Neurotoxicity and Inflammation in the Nasal Airways of Mice Exposed to the Macrocyclic Trichothecene Mycotoxin Roridin A: Kinetics and Potentiation by Bacterial Lipopolysaccharide Coexposure Zahidul Islam,* , † Chidozie J. Amuzie,* , † Jack R. Harkema,† , ‡ and James J. Pestka* , † ,1 *Department of Food Science and Human Nutrition, †Center for Integrative Toxicology, and ‡Department of Pathobiology and Diagnostic Investigation, Michigan State University, East Lansing, Michigan 48824 Received February 7, 2007; accepted April 27, 2007 Macrocyclic trichothecene mycotoxins produced by indoor air molds potentially contribute to symptoms associated with damp building illnesses. The purpose of this investigation was to de- termine (1) the kinetics of nasal inflammation and neurotoxicity after a single intranasal instillation of roridin A (RA), a represen- tative macrocyclic trichothecene; and (2) the capacity of lipopoly- saccharide (LPS) to modulate RA’s effects. C57Bl/6 female mice were intranasally instilled once with 50 ml of RA (500 mg/kg body weight [bw]) in saline or saline only and then nose and brain tissues were collected over 72 h and processed for histopathologic and messenger RNA (mRNA) analysis. RA-induced apoptosis specifically in olfactory sensory neurons (OSNs) after 24 h post- instillation (PI) causing marked atrophy of olfactory epithelium (OE) that was maximal at 72 h PI. Concurrently, there was marked bilateral atrophy of olfactory nerve layer of the olfactory bulbs (OBs) of the brain. In the ethmoid turbinates, upregulated messenger RNA (mRNA) expression of the proapoptotic gene FAS and the proinflammatory cytokines tumor necrosis factor-a, interleukin (IL)-6, IL-1, and macrophage inhibitory protein-2 was observed from 6 to 24 h PI, whereas expression of several other proapoptotic genes (PKR, p53, Bax, and caspase-activated DNAse) was detectable only at 24 h PI. Simultaneous exposure to LPS (500 ng/kg bw) and a lower dose of RA (250 mg/kg bw) magnified RA-induced proinflammatory gene expression, apopto- sis, and inflammation in the nasal tract. Taken together, the results suggest that RA markedly induced FAS and proinflammatory cytokine expression prior to evoking OSN apoptosis and OE atrophy and that RA’s effects were augmented by LPS. Key Words: trichothecene; mycotoxin; apoptosis; neurotoxicity; inflammation; endotoxin. Adverse human health effects are often attributed to damp indoor air environments generated by excessive condensation, failure of water-use devices, or building envelope breach dur- ing flooding (Institute of Medicine, 2004). Symptoms typically reported by affected occupants of water-damaged buildings include cough, rhinosinusitis, exacerbation of asthma, and in- creased susceptibility to respiratory infections. For example, a recent National Institute of Occupational Safety and Health (NIOSH) study (Cox-Ganser et al., 2005) reported that exces- sive respiratory symptoms and physician-diagnosed asthma oc- curred among occupants in an office building that was subject to water incursions. These effects contributed to overall burden of illness delineated in terms of absences, use of breathing medications, and health-related quality of life. Damp building–related illnesses are frequently linked to aberrant growth of mold (Andersson et al., 1997; Fog Nielsen, 2003). In 2004, an Institute of Medicine (IOM) expert com- mittee (Institute of Medicine, 2004) concluded that there are sufficient scientific data for an association between the ex- posure to moldy damp indoor environments and (1) upper respiratory tract symptoms (nasal congestion, sneezing, runny or itchy nose, throat irritation) as well as (2) some lower res- piratory tract symptoms/syndromes (cough, wheeze, asthma exacerbation, hypersensitivity pneumonitis in susceptible peo- ple), but that insufficient evidence exists to date for other sometimes disparate health conditions (airflow obstruction, mu- cous membrane irritation, chronic obstructive pulmonary dis- ease, pulmonary hemorrhage, neurologic effects, and cancer). The black mold Stachybotrys chartarum is a saprophytic fungus that grows on cellulosic building materials including wallboard, ceiling tiles, and cardboard (Andersson et al., 1997; Boutin-Forzano et al., 2004; Tuomi et al., 1998, 2000). It has been postulated that building-related S. chartarum, its tricho- thecene mycotoxins, or other byproducts are possible etiologic contributors to debilitating respiratory illnesses (Fung et al., 1998; Hossain et al., 2004; Jarvis et al., 1986, 1998; Kilburn, 2004) as well as immune dysfunction (Johanning et al., 1996) and cognitive impairment (Gordon, 1999). The aforementioned IOM report (Institute of Medicine, 2004) suggested, however, that while in vitro and in vivo research on S. chartarum suggest that adverse effects in humans are indeed ‘‘biologically plau- sible,’’ establishing a clear association with building-related illnesses will require rigorous study from the perspectives of mechanisms, dose–response and exposure assessment. 1 To whom correspondence should be addressed at 234 G.M. Trout Building, Michigan State University, East Lansing, MI 48824-1224. Fax: (517) 353-8963. E-mail: [email protected]. Ó The Author 2007. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For Permissions, please email: [email protected] TOXICOLOGICAL SCIENCES 98(2), 526–541 (2007) doi:10.1093/toxsci/kfm102 Advance Access publication May 4, 2007 by guest on November 19, 2014 http://toxsci.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neurotoxicity and Inflammation in the Nasal Airways of Mice Exposedto the Macrocyclic Trichothecene Mycotoxin Roridin A: Kinetics and
Potentiation by Bacterial Lipopolysaccharide Coexposure
Zahidul Islam,*,† Chidozie J. Amuzie,*,† Jack R. Harkema,†,‡ and James J. Pestka*,†,1
*Department of Food Science and Human Nutrition, †Center for Integrative Toxicology, and ‡Department of Pathobiology and Diagnostic Investigation,
Michigan State University, East Lansing, Michigan 48824
Received February 7, 2007; accepted April 27, 2007
Macrocyclic trichothecene mycotoxins produced by indoor air
molds potentially contribute to symptoms associated with damp
building illnesses. The purpose of this investigation was to de-
termine (1) the kinetics of nasal inflammation and neurotoxicity
after a single intranasal instillation of roridin A (RA), a represen-
tative macrocyclic trichothecene; and (2) the capacity of lipopoly-
saccharide (LPS) to modulate RA’s effects. C57Bl/6 female mice
were intranasally instilled once with 50 ml of RA (500 mg/kg body
weight [bw]) in saline or saline only and then nose and brain
tissues were collected over 72 h and processed for histopathologic
and messenger RNA (mRNA) analysis. RA-induced apoptosis
specifically in olfactory sensory neurons (OSNs) after 24 h post-
instillation (PI) causing marked atrophy of olfactory epithelium
(OE) that was maximal at 72 h PI. Concurrently, there was
marked bilateral atrophy of olfactory nerve layer of the olfactory
bulbs (OBs) of the brain. In the ethmoid turbinates, upregulated
messenger RNA (mRNA) expression of the proapoptotic gene FAS
and the proinflammatory cytokines tumor necrosis factor-a,interleukin (IL)-6, IL-1, and macrophage inhibitory protein-2
was observed from 6 to 24 h PI, whereas expression of several
other proapoptotic genes (PKR, p53, Bax, and caspase-activated
DNAse) was detectable only at 24 h PI. Simultaneous exposure to
LPS (500 ng/kg bw) and a lower dose of RA (250 mg/kg bw)
magnified RA-induced proinflammatory gene expression, apopto-
sis, and inflammation in the nasal tract. Taken together, the results
suggest that RA markedly induced FAS and proinflammatory
cytokine expression prior to evoking OSN apoptosis and OE
atrophy and that RA’s effects were augmented by LPS.
Key Words: trichothecene; mycotoxin; apoptosis; neurotoxicity;
inflammation; endotoxin.
Adverse human health effects are often attributed to dampindoor air environments generated by excessive condensation,failure of water-use devices, or building envelope breach dur-ing flooding (Institute of Medicine, 2004). Symptoms typicallyreported by affected occupants of water-damaged buildings
include cough, rhinosinusitis, exacerbation of asthma, and in-creased susceptibility to respiratory infections. For example,a recent National Institute of Occupational Safety and Health(NIOSH) study (Cox-Ganser et al., 2005) reported that exces-sive respiratory symptoms and physician-diagnosed asthma oc-curred among occupants in an office building that was subjectto water incursions. These effects contributed to overall burdenof illness delineated in terms of absences, use of breathingmedications, and health-related quality of life.
Damp building–related illnesses are frequently linked toaberrant growth of mold (Andersson et al., 1997; Fog Nielsen,2003). In 2004, an Institute of Medicine (IOM) expert com-mittee (Institute of Medicine, 2004) concluded that there aresufficient scientific data for an association between the ex-posure to moldy damp indoor environments and (1) upperrespiratory tract symptoms (nasal congestion, sneezing, runnyor itchy nose, throat irritation) as well as (2) some lower res-piratory tract symptoms/syndromes (cough, wheeze, asthmaexacerbation, hypersensitivity pneumonitis in susceptible peo-ple), but that insufficient evidence exists to date for othersometimes disparate health conditions (airflow obstruction, mu-cous membrane irritation, chronic obstructive pulmonary dis-ease, pulmonary hemorrhage, neurologic effects, and cancer).
The black mold Stachybotrys chartarum is a saprophyticfungus that grows on cellulosic building materials includingwallboard, ceiling tiles, and cardboard (Andersson et al., 1997;Boutin-Forzano et al., 2004; Tuomi et al., 1998, 2000). It hasbeen postulated that building-related S. chartarum, its tricho-thecene mycotoxins, or other byproducts are possible etiologiccontributors to debilitating respiratory illnesses (Fung et al.,1998; Hossain et al., 2004; Jarvis et al., 1986, 1998; Kilburn,2004) as well as immune dysfunction (Johanning et al., 1996)and cognitive impairment (Gordon, 1999). The aforementionedIOM report (Institute of Medicine, 2004) suggested, however,that while in vitro and in vivo research on S. chartarum suggestthat adverse effects in humans are indeed ‘‘biologically plau-sible,’’ establishing a clear association with building-relatedillnesses will require rigorous study from the perspectives ofmechanisms, dose–response and exposure assessment.
1 To whom correspondence should be addressed at 234 G.M. Trout Building,
Michigan State University, East Lansing, MI 48824-1224. Fax: (517) 353-8963.
E-mail: [email protected].
� The Author 2007. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved.For Permissions, please email: [email protected]
TOXICOLOGICAL SCIENCES 98(2), 526–541 (2007)
doi:10.1093/toxsci/kfm102
Advance Access publication May 4, 2007
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
The trichothecene mycotoxins comprise more than 180structurally related sesquiterpenoid metabolites and are pro-duced by Fusarium, Stachybotrys, Myrothecium, and othermolds (Grove, 1988, 1993, 2000). Trichothecenes are potenttranslational inhibitors that have in common a 9, 10 doublebond, and a 12, 13 epoxide group, but extensive variation existsrelative to ring oxygenation patterns. The Type D or ‘‘macro-cyclic’’ trichothecenes, which have a cyclic diester or triesterring linking C-4 to C-15 (e.g., satratoxins or roridins), areproduced by Stachybotrys and Myrothecium. Macrocyclictrichothecenes occur in the outer plasmalemma surface andthe inner wall layers of fungal conidiospores (Gregory et al.,2004). Dry spores are readily aerosolized and have a respirablemean aerodynamic diameter of approximately 5 lm (Sorensonet al., 1987; Yike and Dearborn, 2004). In addition, nonviable,fine (� 1 lm in diameter) airborne particulates can also containtrichothecenes (Brasel et al., 2005). Thus, human exposure tothese toxins is indeed possible.
Several experimental studies of S. chartarum spores havefocused on the contributions of trichothecenes to airway pa-thology. Rand et al. (2002) determined, in direct intratrachealinstillation studies of mice, that respiratory pathology causedby fungal spores could be reproduced with pure isosatratoxin F.After investigating the effects of a single tracheal instillation offungal spores on survival, growth, histopathology of the lung,and respiration in the rat pup model, Yike et al. (2002) con-cluded that mycotoxins were critical for the hemorrhagic andinflammatory responses. In subsequent work, this group dem-onstrated that toxic S. chartarum spores or associated macro-cyclic trichothecenes initiated acute inflammatory responses inthe rodent lung (Yike et al., 2005; Yike and Dearborn, 2004).Recently, our laboratory has observed that acute intranasalexposure to satratoxin G and isosatratoxin F, macrocyclictrichothecenes produced by S. chartarum, causes inflammationand apoptosis in the murine nose 24 h after instillation (Islamet al., 2006). Both olfactory sensory neurons (OSNs) and theolfactory bulb (OB) of the brain are targets of satratoxinG–induced toxicity. Elevated proinflammatory cytokine geneexpression is also observed at 24 h PI, suggesting that thesemediators might contribute to OSN apoptosis as well as ac-companying rhinitis and mild focal encephalitis observed intoxin-exposed mice.
Occupants of damp buildings are likely to be exposed toother microbial products besides fungi (Institute of Medicine,2004). Notably, endotoxin often co-occurs with molds in dampbuildings (Bornehag et al., 2004; Foto et al., 2005; Heinrichet al., 2003; Kovesi et al., 2006; Sebastian and Larsson, 2003;Sebastian et al., 2005) as was dramatically evidenced in NewOrleans following Hurricane Katrina (Solomon et al., 2006).A recent NIOSH study suggested that occupant respiratoryproblems might result from the combined exposure to fungiand endotoxin and that both agents should be assessed inepidemiological investigations of indoor environments (Parket al., 2006). Oral trichothecene toxicity is amplified by
coexposure to lipopolysaccharide (LPS), the major bioactivecomponent of endotoxin (Chung et al., 2003a,b; Islam et al.,2002; Mbandi and Pestka, 2006; Tai and Pestka, 1988b; Zhouet al., 1999, 2000). LPS might similarly enhance the nasaltoxicity of macrocyclic trichothecenes.
Since satratoxins are not commercially available and theyare extremely difficult to purify from fungal culture, charac-terization of their toxic effects and mechanisms whether aloneor in the context of co-occurring contaminants presents atechnical challenge. One possible alternative would be to em-ploy another macrocyclic trichothecene as a surrogate for thesatratoxins. Roridin A (RA) is one such structurally relatedtrichothecene that is produced in copious amounts by thefungus Myrothecium and that is commercially available. Inpreliminary studies, we have observed that, like the satratoxins,
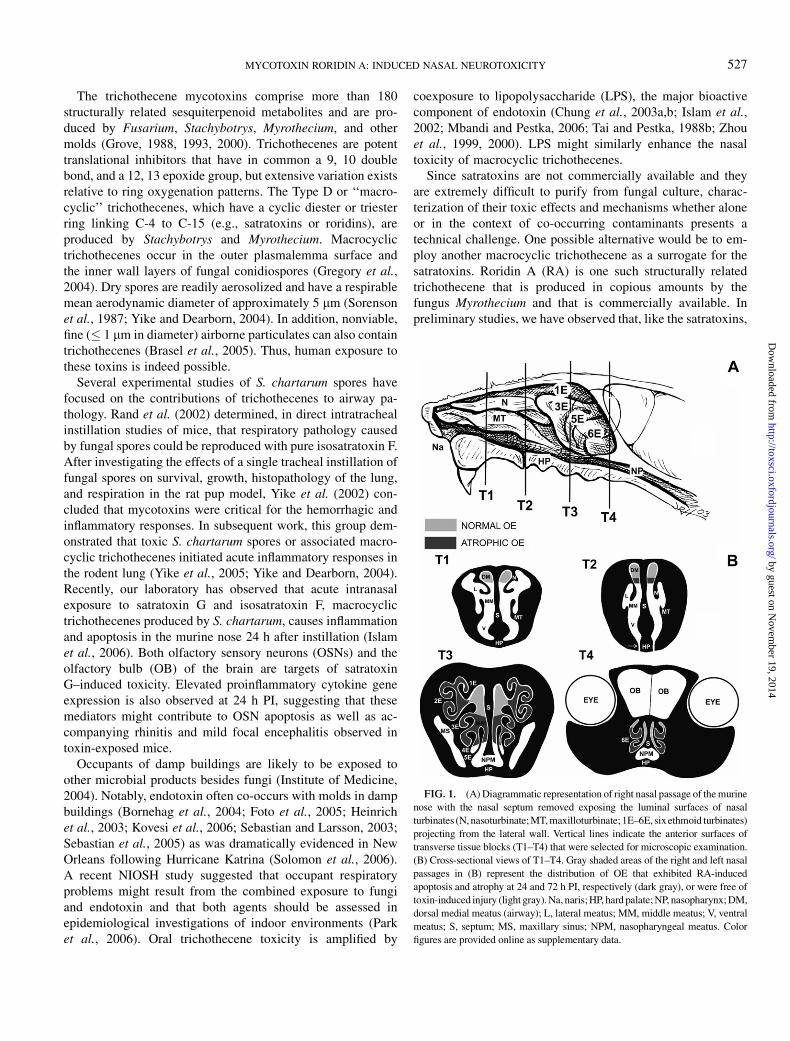
FIG. 1. (A) Diagrammatic representation of right nasal passage of the murine
nose with the nasal septum removed exposing the luminal surfaces of nasal
turbinates (N, nasoturbinate; MT, maxilloturbinate; 1E–6E, sixethmoid turbinates)
projecting from the lateral wall. Vertical lines indicate the anterior surfaces of
transverse tissue blocks (T1–T4) that were selected for microscopic examination.
(B) Cross-sectional views of T1–T4. Gray shaded areas of the right and left nasal
passages in (B) represent the distribution of OE that exhibited RA-induced
apoptosis and atrophy at 24 and 72 h PI, respectively (dark gray), or were free of
toxin-induced injury (light gray). Na, naris; HP, hard palate; NP, nasopharynx; DM,
dorsal medial meatus (airway); L, lateral meatus; MM, middle meatus; V, ventral
meatus; S, septum; MS, maxillary sinus; NPM, nasopharyngeal meatus. Color
figures are provided online as supplementary data.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 527
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
RA can induce inflammation and OSN loss in mice followingintranasal instillation. The purpose of this study was two fold.First, we determined the relative kinetics of proinflammatorycytokine and proapoptotic gene expression following intranasalRA exposure and further related these effects to OSN apoptosisand OE atrophy. Second, we assessed the capacity of LPS tomodulate RA-induced gene expression and toxicity in the nasalairway. This investigation (1) confirmed that RA mimickedsatratoxin G’s toxic effects in the nose, (2) identified fromexpression kinetics those genes likely to contribute to apopto-sis, and (3) demonstrated the capacity of LPS to exacerbateRA’s adverse effects.
MATERIALS AND METHODS
Experimental design. RA was purchased from Sigma Chemical Co.
(St Louis, MO). Toxin purity was > 99% as determined by a single high
performance liquid chromatography peak at 260 nm (Hinkley and Jarvis, 2001).
LPS derived from Escherichia coli serotype 0111:B4 with an activity of 1.5 3
106 EU/mg was purchased from Sigma. Animal studies were conducted in
accordance with National Institutes of Health guidelines as overseen by the All
University Committee on Animal Use and Care at Michigan State University.
Pathogen-free female C57Bl/6 mice (7–8 weeks, Charles River, Portage, MI)
were randomly assigned to experimental groups (n ¼ 6) and housed in
polycarbonate cages containing Cell-Sorb plus bedding (A & W Products,
Cincinnati, OH) and covered with filter bonnets. Mice were provided free
access to food and water. Room lights were set on a 12-h light/dark cycle, and
temperature and relative humidity were maintained between 21�C–24�C and
40–55% humidity, respectively.
For each experiment, mice were anesthetized with 4% halothane and
96% oxygen and then instilled intranasally at 50 ll per mouse with RA (250 or
500 lg/kg body weight [bw]) and/or LPS (500 ng/kg bw) dissolved in a vehicle
of pyrogen-free saline (Abbott Laboratories, IL) or with vehicle(s) alone.
Selection of RA doses was based on capacity to mimic satratoxin G olfactory
effects in preliminary experiments. Mice were euthanized after various time
intervals and selected tissues from the nose and brain were subjected to
histopathologic and messenger RNA (mRNA) analysis as described below.
Animal necropsies and tissue processing for light microscopic examina-
tion. Mice were sacrificed at 6, 12, 24, or 72 h PI. For light morphologic
examination and morphometric analyses of nasal cavity, mice were deeply
anesthetized via ip injection of 0.1 ml of 12% (wt/vol) sodium pentobarbital
and killed via exsanguination by cutting the abdominal aorta. Heads from each
mouse were immediately removed and 1 ml of 10% (vol/vol) neutral buffered
formalin (Fisher Scientific Co., Fairlawn, NJ) was flushed retrograde through
the nasopharyngeal meatus. After the lower jaw, skin, muscles, and dorsal
cranium were removed, the head with the intact brain was immersed and stored
in a large volume of the fixative for at least 24 h prior to further tissue
processing.
After fixation, transverse tissue blocks from the head of these mice were
selected for light microscopy as previously described (Islam et al., 2006). Prior
to sectioning, the heads were decalcified in 13% (vol/vol) formic acid for 7 days
and then rinsed in tap water for at least 4 h. The nasal cavity of each mouse was
transversely sectioned at four specific anatomic locations (Islam et al., 2006;
Mery et al., 1994) (Fig. 1A). The most proximal nasal section was taken
immediately posterior to the upper incisor teeth (proximal, T1); the middle
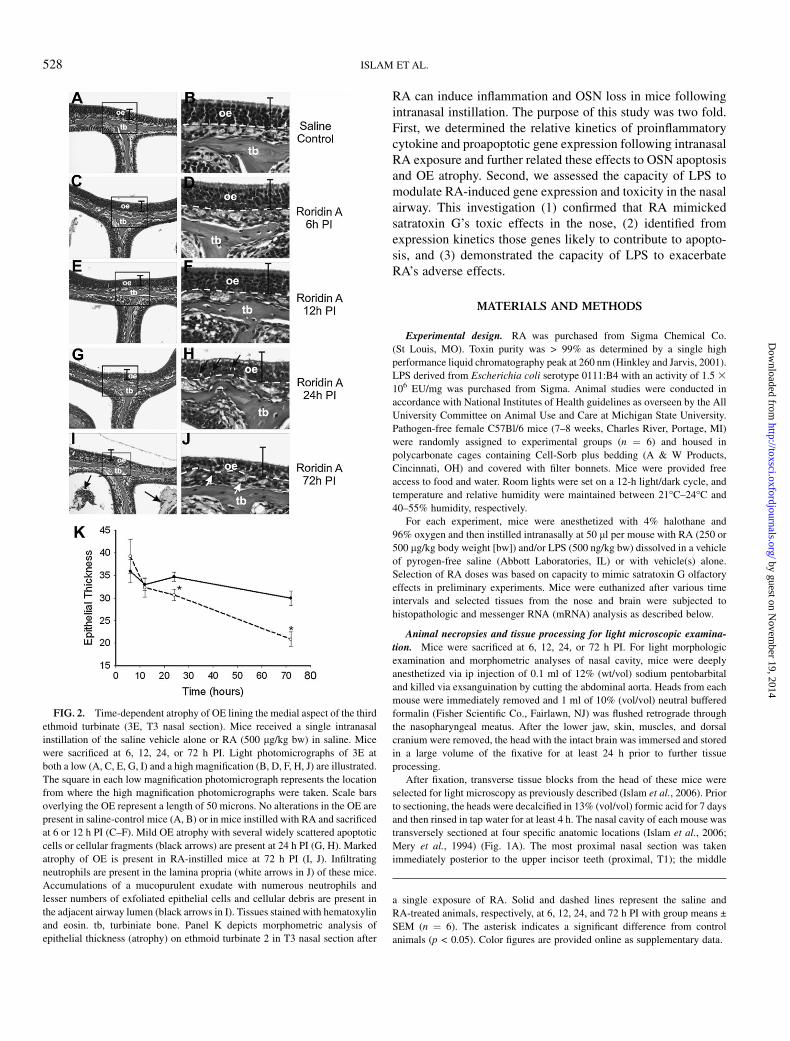
FIG. 2. Time-dependent atrophy of OE lining the medial aspect of the third
ethmoid turbinate (3E, T3 nasal section). Mice received a single intranasal
instillation of the saline vehicle alone or RA (500 lg/kg bw) in saline. Mice
were sacrificed at 6, 12, 24, or 72 h PI. Light photomicrographs of 3E at
both a low (A, C, E, G, I) and a high magnification (B, D, F, H, J) are illustrated.
The square in each low magnification photomicrograph represents the location
from where the high magnification photomicrographs were taken. Scale bars
overlying the OE represent a length of 50 microns. No alterations in the OE are
present in saline-control mice (A, B) or in mice instilled with RA and sacrificed
at 6 or 12 h PI (C–F). Mild OE atrophy with several widely scattered apoptotic
cells or cellular fragments (black arrows) are present at 24 h PI (G, H). Marked
atrophy of OE is present in RA-instilled mice at 72 h PI (I, J). Infiltrating
neutrophils are present in the lamina propria (white arrows in J) of these mice.
Accumulations of a mucopurulent exudate with numerous neutrophils and
lesser numbers of exfoliated epithelial cells and cellular debris are present in
the adjacent airway lumen (black arrows in I). Tissues stained with hematoxylin
and eosin. tb, turbiniate bone. Panel K depicts morphometric analysis of
epithelial thickness (atrophy) on ethmoid turbinate 2 in T3 nasal section after
a single exposure of RA. Solid and dashed lines represent the saline and
RA-treated animals, respectively, at 6, 12, 24, and 72 h PI with group means ±
SEM (n ¼ 6). The asterisk indicates a significant difference from control
animals (p < 0.05). Color figures are provided online as supplementary data.
528 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

section was taken at the level of the incisive papilla of the hard palate (middle,
T2); the third nasal section was taken at the level of the second palatal ridge
(T3); and the most distal nasal section (T4) was taken at the level of the
intersection of the hard and soft palate and through the proximal portion of the
OB of the brain (Fig. 1B). Tissue blocks were embedded in paraffin and
the anterior face of each block was sectioned at a thickness of 5 lm, and stained
with hematoxylin and eosin.
Immunohistochemistry. Unstained and hydrated paraffin sections from
the same nasal blocks (T1–T4) were incubated first with a nonspecific pro-
tein blocking solution containing normal sera (Vector Laboratories Inc.,
Burlingame, CA) and then with specific dilutions of primary polyclonal
antibodies directed against caspase-3 (1:100; rabbit anti-caspase-3 antibody;
Abcam, Inc., Cambridge, MA), olfactory marker protein (OMP) (1:4000; goat
anti-OMP antibody provided by Dr. Frank Margolis, University of Maryland),
or neutrophils infiltrating the nasal mucosa (1:600; rabbit anti-rat neutrophil
antibody provided by Dr Robert Roth, Michigan State University). Tissue
sections used for caspase-3 or OMP detection were pretreated prior to the
blocking solution with 3% (vol/vol) H2O2 in methanol to destroy endoge-
nous peroxidase. Tissue sections were then incubated with biotinylated
antispecies IgG. Immunoreactivity of caspase-3 and OMP was visualized with
Vector R.T.U. Elite ABC-Peroxidase Reagent followed by Nova Red chroma-
gen. Anti-neutrophil antibody treatment was sequentially followed with
biotinylated anti-rabbit IgG, Streptavidin–Phosphatase complex (KPL labora-
tories, Gaithersburg, MD) and Vector Red chromagen. After immunohisto-
chemistry, slides were lightly counterstained with hematoxylin.
Light microscopic morphometry. Thickness of the olfactory epithelium
(OE) lining the medial surface of the second ethmoid turbinates (2E) in T3
(Fig. 1) was morphometrically evaluated as previously described (Islam et al.,
FIG. 3. Loss of OSNs in the OE 72 h PI after a single intranasal instillation of RA (500 lg/kg bw). Light photomicrographs illustrate the immunohistochemical
detection of OMP (marker of mature OSNs; dark intra epithelial) in the OE lining the nasal airways in the T3 nasal section from a saline-instilled control mouse (A,
C) and a mouse intranasally instilled with RA and sacrificed 72 h PI (B, D). (A, B) Photomicrographs of T3 at a low magnification illustrating the bilateral nasal
passages containing ethmoid turbinates (e.g., E1, E2, E3) and the dividing nasal septum (S). (C, D) High magnification photomicrographs of the OE lining the
medial aspect of E2 (see rectangles in A, B) in the saline- and RA-instilled mice, respectively. There is widespread loss of OMP-staining in the OE of the RA-
instilled mouse compared to the saline-instilled control mouse (open arrows in (B) indicate regions with loss of OMP; closed arrows illustrate regions where OMP
is still present). Closed arrows in (C) and (D) indicate the presence and loss, respectively, of OMP-positive cilia of the dendritic portion of the OSNs. DMM,
dorsomedial meatus; LM, lateral meatus; lp, lamina propria, MM, medial meatus; m, mucoproteinaceous material in the nasal airway containing a few
inflammatory cells and epithelial cellular debris; MS, maxillary sinus; NPM, nasopharyngeal meatus; tb, turbinate bone. Tissue sections were counterstained with
hematoxylin. Color figures are provided online as supplementary data.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 529
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

2006). Briefly, measurements were conducted at a final magnification of 31920
using a light microscope (Olympus BX40; Olympus America Inc., Melville,
NY), coupled to a 3.3 megapixel digital color camera (Q-Color 3 Camera,
Quantitative Imaging Corp., Burnaby, BC, Canada), and a PC (Dell Dimension
8200, Austin, TX). The morphometric analyses were performed using a cycloid
grid overlay and software for counting points and intercepts (Stereology
Toolbox, Davis, CA) (Hyde et al., 1990, 1991). The percentage volume density,
Vv, the proportion of the epithelium composed of cytoplasm, nuclei, or
apoptotic nuclear fragments was determined by point counting and calculated
using the following formula:
Vv ¼PP �Pn=Pt
where Pp is the point fraction of Pn, the number of test points hitting the
structure of interest, divided by Pt, the total points hitting the reference space
(OE). The volume of the epithelial component of interest (e.g., apoptotic
nuclei) per unit of basement membrane (Sv) was determined by point and
intercept counting and was calculated using the following formula:
Sy ¼ 2IoLr
where Io is the number of intercepts with the object (epithelial basal lamina) and
Lr is the length of test line in the reference volume (epithelium). To determine
thickness of the OE, a volume per unit area of basal lamina (l3/l2) was then
calculated using the following formula for arithmetic mean thickness:
s¼Vv=Sv
Other standard morphometric and image analysis techniques were used to
determine the numeric cell density of mature OSNs in OE. Morphometric
estimates of the numeric cell density of OSNs immunohistochemically reactive
for OMP (protein indicator of mature OSNs) were determined via light micros-
copy (3790 final magnification) by counting the number of the nuclear profiles
of these immunoreactive neuroepithelial cells in the OE lining the medial
surface of the second ethmoid turbinates (2E) in T3 (Fig. 1) and dividing by the
length of the underlying basal lamina. The length of the basal lamina was
determined from the contour length of a computerized digital image of the basal
lamina using Scion Image program (Scion Corporation, Fredrick, MD). All
numeric cell density data were expressed as the number of OSN nuclei/mm
basal lamina.
Necropsy and tissue processing for PCR. Mice used for real-time PCR
analyses of nasal and brain tissues were anesthetized and killed at designated
times after RA instillation as described above. Immediately upon death, the
head of each mouse was removed from the carcass after the skin, muscles, and
lower jaw were removed from the head, the nasal airways were opened by
splitting the nose in a sagittal plane adjacent to the midline. The nasal septum
was removed thereby exposing the nasal turbinates projecting from the lateral
wall of each nasal passage (Fig. 1). Using a dissecting microscope and oph-
thalmic surgical instruments, all ethmoid turbinates and OB were dissected
from both nasal passages and brain, respectively. These excised tissues were
FIG. 4. RA causes depletion of OSNs in OE. Morphometric analyses were
conducted on ethmoid turbinate 2 in T3 nasal sections 72 h after a single
intranasal instillation of RA (500 lg/kg bw). Data are means of OMP-positive
cells per mm of the basal lamina ± SEM (n¼ 6). Asterisk indicates significantly
different from the saline vehicle control (p < 0.05).
FIG. 5. RA-induced atrophy of olfactory nerve layer (ONL) in the OB at 3
days PI. (A) Diagrammatic representation of nasal tissue section T4 containing
the bilateral OBs, nasal passages separated by the nasal septum (S), the sixth
ethmoid turbinate (6E), the nasopharyngeal meatus (NPM), and the hard palate
(HP). The stars in the OBs represent the sites of RA-induced atrophy. The dotted
square indicates the location from were the light photomicrographs (B, C) were
taken. (B, C) Light photomicrographs of the ONL and glomerular layer (G) in the
OB from mice instilled with the saline-vehicle alone (B) or RA (C). Tissues were
immunohistochemically stained for OMP (dark stain) and counterstained with
hematoxylin. There is marked vacuolation (arrows) and loss of OMP-staining in
(C) compared to (B). Color figures are provided online as supplementary data.
530 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

immediately stored in RNAlater (Ambion Inc., Austin, TX). Total RNA was
isolated using RNeasy Protect Mini kit (Qiagen Inc. Valencia, CA) within
7 days after removal.
Real-time PCR. Real-time PCR for proapoptotic genes (Fas, FasL, PKR,
p53, Bax, Bcl-2, caspase-activated DNAse [CAD]) and proinflammatory
cytokine genes (interleukin [IL]-1a, IL-1b, tumor necrosis factor [TNF]-a,
IL-6, MIP-2) were performed on an ABI PRISM 7900HT using Taqman One-
Step RT-PCR (reverse transcription–polymerase chain reaction) Master Mix
and Assays-on-Demand primer/probe gene expression products according to
the manufacturer’s protocols (Applied Biosystems, Foster City, NY). Relative
quantification of apoptotic and cytokine gene expression was carried out using
an 18S RNA control and an arithmetic formula method (Audige et al., 2003;
Islam et al., 2006).
Statistics. All data were analyzed with SigmaStat v 3.1 (Jandel Scientific;
San Rafael, CA) with the criterion for significance set at p < 0.05. Mor-
phometric and RT-PCR data were subjected to one-way analysis of variance
with Student–Newman–Keuls post hoc test. The capacity of LPS to magnify
RA effects in a nonadditive manner was evaluated by comparing the response
of cotreated mice to the predicted mean additive response (Islam and Pestka,
2006).
RESULTS
Intranasal RA Exposure Induces Apoptosis in OE
Light microscopic examination of the nasal tissues (T1–4)revealed that all mice (6/6) exposed to RA (500 lg/kg bw) andsacrificed at 1 and 3 days PI had conspicuous nasal epithelialand inflammatory lesions in the dorsocaudal half of the nasalpassages that is normally lined by OE (Fig. 1). These lesionswere not apparent in the nasal cavity of saline vehicle–treatedcontrols nor were they observed in RA-instilled mice that weresacrificed after only 6 and 12 h. RA-induced alterations werenot present in regions of the nasal airways lined by other nasalepithelial types including respiratory, transitional, or squamousepithelium.
RA-induced OE lesions at 1 day PI consisted of numerousindividual epithelial cells with morphologic features charac-teristic of apoptosis in the OE lining all the ethmoid turbinatesand the adjacent lateral walls that border the lateral meatus inthe distal regions of both nasal passages (T3 and T4), with themost dorsolateral ethmoid turbinates (1E and 2E) being mostseverely affected (Fig. 1B). RA-induced apoptosis was alsopresent in the OE of the mid and ventral septum lining themiddle medial meatus in T3, but not in T4. In the middle ofthe nasal passages (T2), prior to the distal regions containingthe ethmoid turbinates (T3 and T4), RA-induced apoptoticlesions in the OE were only detected in a small mucosal regionof the lateral walls and septum lining the middle medial meatuswhere the OE meets the respiratory epithelium. RA-inducednasal epithelial lesions were undetectable in the most proximalregions of the nasal passages (T1). Interestingly, the OE liningthe dorsal medial meatus throughout the nasal passages (T1–T4)had no microscopic evidence of RA-induced apoptosis of OEor any other epithelial alterations.
RA Induces OE Atrophy and Loss of OSNs
When effects of RA on OE effects (Fig. 2A–J) were assessedover time, RA-induced OE lesions and OE atrophy were firstdetectable at 24 h PI (Fig. 2 G,H) and to a much greater extentat 72 h PI (Figs. 2I and 2J). Morphometry of OE lining ethmoidturbinate 2 (region 2E, Fig. 1B) confirmed that greater atrophyoccurred in mice sacrificed 3 days PI than in mice sacrificed 1day PI, with an approximately 33% reduction in epithelialthickness compared to control mice (Fig. 2K).
OMP, a specific peptide found only in mature OSNs (Kreamand Margolis, 1984), was dramatically reduced in the OE ofRA-instilled mice compared to saline-instilled control mice(Figs. 3A and 3B). Consistent with OSN loss, immunohisto-chemical examination demonstrated a marked reduction of thenormally dense mat of cilia projecting from the dendritic knobsand lining the surface of the nasal airway lumen (Figs. 3C and3D). There was also noticeable atrophy of the OMP-positiveolfactory nerve bundles located in the lamina propria un-derlying the atrophic OE. Morphometric analysis revealed
FIG. 6. Immunohistochemical detection of activated caspase-3 expression
in the OE at 24 h after a single instillation of RA. Light photomicrographs of
OE lining E2 from mice intranasally instilled with saline-vehicle alone (A) or
RA (B). Tissues were immunohistochemically stained for activated caspase-3
(marker of cells undergoing apoptosis; dark cellular stain in OE) and
counterstained with hematoxylin. Numerous activated caspase-3–positive cells
(arrows) are present in (B), but not in (A). tb, turbinate bone. Color figures are
provided online as supplementary data.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 531
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

approximately a 90% loss in OSNs per mm of OE in the nasalmucosa lining the dorsolateral meatus at 3 days PI when com-pared to the identical region in vehicle-instilled mice (Fig. 4).
Concurrent with the loss of OMP-positive OSNs in the OEand lamina propria in the nasal mucosa, there was also markedbilateral atrophy and vacuolation of the outer tissue layer(olfactory nerve layer) of the OBs (Fig. 5A) in the brains ofthese RA-treated mice at 72 h PI. Loss of OMP-stained axonsof the OSNs in the brain was most evident in the lateral andmedial aspects of each OB (Figs. 5B and 5C). Loss of OMP-staining was also evident in the adjacent glomerular layer of theOB where the axons of OSNs first synapse with other neuronsin the brain.
RA Induces Apoptosis of OSN in OE at 24 h PI
RA-induced conspicuous apoptotic lesions in the OE at 24 h.Toxin-induced apoptosis was defined by light morphologicfeatures of condensation and shrinkage of individual epithelialcells; clumping, fragmentation, and margination of nuclear
chromatin; and numerous widely scattered cellular fragments(apoptotic bodies) (Figs. 2G, 2H, and 6A, 6B). Morphologic de-tection of RA-induced apoptosis in the OE corresponded withthe immunohistochemical detection of the activated caspase-3in many of the apoptotic cells (Figs. 6A and 6B). RA-inducedapoptosis was restricted to OSNs whose cell bodies and nucleireside in the middle nuclear layers of the OE below the distinctapical row of sustentacular (support) cell nuclei and above thebasal cell nuclei near the basal lamina. Prominent anti-caspase-3 staining was also present in olfactory nerve bundles (axons ofOSNs) in the underlying lamina propria of the mice instilledwith RA. Therefore, the marked RA-induced atrophy of OE,that was most prominent at 72 h PI, was likely a result ofwidespread apoptosis of OSN at 24 h with subsequent loss ofthese neuronal cells. Very few apoptotic cells or cellular frag-ments remained in the atrophic OE at 72 h PI, and there was noincrease in immunohistochemical detectable activated caspase-3, above that of saline-instilled controls, in these nasal tissuesat this later time point.
FIG. 7. RA induction of proapoptotic gene mRNAs in ethmoid turbinates and OB. mRNAs for proapoptotic genes Fas, PKR, p53, Bax, and CAD were
determined by real-time PCR of RNA from microisolated ethmoid turbinates of mice treated for various time intervals with saline vehicle (solid circle) or RA (500
lg/kg bw) (open circle). Data are means ± SEM (n ¼ 6). Data points designated with different letters are statistically different (p < 0.05).
532 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

RA Upregulates Proapoptotic gene Expression in theEthmoid Turbinates
Real-time PCR analysis of microdissected OE-lined ethmoidturbinates from RA-treated mice demonstrated a marked up-regulation of the proapoptotic genes Fas, PKR, p53, Bax, andCAD (Fig. 7). Fas mRNA expression was elevated as early as 6h PI, remained significantly elevated up to 24 h PI, but returnedto baseline control levels at 72 h PI. Although there was alsoa trend toward higher mRNA expression for PKR, p53, Bax,and CAD at 12 h, these genes were significantly elevated onlyat 24 h PI and returned to control values at 72 h PI. Thus,apoptotic death of OSNs in the OE was preceded by Fasexpression and concurrent with expression of these otherproapoptotic genes.
RA Induces Neutrophil Infiltration in OE
Along with the marked atrophy of OE at 72 h PI, RA-instilled mice also had prominent accumulations of mucopro-teinaceous material with exfoliated and degenerating cellulardebris from the dendritic portions of the apoptotic OSNs in thenasal airways adjacent to the luminal surfaces of the atrophyingOE. With secondary degeneration of these exfoliated dendritic
fragments, there was accompanying infiltration of numerousphagocytic cells consisting mainly of polymorphonuclear leu-kocytes (neutrophils) and only occasional mononuclear cells(monocytes and macrophages) (Figs. 2I, 3D, and 8A–D). In-filtrating neutrophils were also widely scattered in the atrophicOE and lamina propria of the RA-altered olfactory mucosa(Figs. 8C and 8D).
RA Upregulates Proinflammatory Gene Expression in theEthmoid Turbinates and the OB
In the previous study with satratoxin G, there was a robustinfiltration of mucosubstances filled with neutrophils in thenasal airways and sporadic incidences of neutrophils in the OB(Islam et al., 2006). Therefore, proinflammatory cytokines andthe neutrophil chemoattractive protein MIP-2 were measuredin the tissues from ethmoid turbinate (ET) and OB. The resultsshowed that there was markedly increased expression ofmRNAs for the proinflammatory cytokines TNF-a, IL-1a,IL-1b, IL-6, and MIP-2 in the ethmoid turbinates of miceintranasally instilled with RA (Fig. 9A). Except for IL-1a,upregulation of these mRNAs was evident as early as 6 h PI.Expression of all genes remained elevated at 12 and 24 h PI and
FIG. 8. RA-induced neutrophilic rhinitis at 72 h PI. Light photomicrographs of the OE lining ethmoid turbinate 3 (E3) and the adjacent lateral wall (LW) in the
nasal airways of mice intranasally instilled with saline-vehicle alone (A, B) or RA (C, D). Rectangles in (A) and (C) represent the sites from which (B) and (D) were
taken. Tissues were immunohistochemically stained with an anti-neutrophil–specific antibody to detect infiltrating neutrophils (dark cellular stain) in the nasal
mucosa, and counterstained with hematoxylin. Numerous infiltrating neutrophils (arrows) are present in the nasal mucosa (OE and lamina propria) and nasal
airway lumen (lateral meatus; asterisk) in (C, D) compared to (A, B). Color figures are provided online as supplementary data.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 533
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
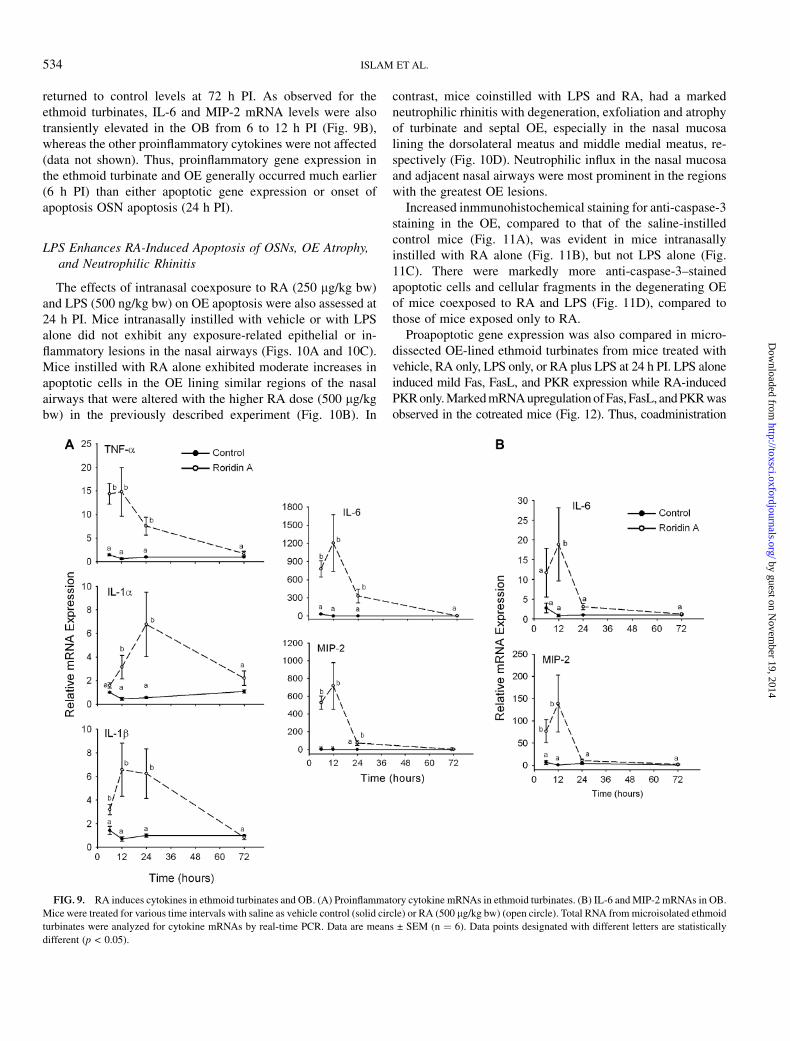
returned to control levels at 72 h PI. As observed for theethmoid turbinates, IL-6 and MIP-2 mRNA levels were alsotransiently elevated in the OB from 6 to 12 h PI (Fig. 9B),whereas the other proinflammatory cytokines were not affected(data not shown). Thus, proinflammatory gene expression inthe ethmoid turbinate and OE generally occurred much earlier(6 h PI) than either apoptotic gene expression or onset ofapoptosis OSN apoptosis (24 h PI).
LPS Enhances RA-Induced Apoptosis of OSNs, OE Atrophy,and Neutrophilic Rhinitis
The effects of intranasal coexposure to RA (250 lg/kg bw)and LPS (500 ng/kg bw) on OE apoptosis were also assessed at24 h PI. Mice intranasally instilled with vehicle or with LPSalone did not exhibit any exposure-related epithelial or in-flammatory lesions in the nasal airways (Figs. 10A and 10C).Mice instilled with RA alone exhibited moderate increases inapoptotic cells in the OE lining similar regions of the nasalairways that were altered with the higher RA dose (500 lg/kgbw) in the previously described experiment (Fig. 10B). In
contrast, mice coinstilled with LPS and RA, had a markedneutrophilic rhinitis with degeneration, exfoliation and atrophyof turbinate and septal OE, especially in the nasal mucosalining the dorsolateral meatus and middle medial meatus, re-spectively (Fig. 10D). Neutrophilic influx in the nasal mucosaand adjacent nasal airways were most prominent in the regionswith the greatest OE lesions.
Increased inmmunohistochemical staining for anti-caspase-3staining in the OE, compared to that of the saline-instilledcontrol mice (Fig. 11A), was evident in mice intranasallyinstilled with RA alone (Fig. 11B), but not LPS alone (Fig.11C). There were markedly more anti-caspase-3–stainedapoptotic cells and cellular fragments in the degenerating OEof mice coexposed to RA and LPS (Fig. 11D), compared tothose of mice exposed only to RA.
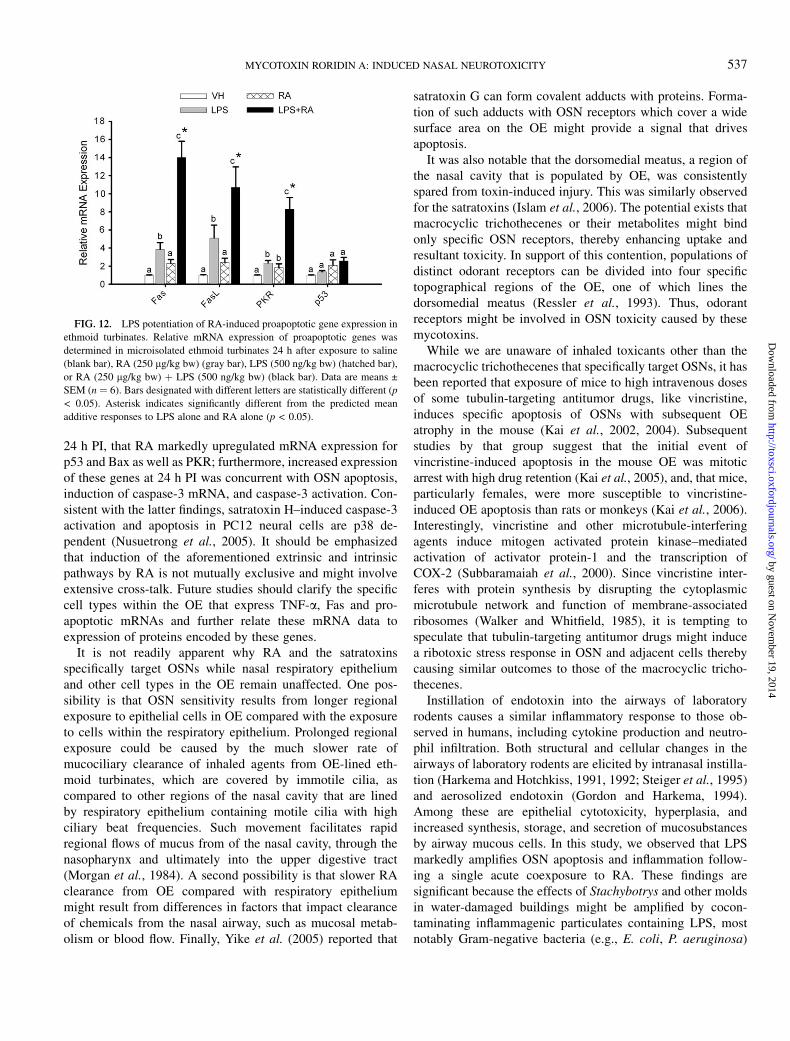
Proapoptotic gene expression was also compared in micro-dissected OE-lined ethmoid turbinates from mice treated withvehicle, RA only, LPS only, or RA plus LPS at 24 h PI. LPS aloneinduced mild Fas, FasL, and PKR expression while RA-inducedPKR only. Marked mRNA upregulation of Fas, FasL, and PKR wasobserved in the cotreated mice (Fig. 12). Thus, coadministration
FIG. 9. RA induces cytokines in ethmoid turbinates and OB. (A) Proinflammatory cytokine mRNAs in ethmoid turbinates. (B) IL-6 and MIP-2 mRNAs in OB.
Mice were treated for various time intervals with saline as vehicle control (solid circle) or RA (500 lg/kg bw) (open circle). Total RNA from microisolated ethmoid
turbinates were analyzed for cytokine mRNAs by real-time PCR. Data are means ± SEM (n ¼ 6). Data points designated with different letters are statistically
different (p < 0.05).
534 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

of RA and LPS caused greater proapoptotic gene expression andapoptosis in the OE than either toxin alone.
Based on the prior kinetic study with RA, 12 h PI was chosento compare proinflammatory cytokine gene expression in miceinstilled with LPS and/or RA (Fig. 13). LPS but not RA aloneinduced TNF-a, IL-1a, and IL-1b in ethmoid turbinates(Fig. 13A) and OB (Fig. 13B). There was a trend towardelevated expression of all three cytokines following cotreat-ment with RA and LPS, but these changes were not significantcompared to LPS alone. IL-6 and MIP-2 mRNAs weremarginally induced by either LPS or RA alone in both ethmoidturbinates (Fig. 13A) and OB (Fig. 13B). However, coexposureto both agents dramatically elevated these genes as comparedto either toxin alone. Thus, LPS could magnify OSN apopto-sis and inflammation following a single acute coexposure to RA.
DISCUSSION
As a portal of entry of inhaled air, the nose is a prime targetfor airborne toxic and infectious agents. Surface epithelial cellpopulations lining the nasal passages are often the initial targetsfor inhaled toxicants. Morphologically and functionally dis-tinct epithelia line the mammalian nasal passages that include(1) squamous epithelium, (2) ciliated, pseudostratified, re-
spiratory epithelium, (3) nonciliated transitional epithelium,and (4) OE, located in the dorsal or dorsoposterior aspect ofthe nasal cavity (Harkema et al., 2006). Very low satratoxinG doses (as little as 25 lg/kg bw or 0.5 lg/mouse) causeapoptosis of OSNs of the OE in the mouse as evidencedby morphologic examination at the light and electron micro-scopic levels, immunohistochemical detection of activatedcaspase-3, and proapoptotic gene expression (Islam et al.,2006). Concurrently, satratoxin G causes rhinitis and a mildfocal encephalitis in the OB of the brain. Yike and Dearborn(2004) estimated that a single S. chartarum spore contains 1 pgequivalent of satratoxin G, suggesting that a single exposure to5 3 105 spores could deliver sufficient toxin in the mouse toevoke olfactory toxicity. The results presented herein indicatethat the related macrocyclic trichothecene RA similarly targetsOSNs and can also initiate inflammatory responses in the noseand OB. It should be noted that approximately 10 times moreRA than satratoxin G was required to achieve equivalent ef-fects. Nevertheless, RA should be a suitable surrogate to un-cover underlying mechanisms of macrocyclic trichothecenetoxicity in the nasal airway and to investigate interactive effectswith endotoxin, a co-occurring airborne toxicant.
The OE and OSNs are vulnerable to injury and death fromdirect exposure to inhaled toxins and infectious agents. OEundergoes continuous neuronal cell renewal throughout the life
FIG. 10. LPS enhancement of RA-induced nasal toxicity at 24 h PI. Light photomicrographs of the OE lining the midseptum in the nasal tissue section T3 from
mice intranasally instilled with saline-vehicle alone (A), RA alone (B), LPS alone (C), or LPS and RA (D). No exposure-related alterations to the OE are present in
(A) or (C). Mild atrophy of OE containing apoptotic cells and cellular fragments (arrows) is present in (B). In (D) there is marked degeneration, exfoliation, and
atrophy of OE with apoptotic cells (arrows) and a concomitant mucopurulent airway exudate (asterisk) containing mainly neutrophils and epithelial cellular debris.
Tissues were stained with hematoxylin and eosin. Scale bars ¼ 50 lm. Color figures are provided online at supplementary data.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 535
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

span of an individual. Its regenerative ability following injuryenables the OE to maintain olfactory sensory function. Com-pared to other neurons, OSNs are unique in that they haverelatively short life spans (30–40 days or longer) and are con-tinuously being replaced through basal cell proliferation anddifferentiation (neuronal regeneration) (Graziadei and Monti-Graziadei, 1978). Similar to other epithelial cells, but unlikemost neuronal cells, OSNs undergo continuous apoptosis andgenesis throughout life as part of the normal turnover of matureOE. Experimentally, OSNs in laboratory animals can beinduced to die via manipulative methods such as olfactorybulbectomy, transection of the olfactory nerve at the cribiformplate, and intranasal exposure to chemicals known to be toxicto the OE, such as zinc sulfate and methyl bromide (Cowan andRoskams, 2002, 2004). While bulbectomy and nerve transec-tion evoke selective apoptosis in death of OSNs, most chemicaltoxins cause necrosis (oncosis) of the OSNs along with otherepithelial cells (sustentacular and basal cells) in the OE. Thus,it is quite remarkable that RA, as described here, and thesatratoxins, as reported previously (Islam et al., 2006), areexquisitely specific in initiating OSN apoptosis and OE atrophyin the nose and in the OB.
RA might induce OSN apoptosis in two ways. One plausiblemechanism is via death receptors. OSN apoptosis has beenpreviously linked to specific genes associated extrinsic deathreceptor pathways involving TNF-a and Fas (Cowan andRoskams, 2002). Both TNF-a and Fas directly induce apopto-sis in in vitro OE organ cultures (Farbman et al., 1999; Suzukiand Farbman, 2000). Consistent with this mechanism, the re-sults presented here indicated that RA-induced TNF-a and FasmRNA expression from 6 to 24 h PI which preceded or wasconcurrent with OSN apoptosis, induction of caspase-3 mRNAand caspase-3 activation at 24 h PI. The origins of inducedTNF-a might be OSNs or adjacent cells in the OE which wouldpromote autocrine or paracrine responses, respectively. For Fasupregulation to have an effect it would have to occur within thesusceptible OSN population. A second potential mechanism isthat RA directly induces apoptosis in the OSNs by initiatingmitochondrial cell death via an intrinsic pathway involving p53and Bax. Previously, we have demonstrated in macrophagesthat the trichothecene deoxynivalenol induced p38-dependentapoptosis involving p53, Bax, and caspase-3 (Zhou et al., 2005)and furthermore, that PKR plays a critical role in p38 activationas well as apoptosis (Zhou et al., 2003). Here, it was shown, at
FIG. 11. LPS enhancement of RA-induced expression of activated caspase-3 in the apoptotic OSNs of the OE at 24 h PI. (A–D) Light photomicrographs of the OE
lining the midseptum in the nasal tissue section T3 from mice intranasally instilled with saline-vehicle alone (A), RA alone (B), LPS alone (C), or LPS and RA (D).
Tissues were immunohistochemically stained for activated caspase-3 (marker of cells undergoing apoptosis; dark cellular stain) and counterstained with hematoxylin.
No exposure-related alterations to the OE are present in (A) or (C) (arrow indicates a single activated caspase-3–positive OSN). Mild atrophy of OE containing a few
activated caspase–positive apoptotic cells and cellular fragments (white arrows) is present in (B). In (D) there is marked degeneration, exfoliation, and atrophy of OE
with numerous activated caspase-3–positive apoptotic cells (white arrows). Olfactory nerve bundles (ON), containing the axonal portion of OSNs, are also positively
stained for activated caspase-3 (black arrows) in (B) and (D), but not in (A) and (C). Scale bars ¼ 50 lm. Color figures are provided online as supplementary data.
536 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
24 h PI, that RA markedly upregulated mRNA expression forp53 and Bax as well as PKR; furthermore, increased expressionof these genes at 24 h PI was concurrent with OSN apoptosis,induction of caspase-3 mRNA, and caspase-3 activation. Con-sistent with the latter findings, satratoxin H–induced caspase-3activation and apoptosis in PC12 neural cells are p38 de-pendent (Nusuetrong et al., 2005). It should be emphasizedthat induction of the aforementioned extrinsic and intrinsicpathways by RA is not mutually exclusive and might involveextensive cross-talk. Future studies should clarify the specificcell types within the OE that express TNF-a, Fas and pro-apoptotic mRNAs and further relate these mRNA data toexpression of proteins encoded by these genes.
It is not readily apparent why RA and the satratoxinsspecifically target OSNs while nasal respiratory epitheliumand other cell types in the OE remain unaffected. One pos-sibility is that OSN sensitivity results from longer regionalexposure to epithelial cells in OE compared with the exposureto cells within the respiratory epithelium. Prolonged regionalexposure could be caused by the much slower rate ofmucociliary clearance of inhaled agents from OE-lined eth-moid turbinates, which are covered by immotile cilia, ascompared to other regions of the nasal cavity that are linedby respiratory epithelium containing motile cilia with highciliary beat frequencies. Such movement facilitates rapidregional flows of mucus from of the nasal cavity, through thenasopharynx and ultimately into the upper digestive tract(Morgan et al., 1984). A second possibility is that slower RAclearance from OE compared with respiratory epitheliummight result from differences in factors that impact clearanceof chemicals from the nasal airway, such as mucosal metab-olism or blood flow. Finally, Yike et al. (2005) reported that
satratoxin G can form covalent adducts with proteins. Forma-tion of such adducts with OSN receptors which cover a widesurface area on the OE might provide a signal that drivesapoptosis.
It was also notable that the dorsomedial meatus, a region ofthe nasal cavity that is populated by OE, was consistentlyspared from toxin-induced injury. This was similarly observedfor the satratoxins (Islam et al., 2006). The potential exists thatmacrocyclic trichothecenes or their metabolites might bindonly specific OSN receptors, thereby enhancing uptake andresultant toxicity. In support of this contention, populations ofdistinct odorant receptors can be divided into four specifictopographical regions of the OE, one of which lines thedorsomedial meatus (Ressler et al., 1993). Thus, odorantreceptors might be involved in OSN toxicity caused by thesemycotoxins.
While we are unaware of inhaled toxicants other than themacrocyclic trichothecenes that specifically target OSNs, it hasbeen reported that exposure of mice to high intravenous dosesof some tubulin-targeting antitumor drugs, like vincristine,induces specific apoptosis of OSNs with subsequent OEatrophy in the mouse (Kai et al., 2002, 2004). Subsequentstudies by that group suggest that the initial event ofvincristine-induced apoptosis in the mouse OE was mitoticarrest with high drug retention (Kai et al., 2005), and, that mice,particularly females, were more susceptible to vincristine-induced OE apoptosis than rats or monkeys (Kai et al., 2006).Interestingly, vincristine and other microtubule-interferingagents induce mitogen activated protein kinase–mediatedactivation of activator protein-1 and the transcription ofCOX-2 (Subbaramaiah et al., 2000). Since vincristine inter-feres with protein synthesis by disrupting the cytoplasmicmicrotubule network and function of membrane-associatedribosomes (Walker and Whitfield, 1985), it is tempting tospeculate that tubulin-targeting antitumor drugs might inducea ribotoxic stress response in OSN and adjacent cells therebycausing similar outcomes to those of the macrocyclic tricho-thecenes.
Instillation of endotoxin into the airways of laboratoryrodents causes a similar inflammatory response to those ob-served in humans, including cytokine production and neutro-phil infiltration. Both structural and cellular changes in theairways of laboratory rodents are elicited by intranasal instilla-tion (Harkema and Hotchkiss, 1991, 1992; Steiger et al., 1995)and aerosolized endotoxin (Gordon and Harkema, 1994).Among these are epithelial cytotoxicity, hyperplasia, andincreased synthesis, storage, and secretion of mucosubstancesby airway mucous cells. In this study, we observed that LPSmarkedly amplifies OSN apoptosis and inflammation follow-ing a single acute coexposure to RA. These findings aresignificant because the effects of Stachybotrys and other moldsin water-damaged buildings might be amplified by cocon-taminating inflammagenic particulates containing LPS, mostnotably Gram-negative bacteria (e.g., E. coli, P. aeruginosa)
FIG. 12. LPS potentiation of RA-induced proapoptotic gene expression in
ethmoid turbinates. Relative mRNA expression of proapoptotic genes was
determined in microisolated ethmoid turbinates 24 h after exposure to saline
(blank bar), RA (250 lg/kg bw) (gray bar), LPS (500 ng/kg bw) (hatched bar),
or RA (250 lg/kg bw) þ LPS (500 ng/kg bw) (black bar). Data are means ±
SEM (n ¼ 6). Bars designated with different letters are statistically different (p
< 0.05). Asterisk indicates significantly different from the predicted mean
additive responses to LPS alone and RA alone (p < 0.05).
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 537
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
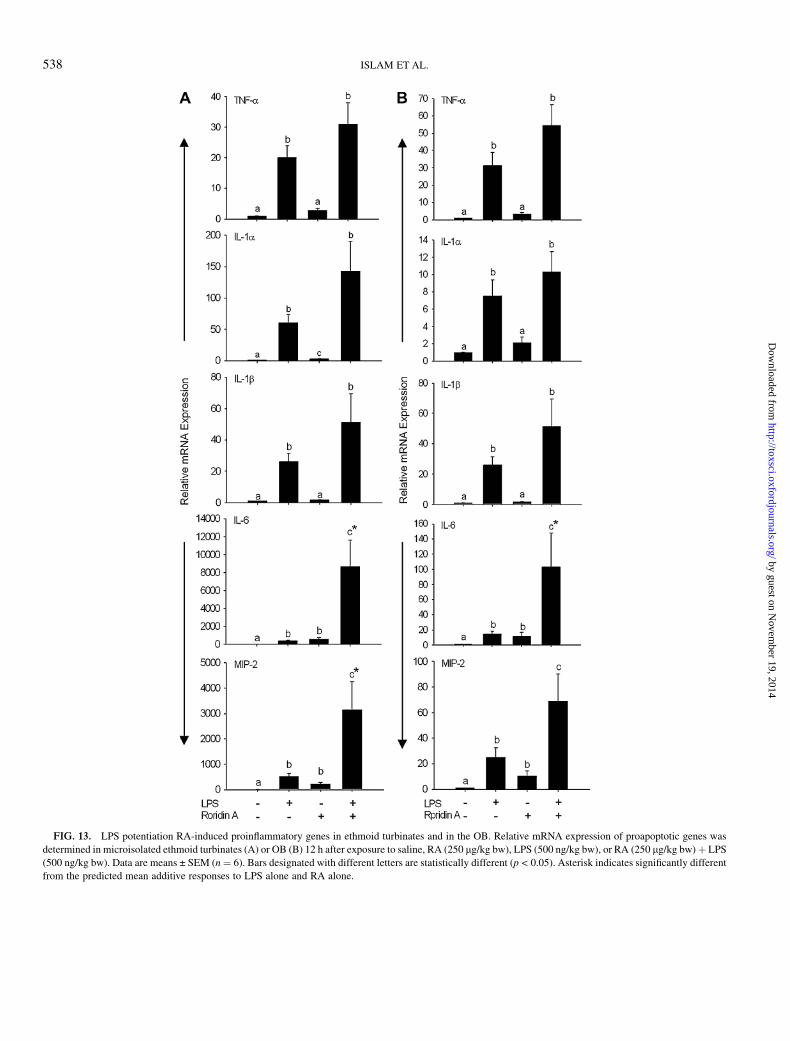
FIG. 13. LPS potentiation RA-induced proinflammatory genes in ethmoid turbinates and in the OB. Relative mRNA expression of proapoptotic genes was
determined in microisolated ethmoid turbinates (A) or OB (B) 12 h after exposure to saline, RA (250 lg/kg bw), LPS (500 ng/kg bw), or RA (250 lg/kg bw) þ LPS
(500 ng/kg bw). Data are means ± SEM (n¼ 6). Bars designated with different letters are statistically different (p < 0.05). Asterisk indicates significantly different
from the predicted mean additive responses to LPS alone and RA alone.
538 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

(Andersson et al., 1997; Park et al., 2006). Potentiation ofapoptosis by LPS appeared to correlate with increased Fas,FasL, and PKR, but not with TNF-a or IL-1 mRNAs, in the OE.The increased neutrophilic rhinitis that occurred with RA andLPS coexposure also correlated with increased IL-6 and MIP-2expression.
Consistent with the findings of this study, the trichothecenesT-2 and deoxynivalenol become more toxic in the presence ofendotoxin, thereby causing elevated tissue injury and mortality(Tai and Pestka, 1988a,b; Taylor et al., 1989, 1991; Zhou et al.,1999, 2000). Coexposure of mice to subtoxic doses of LPS anddeoxynivalenol markedly upregulates proinflammatory cyto-kine expression and subsequently induces apoptosis in thymus,Peyer’s patches, bone marrow, and spleen (Islam et al., 2002).Remarkably, a single LPS exposure can sensitize the innateimmune system to trichothecene-induced lymphoid apoptosisfor at least 24 h (Islam and Pestka, 2006).
Taken together, our observations here and previously that theOE and OB are specific targets of macrocyclic trichothecenesshould be a critical consideration in future studies of dampbuilding–related illnesses and the potential etiologic role ofS. chartarum and other molds. Acute inflammation of nose(allergic rhinitis) and brain (encephalitis) are significant healthproblems that can affect many aspects of daily life because ofphysical discomfort and impairment along with emotionaldistress (Baiardini et al., 2006; Olgar et al., 2006). The genesupregulated here could likely contribute to OSN apoptosis aswell as accompanying rhinitis and even mild encephalitis. Theobservation that LPS exacerbated RA-induced proinflamma-tory gene expression, apoptosis, and inflammation in the OEsuggests that co-occurring inflammagens have the potential tointeract with macrocylic trichothecenes in evoking even moresevere pathologic effects in the nasal cavity. While the datapresented herein and previously (Islam et al., 2006) provideimportant insight into targets and mechanisms, future perspec-tives must include assessment of dose dependency and latencyof recovery in nasal tissue after chronic exposure to macrocy-clic trichothecenes alone, as well as the contributions of sporematrix and co-occurring inflammagens found in indoor air. Itwill also be critically important to understand the extent towhich toxicant-induced inflammation and neuronal injuryoccur in other parts of the brain along the olfactory pathwayand whether such injury could contribute to neurocognitivedysfunction. Finally, all mechanistic findings must be consid-ered in the context of accurate quantitative assessments ofhuman exposure to trichothecenes using both state-of-the-artsampling and analytical methods and relevant biomarkers.
SUPPLEMENTARY DATA
Supplementary data are available online at http://toxsci.oxfordjournals.org/.
ACKNOWLEDGMENTS
This research was funded by a Michigan State University Foundation
Strategic Partnership Grant and Public Health Service Grant ES03358 (J.J.P.)
from the National Institute for Environmental Health Sciences. We thank Lori
Bramble, Sarah Godbehere, Amy Porter, Rick Rosebury, Kathleen Campbell,
and Dr Annette Thelen for technical assistance.
REFERENCES
Andersson, M. A., Nikulin, M., Koljalg, U., Andersson, M. C., Rainey, F.,
Reijula, K., Hintikka, E. L., and Salkinoja-Salonen, M. (1997). Bacteria,
molds, and toxins in water-damaged building materials. Appl. Environ.
Microbiol. 63, 387–393.
Audige, A., Yu, Z. R., Frey, B. M., Uehlinger, D. E., Frey, F. J., and Vogt, B.
(2003). Epithelial sodium channel (ENaC) subunit mRNA and protein
expression in rats with puromycin aminonucleoside-induced nephrotic
syndrome. Clin. Sci. (Lond.) 104, 389–395.
Baiardini, I., Braido, F., Brandi, S., and Canonica, G. W. (2006). Allergic
diseases and their impact on quality of life. Ann. Allergy Asthma Immunol.
97, 419–428.
Bornehag, C. G., Sundell, J., and Sigsgaard, T. (2004). Dampness in buildings
and health (DBH): Report from an ongoing epidemiological investigation on
the association between indoor environmental factors and health effects
among children in Sweden. Indoor Air 14(Suppl. 7), 59–66.
Boutin-Forzano, S., Charpin-Kadouch, C., Chabbi, S., Bennedjai, N., Dumon, H.,
and Charpin, D. (2004). Wall relative humidity: A simple and reliable index
for predicting Stachybotrys chartarum infestation in dwellings. Indoor Air
14, 196–199.
Brasel, T. L., Douglas, D. R., Wilson, S. C., and Straus, D. C. (2005). Detection
of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins
on particulates smaller than conidia. Appl. Environ. Microbiol. 71, 114–122.
Chung, Y. J., Jarvis, B., and Pestka, J. (2003a). Modulation of lipopolysaccha-
ride-induced proinflammatory cytokine production by satratoxins and other
macrocyclic trichothecenes in the murine macrophage. J. Toxicol. Environ.
Health A 66, 379–391.
Chung, Y. J., Yang, G. H., Islam, Z., and Pestka, J. J. (2003b). Up-regulation of
macrophage inflammatory protein-2 and complement 3A receptor by the
trichothecenes deoxynivalenol and satratoxin G. Toxicology 186, 51–65.
Cowan, C. M., and Roskams, A. J. (2002). Apoptosis in the mature and
developing olfactory neuroepithelium. Microsc. Res. Tech. 58, 204–215.
Cowan, C. M., and Roskams, A. J. (2004). Caspase-3 and caspase-9 mediate
developmental apoptosis in the mouse olfactory system. J. Comp. Neurol.
474, 136–148.
Cox-Ganser, J. M., White, S. K., Jones, R., Hilsbos, K., Storey, E., Enright, P. L.,
Rao, C. Y., and Kreiss, K. (2005). Respiratory morbidity in office workers in
a water-damaged building. Environ. Health Perspect. 113, 485–490.
Farbman, A. I., Buchholz, J. A., Suzuki, Y., Coines, A., and Speert, D. (1999).
A molecular basis of cell death in olfactory epithelium. J. Comp. Neurol.
414, 306–314.
Fog Nielsen, K. (2003). Mycotoxin production by indoor molds. Fungal Genet.
Biol. 39, 103–117.
Foto, M., Vrijmoed, L. L., Miller, J. D., Ruest, K., Lawton, M., and Dales, R. E.
(2005). A comparison of airborne ergosterol, glucan and Air-O-Cell data in
relation to physical assessments of mold damage and some other parameters.
Indoor Air 15, 257–266.
Fung, F., Clark, R., and Williams, S. (1998). Stachybotrys, a mycotoxin-
producing fungus of increasing toxicologic importance. J. Toxicol. Clin.
Toxicol. 36, 79–86.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 539
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

Gordon, T., and Harkema, J. R. (1994). Effect of inhaled endotoxin on
intraepithelial mucosubstances in F344 rat nasal and tracheobronchial
airways. Am. J. Respir. Cell Mol. Biol. 10, 177–183.
Gordon, W. A. (1999) Cognitive impairment associated with exposure to
toxigenic fungi. In Bioaerosols, Fungi and Mycotoxins: Health Effects,
Assessment, Prevention and Control, Chapter I (E. Johanning, Ed.), pp.
94–98. Eastern New York Occupational and Environmental Health Center,
Albany, New York.
Graziadei, P. P., and Monti-Graziadei, G. A. (1978). The Olfactory System:
A Model for the Study of Neurogenesis and Axon Regeneration in Mammals.
Raven Press, New York.
Gregory, L., Pestka, J. J., Dearborn, D. G., and Rand, T. G. (2004). Localization
of satratoxin-G in Stachybotrys chartarum spores and spore-impacted mouse
lung using immunocytochemistry. Toxicol. Pathol. 32, 26–34.
Grove, J. F. (1988). Non-macrocyclic trichothecenes. Nat. Prod. Rep. 5, 187–209.
Grove, J. F. (1993). Macrocyclic trichothecenes. Nat. Prod. Rep. 10, 429–448.
Grove, J. F. (2000). Non-macrocyclic trichothecenes. Part 2. Prog. Chem. Org.
Nat. Prod. 69, 1–70.
Harkema, J. R., Carey, S. A., and Wagner, J. G. (2006). The nose revisited:
A brief review of the comparative structure, function, and toxicologic
pathology of the nasal epithelium. Toxicol. Pathol. 34, 252–269.
Harkema, J. R., and Hotchkiss, J. A. (1991). In vivo effects of endotoxin on
nasal epithelial mucosubstances: Quantitative histochemistry. Exp. Lung
Res. 17, 743–761.
Harkema, J. R., and Hotchkiss, J. A. (1992). In vivo effects of endotoxin on
intraepithelial mucosubstances in rat pulmonary airways. Quantitative
histochemistry. Am. J. Pathol. 141, 307–317.
Heinrich, J., Holscher, B., Douwes, J., Richter, K., Koch, A., Bischof, W.,
Fahlbusch, B., Kinne, R. W., and Wichmann, H. E. (2003). Reproducibility
of allergen, endotoxin and fungi measurements in the indoor environment.
J. Expo. Anal. Environ. Epidemiol. 13, 152–160.
Hinkley, S. F., and Jarvis, B. B. (2001). Chromatographic method for
Stachybotrys toxins. Methods Mol. Biol. 157, 173–194.
Hossain, M. A., Ahmed, M. S., and Ghannoum, M. A. (2004). Attributes of
Stachybotrys chartarum and its association with human disease. J. Allergy
Clin. Immunol. 113, 200–208.
Hyde, D. M., Magliano, D. J., and Plopper, C. G. (1991). Morphometric
assessment of pulmonary toxicity in the rodent lung. Toxicol. Pathol. 19(4 Pt 1),
428–446.
Hyde, D. M., Plopper, C. G., St George, J. A., and Harkema, J. R. (1990).
Morphometric cell biology of air space epithelium. In Electron Microscopy
of the Lung, (Schraufnagel DE, Ed.) pp. 1–120. Marcel Dekker, New York.
Institute of Medicine. (2004). Damp Indoor Spaces and Health. National
Academies Press, Washington, D.C.
Islam, Z., Harkema, J. R., and Pestka, J. J. (2006). Satratoxin G from the black
mold Stachybotrys chartarum evokes olfactory sensory neuron loss and
inflammation in the murine nose and brain. Environ. Health Perspect. 114,
1099–1107.
Islam, Z., Moon, Y. S., Zhou, H. R., King, L. E., Fraker, P. J., and Pestka, J. J.
(2002). Endotoxin potentiation of trichothecene-induced lymphocyte apo-
ptosis is mediated by up-regulation of glucocorticoids. Toxicol. Appl.
Pharmacol. 180, 43–55.
Islam, Z., and Pestka, J. J. (2006). LPS priming potentiates and prolongs
proinflammatory cytokine response to the trichothecene deoxynivalenol in
the mouse. Toxicol. Appl. Pharmacol. 211, 53–63.
Jarvis, B. B., Lee, Y. W., Comezoglu, S. N., and Yatawara, C. S. (1986).
Trichothecenes produced by Stachybotrys atra from Eastern Europe. Appl.
Environ. Microbiol. 51, 915–918.
Jarvis, B. B., Sorenson, W. G., Hintikka, E. L., Nikulin, M., Zhou, Y., Jiang, J.,
Wang, S., Hinkley, S., Etzel, R. A., and Dearborn, D. (1998). Study of toxin
production by isolates of Stachybotrys chartarum and Memnoniella echinata
isolated during a study of pulmonary hemosiderosis in infants. Appl. Environ.
Microbiol. 64, 3620–3625.
Johanning, E., Biagini, R., Hull, D., Morey, P., Jarvis, B., and Landsbergis, P.
(1996). Health and immunology study following exposure to toxigenic fungi
(Stachybotrys chartarum) in a water-damaged office environment. Int. Arch.
Occup. Environ. Health 68, 207–218.
Kai, K., Sahto, H., Yoshida, M., Suzuki, T., Shikanai, Y., Kajimura, T., and
Furuhama, K. (2006). Species and sex differences in susceptibility to
olfactory lesions among the mouse, rat and monkey following an intravenous
injection of vincristine sulphate. Toxicol. Pathol. 34, 223–231.
Kai, K., Satoh, H., Kajimura, T., Kato, M., Uchida, K., Yamaguchi, R.,
Tateyama, S., and Furuhama, K. (2004). Olfactory epithelial lesions induced
by various cancer chemotherapeutic agents in mice. Toxicol. Pathol. 32,
701–709.
Kai, K., Satoh, H., Kashimoto, Y., Kajimura, T., and Furuhama, K. (2002).
Olfactory epithelium as a novel toxic target following an intravenous
administration of vincristine to mice. Toxicol. Pathol. 30, 306–311.
Kai, K., Yoshida, M., Sugawara, T., Kato, M., Uchida, K., Yamaguchi, R.,
Tateyama, S., and Furuhuma, K. (2005). Investigation of initial changes in
the mouse olfactory epithelium following a single intravenous injection of
vincristine sulphate. Toxicol. Pathol. 33, 750–759.
Kilburn, K. H. (2004). Role of molds and mycotoxins in being sick in buildings:
Neurobehavioral and pulmonary impairment. Adv. Appl. Microbiol. 55,
339–359.
Kovesi, T., Creery, D., Gilbert, N. L., Dales, R., Fugler, D., Thompson, B.,
Randhawa, N., and Miller, J. D. (2006). Indoor air quality risk factors for
severe lower respiratory tract infections in Inuit infants in Baffin Region,
Nunavut: A pilot study. Indoor Air 16, 266–275.
Kream, R. M., and Margolis, F. L. (1984). Olfactory marker protein: Turn-
over and transport in normal and regenerating neurons. J. Neurosci. 4,
868–879.
Mbandi, E., and Pestka, J. J. (2006). Deoxynivalenol and satratoxin
G potentiate proinflammatory cytokine and macrophage inhibitory protein
2 induction by Listeria and Salmonella in the macrophage. J. Food Prot. 69,
1334–1339.
Mery, S., Gross, E. A., Joyner, D. R., Godo, M., and Morgan, K. T. (1994).
Nasal diagrams: A tool for recording the distribution of nasal lesions in rats
and mice. Toxicol. Pathol. 22, 353–372.
Morgan, K. T., Jiang, X. Z., Patterson, D. L., and Gross, E. A. (1984). The nasal
mucociliary apparatus. Correlation of structure and function in the rat. Am.
Rev. Respir. Dis. 130, 275–281.
Nusuetrong, P., Yoshida, M., Tanitsu, M. A., Kikuchi, H., Mizugaki, M.,
Shimazu, K., Pengsuparp, T., Meksuriyen, D., Oshima, Y., and Nakahata, N.
(2005). Involvement of reactive oxygen species and stress-activated MAPKs
in satratoxin H-induced apoptosis. Eur. J. Pharmacol. 507, 239–246.
Olgar, S., Ertugrul, T., Nisli, K., Aydin, K., and Caliskan, M. (2006). Influenza
A-associated acute necrotizing encephalopathy. Neuropediatrics 37,
166–168.
Park, J. H., Cox-Ganser, J., Rao, C., and Kreiss, K. (2006). Fungal and
endotoxin measurements in dust associated with respiratory symptoms in
a water-damaged office building. Indoor Air 16, 192–203.
Rand, T. G., Mahoney, M., White, K., and Oulton, M. (2002). Microanatomical
changes in alveolar type II cells in juvenile mice intratracheally exposed to
Stachybotrys chartarum spores and toxin. Toxicol. Sci. 65, 239–245.
Ressler, K. J., Sullivan, S. L., and Buck, L. B. (1993). A zonal organization of
odorant receptor gene expression in the olfactory epithelium. Cell 73,
597–609.
Sebastian, A., and Larsson, L. (2003). Characterization of the microbial
community in indoor environments: A chemical-analytical approach. Appl.
Environ. Microbiol. 69, 3103–3109.
540 ISLAM ET AL.
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from

Sebastian, A., Szponar, B., and Larsson, L. (2005). Characterization of the
microbial community in indoor environments by chemical marker analysis:
An update and critical evaluation. Indoor Air 15(Suppl. 9), 20–26.
Solomon, G. M., Hjelmroos-Koski, M., Rotkin-Ellman, M., and Hammond, S. K.
(2006). Airborne mold and endotoxin concentrations in New Orleans,
Louisiana, after flooding, October through November 2005. Environ. Health
Perspect. 114, 1381–1386.
Sorenson, W. G., Frazer, D. G., Jarvis, B. B., Simpson, J., and Robinson, V. A.
(1987). Trichothecene mycotoxins in aerosolized conidia of Stachybotrys atra.
Appl. Environ. Microbiol. 53, 1370–1375.
Steiger, D., Hotchkiss, J., Bajaj, L., Harkema, J., and Basbaum, C. (1995).
Concurrent increases in the storage and release of mucin-like molecules by
rat airway epithelial cells in response to bacterial endotoxin. Am. J. Respir.
Cell Mol. Biol. 12, 307–314.
Subbaramaiah, K., Hart, J. C., Norton, L., and Dannenberg, A. J. (2000).
Microtubule-interfering agents stimulate the transcription of cyclooxyge-
nase-2. Evidence for involvement of ERK1/2 AND p38 mitogen-activated
protein kinase pathways. J. Biol. Chem. 275, 14838–14845.
Suzuki, Y., and Farbman, A. I. (2000). Tumor necrosis factor-alpha-induced
apoptosis in olfactory epithelium in vitro: Possible roles of caspase 1 (ICE),
caspase 2 (ICH-1), and caspase 3 (CPP32). Exp. Neurol. 165, 35–45.
Tai, J. H., and Pestka, J. J. (1988a). Synergistic interaction between the
trichothecene T-2 toxin and Salmonella typhimurium lipopolysaccharide in
C3H/HeN and C3H/HeJ mice. Toxicol. Lett. 44, 191–200.
Tai, J. H., and Pestka, J. J. (1988b). Impaired murine resistance to Salmonella
typhimurium following oral exposure to the trichothecene T-2 toxin. Food
Chem. Toxicol. 26, 691–698.
Taylor, M. J., Lafarge-Frayssinet, C., Luster, M. I., and Frayssinet, C. (1991).
Increased endotoxin sensitivity following T-2 toxin treatment is associated
with increased absorption of endotoxin. Toxicol. Appl. Pharmacol. 109, 51–59.
Taylor, M. J., Smart, R. A., and Sharma, R. P. (1989). Relationship of the
hypothalamic-pituitary-adrenal axis with chemically induced immunomodula-
tion. I. Stress-like response after exposure to T-2 toxin.Toxicology56, 179–195.
Tuomi, T., Reijula, K., Johnsson, T., Hemminki, K., Hintikka, E. L., Lindroos, O.,
Kalso, S., Koukila-Kahkola, P., Mussalo-Rauhamaa, H., and Haahtela, T.
(2000). Mycotoxins in crude building materials from water-damaged
buildings. Appl. Environ. Microbiol. 66, 1899–1904.
Tuomi, T., Saarinen, L., and Reijula, K. (1998). Detection of polar and
macrocyclic trichothecene mycotoxins from indoor environments. Analyst
123, 1835–1841.
Walker, P. R., and Whitfield, J. F. (1985). Cytoplasmic microtubules are
essential for the formation of membrane-bound polyribosomes. J. Biol.
Chem. 260, 765–770.
Yike, I., and Dearborn, D. G. (2004). Pulmonary effects of Stachybotrys
chartarum in animal studies. Adv. Appl. Microbiol. 55, 241–273.
Yike, I., Miller, M. J., Sorenson, W. G., Walenga, R., Tomashefski, J. F., Jr,
and Dearborn, D. G. (2002). Infant animal model of pulmonary myco-
toxicosis induced by Stachybotrys chartarum. Mycopathologia 154,
139–152.
Yike, I., Rand, T. G., and Dearborn, D. G. (2005). Acute inflammatory
responses to Stachybotrys chartarum in the lungs of infant rats: Time course
and possible mechanisms. Toxicol. Sci. 84, 408–417.
Zhou, H. R., Harkema, J. R., Hotchkiss, J. A., Yan, D., Roth, R. A., and Pestka,
J. J. (2000). Lipopolysaccharide and the trichothecene vomitoxin (deoxy-
nivalenol) synergistically induce apoptosis in murine lymphoid organs.
Toxicol. Sci. 53, 253–263.
Zhou, H. R., Harkema, J. R., Yan, D., and Pestka, J. J. (1999). Amplified
proinflammatory cytokine expression and toxicity in mice coexposed to
lipopolysaccharide and the trichothecene vomitoxin (deoxynivalenol).
J. Toxicol. Environ. Health 57, 115–136.
Zhou, H. R., Islam, Z., and Pestka, J. J. (2005). Induction of competing
apoptotic and survival signaling pathways in the macrophage by the
ribotoxic trichothecene deoxynivalenol. Toxicol. Sci. 87, 113–122.
Zhou, H. R., Lau, A. S., and Pestka, J. J. (2003). Role of double-stranded RNA-
activated protein kinase R (PKR) in deoxynivalenol-induced ribotoxic stress
response. Toxicol. Sci. 74, 335–344.
MYCOTOXIN RORIDIN A: INDUCED NASAL NEUROTOXICITY 541
by guest on Novem
ber 19, 2014http://toxsci.oxfordjournals.org/
Dow
nloaded from
Related Documents











