1 Introduction Saurichthys is a highly specialized genus of Triassic actinopetrygian fishes characterized by long and slender body shape. The genus was initially named by Agassiz in 1834 based on a rostrum material from the Middle Triassic of Germany to emphasize its ichthyosaurus-like feature. So far, more than 30 species are attributed to the genus based on materials more than tooth or fragmentary bones (Rieppel, 1985), showing a great morphological diversity and a world-wide distribution since its first appearance. However, most of these species are still poorly known except for a few that are represented by relatively complete materials from the Lower Triassic of Spitzbergen and Madagascar (Stensiö, 1925; Piveteau, 1945; Lehman, 1952; Lehman et al., 1959; Beltan, 1968; Rieppel, 1980), the Middle Triassic of Europe (Griffith, 1959; Rieppel, 1985, 1992) and Australia (Woodward, 1890), and Upper Triassic of Austria (Griffith, 1959, 1962, 1977). Previously Saurichthys was rarely reported in China. There are only two species related to the saurichthyids. One is S. huanshenensis from Hengshan of Shaanxi Province (Chou and Liu, 1957), represented only by a specimen with the caudal region preserved, and the other is Eosaurichthys chaoi from Zhejiang Province based on two specimens assumed to be collected from the Upper Permian Changxing Limestone (Liu and Wei, 1988). Although some authors also mentioned the existence of Saurichthys in some other localities of China (Su and Li, 1983; Chen, 1985; Tong et al., 2007), they did not give any description or figures. In the present paper we report a new species of Saurichthys based on three well-preserved specimens collected from a more than 10-m thick, vertebrate fossiliferous interval near the top of the third member of the Gejiu Formation exposed in the Luoping area of Yunnan Province (Zhang et al., 2008). The complete and good preservation of the specimens allows a thorough taxonomic study and detailed anatomic comparison with other species of the genus. 2 Fossil Locality and Geological Setting The studied material was collected from the upper part of New species of Saurichthys (Actinopterygii: Saurichthyidae) from Middle Triassic (Anisian) of Yunnan Province, China WU Feixiang 1 , SUN Yuanlin 1, * , HAO Weicheng 1 , JIANG Dayong 1 , XU Guanghui 1 , SUN Zuoyu 1 and Andrea TINTORI 2 1 Key Laboratory of Orogenic Belts and Crustal Evolution, School of Earth and Space Sciences, Peking University, Beijing 100871, China 2 Dipartimento di Scienze della Terra, Universita degli Studi di Milano, Via Mangiagalll 34, 20133 Milano, Italy Abstract: Saurichthys dawaziensis sp. nov., from the third member of the Gejiu Formation (Anisian of Middle Triassic) in the vicinity of Luoping, Yunnan Province, is a small to medium- sized saurichthyid fish. It is characterized by the efferent pseudobranchial arteries penetrating on the parasphenoid, the mandible sensory canal with a branch in the angular and the small, ossified haemal arches between pelvic and anal fins without ossified haemal spines. The new species is more derived than the Lower Triassic species in that it has a relatively narrow postorbital region, short preopercular, single ossification of the sclerotic ring, small haemal arches between the pelvic fins and the anal fin without haemal spines, and the absence of quadratojugal. It is more primitive than the Middle and Upper Triassic Saurichthys in that it has fringing fulcra and numerous segments in the unpaired fins and the presence of both prae- and post-zygapophyses in the neural arches. Some anatomic changes related to the vertebral column, unpaired fins and squamation of the genus are also discussed. Key words: Actinopterygian fishes, Saurichthyidae, Saurichthys, Middle Triassic, Yunnan Vol. 83 No. 3 pp. 801–840 ACTA GEOLOGICA SINICA June 2009 2009-4-10打印 * Corresponding author. E-mail: [email protected] 完成作者一校并发出二校。首先请作者签好版权转让协议(见邮件附件),把全部作者名称,文章名称填好,全部作者签字即可(如果 第一作者出差,请其他作者代办此事),扫描后随二校稿一起发送过来。请作者认真校对图件和表铬,确保图件和表格没有被遗漏,同 时图件也和图名对应,文责自负。请作者补充我刊近期发表的参考文献1-2篇具体参考邮件附件。

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 Introduction
Saurichthys is a highly specialized genus of Triassic actinopetrygian fishes characterized by long and slender body shape. The genus was initially named by Agassiz in 1834 based on a rostrum material from the Middle Triassic of Germany to emphasize its ichthyosaurus-like feature. So far, more than 30 species are attributed to the genus based on materials more than tooth or fragmentary bones (Rieppel, 1985), showing a great morphological diversity and a world-wide distribution since its first appearance. However, most of these species are still poorly known except for a few that are represented by relatively complete materials from the Lower Triassic of Spitzbergen and Madagascar (Stensiö, 1925; Piveteau, 1945; Lehman, 1952; Lehman et al., 1959; Beltan, 1968; Rieppel, 1980), the Middle Triassic of Europe (Griffith, 1959; Rieppel, 1985, 1992) and Australia (Woodward, 1890), and Upper Triassic of Austria (Griffith, 1959, 1962, 1977). Previously Saurichthys was rarely reported in China. There are only two species related to the saurichthyids. One is S.
huanshenensis from Hengshan of Shaanxi Province (Chou and Liu, 1957), represented only by a specimen with the caudal region preserved, and the other is Eosaurichthys chaoi from Zhejiang Province based on two specimens assumed to be collected from the Upper Permian Changxing Limestone (Liu and Wei, 1988). Although some authors also mentioned the existence of Saurichthys in some other localities of China (Su and Li, 1983; Chen, 1985; Tong et al., 2007), they did not give any description or figures.
In the present paper we report a new species of Saurichthys based on three well-preserved specimens collected from a more than 10-m thick, vertebrate fossiliferous interval near the top of the third member of the Gejiu Formation exposed in the Luoping area of Yunnan Province (Zhang et al., 2008). The complete and good preservation of the specimens allows a thorough taxonomic study and detailed anatomic comparison with other species of the genus. 2 Fossil Locality and Geological Setting
The studied material was collected from the upper part of
New species of Saurichthys (Actinopterygii: Saurichthyidae) from Middle
Triassic (Anisian) of Yunnan Province, China
WU Feixiang1, SUN Yuanlin1, *, HAO Weicheng1, JIANG Dayong1, XU Guanghui1, SUN Zuoyu1 and Andrea TINTORI2
1 Key Laboratory of Orogenic Belts and Crustal Evolution, School of Earth and Space Sciences, Peking University, Beijing 100871, China 2 Dipartimento di Scienze della Terra, Universita degli Studi di Milano, Via Mangiagalll 34, 20133 Milano, Italy Abstract: Saurichthys dawaziensis sp. nov., from the third member of the Gejiu Formation (Anisian of Middle Triassic) in the vicinity of Luoping, Yunnan Province, is a small to medium- sized saurichthyid fish. It is characterized by the efferent pseudobranchial arteries penetrating on the parasphenoid, the mandible sensory canal with a branch in the angular and the small, ossified haemal arches between pelvic and anal fins without ossified haemal spines. The new species is more derived than the Lower Triassic species in that it has a relatively narrow postorbital region, short preopercular, single ossification of the sclerotic ring, small haemal arches between the pelvic fins and the anal fin without haemal spines, and the absence of quadratojugal. It is more primitive than the Middle and Upper Triassic Saurichthys in that it has fringing fulcra and numerous segments in the unpaired fins and the presence of both prae- and post-zygapophyses in the neural arches. Some anatomic changes related to the vertebral column, unpaired fins and squamation of the genus are also discussed. Key words: Actinopterygian fishes, Saurichthyidae, Saurichthys, Middle Triassic, Yunnan
Vol. 83 No. 3 pp. 801–840 ACTA GEOLOGICA SINICA June 2009
2009-4-10打印
* Corresponding author. E-mail: [email protected]
完成作者一校并发出二校。首先请作者签好版权转让协议(见邮件附件),把全部作者名称,文章名称填好,全部作者签字即可(如果
第一作者出差,请其他作者代办此事),扫描后随二校稿一起发送过来。请作者认真校对图件和表铬,确保图件和表格没有被遗漏,同
时图件也和图名对应,文责自负。请作者补充我刊近期发表的参考文献1-2篇具体参考邮件附件。

802 New species of Saurichthys from Middle Triassic of Yunnan Province Wu et al.
the more than 10-m thick fossiliferous interval of thin-bedded marly limestone near the top of the third member of the Gejiu Formation exposed around Dawazi Villege, Luoping, Yunnan Province (Fig. 1), where abundant fossil fishes and reptiles exist.
The Gejiu Formation is a local lithostratigraphic unit applied formerly for the interval exposed in Yunnan Province ranging from the upper Lower Triassic to the upper Middle Triassic and its third member roughly corresponds with the middle and upper members of the Guanling Formation used in Guizhou Province. The conodont Nicoraella kockeli is recovered in the fossiliferous interval (Fig. 1), suggesting an age of the Anisian of the Middle Triassic. Thus, this new discovered fossil marine vertebrate fauna is of an age very close to that of the Panxian Fauna of Guizhou Province (Sun et al., 2006). 3 Systematic Paleontology Subclass Actinopterygii Cope, 1887 Order Saurichthyiformes Aldinger, 1937 Family Saurichthyidae Stensiö, 1925 Genus Saurichthys Agassiz, 1834 Type specie: Saurichthys apicalis Agassize, 1834
Saurichthys dawaziensis sp. nov. (Fig. 2-4; Plate I)
Diagnosis: A small to medium-sized species of
Saurichthys characterized by the maxilla with a very high trapezoid posterior lobe, a relatively slender boomerang-shaped preopercular with its posterior portion almost vertically against the posterior margin of the maxilla; supraorbital probably present; the efferent pseudobranchial artery penetrating the parasphenoid immediately anterior to the ascending process; the opercular trapezoid-shaped ornamented with characteristic vertical striae; the lepidotrichia of the median fins with numerous segmentation; the large pelvic plates are well-ossified with an evident hook-like structure at the anterolateral end; Mid-dorsal scales lanceolate in shape anterior to the dorsal fin and gradually becoming coulter-shaped in the caudal region. Neural arches with both prae- and postzygapophysis, the haemal arches ossified but without haemal spines in the region between the pelvic and anal fins.
Holotype: GMPKU-P-1524. Other referred specimens: GMPKU-P-1525 and 1526. Type locality: around Dawazi Village, Luoping County,
Yunnan Province, China. Type horizon: Upper member of the Guanling
Formation at the type locality, within the conodont
Nicoraella kockeli Zone, Pelsonian substage of Anisian, Middle Triassic.
Etymology: Refers to the name of the fossil locality. 4 Description
The body is very long and slender, about 38 cm in the
holotype (Plate I-1). The head occupies about 32% of the standard length, while the rostrum is 58.5% of the skull length. The pectoral fin lies close behind the operculum. The pelvic fins are placed at about the mid point between the operculum and caudal fin. The dorsal and anal fins are symmetrically arranged, slightly closer to the caudal fin than to the ventral fins. The caudal fin is abbreviated diphycercal. 4.1 Skull
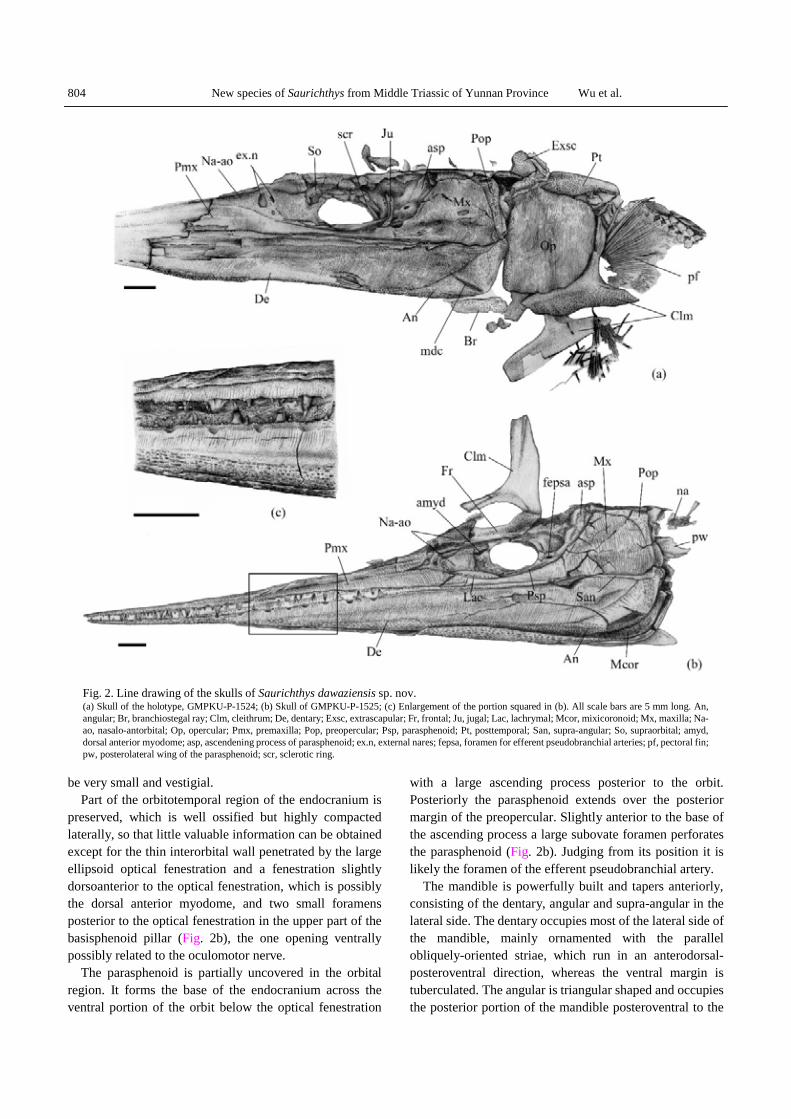
The skull is conspicuously prolonged into a long, sharply pointed beak, which involves the anterior part of the jaws (Plate I–5). Owing to the state of preservation, it is impossible to clearly distinguish the dermal bones in the skull roof except one of the paired medial extrascapulars, which is completely exposed posterior to the dermopterotic in the holotype; however, most of the elements forming the lateral side of the skull can be recognized and shown in Fig. 2a, b.
The rostrum is mainly composed of the premaxilla, which are extensively extended anteriorly and ornamented with tubercles in their anterior one-third length and parallel anterodorsally-obliqued striae on the remaining part. Posteriorly the premaxilla terminates slightly in front of the ventroanterior corner of the orbit. It is in contact with the nasal-antorbital posterodorsally and with the maxilla posteriorly. The nasal-antorbital is a roughly triangular bone with the paired subovate external nares in the same way as in other species of Saurichthys, the anterior one is much larger than the posterior, and is separated by the supraorbital sensory cannel that passes down to meet the infraorbital sensory canal in the internarial commissure, where the infraorbital sensory canal branches off and continues toward the anterior tip of the premaxilla.
Posterior to the nasal-antorbital is the large, subovate orbit, which is confined anteriorly by the nasal-antorbital and ventrally by the maxilla (Fig. 2a, b; Plate I-1, 5). Two infraorbital bones, lacrymal and jugal, can be recognized around the ventral and posterior margin of the orbit. The lanceolate lacrymal is wedged anteriorly between the premaxilla and nasalo-antorbital and posteriorly it confines the anteroventral margin of the orbit. Different to other dermal bones of the skull, the lachrymal is free of ornamentation except for a series of openings of the infraorbital sensory canal. The jugal is crescent-shaped,

Vol. 83 No. 3 ACTA GEOLOGICA SINICA June 2009 803
forming the posteroventral and much of the posterior margin of the orbit (Fig. 2b), with an obvious groove of the infraorbital sensory canal close to its anterior margin and some tubercles on the remaining part. In the holotype, the bone attributed to the supraorbital is located near the anterodorsal margin of the orbit (Fig. 2a). It is crescent-shaped and runs about half of the orbital width with tubercled ornaments. The dermosphnotic in both specimens with the skull is not recognized due to damage in the region posterodorsal to the orbit.
Trace of the sclerotic ring is observed in two specimens. In the holotype, a piece of the sclerotic is, making up about half of the circumference of the orbit, ranging along the dorsal and posterior margin of the optical fenestration in the interorbital wall (Fig. 2a) and in the other specimen the sclerotic ring is displaced some distance dorsoanterior to the orbit and the preserved part shows an ideal arc roughly making up the one-third circumference of the orbit. It is likely that the sclerotic ring in the new species consists of a single ossification, The similar case is known in Saurichthys curionii, Saurichthys striolatus and Saurichthys krambergeri (Rieppel, 1985; Griffith, 1959, 1962), but it is of eight ossified elements in Saurichthys
madagascariensis (Rieppel, 1980). Posterior to the orbital region, the maxilla and
preopercular are the main dermal bones forming the cheek (Fig. 2a, b). The maxilla has a slender anterior part extending forwards below the orbit to meet the premaxilla anteriorly and a highly expanded trapezoid posterior lobe behind the orbit, which is much higher than that of other known species of Saurichthys. The ornamentation of the maxilla consists of the lines radiating from the ossification center levelling with the posterior limit of the orbit and nearly vertical striae on the posterior portion. The maxilla is firmly attached to the preopercular posteriorly and dorsally. The preopercular is boomerang-shaped with radiating ornamentation from the posterodorsal corner and some tubercles on the dorsal margin. Compared with that of other well-known species of Saurichthys, the preopercular of the new species is much more slender in proportion to the size of the maxilla and the posterior part of the preopercular is almost vertical against the posterior margin of the maxilla. A length of the openings of the sensory canal in the preopercular can be observed in the posterior margin of the vertical part of this bone. No quadratojugal can be recognized in the material. If this bone is present, it should
Dawazi
Xinzhai
GuishanWaina
Daheishan
2 km
limestone marlylimestone
chertlimestone
dolostonedolomiticlimestone
bentonitelayer
vertebratefossils
areniticdolostone
S. dawaziensis sp. nov
D50
D37
D54
Nicorella kockeli
Maanshan
10 m 0.5
mFig. 1. Sketched locality map and lithologic sequence of the third member of the Gejiu Formation at Dawazi, Luoping, Yunnan.
804 New species of Saurichthys from Middle Triassic of Yunnan Province Wu et al.
be very small and vestigial. Part of the orbitotemporal region of the endocranium is
preserved, which is well ossified but highly compacted laterally, so that little valuable information can be obtained except for the thin interorbital wall penetrated by the large ellipsoid optical fenestration and a fenestration slightly dorsoanterior to the optical fenestration, which is possibly the dorsal anterior myodome, and two small foramens posterior to the optical fenestration in the upper part of the basisphenoid pillar (Fig. 2b), the one opening ventrally possibly related to the oculomotor nerve.
The parasphenoid is partially uncovered in the orbital region. It forms the base of the endocranium across the ventral portion of the orbit below the optical fenestration
with a large ascending process posterior to the orbit. Posteriorly the parasphenoid extends over the posterior margin of the preopercular. Slightly anterior to the base of the ascending process a large subovate foramen perforates the parasphenoid (Fig. 2b). Judging from its position it is likely the foramen of the efferent pseudobranchial artery.
The mandible is powerfully built and tapers anteriorly, consisting of the dentary, angular and supra-angular in the lateral side. The dentary occupies most of the lateral side of the mandible, mainly ornamented with the parallel obliquely-oriented striae, which run in an anterodorsal-posteroventral direction, whereas the ventral margin is tuberculated. The angular is triangular shaped and occupies the posterior portion of the mandible posteroventral to the
Fig. 2. Line drawing of the skulls of Saurichthys dawaziensis sp. nov. (a) Skull of the holotype, GMPKU-P-1524; (b) Skull of GMPKU-P-1525; (c) Enlargement of the portion squared in (b). All scale bars are 5 mm long. An, angular; Br, branchiostegal ray; Clm, cleithrum; De, dentary; Exsc, extrascapular; Fr, frontal; Ju, jugal; Lac, lachrymal; Mcor, mixicoronoid; Mx, maxilla; Na-ao, nasalo-antorbital; Op, opercular; Pmx, premaxilla; Pop, preopercular; Psp, parasphenoid; Pt, posttemporal; San, supra-angular; So, supraorbital; amyd, dorsal anterior myodome; asp, ascendening process of parasphenoid; ex.n, external nares; fepsa, foramen for efferent pseudobranchial arteries; pf, pectoral fin; pw, posterolateral wing of the parasphenoid; scr, sclerotic ring.

Vol. 83 No. 3 ACTA GEOLOGICA SINICA June 2009 805
dentary. It extends forward along the ventral margin of the dentary beyond the orbit. The angular is decorated with numeous shallow grooves radiating from the posteroventral corner in its lateral surface and with tubercles near its ventral margin. The slender supra-angular forms the posterodorsal portion of the mandible and tapers anteriorly not beyond the level of the posterior margin of the orbit (Fig. 2b). It is in contact with the dentary anteroventrally and with the angular posteroventrally. Unlike the dentary and angular, it is smooth on the surface. The mandible sensory canal occurs as very fine pores along the ventral margin of the angular. It is interesting that the mandible sensory canal gives a branch in the angular from the posteroventral corner, which is anterodorsally directed and restricted between two parallel and closely-spaced ridges. No other species of the genus is known to have such a feature. In saurichthyids, previously only in Acidorhynchus it is known that the mandible sensory canal branches in the angular but it has many branches (Gardiner, 1960; Thies, 1985)
The medial side of the posteroventral corner of the right mandible is partially exposed in the specimen GMPKU-P-1525 (Fig. 2b), where a large piece of dermal bone is in contact with the medial edge of the angular ventrally and posteriorly but its exact extension is not clear due to the overlapping of the left mandible. Its surface is decorated with some irregularly-shaped low ridges radiating from the posteroventral corner. In Saurichthys hamiltoni and S. curionii this bone was called the mixicoronoid and covers the medial side of the mandible considerably (Stensiö, 1925; Rieppel, 1985); however, the position of the radiation centers is variable in these species.
The dentition can be observed in the labial edges of the upper and lower jaws. In the anterior part of the jaw, the dentition consists of one row of main teeth intercalated with some tiny ones in each side. The main teeth are conical, about 0.5–1.2 mm wide at the base and 1.3–2.3 mm high, with height/width ratio ranging from 1.9 to 2.6. Each tooth is composed of a smooth, transparent apical cap distally and a weakly striated basal portion. On the labial edge in the middle part of the dentary, at least eight distinct depressions are very remarkable into which the apices of the main teeth of the upper jaw fitted in occlusion (Fig. 2c). In fact, although not so obvious, two to three similar depressions also can be recognized on the labial edge of the upper jaw. This so-called “incissivlucken” is previously only known in Saurichthys krambergeri (Griffith, 1962), a fragment of jaw tip of an undetermined species of Saurichthys from the Eotrias of South African (Griffith, 1978) and in Acidorhynchus (Gardiner, 1960; Thies, 1985).
Like all other species of the genus, the new species has a single large opercular in each side. It is roughly trapezoid-
shaped (Fig. 2a), more or less similar to that of S. striolatus and Saurichthys calcaratus of Upper Triassic (Griffith, 1959, 1977), about 16 mm wide and 18 mm deep, with a nearly straight anterior and dorsal margin and round curved posteroventral margin. The external surface of the opercular is almost fully covered with closely- spaced vertical striae besides some tubercles in the anterior margin. Similar vertical striate ornament in opercular is previously only known in S. calcaratus (Griffith, 1977).
An elongated branchiostegal ray can be seen slightly ventral to the mandible and ventroanterior to the opercular of the holotype to overlap on its counterpart (Fig. 2a).
4.2 Paired fins and appendicular skeleton
The left pectoral fin is well preserved where it is enclosed by the dorsal and posterior processes of the cleithrum in the holotype (Fig. 2a). It is roughly fan-shaped and contains approximately 34 lepidotrichia, in which the longest one is less than 14 mm long.
Two dermal bones related to the pectoral girdle are recognized in the holotype, including the cleithrum and posttemporal (Fig. 2a). The cleithum is of typically triradiate shape with a slender and long dorsal ramous that reaches about half of the skull depth, and a relatively broad and horizontally-extended ventral portion that consists of long anterior and shorter posterior processes. The anterior process of the cleithrum truncates anteriorly where a clavicle should be originally in contact with but is not preserved in the current material. The left supracleithrum is slightly displaced posteriorly from the dermopterotic but is still partially overlapped by the opercular (Fig. 2a). Its anterior part is overlapped by the left extrascapular and the posterior exposed part is roughly composed of a triangular dorsal face and a crescent lateral face, the former tapering posteriorly and decorated with tubercle ornaments, the latter partially overlapped by the oprcular. The lateral line runs along the boundary of the two faces. Thus, it is possible that it is a compound bone fused with the posttemporal. No trace of the endoskeletal element of the pectoral girdle is found.
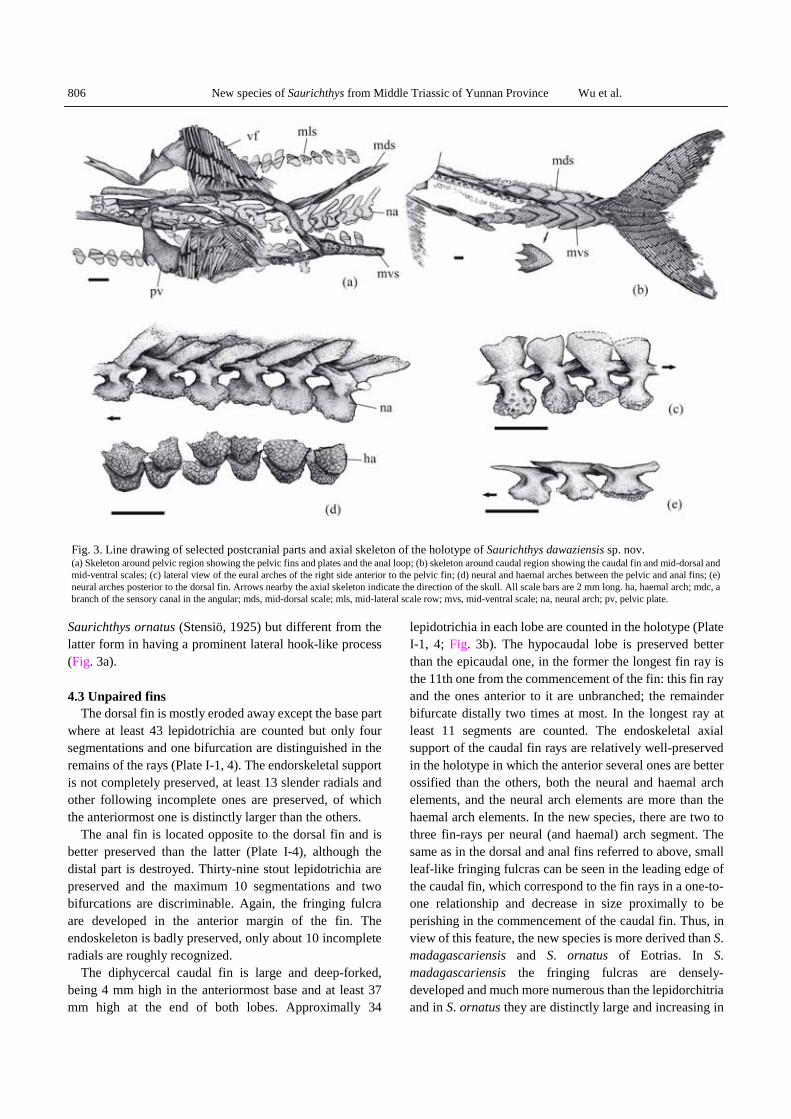
The pelvic fins are roughly situated abdominally in the midpoint between the opercular and the caudal fin (Plate I-1). They are rather small, at least not larger than the pectoral ones, consisting of 23–27 stout lepidotrichia with at least three times segmentation and once distal bifurcation (Fig. 3a). The fringing fulcra are observed in the anterior margin of the left pelvic fin.
The pelvic plates are well-ossified but its posterior portion of them is covered by the fin-rays (Fig. 3a). The exposed part of the pelvic plate is quite large and is of two-shanked structure with a shank directing anteromedially and the other posterolaterally, very similar to that in
806 New species of Saurichthys from Middle Triassic of Yunnan Province Wu et al.
Saurichthys ornatus (Stensiö, 1925) but different from the latter form in having a prominent lateral hook-like process (Fig. 3a). 4.3 Unpaired fins
The dorsal fin is mostly eroded away except the base part where at least 43 lepidotrichia are counted but only four segmentations and one bifurcation are distinguished in the remains of the rays (Plate I-1, 4). The endorskeletal support is not completely preserved, at least 13 slender radials and other following incomplete ones are preserved, of which the anteriormost one is distinctly larger than the others.
The anal fin is located opposite to the dorsal fin and is better preserved than the latter (Plate I-4), although the distal part is destroyed. Thirty-nine stout lepidotrichia are preserved and the maximum 10 segmentations and two bifurcations are discriminable. Again, the fringing fulcra are developed in the anterior margin of the fin. The endoskeleton is badly preserved, only about 10 incomplete radials are roughly recognized.
The diphycercal caudal fin is large and deep-forked, being 4 mm high in the anteriormost base and at least 37 mm high at the end of both lobes. Approximally 34
lepidotrichia in each lobe are counted in the holotype (Plate I-1, 4; Fig. 3b). The hypocaudal lobe is preserved better than the epicaudal one, in the former the longest fin ray is the 11th one from the commencement of the fin: this fin ray and the ones anterior to it are unbranched; the remainder bifurcate distally two times at most. In the longest ray at least 11 segments are counted. The endoskeletal axial support of the caudal fin rays are relatively well-preserved in the holotype in which the anterior several ones are better ossified than the others, both the neural and haemal arch elements, and the neural arch elements are more than the haemal arch elements. In the new species, there are two to three fin-rays per neural (and haemal) arch segment. The same as in the dorsal and anal fins referred to above, small leaf-like fringing fulcras can be seen in the leading edge of the caudal fin, which correspond to the fin rays in a one-to-one relationship and decrease in size proximally to be perishing in the commencement of the caudal fin. Thus, in view of this feature, the new species is more derived than S. madagascariensis and S. ornatus of Eotrias. In S. madagascariensis the fringing fulcras are densely-developed and much more numerous than the lepidorchitria and in S. ornatus they are distinctly large and increasing in
Fig. 3. Line drawing of selected postcranial parts and axial skeleton of the holotype of Saurichthys dawaziensis sp. nov. (a) Skeleton around pelvic region showing the pelvic fins and plates and the anal loop; (b) skeleton around caudal region showing the caudal fin and mid-dorsal and mid-ventral scales; (c) lateral view of the eural arches of the right side anterior to the pelvic fin; (d) neural and haemal arches between the pelvic and anal fins; (e) neural arches posterior to the dorsal fin. Arrows nearby the axial skeleton indicate the direction of the skull. All scale bars are 2 mm long. ha, haemal arch; mdc, a branch of the sensory canal in the angular; mds, mid-dorsal scale; mls, mid-lateral scale row; mvs, mid-ventral scale; na, neural arch; pv, pelvic plate.

Vol. 83 No. 3 ACTA GEOLOGICA SINICA June 2009 807
size proximally.
4.4 Axial skeleton The axial skeleton consists of the neural and haemal
arches applied to the persisting notochord as is typical for the genus. The total number of the neural arch segments between the opercular and caudal fin is 160 ± 1. The neural arches are paired in structure. Each of the neural arches in front of the dorsal fin is differentiated into a plate-like dorsal neural spine, an acetabula-like ventral foot and a laterally convex lateral wall between them (Fig. 3c, d). The neural spine of the neural arches gradually decreases in width but increases in depth posteriorly to the level anterior to the dorsal fin and then suddenly disappeared to make the remaining neural arches in a “T” shape in the caudal pedicle region (Fig. 3e). The medial surface of the neural arch, together with a lamella located medially at the base of the neural spine, encloses a complete circular canal for the spinal cord with the corresponding structures of the other side. A small circular foramen can be observed to occur in the medial surface of every other neural arch. It is possibly related to the dorsal branch of the dorsal intersegmental branch of the aorta or to the dorsal or ventral roots of the spinal nerve. The lamella mentioned above also prolongs both anteriorly and posteriorly along its dorsal edge to form the so-called prea- and post-zygapophyses to articulate with the preceding or subsequent neural arches in the way that its preazygapophyses overlaps on the postzygapophyses of the preceding one. The acetabula-like ventral foot of the neural arch rests on the dorsolateral aspect of the notochord.
The ossified haemal arches are poorly preserved and can only be recognized in region between the pelvic and anal fins, occurring as small semicircular plates without haemal spines (Fig. 3d).
4.5 The squamation
Like the saurichthyid species from the Lower Triassic of Spitzbergen (Stensiö, 1925) and some Upper Triassic species from Europe (Griffith, 1959, 1977), the squamation of the new species is reduced to four longitudinal scale rows, a mid dorsal and a mid ventral plus a lateral row in each side of the trunk (Plate I-1, 3).
The mid-dorsal scale row runs backward from the skull to the base of the epicaudal lobe of the caudal fin, interrupted only by the dorsal fin. The scales anterior to the dorsal fin, are slender lanceolate shaped and slightly increase in length from 6 mm to close to 9 mm in length posteriorly with a length : width ratio of about 10:1 (Fig. 4a, b). The ones posterior to the dorsal fin gradually increase in width to become coulter-shaped with the length : width ratio reaching about 5:3 except the last one and almost fully
covering the posterior part of the caudal region together with the mid-ventral scales (Fig. 3b). The last of these scales is reduced to a small plate covering the anterior base of the epicaudal lobe of the caudal fin. The anterior half of the mid-dorsal scale is excavated to produce a depression for accepting the posterior portion of the preceding scale where there is a keel along the longitudinal axis ventrally (Fig. 4c), while the exposed posterior part of the scale is ornamented with tubercles.
The mid-ventral scale row begins to develop roughly near the mid-point between the pectoral and pelvic fins, from where it runs backward to the caudal region. The mid-ventral scales show similar shape and transformation with the mid-dorsal ones, but they are evidently smaller than the mid-dorsal one anterior to the pelvic fin. In the region of the pelvic fins the mid-ventral scale row branches to form the anal loop between the pelvic fins and each side of the loop is composed of five scales (Fig. 3a).
The mid-lateral scale row runs along the bodylobe through the diphycercal caudal fin. The scales are of roughly triangular to subovate shape with a prominent axial ridge in the external surface that protrudes anteriorly and
Fig. 4. Line drawing of the squamation of Saurichthys dawazien-sis sp. nov. (a) Mid-dorsal scales posterior to the skull in lateral view; (b) mid-dorsal and lateral scale rows anterior to the dorsal fin showing the size proportion of the scales; (c) isolated mid-dorsal scales in dorsal view; (d) lateral scales in lateral view; (e) lateral scale in medial view. Part (c) is from GMPKU-P-1526; the others are from the holotype. All scale bars are 1 mm long.
GMPKU-SUN
Note
插入and ventral view

808 New species of Saurichthys from Middle Triassic of Yunnan Province Wu et al.
posteriorly to articulate with the subsequent and proceeding scales, respectively and subdivides the scale into a dorsal and ventral portion (Fig. 4b, d). The dorsal portion of the scale is decorated with some spines, whereas the ventral portion is free of ornaments. In the medial side of the scale a longitudinal groove can be seen corresponding to the ridge in the external surface (Fig. 4e), which should be related to the lateral line canal. 5 Comparison
As described above, the elongated premaxilla and the peculiar external nostrils within the nasal-antorbital, typical paleoniscoid-type maxilla and large single opercular, as well as the reduced squamation, remotely and symmetrically arranged dorsal and anal fins and the abbreviated diphycercal caudal fin, strongly suggest that the new species should be unquestionably refered to as the genus Saurichthys. It shares some features with the Lower Triassic species of the genus, such as the numerous segments in the lepidotrichia of the unpaired fins and the presence of fringing fulcra in the unpaired fins (Stensiö, 1925; Lehman, 1959) and the presence of the supraorbital, meanwhile it also has some other characters that are more derived than those in the Lower Triassic species but similar to those of some Middle–Late Triassic representatives, such as single ossification in sclerotic ring, narrow postorbital region and relatively high posterior lobe of the maxilla, reduced quadratojugal, shape and ornamentation of the opercular, and unjointed pectoral fin rays (Griffith, 1959, 1962; Rieppel, 1985, 1992). All of these suggest that the new species is an intermediate form between the early and later representatives of the genus. The presence of a branch of the mandible sensory canal in the angular, the ossified haemal arches without haemal spines, as well as the foramen of the efferent pseudobranchial artery in the parasphenoid and variation of the mid-dorsal scales are the unique features of the new species, which allows obvious distinguishing from all others of the genus. 6 Discussion on the Vertebral Column, Unpaired Fins and Squamation in Saurichthys
Saurichthys is a rather conservative genus and few
anatomic pattern changes occur in the dermal bones of the skull. Thus, there have been few changes in the skull reliably linked with phylogeny in the genus, possibly except for the changes of some aspects in the vertebrate column, unpaired fins and squamation, which seem to have potential phylogenetic significance.
6.1 Vertebral column The axial skeleton of Saurichthys consists of paired
neural and haemal arches and has been recorded in some species through the Triassic (Stensiö, 1925; Lehman, 1952; Griffith, 1959, 1962; Rieppel, 1985, 1992; Tintori, 1990). Although the complete pattern of axial skeleton is not well known in most cases, it seems that the ratio of the numbers of the neural arch segments to the haemal arch segments between the pelvic and anal fins in Saurichthys is variable through time. In the Lower Triassic Saurichthys, two neural arches correspond with one haemal arch (Stensiö, 1925; Lehman, 1952), while in Saurichthys dawaziensis sp. nov. and other Middle Triassic species (Rieppel, 1985, 1992), the neural arches keep in a one-to-one relationship to the haemal arches, and the haemal arches anterior to the caudal fin may eventually not be ossified in the Late Triassic species from Austria (Griffith, 1959, 1962). Based on the relationship of the neural arch segments to the haemal arch segments between the pelvic and anal fins in Saurichthys from Spitzbergen, Stensiö (1925) hypothesized that the neural arches of the vertebral column of Saurichthys consists of equally developed basidorsals and interdorsals and possessing the same shape, that is, every other neural arch is the basidorsal and the other is the interdorsal. But this viewpoint was not widely accepted (Lehman, 1952). However, the axial skeleton of Saurichthys dawazienesis sp. nov. provides the crucial evidence to support Stensiö’s hypothesis that the neural arches of Saurichthys consist of equally developed basidorsals and interdorsals. In Saurichthys dawazienesis sp. nov. a circular foramen that possibly related to the dorsal branch of the dorsal intersegmental branch of the aorta to the neural cord or to the dorsal or ventral roots of the spinal nerve regularly consistently occurs in every other neural arch. The distribution pattern of the foramen in the neural arches of the new species strongly suggests that in Saurichthys, one myotome segment corresponds to two neural arches. Thus, we have confidence to conclude that the equally developed basidorsals and interdorsals in the development of the vertebral column is a synapomorph of the genus Saurichthys or the family Saurichthyidae, differing from other actinopterygian fishes. We can also go further from Stensiö’s (1925) hypothesis that the one-to-one relationship of the haemal arches to the neural arches between the pelvic and anal fins in the latter forms (including the new species described in this paper) might represent a secondary splitting of the coalesced basiventral and interventral.
Besides, the so-called postzygapophyses of the neural arches is present in S. ornatus and S. dawaziensis sp. nov., but absent in S. madagascariensis and most of the other Middle and Upper Triassic forms of Saurichthys (Lehman, 1952; Griffith, 1959; Rieppel, 1985, 1992; Tintori, 1990),

Vol. 83 No. 3 ACTA GEOLOGICA SINICA June 2009 809
while in S. krambergeri, both prae- and postzygapophysis are absent (Griffith, 1962). Additionally, the haemal spines are not developed in S. dawaziensis sp. nov.
6.2 Unpaired fins
Numerous segmentation and the presence of the fringing fulcra in the unpaired fins in the Lower Triassic Saurichthys are considered to be the plesiomorph condition (Rieppel, 1992). The unpaired fins have 10 or more segments in S. madagascariensis, S. ornatus and S. dawaziensis sp. nov., less than four or five in most of the Middle Triassic species (Rieppel, 1985, 1992) and the fin rays in the unpaired fins are unjointed in the Upper Triassic S. calcaratus and S. striolatus (Griffith, 1959, 1977). The fringing fulcra are absent in all known Middle-Upper Triassic Saurichthys except for S. dawaziensis sp. nov., in which, as discussed above, it is more derived than in Lower Triassic Saurichthys.
6.3 Squamation
Reduced squamation is a characteristic feature of all representatives of the genus except S. madagascariensis from the Eotrias of Madagascar (Lehman, 1959; Rieppel, 1980) and the earliest records of the saurichthyids, Eosaurichthys chaoi from the Upper Permian of southeastern China (Liu and Wei, 1988), both of which bear the complete squamation. Four longitudinal scale rows are common in most species of the genus (Woodward, 1890; Stensiö, 1925; Griffith, 1959, 1977; Schaeffer and Mangus, 1976) and the new species described herein, whereas six scale rows exist in the four Middle Triassic species from Monte San Giorgio (Rieppel, 1985, 1992) and two rows in S. krambergeri (Upper Triassic) from Austria (Griffith, 1962).
The shape of mid-dorsal scales anterior to the dorsal fin are subovate to ovate in the Lower Triassic Saurichthys from Madagascar and British Columbia (Rieppel, 1980; Schaeffer and Mangus, 1976) S. gigas of the Middle Triassic from Australia (Woodward, 1890), while in other species the mid-dorsal scales are generally elongated or lanceolate in shape, evidently longer than wide (Woodward, 1890; Stensiö, 1925; Griffith, 1959, 1962, 1977; Rieppel, 1985, 1992; Tintori, 1990). The ratio of the mid-dorsal to the mid-lateral scales in number seems to be of the one-to-two relationship among all Lower and Middle Triassic Saurichthys forms with reduced squamation, while in the Upper Triassic representatives there are two different occasions: in S. calcaratus and S. striolatus where contrarily the mid-lateral scales are distinctly larger than the mid-dorsal scales (Griffith, 1959, 1977) and in S. krambergeri, where the mid-lateral scales are totally lost (Griffith, 1962). The mid-lateral scales in most species of
Saurichthys with reduced squamation are generally triangular to subovate in outline; however, the rib-like mid-lateral scales are characteristic of the Middle Triassic S. costasquamosus and S. paucitrichus from Monte San Giorgio (Rieppel, 1985, 1992). 7 Conclusions
(1) Saurichthys dawaziensis sp. nov. is a transition form between the Lower and Middle-Upper Triassic species of Saurichthys, characterized by the efferent pseudobranchial arteries penetrating on the parasphenoid, the mandible sensory canal with a branch in the angular and the ossified haemal arches without haemal spines between pelvic and anal fins. It is more derived than the Lower Triassic species in having a relatively narrow postorbital region, short preopercular, single ossification of the sclerotic ring, the small haemal arches between the pelvic fins and the anal fin without haemal spines, and in the absence of quadratojugal, and more primitive than the Middle and Upper Triassic Saurichthys in having fringing fulcra and numerous segments in the unpaired fins and the presence of both prae- and post-zygapophyses in the neural arches.
(2) The presence of the foramen, possibly for the dorsal intersegmental branch of the aorta or the roots of the spinal nerve, in every other neural arch of the new species provides crucial evidence for Stensiö’s (1925) hypothesis that the neural arches of the vertebral column of Saurichthys consists of equally developed basidorsals and interdorsals possessing the same shape; a unique feature of Saurichthys. The one-to-one relationship of the haemal arches to the neural ones between the pelvic and anal fins in the latter representatives of Saurichthys might reflect a secondary splitting of the coalesced basiventral and interventral.
(3) The segmentation of the unpaired fins is numerous in the Lower Triassic Saurichthys and S. dawaziensis sp. nov., and tends to be fewer in the late representatives of the genus. The absence of the fringing fulcra in the unpaired fins is characterized for most of the Middle-Upper Triassic species of Saurichthys.
(4) The squamation reducing is a general trend in Saurichthys. The variation of the squamation and certain scales among Saurichthys is rather complicated and requires detailed comparative study in different species.
Acknowledgements
The authors thank Professor Chang Mee-mann for her
helpful guide on the anatomic study of the specimens and providing related literature. This work is supported by the NSFC (grants no. 40572004 and 40702001).

810 New species of Saurichthys from Middle Triassic of Yunnan Province Wu et al.
Manuscript received Jan. 9, 2009
accepted Feb. 22, 2009 edited by Jiang Shaoqing
References 请作者补充我刊近期发表的参考文献1-2篇具体参考邮件附件。 Agassiz, L., 1834. Recherches sur les Poissons Fossiles, Vol. 2,
336 ps. Imprimerie de Petitpierre, Neuchâtel. Beltan, L., 1968. La faune ichthyologique de l’Eotrias du N.W. de
Madagascar: le neurocrane, 1–135. C.N.R.S., Paris. Chen Liezu, 1985. Early Triassic ichthyosaurs from Chaoxian,
Anhui. Regional Geology of China, 15(是否有期号?): 139–146. (in Chinese with English abstract?).
Chou Xiaohe and Liu Xianting, 1957. Fossil fishes from Hengshan, Shenxi. Acta Palaeontologica Sinica, 5(2): 295–305.
Gardiner, B.G., 1960. A review of certain actinopterygian and coelacanth fishes, chiefly from the Lower Lias. Bulletin of the British Museum (Natural History). Geology, 4(7): 239–384.
Griffith, J., 1959. On the anatomy of two saurichthyids fishes, Saurichthys striolatus (Bronn) and S. curionii (Bellotii). Proceedings of the Zoological Society of London, 132(4): 587–606.
Griffith, J., 1962. The Triassic fish Saurichthys krambergeri (Bronn). Palaeontology, 5(是否有期号?): 344–354.
Griffith, J., 1977. The Upper Triassic fishes from Polzberg bei Linz, Austria. Zoological Journal of the Linnean Society of London, 60(1): 1–93.
Griffith, J., 1978. A fragmentary specimen of Saurichthys sp. from Upper Beaufort Series of South Africa. Annales of the South African Museum, 76(是否有期号?): 299–307.
Lehman, J.P., 1952. Etude complémentaire des poissons de l’Eotrias de Madagascar. Kungliga Svenska Vetenskapsakademiens Handlinga, 4 series, 2(6): 1–201.
Lehman, J.P., Chateau, C., Laurain, M., and Nauche, M., 1959. Paléotologie de Madagascar. XXVIII. Les poisons de la Sakamena Moyenne. Annales de Paléontologie, Vertébres, 45(是否有期号?): 177–219.
Liu Xianting and Wei Feng, 1988. A new saurichthyid from the upper Permian of Zhejiang, China. Vertebrata Palasiatica, 26(2): 77–89.
Piveteau, J, 1945. Les Poissons du Trias Inférieur la Famille das Saurichthyidés.Paléontologie de Madagascar, XXV. Annales de Paléontologie, 31(是否有期号?): 79–87.
Rieppel, O., 1980. Additional specimens of Saurichthys madagascariensis Piveteau, from the Eotrias of Madagascar. Neues Jahrbuch für Geologie und Palæontologie, Monatshefte, 43–51.
Rieppel, O., 1985. Die Triasfauna der Tessiner Kalkalpen. XXV. Die Gattung Saurichthys (Pisces, Actinopterygii) aus der mittleren Trias des Monte San Giorgio, Kanton Tessin. Schweizerische Paläontologische Abhandlungen, 108(是否有期号?): 1–103.
Rieppel, O., 1992. A new species of the genus Saurichthys (Pisces:Actinopterygii) from the Middle Triassic of Monte San Giorgio (Switzerland), with comments on the phylogenetic interrelationships of the genus. Palaeontographica Abt. A, 221(是否有期号?): 63–94.
Schaeffer, B., and Mangus, M., 1976. An Early Triassic fish
assemblage from British Columbia. Bulletin of the American Museum of the Natural History, 156(是否有期号?): 515–564.
Stensiö. S, 1925. Triassic Fishes from Spitzebergen, Part II. 此文献来自书还是期刊?还是科研报告?按照我刊文献格式列出.
Su Dezao and Li Zuocong, 1983. A new Triassic perleidid fish from Hubei, China. Vertebrata PalAsiatica, 21(是否有期号?): 9–16 (in Chinese with English abstract).
Sun Zuoyu, Sun Yuanlin, Hao Weicheng and Jiang Dayong, 2006. Conodont evidence for the age of the Panxian Fauna, Guizhou, China. Acta Geologica Sinica (English Edition), 80(5): 621–630.
Thies, D., 1985. Funde von Acidorhynchus brevirostris (Woodward 1895) aus dem Posidonienschiefer (Unter-Toarcium) NW-Deutschlands. Palaeontographica Abt. A., 187(是否有期号?): 183–203.
Tintori, A., 1990. The vertebral column of the Triassic fish Saurichthys (Actinopterygii) and its stratigraphical significance. Rivista Italiana di Paleontologia e Stratigrafia, 96(是否有期号?): 93–102.
Tong, J., Zhou, X, Erwin, D.H., Zou, J. and Zhao, L., 2006. Fossil fishes from the Lower Triassic of Majiashan, Chaohu, Anhui Province, China. Journal of Paleontology, 80(是否有期号?): 146–161.
Woodward, A.S., 1890. The fossil fishes of the Hawkesbury Series at Gosford. Memoirs of the Geological. Survey of New South Wales, 4(是否有期号?): 1–55.
Zhang Qiyue, Zhou Changyong, Lü Tao, Xie Tao, Lou Xiongying, Liu Wei, Sun Yuanyuan and Jiang Xinsheng, 2008. Discovery and significance of the Middle Trassic Anisian biota from Luoping, Yunnan Province. Geological Review, 54(4): 523–526 (in Chinese with English abstract).
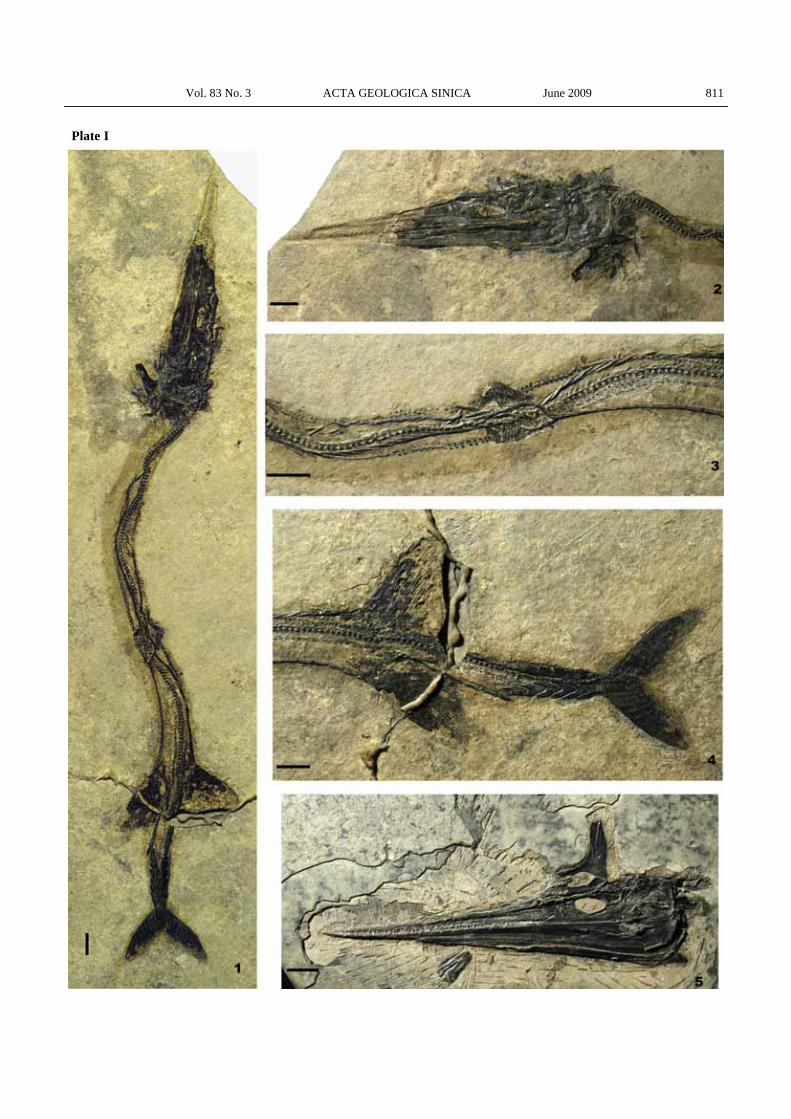
Explanation of plates Plate I Saurichthys dawaziensis sp. nov. All scale bars are 1
cm long. 1. Overview of the holotype, GMPKU-P-1524. 2. Skull of the holotype. 3. Skeleton around the pelvic region of the holotype. 4. Caudal region of the holotype. 5. Skull of GMPKU-P-1525.
Vol. 83 No. 3 ACTA GEOLOGICA SINICA June 2009 811
Plate I
Related Documents











