ORIGINAL ARTICLE Neutrophils sustain effective CD8 + T-cell responses in the respiratory tract following influenza infection Michelle D Tate 1 , Andrew G Brooks 1 , Patrick C Reading 1,3 and Justine D Mintern 2 Neutrophils have an important role in early host protection during influenza A virus infection. Their ability to modulate the virus-specific adaptive immune response is less clear. Here, we have used a mouse model to examine the impact of neutrophils on CD8 + T-cell responses during influenza virus infection. CD8 + T-cell priming, expansion, migration, cytokine secretion and cytotoxic capacity were investigated in the virus-infected airways and secondary lymphoid organs. To do this, we utilised a Ly6G-specific monoclonal antibody (mAb; 1A8) that specifically depletes neutrophils in vivo. Neutrophil depletion early after infection with influenza virus strain HKx31 (H3N2) did not alter influenza virus-derived antigen presentation or naı¨ve CD8 + T-cell expansion in the secondary lymphoid organs. Trafficking of virus-specific CD8 + T cells into the infected pulmonary airways was also unaltered. Instead, early neutropenia reduced both the overall magnitude of influenza virus-specific CD8 + T cells, together with impaired cytokine production and cytotoxic effector function. Therefore, neutrophils are important participants in anti-viral mechanisms that sustain effective CD8 + T-cell responses in the respiratory tract of influenza virus-infected mice. Immunology and Cell Biology advance online publication, 12 April 2011; doi:10.1038/icb.2011.26 Keywords: influenza; CD8 + T cell; neutrophil and lung Neutrophil infiltration is a hallmark of the innate immune response to virus infection. 1 Immediately following influenza A virus infection, neutrophils provide critical protection by acting to limit viral repli- cation. 2–5 An emerging role for neutrophils in immunity is their contribution to the subsequent adaptive immune response. A large body of evidence implicates neutrophils in facilitating antigen pre- sentation to T cells and/or by modulating dendritic cell (DC) recruit- ment 6 and activation. 7,8 In addition, neutrophils may also directly impact T cells through their release of pro-inflammatory cytokines and chemokines. 9 Together, these mechanisms suggest that neutro- phils are a cell type with the potential to bridge innate and adaptive immune pathways. Neutrophil participation in adaptive immunity to influenza A virus infection has been previously examined for antibody responses where they were shown to have a critical role. 4 In contrast, the role of neutrophils in shaping anti-influenza A virus T-cell immunity has not been examined. Herein, we have investigated the hypothesis that neutrophils modulate the influenza-specific CD8 + T-cell response. In murine models of infection, the role of neutrophils is generally studied by employing antibody-mediated neutrophil depletion. Previous studies examining neutrophils in modulating T-cell responses have relied on mAb RB6-8C5. 10–12 Although mAb RB6- 8C5 binds the Ly6G antigen expressed on neutrophils, 13 it also targets the Ly6C antigen, which is expressed on a range of leukocyte populations, including CD8 + T cells. 3,14–16 A more specific protocol utilises the Ly6G-specific mAb 1A8, 17 which is now employed to study the role of neutrophils in models of infection 3,18–20 and wound healing. 17 Our own recent studies have demonstrated that mAb 1A8 recognises Ly6G high neutrophils, but not other leukocytes, such as lung macrophages, DC, CD8 + T cells and CD4 + T cells. 3 The mAb 1A8 therefore allows for specific depletion of neutrophils without com- promising other components of the immune response to viral infec- tion. We have previously utilised mAb1A8 to probe the significance of neutrophils in two major mouse models of viral infection. Confirming a critical role for neutrophils in influenza virus infection, ongoing 1A8-mediated neutrophil depletion during influenza virus infection elicited enhanced respiratory tract viral replication associated with exacerbated pulmonary inflammation, oedema and respiratory dysfunction. 3 In contrast, 1A8-mediated neutrophil depletion follow- ing herpes simplex virus (HSV)-1 infection did not provoke major alterations in HSV-1 titre or disease outcomes, suggesting an insigni- ficant role for neutrophils for HSV-1 resolution. 18,19 In this study, we have employed mAb 1A8 to examine the role of neutrophils in shaping CD8 + T-cell responses following influenza A virus infection of C57BL/6 mice. A potent anti-viral CD8 + T-cell response is a major component of resolving primary influenza A virus infection. 21 Here, we show that mAb 1A8 induced systemic neutro- paenia, but did not directly deplete influenza-specific CD8 + T cells in vivo. Treatment of mice with mAb 1A8 early after influenza virus infection did not alter influenza virus antigen presentation, CD8 + Received 30 September 2010; revised and accepted 23 February 2011 1 Department of Microbiology and Immunology, University of Melbourne, Melbourne, Victoria, Australia; 2 Immunology Division, Walter and Eliza Hall Institute of Medical Research, Melbourne, Victoria, Australia and 3 WHO Collaborating Centre for Reference and Research on Influenza, Melbourne, Victoria, Australia Correspondence: Dr JD Mintern, Immunology Division, Walter and Eliza Hall Institute of Medical Research, Melbourne 3010, Victoria, Australia. E-mail: [email protected] Immunology and Cell Biology (2011), 1–9 & 2011 Australasian Society for Immunology Inc. All rights reserved 0818-9641/11 www.nature.com/icb

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Neutrophils sustain effective CD8+ T-cell responsesin the respiratory tract following influenza infection
Michelle D Tate1, Andrew G Brooks1, Patrick C Reading1,3 and Justine D Mintern2
Neutrophils have an important role in early host protection during influenza A virus infection. Their ability to modulate thevirus-specific adaptive immune response is less clear. Here, we have used a mouse model to examine the impact of neutrophilson CD8+ T-cell responses during influenza virus infection. CD8+ T-cell priming, expansion, migration, cytokine secretion andcytotoxic capacity were investigated in the virus-infected airways and secondary lymphoid organs. To do this, we utiliseda Ly6G-specific monoclonal antibody (mAb; 1A8) that specifically depletes neutrophils in vivo. Neutrophil depletion early afterinfection with influenza virus strain HKx31 (H3N2) did not alter influenza virus-derived antigen presentation or naı̈ve CD8+
T-cell expansion in the secondary lymphoid organs. Trafficking of virus-specific CD8+ T cells into the infected pulmonary airwayswas also unaltered. Instead, early neutropenia reduced both the overall magnitude of influenza virus-specific CD8+ T cells,together with impaired cytokine production and cytotoxic effector function. Therefore, neutrophils are important participantsin anti-viral mechanisms that sustain effective CD8+ T-cell responses in the respiratory tract of influenza virus-infected mice.Immunology and Cell Biology advance online publication, 12 April 2011; doi:10.1038/icb.2011.26
Keywords: influenza; CD8+ T cell; neutrophil and lung
Neutrophil infiltration is a hallmark of the innate immune response tovirus infection.1 Immediately following influenza A virus infection,neutrophils provide critical protection by acting to limit viral repli-cation.2–5 An emerging role for neutrophils in immunity is theircontribution to the subsequent adaptive immune response. A largebody of evidence implicates neutrophils in facilitating antigen pre-sentation to T cells and/or by modulating dendritic cell (DC) recruit-ment6 and activation.7,8 In addition, neutrophils may also directlyimpact T cells through their release of pro-inflammatory cytokinesand chemokines.9 Together, these mechanisms suggest that neutro-phils are a cell type with the potential to bridge innate and adaptiveimmune pathways. Neutrophil participation in adaptive immunity toinfluenza A virus infection has been previously examined for antibodyresponses where they were shown to have a critical role.4 In contrast,the role of neutrophils in shaping anti-influenza A virus T-cellimmunity has not been examined. Herein, we have investigated thehypothesis that neutrophils modulate the influenza-specific CD8+
T-cell response.In murine models of infection, the role of neutrophils is generally
studied by employing antibody-mediated neutrophil depletion.Previous studies examining neutrophils in modulating T-cellresponses have relied on mAb RB6-8C5.10–12 Although mAb RB6-8C5 binds the Ly6G antigen expressed on neutrophils,13 it also targetsthe Ly6C antigen, which is expressed on a range of leukocytepopulations, including CD8+ T cells.3,14–16 A more specific protocol
utilises the Ly6G-specific mAb 1A8,17 which is now employed to studythe role of neutrophils in models of infection3,18–20 and woundhealing.17 Our own recent studies have demonstrated that mAb 1A8recognises Ly6Ghigh neutrophils, but not other leukocytes, such aslung macrophages, DC, CD8+ T cells and CD4+ T cells.3 The mAb 1A8therefore allows for specific depletion of neutrophils without com-promising other components of the immune response to viral infec-tion. We have previously utilised mAb1A8 to probe the significance ofneutrophils in two major mouse models of viral infection. Confirminga critical role for neutrophils in influenza virus infection, ongoing1A8-mediated neutrophil depletion during influenza virus infectionelicited enhanced respiratory tract viral replication associatedwith exacerbated pulmonary inflammation, oedema and respiratorydysfunction.3 In contrast, 1A8-mediated neutrophil depletion follow-ing herpes simplex virus (HSV)-1 infection did not provoke majoralterations in HSV-1 titre or disease outcomes, suggesting an insigni-ficant role for neutrophils for HSV-1 resolution.18,19
In this study, we have employed mAb 1A8 to examine the role ofneutrophils in shaping CD8+ T-cell responses following influenza Avirus infection of C57BL/6 mice. A potent anti-viral CD8+ T-cellresponse is a major component of resolving primary influenza A virusinfection.21 Here, we show that mAb 1A8 induced systemic neutro-paenia, but did not directly deplete influenza-specific CD8+ T cellsin vivo. Treatment of mice with mAb 1A8 early after influenza virusinfection did not alter influenza virus antigen presentation, CD8+
Received 30 September 2010; revised and accepted 23 February 2011
1Department of Microbiology and Immunology, University of Melbourne, Melbourne, Victoria, Australia; 2Immunology Division, Walter and Eliza Hall Institute of Medical Research,Melbourne, Victoria, Australia and 3WHO Collaborating Centre for Reference and Research on Influenza, Melbourne, Victoria, AustraliaCorrespondence: Dr JD Mintern, Immunology Division, Walter and Eliza Hall Institute of Medical Research, Melbourne 3010, Victoria, Australia.E-mail: [email protected]
Immunology and Cell Biology (2011), 1–9& 2011 Australasian Society for Immunology Inc. All rights reserved 0818-9641/11
www.nature.com/icb

T-cell priming or CD8+ T-cell migration. Instead, the early absence ofneutrophils significantly impaired the magnitude of the influenza-specific CD8+ T-cell response in the airways, as well as the ability ofthese cells to produce effector cytokines and cytotoxicity.
RESULTSKinetics of neutrophil and CD8+ T-cell responses followinginfluenza A virus infectionFirst, we examined the kinetics and magnitude of neutrophil andCD8+ T-cell responses in the airways of B6 mice infected withinfluenza A virus. Infection of mice with 102 plaque-forming unit(PFU) of HKx31 was characterised by rapid recruitment of neutro-phils to the airways (Figure 1a) and subsequently by CD8+ T cells(Figure 1b), including CD8+ T cells specific for influenza virus acidpolymerase (PA) presented by H2-Db (DbPA224) and CD8+ T cellsspecific for the influenza A virus nucleoprotein-derived epitopepresented by H2-Db (DbNP366; Figure 1c). DbPA224 and DbNP366represent the two major immunodominant epitopes, followingprimary infection of C57BL/6 mice with HKx31 influenza virus.29
Expansion and accumulation of CD8+ T cells in the airways correlatedwith the control of infection and influenza virus clearance (Figure 1d).
A treatment regime for depletion of neutrophils during the earlyphase of influenza virus infectionOur next objective was to establish an effective regime to depleteneutrophils without direct depletion of virus-specific CD8+ T cells. Wefirst characterised expression of Ly6C, Ly6G and Gr-1 on CD8+ T cellsrecovered from mice infected with 105 PFU of influenza virus strainHKx31. Figure 2 shows binding of mAb RB6-8C5 (anti-Ly6C/G;Gr-1), mAb 1A8 (anti-Ly6G) and mAb AL-21 (anti-Ly6C) to total(Figure 2a) and DbPA224-specific CD8+ T cells (Figure 2b). Gr-1, asdetected by binding of mAb RB6-8C5, was expressed by both totaland virus-specific CD8+ T cells, correlating with Ly6C expression, asdetected by mAb AL-21. In contrast, mAb 1A8 (anti-Ly6G) did notbind to total bronchoalveolar lavage (BAL) and/or spleen CD8+ Tcells from naı̈ve mice, nor to total or DbPA224-specific CD8+ frominfluenza virus-infected mice.Next, we determined whether mAb RB6-8C5 and/or mAb 1A8
directly depleted CD8+ T cells during influenza virus infection.A single treatment of mice with mAb 1A8 at day 7 after infectiondid not alter numbers of total or DbPA224-specific CD8
+ T cells in theairways relative to treatment with control rat immunoglobulin G(IgG). In contrast, significantly reduced numbers of total andDbPA224-specific CD8
+ T cells were recovered from mice that receiveda single treatment with mAb RB6-8C5 (Figure 2c). Together,these data indicate that mAb RB6-8C5 can bind to and depleteCD8+ T cells following influenza infection because of its ability tobind Ly6C expressed by these cells. In contrast, mAb 1A8 does notbind to total or virus-specific CD8+ T cells, and therefore does notdeplete this cell population directly during infection. As such, mAb1A8 is an ideal antibody to utilise for examining the contributionof neutrophils to CD8+ T-cell responses following influenza A virusinfection.To examine the direct role of neutrophils in adaptive immunity
following influenza infection, an optimal neutrophil-depletion proto-col would enable neutrophil depletion in the absence of majordifferences in viral replication. Increases in influenza virus titrewould likely promote more robust anti-viral CD8+ T-cell immunityindependently of neutropenia. We therefore aimed to establish aprotocol that allowed sufficient neutrophil depletion, accompaniedby a limited increase in influenza virus titre. Two major neutrophil
depletion regimes were tested: administration of 1A8 mAb at days!1, +1 and +3 (‘early’) or days !1, +1, +3 and +5 (‘full’) relative toinfluenza virus infection. Our previous studies show that repeatedadministration of 1A8 mAb effectively depletes neutrophils.3 In thecase of the ‘early’ 1A8 mAb treatment protocol, BAL neutrophilnumbers are restored to levels present in control IgG-treated miceat day 7 (data not shown). Examination of lung tissue at day 7showed that both depletion regimes elicited increased influenza virustitre (Figure 2d), with the ‘full’ protocol eliciting a 27.5-fold increaserelative to IgG treatment, compared with a 4.5-fold increase with the‘early’ treatment protocol. The 1A8 treatment also elicited significantmouse weight loss regardless of whether the ‘early’ or ‘full’ treatment
Virus-specific
CD8+
TotalCD8+
00.20.40.60.81.01.21.41.6
Naïve 1 3 5 7 9 11
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Naïve 1 3 5 7 9 11
DbPA224
DbNP366
Vir
al ti
tre
(log 1
0PFU
/sam
ple)
0
1
2
3
4
5
6
1 3 5 7 9 11
Day post-infection
0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Naïve 1 3 5 7 9 11
Tota
l num
ber
cells
(x10
5 )
Tota
l num
ber
cells
(x10
5 )
Tota
l num
ber
cells
(x10
5 )
Neutrophils
Figure 1 Neutrophil and CD8+ T-cell kinetics in response to intranasalinfection with influenza A virus HKx31. B6 mice were i.n. infected with102 PFU of HKx31. Total numbers of BAL (a) CD45+ Gr-1high neutrophils,(b) total CD8+ T cells and (c) DbPA224-and DbNP366-specific CD8+ T cells.(d) Viral load in the lung following infection with 102 PFU of HKx31. Barsrepresent mean cell number±1s.d., n"4–5 mice per group. The detectionlimit of the plaque assay is indicated by the dotted line.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
2
Immunology and Cell Biology
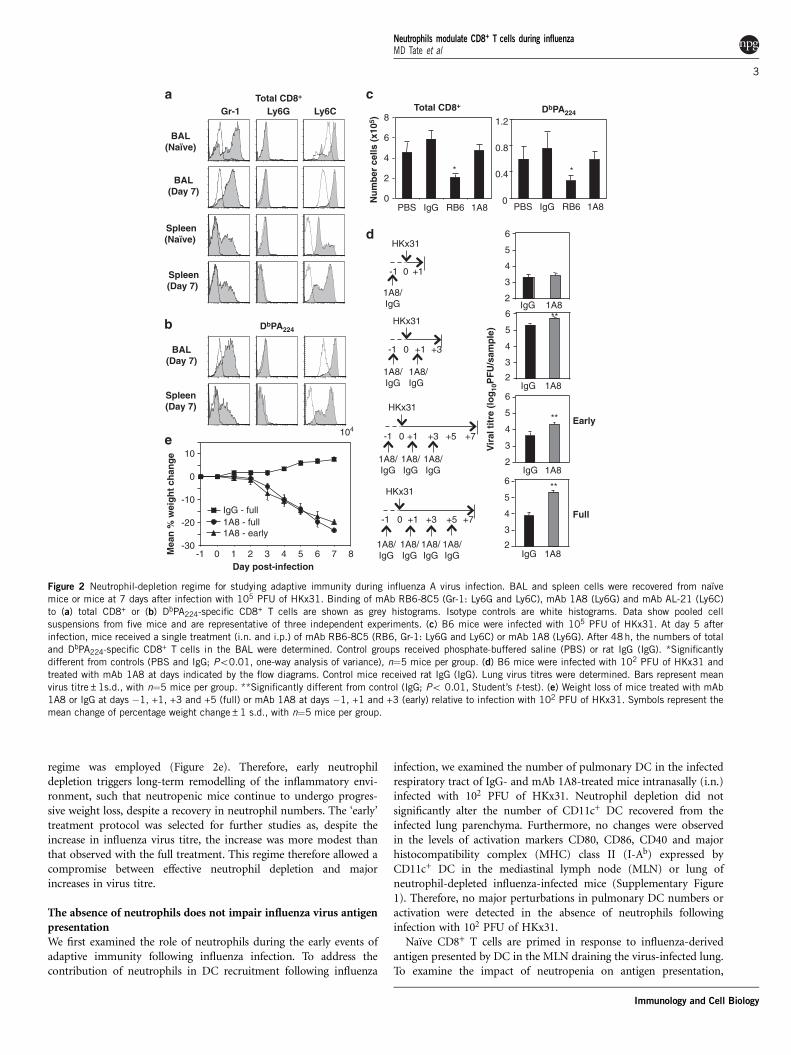
regime was employed (Figure 2e). Therefore, early neutrophildepletion triggers long-term remodelling of the inflammatory envi-ronment, such that neutropenic mice continue to undergo progres-sive weight loss, despite a recovery in neutrophil numbers. The ‘early’treatment protocol was selected for further studies as, despite theincrease in influenza virus titre, the increase was more modest thanthat observed with the full treatment. This regime therefore allowed acompromise between effective neutrophil depletion and majorincreases in virus titre.
The absence of neutrophils does not impair influenza virus antigenpresentationWe first examined the role of neutrophils during the early events ofadaptive immunity following influenza infection. To address thecontribution of neutrophils in DC recruitment following influenza
infection, we examined the number of pulmonary DC in the infectedrespiratory tract of IgG- and mAb 1A8-treated mice intranasally (i.n.)infected with 102 PFU of HKx31. Neutrophil depletion did notsignificantly alter the number of CD11c+ DC recovered from theinfected lung parenchyma. Furthermore, no changes were observedin the levels of activation markers CD80, CD86, CD40 and majorhistocompatibility complex (MHC) class II (I-Ab) expressed byCD11c+ DC in the mediastinal lymph node (MLN) or lung ofneutrophil-depleted influenza-infected mice (Supplementary Figure1). Therefore, no major perturbations in pulmonary DC numbers oractivation were detected in the absence of neutrophils followinginfection with 102 PFU of HKx31.Naı̈ve CD8+ T cells are primed in response to influenza-derived
antigen presented by DC in the MLN draining the virus-infected lung.To examine the impact of neutropenia on antigen presentation,
BAL(Day 7)
Spleen(Day 7)
Gr-1 Ly6G Ly6C
BAL(Naïve)
BAL(Day 7)
Total CD8+
Total CD8+
Spleen(Naïve)
Spleen(Day 7)
DbPA224
Num
ber
cells
(x10
5 )
PBS IgG RB6 1A80
2
4
6
8
*
DbPA224
0
0.4
0.8
1.2
PBS IgG RB6 1A8
*
-30
-20
-10
0
10
-1 0 1 2 3 4 5 6 7 8
104
IgG - full
1A8 - early1A8 - full
Day post-infection
Mea
n %
wei
ght c
hang
e
IgG 1A8
Vir
al ti
tre
(log 1
0PFU
/sam
ple)
**
2
3
4
5
6
Early
IgG 1A8
**
2
3
4
5
6
Full
2
3
4
5
6
IgG 1A8**
2
3
4
5
6
IgG 1A8
+7
1A8/IgG
-1
1A8/IgG
0 +1
HKx31
+3
1A8/IgG
+5
1A8/IgG
1A8/IgG
-1
1A8/IgG
0 +1
HKx31
+3
1A8/IgG
+5 +7
-1
1A8/IgG
0 +1
HKx31
-1
1A8/IgG
1A8/IgG
0 +1
HKx31
+3
Figure 2 Neutrophil-depletion regime for studying adaptive immunity during influenza A virus infection. BAL and spleen cells were recovered from naı̈vemice or mice at 7 days after infection with 105 PFU of HKx31. Binding of mAb RB6-8C5 (Gr-1: Ly6G and Ly6C), mAb 1A8 (Ly6G) and mAb AL-21 (Ly6C)to (a) total CD8+ or (b) DbPA224-specific CD8+ T cells are shown as grey histograms. Isotype controls are white histograms. Data show pooled cellsuspensions from five mice and are representative of three independent experiments. (c) B6 mice were infected with 105 PFU of HKx31. At day 5 afterinfection, mice received a single treatment (i.n. and i.p.) of mAb RB6-8C5 (RB6, Gr-1: Ly6G and Ly6C) or mAb 1A8 (Ly6G). After 48h, the numbers of totaland DbPA224-specific CD8+ T cells in the BAL were determined. Control groups received phosphate-buffered saline (PBS) or rat IgG (IgG). *Significantlydifferent from controls (PBS and IgG; Po0.01, one-way analysis of variance), n"5 mice per group. (d) B6 mice were infected with 102 PFU of HKx31 andtreated with mAb 1A8 at days indicated by the flow diagrams. Control mice received rat IgG (IgG). Lung virus titres were determined. Bars represent meanvirus titre±1s.d., with n"5 mice per group. **Significantly different from control (IgG; Po 0.01, Student’s t-test). (e) Weight loss of mice treated with mAb1A8 or IgG at days !1, +1, +3 and +5 (full) or mAb 1A8 at days !1, +1 and +3 (early) relative to infection with 102 PFU of HKx31. Symbols represent themean change of percentage weight change±1 s.d., with n"5 mice per group.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
3
Immunology and Cell Biology

we utilised an engineered HKx31 virus expressing the OVA257!264
peptide within the neuraminidase stalk (HKx31-OVA). Togetherwith the T cell receptor (TCR) transgenic OT-I CD8+ T cells, thismodel enables assessment of antigen presentation following influenzainfection. Mice were treated with mAb 1A8 (or control rat IgG) at days!1, +1 and +3 relative to challenge with 104 PFU of HKx31-OVA. Inthis case, a higher viral dose was utilised, given the reduced virulenceof the engineered HKx31-OVA virus relative to the parental strain.2#106 carboxyfluorescein succinimidyl ester (CFSE)-labelled OT-Iwere injected intravenously (i.v.) into naive or HKx31-OVA-infectedrecipients at 24h after infection. Antigen-specific division of CD8+
OT-I in response to H2-Kb-OVA254!267 in the lung draining MLN wassimilar in IgG- or mAb 1A8-treated mice (Figure 3a). This indicatedthat presentation of OVA254!267 derived from HKx31-OVA was intactin the absence of neutrophils. In accordance with unimpaired antigenpresentation, CD8+ T-cell priming and expansion in the secondarylymphoid organs were also unaltered by neutropenia. Similar numbersof total virus-specific CD8+ T cells, DbPA224-specific and DbNP366-specific CD8+ T cells, were recovered from the spleen and MLN ofIgG- versus 1A8-treated mice (Figure 3b). The phenotype of virus-specific CD8+ T cells was also similar, as expression of CD69, CD44,CD25, CD62L and PD-1 was not different between IgG- and mAb1A8-treated mice (data not shown). These results indicated thatinfluenza-derived antigen presentation and influenza-specific CD8+
T-cell priming in the secondary lymphoid organs were not signifi-cantly perturbed in the absence of neutrophils.
Neutrophils do not alter CD8+ T-cell trafficking to the infectedairwaysFollowing priming in the lymphoid organs, virus-specific CD8+ T cellsundergo clonal expansion and migration into the infected respiratorytract. Therefore, we next examined whether CD8+ T-cell traffickingwas altered in the absence of neutrophils. 107 in vitro activated OT-Iwere adoptively transferred into mice infected with 102 PFU of HKx31at day 5 after infection. Mice were treated with mAb 1A8 (or controlrat IgG) at days !1, +1 and +3 relative to virus challenge. At 48h afterOT-I transfer, the number of OT-I cells in the lung was determined.As seen in Figure 4a, neutropaenia did not impact the numberof in vitro activated OT-I that migrated into the BAL or lung ofinfluenza-infected IgG- or 1A8-treated mice.A possible caveat to this experiment is that in vitro activation of
OT-I elicits T cells that do not resemble those generated during naturalinfection. Therefore, to further assess whether neutrophils modulateCD8+ T-cell recruitment into the lung, CD8+ T cells were purifiedfrom influenza virus-infected Ly5.1+ mice at day 7 after infection.In all, 106 Ly5.1 influenza-primed CD8+ T cells were transferredinto intact or neutropenic Ly5.2+ mice at day 3 following infectionwith HKx31. No differences were observed in the number of Ly5.1+
DbPA224-or DbNP366-specific CD8
+ T cells recovered from the lung atday 7 after infection (Figure 4b). Conversely, no significant differenceswere noted when purified virus primed CD8+ T cells from neutro-penic or IgG-treated influenza-infected Ly5.1+ mice were transferredinto intact influenza-infected Ly5.2+ mice (Figure 4c). Therefore,
Naïve1A8
HKx31-OVA1A8
HKx31-OVAIgG
DbPA224 DbNP366
Spleen
MLN
Total
0.2
0.6
1.0
IgG 1A80
2
4
6
IgG 1A8
0
2
4
6
8
IgG 1A80
0.4
0.8
1.2
IgG 1A8
Cel
l num
ber
(x10
4 )
Cel
l num
ber
(x10
5 )
Cel
l num
ber
(x10
5 )
Cel
l num
ber
(x10
6 )
0
0.4
0.8
1.2
IgG 1A8
0.2
1.0
1.8
IgG 1A8
CFSE
1A8/IgG
1A8/IgG
-1 0 +1
HKx31-OVA OT-I
+3MLN
analysis
Figure 3 The absence of neutrophils does not impair influenza virus antigen presentation. (a) Mice were treated with mAb 1A8 at day !1 and every 48hthereafter. Control mice received rat IgG (IgG). Mice were inoculated with 104 PFU of HKx31-OVA. 2#106 carboxyfluorescein succinimidyl ester (CFSE)-labelled OT-I cells were injected i.v. into B6 recipients at 24h after infection. MLN were harvested at 60h after transfer and CFSE+ CD8+ Ly5.1+ OT-I cellswere analysed for their CFSE florescence by flow cytometry. OT-I cells were also transferred into 1A8-treated, uninfected mice (naı̈ve) as a control. Data arerepresentative of two independent experiments, with n"3 mice per group. (b) B6 mice were treated with mAb 1A8 on days !1, +1 and +3 relative toinfection with 102 PFU of HKx31. Control mice received rat IgG (IgG). At day 7 after infection, the number of total, DbPA224- and DbNP366-specific CD8+ Tcells in the MLN and spleen was determined. Data shown are from one experiment (n"5 mice per group) and are representative of two independentexperiments.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
4
Immunology and Cell Biology

neutrophils do not exert a major impact on influenza virus-specificCD8+ T-cell trafficking to the airways.
Reduced CD8+ T-cell numbers in the respiratory tract ofneutropenic miceWe next examined CD8+ T-cell responses at the site of infection ininfluenza-infected respiratory tissues. To do this, BAL and lung wereharvested from IgG- or 1A8-treated mice following infection with 102
PFU of HKx31. The magnitude of total CD8+ T cells was determined,
together with the number of DbPA224 DbNP366 or specific CD8+
T cells. mAb 1A8 treatment of mice during the early phase of influenzainfection (that is, !1, +1 and +3 relative to infection) resulted inreduced DbPA224-specific and DbNP366-specific CD8+ T cells reco-vered from the BAL at day 5 (Figure 5a) and reduced DbPA224-specificCD8+ T cells recovered from the BAL at day 7 (Figure 5b). Consistentwith this, early neutrophil depletion also impacted influenza-specificCD8+ T-cell populations recovered from the infected lung paren-chyma (Figure 5c). These results implicate neutrophils in sustaining
1.0
1.5
2.0
2.5
3.0
3.5
IgG 1A8
Num
ber
OT-
I cel
ls(x
105 )
0
0.5
1.0
1.5
2.0IgG1A8
BAL Lung
4
6
8
10
12
IgG 1A8
0
0.5
1.0
1.5
2.0
2.5
DbPA224 DbNP366
DbPA224 DbNP366
IgG1A8
In vitro activated OT-I influenza infected mice (± 1A8)
Influenza primed CD8+ T cells influenza infected mice (± 1A8)
Influenza primed CD8+ T cells (± 1A8) influenza infected mice
-1 0 +1
1A8/IgG
1A8/IgG
+3
1A8/IgG
HKx31
+5
in vitroactivated OT-I
+7
BAL andlung
analysis
-1 0 +1
1A8/IgG
1A8/IgG
+3
1A8/IgG
+5
BALandlung
analysis
0 +7
HKx31
CD8+
enrichment
Ly5.1
Ly5.1
-1 0 +1
1A8/IgG
1A8/IgG
+3
1A8/IgG
HKx31
+7
HKx31
0
CD8+
enrichment
+3
BALandlung
analysis
Num
ber
Ly5.
1+ C
D8+
tetr
amer
+ ce
lls (x
103 )
Num
ber
Ly5.
1+ C
D8+
tetr
amer
+ ce
lls (x
103 )
+5
Figure 4 Neutrophils do not alter CD8+ T-cell trafficking to the infected airways. (a) Ly5.2 mice were depleted of neutrophils via i.n. and i.p. doses ofanti-Ly6G antibody (1A8) at days !1, +1 and +3. Control mice received rat IgG (IgG). Mice were inoculated with 102 PFU of HKx-31 and at day 5after infection, 107 in vitro activated Ly5.1+ OT-I cells were intravenously (i.v.) transferred. At 48h after transfer, the number of CD8+ Ly5.1+ OT-I cells inthe BAL and lung were determined. Data shown are from one experiment (n"5 mice per group) and are representative of two independent experiments.(b) Ly5.1 mice were inoculated with 102 PFU of HKx31. At day 7 after infection, Ly5.1+ CD8+ cells were enriched. 106 virus-primed Ly5.1+ CD8+ cellswere i.v. injected into 1A8- or IgG-treated naı̈ve Ly5.2 recipients. At 48h following transfer, the number of CD8+ Ly5.1+ DbPA224- or D
bNP366-specific CD8+
T cells in the lung was determined. Data shown are from one experiment (n"5 mice per group) and are representative of two independent experiments.(c) Ly5.1 mice were depleted of neutrophils via i.n. and i.p. doses of anti-Ly6G antibody (1A8) at days !1, +1 and +3. Control groups received rat IgG (IgG).At day 7 following infection with 102 PFU of HKx31, virus-primed Ly5.1+ CD8+ cells were enriched (as described in Methods) and transferred into Ly5.2mice inoculated 3 days previously with 102 PFU of HKx31. At 48h post-transfer, the number of CD8+ Ly5.1+ DbPA224- or DbNP366-specific CD8+ T cells inthe lung were determined. Data shown are from one experiment (n"5 mice per group) and are representative of two independent experiments.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
5
Immunology and Cell Biology

virus-specific CD8+ T-cell responses at the site of infection in therespiratory tract.
Impaired effector function of CD8+ T cells in the respiratory tractof neutropenic miceThe residual virus-specific CD8+ T-cell populations present in thelungs of neutropenic influenza-infected mice were examined foraltered phenotype and/or function in the absence of neutrophils.First, the capacity to secrete effector cytokines interferon-g andtumor necrosis factor-a was examined. Previous studies have demon-strated that influenza-specific CD8+ T cells have the capacity toproduce multiple cytokines upon re-stimulation with their cognateMHC-I peptide.27 Within a given influenza antigen-specific CD8+ T-cell population, all cells secrete interferon-g following re-stimulation,whereas only a proportion of this population also produces tumornecrosis factor-a. In our studies, DbNP366- and D
bPA224-specific CD8+
T cells displayed reduced production of interferon-g and tumornecrosis factor-a in the BAL and lung (Figure 6a) of neutrophil-depleted mice following in vitro re-stimulation with PA224 and NP366peptide. Therefore, the influenza-infected lungs of neutropenic micecontained reduced numbers of cytokine-producing effector CD8+
T cells.Virus-specific CD8+ T cells elicit effector function via the
targeted release of cytolytic molecules that mediate apoptosis of the
virus-infected cell. Granzyme B, a serine protease, is the majorcytolytic effector molecule expressed by influenza-specific CD8+
T cells.28 We therefore examined intracellular grzanzyme B contentfor virus-specific CD8+ T cells harvested from BAL of neutrophil-depleted mice. Interestingly, early treatment with mAb 1A8 resulted inimpaired granzyme B expression, relative to IgG-treated controls, forboth DbNP366- and DbPA224-specific CD8
+ T cells (Figure 6b). There-fore, in addition to reducing the overall magnitude of the influenzavirus-specific CD8+ T-cell response, the absence of neutrophils alsorendered the remaining CD8+ T cells less effective due to theirimpaired cytokine and cytotoxic effector functions.
DISCUSSIONNeutrophils are critical participants in anti-microbial innate immu-nity. Here, we have established a regime for defining the role ofneutrophils in modulating CD8+ T-cell adaptive immunity duringinfluenza virus infection. Our analysis employed mAb 1A8 to speci-fically target murine Ly6G-, but not Ly6C-, expressing leukocytes. Thistreatment provided specific depletion of neutrophils but not otherimmune cell types, in particular virus-specific CD8+ T cells. Here, wehave dissected the impact of neutrophils on the priming, expansion,migration and effector phases of CD8+ T-cell immunity in a mouse
DbPA224 DbNP366
Lung
0
1
2
3
4
IgG 1A8
*
0
1
2
IgG 1A8
*
Cel
l num
ber
(x10
2 )
Cel
l num
ber
(x10
3 )
Cel
l num
ber
(x10
4 )
0
1
2
3
IgG 1A8
*
0
1
2
3
4
IgG 1A8
*Day 5BAL
0
2
4
6
8
IgG 1A80
1
2
3
IgG 1A8
*
Day 7BAL
Figure 5 Reduced CD8+ T-cell numbers in the respiratory tract ofneutropenic mice. B6 mice were treated with mAb 1A8 on days !1, +1 and+3 relative to infection with 102 PFU of HKx31. Control groups received ratIgG (IgG). The number of DbPA224-specific or DbNP366-specific CD8+ T cellsrecovered from the (a) BAL (day 5), (b) BAL (day 7) or (c) lung (day 7) wasdetermined. *Significantly different to control (IgG) (po 0.05, Student’s t-test). Data shown are from one experiment (n"5 mice per group) and arerepresentative of two independent experiments.
IgG 1A8
800
300
0
0
2
4
6
8
*
IgG1A8
0
2
4
6
8
10
12
*
IgG1A8
BAL
% IF
N!+
% IF
N!+
IFN!+
% T
NF"
+%
TN
F"+
TNF"+
0
1
2
3
4
5
6
**
IgG1A8
Lung
0
1
2
3
4
5
6
7
*
IgG1A8
*
IgG 1A8
500
300
100
DbPA22
* *
DbPA224 DbNP366
DbPA224 DbNP366DbPA224 DbNP366
DbPA224 DbNP366
DbNP3662
Gzm
Bflo
resc
ence
inte
nsity
Figure 6 Impaired effector function of CD8+ T cells in the respiratory tractof neutropenic mice. B6 mice were treated with mAb 1A8 on days !1, +1and +3 relative to infection with 102 PFU of HKx31. Control mice receivedrat IgG (IgG). At day 7 post-infection, (a) the percentage of CD8+ T cellsexpressing interferon-g (IFNg) or tumor necrosis factor-a (TNFa) in the BALor lung in response to DbPA224 or DbNP366 peptide was determined. Barsrepresent the mean percentage ±1 s.d. (b) Intracellular granzyme B (gzmB) expression was determined for DbPA224- or DbNP366-specific CD8+ T cellsin the BAL. Symbols represent individual mice and the line designates themean. *Significantly different from IgG-treated control, Po0.05, Student’st-test. Data shown are from one experiment (n"5 mice per group) and arerepresentative of two independent experiments.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
6
Immunology and Cell Biology

model of respiratory influenza virus infection. In contrast toprevious reports that implicate a role for neutrophils in antigenpresentation,6–8,12,30–33 the absence of neutrophils during early influ-enza infection did not significantly impact influenza virus-specificantigen presentation or CD8+ T-cell priming in the secondarylymphoid organs. Interestingly, although our neutrophil depletionregime induced an increase in infectious influenza virus titre in therespiratory tissues, it did not elicit a corresponding increase in themagnitude of the CD8+ T-cell response as might be expected. Instead,the absence of neutrophils reduced total numbers of virus-specificCD8+ T cells recovered from the respiratory tract. The remainingvirus-specific CD8+ T cells displayed impaired cytokine productionand cytotoxic effector function. Together these data suggest thatneutrophils can indeed modulate CD8+ adaptive immunity, althoughtheir role in the antigen presentation phase of this response is lesscritical than previously anticipated.Neutrophils have the potential to serve multiple roles in antigen
presentation to T cells, including acting as antigen-presenting cellsthemselves.30–33 Capable of phagocytosing antigen, together with alimited life span, neutrophils could also provide a source of antigenthat is captured by DC for efficient T-cell priming. Indeed, antigentransfer between neutrophils and DC has been reported previously.12
Neutrophils release chemoattractants6,34,35 that signal recruitment ofDC into infected tissue, and activated neutrophils promote increasedDC activation7,8 and cytokine secretion.34 Despite these potential rolesfor neutrophils in antigen presentation, we observed no majordifferences in influenza virus-specific antigen presentation followingneutrophil depletion. Presentation of H2-Kb-OVA254!267 to CD8+ Tcells in the lymph node draining the HKx31-OVA-infected lung wasunaltered in neutropenic mice. In addition, similar numbers of virus-primed DbPA224- and DbNP366-specific CD8
+ T cells were present inthe lung-draining lymph node and spleen. This suggests that althoughneutrophils have the capacity to participate in antigen presentationpathways, their contribution is not essential for the liberation andpresentation of virus-derived antigen. Our data also suggested alimited role for neutrophils in DC recruitment to the infected airways.Numbers and activation status of CD11c+ DC were similar in IgG-and 1A8-treated mice. A possible caveat to this data is that more subtlealterations in specific respiratory DC subsets may occur in the absenceof neutrophils. This would require more detailed and sophisticatedDC analysis. Regardless, together, this data suggest that the absence ofneutrophils had no major impact on the presentation of influenza-derived antigen within the draining lymph node.Following antigen priming in the secondary lymphoid organs,
CD8+ T lymphocytes are recruited to sites of infection in theperipheral tissues. Previous reports indicate that the intensity ofneutrophilic infiltration directly correlates with T-cell recruitment,36
implicating neutrophils as a major source of T-cell chemokines10 or asmediators of chemoattractant release from neighbouring cells. Here,we did not observe any impact of neutropenia on CD8+ T-cellmigration to influenza-infected lung tissue. In vitro activated OT-I Tcells infiltrated the neutropenic lung of virus-infected mice, as didinfluenza-primed endogenous CD8+ T cells. Furthermore, CD8+ Tcells primed in the absence of neutrophils did not display impairedmigration patterns and readily accessed the respiratory airways.Therefore, in the absence of neutrophils, there are sufficient chemo-tactic signals elicited in response to infection that enable CD8+ T-cellrecruitment to the influenza-infected respiratory tract.A major impact of neutrophil depletion during early influenza
infection was the significant reduction in virus-specific CD8+ T cells inthe respiratory airways (lung and BAL). Therefore, despite neutrophils
having little impact on CD8+ T-cell priming and recruitment, theirabsence dampened the overall virus-specific CD8+ T-cell responsedirectly at the site of infection. This suggests that although the earlyabsence of neutrophils does not alter CD8+ T-cell traffic into theinfected respiratory airways, it impairs the establishment of a sus-tained CD8+ T-cell population at this site. In the absence of an earlyneutrophilic response, virus-specific CD8+ T cells may either transitrapidly through the lung or alternatively establish residence only toexhibit rapid death. Interestingly, not only did early neutropaeniaimpair the magnitude of the CD8+ T-cell response at the site ofinfection, it greatly diminished the effector function of the remainingCD8+ T cells. Both the capacity to secrete inflammatory cytokines,together with expression of cytolytic effector molecule grzB, werereduced, indicative of a much dampened influenza-specific CD8+
T-cell response. Consequently, an early neutrophil response to viralinfection has outcomes that later impact the development of robustadaptive immunity. Early neutrophil-mediated liberation of antigenfrom the infected pulmonary tissue, cytokine or chemokine releaseand/or damage to infected tissue may be mechanisms of neutrophilactivity that ultimately shape the infected pulmonary tissue, so thatlater in the response, the tissue environment no longer maintains orpromotes an effective cytotoxic virus-specific CD8+ T-cell responseonce these cells have arrived in the infected lesion. Neutrophils mayalso act as local antigen-presenting cells at the inflammatory site,30–32
providing amplification of the T-cell response by stimulating localCD8+ T-cell division and cytotoxic effector acquisition. Finally, theimpact of neutrophils on developing CD8+ T-cell immunity may bemediated by other cell types present in the lung including alveolarmacrophages.37 Elucidating the specific mechanisms involved willform the basis of future studies.In conclusion, we describe here a defined model for dissecting the
role of neutrophils in the adaptive CD8+ T-cell response duringinfluenza infection. Surprisingly, the early events of antigen presenta-tion and CD8+ T-cell priming were unaltered in the absence ofneutrophils. In contrast, neutrophil depletion significantly impairedthe magnitude and cytotoxic capacity of virus-specific CD8+ T cells inthe infected pulmonary airways. This implicates an important role forneutrophils in sustaining large numbers of cytotoxic effector CD8+ Tcells at sites of infection. Therefore, this study provides furthersupport for the concept of innate and adaptive immune responseinterplay during immunity to infection.
METHODSMice and virusesC57BL/6 (B6) mice were bred and housed in specific pathogen-free conditionsat the Department of Microbiology and Immunology, University of Melbourne,Australia. Male mice that were 6- to 8-weeks-old were used in all experiments.The influenza A virus strain used in this study was HKx31 (H3N2), a high-yielding reassortant of A/PR/8/34 (PR8; H1N1) with the H3N2 virus A/Aichi/2/68. HKx31 bears the surface glycoproteins of the H3N2 virus strain. Virus wasobtained from World Health Organization Collaborating Centre for Referenceand Research on Influenza, Melbourne, Australia. Viruses were grown in 10-dayembryonated hen’s eggs by standard procedures and titrated on Madin–Darbycanine kidney cells as described.22
Infection and treatment of miceMice were anaesthetised and infected with 102 or 105 PFU of HKx31 via theintranasal route in 50ml of phosphate-buffered saline. Mice were weighed daily.Animals that lost X25% of their original body weight and/or displayedevidence of pneumonia were killed. All research complied with the Universityof Melbourne’s Animal Experimentation Ethics guidelines and policies.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
7
Immunology and Cell Biology

To determine virus titres, lungs and nasal tissues were removed from virus-infected mice, homogenised in 2ml of phosphate-buffered saline and clarifiedby centrifugation. Titres of infectious virus in tissue homogenates weredetermined by standard plaque assay on Madin–Darby canine kidney cells inthe presence of trypsin.22
For depletion of neutrophils in vivo, purified anti-Ly6G rat mAb (1A8, a giftfrom Professor Thomas Malek, Department of Microbiology and Immunology,University of Miami, Florida, USA) was administered to mice. Antibody waspurified from hybridoma supernatants via a protein G column (GE Healthcare,Buckinghamshire, UK). A combination of intraperitoneal (i.p.; 0.5mg in0.2ml) and intranasal (0.2mg in 0.05ml) routes were administered to obtain490% neutrophil depletion in the blood and to maintain 480% neutrophildepletion in the airways during influenza virus infection.3 Mice were treated24 h before the infection and every 48 h thereafter. Control animals received asimilar dose of purified whole-rat IgG (Jackson Laboratories, Sacramento, CA,USA). Depletion of neutrophils in the blood and airways was confirmed bydifferential leukocyte counts, as previously described.2
Recovery of leukocytes from miceBAL cells were obtained by flushing lungs of killed mice three times with 1mlof phosphate-buffered saline through a blunted 23-guage needle inserted intothe trachea. Lungs were removed, finely minced with scissors and incubated for30min at 37 1C with 2mgml!1 collagenase A (Roche Diagnostics, Mannheim,Germany) in serum-free RMPI 1640 medium. Single-cell suspensions of thelung, MLN and spleen were prepared by passing samples through a cell strainer(BD Biosciences, San Jose, CA, USA). Samples were treated with Tris–NH4Cl(0.14M NH4Cl in 17mM Tris, adjusted to pH 7.2) to lyse erythrocytes andwashed in RPMI 1640 medium supplemented with 10% fetal calf serum (RF10).Cell number and viability were assessed via trypan blue exclusion.
Flow cytometryFor flow cytometry analysis, single-cell suspensions prepared from BAL, lung,MLN and spleen were incubated on ice for 20min with supernatants fromhybridoma 2.4G2 to block Fc receptors and then stained with appropriatecombinations of fluorescein isothiocyanate (FITC), phycoerythrin (PE),allophycocyanin (APC) or biotinylated monoclonal antibodies to Ly6G(1A8), Gr-1 (RB6-8C5), Ly6C (AL-21), CD45.2 (104), CD8a (53–6.7),CD11c (HL3), CD11b (M1/70), CD40 (FGK45.5), CD45.1 (A20), CD80(16-10A1), CD86 (GL1) and I-Ab (AF6-120.1). In some experiments, cellswere stained with PE or APC-labelled DbPA224 (acid polymerase; SSLEN-FRAYV) or DbNP366 (nucleoprotein; ASNENMETM)-specific MHC-I tetra-mers (a gift from Laureate Professor Peter Doherty, Department ofMicrobiology and Immunology, The University of Melbourne) for 60min atroom temperature before staining with anti-CD8. Live cells were analysed bythe addition of propidium iodide (PI; 10mg/ml) to each sample, and cells wereanalysed on a BD FACSCalibur flow cytometer (BD Biosciences). A minimumof 50 000 viable cells (PI!) were collected for each sample.
Analysis of lymph node and lung dendritic cell populationsDC were isolated as previously described.23 To isolate LN DC populations,MLN were digested with collagenase (type II, Worthington Biochemical,Freehold, NJ, USA) and DNase I (Boehringer Mannheim, Mannheim,Germany), and light density cells selected by centrifugation in 1.082 g cm!3
Nycodenz medium (Nycomed, Oslo, Norway). DCs were identified asCD11chigh by flow cytometry.
For analysis of lung DC, perfused whole lungs were digested with colla-genase and DNase I similar to MLN. Lung suspensions were incubated withthe following purified rat antibodies: anti-CD3 (KT3), anti-Thy1 (T24/31.7),anti-CD19 (ID3), anti-Ly6G (1A8) and anti-erythrocyte (TER-119). DCswere enriched by negative selection by removing antibody-bound cells withgoat anti-rat Ig-coupled magnetic beads (BioMag, Qiagen, Valencia, CA, USA).DCs were positively selected via staining with PE-conjugated anti-CD11cantibody (N418), followed by anti-PE magnetic beads (Miltenyi Biotec,Bergisch Gladbach, Germany). CD11c+ DCs were isolated using a MiltenyiautoMACS magnetic cell sorter (Miltenyi Biotec). Lung DCs were identifiedas CD11c+ CD11b+ by flow cytometry.
Assessment of influenza antigen presentation in vivoMice were infected with 104 PFU of an engineered HKx31 virus expressing theOVA257!264 peptide within the NA stalk (HKx31-OVA) of the influenza virus.24
Naı̈ve OT-I cells were harvested from the lymph nodes of donor Ly5.1 OT-Imice.25 Lymphocytes were stained with CFSE (5mM), as previously described.26
2#106 CFSE-labelled OT-I cells were injected intravenously (i.v., in 200ml) intoB6 Ly5.2 recipients 24h post-infection. MLN were harvested 60 hrs post-OT-Itransfer and CD8+ Ly5.1+ OT-I cells were analysed for their CFSE florescence byflow cytometry.
Assessment of CD8+ T-cell trafficking in vivo5#107 transgenic splenocytes from OT-I/Ly5.1 mice were cultured for 4 days,with 5#107 C57BL/6 splenocytes pulsed with 0.1mg of OVA257!264 peptide,in RF10 supplemented with 6mg lipopolysaccharide (Escherichia coli 0111:B4;Sigma-Aldrich, St Louis, MO, USA). After 48h, splenocytes were cultured inthe presence of 10Uml!1 interleukin-2. On day 4, cells were collected and 107
CD8+ T cells were transferred into recipient mice via i.v. injection.In some experiments, Ly5.1+ CD8+ T cells in the MLN and spleen of HKx31-
infected mice at day 7 following infection were enriched by incubation with thefollowing purified antibodies: anti-CD11b (M1/70), F4/80, anti-erythrocyte(Ter-119), anti-Gr-1 (RB6-8C5), anti-MHC class II (M5/114) and anti-CD4(GK1.5), followed by incubation with anti-rat IgG-coupled magnetic beads(Qiagen). CD8+ T cells were enriched by negative selection by removingantibody-bound cells with goat anti-rat Ig-coupled magnetic beads. T-cellpreparations were 90–95% pure, as determined by flow cytometry. Ly5.1+
CD8+ T cells were transferred into Ly5.2+-recipient mice via i.v. injection.
Intracellular cytokine staining of CD8+ T cellsIntracellular cytokine staining was performed, as previously described.27 BAL orlung cells were incubated for 5 h with or without 1mM of PA224 or NP366peptides in 200ml of RPMI 1640 medium supplemented with 25Uml!1
recombinant human IL-2 and Golgi-Plug (BD Biosciences/BD Pharmingen).Cells were stained with anti-CD8a PerCP-Cy5.5 antibody (BD Pharmingen).Cells were fixed with 2% paraformaldehyde, permeabilised in 0.2% saponinand stained with anti-IFNg-FITC (XMG1.2; BD Pharmingen) and anti-TNFa-APC (MP6-XT22; BD Pharmingen). CD8+ T cells were analysed by flowcytometry.
Intracellular granzyme B staining of CD8+ T cellsIntracellular granzyme B staining was performed, as previously described.28
Lymphocyte preparations were fixed with 2% paraformaldehyde, permeabilisedin 0.2% saponin and stained with anti-granzyme B-APC (MHGB05; Caltag,Buckingham, UK). CD8+ T cells were analysed by flow cytometry.
Statistical analysisFor the comparison of two sets of values, a Student’s t-test (two-tailed,two-sample equal variance) was used. When comparing three or more sets ofvalues, a one-way analysis of variance test was applied. A P-value ofp0.05 wasconsidered statistically significant.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
ACKNOWLEDGEMENTSThis study was supported by Project Grants 509230 and 508905 from TheNational Health and Medical Research Council (NHMRC) of Australia. JDM isan NHMRC CDA Fellow. PCR is an NHMRC RD Wright Research Fellow.The Melbourne WHO Collaborating Centre for Reference and Research onInfluenza is supported by the Australian Government Department of Healthand Ageing.
1 Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol2006; 6: 173–182.
2 Tate MD, Brooks AG, Reading PC. The role of neutrophils in the upper and lowerrespiratory tract during influenza virus infection of mice. Respir Res 2008; 9: 57.
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
8
Immunology and Cell Biology

3 Tate MD, Deng YM, Jones JE, Anderson GP, Brooks AG, Reading PC. Neutrophilsameliorate lung injury and the development of severe disease during influenza infection.J Immunol 2009; 183: 7441–7450.
4 Fujisawa H. Neutrophils play an essential role in cooperation with antibody in bothprotection against and recovery from pulmonary infection with influenza virus in mice.J Virol 2008; 82: 2772–2783.
5 Fujisawa H. Inhibitory role of neutrophils on influenza virus multiplication in the lungsof mice. Microbiol Immunol 2001; 45: 679–688.
6 Bennouna S, Bliss SK, Curiel TJ, Denkers EY. Cross-talk in the innate immune system:neutrophils instruct recruitment and activation of dendritic cells during microbialinfection. J Immunol 2003; 171: 6052–6058.
7 Megiovanni AM, Sanchez F, Robledo-Sarmiento M, Morel C, Gluckman JC, Boudaly S.Polymorphonuclear neutrophils deliver activation signals and antigenic molecules todendritic cells: a new link between leukocytes upstream of T lymphocytes. J LeukocBiol 2006; 79: 977–988.
8 van Gisbergen KP, Sanchez-Hernandez M, Geijtenbeek TB, van Kooyk Y. Neutrophilsmediate immune modulation of dendritic cells through glycosylation-dependentinteractions between Mac-1 and DC-SIGN. J Exp Med 2005; 201: 1281–1292.
9 Appelberg R. Neutrophils and intracellular pathogens: beyond phagocytosis and killing.Trends Microbiol 2007; 15: 87–92.
10 de Oca RM, Buendia AJ, Del Rio L, Sanchez J, Salinas J, Navarro JA. Polymorpho-nuclear neutrophils are necessary for the recruitment of CD8(+) T cells in the liver in apregnant mouse model of Chlamydophila abortus (Chlamydia psittaci serotype 1)infection. Infect Immun 2000; 68: 1746–1751.
11 Grabie N, Hsieh DT, Buono C, Westrich JR, Allen JA, Pang H et al. Neutrophils sustainpathogenic CD8+ T cell responses in the heart. Am J Pathol 2003; 163: 2413–2420.
12 Tvinnereim AR, Hamilton SE, Harty JT. Neutrophil involvement in cross-priming CD8+T cell responses to bacterial antigens. J Immunol 2004; 173: 1994–2002.
13 Fleming TJ, Fleming ML, Malek TR. Selective expression of Ly-6G on myeloid lineagecells in mouse bone marrow. RB6-8C5mAb to granulocyte-differentiation antigen(Gr-1) detects members of the Ly-6 family. J Immunol 1993; 151: 2399–2408.
14 Czuprynski CJ, Brown JF, Maroushek N, Wagner RD, Steinberg H. Administration ofanti-granulocyte mAb RB6-8C5 impairs the resistance of mice to Listeria monocyto-genes infection. J Immunol 1994; 152: 1836–1846.
15 Tumpey TM, Chen SH, Oakes JE, Lausch RN. Neutrophil-mediated suppression of virusreplication after herpes simplex virus type 1 infection of the murine cornea. J Virol1996; 70: 898–904.
16 Matsuzaki J, Tsuji T, Chamoto K, Takeshima T, Sendo F, Nishimura T. Successfulelimination of memory-type CD8+ T cell subsets by the administration of anti-Gr-1monoclonal antibody in vivo. Cell Immunol 2003; 224: 98–105.
17 Daley JM, Thomay AA, Connolly MD, Reichner JS, Albina JE. Use of Ly6G-specificmonoclonal antibody to deplete neutrophils in mice. J Leukoc Biol 2008; 83: 64–70.
18 Wojtasiak M, Pickett DL, Tate MD, Londrigan SL, Bedoui S, Brooks AG et al. Depletionof Gr-1+, but not Ly6G+, immune cells exacerbates virus replication and disease inan intranasal model of herpes simplex virus type 1 infection. J Gen Virol 2010; 91:2158–2166.
19 Wojtasiak M, Pickett DL, Tate MD, Bedoui S, Job ER, Whitney PG et al. Gr-1+ cells, butnot neutrophils, limit virus replication and lesion development following flank infectionof mice with herpes simplex virus type-1. Virology 2010; 407: 143–151.
20 Dunay IR, Fuchs A, Sibley LD. Inflammatory monocytes but not neutrophils arenecessary to control infection with Toxoplasma gondii in mice. Infect Immun 2010;78: 1564–1570.
21 Doherty PC, Turner SJ, Webby RG, Thomas PG. Influenza and the challenge forimmunology. Nat Immunol 2006; 7: 449–455.
22 Anders EM, Hartley CA, Jackson DC. Bovine and mouse serum beta inhibitors ofinfluenza A viruses are mannose-binding lectins. Proc Natl Acad Sci USA 1990; 87:4485–4489.
23 Mintern JD, Bedoui S, Davey GM, Moffat JM, Doherty PC, Turner SJ. Transience of MHCClass I-restricted antigen presentation after influenza A virus infection. Proc Natl AcadSci USA 2009; 106: 6724–6729.
24 Jenkins MR, Webby R, Doherty PC, Turner SJ. Addition of a prominent epitope affectsinfluenza A virus-specific CD8+ T cell immunodominance hierarchies when antigen islimiting. J Immunol 2006; 177: 2917–2925.
25 Clarke SR, Barnden M, Kurts C, Carbone FR, Miller JF, Heath WR. Characterization ofthe ovalbumin-specific TCR transgenic line OT-I: MHC elements for positive andnegative selection. Immunol Cell Biol 2000; 78: 110–117.
26 Mintern J, Li M, Davey GM, Blanas E, Kurts C, Carbone FR et al. The use ofcarboxyfluorescein diacetate succinimidyl ester to determine the site, duration andcell type responsible for antigen presentation in vivo. Immunol Cell Biol 1999; 77:539–543.
27 La Gruta NL, Turner SJ, Doherty PC. Hierarchies in cytokine expression profiles foracute and resolving influenza virus-specific CD8+ T cell responses: correlation ofcytokine profile and TCR avidity. J Immunol 2004; 172: 5553–5560.
28 Moffat JM, Gebhardt T, Doherty PC, Turner SJ, Mintern JD. Granzyme A expressionreveals distinct cytolytic CTL subsets following influenza A virus infection. Eur JImmunol 2009; 39: 1203–1210.
29 Marshall DR, Turner SJ, Belz GT, Wingo S, Andreansky S, Sangster MY et al.Measuringthe diaspora for virus-specific CD8+ T cells. Proc Natl Acad Sci USA 2001; 98:6313–6318.
30 Culshaw S, Millington OR, Brewer JM, McInnes IB. Murine neutrophils present Class IIrestricted antigen. Immunol Lett 2008; 118: 49–54.
31 Oehler L, Majdic O, Pickl WF, Stockl J, Riedl E, Drach J et al. Neutrophil granulocyte-committed cells can be driven to acquire dendritic cell characteristics. J Exp Med1998; 187: 1019–1028.
32 Sandilands GP, Ahmed Z, Perry N, Davison M, Lupton A, Young B. Cross-linking ofneutrophil CD11b results in rapid cell surface expression of molecules required forantigen presentation and T-cell activation. Immunology 2005; 114: 354–368.
33 Beauvillain C, Delneste Y, Scotet M, Peres A, Gascan H, Guermonprez P et al.Neutrophils efficiently cross-prime naive T cells in vivo. Blood 2007; 110:2965–2973.
34 Wittamer V, Bondue B, Guillabert A, Vassart G, Parmentier M, Communi D. Neutrophil-mediated maturation of chemerin: a link between innate and adaptive immunity.J Immunol 2005; 175: 487–493.
35 Charmoy M, Brunner-Agten S, Aebischer D, Auderset F, Launois P, Milon G et al.Neutrophil-derived CCL3 is essential for the rapid recruitment of dendritic cells to thesite of Leishmania major inoculation in resistant mice. PLoS Pathog 2010; 6:e1000755.
36 Engeman T, Gorbachev AV, Kish DD, Fairchild RL. The intensity of neutrophil infiltrationcontrols the number of antigen-primed CD8 T cells recruited into cutaneous antigenchallenge sites. J Leukoc Biol 2004; 76: 941–949.
37 Fujisawa H, Tsuru S, Taniguchi M, Zinnaka Y, Nomoto K. Protective mechanismsagainst pulmonary infection with influenza virus. I. Relative contribution of polymor-phonuclear leukocytes and of alveolar macrophages to protection during the early phaseof intranasal infection. J Gen Virol 1987; 68(Part 2): 425–432.
Supplementary Information accompanies the paper on Immunology and Cell Biology website (http://www.nature.com/icb)
Neutrophils modulate CD8+ T cells during influenzaMD Tate et al
9
Immunology and Cell Biology
Related Documents











