Neurotrophic factors acutely alter the sorting of the vesicular acetyl choline transporter and the vesicular monoamine transporter 2 in bimodal sympathetic neurons Edward Felder and Georg Dechant ⁎ Institute for Neuroscience, Innsbruck Medical University, MZA, Anichstr. 35, 6020 Innsbruck, Austria Received 22 February 2006; revised 31 August 2006; accepted 7 September 2006 Available online 23 October 2006 Individual sympathetic neurons in co-cultures with cardiac myocytes store acetylcholine and noradrenaline in two different populations of synaptic vesicles and release both neurotransmitters from single presynaptic terminals. Neurotrophic factors selectively and acutely stimulate differential release of the two types of neurotransmitters from these bimodal neurons. Here we investigated the acute effects of neurotrophic factors on two pivotal marker proteins for catecholami- nergic and cholinergic synaptic vesicle populations: the vesicular monoamine transporter 2 and the vesicular acetylcholine transporter. We observed that separation of the two fluorescence labeled transporters is not restricted to the varicosities, but can also be observed in the neurites as well as in the cell soma. Application of nerve growth factor, brain-derived neurotrophic factor and ciliary neuronotrophic factor caused acute alterations in transporter segregation. These results point to a novel function of neurotrophic factors during the short-term regulation of synaptic protein sorting in neurons. © 2006 Elsevier Inc. All rights reserved. Introduction Developing sympathetic neurons in mammals and birds have the remarkable potential to switch in vivo from noradrenergic to cholinergic neurotransmission passing through a transitory bimodal state. While in vivo this neurotransmitter phenotype switch occurs during early postnatal stages and is completed within a period of days, sympathetic neurons in co-cultures with cardiac myocytes can maintain a dual neurotransmitter status for prolonged periods of time (Furshpan et al., 1986a,b; Potter et al., 1986; Matsumoto et al., 1987; Landis, 1976; Johnson et al., 1980a). In these co-cultures individual neurons synthesize and release acetylcholine (ACh) as well as noradrenaline (NA). Electron microscopic analyses revealed the presence of two different types of small synaptic vesicles. With the help of electrophysiological and biochemical methods, it could be established that the proportion of released NA correlates with the amount of small granular vesicles, whereas choline-acetyl transferase activity as well as ACh release from varicosities correlate with the proportion of small clear vesicles (Furshpan et al., 1986a; Landis, 1976; Johnson et al., 1980b). Taken together, these findings demonstrate that NA and ACh are separated in the two different populations of synaptic vesicles. Therefore, bimodal sympathetic neurons provide a unique neuronal model to study the presynaptic neurotransmitter release of two different classic neurotransmitters from two types of vesicles present within the same, individual presynaptic terminal. Neurotrophic factors exert strong, fast and specific functions on synapses in the central and peripheral nervous system and sympathetic neurons are in particular sensitive for neurotrophic factor stimulation. Recent studies revealed that bimodal sympathetic neurons acutely change their neurotransmitter release in response to nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and, under certain conditions, also ciliary neuronotrophic factor (CNTF) (Lockhart et al., 1997; Yang et al., 2002; Slonimsky et al., 2002). Neurons that underwent a 10 to 15 min treatment with NGF prior to electrical stimulation preferentially release NA, whereas BDNF treatment induces a shift to ACh release within the same period of time. In order to exert these acute effects on neuro- transmitter release, NGF and BDNF employ two different types of signaling receptors. The NGF effect is mediated via binding to the receptor tyrosine kinase TrkA. In contrast, the effect of BDNF requires the neurotrophin receptor p75 NTR and can be potentiated by overexpression of p75 NTR (Lockhart et al., 1997; Yang et al., 2002; Slonimsky et al., 2002). At present it is not clear by which cellular mechanism NGF and BDNF modify sympathetic transmitter release. A selective mobilization of one of the two synaptic vesicle populations, thereby retaining the other population in the presynaptic terminal, is one likely explanation. But selective transport of vesicular proteins to the synapse or effects on vesicle internalization in the presynapse during repeated cycles of exo- and endocytosis may also account for the observed neurotrophic factor effects. www.elsevier.com/locate/ymcne Mol. Cell. Neurosci. 34 (2007) 1 – 9 ⁎ Corresponding author. Fax: +43 512 504 67 27390. E-mail address: [email protected] (G. Dechant). Available online on ScienceDirect (www.sciencedirect.com). 1044-7431/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.mcn.2006.09.005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 34 (2007) 1–9Neurotrophic factors acutely alter the sorting of the vesicular acetylcholine transporter and the vesicular monoamine transporter 2 inbimodal sympathetic neurons
Edward Felder and Georg Dechant⁎
Institute for Neuroscience, Innsbruck Medical University, MZA, Anichstr. 35, 6020 Innsbruck, Austria
Received 22 February 2006; revised 31 August 2006; accepted 7 September 2006Available online 23 October 2006
Individual sympathetic neurons in co-cultures with cardiac myocytesstore acetylcholine and noradrenaline in two different populations ofsynaptic vesicles and release both neurotransmitters from singlepresynaptic terminals. Neurotrophic factors selectively and acutelystimulate differential release of the two types of neurotransmitters fromthese bimodal neurons. Here we investigated the acute effects ofneurotrophic factors on two pivotal marker proteins for catecholami-nergic and cholinergic synaptic vesicle populations: the vesicularmonoamine transporter 2 and the vesicular acetylcholine transporter.We observed that separation of the two fluorescence labeled transportersis not restricted to the varicosities, but can also be observed in theneurites as well as in the cell soma. Application of nerve growth factor,brain-derived neurotrophic factor and ciliary neuronotrophic factorcaused acute alterations in transporter segregation. These results pointto a novel function of neurotrophic factors during the short-termregulation of synaptic protein sorting in neurons.© 2006 Elsevier Inc. All rights reserved.
Introduction
Developing sympathetic neurons in mammals and birds havethe remarkable potential to switch in vivo from noradrenergic tocholinergic neurotransmission passing through a transitory bimodalstate. While in vivo this neurotransmitter phenotype switch occursduring early postnatal stages and is completed within a period ofdays, sympathetic neurons in co-cultures with cardiac myocytescan maintain a dual neurotransmitter status for prolonged periodsof time (Furshpan et al., 1986a,b; Potter et al., 1986; Matsumoto etal., 1987; Landis, 1976; Johnson et al., 1980a). In these co-culturesindividual neurons synthesize and release acetylcholine (ACh) aswell as noradrenaline (NA). Electron microscopic analysesrevealed the presence of two different types of small synapticvesicles. With the help of electrophysiological and biochemical
⁎ Corresponding author. Fax: +43 512 504 67 27390.E-mail address: [email protected] (G. Dechant).Available online on ScienceDirect (www.sciencedirect.com).
1044-7431/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.mcn.2006.09.005
methods, it could be established that the proportion of released NAcorrelates with the amount of small granular vesicles, whereascholine-acetyl transferase activity as well as ACh release fromvaricosities correlate with the proportion of small clear vesicles(Furshpan et al., 1986a; Landis, 1976; Johnson et al., 1980b).Taken together, these findings demonstrate that NA and ACh areseparated in the two different populations of synaptic vesicles.Therefore, bimodal sympathetic neurons provide a unique neuronalmodel to study the presynaptic neurotransmitter release of twodifferent classic neurotransmitters from two types of vesiclespresent within the same, individual presynaptic terminal.
Neurotrophic factors exert strong, fast and specific functions onsynapses in the central and peripheral nervous system andsympathetic neurons are in particular sensitive for neurotrophicfactor stimulation. Recent studies revealed that bimodal sympatheticneurons acutely change their neurotransmitter release in response tonerve growth factor (NGF), brain-derived neurotrophic factor(BDNF) and, under certain conditions, also ciliary neuronotrophicfactor (CNTF) (Lockhart et al., 1997; Yang et al., 2002; Slonimsky etal., 2002). Neurons that underwent a 10 to 15 min treatment withNGF prior to electrical stimulation preferentially release NA,whereas BDNF treatment induces a shift to ACh release within thesame period of time. In order to exert these acute effects on neuro-transmitter release, NGF and BDNF employ two different types ofsignaling receptors. The NGF effect is mediated via binding to thereceptor tyrosine kinase TrkA. In contrast, the effect of BDNFrequires the neurotrophin receptor p75NTR and can be potentiated byoverexpression of p75NTR (Lockhart et al., 1997; Yang et al., 2002;Slonimsky et al., 2002).
At present it is not clear by which cellular mechanism NGF andBDNF modify sympathetic transmitter release. A selectivemobilization of one of the two synaptic vesicle populations,thereby retaining the other population in the presynaptic terminal,is one likely explanation. But selective transport of vesicularproteins to the synapse or effects on vesicle internalization in thepresynapse during repeated cycles of exo- and endocytosis mayalso account for the observed neurotrophic factor effects.

2 E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
In order to analyze the mechanisms of neurotrophic factor-induced differential neurotransmitter release, we set out to studydynamic changes in the distribution of NA- and ACh-specificsynaptic vesicle markers in bimodal sympathetic neurons byconfocal fluorescence microscopy. For this approach we chose themost characteristic proteins of the two synaptic vesicle populationsin sympathetic neurons, the vesicular monoamine transporter 2(VMAT2) and the vesicular acetylcholine transporter (VAChT).
Results
Neurons in co-cultures with cardiomyocytes were double-transfected with constructs for rat VAChT-pEGFP (FP-vector:pEGFP-N2, Clonetech, Palo Alto) and VMAT2-pDsRed1 (FP-vector: pDsRed1-N1, Clonetech, Palo Alto). Both vectors werekindly provided by M. Takahashi and co-workers (Shoju-Kasai etal., 2002). In these cultures neurons were stained for the synapticmarker protein synapsin I (Figs. 1A and B) to identify the axonalswellings as varicosities. Neurons, double-transfected with fluor-escence-tagged VMAT2 and VAChT had a healthy morphologicalappearance with long neurites. They contained accumulations of thetwo FP-tagged vesicular transporters (Fig. 1C) in the varicosities.
Intracellular separation of the two proteins was determined bycalculating a colocalization index (CI), representing the ratio of thearea of overlapping red/green fluorescence and the total fluores-cence area of each of the two single fluorescence colors. Anincrease of red and green fluorescence separation is reflected by anincrease of the CI, whereby a CI value of 1 would reflect atheoretical perfect colocalization. To determine any unspecificseparation of the fluorescence signal (e.g., due to FP aggregation ordifferent expression levels of the two proteins), CIs after VAChT-pEGFP and VMAT2-pDsRed1 transfection were compared to adouble transfection with constructs for VMAT2-pEGFP (also fromM. Takahashi) and VMAT2-pDsRed1, further referred to as“control transfection”. In this control transfection the two
Fig. 1. Characterization of co-cultures. The overlays of immunostainings for the prneurons form synaptic contacts with cardiac myocytes (A) and other neurons (B).phase contrast image. Both fluorescent proteins are present in the varicosities of a
transfected proteins differ only in their fluorescence tags andtherefore a CI higher than 1 reflects a non-specific separation thatis independent of any protein-specific sorting or targetingmechanism. In contrast, any additional separation of the fluores-cence colors (increase of the CI) in the VAChT-pEGFP andVMAT2-pDsRed1 transfection is explained by the intrinsictargeting properties of VAChT or VMAT2. The significantly lowerCI-values of the control transfection (see below) also demonstratedthat double transfection leads to comparable expression levels ofthe two proteins used. In the soma of the cell the CI (VAChT-GFP/VMAT2-DsRed and control transfection) was always determinedby calculating the mean CI from 3 different confocal sections of thesame cell.
Fig. 2 compares varicosities of VAChT-GFP/VMAT2-DsReddouble-transfected sympathetic neurons with neurons that under-went control transfection. The frequency histograms of colocaliza-tion index (CI) values (Figs. 2A and D) from single varicositiesdemonstrate that CI values from VAChT-GFP/VMAT2-DsRedtransfections (n=34; avg=1.79; SD=0.52) are significantly(p<0.001) different compared to the CI values determined aftercontrol transfections (n=38; avg=1.14; SD=0.14). Fig. 2Brepresents a 1-μm confocal section through varicosities of aVAChT-GFP/VMAT2-DsRed-transfected cell and is compared tovaricosities after a control transfection (Fig. 2E). The confocalsections demonstrate the stronger separation of the fluorescencecolors in Fig. 2B in comparison to the fluorescent structures inpanel 2E. Figs. 2C and F depict 3D reconstructions of the uppervaricosities from Figs. 2B and E. The two fluorescence colors aredigitally separated from each other in the 3D reconstruction, whichreveals a distinctly different shape of the fluorescence intensitythreshold rendered volumes in the VAChT-GFP/VMAT2-DsRed-transfected varicosities (Fig. 2C). In contrast, almost identicallyshaped red and green structures are visible in panel 2F. Thesefindings show that the use of fluorescence-tagged VAChT andVMAT2 is an appropriate methodological approach to detect the
esynaptic marker protein synapsin I and phase contrast images show that thePanel C shows an overlay of VAChT-GFP/VMAT2-DsRed signals with thedouble-transfected neuron.
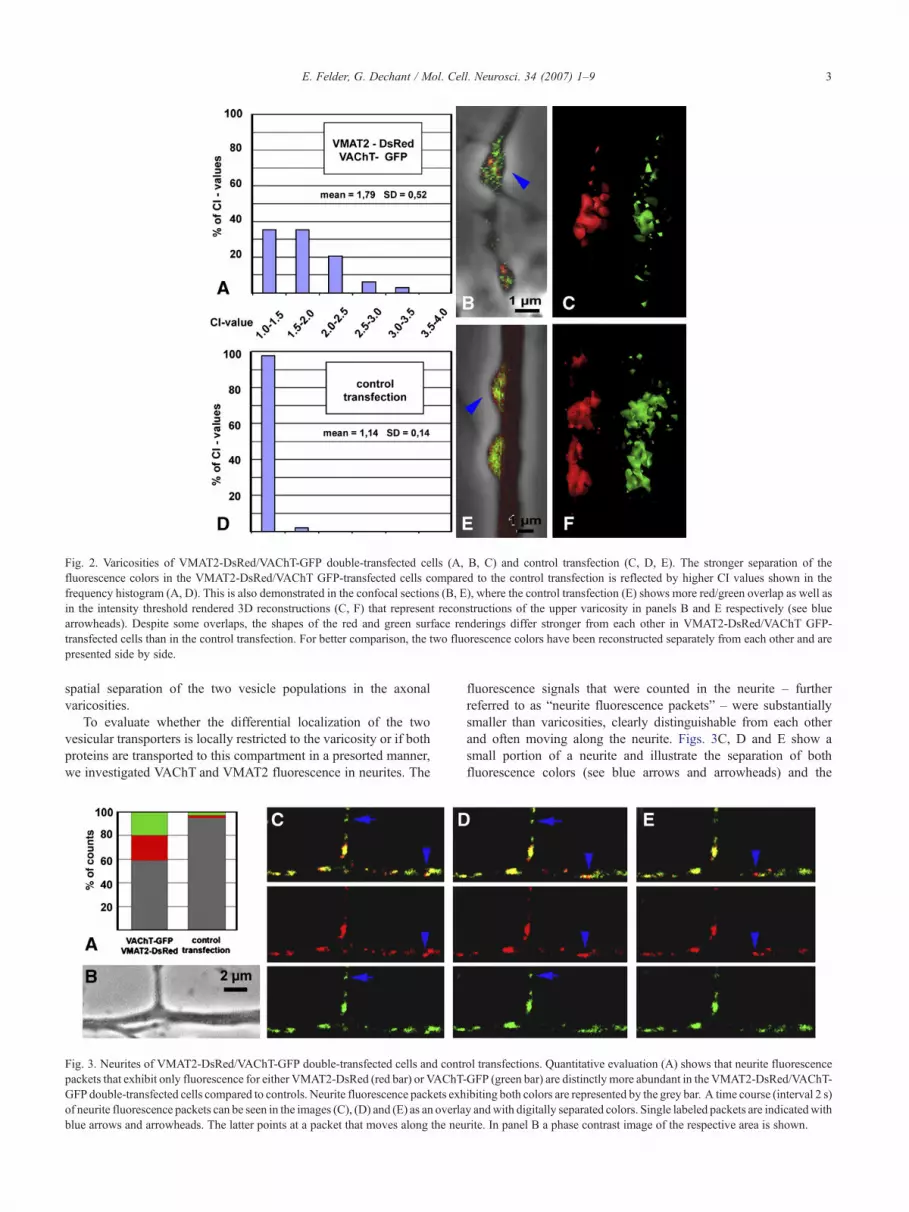
Fig. 2. Varicosities of VMAT2-DsRed/VAChT-GFP double-transfected cells (A, B, C) and control transfection (C, D, E). The stronger separation of thefluorescence colors in the VMAT2-DsRed/VAChT GFP-transfected cells compared to the control transfection is reflected by higher CI values shown in thefrequency histogram (A, D). This is also demonstrated in the confocal sections (B, E), where the control transfection (E) shows more red/green overlap as well asin the intensity threshold rendered 3D reconstructions (C, F) that represent reconstructions of the upper varicosity in panels B and E respectively (see bluearrowheads). Despite some overlaps, the shapes of the red and green surface renderings differ stronger from each other in VMAT2-DsRed/VAChT GFP-transfected cells than in the control transfection. For better comparison, the two fluorescence colors have been reconstructed separately from each other and arepresented side by side.
3E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
spatial separation of the two vesicle populations in the axonalvaricosities.
To evaluate whether the differential localization of the twovesicular transporters is locally restricted to the varicosity or if bothproteins are transported to this compartment in a presorted manner,we investigated VAChT and VMAT2 fluorescence in neurites. The
Fig. 3. Neurites of VMAT2-DsRed/VAChT-GFP double-transfected cells and contrpackets that exhibit only fluorescence for either VMAT2-DsRed (red bar) or VAChT-GFP double-transfected cells compared to controls. Neurite fluorescence packets exhof neurite fluorescence packets can be seen in the images (C), (D) and (E) as an overlablue arrows and arrowheads. The latter points at a packet that moves along the neu
fluorescence signals that were counted in the neurite – furtherreferred to as “neurite fluorescence packets” – were substantiallysmaller than varicosities, clearly distinguishable from each otherand often moving along the neurite. Figs. 3C, D and E show asmall portion of a neurite and illustrate the separation of bothfluorescence colors (see blue arrows and arrowheads) and the
ol transfections. Quantitative evaluation (A) shows that neurite fluorescenceGFP (green bar) are distinctly more abundant in the VMAT2-DsRed/VAChT-ibiting both colors are represented by the grey bar. A time course (interval 2 s)y andwith digitally separated colors. Single labeled packets are indicatedwithrite. In panel B a phase contrast image of the respective area is shown.

4 E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
movement of a neurite fluorescence packet (see arrowheads only)in the time course series from (C) to (E). The counts of thesefluorescence signals were again compared to counts of controltransfections. Fig. 3A shows that only in the VAChT-GFP/VMAT2-DsRed-transfected cells a considerably high number(approximately 40%) of neurite fluorescence packets of singlecolor could be observed, whereas the latter case practically lacksseparation of fluorescence colors. Thus at least a substantialfraction of the two vesicular neurotransmitter transporters reachesthe varicosity in different cargo containers.
In Fig. 4 we demonstrate the existence of a sorting mechanismin the cell soma of the neuron by analyzing the fluorescence signalin this part of the transfected neurons. Statistical analysis of CIs ofconfocal cross-sections through the entire soma of VAChT-GFP/VMAT2-DsRed-transfected cells (n=58; avg=2.01; SD=0.96)and the CIs of control transfections (n=48; avg=1.43; SD=0.28)revealed a highly significant difference (p<0.001). Again, thedistribution of CIs is shown in a frequency histogram (Figs. 4A andE). The confocal cross-sections illustrate the higher degree offluorescence color separation in the VAChT-GFP/VMAT2-DsRed-transfected cells (Figs. 4G and H), compared to the controltransfection (Figs. 4C and D). The nucleus of both cells is locatedin the right portion of the cell and is void of any fluorescence. Thephenomenon of a bright and voluminous “core” region (visible inthe left half of the cell) was observed for both vesicular transportersin most transfected cells. Transfections with fluorescence plasmidsdesigned for labeling subcellular compartments (data not shown)
Fig. 4. Cell soma in control transfections (A–D) and VMAT2-DsRed/VAChT-GFPDsRed/VAChTGFP-transfected cells and the control transfection is again shown inG; with separated fluorescence colors in panels D and H) demonstrate the distribuVMAT2-DsRed is indicated with a blue arrow in panels G and H.
indicate that these core regions comprise the Golgi and ER regions,whereby the latter is adjacent to the Golgi. In the remaining part ofthe cell, readily distinguishable fluorescence packets are visible, inparticular in peripheral regions of the cell in the vicinity of theplasma membrane.
After having demonstrated that both transporters are sorted inthe soma of the cell we tried to exploit our double labelingapproach to gain information about the VAChT/VMAT2 separationmechanism in the cell soma (Fig. 5). Similar to the analysis in theneurite the term “fluorescence packet” is again used to describe asmall, distinct fluorescence signal (an example is indicated by ablue arrow in Figs. 4G and H). Panel A shows the frequencydistribution of the distances between the cell membrane and singlefluorescence packets in single-transfected neurons. We observedthat both VAChT and VMAT2 fluorescence can be found in theplasma membrane, which is represented by the events comprised inthe 0–100 nm bar. VAChT fluorescence packets (n=82) are moreabundant in a region 200–300 nm distant from the membranecompared to VMAT2 fluorescence packets (n=71), where thenumber of fluorescence packets steadily increases towards themembrane. This might indicate a longer dwell time of a vesicularcompartment adjacent to the plasma membrane involved inVAChT trafficking. Next we analyzed the pH of VAChT-GFPand VMAT2-GFP fluorescent packets (Fig. 5B). In the region closeto the plasma membrane (<1 μm), that was considered for thismeasurement, the fluorescence packets of VAChT-GFP-transfectedcells (n=30) show significantly less colocalization (p<0.001) with
double-transfected cells (E–H). The difference of the CIs between VMAT2-the frequency histogram (A, E). Confocal sections (as overlay in panels C andtion in the cell soma. One instance of a fluorescence packet containing only
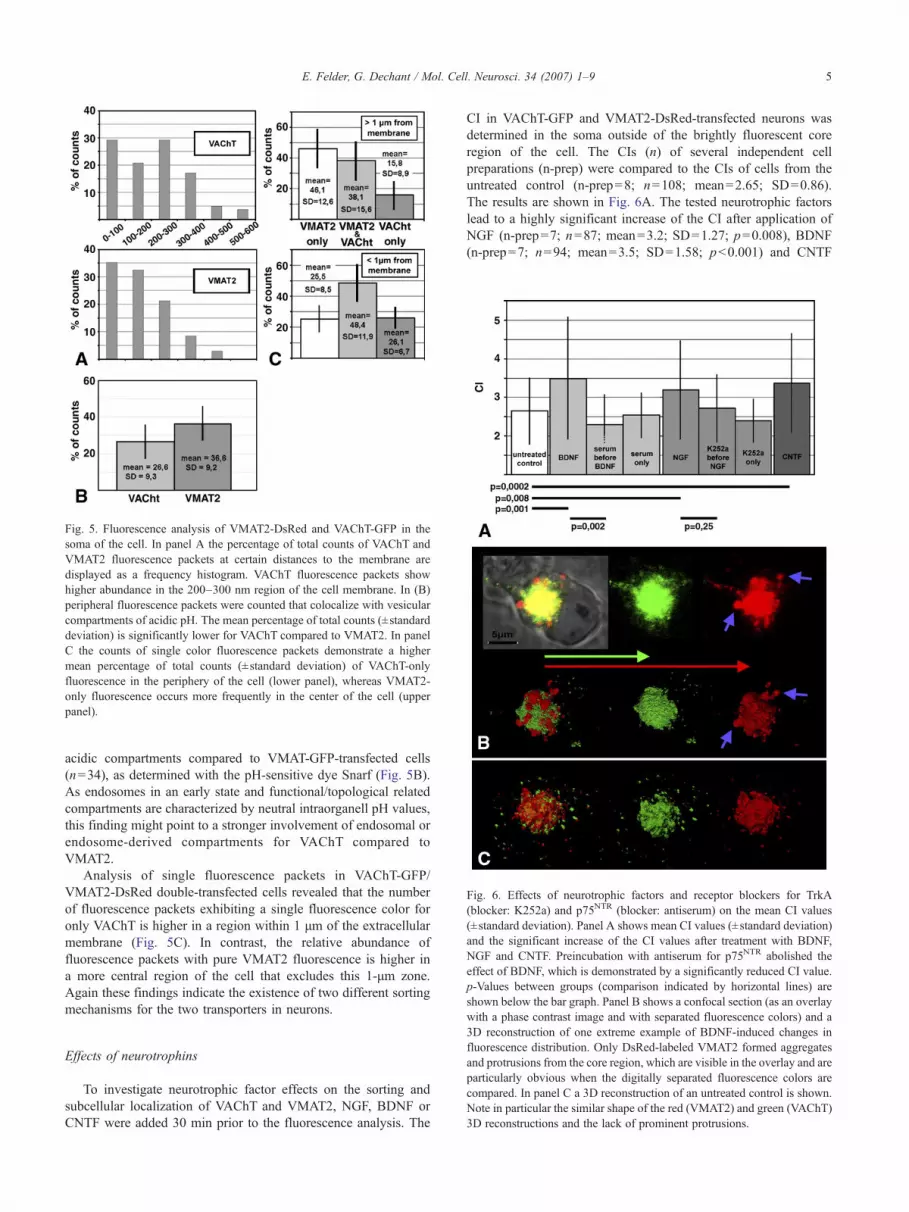
Fig. 5. Fluorescence analysis of VMAT2-DsRed and VAChT-GFP in thesoma of the cell. In panel A the percentage of total counts of VAChT andVMAT2 fluorescence packets at certain distances to the membrane aredisplayed as a frequency histogram. VAChT fluorescence packets showhigher abundance in the 200–300 nm region of the cell membrane. In (B)peripheral fluorescence packets were counted that colocalize with vesicularcompartments of acidic pH. The mean percentage of total counts (±standarddeviation) is significantly lower for VAChT compared to VMAT2. In panelC the counts of single color fluorescence packets demonstrate a highermean percentage of total counts (±standard deviation) of VAChT-onlyfluorescence in the periphery of the cell (lower panel), whereas VMAT2-only fluorescence occurs more frequently in the center of the cell (upperpanel).
Fig. 6. Effects of neurotrophic factors and receptor blockers for TrkA(blocker: K252a) and p75NTR (blocker: antiserum) on the mean CI values(±standard deviation). Panel A shows mean CI values (±standard deviation)and the significant increase of the CI values after treatment with BDNF,NGF and CNTF. Preincubation with antiserum for p75NTR abolished theeffect of BDNF, which is demonstrated by a significantly reduced CI value.p-Values between groups (comparison indicated by horizontal lines) areshown below the bar graph. Panel B shows a confocal section (as an overlaywith a phase contrast image and with separated fluorescence colors) and a
5E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
acidic compartments compared to VMAT-GFP-transfected cells(n=34), as determined with the pH-sensitive dye Snarf (Fig. 5B).As endosomes in an early state and functional/topological relatedcompartments are characterized by neutral intraorganell pH values,this finding might point to a stronger involvement of endosomal orendosome-derived compartments for VAChT compared toVMAT2.
Analysis of single fluorescence packets in VAChT-GFP/VMAT2-DsRed double-transfected cells revealed that the numberof fluorescence packets exhibiting a single fluorescence color foronly VAChT is higher in a region within 1 μm of the extracellularmembrane (Fig. 5C). In contrast, the relative abundance offluorescence packets with pure VMAT2 fluorescence is higher ina more central region of the cell that excludes this 1-μm zone.Again these findings indicate the existence of two different sortingmechanisms for the two transporters in neurons.
3D reconstruction of one extreme example of BDNF-induced changes influorescence distribution. Only DsRed-labeled VMAT2 formed aggregatesand protrusions from the core region, which are visible in the overlay and areparticularly obvious when the digitally separated fluorescence colors arecompared. In panel C a 3D reconstruction of an untreated control is shown.Note in particular the similar shape of the red (VMAT2) and green (VAChT)3D reconstructions and the lack of prominent protrusions.
Effects of neurotrophins
To investigate neurotrophic factor effects on the sorting andsubcellular localization of VAChT and VMAT2, NGF, BDNF orCNTF were added 30 min prior to the fluorescence analysis. The
CI in VAChT-GFP and VMAT2-DsRed-transfected neurons wasdetermined in the soma outside of the brightly fluorescent coreregion of the cell. The CIs (n) of several independent cellpreparations (n-prep) were compared to the CIs of cells from theuntreated control (n-prep=8; n=108; mean=2.65; SD=0.86).The results are shown in Fig. 6A. The tested neurotrophic factorslead to a highly significant increase of the CI after application ofNGF (n-prep=7; n=87; mean=3.2; SD=1.27; p=0.008), BDNF(n-prep=7; n=94; mean=3.5; SD=1.58; p<0.001) and CNTF

6 E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
(n-prep=3; n=49; p=0.001; mean=3.38; SD=1.28). Preincuba-tion with p75NTR antiserum (n-prep=4; n=35; mean=2.3;SD=0.77) reduced the effect of BDNF significantly (p=0.002),whereas K252a (n-prep=4; n=39; mean=2.72; SD=0.88), ablocker of TrkA enzymatic activity, caused only a slight, non-significant reduction of the NGF effect. Control incubationswithout neurotrophic factor but with K252a (n-prep=4; n=23;mean=2.4; SD=0.55) and p75NTR antiserum (n-prep=4; n=39;mean=2.54; SD=0.58) did not result in any significantdifferences compared to the untreated control. Fig. 6B shows aconfocal section of one extreme example of a cell after BDNFtreatment combined with a 3D reconstruction of the same cell.Fig. 6C depicts a 3D reconstruction of an untreated cell. Wecould repeatedly observe that neurotrophic factor treatment (inparticular BDNF and CNTF) led to VMAT2-DsRed fluorescencestructures that protrude from the core region of the cell (see alsoblue arrow in Fig. 4B). No obvious changes were observed forthe green VAChT fluorescence.
Discussion
Our data suggest that bimodal neurons harbor two differentsorting mechanisms for vesicle type specific proteins. Neuro-trophins acutely alter the sorting of vesicular transporters tospecific NA and ACh containing vesicle populations. The effectof BDNF depends on interactions with the p75NTR receptor.
Our study was prompted by the striking finding thatneurotrophic factors can acutely influence the type of neurotrans-mitter released from bimodal sympathetic neurons, where NGFacts through TrkA to facilitate NA release and BDNF mediates itseffects on ACh through p75NTR (Lockhart et al., 1997; Yang et al.,2002). We addressed the question if these neurotrophic factoreffects can be correlated with acute alterations in the spatialdistribution of synaptic vesicular proteins in bimodal sympatheticneurons. We show that separation of the two vesicular transporters– each detectable with a different fluorescence label – occurs in thesoma of the cells. The proteins are then delivered via differentcargo containers in neurites in a presorted manner to the synapse.The degree of separation between VMAT2 and VAChT in the cellsoma significantly increased after NGF, BDNF and CNTF wereacutely applied to the medium. In the case of BDNF and CNTF thisseparation is accompanied by changes in the pattern of thefluorescence VMAT2 distribution. This might reflect a slowingdown of VMAT2 transport or transfer of the protein to anintracellular storage compartment.
The fluorescence analysis of FP-tagged VAChT and VMAT2transporters in neuronal presynaptic structures yielded surprisinglyclear results (Fig. 2). Considering that the vesicles of the adrenergicand cholinergic populations are randomly distributed (as seen inelectron microscopy) and also considering that the distancebetween each vesicle in the varicosity is too small to be resolvedby light microscopy, we presuppose that only a fraction of synapticvesicles was specifically labeled in our experiments. Correcttargeting of VAChT-GFP and VMAT2-DsRed then leads to acertain percentage of red or green labeled vesicles and a remainingfraction of non-labeled vesicles. The latter function as spacersbetween the fluorescent vesicle and enabled us to optically resolvethe labeled vesicles with the two different colors. In contrast, thecontrol transfection leads to individual vesicles containing bothVAChT-GFP and VMAT2-DsRed. Under this condition thedistance between the chromophores is too small to allow separation
with light microscopy, which leads to the observed overlap offluorescence colors and therefore a lower colocalization index.
Differential sorting of VAChT and VMAT2 not only occurs inthe varicosity. The transporters are already spatially segregated inthe soma of the cell and reach the varicosities in a presortedmanner. Although the differences between CI values in the cellsoma are highly significant, we noted that approximately 60% ofthe observed VAChT-GFP/VMAT2-DsRed-transfected cells showa CI between 1 and 2. These CIs are therefore not strikinglydifferent from the values observed in the control transfections. Themost likely explanation for this observation is a heterogeneity inthe bimodal phenotype maturation in our population of primaryneurons. Alternatively, it might also be caused by an unphysio-logical aggregation of fluorescence proteins. The systematiccomparison between the VAChT-GFP/VMAT2-DsRed transfectedand the control transfection was therefore an essential measure todetermine unphysiological separation of fluorescence colors.
Our experiments expand the results of former investigations,according to which both vesicular transporters are separated intodifferent populations of vesicles in PC12 cells (Liu et al., 1994;Erickson et al., 1996a,b; Liu and Edwards, 1997; Varoqui andErickson, 1998; Weihe et al., 1996). These results could not a prioribe extrapolated to neurons, since PC12 are neuroendocrine cellsand contain different types of vesicles compared to sympatheticneurons. Theoretically, selective re-uptake and/or sorting ofVAChT and VMAT2 strictly within the varicosity would sufficeto produce and maintain the two populations of synaptic vesicles.Therefore, the finding of differential VAChT/VMAT2 sorting in thecell soma might contribute to understanding the biogenesis of thetwo populations of synaptic vesicles in bimodal synapses. In bothVMAT and VAChT, the C-terminal (cytoplasmatic) tail of theproteins has been shown to play a crucial role in specific targeting(Varoqui and Erickson, 1998). The sequence KEEKMAIL inVMAT2 and its corresponding sequence in VAChT are a well-characterized signal for internalization. In particular, dipeptidemotifs (LL in VAChT and IL in VMAT) have been shown tomediate internalization in PC12 cells (Tan et al., 1998). Whileinternalization of the vesicular transporters is of fundamentalimportance in the synapse, it is not necessarily involved in thesorting of these molecules outside of this cellular domain. Weobserved that both vesicular transporters are inserted into theplasma membrane of the cell soma, which suggests that subsequentinternalization and endocytotic sorting occurs outside the synapsefor both proteins.
As shown in Fig. 4, differences in the sorting of both proteinsare not restricted to a selective internalization from the plasmamembrane. Despite the fact that both transporters can be found inthe plasma membrane, VAChT fluorescence packets show ahigher abundance in a region adjacent to the membrane (Fig. 5A).This could be interpreted as an early endosomal compartment thatis not (or less) important for the VMAT2 sorting. The idea of twodifferent modes of endosomal sorting for both proteins is furtherstrengthened by the finding that VAChT containing fluorescencepackets in the vicinity of the plasma membrane show moreneutral pH values compared to VMAT2. As recently endocytosedvesicles must again build up their acidic intraluminal pH, thisresult indicates a higher proportion of VAChT in newlyendocytosed vesicles. The quantitative analysis of single labeledfluorescence packets in double-transfected neurons also stands infavor of this hypothesis. VAChT-only fluorescence packets aremore frequent in the very peripheral part of the cell, whereas

7E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
VMAT-only fluorescence is more abundant in the central part ofthe neuron. Especially the latter finding suggests that besidesendosomal sorting another sorting step for VMAT2 might existthat is not associated with the plasma membrane but occurs inother intracellular membraneous compartments, such as the Golgi.Since the double leucine motif mentioned above mediatestrafficking at multiple sites of the secretory pathway, it may alsoinfluence this supposed intracellular VMAT2 sorting step.
For our study of acute affects of neurotrophins we chose thecolocalization index in the soma as the statistically most reliableparameter. We found that the degree of spatial separation of bothvesicular transporters significantly increases within a period of30 min after application of NGF, BDNF or CNTF to the culturemedium. Interestingly, treatment with BDNF and CNTF had aspecific effect on VMAT2, since we frequently observed the acuteformation of VMAT2 fluorescence aggregates in the central part ofthe cell in response to these factors. We found large, sphericalfluorescent VMAT2 containing structures, which were partlyseparated from the supposed Golgi region and partly protrudingfrom it (Fig. 6B). These changes suggest a growth factor-specificmodification of a sorting process in the central part of the neuronfor the VMAT2. Moreover, this finding establishes a correlationbetween the pro-cholinergic effect of BDNF and CNTF and aneffect of these neurotrophic factors on VMAT2.
The acute effects on neurotransmitter release involve bindingto TrkA for NGF and binding to p75NTR for BDNF (Lockhart etal., 1997; Yang et al., 2002). To investigate which receptorsmediate the effect on the sorting of transmitter transporters in ourexperiments, we applied receptor function blocking agents. Forp75NTR we made use of a well-characterized antiserum thatprevents ligand-interactions. The function of TrkA was inhibitedwith K252a, a blocker of tyrosine kinase activity with highspecificity for the Trk receptors. Our experiments indicate animportant function of p75NTR for transporter sorting, since thep75-antiserum eliminated the effect of BDNF, whereas theblocking effect of K252a was not statistically significant. Theprecise signaling mechanisms employed by p75NTR (and possiblyTrkA) in order to acutely alter transporter sorting remain a subjectfor further investigations. A diverse set of proteins is involved invesicle budding and docking, which not only includes manystructural proteins but also a large number of protein kinases.Recently it was demonstrated that p75NTR recruits CaMKII topromote acetylcholine release in bimodal neurons (Slonimsky etal., 2006). It is tempting to speculate that this kinase mightdirectly regulate the phosphorylation status of the two vesicularneurotransmitter transporters. Indeed, the serine at the position −5relative to the LL-motive in VAChT (ser-480) can be phosphory-lated. Mutation of this residue (Krantz et al., 2000) or applicationof phorbol-esther (Shoju-Kasai et al., 2002) leads to alterations intransporter targeting in PC12 cells. For VMAT2, a C-terminalphosphorylation site that can be phosphorylated by casein kinaseI and II (Waites et al., 2001) was identified at positions 512 and514 and casein kinase II is activated by NGF in PC12 cells(Kleijn et al., 1998).
Our data provide evidence for acute effects of neurotrophicfactors on the sorting of synaptic proteins in sympathetic neurons.Experiments that combine electric stimulation of the neuron withsimultaneous monitoring of the two fluorescence-tagged vesicularpopulations within the varicosity will help to further elucidate therelevance of this mechanism for presynaptic transmitter release.Furthermore, it will also be interesting to see whether neurotro-
phins also acutely regulate intracellular sorting of synaptic proteinsin neurons of the central nervous system.
Experimental methods
Cell culture, preparation and transfection
Isolation procedure of cardiac myocytesHearts (ventricle and atrium) were removed from chicken embryos at
embryonic day 11 (E11). The myocard was minced in pieces (0.2–0.5 mm3) and washed with medium HAMTs F-14 (GibcoBRL)supplemented with apotransferrin [100 μg/ml], putrescine [16 μg/ml],progesteron [6 ng/ml] and sodium selenite [8 ng/ml]. The medium alsocontained penicillin [100 mg/l] and streptomycin [60 mg/l] and wasbuffered with 5 mM HEPES buffer (all chemicals were from Sigma unlessstated otherwise). For cell isolation the tissue was incubated in 10 mlmedium with collagenase [1 mg/ml] (Worthington) for 1 h at 37°C, thencarefully triturated with a fire-polished glass pipette and centrifuged at72×g for 3 min in a 50 ml tube. 2×105 cells were plated in a volume of0.5 ml on the glass part of glass bottom dishes (Mattek, P50G-1,5-14-F).The dishes had been coated with poly-DL-ornithine [0.5 mg/ml] for at least1 day followed by a mixture of laminin [0.5 μg/ml] and rat tail collagen[0.2 mg/ml] (Roche) for 4–6 h at 4°C. After 1–2 h most of the cells hadadhered and the dish was filled with additional 2–3 ml medium. Cultureswere incubated at 37°C, 0% CO2 and 90% humidity over night. The cellswere washed with PBS the next day and then further cultured in the old,conditioned medium mixed with fresh preparation medium in equalvolumes. Recombinant human NGF (Genentech) was added to thismedium at a concentration of 10 ng/ml before this feeding step. Thisneurotrophin concentration is sufficient to support the survival of neuronsbut does not elicit acute effects on synaptic transmission (Lockhart et al.,1997).
Isolation procedure and transfection of sympathetic neuronsThe lumbosacral part of the paravertebral sympathetic chain was
dissected from E13 chicken embryos and collected in PBS. Then, excessconnective tissue was removed. The chains were incubated in 2 ml PBSwith trypsin [1 mg/ml] (Worthington) in a 15 ml centrifugation tube at 37°Cfor 15 min. Chains were collected by brief centrifugation, 10–15 chains pertransfection were transferred to an electroporation cuvette (Apollo, 2 mmelectrode distance), filled with 100 μl of PBS. Subsequently, plasmids (seebelow) were added at a concentration of 2+2 μg in double transfections or2 μg in single transfections. The trypsinized ganglia were electroporatedwith the Nucleofector I (Amaxa Biosystems) using the program O-03.During the rough process of electroporation the ganglia partly disintegratedinto smaller fragments, also releasing many single neurons. The content ofthe electroporation cuvette was then plated onto 4–6 glass bottom dishescontaining the 2-day-old cardiomyocyte cultures and incubated at 37°C, 0%CO2 and 90% humidity for approximately 30 h. Immunocytochemicalstaining for synapsin one was performed according to the protocol providedby the manufacturer (Chemicon).
Application of neurotrophic factors for acute effects and blockersFactors were pipetted into the dishes to a final concentration of 100 ng/
ml human NGF (Genentech), 50 ng/ml human BDNF (Amgen-Regeneron)and 10 ng/ml recombinant human CNTF (PeproTech). The dishes were thenincubated at 37°C for 30 min and examined with a confocal microscope. Inthe blocking experiments and the corresponding controls, antiserum againstp75NTR (final dilution of antiserum: 1:2000; Huber and Chao, 1995) and thetyrosine kinase inhibitor K252a (Fermentek, final concentration: 500 nM)were pipetted into the dish for 30 min at 37°C before application of theneurotrophic factors or image acquisition, respectively.
Imaging and fluorescence labelingAll images were acquired with a Zeiss 510 Meta confocal microscope
with a 63× oil immersion objective at room temperature. The pinhole was

8 E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
adjusted to achieve an optical section thickness of 1 μm. Data werecollected from at least 3 (usually more) independent neuronal cell culturepreparations for each experimental approach; the number of preparationsfor the neurotrophin experiments are separately mentioned in the results. 3Dreconstructions were performed with the Amira software (Mercurycomputer systems) and represent surface renderings defined by asubjectively chosen intensity threshold. These reconstructions were onlyused to visualize the spatial distribution of the fluorescence signals. Forquantitative image analysis only confocal cross-sections were considered.For statistical analysis, the data were tested for normal distribution(Kolrogonov–Smirnov test) and p-values were determined either withMann–Whitney (not normally distributed) or with a Student’s t-test(normally distributed).
Colocalization analysis of fluorescent proteinsIntracellular separation of fluorescent proteins was determined either
by counting single, readily distinguishable green or red fluorescencepackets (as in Figs. 3A and 5C) or by calculating a colocalization-index(CI), representing the ratio of the area of overlapping red/greenfluorescence and the total fluorescence area of the two single fluorescencecolors (as in Figs. 2A, D and 4A, E). To achieve a maximum of accuracyin the soma of the cell the CI of VAChT-GFP/VMAT2-DsRedtransfections and control transfection was always determined bycalculating the mean CI from 3 different confocal sections of the samecell.
Distance from membraneNeurons that had been transfected with either VMAT2-GFP or VAChT-
GFP were fixed in 4% paraformaldehyde for 10 min, washed and incubatedin PBS with component G (lipophilic tracer sample kit; Molecular Probes)and dissolved in DMSO/Pluronic (Molecular Probes) for 10 min to labelthe plasma membrane. The distance between single fluorescence packetsand the membrane was determined along a linear intensity profileintersecting the two fluorescence intensity peaks, one representing thevesicle and the other one the plasma membrane. The confocal section withthe most clearly defined membrane cross-section was used for thismeasurement (Fig. 5A).
pH measurements of fluorescence packetsNeurons that had been transfected with either VMAT2-GFP or VAChT-
GFP were incubated with the pH-indicator Snarf-4F (Molecular Probes) for30 min according to the manufacturer’s instructions. The fluorescenceemission ratio at 590 nm/640 nm and the GFP fluorescence were assessedsimultaneously at an excitation wavelength of 488 nm. In linear intensityprofiles, covering the region within 1 μm of the cell membrane, all peaksabove background fluorescence were counted that were either positive foronly GFP or both GFP in combination with a high fluorescence 590 nm/640 nm ratio representing neutral pH (Fig. 5B).
Acknowledgments
We thank Masami Takahashi and Makoto Itakura forgenerously providing us with the vectors used in this study andMoses Chao for his gift of p75NTR antiserum. The excellenttechnical assistance of Heidi Miedl, Sandra Kemmerling and TanjaMassimo is gratefully acknowledged.
References
Erickson, J.D., Schafer, M.K., Bonner, T.I., Eiden, L.E., Weihe, E., 1996a.Distinct pharmacological properties and distribution in neurons andendocrine cells of two isoforms of the human vesicular monoaminetransporter. Proc. Natl. Acad. Sci. U. S. A. 93, 5166–5171.
Erickson, J.D., Weihe, E., Schaefer, M.K.H., Neale, E., Williamson, L.,Bonner, T.I., Tao-Chen, J.H., Eiden, L.E., 1996b. The VAChT/ChAT
“cholinergic gene locus”: new aspects of genetic and vesicular regulationof cholinergic function. Prog. Brain Res. 109, 69–82.
Furshpan, E.J., Landis, S.C., Matsumoto, S.G., Potter, D.D., 1986a.Synaptic functions in rat sympathetic neurons in microcultures: I.Secretion of norepinephrine and acetylcholine. J. Neurosci. 6 (4),1061–1079.
Furshpan, E.J., Potter, D.D., Matsumoto, S.G., 1986b. Synaptic functions inrat sympathetic neurons in microcultures: III. A purinergic effect oncardiac myocytes. J. Neurosci. 6 (4), 1099–1107.
Huber, L.J., Chao, M.V., 1995. Mesenchymal and neuronal cell expressionof the p75 neurotrophin receptor gene occurs by different mechanisms.Dev. Biol. 167, 227–238.
Johnson, M.I., Ross, C.D., Bunge, R.P., 1980a. Morphological andbiochemical studies on the development of cholinergic properties incultures sympathetic neurons: II. Dependence on postnatal age. J. CellBiol. 84, 692–704.
Johnson, M.I., Ross, C.D., Meyers, M., Spitznagel, E.L., Bunge, R.P.,1980b. Morphological and biochemical studies on the development ofcholinergic properties in cultures sympathetic neurons: I. Correlativechanges in choline acetyltransferase and synaptic vesicle cytochemistry.J. Cell Biol. 84, 680–691.
Kleijn, M., Welsh, G.I., Scheper, G.C., Voorma, H.O., Proud, C.G., Thomas,A.A., 1998. Nerve and epidermal growth factor induce protein synthesisand eIF2B activation in PC12 cells. J. Biol. Chem. 273, 5536–5541.
Krantz, D.E., Waites, C., Oorschot, V., Liu, Y., Wilson, R.I., Tan, P.K.,Klumperman, J., Edwards, R.H., 2000. A phosphorylation site regulatessorting of the vesicular acetylcholine transporter to dense core vesicles.J. Cell Biol. 149 (2), 379–395.
Landis, S.C., 1976. Rat sympathetic neurons and cardiac myocytesdeveloping in microcultures: correlation of the fine structure of endingswith neurotransmitter function in single neurons. Proc. Natl. Acad. Sci.U. S. A. 73 (11), 4220–4224.
Liu, Y., Edwards, R.H., 1997. Differential localization of vesicularacetylcholine and monoamine transporters in PC12 cells but not CHOcells. J. Cell Biol. 139 (4), 907–916.
Liu, Y., Schweitzer, E.S., Nirenberg, M.J., Pickel, V.M., Evans, C.J.,Edwards, R.H., 1994. Preferential localization of a vesicular monoaminetransporter to dense core vesicles in PC12 cells. J Cell Biol. 127,1419–1433.
Lockhart, S.T., Turrigiano, G.G., Birren, S.J., 1997. Nerve growth factormodulates synaptic transmission between sympathetic neurons andcardiac myocytes. J. Neurosci. 17 (24), 9573–9582.
Matsumoto, S.G., Sah, D., Potter, D.D., Furshpan, E.J., 1987. Synapticfunctions in rat sympathetic neurons in microcultures: IV. Noradrenergicexcitation of cardiac myocytes and the variety of multiple-transmitterstates. J. Neurosci. 7 (2), 380–390.
Potter, D.D., Landis, S.G., Matsumoto, S.G., Furshpan, E.J., 1986. Synapticfunctions in rat sympathetic neurons in microcultures: II. Adrenergic/cholinergic dual status and plasticity. J. Neurosci. 6 (4), 1080–1098.
Shoju-Kasai, Y., Itakura, M., Kataoka, M., Yamamori, S., Takahashi, M.,2002. Protein kinase C-mediated translocation of secretory vesicles toplasma membrane and enhancement of neurotransmitter release fromPC12 cells. Eur. J. Neurosci. 15, 1390–1394.
Slonimsky, J.D., Yang, B., Hintereder, J.M., Nokes, E.B., Birren, S.J., 2002.BDNF and CNTF regulate cholinergic properties of sympatheticneurons through independent mechanisms. Mol. Cell. Neurosci. 23(4), 648–660.
Slonimsky, J.D., Mattaliano, M.D., Moon, J., Griffith, L.C., Birren, S.J.,2006. Role for calcium/calmodulin-dependent protein kinase II in thep75-mediated regulation of sympathetic cholinergic transmission. Proc.Natl. Acad. Sci. U. S. A. 103, 2915–2919.
Tan, P.K., Waites, C., Liu, Y., Krantz, D.E., Edwards, R.H., 1998. A leucine-based motif mediates the endocytosis of vesicular monoamine andacetylcholine transporters. J. Biol. Chem. 10 (273 (28)), 17351–17360.
Varoqui, H., Erickson, J.D., 1998. The cytoplasmatic tail of the vesicularacetylcholine transporter contains a synaptic vesicle targeting signal.J. Biol. Chem. 273 (15), 9094–9098.

9E. Felder, G. Dechant / Mol. Cell. Neurosci. 34 (2007) 1–9
Waites, C.L., Mehta, A., Tan, P.K., Thomas, G., Edwards, R.H., Krantz, D.E., 2001. An acidic motif retains vesicular monoamine transporter 2 onlarge dense core vesicles. J. Cell Biol. 152 (6), 1159–1168.
Weihe, E., Tao-Cheng, J.H., Schaefer, M.K.H., Erickson, J.D., Eiden, L.E.,1996. Visualization of the vesicular acetylcholine transporter in
cholinergic nerve terminals and its targeting to a specific population ofsmall synaptic vesicles. Proc. Natl. Acad. Sci. U. S. A. 93, 3547–3552.
Yang, B., Slonimski, J.D., Birren, S.J., 2002. A rapid switch in sympatheticneurotransmitter release properties mediated by the p75 receptor. Nat.Neurosci. 5 (6), 539–545.
Related Documents



![Expression cloning of monoamine transporter · abilized CV-1 cells (-5 X 104 cells) expressing MAT. (A) Total uptakeof[3H]5HT(0.4 JLM)at5 minwith5 mMATP(CON),in the presence of 100](https://static.cupdf.com/doc/110x72/5f46a6d6df5f79688c496b19/expression-cloning-of-monoamine-transporter-abilized-cv-1-cells-5-x-104-cells.jpg)






