ARTICLE NEW DATA ON THE MOSCHIDAE (MAMMALIA, RUMINANTIA) FROM THE UPPER MIOCENE OF SPAIN (MN 10- MN 11) ISRAEL M. SA ´ NCHEZ, *,1,2 M. SOLEDAD DOMINGO, 2 and JORGE MORALES 2 1 Department of Zoology, Museum of Zoology, University of Cambridge. Downing Street, Cambridge, CB2 3EJ, United Kingdom, [email protected]; 2 Departamento de Paleobiologı´a. Museo Nacional de Ciencias Naturales-CSIC. C/ Jose ´ Gutie ´ rrez Abascal, 2. 28006 Madrid, Spain ABSTRACT—New data on the last representatives of the Spanish Miocene Moschidae (Mammalia, Ruminantia) are presented and discussed. A new species of Micromeryx, Micromeryx soriae, is described on the basis of fossil material from the locality of La Roma-2 (upper Vallesian; MN 10, Teruel province, Spain, previously assigned to Micromeryx sp.), Batallones-1, and Batallones-10 (upper Vallesian; local zone J, MN 10, Madrid province, Spain). The new species is characterized by possessing relatively hypsodont lower molars of advanced morphology and a unique type of Palaeo- meryx-fold. M. soriae gives insight to the last representatives of the genus, which achieved an overall lower molar morphology that mimics to certain degree that of Hispanomeryx. several characters of the upper molars of the Miocene moschids Micromeryx and Hispanomeryx are described that are useful for characterizing both genera. These characters are used to solve the taxonomic problems of PM-659, an upper molar from Puente Minero (MN 11, Teruel province, Spain) that represents one of the last survivors of the Moschidae in the Iberian Peninsula. The systematic utility of the upper molars of Hispanomeryx and Micromeryx is demonstrated, and the use of size as the only way to distinguish between the two genera is refuted. Finally, the study of the morphological characters of PM-659 clearly supports its generic change from Micromeryx sp. to Hispanomeryx sp., thus recording the presence of Hispanomeryx in the Iberian Peninsula as late as the lower Turolian. INTRODUCTION The Moschidae comprise small hornless pecorans whose only extant representatives are the Asian musk deer, Moschus spp. (Nowak, 1999). Moschids, represented by the genera Hispano- meryx and Micromeryx, ranged in Spain from MN 5 to MN 11 (Sa ´ nchez and Morales, 2006), although there are purported moschid remains dated as late as MN 12 (Daams et al., 1998; Agustı´ et al., 2001). They were very common in the Iberian Peninsula and continental Europe during the middle Miocene, but started to decline relatively quickly after the end of the Vallesian (Sa ´ nchez, 2006; Sa ´ nchez and Morales, 2006). Due to the rarity of their remains, upper Vallesian-lower Turolian Span- ish moschids are less well-known than those of the upper Arago- nian-lower Vallesian (Sa ´nchez and Morales, 2006). The main descriptive work regarding these younger forms is that of Alcala ´ (1994) on the moschid remains from La Roma-2 (upper Valle- sian, MN 10) and Puente Minero (lower Turolian, MN 11) where they are assigned to Micromeryx sp. The present paper aims to update the current state of knowl- edge of Iberian upper Miocene moschids through systematic revision of the more noteworthy fossils from the upper Valle- sian-lower Turolian. First, we study the isolated upper molar of Puente Minero (PM-659) and describe several anatomical char- acters of the upper molars relevant to the taxonomic status of PM-659 that can be used to differentiate the Miocene moschids Micromeryx and Hispanomeryx. Since the description of Hispa- nomeryx in the Vallesian of Spain (Morales et al., 1981), very few attempts have been made to identify the morphological differences between Hispanomeryx and Micromeryx, which sha- re many features. Studies so far have focused mainly on the lower dentition (Morales and Soria, 1981; Azanza, 1986; Vislo- bokova, 2007) or the morphology of the metatarsal sulcus (Alcala ´ , 1994), but the upper molars have never been discussed. The Spanish fossil record of the Miocene Moschidae has been significantly enriched since the original description of Hispano- meryx duriensis (Morales et al., 1981; Azanza, 1986; Azanza et al., 1999, 2004; Sa ´ nchez, 2006; Sa ´ nchez and Morales, 2006; Sa ´ nchez and Morales, 2008). An abundant collection of non- equivocal fossils of Hispanomeryx (Morales et al., 1981; Azanza, 1986; Sa ´ nchez, 2006; Sa ´ nchez and Morales, 2006) allows for the identification of several patterns of anatomical similarities and dissimilarities between Hispanomeryx and Micromeryx. We use the key characters of the upper molars in order to reassess the taxonomical assignment of PM-659 from Micromeryx sp. to His- panomeryx sp., and discuss the usefulness of body size and mor- phology for the systematics of Hispanomeryx and Micromeryx. Finally, we provide here a formal description of Micromeryx soriae sp. nov., from La Roma-2, Batallones-1 and Batallones-10 (upper Vallesian). The specific diversity of Micromeryx has been re-evaluated both in Spain and continental Europe (Sa ´ nchez, 2006; Vislobokova, 2007; Sa ´ nchez and Morales, 2008). Until very recently, only two species of Micromeryx had been described— the type species M. flourensianus Lartet, 1851 from the site of Sansan (France, MN 6), and M. styriacus Thenius, 1950, from the site of Go ¨ riach (Austria, MN 6)—but lately two new species have been erected: M. mirus Vislobokova, 2007 (Kohfidisch, lower Turolian, MN 11, Austria) and M. azanzae Sa ´ nchez and Morales, 2008 (Toril-3, upper Aragonian, MN 7/8, local zone G3, Spain). M. soriae is the more common of the last represen- tatives of the genus in the Iberian Peninsula during the upper Vallesian (see Sa ´ nchez and Morales, 2006). The original fossils of Micromeryx sp. from La Roma-2 (Alcala ´ , 1994) constitute * Corresponding author. Journal of Vertebrate Paleontology 29(2):567–575, June 2009 # 2009 by the Society of Vertebrate Paleontology 567

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE
NEW DATA ON THE MOSCHIDAE (MAMMALIA, RUMINANTIA) FROMTHE UPPER MIOCENE OF SPAIN (MN 10- MN 11)
ISRAEL M. SANCHEZ,*,1,2 M. SOLEDAD DOMINGO,2 and JORGE MORALES21Department of Zoology, Museum of Zoology, University of Cambridge. Downing Street, Cambridge, CB2 3EJ,
United Kingdom, [email protected];2Departamento de Paleobiologıa. Museo Nacional de Ciencias Naturales-CSIC.
C/ Jose Gutierrez Abascal, 2. 28006 Madrid, Spain
ABSTRACT—New data on the last representatives of the Spanish Miocene Moschidae (Mammalia, Ruminantia) arepresented and discussed. A new species of Micromeryx, Micromeryx soriae, is described on the basis of fossil materialfrom the locality of La Roma-2 (upper Vallesian; MN 10, Teruel province, Spain, previously assigned to Micromeryx sp.),Batallones-1, and Batallones-10 (upper Vallesian; local zone J, MN 10, Madrid province, Spain). The new species ischaracterized by possessing relatively hypsodont lower molars of advanced morphology and a unique type of Palaeo-meryx-fold. M. soriae gives insight to the last representatives of the genus, which achieved an overall lower molarmorphology that mimics to certain degree that of Hispanomeryx. several characters of the upper molars of the Miocenemoschids Micromeryx and Hispanomeryx are described that are useful for characterizing both genera. These charactersare used to solve the taxonomic problems of PM-659, an upper molar from Puente Minero (MN 11, Teruel province,Spain) that represents one of the last survivors of the Moschidae in the Iberian Peninsula. The systematic utility of theupper molars of Hispanomeryx and Micromeryx is demonstrated, and the use of size as the only way to distinguishbetween the two genera is refuted. Finally, the study of the morphological characters of PM-659 clearly supports itsgeneric change from Micromeryx sp. to Hispanomeryx sp., thus recording the presence of Hispanomeryx in the IberianPeninsula as late as the lower Turolian.
INTRODUCTION
The Moschidae comprise small hornless pecorans whose onlyextant representatives are the Asian musk deer, Moschus spp.(Nowak, 1999). Moschids, represented by the genera Hispano-meryx and Micromeryx, ranged in Spain from MN 5 to MN 11(Sanchez and Morales, 2006), although there are purportedmoschid remains dated as late as MN 12 (Daams et al., 1998;Agustı et al., 2001). They were very common in the IberianPeninsula and continental Europe during the middle Miocene,but started to decline relatively quickly after the end of theVallesian (Sanchez, 2006; Sanchez and Morales, 2006). Due tothe rarity of their remains, upper Vallesian-lower Turolian Span-ish moschids are less well-known than those of the upper Arago-nian-lower Vallesian (Sanchez and Morales, 2006). The maindescriptive work regarding these younger forms is that of Alcala(1994) on the moschid remains from La Roma-2 (upper Valle-sian, MN 10) and Puente Minero (lower Turolian, MN 11) wherethey are assigned to Micromeryx sp.The present paper aims to update the current state of knowl-
edge of Iberian upper Miocene moschids through systematicrevision of the more noteworthy fossils from the upper Valle-sian-lower Turolian. First, we study the isolated upper molar ofPuente Minero (PM-659) and describe several anatomical char-acters of the upper molars relevant to the taxonomic status ofPM-659 that can be used to differentiate the Miocene moschidsMicromeryx and Hispanomeryx. Since the description of Hispa-nomeryx in the Vallesian of Spain (Morales et al., 1981), veryfew attempts have been made to identify the morphologicaldifferences between Hispanomeryx and Micromeryx, which sha-
re many features. Studies so far have focused mainly on thelower dentition (Morales and Soria, 1981; Azanza, 1986; Vislo-bokova, 2007) or the morphology of the metatarsal sulcus(Alcala, 1994), but the upper molars have never been discussed.The Spanish fossil record of the Miocene Moschidae has beensignificantly enriched since the original description of Hispano-meryx duriensis (Morales et al., 1981; Azanza, 1986; Azanzaet al., 1999, 2004; Sanchez, 2006; Sanchez and Morales, 2006;Sanchez and Morales, 2008). An abundant collection of non-equivocal fossils of Hispanomeryx (Morales et al., 1981; Azanza,1986; Sanchez, 2006; Sanchez and Morales, 2006) allows for theidentification of several patterns of anatomical similarities anddissimilarities between Hispanomeryx and Micromeryx. We usethe key characters of the upper molars in order to reassess thetaxonomical assignment of PM-659 from Micromeryx sp. to His-panomeryx sp., and discuss the usefulness of body size and mor-phology for the systematics of Hispanomeryx and Micromeryx.Finally, we provide here a formal description of Micromeryx
soriae sp. nov., from La Roma-2, Batallones-1 and Batallones-10(upper Vallesian). The specific diversity ofMicromeryx has beenre-evaluated both in Spain and continental Europe (Sanchez,2006; Vislobokova, 2007; Sanchez and Morales, 2008). Until veryrecently, only two species of Micromeryx had been described—the type species M. flourensianus Lartet, 1851 from the site ofSansan (France, MN 6), andM. styriacus Thenius, 1950, from thesite of Goriach (Austria, MN 6)—but lately two new specieshave been erected: M. mirus Vislobokova, 2007 (Kohfidisch,lower Turolian, MN 11, Austria) and M. azanzae Sanchez andMorales, 2008 (Toril-3, upper Aragonian, MN 7/8, local zoneG3, Spain). M. soriae is the more common of the last represen-tatives of the genus in the Iberian Peninsula during the upperVallesian (see Sanchez and Morales, 2006). The original fossilsof Micromeryx sp. from La Roma-2 (Alcala, 1994) constitute*Corresponding author.
Journal of Vertebrate Paleontology 29(2):567–575, June 2009# 2009 by the Society of Vertebrate Paleontology
567

the type series of the new species, whereas the fossil remainsof M. soriae from Cerro de los Batallones are described for thefirst time, providing additional insights into the anatomy of thespecies.
LOCALITIES AND GEOLOGICAL SETTING
La Roma-2
The type locality of Micromeryx soriae sp. nov. is located inthe Alfambra-Teruel Basin (Fig. 1) between the villages ofAlfambra and Peralejos (Teruel province, Spain). Both thefossil mammal assemblage and the stratigraphical section of LaRoma-2 are fully detailed in Alcala (1994) and van Dam (1997).A late Vallesian age has been estimated for La Roma-2 (Fig. 2;MN 10; local zone J3; 8.789 � 0.019 Ma; see Alcala, 1994;van Dam, 1997; Pesquero et al., 2006; and Domingo et al.,2007a for further details). The assemblage includes carnivorans(hyaenids), artiodactyls (suids, moschids and bovids), and peri-ssodactyls (rhinocerotids and equids) in addition to severalfamilies of micromammals (Alcala, 1994; Pesquero et al., 2006).La Roma-2 is also the type locality of the bovid Aragoral mude-jar Alcala and Morales, 1997, the rhinocerotid Alicornops alfam-brense Cerdeno and Alcala, 1989 and the equid Hipparionlaromae Pesquero, Alberdi and Alcala, 2006. With regard to thepaleoenvironment of La Roma-2, Alcala (1994) infers an openand dry savanna-like habitat.
Cerro de los Batallones
Batallones-1—The locality of Batallones-1 is currently themost important of the fossil site-complex of Cerro de los Batal-lones (Batallones Butte), an array of nine localities discovered in1991 and subsequently excavated. Cerro de los Batallones islocated in the Madrid Basin (Fig. 1) near the town of Torrejonde Velasco (southern Madrid province). The fossil associationfound at Batallones indicates a late Vallesian age (Fig. 2; MN 10;local zone J; see Morales et al., 1992, 2000, 2004; Anton et al.,2004; Peigne et al., 2005; Salesa et al., 2005, 2006a, b); the abso-lute age of Batallones-1 has been estimated to be 8.976 � 0.018Ma (Domingo et al., 2007a). Batallones-1 has been interpreted
as a carnivore-trap site because carnivoran remains far outnum-ber those of other macromammalian taxa (Morales et al., 2004).The assemblage includes non-mammals such as freshwater deca-pod crustaceans, teleosteans, anuran lissamphibians, chelonians,squamates, and aves. The mammals include carnivorans (amphi-cyonids, machairodont and feline felids, viverrids, mustelids,ailurids, and ursids), artiodactyls (suids, bovids, moschids, andgiraffids), perissodactyls (hipparionine equids and rhinocero-tids), proboscideans, insectivorans, lagomorphs, and several ro-dent families (Morales, 2004; Morales et al., 2004; Domingoet al., 2007b).The fossiliferous sediments in the butte fill cavities that prob-
ably acted as natural faunal traps; it has been suggested thatthese cavities are piping-like structures where water and car-casses of herbivores attracted carnivores to feed and drink,resulting in their trapping and death (Morales et al., 2004; Pozoet al., 2004).
FIGURE 1. Map illustrating Madrid and Alfambra-Teruel basinsamong Miocene Spanish continental basins. Abbreviations: ATB,Alfambra-Teruel basin; MB, Madrid basin.
FIGURE 2. Chronostratigraphic position of La Roma-2, the sites ofCerro de los Batallones, and Puente Minero (modified from Sanchezand Morales, 2006). Abbreviations: BAT, fossil sites of Cerro de losBatallones; LR2, La Roma-2; PM, Puente Minero.
568 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 29, NO. 2, 2009

The inferred environment for Batallones is a savanna withforested patches that offered refuge for taxa like Paramachair-odus and Simocyon (Anton and Morales, 2000; Salesa, 2002;Peigne et al., 2005; Salesa et al., 2006a, b).Batallones-10—Batallones-10 is the last fossil locality discov-
ered in Cerro de los Batallones, found in 2007. The fossil associ-ation of Batallones-10 is astonishingly rich and totally differentfrom that of Batallones-1; apart from very large chelonians, it iscomposed principally of large mammalian herbivores, mainlygiraffids and hipparionine equids (from colts to fully grown spe-cimens), as well as proboscideans, rhinocerotids, medium-sizedbovids, suids, and moschids, including M. soriae. Carnivoranremains are very scarce.
Puente Minero
The fossil site of Puente Minero is located in the Alfambra-Teruel Basin (Fig. 1), Northeast of Teruel capital town (Alcala,1994). Its age has been estimated to be lower Turolian (Fig. 2;Alcala, 1994), with an absolute age of 7.834 � 0.021 Ma (Dom-ingo et al., 2007a). The rich mammalian fossil fauna of PuenteMinero includes insectivores, bats, rodents, lagomorphs, carni-vorans, artiodactyls, perissodactyls, and proboscideans (Alcala,1994). The paleoenvironment of Puente Minero has been de-scribed as an open area with more humid conditions than in theupper Vallesian sites (Alcala, 1994).
MATERIALS AND METHODS
Measurements—All measurements were taken with digitalcalipers. The measurements set is detailed in Sanchez and Mor-ales (2008). The metric data are detailed in SupplementaryData (Tables 1S–4S; www.vertpaleo.org/publications/JVPContent.cfm).Nomenclature—We follow the terminology of Barone (1999)
for anatomic nomenclature of the cranial and postcranial skele-ton, and Azanza (2000) for the nomenclature of the dentition.Institutional and Fossil Localities Abbreviations—BAT,
Batallones (Upper Vallesian, Madrid, Spain); MNCN-CSIC,Museo Nacional de Ciencias Naturales-CSIC (Madrid, Spain);PM, Puente Minero; RO, La Roma-2 (Upper Vallesian, Teruel,Spain). The fossils from La Roma-2 are curated by the Funda-cion Conjunto Paleontologico de Teruel-Dinopolis (Teruel,Spain), but are currently stored in the MNCN-CSIC (Madrid,Spain). The specimens from Cerro de los Batallones are curatedby the MNCN-CSIC (Madrid, Spain).
SYSTEMATIC PALEONTOLOGY
MAMMALIA Linnaeus, 1758ARTIODACTYLA Owen, 1848RUMINANTIA Scopoli, 1777
PECORA Linnaeus, 1758 sensu Webb and Taylor, 1980MOSCHIDAE Gray, 1821
HISPANOMERYX Morales, Moya-Sola and Soria, 1981
Type Species—Hispanomeryx duriensis Morales, Moya-Solaand Soria, 1981
HISPANOMERYX sp.(Fig. 3)
Micromeryx sp.: Alcala, 1994:260-261.Hispanomeryx sp.: Sanchez and Morales, 2006:540-541.
Locality—Puente Minero, MN 11, local zone K, Teruel prov-ince, Spain.Referred Material—PM-659, upper molar (undetermined, but
probably a M1).
Note—We follow the original assignation of Hispanomeryx tothe Moschidae given by Morales et al. (1981) in the descriptionof the genus. PM-659 is curated by the Conjunto Paleontologicode Teruel-Dinopolis (Teruel, Spain), but is currently stored inthe MNCN-CSIC (Madrid, Spain).Description—PM-659 lacks almost all the posterior lobe, so it
is difficult to assign it to any of the three upper molars; never-theless, both the morphology of the paracone—with the buccalwall slightly misaligned with the metacone (as occurs in Hispa-nomeryx; pers. obs.)—and the overall morphology of the trigonsuggest it represents M1. PM-659 sports a relative high crown, aspreviously noted by Alcala (1994). The parastyle, the mesostyle,and the protoconal rib are moderately developed, narrow andcolumnar (not pyramidal). The metastyle is poorly developed.There is no protoconal fold, nor cingula. All cusps are united bytheir cristas, except the post-protocrista, which does not reachthe buccal wall.Comparisons and Discussion—In his original description,
Alcala (1994) discussed the anatomical aspects of PM-659, butfinally assigned it to the genus Micromeryx on the basis of itssmaller size compared to Hispanomeryx (Alcala, 1994). The up-per dentition of H. aragonensis was unknown when PM-659 wasfirst described leaving the much larger H. duriensis as the onlyknown Hispanomeryx to which it could be compared. Thus, thetaxonomical assignment of Alcala (1994) was not unrealistic
FIGURE 3. Hispanomeryx sp. A, upper molar PM-659 in labial view;B, upper molar PM-659 in occlusal view; C, drawing of PM-659 in labialview; D, Hispanomeryx duriensis, EL-SS, M1 from El Lugarejo (Spain,type locality; Morales et al., 1981); E,Micromeryx flourensianus, Sa 9772,M1 from Sansan (type locality of Micromeryx flourensianus); F, Micro-meryx azanzae, TOR3B-24, M1 from Toril-3 (Spain). Notice the mor-phology of the molar structures in Hispanomeryx and Micromeryx: thebuccal ribs and styles are narrow and columnar in the former (C-D) andpyramidal with broad bases in the latter (E-F). Also, Hispanomeryxshows much more developed cristas and flatter cusps. PM-659 (A-C)clearly shows a Hispanomeryx-like morphology. Abbreviations: Mesost,mesostyle; ParR, paraconal rib; Parst, parastyle. Scale bar applies toA-B; line art drawings not to scale.
SANCHEZ ET AL.—SPANISH UPPER MIOCENE MOSCHIDAE 569

size-wise, since all then-known upper molars of Micromeryxwere far smaller than those of Hispanomeryx. Nevertheless, thenew data regarding the fossil record of the Spanish MioceneMoschidae demonstrates that the larger molars of the medium-sized species of Micromeryx, such as M. azanzae, were nearly aslarge as those of the smaller Hispanomeryx (Table 4S; Sanchezand Morales, 2008). This means that an apparently small isolatedindeterminate molar like PM-659 should not be assigned toMicromeryx only on the basis of size. An additional problemregarding the use of size as the only taxonomical criterion isthe a priori exclusion of the existence of a small species ofHispanomeryx, which would be misidentified if its morphologi-cal features were ignored. Morphology is therefore the onlyway to discriminate between these two genera. Hispanomeryxpossesses thin and columnar parastyles, mesostyles, and proto-conal ribs, whereas Micromeryx has wider and more developedpyramidal styles and ribs with broader bases and pointed tips(Fig. 4). The internal walls of the main cusps are flatter inHispanomeryx than they are in Micromeryx. Also, the cristas ofHispanomeryx merge with the main cusps, and between them,higher in the molar crown than those ofMicromeryx. Finally, theunion between the parastyle and mesostyle with the base ofthe protoconal rib gives it a triangular shape in Micromeryxwhereas it forms a rectangular (or near-rectangular) shape inHispanomeryx, due to the thinner buccal structures of thelatter (Fig. 4). As observed in Figure 4A–C, PM-659 shows allcharacters of Hispanomeryx, clearly distinct from Micromeryx;thus, despite size discussions, the generic reassignment of PM-659 from Micromeryx to Hispanomeryx is strongly supported bymorphology.
Although it is necessary to check the true taxonomic status ofthe purported middle Turolian Spanish ‘Micromeryx’ (MN 12,local zone L; Agustı et al,. 2001; Vislobokova, 2007), the recordof Hispanomeryx in the early Turolian of Spain is presently theyoungest of the genus (Sanchez and Morales, 2006), previouslyestablished in the MN 10 (Daams et al., 1998; Agustı et al.,2001).
MICROMERYX Lartet, 1851
Type Species—Micromeryx flourensianus Lartet, 1851
MICROMERYX SORIAE Sp. nov.(Figs. 5–7)
Micromeryx sp.: Alcala, 1994: 260-261 (specimens from LaRoma-2).Hispanomeryx duriensis: Morales et al., 2000: 186 (specimensfrom Batallones-1).Hispanomeryx duriensis: Morales et al., 2004: 140, table I (speci-mens from Batallones-1).Hispanomeryx sp.: Sanchez and Morales, 2004: 522, fig. 6 (speci-mens from Batallones-1).Micromeryx sp.: Pesquero et al., 2006: 343 (specimens from LaRoma-2).
Holotype—RO-9M, right hemimandible with m1-m2.Etymology—Dedicated to the memory of Dr. Marıa Dolores
Soria.Paratypes—RO-2S, right p3; RO-1S, right dp4.Type Locality and Horizon—La Roma-2, late Vallesian, MN
10, local zone J, Alfambra-Teruel Basin, province of Teruel,Spain.Referred Material—RO-9M, right hemimandible with m1–2;,
RO-2S, right p3; RO-1S, right dp4; BAT1’02.E4-12, right hemi-mandibular fragment with dp4 and m1; B-SS, right distal frag-ment of humerus; BAT1’03.D6-218, right unciform; BAT1’03.D6-262, right magnotrapezoid; B-4843a, left distal fragment oftibia; B-4843b, left malleolar bone; B-4843c, left astragalus (allthree B-4843 specimens are articulated); BAT1’04.F5-30, rightdistal fragment of tibia; B-2260, left astragalus; B-456, right as-tragalus; B-5460, left astragalus; BAT1’03.E6-145, first phalanx;
FIGURE 4. Comparative bucco-occlusal view (line drawing andcorresponding real specimen) of the posterior wall of the protoconid inseveral moschid species, with special remarks on the morphology of thePalaeomeryx-fold. A, m1 of Micromeryx sp. (O’Donnell, MN 5, Madrid)from the specimen OD-6, showing the typical Palaeomeryx-fold, narrowand with well-marked profile. B, m1 of Hispanomeryx duriensis fromspecimen EL-8 (El Lugarejo, MN 9, Avila), showing the flat posteriorwall with absent Palaeomeryx-fold (for ease of comparison, this illustra-tion has been mirrored with respect to the original). C, m2 from Micro-meryx soriae holotype (RO-9M), showing the morphology of thePalaeomeryx-fold present in this species; compare the narrow fold ofA with the low-profiled and elongated structure present in the molars ofMicromeryx soriae. Abbreviations: PalF, Palaeomeryx-fold; Protoc, pro-toconid; PwProtoc, posterior wall of the protoconid.
FIGURE 5. Micromeryx soriae, type series. A, holotype RO-9M inocclusal (above), lingual (middle), and buccal (below) views; B, paratypeRO-1S in occlusal (above), lingual (middle), and buccal (below) views;C, paratype RO-2S in occlusal (above), lingual (middle), and buccal(below) views.
570 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 29, NO. 2, 2009
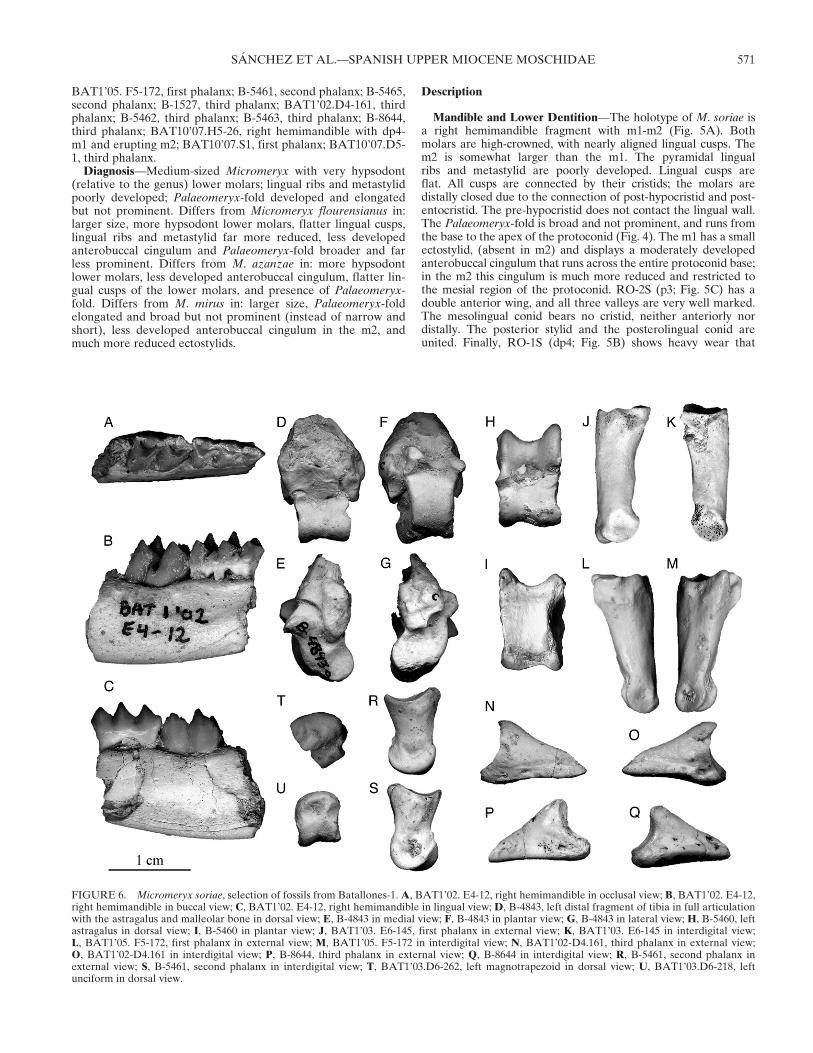
BAT1’05. F5-172, first phalanx; B-5461, second phalanx; B-5465,second phalanx; B-1527, third phalanx; BAT1’02.D4-161, thirdphalanx; B-5462, third phalanx; B-5463, third phalanx; B-8644,third phalanx; BAT10’07.H5-26, right hemimandible with dp4-m1 and erupting m2; BAT10’07.S1, first phalanx; BAT10’07.D5-1, third phalanx.Diagnosis—Medium-sized Micromeryx with very hypsodont
(relative to the genus) lower molars; lingual ribs and metastylidpoorly developed; Palaeomeryx-fold developed and elongatedbut not prominent. Differs from Micromeryx flourensianus in:larger size, more hypsodont lower molars, flatter lingual cusps,lingual ribs and metastylid far more reduced, less developedanterobuccal cingulum and Palaeomeryx-fold broader and farless prominent. Differs from M. azanzae in: more hypsodontlower molars, less developed anterobuccal cingulum, flatter lin-gual cusps of the lower molars, and presence of Palaeomeryx-fold. Differs from M. mirus in: larger size, Palaeomeryx-foldelongated and broad but not prominent (instead of narrow andshort), less developed anterobuccal cingulum in the m2, andmuch more reduced ectostylids.
Description
Mandible and Lower Dentition—The holotype of M. soriae isa right hemimandible fragment with m1-m2 (Fig. 5A). Bothmolars are high-crowned, with nearly aligned lingual cusps. Them2 is somewhat larger than the m1. The pyramidal lingualribs and metastylid are poorly developed. Lingual cusps areflat. All cusps are connected by their cristids; the molars aredistally closed due to the connection of post-hypocristid and post-entocristid. The pre-hypocristid does not contact the lingual wall.The Palaeomeryx-fold is broad and not prominent, and runs fromthe base to the apex of the protoconid (Fig. 4). The m1 has a smallectostylid, (absent in m2) and displays a moderately developedanterobuccal cingulum that runs across the entire protoconid base;in the m2 this cingulum is much more reduced and restricted tothe mesial region of the protoconid. RO-2S (p3; Fig. 5C) has adouble anterior wing, and all three valleys are very well marked.The mesolingual conid bears no cristid, neither anteriorly nordistally. The posterior stylid and the posterolingual conid areunited. Finally, RO-1S (dp4; Fig. 5B) shows heavy wear that
FIGURE 6. Micromeryx soriae, selection of fossils from Batallones-1.A, BAT1’02. E4-12, right hemimandible in occlusal view; B, BAT1’02. E4-12,right hemimandible in buccal view; C, BAT1’02. E4-12, right hemimandible in lingual view; D, B-4843, left distal fragment of tibia in full articulationwith the astragalus and malleolar bone in dorsal view; E, B-4843 in medial view; F, B-4843 in plantar view; G, B-4843 in lateral view; H, B-5460, leftastragalus in dorsal view; I, B-5460 in plantar view; J, BAT1’03. E6-145, first phalanx in external view; K, BAT1’03. E6-145 in interdigital view;L, BAT1’05. F5-172, first phalanx in external view; M, BAT1’05. F5-172 in interdigital view; N, BAT1’02-D4.161, third phalanx in external view;O, BAT1’02-D4.161 in interdigital view; P, B-8644, third phalanx in external view; Q, B-8644 in interdigital view; R, B-5461, second phalanx inexternal view; S, B-5461, second phalanx in interdigital view; T, BAT1’03.D6-262, left magnotrapezoid in dorsal view; U, BAT1’03.D6-218, leftunciform in dorsal view.
SANCHEZ ET AL.—SPANISH UPPER MIOCENE MOSCHIDAE 571

obscures any important occlusal features. The lingual wall is quiteflat and the posterior buccal stylid is very small.
BAT1’02.E4-12 (Fig. 6A–C) is a juvenile hemimandibularfragment bearing dp4-m1 and the corresponding section of thehorizontal mandibular ramus. All cusps of the dp4 are united bytheir cristids, the lingual cusps are aligned, and the anteriorcingulid is poorly developed. The m1 is almost identical to thatof RO-9M, has nearly aligned lingual cusps, and the lingual ribsand stylids are weak. The tooth is as hypsodont as in the holo-type RO-9M, and all cusps are united with the exception of thepre-hypocristid, which does not contact the post-protocristid.
The Palaeomeryx-fold shows the characteristic morphology ob-served in RO-9M. The specimen BAT10’07.H5-26 from Batal-lones-10 (Fig. 7A) preserves the major part of the mandibularsymphysis. The presence of m2 in BAT10’07.H5-26 confirmsthat the m2 is slightly larger relative to the m1 than usual inMicromeryx, as observed in the holotype.Humerus—The only specimen is not well preserved. The me-
dial condyle of the distal trochlea is well developed and extendsproximally. The trochlea shows a slightly inclined proximo-distalaxis. The capitulum is well developed, and sports the near-straight proximal border typical of Micromeryx.Unciform—The facet for the pyramidal is triangular, and is
broader than the facet for the semilunar. The proximal facet forthe magnotrapezoid extends along the proximal border of thebone and presents two articular areas; the anterior articular areais flat and triangular, and the posterior one is elliptic and slightlyconcave. The dorsal facet for the magnotrapezoid is small anddoes not contact the proximal facet for the magnotrapezoid; thepalmar facet for the magnotrapezoid is flat and elliptic. Thefacet for the metacarpal III–IV is triangular and flat, and exhi-bits a short, rounded, non-articular groove.Magnotrapezoid—The articular facet for the semilunate is
smaller than the facet for the scaphoid. The proximodorsal facetfor the unciform is flat and subtriangular and does not contactthe distal border of the bone. The two palmar facets for theunciform are flat and rounded, and do not contact each other.The facet for the metacarpal III–IV is triangular and concave,with an elongated and narrow lateral non-articular groove. It isprobable that specimens BAT1’03.D6-262 (Fig. 6T) andBAT1’03.D6-218 (Fig. 6U) belonged to the same individual,given the special characteristics of Batallones-1, where it is nor-mal to find several skeletal portions of the same individual closeone to another. They match in size, articulate perfectly, and werefound very close to each other.Tibia—The tibial fragment B-4843b belongs to one of the
best-preserved moschid fossils from Batallones-1 (Fig. 7D–G).It is in full articulation with the astragalus B-4843a and the mal-leolar B-4843c. The lateral border of the dorsodistal tibial pro-cess shows the well-marked step characteristic of Micromeryx.The tibia B-4843b is broken above the distal articulation andshows evidence of digestion by carnivore mammals (categories4 and 5 of Andrews, 1990) in the form of glossy and roundededges (Fig. 8; Domingo et al., 2007b). On the other hand, speci-men BAT-1’04.F5-30 shows a strong diagenetic flattening.
FIGURE 7. Micromeryx soriae from Batallones-10. A, BAT10’07.H5-26, right hemimandible in lingual (above) and buccal (below) views; B,BAT10’07.S1, first phalanx in external (left) and interdigital (right)views; C, BAT10’07.D5-1, third phalanx in interdigital (above) and ex-ternal (below) views.
FIGURE 8. Micromeryx soriae sp. nov. Evidence of digestion by mammalian carnivores (following the categories of Andrews, 1990) in theBatallones-1 sample. A, SEM (Scanning Electron Microscope) photograph of specimen B-4843 in lateral view showing the glossy and rounded aspectof broken ends in digested bones. B, SEM photograph of specimen B-2260 in plantar view showing intrusive digestion with tissue loss. C, detail SEMphotograph of specimen B-456 in plantar view showing microscopic splitting and cracking of the bone surface. Scale bar applies to A-B.
572 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 29, NO. 2, 2009

Malleolar—In B-4843c (Fig. 6D–G) the distal surface of thebone is divided into two facets separated by a bony ridge, where-as in Moschus there is only one broad, elliptic and concave distalfacet; both facets are concave and triangular, the dorsal onebeing shorter than the plantar. B-4843b is fully articulated withthe tibia and consequently the proximal articulation remainshidden.Astragalus—The specimens B-456 and B-2260 are small, elon-
gate, and slender, and probably belonged to juveniles. Bothshow clear evidence of digestion by mammalian carnivores, withintrusive digestion that results in heavy tissue loss and micro-scopic splitting and cracking of the bone surface (Fig. 8;Andrews, 1990; Domingo et al., 2007b). Specimens B-4843a andB-5460 (Fig. 6H, I) are complete and well preserved. The lateralcondyle of the proximal trochlea is higher than the medial con-dyle. There is a triangular notch in the proximal part of thedorsolateral border of the distal trochlea. The plantar trochleadoes not contact any of the distal articular surfaces.First Phalanx—The proximal articular sulcus is deep and
broad, and extends to the dorsal border of the phalanx. Theexternal proximal facet is narrow dorsally. The furrow for theinterosseus muscle is elongated and well marked. The dorsalsurface of the diaphysis is flat, but this feature is not as pro-nounced as in typical B-type phalanges, which show perfectlystraight dorsal borders (Kohler, 1993). The insertion area of theinterdigital ligament is well marked. The external face of thediaphysis is almost straight in dorsal view. The proximal borderof the distal articulation surface is visible in dorsal view. Thereare no differences between the Batallones-1 (Fig. 6J–M) andBatallones-10 (Fig. 7B) samples.Second Phalanx—The only specimen (Fig. 6N–Q) shows a
well-developed origin area of the crossed ligaments. The exten-sor process is short and pointed. The insertion area for the su-perficial flexor tendon is well marked. The distal facet extendsdorsally in typical B-type fashion (Kohler, 1993), but does notshow any plantar extension.Third Phalanx—M. soriae has the low-profile and elongate
third phalanges typical of the genus. The dorsal border is longand straight, with a slight inclination near the distal end. Theextensor process is not well developed. There is no postarticularplateau. The articular facet for the second phalanx occupiesalmost all of the proximal surface. The process for the insertionof the deep digital flexor tendon is well developed. The interdi-gital border of the plantar surface is straight. The third phalan-ges from Batallones-1 (Fig. 6R-S) and Batallones-10 (Fig. 7C)are virtually identical.
Discussion
As already pointed out by Alcala (1994), M. soriae is com-parable to Hispanomeryx in terms of relative crown height ofthe lower molars. However, its morphological characters differ:Hispanomeryx has even more poorly developed stylids thanM. soriae, the cristids are straighter and taller, the cusps areflatter, and the lingual ribs are columnar instead of having apyramidal base. Also, the difference in size between m1 and m2of M. soriae is not as developed as in Hispanomeryx. Alcala(1994) also notes the derived morphology of the lower molars—pointing out their great resemblance to the molars of bovids—butdescribes them as lacking a Palaeomeryx-fold. The lower molarsof M. soriae do not show the typical Palaeomeryx-fold (i.e.,narrow, short, and well developed), but have instead a broadand low relief that runs along the entire height of the protoconid.This morphology is unique within the genusMicromeryx (Fig. 4).M. azanzae is the only described species of Micromeryx that hastotally lost the Palaeomeryx-fold, and the morphology of thedistal wall of the protoconid differs from that of M. soriae. Thesame occurs with Hispanomeryx spp., the molars of which have
no Palaeomeryx-fold. In both cases, the distal wall of the proto-conid is entirely flat, indicating that the structure inM. soriae canonly be referred to as a broad and weak Palaeomeryx-fold thatretains its longitudinal extension (Fig. 4). Hence, we do not agreewith the original description of Alcala (1994). The loss of thePalaeomeryx-fold is generally thought to be correlated with thedevelopment of higher-crowned lower molars within a lineage(Moya-Sola, 1986), but Micromeryx does not confirm thathypothesis because the more hypsodont taxa as M. soriae andM. mirus still retain this structure whereas it has been lost in themuch less hypsodont M. azanzae.The ‘hypsodont’ lower molars of M. mirus are more similar to
those of M. soriae than to those of earlier Micromeryx species.The size of the m2 relative to the m1 and the tall cristids are alsosimilar to M. soriae. However, M. mirus retains a regular Palaeo-meryx-fold, has more developed ectostylids and anterobuccalcingula in the m2, and is smaller thanM. soriae (Table 1S). Thesedifferences suggest that M. mirus is more primitive than M. sor-iae. M. soriae and M. mirus indicate that a new type of advancedMicromeryx with a more hypsodont dentition was present in theupper Vallesian-lower Turolian of Europe, characterizing theEuropean moschid fauna of the middle upper Miocene. The den-tition of these two species demonstrates that the achievement ofhigh-crowned molars is a relatively common phenomenonamongst the Miocene Moschidae since it has occurred in bothMicromeryx and Hispanomeryx. Nevertheless, the developmentof this feature clearly differs between these two genera. Even M.soriae and M. mirus show less flattened cusps, less developedcristids and more developed lingual structures than Hispano-meryx. The general morphology of the molars in Hispanomeryxis very bovid-like, featuring flat lower molars with tall cristids andalmost non-existent stylids and lingual ribs; also, the Palaeo-meryx-fold is always absent. The relative increase in crown heightis often correlated with an adaptive trend to more open habitatswith increased aridity levels, and the resulting exposure of teethto a more abrasive diet (Janis, 1988). Thus, the high-crownedmolars of M. soriae and the seasonally dry habitats that thisspecies was thought to inhabit in the Iberian Peninsula (seeAlcala, 1994; van Dam, 1997; Anton and Morales, 2000) couldbe correlated, and probably constituted a competitive advantageagainst the more primitive forms ofMicromeryx.The specific diversity of the Iberian Moschidae reached its
maximum peak between local zone G3 and local zone H, withthe ubiquitous M. azanzae as the dominant form (MN 7/8 toLower MN 9). Those moschid communities disappeared at theend of the local zone H, being replaced by new moschid assem-blages in the upper Vallesian (local zone J, MN 10). In spite ofthat, moschids never resumed their former diversity in theIberian Peninsula (Sanchez and Morales, 2006). M. soriae prob-ably entered the Iberian Peninsula between the demise of theAragonian-lower Vallesian moschid communities and the be-ginning of the upper Vallesian (MN 10). Of the two localitiesin which fossils of this taxon have been found, the slightly oldersites (Batallones-1 and Batallones-10; 8.976 Ma) show a morediverse assemblage of moschid species, with Micromeryx sp.‘large size’ (a still undescribed very large Vallesian species)and Hispanomeryx sp. cf. H. duriensis alongside M. soriae(Sanchez and Morales, 2006; Domingo et al., 2007b). However,only M. soriae appears in the younger site (La Roma-2; 8.789Ma), so this phenomenon can be interpreted as an indicator ofthe general decline of the moschids and the final demise of thegenus Micromeryx, which probably became extinct at the endof the Vallesian in the Iberian Peninsula.
CONCLUSIONS
Our knowledge about the previously poorly known Iberianupper Miocene moschids is updated through the present system-
SANCHEZ ET AL.—SPANISH UPPER MIOCENE MOSCHIDAE 573

atic revision of the more representative fossils from the upperVallesian-lower Turolian of Spain. This revision leads to a set ofsignificant conclusions with regard to the systematics, compara-tive morphology and biochronology of the youngest representa-tives of Hispanomeryx and Micromeryx.
The specimen PM-659 from Puente Minero is the youngestcorroborated record of the Moschidae in the Iberian Peninsula.It was previously assigned to Micromeryx sp. due to its smallsize, but more likely pertains to the genus Hispanomeryx on thebasis of morphology. The systematic utility of the morphology ofupper molars of Micromeryx and Hispanomeryx is clearlydemonstrated. Conversely, the systematic utility of size regard-ing Hispanomeryx and Micromeryx is strongly rebated, becauseof overlapping size ranges. The presence of a member of Hispa-nomeryx in the MN 11 revises the last occurrence of the genusin Spain, extending its biochronological range into the lowerTurolian.
Finally, M. soriae is the last of the recently described newspecies of Micromeryx that have been identified in the IberianPeninsula and continental Europe, demonstrating that this genuswas more diverse than previously thought. The new species ischaracterized by its relative hypsodont lower molars with aPalaeomeryx-fold of unique morphology, and clearly confirmsthe presence in the upper Vallesian of Spain of a type of derivedMicromeryx very similar to that described in the lower Turolian(MN 11) of continental Europe. M. soriae probably enteredthe Iberian Peninsula after the collapse of the Aragonian-lowerVallesian communities of moschids at the end of the local zoneH (early MN 9), and became one of the last representativesof the genus in the Iberian Peninsula. Its derived dentition prob-ably was a competitive advantage against the more primitiverepresentatives of the genus.
ACKNOWLEDGMENTS
We are very grateful to the late M. D. Soria (MNCN-CSIC)for starting this work with us, and for her continuous help andencouragement. We thank L. Alcala (Conjunto Paleontologicode Teruel-Dinopolis) for facilitating use of the fossil materialfrom La Roma-2 and Puente Minero; Ma D. Pesquero (MNCN-CSIC) and Y. Fernandez-Jalvo (MNCN-CSIC) for their assis-tance with the taphonomic aspects of the fossils of Micromeryxfrom Batallones; the reviewers of JVP for their comments andsuggestions; and R. Asher (University of Cambridge) for hisreview of the original manuscript and his useful corrections.Thanks also go to the Comunidad de Madrid-Direccion Generalde Patrimonio Historico and the Comunidad de Aragon-DGAfor their funding support. MSD acknowledges a predoctoralFPU contract from the Spanish Government MICINN. IMS ac-knowledges a postdoctoral contract from the Spanish Govern-ment MICINN. This study is part of the Spanish GovernmentMICINN research projects CGL2006-01773/BTE, CGL-2005-03900 and CGL2004-00400/BTE, and is included in the ResearchGroup CAM-UCM 910607.
LITERATURE CITED
Agustı, J., L. Cabrera, M. Garces, W. Krijgsman, O. Oms, and J. M. Pares.2001. A calibrated mammal scale for the Neogene of Western Eur-ope: state of the art. Earth-Science Reviews 52:247–260.
Alcala, L. 1994. Macromamıferos neogenos de la fosa Alfambra-Teruel.Instituto de Estudios Turolenses-Museo Nacional de CienciasNaturales, Teruel, 554 pp.
Alcala, L., and J. Morales. 1997. A primitive caprine from the UpperVallesian of La Roma 2 (Alfambra, Teruel, Aragon, Spain). Comp-tes Rendus Academie des Sciences, Paris 324:947–953.
Andrews, P. 1990. Owls, Caves and Fossils. Natural History MuseumPublications, London, 231 pp.
Anton, M., and J. Morales. 2000. Inferencias paleoecologicas de laasociacion de carnıvoros del yacimiento de Cerro Batallones; pp.190–201 in J. Morales, M. Nieto, L. Amezua, S. Fraile, E. Gomez,E. Herraez, P. Pelaez-Campomanes, M. J. Salesa, I. M. Sanchez, andD. Soria (eds.), Patrimonio Paleontologico de la Comunidad deMadrid. Serie Arqueologıa, Paleontologıa y Etnografıa 6, Comuni-dad Autonoma de Madrid.
Anton, M., M. J. Salesa, J. Morales, and A. Turner. 2004. First knowncomplete skulls of the scimitar-toothed cat Machairodus aphanistus(Felidae, Carnivora) from the Spanish late Miocene site of Batal-lones-1. Journal of Vertebrate Paleontology 24:957–969.
Azanza, B. 1986. Estudio geologico y paleontologico del Mioceno delsector oeste de la Comarca de Borja. Cuadernos de Estudios Borja-nos XVII–XVIII:63–126.
Azanza, B. 2000. Los Cervidae (Artiodactyla, Mammalia) del Miocenode las cuencas del Duero, Tajo, Calatayud–Teruel y Levante. Mem-orias del Museo Paleontologico de la Universidad de Zaragoza8:1–376.
Azanza, B., J. Morales, X. Murelaga, A. Sanchez, S. Fraile, E. Gomez,M. Nieto, B. Perez, M. J Salesa, I. M. Sanchez, and D. Soria. 1999.The middle Miocene vertebrate assemblage of Toril-3 (Zaragoza,Spain)—preliminary analysis. IV European Workshop on Verte-brate Paleontology. Programme and Abstracts, Field Guide 18.
Azanza, B., A. Alonso-Zarza, M. A. Alvarez-Sierra, J. P. Calvo, S. Fraile,I. Garcıa-Paredes, E. Gomez, M. Hernandez-Fernandez, A. van derMeulen, D. de Miguel, P. Montoya, J. Morales, X. Murelaga,P. Pelaez-Campomanes, B. Perez, V. Quiralte, M. J. Salesa, I. M.Sanchez, A. Sanchez-Marco, and D. Soria. 2004. Los yacimientosde vertebrados continentales de Aragoniense superior (Miocenomedio) de Toril, Cuenca de Calatayud-Daroca. GEO-TEMAS6:271–274.
Barone, R. 1999. Anatomie Comparee des Mammiferes Domestiques.Tome 1. Osteologie. Quatrieme edition, Vigot Freres Editeurs,Paris, 761 pp.
Cerdeno, E., and L. Alcala. 1989. Aceratherium alfambrense sp. n., nuevorinocerotido del Vallesiense superior de Teruel (Espana). RevistaEspanola de Paleontologıa 4:39–51.
Daams, R., L. Alcala, M. A. Alvarez Sierra, B. Azanza, J. A. van Dam,A.-J. van der Meulen, J. Morales, M. Nieto, P. Pelaez-Campomanes,and D. Soria. 1998. A stratigraphical framework for Miocene /Mn 4-MN 13) continental sediments of Central Spain. Comptes RendusAcademie des Sciences, Paris 327:625–631.
Domingo, M. S., M. T. Alberdi, and B. Azanza. 2007a. A new quantita-tive biochronological ordination for the Upper Neogene mammali-an localities of Spain. Palaeogeography, Palaeoclimatology,Palaeoecology 255:361–376.
Domingo, M. S., I. M. Sanchez, J. Morales, and M. T. Alberdi. 2007b.Miocene prey and predators: diversity of the Moschidae (Ruminan-tia; Pecora) from the carnivore–trap site of Batallones-1 (Late Val-lesian; MN 10; Madrid Basin; Spain) and evidence of carnivoretrophic activity on moschids. Journal of Vertebrate Paleontology 27(3, Supplement):69A.
Gray, J. E. 1821. On the natural arrangement of the vertebrose animals.The London Medical Repository Journal and Review 15:296–310.
Janis, C. M. 1988. An estimation of tooth volume and hypsodonty indicesin ungulate mammals, and the correlation of these factors withdietary preferences; pp. 367–387 in D. E. Russell, J. P. Santoro, andD. Sigogneau-Russell (eds.), Teeth Revisited. Memoires du Muse-um National d’Histoire Naturelle (Serie C) 53. Paris: Editions duMuseum.
Kohler, M. 1993. Skeleton and Habitat of Recent and Fossil Ruminants.Munchner Geowissenschaftliche Abhandlungen 25:1–88.
Lartet, E. 1851. Notice sur la Colline de Sansan, suvie d’une recapitula-tion des diverses especes d’animaux vertebres fossiles trouves soit aSansan, soit d’autres gisements du terrain tertiaire Miocene dans leBassin Sous-Pyreneen. J.-A. Portes: Imprimeur de la Prefecture etLibraire, 45 pp.
Linnaeus, C. 1758. Systema Naturae, Ed. X (Systema naturae per regnatria naturae, secundum classes, ordines, genera, species, cum char-acteribus, diferentiis, synonymis, locis. Tomus I. Editio decimareformata). Homiae (Stockholm), Sweden, 824 pp.
Morales, J. 2004. El yacimiento paleontologico de Cerro de los Batal-lones (Torrejon de Velasco, Madrid); pp. 245–277 in J. L. Sanz,(ed.), Aportaciones recientes en el conocimiento de la historia de laVida. Fundacion de Cultura, Ciudad de Cuenca.
574 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 29, NO. 2, 2009

Morales, J., and D. Soria. 1981. Los artiodactilos de Los Valles de Fuen-tiduena (Segovia). Estudios Geologicos 37:477–501.
Morales, J., S. Moya-Sola, and D. Soria. 1981. Presencia de la familiaMoschidae (Artiodactyla, Mammalia) en el Vallesiense de Espana:Hispanomeryx duriensis novo gen. nova sp. Estudios Geologicos37:467–475.
Morales, J., J. Capitan, J. P. Calvo, and D. Soria. 1992. Nuevo yacimientode vertebrados del Mioceno Superior al Sur de Madrid (CerroBatallones, Torrejon de Velasco). Geogaceta 12:77–80.
Morales, J., L. Alcala, L. Amezua, M. Anton, S. Fraile, E. Gomez,P. Montoya, M. Nieto, B. Perez, M. J. Salesa, and I. M. Sanchez.2000. Mioceno Superior: El Yacimiento de Cerro de los Batallones;pp. 178–190 in J. Morales, M. Nieto, L Amezua, S. Fraile, E. Gomez,E. Herraez, P. Pelaez-Campomanes, M. J. Salesa, I. M. Sanchez, andD. Soria (eds.), Patrimonio Paleontologico de la Comunidad deMadrid. Serie Arqueologıa, Paleontologıa y Etnografıa 6, Comuni-dad Autonoma de Madrid.
Morales, J., L. Alcala, MA. Alvarez-Sierra, M. Anton, B. Azanza,J. P. Calvo, P. Carrasco, S. Fraile, I. Garcıa-Paredes, E. Gomez,M. Hernandez-Fernandez, L. Merino, A. van der Meulen, C. MartınEscorza, P. Montoya, M. Nieto, S. Peigne, B. Perez, P. Pelaez-Campomanes, M. Pozo, V. Quiralte, M. J. Salesa, I. M. Sanchez,A. Sanchez-Marco, P. G. Silva, M. D. Soria, and A. Turner. 2004.Paleontologıa del sistema de yacimientos de mamıferos miocenos delCerro de los Batallones, Cuenca de Madrid. Geogaceta 35:139–142.
Moya-Sola, S. 1986. El genero Hispanomeryx Morales et al., (1981):posicion filogenetica y sistematica. Su contribucion al conocimientode la evolucion de los Pecora (Artiodactyla, Mammalia). Paleonto-logıa i Evolucio 20:267–287.
Nowak, R. M. 1999. Walker’s Mammals of the World. Sixth Edition.Volume II:The Johns Hopkins University Press, Baltimore andLondon, 1921 pp.
Owen, R. 1848. Description of teeth and portions of jaws of two extinctAnthracotherioid quadrupeds (Hyopotamus vectianus and Hyop.bovinus) discovered by the Marchioness of Hastings in the Eocenedeposits on the N. W. coast of the Isle of Wight: with an attempt todevelop Cuvier’s idea of the classification of pachyderms by thenumber of their toes. Quarterly Journal of the Geological Societyof London 4:103–141.
Peigne, S., M. J. Salesa, M. Anton, and J. Morales. 2005. Ailurid carni-voran mammal Simocyon from the late Miocene of Spain and thesystematics of the genus. Acta Palaeontologica Polonica 50:219–238.
Pesquero, M. D., M. T. Alberdi, and L. Alcala. 2006. New species ofHipparion from La Roma 2 (Late Vallesian; Teruel, Spain): a studyof the morphological and biometric variability of Hipparion primi-genium. Journal of Paleontology 80:343–356.
Pozo, M., J. P. Calvo, P. G. Silva, J. Morales, P. Pelaez-Campomanes,and M. Nieto. 2004. Geologıa del sistema de yacimientos de mamı-feros miocenos del Cerro de los Batallones, Cuenca de Madrid.Geogaceta 35:143–146.
Salesa, M. J. 2002. Estudio anatomico, biomecanico, paleoecologico yfilogenetico de Paramachairodus ogygia (Kaup, 1832) Pilgrim, 1913(Felidae, Machairodontinae) del yacimiento vallesiense (Miocenosuperior) de Batallones-1 (Torrejon de Velasco, Madrid). Ph. D.dissertation Universidad Complutense de Madrid, 370 pp.
Salesa, M. J., M. Anton, A. Turner, and J. Morales. 2005. Aspects of thefunctional morphology of the cranial and cervical skeleton of thesabre-toothed cat Paramachairodus ogygia (Kaup, 1832) (Felidae,Machairodontinae) from the late Miocene of Spain: implications forthe origin of the machairodont killing bite. Zoological Journal of theLinnean Society 144:363–377.
Salesa, M. J., M. Anton, S. Peigne, and J. Morales. 2006a. Evidence of afalse thumb in a fossil carnivore clarifies the evolution of pandas.Proceedings of the National Academy of Sciences 103:379–382.
Salesa, M. J., M. Anton, A. Turner, and J. Morales. 2006b. Inferredbehaviour and ecology of the primitive sabre-toothed cat Parama-chairodus ogygia (Felidae, Machairodontinae) from the late Mio-cene of Spain. Journal of Zoology 208:243–254.
Sanchez, I. M. 2006. Evolucion y Sistematica de losMoschidae (Mammalia;Artiodactyla) del Mioceno medio y superior de Espana. Ph. D. disser-tation, Universidad Complutense de Madrid, Madrid, Spain, 513 pp.
Sanchez, I. M., and J. Morales. 2004. Los rumiantes inermes (Artiodactyla;Mammalia) de la Comunidad de Madrid; pp. 519–524 in Zona Arqueo-logica. Miscelanea en homenaje a Emiliano Aguirre: Volumen II,Paleontologıa. Museo Arqueologico Regional, Alcala de Henares.
Sanchez, I. M., and J. Morales. 2006. Distribucion biocronologica de losMoschidae (Mammalia; Ruminantia) en Espana. Estudios Geologi-cos 62:533–546.
Sanchez, I. M., and J. Morales. 2008. Micromeryx azanzae sp. nov.(Ruminantia, Moschidae) from the Middle-Upper Miocene ofSpain, and the first description of the cranium of Micromeryx. Jour-nal of Vertebrate Paleontology 28:000–000.
Scopoli, J. A. 1777. Introdvctio ad historiam natvralem sistens generalapidvm, platarvm, et animalivm hactenvs detecta, caracteribvsessentialibvs donata, in tribvs divisa, svbinde ad leges natvrae.Pragae 506.
Thenius, E. 1950. Die tertiaren lagomeryciden und cerviden derSteiermark. Stzungsberichte der Oesterreichischen Akademie derWissenschaften, Mathematisch-Naturwissenschaftliche Klasse I. 159:219–254.
van Dam, J. A. 1997. The small mammals from the Upper Miocene ofthe Teruel-Alfambra region (Spain): paleobiology and paleoclimaticreconstructions. Geologica Ultraiectina 156:1–204.
Vislobokova, I. A. 2007. New data on Late Miocene Mammals of Kohfi-disch, Austria. Paleontological Journal 41:451–460.
Webb, S. D., and B. E. Taylor. 1980. The phylogeny of hornless rumi-nants and a description of the cranium of Archaeomeryx. Bulletin ofAmerican Museum of Natural History 167:117–158.
Submitted July 16, 2008; accepted October 20, 2008.
SANCHEZ ET AL.—SPANISH UPPER MIOCENE MOSCHIDAE 575
Related Documents











