard diet. Anandamide caused acute glucose intolerance and insulin resistance in wild-type and htgCB1R-/-, but not in CB1R-/- mice, providing clear evidence that these effects are mediated by hepatic CB1R (Fig. 3A). Interestingly, placing these mice on HFD for 14 weeks produced the same pattern of effects: glucose intolerance and insulin resistance in wild-type and htgCB1R-/- but not in CB1R-/- mice (Fig. 3B). HtgCB1R-/- mice on normal diet had elevated blood glucose and insulin levels relative to their CB1R-/- littermates, while the rest of their hormonal metabolic profile and body composition were similar20. When maintained on HFD, htgCB1R-/- mice remained lean with only a modest increase in liver fat. The similar effects of acute anandamide treatment and chronic HFD further support the notion that diet-induced insulin resistance is mediated by hepatic CB1R.
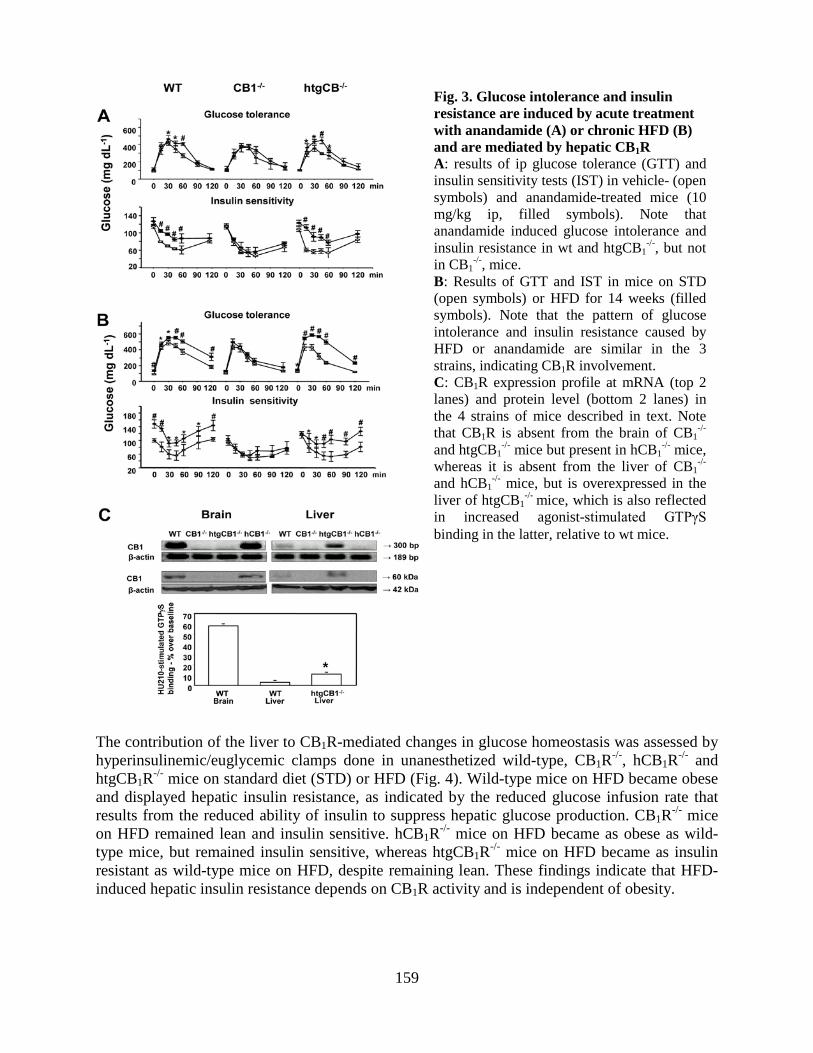
159
Fig. 3. Glucose intolerance and insulin resistance are induced by acute treatment with anandamide (A) or chronic HFD (B) and are mediated by hepatic CB1R A: results of ip glucose tolerance (GTT) and insulin sensitivity tests (IST) in vehicle- (open symbols) and anandamide-treated mice (10 mg/kg ip, filled symbols). Note that anandamide induced glucose intolerance and insulin resistance in wt and htgCB1
-/-, but not in CB1
-/-, mice. B: Results of GTT and IST in mice on STD (open symbols) or HFD for 14 weeks (filled symbols). Note that the pattern of glucose intolerance and insulin resistance caused by HFD or anandamide are similar in the 3 strains, indicating CB1R involvement. C: CB1R expression profile at mRNA (top 2 lanes) and protein level (bottom 2 lanes) in the 4 strains of mice described in text. Note that CB1R is absent from the brain of CB1
-/- and htgCB1
-/- mice but present in hCB1-/- mice,
whereas it is absent from the liver of CB1-/-
and hCB1-/- mice, but is overexpressed in the
liver of htgCB1-/- mice, which is also reflected
in increased agonist-stimulated GTPγS binding in the latter, relative to wt mice.
The contribution of the liver to CB1R-mediated changes in glucose homeostasis was assessed by hyperinsulinemic/euglycemic clamps done in unanesthetized wild-type, CB1R-/-, hCB1R-/- and htgCB1R-/- mice on standard diet (STD) or HFD (Fig. 4). Wild-type mice on HFD became obese and displayed hepatic insulin resistance, as indicated by the reduced glucose infusion rate that results from the reduced ability of insulin to suppress hepatic glucose production. CB1R-/- mice on HFD remained lean and insulin sensitive. hCB1R-/- mice on HFD became as obese as wild-type mice, but remained insulin sensitive, whereas htgCB1R-/- mice on HFD became as insulin resistant as wild-type mice on HFD, despite remaining lean. These findings indicate that HFD-induced hepatic insulin resistance depends on CB1R activity and is independent of obesity.
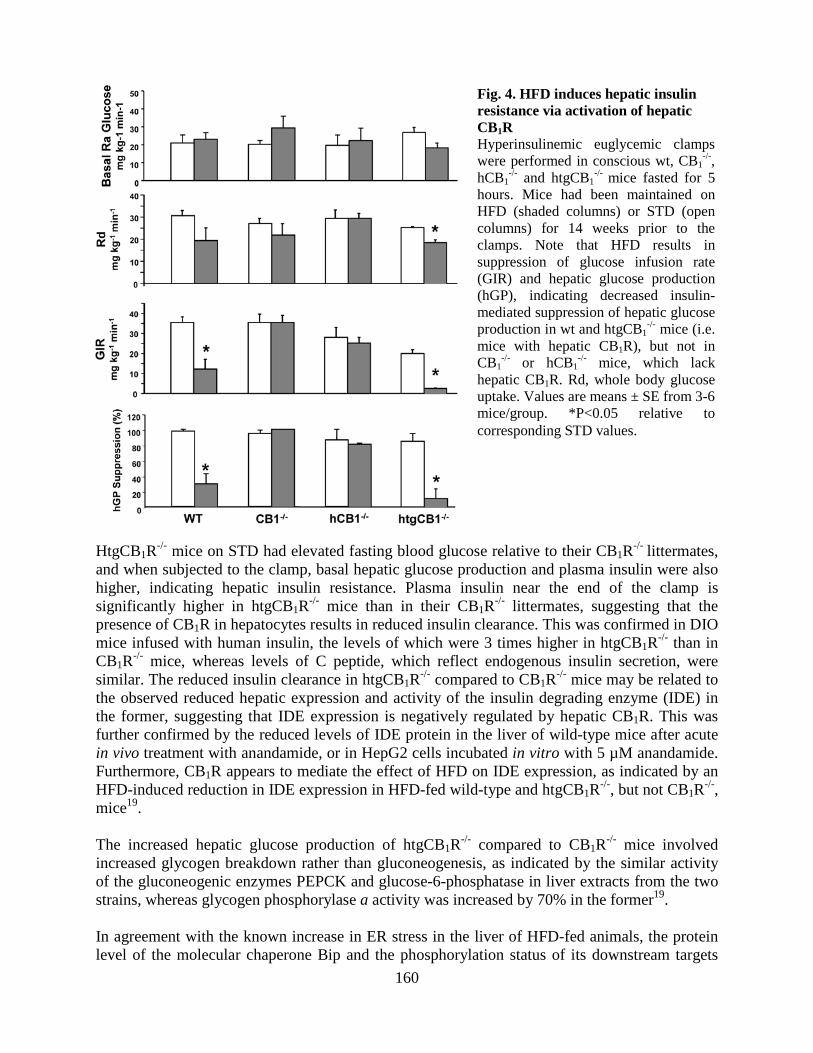
160
HtgCB1R-/- mice on STD had elevated fasting blood glucose relative to their CB1R-/- littermates, and when subjected to the clamp, basal hepatic glucose production and plasma insulin were also higher, indicating hepatic insulin resistance. Plasma insulin near the end of the clamp is significantly higher in htgCB1R-/- mice than in their CB1R-/- littermates, suggesting that the presence of CB1R in hepatocytes results in reduced insulin clearance. This was confirmed in DIO mice infused with human insulin, the levels of which were 3 times higher in htgCB1R-/- than in CB1R-/- mice, whereas levels of C peptide, which reflect endogenous insulin secretion, were similar. The reduced insulin clearance in htgCB1R-/- compared to CB1R-/- mice may be related to the observed reduced hepatic expression and activity of the insulin degrading enzyme (IDE) in the former, suggesting that IDE expression is negatively regulated by hepatic CB1R. This was further confirmed by the reduced levels of IDE protein in the liver of wild-type mice after acute in vivo treatment with anandamide, or in HepG2 cells incubated in vitro with 5 µM anandamide. Furthermore, CB1R appears to mediate the effect of HFD on IDE expression, as indicated by an HFD-induced reduction in IDE expression in HFD-fed wild-type and htgCB1R-/-, but not CB1R-/-, mice19. The increased hepatic glucose production of htgCB1R-/- compared to CB1R-/- mice involved increased glycogen breakdown rather than gluconeogenesis, as indicated by the similar activity of the gluconeogenic enzymes PEPCK and glucose-6-phosphatase in liver extracts from the two strains, whereas glycogen phosphorylase a activity was increased by 70% in the former19. In agreement with the known increase in ER stress in the liver of HFD-fed animals, the protein level of the molecular chaperone Bip and the phosphorylation status of its downstream targets
Fig. 4. HFD induces hepatic insulin resistance via activation of hepatic CB1R Hyperinsulinemic euglycemic clamps were performed in conscious wt, CB1
-/-, hCB1
-/- and htgCB1-/- mice fasted for 5
hours. Mice had been maintained on HFD (shaded columns) or STD (open columns) for 14 weeks prior to the clamps. Note that HFD results in suppression of glucose infusion rate (GIR) and hepatic glucose production (hGP), indicating decreased insulin-mediated suppression of hepatic glucose production in wt and htgCB1
-/- mice (i.e. mice with hepatic CB1R), but not in CB1
-/- or hCB1-/- mice, which lack
hepatic CB1R. Rd, whole body glucose uptake. Values are means ± SE from 3-6 mice/group. *P<0.05 relative to corresponding STD values.

161
PERK and the α subunit of translation initiation factor-2 (eIF2α) were increased in the liver of wild-type mice by HFD or anandamide treatment. Similar changes in the liver of htgCB1R-/-, but not CB1R-/-, mice indicated the role of hepatic CB1R in these effects, which reflect one of the pathways of ER stress suppressing protein translation, whereas ATF6 and XBP1, key components of two other arms of the ER stress response involved in protein folding and degradation, respectively, remained unaffected19. Insulin signaling in the liver is via phosphorylation of protein kinase B/akt-2 which, in turn, has a key role in down-regulating gluconeogenic enzyme expression and increasing glycogen synthase activity. Studies in primary cultured mouse and human hepatocytes indicated that activation of CB1R inhibits insulin-induced phosphorylation of akt-2 by two mechanisms: first, it increases serine-307 phosphorylation of IRS-1, which is known to inhibit the phosphorylation of akt-221; second, it increases the expression of Phllp1, a serine/threonine phosphatase involved in the dephosphorylation of akt-222. The role of ER stress in these effects is indicated by the finding that shRNA knockdown of eIF2α abrogated the ability of CB1R agonists to inhibit akt-2 phosphorylation or to increase Phlpp1 expression. Thus, activation of hepatic CB1R inhibits the antiglycemic action of insulin by inducing ER stress, which inhibits insulin signaling by suppressing p-akt-2 via serine-307 phosphorylation of IRS1 and at the same time promoting the dephosphorylation of akt-2 via activation of Phlpp119. In summary, the findings in this study indicate that activation of hepatic CB1R by endocannabinoids is both necessary and sufficient to account for HFD-induced insulin resistance. Endocannabinoids induce insulin resistance via ER stress-dependent suppression of insulin signaling through p-akt-2 and cause hyperinsulinemia by inhibiting insulin clearance through reducing the expression of the insulin degrading enzyme in the liver. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via CB1R
in the regenerating liver23 The mammalian liver can fully regenerate after tissue loss or injury. Endocannabinoids and CB1R are present in the liver where they promote lipogenesis5, a process also activated during regeneration24, and CB1R activation has been shown to promote neural progenitor cell proliferation25. The ability of endocannabinoids to promote cell proliferation and to stimulate hepatic lipogenesis led us to explore their potential involvement in the early stages of the regenerative response of the liver, using the 2/3 partial hepatectomy (PHX) model of liver regeneration. To explore the role of CB1R in the early regenerative response, proliferative cells were imaged by bromo-deoxyuridine (BrDU) staining 40 h post-PHX. The PHX-induced marked increase in BrDU-positive cells in the remnant livers, as observed in wild-type mice, was severely blunted in CB1R-/- and hCB1R-/- mice, and in wild-type mice treated with the CB1R antagonist rimonabant prior to and following PHX. This suggested a permissive role of hepatic CB1R in the proliferative response. In accordance with such a role, the expression of CB1R was upregulated in the post-PHX liver of wild-type mice. PHX also resulted in a dramatic, ~200-fold increase in anandamide, but not 2-AG, levels in the remnant liver. Anandamide (arachidonoyl ethanolamide) is normally degraded by FAAH, yielding arachidonic acid and ethanolamine. However, treatment of post-PHX mice with the FAAH inhibitor URB597 blunted rather than potentiated the increase in hepatic anandamide, and PHX similarly failed to increase hepatic anandamide levels in FAAH-/- mice. This suggested that in the post-PHX remnant liver, FAAH operates in reverse, synthesizing anandamide by the
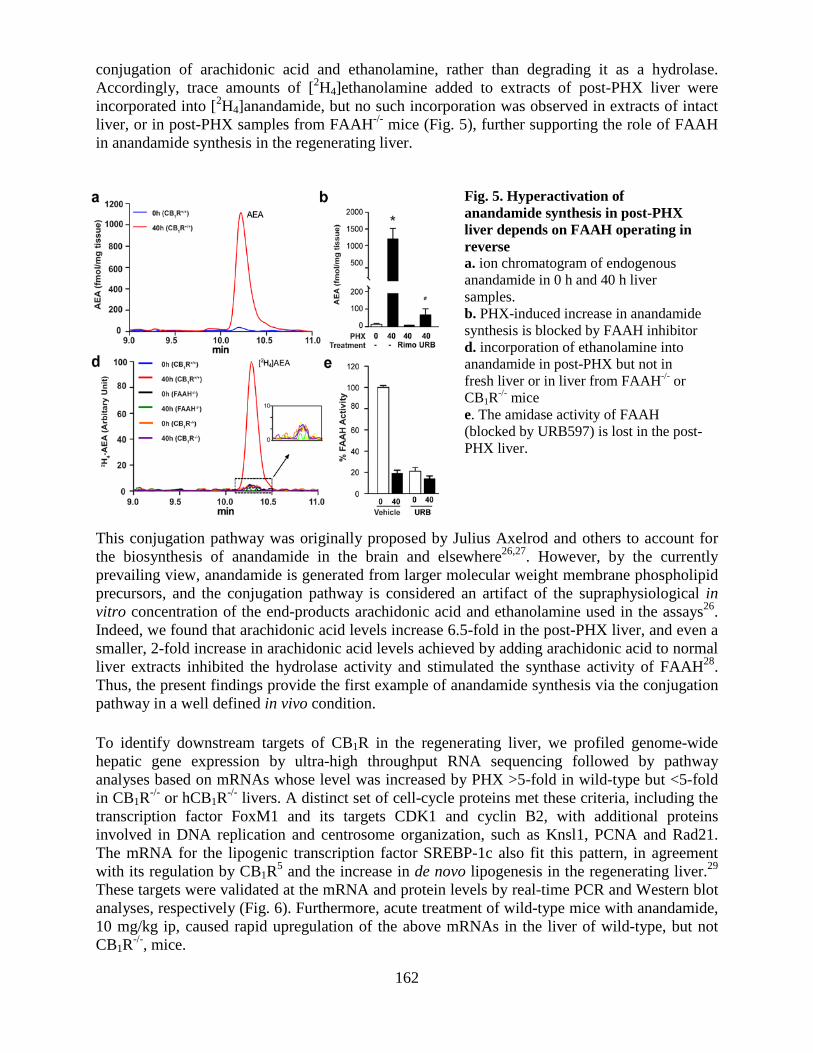
162
conjugation of arachidonic acid and ethanolamine, rather than degrading it as a hydrolase. Accordingly, trace amounts of [2H4]ethanolamine added to extracts of post-PHX liver were incorporated into [2H4]anandamide, but no such incorporation was observed in extracts of intact liver, or in post-PHX samples from FAAH-/- mice (Fig. 5), further supporting the role of FAAH in anandamide synthesis in the regenerating liver.
This conjugation pathway was originally proposed by Julius Axelrod and others to account for the biosynthesis of anandamide in the brain and elsewhere26,27. However, by the currently prevailing view, anandamide is generated from larger molecular weight membrane phospholipid precursors, and the conjugation pathway is considered an artifact of the supraphysiological in vitro concentration of the end-products arachidonic acid and ethanolamine used in the assays26. Indeed, we found that arachidonic acid levels increase 6.5-fold in the post-PHX liver, and even a smaller, 2-fold increase in arachidonic acid levels achieved by adding arachidonic acid to normal liver extracts inhibited the hydrolase activity and stimulated the synthase activity of FAAH28. Thus, the present findings provide the first example of anandamide synthesis via the conjugation pathway in a well defined in vivo condition.
To identify downstream targets of CB1R in the regenerating liver, we profiled genome-wide hepatic gene expression by ultra-high throughput RNA sequencing followed by pathway analyses based on mRNAs whose level was increased by PHX >5-fold in wild-type but <5-fold in CB1R-/- or hCB1R-/- livers. A distinct set of cell-cycle proteins met these criteria, including the transcription factor FoxM1 and its targets CDK1 and cyclin B2, with additional proteins involved in DNA replication and centrosome organization, such as Knsl1, PCNA and Rad21. The mRNA for the lipogenic transcription factor SREBP-1c also fit this pattern, in agreement with its regulation by CB1R5 and the increase in de novo lipogenesis in the regenerating liver.29 These targets were validated at the mRNA and protein levels by real-time PCR and Western blot analyses, respectively (Fig. 6). Furthermore, acute treatment of wild-type mice with anandamide, 10 mg/kg ip, caused rapid upregulation of the above mRNAs in the liver of wild-type, but not CB1R-/-, mice.
Fig. 5. Hyperactivation of anandamide synthesis in post-PHX liver depends on FAAH operating in reverse a. ion chromatogram of endogenous anandamide in 0 h and 40 h liver samples. b. PHX-induced increase in anandamide synthesis is blocked by FAAH inhibitor d. incorporation of ethanolamine into anandamide in post-PHX but not in fresh liver or in liver from FAAH-/- or CB1R-/- mice e. The amidase activity of FAAH (blocked by URB597) is lost in the post-PHX liver.
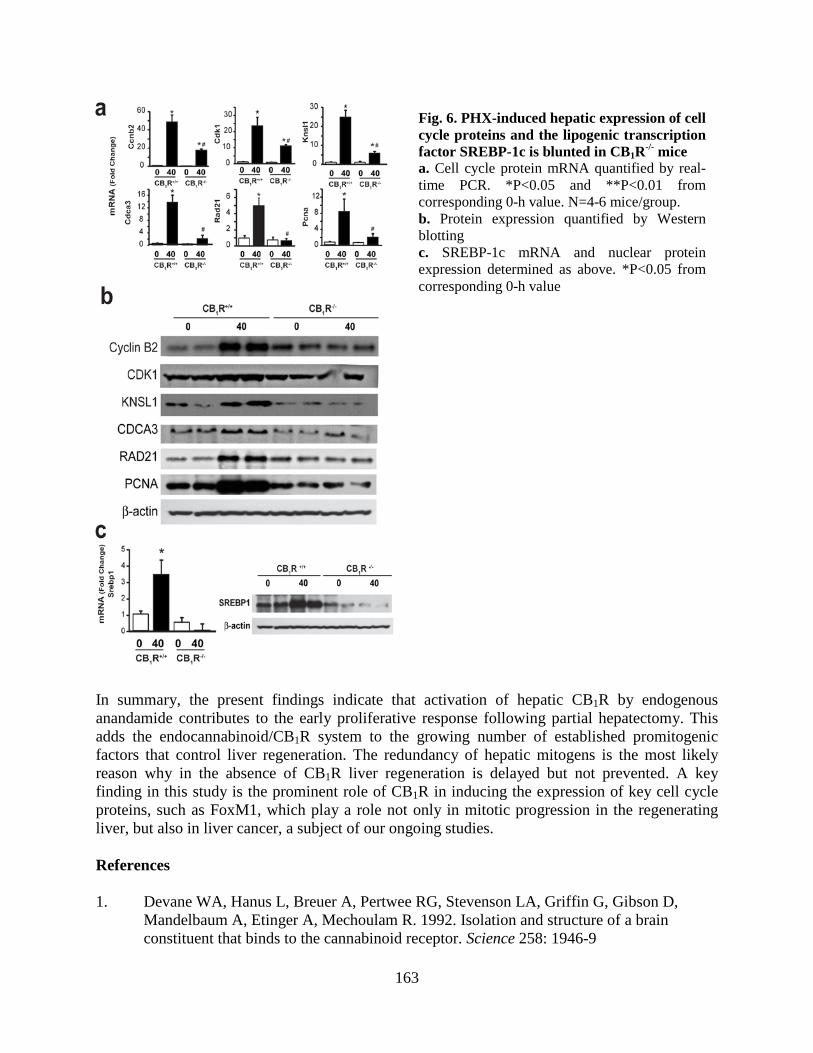
163
In summary, the present findings indicate that activation of hepatic CB1R by endogenous anandamide contributes to the early proliferative response following partial hepatectomy. This adds the endocannabinoid/CB1R system to the growing number of established promitogenic factors that control liver regeneration. The redundancy of hepatic mitogens is the most likely reason why in the absence of CB1R liver regeneration is delayed but not prevented. A key finding in this study is the prominent role of CB1R in inducing the expression of key cell cycle proteins, such as FoxM1, which play a role not only in mitotic progression in the regenerating liver, but also in liver cancer, a subject of our ongoing studies. References 1. Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D,
Mandelbaum A, Etinger A, Mechoulam R. 1992. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258: 1946-9
Fig. 6. PHX-induced hepatic expression of cell cycle proteins and the lipogenic transcription factor SREBP-1c is blunted in CB1R-/- mice a. Cell cycle protein mRNA quantified by real-time PCR. *P<0.05 and **P<0.01 from corresponding 0-h value. N=4-6 mice/group. b. Protein expression quantified by Western blotting c. SREBP-1c mRNA and nuclear protein expression determined as above. *P<0.05 from corresponding 0-h value

164
2. Pacher P, Batkai S, Kunos G. 2006. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev 58: 389-462
3. Di Marzo V, Goparaju SK, Wang L, Liu J, Batkai S, Jarai Z, Fezza F, Miura GI, Palmiter RD, Sugiura T, Kunos G. 2001. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature 410: 822-5
4. Wang L, Liu J, Harvey-White J, Zimmer A, Kunos G. 2003. Endocannabinoid signaling via cannabinoid receptor 1 is involved in ethanol preference and its age-dependent decline in mice. Proc Natl Acad Sci U S A 100: 1393-8
5. Osei-Hyiaman D, DePetrillo M, Pacher P, Liu J, Radaeva S, Batkai S, Harvey-White J, Mackie K, Offertaler L, Wang L, Kunos G. 2005. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J Clin Invest 115: 1298-305
6. Osei-Hyiaman D, Liu J, Zhou L, Godlewski G, Harvey-White J, Jeong WI, Batkai S, Marsicano G, Lutz B, Buettner C, Kunos G. 2008. Hepatic CB(1) receptor is required for development of diet-induced steatosis, dyslipidemia, and insulin and leptin resistance in mice. J Clin Invest 118: 3160-9
7. Despres JP, Golay A, Sjostrom L. 2005. Effects of rimonabant on metabolic risk factors in overweight patients with dyslipidemia. N Engl J Med 353: 2121-34
8. Le Foll B, Gorelick DA, Goldberg SR. 2009. The future of endocannabinoid-oriented clinical research after CB1 antagonists. Psychopharmacology (Berl) 205: 171-4
9. Tam J, Vemuri VK, Liu J, Batkai S, Mukhopadhyay B, Godlewski G, Osei-Hyiaman D, Ohnuma S, Ambudkar SV, Pickel J, Makriyannis A, Kunos G. 2010. Peripheral CB1 cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J Clin Invest 120: 2953-66
10. Tam J, Cinar R, Liu J, Godlewski G, Wesley D, Jourdan T, Szanda G, Mukhopadhyay B, Chedester L, Liow JS, Innis RB, Cheng K, Rice KC, Deschamps JR, Chorvat RJ, McElroy JF, Kunos G. 2012. Peripheral cannabinoid-1 receptor inverse agonism reduces obesity by reversing leptin resistance. Cell Metab 16: 167-79
11. Knight ZA, Hannan KS, Greenberg ML, Friedman JM. 2010. Hyperleptinemia is required for the development of leptin resistance. PLoS One 5: e11376
12. Gettys TW, Harkness PJ, Watson PM. 1996. The beta 3-adrenergic receptor inhibits insulin-stimulated leptin secretion from isolated rat adipocytes. Endocrinology 137: 4054-7
13. Mantzoros CS, Qu D, Frederich RC, Susulic VS, Lowell BB, Maratos-Flier E, Flier JS. 1996. Activation of beta(3) adrenergic receptors suppresses leptin expression and mediates a leptin-independent inhibition of food intake in mice. Diabetes 45: 909-14
14. Ceccarini G, Flavell RR, Butelman ER, Synan M, Willnow TE, Bar-Dagan M, Goldsmith SJ, Kreek MJ, Kothari P, Vallabhajosula S, Muir TW, Friedman JM. 2009. PET imaging of leptin biodistribution and metabolism in rodents and primates. Cell Metab 10: 148-59

165
15. Lim JC, Lim SK, Han HJ, Park SH. 2010. Cannabinoid receptor 1 mediates palmitic acid-induced apoptosis via endoplasmic reticulum stress in human renal proximal tubular cells. J Cell Physiol 225: 654-63
16. Barutta F, Corbelli A, Mastrocola R, Gambino R, Di Marzo V, Pinach S, Rastaldi MP, Perin PC, Gruden G. 2010. Cannabinoid receptor 1 blockade ameliorates albuminuria in experimental diabetic nephropathy. Diabetes 59: 1046-54
17. Koura Y, Ichihara A, Tada Y, Kaneshiro Y, Okada H, Temm CJ, Hayashi M, Saruta T. 2004. Anandamide decreases glomerular filtration rate through predominant vasodilation of efferent arterioles in rat kidneys. J Am Soc Nephrol 15: 1488-94
18. Bennetzen MF, Wellner N, Ahmed SS, Ahmed SM, Diep TA, Hansen HS, Richelsen B, Pedersen SB. 2011. Investigations of the human endocannabinoid system in two subcutaneous adipose tissue depots in lean subjects and in obese subjects before and after weight loss. Int J Obes (Lond) 35: 1377-84
19. Liu J, Zhou L, Xiong K, Godlewski G, Mukhopadhyay B, Tam J, Yin S, Gao P, Shan X, Pickel J, Bataller R, O'Hare J, Scherer T, Buettner C, Kunos G. 2012. Hepatic cannabinoid receptor-1 mediates diet-induced insulin resistance via inhibition of insulin signaling and clearance in mice. Gastroenterology 142: 1218-28
20. Liu J, Zhou L, Xiong K, Godlewski G, Mukhopadhyay B, Tam J, Yin S, Gao P, Shan X, Pickel J, Bataller R, O'Hare J, Scherer T, Buettner C, Kunos G. 2012. Hepatic Cannabinoid Receptor-1 Mediates Diet-Induced Insulin Resistance via Inhibition of Insulin Signaling and Clearance in Mice. Gastroenterology 142: 1218-28 e1
21. Hotamisligil GS, Peraldi P, Budavari A, Ellis R, White MF, Spiegelman BM. 1996. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science 271: 665-8
22. Gao T, Furnari F, Newton AC. 2005. PHLPP: a phosphatase that directly dephosphorylates Akt, promotes apoptosis, and suppresses tumor growth. Mol Cell 18: 13-24
23. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, Godlewski G, Harvey-White J, Mordi I, Cravatt BF, Lotersztajn S, Gao B, Yuan Q, Schuebel K, Goldman D, Kunos G. 2011. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proc Natl Acad Sci U S A 108: 6323-8
24. Schofield PS, Sugden MC, Corstorphine CG, Zammit VA. 1987. Altered interactions between lipogenesis and fatty acid oxidation in regenerating rat liver. Biochem J 241: 469-74
25. Molina-Holgado E, Vela JM, Arevalo-Martin A, Almazan G, Molina-Holgado F, Borrell J, Guaza C. 2002. Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/Akt signaling. J Neurosci 22: 9742-53

166
26. Devane WA, Axelrod J. 1994. Enzymatic synthesis of anandamide, an endogenous ligand for the cannabinoid receptor, by brain membranes. Proc Natl Acad Sci U S A 91: 6698-701
27. Deutsch DG, Chin SA. 1993. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem Pharmacol 46: 791-6
28. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, Godlewski G, Harvey-White J, Mordi I, Cravatt BF, Lotersztajn S, Gao B, Yuan Q, Schuebel K, Goldman D, Kunos G. 2011. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proc Natl Acad Sci U S A
29. Gove CD, Hems DA. 1978. Fatty acid synthesis in the regenerating liver of the rat. Biochem J 170: 1-8
Publications 2011-2012 1. Heilig M, Warren KR, Kunos G, Silverman PB, Hewitt BG. 2011. Addiction Research
Centres and the Nurturing of Creativity. Addiction 106: 1052-60
2. Thanos PK, Gopez V, Delis F, Michaelides M, Grandy DK, Wang GJ, Kunos G, Volkow ND. 2011. Upregulation of Cannabinoid Type 1 Receptors in Dopamine D2 Receptor Knockout Mice Is Reversed by Chronic Forced Ethanol Consumption. Alcoholism-Clinical and Experimental Research 35: 19-27
3. Tam J, Liu J, Mukhopadhyay B, Cinar R, Godlewski G, Kunos G. 2011.
Endocannabinoids in Liver Disease. Hepatology 53: 346-55 (Invited Review)
4. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, Godlewski G, Harvey-White J, Mordi I, Cravatt BF, Lotersztajn S, Gao B, Yuan QP, Schuebel K, Goldman D, Kunos G. 2011. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proceedings of the National Academy of Sciences of the United States of America 108: 6323-8
Subject of editorial, PNAS 108:6339-40, 2011.
5. Kunos G, Tam J. 2011. The case for peripheral CB1 receptor blockade in the treatment of visceral obesity and its cardiometabolic complications. British Journal of Pharmacology 163: 1423-31
6. Bashashati M, Storr MA, Nikas SP, Wood JT, Godlewski G, Liu J, Ho W, Keenan CM,
Zhang H, Alapafuja SO, Cravatt BF, Lutz B, Mackie K, Kunos G, Patel KD, Makriyannis A, Davison JS, Sharkey KA. 2012. Inhibiting fatty acid amide hydrolase normalizes endotoxin-induced enhanced gastrointestinal motility in mice. British Journal of Pharmacology 165: 1556-71
7. Seif T, Makriyannis A, Kunos G, Bonci A, Hopf FW. 2011. The Endocannabinoid 2-
Arachidonoylglycerol Mediates D1 and D2 Receptor Cooperative Enhancement of Rat Nucleus Accumbens Core Neuron Firing. Neuroscience 193: 21-33

167
8. Liu J, Zhou L, Xiong KM, Godlewski G, Mukhopadhyay B, Tam J, Yin S, Gao P, Shan X, Pickel J, Bataller R, O'Hare J, Scherer T, Buettner C, Kunos G. 2012. Hepatic Cannabinoid Receptor-1 Mediates Diet-Induced Insulin Resistance via Inhibition of Insulin Signaling and Clearance in Mice. Gastroenterology 142: 1218-28
Subject of editorial on page 1063, also in ‘Covering the Cover’, p. 1049 of same issue. Evaluated by Faculty of 1000: Stieger B: 2012. F1000.com/716247804#eval791552904
9. Tam J, Cinar R, Liu J, Godlewski G, Wesley D, Jourdan T, Szanda G, Mukhopadhyay B, Chedester L, Liow JS, Innis RB, Cheng KJ, Rice KC, Deschamps JR, Chorvat RJ, McElroy JF, Kunos G. 2012. Peripheral Cannabinoid-1 Receptor Inverse Agonism Reduces Obesity by Reversing Leptin Resistance. Cell Metabolism 16: 167-79 Subject of press release by journal; highlighted in Nature Rev Endocrinology; Nature Rev Drug Discovery.
10. Gunduz-Cinar O, Martin KP, Cinar R, Sugden K, Williams B, Gorka AX, Manuck SB,
Makriyannis A, Poulton R, Hariri AR, Caspi A, Moffitt TE, Kunos G, Holmes A. 2012. Enhanced endocannabinoids and reduced fear in mouse and man. Molecular Psychiatry
June 12, 2012. doi:10.1038/mp.2012.72 [epub ahead of print] Evaluated by Faculty of 1000.
11. Engeli S, Bluher M, Jumpertz R, Wiesner T, Wirtz H, Bosse-Henck A, Stumvoll M, Batkai S, Pacher P, Harvey-White J, Kunos G, Jordan J. 2012. Circulating anandamide and blood pressure in patients with obstructive sleep apnea. Journal of Hypertension 30: 2345-51

168
Collaborators
Markus Heilig, MD,PhD, Laboratory of Clnical and Translational Studies, NIAAA Bin Gao, MD, PhD, Laboratory of Liver Diseases, NIAAA Pal Pacher, MD, PhD, Section on Oxydative Stress and Tissue Injury, LPS, NIAAA Peter Thanos, PhD, Laboratory of Neuroimaging, NIAAA Nora Volkow, MD, Laboratory of Neuroimaging, NIAAA James Pickel, PhD, Laboratory of Genetics, NIMH Antonello Bonci, MD, Scientific Director, NIDA Andrew Holmes, PhD, Laboratory of Behavioral and Genomic Neuroscience, NIAAA Robert B. Innis, MD, Molecular Imaging Branch, NIMH Monica Skarulis, MD, Clinical Endocrine Section, NIIDDK Marilyn Huestis, PhD, Chemistry and Drug Metabolism Section, NIDA David Goldman, MD, Laboratory of Neurogenetics, NIAAA Kornel Schuebel, PhD, Laboratory of Neurogenetics, NIAAA Christoph Buettner, MD, Mount Sinai School of Medicine, New York, NY Keith Sharkey, PhD, University of Calgary, Calgary, Canada Samuel S Lee, MD, University of Calgary, Calgary, Canada Benjamin F Cravatt, PhD, Scripps Institute, La Jolla, CA Alexandros Makriyannis, PhD, Northeastern University, Boston, MA Sophie Lotersztajn, PhD, INSERM, Creteil, France Jeffery Deschamps, PhD, Naval Research Labooratory, Washington DC Ramon Bataller, MD, University of Barcelona, Spain Woody Hopf, PhD, University of California at San Francisco, CA Stefan Engeli, MD, University of Hanover, Hanover, Germany Dr. Kunos: [email protected]

169
______________________________________________________________________
Section on Oxidative Stress – Tissue Injury ______________________________________________________________________
Pal Pacher, MD, PhD, Chief Partha Mukhopadhyay, PhD, Staff Scientist Zongxian Cao, MD., PhD, Postdoctoral IRTA Katalin Erdelyi, PhD, Postdoctoral IRTA
Mechanisms of Oxidative-Nitrosative Stress and Inflammation-Induced Tissue Injury Overwhelming evidence suggests that oxidative-nitrosative/nitrative stress and inflammation areinvolved in essentially all major pathologicalprocesses affecting humans, including those inducedby excessive alcohol consumption. The research focusof SOSTI is to understand the cellular and molecular mechanisms underlying oxidative/nitrosative/nitrative stress, inflammation, and their downstream effector pathways using clinically relevant animal models of disease (e.g., ischemia reperfusion injury, cardiomyopathy/heart failure), and to identify novel therapeutic targets against these pathologies. Interplay of oxidative-nitrosative/nitrative stress and poly(ADP-ribose) polymerase in tissue injury (e.g., cardiovascular pathophysiology, ischemia/reperfusion injury, shock, nephropathy and diabetic complications): cellular and molecular mechanisms Oxidative-nitrosative stress and consequent poly(ADP-ribose) polymerase (PARP) activation are key events in the development of tissue injury and organ dysfunction in virtually all disease states affecting humans (Fig. 1; Pacher et al. 2002, 2003, 2007; Moon et al. 2008; Mukhopadhyay et al. 2009; Stasch et al. 2011). Novel drug candidates targeting this pathway are entering or being evaluated in clinical trials for a variety of diseases associated with tissue injury and inflammation, as well as in cancer. In collaboration with Drs. Kalyanaraman, Zsengeller and Stillman we have recently studied the potential role of the mitochondrial ROS generation-PARP-1 pathway in the pro-inflammatory response associated with hepatic ischemia/reperfusion injury and nephropathy induced by a widely used chemotherapeutic drug cisplatin. These studies demonstrated that mitochondrially generated oxidants and PARP-1 activation play a central role in triggering the deleterious cascade of events associated with liver and kidney injury, which could be targeted by novel antioxidants or PARP inhibitors for therapeutic advantage (Mukhopadhyay et al. 2011a; 2012a,b; Zsengeller et al. 2012). We have also shown that treatment of mice with sulforaphan, a natural constituent of broccoli and other cruciferous vegetables, prevented or markedly attenuated the cisplatin-induced oxidative/nitrosative stress, inflammation, and cell death in the kidney, and improved renal function, suggesting promising new protective strategies against cisplatin-induced nephrotoxicity (Guerrero-Beltrán et al. 2012). These results are exciting, because PARP inhibitors are in clinical trials to treat various malignancies and sulforaphan has been reported to exert anticancer effects in preclinical models
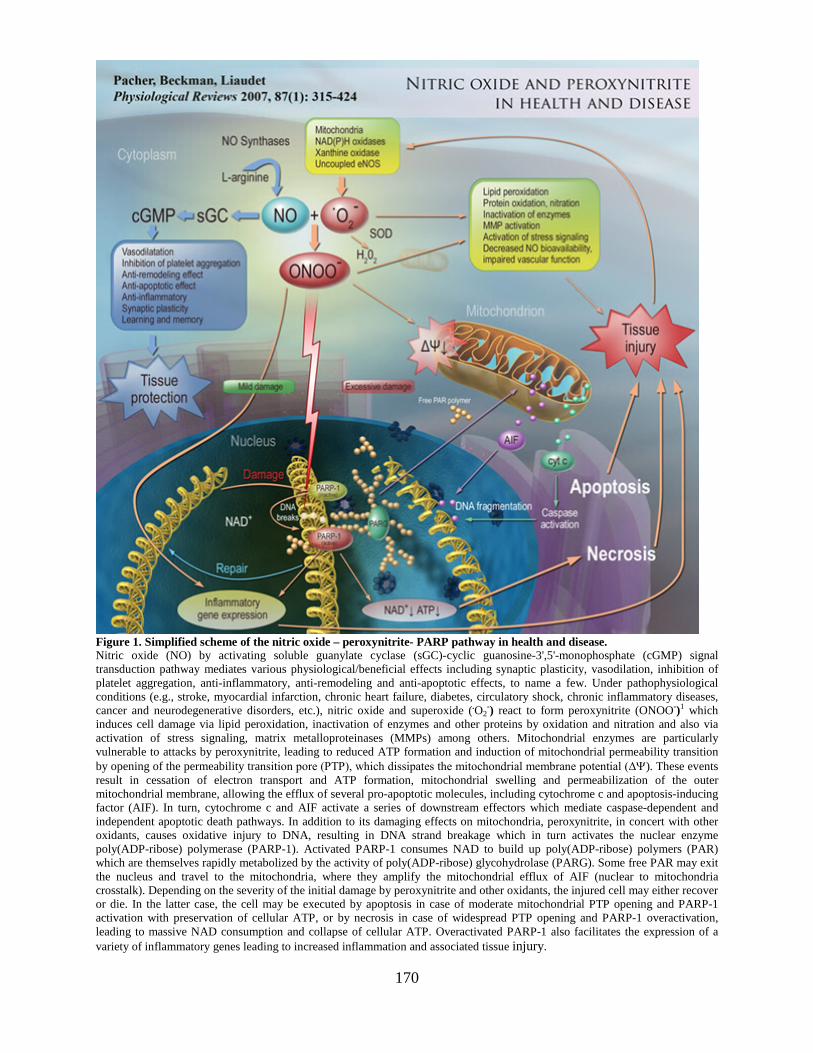
170
Figure 1. Simplified scheme of the nitric oxide – peroxynitrite- PARP pathway in health and disease. Nitric oxide (NO) by activating soluble guanylate cyclase (sGC)-cyclic guanosine-3',5'-monophosphate (cGMP) signal transduction pathway mediates various physiological/beneficial effects including synaptic plasticity, vasodilation, inhibition of platelet aggregation, anti-inflammatory, anti-remodeling and anti-apoptotic effects, to name a few. Under pathophysiological conditions (e.g., stroke, myocardial infarction, chronic heart failure, diabetes, circulatory shock, chronic inflammatory diseases, cancer and neurodegenerative disorders, etc.), nitric oxide and superoxide (.O2
-) react to form peroxynitrite (ONOO-)1 which induces cell damage via lipid peroxidation, inactivation of enzymes and other proteins by oxidation and nitration and also via activation of stress signaling, matrix metalloproteinases (MMPs) among others. Mitochondrial enzymes are particularly vulnerable to attacks by peroxynitrite, leading to reduced ATP formation and induction of mitochondrial permeability transition by opening of the permeability transition pore (PTP), which dissipates the mitochondrial membrane potential (ΔΨ). These events result in cessation of electron transport and ATP formation, mitochondrial swelling and permeabilization of the outer mitochondrial membrane, allowing the efflux of several pro-apoptotic molecules, including cytochrome c and apoptosis-inducing factor (AIF). In turn, cytochrome c and AIF activate a series of downstream effectors which mediate caspase-dependent and independent apoptotic death pathways. In addition to its damaging effects on mitochondria, peroxynitrite, in concert with other oxidants, causes oxidative injury to DNA, resulting in DNA strand breakage which in turn activates the nuclear enzyme poly(ADP-ribose) polymerase (PARP-1). Activated PARP-1 consumes NAD to build up poly(ADP-ribose) polymers (PAR) which are themselves rapidly metabolized by the activity of poly(ADP-ribose) glycohydrolase (PARG). Some free PAR may exit the nucleus and travel to the mitochondria, where they amplify the mitochondrial efflux of AIF (nuclear to mitochondria crosstalk). Depending on the severity of the initial damage by peroxynitrite and other oxidants, the injured cell may either recover or die. In the latter case, the cell may be executed by apoptosis in case of moderate mitochondrial PTP opening and PARP-1 activation with preservation of cellular ATP, or by necrosis in case of widespread PTP opening and PARP-1 overactivation, leading to massive NAD consumption and collapse of cellular ATP. Overactivated PARP-1 also facilitates the expression of a variety of inflammatory genes leading to increased inflammation and associated tissue injury.

171
High-mobility group box 1 (HMGB1) is a nuclear protein actively secreted by immune cells and passively released by necrotic cells that initiates pro-inflammatory signaling through binding to the receptor for advance glycation end-products. HMGB1 has been established as a key inflammatory mediator during myocardial infarction, but the proximal mechanisms responsible for myocardial HMGB1 expression and release in this setting remain unclear. In collaboration with Dr. Liaudet we have recently investigated the possible involvement of peroxynitrite, a potent cytotoxic oxidant formed during myocardial infarction, in these processes. Our findings indicated that peroxynitrite is a key mediator of HMGB1 overexpression and release by cardiac cells which provide a novel mechanism linking myocardial oxidative/nitrative stress with post-infarction myocardial inflammation (Loukili et al. 2011).
In collaboration with Dr. Hasko we have explored the role of A2A/B adenosine receptors in alternative macrophage activation (Hasko et al. 2008; Csoka et al. 2012; Hasko and Pacher 2012) and IL-10 production in microglia cells (Kocso et al. 2012), as well as the role of ecto-5’-nucleotidase (CD73) in organ injury associated with sepsis (Hasko e al. 2011; Szabo and Pacher 2012). We found that adenosine promoted alternative macrophage activation via A2A and A2B receptors, and augmented IL-10 production by microglia cells through A2B receptor. We also found that CD73 decreased mortality and organ injury in a model of sepsis. Interplay of oxidative/nitrative stress, inflammation with the endocannabinoid system (ES), and natural plant derived cannabinoids in tissue injury. During the last couple of years the main focus of SOSTI has been to understand the interplay of oxidative/nitrosative stress and inflammation with the novel lipid signaling endocannabinoid system (ES; an emerging very promising therapeutic target against various inflammatory and other diseases; Pacher et al. 2006)). We have explored the role of the ES in tissue injury associated with ischemia/reperfusion (I/R), heart failure and various in vitro and in vivo models of cardiovascular inflammation, nephropathy, as well as other pathologies associated with inflammation and tissue injury. These studies have established a new concept that oxidative/nitrosative /nitrative stress is involved in the activation of the ES, and that modulation of peripheral CB2 cannabinoid receptors protects against I/R-induced tissue injury and/or vascular inflammation by decreasing endothelial cell activation, inflammatory response and reactive oxygen species (ROS) generation (Batkai et al. 2007; Rajesh et al. 2007; Pacher and Mechoulam 2011; Figure 2). We have also discovered an important signaling role of CB2 receptors in human coronary artery endothelial and vascular smooth muscle cells (Rajesh et al. 2007; Pacher and Mechoulam, 2011). The CB2 receptor is a very appealing target for drug development for various inflammatory diseases and ischemia reperfusion injury, in which these receptors on immune cells attenuate inflammatory response and tissue injury (Pacher and Mechoulam 2011). However, most currently available CB2 agonists have problems related to their long term stability, in vivo efficacy, and their potential modulatory effects on CB1 signaling at higher doses, which often result in opposing cellular consequences on inflammation when the dose is increased (bell-shaped dose-response relationship with many cannabinergic ligands has been reported). We have aimed with Professor Raphael Mechoulam at Hebrew University to develop better leads with improved in vivo efficacy and selectivity. One of the best compounds from this new series, HU910, was recently characterized in detail in vitro, and was proven to be considerably better in its in vivo efficacy in models of hepatic I/R injury compared to other CB2 agonists (Horvath et al. 2012a). During these studies we unexpectedly revealed an important function of CB2

172
receptors in Kupffer cells in regulating inflammatory responses in the liver (Horvath et al. 2012a). In collaboration with Professor Pertwee we have discovered that delta(8)-tetrahydrocannabivarin delta(8)-THCV), a stable synthetic analog of the plant-derived cannabinoid delta(9)-tetrahydrocannabivarin, which exerts some anti-inflammatory effects in rodents, is a potent CB2 agonist both in vitro and in vivo with anti-inflammatory effects in hepatic I/R injury (Batkai et al. 2012). We hope that these new ligands may serve as a basis for the development of a whole new class of novel compounds with anti-inflammatory activities. Endothelial and smooth muscle cell activation and inflammatory response play a pivotal role in the development of atherosclerosis and restenosis following vascular surgery. Our previous in vitro studies have identified opposing functional regulatory role of CB1/2 receptors on human vascular smooth muscle proliferation and migration triggered by inflammatory mediators and/or mitogens (Rajesh et al. 2007, 2010; Pacher and Mechoulam, 2011). In collaboration with Dr. Sabine Steffens we have recently discovered that CB2 receptors play an important role in limiting the pathological vascular smooth muscle proliferation and macrophage infiltration during vascular injury (Molica et al. 2012). These results suggest that CB2 receptor agonists may be beneficial in attenuating pathological vascular remodeling following coronary bypass surgery (Steffens and Pacher, 2012). Our recent studies also suggest that certain purified plant (not marijuana) derived oils which are widely used in alternative medicine, contain powerful anti-inflammatory compounds which are potent natural CB2 agonists. In collaboration with Dr. Gertsch we recently found that β-caryophyllene (BCP), a natural sesquiterpene found in many essential oils of spice, dose-dependently ameliorated cisplatin-induced kidney dysfunction, morphological damage, and renal inflammatory response induced by cisplatin (Mukhopadhyay et al. 2010; Horvath et al. 2012b). It also markedly mitigated oxidative/nitrative stress and cell death. The protective effects of BCP against biochemical and histological markers of nephropathy were absent in CB2 knockout mice (Horvath et al. 2012b). Thus, BCP may be an excellent therapeutic agent to prevent cisplatin-induced nephrotoxicity through a CB2 receptor-dependent pathway. Given the excellent safety profile of BCP in humans it has tremendous therapeutic potential in a multitude of diseases associated with inflammation and oxidative stress. There is accumulating evidence indicating that certain non-psychoactive plant-derived natural cannabinoids such as cannabidiol (CBD) may exert potent protective effects independent from classical cannabinoid 1 and 2 receptors. Previously we showed that CBD attenuated high glucose-induced endothelial cell activation and barrier disruption, which are crucial early events underlying the development of various diabetic complications and atherosclerosis, and protected mice from the development of diabetic cardiomyopathy (Rajesh et al. 2010). Recently we found that CBD, significantly reduced the extent of liver inflammation, oxidative/nitrative stress, and cell death associated with hepatic I/R (Mukhopadhyay et al. 2011c). CBD also attenuated the bacterial endotoxin-triggered NF-kappaB activation and TNF-alpha production in isolated Kupffer cells in vitro, likewise the adhesion molecule expression in human liver sinusoidal endothelial cells stimulated with TNF-alpha, and attachment of human neutrophils to the activated endothelium. These protective effects were preserved in CB2 knockout mice and were not prevented by CB1/2 antagonists in vitro (Mukhopadhyay, 2011c).
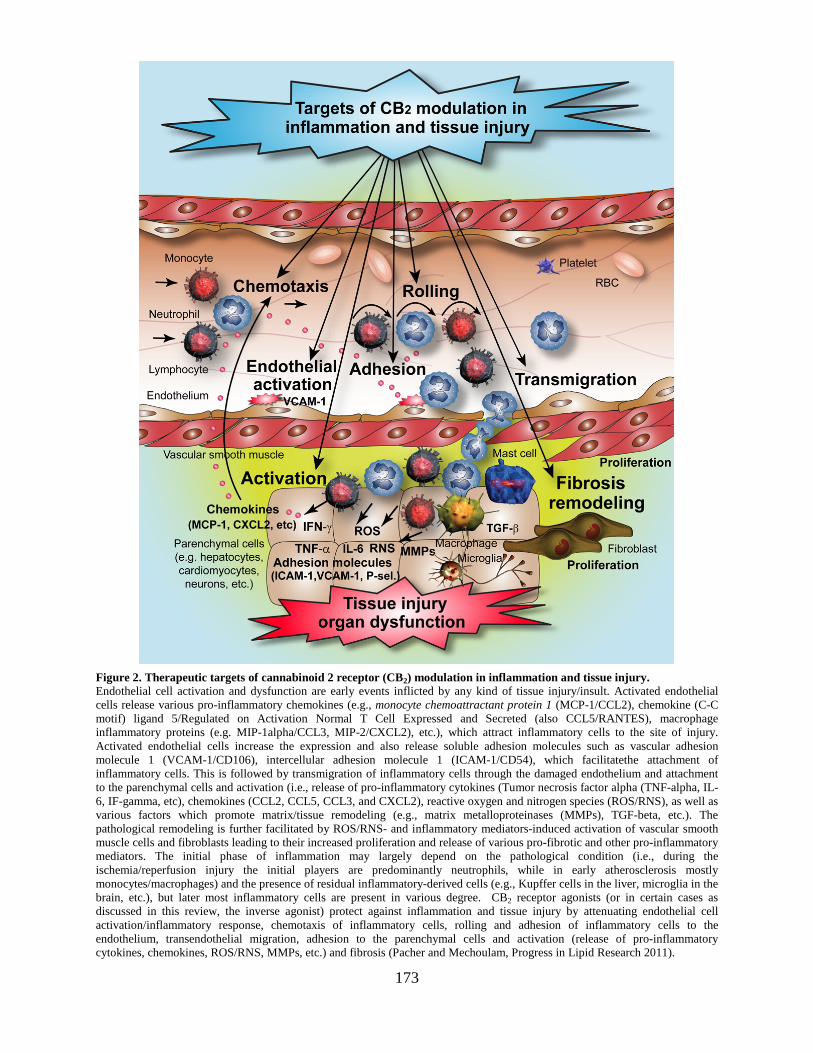
173
Figure 2. Therapeutic targets of cannabinoid 2 receptor (CB2) modulation in inflammation and tissue injury. Endothelial cell activation and dysfunction are early events inflicted by any kind of tissue injury/insult. Activated endothelial cells release various pro-inflammatory chemokines (e.g., monocyte chemoattractant protein 1 (MCP-1/CCL2), chemokine (C-C motif) ligand 5/Regulated on Activation Normal T Cell Expressed and Secreted (also CCL5/RANTES), macrophage inflammatory proteins (e.g. MIP-1alpha/CCL3, MIP-2/CXCL2), etc.), which attract inflammatory cells to the site of injury. Activated endothelial cells increase the expression and also release soluble adhesion molecules such as vascular adhesion molecule 1 (VCAM-1/CD106), intercellular adhesion molecule 1 (ICAM-1/CD54), which facilitatethe attachment of inflammatory cells. This is followed by transmigration of inflammatory cells through the damaged endothelium and attachment to the parenchymal cells and activation (i.e., release of pro-inflammatory cytokines (Tumor necrosis factor alpha (TNF-alpha, IL-6, IF-gamma, etc), chemokines (CCL2, CCL5, CCL3, and CXCL2), reactive oxygen and nitrogen species (ROS/RNS), as well as various factors which promote matrix/tissue remodeling (e.g., matrix metalloproteinases (MMPs), TGF-beta, etc.). The pathological remodeling is further facilitated by ROS/RNS- and inflammatory mediators-induced activation of vascular smooth muscle cells and fibroblasts leading to their increased proliferation and release of various pro-fibrotic and other pro-inflammatory mediators. The initial phase of inflammation may largely depend on the pathological condition (i.e., during the ischemia/reperfusion injury the initial players are predominantly neutrophils, while in early atherosclerosis mostly monocytes/macrophages) and the presence of residual inflammatory-derived cells (e.g., Kupffer cells in the liver, microglia in the brain, etc.), but later most inflammatory cells are present in various degree. CB2 receptor agonists (or in certain cases as discussed in this review, the inverse agonist) protect against inflammation and tissue injury by attenuating endothelial cell activation/inflammatory response, chemotaxis of inflammatory cells, rolling and adhesion of inflammatory cells to the endothelium, transendothelial migration, adhesion to the parenchymal cells and activation (release of pro-inflammatory cytokines, chemokines, ROS/RNS, MMPs, etc.) and fibrosis (Pacher and Mechoulam, Progress in Lipid Research 2011).

174
Cardiovascular complications are the most common cause of morbidity and mortality in diabetic patients. Diabetic vascular dysfunction is a major clinical problem which can lead to retinopathy, nephropathy, neuropathy and increased risk of stroke, hypertension and myocardial infarction. The presence of myocardial left ventricular dysfunction (both diastolic and later systolic) independent of atherosclerosis, coronary artery disease in diabetes, defined as ‘diabetic cardiomyopathy’, has been well documented in both human beings and animals. Our previous studies suggested that cannabinoid 1 receptor (CB1) activation by endocannabinoid anandamide (AEA) in human coronary endothelial cells and cardiomyocytes may amplify the reactive oxygen species (ROS)-MAPK activation-cell death pathway in pathological conditions when the endocannabinoid biosynthetic or metabolic pathways are dysregulated by excessive inflammation and/or oxidative/nitrosative stress, thereby contributing to the development of endothelial dysfunction and pathophysiology of multiple cardiovascular diseases (Mukhopadhyay et al. 2007a; Rajesh et al. 2010). To test the in vivo relevance of the above findings in endothelial cells and cardiomyocytes, we investigated the effects of genetic or pharmacological inhibition of CB1 in a clinically relevant diabetic retinopathy and cardiomyopathy model (El-Remessy et al. 2011; Rajesh et al. 2012). Deletion of CB1 or treatment of diabetic mice with CB1 antagonist SR141716 almost completely prevented retinal cell death. Treatment of diabetic mice or human primary retinal endothelial cells (HREC) exposed to high glucose with CB1 antagonist SR141716 attenuated the oxidative and nitrative stress, reduced nuclear factor kappa B activation, and adhesion molecules expression. In addition, SR141716 attenuated the diabetes- or high glucose-induced proapoptotic activation of MAPKs and retinal vascular cell death (El-Remessy et al. 2011). We also discovered novel interactions of cardiovascular CB1 with angiotensin II receptor type 1 AT1R and its downstream signaling via p47(phox)/ROS-generating NADPH enzyme isoform, advanced glycation end product (AGE)-receptor (RAGE), and other pro-inflammatory/pro-oxidant signaling pathways in diabetic cardiovascular tissues, which may underlie the development of multiple diabetic complications (Rajesh et al. 2012). We found that diabetic cardiomyopathy was characterized by increased myocardial endocannabinoid anandamide levels, oxidative/nitrative stress, activation of p38/Jun MAPKs, enhanced inflammation, increased expression of CB1, RAGE, AT1R, p47(phox) NADPH oxidase subunit, β-myosin heavy chain isozyme switch, accumulation of AGE, fibrosis, and decreased expression of sarcoplasmic/endoplasmic reticulum Ca2+-ATPase. Pharmacological inhibition or genetic deletion of CB1 attenuated the diabetes-induced cardiac dysfunction, measured by pressure-volume approach (Pacher et al. 2008), and the above-mentioned pathological alterations (Rajesh et al. 2012). To further explore the potential mechanisms of AEA-induced cardiovascular injury and potential interplay of the endocannabinoid signaling with the oxidative/nitrative stress, we have investigated the role of the AEA-metabolizing enzyme, fatty acid amide hydrolase (FAAH) using cardiomyopathy models induced by Doxorubicin (DOX) in mice. The DOX-induced myocardial oxidative/nitrative stress positively correlated with multiple cell death markers, which were enhanced in FAAH knockout mice exhibiting significantly increased DOX-induced mortality and cardiac dysfunction compared to their wild type littermates. The effects of DOX in FAAH knockouts were partially attenuated by CB1 receptor antagonists (Mukhopadhyay et al. 2011b). Furthermore, anandamide enhanced cell death in human cardiomyocytes pretreated with FAAH inhibitor and enhanced sensitivity to ROS generation in inflammatory cells of FAAH knockouts (Mukhopadhyay et al. 2011b).
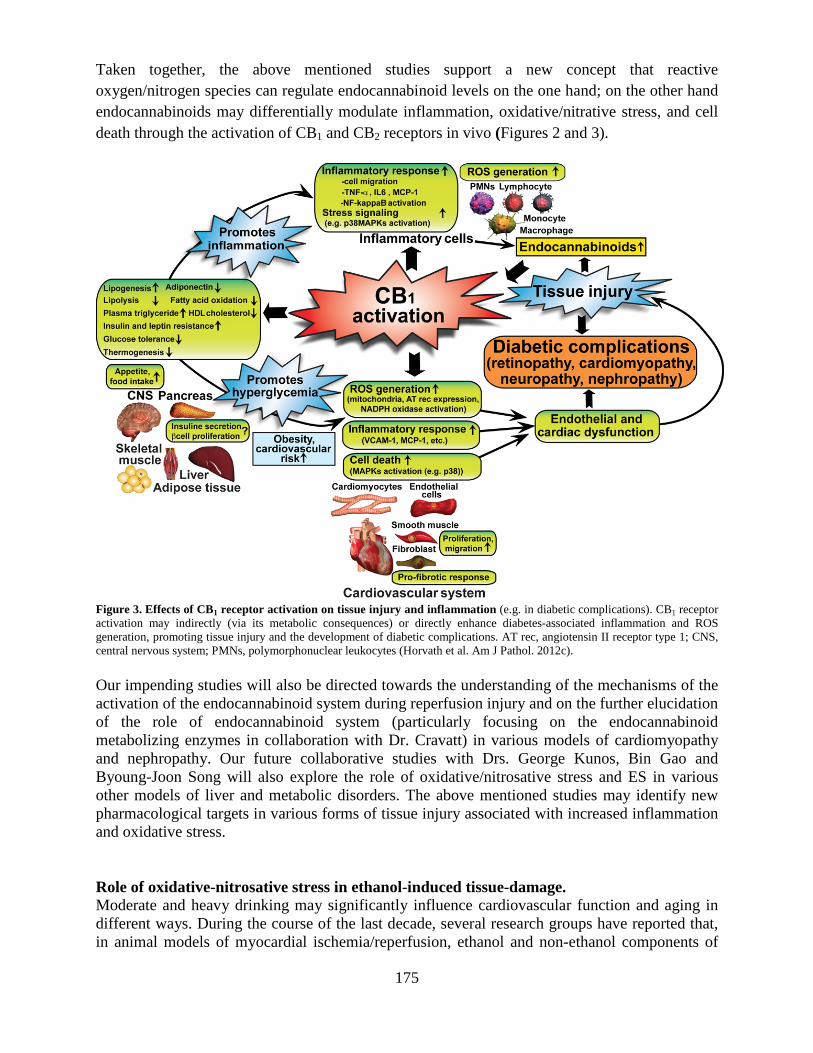
175
Taken together, the above mentioned studies support a new concept that reactive oxygen/nitrogen species can regulate endocannabinoid levels on the one hand; on the other hand endocannabinoids may differentially modulate inflammation, oxidative/nitrative stress, and cell death through the activation of CB1 and CB2 receptors in vivo (Figures 2 and 3).
Figure 3. Effects of CB1 receptor activation on tissue injury and inflammation (e.g. in diabetic complications). CB1 receptor activation may indirectly (via its metabolic consequences) or directly enhance diabetes-associated inflammation and ROS generation, promoting tissue injury and the development of diabetic complications. AT rec, angiotensin II receptor type 1; CNS, central nervous system; PMNs, polymorphonuclear leukocytes (Horvath et al. Am J Pathol. 2012c). Our impending studies will also be directed towards the understanding of the mechanisms of the activation of the endocannabinoid system during reperfusion injury and on the further elucidation of the role of endocannabinoid system (particularly focusing on the endocannabinoid metabolizing enzymes in collaboration with Dr. Cravatt) in various models of cardiomyopathy and nephropathy. Our future collaborative studies with Drs. George Kunos, Bin Gao and Byoung-Joon Song will also explore the role of oxidative/nitrosative stress and ES in various other models of liver and metabolic disorders. The above mentioned studies may identify new pharmacological targets in various forms of tissue injury associated with increased inflammation and oxidative stress. Role of oxidative-nitrosative stress in ethanol-induced tissue-damage. Moderate and heavy drinking may significantly influence cardiovascular function and aging in different ways. During the course of the last decade, several research groups have reported that, in animal models of myocardial ischemia/reperfusion, ethanol and non-ethanol components of

176
wine may have a specific protective effect on the myocardium, independent of the classical risk factors implicated in vascular atherosclerosis and thrombosis. Apoptosis is a mechanism of cell death implicated in the pathogenesis of alcohol-induced organ damage. Experimental studies have suggested alcohol-mediated apoptosis in cardiac muscle, and there is also evidence of skeletal muscle apoptosis in long-term high-dose alcohol drinkers. Apoptosis is present to a similar degree in the heart muscle of high-dose alcohol drinkers and long-standing hypertensive subjects and is related to structural damage. Resveratrol, a polyphenolic antioxidant, is a putative wine constituent to provide an explanation for the phenomenon known as the “French paradox” (French people suffer a relatively low incidence of coronary heart disease presumably because of the consumption of red wine). We have previously demonstrated that resveratrol induced mitochondrial biogenesis in coronary artery endothelial cells and attenuated mitochondrial oxidative stress. We have also showed that resveratrol attenuated azidothymidine (an important anti-retroviral drug to treat HIV)-induced cardiotoxicity by decreasing mitochondrial reactive oxygen species generation in human cardiomyocytes (Gao et al. 2011). Our recent studies are also focused on the understanding of the mechanisms of ethanol-induced oxidative/nitrosative stress and apoptosis in the cardiovascular system and also in other organ systems. Using a clinically relevant model of aging (Fisher rats developed by the National Institute on Aging) we try to address the effects of ethanol on the course of oxidative/nitrosative stress and inflammation associated with aging. Through collaboration with Drs. Emanuel Rubin and Gyorgy Hajnoczky at Dept. of Pathology, Thomas Jefferson University, we will be able to access unique tissue samples from alcoholics to investigate the role of oxidative stress and inflammatory signaling in alcohol-induced myocardial and skeletal muscle injury. Our ongoing studies are also focused on the understanding of the mechanisms of ethanol-induced oxidative/nitrative stress, inflammation and cell death in the cardiovascular system and in the liver during pathological processes (e.g. associated with aging). Prior key references
Batkai S, Osei-Hyiaman D, Pan H, El-Assal O, Rajesh M, et al. 2007. Cannabinoid-2 receptor mediates protection against hepatic ischemia/reperfusion injury. FASEB J 21:1788-800
Hasko G, Linden J, Cronstein B, Pacher P. 2008. Adenosine receptors: therapeutic aspects for inflammatory and immune diseases. Nat Rev Drug Discov 7:759-70
Moon KH, Hood BL, Mukhopadhyay P, Rajesh M, Abdelmegeed MA, et al. 2008. Oxidative inactivation of key mitochondrial proteins leads to dysfunction and injury in hepatic ischemia reperfusion. Gastroenterology 135:1344-57
Mukhopadhyay P, Batkai S, Rajesh M, Czifra N, Harvey-White J, et al. 2007a. Pharmacological inhibition of CB1 cannabinoid receptor protects against doxorubicin-induced cardiotoxicity. J Am Coll Cardiol 50:528-36
Mukhopadhyay P, Rajesh M, Batkai S, Kashiwaya Y, Hasko G, et al. 2009. Role of superoxide, nitric oxide, and peroxynitrite in doxorubicin-induced cell death in vivo and in vitro. Am J Physiol-Heart and Circulatory Physiology 296:H1466-H83

177
Mukhopadhyay P, Rajesh M, Hasko G, Hawkins BJ, Madesh M, Pacher P. 2007b. Simultaneous detection of apoptosis and mitochondrial superoxide production in live cells by flow cytometry and confocal microscopy. Nat Protoc 2:2295-301
Mukhopadhyay P, Rajesh M, Pan H, Patel V, Mukhopadhyay B, et al. 2010. Cannabinoid-2 receptor limits inflammation, oxidative/nitrosative stress, and cell death in nephropathy. Free Radical Biology and Medicine 48:457-67
Pacher P, Batkai S, Kunos G. 2006. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev 58:389-462
Pacher P, Beckman JS, Liaudet L. 2007. Nitric oxide and peroxynitrite in health and disease. Physiol Rev 87:315-424
Pacher P, Liaudet L, Bai P, Mabley JG, Kaminski PM, et al. 2003. Potent metalloporphyrin peroxynitrite decomposition catalyst protects against the development of doxorubicin-induced cardiac dysfunction. Circulation 107:896-904
Pacher P, Liaudet L, Soriano FG, Mabley JG, Szabo E, Szabo C. 2002. The role of poly(ADP-ribose) polymerase activation in the development of myocardial and endothelial dysfunction in diabetes. Diabetes 51:514-21
Pacher P, Nagayama T, Mukhopadhyay P, Batkai S, Kass DA. 2008. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat Protoc 3:1422-34
Rajesh M, Mukhopadhyay P, Batkai S, Hasko G, Liaudet L, et al. 2007. CB2-receptor stimulation attenuates TNF-alpha-induced human endothelial cell activation, transendothelial migration of monocytes, and monocyte-endothelial adhesion. Am J Physiol Heart Circ Physiol 293:H2210-8
Rajesh M, Mukhopadhyay P, Batkai S, Patel V, Saito K, et al. 2010. Cannabidiol attenuates cardiac dysfunction, oxidative stress, fibrosis, and inflammatory and cell death signaling pathways in diabetic cardiomyopathy. J Am Coll Cardiol 56:2115-25
Rajesh M, Mukhopadhyay P, Hasko G, Liaudet L, Mackie K, Pacher P. 2010. Cannabinoid-1 receptor activation induces reactive oxygen species-dependent and -independent mitogen-activated protein kinase activation and cell death in human coronary artery endothelial cells. British Journal of Pharmacology 160:688-700
Publications 2011-2012: 1. Chandra R, Federici S, Nemeth ZH, Horvath B, Pacher P, Hasko G, Deitch EA, Spolarics
Z. 2011. Female X-chromosome mosaicism for NOX2 deficiency presents unique inflammatory phenotype and improves outcome in polymicrobial sepsis. J Immunol 186: 6465-73
2. Drel VR, Pacher P, Stavniichuk R, Xu W, Zhang J, Kuchmerovska TM, Slusher B,
Obrosova IG. 2011. Poly(ADP-ribose)polymerase inhibition counteracts renal

178
hypertrophy and multiple manifestations of peripheral neuropathy in diabetic Akita mice. Int J Mol Med 28: 629-35
3. El-Remessy AB, Rajesh M, Mukhopadhyay P, Horvath B, Patel V, Al-Gayyar MM, Pillai
BA, Pacher P. 2011. Cannabinoid 1 receptor activation contributes to vascular inflammation and cell death in a mouse model of diabetic retinopathy and a human retinal cell line. Diabetologia 54: 1567-78
4. Gao RY, Mukhopadhyay P, Mohanraj R, Wang H, Horvath B, Yin S, Pacher P. 2011.
Resveratrol attenuates azidothymidine-induced cardiotoxicity by decreasing mitochondrial reactive oxygen species generation in human cardiomyocytes. Mol Med Report 4: 151-5
5. Hasko G, Csoka B, Koscso B, Chandra R, Pacher P, Thompson LF, Deitch EA, Spolarics
Z, Virag L, Gergely P, Rolandelli RH, Nemeth ZH. 2011. Ecto-5'-nucleotidase (CD73) decreases mortality and organ injury in sepsis. J Immunol 187: 4256-67
6. Hasko G, Pacher P. 2011. Suppression of tumorigenicity 2: a janus-faced player in sepsis.
Am J Respir Crit Care Med 183: 841-3 7. Koscso B, Csoka B, Pacher P, Hasko G. 2011. Investigational A(3) adenosine receptor
targeting agents. Expert Opin Investig Drugs 20: 757-68 8. Loukili N, Rosenblatt-Velin N, Li J, Clerc S, Pacher P, Feihl F, Waeber B, Liaudet L.
2011. Peroxynitrite induces HMGB1 release by cardiac cells in vitro and HMGB1 upregulation in the infarcted myocardium in vivo. Cardiovasc Res 89: 586-94
9. Mabley J, Gordon S, Pacher P. 2011. Nicotine exerts an anti-inflammatory effect in a
murine model of acute lung injury. Inflammation 34: 231-7 10. Mukhopadhyay P, Horvath B, Kechrid M, Tanchian G, Rajesh M, Naura AS, Boulares
AH, Pacher P. 2011a. Poly(ADP-ribose) polymerase-1 is a key mediator of cisplatin-induced kidney inflammation and injury. Free Radic Biol Med 51: 1774-88
11. Mukhopadhyay P, Horvath B, Rajesh M, Matsumoto S, Saito K, Batkai S, Patel V,
Tanchian G, Gao RY, Cravatt BF, Hasko G, Pacher P. 2011b. Fatty acid amide hydrolase is a key regulator of endocannabinoid-induced myocardial tissue injury. Free Radic Biol Med 50: 179-95
12. Mukhopadhyay P, Rajesh M, Horvath B, Batkai S, Park O, Tanchian G, Gao RY, Patel
V, Wink DA, Liaudet L, Hasko G, Mechoulam R, Pacher P. 2011c. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radic Biol Med 50: 1368-81
13. Pacher P, Mechoulam R. 2011. Is lipid signaling through cannabinoid 2 receptors part of
a protective system? Prog Lipid Res 50: 193-211 14. Soriano FG, Lorigados CB, Pacher P, Szabo C. 2011. Effects of a potent peroxynitrite
decomposition catalyst in murine models of endotoxemia and sepsis. Shock 35: 560-6

179
15. Stasch JP, Pacher P, Evgenov OV. 2011. Soluble guanylate cyclase as an emerging
therapeutic target in cardiopulmonary disease. Circulation 123: 2263-73 16. Szebeni A, Szentandrassy N, Pacher P, Simko J, Nanasi PP, Kecskemeti V. 2011. Can
the electrophysiological action of rosiglitazone explain its cardiac side effects? Curr Med Chem 18: 3720-8
17. Watcho P, Stavniichuk R, Tane P, Shevalye H, Maksimchyk Y, Pacher P, Obrosova IG.
2011. Evaluation of PMI-5011, an ethanolic extract of Artemisia dracunculus L., on peripheral neuropathy in streptozotocin-diabetic mice. Int J Mol Med 27: 299-307
18. Balamayooran G, Batra S, Theivanthiran B, Cai S, Pacher P, Jeyaseelan S. 2012.
Intrapulmonary G-CSF rescues neutrophil recruitment to the lung and neutrophil release to blood in gram-negative bacterial infection in MCP-1-/- mice. J Immunol 189: 5849-59
19. Batkai S, Mukhopadhyay P, Horvath B, Rajesh M, Gao RY, Mahadevan A, Amere M,
Battista N, Lichtman AH, Gauson LA, Maccarrone M, Pertwee RG, Pacher P. 2012. Delta8-Tetrahydrocannabivarin prevents hepatic ischaemia/reperfusion injury by decreasing oxidative stress and inflammatory responses through cannabinoid CB2 receptors. Br J Pharmacol 165: 2450-61
20. Cai S, Batra S, Wakamatsu N, Pacher P, Jeyaseelan S. 2012. NLRC4 inflammasome-
mediated production of IL-1beta modulates mucosal immunity in the lung against gram-negative bacterial infection. J Immunol 188: 5623-35
21. Csoka B, Selmeczy Z, Koscso B, Nemeth ZH, Pacher P, Murray PJ, Kepka-Lenhart D,
Morris SM, Jr., Gause WC, Leibovich SJ, Hasko G. 2012. Adenosine promotes alternative macrophage activation via A2A and A2B receptors. FASEB J 26: 376-86
22. Engeli S, Bluher M, Jumpertz R, Wiesner T, Wirtz H, Bosse-Henck A, Stumvoll M,
Batkai S, Pacher P, Harvey-White J, Kunos G, Jordan J. 2012. Circulating anandamide and blood pressure in patients with obstructive sleep apnea. J Hypertens 30: 2345-51
23. Guerrero-Beltran CE, Mukhopadhyay P, Horvath B, Rajesh M, Tapia E, Garcia-Torres I,
Pedraza-Chaverri J, Pacher P. 2012. Sulforaphane, a natural constituent of broccoli, prevents cell death and inflammation in nephropathy. J Nutr Biochem 23: 494-500
24. Hasko G, Pacher P. 2012. Regulation of macrophage function by adenosine. Arterioscler
Thromb Vasc Biol 32: 865-9 25. Horvath B, Magid L, Mukhopadhyay P, Batkai S, Rajesh M, Park O, Tanchian G, Gao
RY, Goodfellow CE, Glass M, Mechoulam R, Pacher P. 2012a. A new cannabinoid CB2 receptor agonist HU-910 attenuates oxidative stress, inflammation and cell death associated with hepatic ischaemia/reperfusion injury. Br J Pharmacol 165: 2462-78
26. Horvath B, Mukhopadhyay P, Hasko G, Pacher P. 2012c. The endocannabinoid system
and plant-derived cannabinoids in diabetes and diabetic complications. Am J Pathol 180: 432-42

180
27. Horvath B, Mukhopadhyay P, Kechrid M, Patel V, Tanchian G, Wink DA, Gertsch J,
Pacher P. 2012b. beta-Caryophyllene ameliorates cisplatin-induced nephrotoxicity in a cannabinoid 2 receptor-dependent manner. Free Radic Biol Med 52: 1325-33
28. Koscso B, Csoka B, Selmeczy Z, Himer L, Pacher P, Virag L, Hasko G. 2012. Adenosine
augments IL-10 production by microglial cells through an A2B adenosine receptor-mediated process. J Immunol 188: 445-53
29. Lupachyk S, Stavniichuk R, Komissarenko JI, Drel VR, Obrosov AA, El-Remessy AB,
Pacher P, Obrosova IG. 2012. Na+/H+-exchanger-1 inhibition counteracts diabetic cataract formation and retinal oxidative-nitrative stress and apoptosis. Int J Mol Med 29: 989-98
30. Molica F, Matter CM, Burger F, Pelli G, Lenglet S, Zimmer A, Pacher P, Steffens S.
2012. Cannabinoid receptor CB2 protects against balloon-induced neointima formation. Am J Physiol Heart Circ Physiol 302: H1064-74
31. Mukhopadhyay P, Horvath B, Zsengeller Z, Batkai S, Cao Z, Kechrid M, Holovac E,
Erdelyi K, Tanchian G, Liaudet L, Stillman IE, Joseph J, Kalyanaraman B, Pacher P. 2012a. Mitochondrial reactive oxygen species generation triggers inflammatory response and tissue injury associated with hepatic ischemia-reperfusion: therapeutic potential of mitochondrially targeted antioxidants. Free Radic Biol Med 53: 1123-38
32. Mukhopadhyay P, Horvath B, Zsengeller Z, Zielonka J, Tanchian G, Holovac E, Kechrid
M, Patel V, Stillman IE, Parikh SM, Joseph J, Kalyanaraman B, Pacher P. 2012b. Mitochondrial-targeted antioxidants represent a promising approach for prevention of cisplatin-induced nephropathy. Free Radic Biol Med 52: 497-506
33. Pacher P, Mackie K. 2012. Interplay of cannabinoid 2 (CB2) receptors with nitric oxide
synthases, oxidative and nitrative stress, and cell death during remote neurodegeneration. J Mol Med (Berl) 90: 347-51
34. Rajesh M, Batkai S, Kechrid M, Mukhopadhyay P, Lee WS, Horvath B, Holovac E,
Cinar R, Liaudet L, Mackie K, Hasko G, Pacher P. 2012. Cannabinoid 1 receptor promotes cardiac dysfunction, oxidative stress, inflammation, and fibrosis in diabetic cardiomyopathy. Diabetes 61: 716-27
35. Silverman MN, Mukhopadhyay P, Belyavskaya E, Tonelli LH, Revenis BD, Doran JH,
Ballard BE, Tam J, Pacher P, Sternberg EM. 2012. Glucocorticoid receptor dimerization is required for proper recovery of LPS-induced inflammation, sickness behavior and metabolism in mice. Mol Psychiatry
36. Steffens S, Pacher P. 2012. Targeting cannabinoid receptor CB(2) in cardiovascular
disorders: promises and controversies. Br J Pharmacol 167: 313-23 37. Szabo C, Pacher P. 2012. The outsiders: emerging roles of ectonucleotidases in
inflammation. Sci Transl Med 4: 146ps14

181
38. Zsengeller ZK, Ellezian L, Brown D, Horvath B, Mukhopadhyay P, Kalyanaraman B, Parikh SM, Karumanchi SA, Stillman IE, Pacher P. 2012. Cisplatin nephrotoxicity involves mitochondrial injury with impaired tubular mitochondrial enzyme activity. J Histochem Cytochem 60: 521-9
Collaborators
George Kunos, MD, PhD, Scientific Director, NIAAA/NIH Bin Gao, MD, PhD, Section Chief, Liver Biology, NIAAA/NIH B.J. Song, PhD, Section Chief, Laboratory of Membrane Biochemistry and Biophysics, NIAAA Yoshihiro Kashiwaya, MD, PhD, NIAAA/NIH Esther M. Sternberg, MD, Director Integrative Neural Immune Program Section on Neuro-
endocrine Immunology & Behavior, NIMH/NIH Benjamin F. Cravatt, PhD, Departments of Cell Biology and Chemistry, The Scripps Research
Institute, La Jolla, CA Lucas Liaudet, MD, Critical Care Division, Department of Internal Medicine, University
Hospital, Lausanne, Switzerland Zoltan Ungvari, MD, PhD, Reynolds Oklahoma Center on Aging, University of Oklahoma
Health Sciences Center Irina Obrosova, PhD, Division of Nutrition and Chronic Disease, Pennington Biomedical
Research Center, Louisiana State University, Baton Rouge, LA 70808 Gyorgy Hasko, MD, PhD, Department of Surgery, UMD NJ-New Jersey Medical School,
Newark, NJ Oleg V. Evgenov, MD, PhD, Department of Anesthesia & Critical Care Massachusetts General
Hospital, Harvard Medical School, Boston, MA Ken Mackie, MD, PhD, Department of Psychological and Brain Sciences and Program in
Neuroscience, Indiana University, Bloomington Csaba Szabo, MD, PhD, Department of Anesthesiology, U of Texas Medical Branch, Galveston David Wink, PhD, Radiology Branch, NCI/NIH Murali Cherukuri, PhD, Radiology Branch, NCI/NIH Sabine Steffens, PhD, Department of Medicine, Geneva University Hospital, Foundation for Medical Researches, Geneva, Switzerland José Pedraza-Chaverri, PhD, Universidad Nacional Autónoma de México, 04510 México, DF, Mexico Azza El-Remessy, PhD, The University of Georgia College of Pharmacy Aristidis Veves, MD, PhD, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts, USA Roger Pertwee, PhD, University of Aberdeen, Foresterhill, Aberdeen, Scotland, UK Raphael Mechoulam, PhD, Department of Medicinal Chemistry and Natural Products, Pharmacy
School, Medical Faculty, Hebrew University, 91120 Jerusalem, Israel Dr. Pacher: [email protected]

182
______________________________________________________________________
Laboratory of Molecular Physiology ______________________________________________________________________
Stephen R. Ikeda, MD, PhD, Chief The goals of the Laboratory are to explore molecular mechanisms contributing to synaptic transmission and neuronal excitability at the molecular, cellular and organismic level. An emphasis is placed on developing new technologies to explore neuronal cellular and subcellular signaling pathways based on advanced optical techniques. The Section on Transmitter Signaling, headed by Dr. Stephen R. Ikeda, focuses primarily on determining the molecular mechanisms underlying G-protein coupled receptor (GPCR) modulation of neuronal high-threshold Ca2+ channels in neuronal systems using electrophysiological and molecular techniques. A second goal is the development of optical sensors based on fluorescence resonance energy transfer (FRET) and reporter protein complementation to examine GPCR activity in living cells. Techniques in the laboratory include whole-cell voltage-clamp, heterologous expression in neurons by microinjection and viral techniques, wide-field FRET imaging, and total internal reflectance fluorescence microscopy. GPCRs (e.g., CB1, CRF, mGluR, NPY, and nociceptin receptors) comprise major cellular targets for pharmaceuticals used in the treatment of alcoholism and other addictive disorders. Additionally, N-type Ca2+ channels and heterotrimeric G-protein signaling pathways utilizing Gβγ have been directly implicated in ethanol reward and consumption mechanisms. Thus, increasing our knowledge of the basic mechanisms underlying GPCR modulation of Ca2+ and other ion channels should help clarify molecular mechanisms underlying the therapeutic efficacy of agents used for the treatment of alcoholism and addiction.
The Section on Cellular Biophotonics, headed by Dr. Steven S. Vogel, is focused on understanding how cells integrate exocytosis and endocytosis to regulate the abundance and assembly of membrane proteins on the cell surface. Toward this end his section has been developing advanced imaging techniques for the determination of protein-protein interaction and protein complex stoichiometry in living cells. Techniques available in the section include single- and two-photon laser scanning confocal microscopy, FRET imaging, spectral imaging, fluorescence lifetime imaging (FLIM), time-resolved and steady state anisotropy, and fluctuation correlation spectroscopy (FCS). Model systems used in the laboratory include: Developing sea urchin eggs, transfected cell lines, and primary hippocampal neurons in culture. One project in the Section is investigating conformational changes in the calcium calmodulin dependent protein kinase II (CaMKII) holoenzyme before and after activation. The second project involves using fiber optics for deep brain photometry in living mice. The objective of this project is to use FRET to study protein-protein interactions in synapses as the mice perform operant tasks.
The Section on Model Synaptic Systems, headed by Dr. Fumihito Ono, focuses on the cellular and molecular mechanisms underlying synaptic transmission in the vertebrate model organism Danio rerio (zebrafish). Zebrafish are genetically malleable, optically transparent at early stages

183
of development, and amenable to higher throughput methodologies. Extensive regions of synteny between zebrafish and human genomes make Danio an ideal model for translational research. The transparent nature of the early zebrafish embryo meshes well with the advanced imaging and electrophysiological expertise within the Laboratory.
The integration of the three Sections is designed to bring together investigators with common interests in molecules involved in neuronal excitability and synaptic transmission yet expertise in diverse but complementary techniques. The emphasis on model systems facilitates the application, from the cellular to organismic level, of new technologies in an efficient and cost-effective manner.
Laboratory of Molecular Physiology
Back row: (left to right) Henry Puhl, Pabak Sarker, Van Lu, Ho-Won Kang, Steve Vogel, Fumi Ono, Steve Ikeda
Middle: (far left) Tuan Nguyen, (far right) Liz Kirby
Front: (left to right): Irina Ivanova, Yu-Jin Won, Kaitlin Davis, Megan Mott, Hiromi Ikeda, Jee-Young Park, Jithesh Velichamthotu Veetil
Dr. Ikeda: [email protected]

184
______________________________________________________________________
Section on Transmitter Signaling ______________________________________________________________________
Stephen R. Ikeda, MD, PhD, Chief Current members Henry L. Puhl III, PhD, Staff Scientist Yu-Jin Won, PhD, Postdoctoral Fellow Van B. Lu, PhD, Postdoctoral Fellow Ho-Won Kang, PhD, Postdoctoral Fellow Irina Ivanova, Technical IRTA Elizabeth Kirby, Administrative Laboratory Manager
Ion Channel Modulation by Second Messenger Systems
The Section on Transmitter Signaling focuses primarily on determining the molecular mechanisms underlying G-protein coupled receptor (GPCR) modulation of voltage-gated Ca2+ channels in neuronal systems using electrophysiological, optical, molecular, and biochemical techniques. A consequence of modulation, which usually manifests as a decrease in current flow through the channel, neuronal excitability and neurotransmitter release at synapses is modified. Although several signaling pathways have been identified, the best-studied is a direct inhibition of the ion channel by G-protein βγ-subunits liberated from the G-protein heterotrimer following agonist-mediated receptor activation. This canonical pathway is shared among the high-voltage activated Ca2+ channels of the CaV2.x family (CaV2.1–2.3; P/Q-, N-, and R-type, respectively)

185
and represents one of the most widely studied and best understood mechanism of presynaptic inhibition. GPCRs (e.g., CB1, CRF, mGluR, NPY, and nociceptin) comprise major cellular targets for pharmaceuticals used in the treatment of alcoholism and other addictive disorders. Additionally, N-type Ca2+ channels and heterotrimeric G-protein signaling pathways utilizing Gβγ have been directly implicated in ethanol reward and consumption mechanisms.
Characterization of Na+ and Ca2+ channels in zebrafish dorsal root ganglion neurons Won, in collaboration with Ono Dorsal root ganglia (DRG) somata from rodents have provided an excellent model system to study ion channel properties and modulation using electrophysiological investigation. As in other vertebrates, zebrafish (Danio rerio) DRG are organized segmentally and possess peripheral axons that bifurcate into each body segment. However, the electrical properties of zebrafish DRG sensory neurons, as compared with their mammalian counterparts, are relatively unexplored because a preparation suitable for electrophysiological studies has not been available. We show enzymatically dissociated DRG neurons from juvenile zebrafish expressing Isl2b-promoter driven EGFP were easily identified with fluorescence microscopy and amenable to conventional whole-cell patch-clamp studies. Two kinetically distinct TTX-sensitive Na+ currents (rapidly- and slowly-inactivating) were discovered. Rapidly-inactivating INa were preferentially expressed in relatively large neurons, while slowly-inactivating INa was more prevalent in smaller DRG neurons. RT-PCR analysis suggests zscn1aa/ab, zscn8aa/ab, zscn4ab and zscn5Laa are possible candidates for these INa components. Voltage-gated Ca2+ currents (ICa) were primarily (87%) comprised of a high-voltage activated component arising from ω-conotoxin GVIA-sensitive CaV2.2 (N-type) Ca2+ channels. A few DRG neurons (8%) displayed a miniscule low-voltage-activated component. ICa in zebrafish DRG neurons were modulated by neurotransmitters via either voltage-dependent or -independent G-protein signaling pathway with large cell-to-cell response variability. Our present results indicate that, as in higher vertebrates, zebrafish DRG neurons are heterogeneous being composed of functionally distinct subpopulations that may correlate with different sensory modalities. These findings provide the first comparison of zebrafish and rodent DRG neuron electrical properties and thus provide a basis for future studies. (Won et al., 2011) Nr4a1-eGFP Is a Marker of Striosome-Matrix Architecture, Development and Activity in the Extended Striatum Puhl, in collaboration with Davis Transgenic mice expressing eGFP under population specific promoters are widely used in neuroscience to identify specific subsets of neurons in situ and as sensors of neuronal activity in vivo. Mice expressing eGFP from a bacterial artificial chromosome under the Nr4a1 promoter have high expression within the basal ganglia, particularly within the striosome compartments and striatal-like regions of the extended amygdala (bed nucleus of the stria terminalis, striatal fundus, central amygdaloid nucleus and intercalated cells). Grossly, eGFP expression is inverse to the matrix marker calbindin 28K and overlaps with mu-opioid receptor immunoreactivity in the striatum. This pattern of expression is similar to Drd1, but not Drd2, dopamine receptor driven eGFP expression in structures targeted by medium spiny neuron afferents. Striosomal expression is strong developmentally where Nr4a1-eGFP expression overlaps with Drd1, TrkB, tyrosine hydroxylase and phospho-ERK, but not phospho-CREB, immunoreactivity in ‘‘dopamine islands’’. Exposure of adolescent mice to methylphenidate resulted in an increase in eGFP in both compartments in the dorsolateral striatum but eGFP expression remained brighter in the striosomes. To address the role of activity in Nr4a1-eGFP expression, primary striatal

186
cultures were prepared from neonatal mice and treated with forskolin, BDNF, SKF-83822 or high extracellular potassium and eGFP was measured fluorometrically in lysates. eGFP was induced in both neurons and contaminating glia in response to forskolin but SKF-83822, brain derived neurotrophic factor and depolarization increased eGFP in neuronal-like cells selectively. High levels of eGFP were primarily associated with Drd1+ neurons in vitro detected by immunofluorescence; however, 15% of the brightly expressing cells contained punctate met-enkephalin immunoreactivity. The Nr4a1-GFP mouse strain will be a useful model for examining the connectivity, physiology, activity and development of the striosome system. (Davis and Puhl, 2012) N-arachidonyl glycine (NAGly does not activate G protein-coupled receptor 18 (GPR18) signaling via canonical pathways Lu, Puhl Recent studies propose that N-arachidonyl glycine (NAGly), a carboxylic analog of anandamide, is an endogenous ligand of the Gαi/o -coupled receptor 18 (GPR18). However, a high-throughput β-arrestin-based screen failed to detect activation of GPR18 by NAGly (Yin et al., 2009; JBC, 18:12328). To address this inconsistency, we investigated GPR18 coupling in a native neuronal system with endogenous signaling pathways and effectors. We heterologously expressed GPR18 in rat sympathetic neurons and examined the modulation of N-type (Cav2.2) calcium channels. Proper expression and trafficking of receptor was confirmed by the “rim-like” fluorescence of fluorescently-tagged receptor and the positive staining of external hemagglutinin-tagged GPR18-expressing cells. Application of NAGly on GPR18-expressing neurons did not inhibit calcium currents, but instead potentiated currents in a voltage-dependent manner, similar to what has previously been reported (Guo et al., 2008; J Neurophysiol, 100:1147). Other proposed agonists of GPR18, including anandamide and abnormal cannabidiol, also failed to induce inhibition of calcium currents. Mutants of GPR18, designed to constitutively activate receptors, did not tonically inhibit calcium currents indicating a lack of GPR18 activation or coupling to endogenous G proteins. Other downstream effectors of Gαi/o-coupled receptors, G protein-coupled inwardly-rectifying potassium channels and adenylate cyclase, were not modulated by GPR18 signaling. Furthermore, GPR18 did not couple to other G proteins tested: Gαs, Gαz and Gα15. These results suggest NAGly is not an agonist for GPR18 or that GPR18 signaling involves non-canonical pathways not examined in these studies. (Lu et al., 2013) Publications 2011-2012 1. Won YJ, Ono F, Ikeda SR. 2011. Identification and Modulation of Voltage-gated Ca2+
currents in Zebrafish Rohon-Beard Neurons. Journal of Neurophysiology 105:442-53
2. Davis, YJ, Puhl HL 2011. Nr4a1-eGFP is a marker of striosome-matrix architecture, development and activity in the extended striatum. PLoS One 6:e16619
3. Won YJ, Ono F, Ikeda SR. 2012. Characterization of Na+ and Ca2+ channels in zebrafish dorsal root ganglion neurons. PLoS One 6:e16619
4. Lu VB, Puhl HL, Ikeda SR. 2013. N-Arachidonyl glycine does not activate G protein-coupled receptor 18 signaling via canonical pathways. Mol Pharmacol. 83:267-282.
Dr. Ikeda: [email protected]

187
__________________________________________________________________
Section on Model Synaptic Systems ___________________________________________________________________
Fumihito Ono, MD, PhD, Acting Chief Jee-Young Park, PhD, Postdoc IRTA Hiromi Ikeda, PhD, Postdoc IRTA Meghan Mott, PhD, Postdoc IRTA Vivek Gupta, Summer Intern Nao Kawahori, Special volunteer
Zebrafish as a model to study nervous system The Section on Model Synaptic Systems started in 2007, when Fumihito Ono moved to NIH/NIAAA from the University of Florida. We are currently studying the nervous system at several levels. An experimental paradigm we have used heavily for the past several years is the neuromuscular junction (NMJ), which is a synapse between motor neuron and muscle. Not only is this synapse directly linked to various diseases arising from genetic defects or autoimmune conditions, it also offers an exceptional accessibility for an array of experimental techniques. We have recently broadened our scope of research and are also studying neural networks in the central nervous system and its response to ethanol. Neuromuscular junction as a model system of nicotinic synapse
Our projects on NMJ center around locomotion mutants we discovered to have defects in two key molecules of the neuromuscular synapse. One mutant lacks acetylcholine receptors (AChR) in the muscle. As a result, the fish cannot mount a movement when the motor neuron releases ACh. Another mutant has a dysfunctional rapsyn. Rapsyn is a post-synaptic protein that brings
l to r:, JY Park, L Kirby, F Ono, M Mott, H Ikeda
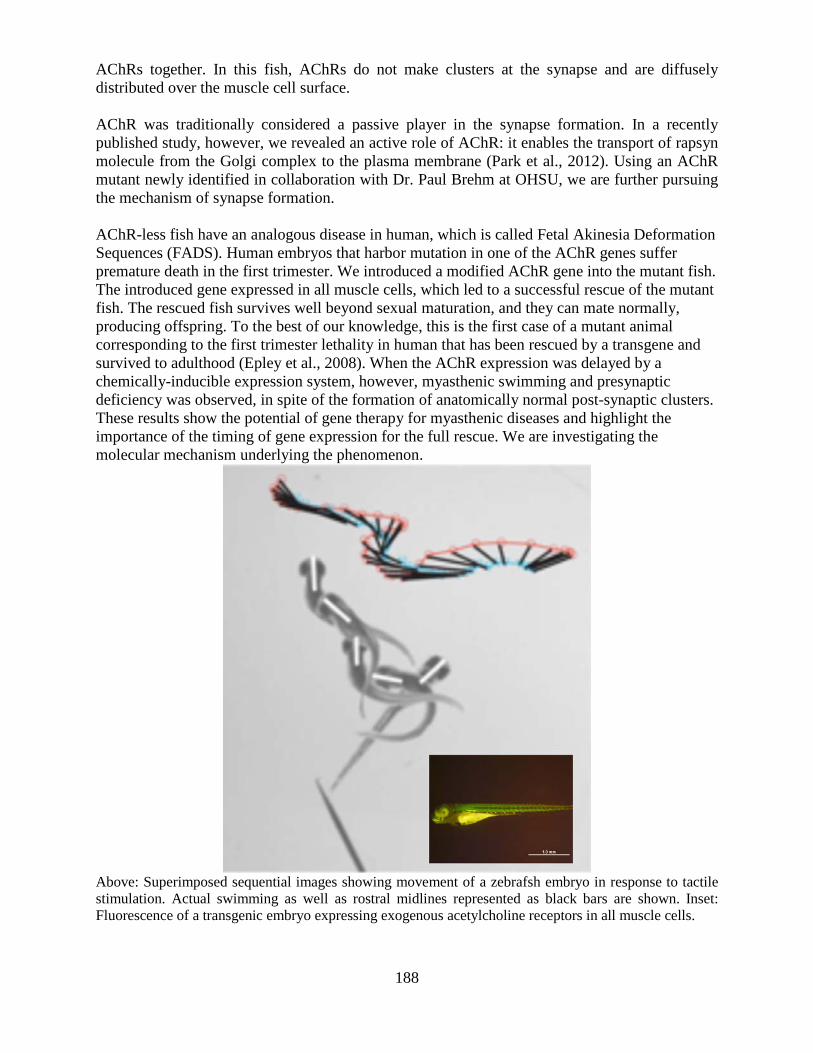
188
AChRs together. In this fish, AChRs do not make clusters at the synapse and are diffusely distributed over the muscle cell surface. AChR was traditionally considered a passive player in the synapse formation. In a recently published study, however, we revealed an active role of AChR: it enables the transport of rapsyn molecule from the Golgi complex to the plasma membrane (Park et al., 2012). Using an AChR mutant newly identified in collaboration with Dr. Paul Brehm at OHSU, we are further pursuing the mechanism of synapse formation. AChR-less fish have an analogous disease in human, which is called Fetal Akinesia Deformation Sequences (FADS). Human embryos that harbor mutation in one of the AChR genes suffer premature death in the first trimester. We introduced a modified AChR gene into the mutant fish. The introduced gene expressed in all muscle cells, which led to a successful rescue of the mutant fish. The rescued fish survives well beyond sexual maturation, and they can mate normally, producing offspring. To the best of our knowledge, this is the first case of a mutant animal corresponding to the first trimester lethality in human that has been rescued by a transgene and survived to adulthood (Epley et al., 2008). When the AChR expression was delayed by a chemically-inducible expression system, however, myasthenic swimming and presynaptic deficiency was observed, in spite of the formation of anatomically normal post-synaptic clusters. These results show the potential of gene therapy for myasthenic diseases and highlight the importance of the timing of gene expression for the full rescue. We are investigating the molecular mechanism underlying the phenomenon.
Above: Superimposed sequential images showing movement of a zebrafsh embryo in response to tactile stimulation. Actual swimming as well as rostral midlines represented as black bars are shown. Inset: Fluorescence of a transgenic embryo expressing exogenous acetylcholine receptors in all muscle cells.

189
Response of neurons to ethanol We study effects of ethanol on the zebrafish brain, by analyzing intoxicated embryos at the cellular level. We obtained two interesting findings from these studies. First, zebrafish larvae show unusual resistance to a high concentration of ethanol. Second, their neurons rapidly adapt to ethanol, which may underlie the development of acute tolerance to alcohol. We are currently investigating the molecular mechanisms for these phenomena, with an emphasis on nicotinic ACh receptors. References Epley KE, Urban JM, Ikenaga T, Ono F. 2008. A modified acetylcholine receptor delta-
subunit enables a null mutant to survive beyond sexual maturation. J Neurosci 28:13223-31
Ono F. 2008. An emerging picture of synapse formation: a balance of two opposing pathways. Sci Signal 1:pe3
Park J, Ikeda H, Ikenaga T, Ono F. 2012. Acetylcholine receptors enable the transport of rapsyn from the Golgi complex to the plasma membrane. J Neurosci 32:7356-7363.
Publications 2011-20102 1. Won Y, Ono F, Ikeda S. 2011. Identification and modulation of voltage-gated Ca2+
current in zebrafish Rohon-Beard neurons. J Neurophys 105: 442-453. 2. Diep C, Ma D, Arora N, Wingert R, Bollig F, Djordjevic G, Lichman B, Zhu H, Ikenaga
T, Ono F, Englert C, Hukriede N, Handin R, Davidson A. 2011. Identification of adult renal progenitor cells capable of nephron formation and regeneration in zebrafish. Nature 470: 95-100.
3. Ikenaga T, Urban J, Gebhart N, Hatta K, Kawakami K, Ono F. 2011. Formation of the
spinal network in zebrafish determined by domain-specific Pax genes. J Comp Neurol 519: 1562-1579.
4. Mongeon R, Walogorsky M, Urban J, Mandel G, Ono F, Brehm P. 2011. An
acetylcholine receptor lacking both γ and ε subunits mediates transmission in zebrafish slow muscle synapses. J Gen Phys 138: 353-366.
5. Nakayama S, Ikenaga T, Kawakami K, Ono F, Hatta K. 2012. A Gal4 gene trap line
useful for studying the morphogenesis of craniofacial perichondrium, vascular endothelium-associated cells, floor plate, and dorsal midline radial glia during zebrafish development. Dev Gro Diff 54: 202-215.
6. Papke R, Ono F, Stokes C, Urban J, Boyd T. 2012 The nicotinic acetylcholine receptors
of zebrafish and an evaluation of pharmacological tools used for their study. Biochem Pharmacol 84:352-365.

190
7. Park J, Ikeda H, Ikenaga T, Ono F. 2012. Acetylcholine receptors enable the transport of rapsyn from the Golgi complex to the plasma membrane. J Neurosci 32:7356-7363.
8. Won Y, Ono F, Ikeda S. 2012. Characterization of Na+ and Ca2+ channels in zebrafish
dorsal root ganglion neurons. PLoS ONE 7: e42602.
9. Walogorsky M, Wen H, Mongeon R, Urban J, Ono F, Mandel G, Brehm P. 2012. A zebrafish model of congenital myasthenic syndrome reveals mechanisms causal to developmental recovery. PNAS 109: 17711-17716.
Collaborators Stephen Ikeda, LMP, NIAAA Stephen Vogel, LMP, NIAAA Ralph Nelson, NINDS Kevin Briggman, NINDS Harold Burgess, NICHD Fielding Hejtmancik, NEI Gerald Downes, University of Massachusetts Roger Papke, University of Florida Alan Davidson, University of Auckland Paul Brehm, Oregon Health and Science University Koichi Kawakami, National Institute of Genetics, Japan Kohei Hatta, University of Hyogo, Japan Dr. Ono: [email protected]

191
______________________________________________________________________
Section on Cellular Biophotonics ______________________________________________________________________
Steven S. Vogel, PhD, Acting Chief Tuan Nguyen, PhD, Research Fellow Pabak Sarkar, PhD, Visiting Fellow Jithesh Velichamthotu Veetil, PhD, Visiting Fellow Kaitlin Davis, BS, Grad Student NIH/Johns Hopkins GPP
FRET imaging of protein-protein interactions in
living cells The primary objective of the Section on Cellular Biophotonics of the Laboratory of Molecular Biophysics ((SCB/LMP) is to develop new forms of microscopy to study protein-protein interactions under physiological conditions. We are specifically interested in the interactions of synaptic proteins that are involved in regulating memory, behavior, and addiction.
In cells, proteins rarely function in isolation; typically they act together to form assemblies that mediate cellular processes. Considering how ubiquitous protein complexes are, as well as the vast array of functions they perform, it is no surprise that many human diseases arise in part from inappropriate protein interaction. A major obstacle to understanding the basis of these diseases is the paucity of robust methods for studying both normal and abnormal protein interactions under physiological conditions. Since its formation the Section on Cellular Biophotonics has been developing new forms of live-cell microscopy based primarily on Förster resonance energy transfer (FRET) to monitor protein interactions. Ultimately, we would like to apply these approaches to facilitate the identification of drugs that can target and specifically correct abnormal protein interactions.
We currently have three major projects in the laboratory. The first project is using a method recently developed in our Section, fluorescence polarization and fluctuation analysis (FPFA), to monitor the conformational changes associated with the activation and translocation of the calcium calmodulin dependent protein kinase-II (CaMKII). FPFA is a hybrid single-molecule based assay for measuring FRET, rotational and lateral diffusion, and the number of subunits in a holoenzyme. FPFA can be used for solution samples as well as in living cells, and is based on time resolved anisotropy and fluorescence correlation spectroscopy (FCS).
The second project also utilizes FPFA microscopy, but in conjunction with scanning mutagenesis. The goal of this project is to adapt FPFA to identify and map all of the sites on a protein that is involved in protein-protein interactions. Single-point mutations will be randomly inserted throughout a protein of interest, and then automate FPFA microscopy will be used to

192
screen these samples for changes in FRET, diffusion, and stoichiometry. The specific aim of the third project is to develop fiber optic technologies that can detect
changes in protein interactions occurring deep inside the brain of mice performing motor tasks. Our strategy involves measuring hetero-FRET using a fluorescence lifetime based FRET system (FLIM-FRET) between a donor fluorescent protein and a ‘fluorescent’-protein dark absorber as a FRET acceptor while simultaneously measuring FCS, all through a fiber optic. A sub-femto-liter excitation volume at the fiber tip (required for FCS) will be generated by using two-photon excitation. Like FPFA, by combining FRET measurements with FCS measurements FLFA (fluorescence lifetime and fluctuation analysis) should be able to control for many of the problems associated with standard FRET analysis.
Publications 2011-2012
1. Nguyen TA, Sarkar P, Veetil JV, Koushik SV, Vogel SS. 2012. Fluorescence Polarization and Fluctuation Analysis Monitors Subunit Proximity, Stoichiometry, and Protein Complex Hydrodynamics. Plos One 7:e38209
2. Vogel SS, Nguyen TA, van der Meer BW, Blank PS. 2012. The Impact of Heterogeneity and Dark Acceptor States on FRET: Implications for Using Fluorescent Protein Donors and Acceptors. Plos One 7:e49593
3. Cui G, Jun S, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM. Concurrent Activation of Striatal Direct and Indirect Pathways During Action Initiation. Nature, in Press.
4. van der Meer BW, van der Meer DM, Vogel SS. (Chapter in Press). Chapter 3: Optimizing the Orientation Factor KAPPA-SQUARED for More Accurate FRET Measurements in FRET – Förster Resonance Energy Transfer: From Theory to Applications, John Wiley & Sons.
Collaborators: Dr. Paul Blank, NICHD Dr. David Lovinger, NIAAA Dr. Fumihito Ono, NIAAA Dr. Eric O. Long, NIAID Dr. Richard Youle, NINDS Dr. Rui Costa, Instituto Gulbenkian de Ciencia, Portugal Dr. Wieb van der Meer, Western Kentucky University Dr. Anne Kenworthy, Vanderbilt University Dr.Vogel: [email protected]

193
______________________________________________________________________
Laboratory of Metabolic Control ______________________________________________________________________ Richard L. Veech, MD, DPhil, Chief Robert Pawlosky, PhD, Staff Scientist Martin Kemper, PhD, Postdoc IRTA M. Todd King, MS, Chemist Mellisa Aplan, BS, Postbac IRTA Control of Cellular Redox States Veech Central to energy homeostasis is the control of cellular redox states and the linked phosphorylation potential. The redox state is a measure of the ability of metabolic reactions to donate or accept electrons. Within the cell there are many redox couples with varying potentials. For example the free cytosolic NAD couple readily accepts electrons from the glycolytic pathway and has a potential of about -0.19 V. In contrast, the free cytosolic NADP couple, used for the synthesis of lipids and other reactions, has a much more negative and therefore powerful redox potential of -0.42 V, allowing synthetic reductions to be driven to completion. Intermediate between these cytosolic co-factor redox couples is the mitochondrial NAD couple whose potential is usually about -0.28 V. This intermediate potential allows the mitochondria to synthesize ATP in the combined reactions of electron transport and ATP synthesis. Linked to and determined by the NAD redox couples of cytoplasm and mitochondria is the phosphorylation potential, the ATP/ADPxPi ratio, or the energy of the ATP bond. Depending upon the redox energies available between the mitochondrial NAD couple and the terminal electron acceptor, molecular O2, the energy of cellular ATP can vary particularly in disease states. In non pathological states, the energy of ATP hydrolysis is tightly regulated, varying only about 10%, between -53 to -60 kJ/ mole of ATP hydrolyzed. The cell possesses many other redox couples wherein common co-factors link and integrate multiple reactions sharing the common co-factors. Examples of such co-factor determined redox states are the mitochondrial co-enzyme Q/co-enzyme QH2 couple and the folate couples to name only a couple. The former mitochondrial couple is of particular importance for two reasons. Firstly, the distance between the redox states of the mitochondrial NAD and Q couples determines the energy of ATP synthesized by the mitochondria. Secondly, the major source of cellular free radical production is the spontaneous reaction of the intermediate Q semiquinone with oxygen. Oxidation of the Q couple therefore decreases the production of free radicals, an important etiological factor in many disease states. ATP hydrolysis has been called “the energy currency” of the cell. Many disease states are characterized by a lowering of the energy of ATP hydrolysis. One the most important links between the energy of ATP hydrolysis and other cellular reactions is the extent of inorganic ion gradients between the extra and intracellular phase of the cell. These gradients between Na+ and

194
Ca2+ between the phases are determined by the energy of ATP hydrolysis as are the gradients of the 9 major inorganic ions. In turn the gradients of inorganic ions determines the distribution of water between intra and extracellular phases accounting for the universal phenomena of the loss of intracellular K+ and the gain of intracellular Na+ and Ca2+ during any type of injury to the cell. Cellular redox and phosphorylation states can vary with the type of substrate being metabolized. This variability in energy available from the metabolism of substrates is inherent in the difference between the heats of combustion of the various metabolic substrates being combusted to H2O and CO2 in the citric acid cycle and the mitochondrial processes of oxidative phosphorylation. Redox and phosphorylation states can also be changed by variance in hormonal or disease status of the organism. The energy of ATP hydrolysis is also changed by changes in the intracellular free [Mg2+] concentrations. The quantitation of these changes is the subject of a recent publication (Bergman et al, 2010). Recognition that change of substrate being used for the generation of metabolic energy production can alter these central controlling cellular co-factor couples presents new and unexpected therapeutic opportunities in a variety of diverse disease states (Veech, 1986). It is also of importance in the design of resuscitation fluids (Valeri et al, 2008) Metabolomics Pawlosky, Veech The determination of cellular redox and phosphorylation states cannot be determined by direct measurement of the nucleotides involved such as NAD, NADH, ADP, AMP and the like for reasons previously reviewed (Veech, 2006). Instead, various intermediary metabolites of enzymatic reactions of high capacity relative to flux must be measured to determine the redox or phosphorylation state in the compartment of interest. This can be accomplished by established methods of enzymatic analysis. However these methods are slow and labor intensive. Accordingly, we have developed new methods for the analysis of new samples of tissue using capillary electrophoresis combined with mass spectrometry which allows the quantitative determination of the products and reactants of the enzymes comprising a metabolic pathway of interest as well as the determination of the flux through that pathway using stable isotopes. Such measurements, when combined with knowledge or determination of the kinetic constants for the enzymes involved and the equilibrium constant for each reaction under conditions appropriate to the in vivo situation allow for the determination of the degree of control of flux exerted by each enzyme within the pathway. It is the alteration of flux through metabolic pathways that define the disease phenotype. Knowledge of the control strength of each step suggests new approaches where the disease phenotype may be returned to normal (Volkow et al, 2006). Of particular interest to the programs of NIAAA are the metabolic effects of acetate, the primary metabolite of ethanol. Ethanol is almost totally converted to acetate in the liver. The acetate formed then leaves liver where it is metabolized in extra hepatic tissues, including brain. Relatively little attention has been paid to the effects of acetate metabolism upon brain. Using techniques which are unique to this laboratory, namely brain blowing, which eliminates post mortal changes in energy metabolites, and a combination of enzymatic analysis with Capillary Electrophoresis and Gas Chromatographic Mass Spectrometry we have now completed a metabolomic survey of the changes in brain metabolism resulting from the metabolism of acetate. The most striking finding is that the metabolism of acetate by brain leads to a lowering of ∆G’ of ATP hydrolysis (Pawlosky et al, 2010).

195
Ketosis, Neurological Disease, Insulin Resistance and Obesity King, Pawlosky, Clarke, Veech A ketogenic diet has been used for over 100 years in the treatment of refractory epilepsy. However such high fat, low carbohydrate diets are not suitable for treatment of patients over 17 years of age because of poor patient tolerance and the atherogenic potential and other abnormalities resulting from elevation of blood cholesterol, triglyceride and free fatty acids. Accordingly, in collaboration with Oxford University we have been developing an oral form of ketone bodies suitable for human use. Because the metabolism of the ketone body, D-β-hydroxybutyrate, leads to elevation of the energy of ATP hydrolysis above that which results from the metabolism of glucose, a ketone diet has potential application in many disease phenotypes beyond the treatment of epilepsy. An important group of this class of disease phenotypes is those of neurodegeneration, among which is Parkinson’s disease. Several years ago we demonstrated that administration of D-β-hydroxybutyrate preserved mesencephalic neurons from death caused by MPTP, a neurotoxin resulting in immediate Parkinsonism in humans. It has subsequently been shown that treatment of C57 black mice with a ketogenic diet prevented mesencephalic damage from administered MPTP. Further, a ketogenic diet has been shown to reduce tremor and rigidity in a small study of human patients by about 60% while improving mentation. More recently we have shown that partial inhibition of NADH dehydrogenase (Complex I) by rotenone in cultured dopaminergic neuroblastoma cells could be overcome by D-β-hydroxybutyrate. Abnormalities in complex I have long been reported in Parkinsonism and recent 31P-MRS studies of occipital cortex shows a lowered PCr/ATP ratio or energy of ATP hydrolysis in this area, not generally thought to be involved in Parkinsonism. These observations suggest that elevation of ketones deserve a wider therapeutic test in this disease (Imamura et al, 2006; Veech, 2004). In attempts to by-pass a number of the complications of inducing ketosis without feeding ketogenic diet, we have developed ketone esters suitable for oral administration. We have completed animal toxicity of newly synthesized ketone esters. We have taken the necessary steps to obtain FDA approval to feed this new food to human subjects. Human toxicity studies have now been completed in first in human trials and no toxic effects were observed. Studies of the effects of these esters on physical and cognitive performance in human subjects are underway at Oxford University under the direction of Dr. Kieran Clarke. Three significant advances recently reported from the lab include: 1) demonstration that in the triple transgenic mouse model of Alzheimer’s disease, feeding of the ketone ester diet resulted in a decrease in accumulation in brain of amyloid and phosphorylated tau and an improvement in cognitive function. 2) publication of a hypothesis which suggests that traumatic brain injury results from a loss of cerebral energy secondary to the opening of the mitochondrial voltage dependent pore and that this defect could be corrected by ketosis. 3) demonstration that feeding ketone ester diets to mice results in an increase in brown fat mitochondria and uncoupling protein and the activation of brown fat which would be of significance in combating the epidemic of obesity currently adding the cost of health care in the West as well as adding to morbidity and premature mortality.

196
In addition, significant advances have been made in developing methods for producing ketone esters in quantities sufficient to meet national needs at an affordable price. What is presently needed is: 1) an industrial partner capable of producing significant quantities of ketone esters, 2) further approvals from FDA to administer ketone ester, now ruled as a GRAS food for prolonged periods of time, and 3) clinical trials of ester in Alzheimer’ disease, Parkinson’s disease, traumatic brain injury, type 1 and 2 diabetes, obesity and a variety of rare diseases including amytrophic lateral sclerosis, GLUT 1 deficiency and Leprechaunism and resuscitation injury. References
Bergman C, Kashiwaya Y, Veech RL. 2010. The effect of pH and free Mg2+ on ATP linked enzymes and the calculation of Gibbs free energy of ATP hydrolysis. J Phys Chem B.114:16137-46
Imamura K, Takeshima T, Kashiwaya Y, Nakaso K, Nakashima K. 2006. D-beta-hydroxybutyrate protects dopaminergic SH-SY5Y cells in a rotenone model of Parkinson's disease. J Neurosci Res 84:1376-84
Kashiwaya Y, Pawlosky R, Markis W, King MT, Bergman C, Srivastava S, Murray A, Clarke K, Veech RL. 2010. A ketone ester diet increases brain malonyl-CoA and Uncoupling proteins 4 and 5 while decreasing food intake in the normal Wistar Rat. The Journal of biological chemistry 285: 25950-6 Pawlosky RJ, Kashiwaya Y, Srivastava S, King MT, Crutchfield C, Volkow N, Kunos G, Li TK, Veech RL. 2010. Alterations in brain glucose utilization accompanying elevations in blood ethanol and acetate concentrations in the rat. Alcohol Clin Exp Res. 34:375-81.
Valeri RC, Ragno G, Veech RL. 2008. Severe adverse events associated with hemoglobin based oxygen carriers: role of resuscitative fluids and liquid preserved RBC. Transfus Apher Sci 39:205-11
Veech RL. 1986. The toxic impact of parenteral solutions on the metabolism of cells: a hypothesis for physiological parenteral therapy. Am J Clin Nutr 44:519-51
Veech RL. 2004. The therapeutic implications of ketone bodies: the effects of ketone bodies in pathological conditions: ketosis, ketogenic diet, redox states, insulin resistance, and mitochondrial metabolism. Prostaglandins Leukot Essent Fatty Acids 70:309-19
Veech RL. 2006. The determination of the redox states and phosphorylation potential in living tissues and their relationship to metabolic control of disease phenotypes. Biochem Mol Biol Edu 34:168-79
Volkow ND, Wang GJ, Franceschi D, Fowler JS, Thanos PP, et al. 2006. Low doses of alcohol substantially decrease glucose metabolism in the human brain. Neuroimage 29:295-301

197
Publications 2011-2012 1. Kashiwaya Y, Bergman C, Lee JH, Wan R, Todd King M, Mughal MR, Okun E, Clarke
K, Mattson MP, Veech RL. 2012. A ketone ester diet exhibits anxiolytic and cognition-sparing properties, and lessens amyloid and tau pathologies in a mouse model of Alzheimer's disease. Neurobiology of aging
2. Veech RL, Valeri CR, Vanitallie TB. 2012. The mitochondrial permeability transition pore provides a key to the diagnosis and treatment of traumatic brain injury. IUBMB Life 64: 203-7
3. Srivastava S, Kashiwaya Y, King MT, Baxa U, Tam J, Niu G, Chen X, Clarke K, Veech RL. 2012. Mitochondrial biogenesis and increased uncoupling protein 1 in brown adipose tissue of mice fed a ketone ester diet. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 26: 2351-62
Collaborators Prof. Kieran Clarke, Dept of Physiology, Oxford Dr. C. Robert Valeri, Naval Blood Research Laboratory Prof. Theodore VanItallie, St Lukes Hospital, Columbia U Medical School.
Dr Veech: [email protected]
l to r: R Pawlosky, M Aplan, R Veech, T King

198
______________________________________________________________________
Laboratory of Neuroimaging ______________________________________________________________________ Nora Volkow, MD, Chief Dardo Tomasi, PhD, Staff Scientist Panayotis Thanos, PhD, Staff Scientist Sunny Kim, PhD, Staff Scientist Millard Jayne, Clinical Research Nurse Lisa Muench, Chemical Engineer Karen Apelskog, Research Coordinator Cristopher Wong, Data Manager This laboratory utilizes brain imaging to investigate the neurobiology of addictive behaviors, compulsive overeating and obesity, attention deficit hyperactivity disorder (ADHD) and the effects that conditions such as sleep deprivation can have on brain function. Also being conducted are basic studies of the organization of the normal human brain. Specific aims are: 1) To study the effects of drugs on the physiology, metabolism and function of the normal
human brain 2) To characterize the molecular changes underlying addiction and their relationship to brain
function and vulnerability. 3) To explore common and distinct neural mechanisms underlying obesity and addiction. 4) To identify the neurobiological underpinnings of ADHD and the mechanism underlying the
therapeutic effects of stimulant drugs. 5) To explore physiological processes underlying human brain function.
Model of brain circuits involved with addiction: reward/motivation, conditioning/memory, and executive function/inhibitory control.

199
1. Drug Effects
Effects of acute and chronic alcohol on brain acetate metabolism Alcohol intoxication results in marked reductions in brain glucose metabolism, which we hypothesized reflect not just its GABAergic enhancing effects but also the metabolism of acetate as an alternative brain energy source. To test this hypothesis we separately assessed the effects of alcohol intoxication on brain glucose and acetate metabolism using PET. We found that alcohol intoxication significantly decreased whole brain glucose metabolism (measured with fludeoxyglucose (FDG)) with the largest decrements in cerebellum and occipital cortex and the smallest in the thalamus. In contrast, alcohol intoxication caused a significant increase in 11C-acetate brain uptake (measured as standard uptake value, SUV), with the largest increases occurring in the cerebellum and the smallest in the thalamus. In heavy alcohol drinkers 11C-acetate brain uptake during alcohol challenge tended to be higher than in occasional drinkers (p<0.06) and the increases in 11C-acetate uptake in cerebellum with alcohol were positively associated with the reported amount of alcohol consumed (r=0.66, p<0.01). Our findings corroborate a reduction of brain glucose metabolism during intoxication and document an increase in brain acetate uptake. The opposite changes observed between regional brain metabolic decrements and regional increases in 11C-acetate uptake support the hypothesis that during alcohol intoxication the brain may rely on acetate as an alternative brain energy source and provides preliminary evidence that heavy alcohol exposures may facilitate the use of acetate as an energy substrate. These findings raise the question of the potential therapeutic benefits that increasing plasma acetate concentration (i.e. ketogenic diets) may have in alcoholics undergoing alcohol detoxification as well as the role that energy deprivation during acute withdrawal may have on its detrimental effects to the human brain.
2. Addiction
Decreased activity in brain "control networks" after exposure to cocaine-cues in female cocaine abusers. Gender differences in vulnerability for cocaine addiction have been reported. Though the mechanisms are not understood, here we hypothesize that gender differences in reactivity to conditioned-cues, which contributes to relapse, are involved. To test this we compared brain metabolism (using positron emission tomography (PET) and FDG) between female (n = 10) and male (n=16) active cocaine abusers when they watched a neutral video (nature scenes) versus a cocaine-cues video. We show that self-reports of craving increased with the cocaine-cue video but responses did not differ between genders. In contrast, changes in whole brain metabolism with cocaine-cues differed by gender (p<0.05); females significantly decreased metabolism (-8.6%±10) whereas males tended to increase it (+5.5%±18). Statistical Parameteric Mapping analysis (Cocaine-cues vs Neutral) in females revealed decreases in frontal, cingulate and parietal cortices, thalamus and midbrain (p<0.001) whereas males showed increases in right inferior frontal gyrus (Brodmann Area (BA) 44/45) (only at p<0.005). The gender-cue interaction showed greater decrements with Cocaine-cues in females than males (p<0.001) in frontal (BA 8, 9, 10), anterior cingulate (BA 24, 32), posterior cingulate (BA 23, 31), inferior parietal (BA 40) and thalamus (dorsomedial nucleus). Females showed greater brain reactivity to cocaine-cues than males but no differences in craving, suggesting that there may be gender differences in response to cues that are not linked with craving but could affect subsequent drug use. Specifically deactivation of brain regions from "control networks" (prefrontal, cingulate, inferior parietal, thalamus) in females could increase their vulnerability to relapse since it would

200
interfere with executive function (cognitive inhibition). This highlights the importance of gender tailored interventions in female cocaine abusers to improve the function of “control brain networks” as a means to prevent relapse from cues exposures.
Decreased dopamine activity predicts relapse in methamphetamine abusers. Methamphetamine (METH) is harmful to dopamine (DA) cells but brain PET imaging studies have reported significant recovery in METH abusers that are successful at remaining abstinent for long time periods. However the extent to which differences in brain DA activity enabled them to remain drug free in contrast to those who did not succeed is unclear. Moreover, the extent to which striatal DA function in METH abusers predicts recovery has not been evaluated. Here we assessed whether striatal DA activity in METH abusers is associated with clinical outcomes. Brain DA D2 receptor (D2R) availability was measured with positron emission tomography and 11C-raclopride in 16 METH abusers, both after placebo and after challenge with 60 mg oral methylphenidate (MP) (to measure DA release) to assess whether it predicted clinical outcomes. For this purpose, METH abusers were tested within 6 months of last METH use and then followed up for 9 months of abstinence. In parallel, 15 healthy controls were tested. METH abusers had lower D2R availability in caudate than in controls. Both METH abusers and controls showed decreased striatal D2R availability after MPH and these decreases were smaller in METH than in controls in left putamen. The six METH abusers who relapsed during the follow-up period had lower D2R availability in dorsal striatum than in controls, and had no D2R changes after MP challenge. The 10 METH abusers who completed detoxification did not differ from controls neither in striatal D2R availability nor in MP-induced striatal DA changes. These results provide preliminary evidence that low striatal DA function in METH abusers is associated with a greater likelihood of relapse during treatment.
Common neural substrates underlying protective personality factors and SUD The personality construct of positive emotionality (PEM) (of well-being, achievement/motivation, social and closeness) is considered a trait that protects against substance use disorders (SUD). Higher scores have been associated with higher striatal dopamine D2 receptor availability in healthy controls. As high striatal D2 receptor (D2R) availability is protective against SUD and D2R modulate the activity in the orbitofrontal cortex (OFC) and anterior cingulate gyrus (brain regions that process natural and drug rewards), we hypothesized that these regions underlie PEM. To test this, we assessed the correlation between baseline brain glucose metabolism (measured with PET and DSG and scores on PEM (obtained from the multidimensional personality questionnaire or MPQ) in healthy controls (n = 47). Statistical parametric mapping (SPM) analyses revealed that PEM was positively correlated (P(c)<0.05, voxel corrected) with metabolism in various cortical regions that included orbitofrontal (BA 11, 47) and cingulate (BA 23, 32) and other frontal (BA 10, 9), parietal (precuneus, BA 40) and temporal (BA 20, 21) regions that overlap with the brain's default mode network (DMN). Correlations with the other two main MPQ personality dimensions (negative emotionality and constraint) were not significant (SPM P(c)<0.05). Our results corroborate an involvement of orbitofrontal and cingulate regions in PEM, which might mediate the protective effects against substance use disorders. As dysfunction of OFC and cingulate is a hallmark of addiction, these findings support a common neural basis underlying protective personality factors and brain dysfunction underlying SUD. In addition, we also uncovered an association between PEM and baseline metabolism in regions from the DMN, which suggests that PEM may relate to global cortical processes that are active during resting conditions (introspection, mind wandering).

201
3. Obesity Enhanced striatal DA release during food stimulation in binge eating disorder. Subjects with binge eating disorder (BED) regularly consume large amounts of food in short time periods. The neurobiology of BED is poorly understood. Brain dopamine, which regulates motivation for food intake, is likely to be involved. We assessed the involvement of brain dopamine in the motivation for food consumption in binge eaters. Positron emission tomography (PET) scans with 11C-raclopride were done in 10 obese BED and 8 obese subjects without BED. Changes in extracellular dopamine in the striatum in response to food stimulation in food-deprived subjects were evaluated after placebo and after oral methylphenidate (MP), a drug that blocks the dopamine reuptake transporter and thus amplifies dopamine signals. Neither the neutral stimuli (with or without MP) nor the food stimuli when given with placebo increased extracellular dopamine. The food stimuli when given with MP significantly increased dopamine in the caudate and putamen in the binge eaters but not in the nonbinge eaters. Dopamine increases in the caudate were significantly correlated with the binge eating scores but not with BMI. These results identify dopamine neurotransmission in the caudate as being of relevance to the neurobiology of BED. The lack of correlation between BMI and dopamine changes suggests that dopamine release per se does not predict BMI within a group of obese individuals but that it predicts binge eating
Striatal D2R predict future body weight and cocaine preference in rodents. Deficits in striatal dopamine D2/D3 receptor (D2R/D3R) binding availability using PET imaging have been reported in obese humans and rodents. Similar deficits have been reported in cocaine-addicts and cocaine-exposed primates. Here we use PET to assess if striatal D2R/D3R predicted weight gain and alcohol consumption in drug-naïve chow fed rats. We found that D2R/D3R binding availability negatively correlated with measures of body weight at the time of scan (ventral striatum) and at 1 month (ventral striatum) and 2 months (dorsal and ventral striatum) post scan measurements. We also showed that cocaine preference was negatively correlated with D2R/D3R binding availability at 2 months (ventral striatum) post scan measurements. Our findings suggest that inherent deficits in striatal D2R/D3R signaling may increase the vulnerability the rewarding effects of both food and drugs and that ventral and dorsal striatum might serve dissociable roles in modulating weight gain and cocaine preference. Alcohol reward is increased after Roux-en-Y gastric bypass in dietary obese rats: ghrelin’s involvement. Roux-en-Y gastric bypass (RYGB) is one of the most successful treatments for severe obesity and associated comorbidities. One potential adverse outcome, however, is increased risk for alcohol use. As such, we tested whether RYGB alters motivation to self-administer alcohol in outbred dietary obese rats, and investigated the involvement of the ghrelin system as a potential underlying mechanism. High fat (60% cal from fat) diet-induced obese, non-diabetic male Sprague Dawley rats underwent RYGB (n=9) or sham operation (Sham, n=9) and were tested 4 months after surgery on a progressive ratio-10 (PR10) schedule of reinforcement operant task for 2, 4, and 8% ethanol. In addition, the effects of the ghrelin-1a-receptor antagonist D-[Lys3]-GHRP-6 (50, 100 nmol/kg, IP) were tested on PR10 responding for 4% ethanol. Compared to Sham, RYGB rats made significantly more active spout responses to earn reward, more consummatory licks on the ethanol spout, and achieved higher breakpoints. Pretreatment with a single peripheral injection of D-[Lys3]-GHRP-6 at either dose was ineffective in altering

202
appetitive or consummatory responses to 4% ethanol in the Sham group. In contrast, RYGB rats demonstrated reduced operant performance to earn alcohol reward on the test day and reduced consummatory responses for two subsequent days following the drug. Sensitivity to threshold doses of D-[Lys3]-GHRP-6 suggests that an augmented ghrelin system may contribute to increased alcohol reward in RYGB. 4. ADHD and Therapeutic Mechanisms for Stimulants Abnormal functional connectivity in children with attention-deficit/hyperactivity disorder. Attention-deficit/hyperactivity disorder (ADHD) is typically characterized by symptoms of inattention and hyperactivity/impulsivity, but there is increased recognition of a motivation deficit too. This neuropathology may reflect dysfunction of both attention and reward-motivation networks. To test this hypothesis, we compared the functional connectivity density between 247 ADHD and 304 typically developing control children from a public magnetic resonance imaging database. We quantified short- and long-range functional connectivity density in the brain using an ultrafast data-driven approach. We showed that children with ADHD had lower connectivity (short- and long-range) in regions of the dorsal attention (superior parietal cortex) and default-mode (precuneus) networks and in cerebellum and higher connectivity (short-range) in reward-motivation regions (ventral striatum and orbitofrontal cortex) than control subjects. In ADHD children, the orbitofrontal cortex (region involved in salience attribution) had higher connectivity with reward-motivation regions (striatum and anterior cingulate) and lower connectivity with superior parietal cortex (region involved in attention processing). The enhanced connectivity within reward-motivation regions and their decreased connectivity with regions from the default-mode and dorsal attention networks suggest impaired interactions between control and reward pathways in ADHD that might underlie attention and motivation deficits in ADHD. Functional connectivity of midbrain DA nuclei: maturation and effects of ADHD. Dopaminergic pathways play crucial roles in brain function and their disruption is implicated in various neuropsychiatric diseases. Here, we demonstrate in 402 healthy children/adolescents (12 ± 3 years) and 704 healthy young adults (23 ± 5 years) that the functional connectivity of DA pathways matures significantly from childhood to adulthood and is different for healthy children and children with ADHD; N = 203; 12 ± 3 years). This transition is characterized by age-related increases in the functional connectivity of the ventral tegmental area (VTA) with limbic regions and with the default mode network and by decreases in the connectivity of the substantia nigra (SN) with motor and medial temporal cortices. The changes from a predominant influence of SN in childhood/adolescence to a combined influence of SN and VTA in young adulthood might explain the increased vulnerability to psychiatric disorders, such as ADHD, early in life. We also show that VTA and SN connectivity networks were highly reproducible, which highlights their potential value as biomarkers for evaluating dopaminergic dysfunction in neuropsychiatric disorders. Methylphenidate-elicited DA increases in ventral striatum are associated with symptom improvement in ADHD. Stimulant medications, such as methylphenidate (MP), which are effective treatments for ADHD, enhance brain DA signaling. However, the relationship between regional brain DA enhancement and treatment response has not been evaluated. Here, we assessed whether the DA increases elicited by MP are associated with long-term clinical response. We used a prospective

203
design to study 20 treatment-naive adults with ADHD who were evaluated before treatment initiation and after 12 months of clinical treatment with a titrated regimen of oral MP. MP-induced DA changes were evaluated with PET and 11C-raclopride (D(2)/D(3) receptor radioligand sensitive to competition with endogenous DA). Clinical responses were assessed using the Conners' Adult ADHD Rating Scale and revealed a significant reduction in symptoms of inattention and hyperactivity with long-term methylphenidate treatment. A challenge dose of 0.5 mg/kg intravenous MP significantly increased DA in striatum (assessed as decreases in D(2)/D(3) receptor availability). In the ventral striatum, these DA increases were associated with the reductions in ratings of symptoms of inattention with clinical treatment. Statistical parametric mapping additionally showed DA increases in prefrontal and temporal cortices with intravenous MP that were also associated with decreases in symptoms of inattention. Our findings indicate that DA enhancement in ventral striatum (the brain region involved with reward and motivation) was associated with therapeutic response to MP, further corroborating the relevance of the DA reward/motivation circuitry in ADHD. It also provides preliminary evidence that MP-elicited DA increases in prefrontal and temporal cortices may also contribute to the clinical response. Methylphenidate enhances brain activation and deactivation responses to visual attention and working memory tasks in healthy controls. Previous studies have suggested DA to be involved in error monitoring/processing, possibly through impact on reinforcement learning. The current study tested whether methylphenidate (MP), an indirect DA agonist, modulates brain and behavioral responses to error, and whether such modulation is more pronounced in cocaine-addicted individuals, in whom dopamine neurotransmission is disrupted. After receiving oral MP (20 mg) or placebo (counterbalanced), 15 healthy human volunteers and 16 cocaine-addicted individuals completed a task of executive function (the Stroop color word) during functional magnetic resonance imaging (fMRI). During MP, despite not showing differences on percent accuracy and reaction time, all subjects committed fewer total errors and slowed down more after committing errors, suggestive of more careful responding. In parallel, during MP all subjects showed reduced dorsal anterior cingulate cortex response to the fMRI contrast error>correct. In the cocaine subjects only, MP also reduced error>correct activity in the dorsolateral prefrontal cortex (controls instead showed lower error>correct response in this region during placebo). Taken together, MP modulated dopaminergically innervated prefrontal cortical areas involved in error-related processing, and such modulation was accentuated in the cocaine subjects. These results are consistent with a dopaminergic contribution to error-related processing during a cognitive control task.
Methylphenidate induced dopamine increase and associated stimulation of D2R is involved in its arousing effects during sleep deprivation. Stimulant drugs, which enhance DA signaling through DA transporter (DAT) blockade (methylphenidate, modafinil) or by releasing DA (amphetamine) increase wakefulness in human subjects and cognitive performance in sleep deprived subjects. This may reflect stimulation of DA D2 receptors (D2R) since they are involved with wakefulness. We had shown that sleep deprivation reduced DA D2/D3R availability (measured with PET and 11C-raclopride in controls) in striatum, but could not determine whether this reflected DA increases (11C-raclopride competes with dopamine for D2/D3R binding) or receptor downregulation. Since decreases in D2/D3 availability were associated with cognitive impairment rather than an expected improvement if they reflect DA increases, we compared these responses with and without the administration of methylphenidate (MP). Specifically, we compared the DA increases induced by MP during sleep deprivation versus rested sleep, with the assumption that MP's effects would be greater if, indeed, DA release was increased during sleep deprivation and that DA increases

204
would be associated with improvement in performance. We scanned 20 controls with 11C-raclopride after rested sleep and after 1 night of sleep deprivation; both after placebo and after MP. We corroborated a decrease in D2/D3R availability in the ventral striatum with sleep deprivation (compared with rested sleep) that was associated with reduced alertness and increased sleepiness. However, the DA increases induced by MP (measured as decreases in D2/D3 receptor availability compared with placebo) did not differ between rested sleep and sleep deprivation, and were associated with the increased alertness and reduced sleepiness when MP was administered after sleep deprivation. These findings are consistent with a downregulation of D2/D3 receptors in ventral striatum with sleep deprivation that may contribute to the associated decreased wakefulness and show that MP-induced enhancement of D2 receptor signaling is involved in the arousing effects of MP in humans. 5. Function of the normal human brain Effects of cell phone radiofrequency signal exposure on brain glucose metabolism. The dramatic increase in use of cellular telephones has generated concern about possible negative effects of radiofrequency signals delivered to the brain. However, whether acute cell phone exposure affects the human brain is unclear. This study evaluated if acute cell phone exposure affects brain glucose metabolism, a marker of brain activity. For this purpose we conducted a randomized crossover study conducted in 47 healthy participants. Cell phones were placed on the left and right ears and brain glucose metabolism was measured with PET and FDG. Subjects were tested twice, once with the right cell phone activated (sound muted) for 50 minutes ("on" condition) and once with both cell phones deactivated ("off" condition). Statistical parametric mapping was used to compare metabolism between on and off conditions using paired t tests, and Pearson linear correlations were used to verify the association of metabolism and estimated amplitude of radiofrequency-modulated electromagnetic waves emitted by the cell phone. Clusters with at least 1000 voxels (volume >8 cm) and P < .05 (corrected for multiple comparisons) were considered significant. We showed that whole-brain metabolism did not differ between on and off conditions. In contrast, metabolism in the region closest to the antenna (orbitofrontal cortex and temporal pole) was significantly higher for on than off conditions (35.7 vs 33.3 μmol/100 g per minute; mean difference, 2.4 [95% confidence interval, 0.67-4.2]; P = .004). The increases were significantly correlated with the estimated electromagnetic field amplitudes both for absolute metabolism (R = 0.95, P < .001) and normalized metabolism (R = 0.89; P < .001). In healthy participants and compared with no exposure, we show that 50-minute cell phone exposure was associated with increased brain glucose metabolism in the region closest to the antenna. This finding indicates that the human brain is sensitive to cell phone radiation though the clinical significance of this is unclear.
Brain functional connectivity of the normal human brain; organization into functional networks and the effects of gender and age. In order to investigate the organization of the normal brain we have analyzed resting-state data sets corresponding to 1031 healthy subjects from a public magnetic resonance imaging database (1000 Functional Connectomes) using functional connectivity density (FCD) mapping, a voxelwise and data-driven approach, together with parallel computing. We used the method to map local FCD and long range FCD.
To study the reliance of brain networks in local and long range connectivity we localized the regions with high functional connectivity density (hubs). We show that the strongest hubs

205
were consistently located in ventral precuneus/cingulate gyrus (previously identified by other analytical methods) and in primary visual cortex (BA 17/18), which highlights their centrality to resting connectivity networks. In contrast and after rescaling, hubs in prefrontal regions had lower long range FCD than local FCD, which suggests that their local functional connectivity (as opposed to long-range connectivity) prevails in the resting state. The power scaling of the probability distribution of local and long range FCD hubs was consistent across research centers further corroborating the "scale-free" topology of brain networks.
To assess the laterality patterns in FCD we quantified the local- and long-range functional connectivity densities in 913 healthy subjects. Local-range connectivity was rightward lateralized and most asymmetrical in areas around the lateral sulcus, whereas long-range connectivity was rightward lateralized in lateral sulcus and leftward lateralizated in inferior prefrontal cortex and angular gyrus. The posterior inferior occipital cortex was leftward lateralized (local- and long-range connectivity). Males had greater rightward lateralization of brain connectivity in superior temporal, inferior frontal, and inferior occipital cortices, whereas females had greater leftward lateralization in the inferior frontal cortex. The greater lateralization of the male's brain (rightward and predominantly local-range) may underlie their greater vulnerability to disorders with disrupted brain asymmetries (schizophrenia, autism).
To assess the effects of gender on RFC we compared the brain imaging data from 336 women and 225 men from this data set. We show gender dimorphism in the functional organization of the brain. Specifically, women had 14% higher short-range FCD and up to 5% higher gray matter density than men in cortical and subcortical regions. The negative power scaling of the short-range FCD was steeper for men than for women, suggesting that the balance between strongly and weakly connected nodes in the brain is different across genders. The more distributed organization of the male brain than that of the female brain could help explain the gender differences in cognitive style and behaviors and in the prevalence of neuropsychiatric diseases (i.e., autism spectrum disorder).
To assess the effects of age on RFC we evaluated brain imaging data sets from 913 healthy subjects. Aging was associated with pronounced long-range FCD decreases in default mode network (DMN) and dorsal attention network (DAN) and with increases in somatosensory and subcortical networks. Aging effects in these networks were stronger for long-range than for short-range FCD and were also detected at the level of the main functional hubs. These findings suggest that long-range connections may be more vulnerable to aging effects than short-range connections and that, in addition to the DMN, the DAN is also sensitive to aging effects, which could underlie the deterioration of attention processes that occurs with aging. Publications 2011-2012
Original studies
1. Alia-Klein N, Parvaz MA, Woicik PA, Konova AB, Maloney T, Shumay E, Wang R, Telang F, Biegon A, Wang GJ, Fowler JS, Tomasi D, Volkow ND, Goldstein RZ. 2011. Gene x disease interaction on orbitofrontal gray matter in cocaine addiction. Arch Gen Psychiatry 68: 283-94
2. Alzghoul L, Bortolato M, Delis F, Thanos PK, Darling RD, Godar SC, Zhang JL, Grant S, Wang GJ, Simpson KL, Chen K, Volkow ND, Lin RCS, Shih JC. 2012. Altered cerebellar organization and function in monoamine oxidase A hypomorphic mice. Neuropharmacology 63: 1208-17

206
3. Delis F, Benveniste H, Xenos M, Grandy D, Wang GJ, Volkow ND, Thanos PK. 2012. Loss of Dopamine D2 Receptors Induces Atrophy in the Temporal and Parietal Cortices and the Caudal Thalamus of Ethanol-Consuming Mice. Alcoholism-Clinical and Experimental Research 36: 815-25
4. Delis F, Thanos PK, Rombola C, Rosko L, Grandy D, Wang GJ, Volkow ND. 2012. Chronic mild stress increases alcohol intake in mice with low dopamine D2 receptor levels. Behav Neurosci doi: 10.1037/a0030750
5. Dunn JP, Kessler RM, Feurer ID, Volkow ND, Patterson BW, Ansari MS, Li R, Marks-Shulman P, Abumrad NN. 2012. Relationship of dopamine type 2 receptor binding potential with fasting neuroendocrine hormones and insulin sensitivity in human obesity. Diabetes Care 35: 1105-11
6. Dunning JP, Parvaz MA, Hajcak G, Maloney T, Alia-Klein N, Woicik PA, Telang F, Wang GJ, Volkow ND, Goldstein RZ. 2011. Motivated attention to cocaine and emotional cues in abstinent and current cocaine users--an ERP study. Eur J Neurosci 33: 1716-23
7. Goldstein RZ, Volkow ND. 2011. Oral Methylphenidate Normalizes Cingulate Activity and Decreases Impulsivity in Cocaine Addiction During an Emotionally Salient Cognitive Task. Neuropsychopharmacology 36: 366-7
8. Gonzalez S, Rangel-Barajas C, Peper M, Lorenzo R, Moreno E, Ciruela F, Borycz J, Ortiz J, Lluis C, Franco R, McCormick PJ, Volkow ND, Rubinstein M, Floran B, Ferre S. 2012. Dopamine D4 receptor, but not the ADHD-associated D4.7 variant, forms functional heteromers with the dopamine D2S receptor in the brain. Mol Psychiatry 17: 650-62
9. Hajnal A, Zharikov A, Polston JE, Fields MR, Tomasko J, Rogers AM, Volkow ND, Thanos PK. 2012. Alcohol Reward Is Increased after Roux-en-Y Gastric Bypass in Dietary Obese Rats with Differential Effects following Ghrelin Antagonism. Plos One 7
10. Hirvonen J, Goodwin RS, Li CT, Terry GE, Zoghbi SS, Morse C, Pike VW, Volkow ND, Huestis MA, Innis RB. 2012. Reversible and regionally selective downregulation of brain cannabinoid CB1 receptors in chronic daily cannabis smokers. Mol Psychiatry 17: 642-9
11. Komatsu DE, Thanos PK, Mary MN, Janda HA, John CM, Robison L, Ananth M, Swanson JM, Volkow ND, Hadjiargyrou M. 2012. Chronic exposure to methylphenidate impairs appendicular bone quality in young rats. Bone 50: 1214-22
12. Konova AB, Moeller SJ, Tomasi D, Parvaz MA, Alia-Klein N, Volkow ND, Goldstein RZ. 2012. Structural and behavioral correlates of abnormal encoding of money value in the sensorimotor striatum in cocaine addiction. Eur J Neurosci 36: 2979-88
13. Krasnova IN, Ladenheim B, Hodges AB, Volkow ND, Cadet JL. 2011. Chronic methamphetamine administration causes differential regulation of transcription factors in the rat midbrain. Plos One 6: e19179

207
14. Li ZZ, Xu YW, Warner D, Volkow ND. 2012. Alcohol ADME in Primates Studied with Positron Emission Tomography. Plos One 7
15. Luo ZC, Volkow ND, Heintz N, Pan YT, Du CW. 2011. Acute Cocaine Induces Fast Activation of D1 Receptor and Progressive Deactivation of D2 Receptor Striatal Neurons: In Vivo Optical Microprobe Ca2+ (i) Imaging. Journal of Neuroscience 31: 13180-90
16. Michaelides M, Thanos PK, Kim R, Cho J, Ananth M, Wang GJ, Volkow ND. 2012. PET imaging predicts future body weight and cocaine preference. Neuroimage 59: 1508-13
17. Moeller SJ, Hajcak G, Parvaz MA, Dunning JP, Volkow ND, Goldstein RZ. 2012. Psychophysiological prediction of choice: relevance to insight and drug addiction. Brain 135: 3481-94
18. Moeller SJ, Honorio J, Tomasi D, Parvaz MA, Woicik PA, Volkow ND, Goldstein RZ. 2012. Methylphenidate Enhances Executive Function and Optimizes Prefrontal Function in Both Health and Cocaine Addiction. Cereb Cortex
19. Moeller SJ, Tomasi D, Honorio J, Volkow ND, Goldstein RZ. 2012. Dopaminergic involvement during mental fatigue in health and cocaine addiction. Translational Psychiatry 2
20. Moeller SJ, Tomasi D, Woicik PA, Maloney T, Alia-Klein N, Honorio J, Telang F, Wang GJ, Wang RL, Sinha R, Carise D, Astone-Twerell J, Bolger J, Volkow ND, Goldstein RZ. 2012. Enhanced midbrain response at 6-month follow-up in cocaine addiction, association with reduced drug-related choice. Addiction Biology 17: 1013-25
21. Parvaz MA, Konova AB, Tomasi D, Volkow ND, Goldstein RZ. 2012. Structural integrity of the prefrontal cortex modulates electrocortical sensitivity to reward. J Cogn Neurosci 24: 1560-70
22. Parvaz MA, Maloney T, Moeller SJ, Woicik PA, Alia-Klein N, Telang F, Wang GJ, Squires NK, Volkow ND, Goldstein RZ. 2012. Sensitivity to monetary reward is most severely compromised in recently abstaining cocaine addicted individuals: A cross-sectional ERP study. Psychiatry Research-Neuroimaging 203: 75-82
23. Ren H, Du C, Yuan Z, Park K, Volkow ND, Pan Y. 2012. Cocaine-induced cortical microischemia in the rodent brain: clinical implications. Mol Psychiatry 17: 1017-25
24. Ren HG, Du CW, Park K, Volkow ND, Pan YT. 2012. Quantitative imaging of red blood cell velocity in vivo using optical coherence Doppler tomography. Applied Physics Letters 100
25. Shumay E, Chen J, Fowler JS, Volkow ND. 2011. Genotype and ancestry modulate brain's DAT availability in healthy humans. Plos One 6: e22754
26. Shumay E, Fowler JS, Wang GJ, Logan J, Alia-Klein N, Goldstein RZ, Maloney T, Wong C, Volkow ND. 2012. Repeat variation in the human PER2 gene as a new genetic

208
marker associated with cocaine addiction and brain dopamine D2 receptor availability. Transl Psychiatry 2: e86
27. Shumay E, Logan J, Volkow ND, Fowler JS. 2012. Evidence that the methylation state of the monoamine oxidase A (MAOA) gene predicts brain activity of MAO A enzyme in healthy men. Epigenetics 7: 1151-60
28. Thanos PK, Bermeo C, Wang GJ, Volkow ND. 2011. D-Cycloserine Facilitates Extinction of Cocaine Self-Administration in Rats. Synapse 65: 938-44
29. Thanos PK, Cho J, Kim R, Michaelides M, Primeaux S, Bray G, Wang GJ, Volkow ND. 2011. Bromocriptine increased operant responding for high fat food but decreased chow intake in both obesity-prone and resistant rats. Behavioural Brain Research 217: 165-70
30. Thanos PK, Gopez V, Delis F, Michaelides M, Grandy DK, Wang GJ, Kunos G, Volkow ND. 2011. Upregulation of Cannabinoid Type 1 Receptors in Dopamine D2 Receptor Knockout Mice Is Reversed by Chronic Forced Ethanol Consumption. Alcoholism-Clinical and Experimental Research 35: 19-27
31. Thanos PK, Robison LS, Robinson JK, Michaelides M, Wang GJ, Volkow ND. 2012. Obese rats with deficient leptin signaling exhibit heightened sensitivity to olfactory food cues. Synapse 67: 171-8 DOI: 10.1002/syn.21627
32. Thanos PK, Subrize M, Delis F, Cooney RN, Culnan D, Sun MJ, Wang GJ, Volkow ND, Hajnal A. 2012. Gastric Bypass Increases Ethanol and Water Consumption in Diet-Induced Obese Rats. Obesity Surgery 22: 1884-92
33. Thanos PK, Subrize M, Lui W, Puca Z, Ananth M, Michaelides M, Wang GJ, Volkow ND. 2011. D-Cycloserine Facilitates Extinction of Cocaine Self-Administration in c57 Mice. Synapse 65: 1099-105
34. Tomasi D, Volkow ND. 2011. Association between Functional Connectivity Hubs and Brain Networks. Cerebral Cortex 21: 2003-13
35. Tomasi D, Volkow ND. 2011. Functional connectivity hubs in the human brain. Neuroimage 57: 908-17
36. Tomasi D, Volkow ND. 2012. Resting functional connectivity of language networks: characterization and reproducibility. Molecular Psychiatry 17: 841-54
37. Tomasi D, Volkow ND. 2012. Laterality Patterns of Brain Functional Connectivity: Gender Effects. Cerebral Cortex 22: 1455-62
38. Tomasi D, Volkow ND. 2012. Aging and functional brain networks. Molecular Psychiatry 17: 549-58
39. Tomasi D, Volkow ND. 2012. Gender differences in brain functional connectivity density. Human Brain Mapping 33: 849-60
40. Tomasi D, Volkow ND. 2012. Abnormal Functional Connectivity in Children with Attention-Deficit/Hyperactivity Disorder. Biological Psychiatry 71: 443-50

209
41. Tomasi D, Volkow ND. 2012. Language network: segregation, laterality and connectivity. Mol Psychiatry 17: 759
42. Tomasi D, Volkow ND. 2012. Aging and functional brain networks. Mol Psychiatry 17: 471, 549-58
43. Tomasi D, Volkow ND. 2012. Functional Connectivity of Substantia Nigra and Ventral Tegmental Area: Maturation During Adolescence and Effects of ADHD. Cereb Cortex
44. Tomasi D, Volkow ND, Wang GJ, Wang R, Telang F, Caparelli EC, Wong C, Jayne M, Fowler JS. 2011. Methylphenidate enhances brain activation and deactivation responses to visual attention and working memory tasks in healthy controls. Neuroimage 54: 3101-10
45. Volkow ND, McLellan TA, Cotto JH, Karithanom M, Weiss SR. 2011. Characteristics of opioid prescriptions in 2009. JAMA 305: 1299-301
46. Volkow ND, Tomasi D, Vaska P. 2011. Cell Phone Activation and Brain Glucose Metabolism Reply. Jama-Journal of the American Medical Association 305: 2067-8
47. Volkow ND, Tomasi D, Wang GJ, Fowler JS, Telang F, Goldstein RZ, Alia-Klein N, Woicik P, Wong C, Logan J, Millard J, Alexoff D. 2011. Positive emotionality is associated with baseline metabolism in orbitofrontal cortex and in regions of the default network. Molecular Psychiatry 16: 818-25
48. Volkow ND, Tomasi D, Wang GJ, Fowler JS, Telang F, Goldstein RZ, Alia-Klein N, Wong C. 2011. Reduced Metabolism in Brain "Control Networks" following Cocaine-Cues Exposure in Female Cocaine Abusers. Plos One 6
49. Volkow ND, Tomasi D, Wang GJ, Telang F, Fowler JS, Logan J, Benveniste H, Kim R, Thanos PK, Ferre S. 2012. Evidence That Sleep Deprivation Downregulates Dopamine D2R in Ventral Striatum in the Human Brain. Journal of Neuroscience 32: 6711-7
50. Volkow ND, Tomasi D, Wang GJ, Vaska P, Fowler JS, Telang F, Alexoff D, Logan J, Wong C. 2011. Effects of Cell Phone Radiofrequency Signal Exposure on Brain Glucose Metabolism. Jama-Journal of the American Medical Association 305: 808-13
51. Volkow ND, Wang GJ, Newcorn JH, Kollins SH, Wigal TL, Telang F, Fowler JS, Goldstein RZ, Klein N, Logan J, Wong C, Swanson JM. 2011. Motivation deficit in ADHD is associated with dysfunction of the dopamine reward pathway. Molecular Psychiatry 16: 1147-54
52. Volkow ND, Wang GJ, Tomasi D, Kollins SH, Wigal TL, Newcorn JH, Telang FW, Fowler JS, Logan J, Wong CT, Swanson JM. 2012. Methylphenidate-elicited dopamine increases in ventral striatum are associated with long-term symptom improvement in adults with attention deficit hyperactivity disorder. J Neurosci 32: 841-9
53. Wang GJ, Geliebter A, Volkow ND, Telang FW, Logan J, Jayne MC, Galanti K, Selig PA, Han H, Zhu W, Wong CT, Fowler JS. 2011. Enhanced Striatal Dopamine Release During Food Stimulation in Binge Eating Disorder. Obesity 19: 1601-8

210
54. Wang GJ, Smith L, Volkow ND, Telang F, Logan J, Tomasi D, Wong CT, Hoffman W, Jayne M, Alia-Klein N, Thanos P, Fowler JS. 2012. Decreased dopamine activity predicts relapse in methamphetamine abusers. Molecular Psychiatry 17: 918-25
55. Woicik PA, Urban C, Alia-Klein N, Henry A, Maloney T, Telang F, Wang GJ, Volkow ND, Goldstein RZ. 2011. A pattern of perseveration in cocaine addiction may reveal neurocognitive processes implicit in the Wisconsin Card Sorting Test. Neuropsychologia 49: 1660-9
56. Yuan Z, Luo Z, Volkow ND, Pan Y, Du C. 2011. Imaging separation of neuronal from vascular effects of cocaine on rat cortical brain in vivo. Neuroimage 54: 1130-9
57. Zhang S, Huang J, Uzunbas M, Shen T, Delis F, Huang X, Volkow N, Thanos P, Metaxas DN. 2011. 3D segmentation of rodent brain structures using hierarchical shape priors and deformable models. Med Image Comput Comput Assist Interv 14: 611-8
Review and theoretical papers and editorials 2011-2012
1. Baler RD, Volkow ND. 2011. Addiction as a systems failure: focus on adolescence and smoking. J Am Acad Child Adolesc Psychiatry 50: 329-39
2. Goldstein RZ, Volkow ND. 2011. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nature Reviews Neuroscience 12: 652-69
3. Javitt DC, Schoepp D, Kalivas PW, Volkow ND, Zarate C, Merchant K, Bear MF, Umbricht D, Hajos M, Potter WZ, Lee CM. 2011. Translating glutamate: from pathophysiology to treatment. Sci Transl Med 3: 102mr2
4. Kalivas PW, Volkow ND. 2011. New medications for drug addiction hiding in glutamatergic neuroplasticity. Mol Psychiatry 16: 974-86
5. Michaelides M, Thanos PK, Volkow ND, Wang GJ. 2012. Dopamine-related frontostriatal abnormalities in obesity and binge-eating disorder: Emerging evidence for developmental psychopathology. International Review of Psychiatry 24: 211-8
6. Parvaz MA, Alia-Klein N, Woicik PA, Volkow ND, Goldstein RZ. 2011. Neuroimaging for drug addiction and related behaviors. Rev Neurosci 22: 609-24
7. Skolnick P, Volkow ND. 2011. Editorial: magic bullets and arrows: biologic approaches to treat substance use disorders. CNS Neurol Disord Drug Targets 10: 864
8. Skolnick P, Volkow ND. 2012. Addiction therapeutics: obstacles and opportunities. Biol Psychiatry 72: 890-1
9. Swanson J, Baler RD, Volkow ND. 2011. Understanding the effects of stimulant medications on cognition in individuals with attention-deficit hyperactivity disorder: a decade of progress. Neuropsychopharmacology 36: 207-26

211
10. Swanson JM, Wigal TL, Volkow ND. 2011. Contrast of medical and nonmedical use of stimulant drugs, basis for the distinction, and risk of addiction: comment on Smith and Farah (2011). Psychol Bull 137: 742-8
11. Tomasi D, Volkow ND. 2012. Striatocortical pathway dysfunction in addiction and obesity: differences and similarities. Crit Rev Biochem Mol Biol DOI: 10.3109/10409238.2012.735642
12. Volkow ND. 2011. Epigenetics of nicotine: another nail in the coughing. Sci Transl Med 3: 107ps43
13. Volkow ND. 2012. Long-term safety of stimulant use for ADHD: findings from nonhuman primates. Neuropsychopharmacology 37: 2551-2
14. Volkow ND, Baler RD. 2012. Neuroscience. To stop or not to stop? Science 335: 546-8
15. Volkow ND, Baler RD, Goldstein RZ. 2011. Addiction: pulling at the neural threads of social behaviors. Neuron 69: 599-602
16. Volkow ND, Baler RD, Normand JL. 2011. The unrealized potential of addiction science in curbing the HIV epidemic. Curr HIV Res 9: 393-5
17. Volkow ND, McLellan TA. 2011. Curtailing diversion and abuse of opioid analgesics without jeopardizing pain treatment. JAMA 305: 1346-7
18. Volkow ND, Montaner J. 2011. The urgency of providing comprehensive and integrated treatment for substance abusers with HIV. Health Aff (Millwood) 30: 1411-9
19. Volkow ND, Muenke M. 2012. The genetics of addiction. Hum Genet 131: 773-7
20. Volkow ND, Skolnick P. 2012. New medications for substance use disorders: challenges and opportunities. Neuropsychopharmacology 37: 290-2
21. Volkow ND, Wang GJ, Baler RD. 2011. Reward, dopamine and the control of food intake: implications for obesity. Trends Cogn Sci 15: 37-46
22. Volkow ND, Wang GJ, Fowler JS, Tomasi D. 2012. Addiction circuitry in the human brain. Annu Rev Pharmacol Toxicol 52: 321-36
23. Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baler R. 2012. Food and drug reward: overlapping circuits in human obesity and addiction. Curr Top Behav Neurosci 11: 1-24
24. Volkow ND, Wang GJ, Fowler JS, Tomasi D, Telang F. 2011. Addiction: Beyond dopamine reward circuitry. Proceedings of the National Academy of Sciences of the United States of America 108: 15037-42
Dr. Volkow: [email protected]

212
______________________________________________________________________
Laboratory of Epidemiology and Biometry ______________________________________________________________________ Bridget F. Grant, PhD, PhD, Chief Risë B. Goldstein, PhD, Staff Scientist Jeesun Jung, PhD, Staff Scientist Haitao Zhang, PhD, Staff Scientist S. Patricia Chou, PhD, Mathematical Statistician Tulshi D. Saha, PhD, Survey Statistician Boji Huang, PhD, MD, Epidemiologist Sharon M. Smith, PhD, Epidemiologist Roger P. Pickering, MS, Survey Statistician W. June Ruan, MA, Survey Statistician Veronica Wilson, Staff Assistant The Laboratory of Epidemiology and Biometry (LEB) develops, designs, implements, and directs a large multidisciplinary epidemiologic research program of national scope and complexity. In addition to the design, collection, cleaning and dissemination of large-scale, nationally representative survey data on alcohol use and its related conditions, the Laboratory: (1) plans, directs and implements large complex national epidemiologic, including genetic epidemiologic surveys on alcohol and drug use disorders and their associated physical and psychiatric disabilities; (2) develops, plans, monitors and provides management and statistical support to numerous international epidemiologic surveys; (3) collects, processes, compiles and analyzes national and international epidemiologic survey data; (4) provides expert consultation to NIAAA Extramural Divisions related to statistics, epidemiology, and survey research; (5) serves as principal advisor and consultant to Director, NIAAA on all areas of statistics, epidemiology, and survey methodology necessary to achieve the mission and goals of NIAAA and NIH; (6) conducts an independent research program in statistical, survey and epidemiologic arenas; (7) reviews, develops and assesses need for additional support and funds for areas of program responsibilities; (8) analyzes epidemiologic data on alcohol use, abuse and dependence and their associated disabilities and prepares scientific reports and manuscripts for publication in peer-reviewed journals, books, monographs and extensive compendia; (9) adapts state-of-the-art statistical and survey methodology to the alcohol field; (10) collaborates with other agencies and organization to promote the application of epidemiologic research nationally and internationally; (11) maintains national statistics on alcohol use disorders and their related conditions and disabilities, and consequences; (12) sponsors, develops, and participates in scientific conferences, meetings, workshops and symposia to exchange information and disseminate new knowledge; (13) collaborates with other National Institutes of Health and other government and outside agencies to foster shared interests and goals; (14) collaborates with national and international agencies and organizations on complex issues associated with the global burden of disease as it relates to alcohol use disorders and alcohol as a risk factor of numerous physical and mental conditions; and (15) supports a full range of interagency agreements, contracts, and collaborative grants to implement the LEB and NIAAA mission. The primary focus of LEB’s work during 2011 and 2012 has been directed toward the planning, preparation and fielding of the National Epidemiologic Survey on Alcohol and Related

213
Conditions-III (NESARC-III). The target population of the NESARC-III is the noninstitutionalized population, 18 years and older, residing in households and noninstitutional group quarters in the United States, including the District of Columbia, Alaska and Hawaii. The NESARC-III design called for oversampling on Blacks, Hispanics and Asians. Designed to measure a broad range of substance use and other psychiatric disorders based on the Diagnostic and Statistical Manual of Mental Disorders–Fifth Edition (DSM-5), the NESARC-III is the largest national survey on alcohol use disorder and related conditions to additionally collect DNA for whole genome sequencing ever conducted in the United States. Using the NIAAA Alcohol Use Disorder and Associated Disabilities Interview Schedule-DSM-5 version (AUDADIS-5), comprehensive information is collected on alcohol consumption, drug use, alcohol and 10 drug use disorders, as well as major mood (e.g., major depressive disorder), anxiety (e.g., generalized anxiety disorder), eating (e.g., anorexia nervosa) and personality (e.g., antisocial personality) disorders. The NESARC-III also introduced a new battery of risk factors including race-ethnic orientation, acculturation, perceived discrimination, stressful life events, sexual orientation, attraction and preference, alcohol-related medical conditions, family history of substance use and associated disorders, social networks, adverse childhood experiences and a variety of sociodemographic factors. During 2011 and 2012, LEB published 80 papers primarily using NIAAA’s prior longitudinal surveys, the 2001-2002 and 2004-2005 Waves 1 and 2 NESARC. These studies focused on several areas including: (1) alcohol abuse and dependence: recovery/relapse/early onset/course/ comorbidity/treatment/stigma/discrimination; (2) alcohol consumption/drinking guidelines/ screening for alcohol and drug use disorders; (3) research examining the nature of DSM-IV and DSM-5 diagnoses; (4) syndromes of antisocial behavior; (5) intimate partner violence (IPV); (6) special populations; (7) cross-national research; (8) suicide/suicidal ideation; and (9) posttraumatic stress disorder. Dimensionality of DSM-IV Substance Use Disorders One major change to substance use disorders in the upcoming DSM-5 will be to combine DSM-IV substance abuse and dependence criteria to create one single diagnosis of substance use disorder for each drug class appearing in the Manual. Another change proposed for the substance use disorder definitions to appear in the DSM-5 is to eliminate the legal problems criterion. Waves 1 and 2 NESARC data were used to determine the dimensionality of DSM-IV alcohol and drug use disorders, the effect of eliminating the legal problem criterion on the informational value of the diagnoses and the applicability of the proposed diagnoses for sex, age and race-ethnic subgroups of the population. These studies found support for the dimensionality of alcohol, nicotine, amphetamine, cocaine, sedative, tranquilizer, opioid, inhalant/solvent and hallucinogen abuse and dependence criteria. The 1-factor solution was shown to be the best fit for abuse and dependence criteria for alcohol and each drug, consistent with the corresponding Item Response Theory (IRT) analyses. Comparisons of IRT models including and excluding the legal problems criterion showed models without the criterion to have a significantly better fit than those with the criterion for alcohol and all drugs examined. Aggregate criterion information functions for each drug visually depicted a slight reduction in information from the models that included the legal problems criterion to models that excluded that criterion, especially at the severe end of the continuum. However, there were no statistical differences in the information value of the IRT models with and without the legal problems criterion, supporting the elimination of that criterion. All alcohol and drug use disorder abuse and dependence criteria demonstrated no differential criterion functioning across important subgroups of the population

214
defined by sex, age and race-ethnicity. This study showed that alcohol, nicotine, amphetamine, cocaine, sedative, tranquilizer, opioid, inhalant/solvent, and hallucinogen abuse and dependence criteria reflected underlying unitary dimensions of severity. The legal problems criterion associated with each of these disorders could also be eliminated with no loss of informational value and an advantage of parsimony. Importantly, DSM-IV abuse and dependence criteria demonstrated invariance and lack of bias of the alcohol and drug use disorder diagnoses across subgroups of the population. Taken together, these findings support the changes to substance use disorder diagnoses recommended by the American Psychiatric Association’s DSM-5 Substance and Related Disorders Workgroup. Representative publications: Kerridge BT, Saha TD, Smith S, Chou PS, Pickering RP, et al. 2011. Dimensionality of hallucinogen and inhalant/solvent abuse and dependence criteria: implications for the Diagnostic and Statistical Manual of Mental Disorders-Fifth Edition. Addict Behav 36:912-918. Keyes KM, Krueger RF, Grant BF, Hasin DS, 2011. Alcohol craving and the dimensionality of alcohol disorders. Psychol Med 41:629-640. Saha TD, Compton WM, Chou SP, Smith S, Ruan WJ, et al. 2012. Analyses related to the development of DSM-5 criteria for substance use related disorders 1. Toward amphetamine, cocaine and prescription drug use disorder continua using Item Response Theory. Drug Alcohol Depend 122:38-46. Shmulewitz D, Keyes KM, Wall MM, Aharonovich E, Aivadyan C, et al. 2011. Nicotine dependence, abuse and craving: dimensionality in an Israeli sample. Addiction 106:1675-1686. U.S. Drinking Guidelines The U.S. is one of only a handful of countries with 2 sets of drinking guidelines. The Dietary Guidelines for Americans define moderate drinking as no more than 2 drinks per day for men and no more than 1 drink per day for women, whereas the NIAAA low-risk drinking guidelines define low-risk drinking as no more than 14 drinks per week and no more than 4 on any day for men and no more than 7 drinks per week and no more than 3 on any day for women. We addressed the “gray area” of consumption between these 2 sets of limits, using data from Waves 1 and 2 of the NESARC, by examining the risks of various outcomes among individuals who exceeded the Dietary Guidelines moderate drinking limits but drank within the NIAAA low-risk drinking guidelines. The results showed that drinkers were evenly divided among those who drank less than the Dietary Guidelines limits, those in the gray area of consumption and those who exceeded the NIAAA low-risk limits (32%, 29% and 39%, respectively). Among those in the gray area of consumption, there was a slightly more than 2-fold increase in the risk of concurrent alcohol dependence. However, the absolute prevalence of dependence was very low and gray area drinkers accordingly accounted for less than 3% of all cases of dependence. Gray area drinkers also had a slightly increased risk of job loss, but had a decreased risk of concurrent injury. The prospective risk of developing alcohol dependence over the follow-up interval was increased by 77% (OR = 1.77) among gray area drinkers, accounting for 8.7% of incident cases of dependence; however, almost all of these occurred among individuals whose consumption had

215
increased enough that they no longer fell into the gray area of consumption by the onset of dependence. Although the harms associated with the gray level of consumption reflected either a low absolute risk of harm or a low level of relative risk, the large number of gray area drinkers resulted in increased levels of harm that were not altogether negligible. For example, almost 215,000 cases of alcohol dependence were attributable to gray area consumption. However, the great majority of prospective harm, for which causal inferences are more strongly supported, occurred among drinkers who consumption had increased beyond the gray area into risk drinking levels. Thus one of the greatest risks associated with gray area consumption is the risk of increasing consumption to risk levels over time. In another study, a novel statistical approach was applied to data from the Waves 1 and 2 NESARC to compare 27 alternative sets of drinking guidelines based on a wide range of weekly and daily drinking limits in terms of their ability to predict 9 different types of concurrent and prospective alcohol-related harm. The study was unique in looking at a wide range of performance measures, including sensitivity and specificity, ORs, the proportion of variance explained, and various other statistics, all of which were rescaled so that they could be averaged without favoring one measure of performance over another. This study found that for all types of harm combined, daily-only limits of 4 drinks for men and 3 drinks for women performed almost as well as when these daily limits were combined with weekly limits of 14 drinks for both men and women. The performance of the various drinking limits varied by type of harm, with no one combination of weekly and daily limits best predicting all types of harm. The guidelines that best predicted concurrent harm consisted of daily-only limits of 4 drinks per day for men and 3 drinks per day for women; but gender-invariant limits of 4 drinks per day for men and women also performed well. Further, the guidelines that best predicted prospective harm included daily limits of 4 drinks for both men and women combined with weekly limits of 14 drinks for men and 7 drinks for women. This paper provided a template though which different countries could evaluate their own drinking guidelines and was featured in a special edition of Drug and Alcohol Review devoted to international drinking guidelines. Its findings showed that the NIAAA low-risk drinking guidelines (no more than 14 drinks per week and no more than 4 drinks on any day for men; no more than 7 drinks per week and no more than 4 drinks on any day for women), originally developed to reduce the risk of AUDs, performed very well in predicting other types of harm as well. At the same time, they indicated that the guidelines could be simplified by eliminating the weekly limits with little loss of predictive ability. Representative publications: Dawson DA. 2011. Defining risk drinking. Alcohol Res Health 34:144-156. Dawson DA, Grant BF, 2011. The “gray area” of consumption between moderate and risk drinking. J Stud Alcohol Drugs 72:453-458. Dawson DA, Smith SM, Pickering RP, Grant BF, 2012. Am empirical approach to evaluating the validity of alternative low-risk drinking guidelines. Drug Alcohol Rev 31:141-150.

216
Updating Mortality and Potential Terms of Life Lost Attributable to Alcohol in the U.S./Global Burden of Disease Estimates for Alcohol as a Risk Factor and Alcohol Use Disorders DALYs Alcohol has been linked to health disparities between races in the U.S.; however, race-specific alcohol-attributable mortality has never been estimated. The objective of this paper was to estimate premature mortality attributable to alcohol in the U.S. in 2005, differentiated by race, age and sex for people 15 to 64 years of age. Mortality attributable to alcohol was estimated based on alcohol-attributable fractions using indicators of exposure from the National Epidemiologic Survey on Alcohol and Related Conditions and risk relations from the WHO Comparative Risk Assessment study. Consumption data were corrected for undercoverage (the observed underreporting of alcohol consumption when using survey as compared to sales data) using adult per capita consumption from WHO databases. Mortality data by cause of death were obtained from the US Department of Health and Human Services. For people 15 to 64 years of age in the US in 2005, alcohol was responsible for 55,974 deaths (46,461 for men; 9,513 for women) representing 9.0% of all deaths, and 1,288,700 Potential Years of Life Lost (PYLL) (1,087,280 for men; 201,420 for women) representing 10.7% of all PYLL. Per 100,000 people, this represents 29 deaths (29 for White; 40 for Black; 82 for Native Americans; 6 for Asian/Pacific Islander) and 670 PYLL (673 for White; 808 for Black; 1,808 for Native American; 158 for Asian/Pacific Islander). Sensitivity analyses showed a lower but still substantial burden without adjusting for undercoverage. In conclusion, the burden of mortality attributable to alcohol in the US is unequal among people of different races and between men and women. Racial differences in alcohol consumption and the resulting harms explain in part the observed disparities in the premature mortality burden between races, suggesting the need for interventions for specific subgroups of the population such as Native Americans. LEB staff also served on the International Expert Alcohol Group of the 2010 Global Burden of Disease (GBD) Study to update global burden of disease estimates for alcohol as a risk factor and estimates of alcohol use disorders. Quantification of the disease burden caused by different risks informs prevention by providing an account of health loss different to that provided by a disease-by-disease analysis. No complete revision of global disease burden caused by alcohol use has been done since the comparative risk assessment in 2000. The 2010 GBD Study estimated deaths and disability-adjusted life years (DALYs) attributable to the independent effects of 67 factors and clusters of risk factors including alcohol for 21 regions in 1990 and 2010. Exposure distributions for each year, region, sex, and age group, and relative risks per unit of exposure were estimated by systematically reviewing and synthesizing published and unpublished data. These estimates, together with estimates of cause-specific deaths and DALYs from the Global Burden of Disease Study 2010, were used to calculate the burden attributable to each risk factor exposure compared with the theoretical-minimum-risk exposure. Uncertainty in disease burden, relative risks, and exposures were incorporated into estimates of attributable burden. In 2010, the three leading risk factors for global disease burden were high blood pressure (7·0% [95% uncertainty interval 6·2-7·7] of global DALYs), tobacco smoking including second-hand smoke (6·3% [5·5-7·0]), and alcohol use (5·5% [5·0-5·9]). In 1990, alcohol use was the sixth leading risk factor, 28% lower than in 2010. Between 1990 and 2010, the contribution of alcohol

217
use to disease burden has substantially increased, consistent with an overall shift in global burden away from risks for communicable diseases in children towards those for noncommunicable diseases in adults. LEB staff also substantially contributed to the 2010 GBD study to update global burden of disease estimates for alcohol use disorder. Measuring disease burden, including alcohol use disorders, in populations requires a composite metric that captures both premature mortality and the prevalence and severity of ill-health. No comprehensive update of disease burden worldwide incorporating a systematic reassessment of disease-specific epidemiology has been done since the 1990 GBD study. Disease burden was calculated worldwide and for 21 regions for 1990 and 2010 with methods to enable meaningful comparisons over time. DALYs were calculated as the sum of years of life lost (YLLs) and years lived with disability (YLDs). DALYs were calculated for 291 causes, 20 age groups, both sexes, and for 187 countries, and aggregated to regional and global estimates of disease burden for three points in time with strictly comparable definitions and methods. YLLs were calculated from age-sex-country-time-specific estimates of mortality by cause, with death by standardized lost life expectancy at each age. YLDs were calculated by age, sex, and cause, and weighted by new disability weights for each health state. Neither YLLs nor YLDs were age-weighted or discounted. Uncertainty around cause-specific DALYs was calculated for levels of all-cause mortality, cause-specific mortality, prevalence, and disability weights. Although global all cause DALYs remained stable from 1990 (2.503 billion) to 2010 (2.490 billion), alcohol use disorder DALYs rose from 13.143 million to 17.644 million between those years. These major findings support the epidemiological transition from communicable to noncommunicable diseases. The rising global burden of alcohol use disorders will impose new challenges for health infrastructures worldwide. Representative publications: Lim SS, Vos T, Flaxman AD, Danaei G, Shibuya K, et al. 2012. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study. Lancet 380:2224-2260. Shield K, Gmel G, Kehoe-Chan T, Dawson DA, Grant BF, Rehm J. 2013. Mortality and potential years of life lost attributable to alcohol consumption by race and sex in the United States in 2005. pLOS One 8:e51923. doi:10.1371/journal.pone.0051923. Murray CJL, Vos T, Lozano R, Naghavi M, Flaxman AD et al. 2012. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis of the Global Burden of Disease Study. Lancet 380:2197-2223. International Studies Despite a large body of evidence documenting the associations between alcohol and injuries, data from Asian countries including South Korea are sparse. The aim of the first comparative study was to investigate the associations between episodic heavy past-year drinking, problem drinking symptomatic of alcohol dependence and alcohol-related and intentional injuries. Data

218
from 1989 injured patients recruited for the WHO/NIAAA Collaborative Study on Alcohol and Injury in South Korea were analyzed with respect to the prevalence rates and associations between injuries and frequency of past-year episodic heavy drinking and problem drinking. In estimating the odds ratios (ORs) and the associated 95% confidence intervals between alcohol intake and injuries multivariable logistic models were employed to adjust for sociodemographic characteristics and selected drinking variables. Findings of this study were consistent with prior studies: the risk of alcohol-related or intentional injury was positively associated with the frequency of episodic heavy drinking. The magnitudes of the associations were large with frequent consumption of 5+ drinks (OR=4.0 approximately), greater than the frequent consumption of 12+ drinks (OR=3.1). Strong associations were also noted between RAPS4-assessed alcohol dependence and alcohol-related and intentional injuries. Further, the prevalence of intentional injury and its association with alcohol increased sharply once the acute alcohol intake exceeded 90 ml. Our results were consistent with prior studies that episodic heavy consumption, acute intoxication and problem drinking are pervasive among emergency room patients. Results of our study also lent support for administering a single-item screener querying consumption of 5+ drinks at a sitting in the past 12 months as triage tool in Korea. A second comparative study examined the pattern of comorbidity of alcohol use disorders (AUDs) and nicotine dependence (ND) between the culturally diverse countries of the United States and South Korea. Using the nationally representative samples of the U.S. and Korean general populations, we directly compared rates and comorbidity patterns of AUDs, ND, and mood and anxiety disorders between the 2 countries. We further examined the rates and the comorbidity pattern among individuals with AUDs who sought treatment in the last 12 months. The 12-month prevalence rates of AUDs, ND, and any mood disorder and any anxiety disorder were 9.7, 14.4, 9.5, and 11.9% among Americans, whereas the corresponding rates were 7.1, 6.6, 2.0, and 5.2% among Koreans. These rates were significantly greater (except for any AUD) among Americans than among their Korean counterparts. With respect to comorbidity, both countries showed comparable patterns: the prevalence rates of mood and anxiety disorders were consistently the highest among persons with alcohol dependence (AD). Also, a disparate pattern was observed in Korea in that the prevalence rates of mood and anxiety disorders were generally lower among individuals with ND than among those with alcohol abuse and AD. Furthermore, despite significantly greater prevalence of AD in Korea (5.1%) than in the United States (4.4%), alcohol-dependent Americans were 4 times (OR=3.93) more likely to seek treatment compared to their Korean counterparts. The results indicated that the prevalence of AD in Korea was substantially greater than that in both Western and other Asian countries, suggesting a maladaptive pattern of alcohol use in Korea, which is different from the general use pattern of other East Asian countries. The low rate of treatment utilization among Koreans might be attributable to perceived social stigma toward SUDs or mental health problems despite the fact that the Korean government offers national health insurance. Representative publications: Chou, SP, Chun S, Smith S, Ruan J, Li T-K, Grant BF. 2012. Episodic heavy drinking, problem drinking and injuries – results of the WHO/NIAAA Collaborative Emergency Room Study in South Korea. Alcohol 46:407-413.

219
Chou SP, Lee HK, Cho MJ, Park JI, Dawson DA, Grant BF. 2012. Alcohol use disorders, nicotine dependence, and co-occurring mood and anxiety disorders in the United States and South Korea – a cross-national comparison. Alcohol Clin Exp Res 36:654-662. 2011-2012 Publications 1. Keyes KM, Hatzenbuehler ML, Hasin DS. 2011. Stressful life experience, alcohol
consumption and alcohol use disorders: epidemiologic evidence for four main types of stressors. Psychopharmacol 218:1-17.
2. Wang K, Diskin SJ, Zhang H, Attiyeh EF, Winter C, et al. 2011. Integrative genomics identifies LMO1 as a neuroblastoma oncogene. Nature 469:216-220.
3. Zhao J, Bradfield JP, Zhang H, Sleiman PM, Kim CE, et al. 2011. Role of BMI-associated loci identified in GWAS meta-analyses in the context of common childhood obesity in European Americans. Obesity 19:2436-2439
4. Baca-Garcia E, Perez-Rodriguez MM, Oquendo MA, Keyes KM, Hasin DS, et al. 2011. Estimating risk for suicide attempt: are we asking the right questions? Passive suicidal ideation as a marker for suicidal behavior. J Affect Disord 134:327-332.
5. Hasin D, Fenton MC, Skodol A, Krueger R, Keyes K, et al. 2011. Personality disorders and the 3-year course of alcohol, drug, and nicotine use disorders. Arch Gen Psychiatry 68:1158-1167.
6. Kerridge BT, Saha TD, Smith S, Chou PS, Pickering RP, et al. 2011. Dimensionality of hallucinogen and inhalant/solvent abuse and dependence criteria: implications for the Diagnostic and Statistical Manual of Mental Disorders-Fifth Edition. Addict Behav 36:912-918.
7. Shmulewitz D, Keyes KM, Wall MM, Aharonovich E, Aivadyan C, et al. 2011. Nicotine dependence, abuse and craving: dimensionality in an Israeli sample. Addiction 106:1675-1686.
8. Dawson DA. 2011. Defining risk drinking. Alcohol Res Health 34:144-156.
9. Xu Y, Okuda M, Hser YI, Hasin D, Liu SM, et al. 2011. Twelve-month prevalence of psychiatric disorders and treatment-seeking among Asian Americans/Pacific Islanders in the United States: results from the National Epidemiological Survey on Alcohol and Related Conditions. J Psychiatr Res 45:910-918.
10. Hu Y, Liu Y, Jung J, Dunker AK, Wang Y. 2011. Changes in predicted protein disorder tendency may contribute to disease risk. BMC Genomics 12 Suppl 5:S2.
11. Pickering RP, Goldstein RB, Hasin DS, Blanco C, Smith SM, et al. 2011. Temporal relationships between overweight and obesity and DSM–IV substance use, mood, and anxiety disorders: results from a prospective study, the National Epidemiologic Survey on Alcohol and Related Conditions. J Clin Psychiatry 72:1494-1502.
12. Jung J, Dantzer J, Liu Y. 2011. Identification of multiple rare variants associated with a disease. BMC Proc 5:S103.
13. Teng M, Wang Y, Wang G, Jung J, Edenberg HJ, et al. 2011. Prioritizing single

220
nucleotide variations potentially regulating alternative splicing. BMC Proc 5:S40.
14. Wang K, Zhang H, Bloss CS, Duvvuri V, Kaye W, et al. 2011. A genome-wide association study on common SNPs and rare CNVs in anorexia nervosa. Mol Psychiatry 16:949-959.
15. Bradfield JP, Qu HQ, Wang K, Zhang H, Sleiman PM, et al. 2011. A genome-wide meta-analysis of six type 1 diabetes cohorts identifies multiple associated loci. PLoS Genet 7:e1002293.
16. Okuda M, Olfson M, Hasin D, Grant BF, Lin KH, Blanco C. 2011. Mental health of victims of intimate partner violence: results from a national epidemiologic survey. Psychiatr Serv 62:959-962.
17. Dawson DA, Grant BF. 2011. The “gray area” of consumption between moderate and risk drinking. J Stud Alcohol Drugs 72:453-458.
18. Dawson DA. 2011. The challenge of distinguishing high volume and frequent risky drinking. Addiction 106:1046-1047.
19. López-Quintero C, Hasin DS, Pérez de los Cobos J, Pines A, Wang S, et al. 2011. Probability and predictors of remission from lifetime nicotine, alcohol, cannabis, or cocaine dependence: results from the National Epidemiologic Survey on Alcohol and Related Conditions. Addiction 106:657-659.
20. Skodol AE, Grilo CM, Keyes KM, Geier T, Grant BF, Hasin DS. 2011. Relationship of personality disorders to the course of major depressive disorder in a nationally representative sample. Am J Psychiatry 168:257-264.
21. Cox BJ, Turnbull DL, Robinson JA, Grant BF, Stein MB. 2011. The effect of avoidant personality disorder on the persistence of generalized social anxiety disorder in the general population: results from a longitudinal, nationally representative mental health survey. Depress Anxiety 28:250-255.
22. Bailey-Wilson JE, Brennan JS, Bull SB, Culverhouse R, Kim Y, et al. 2011. Regression and data mining methods for analyses of multiple rare variants in the Genetic Analysis Workshop 17 Mini-Exome Data. Genet Epidemiol 35 Suppl 1:S92-100.
23. Comer JS, Blanco C, Hasin DS, Liu SM, Grant BF, et al. 2011. Health-related quality of life across the anxiety disorders: results from the National Epidemiologic Survey on Alcohol and Related Conditions. J Clin Psychiatry 72:43-50.
24. Baca-Garcia E, Perez-Rodriguez MM, Keyes KM, Oquendo MA, Hasin DS, et al. 2011. Suicidal ideation and suicide attempts among Hispanic subgroups in the United States: 1991-1992 and 2001-2002. J Psychiatr Res 45:512-518.
25. Lopez-Quintero C, Cobos JP, Hasin DS, Okuda M, Wang S, et al. 2011. Probability and predictors of transition from first use to dependence on nicotine, alcohol, cannabis, and cocaine: results of the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). Drug Alcohol Depend 115:120-130.
26. Zhao J, Bradfield JP, Li M, Zhang H, Mentch FD, et al. 2011. BMD-associated variation at the Osterix locus is correlated with childhood obesity in females. Obesity 19:1311-1314.
27. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Medical comorbidity of full

221
and partial posttraumatic stress disorder in United States adults: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Psychosom Med 73:697-707.
28. Jung J, Basile DP, Pratt JH. 2011. Sodium reabsorption in the thick ascending limb in relation to blood pressure: a clinical perspective. Hypertension 57:873-879.
29. Grant BF, Dawson DA, Moss HB. 2011. Disaggregating the burden of substance dependence in the United States. Alcohol Clin Exp Res 35:387-388.
30. Tu W, Eckert GJ, Dimeglio LA, Yu Z, Jung J, Pratt JH. 2011. Intensified effect of adiposity on blood pressure in overweight and obese children. Hypertension 58:818-824.
31. International Multiple Sclerosis Genetics Consortium; Wellcome Trust Case Control Consortium, Sawcer S, Hellenthal G, Pirinen M, et al. 2011. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 476:214-219.
32. Zanarini MC, Horwood J, Wolke D, Waylen A, Fitzmaurice G, Grant BF. 2011. Prevalence of DSM-IV borderline personality disorder in two community samples: 6,330 English 11-year-olds and 34,653 American adults. J Pers Disord 25:607-619.
33. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Prevalence and Axis I comorbidity of full and partial posttraumatic stress disorder in the United States: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. J Anxiety Disord 25:456-465.
34. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Personality disorders associated with full and partial posttraumatic stress disorder in the U.S. population: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. J Psychiatr Res 45:678-686.
35. Benson MD, Teague SD, Kovacs R, Feigenbaum H, Jung J, Kincaid JC. 2011. Rate of progression of transthyretin amyloidosis. Am J Cardiol 108:285-289.
36. Eaton NR, Krueger RF, Keyes KM, Skodol AE, Markon KE, et al. 2011. Borderline personality disorder co-morbidity: relationship to the internalizing-externalizing structure of common mental disorders. Psychol Med 41:1041-1050.
37. Keyes KM, Krueger RF, Grant BF, Hasin DS. 2011. Alcohol craving and the dimensionality of alcohol disorders. Psychol Med 41:629-640.
38. Lanktree MB, Guo Y, Murtaza M, Glessner JT, Bailey SD, et al. 2011. Meta-analysis of dense denecentric association studies reveals common and uncommon variants associated with height. Am J Hum Genet 88:6-18.
39. Elia J, Glessner JT, Wang K, Takahashi N, Shtir CJ, et al. 2011. Genome-wide copy number variation study associates metabotropic glutamate receptor gene networks with attention deficit hyperactivity disorder. Nat Genet 44:78-84.
40. Dawson DA, Goldstein RB, Grant BF. In press. Prospective correlates of drinking cessation: variation across the life course. Addiction.
41. Rubinsky AD, Dawson DA, Kivlahan DR, Williams EC, Etzioni R, Bradley KA. In press. AUDIT-C scores as a scaled marker of mean daily drinking, alcohol use disorder severity, and probability of alcohol dependence in a U.S. general population sample of drinkers. Alcohol Clin Exp Res.

222
42. Compton WM, Dawson DA, Conway KP, Brodsky M, Grant BF. In press. Transitions in drug use status over 3 years: a prospective analysis of a general population sample. Am J Psychiatry.
43. Lim SS, Vos T, Flaxman AD, Danaei G, Shibuya K, et al. 2012. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380:2224–2260.
44. Murray CJL, Vos T, Lozano R, Naghavi M, Flaxman AD, et al. 2012. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380:2197–2223.
45. Horikoshi M, Yaghootkar H, Mook-Kanamori DO, Sovio U, Taal HR, et al. 2012. New loci associated with birth weight identify genetic links between intrauterine growth and adult height and metabolism. Nat Genet Dec 2. [Epub ahead of print]
46. Dawson DA, Smith SM, Saha TD, Rubinsky AD, Grant BF. 2012. Comparative performance of the AUDIT-C in screening for DSM-IV and DSM-5 alcohol use disorders, 2012. Drug Alcohol Depend 126:384-388.
47. Goldstein RB, Dawson DA, Chou SP, Grant BF. 2012. Sex differences in prevalence and comorbidity of alcohol and drug use disorders: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. J Stud Alcohol Drugs 73:938-950.
48. Khan SS, Secades-Villa R, Okuda M, Wang S, Pérez-Fuentes G, et al. 2012. Gender differences in cannabis use disorders. Drug Alcohol Depend Nov 22. [Epub ahead of print]
49. Keyes KM, Eaton NR, Krueger RF, Skodol AE, Wall MM, et al. 2012. Thought disorder in the meta-structure of psychopathology. Psychol Med Nov 21. [Epub ahead of print]
50. Jung J, Barrett PQ, Eckert GJ, Edenberg HJ, Xuei X,et al. 2012. Association of the potassium channel gene, KCNK9, with blood pressure and aldosterone production. J Clin Endocrinol Metab 97:E2160-2167.
51. Kerridge BT, Saha TD, Gmel G, Rehm JT. 2012. Taxometic analysis of DSM-IV and DSM-5 alcohol use disorders. Drug Alcohol Depend Oct 31. [Epub ahead of print]
52. Eaton NR, Keyes KM, Krueger RF, Noordhof A, Skodol AE, et al. 2012. Ethnicity and psychiatric comorbidity in a national sample: evidence for latent liability spectrum invariance and connections with disorder prevalence. Soc Psychiatry Psychiatr Epidemiol Sep 30. [Epub ahead of print]
53. Dawson DA, Goldstein RB, Grant BF. 2012. Differences in the profiles of DSM-IV and DSM-5 alcohol use disorders: implications for clinicians. Alcohol Clin Exp Res Sep 13. [Epub ahead of print]
54. Eaton NR, Krueger RF, Markon KE, Keyes KM, Skodol AE, et al. 2012. The structure and predictive validity of the internalizing disorders. J Abnorm Psychol Aug 20. [Epub ahead of print]
55. Fenton MC, Geier T, Keyes K, Skodol A, Grant BF, Hasin D. 2012. Combined role of

223
childhood maltreatment, family history and gender in the risk for adult alcohol dependence in the United States. Psychol Med Aug 10. [Epub ahead of print]
56. Cox BJ, Clara IP, Worobec LM, Grant BF. 2012. An empirical evaluation of the structure of DSM-IV personality disorders in a nationally representative sample: results of confirmatory factor Analysis in the National Epidemiologic Survey on Alcohol and Related Conditions Waves 1 and 2. J Pers Disord Aug 1. [Epub ahead of print]
57. Sugaya L, Hasin DS, Olfson M, Lin KH, Grant BF, Blanco C. 2012. Child physical abuse and adult mental health: a national study. J Trauma Stress 25:384-392.
58. Dawson DA, Goldstein RB, Grant BF. 2012. Factors associated with first utilization of different types of care for alcohol problems. J Stud Alcohol Drugs 73:647-656.
59. Cherpitel CJ, Ye Y, Bond J, Borges G, Chou P, et al. 2012. Multi-level analysis of alcohol-related injury and drinking pattern: emergency department data from 19 countries. Addiction 107:1263-1272.
60. Moreno C, Hasin DS, Arango C, Oquendo MA, Vieta E, et al. 2012. The bipolar-depressive continuum in the National Epidemiologic Survey on Alcohol and Related Conditions. Bipolar Disord Dec 12. [Epub ahead of print]
61. Moreno C, Hasin DS, Arango C, Oquendo MA, Vieta E, et al. 2012. Depression in bipolar versus major depressive disorder: results from the National Epidemiologic Survey on Alcohol and Related Conditions. Bipolar Disord 14:271-282
62. Haas DM, Lehmann AS, Skaar T, Philips S, McCormick CL, et al. 2012. The impact of drug metabolizing enzyme polymorphisms on outcomes after antenatal corticosteroid use. Am J Obstet Gynecol 206:447.e17-e24.
63. Latorre V, Diskin SJ, Diamond MA, Zhang H, Hakonarson H, et al. 2012. Replication of neuroblastoma SNP association at the BARD1 locus in African-Americans. Cancer Epidemiol Biomarkers Prev 21:658-663.
64. Snell LD, Ramchandani VA, Saba L, Herion D, Heilig M, et al. 2012. WHO/ISBRA Study on State and Trait Markers of Alcohol Use and Dependence Investigators. The biometric measurement of alcohol consumption. Alcohol Clin Exp Res 36:332-41.
65. Chou SP, Chun S, Smith S, Ruan J, Li T-K, Grant BF. 2012. Episodic heavy drinking, problem drinking and injuries – results of the WHO/NIAAA Collaborative Emergency Room Study in South Korea. Alcohol 46:407-413
66. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2012. Psychiatric comorbidity of full and partial posttraumatic stress disorder among older adults in the United States: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Am J Geriatr Psychiatry 20:380-390.
67. Saha TD, Harford T, Goldstein RB, Kerridge BT, Hasin D. 2012. Relationship of substance abuse to dependence in the U.S. general population. J Stud Alcohol Drugs 73:368-378.
68. Saha TD, Compton WM, Chou SP, Smith S, Ruan WJ, et al. 2012. Analyses related to the development of DSM-5 criteria for substance use related disorders 1. Toward amphetamine, cocaine and prescription drug use disorder continua using item response theory. Drug Alcohol Depend 122:38-46.

224
69. Chou SP, Lee HK, Cho MJ, Park JI, Dawson DA, Grant BF. 2012. Alcohol use disorders, nicotine dependence, and co-occurring mood and anxiety disorders in the United States and South Korea – a cross-national comparison. Alcohol Clin Exp Res 36:654-662.
70. Harford TC, Chen CM, Saha TD, Smith SM, Hasin DS, Grant BF. 2012. An item response theory analysis of DSM-IV diagnostic criteria for personality disorders: findings from the National Epidemiologic Survey on Alcohol and Related Conditions. Personal Disord Mar 26. [Epub ahead of print]
71. Lopes M, Olfson M, Rabkin J, Hasin DS, Alegría AA, et al. 2012. Gender, HIV status, and psychiatric disorders: results from the National Epidemiologic Survey on Alcohol and Related Conditions. J Clin Psychiatry 73:384-391.
72. Goldstein RB, Dawson DA, Smith SM, Grant BF. 2012. Antisocial behavioral syndromes and three-year quality of life outcomes in United States adults. Acta Psychiatr Scand 126(2):137-50.
73. Blanco C, Vesga-López O, Stewart JW, Liu SM, Grant BF, Hasin DS. 2012. Epidemiology of major depression with atypical features: results from the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). J Clin Psychiatry 73:224-232.
74. Dawson DA, Goldstein RB, Ruan WJ, Grant BF. 2012. Correlates of recovery from alcohol dependence: a prospective study over a 3-year follow-up interval. Alcohol Clin Exp Res 36:1268-1277.
75. Olfson M, Liu SM, Grant BF, Blanco C. 2012. Influence of comorbid mental disorders on time to seeking treatment for major depressive disorder. Med Care 50:227-232.
76. Keyes KM, Eaton NR, Krueger RF, McLaughlin KA, Wall MM, et al. 2012. Child maltreatment and the structure of common psychiatric disorders. Br J Psychiatry 200:107-115.
77. Fenton MC, Keyes K, Geier T, Greenstein E, Skodol A, et al. 2012. Psychiatric comorbidity and the persistence of drug use disorders in the United States. Addiction 107:599-609.
78. Eaton NR, Keyes KM, Krueger RF, Balsis S, Skodol AE, et al. 2012. An invariant dimensional liability model of gender differences in mental disorder prevalence: evidence from a national sample. J Abnorm Psychol 121:282-288.
79. Dawson DA, Smith SM, Pickering RP, Grant BF. 2012. An empirical approach to evaluating the validity of alternative low-risk drinking guidelines. Drug Alcohol Rev 31:141-150.
80. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2012. Physical health conditions associated with posttraumatic stress disorder in U.S. older adults: results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. J Am Geriatric Soc 60:296-303.

225
Collaborators Christina Aivadyan, PhD, Columbia University, New York, NY. Michelle Amsbary, PhD, Westat, Rockville, MD. Marc Auriacombe, MD, Univ de Bordeaux, Bordeaux Cedex, France. Enrique Baca-Garcia, MD, Univ Complutense de Madrid, Madrid, Spain. Martha Beslin, PhD, Westat, Rockville, MD. Carlos Blanco, MD, PhD, Columbia Univ and NY State Psychiat Inst, New York, NY. Guilherme Borges, MD, DSc, Inst Nac de Psiquiatria, Univ Autonoma Metropolitana -
Xochimilco, Mexico. Andrew Brooks, PhD, Rutgers University, Piscataway, NJ. Kathleen Bucholz, PhD, Midwest Alc Res Ctr, Washington Univ, St. Louis, MO. Alan Budney, PhD, Univ of Arkansas, Little Rock, AR. Cheryl Cherpital, PhD, Alcohol Research Group, Emeryville, CA. Sung Soo Chun, PhD, MPH, Korean Inst of Alc Probl, Sahmyook Univ, Seoul, Korea. Wilson Compton, MD, MPE, NIDA, NIH, Bethesda, MD. Kevin Conway, PhD, NIDA, NIH, Bethesda, MD. Brian Cox, MD, Univ of Manitoba, Winnipeg, Canada. Nicholas Eaton, PhD, State University of New York, Stony Brook, NY. Stephanie Fram, PhD, Rutgers University, Piscataway, NJ. Gerhard Gmel, PhD, Swiss Institute of Prevention, Lausanne, Switzerland. Wei Hao, PhD, Central South University, Changsha, China. Thomas Harford, PhD, Boston Univ, Boston, MA and CSR, Inc, Arlington, VA. Deborah S. Hasin, PhD, Columbia Univ and NY State Psychiat Inst, New York, NY. Susumu Higuchi, LM.D., Hakodate Municipal General Hospital, Japan. Ronald Hirshhorn, PhD, Westat, Rockville, MD. Ming-Chyi Huang, Taipei City Psych Center, Taipei, Taiwan. Kenneth Kaplan, BS, KDK Consulting Co, Frederick, MD. Katherine Keys, MPhil, NY State Psychiatric Inst, New York, NY. Robert Krueger, PhD, University of Minnesota, Minneapolis, MN. Hae-Kook Lee, MD, PhD, Catholic Medical University of Korea, Seoul, Korea. Xu Lin, PhD, Inst for Nutr Biol Sci, Chinese Acad Sci, Shanghai, China. Walter Ling, MD, University of California, Los Angeles, CA. Sang-Min Liu, MS, NY State Psychiatric Inst, New York, NY. Xinhua Liu, PhD, Columbia University, New York, NY. Catalina Lopez-Quintero, MD, Hebrew University, Jerusalem, Israel. David Maklan, PhD, Westat, Rockville, MD. Jose Moreno, MD, Univ Complutense de Madrid, Madrid, Spain. Man Nguyen, MD, Department of Health, Saigon, Vietnam. Charles O’Brien, MD, PhD, University of Pennsylvania, Philadelphia, PA. Mayumi Okuda, MD, Department of Psychiatry, Columbia University, New York, NY. Mark Olfson, MD, Columbia University, New York, NY. Maria Oquendo, MD, Columbia University, New York, NY. Jong-Ik Park, MD, MD, PhD, Kangwon National University, Seoul, Korea. Jung Yeon Park, MD, New York State Psychiatric Institute, New York, NY. Mercedes Perez-Rodriquez, MD, University Autonoma de Madrid, Madrid, Spain. Nancy Petry, PhD, University of Connecticut, Farmington, CT. Darrell Regier, MD, MPH, Am Psychiatric Assn, Arlington, VA. Juergen Rehm, PhD, University of Toronto, Toronto, Canada.

226
Robin Room, PhD, Turning Point Alc and Drug Ctr, Melbourne, Australia. Jitendar Sareen, MD, University of Manitoba, Winnipeg, Canada. Marc Schuckit, MD, UCSD, San Diego, CA. Dvora Shmulewitz, PhD, Columbia University, New York, NY. Andrew Skodol, MD, Columbia University, New York, NY. Thaksaphon Thamarangsi, M.D., Ph.D., Lampang Prinvicial Gen. Hosp., Lampang, Thailand. Jay Tischfield, PhD, Rutgers University, Piscataway, NJ. Oriana Vesga-Lopez, BA, NY State Psychiatric Inst, New York, NY. Shuai Wang, PhD, Columbia University, New York, NY. Steve Williamson, PhD, Westat, Rockville, MD. Chuanjun Zhou, MD Anning Hospital of Tianjin City, Tianjin City, China. Dr. Grant: [email protected]

227
______________________________________________________________________
Laboratory of Behavioral and Genomic Neuroscience
______________________________________________________________________ Andrew Holmes, PhD, Chief Ozge Gunduz-Cinar, PhD, Postdoctoral IRTA Maxine Reger, PhD, Postdoctoral IRTA Nicolas Jury, PhD, Postdoctoral IRTA Charles Pickens, PhD, Research Fellow Anne Radke, PhD, Postdoctoral IRTA Courtney Pinard, PhD, Postdoctoral IRTA Erica Busch, BSc, Predoctoral IRTA Mission statement The mission of the Laboratory of Behavioral and Genomic Neuroscience is to contribute to a deeper understanding of the causes of alcoholism and comorbid neuropsychiatric conditions such as mood and anxiety disorders. Our goal is to help identify new directions for the prevention and effective treatment of these devastating diseases. To this end, we are using models of chronic alcohol exposure and chronic stress to examine how these environmental insults reshape brain circuits to modify behavior, and why they do so in a manner that varies greatly from individual to individual as a function of genetics, sex and age. A major current focus of our work is how alcohol and stress affect the structure and function of circuits interconnecting the prefrontal cortex with limbic and dorsal striatal regions that are critical for the regulation of emotion, cognition and executive control over drug-seeking. Research projects Convergent technologies to study alcoholism Over the past decade, rapid advances in the application of molecular genetics techniques to mammalian model systems has led to an explosion in the use of mice in behavioral neuroscience. Complex genetic populations, targeted gene mutation and optogenetics are just some of the technological advances that allow us to precisely manipulate specific brain circuits and molecules to the pathophysiology of neuropsychiatric disorders. To fully utilize these tools, we are also actively developing equally sophisticated models of alcoholism and new methods to assess mouse behavior. To give one example, the mouse ‘touchscreen’ operant system has been developed in close collaboration with our colleagues in Cambridge
Figure 1: A mouse eye view of the ‘touchscreen’ system for measuring executive function. The system displays computer-generated visual stimuli on an LCD monitor screen, and allows the mouse to select stimuli by touching the screen with its nose.
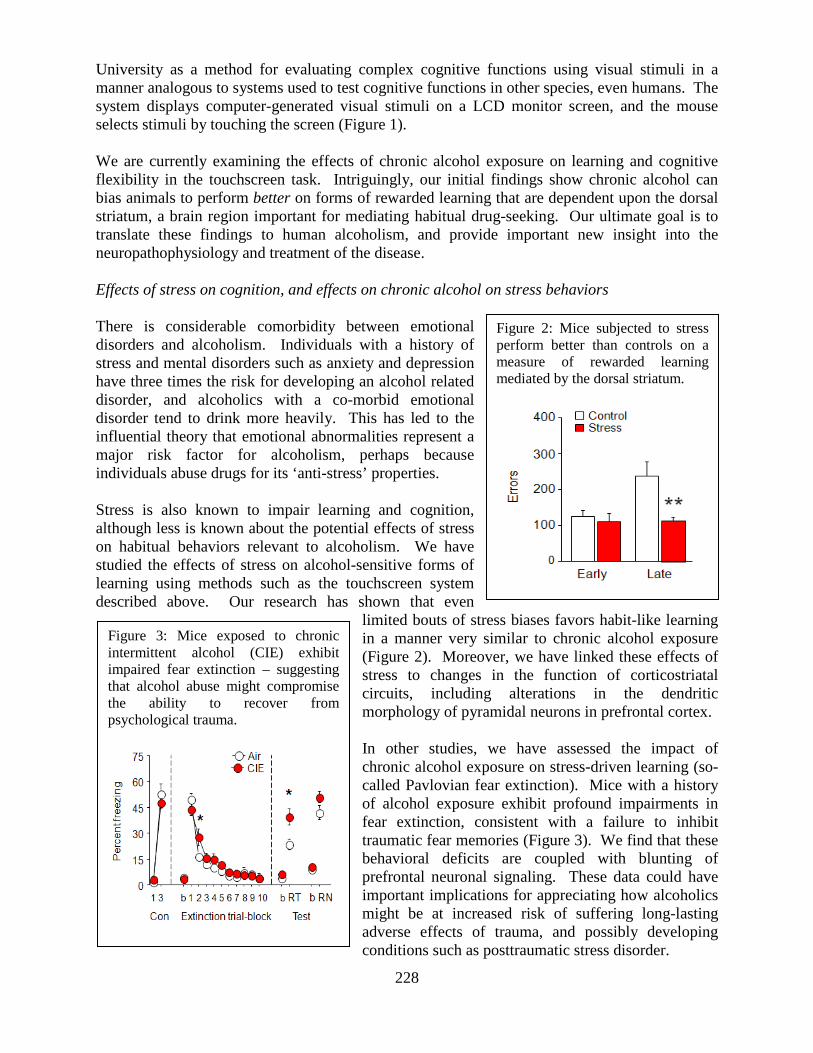
228
University as a method for evaluating complex cognitive functions using visual stimuli in a manner analogous to systems used to test cognitive functions in other species, even humans. The system displays computer-generated visual stimuli on a LCD monitor screen, and the mouse selects stimuli by touching the screen (Figure 1). We are currently examining the effects of chronic alcohol exposure on learning and cognitive flexibility in the touchscreen task. Intriguingly, our initial findings show chronic alcohol can bias animals to perform better on forms of rewarded learning that are dependent upon the dorsal striatum, a brain region important for mediating habitual drug-seeking. Our ultimate goal is to translate these findings to human alcoholism, and provide important new insight into the neuropathophysiology and treatment of the disease. Effects of stress on cognition, and effects on chronic alcohol on stress behaviors There is considerable comorbidity between emotional disorders and alcoholism. Individuals with a history of stress and mental disorders such as anxiety and depression have three times the risk for developing an alcohol related disorder, and alcoholics with a co-morbid emotional disorder tend to drink more heavily. This has led to the influential theory that emotional abnormalities represent a major risk factor for alcoholism, perhaps because individuals abuse drugs for its ‘anti-stress’ properties.
Figure 2: Mice subjected to stress perform better than controls on a measure of rewarded learning mediated by the dorsal striatum.
Stress is also known to impair learning and cognition, although less is known about the potential effects of stress on habitual behaviors relevant to alcoholism. We have studied the effects of stress on alcohol-sensitive forms of learning using methods such as the touchscreen system described above. Our research has shown that even
limited bouts of stress biases favors habit-like learning in a manner very similar to chronic alcohol exposure (Figure 2). Moreover, we have linked these effects of stress to changes in the function of corticostriatal circuits, including alterations in the dendritic morphology of pyramidal neurons in prefrontal cortex. In other studies, we have assessed the impact of chronic alcohol exposure on stress-driven learning (so-called Pavlovian fear extinction). Mice with a history of alcohol exposure exhibit profound impairments in fear extinction, consistent with a failure to inhibit traumatic fear memories (Figure 3). We find that these behavioral deficits are coupled with blunting of prefrontal neuronal signaling. These data could have important implications for appreciating how alcoholics might be at increased risk of suffering long-lasting adverse effects of trauma, and possibly developing conditions such as posttraumatic stress disorder.
Figure 3: Mice exposed to chronic intermittent alcohol (CIE) exhibit impaired fear extinction – suggesting that alcohol abuse might compromise the ability to recover from psychological trauma.

229
l to r: A Radke, C Pickens, O Gunduz-Cinar, N Jury, S Flynn, E Sagalyn, O Bukalo, C Pinard, E Busch Publications 2011-2012 1. Lederle L, Weber S, Wright T, Feyder M, Brigman JL, Crombag H, Saksida LM, Bussey TJ,
Holmes A. 2011. Reward-related behavioral paradigms for addiction research in the mouse: performance of common inbred strains. PLoS One 6: e15536
2. Camp M, Feyder M, Ihne J, Palachick B, Hurd B, et al 2011. A novel role for PSD-95 in
mediating ethanol intoxication, drinking and place preference. Addiction Biology 16:428-439 3. Kiselycznyk C, Svenningson P, Delpire E, Holmes A. 2011. Genetic, pharmacological and
lesion analyses reveal a selective role for corticohippocampal GluN2B in a novel repeated swim stress paradigm. Neuroscience 193:259-268
4. Kiselycznyk C, Holmes A. 2011. All (C57BL/6) mice are not created equal. Frontiers in
Neuroscience 5:10 5. Graybeal C, Schulman E, Saksida LM, Bussey TJ, Feyder M, et al. 2011. Paradoxical
reversal learning enhancement by stress or prefrontal cortical damage: rescue with BDNF. Nature Neuroscience 14:1507-1509
6. Ihne J, Fitzgerald PJ, Hefner K, Holmes A. 2012. Pharmacological manipulation of stress-
induced C57BL/6J anxiety-related behavior. Neuropharmacology 62:464-473 7. Barkus C, Bannerman DM, Feyder M, Graybeal G, Wright T, et al. 2012. GluA1 knockout
mice exhibit behavioral abnormalities relevant to the negative symptoms of schizophrenia and schizoaffective disorder. Neuropharmacology 62:1263-1272

230
8. Kiselycznyk C, Hoffman D, Holmes A (2012) Effects of genetic deletion of the Kv4.2
voltage-gated potassium channel on murine anxiety-, fear- and stress-related behaviors. Biology of Mood and Anxiety Disorders 2:5
9. Camp M, Martin K, Graybeal C, Lederle L, Gaburro S, et al. 2012. Genetic strain differences
in learned fear inhibition associate with variation in neuroendocrine, autonomic and amygdala dendritic phenotype. Neuropsychopharmacology 37:1534-1547
10. Hartley CA, McKenna MC, Salman R, Holmes A, Casey BJ, et al. 2012. A human serotonin
transporter polyadenylation polymorphism selectively modulates the retention of fear extinction memory. Proceedings of the National Academy of Sciences, USA 109:5493-5498
11. Maroun M, Birger O, Holmes A, Wellman C, Montanis H. 2012. Enhanced extinction of
aversive memories by high-frequency stimulation of the rat infralimbic cortex. PLoS One 7:e35853
12. Chesler EJ, Plitt A, Fisher D, Hurd B, Lederle L, Kiselycznyk C, Holmes A. 2012.
Quantitative trait loci for sensitivity to ethanol intoxication in a C57BL/6J x 129S1/SvImJ inbred mouse cross. Mammalian Genome 23:305-321
13. Karlsson RM, Adermark L, Molander A, Pereau-Lenz S, Singley E, et al. 2012. Reduced
alcohol intake, lack of alcohol reward and impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology 63:181-189
14. Whittle N Gunduz-Cinar O, Holmes A Singewald N. 2012. Deep brain stimulation and
adjunctive drug treatments rescue impaired fear extinction in a genetic mouse model. Neuropharmacology 64:414-423
15. Holmes A, Fitzgerald PJ, MacPherson K, DeBrouse L, Colacicco G, et al. 2012. Chronic
alcohol remodels prefrontal neurons and disrupts NMDA receptor-mediated fear extinction encoding. Nature Neuroscience 15:1359-1361
16. Bussey TJ, Holmes A, Lyon L, Mar A, McAllister K, et al. 2012. New translational assays
for preclinical modelling of cognition in schizophrenia: the touchscreen testing method for mice and rats. Neuropharmacology 62:1191-1203
17. Graybeal C, Kiselycznyk C, Holmes A. 2012. Stress-induced impairments in prefrontal-
mediated behaviors and the role of the N-methyl-D-aspartate receptor. Neuroscience 211:28-38
18. Holmes A. 2011. Merger fever: can two mechanisms work together to explain alcoholism?
Biological Psychiatry 69:1043-1051
Dr.Holmes: [email protected]

231
______________________________________________________________________
Laboratory of Molecular Signaling ______________________________________________________________________ Hee-Yong Kim, PhD, Chief Mohammed Akbar, PhD, Staff Scientist Xudong Huang, PhD, Senior Chemist Karl Kevala, MS, Chemist Atsuko Kimura, PhD, Visiting Fellow Vishaldeep Sidhu, PhD, Visiting Fellow Ji-Won Lee, PhD, Visiting Fellow Mohammad Rashid, PhD, Visiting Fellow Abhishek Desai, PhD, Visiting Fellow Giorgi Kharebava, PhD, HJF Fellow Kenneth Newcomer, BS, Postbac IRTA Jaquel Barnes, BS, Postbac IRTA Arthur Spector, MD, Adj Investigator Rachel Quade, Administrative Assistant Neurotrophic Effects of Polyunsaturated Fatty Acids The principal objective of the research in Laboratory of Molecular Signaling is to elucidate biological and metabolic functions of polyunsaturated fatty acids, especially docosahexaenoic acid (DHA, 22:6n-3) in the nervous system with particular reference to their modulation by ethanol. We investigate biotransformation of polyunsaturates to bioactive mediators for neurodevelopment and function. We also investigate biochemical mechanisms by which polyunsaturated lipids and ethanol modify neuronal cell membranes, and characterize membrane-related signaling mechanisms involved in neuroprotection. To this end, we develop multidisciplinary approaches using molecular, cellular and modern instrumental techniques such as mass spectrometry and biomolecular interaction analysis. Current research efforts are: 1) to elucidate cellular and molecular mechanisms of DHA-dependent neurodevelopment and neuroprotection; 2) to probe effects of ethanol on DHA-dependent neuroprotective signaling processes; 3) to discover inhibitors/modulators targeting molecular mechanisms underlying DHA-dependent neuroprotection; 4) to characterize DHA- and ethanol-dependent neuronal membrane modification; 5) to identify DHA- and ethanol-dependent modulation of synaptic membrane proteome; 6) to develop strategies to probe protein-protein and protein-membrane interactions, particularly conformational changes of integral and/or membrane-interacting proteins; 7) to establish a strategy to discover novel bioactive metabolites; and 8) to evaluate in vivo neuroprotective effects of DHA, bioactive DHA metabolites and their analogues. DHA is the major polyunsaturated fatty acid that accumulates in the brain during development, which has been implicated in cognitive development and memory-related learning ability. We found that DHA increases neural phosphatidylserine (PS) and prevents apoptotic neuronal cell death in a PS- and PI3 kinase-dependent manner (Kim et al., 2000; Akbar et. al., 2005). We have also established that DHA (Calderon and Kim, 2004; Cao et al., 2009) and its metabolite docosahexaenoylethanolamide promotes neurite outgrowth and synaptogenesis in hippocampal neurons, and improves synaptic function (Kim et al., 2011). N-3 fatty acid deficiency or long
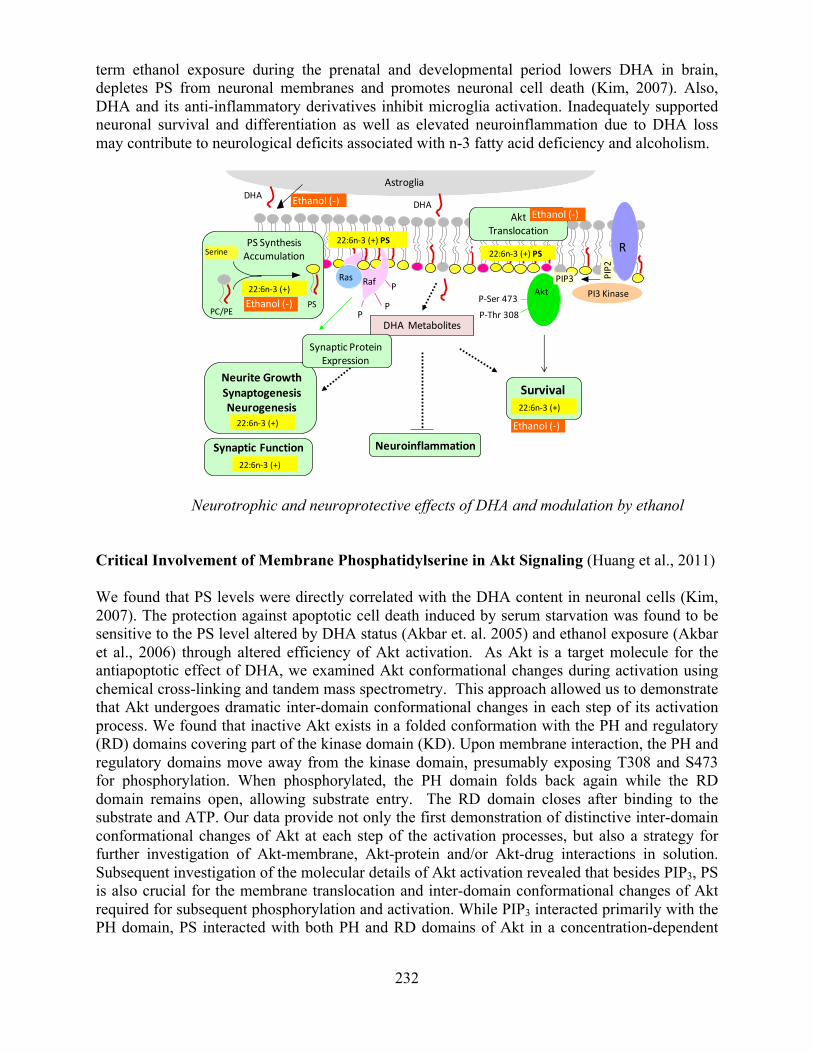
232
term ethanol exposure during the prenatal and developmental period lowers DHA in brain, depletes PS from neuronal membranes and promotes neuronal cell death (Kim, 2007). Also, DHA and its anti-inflammatory derivatives inhibit microglia activation. Inadequately supported neuronal survival and differentiation as well as elevated neuroinflammation due to DHA loss may contribute to neurological deficits associated with n-3 fatty acid deficiency and alcoholism.
Neurotrophic and neuroprotective effects of DHA and modulation by ethanol
Ras Raf P AktPIP3
P-‐Ser 473
P-‐Thr 308
22:6n-‐3 (+) PS R
PI3 Kinase
PS Synthesis Accumulation
PC/PEPS
Serine
P
Astroglia
22:6n-‐3 (+) PS
Neurite GrowthSynaptogenesisNeurogenesis
22:6n-‐3 (+)
P
PIP2
Survival22:6n-‐3 (+)
Akt Translocation
22:6n-‐3 (+)
Synaptic Function22:6n-‐3 (+)
DHA Metabolites
Synaptic Protein Expression
DHADHA
Neuroinflammation
Ethanol (-‐)
Ethanol (-‐)
Ethanol (-‐)
Ethanol (-‐)
Critical Involvement of Membrane Phosphatidylserine in Akt Signaling (Huang et al., 2011) We found that PS levels were directly correlated with the DHA content in neuronal cells (Kim, 2007). The protection against apoptotic cell death induced by serum starvation was found to be sensitive to the PS level altered by DHA status (Akbar et. al. 2005) and ethanol exposure (Akbar et al., 2006) through altered efficiency of Akt activation. As Akt is a target molecule for the antiapoptotic effect of DHA, we examined Akt conformational changes during activation using chemical cross-linking and tandem mass spectrometry. This approach allowed us to demonstrate that Akt undergoes dramatic inter-domain conformational changes in each step of its activation process. We found that inactive Akt exists in a folded conformation with the PH and regulatory (RD) domains covering part of the kinase domain (KD). Upon membrane interaction, the PH and regulatory domains move away from the kinase domain, presumably exposing T308 and S473 for phosphorylation. When phosphorylated, the PH domain folds back again while the RD domain remains open, allowing substrate entry. The RD domain closes after binding to the substrate and ATP. Our data provide not only the first demonstration of distinctive inter-domain conformational changes of Akt at each step of the activation processes, but also a strategy for further investigation of Akt-membrane, Akt-protein and/or Akt-drug interactions in solution. Subsequent investigation of the molecular details of Akt activation revealed that besides PIP3, PS is also crucial for the membrane translocation and inter-domain conformational changes of Akt required for subsequent phosphorylation and activation. While PIP3 interacted primarily with the PH domain, PS interacted with both PH and RD domains of Akt in a concentration-dependent

233
manner, augmenting Akt translocation, phosphorylation and activation triggered by PIP3 in living cells.
b.
c.
d.
e.
f.
2400 2600 2800 3000 3200 3400 3600
2458 3465
K284-K426 (-)
K284-K426 (-)
K284-K426 (-)K30-K389 (-)
K30-K389 (+)
K30-K389 (+)
K30-K389 (+)
K30-K389 (+) K284-K426 (+)
K284-K426 (+)
Mass, Da
* ** ** * **
*
* *
**
*
K268 capping K111-K112
*
*
b. Crosslinked no liposomes
e. PIP3/PE/PC
d. PS/PE/PC
c. PE/PC
(30%/50%/20%)
(50%/50%)
a. Akt no crosslinking
Mass spectra Schematic depiction of conformational changes
PH RD
KD
T308S473
Membrane PS also influenced the downstream events of Akt activation such as the phosphorylation of Foxo1(T24), GSK-3β(S9) and BAD(S136), consequently promoting cell survival. We identified specific PS-interacting residues in the PH and RD domains. Mutation of these PS-interacting sites from the PH and RD domains prevented the Akt activation. We found that Akt interaction with R15 and K20 in the PH domain outside the PIP3 binding pocket is critical for Akt translocation and activation. Binding of PS to the RD domain, particularly at K419 and K420, significantly influenced Akt conformational changes for phosphorylation at S473 by mTORC2, augmenting Akt activation triggered by PIP3 in living cells.
Effects of membrane interaction on inter-domain conformational changes of Akt probed by mass spectrometry.
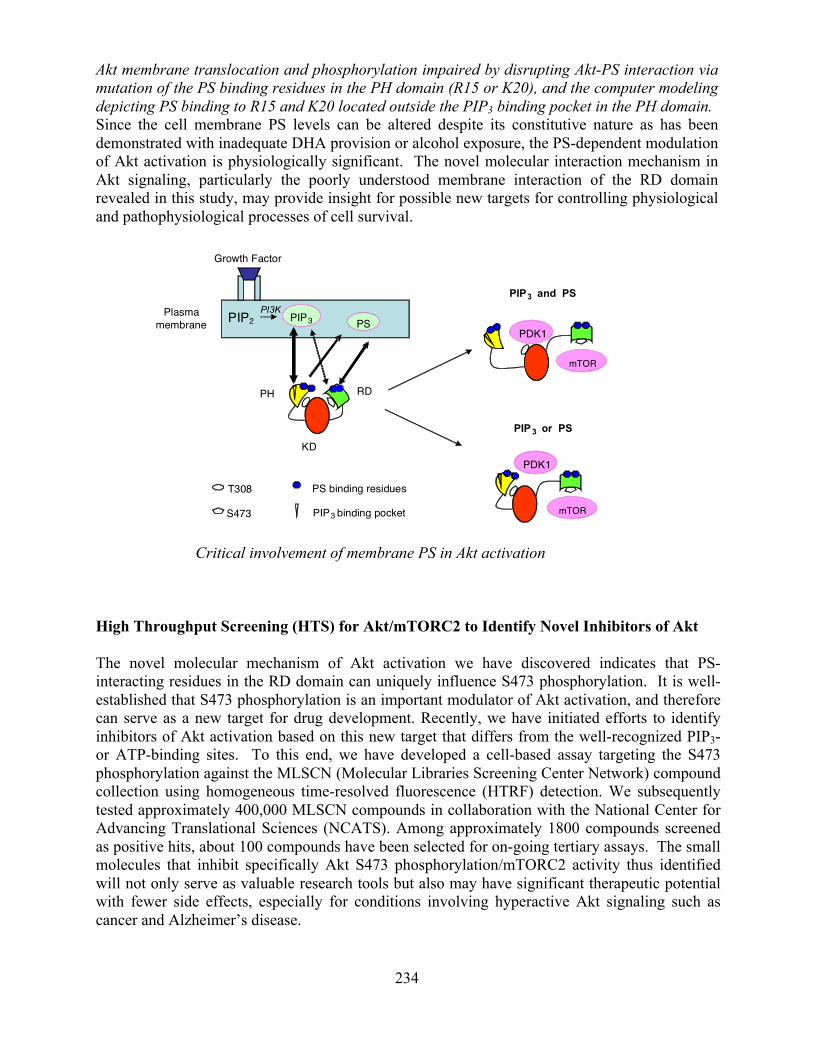
234
Akt membrane translocation and phosphorylation impaired by disrupting Akt-PS interaction via mutation of the PS binding residues in the PH domain (R15 or K20), and the computer modeling depicting PS binding to R15 and K20 located outside the PIP3 binding pocket in the PH domain. Since the cell membrane PS levels can be altered despite its constitutive nature as has been demonstrated with inadequate DHA provision or alcohol exposure, the PS-dependent modulation of Akt activation is physiologically significant. The novel molecular interaction mechanism in Akt signaling, particularly the poorly understood membrane interaction of the RD domain revealed in this study, may provide insight for possible new targets for controlling physiological and pathophysiological processes of cell survival.
Critical involvement of membrane PS in Akt activation
Growth Factor
PH RD
KD
S473 T308
PIP 2 PI3K Plasma
membrane
PS binding residues
PIP 3 PS
PIP 3 binding pocket
PDK1
mTOR
PIP 3 and PS
mTOR
PIP 3 or PS
PDK1
High Throughput Screening (HTS) for Akt/mTORC2 to Identify Novel Inhibitors of Akt The novel molecular mechanism of Akt activation we have discovered indicates that PS-interacting residues in the RD domain can uniquely influence S473 phosphorylation. It is well-established that S473 phosphorylation is an important modulator of Akt activation, and therefore can serve as a new target for drug development. Recently, we have initiated efforts to identify inhibitors of Akt activation based on this new target that differs from the well-recognized PIP3- or ATP-binding sites. To this end, we have developed a cell-based assay targeting the S473 phosphorylation against the MLSCN (Molecular Libraries Screening Center Network) compound collection using homogeneous time-resolved fluorescence (HTRF) detection. We subsequently tested approximately 400,000 MLSCN compounds in collaboration with the National Center for Advancing Translational Sciences (NCATS). Among approximately 1800 compounds screened as positive hits, about 100 compounds have been selected for on-going tertiary assays. The small molecules that inhibit specifically Akt S473 phosphorylation/mTORC2 activity thus identified will not only serve as valuable research tools but also may have significant therapeutic potential with fewer side effects, especially for conditions involving hyperactive Akt signaling such as cancer and Alzheimer’s disease.

235
Ethanol-induced Impairment of Akt Activation (Huang and Kim, 2012) Although PI3K/Akt signaling that regulates neuronal survival has been implicated in the deleterious effects of ethanol on the central nervous system, the underlying molecular mechanisms have not been fully elucidated. Our research indicates at least two mechanisms; 1) neuron specific mechanism where ethanol reduces PS levels in neuronal membranes, and thereby hampering Akt-PS interaction that is necessary for Akt activation (Wen and Kim, 2004); 2) ethanol inhibits Akt activation by direct interaction with Akt. Akt-membrane interaction is a prerequisite step for Akt activation since it induces inter-domain conformational changes to an open conformer that allows Akt phosphorylation by upstream kinases. By quantitatively probing Akt conformation using chemical cross-linking, 18O labeling and mass spectrometry, we found that ethanol at pharmacologically relevant concentrations (20 or 170 mM) directly interacts with Akt and alters the local PH domain configuration near the PIP3-binding site. We also found that ethanol significantly impairs subsequent membrane-induced inter-domain conformational changes to an open conformer needed for Akt activation. The observed alteration of Akt conformation caused by ethanol during the activation sequence provides a new molecular basis for the effects of ethanol on Akt signaling. The in vitro conformation-based approach employed in this study should also be useful in probing the molecular mechanisms for the action of ethanol or drugs on other signaling proteins, particularly for those undergoing dramatic conformational change during activation processes such as members of AGC kinase super family.
Anti-‐Akt-‐d2
Anti-‐pAkt (S473)
IGF
IGF-‐ReceptorDiagram of the HTRF pAktdetection assay. After stimulation with IGF, cells are lysed and incubated with anti-Akt MAb labeled with acceptor fluorophore d2 and anti-pS473-Akt MAb labeled with Europium cryptate. Two antibodies bind to Akt at different sites. AktpS473 is detected by the fluorescence resonance energy transfer (FRET) between two fluorophores.
Ethanol impairs membrane-induced inter-domain conformational changes to an open conformer required for Akt activation.
- EtOH
+ EtOH
16O-labeled membrane interacted Akt
Membrane interaction
EtOH
T308
S473
Open Conformer Folded Conformer
PH RD
KD
RD-KDPH-KD
3473
.7
3465
.7
3473
.7
3465
.7
2467
.5
2459
.5
2467
.5
2459
.5
18O-labeled inactive Akt
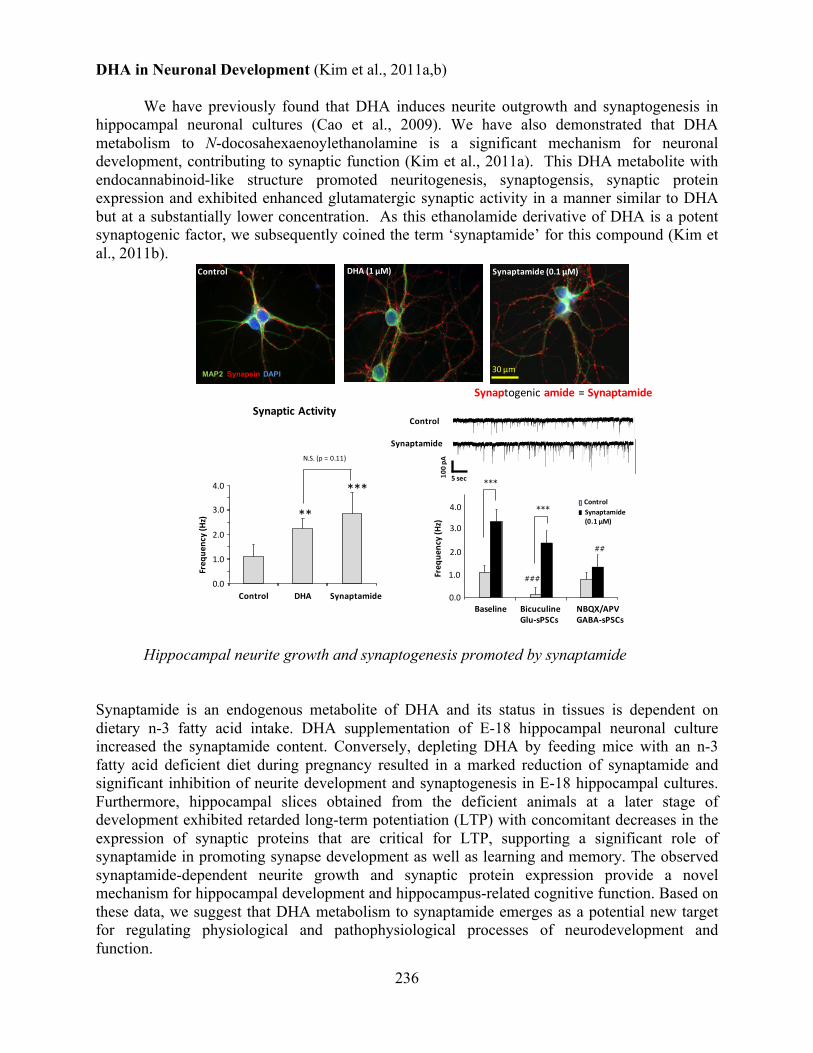
236
DHA in Neuronal Development (Kim et al., 2011a,b)
We have previously found that DHA induces neurite outgrowth and synaptogenesis in hippocampal neuronal cultures (Cao et al., 2009). We have also demonstrated that DHA metabolism to N-docosahexaenoylethanolamine is a significant mechanism for neuronal development, contributing to synaptic function (Kim et al., 2011a). This DHA metabolite with endocannabinoid-like structure promoted neuritogenesis, synaptogensis, synaptic protein expression and exhibited enhanced glutamatergic synaptic activity in a manner similar to DHA but at a substantially lower concentration. As this ethanolamide derivative of DHA is a potent synaptogenic factor, we subsequently coined the term ‘synaptamide’ for this compound (Kim et al., 2011b).
0.0
1.0
2.0
3.0
4.0
Baseline BicuculineGlu-‐sPSCs
NBQX/APVGABA-‐sPSCs
ControlSynaptamide (0.1 µM)
###
##
***
***
Freq
uency (Hz)
Control
Synaptamide10
0 pA
5 sec
Control DHA (1 µM) Synaptamide (0.1 µM)
0.0
1.0
2.0
3.0
4.0
Control DHA Synaptamide
N.S. (p = 0.11)
**
***
Freq
uency (Hz)
Synaptic Activity
MAP2/ Synapsin DAPI
Synaptogenic amide = Synaptamide
30 µm
Hippocampal neurite growth and synaptogenesis promoted by synaptamide
Synaptamide is an endogenous metabolite of DHA and its status in tissues is dependent on dietary n-3 fatty acid intake. DHA supplementation of E-18 hippocampal neuronal culture increased the synaptamide content. Conversely, depleting DHA by feeding mice with an n-3 fatty acid deficient diet during pregnancy resulted in a marked reduction of synaptamide and significant inhibition of neurite development and synaptogenesis in E-18 hippocampal cultures. Furthermore, hippocampal slices obtained from the deficient animals at a later stage of development exhibited retarded long-term potentiation (LTP) with concomitant decreases in the expression of synaptic proteins that are critical for LTP, supporting a significant role of synaptamide in promoting synapse development as well as learning and memory. The observed synaptamide-dependent neurite growth and synaptic protein expression provide a novel mechanism for hippocampal development and hippocampus-related cognitive function. Based on these data, we suggest that DHA metabolism to synaptamide emerges as a potential new target for regulating physiological and pathophysiological processes of neurodevelopment and function.
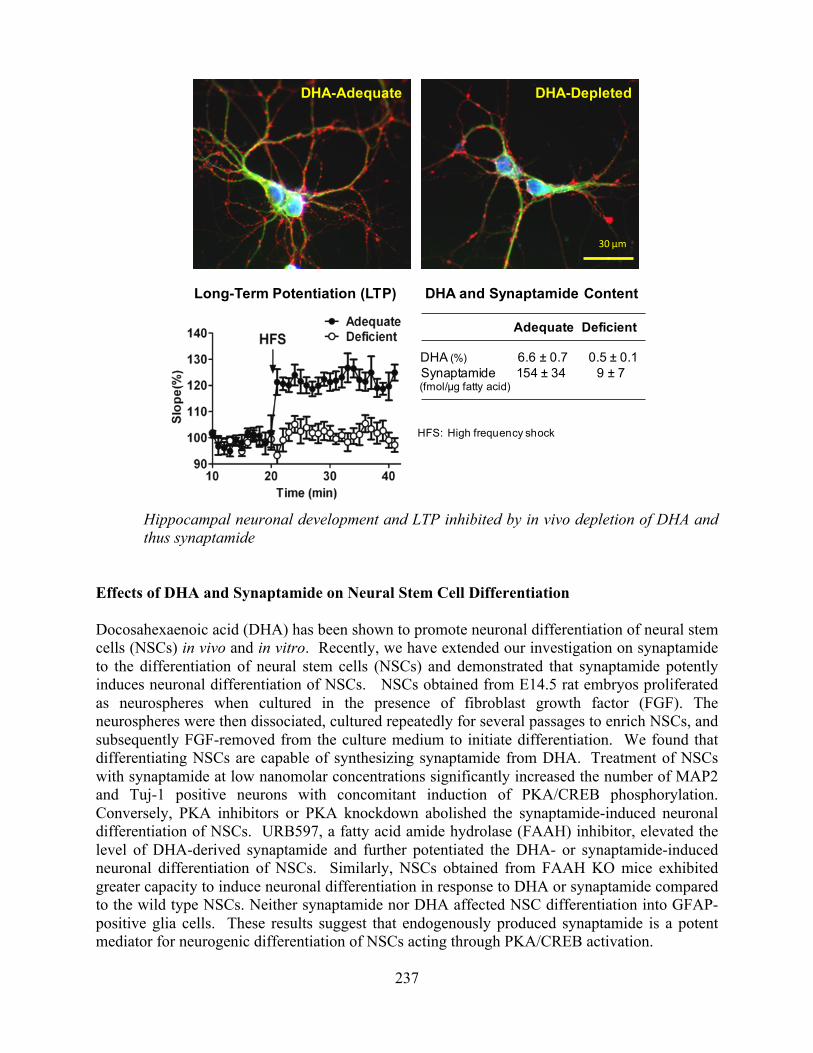
237
30 µm
DHA-DepletedDHA-Adequate
Long-Term Potentiation (LTP)
DHA (%) 6.6 ± 0.7 0.5 ± 0.1 Synaptamide 154 ± 34 9 ± 7(fmol/µg fatty acid)
Adequate Deficient
DHA and Synaptamide Content
HFS: High frequency shock
Hippocampal neuronal development and LTP inhibited by in vivo depletion of DHA and thus synaptamide
Effects of DHA and Synaptamide on Neural Stem Cell Differentiation Docosahexaenoic acid (DHA) has been shown to promote neuronal differentiation of neural stem cells (NSCs) in vivo and in vitro. Recently, we have extended our investigation on synaptamide to the differentiation of neural stem cells (NSCs) and demonstrated that synaptamide potently induces neuronal differentiation of NSCs. NSCs obtained from E14.5 rat embryos proliferated as neurospheres when cultured in the presence of fibroblast growth factor (FGF). The neurospheres were then dissociated, cultured repeatedly for several passages to enrich NSCs, and subsequently FGF-removed from the culture medium to initiate differentiation. We found that differentiating NSCs are capable of synthesizing synaptamide from DHA. Treatment of NSCs with synaptamide at low nanomolar concentrations significantly increased the number of MAP2 and Tuj-1 positive neurons with concomitant induction of PKA/CREB phosphorylation. Conversely, PKA inhibitors or PKA knockdown abolished the synaptamide-induced neuronal differentiation of NSCs. URB597, a fatty acid amide hydrolase (FAAH) inhibitor, elevated the level of DHA-derived synaptamide and further potentiated the DHA- or synaptamide-induced neuronal differentiation of NSCs. Similarly, NSCs obtained from FAAH KO mice exhibited greater capacity to induce neuronal differentiation in response to DHA or synaptamide compared to the wild type NSCs. Neither synaptamide nor DHA affected NSC differentiation into GFAP-positive glia cells. These results suggest that endogenously produced synaptamide is a potent mediator for neurogenic differentiation of NSCs acting through PKA/CREB activation.
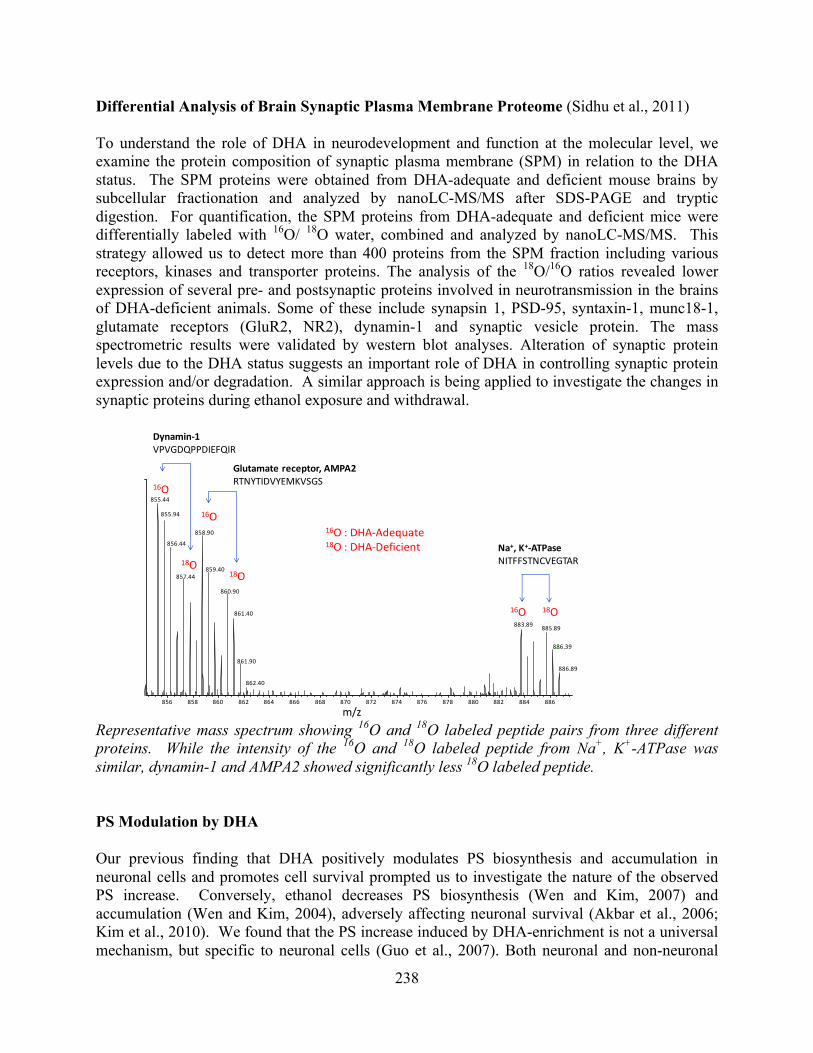
238
Differential Analysis of Brain Synaptic Plasma Membrane Proteome (Sidhu et al., 2011) To understand the role of DHA in neurodevelopment and function at the molecular level, we examine the protein composition of synaptic plasma membrane (SPM) in relation to the DHA status. The SPM proteins were obtained from DHA-adequate and deficient mouse brains by subcellular fractionation and analyzed by nanoLC-MS/MS after SDS-PAGE and tryptic digestion. For quantification, the SPM proteins from DHA-adequate and deficient mice were differentially labeled with 16O/ 18O water, combined and analyzed by nanoLC-MS/MS. This strategy allowed us to detect more than 400 proteins from the SPM fraction including various receptors, kinases and transporter proteins. The analysis of the 18O/16O ratios revealed lower expression of several pre- and postsynaptic proteins involved in neurotransmission in the brains of DHA-deficient animals. Some of these include synapsin 1, PSD-95, syntaxin-1, munc18-1, glutamate receptors (GluR2, NR2), dynamin-1 and synaptic vesicle protein. The mass spectrometric results were validated by western blot analyses. Alteration of synaptic protein levels due to the DHA status suggests an important role of DHA in controlling synaptic protein expression and/or degradation. A similar approach is being applied to investigate the changes in synaptic proteins during ethanol exposure and withdrawal.
Representative mass spectrum showing 16O and 18O labeled peptide pairs from three different proteins. While the intensity of the 16O and 18O labeled peptide from Na+, K+-ATPase was similar, dynamin-1 and AMPA2 showed significantly less 18O labeled peptide. PS Modulation by DHA Our previous finding that DHA positively modulates PS biosynthesis and accumulation in neuronal cells and promotes cell survival prompted us to investigate the nature of the observed PS increase. Conversely, ethanol decreases PS biosynthesis (Wen and Kim, 2007) and accumulation (Wen and Kim, 2004), adversely affecting neuronal survival (Akbar et al., 2006; Kim et al., 2010). We found that the PS increase induced by DHA-enrichment is not a universal mechanism, but specific to neuronal cells (Guo et al., 2007). Both neuronal and non-neuronal
Dynamin-‐1VPVGDQPPDIEFQIR
Glutamate receptor, AMPA2RTNYTIDVYEMKVSGS
Na+, K+-‐ATPaseNITFFSTNCVEGTAR
855.44
855.94
858.90
856.44
859.40857.44
860.90
861.40
883.89 885.89
886.39
861.90886.89
856 858 860 862 864 866 868 870 872 874 876 878 880 882 884 886
862.40
16O
18O
16O
18O
16O 18O
16O : DHA-‐Adequate18O : DHA-‐Deficient
m/z

239
cells used the 18:0, 22:6 molecular species as the best substrate for PS biosynthesis (Kim et al., 2004). However, non-neuronal cells compensate for the increase of DHA-PS species by a decrease in monoene-PS species, presumably due to product inhibition. DHA enrichment did not affect the mRNA levels of PSS1 and PSS2, proteins primarily responsible for PS synthesis in mammalian cells. Over-expression of genes encoding PSS1 or PSS2 altered neither the PS level nor the DHA-induced PS increase in both neuronal and non-neuronal cells. From these results it is concluded that the PS increase produced by DHA, specifically observed in neuronal cells, may represent a unique mechanism of expanding the PS pool in mammalian cells. To characterize the neuronal specific PS increase produced by DHA, the molecular structure and substrate specificity of PSS enzymes and their alteration by DHA and ethanol are investigated. We established a large scale expression system for PSS2, an integral protein, by expressing Flag-PSS2 in HEK cells in suspension culture. Subsequently, we successfully immunopurified functionally active PSS2 reconstituted in liposomes. We found that purified PSS2 also produced DHA-PS most favorably versus other fatty acid-containing PS species, as was observed previously with microsomes (Kimura and Kim, 2012). This specificity may contribute to the high accumulation of PS in neuronal cells where DHA is highly concentrated. Biochemical and structural details of PSS2 conferring this specificity are under investigation. Novel Metabolite Search using Stable Isotope Labeled Docosahexaenoic Acid and HPLC/MS/MS Identifying novel metabolites in complex biological samples is challenging and requires an integrated analytical strategy. Stable isotopes have provided a powerful tool for investigating metabolism as an alternative to radioisotope approach. The goal of our study is to propose an approach to identify novel metabolites derived from labeled substrates using LC/MS/MS and computer-assisted peak finding routine. As a model system, we first characterized neuronal metabolites formed from unlabeled DHA and uniformly labeled 13C-DHA. High mass accuracy data were generated using a Q-Exactive mass spectrometer operating in negative or positive ion mode with data-dependent scanning. Peak finding was achieved by comparing mass differences between labeled and non-labeled counterparts in consideration of possible biochemical transformation as well as by using other constraints such as the elemental composition, isotope mass profiles, and retention time. Using this approach we found the transformation of DHA to N-docosahexaenoylethanolamine (synaptamide) in primary neuronal cells in culture. The homogenate of fetal brains also actively converted 13C-DHA to 13C-synaptamide. Stable isotope-assisted assignment offered unambiguous identification of metabolites among number of potential candidates without the need for standard compounds. Based on the high mass accuracy and unique isotope profile of 13C-DHA-derived metabolites together with the predicted mass differences from the corresponding 12C-DHA-derived metabolites, a broader range of DHA metabolites is being surveyed. This approach will also be extended to other group of bioactive metabolites for which stable-isotopes are available with the help of automatic peak finding algorithm. DHA Status and Traumatic Brain Injury
Based on the neurotrophic mechanisms of DHA established in our laboratory, we develop strategies to improve neural resilience to brain injuries and injury outcome. We investigate
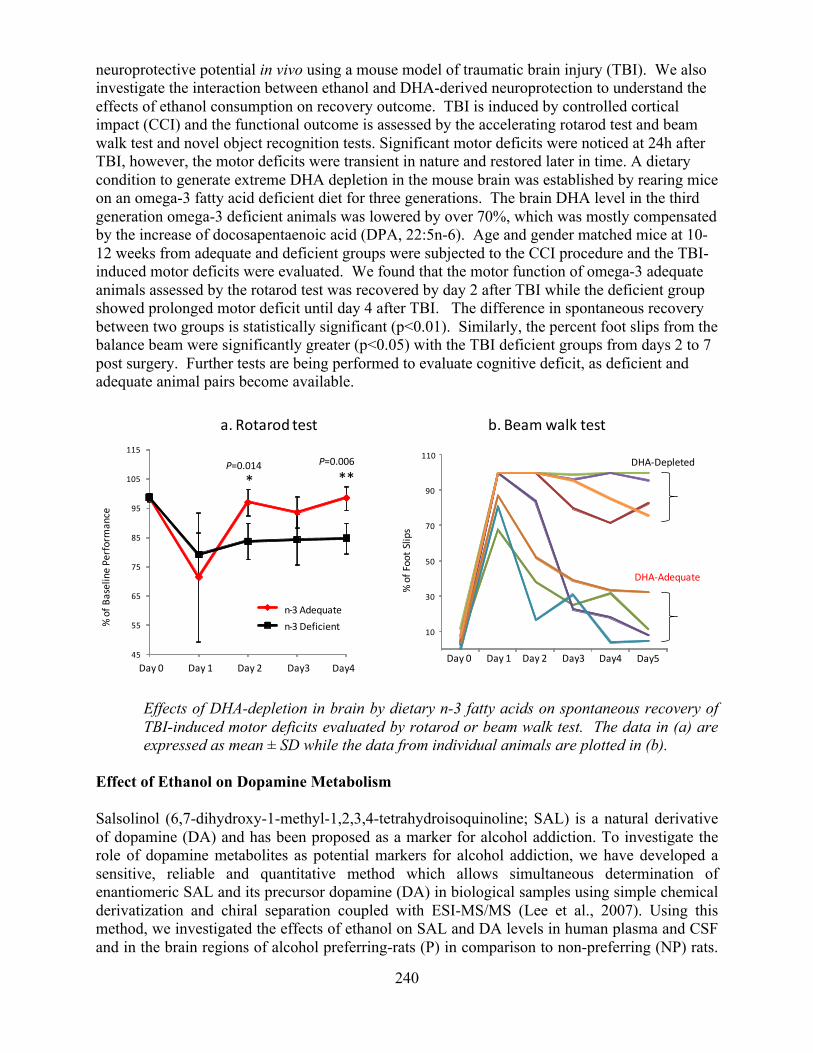
240
neuroprotective potential in vivo using a mouse model of traumatic brain injury (TBI). We also investigate the interaction between ethanol and DHA-derived neuroprotection to understand the effects of ethanol consumption on recovery outcome. TBI is induced by controlled cortical impact (CCI) and the functional outcome is assessed by the accelerating rotarod test and beam walk test and novel object recognition tests. Significant motor deficits were noticed at 24h after TBI, however, the motor deficits were transient in nature and restored later in time. A dietary condition to generate extreme DHA depletion in the mouse brain was established by rearing mice on an omega-3 fatty acid deficient diet for three generations. The brain DHA level in the third generation omega-3 deficient animals was lowered by over 70%, which was mostly compensated by the increase of docosapentaenoic acid (DPA, 22:5n-6). Age and gender matched mice at 10-12 weeks from adequate and deficient groups were subjected to the CCI procedure and the TBI-induced motor deficits were evaluated. We found that the motor function of omega-3 adequate animals assessed by the rotarod test was recovered by day 2 after TBI while the deficient group showed prolonged motor deficit until day 4 after TBI. The difference in spontaneous recovery between two groups is statistically significant (p<0.01). Similarly, the percent foot slips from the balance beam were significantly greater (p<0.05) with the TBI deficient groups from days 2 to 7 post surgery. Further tests are being performed to evaluate cognitive deficit, as deficient and adequate animal pairs become available.
45
55
65
75
85
95
105
115
Day 0 Day 1 Day 2 Day3
n-‐3 Adequaten-‐3 Deficient
P=0.014*
P=0.006
**
Day4
% of B
aseline Performance
a. Rotarod test
10
30
50
70
90
110
Day 0 Day 1 Day 2 Day3 Day4 Day5
% of Foot S
lips
DHA-‐Adequate
DHA-‐Depleted
b. Beam walk test
Effects of DHA-depletion in brain by dietary n-3 fatty acids on spontaneous recovery of TBI-induced motor deficits evaluated by rotarod or beam walk test. The data in (a) are expressed as mean ± SD while the data from individual animals are plotted in (b).
Effect of Ethanol on Dopamine Metabolism Salsolinol (6,7-dihydroxy-1-methyl-1,2,3,4-tetrahydroisoquinoline; SAL) is a natural derivative of dopamine (DA) and has been proposed as a marker for alcohol addiction. To investigate the role of dopamine metabolites as potential markers for alcohol addiction, we have developed a sensitive, reliable and quantitative method which allows simultaneous determination of enantiomeric SAL and its precursor dopamine (DA) in biological samples using simple chemical derivatization and chiral separation coupled with ESI-MS/MS (Lee et al., 2007). Using this method, we investigated the effects of ethanol on SAL and DA levels in human plasma and CSF and in the brain regions of alcohol preferring-rats (P) in comparison to non-preferring (NP) rats.

241
Contrary to the previous findings, we observed that ethanol exposure did not increase, but significantly decreased the SAL or DA levels in the striatum and nucleus accumbens in P rats, suggesting altered biosynthesis/ metabolism of DA (Lee et al., 2010). As a continuing effort to evaluate the possible contribution of ethanol to addiction processes, we extended our analysis to tetrahydropapaveroline (THP), a condensation product of DOPAL and DA. DA is metabolized by monoamine oxidase (MAO) to 3,4-dihydroxyphenylacetaldehyde (dopaldehyde, DOPAL) which is a substrate for aldehyde dehydrogenase-2 (ALDH2). We established a sensitive mass spectrometric analysis method to simultaneously determine THP, DA and SAL using the similar isolation and derivatization approaches. The recovery of THP from stripped plasma and brain tissues was greater than 80% at low pg levels. This approach allowed us to determine their levels in various regions of the rat brain. In a rat relapse model where an ALDH2 inhibitor has been shown to prevent cocaine-induced reinstatement, we found concomitant increases of THP in the VTA, substantia nigra and nucleus accumbens. These data suggest that increasing THP by selective ALDH-2 inhibition may have therapeutic potential for human cocaine addiction and preventing relapse (Yao et al., 2010). References
Akbar M, Baick J, Calderon F, Wen Z, Kim HY. 2006. Ethanol promotes neuronal apoptosis by inhibiting phosphatidylserine accumulation. J Neurosci Res 83:432-440.
Akbar M, Calderon F, Wen Z, Kim HY. 2005. Docosahexaenoic acid: a positive modulator of Akt signaling in neuronal survival. Proc Natl Acad Sci USA 102:10858-10863.
Calderon F, Kim HY. 2004. Docosahexaenoic acid promotes neurite growth in hippocampal neurons. J Neurochem 90:979-988.
Cao DH, Kevala K, Kim J, Moon HS, Jun SB, et al. 2009. Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J Neurochem 11:510-521.
Guo M, Stockert L, Akbar M, Kim HY. 2007. Neuronal specific increase of phosphatidylserine by docosahexaenoic acid. J Mol Neurosci 33:67-73.
Huang BX, Kim HY. 2006. Interdomain conformational changes in Akt activation revealed by chemical cross-linking and tandem mass spectrometry. Mol Cell Proteomics 5:1045-53.
Huang BX, Akbar M, Kevala K, Kim HY. 2011. Phosphatidylserine as a critical modulator of Akt activation. J Cell Biol 192:979-992.
Huang BX, Kim HY. 2012. Effects of ethanol on conformational changes of Akt studied by chemical cross-linking, mass spectrometry and 18O labeling. ACS Chem Biol 7:387−394.
Kim HY, Akbar M, Kim YS. 2010. Phosphatidylserine-dependent neuroprotective signaling promoted by docosahexaenoic acid. Prostaglandins Leukot and Essent Fatty Acids 82:165-172.
Kim HY, Akbar M, Lau A, Edsall L. 2000. Inhibition of neuronal apoptosis by docosahexaenoic acid (22:6n-3). Role of phosphatidylserine in antiapoptotic effect. J Biol Chem 275:35215-35223.

242
Kim HY, Bigelow J, Kevala JH. 2004. Substrate preference in phosphatidylserine biosynthesis for docosahexaenoic acid containing species. Biochemistry 43:1030-1036.
Kim HY, Lee J, Moon HS, Cao D, Kevala K, et al. 2011a. N-docosahexaenoylethanolamine promotes development of hippocampal neurons. Biochem J 435:327-336.
Kim HY, Spector A, Xiong Z. 2011b. A synaptogenic amide N-docosahexaenoylethanolamine promotes hippocampal development. Prostaglandins Other Lipid Med 96:114-120.
Kim HY. 2007. Novel metabolism of docosahexaenoic acid in neural cells. J Biol Chem 282:18661-18665.
Kimura AK, Kim HY. Phosphatidylserine synthase 2: high efficiency for synthesizing phosphatidylserine containing docosahexaenoic acid. J Lipid Res [Epub 2012 Oct 15].
Lee J, Huang BX, Yuan Z, Kim HY. 2007. Simultaneous determination of salsolinol enantiomers and dopamine in human plasma and cerebrospinal fluid by chemical derivatization coupled to chiral liquid chromatography/electrospray ionization-tandem mass spectrometry. Anal Chem 79:9166-9173.
Lee J, Ramchandani VA, Hamazaki K, Engleman EA, McBride WJ, et al. 2010. A Critical Evaluation of Influence of Ethanol and Diet on Salsolinol Enantiomers in Humans and Rats. Alcoholism-Clinical and Experimental Research 34:242-250.
Sidhu VK, Huang BX, Kim HY. 2011. Effects of docosahexaenoic acid on mouse brain synaptic plasma membrane proteome analyzed by mass spectrometry and 16O/18O labeling. J Proteome Res 10:5472-5480.
Wen Z, Kim HY. 2004. Alterations in hippocampal phospholipid profile by prenatal exposure to ethanol. J Neurochem 89:1368-1377.
Wen Z, Kim HY. 2007. Inhibition of phosphatidylserine biosynthesis in developing rat brain by maternal exposure to ethanol. J Neurosci Res 85:1568-1578.
Yao LN, Fan PD, Arolfo M, Jiang Z, Olive MF, et al. 2010. Inhibition of aldehyde dehydrogenase-2 suppresses cocaine seeking by generating THP, a cocaine use-dependent inhibitor of dopamine synthesis. Nature Medicine 16:1024-1028.
Publications 2011-2012 1. Huang BX, Akbar M, Kevala K, Kim HY. 2011. Phosphatidylserine as a critical
modulator of Akt activation. J Cell Biol 192:979-992. 2. Kim HY, Lee J, Moon HS, Cao D, Kevala K, et al. 2011.
N-docosahexaenoylethanolamine promotes development of hippocampal neurons. Biochem J 435:327-336.
3. O’Konek JJ, Illarionov P, Khursigara DS, Ambrosino E, Izhak L, et al. 2011. Novel
agonist of mouse and human iNKT cells reveals a new mechanism of tumor immunity. J Clin Inv 121: 683-694.

243
4. Kim HY, Bierbaum V, 2011. Focus in Honor of Marvin L. Vestal, Recipient of the 2010
Award for a Distinguished Contribution in Mass Spectrometry. J Am Soc Mass Spectrom 22:949-952.
5. Hankin JA, Farias S, Barkley RM, Heidenreich K, Frey LC et al.2011. MALDI Mass Spectrometric Imaging of Lipids in Rat Brain Injury Models. J Am Soc Mass Spectrom 22:1014-1021.
6. Sidhu VK, Huang BX, Kim HY. 2011. Effects of docosahexaenoic acid on mouse brain synaptic plasma membrane proteome analyzed by mass spectrometry and 16O/18O labeling. J Proteome Res 10:5472-5480.
7. Kim HY, Spector A, Xiong Z. 2011. A synaptogenic amide N-docosahexaenoylethanolamine promotes hippocampal development. Prostaglandins Other Lipid Med 96:114-120.
8. Park JH, Johansson HE, Aoki H, Huang BX, Kim HY et al. 2012. Post-translational modification by β[beta]-lysylation is required for the activity of E. coli Elongation Factor P (EF-P). J Biol Chem 2579-90.
9. Huang BX, Kim HY. 2012. Effects of ethanol on conformational changes of Akt studied by chemical cross-linking, mass spectrometry and 18O labeling. ACS Chem Biol 7: 387−394.
10. Kim HY, Spector AA. Synaptamide, endocannabinoid-like derivative of docosahexaenoic acid with cannabinoid-independent function. Prostaglandins Leukot Essent Fatty Acids [Epub 2012 Sep 5].
11. Kimura AK, Kim HY. Phosphatidylserine synthase 2: high efficiency for synthesizing phosphatidylserine containing docosahexaenoic acid. J Lipid Res [Epub 2012 Oct 15]. Collaborators Dr. William McBride, Indiana University, Indianapolis, IN Dr. Robert Chapkin, Texas A&M, College Station, TX Dr. Stephen Cunnane, University of Sherbrook, Canada Dr. Ivan Diamond, CV Therapeutics, Palo Alto, CA Dr. Robert Murphy, University of Colorado, Denver, CO Dr. Marck Rasenik, University of Illinois, Chicago, IL Dr. Won-Hwa Cho, University of Illinois, Chicago, IL Dr. Jay Berzofsky, Vaccine Branch, NCI, NIH Dr. Juan Marugan, NIH NCATS Dr. Marc Ferrer, NIH NCATS Dr. Myung-Hee Park, NIDCR, NIH Dr. David Lovinger, NIAAA, NIH Dr. David Goldman, NIAAA, NIH Dr. Margaret Davis, NIAAA, NIH

244
Dr. Stephen Ikeda, NIAAA, NIH Dr. Vijay Ramchandani, NIAAA, NIH
Laboratory of Molecular Signaling
Karl Kevala Bill Huang
Hee -‐ Yong Kim
Ji -‐ Won Lee
Vishaldeep Sidhu
Atsuko Kimura
M . Abdur Rashid
Giorgi Kharebava Abhishek Desai Zhiyi Qiang
Mohammed Akbar
Arthur Spector Rachel Quade
Dr. Kim: [email protected]

245
______________________________________________________________________
Laboratory of Liver Diseases ______________________________________________________________________
Bin Gao, MD, PhD, Chief Hua Wang, MD, PhD, Research Fellow Ogyi Park, PhD, Research Fellow Adeline Bertola, PhD, Visiting Fellow Mingjiang Xu, MD, PhD, Visiting Fellow Dechun Feng, PhD, Visiting Fellow Yung Won, PhD, Visiting Fellow Hyojung Kwun, DMV, PhD, Visiting Fellow Stephanie Mathews, PhD, Postdoctoral Fellow Yan Wang, MD, PhD, Guest Researcher Yongmei Li, MD, PhD, Special Volunteer The mission of the Laboratory of Liver Diseases is to investigate the immunological aspects and molecular pathogenesis of liver diseases, focusing on alcoholic liver disease (ALD), and to explore novel therapeutic targets to treat ALD. ALD is a major cause of chronic liver disease worldwide and can lead to fibrosis and cirrhosis. The latest surveillance report published by the National Institute on Alcohol Abuse and Alcoholism showed that liver cirrhosis was the 12th leading cause of death in the United States, with a total of 29,925 deaths in 2007, 48% of which were alcohol related. The spectrum of ALD includes simple steatosis, alcoholic hepatitis, fibrosis, cirrhosis, and superimposed hepatocellular carcinoma (Gao and Bataller, 2011). During the last two years, we have been actively investigating the cellular and molecular mechanisms of various stages of ALD and explored the therapeutic targets for the treatment of ALD with focus on several projects listed below.
Hepatoprotective effects and therapeutic potential of IL-22 Feng D, Park O, Kong X, Wang H, and Gao B in collaboration with Young H, Kolls J. We have previously demonstrated that IL-22 plays an important role in preventing liver injury induced by T cell hepatitis and IL-22 treatment prevents liver injury induced by various hepatic toxins including alcohol feeding. We have also demonstrated that hepatic expression of IL-22 is highly upregulated in patients with viral hepatitis; however, hepatic IL-22 is not significantly elevated in mouse models of chronic liver injury. During the last two years, we developed liver-specific IL-22 transgenic mice with overexpression of IL-22 in the liver, which mimics the high hepatic levels of IL-22 in patients with viral hepatitis. By using IL-22 transgenic mice, we have demonstrated that IL-22 protects against Con A-induced T cell hepatitis, inhibits liver fibrosis by inducing hepatic stellate cell senescence, and promotes liver progenitor cell proliferation. Because IL-22 receptor expression is restricted to epithelial cells, IL-22 treatment may generate few side effects. Our findings suggest that IL-22 treatment is a potential therapeutic option to treat ALD, due to its antioxidant, antiapoptotic, anti-steatotic, proliferative, and antimicrobial effects with the added benefit of potentially few side effects due to restricted expression of IL-22R. Indeed, IL-22 is currently being evaluated in phase I clinical trials with healthy volunteers by Generon (Shanghai) Corporation Ltd. Generon also plans to conduct the phase II trial with IL-22 for the treatment of alcoholic hepatitis. (Feng et al., 2012; Kong et al. 2012; Park et al., 2011)

246
Inflammation-associated IL-6/STAT3 activation ameliorates alcoholic and nonalcoholic fatty liver diseases in IL-10 deficient mice Miller A, Wang H, Park O, Yin S, Horiguchi N, Ki S, Lafdil F, and Gao B
Alcoholic and nonalcoholic steatohepatitis are characterized by fatty liver plus inflammation. It is generally believed that steatosis promotes inflammation, while inflammation in turn aggravates steatosis. Thus, we hypothesized the deletion of IL-10, a key anti-inflammatory cytokine, exacerbates liver inflammation, steatosis, and hepatocellular damage after feeding with an ethanol diet or a high fat diet. During the last two years, we tested this hypothesis by examining alcoholic and nonalcoholic fatty livers in IL-10 knockout (IL-10-/-) mice and several other strains of genetically modified mice. We demonstrated that IL-10-/- mice had greater liver inflammation with higher levels of IL-6 and hepatic STAT3 activation, but less steatosis and hepatocellular damage after alcohol or high fat diet feeding. An additional deletion of IL-6 or hepatic STAT3 restored steatosis and hepatocellular damage but further enhanced liver inflammation in IL-10-/- mice. In addition, the hepatic expression of SREBP1c and key downstream lipogenic proteins and enzymes in fatty acid synthesis were downregulated in IL-10-
/- mice. Conversely, IL-10-/- mice displayed enhanced levels of phosphorylated (activated) AMPK, phosphorylated (inhibited) ACC1 and CPT-1 in the liver. Such dysregulations were corrected in IL-10-/-IL-6-/- or IL-10-/- STAT3Hep-/- double knockout mice. Our findings suggest that IL-10-/- mice are prone to liver inflammation but resistant to steatosis and hepatocellular damage induced by ethanol or high fat diet feeding. Resistance to steatosis in these mice is attributable to elevation of inflammation-associated hepatic IL-6/STAT3 activation that subsequently downregulates lipogenic genes but upregulates fatty acid oxidation-associated genes in the liver (Miller et al. 2011).
Chronic and binge ethanol feeding synergistically accelerate alcoholic liver injury and inflammation: a critical role for E-selectin Bertola A, Mathews S., Park O, Li Y, Wang H, and Gao B
More than 95% of heavy drinkers develop fatty liver, but only 20-40% of them develop steatohepatitis and progress to cirrhosis and hepatocellular carcinoma. The underlying mechanisms that render some individuals more susceptible to severe forms of ALD are not clear, and many risk factors may be involved. These potential risk factors include sex, obesity, dietary factors, smoking, and non-sex-linked genetic factors. Additionally, several epidemiologic studies have suggested that the drinking pattern also significantly influences the development and progression of ALD in humans. For example, drinking outside mealtime, weekend drinking, the consumption of multiple types of drinks, and a mixed drinking pattern were significantly associated with an increased risk of ALD. However, the molecular mechanisms through which drinking pattern affects ALD pathogenesis remain unknown. Previously, we have demonstrated that chronic ethanol feeding for 10 days plus a single binge dose of ethanol delivered by gavage synergistically elevated serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), induced steatosis, and upregulated the expression of proinflammatory cytokines (Ki et al., 2010). This pattern of chronic-binge ethanol feeding reproduces closely the chronic plus binge consumption patterns in many heavy drinkers, which may be associated with an increased risk of ALD. During the last two years, we have further characterized liver inflammation in this mouse model of chronic-binge ethanol feeding. We show that chronic plus binge ethanol feeding synergistically upregulated the hepatic expression of IL-1β and TNF-α

247
and induced neutrophil accumulation in the liver compared to chronic or binge feeding alone. The in vivo depletion of neutrophils through the administration of an anti-Ly6G antibody markedly reduced chronic-binge ethanol feeding-induced liver injury. Real-time PCR analyses revealed that hepatic E-selectin expression was upregulated 10-fold, whereas the expression of other neutrophil infiltration-related adhesion molecules (such as P-selectin, ICAM-1, and VCAM-1) was slightly upregulated or downregulated in this chronic-binge model. The genetic deletion of E-selectin prevented chronic-binge ethanol-induced hepatic neutrophil infiltration and the elevation of serum transaminases without affecting ethanol-induced steatosis. In addition, E-selectin-deficient mice showed reduced hepatic expression of several proinflammatory cytokines, chemokines, and adhesion molecules compared to wild-type mice after chronic-binge ethanol feeding. Our findings suggest that chronic-binge ethanol feeding synergistically upregulates the expression of proinflammatory cytokines, followed by the induction of E-selectin. Elevated E-selectin plays an important role in neutrophil recruitment, inflammation, and injury in the alcoholic liver injury induced by chronic-binge ethanol feeding. (Bertola et al., 2013a,b).
Hepatoprotective versus oncogenic functions of signal transducer and activator of transcription 3 in liver tumorigenesis Wang H, Lafdil F, Park O, Yin S, Miller A, Gao B in collaboration with Sun Z
Aberrantly hyperactivated signal transducer and activator of transcription 3 (STAT3) has been found in human liver cancers including alcoholic liver cancer as an oncogene; however, STAT3 has also been shown to exert hepatoprotective effects during liver injury (Wang et al., 2011). The balancing act that STAT3 plays, between hepatoprotection and liver tumorigenesis, remains poorly defined. During the last two years, we have used both the diethylnitrosamine (DEN)-induced liver tumor model and the chronic carbon tetrachloride (CCl4)-induced liver fibrosis model to investigate the role of STAT3 in liver tumorigenesis. We demonstrated that hepatocyte-specific STAT3 knockout mice were resistant to liver tumorigenesis induced by a single dose of DEN injection, whose tumorigenesis was associated with minimal chronic liver inflammation, injury, and fibrosis. In contrast, chronic CCl4 treatment resulted in severe hepatic oxidative damage, inflammation, fibrosis, but rarely induced liver tumor formation in wild-type mice. Surprisingly, despite the oncogenic function of STAT3 in DEN-induced liver tumor, hepatocyte-specific STAT3 knockout mice were more susceptible to liver tumorigenesis after 16 weeks of CCl4 injection, which was associated with higher levels of liver injury, inflammation, fibrosis, and oxidative DNA damage compared with wild-type mice. Our findings suggest that hepatoprotective feature of STAT3 prevents hepatic damage and fibrosis under the condition of persist inflammatory stress, consequently suppressing liver tumorigenesis (Wang et al., 2011).
Suppression of innate immunity (NK/IFN-γ) in the advanced stages of liver fibrosis in mice Wang H, Park O, Miller A, Gao B in collaboration with Jeong W
Activation of innate immunity (natural killer [NK] cell/interferon-γ [IFN-γ]) has been shown to play an important role in antiviral and antitumor defenses. Our previous studies suggest that activation of NK cells ameliorates liver fibrosis by killing the activated hepatic stellate cells (HSCs) and producing IFN-γ that induces HSC cell cycle arrest and apoptosis (Radaeva et al. 2006) However, little is known about the regulation of innate immunity during chronic liver injury. In the last two years, we have compared the functions of NK cells in early and advanced liver fibrosis induced by a 2-week or a 10-week carbon tetrachloride (CCl4) challenge, respectively. Injection of polyinosinic-polycytidylic acid (poly I:C) or IFN-γ induced NK cell activation and NK cell killing of hepatic stellate cells (HSCs) in the 2-week CCl4 model. Such

248
activation was diminished in the 10-week CCl4 model. Consistent with these findings, the inhibitory effect of poly I:C and IFN-γ on liver fibrosis was markedly reduced in the 10-week versus the 2-week CCl4 model. In vitro coculture experiments demonstrated that 4-day cultured (early activated) HSCs induce NK cell activation via an NK group 2 member D/retinoic acid-induced early gene 1-dependent mechanism. Such activation was reduced when cocultured with 8-day cultured (intermediately activated) HSCs due to the production of transforming growth factor-β (TGF-β) by HSCs. Moreover, early activated HSCs were sensitive, whereas intermediately activated HSCs were resistant to IFN-γ-mediated inhibition of cell proliferation, likely due to elevated expression of suppressor of cytokine signaling 1 (SOCS1). Disruption of the SOCS1 gene restored the IFN-γ inhibition of cell proliferation in intermediately activated HSCs. Production of retinol metabolites by HSCs contributed to SOCS1 induction and subsequently inhibited IFN-γ signaling and functioning, whereas production of TGF-β by HSCs inhibited NK cell function and cytotoxicity against HSCs. Our findings suggest that the antifibrogenic effects of NK cell/IFN-γ are suppressed during advanced liver injury, which is likely due to increased production of TGF-β and expression of SOCS1 in intermediately activated HSCs. (Jeong et al., 2011).
Key prior publications
1. Hong F, Jaruga B, Kim W, Radaeva S, Tian Z, et al.: Opposing roles of STAT1 and STAT3 in T cell-mediated hepatitis: regulation by SOCS. J. Clin. Invest. 2002, 110: 1503-1513
2. Radaeva R, Sun R, Pan H, Hong F, Gao B.: Interleukin-22 (IL-22) plays a protective role in T cell hepatitis: IL-22 is a survival factor for hepatocytes via activation of STAT3. Hepatology 2004, 39:1332-1342
3. Ki S, Park O, Zheng M, Kolls J, Batller R, Gao B.: IL-22 treatment ameliorates alcoholic liver injury in a murine model of chronic plus binge drinking. Hepatology 2010, 52:1291-300
Publications 2011-2012
Original Articles:
1. Yin S, Wang H, Park O, Wei W, Shen, J, Gao B. 2011. Enhanced liver regeneration in IL-10 deficient mice after partial hepatectomy by stimulating inflammatory response and activating hepatocyte STAT3. Am. J. Pathol. 178:1614-1621
2. Wang H, Lafdil F, Wang L, Yin S, Feng D, Gao B. 2011. Tissue inhibitor of metalloproteinase 1 (TIMP-1) deficiency exacerbates CCl4-induced liver injury and fibrosis in mice: involvement of hepatocyte STAT3 and TIMP-1 production. Cell & Bioscience 1:14 doi:10.1186/2045-3701
3. Zhang Z, Zhang S, Zou Z, Fan R, Zhao J, et al. 2011. Roles of NK cells in liver injury and fibrosis in patients with chronic HBV infection. Hepatology 53:73-85
4. Jeong W, Park O, Wang L, Suh Y, Byun J, et al. 2011. Suppression of innate immunity (natural killer cells and IFN-γ) in the late stages of liver injury. Hepatology 53:1342-1351
5. Li Y, Xu S, Mihaylova M, Zheng B, Hou X, et al. 2011. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin resistant mice. Cell Metabolism 13:376-88

249
6. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, et al. 2011. Hyperactivation of anandamide synthesis and regulation of cell cycle progression via CB1 receptors in the regenerating liver. Proc. Natl. Acad. Sci. USA 108:6323-8
7. Park O, Wang H, Weng H, Feigenbaum L, Li H, et al. 2011. In vivo consequences of liver-specific interleukin-22 expression in mice: implications for human liver disease progression. Hepatology 54:252-261
8. Wang H, Lafdil F, Wang L, Park O, Yin S, et al. 2011. Hepatoprotective versus oncogenic functions of STAT3 in liver tumorigenesis in mice. Am. J. Pathol. 179:714-724
9. Miller A, Wang H, Park O, Horiguchi N, Ki S, et al. 2011. Inflammation-associated hepatic IL-6/STAT3 activation ameliorates alcoholic and nonalcoholic fatty liver diseases in IL-10 deficient mice. Hepatology 54:846-856
10. Feng D, Park O, Radaeva S, Wang H, Yin S, et al. 2012. Interleukin-22 ameliorates cerulein-induced pancreatitis in mice via inhibition of the autophagic pathway. Int. J. Biol. Sci. 8:249-257
11. Ji Y, Xia S, Yang L, Sun S, Bhargava P, et al. 2012. Lipid-sensing NKT cells in adipose tissue promote M2 macrophage polarization and glucose homeostasis via IL-4/STAT6 in obesity. J. Biol. Chem. 287:13561-13571
12. Kong X, Feng D, Wang H, Hong F, Bertola A, et al. 2012. Interleukin-22 induces hepatic stellate cell senescence and restricts liver fibrosis. Hepatology 56:1150-1159
13. Feng D, Kong X, Weng H, Park O, Wang H, et al. 2012. Interleukin-22 promotes liver progenitor cell proliferation in patients with viral hepatitis and in mice. Gastroenterology 143:188-198
14. Hisada M, Ota Y, Zhang X, Cameron A, Gao B, et al. 2012. Successful transplantation of reduced sized rat alcoholic fatty livers made possible by mobilization of host stem cells. Am. J. Transplant. 12:3246-56.
15. Xiao C, Wang R, Lahusen T, Park O, Bertola A, et al. 2012. Progression of chronic liver inflammation and fibrosis driven by activation of c-Jun signaling in Sirt6 mutant mice. J. Biol. Chem. 287:41903-13
Reviews, Editorials, Book Chapters:
16. Gao B, Seki E, Brenner D, Friedman S, Cohen J, et al. 2011. Innate immunity in alcoholic liver disease (Review). Am. J. Physiol. (GI and Liver) 300:G516-25
17. Gao B, Bataller R. 2011. Alcoholic liver disease: novel concepts and therapeutic targets (Review). Gastroenterology 141:1572-1585
18. Gao B, Wang H, Lafdil F, Feng D. 2012. The STAT proteins: key regulators of antiviral response, inflammation and tumorigenesis in the liver (Review). Journal of Hepatology 57:430-441
19. Wang H, Gao B, Zakhari S, Nagy L. 2012. Inflammation in alcoholic liver disease (Review). Annual Reviews of Nutrition 32:343-68
20. Miller A, Horiguchi N, Jeong W, Radaeva S, Gao B. 2011. Molecular mechanisms of alcoholic liver disease: Role of IL-6/STAT3 and natural killer cells (Review). Alcohol Clin. Exp. Res. 35:787-793
21. Wang H, Lafdil F, Kong X, Gao B. 2011. Signal transducer and activator of transcription 3 in liver disease: a novel therapeutic target (Review). Int J Biol Sci. 7:536-550

250
22. Gao B. 2012. Hepatoprotective and antiinflammatory cytokines in alcoholic liver disease
(Review). J. Gastro. Hepatol. Suppl 2:89-93 23. Kong X, Horiguchi N, Mori M, Gao B. 2012. Cytokines, STATs, and liver fibrosis
(Review). Frontiers in Bioscience 3:1-5 24. Wang Y, Gao B. 2012. Functions of signal transducers and activators of transcription
(STAT) in the liver (Review). J. Clinical Hepatol. 25. Zakari S, Gao B, Hoek J. 2012. Alcoholic liver disease. Drug Abuse & Addition in Medical
Illness edited by Joris C Verster (book chapter) 26. Gao B, Bertola A. 2011. NK cells take two Tolls to destruct bile ducts (Editorial).
Hepatology 53:1076-1079 27. Gao B, Waisman A. 2012. Th17 cells regulate liver fibrosis by targeting multiple cell
types: Hit many birds with one stone (Editorial). Gastroenterology 143:536-539 Collaborators: Dr. George Kunos, NIAAA, NIH Dr. Pal Pacher, NIAAA, NIH Dr. BJ Song, NIAAA, NIH Dr. Howard Young, NCI, NIH Dr. Yingzi Yang, NHGRI, NIH Dr. Lothar Hennighausen, NIDDK, NIH Dr. Chuxia Deng, NIDDK, NIH Dr. Ray Donnelly, FDA Dr. Ramon Bataller, UNC, NC Dr. Zhaoli Sun, Johns Hopkins, MD Dr. Mengwei Zang, Boston University, MA Dr. Jay Kolls, University of Pittsburgh, PA Dr. Lin Qi, Cornell University, NY Dr. Eric Gershwin, UC Davis, CA Dr. Geoffrey Thiele, Nebraska Medical Center, NE Dr. Vasilis Vasiliou, University of Colorado, CO Dr. Jack Cowland, University of Copenhagen, Denmark Dr. Honglei Weng, Manheim, Germany Dr. Steven Dooley, Manheim, Germany Dr. Fu-sheng Wang, Beijing, China Dr. Won-IL Jeong, KIAST, Korea Dr. Philippe Gual, INSERM U895, Nice, France
Dr.Gao: [email protected]

251
______________________________________________________________________
Chemical Biology Research Branch, Drug Design and Synthesis Section (a joint NIAAA-NIDA laboratory)
______________________________________________________________________ Kenner C. Rice, PhD, Chief Mariena Mattson, AA, Lab Manager Kejun Cheng, PhD, Staff Scientist Arthur Jacobson, PhD, Contractor/Consultant Joshua Antoline, PhD, IRTA Fellow Peter DeMatteo, Ph D, IRTA Fellow Mohd Kabir, PhD, IRTA Fellow Fuying Li, PhD, Visiting Fellow Brandon Selfridge, PhD, IRTA Fellow Masaki Suzuki, PhD, Special Volunteer Background and Introduction The present Chemical Biology Research Branch (CBRB), National Institute on Drug Abuse and National Institute on Alcohol Abuse and Alcoholism is one of the oldest continuous programs at NIH. Its original predecessor was established in 1929 under the auspices of the National Academy of Sciences, National Research Council, as The Drug Addiction Laboratory with Dr. L. F. Small as Chief. This was the chemical arm of a program aimed at the development of a strong analgesic without the undesirable side effects of morphine, and further understanding of the biological basis of drug addiction. The original program also included a pharmacology group under Dr. Nathan Eddy at the University of Michigan, Ann Arbor, that is still active under the direction of Dr. James Woods. The third arm of this program was a clinical group originally headed by Dr. Clifton Himmelsbach in Lexington, KY that became the present-day NIDA Addiction Research Center in Baltimore. Many fundamental contributions to the chemistry and pharmacology of drugs of abuse have resulted from work originated in CBRB and its predecessors. The following are selected, much-abbreviated examples. From the early work of L. F. Small and associates: extensive studies on the chemistry of the opium alkaloids and their transformation products that defined many important reaction pathways and largely the chemical character of reactivity in this series; Small developed the foundation of modern structure activity relationships (SAR) in the opiate series by systematic study relating structure and pharmacologic activity of opium alkaloids and their derivatives. This work clearly showed that chemical modification of opium alkaloids could produce frequently predictable changes in the pharmacologic activity of the resulting drug. His discovery of the opioid agonist metopon and its evaluation in humans provided robust proof of principle of the original program hypothesis that chemical modification of the morphine molecule could result in separation of analgesia from tolerance and dependence. This guiding principle still permeates opioid research today. Later examples from the work of E. L. May and associates include: definitive chemical synthesis and pharmacological study of the methadone and methadol stereoisomers leading to the development of methadone for the treatment of heroin addiction; the discovery of the 5-phenylmorphans and introduction of the 6,7-benzomorphans as analgesics leading to clinically useful drugs and valuable research tools for study of the opioid receptor-endorphin system. This work enabled the agonist-antagonist approach to analgesic development and ultimately resulted in others obtaining the state of the art agonist-antagonist drugs buprenorphine, butorphanol and nalbuphine. May

252
also discovered a bromophenanthrene antimalarial that was widely used as a life-saving drug during the Vietnam War. His discovery of nonnarcotic strong analgesics in the cannabinoid series led others to extend this work and to the biochemical identification of the cannabinoid receptor. More recent advances emanating from this program under the present leadership include: the chemical synthesis and introduction of unnatural opiates as potential drugs and as research tools for the opioid receptor-endorphin system and the toll-4 receptor system; the development of highly selective affinity labels for the mu, delta and kappa opioid receptor subtypes as essential research tools for studies of the opioid receptor-endorphin system; purification of the delta opioid receptor to homogeneity rendering it available for study seven years prior to its availability by cloning. Definitive autoradiographic studies of cannabinoid receptors provided detailed anatomical localization in a number of species including humans. This work explained the almost complete lack of acute toxicity of marijuana and led to the identification by others of the cloned cannabinoid receptor. The discovery of the delta opioid receptor agonist SNC-80 provided a now widely used delta opioid receptor research tool. Our introduction of the oxide-bridged 5-phenylmorphans has provided novel research tools for investigation of opioid receptor subtypes. We developed the fluorinated narcotic antagonist cyclofoxy as a ligand for positron emission tomography (PET) imaging and obtained the first images of opioid receptor occupancy in the living primate. We also developed cyclofoxy for human study and it remains presently available. Cyclofoxy was the first PET ligand developed solely at a single institution from conceptualization, to chemical synthesis, to labeling with a positron emitting isotope and to successful use as an imaging agent in animals and humans. We advanced the corticotropin releasing hormone receptor type 1 (CRH1) antagonist antalarmin for study of stress-related disorders and it has proven to be a valuable research tool in many paradigms. A practical process for kilogram synthesis of antalarmin was developed in CBRB that provided material for primate studies and numerous in vivo and in vitro studies. Novel drugs were designed and synthesized for study of the benzodiazepine receptors, phencyclidine receptors, dopamine transporters, several enzymes and numerous other biological targets. Potential medications for cocaine and methamphetamine abuse were developed that include several drugs that largely prevent cocaine self-administration in primates without affecting feeding, the standard measure of normal behavior. One such drug showed this profile for as long as a month with a single dose. Finally, the CBRB development of the NIH Opiate Total Synthesis has provided a practical process for the chemical synthesis of opium-derived narcotics and antagonists and their unnatural enantiomers. Process development of this patented, nonchromatographic methodology has now been completed by Mallinckrodt Medical Narcotics Division for synthetic production of medical opiates and their antagonists. It is now economically competitive with production from opium for those products targeted for production by the NIH Opiate Synthesis. The process also offers independence from opium and opium poppy eradication as a strategy to eliminate heroin production. It is the only methodology for this purpose despite continuing attempts by many chemical research groups to develop such a process over the last 82 years, and major advances in organic chemical synthesis made during this time. With the relocation of CBRB to NIDA and NIAAA in 2006, two of the three original arms of the program established in 1929 are reunited. Present Research Design and Synthesis of Novel Drugs for the Treatment, Prevention and Understanding of Drug Abuse and Related Disorders: The goals and objectives of this program are the discovery of novel drugs for the treatment, prevention and understanding of drug abuse and dependence, and related disorders. These goals require (1) the rational design and chemical synthesis of diverse,

253
structurally complex new chemical entities as potential medications, and as research tools to advance understanding of the actions of drugs of abuse, and (2) multidisciplinary collaborative studies with a wide number of outstanding biologists, pharmacologists, basic behavioral researchers and clinicians for studying compounds of mutual interest. The systems that mediate the actions of opiates, cannabinoids, alcohol, hallucinogens, and designer drugs present in the widely abused “bath salts” and similar preparations are now under study. Novel Opioid Receptor Ligands as Research Tools and Potential Drugs The opioid receptor endorphin system consists of saturable, enantioselective, high affinity mu, delta and kappa opioid receptor types located in anatomically well defined areas of the mammalian CNS. Numerous endogenous opioid peptides (endorphins) function as the endogenous ligands for these receptors. This system mediates the analgesic, euphoric and addictive effects of narcotic drugs and contributes to regulation of numerous physiologic and behavioral functions in its normal state. This system is dysregulated by the abuse of prescription opioids, heroin, and other drugs. The abuse of prescription narcotic drugs alone in the U.S. is presently an enormous and growing problem. In 2010, about 360,000 emergency room episodes (nearly 1000/day) resulted from the abuse of prescription opioids, an increase of 79% from 2006. Landmark biophysical advances from several laboratories in 2012 have resulted in the crystallization and X-ray structural determination of the mu, delta and kappa opioid receptors. These are enormously important results that will enable a much more precise understanding of how different classes of opioid drugs interact with the receptors to elicit their effects, both desirable and undesirable. In addition, recent pharmacologic advances have shown (a) that highly selective delta receptor antagonists might be valuable medications for the treatment and prevention of human narcotic abuse and (b) that a drug showing a mu agonist-delta antagonist profile might produce strong analgesia without producing tolerance and dependence thus allowing continuous treatment of chronic pain. Optimal exploitation of these and other similarly intriguing observations now requires novel, exquisitely selective, nonpeptide ligands as research tools and potential medications. These new tools will enable the study of many questions of fundamental importance concerning the function of mu, delta and kappa opioid receptor subtypes and how drugs interact with their receptors to elicit these functions. We have, and are continuing to design, synthesize and evaluate novel drugs for these purposes. We earlier identified a mu agonist-delta antagonist and a delta inverse agonist in the 5-phenylmorphan series. We recently identified a morphine-like mu agonist and also a mu antagonist in a series of conformationally restrained 5-phenylmorphans (see below). The diverse profiles obtained in this series illustrate the importance of subtle changes in the carbon-nitrogen skeleton and careful attention to stereochemical detail, and can provide important leads toward novel pain medications with reduced side effects and further understanding of drug-receptor interactions. Collaborative computer assisted molecular modeling and ab initio quantum mechanical chemical methods are being employed in the design of these compounds. We have also developed a diastereoselective one-pot synthesis of 7- and 8-substituted 5-phenylmorphans and have utilized it to develop novel, high affinity (subnanomolar) opioid ligands. These and other novel drugs prepared in our studies are or will be studied in the appropriate in vitro binding assays in native and cloned systems, smooth muscle assays, and in vivo assays in studies aimed at gaining further insight into the function of the opioid receptor endorphin system.
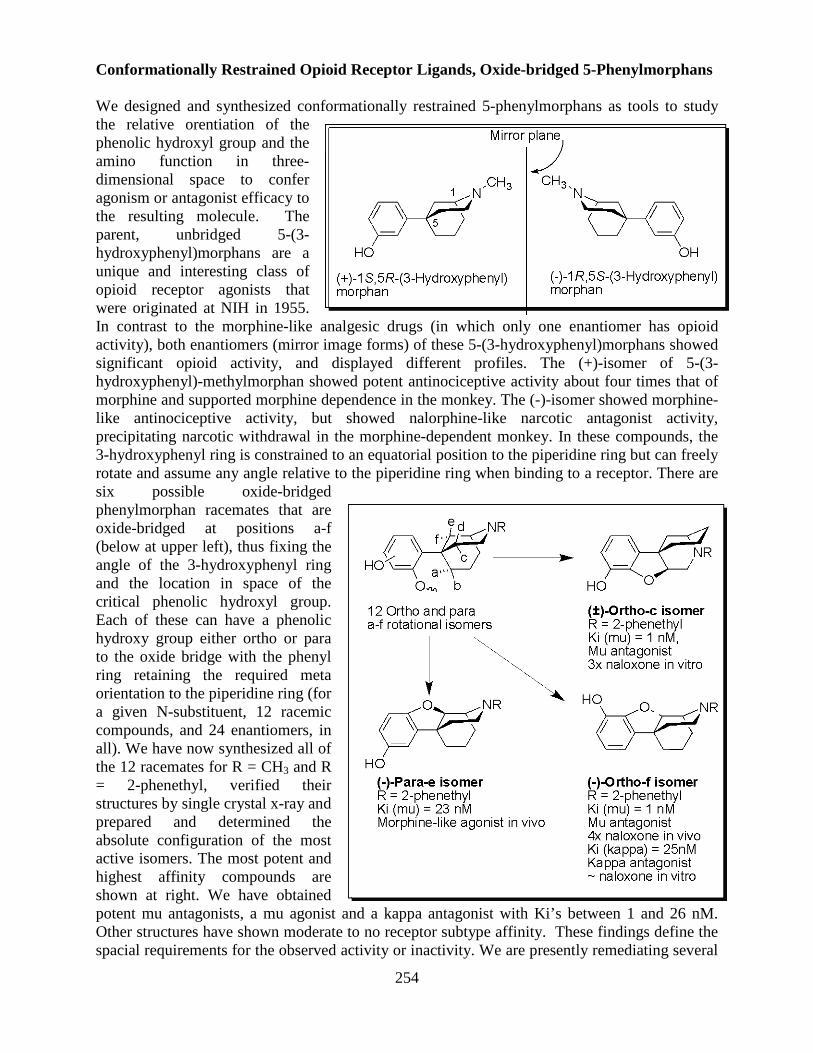
254
Conformationally Restrained Opioid Receptor Ligands, Oxide-bridged 5-Phenylmorphans We designed and synthesized conformationally restrained 5-phenylmorphans as tools to study the relative orentiation of the phenolic hydroxyl group and the amino function in three-dimensional space to confer agonism or antagonist efficacy to the resulting molecule. The parent, unbridged 5-(3-hydroxyphenyl)morphans are a unique and interesting class of opioid receptor agonists that were originated at NIH in 1955. In contrast to the morphine-like analgesic drugs (in which only one enantiomer has opioid activity), both enantiomers (mirror image forms) of these 5-(3-hydroxyphenyl)morphans showed significant opioid activity, and displayed different profiles. The (+)-isomer of 5-(3-hydroxyphenyl)-methylmorphan showed potent antinociceptive activity about four times that of morphine and supported morphine dependence in the monkey. The (-)-isomer showed morphine-like antinociceptive activity, but showed nalorphine-like narcotic antagonist activity, precipitating narcotic withdrawal in the morphine-dependent monkey. In these compounds, the 3-hydroxyphenyl ring is constrained to an equatorial position to the piperidine ring but can freely rotate and assume any angle relative to the piperidine ring when binding to a receptor. There are six possible oxide-bridged phenylmorphan racemates that are oxide-bridged at positions a-f (below at upper left), thus fixing the angle of the 3-hydroxyphenyl ring and the location in space of the critical phenolic hydroxyl group. Each of these can have a phenolic hydroxy group either ortho or para to the oxide bridge with the phenyl ring retaining the required meta orientation to the piperidine ring (for a given N-substituent, 12 racemic compounds, and 24 enantiomers, in all). We have now synthesized all of the 12 racemates for R = CH3 and R = 2-phenethyl, verified their structures by single crystal x-ray and prepared and determined the absolute configuration of the most active isomers. The most potent and highest affinity compounds are shown at right. We have obtained potent mu antagonists, a mu agonist and a kappa antagonist with Ki’s between 1 and 26 nM. Other structures have shown moderate to no receptor subtype affinity. These findings define the spacial requirements for the observed activity or inactivity. We are presently remediating several

255
low yielding steps in the synthesis of the ortho-c isomer to enable the synthesis of sufficient material for in vivo study. The optical resolution of the racemic N-phenethyl ortho-c isomer (Ki = 1 nM at mu and potent mu antagonist in vitro) has provided one enantiomer with subnanomolar mu affinity (0.5 nM). This is expected to constitute a very valuable systemically active selective mu antagonist and an important lead compound for further elaboration. At present, the only highly selective mu antagonists are peptides that are only active when administered directly into the brain. In collaboration with Dr. Yong Lee (CIT, NIH), an ab initio quantum chemical study of the 24 oxide-bridged phenylmorphan ligands has been carried out. These calculations clearly show the approximately 60° turn of the phenolic ring in the a-f oxide-bridged phenylmorphan isomers. The determination of the pharmacology of the synthesized isomers is continuing. We are now extending these studies to partial structures and more flexible structural analogs. We conceptualized oxide-bridged phenylmorphans as topologically distinct, structurally rigid ligands with 3-dimensional shapes that could not be appreciably modified on interaction with opioid receptors. An enantiomer of the N-phenethyl-substituted ortho-f isomer was found to have high affinity for the mu receptor and was about four times more potent than naloxone as an antagonist. In order to examine the effect of the introduction of a small amount of flexibility into these molecules, we have replaced the rigid 5-membered oxide ring with a more flexible 6-membered carbon ring structures shown below. Synthesis of the new N-phenethyl-substituted tricyclic N-substituted-2,3,4,9,10,10a-hexahydro-1H-1,4a-(epiminoethano)phenanthren-6- and 8-ols resulted in a two carbon-bridged relative of the f-isomers, the dihydrofuran ring being replaced by a cyclohexene ring. The carbocyclic compounds had much higher affinity and greater selectivity for the mu receptor than the f-oxide-bridged phenylmorphans. They were also much more potent mu antagonists than naloxone, with activities comparable to naltrexone in the [35S]GTP-Gamma-S assay for narcotic antagonism. Unnatural Enantiomers of (+)-Naloxone and Related Compounds as Functional Antagonists of Toll-like 4 Receptors Earlier, in collaboration with Linda Watkins at the University of Colorado, Boulder, we introduced the concept of toll-like receptor (TLR)-mediated glial activation as central to all of the following: neuropathic pain, compromised acute opioid analgesia, and unwanted opioid side effects (tolerance, dependence, and reward). Attenuation of glial activation was demonstrated both to alleviate exaggerated pain states induced by experimental pain models and to reduce the development of opioid tolerance. We first synthesized and characterized unnatural (+)-naloxone in 1978 as an opioid receptor inert research tool useful for detecting opioid receptor mediated effects when used in conjunction with (-)-naloxone, a high affinity, clinical useful narcotic antagonist. We know now that (+)-naloxone

256
and (+)-naltrexone, long thought to be inert compounds, are functional antagonists of TLR-4 receptors and that selective acute functional antagonism of TLR4 by (+)-naloxone results in reversal of neuropathic pain as well as potentiation of opioid analgesia. Opioid action has been thought to exert reinforcing effects solely via the initial agonist response of opioid receptors. We now have evidence for an additional novel contributor to opioid reward: the innate immune pattern-recognition receptor, toll-like receptor 4 (TLR4)/mylenoid differentiation factor (MD2), and its gene-dependent myeloid differentiation primary response gene 88 (MyD88) signaling. Blockade of TLR4/MD2 by administration of the nonopioid, (+)-naloxone (rats), or two independent genetic knock-outs of MyD88-TLR4-dependent signaling (mice), suppressed opioid-induced conditioned place preference. (+)-Naloxone also reduced opioid (remifentanil) self-administration (rats), another commonly used behavioral measure of drug reward. Moreover, pharmacological blockade of morphine-TLR4/MD2 activity potently reduced morphine-induced elevations of extracellular dopamine in rat nucleus accumbens, a region critical for opioid reinforcement. Importantly, opioid-TLR4 actions are not a unidirectional influence on opioid pharmacodynamics, since TLR4(-/-) mice had reduced oxycodone-induced p38 and JNK phosphorylation, while displaying potentiated analgesia. Similar to our recent reports of morphine-TLR4/MD2 binding, we have provided a combination of in silico and biophysical data to support (+)-naloxone and remifentanil binding to TLR4/MD2. Collectively, these data indicate that the actions of opioids at classical opioid receptors, together with their newly identified TLR4/MD2 actions, affect the mesolimbic dopamine system that amplifies opioid-induced elevations in extracellular dopamine levels, therefore possibly explaining altered opioid reward behaviors. Thus, the discovery of TLR4/MD2 recognition of opioids as foreign xenobiotic substances adds to the existing hypothesized neuronal reinforcement mechanisms, identifies a new drug target in TLR4/MD2 for the treatment of addictions, and provides further evidence supporting a role for central proinflammatory immune signaling in drug reward. We previously showed that both the opioid antagonist (-)-naloxone and the non-opioid (+)-naloxone inhibit toll-like receptor 4 (TLR4) signaling and reverse neuropathic pain expressed shortly after chronic constriction injury. We have now found that the TLR4 contributes to neuropathic pain in another major model (spinal nerve ligation) and to long established (2-4 months) neuropathic pain, not just to pain shortly after nerve damage. Analyses of plasma levels of (+)-naloxone after subcutaneous administration indicate that (+)-naloxone has comparable pharmacokinetics to (-)-naloxone with a relatively short half-life. This finding accounts for the rapid onset and short duration of allodynia reversal produced by subcutaneous (+)-naloxone. Since toll-like receptor 2 (TLR2) has also recently been implicated in neuropathic pain, cell lines transfected with either TLR4 or TLR2, necessary co-signaling molecules, and a reporter gene were used to define whether (+)-naloxone effects could be accounted for by actions at TLR2 in addition to TLR4. (+)-Naloxone inhibited signaling by TLR4 but not TLR2. These studies provide evidence for broad involvement of TLR4 in neuropathic pain, both early after nerve damage and months later. In addition, they provide further support for the TLR4 inhibitor (+)-naloxone as a novel candidate for the treatment of neuropathic pain. Both (+)-naloxone and (+)-naltrexone were inactive in the NOVA screen for 63 receptors and enzymes indicating there will be few, if any, off-target side effects. In summary, our studies demonstrated that (+)-naloxone, a systemically available, blood-brain barrier permeable, small molecule TLR4 inhibitor can reverse neuropathic pain in rats, even months after nerve injury. These findings suggest that (+)-naloxone, or similar compounds, be considered as a novel, first-in-class treatment for

257
neuropathic pain. We are continuing to synthesize unnatural opioids and antagonists and study their effects on TLR-4 receptors. Opioid Program Summary Current efforts and future goals for the opioid program consist of the design, chemical synthesis and pharmacological study of novel opioid receptor subtype selective agonists and antagonists. These compounds will serve as tools for further elucidation of the structure and function of opioid receptor subtypes and as drugs for their manipulation. One example briefly mentioned above is the observation that a drug with a mu agonist and delta antagonist or delta agonist component may constitute a clinically useful strong analgesic drug with an attenuated or eliminated propensity to produce tolerance and dependence. However, a major unanswered question is the optimum degree of mu agonism and either delta agonism or antagonism in a single compound for the ideal strong analgesic. We are presently pursuing this question in primate studies in collaboration with Steve Negus at the Medical College of Virginia who is utilizing the delta agonist SNC80 and related compounds we introduced earlier in combination with fentanyl and other strong mu agonists. We have also identified systemically active drugs in the unbridged phenylmorphan series that show mu agonism and delta antagonist activity in the same compound. We are presently extending this work in (a) oxide-bridged and (b) nonbridged 5-phenylmorphans, (c) other appropriate structures. We will also continue to extend the structure activity relations (SAR) in the opioid (+)-enantiomers related to (+)-naloxone and characterize the effects of functional antagonism of TLR-4 receptors. Collaborative computer assisted molecular modeling and ab initio quantum chemical methods will be employed in the design of these compounds. These novel drugs will be studied in the appropriate in vitro binding assays in native and cloned systems, smooth muscle assays, in vivo assays in small animals, and self-administration and other studies in rhesus monkeys. Hallucinogenic Drugs The abuse of phenethylamine and tryptamine based hallucinogenic drugs continues to be a serious problem that is now rivaled by the appearance of numerous cannabinoid agonists and cathinones (bath salts). The problem has been exacerbated by the publication of cookbook chemical syntheses and detailed accounts of the doses used and hallucinogenic effects seen in humans for almost all of the 179 phenethylamine and 53 tryptamine analogs that were synthesized and self-administered by A. T. Shulgin and his associates during more than 30 years. These are disturbing developments that substantially aid and encourage clandestine drug production and may presage a resurgence in hallucinogenic drug abuse. We have begun a program to synthesize and evaluate a number of these hallucinogenic agents and their antagonists. MDMA (racemic 3,4-methylenedioxymethamphetamine) also known as Ecstasy is a widely abused street drug. We studied the effects of racemic and R-(-)-MDMA on plasma prolactin secretion by plasma analysis and extracellular serotonin in the monkey striatum by in vivo microdialysis. Pretreatment with fluoxetine attenuated serotonin release by racemic or R-(-)-MDMA. Neither fluoxetine nor MDL100907 alone suppressed prolactin secretion by (-)-MDMA. However, coadministration of both of these drugs attenuated prolactin release suggesting that both serotonin release and 5HT2A agonism is required for prolactin secretion. Our results provide insight into the complex pharmacology of racemic MDMA.

258
Sigma Receptors and Binge Eating Sigma receptors are high affinity binding sites widely distributed in the mammalian central nervous system and periphery. Two subtypes, sigma-1 and sigma-2, have been characterized and shown to be involved in numerous physiologic functions and in some aspects of drug abuse. Earlier, we introduced BD-1063 as a selective sigma-1 antagonist that attenuates drug reinforcement in some assays. Binge eating is a disorder that resembles drug self-administration as a compulsive behavior regardless of predictable negative consequences. We examined the question of whether BD1063 can block binge eating in a rat model. BD1063 reduced food-seeking behavior, the quantity of food consumed and the rate of eating vs. a control group. Furthermore, sigma-1 mRNA was altered in several brain regions in the binge group compared to the control group. Our findings indicate the involvement of the sigma-1 receptor in binge eating in the rat and in related neurobiological adaptations. Sigma Receptors and Alcohol Self-Administration Sigma receptors have been implicated in the behavioral and motivational effects of alcohol and psychostimulants. Sigma receptor antagonists reduce the reinforcing effects of alcohol and excessive alcohol intake in both genetic (alcohol-preferring rats) and environmental (chronic alcohol-induced) models of alcoholism. We tested the hypothesis that pharmacological activation of sigma-receptors facilitates ethanol reinforcement and induces excessive, binge-like ethanol intake. The effects of repeated subcutaneous treatment with the selective sigma-receptor agonist 1,3-di-(2-tolyl)guanidine (DTG; 15 mg/kg, twice a day for 7 days) on operant ethanol (10%) self-administration were studied in Sardinian alcohol-preferring (sP) rats. To confirm that the effect of DTG was mediated by sigma-receptors, the effects of pretreatment with the selective sigma-receptor antagonist BD-1063 (7mg/kg, subcutaneously) were determined. To assess the specificity of action, the effects of DTG on the self-administration of equally reinforcing solutions of saccharin or sucrose were also determined. Finally, gene expression of opioid receptors in brain areas implicated in ethanol reinforcement was analyzed in ethanol-naive sP rats treated acutely or repeatedly with DTG, because of the well-established role of the opioid system in alcohol reinforcement and addiction. Repeatedly administered DTG progressively and dramatically increased ethanol self-administration in sP rats and increased blood alcohol levels, which reached mean values close to 100 mg% in 1h drinking sessions. Repeated DTG treatment also increased the rats' motivation to work for alcohol under a progressive-ratio schedule of reinforcement. BD-1063 prevented the effects of DTG, confirming that sigma-receptors mediate the effects of DTG. Repeated DTG treatment also increased the self-administration of the non-drug reinforcers saccharin and sucrose. Naive sP rats repeatedly treated with DTG showed increased mRNA expression of mu and delta-opioid receptors in the ventral tegmental area. These results suggest a key facilitatory role for sigma-receptors in the reinforcing effects of alcohol and identify a potential mechanism that contributes to binge-like and excessive drinking. Psychomotor Stimulants The abuse of psychomotor stimulants such as cocaine and methamphetamine is a continuing and very serious problem worldwide. In our program to synthesize and evaluate a number of 5-HT agonists and their antagonists, we previously developed a practical nonchromatographic chemical synthesis of the 5-HT2A receptor antagonist MDL100,907 that is providing multigram

259
amounts of this critical research tool. We have also studied the discriminative stimulus effects of MDL100,907 and several other drugs in order to gain further insight into their 5-HT receptor subtype(s) selectivity and the possible receptor role in certain neuropsychiatric disorders. Relapse to cocaine self-administration is a major obstacle to overcome in the successful treatment of human cocaine addicts. Such relapse can be triggered by impulsivity or cue reactivity. It is well established that the dopamine and serotonin (5-HT) receptor systems are involved in the regulation of impulsive behavior in animals and humans and prior studies have implicated the 5HT2A receptor subtype. We used the 5HT2A receptor antagonist MDL100907 to examine the question of whether this drug would suppress cocaine-induced impulsivity in two established rat models of impulsive behavior. MDL100907 attenuated impulsivity in the differential reinforcement of low rate (DRL) task and the one-choice serial reaction time (1-CSRT) task. Our results suggest that 5-HT2A receptor antagonists will be useful in further study of cocaine-induced impulsivity and may be therapeutically useful in the treatment of cocaine abuse and other impulse-control disorders. Preclinical studies have now shown that impulsivity and cocaine seeking behavior can be suppressed by relatively large doses of either a selective 5-HT2A antagonist or a selective 5HT2C agonist. We evaluated the possibility that low doses (to avoid side effects) of both a 5-HT2A antagonist MDL100907 and a 5HT2C agonist WAY163909 can be effective and offer advantages over single drug treatment in preventing relapse. Our results showed that low doses of both drugs in combination were effective when the same doses separately had no effect. The drug mixture suppressed cocaine-induced inherent and cocaine-evoked impulsive action, as well as cue- and cocaine-primed reinstatement of cocaine-seeking behavior. This is an important finding that suggests that a single drug showing both profiles in the optimum ratio could constitute a valuable medication for the prevention of cocaine relapse. TLR-4 receptors in the study of Alcohol Abuse Emerging evidence implicates a role for toll-like receptor 4 (TLR4) in the CNS effects of alcohol. We examined the question of whether TLR4-MyD88-dependent signalling is involved in the acute behavioural actions of alcohol and if alcohol can activate TLR4-downstream MAPK and NF-κB pathways. The TLR4 pathway was evaluated using the TLR4 antagonist (+)-naloxone (opioid receptor-inactive isomer) and mice with null mutations in the TLR4 and MyD88 genes. Sedation and motor impairment induced by a single dose of alcohol were assessed by loss of righting reflex (LORR) and rotarod tests, separately. The phosphorylation of JNK, ERK and p38, and levels of IκBα were measured to determine the effects of acute alcohol exposure on MAPK and NF-κB signaling. After a single dose of alcohol, both pharmacological inhibition of TLR4 signalling with (+)-naloxone and genetic deficiency of TLR4 or MyD88 significantly (P < 0.0001) reduced the duration of LORR by 45-78% and significantly decreased motor impairment recovery time to 62-88% of controls. These behavioural actions were not due to changes in the peripheral or central alcohol pharmacokinetics. IκBα levels responded to alcohol by 30 min in mixed hippocampal cell samples, from wild-type mice, but not in cells from TLR4- or MyD88-deficient mice. Our data provide new evidence that TLR4-MyD88 signalling is involved in the acute behavioral actions of alcohol in mice.
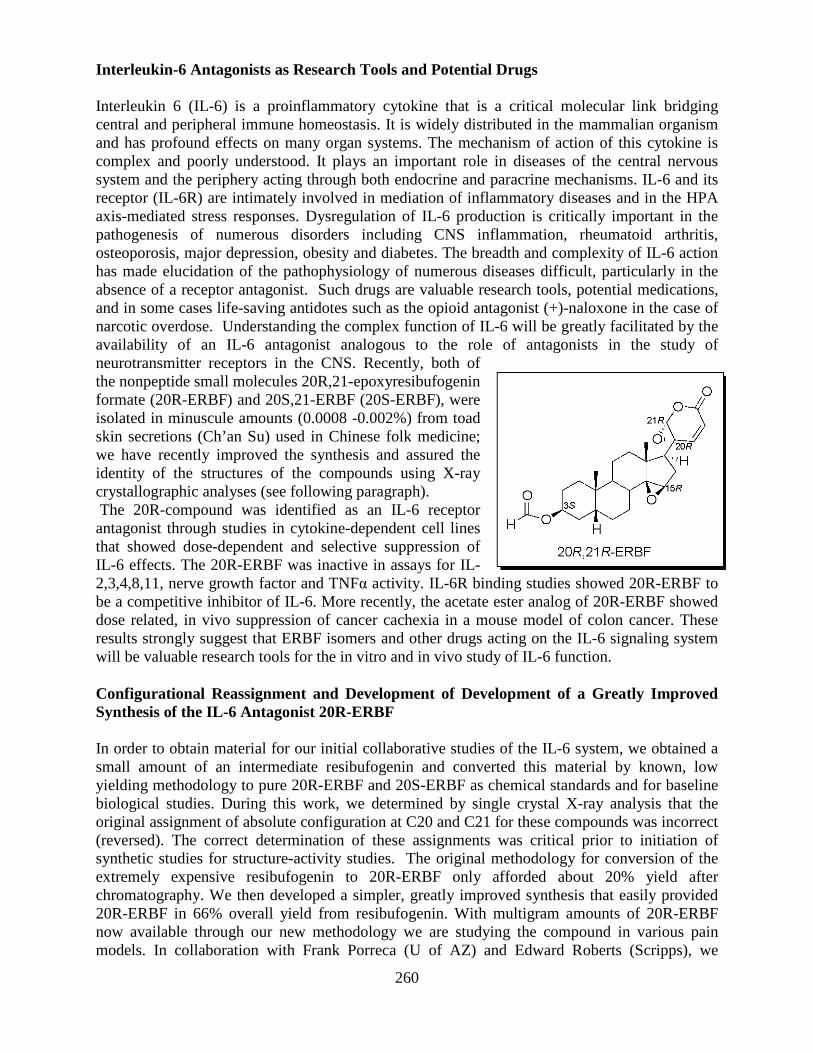
260
Interleukin-6 Antagonists as Research Tools and Potential Drugs Interleukin 6 (IL-6) is a proinflammatory cytokine that is a critical molecular link bridging central and peripheral immune homeostasis. It is widely distributed in the mammalian organism and has profound effects on many organ systems. The mechanism of action of this cytokine is complex and poorly understood. It plays an important role in diseases of the central nervous system and the periphery acting through both endocrine and paracrine mechanisms. IL-6 and its receptor (IL-6R) are intimately involved in mediation of inflammatory diseases and in the HPA axis-mediated stress responses. Dysregulation of IL-6 production is critically important in the pathogenesis of numerous disorders including CNS inflammation, rheumatoid arthritis, osteoporosis, major depression, obesity and diabetes. The breadth and complexity of IL-6 action has made elucidation of the pathophysiology of numerous diseases difficult, particularly in the absence of a receptor antagonist. Such drugs are valuable research tools, potential medications, and in some cases life-saving antidotes such as the opioid antagonist (+)-naloxone in the case of narcotic overdose. Understanding the complex function of IL-6 will be greatly facilitated by the availability of an IL-6 antagonist analogous to the role of antagonists in the study of neurotransmitter receptors in the CNS. Recently, both of the nonpeptide small molecules 20R,21-epoxyresibufogenin formate (20R-ERBF) and 20S,21-ERBF (20S-ERBF), were isolated in minuscule amounts (0.0008 -0.002%) from toad skin secretions (Ch’an Su) used in Chinese folk medicine; we have recently improved the synthesis and assured the identity of the structures of the compounds using X-ray crystallographic analyses (see following paragraph). The 20R-compound was identified as an IL-6 receptor antagonist through studies in cytokine-dependent cell lines that showed dose-dependent and selective suppression of IL-6 effects. The 20R-ERBF was inactive in assays for IL-2,3,4,8,11, nerve growth factor and TNFα activity. IL-6R binding studies showed 20R-ERBF to be a competitive inhibitor of IL-6. More recently, the acetate ester analog of 20R-ERBF showed dose related, in vivo suppression of cancer cachexia in a mouse model of colon cancer. These results strongly suggest that ERBF isomers and other drugs acting on the IL-6 signaling system will be valuable research tools for the in vitro and in vivo study of IL-6 function. Configurational Reassignment and Development of Development of a Greatly Improved Synthesis of the IL-6 Antagonist 20R-ERBF In order to obtain material for our initial collaborative studies of the IL-6 system, we obtained a small amount of an intermediate resibufogenin and converted this material by known, low yielding methodology to pure 20R-ERBF and 20S-ERBF as chemical standards and for baseline biological studies. During this work, we determined by single crystal X-ray analysis that the original assignment of absolute configuration at C20 and C21 for these compounds was incorrect (reversed). The correct determination of these assignments was critical prior to initiation of synthetic studies for structure-activity studies. The original methodology for conversion of the extremely expensive resibufogenin to 20R-ERBF only afforded about 20% yield after chromatography. We then developed a simpler, greatly improved synthesis that easily provided 20R-ERBF in 66% overall yield from resibufogenin. With multigram amounts of 20R-ERBF now available through our new methodology we are studying the compound in various pain models. In collaboration with Frank Porreca (U of AZ) and Edward Roberts (Scripps), we

261
recently found that 20R-ERBF (TB-2-081) is systemically available and orally effective in blocking chronic pain in animal models of bone cancer pain and pancreatitis by antagonizing IL-6. We characterized it as a reversible IL-6 receptor antagonist with femtomolar affinity that acts through peripheral mechanisms. In collaboration with David Clark and Wade Kingery (VA
Health Care, Palo Alto), 20R-ERBF was also shown to partially reverse post-fracture pain behaviors in wildtype mice in a tibia fracture model of complex regional pain syndrome. These results are highly significant since the only presently available human therapy for IL-6 related disorders is tocilizumab, an expensive (15-20K/year/patient), monoclonal antibody against the IL-6 receptor itself. Nevertheless, the clinical effectiveness of tocilizumab validates the mechanism of suppressing IL-6 function and the notion that a small molecule antagonist of the IL-6 should be a much more advantageous treatment for inflammatory pain than an antibody against the IL-6 receptor. In addition, it is well known that excessive levels of IL-6 play a role in the pathogenesis of a number of disorders including rheumatoid arthritis, Crohn's disease, multiple sclerosis, Castleman's disease, and certain types of leukemias and tumors, as well as depression, cardiomyopathy, and atherosclerotic disease, osteoporosis, diabetic neuropathy. A clinically acceptable, small molecule IL-6 antagonist should also be useful in treating these disorders and offer great advantages over tocilizumab therapy. Such treatment could offer distinct advantages to the use of narcotics that produce dependence, tolerance and in some cases life-threatening constipation. Combination of the IL-6 antagonist with narcotic therapy for severe pain should lower the dose of narcotic required and reduce narcotic-mediated side effects. IL-6 Summary We have identified 20R-ERBF as an orally effective IL-6 receptor antagonist that blocks chronic pain in three animal models and provides an important lead to novel nonnarcotic agents for the treatment of pain. This compound or a derivative may also offer great advantages over an expensive biological treatment to block the actions of IL-6 and provide a novel therapeutic for a number of disease states.

262
2011-2012 refereed publications 1. Koffarnus MN, Newman AH, Grundt P, Rice KC, Woods JH. 2011. Assessing Animal Models of
Impulsive Behavior: Effects of Selective Dopaminergic Compounds on a Delay Discounting Task. Behav Pharmacol 22:300-11.
2. Jagoda EM, Lang L, McCullough K, Contoreggi C, Kim BM, et al. 2011. [76Br]BMK-152, A
Non-Peptide Analogue, With High Affinity and Low Non-Specific Binding for the Corticotropin-Releasing Factor Type 1 Receptor. Synapse 65:910-18.
3. Jutkiewicz EM, Brooks EA, Kynaston AD, Rice KC, Woods JH. 2011. Patterns of Nicotinic
Receptor Antagonism: Nicotine Discrimination Studies. J Pharmacol Exp Ther 339:194-202.
4. Sabino V, Cottone P, Blasio A, Iyer MR, Steardo L, et al. 2011. Activation of Sigma Receptors Induces Binge-like Drinking in Sardinian Alcohol-Preferring Rats. Neuropsychopharmacology 36:1207-18.
5. Banks ML, Roma PG, Folk JE, Rice KC, Negus SS. 2011. Effects of The Delta-Opioid Agonist
SNC80 on the Abuse Liability of Methadone in Rhesus Monkeys: A Behavioral Economic Analysis. Psychopharmacology 216:431-39.
6. Anastasio NC, Stoffel EC, Fox RG, Bubar MJ, Rice KC, et al. 2011. The Serotonin (5-HT) 5-
HT2a Receptor: Association with Inherent and Cocaine-Evoked Behavioral Disinhibition in Rats. Behav Pharmacol 22:248-61.
7. Li J-X, Koek W, Rice KC, France CP. 2011. Effects of Direct- and Indirect-Acting Serotonin
Receptor Agonists on The Antinociceptive And Discriminative Stimulus Effects of Morphine in Rhesus Monkeys. Neuropsychopharmacology 36:940-49.
8. Cheng K, Lee YS, Rothman RB, Dersch CM, Bittman RW, et al. 2011. Probes for Narcotic
Receptor Mediated Phenomena. 41. Unusual Inverse μ-Agonists and Potent μ-Opioid Antagonists by Modification of the N-Substituent in Enantiomeric 5-(3-Hydroxyphenyl)morphans. J Med Chem 54:957-69.
9. Li J-X, Crocker C, Koek W, Rice KC, France CP. 2011. Effects of Serotonin (5-HT)1A and 5-
HT2A Receptor Agonists on Schedule-Controlled Responding in Rats: Drug Combination Studies. Psychopharmacology 213:489-97. In Special Issue: "Serotonin Revisited".
10. Schank JR, Pickens CL, Rowe KE, Cheng K, Thorsell A, et al. 2011. Stress-Induced
Reinstatement of Alcohol Seeking in Rats is Selectively Suppressed by the Neurokinin 1(NK1) Antagonist L822429. Psychopharmacology 218:111-19.
11. Xiong W, Cheng K, Cui T, Godlewski G, Rice KC, et al. 2011. Cannabinoid Potentiation of
Glycine Receptors Contributes to Cannabis-Induced Analgesia. Nat Chem Biol 7:296-303. See Commentary p. 249-250.
12. Kim J-H, Deschamps JR, Rothman RB, Dersch CM, Folk JE, et al. 2011. Probes For Narcotic
Receptor Mediated Phenomena. 42. Synthesis and In Vitro Pharmacological Characterization of the N-Methyl and N-Phenethyl Analogues of Rac-(3R,6aS,11aS)-1,3,4,5,6,11a-Hexahydro-2H-

263
3,6a-methanobenzofuro[2,3-c]azocin-10-ol and -8-ol, The ortho- and para-C Oxide-Bridged Phenylmorphans. Bioorg Med Chem 19:3434-43.
13. Li F, Folk JE, Cheng K, Kurimura M, Deck JA, et al. 2011. Probes for Narcotic Receptor
Mediated Phenomena. 43. Synthesis of The ortho-A and para-A, and Improved Synthesis and Optical Resolution of the ortho-B and para–B Oxide-Bridged Phenylmorphans: Compounds with Moderate to Low Opioid-Receptor Affinity. Bioorg Med Chem 19:4330-37.
14. Winter JC, Amorosi DJ, Rice KC, Cheng K, Yu A-M. 2011. Stimulus Control by 5-Methoxy-
N,N-Dimethyltryptamine in Wild-Type and CYP2D6-Humanized Mice. Pharmacol Biochem Behav 99:311-15.
15. Lim HJ, Deschamps JR, Jacobson AE, Rice KC. 2011. Diastereoselective One-Pot Synthesis of
7- and 8-Substituted 5-Phenylmorphans. Org Lett 13:5322-25.
16. Hensler JG, Advani T, Burke TF, Cheng K, Rice KC, Koek W. 2012. GABAB Receptor-Positive Modulators: Brain Region-Dependent Effects. J Pharmacol Exp Ther 340:19-26.
17. Wu Y, Lousberg EL, Moldenhauer LM, Hayball JD, Robertson SA, et al. 2012. Inhibiting the
TLR4-Myd88 Signalling Cascade by Genetic or Pharmacological Strategies Reduces Acute Alcohol-Induced Sedation and Motor Impairment in Mice. Br J Pharmacol 165:1319-29.
18. Negus SS, O'Connell R, Morrissey E, Cheng K, Rice KC. 2012. Effects of Peripherally Restricted
Kappa Opioid Receptor Agonists on Pain-Stimulated and Pain-Depressed Behavior in Rats. J Pharmacol Exp Ther 340:501-09.
19. Kimura T, Yeliseev AA, Vukoti K, Rhodes SD, Cheng K, et al. 2012. Recombinant Cannabinoid
Type 2 Receptor in Liposome Model Activates G Protein in Response to Anionic Lipid Constituents. J Biol Chem 287:4076-87.
20. Murname KS, Kimmel HL, Rice KC, Howell LL. 2012. The Neuropharmacology Of Prolactin
Secretion Elicited By 3,4-Methylenedioxymethamphetamine ("Ecstasy"): A Concurrent Microdialysis And Plasma Analysis Study. Horm Behav 61:181-90.
21. Iyer MR, Lee YS, Deschamps JR, Dersch CM, Rothman RB, et al. 2012. Probes For Narcotic
Receptor Mediated Phenomena. 44. Synthesis of an N-Substituted 4-Hydroxy-5-(3-Hydroxyphenyl)morphan with High Affinity And Selective Mu-Antagonist Activity. Eur J Med Chem 50:44-54.
22. Sawyer EK, Mun J, Nye JA, Kimmel HL, Rice KC, et al. 2012. Neurobiological Changes
Mediating The Effects Of Chronic Fluoxetine On Cocaine Use. Neuropsychopharmacology 37:1816-24.
23. Lewis SS, Loram LC, Hutchinson MR, Zhang Y, Maier SF, et al. 2012. (+)-Naloxone, an Opioid-
Inactive Toll-Like Receptor 4 Inhibitor, Reverses Multiple Models Of Chronic Neuropathic Pain In Rats. J Pain 13:498-506.
24. Rissman RR, Staup MA, Lee AR, Justice NJ, Rice KC, et al. 2012. Corticotropin-Releasing
Factor Receptor-Dependent Effects of Repeated Stress on Tau Phosphorylation, Solubility and Aggregation. Proc Natl Acad Sci USA 109:6277-82.

264
25. Negus SS, Marisa B, Rosenberg BS, Altarifi AA, O'Connell RH, et al. 2012. Effects of the Delta Opioid Receptor Agonist SNC80 on Pain-Related Depression of Intracranial Self-Stimulation (ICSS) in Rats. J Pain 13:317-27.
26. Guo T-Z, Wei T, Shi X, Li W-W, Hou S, et al. 2012. Neuropeptide Deficient Mice Have
Attenuated Nociceptive, Vascular, and Inflammatory Changes in a Tibia Fracture Model of Complex Regional Pain Syndrome. Mol Pain 8:85-99.
27. Yekkirala AS, Banks, ML, Lunzer MM, Negus SS, Rice KC, Portoghese PS. 2012. Clinically
Employed Opioid Analgesics Produce Antinociception via µ-δ Opioid Receptor Heteromers In Rhesus Monkeys. ACS Chem Neurosci 3:720-27.
28. Hutchinson MR, Northcutt AL, Hiranita T, Wang X, Lewis S, et al. 2012. Opioid Activation of
Toll-like Receptor 4 Contributes to Drug Reinforcement. J Neurosci 32:11187-200.
29. Cottone P, Wang X, Park JW, Valenza M, Blasio A, et al. 2012. Antagonism Of Sigma-1 Receptors Blocks Compulsive-like Eating. Neuropsychopharmacology 37:2593-604.
30. Boos TL, Cheng K, Greiner E, Deschamps JR, Jacobson AE, Rice KC. 2012. Configurational
Reassignment and Improved Preparation of the Competitive IL-6 Receptor Antagonist 20R,21R-Epoxyresibufogenin-3-formate. J Nat Prod 75:661-68.
31. Koek W, France CP, Cheng K, Rice KC. 2012. Effects of the GABAB receptor-positive
modulators CGP7930 and rac-BHFF in baclofen- and in GHB-discriminating pigeons. J Pharmacol Exp Ther 341:369-76.
32. Negus SS, Morrissey EM, Folk JE, Rice, KC. 2012. Interaction Between Mu and Delta Opioid
Receptor Agonists in an Assay of Capsaicin-Induced Thermal Allodynia in Rhesus Monkeys. Pain Res Treat epub May 15, doi 10.1155/2012/867067.
33. Koffarnus MN, Collins GT, Rice KC, Chen J, Woods JH, Winger G. 2012. Self-Administration
of Agonists Selective for Dopamine D2, D3, and D4 Receptors by Rhesus Monkeys. Behav Pharmacol 23:331-38.
34. Serafine KM, Briscione MA, Rice KC, Riley AL. 2012. Dopamine Mediates Cocaine-induced
Conditioned Taste Aversions as Demonstrated with Cross-drug Pre-exposure to GBR 12909. Pharmacol Biochem Behav 102:269-74.
35. Sabino V, Kwak J, Rice KC, Cottone P. 2012. Pharmacological Characterization of the 20%
Alcohol Intermittent Access Model In Sardinian Alcohol-Preferring Rats: A Model Of Binge-Like Drinking. Alcohol Clin Exp Res epub Nov 5, doi: 10.1111/acer.12008.
36. Sulima A, Cheng K, Jacobson AE, Rice KC, Gawrisch K, Lee YS. 2012. Z and E Rotamers of N-
formyl-1-bromo-4-hydroxy-3-methoxymorphinan-6-one and their Interconversion as Studied by 1H/13C NMR Spectroscopy and Quantum Chemical Calculations. Magn Reson Chem epub Dec 11, doi: 10.1002/mrc.3909.
37. Cunningham KA, Anastasio NC, Fox RG, Stutz SJ, Bubar MJ, et al. 2012. Synergism Between a
Serotonin 5-HT2A Receptor (5-HT2AR) Antagonist and 5-HT2CR Agonist Suggests New Pharmacotherapeutics for Cocaine Addiction. ACS Chem Neurosci epub Aug 11, doi: 10.1021/cn300072u.

265
38. Barbier E, Vendruscolo LF, Schlosburg JE, Edwards S, Juergens N, et al. 2012. The NK1 Receptor Antagonist L822429 Reduces Heroin Reinforcement. Neuropsychopharmacology epub Dec18, doi: 10.1038/npp.2012.261.
39. Parylak SL, Cottone P, Sabino V, Rice KC, Zorrilla EP. 2012. Effects of CB1 and CRF1
Receptor Antagonists on Binge-Like Eating in Rats With Limited Access to a Sweet Fat Diet: Lack of Withdrawal-Like Responses. Physiol Behav 107(2):231-42.
40. Tam J, Cinar R, Liu J, Godlewski G, Wesley D, et al. 2012. Peripheral Cannabinoid-1 (CB1)
Receptor Inverse Agonism Reduces Obesity by Reversing Leptin Resistance. Cell Metab 16:167-79.
41. Li J-X, Shah AP, Patel SK, Rice KC, France CP. 2012. Modification of the Behavioral Effects of
Morphine in Rats by Serotonin (5-HT)1A and 5-HT2 Receptor Agonists: Antinociception, Drug Discrimination, and Locomotor Activity: Antinociception, Drug Discrimination, and Locomotor Activity. Psychopharmacology epub Sep 20, doi: 10.1007/s00213-012-2870-2.
42. Li F, Deck JA, Dersch CM, Rothman RB, Deschamps JR, et al. 2012. Probes for Narcotic
Receptor Mediated Phenomena. 46. N-Substituted-2,3,4,9,10,10a-Hexahydro-1H-1,4a-(Epiminoethano)Phenanthren-6- And 8-ols - Carbocyclic Relatives of f-Oxide Bridged Phenylmorphans. Eur J Med Chem 58:557-67.
43. Koek W, Cheng K, Rice KC. 2012. Discriminative Stimulus Effects of The GABAB Receptor-
Positive Modulator rac-BHFF: Comparison with GABAB Receptor Agonists and Drugs of Abuse. J Pharmacol Exp Ther epub, Dec 28.
44. Fantegrossi WE, Gannon BM, Zimmerman SM, Rice KC. 2012. In Vivo Effects of Abused “Bath
Salt” Constituent 3,4-Methylenedioxypyrovalerone (MDPV) in Mice: Drug Discrimination, Thermoregulation, and Locomotor Activity. Neuropsychopharmacology epub Nov 8, doi: 10.1038/npp.2012.233.
45. Xiong W, Wu X, Lovinger DM, Zhang L. on pages 5200–5208 of the April 11, 2012 issue, A
Common Molecular Basis for Exogenous and Endogenous Cannabinoid Potentiation of Glycine Receptors. Xiong W, Wu X, Li F, Cheng K, Rice KC, Lovinger DM, Zhang L. J. Neurosci. 2012, 32, 12979. Corrected author list.
Plenary lecture
International Narcotic Research Conference, Kansas City, MO., July 2012. Plenary Lecture. Topic: “Medicinal Chemistry in Drug Abuse Research at the NIH - Looking Back”.
Provisional patent application
Kunos G, Iyer MR, Resat C, Rice KC. 2012. Novel Peripherally Restricted CB1 Receptor Inverse Agonists for the Treatment of Diabetes and Obesity. U.S. Provisional Patent Application filed Nov 13.
Review Article
Hutchinson MR, Shavit Y, Grace PM, Rice KC, Maier SF, Watkins LR. 2011 Exploring the Neuroimmunopharmacology of Opioids: An Integrative Review of Mechanisms of Central Immune Signaling and Their Implications for Opioid Analgesia. Pharmacol Rev 63:772-810.

266
Drug Design and Synthesis Section, NIDA and NIAAA, March 13, 2013 L to R seated: Arthur Jacobson, Mariena Mattson, Kenner Rice. L to R Standing: Masaki Suzuki, Fuying Li, Peter DeMatteo, Mohd Kabir, Josh Antoline, Malliga Iyer, Brandon Selfridge, Kejun Cheng Dr. Rice: [email protected]

267
DICBR PUBLICATIONS 2011
1. Abdelmegeed MA, Yoo SH, Henderson LE, Gonzalez FJ, Woodcroft KJ, Song BJ. 2011.
PPAR alpha Expression Protects Male Mice from High Fat-Induced Nonalcoholic Fatty Liver. Journal of Nutrition 141: 603-10
2. Abraham J, Balbo S, Crabb D, Brooks PJ. 2011. Alcohol Metabolism in Human Cells Causes DNA Damage and Activates the Fanconi Anemia-Breast Cancer Susceptibility (FA-BRCA) DNA Damage Response Network. Alcoholism-Clinical and Experimental Research 35: 2113-20
3. Abraham J, Brooks PJ. 2011. Divergent Effects of Oxidatively Induced Modification to the C8 of 2 '-Deoxyadenosine on Transcription Factor Binding: 8,5 '(S)-Cyclo-2 '-deoxyadenosine Inhibits the Binding of Multiple Sequence Specific Transcription Factors, While 8-Oxo-2 '-deoxyadenosine Increases Binding of CREB and NF-kappa B to DNA. Environmental and Molecular Mutagenesis 52: 287-95
4. Baca-Garcia E, Perez-Rodriguez MM, Keyes KM, Oquendo MA, Hasin DS, Grant BF, Blanco C. 2011. Suicidal ideation and suicide attempts among Hispanic subgroups in the United States: 1991-1992 and 2001-2002. Journal of Psychiatric Research 45: 512-8
5. Baca-Garcia E, Perez-Rodriguez MM, Oquendo MA, Keyes KM, Hasin DS, Grant BF, Blanco C. 2011. Estimating risk for suicide attempt: Are we asking the right questions? Passive suicidal ideation as a marker for suicidal behavior. Journal of Affective Disorders 134: 327-32
6. Barnett JH, Xu K, Heron J, Goldman D, Jones PB. 2011. Cognitive Effects of Genetic Variation in Monoamine Neurotransmitter Systems: A Population-Based Study of COMT, MAOA, and 5HTTLPR. American Journal of Medical Genetics Part B-Neuropsychiatric Genetics 156B: 158-67
7. Bello EP, Mateo Y, Gelman DM, Noain D, Shin JH, Low MJ, Alvarez VA, Lovinger DM, Rubinstein M. 2011. Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D-2 autoreceptors. Nature Neuroscience 14: 1033-U128
8. Bevilacqua L, Goldman D. 2011. Genetics of emotion. Trends in Cognitive Sciences 15: 401-8
9. Blasbalg TL, Hibbeln JR. 2011. Waste and hydrogenation lower omega-3 and omega-6 fat intake estimates for soybean oil Reply. American Journal of Clinical Nutrition 94: 1153-5
10. Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. 2011. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. American Journal of Clinical Nutrition 93: 950-62
11. Bletzer KV, Yuan NP, Koss MP, Polacca M, Eaves ER, Goldman D. 2011. Taking Humor Seriously: Talking about Drinking in Native American Focus Groups. Medical Anthropology 30: 295-318
12. Breese GR, Sinha R, Heilig M. 2011. Chronic alcohol neuroadaptation and stress contribute to susceptibility for alcohol craving and relapse. Pharmacology & Therapeutics 129: 149-71
13. Buydens-Branchey L, Branchey M, Hibbeln JR. 2011. Higher n-3 fatty acids are associated with more intense fenfluramine-induced ACTH and cortisol responses among cocaine-abusing men. Psychiatry Research 188: 422-7
14. Camp MC, Feyder M, Ihne J, Palachick B, Hurd B, Karlsson RM, Noronha B, Chen YC, Coba MP, Grant SGN, Holmes A. 2011. A novel role for PSD-95 in mediating ethanol intoxication, drinking and place preference. Addiction Biology 16: 428-39

268
15. Carli V, Roy A, Bevilacqua L, Maggi S, Cesaro C, Sarchiapone M. 2011. Insomnia and suicidal behaviour in prisoners. Psychiatry Research 185: 141-4
16. Carlson VCC, Seabold GK, Helms CM, Garg N, Odagiri M, Rau AR, Daunais J, Alvarez VA, Lovinger DM, Grant KA. 2011. Synaptic and Morphological Neuroadaptations in the Putamen Associated with Long-Term, Relapsing Alcohol Drinking in Primates. Neuropsychopharmacology 36: 2513-28
17. Chandra R, Federici S, Nemeth ZH, Horvath B, Pacher P, Hasko G, Deitch EA, Spolarics Z. 2011. Female X-Chromosome Mosaicism for NOX2 Deficiency Presents Unique Inflammatory Phenotype and Improves Outcome in Polymicrobial Sepsis. Journal of Immunology 186: 6465-73
18. Chen G, Carlson VCC, Wang J, Beck A, Heinz A, Ron D, Lovinger DM, Buck KJ. 2011. Striatal Involvement in Human Alcoholism and Alcohol Consumption, and Withdrawal in Animal Models. Alcoholism-Clinical and Experimental Research 35: 1739-48
19. Cheng KJ, Lee YS, Rothman RB, Dersch CM, Bittman RW, Jacobson AE, Rice KC. 2011. Probes for Narcotic Receptor Mediated Phenomena. 41. Unusual Inverse mu-Agonists and Potent mu-Opioid Antagonists by Modification of the N-Substituent in Enantiomeric 5-(3-Hydroxyphenyl)morphans. Journal of Medicinal Chemistry 54: 957-69
20. Cippitelli A, Rezvani AH, Robinson JE, Eisenberg L, Levin ED, Bonaventure P, Motley ST, Lovenberg TW, Heilig M, Thorsell A. 2011. The novel, selective, brain-penetrant neuropeptide Y Y2 receptor antagonist, JNJ-31020028, tested in animal models of alcohol consumption, relapse, and anxiety. Alcohol 45: 567-76
21. Cirulli F, Reif A, Herterich S, Lesch KP, Berry A, Francia N, Aloe L, Barr CS, Suomi SJ, Alleva E. 2011. A novel BDNF polymorphism affects plasma protein levels in interaction with early adversity in rhesus macaques. Psychoneuroendocrinology 36: 372-9
22. Copeland WE, Magnusson A, Goransson M, Heilig MA. 2011. Genetic moderators and psychiatric mediators of the link between sexual abuse and alcohol dependence. Drug and Alcohol Dependence 115: 183-9
23. Copeland WE, Sun H, Costello EJ, Angold A, Heilig MA, Barr CS. 2011. Child mu-Opioid Receptor Gene Variant Influences Parent-Child Relations. Neuropsychopharmacology 36: 1165-70
24. Davis MI, Puhl HL. 2011. Nr4a1-eGFP Is a Marker of Striosome-Matrix Architecture, Development and Activity in the Extended Striatum. Plos One 6
25. Dawson DA. 2011. Risky Single-Occasion Drinking. Addiction 106: 1709-10 26. Dawson DA. 2011. The Challenge of Distinguishing High-Volume and Frequent Risky
Drinking. Addiction 106: 1046-7 27. Dawson DA. 2011. Defining Risk Drinking. Alcohol Research & Health 34: 144-56 28. Dawson DA, Grant BF. 2011. The "Gray Area" of Consumption Between Moderate and
Risk Drinking. Journal of Studies on Alcohol and Drugs 72: 453-8 29. Diep CQ, Ma DD, Deo RC, Holm TM, Naylor RW, Arora N, Wingert RA, Bollig F,
Djordjevic G, Lichman B, Zhu H, Ikenaga T, Ono F, Englert C, Cowan CA, Hukriede NA, Handin RI, Davidson AJ. 2011. Identification of adult nephron progenitors capable of kidney regeneration in zebrafish. Nature 470: 95-U108
30. Dlugos AM, Hamidovic A, Hodgkinson C, Shen PH, Goldman D, Palmer AA, de Wit H. 2011. OPRM1 gene variants modulate amphetamine-induced euphoria in humans. Genes Brain and Behavior 10: 199-209
31. Dobi A, Seabold GK, Christensen CH, Bock R, Alvarez VA. 2011. Cocaine-Induced Plasticity in the Nucleus Accumbens Is Cell Specific and Develops without Prolonged Withdrawal. Journal of Neuroscience 31: 1895-904

269
32. Drel VR, Pacher P, Stavniichuk R, Xu WZ, Zhang J, Kuchmerovska TM, Slusher B, Obrosova IG. 2011. Poly(ADP-ribose)polymerase inhibition counteracts renal hypertrophy and multiple manifestations of peripheral neuropathy in diabetic Akita mice. International Journal of Molecular Medicine 28: 629-35
33. El-Remessy AB, Rajesh M, Mukhopadhyay P, Horvath B, Patel V, Al-Gayyar MMH, Pillai BA, Pacher P. 2011. Cannabinoid 1 receptor activation contributes to vascular inflammation and cell death in a mouse model of diabetic retinopathy and a human retinal cell line. Diabetologia 54: 1567-78
34. Enoch MA. 2011. The role of early life stress as a predictor for alcohol and drug dependence. Psychopharmacology 214: 17-31
35. Enoch MA, Gorodetsky E, Hodgkinson C, Roy A, Goldman D. 2011. Functional genetic variants that increase synaptic serotonin and 5-HT3 receptor sensitivity predict alcohol and drug dependence. Molecular Psychiatry 16: 1139-46
36. Fitzgerald PJ. 2011. A neurochemical yin and yang: does serotonin activate and norepinephrine deactivate the prefrontal cortex? Psychopharmacology 213: 171-82
37. Freeman MP, Hibbeln JR, Silver M, Hirschberg AM, Wang B, Yule AM, Petrillo LF, Pascuillo E, Economou NI, Joffe H, Cohen LS. 2011. Omega-3 fatty acids for major depressive disorder associated with the menopausal transition: a preliminary open trial. Menopause-the Journal of the North American Menopause Society 18: 279-84
38. Gao B, Bataller R. 2011. Alcoholic Liver Disease: Pathogenesis and New Therapeutic Targets. Gastroenterology 141: 1572-85
39. Gao B, Bertola A. 2011. Natural Killer Cells Take Two Tolls to Destruct Bile Ducts. Hepatology 53: 1076-9
40. Gao B, Seki E, Brenner DA, Friedman S, Cohen JI, Nagy L, Szabo G, Zakhari S. 2011. Innate immunity in alcoholic liver disease. American Journal of Physiology-Gastrointestinal and Liver Physiology 300: G516-G25
41. Gao RY, Mukhopadhyay P, Mohanraj R, Wang H, Horvath B, Yin S, Pacher P. 2011. Resveratrol attenuates azidothymidine-induced cardiotoxicity by decreasing mitochondrial reactive oxygen species generation in human cardiomyocytes. Molecular Medicine Reports 4: 151-5
42. George DT, Phillips MJ, Lifshitz M, Lionetti TA, Spero DE, Ghassemzedeh N, Doty L, Umhau JC, Rawlings RR. 2011. Fluoxetine Treatment of Alcoholic Perpetrators of Domestic Violence: A 12-Week, Double-Blind, Randomized, Placebo-Controlled Intervention Study. Journal of Clinical Psychiatry 72: 60-5
43. Goldman D. 2011. Molecular Etiologies of Schizophrenia: Are We Almost There Yet? American Journal of Psychiatry 168: 879-81
44. Goldstein RZ, Volkow ND. 2011. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nature Reviews Neuroscience 12: 652-69
45. Goldstein RZ, Volkow ND. 2011. Oral Methylphenidate Normalizes Cingulate Activity and Decreases Impulsivity in Cocaine Addiction During an Emotionally Salient Cognitive Task. Neuropsychopharmacology 36: 366-7
46. Grant BF, Dawson DA, Moss HB. 2011. Disaggregating the Burden of Substance Dependence in the United States. Alcoholism-Clinical and Experimental Research 35: 387-8
47. Graybeal C, Feyder M, Schulman E, Saksida LM, Bussey TJ, Brigman JL, Holmes A. 2011. Paradoxical reversal learning enhancement by stress or prefrontal cortical damage: rescue with BDNF. Nature Neuroscience 14: 1507-9

270
48. Gremel CM, Young EA, Cunningham CL. 2011. Blockade of opioid receptors in anterior cingulate cortex disrupts ethanol-seeking behavior in mice. Behavioural Brain Research 219: 358-62
49. Hankin JA, Farias SE, Barkley R, Heidenreich K, Frey LC, Hamazaki K, Kim HY, Murphy RC. 2011. MALDI Mass Spectrometric Imaging of Lipids in Rat Brain Injury Models. Journal of the American Society for Mass Spectrometry 22: 1014-21
50. Hansson AC, Rimondini R, Heilig M, Mathe AA, Sommer WH. 2011. Dissociation of antidepressant-like activity of escitalopram and nortriptyline on behaviour and hippocampal BDNF expression in female rats. Journal of Psychopharmacology 25: 1378-87
51. Harper KN, Hibbeln JR, Deckelbaum R, Quesenberry CP, Schaefer CA, Brown AS. 2011. Maternal serum docosahexaenoic acid and schizophrenia spectrum disorders in adult offspring. Schizophrenia Research 128: 30-6
52. Hasin D, Fenton MC, Skodol A, Krueger R, Keyes K, Geier T, Greenstein E, Blanco C, Grant B. 2011. Personality Disorders and the 3-Year Course of Alcohol, Drug, and Nicotine Use Disorders. Archives of General Psychiatry 68: 1158-67
53. Hasko G, Csoka B, Koscso B, Chandra R, Pacher P, Thompson LF, Deitch EA, Spolarics Z, Virag L, Gergely P, Rolandelli RH, Nemeth ZH. 2011. Ecto-5 '-Nucleotidase (CD73) Decreases Mortality and Organ Injury in Sepsis. Journal of Immunology 187: 4256-67
54. Hasko G, Pacher P. 2011. Suppression of Tumorigenicity 2 A Janus-faced Player in Sepsis. American Journal of Respiratory and Critical Care Medicine 183: 841-3
55. Heilig M. 2011. A WELCOME CHANGE THAT STOPS SHORT OF BEING FULLY SATISFYING. Addiction 106: 874-5
56. Heilig M, Goldman D, Berrettini W, O'Brien CP. 2011. Pharmacogenetic approaches to the treatment of alcohol addiction. Nature Reviews Neuroscience 12: 670-84
57. Heilig M, Warren KR, Kunos G, Silverman PB, Hewitt BG. 2011. Addiction Research Centres and the Nurturing of Creativity. Addiction 106: 1052-60
58. Higham JP, Barr CS, Hoffman CL, Mandalaywala TM, Parker KJ, Maestripieri D. 2011. Mu-opioid Receptor (OPRM1) Variation, Oxytocin Levels and Maternal Attachment in Free-Ranging Rhesus Macaques Macaca mulatta. Behavioral Neuroscience 125: 131-6
59. Holmes A. 2011. Merger Fever: Can Two Separate Mechanisms Work Together to Explain Why We Drink? Biological Psychiatry 69: 1015-6
60. Hommer DW, Bjork JM, Gilman JM. 2011. Imaging brain response to reward in addictive disorders. In Addiction Reviews, ed. GR Uhl, pp. 50-61
61. Hong LE, Yang X, Wonodi I, Hodgkinson CA, Goldman D, Stine OC, Stein ES, Thaker GK. 2011. A CHRNA5 allele related to nicotine addiction and schizophrenia. Genes Brain and Behavior 10: 530-5
62. Horibata K, Saijo M, Bay MN, Lan L, Kuraoka I, Brooks PJ, Honma M, Nohmi T, Yasui A, Tanaka K. 2011. Mutant Cockayne syndrome group B protein inhibits repair of DNA topoisomerase I-DNA covalent complex. Genes to Cells 16: 101-14
63. Horiguchi N, Gao B. 2011. Inflammation Does Not Always Kill Hepatocytes During Liver Damage Reply. Hepatology 54: 367-
64. Huang BX, Akbar M, Kevala K, Kim HY. 2011. Phosphatidylserine is a critical modulator for Akt activation. Journal of Cell Biology 192: 979-92
65. Hutchinson MR, Shavit Y, Grace PM, Rice KC, Maier SF, Watkins LR. 2011. Exploring the Neuroimmunopharmacology of Opioids: An Integrative Review of Mechanisms of Central Immune Signaling and Their Implications for Opioid Analgesia. Pharmacological Reviews 63: 772-810

271
66. Ikenaga T, Urban JM, Gebhart N, Hatta K, Kawakami K, Ono F. 2011. Formation of the Spinal Network in Zebrafish Determined by Domain-Specific Pax Genes. Journal of Comparative Neurology 519: 1562-79
67. Jeong WI, Park O, Suh YG, Byun JS, Park SY, Choi E, Kim JK, Ko H, Wang H, Miller AM, Gao B. 2011. Suppression of Innate Immunity (Natural Killer Cell/Interferon-gamma) in the Advanced Stages of Liver Fibrosis in Mice. Hepatology 53: 1342-51
68. Jonassaint CR, Szatkiewicz JP, Bulik CM, Thornton LM, Bloss C, Berrettini WH, Kaye WH, Bergen AW, Magistretti P, Strober M, Keel PK, Brandt H, Crawford S, Crow S, Fichter MM, Goldman D, Halmi KA, Johnson C, Kaplan AS, Klump KL, La Via M, Mitchell JE, Rotondo A, Treasure J, Woodside DB. 2011. Absence of Association Between Specific Common Variants of the Obesity-Related FTO Gene and Psychological and Behavioral Eating Disorder Phenotypes. American Journal of Medical Genetics Part B-Neuropsychiatric Genetics 156B: 454-61
69. Jutkiewicz EM, Brooks EA, Kynaston AD, Rice KC, Woods JH. 2011. Patterns of Nicotinic Receptor Antagonism: Nicotine Discrimination Studies. Journal of Pharmacology and Experimental Therapeutics 339: 194-202
70. Kerridge BT, Saha TD, Smith S, Chou PS, Pickering RP, Huang B, Ruan JW, Pulay AJ. 2011. Dimensionality of hallucinogen and inhalant/solvent abuse and dependence criteria: Implications for the diagnostic and statistical manual of mental disorders-Fifth edition. Addictive Behaviors 36: 912-8
71. Keyes KM, Krueger RF, Grant BF, Hasin DS. 2011. Alcohol craving and the dimensionality of alcohol disorders. Psychological Medicine 41: 629-40
72. Kim HY, Bierbaum VM. 2011. Focus in Honor of Marvin L. Vestal, Recipient of the 2010 Award for a Distinguished Contribution in Mass Spectrometry. Journal of the American Society for Mass Spectrometry 22: 949-52
73. Kim HY, Moon HS, Cao DH, Lee J, Kevala K, Jun SB, Lovinger DM, Akbar M, Huang BX. 2011. N-Docosahexaenoylethanolamide promotes development of hippocampal neurons. Biochemical Journal 435: 327-36
74. Kim HY, Spector AA, Xiong ZM. 2011. A synaptogenic amide N-docosahexaenoylethanolamide promotes hippocampal development. Prostaglandins & Other Lipid Mediators 96: 114-20
75. Kim JH, Deschamps JR, Rothman RB, Dersch CM, Folk JE, Cheng KJ, Jacobson AE, Rice KC. 2011. Probes for narcotic receptor mediated phenomena. Part 42: Synthesis and in vitro pharmacological characterization of the N-methyl and N-phenethyl analogues of the racemic ortho-c and para-c oxide-bridged phenylmorphans. Bioorganic & Medicinal Chemistry 19: 3434-43
76. Kimura M, Higuchi S. 2011. Genetics of alcohol dependence. Psychiatry and Clinical Neurosciences 65: 213-25
77. Kiselycznyk C, Svenningsson P, Delpire E, Holmes A. 2011. Genetic, Pharmacological and Lesion Analyses Reveal a Selective Role for Corticohippocampal Glun2B in a Novel Repeated Swim Stress Paradigm. Neuroscience 193: 259-68
78. Koffarnus MN, Newman AH, Grundt P, Rice KC, Woods JH. 2011. Effects of selective dopaminergic compounds on a delay-discounting task. Behavioural Pharmacology 22: 300-11
79. Koscso B, Csoka B, Pacher P, Hasko G. 2011. Investigational A(3) adenosine receptor targeting agents. Expert Opinion on Investigational Drugs 20: 757-68
80. Kramer PF, Christensen CH, Hazelwood LA, Dobi A, Bock R, Sibley DR, Mateo Y, Alvarez VA. 2011. Dopamine D2 Receptor Overexpression Alters Behavior and Physiology in Drd2-EGFP Mice. Journal of Neuroscience 31: 126-32

272
81. Krueger F, Pardini M, Huey ED, Raymont V, Solomon J, Lipsky RH, Hodgkinson CA, Goldman D, Grafman J. 2011. The Role of the Met66 Brain-Derived Neurotrophic Factor Allele in the Recovery of Executive Functioning after Combat-Related Traumatic Brain Injury. Journal of Neuroscience 31: 598-606
82. Kunos G, Tam J. 2011. The case for peripheral CB1 receptor blockade in the treatment of visceral obesity and its cardiometabolic complications. British Journal of Pharmacology 163: 1423-31
83. Lederle L, Weber S, Wright T, Feyder M, Brigman JL, Crombag HS, Saksida LM, Bussey TJ, Holmes A. 2011. Reward-Related Behavioral Paradigms for Addiction Research in the Mouse: Performance of Common Inbred Strains. Plos One 6
84. Lee MR, Gallen CL, Zhang XC, Hodgkinson CA, Goldman D, Stein EA, Barr CS. 2011. Functional Polymorphism of the Mu-Opioid Receptor Gene (OPRM1) Influences Reinforcement Learning in Humans. Plos One 6
85. Lee SB, Park JH, Folk JE, Deck JA, Pegg AE, Sokabe M, Fraser CS, Park MH. 2011. Inactivation of eukaryotic initiation factor 5A (eIF5A) by specific acetylation of its hypusine residue by spermidine/spermine acetyltransferase 1 (SSAT1). Biochemical Journal 433: 205-13
86. Leggio L, Addolorato G, Cippitelli A, Jerlhag E, Kampov-Polevoy AB, Swift RM. 2011. Role of Feeding-Related Pathways in Alcohol Dependence: A Focus on Sweet Preference, NPY, and Ghrelin. Alcoholism-Clinical and Experimental Research 35: 194-202
87. Lewis MD, Hibbeln JR, Johnson JE, Lin YH, Hyun DY, Loewke JD. 2011. Suicide Deaths of Active-Duty US Military and Omega-3 Fatty-Acid Status: A Case-Control Comparison. Journal of Clinical Psychiatry 72: 1585-90
88. Li F, Folk JE, Cheng KJ, Kurimura M, Deck JA, Deschamps JR, Rothman RB, Dersch CM, Jacobson AE, Rice KC. 2011. Probes for narcotic receptor mediated phenomena. 43. Synthesis of the ortho-a and para-a, and improved synthesis and optical resolution of the ortho-b and para-b oxide-bridged phenylmorphans: Compounds with moderate to low opioid-receptor affinity. Bioorganic & Medicinal Chemistry 19: 4330-7
89. Li JX, Crocker C, Koek W, Rice KC, France CP. 2011. Effects of serotonin (5-HT)(1A) and 5-HT2A receptor agonists on schedule-controlled responding in rats: drug combination studies. Psychopharmacology 213: 489-97
90. Li JX, Koek W, Rice KC, France CP. 2011. Effects of Direct- and Indirect-Acting Serotonin Receptor Agonists on the Antinociceptive and Discriminative Stimulus Effects of Morphine in Rhesus Monkeys. Neuropsychopharmacology 36: 940-9
91. Li Y, Xu SQ, Mihaylova MM, Zheng B, Hou XY, Jiang BB, Park O, Luo ZJ, Lefai E, Shyy JYJ, Gao B, Wierzbicki M, Verbeuren TJ, Shaw RJ, Cohen RA, Zang MW. 2011. AMPK Phosphorylates and Inhibits SREBP Activity to Attenuate Hepatic Steatosis and Atherosclerosis in Diet-Induced Insulin-Resistant Mice. Cell Metabolism 13: 376-88
92. Lim HJ, Deschamps JR, Jacobson AE, Rice KC. 2011. Diastereoselective One-Pot Synthesis of 7-and 8-Substituted 5-Phenylmorphans. Organic Letters 13: 5322-5
93. Lim JH, Kim TW, Park SJ, Song IB, Kim MS, Kwon HJ, Cho ES, Son HY, Lee SW, Suh JW, Kim JW, Yun HI. 2011. Protective Effects of Platycodon grandiflorum Aqueous Extract on Thioacetamide-induced Fulminant Hepatic Failure in Mice. Journal of Toxicologic Pathology 24: 223-8
94. Lin YH, Shah S, Salem N. 2011. Altered essential fatty acid metabolism and composition in rat liver, plasma, heart and brain after microalgal DHA addition to the diet. Journal of Nutritional Biochemistry 22: 758-65

273
95. Liu JX, Zhou ZF, Hodgkinson CA, Yuan QP, Shen PH, Mulligan CJ, Wang A, Gray RR, Roy A, Virkkunen M, Goldman D, Enoch MA. 2011. Haplotype-Based Study of the Association of Alcohol-Metabolizing Genes With Alcohol Dependence in Four Independent Populations. Alcoholism-Clinical and Experimental Research 35: 304-16
96. Lopez-Quintero C, de los Cobos JP, Hasin DS, Okuda M, Wang S, Grant BF, Blanco C. 2011. Probability and predictors of transition from first use to dependence on nicotine, alcohol, cannabis, and cocaine: Results of the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). Drug and Alcohol Dependence 115: 120-30
97. Lopez-Quintero C, Hasin DS, de los Cobos JP, Pines A, Wang SA, Grant BF, Blanco C. 2011. Probability and predictors of remission from life-time nicotine, alcohol, cannabis or cocaine dependence: results from the National Epidemiologic Survey on Alcohol and Related Conditions. Addiction 106: 657-69
98. Luo ZC, Volkow ND, Heintz N, Pan YT, Du CW. 2011. Acute Cocaine Induces Fast Activation of D1 Receptor and Progressive Deactivation of D2 Receptor Striatal Neurons: In Vivo Optical Microprobe Ca2+ (i) Imaging. Journal of Neuroscience 31: 13180-90
99. Mabley J, Gordon S, Pacher P. 2011. Nicotine Exerts an Anti-inflammatory Effect in a Murine Model of Acute Lung Injury. Inflammation 34: 231-7
100. Marsh AA, Crowe SL, Yu HH, Gorodetsky EK, Goldman D, Blair RJR. 2011. Serotonin Transporter Genotype (5-HTTLPR) Predicts Utilitarian Moral Judgments. Plos One 6
101. Mathur BN, Capik NA, Alvarez VA, Lovinger DM. 2011. Serotonin Induces Long-Term Depression at Corticostriatal Synapses. Journal of Neuroscience 31: 7402-11
102. Mihailescu M, Soubias O, Worcester D, White SH, Gawrisch K. 2011. Structure and Dynamics of Cholesterol-Containing Polyunsaturated Lipid Membranes Studied by Neutron Diffraction and NMR. Journal of Membrane Biology 239: 63-71
103. Miller AM, Horiguchi N, Jeong WI, Radaeva S, Gao B. 2011. Molecular Mechanisms of Alcoholic Liver Disease: Innate Immunity and Cytokines. Alcoholism-Clinical and Experimental Research 35: 787-93
104. Miller AM, Wang H, Bertola A, Park O, Horiguchi N, Ki SH, Yin S, Lafdil F, Gao B. 2011. Inflammation-Associated Interleukin-6/Signal Transducer and Activator of Transcription 3 Activation Ameliorates Alcoholic and Nonalcoholic Fatty Liver Diseases in Interleukin-10-Deficient Mice. Hepatology 54: 846-56
105. Mongeon R, Walogorsky M, Urban J, Mandel G, Ono F, Brehm P. 2011. An acetylcholine receptor lacking both gamma and epsilon subunits mediates transmission in zebrafish slow muscle synapses. Journal of General Physiology 138: 353-66
106. Morton RA, Luo GX, Davis MI, Hales TG, Lovinger DM. 2011. Fluorophore assisted light inactivation (FALI) of recombinant 5-HT(3)A receptor constitutive internalization and function. Molecular and Cellular Neuroscience 47: 79-92
107. Mukhopadhyay B, Cinar R, Yin S, Liu J, Tam J, Godlewski G, Harvey-White J, Mordi I, Cravatt BF, Lotersztajn S, Gao B, Yuan QP, Schuebel K, Goldman D, Kunos G. 2011. Hyperactivation of anandamide synthesis and regulation of cell-cycle progression via cannabinoid type 1 (CB1) receptors in the regenerating liver. Proceedings of the National Academy of Sciences of the United States of America 108: 6323-8
108. Mukhopadhyay P, Horvath B, Kechrid M, Tanchian G, Rajesh M, Naura AS, Boulares AH, Pacher P. 2011. Poly(ADP-ribose) polymerase-1 is a key mediator of cisplatin-induced kidney inflammation and injury. Free Radical Biology and Medicine 51: 1774-88
109. Mukhopadhyay P, Horvath B, Rajesh M, Matsumoto S, Saito K, Batkai S, Patel V, Tanchian G, Gao RY, Cravatt BF, Hasko G, Pacher P. 2011. Fatty acid amide hydrolase

274
is a key regulator of endocannabinoid-induced myocardial tissue injury. Free Radical Biology and Medicine 50: 179-95
110. Mukhopadhyay P, Pacher P, Das DK. 2011. MicroRNA signatures of resveratrol in the ischemic heart. In Resveratrol and Health, ed. O Vang, DK Das, pp. 109-16
111. Mukhopadhyay P, Rajesh M, Horvath B, Batkai S, Park O, Tanchian G, Gao RY, Patel V, Wink DA, Liaudet L, Hasko G, Mechoulam R, Pacher P. 2011. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radical Biology and Medicine 50: 1368-81
112. O'Konek JJ, Illarionov P, Khursigara DS, Ambrosino E, Lzhak L, Castillo BF, Raju R, Khalili M, Kim HY, Howel AR, Besra GS, Porcelli SA, Berzofsky JA, Terabe M. 2011. Mouse and human iNKT cell agonist beta-mannosylceramide reveals a distinct mechanism of tumor immunity. Journal of Clinical Investigation 121: 683-94
113. Okuda M, Olfson M, Hasin D, Grant BF, Lin KH, Blanco C. 2011. Mental Health of Victims of Intimate Partner Violence: Results From a National Epidemiologic Survey. Psychiatric Services 62: 959-62
114. Pacher P, Mechoulam R. 2011. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Progress in Lipid Research 50: 193-211
115. Pardini M, Krueger F, Hodgkinson C, Raymont V, Ferrier C, Goldman D, Strenziok M, Guida S, Grafman J. 2011. Prefrontal cortex lesions and MAO-A modulate aggression in penetrating traumatic brain injury. Neurology 76: 1038-45
116. Park O, Wang H, Weng HL, Feigenbaum L, Li H, Yin S, Ki SH, Yoo SH, Dooley S, Wang FS, Young HA, Gao B. 2011. In Vivo Consequences of Liver-Specific Interleukin-22 Expression in Mice: Implications for Human Liver Disease Progression. Hepatology 54: 252-61
117. Pickering RP, Goldstein RB, Hasin DS, Blanco C, Smith SM, Huang BJ, Pulay AJ, Ruan WJ, Saha TD, Stinson FS, Dawson DA, Chou SP, Grant BF. 2011. Temporal Relationships Between Overweight and Obesity and DSM-IV Substance Use, Mood, and Anxiety Disorders: Results From a Prospective Study, the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Clinical Psychiatry 72: 1494-502
118. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Medical Comorbidity of Full and Partial Posttraumatic Stress Disorder in US Adults: Results From Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Psychosomatic Medicine 73: 697-707
119. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Personality disorders associated with full and partial posttraumatic stress disorder in the US population: Results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Psychiatric Research 45: 678-86
120. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2011. Prevalence and Axis I comorbidity of full and partial posttraumatic stress disorder in the United States: Results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Anxiety Disorders 25: 456-65
121. Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, Damadzic R, Eskay R, Schoor M, Thorsell A, Schwandt ML, Sommer WH, George DT, Parsons LH, Herscovitch P, Hommer D, Heilig M. 2011. A genetic determinant of the striatal dopamine response to alcohol in men. Molecular Psychiatry 16: 809-17
122. Ramsden CE, Hibbeln JR, Majchrzak-Hong SF, Davis JM. 2011. Don't disregard the essential distinction between PUFA species. British Journal of Nutrition 106: 953-7

275
123. Ramsden CE, Hibbeln JR, Majchrzak-Hong SF, Davis JM. 2011. Don't disregard the essential distinction between PUFA species Comment on Ramsden et al Response. British Journal of Nutrition 106: 959-60
124. Ramsden CE, Mann JD, Faurot KR, Lynch C, Imam ST, MacIntosh BA, Hibbeln JR, Loewke J, Smith S, Coble R, Suchindran C, Gaylord SA. 2011. Low omega-6 vs. low omega-6 plus high omega-3 dietary intervention for Chronic Daily Headache: Protocol for a randomized clinical trial. Trials 12
125. Reid AE, Kim SW, Seiner B, Fowler FW, Hooker J, Ferrieri R, Babst B, Fowler JS. 2011. Radiosynthesis of C-11 labeled auxin (3-indolyl 1-C-11 acetic acid) and its derivatives from gramine. Journal of Labelled Compounds & Radiopharmaceuticals 54: 433-7
126. Reimold M, Knobel A, Rapp MA, Batra A, Wiedemann K, Strohle A, Zimmer A, Schonknecht P, Smolka MN, Weinberger DR, Goldman D, Machulla HJ, Bares R, Heinz A. 2011. Central serotonin transporter levels are associated with stress hormone response and anxiety. Psychopharmacology 213: 563-72
127. Rinker JA, Hutchison MA, Chen SA, Thorsell A, Heilig M, Riley AL. 2011. Exposure to nicotine during periadolescence or early adulthood alters aversive and physiological effects induced by ethanol. Pharmacology Biochemistry and Behavior 99: 7-16
128. Roh S, Matsushita S, Hara S, Maesato H, Matsui T, Suzuki G, Miyakawa T, Ramchandani VA, Li TK, Higuchi S. 2011. Role of GABRA2 in Moderating Subjective Responses to Alcohol. Alcoholism-Clinical and Experimental Research 35: 400-7
129. Root TL, Szatkiewicz JP, Jonassaint CR, Thornton LM, Pinheiro AP, Strober M, Bloss C, Berrettini W, Schork NJ, Kaye WH, Bergen AW, Magistretti P, Brandt H, Crawford S, Crow S, Fichter MM, Goldman D, Halmi KA, Johnson C, Kaplan AS, Keel PK, Klump KL, La Via M, Mitchell JE, Rotondo A, Treasure J, Woodside DB, Bulik CM. 2011. Association of Candidate Genes with Phenotypic Traits Relevant to Anorexia Nervosa. European Eating Disorders Review 19: 487-93
130. Rostami E, Krueger F, Zoubak S, Dal Monte O, Raymont V, Pardini M, Hodgkinson CA, Goldman D, Risling M, Grafman J. 2011. BDNF Polymorphism Predicts General Intelligence after Penetrating Traumatic Brain Injury. Plos One 6
131. Roy A, Roy M, Goldman D. 2011. Childhood Trauma and Depressive Symptoms in Type 1 Diabetes. Journal of Clinical Psychiatry 72: 1049-53
132. Sabino V, Cottone P, Blasio A, Iyer MR, Steardo L, Rice KC, Conti B, Koob GF, Zorrilla EP. 2011. Activation of sigma-Receptors Induces Binge-like Drinking in Sardinian Alcohol-Preferring Rats. Neuropsychopharmacology 36: 1207-18
133. Schank JR, Pickens CL, Rowe KE, Cheng K, Thorsell A, Rice KC, Shaham Y, Heilig M. 2011. Stress-induced reinstatement of alcohol-seeking in rats is selectively suppressed by the neurokinin 1 (NK1) antagonist L822429. Psychopharmacology 218: 111-9
134. Schneider ML, Moore CF, Barr CS, Larson JA, Kraemer GW. 2011. Moderate Prenatal Alcohol Exposure and Serotonin Genotype Interact to Alter CNS Serotonin Function in Rhesus Monkey Offspring. Alcoholism-Clinical and Experimental Research 35: 912-20
135. Schuckit MA, Smith TL, Heron J, Hickman M, Macleod J, Lewis G, Davis JM, Hibbeln JR, Brown S, Zuccolo L, Miller LL, Davey-Smith G. 2011. Testing a Level of Response to Alcohol-Based Model of Heavy Drinking and Alcohol Problems in 1,905 17-year-olds. Alcoholism-Clinical and Experimental Research 35: 1897-904
136. Schwandt ML, Lindell SG, Higley JD, Suomi SJ, Heilig M, Barr CS. 2011. OPRM1 gene variation influences hypothalamic-pituitary-adrenal axis function in response to a variety of stressors in rhesus macaques. Psychoneuroendocrinology 36: 1303-11

276
137. Seif T, Makriyannis A, Kunos G, Bonci A, Hopf FW. 2011. THE Endocannabinoid 2-Arachidonoylglycerol Mediates D1 and D2 Receptor Cooperative Enhancement of Rat Nucleus Accumbens Core Neuron Firing. Neuroscience 193: 21-33
138. Sidhu VK, Huang BX, Kim HY. 2011. Effects of Docosahexaenoic Acid on Mouse Brain Synaptic Plasma Membrane Proteome Analyzed by Mass Spectrometry and O-16/O-18 Labeling. Journal of Proteome Research 10: 5472-80
139. Sinha R, Shaham Y, Heilig M. 2011. Translational and reverse translational research on the role of stress in drug craving and relapse. Psychopharmacology 218: 69-82
140. Song BJ, Abdelmegeed MA, Yoo SH, Kim BJ, Jo SA, Jo I, Moon KH. 2011. Post-translational modifications of mitochondrial aldehyde dehydrogenase and biomedical implications. Journal of Proteomics 74: 2691-702
141. Soriano FG, Lorigados CB, Pacher P, Szabo C. 2011. Effects of a Potent Peroxynitrite Decomposition Catalyst in Murine Models of Endotoxemia and Sepsis. Shock 35: 560-6
142. Spector AA, Campbell WB, Zeldin DC. 2011. Special Issue Eicosanoids and disease Introduction. Prostaglandins & Other Lipid Mediators 96: 1-2
143. Sreedharan S, Almen MS, Carlini VP, Haitina T, Stephansson O, Sommer WH, Heilig M, de Barioglio SR, Fredriksson R, Schioth HB. 2011. The G protein coupled receptor Gpr153 shares common evolutionary origin with Gpr162 and is highly expressed in central regions including the thalamus, cerebellum and the arcuate nucleus. Febs Journal 278: 4881-94
144. Stangl B, Hirshman E, Verbalis J. 2011. Administration of Dehydroepiandrosterone (DHEA) Enhances Visual-Spatial Performance in Postmenopausal Women. Behavioral Neuroscience 125: 742-52
145. Stasch JP, Pacher P, Evgenov OV. 2011. Soluble Guanylate Cyclase as an Emerging Therapeutic Target in Cardiopulmonary Disease. Circulation 123: 2263-73
146. Stopponi S, Somaini L, Cippitelli A, Cannella N, Braconi S, Kallupi M, Ruggeri B, Heilig M, Demopulos G, Gaitanaris G, Massi M, Ciccocioppo R. 2011. Activation of Nuclear PPAR gamma Receptors by the Antidiabetic Agent Pioglitazone Suppresses Alcohol Drinking and Relapse to Alcohol Seeking. Biological Psychiatry 69: 642-9
147. Tam J, Liu J, Mukhopadhyay B, Cinar R, Godlewski G, Kunos G. 2011. Endocannabinoids in Liver Disease. Hepatology 53: 346-55
148. Thanos PK, Bermeo C, Wang GJ, Volkow ND. 2011. D-Cycloserine Facilitates Extinction of Cocaine Self-Administration in Rats. Synapse 65: 938-44
149. Thanos PK, Cho J, Kim R, Michaelides M, Primeaux S, Bray G, Wang GJ, Volkow ND. 2011. Bromocriptine increased operant responding for high fat food but decreased chow intake in both obesity-prone and resistant rats. Behavioural Brain Research 217: 165-70
150. Thanos PK, Gopez V, Delis F, Michaelides M, Grandy DK, Wang GJ, Kunos G, Volkow ND. 2011. Upregulation of Cannabinoid Type 1 Receptors in Dopamine D2 Receptor Knockout Mice Is Reversed by Chronic Forced Ethanol Consumption. Alcoholism-Clinical and Experimental Research 35: 19-27
151. Thanos PK, Subrize M, Lui W, Puca Z, Ananth M, Michaelides M, Wang GJ, Volkow ND. 2011. D-Cycloserine Facilitates Extinction of Cocaine Self-Administration in c57 Mice. Synapse 65: 1099-105
152. Tikkanen R, Auvinen-Lintunen L, Ducci F, Sjoberg RL, Goldman D, Tiihonen J, Ojansuu I, Virkkunen M. 2011. Psychopathy, PCL-R, and MAOA genotype as predictors of violent reconvictions. Psychiatry Research 185: 382-6
153. Tomasi D, Volkow ND. 2011. Association between Functional Connectivity Hubs and Brain Networks. Cerebral Cortex 21: 2003-13

277
154. Tomasi D, Volkow ND. 2011. Functional connectivity hubs in the human brain. Neuroimage 57: 908-17
155. Tomasi D, Volkow ND, Wang GJ, Wang R, Telang F, Caparelli EC, Wong C, Jayne M, Fowler JS. 2011. Methylphenidate enhances brain activation and deactivation responses to visual attention and working memory tasks in healthy controls. Neuroimage 54: 3101-10
156. Torres-Altoro MI, Mathur BN, Drerup JM, Thomas R, Lovinger DM, O'Callaghan JP, Bibb JA. 2011. Organophosphates dysregulate dopamine signaling, glutamatergic neurotransmission, and induce neuronal injury markers in striatum. Journal of Neurochemistry 119: 303-13
157. Tu WZ, Eckert GJ, DiMeglio LA, Yu ZS, Jung J, Pratt JH. 2011. Intensified Effect of Adiposity on Blood Pressure in Overweight and Obese Children. Hypertension 58: 818-24
158. Umhau JC. 2011. Docosahexaenoic Acid Supplementation and Alzheimer Disease. Jama-Journal of the American Medical Association 305: 672-
159. Umhau JC, Schwandt ML, Usala J, Geyer C, Singley E, George DT, Heilig M. 2011. Pharmacologically induced alcohol craving in treatment seeking alcoholics correlates with alcoholism severity, but is insensitive to acamprosate. Neuropsychopharmacology 36: 1178-86
160. Upreti VV, Moon KH, Yu LR, Lee IJ, Eddington ND, Ye X, Veenstra TD, Song BJ. 2011. Increased oxidative-modifications of cytosolic proteins in 3,4-methylenedioxymethamphetamine (MDMA, ecstasy)-exposed rat liver. Proteomics 11: 202-11
161. Volkow ND, Tomasi D, Vaska P. 2011. Cell Phone Activation and Brain Glucose Metabolism Reply. Jama-Journal of the American Medical Association 305: 2067-8
162. Volkow ND, Tomasi D, Wang GJ, Fowler JS, Telang F, Goldstein RZ, Alia-Klein N, Woicik P, Wong C, Logan J, Millard J, Alexoff D. 2011. Positive emotionality is associated with baseline metabolism in orbitofrontal cortex and in regions of the default network. Molecular Psychiatry 16: 818-25
163. Volkow ND, Tomasi D, Wang GJ, Fowler JS, Telang F, Goldstein RZ, Alia-Klein N, Wong C. 2011. Reduced Metabolism in Brain "Control Networks" following Cocaine-Cues Exposure in Female Cocaine Abusers. Plos One 6
164. Volkow ND, Tomasi D, Wang GJ, Vaska P, Fowler JS, Telang F, Alexoff D, Logan J, Wong C. 2011. Effects of Cell Phone Radiofrequency Signal Exposure on Brain Glucose Metabolism. Jama-Journal of the American Medical Association 305: 808-13
165. Volkow ND, Wang GJ, Fowler JS, Tomasi D, Telang F. 2011. Addiction: Beyond dopamine reward circuitry. Proceedings of the National Academy of Sciences of the United States of America 108: 15037-42
166. Volkow ND, Wang GJ, Newcorn JH, Kollins SH, Wigal TL, Telang F, Fowler JS, Goldstein RZ, Klein N, Logan J, Wong C, Swanson JM. 2011. Motivation deficit in ADHD is associated with dysfunction of the dopamine reward pathway. Molecular Psychiatry 16: 1147-54
167. Wang GJ, Geliebter A, Volkow ND, Telang FW, Logan J, Jayne MC, Galanti K, Selig PA, Han H, Zhu W, Wong CT, Fowler JS. 2011. Enhanced Striatal Dopamine Release During Food Stimulation in Binge Eating Disorder. Obesity 19: 1601-8
168. Wang H, Lafdil F, Kong XN, Gao B. 2011. Signal Transducer and Activator of Transcription 3 in Liver Diseases: A Novel Therapeutic Target. International Journal of Biological Sciences 7: 536-50

278
169. Wang H, Lafdil F, Wang L, Park O, Yin S, Niu JY, Miller AM, Sun ZL, Gao B. 2011. Hepatoprotective versus Oncogenic Functions of STAT3 in Liver Tumorigenesis. American Journal of Pathology 179: 714-24
170. Wang H, Lafdil F, Wang L, Yin S, Feng DC, Gao B. 2011. Tissue inhibitor of metalloproteinase 1 (TIMP-1) deficiency exacerbates carbon tetrachloride-induced liver injury and fibrosis in mice: involvement of hepatocyte STAT3 in TIMP-1 production. Cell and Bioscience 1
171. Wang H, Park O, Gao B. 2011. NKT cells in liver fibrosis: Controversies or complexities. Journal of Hepatology 55: 1166-
172. Watcho P, Stavniichuk R, Tane P, Shevalye H, Maksimchyk Y, Pacher P, Obrosova IG. 2011. Evaluation of PMI-5011, an ethanolic extract of Artemisia dracunculus L., on peripheral neuropathy in streptozotocin-diabetic mice. International Journal of Molecular Medicine 27: 299-307
173. Wells EM, Jarrett JM, Lin YH, Caldwell KL, Hibbeln JR, Apelberg BJ, Herbstman J, Halden RU, Witter FR, Goldman LR. 2011. Body burdens of mercury, lead, selenium and copper among Baltimore newborns. Environmental Research 111: 411-7
174. Winder D, Lovinger DM. 2011. Introduction to the special issue on synaptic plasticity and addiction. Neuropharmacology 61: 1051-
175. Won YJ, Ono F, Ikeda SR. 2011. Identification and Modulation of Voltage-Gated Ca2+ Currents in Zebrafish Rohon-Beard Neurons. Journal of Neurophysiology 105: 442-53
176. Xiong W, Koo BN, Morton R, Zhang L. 2011. Psychotropic and nonpsychotropic Cannabis Derivatives Inhibit Human 5-HT3A Receptors Through a Receptor Desensitization-Dependent Mechanism. Neuroscience 184: 28-37
177. Xu Y, Okuda M, Hser YI, Hasin D, Liu SM, Grant BF, Blanco C. 2011. Twelve-month prevalence of psychiatric disorders and treatment-seeking among Asian Americans/Pacific Islanders in the United States: Results from the National Epidemiological Survey on Alcohol and Related Conditions. Journal of Psychiatric Research 45: 910-8
178. Yi JM, Dhir M, Van Neste L, Downing SR, Jeschke J, Glockner SC, Calmon MD, Hooker CM, Funes JM, Boshoff C, Smits KM, van Engeland M, Weijenberg MP, Iacobuzio-Donahue CA, Herman JG, Schuebel KE, Baylin SB, Ahuja N. 2011. Genomic and Epigenomic Integration Identifies a Prognostic Signature in Colon Cancer. Clinical Cancer Research 17: 1535-45
179. Yin S, Wang H, Park O, Wei W, Shen JL, Gao B. 2011. Enhanced Liver Regeneration in IL-10-Deficient Mice after Partial Hepatectomy via Stimulating Inflammatory Response and Activating Hepatocyte STAT3. American Journal of Pathology 178: 1614-21
180. Yoo SH, Park O, Henderson LE, Abdelmegeed MA, Moon KH, Song BJ. 2011. Lack of PPAR alpha exacerbates lipopolysaccharide-induced liver toxicity through STAT1 inflammatory signaling and increased oxidative/nitrosative stress. Toxicology Letters 202: 23-9
181. Young-Wolff KC, Enoch MA, Prescott CA. 2011. The influence of gene-environment interactions on alcohol consumption and alcohol use disorders: A comprehensive review. Clinical Psychology Review 31: 800-16
182. Zanarini MC, Horwood J, Wolke D, Waylen A, Fitzmaurice G, Grant BF. 2011. Prevalence of DSM-IV Borderline Personality Disorder in Two Community Samples: 6,330 English 11-Year-Olds and 34,653 American Adults. Journal of Personality Disorders 25: 607-19
183. Zhang Z, Zhang SY, Zou ZS, Shi JF, Zhao JJ, Fan R, Qin EQ, Li BS, Li ZW, Xu XS, Fu JL, Zhang JY, Gao B, Tian ZG, Wang FS. 2011. Hypercytolytic Activity of Hepatic

279
Natural Killer Cells Correlates With Liver Injury in Chronic Hepatitis B Patients. Hepatology 53: 73-85
184. Zhou ZF, Yuan QP, Mash DC, Goldman D. 2011. Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol. Proceedings of the National Academy of Sciences of the United States of America 108: 6626-31
185. Zoerner AA, Stichtenoth DO, Engeli S, Batkai S, Winkler C, Schaumann F, Janke J, Holz O, Krug N, Tsikas D, Jordan J, Hohlfeld JM. 2011. Allergen Challenge Increases Anandamide in Bronchoalveolar Fluid of Patients With Allergic Asthma. Clinical Pharmacology & Therapeutics 90: 388-91
DICBR PUBLICATIONS 2012
1. Abdelmegeed MA, Banerjee A, Yoo SH, Jang S, Gonzalez FJ, Song BJ. 2012. Critical
role of cytochrome P450 2E1 (CYP2E1) in the development of high fat-induced non-alcoholic steatohepatitis. Journal of Hepatology 57: 860-6
2. Alvheim AR, Malde MK, Osei-Hyiaman D, Lin YH, Pawlosky RJ, Madsen L, Kristiansen K, Froyland L, Hibbeln JR. 2012. Dietary Linoleic Acid Elevates Endogenous 2-AG and Anandamide and Induces Obesity. Obesity 20: 1984-94
3. Alzghoul L, Bortolato M, Delis F, Thanos PK, Darling RD, Godar SC, Zhang JL, Grant S, Wang GJ, Simpson KL, Chen K, Volkow ND, Lin RCS, Shih JC. 2012. Altered cerebellar organization and function in monoamine oxidase A hypomorphic mice. Neuropharmacology 63: 1208-17
4. Armagan G, Bojnik E, Turunc E, Kanit L, Cinar OG, Benyhe S, Borsodi A, Yalcin A. 2012. Kainic acid-induced changes in the opioid/nociceptin system and the stress/toxicity pathways in the rat hippocampus. Neurochemistry International 60: 555-64
5. Balamayooran G, Batra S, Theivanthiran B, Cai SS, Pacher P, Jeyaseelan S. 2012. Intrapulmonary G-CSF Rescues Neutrophil Recruitment to the Lung and Neutrophil Release to Blood in Gram-Negative Bacterial Infection in MCP-1(-/-) Mice. Journal of Immunology 189: 5849-59
6. Bashashati M, Storr MA, Nikas SP, Wood JT, Godlewski G, Liu J, Ho W, Keenan CM, Zhang H, Alapafuja SO, Cravatt BF, Lutz B, Mackie K, Kunos G, Patel KD, Makriyannis A, Davison JS, Sharkey KA. 2012. Inhibiting fatty acid amide hydrolase normalizes endotoxin-induced enhanced gastrointestinal motility in mice. British Journal of Pharmacology 165: 1556-71
7. Batkai S, Mukhopadhyay P, Horvath B, Rajesh M, Gao RY, Mahadevan A, Amere M, Battista N, Lichtman AH, Gauson LA, Maccarrone M, Pertwee RG, Pacher P. 2012. Delta(8)-Tetrahydrocannabivarin prevents hepatic ischaemia/reperfusion injury by decreasing oxidative stress and inflammatory responses through cannabinoid CB2 receptors. British Journal of Pharmacology 165: 2450-61
8. Bevilacqua L, Carli V, Sarchiapone M, George DK, Goldman D, Roy A, Enoch MA. 2012. Interaction Between FKBP5 and Childhood Trauma and Risk of Aggressive Behavior. Archives of General Psychiatry 69: 62-70
9. Bjork JM, Chen G, Hommer DW. 2012. Psychopathic tendencies and mesolimbic recruitment by cues for instrumental and passively obtained rewards. Biological Psychology 89: 408-15
10. Bjork JM, Smith AR, Chen G, Hommer DW. 2012. Mesolimbic recruitment by nondrug rewards in detoxified alcoholics: Effort anticipation, reward anticipation, and reward delivery. Human Brain Mapping 33: 2174-88

280
11. Blanco C, Vesga-Lopez O, Stewart JW, Liu SM, Grant BF, Hasin DS. 2012. Epidemiology of Major Depression With Atypical Features: Results From the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). Journal of Clinical Psychiatry 73: 224-32
12. Boos TL, Cheng KJ, Greiner E, Deschamps JR, Jacobson AE, Rice KC. 2012. Configurational Reassignment and Improved Preparation of the Competitive IL-6 Receptor Antagonist 20R,21R-Epoxyresibufogenin-3-formate. Journal of Natural Products 75: 661-8
13. Borghese CM, Blednov YA, Quan Y, Iyer SV, Xiong W, Mihic SJ, Zhang L, Lovinger DM, Trudell JR, Homanics GE, Harris RA. 2012. Characterization of Two Mutations, M287L and Q266I, in the alpha 1 Glycine Receptor Subunit That Modify Sensitivity to Alcohols. Journal of Pharmacology and Experimental Therapeutics 340: 304-16
14. Borghese CM, Xiong W, Oh SI, Ho A, Mihic SJ, Zhang L, Lovinger DM, Homanics GE, Eger EI, Harris RA. 2012. Mutations M287L and Q266I in the Glycine Receptor alpha 1 Subunit Change Sensitivity to Volatile Anesthetics in Oocytes and Neurons, but Not the Minimal Alveolar Concentration in Knockin Mice. Anesthesiology 117: 765-71
15. Burghardt PR, Love TM, Stohler CS, Hodgkinson C, Shen PH, Enoch MA, Goldman D, Zubieta JK. 2012. Leptin Regulates Dopamine Responses to Sustained Stress in Humans. Journal of Neuroscience 32: 15369-76
16. Bussey TJ, Holmes A, Lyon L, Mar AC, McAllister KAL, Nithianantharajah J, Oomen CA, Saksida LM. 2012. New translational assays for preclinical modelling of cognition in schizophrenia: The touchscreen testing method for mice and rats. Neuropharmacology 62: 1191-203
17. Byun K, Bayarsaikhan E, Kim D, Kim CY, Mook-Jung I, Paek SH, Kim SU, Yamamoto T, Won MH, Song BJ, Park YM, Lee B. 2012. Induction of Neuronal Death by Microglial AGE-Albumin: Implications for Alzheimer's Disease. Plos One 7
18. Cachope R, Mateo Y, Mathur BN, Irving J, Wang HL, Morales M, Lovinger DM, Cheer JF. 2012. Selective Activation of Cholinergic Interneurons Enhances Accumbal Phasic Dopamine Release: Setting the Tone for Reward Processing. Cell Reports 2: 33-41
19. Cai SS, Batra S, Wakamatsu N, Pacher P, Jeyaseean S. 2012. NLRC4 Inflammasome-Mediated Production of IL-1 beta Modulates Mucosal Immunity in the Lung against Gram-Negative Bacterial Infection. Journal of Immunology 188: 5623-35
20. Camp MC, MacPherson KP, Lederle L, Graybeal C, Gaburro S, DeBrouse LM, Ihne JL, Bravo JA, O'Connor RM, Ciocchi S, Wellman CL, Luthi A, Cryan JF, Singewald N, Holmes A. 2012. Genetic Strain Differences in Learned Fear Inhibition Associated with Variation in Neuroendocrine, Autonomic, and Amygdala Dendritic Phenotypes. Neuropsychopharmacology 37: 1534-47
21. Chen CH, Walker J, Momenan R, Rawlings R, Heilig M, Hommer DW. 2012. Relationship Between Liver Function and Brain Shrinkage in Patients with Alcohol Dependence. Alcoholism-Clinical and Experimental Research 36: 625-32
22. Cherpitel CJ, Ye Y, Bond J, Borges G, Chou P, Nilsen P, Ruan JN, Xiang XJ. 2012. Multi-level analysis of alcohol-related injury and drinking pattern: emergency department data from 19 countries. Addiction 107: 1263-72
23. Chesler EJ, Plitt A, Fisher D, Hurd B, Lederle L, Bubier JA, Kiselycznyk C, Holmes A. 2012. Quantitative trait loci for sensitivity to ethanol intoxication in a C57BL/6J x 129S1/SvImJ inbred mouse cross. Mammalian Genome 23: 305-21
24. Chou SP, Chun S, Smith S, Ruan J, Li TK, Grant BF. 2012. Episodic heavy drinking, problem drinking and injuries - Results of the WHO/NIAAA collaborative emergency room study in South Korea. Alcohol 46: 407-13

281
25. Chou SP, Lee HK, Cho MJ, Park JI, Dawson DA, Grant BF. 2012. Alcohol Use Disorders, Nicotine Dependence, and Co-Occurring Mood and Anxiety Disorders in the United States and South KoreauA Cross-National Comparison. Alcoholism-Clinical and Experimental Research 36: 654-62
26. Cippitelli A, Damadzic R, Singley E, Thorsell A, Ciccocioppo R, Eskay RL, Heilig M. 2012. Pharmacological blockade of corticotropin-releasing hormone receptor 1 (CRH1R) reduces voluntary consumption of high alcohol concentrations in non-dependent Wistar rats. Pharmacology Biochemistry and Behavior 100: 522-9
27. Clarke K, Tchabanenko K, Pawlosky R, Carter E, King MT, Musa-Veloso K, Ho M, Roberts A, Robertson J, VanItallie TB, Veech RL. 2012. Kinetics, safety and tolerability of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate in healthy adult subjects. Regulatory Toxicology and Pharmacology 63: 401-8
28. Clarke K, Tchabanenko K, Pawlosky R, Carter E, Knight NS, Murray AJ, Cochlin LE, King MT, Wong AW, Roberts A, Robertson J, Veech RL. 2012. Oral 28-day and developmental toxicity studies of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate. Regulatory Toxicology and Pharmacology 63: 196-208
29. Cottone P, Wang XF, Park JW, Valenza M, Blasio A, Kwak J, Iyer MR, Steardo L, Rice KC, Hayashi T, Sabino V. 2012. Antagonism of Sigma-1 Receptors Blocks Compulsive-Like Eating. Neuropsychopharmacology 37: 2593-604
30. Csoka B, Selmeczy Z, Koscso B, Nemeth ZH, Pacher P, Murray PJ, Kepka-Lenhart D, Morris SM, Gause WC, Leibovich SJ, Hasko G. 2012. Adenosine promotes alternative macrophage activation via A(2A) and A(2B) receptors. Faseb Journal 26: 376-86
31. Dang MT, Yokoi F, Cheetham CC, Lu J, Vo V, Lovinger DM, Li YQ. 2012. An anticholinergic reverses motor control and corticostriatal LTD deficits in Dyt1 Delta GAG knock-in mice. Behavioural Brain Research 226: 465-72
32. Dawson DA, Goldstein RB, Grant BF. 2012. Factors Associated With First Utilization of Different Types of Care for Alcohol Problems. Journal of Studies on Alcohol and Drugs 73: 647-56
33. Dawson DA, Goldstein RB, Ruan WJJ, Grant BF. 2012. Correlates of Recovery from Alcohol Dependence: A Prospective Study Over a 3-Year Follow-Up Interval. Alcoholism-Clinical and Experimental Research 36: 1268-77
34. Dawson DA, Smith SM, Pickering RP, Grant BF. 2012. An empirical approach to evaluating the validity of alternative low-risk drinking guidelines. Drug and Alcohol Review 31: 141-50
35. Dawson DA, Smith SM, Saha TD, Rubinsky AD, Grant BF. 2012. Comparative performance of the AUDIT-C in screening for DSM-IV and DSM-5 alcohol use disorders. Drug and Alcohol Dependence 126: 384-8
36. Delis F, Benveniste H, Xenos M, Grandy D, Wang GJ, Volkow ND, Thanos PK. 2012. Loss of Dopamine D2 Receptors Induces Atrophy in the Temporal and Parietal Cortices and the Caudal Thalamus of Ethanol-Consuming Mice. Alcoholism-Clinical and Experimental Research 36: 815-25
37. Dilmanian FA, Rusek A, Fois GR, Olschowka J, Desnoyers NR, Park JY, Dioszegi I, Dane B, Wang RL, Tomasi D, Lee H, Hurley SD, Coyle PK, Meek AG, O'Banion MK. 2012. Interleaved Carbon Minibeams: An Experimental Radiosurgery Method With Clinical Potential. International Journal of Radiation Oncology Biology Physics 84: 514-9
38. Ducci F, Goldman D. 2012. The Genetic Basis of Addictive Disorders. Psychiatric Clinics of North America 35: 495-+

282
39. Eaton NR, Keyes KM, Krueger RF, Balsis S, Skodol AE, Markon KE, Grant BF, Hasin DS. 2012. An Invariant Dimensional Liability Model of Gender Differences in Mental Disorder Prevalence: Evidence From a National Sample. Journal of Abnormal Psychology 121: 282-8
40. Egli M, Koob GF, Edwards S. 2012. Alcohol dependence as a chronic pain disorder. Neuroscience and Biobehavioral Reviews 36: 2179-92
41. Engeli S, Bluher M, Jumpertz R, Wiesner T, Wirtz H, Bosse-Henck A, Stumvoll M, Batkai S, Pacher P, Harvey-White J, Kunos G, Jordan J. 2012. Circulating anandamide and blood pressure in patients with obstructive sleep apnea. Journal of Hypertension 30: 2345-51
42. Enoch MA. 2012. The Influence of Gene-Environment Interactions on the Development of Alcoholism and Drug Dependence. Current Psychiatry Reports 14: 150-8
43. Enoch MA, Zhou ZF, Kimura M, Mash DC, Yuan QP, Goldman D. 2012. GABAergic Gene Expression in Postmortem Hippocampus from Alcoholics and Cocaine Addicts; Corresponding Findings in Alcohol-Naive P and NP Rats. Plos One 7
44. Feng DC, Kong XN, Weng HL, Park O, Wang H, Dooley S, Gershwin ME, Gao B. 2012. Interleukin-22 Promotes Proliferation of Liver Stem/Progenitor Cells in Mice and Patients With Chronic Hepatitis B Virus Infection. Gastroenterology 143: 188-U375
45. Feng DC, Park O, Radaeva S, Wang H, Yin S, Kong XN, Zheng MQ, Zakhari S, Kolls JK, Gao B. 2012. Interleukin-22 Ameliorates Cerulein-Induced Pancreatitis in Mice by Inhibiting the Autophagic Pathway. International Journal of Biological Sciences 8: 249-57
46. Fenton MC, Keyes K, Geier T, Greenstein E, Skodol A, Krueger B, Grant BF, Hasin DS. 2012. Psychiatric comorbidity and the persistence of drug use disorders in the United States. Addiction 107: 599-609
47. Fitzgerald PJ. 2012. Whose side are you on: Does serotonin preferentially activate the right hemisphere and norepinephrine the left? Medical Hypotheses 79: 250-4
48. Fitzgerald PJ. 2012. Neurodrinking: Is alcohol a substrate in a novel, endogenous synthetic pathway for norepinephrine? Medical Hypotheses 78: 760-2
49. Fitzgerald PJ. 2012. The NMDA receptor may participate in widespread suppression of circuit level neural activity, in addition to a similarly prominent role in circuit level activation. Behavioural Brain Research 230: 291-8
50. Forbes CE, Poore JC, Barbey AK, Krueger F, Solomon J, Lipsky RH, Hodgkinson CA, Goldman D, Grafman J. 2012. BDNF Polymorphism-Dependent OFC and DLPFC Plasticity Differentially Moderates Implicit and Explicit Bias. Cerebral Cortex 22: 2602-9
51. French CA, Jin X, Campbell TG, Gerfen E, Groszer M, Fisher SE, Costa RM. 2012. An aetiological Foxp2 mutation causes aberrant striatal activity and alters plasticity during skill learning. Molecular Psychiatry 17: 1077-85
52. Gao B. 2012. Hepatoprotective and anti-inflammatory cytokines in alcoholic liver disease. Journal of Gastroenterology and Hepatology 27: 89-93
53. Gao B, Waisman A. 2012. Th17 Cells Regulate Liver Fibrosis by Targeting Multiple Cell Types: Many Birds With One Stone. Gastroenterology 143: 536-9
54. Gao B, Wang H, Lafdil F, Feng DC. 2012. STAT proteins - Key regulators of anti-viral responses, inflammation, and tumorigenesis in the liver. Journal of Hepatology 57: 430-41
55. Gilman JM, Ramchandani VA, Crouss T, Hommer DW. 2012. Subjective and Neural Responses to Intravenous Alcohol in Young Adults with Light and Heavy Drinking Patterns. Neuropsychopharmacology 37: 467-77

283
56. Gilman JM, Smith AR, Ramchandani VA, Momenan R, Hommer DW. 2012. The effect of intravenous alcohol on the neural correlates of risky decision making in healthy social drinkers. Addiction Biology 17: 465-78
57. Goldstein RB, Dawson DA, Chou P, Grant BF. 2012. Sex Differences in Prevalence and Comorbidity of Alcohol and Drug Use Disorders: Results From Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Studies on Alcohol and Drugs 73: 938-50
58. Goldstein RB, Dawson DA, Smith SM, Grant BF. 2012. Antisocial behavioral syndromes and 3-year quality-of-life outcomes in United States adults. Acta Psychiatrica Scandinavica 126: 137-50
59. Graybeal C, Kiselycznyk C, Holmes A. 2012. Stress-Induced Impairments in Prefrontal-mediated Behaviors and the Role of the N-Methyl-D-Aspartate Receptor. Neuroscience 211: 28-38
60. Guerrero-Beltran CE, Mukhopadhyay P, Horvath B, Rajesh M, Tapia E, Garcia-Torres I, Pedraza-Chaverri J, Pacher P. 2012. Sulforaphane, a natural constituent of broccoli, prevents cell death and inflammation in nephropathy. Journal of Nutritional Biochemistry 23: 494-500
61. Guo L, Yeh ML, Carlson VCC, Johnson-Venkatesh EM, Yeh HH. 2012. Nerve Growth Factor in the Hippocamposeptal System: Evidence for Activity-Dependent Anterograde Delivery and Modulation of Synaptic Activity. Journal of Neuroscience 32: 7701-10
62. Guo TZ, Wei TP, Shi XY, Li WW, Hou SY, Wang LP, Tsujikawa K, Rice KC, Cheng KJ, Clark DJ, Kingery WS. 2012. Neuropeptide deficient mice have attenuated nociceptive, vascular, and inflammatory changes in a tibia fracture model of complex regional pain syndrome. Molecular Pain 8
63. Hajnal A, Zharikov A, Polston JE, Fields MR, Tomasko J, Rogers AM, Volkow ND, Thanos PK. 2012. Alcohol Reward Is Increased after Roux-en-Y Gastric Bypass in Dietary Obese Rats with Differential Effects following Ghrelin Antagonism. Plos One 7
64. Hartley CA, McKenna MC, Salman R, Holmes A, Casey BJ, Phelps EA, Glatt CE. 2012. Serotonin transporter polyadenylation polymorphism modulates the retention of fear extinction memory. Proceedings of the National Academy of Sciences of the United States of America 109: 5493-8
65. Hasko G, Pacher P. 2012. Regulation of Macrophage Function by Adenosine. Arteriosclerosis Thrombosis and Vascular Biology 32: 865-9
66. Hensler JG, Advani T, Burke TF, Cheng K, Rice KC, Koek W. 2012. GABA(B) Receptor-Positive Modulators: Brain Region-Dependent Effects. Journal of Pharmacology and Experimental Therapeutics 340: 19-26
67. Hilario M, Holloway T, Jin X, Costa RM. 2012. Different dorsal striatum circuits mediate action discrimination and action generalization. European Journal of Neuroscience 35: 1105-14
68. Hisada M, Ota Y, Zhang X, Cameron AM, Gao B, Montgomery RA, Williams GM, Sun Z. 2012. Successful Transplantation of Reduced-Sized Rat Alcoholic Fatty Livers Made Possible by Mobilization of Host Stem Cells. American Journal of Transplantation 12: 3246-56
69. Holmes A, Fitzgerald PJ, MacPherson KP, DeBrouse L, Colacicco G, Flynn SM, Masneuf S, Pleil KE, Li C, Marcinkiewcz CA, Kash TL, Gunduz-Cinar O, Camp M. 2012. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-mediated fear extinction encoding. Nature Neuroscience 15: 1359-61
70. Honorio J, Tomasi D, Goldstein RZ, Leung HC, Samaras D. 2012. Can a Single Brain Region Predict a Disorder? Ieee Transactions on Medical Imaging 31: 2062-72

284
71. Horvath B, Magid L, Mukhopadhyay P, Batkai S, Rajesh M, Park O, Tanchian G, Gao RY, Goodfellow CE, Glass M, Mechoulam R, Pacher P. 2012. A new cannabinoid CB2 receptor agonist HU-910 attenuates oxidative stress, inflammation and cell death associated with hepatic ischaemia/reperfusion injury. British Journal of Pharmacology 165: 2462-78
72. Horvath B, Mukhopadhyay P, Hasko G, Pacher P. 2012. The Endocannabinoid System and Plant-Derived Cannabinoids in Diabetes and Diabetic Complications. American Journal of Pathology 180: 432-42
73. Horvath B, Mukhopadhyay P, Kechrid M, Patel V, Tanchian G, Wink DA, Gertsch J, Pacher P. 2012. beta-Caryophyllene ameliorates cisplatin-induced nephrotoxicity in a cannabinoid 2 receptor-dependent manner. Free Radical Biology and Medicine 52: 1325-33
74. Hsu DT, Mickey BJ, Langenecker SA, Heitzeg MM, Love TM, Wang H, Kennedy SE, Pecina M, Shafir T, Hodgkinson CA, Enoch MA, Goldman D, Zubieta JK. 2012. Variation in the Corticotropin-Releasing Hormone Receptor 1 (CRHR1) Gene Influences fMRI Signal Responses during Emotional Stimulus Processing. Journal of Neuroscience 32: 3253-60
75. Huang BX, Kim HY. 2012. Effects of Ethanol on Conformational Changes of Akt Studied by Chemical Cross-Linking, Mass Spectrometry, and O-18 Labeling. Acs Chemical Biology 7: 386-93
76. Huang MC, Schwandt ML, Ramchandani VA, George DT, Heilig M. 2012. Impact of Multiple Types of Childhood Trauma Exposure on Risk of Psychiatric Comorbidity Among Alcoholic Inpatients. Alcoholism-Clinical and Experimental Research 36: 598-606
77. Hutchinson MR, Northcutt AL, Hiranita T, Wang X, Lewis SS, Thomas J, van Steeg K, Kopajtic TA, Loram LC, Sfregola C, Galer E, Miles NE, Bland ST, Amat J, Rozeske RR, Maslanik T, Chapman TR, Strand KA, Fleshner M, Bachtell RK, Somogyi AA, Yin H, Katz JL, Rice KC, Maier SF, Watkins LR. 2012. Opioid Activation of Toll-Like Receptor 4 Contributes to Drug Reinforcement. Journal of Neuroscience 32: 11187-200
78. Ihne JL, Fitzgerald PJ, Hefner KR, Holmes A. 2012. Pharmacological modulation of stress-induced behavioral changes in the light/dark exploration test in male C57BL/6J mice. Neuropharmacology 62: 464-73
79. Iyer MR, Lee YS, Deschamps JR, Dersch CM, Rothman RB, Jacobson AE, Rice KC. 2012. Probes for narcotic receptor mediated phenomena. 44. Synthesis of an N-substituted 4-hydroxy-5-(3-hydroxyphenyl)morphan with high affinity and selective mu-antagonist activity. European Journal of Medicinal Chemistry 50: 44-54
80. Ji YW, Sun SY, Xu AM, Bhargava P, Yang L, Lam KSL, Gao B, Lee CH, Kersten S, Qi L. 2012. Activation of Natural Killer T Cells Promotes M2 Macrophage Polarization in Adipose Tissue and Improves Systemic Glucose Tolerance via Interleukin-4 (IL-4)/STAT6 Protein Signaling Axis in Obesity. Journal of Biological Chemistry 287: 13561-71
81. Kalu N, Ramchandani VA, Marshall V, Scott D, Ferguson C, Cain G, Taylor R. 2012. Heritability of Level of Response and Association with Recent Drinking History in Nonalcohol-Dependent Drinkers. Alcoholism-Clinical and Experimental Research 36: 522-9
82. Kamakura N, Yamamoto J, Brooks PJ, Iwai S, Kuraoka I. 2012. Effects of 5 ',8-Cyclodeoxyadenosine Triphosphates on DNA Synthesis. Chemical Research in Toxicology 25: 2718-24

285
83. Karlsson C, Zook M, Ciccocioppo R, Gehlert DR, Thorsell A, Heilig M, Cippitelli A. 2012. Melanin-concentrating hormone receptor 1 (MCH1-R) antagonism: Reduced appetite for calories and suppression of addictive-like behaviors. Pharmacology Biochemistry and Behavior 102: 400-6
84. Karlsson RM, Adermark L, Molander A, Perreau-Lenz S, Singley E, Solomon M, Holmes A, Tanaka K, Lovinger DM, Spanagel R, Heilig M. 2012. Reduced alcohol intake and reward associated with impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology 63: 181-9
85. Kenna GA, Swift RM, Hillemacher T, Leggio L. 2012. The Relationship of Appetitive, Reproductive and Posterior Pituitary Hormones to Alcoholism and Craving in Humans. Neuropsychology Review 22: 211-28
86. Keyes KM, Eaton NR, Krueger RF, McLaughlin KA, Wall MM, Grant BF, Hasin DS. 2012. Childhood maltreatment and the structure of common psychiatric disorders. British Journal of Psychiatry 200: 107-15
87. Keyes KM, Hatzenbuehler ML, Grant BF, Hasin DS. 2012. Stress and Alcohol Epidemiologic Evidence. Alcohol Research & Health 34: 391-400
88. Kimura T, Yeliseev AA, Vukoti K, Rhodes SD, Cheng KJ, Rice KC, Gawrisch K. 2012. Recombinant Cannabinoid Type 2 Receptor in Liposome Model Activates G Protein in Response to Anionic Lipid Constituents. Journal of Biological Chemistry 287: 4076-87
89. Klug JR, Mathur BN, Kash TL, Wang HD, Matthews RT, Robison AJ, Anderson ME, Deutch AY, Lovinger DM, Colbran RJ, Winder DG. 2012. Genetic Inhibition of CaMKII in Dorsal Striatal Medium Spiny Neurons Reduces Functional Excitatory Synapses and Enhances Intrinsic Excitability. Plos One 7
90. Koek W, France CP, Cheng KJ, Rice KC. 2012. Effects of the GABA(B) Receptor-Positive Modulators CGP7930 and rac-BHFF in Baclofen- and gamma-Hydroxybutyrate-Discriminating Pigeons. Journal of Pharmacology and Experimental Therapeutics 341: 369-76
91. Komatsu DE, Thanos PK, Mary MN, Janda HA, John CM, Robison L, Ananth M, Swanson JM, Volkow ND, Hadjiargyrou M. 2012. Chronic exposure to methylphenidate impairs appendicular bone quality in young rats. Bone 50: 1214-22
92. Kong XN, Feng DC, Wang H, Hong F, Bertola A, Wang FS, Gao B. 2012. Interleukin-22 induces hepatic stellate cell senescence and restricts liver fibrosis in mice. Hepatology 56: 1150-9
93. Koralek AC, Jin X, Ii JDL, Costa RM, Carmena JM. 2012. Corticostriatal plasticity is necessary for learning intentional neuroprosthetic skills. Nature 483: 331-5
94. Koscso B, Csoka B, Selmeczy Z, Himer L, Pacher P, Virag L, Hasko G. 2012. Adenosine Augments IL-10 Production by Microglial Cells through an A(2B) Adenosine Receptor-Mediated Process. Journal of Immunology 188: 445-53
95. Krepkiy D, Gawrisch K, Swartz KJ. 2012. Structural Interactions between Lipids, Water and S1-S4 Voltage-Sensing Domains. Journal of Molecular Biology 423: 632-47
96. Lands B, Lamoreaux E. 2012. Using 3-6 differences in essential fatty acids rather than 3/6 ratios gives useful food balance scores. Nutrition & Metabolism 9
97. Lewis SS, Loram LC, Hutchinson MR, Li CM, Zhang YN, Maier SF, Huang Y, Rice KC, Watkins LR. 2012. (+)-Naloxone, an Opioid-Inactive Toll-Like Receptor 4 Signaling Inhibitor, Reverses Multiple Models of Chronic Neuropathic Pain in Rats. Journal of Pain 13: 498-506
98. Li FY, Deck JA, Dersch CM, Rothman RB, Deschamps JR, Jacobson AE, Rice KC. 2012. Probes for narcotic receptor mediated phenomena. 46. N-substituted-2,3,4,9,10,10a-hexahydro-1H-1,4a-(epiminoethano)phenanthre n-6-and 8-ols -

286
Carbocyclic relatives of f-oxide-bridged phenylmorphans. European Journal of Medicinal Chemistry 58: 557-67
99. Li ZZ, Xu YW, Warner D, Volkow ND. 2012. Alcohol ADME in Primates Studied with Positron Emission Tomography. Plos One 7
100. Lin X, Parisiadou L, Sgobio C, Liu GX, Yu J, Sun LX, Shim H, Gu XL, Luo J, Long CX, Ding JH, Mateo Y, Sullivan PH, Wu LG, Goldstein DS, Lovinger D, Cai HB. 2012. Conditional Expression of Parkinson's Disease-Related Mutant alpha-Synuclein in the Midbrain Dopaminergic Neurons Causes Progressive Neurodegeneration and Degradation of Transcription Factor Nuclear Receptor Related 1. Journal of Neuroscience 32: 9248-64
101. Lin YH, Loewke JD, Hyun DY, Leazer J, Hibbeln JR. 2012. Fast Transmethylation of Serum Lipids Using Microwave Irradiation. Lipids 47: 1109-17
102. Lin YH, Salem N, Wells EM, Zhou WY, Loewke JD, Brown JA, Lands WEM, Goldman LR, Hibbeln JR. 2012. Automated High-Throughput Fatty Acid Analysis of Umbilical Cord Serum and Application to an Epidemiological Study. Lipids 47: 527-39
103. Lindell SG, Yuan QP, Zhou ZF, Goldman D, Thompson RC, Lopez JF, Suomi SJ, Higley JD, Barr CS. 2012. The serotonin transporter gene is a substrate for age and stress dependent epigenetic regulation in rhesus macaque brain: Potential roles in genetic selection and Gene x Environment interactions. Development and Psychopathology 24: 1391-400
104. Litten RZ, Egli M, Heilig M, Cui CH, Fertig JB, Ryan ML, Falk DE, Moss H, Huebner R, Noronha A. 2012. Medications development to treat alcohol dependence: a vision for the next decade. Addiction Biology 17: 513-27
105. Liu J, Zhou L, Xiong KM, Godlewski G, Mukhopadhyay B, Tam J, Yin S, Gao P, Shan X, Pickel J, Bataller R, O'Hare J, Scherer T, Buettner C, Kunos G. 2012. Hepatic Cannabinoid Receptor-1 Mediates Diet-Induced Insulin Resistance via Inhibition of Insulin Signaling and Clearance in Mice. Gastroenterology 142: 1218-+
106. Logan J, Carruthers NI, Letavic MA, Sands S, Jiang XH, Shea C, Muench L, Xu YW, Carter P, King P, Fowler JS. 2012. Blockade of the brain histamine H3 receptor by JNJ-39220675: preclinical PET studies with C-11 GSK189254 in anesthetized baboon. Psychopharmacology 223: 447-55
107. Lopes M, Olfson M, Rabkin J, Hasin DS, Alegria AA, Lin KH, Grant BF, Blanco C. 2012. Gender, HIV Status, and Psychiatric Disorders: Results From the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Clinical Psychiatry 73: 384-91
108. Love TM, Enoch MA, Hodgkinson CA, Pecina M, Mickey B, Koeppe RA, Stohler CS, Goldman D, Zubieta JK. 2012. Oxytocin Gene Polymorphisms Influence Human Dopaminergic Function in a Sex-Dependent Manner. Biological Psychiatry 72: 198-206
109. Lovinger DM. 2012. New twist on orphan receptor GPR88 function. Nature Neuroscience 15: 1469-70
110. Lovinger DM. 2012. Young investigators stress alcohol-induced neuroadaptations in extended amygdala. Alcohol 46: 299-300
111. Lu VB, Smith PA, Rashiq S. 2012. The excitability of dorsal horn neurons is affected by cerebrospinal fluid from humans with osteoarthritis. Canadian Journal of Physiology and Pharmacology 90: 783-90
112. Lupachyk S, Stavniichuk R, Komissarenko JI, Drel VR, Obrosov AA, El-Remessy AB, Pacher P, Obrosova IG. 2012. Na+/H+-exchanger-1 inhibition counteracts diabetic cataract formation and retinal oxidative-nitrative stress and apoptosis. International Journal of Molecular Medicine 29: 989-98

287
113. Magnusson A, Lundholm C, Goransson M, Copeland W, Heilig M, Pedersen NL. 2012. Familial influence and childhood trauma in female alcoholism. Psychological Medicine 42: 381-9
114. Maroun M, Kavushansky A, Holmes A, Wellman C, Motanis H. 2012. Enhanced Extinction of Aversive Memories by High-Frequency Stimulation of the Rat Infralimbic Cortex. Plos One 7
115. Marsh AA, Yu HH, Pine DS, Gorodetsky EK, Goldman D, Blair RJR. 2012. The influence of oxytocin administration on responses to infant faces and potential moderation by OXTR genotype. Psychopharmacology 224: 469-76
116. Merritt P, Stangl B, Hirshman E, Verbalis J. 2012. Administration of dehydroepiandrosterone (DHEA) increases serum levels of androgens and estrogens but does not enhance short-term memory in post-menopausal women. Brain Research 1483: 54-62
117. Michaelides M, Thanos PK, Volkow ND, Wang GJ. 2012. Dopamine-related frontostriatal abnormalities in obesity and binge-eating disorder: Emerging evidence for developmental psychopathology. International Review of Psychiatry 24: 211-8
118. Mickey BJ, Sanford BJ, Love TM, Shen PH, Hodgkinson CA, Stohler CS, Goldman D, Zubieta JK. 2012. Striatal Dopamine Release and Genetic Variation of the Serotonin 2C Receptor in Humans. Journal of Neuroscience 32: 9344-50
119. Mitchell DC, Niu SL, Litman BJ. 2012. Quantifying the differential effects of DHA and DPA on the early events in visual signal transduction. Chemistry and Physics of Lipids 165: 393-400
120. Moeller SJ, Hajcak G, Parvaz MA, Dunning JP, Volkow ND, Goldstein RZ. 2012. Psychophysiological prediction of choice: relevance to insight and drug addiction. Brain 135: 3481-94
121. Moeller SJ, Tomasi D, Honorio J, Volkow ND, Goldstein RZ. 2012. Dopaminergic involvement during mental fatigue in health and cocaine addiction. Translational Psychiatry 2
122. Moeller SJ, Tomasi D, Woicik PA, Maloney T, Alia-Klein N, Honorio J, Telang F, Wang GJ, Wang RL, Sinha R, Carise D, Astone-Twerell J, Bolger J, Volkow ND, Goldstein RZ. 2012. Enhanced midbrain response at 6-month follow-up in cocaine addiction, association with reduced drug-related choice. Addiction Biology 17: 1013-25
123. Molander A, Vengeliene V, Heilig M, Wurst W, Deussing JM, Spanagel R. 2012. Brain-Specific Inactivation of the Crhr1 Gene Inhibits Post-Dependent and Stress-Induced Alcohol Intake, but Does Not Affect Relapse-Like Drinking. Neuropsychopharmacology 37: 1047-56
124. Molica F, Matter CM, Burger F, Pelli G, Lenglet S, Zimmer A, Pacher P, Steffens S. 2012. Cannabinoid receptor CB2 protects against balloon-induced neointima formation. American Journal of Physiology-Heart and Circulatory Physiology 302: H1064-H74
125. Momenan R, Steckler LE, Saad ZS, van Rafelghem S, Kerich MJ, Hommer DW. 2012. Effects of alcohol dependence on cortical thickness as determined by magnetic resonance imaging. Psychiatry Research-Neuroimaging 204: 101-11
126. Moreno C, Hasin DS, Arango C, Oquendo MA, Vieta E, Liu SM, Grant BF, Blanco C. 2012. Depression in bipolar disorder versus major depressive disorder: results from the National Epidemiologic Survey on Alcohol and Related Conditions. Bipolar Disorders 14: 271-82
127. Morozova TV, Goldman D, Mackay TFC, Anholt RRH. 2012. The genetic basis of alcoholism: multiple phenotypes, many genes, complex networks. Genome Biology 13

288
128. Mukhopadhyay P, Das S, Ahsan MK, Otani H, Das DK. 2012. Modulation of microRNA 20b with resveratrol and longevinex is linked with their potent anti-angiogenic action in the ischaemic myocardium and synergestic effects of resveratrol and γ-tocotrienol. Journal of Cellular and Molecular Medicine 16: 2504-17
129. Mukhopadhyay P, Horvath B, Zsengeller Z, Batkai S, Cao ZX, Kechrid M, Holovac E, Erdelyi K, Tanchian G, Liaudet L, Stillman IE, Joseph J, Kalyanaraman B, Pacher P. 2012. Mitochondrial reactive oxygen species generation triggers inflammatory response and tissue injury associated with hepatic ischemia-reperfusion: Therapeutic potential of mitochondrially targeted antioxidants. Free Radical Biology and Medicine 53: 1123-38
130. Mukhopadhyay P, Horvath B, Zsengeller Z, Zielonka J, Tanchian G, Holovac E, Kechrid M, Patel V, Stillman IE, Parikh SM, Joseph J, Kalyanaraman B, Pacher P. 2012. Mitochondrial-targeted antioxidants represent a promising approach for prevention of cisplatin-induced nephropathy. Free Radical Biology and Medicine 52: 497-506
131. Murnane KS, Kimmel HL, Rice KC, Howell LL. 2012. The neuropharmacology of prolactin secretion elicited by 3,4-methylenedioxymethamphetamine ("ecstasy"): A concurrent microdialysis and plasma analysis study. Hormones and Behavior 61: 181-90
132. Nakayama S, Ikenaga T, Kawakami K, Ono F, Hatta K. 2012. Transgenic line with gal4 insertion useful to study morphogenesis of craniofacial perichondrium, vascular endothelium-associated cells, floor plate, and dorsal midline radial glia during zebrafish development. Development Growth & Differentiation 54: 202-15
133. Negus SS, O'Connell R, Morrissey E, Cheng KJ, Rice KC. 2012. Effects of Peripherally Restricted kappa Opioid Receptor Agonists on Pain-Related Stimulation and Depression of Behavior in Rats. Journal of Pharmacology and Experimental Therapeutics 340: 501-9
134. Negus SS, Rosenberg MB, Altarifi AA, O'Connell RH, Folk JE, Rice KC. 2012. Effects of the Delta Opioid Receptor Agonist SNC80 on Pain-Related Depression of Intracranial Self-Stimulation (ICSS) in Rats. Journal of Pain 13: 317-27
135. Nguyen TA, Sarkar P, Veetil JV, Koushik SV, Vogel SS. 2012. Fluorescence Polarization and Fluctuation Analysis Monitors Subunit Proximity, Stoichiometry, and Protein Complex Hydrodynamics. Plos One 7
136. Oleski J, Cox BJ, Robinson J, Grant B. 2012. The predictive validity of cluster C personality disorders on the persistence of major depression in the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of Personality Disorders 26: 322-33
137. Olfson M, Liu SM, Grant BF, Blanco C. 2012. Influence of Comorbid Mental Disorders on Time to Seeking Treatment for Major Depressive Disorder. Medical Care 50: 227-32
138. Pacher P, Mackie K. 2012. Interplay of cannabinoid 2 (CB2) receptors with nitric oxide synthases, oxidative and nitrative stress, and cell death during remote neurodegeneration. Journal of Molecular Medicine-Jmm 90: 347-51
139. Papke RL, Ono F, Stokes C, Urban JM, Boyd RT. 2012. The nicotinic acetylcholine receptors of zebrafish and an evaluation of pharmacological tools used for their study. Biochemical Pharmacology 84: 352-65
140. Pardini M, Krueger F, Koenigs M, Raymont V, Hodgkinson C, Zoubak S, Goldman D, Grafman J. 2012. Fatty-acid amide hydrolase polymorphisms and post-traumatic stress disorder after penetrating brain injury. Translational Psychiatry 2
141. Park JH, Johansson HE, Aoki H, Huang BX, Kim HY, Ganoza MC, Park MH. 2012. Post-translational Modification by beta-Lysylation Is Required for Activity of Escherichia coli Elongation Factor P (EF-P). Journal of Biological Chemistry 287: 2579-90

289
142. Park JY, Ikeda H, Ikenaga T, Ono F. 2012. Acetylcholine Receptors Enable the Transport of Rapsyn from the Golgi Complex to the Plasma Membrane. J Neurosci 32: 7356-63
143. Parvaz MA, Maloney T, Moeller SJ, Woicik PA, Alia-Klein N, Telang F, Wang GJ, Squires NK, Volkow ND, Goldstein RZ. 2012. Sensitivity to monetary reward is most severely compromised in recently abstaining cocaine addicted individuals: A cross-sectional ERP study. Psychiatry Research-Neuroimaging 203: 75-82
144. Parylak SL, Cottone P, Sabino V, Rice KC, Zorrilla EP. 2012. Effects of CB1 and CRF1 receptor antagonists on binge-like eating in rats with limited access to a sweet fat diet: Lack of withdrawal-like responses. Physiology & Behavior 107: 231-42
145. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2012. Psychiatric Comorbidity of Full and Partial Posttraumatic Stress Disorder Among Older Adults in the United States: Results From Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. American Journal of Geriatric Psychiatry 20: 380-90
146. Pietrzak RH, Goldstein RB, Southwick SM, Grant BF. 2012. Physical Health Conditions Associated with Posttraumatic Stress Disorder in US Older Adults: Results from Wave 2 of the National Epidemiologic Survey on Alcohol and Related Conditions. Journal of the American Geriatrics Society 60: 296-303
147. Rajesh M, Batkai S, Kechrid M, Mukhopadhyay P, Lee WS, Horvath B, Holovac E, Cinar R, Liaudet L, Mackie K, Hasko G, Pacher P. 2012. Cannabinoid 1 Receptor Promotes Cardiac Dysfunction, Oxidative Stress, Inflammation, and Fibrosis in Diabetic Cardiomyopathy. Diabetes 61: 716-27
148. Ramsden CE, Ringel A, Feldstein AE, Taha AY, MacIntosh BA, Hibbeln JR, Majchrzak-Hong SF, Faurot KR, Rapoport SI, Cheon Y, Chung YM, Berk M, Mann JD. 2012. Lowering dietary linoleic acid reduces bioactive oxidized linoleic acid metabolites in humans. Prostaglandins Leukotrienes and Essential Fatty Acids 87: 135-41
149. Ray LA, Barr CS, Blendy JA, Oslin D, Goldman D, Anton RF. 2012. The Role of the Asn40Asp Polymorphism of the Mu Opioid Receptor Gene (OPRM1) on Alcoholism Etiology and Treatment: A Critical Review. Alcoholism-Clin Exper Res 36: 385-94
150. Ren HG, Du CW, Park K, Volkow ND, Pan YT. 2012. Quantitative imaging of red blood cell velocity in vivo using optical coherence Doppler tomography. Applied Physics Letters 100
151. Robinson JE, Fish EW, Krouse MC, Thorsell A, Heilig M, Malanga CJ. 2012. Potentiation of brain stimulation reward by morphine: effects of neurokinin-1 receptor antagonism. Psychopharmacology 220: 215-24
152. Roiser JP, Levy J, Fromm SJ, Goldman D, Hodgkinson CA, Hasler G, Sahakian BJ, Drevets WC. 2012. Serotonin transporter genotype differentially modulates neural responses to emotional words following tryptophan depletion in patients recovered from depression and healthy volunteers. Journal of Psychopharmacology 26: 1434-42
153. Roy A, Hodgkinson CA, DeLuca V, Goldman D, Enoch MA. 2012. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. Journal of Psychiatric Research 46: 72-9
154. Saha TD, Compton WM, Chou SP, Smith S, Ruan WJ, Huang B, Pickering RP, Grant BF. 2012. Analyses related to the development of DSM-5 criteria for substance use related disorders 1. Toward amphetamine, cocaine and prescription drug use disorder continua using Item Response Theory. Drug and Alcohol Dependence 122: 38-46
155. Saha TD, Harford T, Goldstein RB, Kerridge BT, Hasin D. 2012. Relationship of Substance Abuse to Dependence in the U.S. General Population. Journal of Studies on Alcohol and Drugs 73: 368-78

290
156. Sawyer EK, Mun J, Nye JA, Kimmel HL, Voll RJ, Stehouwer JS, Rice KC, Goodman MM, Howell LL. 2012. Neurobiological Changes Mediating the Effects of Chronic Fluoxetine on Cocaine Use. Neuropsychopharmacology 37: 1816-24
157. Schank JR, Goldstein AL, Rowe KE, King CE, Marusich JA, Wiley JL, Carroll FI, Thorsell A, Heilig M. 2012. The kappa opioid receptor antagonist JDTic attenuates alcohol seeking and withdrawal anxiety. Addiction Biology 17: 634-47
158. Schank JR, Ryabinin AE, Giardino WJ, Ciccocioppo R, Heilig M. 2012. Stress-Related Neuropeptides and Addictive Behaviors: Beyond the Usual Suspects. Neuron 76: 192-208
159. Serafine KM, Briscione MA, Rice KC, Riley AL. 2012. Dopamine mediates cocaine-induced conditioned taste aversions as demonstrated with cross-drug preexposure to GBR 12909. Pharmacology Biochemistry and Behavior 102: 269-74
160. Snell LD, Ramchandani VA, Saba L, Herion D, Heilig M, George DT, Pridzun L, Helander A, Schwandt ML, Phillips MJ, Hoffman PL, Tabakoff B, Trait WISS. 2012. The Biometric Measurement of Alcohol Consumption. Alcoholism-Clinical and Experimental Research 36: 332-41
161. Soubias O, Gawrisch K. 2012. The role of the lipid matrix for structure and function of the GPCR rhodopsin. Biochimica Et Biophysica Acta-Biomembranes 1818: 234-40
162. Srivastava S, Kashiwaya Y, Chen XS, Geiger JD, Pawlosky R, Veech RL. 2012. Microwave irradiation decreases ATP, increases free Mg2+ , and alters in vivo intracellular reactions in rat brain. Journal of Neurochemistry 123: 668-75
163. Srivastava S, Kashiwaya Y, King MT, Baxa U, Tam J, Niu G, Chen XY, Clarke K, Veech RL. 2012. Mitochondrial biogenesis and increased uncoupling protein 1 in brown adipose tissue of mice fed a ketone ester diet. Faseb Journal 26: 2351-62
164. Steer CD, Hibbeln JR, Golding J, Smith GD. 2012. Polyunsaturated fatty acid levels in blood during pregnancy, at birth and at 7 years: their associations with two common FADS2 polymorphisms. Human Molecular Genetics 21: 1504-12
165. Steffens S, Pacher P. 2012. Targeting cannabinoid receptor CB2 in cardiovascular disorders: promises and controversies. British Journal of Pharmacology 167: 313-23
166. Sugaya L, Hasin DS, Olfson M, Lin KH, Grant BF, Blanco C. 2012. Child physical abuse and adult mental health: A national study. Journal of Traumatic Stress 25: 384-92
167. Szabo C, Pacher P. 2012. The Outsiders: Emerging Roles of Ectonucleotidases in Inflammation. Science Translational Medicine 4
168. Tam J, Cinar R, Liu J, Godlewski G, Wesley D, Jourdan T, Szanda G, Mukhopadhyay B, Chedester L, Liow JS, Innis RB, Cheng KJ, Rice KC, Deschamps JR, Chorvat RJ, McElroy JF, Kunos G. 2012. Peripheral Cannabinoid-1 Receptor Inverse Agonism Reduces Obesity by Reversing Leptin Resistance. Cell Metabolism 16: 167-79
169. Thambisetty M, Gallardo KA, Liow JS, Beason-Held LL, Umhau JC, Bhattacharjee AK, Der M, Herscovitch P, Rapoport JL, Rapoport SI. 2012. The utility of C-11-arachidonate PET to study in vivo dopaminergic neurotransmission in humans. Journal of Cerebral Blood Flow and Metabolism 32: 676-84
170. Thanos PK, Subrize M, Delis F, Cooney RN, Culnan D, Sun MJ, Wang GJ, Volkow ND, Hajnal A. 2012. Gastric Bypass Increases Ethanol and Water Consumption in Diet-Induced Obese Rats. Obesity Surgery 22: 1884-92
171. Tomasi D, Volkow ND. 2012. Resting functional connectivity of language networks: characterization and reproducibility. Molecular Psychiatry 17: 841-54
172. Tomasi D, Volkow ND. 2012. Laterality Patterns of Brain Functional Connectivity: Gender Effects. Cerebral Cortex 22: 1455-62

291
173. Tomasi D, Volkow ND. 2012. Aging and functional brain networks. Molecular Psychiatry 17: 549-58
174. Tomasi D, Volkow ND. 2012. Gender differences in brain functional connectivity density. Human Brain Mapping 33: 849-60
175. Tomasi D, Volkow ND. 2012. Abnormal Functional Connectivity in Children with Attention-Deficit/Hyperactivity Disorder. Biological Psychiatry 71: 443-50
176. Umhau JC, Garg K, Woodward AM. 2012. Dietary Supplements and Their Future in Health Care: Commentary on Draft Guidelines Proposed by the Food and Drug Administration. Antioxidants & Redox Signaling 16: 461-2
177. Umhau JC, Trandem K, Shah M, George DT. 2012. The physician's unique role in preventing violence: a neglected opportunity? Bmc Medicine 10
178. Valeri CR, Veech RL. 2012. The unrecognized effects of the volume and composition of the resuscitation fluid used during the administration of blood products. Transfusion and Apheresis Science 46: 121-3
179. Vatsalya V, Issa JE, Hommer DW, Ramchandani VA. 2012. Pharmacodynamic Effects of Intravenous Alcohol on Hepatic and Gonadal Hormones: Influence of Age and Sex. Alcoholism-Clinical and Experimental Research 36: 207-13
180. Veech RL, Valeri CR, VanItallie TB. 2012. The mitochondrial permeability transition pore provides a key to the diagnosis and treatment of traumatic brain injury. Iubmb Life 64: 203-7
181. Vendruscolo LF, Barbier E, Schlosburg JE, Misra KK, Whitfield TW, Logrip ML, Rivier C, Repunte-Canonigo V, Zorrilla EP, Sanna PP, Heilig M, Koob GF. 2012. Corticosteroid-Dependent Plasticity Mediates Compulsive Alcohol Drinking in Rats. Journal of Neuroscience 32: 7563-71
182. Vogel SS, Nguyen TA, van der Meer BW, Blank PS. 2012. The Impact of Heterogeneity and Dark Acceptor States on FRET: Implications for Using Fluorescent Protein Donors and Acceptors. Plos One 7
183. Volkow ND, Tomasi D, Wang GJ, Telang F, Fowler JS, Logan J, Benveniste H, Kim R, Thanos PK, Ferre S. 2012. Evidence That Sleep Deprivation Downregulates Dopamine D2R in Ventral Striatum in the Human Brain. Journal of Neuroscience 32: 6711-7
184. Volkow ND, Wang GJ, Fowler JS, Tomasi D. 2012. Addiction Circuitry in the Human Brain. In Annual Review of Pharmacology and Toxicology, Vol 52, ed. PA Insel, SG Amara, TF Blaschke, pp. 321-36
185. Volkow ND, Warren KR. 2012. Advancing American Indian/Alaska Native Substance Abuse Research. American Journal of Drug and Alcohol Abuse 38: 371-
186. Vukoti K, Kimura T, Macke L, Gawrisch K, Yeliseev A. 2012. Stabilization of Functional Recombinant Cannabinoid Receptor CB2 in Detergent Micelles and Lipid Bilayers. Plos One 7
187. Walogorsky M, Mongeon R, Wen H, Nelson NR, Urban JM, Ono F, Mandel G, Brehm P. 2012. Zebrafish model for congenital myasthenic syndrome reveals mechanisms causal to developmental recovery. Proceedings of the National Academy of Sciences of the United States of America 109: 17711-6
188. Wang D, Wong HK, Zhang L, McAlonan GM, Wang XM, Sze SCW, Feng YB, Zhang ZJ. 2012. Not only dopamine D-2 receptors involved in Peony-Glycyrrhiza Decoction, an herbal preparation against antipsychotic-associated hyperprolactinemia. Progress in Neuro-Psychopharmacology & Biological Psychiatry 39: 332-8
189. Wang GJ, Smith L, Volkow ND, Telang F, Logan J, Tomasi D, Wong CT, Hoffman W, Jayne M, Alia-Klein N, Thanos P, Fowler JS. 2012. Decreased dopamine activity predicts relapse in methamphetamine abusers. Molecular Psychiatry 17: 918-25

292
190. Wang HJ, Gao B, Zakhari S, Nagy LE. 2012. Inflammation in Alcoholic Liver Disease. In Annual Review of Nutrition, Vol 32, ed. RJ Cousins, pp. 343-+
191. Wang J, Cao HC, You CJ, Yuan BF, Bahde R, Gupta S, Nishigori C, Niedernhofer LJ, Brooks PJ, Wang YS. 2012. Endogenous formation and repair of oxidatively induced G 8-5 m T intrastrand cross-link lesion. Nucleic Acids Research 40: 7368-74
192. Wilson ST, Stanley B, Brent DA, Oquendo MA, Huang YY, Haghighi F, Hodgkinson CA, Mann JJ. 2012. Interaction between tryptophan hydroxylase I polymorphisms and childhood abuse is associated with increased risk for borderline personality disorder in adulthood. Psychiatric Genetics 22: 15-24
193. Won YJ, Ono F, Ikeda SR. 2012. Characterization of Na+ and Ca2+ Channels in Zebrafish Dorsal Root Ganglion Neurons. Plos One 7
194. Wu Y, Lousberg EL, Moldenhauer LM, Hayball JD, Coller JK, Rice KC, Watkins LR, Somogyi AA, Hutchinson MR. 2012. Inhibiting the TLR4-MyD88 signalling cascade by genetic or pharmacological strategies reduces acute alcohol-induced sedation and motor impairment in mice. British Journal of Pharmacology 165: 1319-29
195. Xiao CY, Wang RH, Lahusen TJ, Park O, Bertola A, Maruyama T, Reynolds D, Chen Q, Xu XL, Young HA, Chen WJ, Gao B, Deng CX. 2012. Progression of Chronic Liver Inflammation and Fibrosis Driven by Activation of c-JUN Signaling in Sirt6 Mutant Mice. Journal of Biological Chemistry 287: 41903-13
196. Xiong W, Cui TX, Cheng KJ, Yang F, Chen SR, Willenbring D, Guan Y, Pan HL, Ren K, Xu Y, Zhang L. 2012. Cannabinoids suppress inflammatory and neuropathic pain by targeting alpha 3 glycine receptors. Journal of Experimental Medicine 209: 1121-34
197. Xiong W, Wu XW, Lovinger DM, Zhang L. 2012. A Common Molecular Basis for Exogenous and Endogenous Cannabinoid Potentiation of Glycine Receptors. Journal of Neuroscience 32: 5200-8
198. Xu K, Hong KA, Zhou ZF, Hauger RL, Goldman D, Sinha R. 2012. Genetic modulation of plasma NPY stress response is suppressed in substance abuse: Association with clinical outcomes. Psychoneuroendocrinology 37: 554-64
199. Yekkirala AS, Banks ML, Lunzer MM, Negus SS, Rice KC, Portoghese PS. 2012. Clinically Employed Opioid Analgesics Produce Antinociception via mu-delta Opioid Receptor Heteromers in Rhesus Monkeys. Acs Chemical Neuroscience 3: 720-7
200. You CJ, Dai XX, Yuan BF, Wang J, Wang JS, Brooks PJ, Niedernhofer LJ, Wang YS. 2012. A quantitative assay for assessing the effects of DNA lesions on transcription. Nature Chemical Biology 8: 817-22
201. Yuan QP, Zhou ZF, Lindell SG, Higley JD, Ferguson B, Thompson RC, Lopez JF, Suomi SJ, Baghal B, Baker M, Mash DC, Barr CS, Goldman D. 2012. The rhesus macaque is three times as diverse but more closely equivalent in damaging coding variation as compared to the human. Bmc Genetics 13
202. Zoerner AA, Batkai S, Suchy MT, Gutzki FM, Engeli S, Jordan J, Tsikas D. 2012. Simultaneous UPLC-MS/MS quantification of the endocannabinoids 2-arachidonoyl glycerol (2AG), 1-arachidonoyl glycerol (1AG), and anandamide in human plasma: Minimization of matrix-effects, 2AG/1AG isomerization and degradation by toluene solvent extraction. Journal of Chromatography B-Analytical Technologies in the Biomedical and Life Sciences 883: 161-71
203. Zsengeller ZK, Ellezian L, Brown D, Horvath B, Mukhopadhyay P, Kalyanaraman B, Parikh SM, Karumanchi SA, Stillman IE, Pacher P. 2012. Cisplatin Nephrotoxicity Involves Mitochondrial Injury with Impaired Tubular Mitochondrial Enzyme Activity. Journal of Histochemistry & Cytochemistry 60: 521-9
293
Related Documents










