NEW MULTITUBERCULATE MAMMALS FROM THE HAUTERIVIAN ⁄ BARREMIAN TRANSITION OF EUROPE (IBERIAN PENINSULA) by AINARA BADIOLA , JOSE ´ I. CANUDO and GLORIA CUENCA-BESCO ´ S Grupo Aragosaurus, Paleontologı´a, Facultad de Ciencias, Universidad de Zaragoza, Pedro Cerbuna 12, E-50009 Zaragoza, Spain; e-mails: [email protected], [email protected], [email protected] Typescript received 18 March 2008; accepted in revised form 5 August 2008 Abstract: New multituberculate mammals from the Hauter- ivian ⁄ Barremian transition of Europe are described. They were found in the late Hauterivian-early Barremian fossilifer- ous locality of La Cantalera (Josa, Teruel, Spain), one of the Early Cretaceous sites in the Aragonese branch of the Iberian Ranges, in northeastern Iberia. The fossils have been assigned to at least three taxa on the basis of nine isolated teeth: a new pinheirodontid taxon, Cantalera abadi gen. et sp. nov.; a representative of the eobaatarid Eobaatar; a taxon described as Plagiaulacidae or Eobaataridae gen. et sp. indet.; and other as Plagiaulacida indet. These fossils have increased the reso- lution of European Early Cretaceous multituberculate mam- malian biostratigraphy and palaeobiogeography: the oldest representative of Eobaatar is described here; a taxon is assigned to ?Plagiaulacidae, in which case it would be the first of this family in the Iberian Peninsula; and the discovery of a new late Hauterivian pinheirodontid taxon demonstrates greater biodiversity and a wider distribution for these multi- tuberculates than was previously known. The mutituberculate fauna of La Cantalera consists of endemic taxa (Pin- heirodontidae), which were restricted to what is now Wes- tern Europe, and others (Eobaataridae) which have also been described in Asia. Consistent with the Iberian record of late Barremian gobiconodontid mammals, the presence of Eobaa- tar in Iberia with representatives from the late Hauterivian to late Barremian, as well as in the Aptian or Albian of Mon- golia, indicates that faunal exchanges between Europe and Asia could have existed for most of the Early Cretaceous, either sporadically or constantly. Key words: Pinheirodontidae, Eobaataridae, ?Plagiaulacidae, Early Cretaceous, Europe, systematics, palaeobiogeograhy. When Pangaea broke up at the end of the Jurassic, a significant separation between Laurasia and Gondwana took place, causing increased differentiation of their respective faunas. In the Early Cretaceous Laurasia was divided into three palaeobiogeographic units: North America, Europe and Central Asia (Russell 1993). These landmasses, rather than being continuous, formed an archipelago. The combined effect of plate movements and eustatic sea level changes during the Early Cretaceous (Skelton et al. 2003) allowed the coastal or shallow sea areas to be used during sea level lows by the terrestrial tetrapods to move between these three major continental areas. The position of Europe, with its connections to Asia, North America and Gondwana, makes it a challenge to unravel the palaeobiogeography of terrestrial tetrapods at this time. The expression ‘biogeographic ambiguity of Europe’ defines this situation very well (Russell 1993; Canudo 2006). The Early Cretaceous, therefore, is a key period in investigating the geographical distribution, ori- gin and evolution of many continental vertebrate faunas, including mammals. To date, the most complete and con- tinuous Early Cretaceous mammalian fossil record in Europe has been found in the Iberian Peninsula, where immigrant (Gobiconodontidae Chow and Rich, 1984) and endemic faunas (the Dryolestidae Crusafontia cuen- cana Henkel and Krebs, 1969) were present (Cuenca- Besco ´ s and Canudo 2003, 2004). Multituberculate fossils dominate the mammal assemblages of the Iberian Penin- sula (Canudo and Cuenca-Besco ´s 1996; Crusafont and Gibert 1976; Cuenca-Besco ´s et al. 1996; Hahn and Hahn 1992, 1999, 2000, 2001, 2002), which also consist both of endemic taxa (Pinheirodontidae Hahn and Hahn, 1999 and Paulchoffatiidae Hahn, 1969), with representatives from the Late Jurassic, and others which, to date, are only known in western Europe and Asia (Eobaataridae Kielan- Jaworowska et al. 1987). New multituberculate finds from the Iberian Peninsula (Canudo and Cuenca-Besco ´ s 1996; Cuenca-Besco ´ s and Canudo 2004; Hahn and Hahn 2006; Badiola et al. 2007a, b), together with those from Britain (Sigogneau-Russell and Kielan-Jaworowska 2002; Butler [Palaeontology, Vol. 51, Part 6, 2008, pp. 1455–1469] ª The Palaeontological Association doi: 10.1111/j.1475-4983.2008.00822.x 1455

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

NEW MULTITUBERCULATE MAMMALS FROM THE
HAUTERIVIAN ⁄ BARREMIAN TRANSITION OF
EUROPE (IBERIAN PENINSULA)
by AINARA BADIOLA , JOSE I. CANUDO and GLORIA CUENCA-BESCOSGrupo Aragosaurus, Paleontologıa, Facultad de Ciencias, Universidad de Zaragoza, Pedro Cerbuna 12, E-50009 Zaragoza, Spain; e-mails: [email protected],
[email protected], [email protected]
Typescript received 18 March 2008; accepted in revised form 5 August 2008
Abstract: New multituberculate mammals from the Hauter-
ivian ⁄ Barremian transition of Europe are described. They
were found in the late Hauterivian-early Barremian fossilifer-
ous locality of La Cantalera (Josa, Teruel, Spain), one of the
Early Cretaceous sites in the Aragonese branch of the Iberian
Ranges, in northeastern Iberia. The fossils have been assigned
to at least three taxa on the basis of nine isolated teeth: a
new pinheirodontid taxon, Cantalera abadi gen. et sp. nov.;
a representative of the eobaatarid Eobaatar; a taxon described
as Plagiaulacidae or Eobaataridae gen. et sp. indet.; and other
as Plagiaulacida indet. These fossils have increased the reso-
lution of European Early Cretaceous multituberculate mam-
malian biostratigraphy and palaeobiogeography: the oldest
representative of Eobaatar is described here; a taxon is
assigned to ?Plagiaulacidae, in which case it would be the
first of this family in the Iberian Peninsula; and the discovery
of a new late Hauterivian pinheirodontid taxon demonstrates
greater biodiversity and a wider distribution for these multi-
tuberculates than was previously known. The mutituberculate
fauna of La Cantalera consists of endemic taxa (Pin-
heirodontidae), which were restricted to what is now Wes-
tern Europe, and others (Eobaataridae) which have also been
described in Asia. Consistent with the Iberian record of late
Barremian gobiconodontid mammals, the presence of Eobaa-
tar in Iberia with representatives from the late Hauterivian
to late Barremian, as well as in the Aptian or Albian of Mon-
golia, indicates that faunal exchanges between Europe and
Asia could have existed for most of the Early Cretaceous,
either sporadically or constantly.
Key words: Pinheirodontidae, Eobaataridae, ?Plagiaulacidae,
Early Cretaceous, Europe, systematics, palaeobiogeograhy.
When Pangaea broke up at the end of the Jurassic, a
significant separation between Laurasia and Gondwana
took place, causing increased differentiation of their
respective faunas. In the Early Cretaceous Laurasia was
divided into three palaeobiogeographic units: North
America, Europe and Central Asia (Russell 1993). These
landmasses, rather than being continuous, formed an
archipelago. The combined effect of plate movements and
eustatic sea level changes during the Early Cretaceous
(Skelton et al. 2003) allowed the coastal or shallow sea
areas to be used during sea level lows by the terrestrial
tetrapods to move between these three major continental
areas. The position of Europe, with its connections to
Asia, North America and Gondwana, makes it a challenge
to unravel the palaeobiogeography of terrestrial tetrapods
at this time. The expression ‘biogeographic ambiguity of
Europe’ defines this situation very well (Russell 1993;
Canudo 2006). The Early Cretaceous, therefore, is a key
period in investigating the geographical distribution, ori-
gin and evolution of many continental vertebrate faunas,
including mammals. To date, the most complete and con-
tinuous Early Cretaceous mammalian fossil record in
Europe has been found in the Iberian Peninsula, where
immigrant (Gobiconodontidae Chow and Rich, 1984)
and endemic faunas (the Dryolestidae Crusafontia cuen-
cana Henkel and Krebs, 1969) were present (Cuenca-
Bescos and Canudo 2003, 2004). Multituberculate fossils
dominate the mammal assemblages of the Iberian Penin-
sula (Canudo and Cuenca-Bescos 1996; Crusafont and
Gibert 1976; Cuenca-Bescos et al. 1996; Hahn and Hahn
1992, 1999, 2000, 2001, 2002), which also consist both of
endemic taxa (Pinheirodontidae Hahn and Hahn, 1999
and Paulchoffatiidae Hahn, 1969), with representatives
from the Late Jurassic, and others which, to date, are only
known in western Europe and Asia (Eobaataridae Kielan-
Jaworowska et al. 1987). New multituberculate finds from
the Iberian Peninsula (Canudo and Cuenca-Bescos 1996;
Cuenca-Bescos and Canudo 2004; Hahn and Hahn 2006;
Badiola et al. 2007a, b), together with those from Britain
(Sigogneau-Russell and Kielan-Jaworowska 2002; Butler
[Palaeontology, Vol. 51, Part 6, 2008, pp. 1455–1469]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2008.00822.x 1455

and Hooker 2005; Sweetman 2006), have increased the
resolution of the Middle and Late Jurassic and Early
Cretaceous multituberculate biostratigraphy and biogeo-
graphy.
The most abundant and diverse Early Cretaceous multi-
tuberculate fossils are recorded in Western Europe, from
the Berriasian of Portugal and England and from the
Barremian of Spain. Fossils from the Valanginian are
scarce, and no Hauterivian specimens have previously
been described (Kielan-Jaworowska et al. 2004; Hahn
and Hahn 2006). In this paper, the first record of late
Hauterivian multituberculates in Europe is reported.
These have been recovered in the late Hauterivian-early
Barremian site of La Cantalera (Josa, province of Teruel),
one of the sites in the Aragonese branch of the Iberian
Ranges. The latter is becoming one of the most impor-
tant areas in the study of Early Cretaceous vertebrate
faunas, mainly multituberculates, because of the abun-
dant and stratigraphically well-correlated fossiliferous
beds. To date, late Valanginian, late Hauterivian and
early and late Barremian multituberculate fossils have
been recovered (Badiola et al. 2007a, b), of which those
of the Barremian are the most abundant and diverse
throughout the world (Canudo and Cuenca-Bescos 1996;
Cuenca-Bescos and Canudo 2004; Cuenca-Bescos et al.
1996; Ruiz-Omenaca et al. 2004). Further sampling in
the Iberian Range, which is currently being undertaken,
is expected to yield crucial information that will increase
what is known about the phylogeny, systematics, palaeo-
biology and distribution of the Early Cretaceous multi-
tuberculate faunas.
Geologically, La Cantalera site is an outcrop of Early
Cretaceous clays (‘Wealden facies’), specifically clays from
the limestones and marls of the Blesa Formation, located
in the Oliete Sub-Basin, one of the sub-basins of the Early
Cretaceous Maestrazgo Basin (Text-Fig. 1). This unit has
had a complex history, including a lower part with allu-
vial to lacustrine sedimentation (the site of La Cantalera).
The fossiliferous beds comprise grey clays with abundant
vegetal remains, ostracodes (Cypridea [Ulwellia] soriana
Kneuper-Haack, 1966) and gastropods (Viviparidae
indet., Basamotophora indet.) that are reported to be
TEXT -F IG . 1 . Geographical location and geological setting of the late Hauterivian–early Barremian site of La Cantalera (Oliete Sub-
Basin, Maestrazgo Basin). Modified from Aurell et al. 2004.
1456 P A L A E O N T O L O G Y , V O L U M E 5 1

resistant to periods of drought (Brookes Knight et al.
1960; Mayoral and Sequeiros 1983), and abundant car-
bonate nodules that were formed in palaeosols (Aurell
et al. 2004). There are also charophytes (Atopochara tri-
volvis triqueta Grambast, 1968 nov. com. Martın-Closas,
1996, Hemiclavator adnatus Martın-Closas and Grambast-
Fessard, 1986 nov. com. Schudack, 1989) and other mac-
roplant fossil remains. The presence of the charophyte
Atopochara trivolvis triqueta, subzone triqueta, indicates
that the fossiliferous beds are late Hauterivian–early Bar-
remian in age (Canudo et al. 2002). The site was probably
formed in a lucustrine environment with periodic
droughts. It is likely to have been a rich and diverse ter-
restrial ecosystem with a complex and varied faunal
assemblage and abundant vegetation in the surrounding
area (Ruiz-Omenaca et al. 1997). The highly diverse ver-
tebrate fossils are composed mainly of isolated teeth, dis-
articulated, complete or fragmented bones, eggshell
fragments, and coprolites. The vertebrate assemblage con-
sists of fishes (scarce fossils), frogs, lizards, crocodiles,
turtles, pterosaurs, dinosaurs and mammals (Ruiz-
Omenaca et al. 1997, 2005; Canudo et al. 2002). The
study of the vertebrate fossils from La Cantalera is still in
progress. To date, nine isolated teeth of multituberculate
mammals have been recovered, which are here assigned
to at least three different taxa.
Abbreviations
Institutional. FCPT, Fundacion Conjunto Paleontologico de Ter-
uel-Dinopolis, Teruel, Spain.
Localities. CAN, La Cantalera site.
Dentition. We use capital letters (I, C, P, M) for upper dentition
and lower-case letters (i, c, p, m) for lower dentition. Cusps are
labelled following Kielan-Jaworowska et al. (2004, text-fig. 8.28,
modified from Hahn and Hahn 1998a, text-fig. 1a): cusps of the
labial (b, B) and lingual (l, L) rows are numbered mesio-distally,
indicating their corresponding number and letter; a capital letter
(L, B) in the upper cheek tooth row and a lower-case letter (l,
b) in the lower cheek tooth row (e.g. B3: third labial cusp of an
upper tooth; l2: second lingual cusp of a lower tooth). The cusp
formula is that proposed by Kielan-Jaworowska et al. (2004,
p. 279), which is indicated by the number of cusps in consecu-
tive rows given from labial to lingual, separated by a colon. We
have here added the corresponding letter for each side for an
easier reading (e.g. 3B:4L).
Measurements. L, Length; W, width, which are given in mm. L
corresponds to the mesio-distal length measurement of the
crown, taken as oriented parallel to the mesio-distal axis,
whereas W is the maximum width measurement between the
labial and lingual margins of the crown.
Systematic use. The heterogeneous knowledge of multi-
tuberculates results in important unresolved problems
regarding their systematics and the interrelationships
among the different groups. In some of them, the whole
dentition is known in situ, whereas in others only a few
isolated teeth are present. In contrast, some taxa are rep-
resented by complete skeletons (Kielan-Jaworowska and
Gambaryan 1994). Consequently, cladistic analysis does
not produce reliable results, and different systematic
propositions have been addressed (Kielan-Jaworowska
and Hurum 2001 and references therein). Within the sub-
order Plagiaulacida, all authors consider that the families
are the real evolutionary and systematic units, definable
in each case by their special apomorphies. Here, therefore,
we shall not follow any particular systematic proposition
for plagiaulacids to explain the relationships between the
different families (e.g. Kielan-Jaworowska and Hurum
2001; Hahn and Hahn 2006) until new finds allow us to
make a more extensive and reliable phylogenetic study.
SYSTEMATIC PALAEONTOLOGY
Order MULTITUBERCULATA Cope, 1884
Suborder PLAGIAULACIDA McKenna, 1971
[pro Plagiaulacoidea Ameghino, 1889]
Family PINHEIRODONTIDAE Hahn and Hahn, 1999
Genus CANTALERA gen. nov.
Type species. Cantalera abadi sp. nov.
Derivation of name. After the name of the river nearby.
Diagnosis. A pinheirodontid described on the basis of
P4 ⁄ 5 and M1, which are characterized by having abun-
dant, long, and strongly-marked radiating enamel ridges
from the tip to the base of the cusps, which intermesh at
their distal ends with the neighbouring ridges; these form
a labyrinth-like structure on the floor of the central valley
on M1. P4 ⁄ 5 rectangular in outline and considerably
lengthened, a tooth longer than it is broad, with both
rows of cusps, 2B:4L, arranged clearly parallel to the
mesio-distal axis. M1 with a 2B:4L cusp formula, without
B3, with central valley open disto-labially, and with
prominent and small cuspules on the disto-lingual and
mesio-labial corners of the crown; the first one located
above a cingulum-like structure, which is strongly later-
ally protruding, and the second one placed above the
thick cingulum that encircles the mesial margin.
Differential diagnosis. In addition to the differences with
respect to other pinheirodontids apparent in the cusp
B A D I O L A E T A L . : N E W M U L T I T U B E R C U L A T E M A M M A L S F R O M T H E H A U T E R I V I A N ⁄ B A R R E M I A N T R A N S I T I O N 1457

formula and the presence of abundant, long and strongly-
marked radiating enamel ridges throughout the height of
the cusps on P4 ⁄ 5 and M1, which intermesh basally with
the neighbouring ridges, and form a labyrinth-like struc-
ture on the floor of the central valley on M1, Cantalera
gen. nov. can be distinguished from ?P5 of Sunnyodon
Kielan-Jaworowska and Ensom, 1992 by its larger size,
and by the absence of two labial cusps arranged symmet-
rically in the middle of the length of the tooth, with two
small mesial and distal cuspules; from P4 of the Pin-
heirodontidae gen. et sp. indet. from Portugal (Hahn and
Hahn 1999) by its larger size, and by the absence of a
labial row of cusps covering the whole crown longitudi-
nally, with three cusps instead of two; and from M1 of
Iberodon Hahn and Hahn, 1999 by its larger size, and by
the absence of an enlarged L3 and an additional row of
small cuspules on the lingual wall of the crown. Lavocatia
Canudo and Cuenca-Bescos, 1996 is like Cantalera gen.
nov. in having cusps covered by prominent radiating
enamel ridges but they do not intermesh basally with the
neighbouring ridges. Moreover, Cantalera is larger in size
than Lavocatia.
Cantalera abadi sp. nov.
Plate 1
Derivation of name. Dedicated to Jose Marıa Abad, who found
the site of La Cantalera.
Holotype. Left M1, FCPT (CAN 1 ⁄ 1609).
Paratype. Left P4 ⁄ 5, FCPT (CAN 1 ⁄ 934).
Referred material. A fragment of right M1, FCPT (CAN
1 ⁄ 1610).
Diagnosis. As for genus, monotypic.
Description
M1. FCPT (CAN 1 ⁄ 1609) (holotype; L: 2.15; W: 1.52) exhibits a
well-preserved crown. It is nearly rectangular in outline, with a
2B:4L cusp formula. The lingual row of cusps is slightly broad-
ened distally. The labial row of cusps is shortened, with only
two cusps. B3 is missing; consequently, the central valley opens
disto-labially. The strongly-marked and long radiating enamel
ridges, which extend from the tip to the base of the cusps, inter-
mesh basally with the neighbouring ridges, forming a labyrinth-
like structure on the floor of the central valley. At the base of
the disto-lingual corner of the crown, there is a strongly laterally
protruding cingulum-like structure with a small cuspule. A small
cuspule is also present mesio-labially above the thick cingulum
which encircles the mesial margin. FCPT (CAN 1 ⁄ 1610) is a
fragment of a right M1. There is only the mesial portion of the
crown preserved, which is very similar in morphology to that of
the complete M1 described above. The first two lingual cusps
(L1 and L2) and the first labial one (B1) are present. These are
densely covered by thick and prominent radiating enamel ridges.
P4 ⁄ 5. FCPT (CAN 1 ⁄ 934) (paratype; L: 2.16; W: 1.33) is a P4
or a P5. Small portions of its mesio-labial, disto-labial and dis-
to-lingual corners are broken. Neither of the roots is preserved.
The tooth is rectangular in outline in occlusal view and consid-
erably lengthened mesio-distally. The cusp formula is 2B:4L.
Both rows of cusps are oriented clearly parallel to the mesio-dis-
tal axis. The apices in some cusps are also partially broken (B2
and L2). The lingual row of cusps covers the whole crown longi-
tudinally, whereas the labial one is considerably shortened, with
only two cusps confined to the mesial half of the crown. On the
distal margin of the crown, only the last lingual cusp (L4) is
present. This is confined to the disto-lingual corner, but due to
its large size it occupies almost the whole distal margin. The
cusps exhibit different sizes; their heights increases considerably
distally; L4 and B2 are the widest and highest cusps. Cuspules
are absent. The radiating enamel ridges are strongly developed.
They are long and prominent and cover the cusps densely from
the tip to the floor, intermeshing basally with the neighbouring
ridges. The tooth is wider in the middle than at the mesial and
distal margins. Both mesial and distal margins are gently
rounded. The mesial margin bears a thick but faint cingulum.
Comparison. The absence of the disto-lingual ridge on M1 pre-
vents it from being assigned to Plagiaulacidae Gill, 1872, Eobaa-
taridae, Allodontidae Marsh, 1889, Zofiabaataridae Bakker, 1992,
Arginbaataridae Hahn and Hahn, 1983, or to genus Glirodon
Engelmann and Callison 1999 (family incertae sedis). The disto-
lingual ridge is lacking, by contrast, in Paulchoffatiidae and
Pinheirodontidae, as in the M1 from La Cantalera. The latter
exhibits a 2B:4L cusp formula, with the absence of B3, and the
central valley opens disto-labially. These dental features are also
present on M1 of the paulchoffatiids Kuehneodon simpsoni Hahn,
1969 and Pseudobolodon oreas Hahn, 1977, and in M1 of the pin-
heirodontid Iberodon quadrituberculatus Hahn and Hahn, 1999.
These multituberculate fossils were found in the Late Jurassic and
Early Cretaceous (Berriasian) deposits from Portugal, respec-
tively. The morphology of the M1 from La Cantalera, however, is
closer to the pinheirodontid taxon than the paulchoffatiid taxa
EXPLANATION OF PLATE 1
Scanning electron micrographs of gold-palladium-coated teeth of Cantalera abadi gen. et sp. nov. (Pinheirodontidae,
Multituberculata) from La Cantalera (Josa, Teruel), Spain. A–E, left M1, holotype FCPT (CAN 1 ⁄ 1609). F–J, left P4 ⁄ 5, paratype FCPT
(CAN 1 ⁄ 934). A, F, occlusal; B, G, mesial; C, H, distal; D, I, lingual; and E, J, labial views.
1458 P A L A E O N T O L O G Y , V O L U M E 5 1
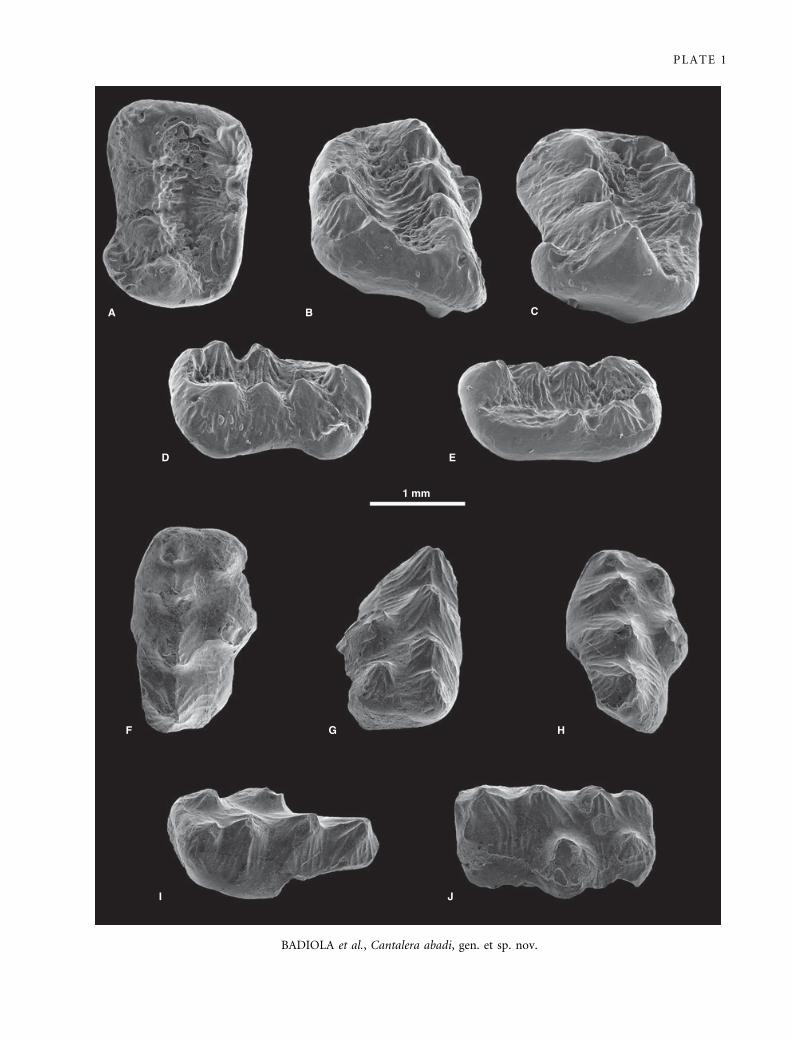
PLATE 1
BADIOLA et al., Cantalera abadi, gen. et sp. nov.
A B C
D
F G H
I J
E
1 mm

mentioned above. One of the diagnostic characters of pinheir-
odontids is the presence of the last cusp in each row of M1 either
enlarged or reduced. Both are reduced in I. quadrituberculatus,
whereas in the M1 under study, L4 is enlarged and B2 reduced.
There are other differences between the M1 of I. quadritubercula-
tus and the specimen from La Cantalera (see differential diagno-
sis). The low cusp which is present above the cingulum-like
structure in the M1 under study, located in the disto-lingual cor-
ner of the crown, could be an incipient radiating ridge like that
of the Berriasian pinheirodontid Sunnyodon notleti Kielan-Jaw-
orowska and Ensom, 1992 of the Purbeck Group of Dorset, Eng-
land, or an incipient row of small cuspules like that of I.
quadrituberculatus. It seems evident that it is not the typical fully
developed disto-lingual ridge of the eobaatarids. Moreover, eoba-
atarids have an M1 that is 3B:4L instead of 2B:4L.
Elongate and molariform upper premolars usually belong to
P4 or P5. The basic cusp formula of P4 in the suborder Plagiau-
lacida is 2-3B:4L, although most of them exhibit 3B:4L on P4
and P5, with the exception of Paulchoffatiidae, which have the
third row of cusps (row bb sensu Hahn and Hahn 2004: 120) in
P4 and P5, and of Pinheirodontidae, Albionbaataridae Kielan-
Jaworowska and Ensom, 1994, and the Allodontidae Morrison-
odon brentbaatar (Bakker, 1998), which exhibit a P5 with the
additional cusp row bb. Therefore, our specimen without row
bb and cuspules, and with a cusp formula of 2B:4L, could
belong to a P4 from a pinheirodontid taxon, or to a P4 or P5
from one of the following allodontid and eobaatarid taxa: the
Late Jurassic (Tithonian) allodontid Psalodon potens (Marsh,
1887) of North America, the Early Cretaceous eobaatarid Eobaa-
tar Kielan-Jaworowska et al. 1987 of the Iberian Peninsula (early
and late Barremian) and Asia (Aptian or Albian), or the late
Early Cretaceous (Albian ⁄ Cenomanian) eobaatarid Janumys ere-
bos Eaton and Cifelli, 2001 of North America. The presence of a
rectangular outline and cusps with considerably different heights,
however, allows us to assign the upper premolar of La Cantalera
to Pinheirodontidae instead of the other families mentioned
above. There are other differences too. In the P5 of P. potens the
inner flank of the lingual row is worn, forming a shearing edge;
the P5 of Eobaatar is characterized by having its main row of
cusps (the lingual one) obliquely oriented to the mesio-distal
axis, forming a cutting edge: consequently, the distal margin of
the crown is formed only by the lingual row of cusps; and the
P4 of J. erebos is clearly shorter and narrower than the tooth
from La Cantalera, and its cusps are less distinctly separated
from one another. The upper premolars of pinheirodontids
which have been interpreted as P4 do not have the third addi-
tional cusp row bb. They exhibit two rows of cusps, of which
the labial one is shorter than the lingual one, as in the premolar
from La Cantalera. We therefore assign it to the family
Pinheirodontidae. However, our specimen could belong either to
a P4 or a P5, because there is an upper premolar tentatively
assigned to the fifth premolar in the pinheirodontid Sunnyodon
notleti that does not have the row bb (Kielan-Jaworowska and
Ensom 1992, p. 107). We thus assign the premolar under study
to P4 ⁄ 5. The occlusal morphology of the P4 ⁄ 5 from La Cantal-
era is close to the Berriasian taxa S. notleti and Pinheirodontidae
gen. et sp. indet. from Portugal (Hahn and Hahn 1999) in hav-
ing cusp-rows oriented parallel to the mesio-distal axis, though
this character is more accentuated in the premolar from La Can-
talera. Other differences between them have also been described
(see differential diagnosis).
The great density and prominence of the radiating enamel
ridges observed in the teeth of Cantalera abadi gen. et sp. nov.
also exist in M1 of the albionbaatarid Proalbionbaatar plagiocyr-
tus Hahn and Hahn, 1998b and in P5 of the pinheirodontid Lav-
ocatia alfambrensis Canudo and Cuenca-Bescos, 1996, which
come from the Late Jurassic of Portugal and Early Cretaceous
(early Barremian) of Spain, respectively. However, the upper
premolar from La Cantalera differs from that of L. alfambrensis
(see differential diagnosis), and the M1 of La Cantalera can also
be distinguished from that of P. plagiocyrtus by its clearly differ-
ent number and arrangement of cusps. P. plagiocyrtus has a
greater number of cusps, and their apices are displaced some-
what to the labial side. The specimens under study belong to a
new pinheirodontid genus, which is characterized by a number
of autopomorphic characters (see diagnosis).
Family EOBAATARIDAE Kielan-Jaworowska et al. 1987
Genus EOBAATAR Kielan-Jaworowska et al. 1987
Eobaatar sp.
Plate 2
Referred specimens. Left P5, FCPT (CAN 1 ⁄ 935), and other left
P5 partially worn FCPT (CAN 1 ⁄ 1611).
Description
P5. FCPT (CAN 1 ⁄ 935) (L: 1.63; W: 1.35) exhibits a well-pre-
served crown, except that the apices of some cusps (L2-L4 and
B2) are slightly broken. Neither of the roots is preserved. The
cusp formula is 2B:4L. The main row of cusps, the lingual one,
is obliquely oriented to the mesio-distal axis. It extends from the
mesio-lingual corner towards the disto-labial corner of the
crown. The labial row of cusps is shortened, with only two cusps
(B3 is missing), and the disto-labial portion of the crown is con-
siderably narrowed, comprising only the last lingual cusp (L4).
Consequently, the occlusal surface is distinctly narrow, with a
EXPLANATION OF PLATE 2
Scanning electron micrographs of gold-palladium-coated teeth of Plagiaulacidae or Eobaataridae gen. et sp. indet. (Multituberculata)
from La Cantalera. A–E, left P1 ⁄ 3, FCPT (CAN 1 ⁄ 936). F–J, fragment of a right p4, FCPT (CAN 1 ⁄ 937). A, F, occlusal; B, mesial; C,
H, distal; G, mesio-labial; D, I, lingual; and E, J, labial views.
1460 P A L A E O N T O L O G Y , V O L U M E 5 1
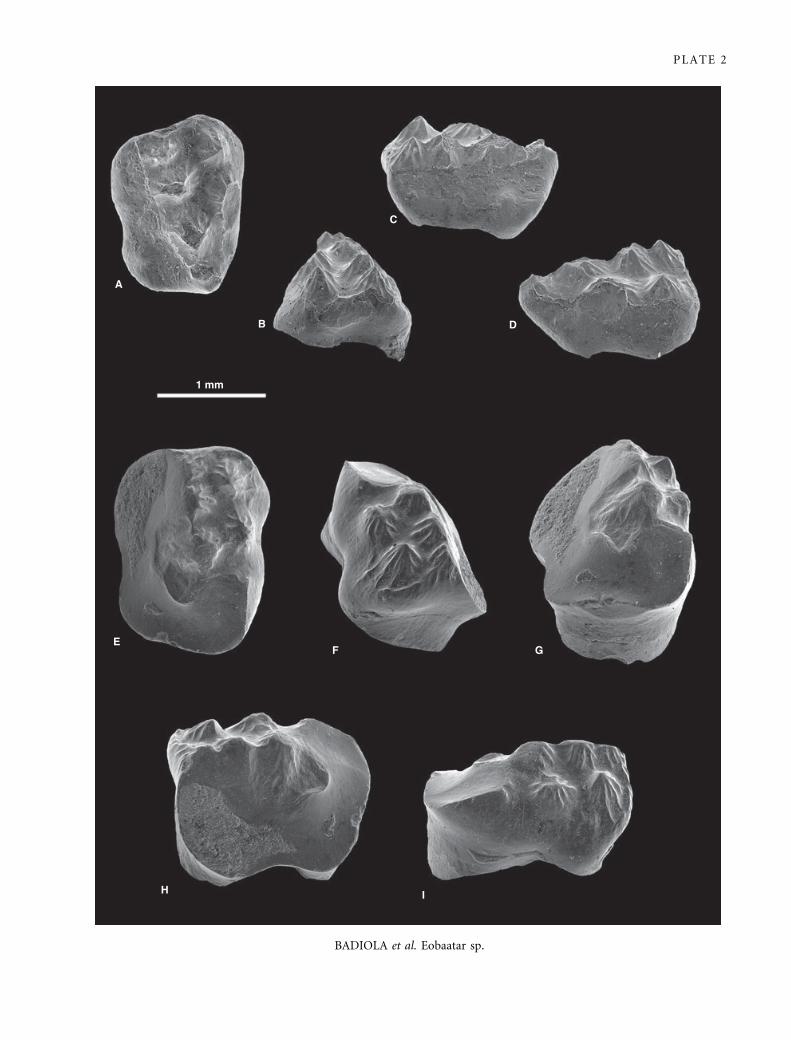
PLATE 2
BADIOLA et al. Eobaatar sp.
A
B
C
D
F G
H I
E
1 mm

triangular outline. In the lingual row of cusps, L2 and L3 are the
widest and highest cusps, whereas L1 and L4 are similar to one
another in size. In the labial row, B2 is considerably bigger than
B1. The cusps are conical, well separated from one another, and
covered by short and thin radiating enamel ridges. The latter are
only present on the cusps. There are no cuspules. The lingual
wall of the tooth extends below the cusps to form a smooth and
wide surface or wing (sensu Kielan-Jaworowska et al. 1987,
p. 12), which is strongly laterally protruding. Lateral to the two
labial cusps, the labial wall likewise forms a smooth wing,
although it is narrower and only mesio-labially protruding. The
mesial margin of the crown is somewhat concave in the middle,
whereas the distal margin is convex. Both the labial and lingual
walls are roughly concave in the middle. FCPT (CAN 1 ⁄ 1611) is
the other left P5, with severe wear on the lingual and distal mar-
gins of the crown. The last lingual cusp (L4) is worn away. Con-
sequently, the crown length value could be smaller than the
actual measurement (L: less than 1.96; W: 1.33). The morpho-
logical features of this specimen, however, are similar to those of
the P5 described above.
Comparison. The crown outline, and the cusp formula and cusp
arrangement of the P5 teeth under study are typical of the eo-
baatarid Eobaatar: the cusp formula is 2B:4L; the lingual row of
cusps is obliquely oriented to mesio-distal axis; the labial row of
cusps is shortened, and the distal portion of the crown is consid-
erably narrowed, comprising only the lingual row of cusps; and
lateral to the cusps of the labial row there is a small wing. The
size of the better-preserved P5 from La Cantalera (L: 1.63; W:
1.35) is similar to that of Eobaatar magnus Kielan-Jaworowska
et al. 1987 (L: 1.7; W: 1.2 in each of the four specimens) from
the Aptian or Albian beds at Khovboor (Gobi Desert, Mongolia),
but the eobaatarid of La Cantalera differs from E. magnus by the
absence of lingual cusps distinctly increasing in height distally,
and by having lateral to the cusps of the lingual row a smooth
and wide wing that covers the whole lingual margin longitudi-
nally and is strongly laterally protruding. The lingual wing in
E. magnus is not present or is incipiently developed disto-
lingually, though it is clearly less laterally protruding than the
one from La Cantalera. The P5 from La Cantalera seems smaller
than the holotype specimen of Eobaatar hispanicus Hahn and
Hahn, 1992 (Hahn and Hahn 1992: text-fig. 5; L: 1.92, W: 1.2)
of Galve (early Barremian), and differs from it in the absence of
lingual cusps distinctly increasing in size distally, and by having
a narrower occlusal surface, which exhibits a clearly developed
triangular outline, and strongly laterally protruding lingual and
labial wings. However, because the differences between the P5
specimens from La Cantalera and the P5 of E. magnus and
E. hispanicus are only slight or the features above mentioned
could be variable intraspecifically, other lower and upper teeth
must be studied in detail in order to be able to conclude that
the morphological differences observed between the premolars
from La Cantalera and those of E. hispanicus and E. magnus are
sufficiently great to regard the studied specimens as belonging to
a different species.
Plagiaulacidae or Eobaataridae gen. et sp. indet.
Plate 3
Referred specimens. Left P1 ⁄ 3, FCPT (CAN 1 ⁄ 936), and frag-
ment of a right p4, FCPT (CAN 1 ⁄ 937).
Description
P1 ⁄ 3. FCPT (CAN 1 ⁄ 936) is an anterior upper premolar, proba-
bly P2 or P3 (L: 1.06; W: 0.68). A small portion of both roots is
preserved. The crown is well preserved. It is roughly rectangular
in shape in occlusal view, with labial and lingual distal bulges,
and somewhat elongate mesio-distally. There are two lingual and
two labial cusps, which do not form clear longitudinal lingual
and labial rows. The cusps are similar in size, although B1 is
slightly smaller than the others and is displaced towards the
mesio-labial corner of the crown. B2 is confined to the midpoint
of the labial margin and placed transversally between the two
lingual cusps. The cusps are covered by thin and radiating
enamel ridges. These are short and present only on the cusps.
The mesial margin is roughly rounded, and two very small cus-
pules are present. The distal margin is convex due to the pres-
ence of a talonid-like ‘‘heel’’. The lingual margin is somewhat
concave in the middle, whereas the labial one is concave between
B1 and B2 and between the B2 and the talonid-like ‘‘heel’’.
p4. FCPT (CAN 1 ⁄ 937) is a fragment of a right p4 (L: bigger
than 2.26; W: 1.24). It cannot be known exactly how many cusps
and serrations are present on the cutting edge, because the tooth
is broken mesially. The projection of the first serration forwards
with a reasonable downward curvature suggests that at least a
quarter part of the mesial margin may be missing. The preserved
serrations are roughly of the same length, have the same distance
between them, and are bent similarly downwards. The first two
and last two serrations are worn or less marked than the others.
A prominent and dorsally worn shelf-like concave structure is
located close to the disto-labial corner. This structure has been
called the posterior wear shelf by Kielan-Jaworowska et al.
(1987: 6), and it seems to belong to several worn labial cusps
instead of only one. These could have been small and fewer in
number than the cusps on the cutting edge. The absence of
EXPLANATION OF PLATE 3
Scanning electron micrographs of gold-palladium-coated teeth of Eobaatar sp. (Eobaataridae, Multituberculata) from La Cantalera.
A–D, left P5, FCPT (CAN 1 ⁄ 935). E–I, left P5 with severe wear on the lingual and distal margins of the crown, FCPT (CAN 1 ⁄ 1611).
A, E, occlusal; B, F, mesial; G, distal; C, H, lingual; and D, I, labial views.
1462 P A L A E O N T O L O G Y , V O L U M E 5 1
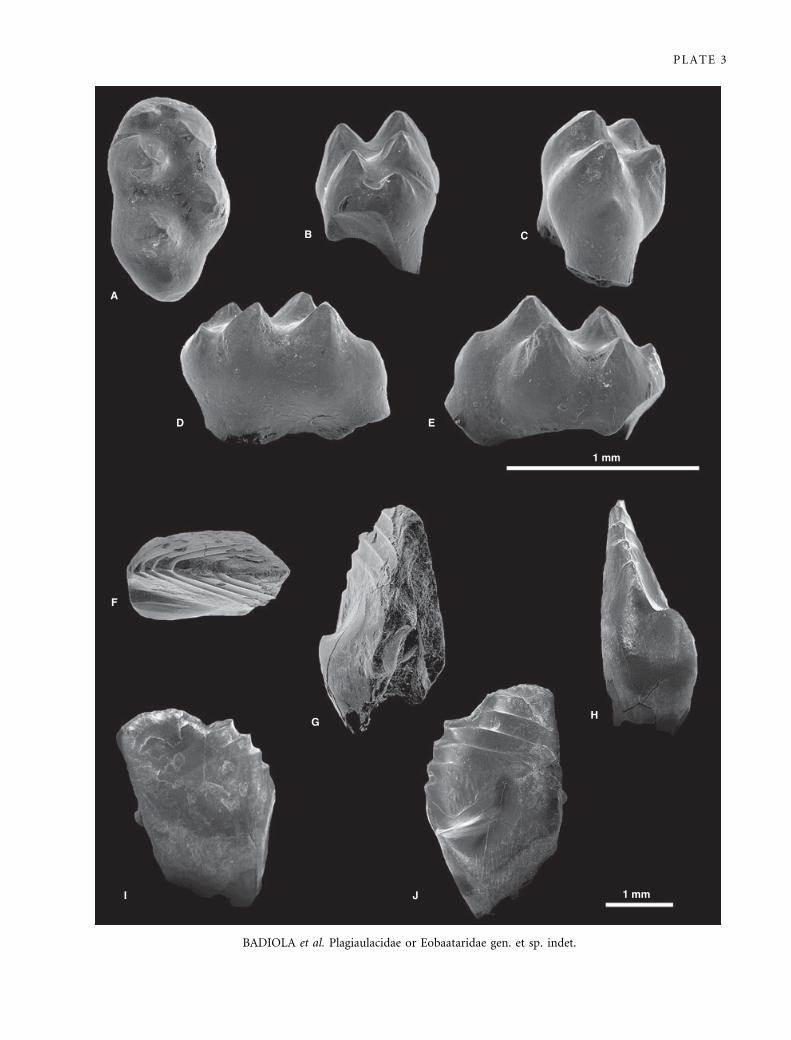
PLATE 3
BADIOLA et al. Plagiaulacidae or Eobaataridae gen. et sp. indet.
A
B C
D
F
GH
I J
E
1 mm
1 mm
horizontal erosion on the cutting edge indicates that the premo-
lar could have been used for cutting, instead of for shearing.
Comparison. The morphology, outline and number of cusps of
the anterior upper premolars are quite similar in the Plagiaulaci-
dae, Eobaataridae, Allodontidae and Arginbaataridae, where they
are short, rounded and tricuspid, whereas in the Paulchoffatiidae
and Pinheirodontidae they are tri- to tetracuspid, with or with-
out additional very small cuspules. Anterior upper premolars are
unknown in Hahnodontidae Sigogneau-Russell, 1991 (Hahn and
Hahn 2004, tables 1, 7). Nevertheless, some exceptions with
respect to the number of cusps and the arrangement of cusp-
rows have been described. Pentacuspid or hexacuspid P1 ⁄ 3 are
present in ?Plagiaulacidae gen. et sp. indet. (Kielan-Jaworowska
and Ensom 1992, pl. 5, figs. 5–6) and in the paulchoffatiid Kie-
lanodon hopsoni (Hahn, 1987), whereas in others there appears a
transitional stage towards P4, with two clear longitudinal rows of
cusps (usually in paulchoffatiids, e.g. Pseudobolodon krebsi Hahn
and Hahn, 1994) or even with three longitudinal cusp-rows (in
albionbaatarids). However, only the plagiaulacid Parabolodon
elongatus (Simpson, 1928), the eobaatarid Eobaatar? pajaronensis
Hahn and Hahn, 2001, and the specimen described here exhibit
a P1 ⁄ 3 mesio-distally lengthened with cusps not arranged in
clear longitudinal rows. The morphology of the P1 ⁄ 3 from La
Cantalera differs from P. elongatus and Eobaatar? pajaronensis for
the reasons set out below, and seems to belong to a different
taxon. The P1 ⁄ 3 under study can be assigned either to Eobaa-
taridae or to Plagiaulacidae. The size of the P1 ⁄ 3 from La Can-
talera is similar to that of E.? pajaronensis, but it differs from it
in having four cusps instead of three, a distal margin that forms
a fully developed talonid-like ‘heel’, which is missing in E.? paja-
ronensis, and the last lingual cusp confined to near the disto-lin-
gual corner. In E.? pajaronensis, by contrast, this has been
displaced towards the middle, alone forming the distal margin of
the crown. P. elongatus is like the P1 ⁄ 3 from La Cantalera in
having a similar talonid-like ‘heel’ behind the cusps, and in the
last lingual cusp being confined to near the disto-lingual corner.
However, the premolar from La Cantalera is smaller than the
smallest premolar (P3) of P. elongatus, and there are four cusps
instead of three. The presence of more than four cusps, but less
than 15, on the cutting edge in the p4 fragment from La Cantal-
era and the fact that the second serrations that is not distinctly
longer than the others allow us to assign it to Eobaataridae or
Plagiaulacidae, as opposed to Paulchoffatiidae, Pinheirodontidae,
Allodontidae, Zofiabaataridae, Glirodon, or Arginbaataridae. The
morphology of the p4 fragment, however, is closer to a p4 of the
Plagiaulacidae than of the Eobaataridae. In the p4 specimens
known within the family Eobaataridae (Eobaatar and Sinobaatar
Hu and Wang, 2002), only one labial cusp is located close to the
distal border of the crown. In contrast, the size of the worn
shelf-like concave structure of the p4 from La Cantalera suggests
that possibly more than one labial cusp existed in the original
unworn tooth, as is the case in plagiaulacids. Therefore, the p4
and the P1 ⁄ 3 from La Cantalera could belong to the family Pla-
giaulacidae rather than to the Eobaataridae. However, because of
the fragmentary state of the p4 under study and the absence of
other tooth specimens, we have assigned the P1 ⁄ 3 and p4 in
question neither to a specific family nor to a specific taxon.
Plagiaulacida indet.
Text-fig. 2
Referred specimens. Fragment of an I2, FCPT (CAN 1 ⁄ 939), and
fragment of a ?ix, FCPT (CAN 1 ⁄ 938).
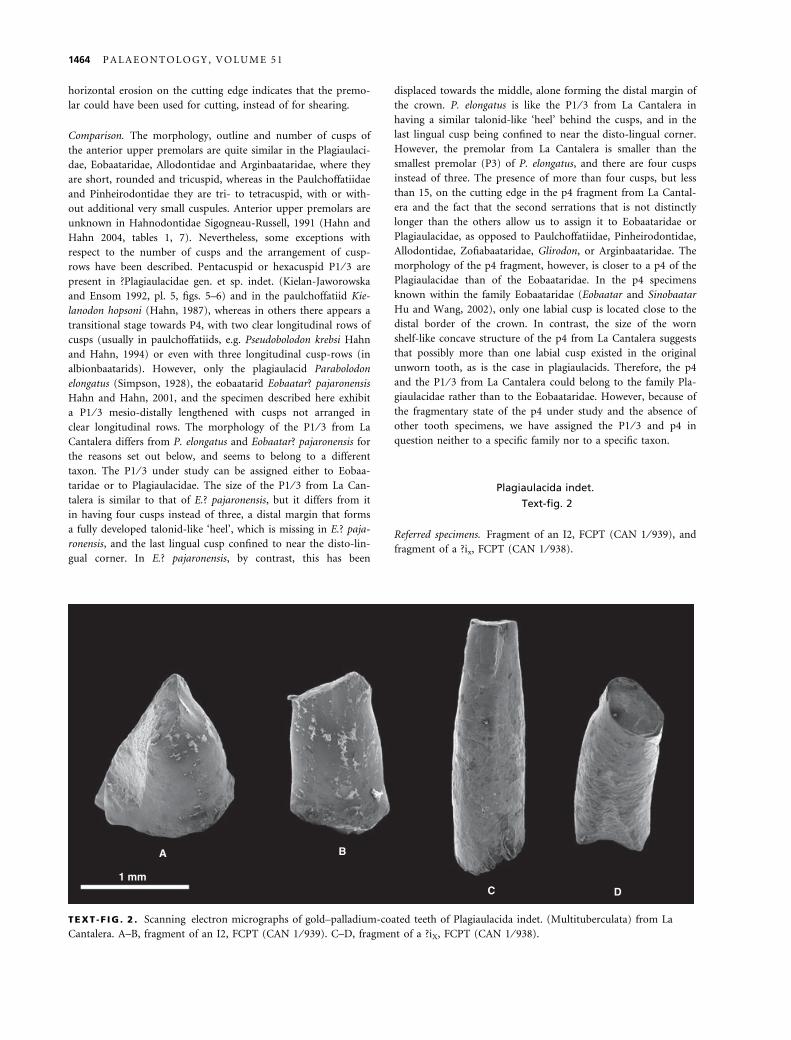
A B
1 mmC D
TEXT -F IG . 2 . Scanning electron micrographs of gold–palladium-coated teeth of Plagiaulacida indet. (Multituberculata) from La
Cantalera. A–B, fragment of an I2, FCPT (CAN 1 ⁄ 939). C–D, fragment of a ?iX, FCPT (CAN 1 ⁄ 938).
1464 P A L A E O N T O L O G Y , V O L U M E 5 1

Description
I2. FCPT (CAN 1 ⁄ 939) is a fragment of an I2 (L: c. 0.94; W:
0.53). Its distal margin is broken, and we cannot see how many
cusps there were. The cusp of the mesial margin is the highest
and widest cusp, and this seems the only functional one. The
labial flank of the crown is convex, whereas the lingual one is
broken.
ix. FCPT (CAN 1 ⁄ 938) is a fragment of an ix. There is a very
small fragment of the crown and another of the root preserved.
Neither the enamel development nor the crown morphology can
be described in this specimen.
Comparison. The I2 and ix seem to correspond in size to the
same multituberculate taxon described above for P1 ⁄ 3 and p4 or
for P5. However, they are too fragmentary for it to be possible
to observe the diagnostic characters of these incisors (the num-
ber of distal cusps on the I2 and the enamel development in the
?ix) in order to assign them to a specific family within the sub-
order Plagiaulacida. Therefore, they are provisionally described
as Plagiaulacida indet.
PALAEOBIOGEOGRAPHICALIMPLICATIONS
The late Hauterivian multituberculate fossil assemblage
from La Cantalera consists of some taxa (Pinheirodonti-
dae and probably ?Plagiaulacidae) that were restricted to
what is now Western Europe, whereas others (Eobaatari-
dae) have also been described from Asia. The two premo-
lars described as Plagiaulacidae or Eobaataridae gen. et
sp. indet. could belong to a new plagiaulacid taxon, in
which case it would be the first in the Iberian Peninsula.
To date, these multituberculates have only been described
from the Berriasian deposits of England (Owen 1871;
Simpson 1928; Kielan-Jaworowska and Ensom 1992,
1994; Sigogneau-Russell and Kielan-Jaworowska 2002;
Hahn and Hahn 2004). Pinheirodontids are described
from the Berriasian deposits of England and Portugal
(Kielan-Jaworowska and Ensom 1992; Hahn and Hahn
1999), and in the late Hauterivian (this paper) and early
and late Barremian deposits of Spain (Canudo and
Cuenca-Bescos, 1996; Cuenca-Bescos et al. 1996; Badiola
et al. 2007a). Eobaatarids are present in Europe (England
and Spain) and Asia (China, Japan and Mongolia). Those
from England are located in the Early Cretaceous (Weal-
den) deposits of Cliff End (Wadhurst Fm., south Eng-
land) and of the Isle of Wight (Wessex Fm.), which are
dated as Valanginian and Barremian, respectively: at the
Cliff End site, Loxaulax valdensis (Woodward 1911) has
been described from a few specimens (Clemens 1963,
1971; Clemens and Lees 1971), and two Barremian speci-
mens from the Wessex Formation of the Isle of Wight
have also been assigned to the genus Loxaulax Simpson,
1928, without specific determination, by Butler and Ford
(1975). New multituberculate fossils have been recovered
in this area, the study of which by Steven C. Sweetman is
still in progress. Two species of the eobaatarid Eobaatar
are described from the early Barremian of Spain: Eobaatar
hispanicus and Eobaatar? pajaronensis (Hahn and Hahn
1992, 2001, 2002). New specimens which belong to Eoba-
atar have recently been identified in the late Barremian
bed at Vallipon (province of Teruel), in the Aragonese
branch of the Iberian Ranges, in northeastern Iberia
(Badiola et al. 2007a). Some of these were previously
described as Eobaatarinae indet. (Cuenca-Bescos et al.
1996). In this paper, the oldest representative of Eobaatar
is described. It comes from the late Hauterivian–early
Barremian site of La Cantalera (Josa, Teruel), another site
in the Aragonese branch of the Iberian Ranges. The age
of the Asiatic localities that have yielded multituberculate
fossils is still subject to debate. Such fossils come from
the Jehol and Tetori Groups of China and Japan, and
from the Gobi Desert in Mongolia. The only eobaatarid
taxon described from the Jehol Group, in the Yixian For-
mation, is Sinobaatar lingyuanensis Hu and Wang, 2002.
The Jehol Biota is currently thought to date from about
130–120 Myr (Meng et al. 2006 and references therein),
and so is Barremian to Aptian in age (Gradstein et al.
2004). There are also two other formations (the Shahai
and Fuxin formations; e.g. Wang et al. 1995) that are
stratigraphically younger than the Yixian Formation,
which have yielded some multituberculate fossils that are
still unpublished (Kusuhashi, pers. comm. 2007). Two
other species of Eobaatar are described from the Aptian
or Albian beds at Khovboor (Gobi Desert, Mongolia; Kie-
lan-Jaworowska et al. 1987): Eobaatar magnus and Eobaa-
tar minor Kielan-Jaworowska et al. 1987. The age of the
fossiliferous beds located in the Japanese Tetori Group is
more obscure, because there are no good radiometric
dates available and reliable index fossils are rare. The age
of the Kuwajima Formation ranges from Valanginian to
Aptian (Matsukawa et al. 2006; Rougier et al. 2007),
although the Zircon U-Pb ages collected from tuff beds
indicate a shorter period, Barremian or early Aptian
(Kusuhashi et al. 2006). The multituberculate remains of
the Kuwajima Formation were previously described as Eo-
baataridae indet. (Takada et al. 2001), but recently have
been assigned to new eobaatarid taxa (Kusuhashi 2008).
The occurrence of Eobaatar in the Iberian Peninsula,
from the late Hauterivian to late Barremian, and in Asia
during the Aptian or Albian, indicates that some faunal
exchange could have taken place between these areas dur-
ing the Neocomian. The palaeogeographic models indi-
cate that it was possible (Smith et al. 1994; Smith and
Rush 1997). Other palaeontological data support this
hypothesis, as well as the vertebrate faunal exchange
B A D I O L A E T A L . : N E W M U L T I T U B E R C U L A T E M A M M A L S F R O M T H E H A U T E R I V I A N ⁄ B A R R E M I A N T R A N S I T I O N 1465

between other parts of Laurasia and Gondwana. The
mammalian family Gobiconodontidae is recorded in the
?Berriasian of Africa (Morocco), in the early and late
Barremian of Europe (England and Spain), in the Hauter-
ivian to Aptian of eastern and central Asia, and in the
Aptian–Albian of North America (Cuenca-Bescos and
Canudo 2003; Sweetman 2006; and references therein),
and some Barremian dinosaur taxa (at generic level) are
observed to be common to the Iberian Peninsula, Asia
and North America, indicating intermittent faunal
exchange between these areas of Laurasia during the Bar-
remian (Milner et al. 2000; Cuenca-Bescos and Canudo
2003; Canudo 2006). However, the presence of Eobaatar,
together with the presence of an euhelopodid dinosaur, in
the late Hauterivian deposits of La Cantalera, in the Ibe-
rian Peninsula (Canudo et al. 2002; Ruiz-Omenaca et al.
2005), suggests that an Early Cretaceous geographical
connection between Europe and Asia could have existed
from the Hauterivian to the Barremian (for c. 11 myr),
either sporadically or constantly for most of the Early
Cretaceous, thus making mammalian and dinosaur faunal
exchange possible.
The area of origin of the family Eobaataridae is still
unknown. The paucity of the fossil record for these multi-
tuberculates, and the absence of well-dated deposits in
Asia, currently inhibits the use of cladistic analysis for
investigating the phylogenetic and palaeobiogeogaphic
relationships between the different taxa. However, the
oldest representative of Eobaatar at La Cantalera, together
with those of the early and late Barremian found in other
Iberian localities, as well as the species from Asia, will be
essential in studying the area of origin and the dispersals
of this eobaatarid.
CONCLUSIONS
The multituberculate fossil assemblage of La Cantalera
(Teruel, Aragon), one of the sites in the Aragonese
branch of the Iberian Ranges in northeastern Iberia,
indicates that the Early Cretaceous (late Hauterivian)
multituberculate fauna of the Iberian Peninsula was
fairly diverse. The fossils consist of endemic western
European taxa (Pinheirodontidae) that are known from
the Late Jurassic and other taxa (the eobaatarid Eobaa-
tar) that were originally described from the Aptian-Al-
bian deposits of Mongolia, Asia. On the basis of the
palaeobiogeographic interpretation of certain mammals
(Gobiconodontidae) and dinosaurs (some Ornithopoda)
a European-Asian connection during the late Barremian
has been previously suggested. New multituberculate
finds from the Hauterivian ⁄ Barremian transition of the
Iberian Peninsula, at La Cantalera site, suggest that the
Early Cretaceous geographical connection between these
areas could have existed from the late Hauterivian,
either sporadically or constantly for most of the Early
Cretaceous, thus making mammalian and dinosaur fau-
nal exchange possible.
Acknowledgements. We thank Drs A. Currant and J. J. Hooker
for their help and giving access to specimens in the Natural His-
tory Museum of London. The journal referees (one of them was
Dr Zofia Kielan-Jaworowska) of this paper provided helpful
comments on an earlier version of the manuscript. Dr Nao
Kusuhashi (Institute of Vertebrate Palaeontology and Palaeoan-
thropology, Chinese Academy of Sciences, Beijing, China) pro-
vided information on the Asian multituberculate faunas.
Financial support was provided by the Ministerio de Educacion
y Ciencia (MEC) (research projects CGL2004 ⁄ 03393 ⁄ BTE and
CGL2007 ⁄ 62469 ⁄ BTE), the Universidad de Zaragoza (‘Financ-
iacion de Grupos Consolidados’ 2008), and the Gobierno de Ar-
agon (Direccion General de Patrimonio Cultural de Gobierno de
Aragon and Programa Europa XXI de Estancias de Investigacion
CAI-Gobierno de Aragon) of Spain. A. Badiola also acknow-
ledges support from the Programa Juan de la Cierva of the
Ministerio de Educacion y Ciencia of Spain.
REFERENCES
A M E GH I N O, F. 1889. Contribution al conocimiento de los
mamıferos fosiles de la Republica Argentina. Actas Academia
Nacional de Ciencias Cordoba, Buenos Aires, 6, 1–207.
A U R E L L , M., B A DE N A S , B., C A N UD O , J. I. and R U I Z -
O M E N A C A , J. I. 2004. Evolucion tectosedimentaria de la
Formacion Blesa (Cretacico Inferior) en el entorno del yacimi-
ento de vertebrados de La Cantalera (Josa, Teruel). Geogaceta,
35, 11–14.
B A DI OL A , A., C A N UD O, J. I. and CU E N CA - B E S C O S , G.
2007a. Los fosiles de multituberculados del Hauteriviense y
Barremiense de la Penınsula Iberica. In IV Jornadas Internacio-
nales sobre Paleontologıa de Dinosaurios y su Entorno, Abstracts.
Sala de los Infantes, Burgos, Spain, 47–48.
—— —— —— 2007b. Multituberculate fossils from the Early
Cretaceous of La Cantalera (Teruel, Spain). Journal of Verte-
brate Paleontology, 27 (Suppl. 3), 42A.
B A K KE R , R. T. 1992. Zofiabaataridae, a new family of multi-
tuberculate mammals from the Breakfast Bench fauna at
Como Bluff. Hunteria, 2, 24.
—— 1998. Dinosaur mid-life crisis: the Jurassic–Cretaceous tran-
sition in Wyoming and Colorado. New Mexico Museum of
Natural History and Science Bulletin, 14, 67–77.
B R OO K E S KN I G HT , J., B A T T E N , R. L., Y O CH E L S O N ,
E. L. and C OX , L. R. 1960. Paleozoic and some Mesozoic
Caenogastropoda and Opistobranchia. 310–331. In M O O R E , C.
(ed.). Treatise on Invertebrate Paleontology, Part 1, Mollusca 1.
Geological Society of America and University of Kansas Press,
Lawrence.
B UT L E R , P. M. and F O R D, R. 1975. Discovery of Cretaceous
mammals on the Isle of Wight. Proceedings of the Isle of Wight,
Natural History and Archaeological Society, 7, 662–663.
1466 P A L A E O N T O L O G Y , V O L U M E 5 1

—— and HO O KE R , J. J. 2005. New teeth of allotherian mam-
mals from the English Bathonian, including the earliest multi-
tuberculates. Acta Palaeontologica Polonica, 50, 185–207.
C A N U DO , J. I. 2006. La ambiguedad paleobiogeografica de los
dinosaurios ibericos durante el Cretacico Inferior. 21–45. In
COLECTIVO ARQUEOLOGICO SALENSE (ed.). Actas de
las III Jornadas sobre Dinosaurios y su Entorno. Sala de los
Infantes, Burgos, Spain.
—— and C U E N C A - B E S C O S , G. 1996. Two new mamma-
lian teeth (Multituberculata and Peramura) from the Lower
Cretaceous (Barremian) of Spain. Cretaceous Research, 17,
215–228.
—— R U I Z - O M E N A C A , J. I., B A R C O, J. L. and R OY O-
T OR R E S , R. 2002. ¿Sauropodos asiaticos en el Barremiense
inferior (Cretacico Inferior) de Espana? Ameghiniana, 39, 443–
452.
C H OW , M. and R I CH , T. H. 1984. A new triconodontan
(Mammalia) from the Jurassic of China. Journal of Vertebrate
Paleontology, 3, 226–231.
C L E M E N S , W. A. 1963. Wealden mammalian fossils. Palaeon-
tology, 6, 55–69.
—— 1971. Mammalian evolution in the Cretaceous. Zoological
Journal of the Linnean Society, 50, 165–180.
—— and L E E S , P. M. 1971. A review of English Early Creta-
ceous mammals. Zoological Journal of the Linnean Society, 50,
117–130.
C OP E , E. D. 1884. The Tertiary Marsupialia. American Natural-
ist, 18, 686–697.
C R US A FO N T , M. and G I B E R T , J. 1976. Los primeros
Multituberculados de Espana. Nota preliminary. Acta Geolo-
gica Hispanica, 11, 57–64.
C U E N C A - B E S C OS , G. and C A N UD O, J. I. 2003. A new
gobiconodontid mammal from the Early Cretaceous of Spain
and its palaeobiogeographical implications. Acta Palaeontolog-
ica Polonica, 48, 575–582.
—— —— 2004. Los mamıferos del Cretacico Inferior de Espana.
Geo-Temas, 6, 35–38.
—— —— and R UI Z - OM E N A C A , J. I. 1996. Los mamıferos
del Barremiense superior (Cretacico Inferior) de Vallipon
(Castellote, Teruel). Mas de las Matas, 15, 105–137.
E A T O N , J. G. and C I F E L L I , R. L. 2001. Multituberculate
mammals from near the Early–Late Cretaceous boundary,
Cedar Mountain Formation, Utah. Acta Palaeontologica Polo-
nica, 46, 453–518.
E N G E L M A N N , G. F. and C A L L I S O N , G. 1999. Glirodon
grandis, a new multituberculate mammal from the Upper
Jurassic Morrison Formation. 161–177. In G I L L E T E , D. D.
(ed.). Vertebrate Paleontology in Utah. Utah Geological Survey,
Special Publication, 99, Salt Lake City.
G I L L , T. H. 1872. Arrangement of the familias of mammals.
With analytical tables. Prepared for the Smithsonian Institu-
tion. Smiithsonian Miscellaneous Collection, 230, 1–98.
G R A D S T E I N , F. M., O G G, J. G., S M I T H , A. G., A GT E R -
B E R G, F. P., B L E E KE R, W., CO O PE R , R. A., D A V Y -
DO V , V., GI B BA R D , P., HI N N O V , L., H OU S E , M. R.,
L OU R E N S , L., L U T E R BA C H E R , H.-P., M C A R T H UR ,
J., M E L C H I N , M. J., R O B B, L. J., S H E R G OL D , J.,
V I L L E N E U V E , M., W A R D LA W , B. R., A LI , J.,
B RI N K HU I S , H., H I LG E N , F. J., HO O KE R , J. J., H O-
W A R T H , R. J., KN O L L , A. H., L A S KA R , J., M ON E -
CH I , S., P O W E L L , J., PL U M B , K. A., R A FF I , I., R OH L ,
U., S A N FI L I P PO , A., S C HM I T Z , B., S H A C KL E T O N ,
N. J., S H I E L D S , G. A., S T R A U S S , H., V A N D A M , J.,
V E I Z E R , J., V A N KO L F S C HO T E N , T. H. and W I L -
S O N , D. 2004. A Geologic Time Scale. Cambridge University
Press, Cambridge, 589 pp.
G R A M B A S T , L. 1968. Evolution of the utricule in the Charo-
phyte genera Perimneste Harris and Atopochara Peck. Journal
of the Linnean Society of London (Botany), 61, 5–11.
H A H N , G. 1969. Beitrage zur Fauna der Grube Guimarota Nr.
3. Die Multituberculata. Palaeontographica, 133, 1–100.
—— 1977. Neue Schadel-Reste von Mulituberculaten (Mamm.)
aus dem Malm Portugals. Geologica et Paleontologica, 11, 161–
168.
—— 1987. Neue Beobachtungen zum Schadel- und Gebiss-Bau
del Paulchoffatiidae (Multituberculata, Ober-Jura). Palaeo-
vertebrata, 17, 155–196.
—— and HA H N , R. 1983. Multituberculata. Fossilium Catalo-
gus. I. Animalia, 127, 1–409.
—— —— 1992. Neue Multituberculaten-Zahne aus der Unter-
Kreide (Barremium) von Spanien (Galve und Una). Geologica
et Paleontologica, 26, 143–162.
—— —— 1994. Nachweis des Septomaxillaire bei Pseudobolodon
krebsi n. sp. (Multituberculata) aus dem Malm Portugals.
Berliner geowissenschaftliche Abhandlungen E, 13, 9–29.
—— —— 1998a. Neue Beobachtungen an Plagiaulacoidea
(Multituberculata) des Ober-Juras. 3. Der Bau der Molares bei
den Paulchoffatiidae. Berliner geowissenschaftliche Abhand-
lungen E, 28, 39–84.
—— —— 1998b. Neue Beobachtungen an Plagiaulacoidea (Mul-
tituberculata) des Ober-Juras. 4. Ein Vertreter der Albionbaa-
taridae im Lusitanien Portugals. Berliner geowissenschaftliche
Abhandlungen E, 28, 85–89.
—— —— 1999. Pinheirodontidae n. fam. (Multituberculata,
Mammalia) aus der tiefen Unter-Kreide Portugals. Palaeonto-
graphica, 253, 77–222.
————2000. The multituberculates from the Guimarota mine.
97–108. In M A RT I N , T. and K R E BS , B. (eds). Guimarota.
A Jurassic Ecosystem, Verlang Dr. Friedrich Pfeil, Munchen,
Germany.
—— —— 2001. Multituberculaten-Zahne aus der Unter-Kreide
(Barremium) von Pie Pajaron (Prov. Cuenca, Spanien). Pala-
ontologische Zeitschrift, 74, 587–589.
—— —— 2002. Neue Multituberculaten-Zahne aus dem Barre-
mium (Unter-Kreide) von Galve (Spanien). Palaontologische
Zeitschrift, 76, 257–259.
—— —— 2004. The dentition of the Plagiaulacida (Multituber-
culata, Late Jurassic to Early Cretaceous). Geologica et Palaeon-
tologica, 38, 119–159.
—— —— 2006. Fossilium Catalogus. I. Animalia. Catalogus
Plagiaulacidorum cum figuris (Multituberculata suprajurassica
et subcretacea) 140. Backhuys Publishers, Leiden, 344 pp.
H E N K E L , S. and K R E BS , B. 1969. Zwei Saugetier-Unterkiefer
aus der Unteren Kreide von Una (Prov. Cuenca, Spanien).
Neues Jahrbuch fur Geologie und Palaontologie Monatshefte, 8,
449–463.
B A D I O L A E T A L . : N E W M U L T I T U B E R C U L A T E M A M M A L S F R O M T H E H A U T E R I V I A N ⁄ B A R R E M I A N T R A N S I T I O N 1467

H U, Y. and W A N G , Y. 2002. Sinobaatar gen. nov.: first multi-
tuberculate from the Jehol Biota of Liaoning, northeast China.
Chinese Science Bulletin, 47, 382–386 (in Chinese with English
summary).
K I E L A N - J A W O RO W S K A , Z. and E N S O M , P. C. 1992.
Multituberculate mammals from the Upper Jurassic Purbeck
Limestone Formation of southern England. Palaeontology, 35,
95–126.
—— —— 1994. Tiny plagiaulacoid multituberculate mammals
from the Purbeck Limestone Formation of Dorset, England.
Palaeontology, 37, 17–31.
—— and G A M BA R Y A N , P. 1994. Postcranial anatomy and
habits of Asian multituberculate mammals. Fossil and Strata,
36, 1–92.
—— and H UR U M , J. H. 2001. Phylogeny and Systematics of
multituberculate mammals. Palaeontology, 44, 389–429.
—— D A S H Z E V E G , D. and T R OF I M O V , B. A. 1987. Early
Cretaceous multituberculates from Mongolia and a compari-
son with Late Jurassic forms. Acta Palaeontologica Polonica,
32, 3–47.
—— C I F E L L I , R. and LU O , Z. H. 2004 (eds). Chapter 8.
Allotherians. 249–342. In Mammals from the age of Dinosaurs.
Origin, Evolution and Structure. Columbia University Press,
New York.
K N E UP E R - H A A C K, F. 1966. Ostracoden aus dem Wealden
der Sierra de los Cameros. 165–209. In BE U T HE R , A.,
DA H M , H. and KN E UP E R - HA A CK , F. Der Jura und
Wealden in Nordost-Spanien. Bundesanstalt fur Bodenforsch-
nung und den Geologischen Landesamtern der Bundesrepub-
lik Deutschland (Beihefte zum Geologischen Jahrbuch, 44),
Hannover.
K US U H A S H I , N. 2008. Early Cretaceous multituberculate
mammals from the Kuwajima Formation (Tetori Group), cen-
tral Japan. Acta Palaeontologica Polonica, 53, 379–390.
—— M A T S U M O T O, A., M UR A K A M I , M., T A G A M I , T.,
HI R A T A , T., I I ZU K A , T., HA N DA , T. and M A T S -
UO KA , H. 2006. Zircon U-Pb ages from tuff beds of the
upper Mesozoic Tetori Group in the Shokawa district, Gifu
Prefecture, central Japan. Island Arc, 15, 378–390.
M C KE N N A , M. C. 1971. Multituberculata. 736–737. In
McGraw-Hill Enciclopedia of Science and Technology 8.
McGraw-Hill Book Company, New York.
M A R S H, O. C. 1887. American Jurassic mammals. American
Journal of Science, Series, 3, 33, 227–348.
—— 1889. Discovery of Cretaceous Mammalia. American Journal
of Science, Series, 3, 38, 81–92.
M A R TI N - CL O S A S , C. 1996. A phylogenetic system of Clava-
toracea (fossil Charophyta). Review of Palaeobotany and Paly-
nology, 94, 259–293.
—— and G R A M B A S T - F E S S A R D , N. 1986. Les charophytes
du Cretace inferieur de la region du Maestrat (Chaıne Iberi-
que, Catalanides, Espagne). Paleobiologie Continentale, 15, 1–
66.
M A TS U K A W A , M., I T O , M., N I S H I D A , N., K O A R A I ,
K., L OC K L E Y , M. G. and N I CH O LS , D. J. 2006. The Cre-
taceous Tetori biota in Japan and its evolutionary significance
for terrestrial ecosystems in Asia. Cretaceous Research, 27,
199–225.
M A Y OR A L , E. and S E Q UE I R OS , L. 1983. El Cretacico Infe-
rior en la region de Plou-Cortes de Aragon-Josa (Teruel).
Boletın de la Real Sociedad Espanola de Historia Natural
(Geologıa), 81, 111–123.
M E N G, J., H U, Y., L I , C. and W A N G , Y. 2006. The mammal
fauna in the Early Cretaceous Jehol Biota: implications for
diversity and biology of Mesozoic mammals. Geological Jour-
nal, 41, 439–463.
M I L N E R , A. C., M I L N E R , A. R. and E V A N S , S. E. 2000.
Amphibians, reptiles and birds: a biogeographic review. 316–
332. In C UL V E R , S. J. and R A W S O N , P. F. (eds). Biotic
Response to Global Change – The Last 145 Million Years. Cam-
bridge University Press, Cambridge.
O W E N , R. 1871. Monograph of the fossil Mammalia of the
Mesozoic Formations. Palaeontographical Society, 33, 1–115.
R O UG I E R , G. W., I S A J I , S. and M A N A B E , M. 2007. An
early Cretaceous mammal from the Kuwajima Formation
(Tetori Group), Japan, and a reassessment of triconodont
phylogeny. Annals of Carnegie Museum, 76, 73–115.
R U I Z - O M E N A C A , J. I., C A N UD O , J. I. and C UE N CA -
B E S CO S , G. 1997. Primera evidencia de un area de alimen-
tacion de dinosaurios herbıvoros en el Cretacico Inferior de
Espana (Teruel). Monografıas de la Academia de Ciencias Exac-
tas, Fısicas, Quımicas y Naturales de Zaragoza, 10, 1–48.
—— —— A U R E L L , M., BA D E N A S , B., B A R C O, J. L.,
C U E N C A - B E S C O S , G. and I PA S , J. 2004. Estado de las
investigaciones sobre los vertebrados del Jurasico Superior y
Cretacico Inferior de Galve (Teruel). Estudios Geologicos, 60,
179–202.
—— —— CR U Z A DO - CA BA L L E R O , P., I N F A N T E , P.
and M O R E N O - A Z A N Z A , M. 2005. Baryonychine teeth
(Theropoda: Spinosauridae) from the Lower Cretaceous of La
Cantalera (Josa, NE Spain). Kaupia, 14, 59–63.
R U S S E L L , D. A. 1993. The role of Central Asia in dinosaurian
biogeography. Canadian Journal of Earth Sciences, 30, 2002–
2012.
S C H UD A CK , M. E. 1989. Charophytenflores aude den unter-
kretazischen Vertebraten-Fundschicten bei Galve und Una
(Ostspanien). Berliner geowissenschaftliche Abhandlungen Reihe
A (Geologie und Palaontologie), 106, 409–443.
S I GO G N E A U - R US S E LL , D. 1991. First evidence of Multi-
tuberculata (Mammalia) in the Mesozoic of Africa. Neues
Jahrbuch fur Geologie und Palaontologie, Monatshefte, 2, 119–
125.
—— and K I E L A N - J A W O R OW S KA , Z. 2002. Mammals
from the Purbeck Limestone Group of Dorset, southern Eng-
land. Special Papers in Palaeontology, 68, 241–255.
S I M P S ON , G. G. 1928. A catalogue of the Mesozoic Mammalia
in the Geological Department of the British Museum, British
Museum Natural History, London. 215 pp.
S K E L T ON , P. W., S PI CE R , R. A., K E L L E Y , S. P. and G I L -
M O UR , I. 2003. The Cretaceous world. Cambridge University
Press, Cambridge. 360 pp.
S M I T H , A. G. and R US H , L. A. 1997. Time trek. Cambridge
Paleo-Map Services. http://www.the-conference.com/CPSL.
—— S M I T H, D. G. and FU N N E L L , B. M. 1994. Atlas of
Mesozoic and Cenozoic coastlines. Cambridge University Press,
Cambridge, 99 pp.
1468 P A L A E O N T O L O G Y , V O L U M E 5 1

S W E E T M A N , S. C. 2006. A gobiconodontid (Mammalia, Eu-
triconodonta) from the Early Cretaceous (Barremian) Wessex
Formation of the Isle of Wight, southern England. Palaeontol-
ogy, 49, 889–897.
T A K A DA , T., M A T S UO KA , H. and S E T OG U CH I , T.
2001. The first multituberculate from Japan. 55–58. In
DE N G , T. and W A N G, Y. (eds). Proceedings of the Eight
Annual Meeting of the Chinese Society of Vertebrate Paleontol-
ogy, China Ocean Press, Beijing.
W A N G, Y., H U , Y., Z HO U , M. and L I , C. 1995. Mesozoic
mammal localities in western Liaoning, northeast China. 221–
227. In S U N , A. and W A N G, Y. (eds). Sixth Symposium on
Mesozoic Terrestrial Ecosystems and Biota, Short Papers, China
Ocean Press, Beijing.
W O O DW A R D , A. S. 1911. One some mammalian teeth of the
Wealden of Hastings. Quaterly Journal of the Geological Society,
London, 67, 278–281.
B A D I O L A E T A L . : N E W M U L T I T U B E R C U L A T E M A M M A L S F R O M T H E H A U T E R I V I A N ⁄ B A R R E M I A N T R A N S I T I O N 1469
Related Documents











