Neuroprotective effects of brimonidine treatment in a rodent model of ischemic optic neuropathy Nataliya O. Danylkova a , Sandra R. Alcala a,b , Howard D. Pomeranz a,1 , Linda K. McLoon a,b, * a Department of Ophthalmology, University of Minnesota, Minneapolis, MN 55455, USA b Department of Neuroscience, University of Minnesota, Minneapolis, MN 55455, USA Received 30 May 2006; accepted in revised form 2 October 2006 Available online 16 November 2006 Abstract Ischemic optic neuropathy (ION) is a common disorder caused by disruption of the arterial blood supply to the optic nerve. It can result in significant loss of visual acuity and/or visual field. An ischemic optic nerve injury was produced in rats by intravenous injection of Rose Bengal dye followed by argon green laser application to the retinal arteries overlying the optic nerve, causing a coagulopathy within the blood vessels and disruption of optic nerve and retinal perfusion. The effect of brimonidine tartrate eye drops on survival of retinal ganglion cell axons in this experimental paradigm was studied. One eye was treated and the contralateral eye served as a control. Four groups of animals were used for this study. Group 1 received 7 days of treatment with 0.15% brimonidine tartrate eye drops twice a day prior to the ischemic injury. Group 2 animals received 0.15% brimonidine tartrate eye drops twice a day for 14 days after photocoagulation injury. Animal groups 3 and 4 received eye drops of 0.9% NaCl twice a day either daily for 7 days before injury or daily for 14 days, respectively. All rats were sacrificed 5 months after the injury to ascertain long-term optic axon survival. Coagulopathy-induced optic nerve ischemia resulted in a 71% loss of optic axons. Treatment with brimonidine daily for the 7 days prior to the injury resulted in a greater survival of optic axons, with only a 56.1% loss compared to control. Brimonidine treatment every day for 14 days after the ischemic injury did not result in a significant rescue of optic axons compared to injury alone. In summary, the application of brimonidine eye drops for one week prior to an ischemic injury resulted in a statistically significant increase in survival of optic axons within the injured optic nerves. Brimonidine treatment of the eye after the ischemic injury did not result in axon rescue, and axon loss was similar to the injured optic nerves treated with saline only. These results suggest that brimonidine may have potential use for prevention of ION in at-risk patients. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: neuroprotection; optic nerve; ischemia; hypoxia; a2-agonists 1. Introduction Ischemic optic neuropathy is one of the most common optic nerve disorders in the elderly. It is characterized by disruption of blood supply to the optic nerve by branches of the posterior ciliary arteries (Hayreh, 1985, 1996). The resultant transient ischemia leads to an alteration of retinal cell metabolism, in- cluding changes in extracellular ion concentrations, depletion of growth factors, altered release of neurotransmitters, and increases in free radicals. These processes lead to axonal de- generation and progressive neuronal cell loss via apoptosis, which ultimately results in significant and permanent vision loss (Levin and Louhab, 1996; Salazar et al., 2000). Due to the complexity of the pathologic processes in the development of ischemic optic neuropathy (ION), many different treatment approaches have been advocated. However, all of the current * Corresponding author. Department of Ophthalmology, University of Min- nesota, Room 374 Lions Research Building, 2001 6th Street SE, Minneapolis, MN 55455, USA. Tel.: þ1 612 626 0777 (Office). E-mail address: [email protected] (L.K. McLoon). 1 Present Address: Department of Ophthalmology, North Shore Long Island Jewish Health System, 600 Northern Blvd Suite 214, Great Neck, NY 11021, USA. 0014-4835/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.exer.2006.10.002 Experimental Eye Research 84 (2007) 293e301 www.elsevier.com/locate/yexer

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Eye Research 84 (2007) 293e301www.elsevier.com/locate/yexer
Neuroprotective effects of brimonidine treatment in arodent model of ischemic optic neuropathy
Nataliya O. Danylkova a, Sandra R. Alcala a,b, Howard D. Pomeranz a,1, Linda K. McLoon a,b,*
a Department of Ophthalmology, University of Minnesota, Minneapolis, MN 55455, USAb Department of Neuroscience, University of Minnesota, Minneapolis, MN 55455, USA
Received 30 May 2006; accepted in revised form 2 October 2006
Available online 16 November 2006
Abstract
Ischemic optic neuropathy (ION) is a common disorder caused by disruption of the arterial blood supply to the optic nerve. It can result insignificant loss of visual acuity and/or visual field. An ischemic optic nerve injury was produced in rats by intravenous injection of Rose Bengaldye followed by argon green laser application to the retinal arteries overlying the optic nerve, causing a coagulopathy within the blood vesselsand disruption of optic nerve and retinal perfusion. The effect of brimonidine tartrate eye drops on survival of retinal ganglion cell axons in thisexperimental paradigm was studied. One eye was treated and the contralateral eye served as a control. Four groups of animals were used for thisstudy. Group 1 received 7 days of treatment with 0.15% brimonidine tartrate eye drops twice a day prior to the ischemic injury. Group 2 animalsreceived 0.15% brimonidine tartrate eye drops twice a day for 14 days after photocoagulation injury. Animal groups 3 and 4 received eye dropsof 0.9% NaCl twice a day either daily for 7 days before injury or daily for 14 days, respectively. All rats were sacrificed 5 months after the injuryto ascertain long-term optic axon survival. Coagulopathy-induced optic nerve ischemia resulted in a 71% loss of optic axons. Treatment withbrimonidine daily for the 7 days prior to the injury resulted in a greater survival of optic axons, with only a 56.1% loss compared to control.Brimonidine treatment every day for 14 days after the ischemic injury did not result in a significant rescue of optic axons compared to injuryalone. In summary, the application of brimonidine eye drops for one week prior to an ischemic injury resulted in a statistically significantincrease in survival of optic axons within the injured optic nerves. Brimonidine treatment of the eye after the ischemic injury did not resultin axon rescue, and axon loss was similar to the injured optic nerves treated with saline only. These results suggest that brimonidine mayhave potential use for prevention of ION in at-risk patients.� 2006 Elsevier Ltd. All rights reserved.
Keywords: neuroprotection; optic nerve; ischemia; hypoxia; a2-agonists
1. Introduction
Ischemic optic neuropathy is one of the most common opticnerve disorders in the elderly. It is characterized by disruption
* Corresponding author. Department of Ophthalmology, University of Min-
nesota, Room 374 Lions Research Building, 2001 6th Street SE, Minneapolis,
MN 55455, USA. Tel.: þ1 612 626 0777 (Office).
E-mail address: [email protected] (L.K. McLoon).1 Present Address: Department of Ophthalmology, North Shore Long Island
Jewish Health System, 600 Northern Blvd Suite 214, Great Neck, NY 11021,
USA.
0014-4835/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.exer.2006.10.002
of blood supply to the optic nerve by branches of the posteriorciliary arteries (Hayreh, 1985, 1996). The resultant transientischemia leads to an alteration of retinal cell metabolism, in-cluding changes in extracellular ion concentrations, depletionof growth factors, altered release of neurotransmitters, andincreases in free radicals. These processes lead to axonal de-generation and progressive neuronal cell loss via apoptosis,which ultimately results in significant and permanent visionloss (Levin and Louhab, 1996; Salazar et al., 2000). Due tothe complexity of the pathologic processes in the developmentof ischemic optic neuropathy (ION), many different treatmentapproaches have been advocated. However, all of the current

294 N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
treatment methods fail to result in any significant improvementin vision in patients with ION.
Alpha 2-adrenergic receptors play an important role invascular autoregulation (Faber and Meininger, 1990; McGilliv-ray-Anderson and Faber, 1991). Activation of alpha 2-adrenergic receptors inhibits adenylate cyclase activity (Jakobs,1979; Osborne, 1991), inhibits calcium channels (Han and Wu,2002), activates opening of the potassium channels in the cells(Debock et al., 2003), and inhibits pro-apoptotic mitochondrialsignaling (Tatton et al., 2001a,b). Alpha 2-adrenergic receptorsare present in the retina (Elena et al., 1989; Matsuo andCynader, 1992), specifically localized in rat retina to theinner plexiform and ganglion cell layers (Zarbin et al., 1986;Wheeler et al., 2001).
A number of studies demonstrated the efficacy of alpha2-agonists in reducing the negative effects of brain ischemia.Alpha 2-agonists such as dexmedetomidine (Maier et al.,1993), and clonidine (Yuan et al., 2001; Zhang, 2004) pro-vided neuroprotection in animal models of CNS ischemia.Another promising candidate for therapeutic neuroprotectiveeffects following transient ischemia is the alpha 2-agonist,brimonidine. Currently, brimonidine tartrate is used clini-cally as a topical ocular hypotensive agent in glaucoma pa-tients (Gandolfi et al., 2003), as well as in postoperativepatients, in order to control intraocular pressure (Katsimpriset al., 2003). Brimonidine is thought to lower intraocularpressure by a combination of reducing aqueous humor pro-duction and increasing uveoscleral outflow (Toris et al.,1995; Greenfield et al., 1997). Topical application of brimo-nidine can achieve a concentration sufficient to activatealpha2-adrenergic receptors within ocular tissues (Acheam-pong et al., 2002).
A number of studies have demonstrated the efficacy ofbrimonidine in increasing survival of retinal ganglion cellsafter various types of injury. Intraperitoneal pretreatmentwith brimonidine tartrate significantly increased ganglioncell survival and retinal function after optic nerve crush (Yoleset al., 1999; Wheeler et al., 1999; Lafuente et al., 2001). Pre-vention of an early loss of retinal ganglion cells was demon-strated after topical administration of brimonidine prior totransient retinal ischemia induced by ophthalmic vessel liga-tion for 60e90 min (Lafuente et al., 2001). The mechanismfor this protection is unclear, but brimonidine appears to resultin inhibition of glutamate and aspartate accumulation (Donelloet al., 2001), the up-regulation of anti-apoptotic genes such asbcl-2 and bcl-xl and neuroprotective molecules such as fibro-blast growth factor (Lai et al., 2002).
The main objective of this study was to determine the effectof brimonidine treatment on the survival of the optic axons andneurons within the retinal ganglion cell layer in the ischemia-compromised eyes. Brimonidine eye drops were applied topi-cally before and/or after induction of ischemia-reperfusiondamage to the optic nerve and retina using a coagulopathymethod that is a rodent model of ION (Bernstein et al.,2003; Danylkova et al., 2006). Optic nerve survival was deter-mined by morphometric analysis of the optic nerve 5 monthsafter injury and treatment.
2. Methods
All procedures were approved by the Animal Care Com-mittee of the University of Minnesota and conformed to theNational Institute of Health Guide for the Care and Use ofLaboratory Animals and the ARVO Statement on the Use ofAnimals in Ophthalmic Research. Adult Long Evans rats,250e300 g, were housed with Research Animal Resourcesat the University of Minnesota on a 12-h light/dark cyclewith food and water ad libitum. Rats were anesthetized with0.224 g/kg tribromoethanol (SigmaeAldrich, St. Louis, MO)and kept on a heating pad for the duration of anesthesia andany experimental manipulations.
2.1. Induction of the ION
Pupils were dilated by instillation of 2 drops of a 2% cy-clogyl and 2.5% phenylephrine hydrochloride ophthalmicsolution (Bausch & Lomb, Tampa, FL) 5 min before laser ap-plication. GenTeal, a lubricant gel (CibaVision, Duluth, GA),was applied to the rat’s right eye to prevent eye irritationand increase the optical power of the lens. A transparent con-tact lens was placed on the animal’s right eye.
Transient ischemia was induced as described previously(Bernstein et al., 2003; Danylkova et al., 2006). Briefly,0.25 ml of a 2.5 mM solution of Rose Bengal dye, (SigmaeAldrich, St. Louis, MO) was injected into the rat’s tail vein,followed immediately by application of a 500 mm laser beamcovering the entire optic disc area and emerging vessels. Laserapplication was within 30 s of Rose Bengal application, andthus primarily in the arteries. However, both arteries and veinsat the optic nerve head were subjected to laser application.Twelve pulses of 1 s duration each were applied with an inten-sity of 100 mW from an argon green laser at a 514 nm wave-length (Coherent Novus 2000). The opposite eye served asa control.
2.2. Brimonidine administration
Four groups of animals were prepared for this study. Thefirst group (n¼ 6) received topical treatment with two drops(5 ml) of 0.15% brimonidine tartrate twice a day for 7 days be-fore the injury. The second group (n¼ 6) received topicaltreatment of 0.15% brimonidine tartrate twice a day for 14days following the induction of optic nerve ischemia. Groups 3(n¼ 3) and 4 (n¼ 3) received a sham topical treatment of 0.9%NaCl, twice a day, either 7 days before or 14 days after injury.
2.3. Histology
Five months after the injury, a time when both the acute andchronic phases of degeneration are complete (Danylkova et al.,2006), rats were deeply anesthetized with tribromoethanol andintracardially perfused with 4% paraformaldehyde in phos-phate buffered saline (PBS). Globes with optic nerves attachedwere removed and post-fixed in 4% paraformaldehyde over-night at room temperature. After 12 h, the tissues were rinsed

295N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
in PBS, and optic nerves were carefully separated 1 mm fromthe globe with a scalpel. The globes were embedded in paraf-fin, sectioned at 10 mm, and stained with cresyl violet for mor-phological analysis. Optic nerves were placed in 2% osmiumtetroxide in saline for 1 h, rinsed in PBS, dehydrated in alco-hol, and embedded in epoxy resin. Subsequently, 1-mm thickcross-sections were cut on an ultramicrotome and stainedwith toluidine blue for light microscopic examination andmorphometric analysis.
2.4. Morphometric analysis
Axonal loss in the optic nerve was determined by exam-ining 3e6 optic nerve cross-sections from each collectedoptic nerve at 100� using oil immersion. This results invery clear visualization of even thinly myelinated opticaxons. Axonal counts were performed by using BioquantNova Prime software (Nashville, TN). Axonal counts wereperformed by counting all the myelinated axons in a contin-uous strip across the entire diameter of each optic nerve us-ing the Bioquant Nova Prime software (Nashville, TN).Total numbers of axons in the area counted were deter-mined, and these were recalculated as total axon number/mm2 (Danylkova et al., 2006). We counted approximately20,000 axons in each control optic nerve; thus, the areacounted represented approximately 20% of the total opticaxons (Cepurna et al., 2005). It is possible that there issome experimental error in these counts due to the patchyloss in the injured optic nerves. In order to control for this,we counted 3e6 sections for each nerve and averaged thecounts for each experimental animal. We have previouslyshown that laser alone and Rose Bengal administration alonedo not cause optic nerve injury (Danylkova et al., 2006), sothose controls are not included. All data are presented asmean� SEM. Significance was assessed using Student’spaired two-tailed t-test and Dunn’s multiple comparison testsaided by the Prism and Statmate software (Graphpad, SanDiego, CA), and data were considered statistically signifi-cant if p< 0.05.
3. Results
3.1. No systemic side effects of brimonidine
The rats showed no evidence of adverse effects from thebrimonidine treatment. We had no animals die, nor did any ofthe treated rats show signs of pain or discomfort as manifestedby changes in activity level, changes in eating patterns and thelike.
3.2. Morphology of the optic nerve
Changes of the optic axons within the optic nerve wereevaluated 5 months after induction of the ischemic injury.Compared to the normal optic nerve (Fig. 1A), the vehicle-treated groups of rats displayed a severe loss of optic axons(Fig. 1B). There was a tendency toward increased loss in the
more central regions of the injured optic nerves, with sparingin the more peripheral portions of the optic nerve cross-sections.
In the group of animals pretreated topically with brimoni-dine 7 days before injury, then subjected to photocoagula-tion-induced ischemia of the optic nerve, there wasa significant reduction in axonal loss compared to the vehi-cle-treated group (Fig. 1C and D). The central area of the opticnerve contained scattered myelinated axons and the peripheryof the optic nerve had large areas with preservation of normal-appearing axons. In contrast, in animals that received a 14-daytreatment of topical brimonidine following ischemic injury,there appeared to be a modest reduction in the axonal degen-eration in the periphery of the optic nerve (Fig. 1E) andabsence of myelinated axons in the central portion of the opticnerve, similar to the control group of optic nerves treated withsaline only. Photomicrographs at higher power demonstratethe clarity with which even thinly myelinated axons can bevisualized in the plastic-embedded optic nerves (Fig. 2AeC)from control animals (Fig. 2A), animals treated with brimoni-dine 7 days prior to injury (Fig. 2B), animals treated with bri-monidine for 14 days after injury (Fig. 2C), and animals thatreceived an ischemic injury and were treated with salineonly (Fig. 2D).
3.3. Axonal counting
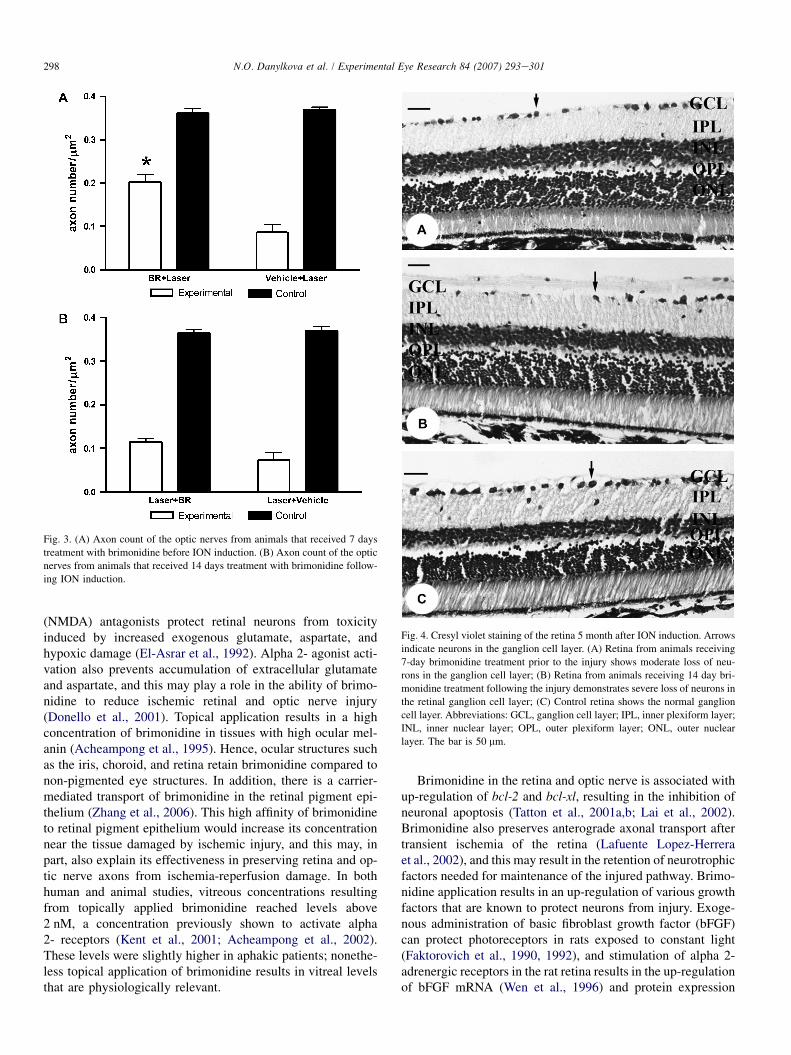
Normal control optic nerves of rats in our study had an op-tic axon density of 0.362� 0.011 axons per mm2 (n¼ 6)(Fig. 3). In rats subjected to the coagulopathy injury, therewas a significant loss of optic axons, with 0.087� 0.018 axonsper mm2 (n¼ 3). This represents a survival of only 23.5% ofthe axons after the ischemic injury. The number of axonsper sampled area in cross-sections from rats treated with top-ical brimonidine daily for 7 days prior to optic nerve ischemiawas 0.203� 0.016 per mm2 (n¼ 6), which represents a 56.1%long-term axonal survival compared to control eyes (Fig. 3).Topical treatment with brimonidine prior to ischemic insultto the optic nerve and retina resulted in significant rescue ofoptic axons compared to injury alone when examined 5months after the injury.
In contrast, the optic nerves of rats treated with topicalbrimonidine for 14 days after the photocoagulation-inducedoptic nerve ischemia contained 0.115� 0.008 axons permm2 (n¼ 6) while optic nerves from control eyes contained0.365� 0.008 per mm2 (n¼ 6) (Fig. 3). This corresponds to30.7% axonal survival in the injured optic nerves treatedwith brimonidine compared to control optic nerves. No sig-nificant difference was observed between groups of animalstreated with 0.9% NaCl either 7 days prior to induction ofthe ION or 14 days following the photocoagulation injury.Specifically, there were 0.073� 0.018 axons per mm2
(n¼ 3) in the group of animals treated with 0.9% NaClfor 14 days after the injury, which is a survival rate of19.7% when compared to normal control (0.370� 0.010per mm2; n¼ 6).
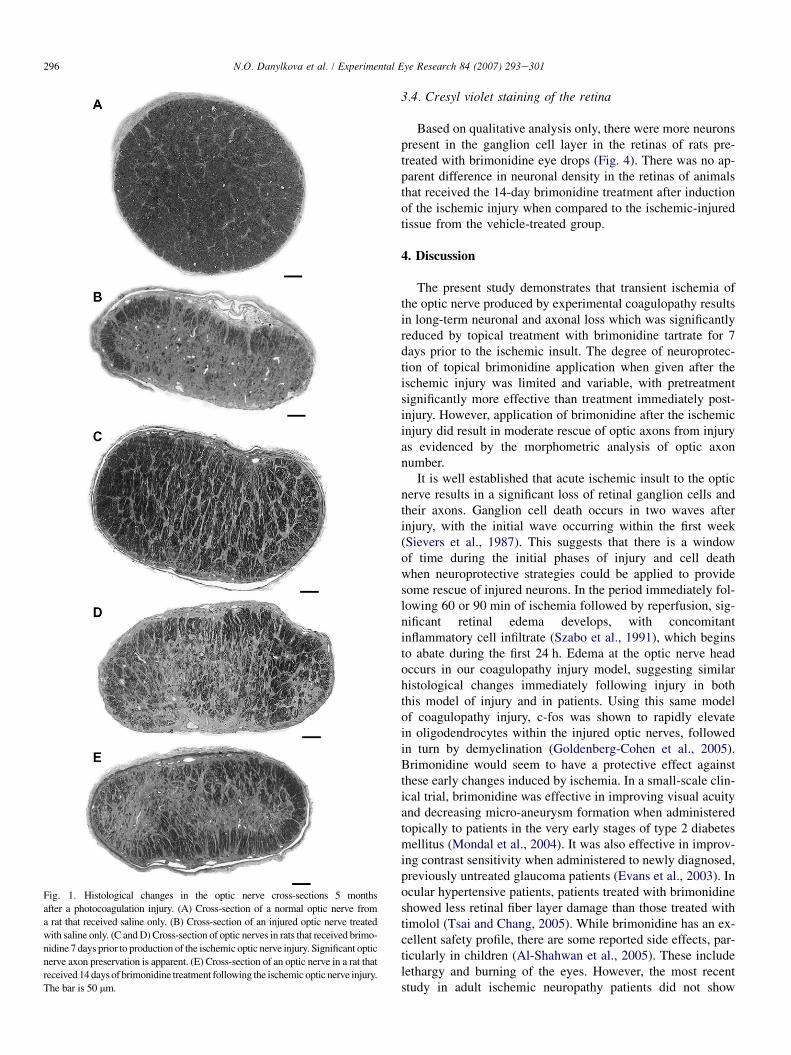
296 N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
Fig. 1. Histological changes in the optic nerve cross-sections 5 months
after a photocoagulation injury. (A) Cross-section of a normal optic nerve from
a rat that received saline only. (B) Cross-section of an injured optic nerve treated
with saline only. (C and D) Cross-section of optic nerves in rats that received brimo-
nidine 7 days prior to production of the ischemic optic nerve injury. Significant optic
nerve axon preservation is apparent. (E) Cross-section of an optic nerve in a rat that
received 14 days of brimonidine treatment following the ischemic optic nerve injury.
The bar is 50 mm.
3.4. Cresyl violet staining of the retina
Based on qualitative analysis only, there were more neuronspresent in the ganglion cell layer in the retinas of rats pre-treated with brimonidine eye drops (Fig. 4). There was no ap-parent difference in neuronal density in the retinas of animalsthat received the 14-day brimonidine treatment after inductionof the ischemic injury when compared to the ischemic-injuredtissue from the vehicle-treated group.
4. Discussion
The present study demonstrates that transient ischemia ofthe optic nerve produced by experimental coagulopathy resultsin long-term neuronal and axonal loss which was significantlyreduced by topical treatment with brimonidine tartrate for 7days prior to the ischemic insult. The degree of neuroprotec-tion of topical brimonidine application when given after theischemic injury was limited and variable, with pretreatmentsignificantly more effective than treatment immediately post-injury. However, application of brimonidine after the ischemicinjury did result in moderate rescue of optic axons from injuryas evidenced by the morphometric analysis of optic axonnumber.
It is well established that acute ischemic insult to the opticnerve results in a significant loss of retinal ganglion cells andtheir axons. Ganglion cell death occurs in two waves afterinjury, with the initial wave occurring within the first week(Sievers et al., 1987). This suggests that there is a windowof time during the initial phases of injury and cell deathwhen neuroprotective strategies could be applied to providesome rescue of injured neurons. In the period immediately fol-lowing 60 or 90 min of ischemia followed by reperfusion, sig-nificant retinal edema develops, with concomitantinflammatory cell infiltrate (Szabo et al., 1991), which beginsto abate during the first 24 h. Edema at the optic nerve headoccurs in our coagulopathy injury model, suggesting similarhistological changes immediately following injury in boththis model of injury and in patients. Using this same modelof coagulopathy injury, c-fos was shown to rapidly elevatein oligodendrocytes within the injured optic nerves, followedin turn by demyelination (Goldenberg-Cohen et al., 2005).Brimonidine would seem to have a protective effect againstthese early changes induced by ischemia. In a small-scale clin-ical trial, brimonidine was effective in improving visual acuityand decreasing micro-aneurysm formation when administeredtopically to patients in the very early stages of type 2 diabetesmellitus (Mondal et al., 2004). It was also effective in improv-ing contrast sensitivity when administered to newly diagnosed,previously untreated glaucoma patients (Evans et al., 2003). Inocular hypertensive patients, patients treated with brimonidineshowed less retinal fiber layer damage than those treated withtimolol (Tsai and Chang, 2005). While brimonidine has an ex-cellent safety profile, there are some reported side effects, par-ticularly in children (Al-Shahwan et al., 2005). These includelethargy and burning of the eyes. However, the most recentstudy in adult ischemic neuropathy patients did not show
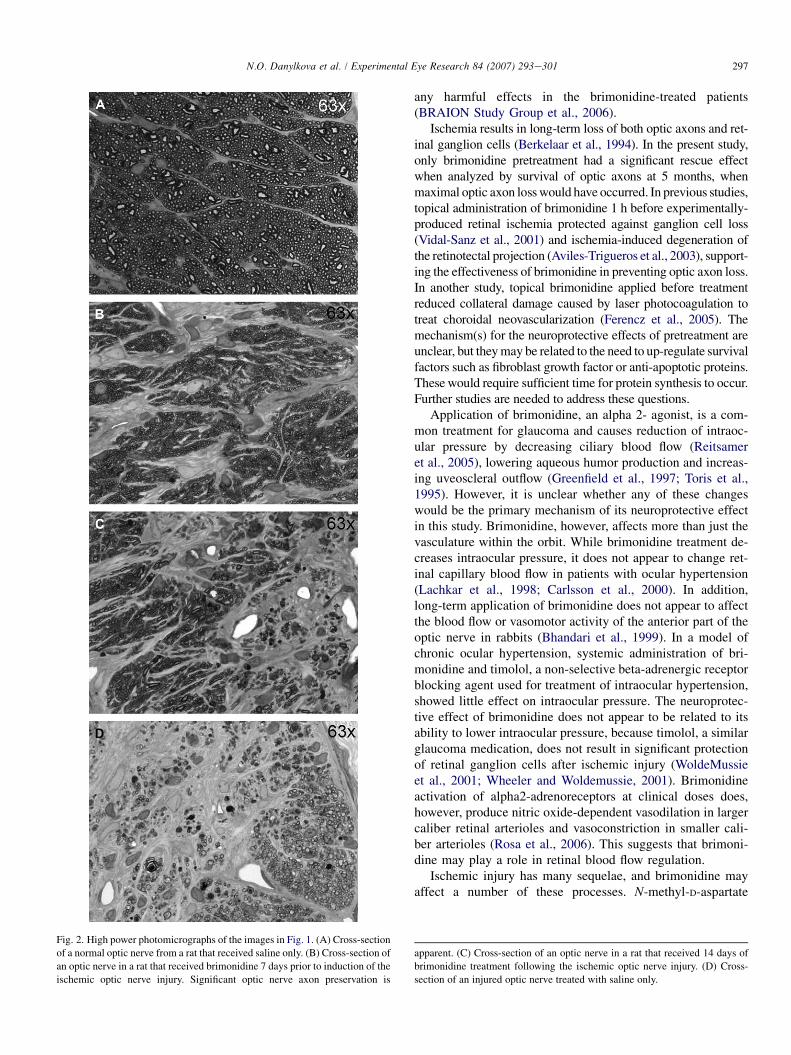
297N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
Fig. 2. High power photomicrographs of the images in Fig. 1. (A) Cross-section
of a normal optic nerve from a rat that received saline only. (B) Cross-section of
an optic nerve in a rat that received brimonidine 7 days prior to induction of the
ischemic optic nerve injury. Significant optic nerve axon preservation is
any harmful effects in the brimonidine-treated patients(BRAION Study Group et al., 2006).
Ischemia results in long-term loss of both optic axons and ret-inal ganglion cells (Berkelaar et al., 1994). In the present study,only brimonidine pretreatment had a significant rescue effectwhen analyzed by survival of optic axons at 5 months, whenmaximal optic axon loss would have occurred. In previous studies,topical administration of brimonidine 1 h before experimentally-produced retinal ischemia protected against ganglion cell loss(Vidal-Sanz et al., 2001) and ischemia-induced degeneration ofthe retinotectal projection (Aviles-Trigueros et al., 2003), support-ing the effectiveness of brimonidine in preventing optic axon loss.In another study, topical brimonidine applied before treatmentreduced collateral damage caused by laser photocoagulation totreat choroidal neovascularization (Ferencz et al., 2005). Themechanism(s) for the neuroprotective effects of pretreatment areunclear, but they may be related to the need to up-regulate survivalfactors such as fibroblast growth factor or anti-apoptotic proteins.These would require sufficient time for protein synthesis to occur.Further studies are needed to address these questions.
Application of brimonidine, an alpha 2- agonist, is a com-mon treatment for glaucoma and causes reduction of intraoc-ular pressure by decreasing ciliary blood flow (Reitsameret al., 2005), lowering aqueous humor production and increas-ing uveoscleral outflow (Greenfield et al., 1997; Toris et al.,1995). However, it is unclear whether any of these changeswould be the primary mechanism of its neuroprotective effectin this study. Brimonidine, however, affects more than just thevasculature within the orbit. While brimonidine treatment de-creases intraocular pressure, it does not appear to change ret-inal capillary blood flow in patients with ocular hypertension(Lachkar et al., 1998; Carlsson et al., 2000). In addition,long-term application of brimonidine does not appear to affectthe blood flow or vasomotor activity of the anterior part of theoptic nerve in rabbits (Bhandari et al., 1999). In a model ofchronic ocular hypertension, systemic administration of bri-monidine and timolol, a non-selective beta-adrenergic receptorblocking agent used for treatment of intraocular hypertension,showed little effect on intraocular pressure. The neuroprotec-tive effect of brimonidine does not appear to be related to itsability to lower intraocular pressure, because timolol, a similarglaucoma medication, does not result in significant protectionof retinal ganglion cells after ischemic injury (WoldeMussieet al., 2001; Wheeler and Woldemussie, 2001). Brimonidineactivation of alpha2-adrenoreceptors at clinical doses does,however, produce nitric oxide-dependent vasodilation in largercaliber retinal arterioles and vasoconstriction in smaller cali-ber arterioles (Rosa et al., 2006). This suggests that brimoni-dine may play a role in retinal blood flow regulation.
Ischemic injury has many sequelae, and brimonidine mayaffect a number of these processes. N-methyl-D-aspartate
apparent. (C) Cross-section of an optic nerve in a rat that received 14 days of
brimonidine treatment following the ischemic optic nerve injury. (D) Cross-
section of an injured optic nerve treated with saline only.
298 N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
(NMDA) antagonists protect retinal neurons from toxicityinduced by increased exogenous glutamate, aspartate, andhypoxic damage (El-Asrar et al., 1992). Alpha 2- agonist acti-vation also prevents accumulation of extracellular glutamateand aspartate, and this may play a role in the ability of brimo-nidine to reduce ischemic retinal and optic nerve injury(Donello et al., 2001). Topical application results in a highconcentration of brimonidine in tissues with high ocular mel-anin (Acheampong et al., 1995). Hence, ocular structures suchas the iris, choroid, and retina retain brimonidine compared tonon-pigmented eye structures. In addition, there is a carrier-mediated transport of brimonidine in the retinal pigment epi-thelium (Zhang et al., 2006). This high affinity of brimonidineto retinal pigment epithelium would increase its concentrationnear the tissue damaged by ischemic injury, and this may, inpart, also explain its effectiveness in preserving retina and op-tic nerve axons from ischemia-reperfusion damage. In bothhuman and animal studies, vitreous concentrations resultingfrom topically applied brimonidine reached levels above2 nM, a concentration previously shown to activate alpha2- receptors (Kent et al., 2001; Acheampong et al., 2002).These levels were slightly higher in aphakic patients; nonethe-less topical application of brimonidine results in vitreal levelsthat are physiologically relevant.
Fig. 3. (A) Axon count of the optic nerves from animals that received 7 days
treatment with brimonidine before ION induction. (B) Axon count of the optic
nerves from animals that received 14 days treatment with brimonidine follow-
ing ION induction.
Brimonidine in the retina and optic nerve is associated withup-regulation of bcl-2 and bcl-xl, resulting in the inhibition ofneuronal apoptosis (Tatton et al., 2001a,b; Lai et al., 2002).Brimonidine also preserves anterograde axonal transport aftertransient ischemia of the retina (Lafuente Lopez-Herreraet al., 2002), and this may result in the retention of neurotrophicfactors needed for maintenance of the injured pathway. Brimo-nidine application results in an up-regulation of various growthfactors that are known to protect neurons from injury. Exoge-nous administration of basic fibroblast growth factor (bFGF)can protect photoreceptors in rats exposed to constant light(Faktorovich et al., 1990, 1992), and stimulation of alpha 2-adrenergic receptors in the rat retina results in the up-regulationof bFGF mRNA (Wen et al., 1996) and protein expression
Fig. 4. Cresyl violet staining of the retina 5 month after ION induction. Arrows
indicate neurons in the ganglion cell layer. (A) Retina from animals receiving
7-day brimonidine treatment prior to the injury shows moderate loss of neu-
rons in the ganglion cell layer; (B) Retina from animals receiving 14 day bri-
monidine treatment following the injury demonstrates severe loss of neurons in
the retinal ganglion cell layer; (C) Control retina shows the normal ganglion
cell layer. Abbreviations: GCL, ganglion cell layer; IPL, inner plexiform layer;
INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear
layer. The bar is 50 mm.

299N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
(Lai et al., 2002). In addition, intravitreal application of brimo-nidine significantly increases endogenous expression of brain-derived neurotrophic factor (BDNF) in the retinal ganglion cells(Gao et al., 2002). Both bFGF and BDNF have significant neuro-protective actions, and thus the elevation of these neurotrophicfactors by local brimonidine treatment prior to the actual injurymay be responsible for the neuronal rescue action of brimoni-dine. Current studies are examining changes in neurotrophicfactor levels. The advantage of brimonidine use for the treatmentof optic nerve ischemia is that it is easy to administer topically,while traditional methods of systemic delivery or direct intraoc-ular injection often do not result in therapeutic doses ofneurotrophic factors and other drugs within the retina and opticnerve. Additional studies are needed to delineate further themechanism by which pretreatment of tissue with brimonidineprotects retina and optic nerve from ischemic injury. We arecurrently investigating intranasal delivery of neurotrophicfactors, likely candidates for rescue of injured neurons andaxons. Preliminary results demonstrate that this method ofdrug delivery results in therapeutic doses of neuroprotectiveagents to the retina and optic nerve (Alcala et al., 2006).
There are several subtypes of alpha 2- adrenergic receptors.While the anterior segment of the human eye possesses onlyalpha 2b- and alpha 2c- adrenergic receptor subtypes, rabbiteyes have all 3 subtypes, 2a-, 2b- and 2c- adrenergic receptors(Huang et al., 1995). It is possible that the rodent retina alsodiffers from the human complement of alpha 2- adrenergicreceptors. It is known that the alpha 2- adrenergic receptorsof the rat and human have different molecular and structuralcharacteristics (Wypijewski et al., 1995). This is important,as small-scale examination of brimonidine in human IONpatients did not appear to be effective when administered afterthe development of ION (Fazzone et al., 2003). These differ-ences should be taken into consideration when interpretingresults of ION treatment in animal models and further devel-opment of neuroprotective drugs.
There are differences between the microcirculation of theoptic nerve head region in the rat and human that should benoted relative to any model of ischemic injury to this region.In humans, the majority of the blood supply to the optic discand anterior optic nerve is from the short posterior ciliary ar-teries (Risco et al., 1981; Onda et al., 1995). These studies dis-agree about the role of the peripapillary choroid in anterior opticnerve blood supply. In the rat, the peripapillary choroid playsa significant role in the blood supply and venous drainage ofthe optic nerve head (Sugiyama et al., 1999; Morrison et al.,1999). This does not negate the protective effects of pretreat-ment with brimonidine after this mode of ischemic injury tothe optic nerve. However, it does make comparisons with otherischemic injury models important in determining whether theneuroprotective effect is dependent on the particular vesselsand particular areas of the optic nerve head that are injured inboth experimental animal models and human patients.
Our study suggests that brimonidine treatment promotesaxonal survival in the optic nerve of rat eyes subjected to is-chemia-reperfusion injury if used prior to the ischemic epi-sode. Treatment with brimonidine prior to the ischemic
injury in rats resulted in a significant increase in axonal sur-vival rate compared to brimonidine treatment given after theinduction of optic nerve ischemia. While the clinical trials inhuman patients have been equivocal, the prophylactic use ofbrimonidine is worth further investigation. These results sug-gest that brimonidine may be useful as a preventive measurefor patients with a high risk of developing ION and for thosedeveloping such a condition in the fellow eye.
Acknowledgements
Supported by the Neuro-ophthalmology Research Fund, theMinnesota Lions and Lionesses, Lew Wasserman Mid-CareerResearch to Prevent Blindness Merit Award (LKM) and anunrestricted grant to the Department of Ophthalmology fromResearch to Prevent Blindness Inc.
References
Acheampong, A.A., Shackleton, M., John, B., Burke, J., Wheeler, L., Tang-
Liu, D., 2002. Distribution of brimonidine into anterior and posterior tis-
sues of monkey, rabbit, and rat eyes. Drug Metab. Dispos. 30, 421e429.
Acheampong, A.A., Shackleton, M., Tang-Liu, D.D., 1995. Comparative ocu-
lar pharmacokinetics of brimonidine after a single dose application to the
eyes of albino and pigmented rabbits. Drug Metab. Dispos. 23, 708e712.
Alcala, S.R., Danylkova, N., Hoekman, J., Hanson, L., Frey, W.,
McLoon, L.K., 2006. Intranasal delivery of insulin-like growth factor-1
distributes to the optic nerve and retina in the rat. Investig. Ophthalmol.
Vis. Sci. 47, 728 (ARVO abstract).
Al-Shahwan, S., Al-Torbak, A.A., Turkmani, S., Al-Omran, M., Al-Jadaan, I.,
Edward, D.P., 2005. Side-effect profile of brimonidine tartrate in children.
Ophthalmology 112, 2143.
Aviles-Trigueros, M., Mayor-Torroglosa, S., Garcia-Aviles, A., Lafuente, M.P.,
Rodriguez, M.E., Miralles de Imperial, J., Villegas-Perez, M.P., Vidal-
Sanz, M., 2003. Transient ischemia of the retina results in massive degen-
eration of the retinotectal projection: long-term neuroprotection with
brimonidine. Exp. Neurol. 184, 767e777.
Berkelaar, M., Clarke, D.B., Wang, Y.C., Bray, G.M., Aguayo, A.J., 1994.
Axotomy results in delayed death and apoptosis of retinal ganglion cells
in adult rats. J. Neurosci. 14, 4368e4374.
Bernstein, S.L., Guo, Y., Kelman, S.E., Flower, R.W., Johnson, M.A., 2003.
Functional and cellular responses in a novel rodent model of anterior ische-
mic optic neuropathy. Investig. Ophthalmol. Vis. Sci. 44, 4153e4162.
Bhandari, A., Cioffi, G.A., Van Buskirk, E.M., Orgul, S., Wang, L., 1999. Ef-
fect of brimonidine on optic nerve blood flow in rabbits. Am. J. Ophthal-
mol. 128, 601e605.
BRAION Study Group, Wilhem, B., Ludtke, H., Wilhem, H., 2006. Efficacy
and tolerability of 0.2% brimonidine tartrate for the treatment of acute
non-arteritic anterior ischemic optic neuropathy (NAION): a 3-month,
double-masked, randomized, placebo-controlled trial. Graefe’s Arch.
Clin. Exp. Ophthalmol. 244, 551e558.
Carlsson, A.M., Chauhan, B.C., Lee, A.A., LeBlanc, R.P., 2000. The effect of
brimonidine tartrate on retinal blood flow in patients with ocular hyperten-
sion. Am. J. Ophthalmol. 129, 297e301.
Cepurna, W.O., Kayton, R.J., Johnson, E.C., Morrison, J.C., 2005. Age related
optic nerve axonal loss in adult Brown Norway rats. Exp. Eye Res. 80,
877e884.
Danylkova, N.O., Pomeranz, H.D., Alcala, S.R., McLoon, L.K., 2006. Histo-
logical and morphometric evaluation of transient retinal and optic nerve
ischemia in rat. Brain Res. 1096, 20e29.
Debock, F., Kurz, J., Azad, S.C., Parsons, C.G., Hapfelmeier, G.,
Zieglgansberger, W., Rammes, G., 2003. Alpha 2-adrenoreceptor activa-
tion inhibits LTP and LTD in the basolateral amygdale: involvement of

300 N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
Gieo-protein-mediated modulation of Ca2þ-channels and inwardly recti-
fying Kþ-channels in LTD. Eur. J. Neurosci. 17, 1411e1424.
Donello, J.E., Padillo, E.U., Webster, M.L., Wheeler, L.A., Gil, D.W., 2001.
Alpha (2)-adrenoceptor agonists inhibit vitreal glutamate and aspartate
accumulation and preserve retinal function after transient ischemia.
J. Pharmacol. Exp. Ther. 296, 216e223.
El-Asrar, A.M., Morse, P.H., Maimone, D., Torczynski, E., Reder, A.T., 1992.
MK-801 protects retinal neurons from hypoxia and the toxicity of gluta-
mate and aspartate. Investig. Ophthalmol. Vis. Sci. 33, 3463e3468.
Elena, P.P., Kosina-Boix, M., Denis, P., Moulin, G., Lapalus, P., 1989. Alpha 2-
adrenergic receptors in rat and rabbit eye: a tritium-sensitive film autora-
diography. Ophthalmic. Res. 21, 309e314.
Evans, D.W., Hosking, S.L., Gherghel, D., Barlett, J.D., 2003. Contrast sensi-
tivity improves after brimonidine therapy in primary open angle glaucoma:
a case for neuroprotection. Br. J. Ophthalmol. 87, 1463e1465.
Faber, J.E., Meininger, G.A., 1990. Selective interaction of alpha-adrenorecep-
tors with myogenic regulation of microvascular smooth muscle. Am. J.
Physiol. 259, H1126eH1133.
Faktorovich, E.G., Steinberg, R.H., Yasumura, D., Matthes, M.T.,
LaVail, M.M., 1992. Basic fibroblast growth factor and local injury protect
photoreceptors from light damage in the rat. J. Neurosci. 12, 3554e3567.
Faktorovich, E.G., Steinberg, R.H., Yasumura, D., Matthes, M.T.,
LaVail, M.M., 1990. Photoreceptor degeneration in inherited retinal dys-
trophy delayed by basic fibroblast growth factor. Nature 347, 83e86.
Fazzone, H.E., Kupersmith, M.J., Leibman, J., 2003. Does topical brimonidine
tartrate help NAION? Br. J. Ophthalmol. 87, 1193e1194.
Ferencz, J.R., Gilady, G., Harel, O., Belkin, M., Assia, E.I., 2005. Topical bri-
monidine reduces collateral damage caused by laser photocoagulation for
choroidal neovascularization. Graefe’s Arch. Clin. Exp. Ophthalmol. 243,
877e880.
Gandolfi, S.A., Cimino, L., Mora, P., 2003. Effect of brimonidine on intraoc-
ular pressure in normal tension glaucoma: a short term clinical trial. Eur. J.
Ophthalmol. 13, 611e615.
Gao, H., Qiao, X., Cantor, L.B., WuDunn, D., 2002. Up-regulation of brain-
derived neurotrophic factor expression by brimonidine in rat retinal
ganglion cells. Arch. Ophthalmol. 120, 797e803.
Goldenberg-Cohen, N., Guo, Y., Margolis, F., Cohen, Y., Miller, N.R.,
Bernstein, S.L., 2005. Oligodendrocyte dysfunction after induction of ex-
perimental anterior optic nerve ischemia. Investig. Ophthalmol. Vis. Sci.
46, 2716e2725.
Greenfield, D.S., Liebmann, J.M., Ritch, R., 1997. Brimonidine: a new alpha 2-
adrenoreceptor agonist for glaucoma treatment. J. Glaucoma 6, 250e258.
Han, Y., Wu, S.M., 2002. NMDA-evoked [Ca2þ] increase in salamander ret-
inal ganglion cells: modulation by PKA and adrenergic receptors. Vis.
Neurosci. 19, 249e256.
Hayreh, S.S., 1985. Inter-individual variation in blood supply of the optic
nerve head. Its importance in various ischemic disorders of the optic nerve
head, and glaucoma, low-tension glaucoma and allied disorders. Doc.
Ophthalmol. 59, 217e246.
Hayreh, S.S., 1996. Acute ischemic disorders of the optic nerve: pathogenesis,
clinical manifestations and management. Ophthalmol. Clin. North Am. 9,
407e442.
Huang, Y., Gil, D.W., Vanscheeuwijck, P., Stamer, W.D., Regan, J.W., 1995.
Localization of alpha 2-adrenergic receptor subtypes in the anterior seg-
ment of the human eye with selective antibodies. Investig. Ophthalmol.
Vis. Sci. 36, 2729e2739.
Jakobs, K.H., 1979. Inhibition of adenylate cyclase by hormones and neuro-
transmitters. Mol. Cell Endocrinol. 16, 147e156.
Katsimpris, J.M., Siganos, D., Konstas, A.G., Kozobolis, V., Georgiadis, N.,
2003. Efficacy of brimonidine 0.2% in controlling acute postoperative in-
traocular pressure elevation after phacoemulsification. J. Cataract Refract.
Surg. 29, 2288e2294.
Kent, A.R., Nussdorf, J.D., David, R., Tyson, F., Small, D., Fellows, D., 2001.
Vitreous concentration of topically applied brimonidine tartrate 0.2%.
Ophthalmology 108, 784e787.
Lachkar, Y., Migdal, C., Dhanjil, S., 1998. Effect of brimonidine tartrate
on ocular hemodynamic measurements. Arch. Ophthalmol. 116,
1591e1594.
Lafuente Lopez-Herrera, M.P., Mayor-Torroglosa, S., Miralles de Imperial, J.,
Villegas-Perez, M.P., Vidal-Sanz, M., 2002. Transient ischemia of the ret-
ina results in altered retrograde axoplasmic transport: neuroprotection with
brimonidine. Exp. Neurol. 178, 243e258.
Lafuente, M.P., Villegas-Perez, M.P., Sobrado-Calvo, P., Garcia-Aviles, A.,
Miralles de Imperial, J., Vidal-Sanz, M., 2001. Neuroprotective effects
of alpha (2)-selective adrenergic agonists against ischemia-induced retinal
ganglion cell death. Investig. Ophthalmol. Vis. Sci. 42, 2074e2084.
Lai, R.K., Chun, T., Hasson, D., Lee, S., Mehrbod, F., Wheeler, L., 2002.
Alpha 2 adrenoreceptor agonist protects retinal function after a retinal
ischemic injury in the rat. Vis. Neurosci. 19, 175e185.
Levin, L.A., Louhab, A., 1996. Apoptosis of retinal ganglion cells in anterior
ischemic optic neuropathy. Arch. Ophthalmol. 114, 488e491.
Maier, C., Steinberg, G.K., Sun, G.H., Zhi, G.T., Maze, M., 1993. Neuropro-
tection by the alpha 2-adrenoreceptor agonist dexmedetomidine in a focal
model of cerebral ischemia. Anesthesiology 79, 306e312.
Matsuo, T., Cynader, M.S., 1992. Localization of alpha-2 adrenergic receptors
in the human eye. Ophthalmic Res. 24, 213e219.
McGillivray-Anderson, K.M., Faber, J.E., 1991. Effect of reduced blood flow
on alpha 1- and alpha 2-adrenoceptor constriction of rat skeletal muscle
microvessels. Circ. Res. 69, 165e173.
Mondal, L.K., Baidya, K.P., Bhattacharya, B., Chatterjee, P.R., Bhaduri, G.,
2004. The efficacy of topical administration of brimonidine to reduce
ischaemia in the very early stage of diabetic retinopathy in good controlled
type-2 diabetes mellitus. J. Indian Med. Assoc. 102, 724e725.
Morrison, J.C., Johnson, E.C., Cepurna, W.O., Funk, R.H.W., 1999. Microvas-
culature of the rat optic nerve head. Investig. Ophthalmol. Vis. Sci. 40,
1702e1709.
Onda, E., Cioffi, G.A., Bacon, D.R., Van Buskirk, E.M., 1995. Microvascula-
ture of the human optic nerve. Am. J. Ophthalmol. 120, 92e102.
Osborne, N.N., 1991. Inhibition of cAMP production by alpha 2-adrenoceptor
stimulation in rabbit retina. Brain Res. 553, 84e88.
Reitsamer, H.A., Posey, M., Kiel, J.W., 2005. Effects of a topical alpha 2 ad-
renergic agonist on ciliary blood flow and aqueous production in rabbits.
Exp. Eye Res. 82, 405e415.
Risco, J.M., Grimson, B.S., Johnson, P.T., 1981. Angioarchitecture of the cil-
iary artery of the posterior pole. Arch. Ophthalmol. 99, 864e868.
Rosa Jr., R.H., Hein, T.W., Yuan, Z., Xu, W., Pechal, M.I., Geraets, R.L.,
Newman, J.M., Kuo, L., 2006. Brimonidine evokes heterogeneous vaso-
motor response of retinal arterioles: diminished nitric oxide-mediated va-
sodilation when size goes small. Am. J. Physiol. Heart Circ. Physiol. (Feb
17 Epub ahead of print).
Salazar, J.J., Ramirez, A.I., de Hoz, R., Rojas, B., Trivino, A., Ramirez, J.M.,
2000. Apoptosis in ischemic optic neuropathy. Arch. Soc. Esp. Oftalmol.
75, 819e824.
Sievers, J., Hausmann, B., Unsicker, K., Berry, M., 1987. Fibroblast growth
factors promote the survival of adult rat retinal ganglion cells after transec-
tion of the optic nerve. Neurosci. Lett. 76, 157e162.
Sugiyama, K., Gu, Z.B., Kawase, C., Yamamoto, T., Kitazawa, Y., 1999. Optic
nerve and peripapillary choroidal microvasculature of the rat eye. Investig.
Ophthalmol. Vis. Sci. 40, 3084e3090.
Szabo, M.E., Droy-Lefaix, M.T., Doly, M., Carre, C., Braquet, P., 1991. Ische-
mia and reperfusion-induced histologic changes in the rat retina. Demon-
stration of a free radical-mediated mechanism. Investig. Ophthalmol. Vis.
Sci. 32, 1471e1478.
Tatton, W.G., Chalmers-Redman, R.M., Tatton, N.A., 2001a. Apoptosis and
anti-apoptosis signaling in glaucomatous retinopathy. Eur. J. Ophthalmol.
11 (Suppl.), S12eS22.
Tatton, W.G., Chalmers-Redman, R.M., Sud, A., Podos, S.M., Mittag, T.W.,
2001b. Maintaining mitochondrial membrane impermeability. an opportunity
for new therapy in glaucoma? Surv. Ophthalmol. 45 (Suppl. 3), S277eS283.
Toris, C.B., Gleason, M.L., Camras, C.B., Yablonski, M.E., 1995. Effects of
brimonidine on aqueous humor dynamics in human eyes. Arch. Ophthal-
mol. 113, 1514e1517.
Tsai, J.C., Chang, H.W., 2005. Comparison of the effects of brimonidine 0.2%
and timolol 0.5% on retinal nerve fiber layer thickness in ocular hyperten-
sive patients: a prospective, unmasked study. J. Ocul. Pharmacol. Ther. 21,
475e482.

301N.O. Danylkova et al. / Experimental Eye Research 84 (2007) 293e301
Vidal-Sanz, M., Lafuente, M.P., Mayor-Torroglosa, S., Aguilera, M.E., Mir-
alles de Imperial, J., Villegas-Perez, M.P., 2001. Brimonidine’s neuropro-
tective effects against transient ischemia-induced retinal ganglion cell
death. Eur. J. Ophthalmol. 11 (Suppl. 2), S36eS40.
Wen, R., Cheng, T., Li, Y., Cao, W., Steinberg, R.H., 1996. Alpha 2-adrenergic
agonists induce basic fibroblast growth factor expression in photoreceptors
in vivo and ameliorate light damage. J. Neurosci. 16, 5986e5992.
Wheeler, L.A., Gil, D.W., WoldeMussie, E., 2001. Role of alpha-2 adrenergic
receptors in neuroprotection and glaucoma. Surv. Ophthalmol. 45 (Suppl.
3), S290eS294.
Wheeler, L.A., Lai, R., Woldemussie, E., 1999. From the lab to the clinic:
activation of an alpha-2 agonist pathway is neuroprotective in models
of retinal and optic nerve injury. Eur. J. Ophthalmol. 9 (Suppl. 1),
S17eS21.
Wheeler, L.A., Woldemussie, E., 2001. Alpha-2 adrenergic receptor agonists
are neuroprotective in experimental models of glaucoma. Eur. J. Ophthal-
mol. 11 (Suppl. 2), S30eS35.
WoldeMussie, E., Ruiz, G., Wijono, M., Wheeler, L.A., 2001. Neuroprotection
of retinal ganglion cells by brimonidine in rats with laser-induced chronic
ocular hypertension. Investig. Ophthalmol. Vis. Sci. 42, 2849e2855.
Wypijewski, K., Duda, T., Sharma, R.K., 1995. Structural, genetic and
pharmacological identity of the rat alpha 2-adrenergic receptor subtype
cA2-47 and its molecular characterization in rat adrenal, adrenocortical
carcinoma and bovine retina. Mol. Cell Biochem. 144, 181e190.
Yoles, E., Wheeler, L.A., Schwartz, M., 1999. Alpha 2-adrenoreceptor
agonists are neuroprotective in a rat model of optic nerve degeneration.
Investig. Ophthalmol. Vis. Sci. 40, 65e73.
Yuan, S.Z., Runold, M., Hagberg, H., Bona, E., Lagercrantz, H., 2001. Hyp-
oxic-ischaemic brain damage in immature rats: effects of adrenoreceptor
modulation. Eur. J. Paediatr. Neurol. 5, 29e35.
Zarbin, M.A., Wamsley, J.K., Palacios, J.M., Kuhar, M.J., 1986. Autoradio-
graphic localization of high affinity GABA, benzodiazepine, dopaminer-
gic, adrenergic and muscarinic cholinergic receptors in the rat, monkey
and human retina. Brain Res. 374, 75e92.
Zhang, Y., 2004. Clonidine preconditioning decreases infarct size and im-
proves neurological outcome from transient forebrain ischemia in the
rat. Neuroscience 125, 625e631.
Zhang, N., Kannan, R., Okamoto, C.T., Ryan, S.J., Lee, V.H., Hinton, D.R.,
2006. Characterization of brimonidine transport in retinal pigment epithe-
lium. Investig. Ophthalmol. Vis. Sci. 47, 287e294.
Related Documents











