Neuropeptidomic analysis of the brain and thoracic ganglion from the Jonah crab, Cancer borealis Jurgen Huybrechts, a, * Michael P. Nusbaum, b Luc Vanden Bosch, a Geert Baggerman, a Arnold De Loof, a and Liliane Schoofs a a Laboratory of Developmental Physiology and Molecular Biology, 59 Naamsestraat, BE-3000 K.U. Leuven, Belgium b Department of Neuroscience, University of Pennsylvania, School of Medicine, Philadelphia, PA, USA Received 2 July 2003 Abstract Mass spectrometric methods were applied to determine the peptidome of the brain and thoracic ganglion of the Jonah crab (Cancer borealis). Fractions obtained by high performance liquid chromatography were characterized using MALDI-TOF MS and ESI-Q-TOF MS/MS. In total, 28 peptides were identified within the molecular mass range 750–3000 Da. Comparison of the mo- lecular masses obtained with MALDI-TOF MS with the calculated molecular masses of known crustacean peptides revealed the presence of at least nine allatostatins, three orcokinin precursor derived peptides, namely FDAFTTGFGHS, [Ala 13 ]-orcokinin, and [Val 13 ]-orcokinin, and two kinins, a tachykinin-related peptide and four FMRFamide-related peptides. Eight other peptides were de novo sequenced by collision induced dissociation on the Q-TOF system and yielded AYNRSFLRFamide, PELDHVFLRFamide or EPLDHVFLRFamide, APQRNFLRFamide, LNPFLRFamide, DVRTPALRLRFamide, and LRNLRFamide, which belong to the FMRFamide related peptide family, as well as NFDEIDRSGFA and NFDEIDRSSFGFV, which display high sequence similarity to peptide sequences within the orcokinin precursor of Orconectes limosus. Our paper is the first (neuro)peptidomic analysis of the crustacean nervous system. Ó 2003 Elsevier Inc. All rights reserved. Keywords: Q-TOF; MALDI-TOF imaging; Mass spectrometry; Peptidomics; Cancer borealis; Orcokinin; FMRFamide-related peptides; Allatostatins Neuropeptides are important messenger molecules that selectively influence most, if not all physiological processes including behavior. They act as neurotrans- mitters, modulators, and classical hormones. When re- leased into the circulation they often display pleiotropic functions. Our knowledge of the structure and function of arthropod neuropeptides has been gradually in- creasing over the past three decades. A dramatic in- crease was seen shortly after the completion and publication of the Drosophila genome project in 2000 [1]. To illustrate this revolution: in 1975 not less than 125,000 cockroaches were sacrificed to purify and characterize a single neuropeptide (proctolin) [2]. In contrast, very recently (mid 2002), biochemical evidence was provided for the presence of 28 neuropeptides in the larval Drosophila central nervous system, using a com- bination of MS and bioinformatics [3]. In this recent study, only 50 CNSs of the fruit fly were needed to obtain enough material to be able to characterize these peptides. The genome project is not the only driving force of neuropeptide research. In the 1980s Electrospray Ioni- zation [4] and Matrix Assisted Laser Desorption/Ioni- zation [5] were introduced in mass spectrometry applications. These so called “soft” ionization tech- niques made the volatilization of biomolecules possible without dissociating them. By applying these techniques to peptide research, peptides can be isolated and iden- tified from smaller tissue extracts [6]. Even entire peptide profiles from specified organs can be studied [3,7,8]. Tissue extraction is not always necessary because small organs [9], single cells [10] or even individual cell Biochemical and Biophysical Research Communications 308 (2003) 535–544 www.elsevier.com/locate/ybbrc BBRC * Corresponding author. Fax: +32-16-32-39-02. E-mail address: [email protected] (J. Hu- ybrechts). 0006-291X/$ - see front matter Ó 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0006-291X(03)01426-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical and Biophysical Research Communications 308 (2003) 535–544
www.elsevier.com/locate/ybbrc
BBRC
Neuropeptidomic analysis of the brain and thoracic ganglionfrom the Jonah crab, Cancer borealis
Jurgen Huybrechts,a,* Michael P. Nusbaum,b Luc Vanden Bosch,a Geert Baggerman,a
Arnold De Loof,a and Liliane Schoofsa
a Laboratory of Developmental Physiology and Molecular Biology, 59 Naamsestraat, BE-3000 K.U. Leuven, Belgiumb Department of Neuroscience, University of Pennsylvania, School of Medicine, Philadelphia, PA, USA
Received 2 July 2003
Abstract
Mass spectrometric methods were applied to determine the peptidome of the brain and thoracic ganglion of the Jonah crab
(Cancer borealis). Fractions obtained by high performance liquid chromatography were characterized using MALDI-TOF MS and
ESI-Q-TOF MS/MS. In total, 28 peptides were identified within the molecular mass range 750–3000 Da. Comparison of the mo-
lecular masses obtained with MALDI-TOF MS with the calculated molecular masses of known crustacean peptides revealed the
presence of at least nine allatostatins, three orcokinin precursor derived peptides, namely FDAFTTGFGHS, [Ala13]-orcokinin, and
[Val13]-orcokinin, and two kinins, a tachykinin-related peptide and four FMRFamide-related peptides. Eight other peptides were de
novo sequenced by collision induced dissociation on the Q-TOF system and yielded AYNRSFLRFamide, PELDHVFLRFamide or
EPLDHVFLRFamide, APQRNFLRFamide, LNPFLRFamide, DVRTPALRLRFamide, and LRNLRFamide, which belong to
the FMRFamide related peptide family, as well as NFDEIDRSGFA and NFDEIDRSSFGFV, which display high sequence
similarity to peptide sequences within the orcokinin precursor of Orconectes limosus. Our paper is the first (neuro)peptidomic
analysis of the crustacean nervous system.
� 2003 Elsevier Inc. All rights reserved.
Keywords: Q-TOF; MALDI-TOF imaging; Mass spectrometry; Peptidomics; Cancer borealis; Orcokinin; FMRFamide-related peptides;
Allatostatins
Neuropeptides are important messenger molecules
that selectively influence most, if not all physiological
processes including behavior. They act as neurotrans-
mitters, modulators, and classical hormones. When re-
leased into the circulation they often display pleiotropic
functions. Our knowledge of the structure and function
of arthropod neuropeptides has been gradually in-
creasing over the past three decades. A dramatic in-crease was seen shortly after the completion and
publication of the Drosophila genome project in 2000 [1].
To illustrate this revolution: in 1975 not less than
125,000 cockroaches were sacrificed to purify and
characterize a single neuropeptide (proctolin) [2]. In
contrast, very recently (mid 2002), biochemical evidence
* Corresponding author. Fax: +32-16-32-39-02.
E-mail address: [email protected] (J. Hu-
ybrechts).
0006-291X/$ - see front matter � 2003 Elsevier Inc. All rights reserved.
doi:10.1016/S0006-291X(03)01426-8
was provided for the presence of 28 neuropeptides in the
larval Drosophila central nervous system, using a com-
bination of MS and bioinformatics [3]. In this recent
study, only 50 CNSs of the fruit fly were needed to
obtain enough material to be able to characterize these
peptides.
The genome project is not the only driving force of
neuropeptide research. In the 1980s Electrospray Ioni-zation [4] and Matrix Assisted Laser Desorption/Ioni-
zation [5] were introduced in mass spectrometry
applications. These so called “soft” ionization tech-
niques made the volatilization of biomolecules possible
without dissociating them. By applying these techniques
to peptide research, peptides can be isolated and iden-
tified from smaller tissue extracts [6]. Even entire peptide
profiles from specified organs can be studied [3,7,8].Tissue extraction is not always necessary because small
organs [9], single cells [10] or even individual cell
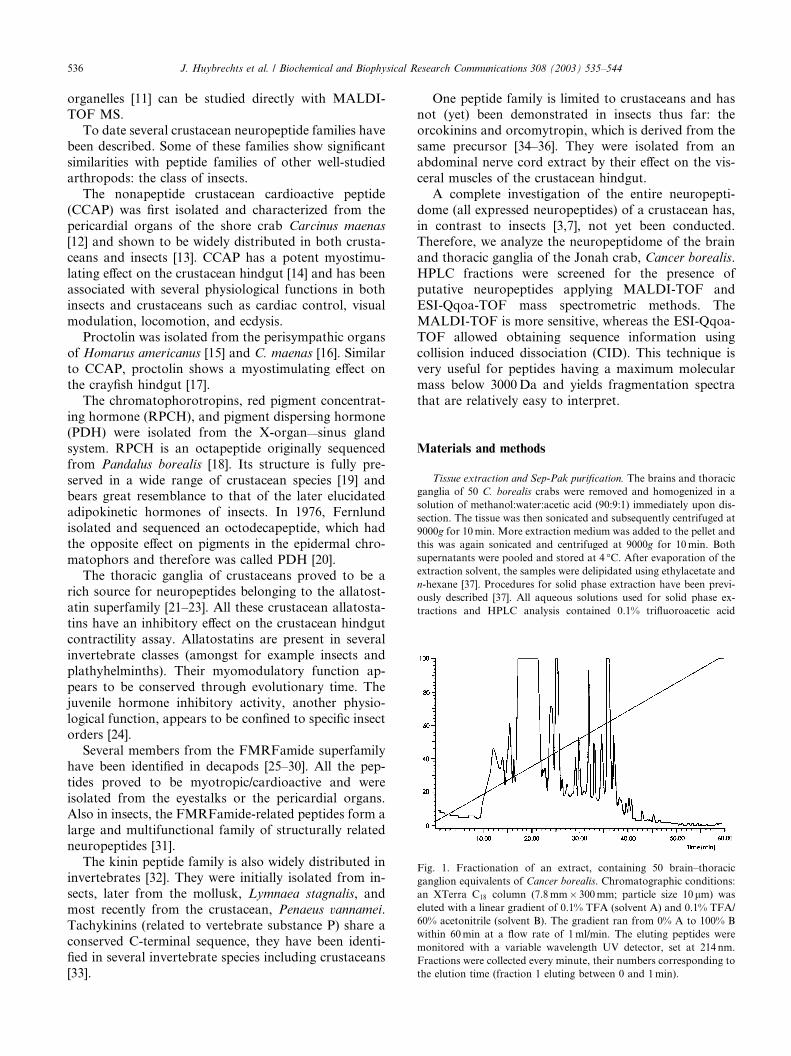
Fig. 1. Fractionation of an extract, containing 50 brain–thoracic
ganglion equivalents of Cancer borealis. Chromatographic conditions:
an XTerra C18 column (7.8 mm� 300mm; particle size 10lm) was
eluted with a linear gradient of 0.1% TFA (solvent A) and 0.1% TFA/
60% acetonitrile (solvent B). The gradient ran from 0% A to 100% B
within 60min at a flow rate of 1 ml/min. The eluting peptides were
monitored with a variable wavelength UV detector, set at 214 nm.
Fractions were collected every minute, their numbers corresponding to
the elution time (fraction 1 eluting between 0 and 1min).
536 J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544
organelles [11] can be studied directly with MALDI-TOF MS.
To date several crustacean neuropeptide families have
been described. Some of these families show significant
similarities with peptide families of other well-studied
arthropods: the class of insects.
The nonapeptide crustacean cardioactive peptide
(CCAP) was first isolated and characterized from the
pericardial organs of the shore crab Carcinus maenas
[12] and shown to be widely distributed in both crusta-
ceans and insects [13]. CCAP has a potent myostimu-
lating effect on the crustacean hindgut [14] and has been
associated with several physiological functions in both
insects and crustaceans such as cardiac control, visual
modulation, locomotion, and ecdysis.
Proctolin was isolated from the perisympathic organs
of Homarus americanus [15] and C. maenas [16]. Similarto CCAP, proctolin shows a myostimulating effect on
the crayfish hindgut [17].
The chromatophorotropins, red pigment concentrat-
ing hormone (RPCH), and pigment dispersing hormone
(PDH) were isolated from the X-organ—sinus gland
system. RPCH is an octapeptide originally sequenced
from Pandalus borealis [18]. Its structure is fully pre-
served in a wide range of crustacean species [19] andbears great resemblance to that of the later elucidated
adipokinetic hormones of insects. In 1976, Fernlund
isolated and sequenced an octodecapeptide, which had
the opposite effect on pigments in the epidermal chro-
matophors and therefore was called PDH [20].
The thoracic ganglia of crustaceans proved to be a
rich source for neuropeptides belonging to the allatost-
atin superfamily [21–23]. All these crustacean allatosta-tins have an inhibitory effect on the crustacean hindgut
contractility assay. Allatostatins are present in several
invertebrate classes (amongst for example insects and
plathyhelminths). Their myomodulatory function ap-
pears to be conserved through evolutionary time. The
juvenile hormone inhibitory activity, another physio-
logical function, appears to be confined to specific insect
orders [24].Several members from the FMRFamide superfamily
have been identified in decapods [25–30]. All the pep-
tides proved to be myotropic/cardioactive and were
isolated from the eyestalks or the pericardial organs.
Also in insects, the FMRFamide-related peptides form a
large and multifunctional family of structurally related
neuropeptides [31].
The kinin peptide family is also widely distributed ininvertebrates [32]. They were initially isolated from in-
sects, later from the mollusk, Lymnaea stagnalis, and
most recently from the crustacean, Penaeus vannamei.
Tachykinins (related to vertebrate substance P) share a
conserved C-terminal sequence, they have been identi-
fied in several invertebrate species including crustaceans
[33].
One peptide family is limited to crustaceans and hasnot (yet) been demonstrated in insects thus far: the
orcokinins and orcomytropin, which is derived from the
same precursor [34–36]. They were isolated from an
abdominal nerve cord extract by their effect on the vis-
ceral muscles of the crustacean hindgut.
A complete investigation of the entire neuropepti-
dome (all expressed neuropeptides) of a crustacean has,
in contrast to insects [3,7], not yet been conducted.Therefore, we analyze the neuropeptidome of the brain
and thoracic ganglia of the Jonah crab, Cancer borealis.
HPLC fractions were screened for the presence of
putative neuropeptides applying MALDI-TOF and
ESI-Qqoa-TOF mass spectrometric methods. The
MALDI-TOF is more sensitive, whereas the ESI-Qqoa-
TOF allowed obtaining sequence information using
collision induced dissociation (CID). This technique isvery useful for peptides having a maximum molecular
mass below 3000 Da and yields fragmentation spectra
that are relatively easy to interpret.
Materials and methods
Tissue extraction and Sep-Pak purification. The brains and thoracic
ganglia of 50 C. borealis crabs were removed and homogenized in a
solution of methanol:water:acetic acid (90:9:1) immediately upon dis-
section. The tissue was then sonicated and subsequently centrifuged at
9000g for 10min. More extraction medium was added to the pellet and
this was again sonicated and centrifuged at 9000g for 10min. Both
supernatants were pooled and stored at 4 �C. After evaporation of the
extraction solvent, the samples were delipidated using ethylacetate and
n-hexane [37]. Procedures for solid phase extraction have been previ-
ously described [37]. All aqueous solutions used for solid phase ex-
tractions and HPLC analysis contained 0.1% trifluoroacetic acid
J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544 537
(TFA), as ion paring agent. The dry residue of the extracted material
of 50 brain and thoracic ganglion equivalents was dissolved in 5 ml of
0.1% TFA and applied to Sep-Pak cartridge. This cartridge was eluted
with 5 ml of 60% acetonitrile/water and the resulting fraction was
lyophilized.
High performance liquid chromatography. HPLC analysis was per-
formed on a Gilson liquid chromatograph equipped with two pumps
and a solvent mixing unit. The eluting peptides were detected with a
variable wavelength UV detector, set at 214 nm (Waters 486 Tunable
Absorbance detector). The separation of the compounds was carried
out on a semi-preparative Waters XTerra C18 column (7.8mm�300mm; particle size 10lm) at ambient temperature and with a sol-
vent flow rate of 2ml/min. The lyophilized residue from 50 brain and
thoracic ganglion equivalents was dissolved in 1.5ml of 0.1% TFA and
transferred to the injector after filtering through a Millipore 0.22 lm
filter. Immediately after injection a linear gradient from 0% to 60%
acetonitrile containing 0.1% TFA was initiated. Final concentration
(60% acetonitrile) was achieved after 60min. Fractions containing 2ml
were collected every minute starting at 0 until 60min.
MALDI-TOF mass spectrometry. Matrix-assisted laser desorption/
ionization (MALDI) time-of-flight (TOF) mass spectrometry (MS)
was performed on a Reflex IV (Bruker Daltonic GmbH, Germany),
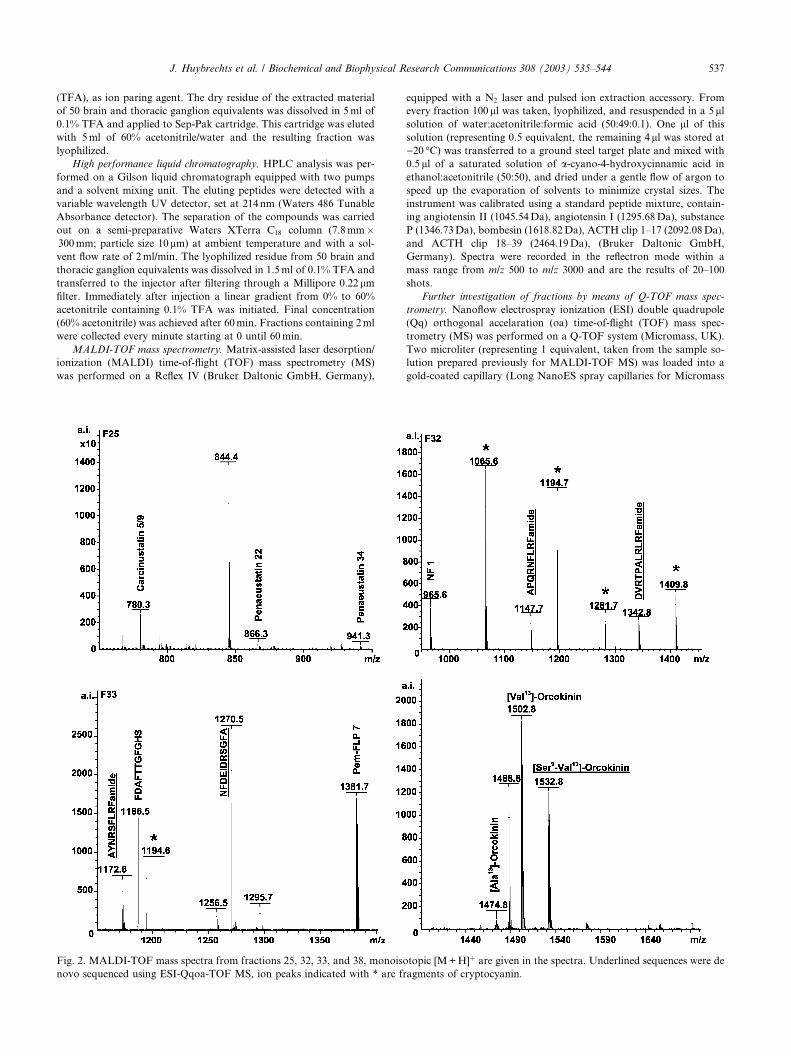
Fig. 2. MALDI-TOF mass spectra from fractions 25, 32, 33, and 38, monoiso
novo sequenced using ESI-Qqoa-TOF MS, ion peaks indicated with * are fr
equipped with a N2 laser and pulsed ion extraction accessory. From
every fraction 100ll was taken, lyophilized, and resuspended in a 5ll
solution of water:acetonitrile:formic acid (50:49:0.1). One ll of this
solution (representing 0.5 equivalent, the remaining 4ll was stored at
)20 �C) was transferred to a ground steel target plate and mixed with
0.5 ll of a saturated solution of a-cyano-4-hydroxycinnamic acid in
ethanol:acetonitrile (50:50), and dried under a gentle flow of argon to
speed up the evaporation of solvents to minimize crystal sizes. The
instrument was calibrated using a standard peptide mixture, contain-
ing angiotensin II (1045.54Da), angiotensin I (1295.68Da), substance
P (1346.73Da), bombesin (1618.82Da), ACTH clip 1–17 (2092.08Da),
and ACTH clip 18–39 (2464.19Da), (Bruker Daltonic GmbH,
Germany). Spectra were recorded in the reflectron mode within a
mass range from m/z 500 to m/z 3000 and are the results of 20–100
shots.
Further investigation of fractions by means of Q-TOF mass spec-
trometry. Nanoflow electrospray ionization (ESI) double quadrupole
(Qq) orthogonal accelaration (oa) time-of-flight (TOF) mass spec-
trometry (MS) was performed on a Q-TOF system (Micromass, UK).
Two microliter (representing 1 equivalent, taken from the sample so-
lution prepared previously for MALDI-TOF MS) was loaded into a
gold-coated capillary (Long NanoES spray capillaries for Micromass
topic [M+H]þ are given in the spectra. Underlined sequences were de
agments of cryptocyanin.

Table 1
Peptides identified in a brain–thoracic ganglion extract from Cancer borealis by mass spectrometric techniques
Peptide name Peptide sequence Molecular mass (Monoisotopic) Fraction
Measured Theoretical
Allatostatins
Carcinustatin 2 EAYAFGLamide 768.3 768.4 41
Carcinustatin 5 NPYAFGLamide 779.3 779.4 22/25
Carcinustatin 8 AGPYAFGLamide 793.4 793.4 27
Carcinustatin 9 GGPYAFGLamide 779.3 779.4 22/25
Carcinustatin 19 APTDMYSFGLamide 1099.5 1099.5 24
Penaeustatin 12 SDYAFGLamide 770.4 770.4 43
Penaeustatin 19 GHYNFGLamide 805.4 805.4 17
Penaeustatin 22 AAPYEFGLamide 865.3 865.4 25
Penaeustatin 34 pQNMYSFGLamide 940.3 940.4 25
RFamides
NF 1 NRNFLRFamide 964.6 964.5 31/32
Cab-FLP 1 TNRNFLRFamide 1065.6 1065.6 31
Pem-FLP 5 SMPSLRLRFamide 1104.5 1104.6 34
[Val2]-FLP 6 DVRTPALRLRFamide 1341.9 1341.8 32
Pem-FLP 7 GYRKPPFNGSIFamide 1380.7 1380.7 33
Pem-PYF 3 YAIAGRPRFamide 1048.7 1048.6 18
[Ala1]-FaRP AYNRSFLRFamide 1171.9 1171.6 33
[Glu2–Leu3]-SchistoFLRFa PELDHVFLRFamide 1270.8 1270.7 39
APQRNFLRFamide 1146.8 1146.6 32
LNPFLRFamide 904.6 904.5 28
LRNLRFamide 816.6 816.5 30
Kinins
Pev-Kinin 2 DFSAWAamide 694.4 694.3 48
Pev-Kinin 4 VAFSPWGamide 761.3 761.4 16
Tachykinin related peptide APSGFLGMRamide 933.50 933.49 31
Orcokinin like peptides
FDAFTTGFGHS 1185.5 1185.5 33
NFDEIDRSGFA 1269.8 1269.6 33
[Ala13]-orcokinin NFDEIDRSGFGFA 1473.7 1473.7 36
[Val13]-orcokinin NFDEIDRSGFGFV 1501.8 1501.7 38
[Ser9–Val13]-orcokinin FDEIDRSSFGFV 1531.8 1531.7 38
Peptide sequences in bold were sequenced using ESI-Qqoa-TOF MS/MS, underlined sequences were sequenced de novo. All measured molecular
masses that were present in MALDI-TOF spectra are in italics.
538 J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544
Q-TOF, Proxeon Biosystems A/S, Denmark). The sample was sprayed
at a typical flow rate of 30 nl/min giving extended analysis time in
which MS spectra as well as several MS2 spectra were acquired. During
MS2 or tandem mass spectrometry, fragment ions are generated from a
selected precursor ion by collision-induced dissociation (CID). Since
not all peptide ions fragment with the same efficiency the collision
energy is typically varied between 20 and 35V so that the parent ion is
fragmented into a satisfying number of different daughter ions. The
amino acid sequence is determined by calculating the m/z difference
(which corresponds to the mass of an amino acid residue) between the
adjacent y-ion peaks and/or b-ion peaks.
Results and discussion
HPLC separation on the XTerra C18 column of an
extract of 50 brain–thoracic ganglion equivalents, ini-
tially prepurified on a C18 reverse phase Sep-pak, yielded
a satisfying separation of components, bearing in mindthat no pure fractions were necessary for further anal-
ysis (Fig. 1). All fractions were screened for the presence
of neuropeptides using MALDI-TOF-MS. Examples of
spectra of individual fractions are shown in Fig. 2.
Allatostatins
So far 58 different naturally occurring isoforms of
allatostatins are identified in crustaceans. Twenty alla-
tostatins were isolated from the shore crab, C. maenas
[21], 39 in the tiger prawn, Penaeus monodon [22], andthree in the crayfish Orconectes limosus [24]. Based on
mass spectral data we conclude that in C. borealis at
least nine putative isoforms of allatostatins are present,
five of them being identical to those previously identified
in C. maenas and four identical to those previously
identified in P. monodon. These peptides occurred as low
intensity ion peaks in the MALDI-TOF spectra and
were unfortunately not seen in ESI-Qqoa-TOF spectra,nor did any of the ions selected for fragmentation lead
to the identification of new allatostatin isoforms. Other
peptides identified based solely on the appearance of

Fig. 3. Collision induced dissociation spectra of two de novo sequenced members of the RFamide neuropeptide family. The upper panel showing the
fragmentation of LNPFLRFamide and the lower panel showing the fragmentation of LRNLRFamide. a-type, b-type, y-type, and z-type fragment
ions are indicated in the tables. The theoretical fragment ion masses found in the spectrum are indicated in bold. Mass differences between expected
and observed fragment ions are indicated by d.
J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544 539
their corresponding ions in the MALDI TOF survey
were Pev-Kinin 2, Pev-Kinin 4, and the tachykinin
related peptide (Table 1).
FMRFamide-related peptides
One of the most phylogenetically diverse, but struc-
turally similar, family of peptides is the family that
contains the FMRFamide-related peptides. The tetra-peptide, FMRFamide, was the first member to be se-
quenced and was identified as a cardioacceleratory
peptide in the clam, Macrocalista nimbosa [38].
FMRFamide is now regarded as the primary member of
an extensive family of peptides with diverse biological
functions. In crustaceans FMRFamide-like peptides
have been isolated and characterized from pericardial
organs, thoracic ganglia, and from eyestalks [25–30].
TNRNFLRFamide and SDRNFLRFamide were iso-
lated and sequenced from the stomatogastric nervous
system of the crab C. borealis [39]. All these peptides
display versatile physiological functions in crustaceans[40]. Here we provide biochemical evidence for the
presence of at least 11 FMRFamide-related peptides in a
brain–thoracic ganglion extract of C. borealis. NR
NFLRFamide (F32, m/z: 965.6, Fig. 2), TNRNFLR
Famide (F31, m/z: 1066.6), SMPSLRLRFamide, GYR
KPPFNGSIFamide (F33, m/z: 1381.7), and YA-
IAGRPRFamide (F18, m/z: 1049.7) have been charac-
terized previously in other crustacean species (onlyNRNFLRFamide was identified in C. borealis). Most of
these peptides occurred also as double charged ions in
the Q-TOF spectra and were selected for fragmentation.
The resulting fragmention spectra (raw data treated with
MaxEnt3 Software, Micromass, UK) confirmed the

Fig. 4. Collision induced dissociation spectra of two de novo sequenced members of the RFamide neuropeptide family. The upper panel showing the
fragmentation of AYNRSFLRFamide and the lower panel showing the fragmentation of DVRTPALRLRFamide. a-type, b-type, y-type, and z-type
fragment ions are indicated in the tables. The theoretical fragment ion masses found in the spectrum are indicated in bold. Mass differences between
expected and observed fragment ions are indicated by d.
540 J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544
results obtained by MALDI-TOF MS (Table 1). In
addition, six members of the FMRFamide-related pep-
tide family were sequenced de novo using this approach.
These sequences were assigned as LNPFLRFamide
(Fig. 3 upper panel), LRNLRFamide (Fig. 3 lower pa-
nel), AYNRSFLRFamide (Fig. 4 upper panel), DVRTP
ALRLRFamide (Fig. 4 lower panel), PELDHVFLR
Famide or EPLDHVFLRFamide (Fig. 5 upper panel),and APQRNFLRFamide (Fig. 5 lower panel). AYN
RSFLRFamide and DVRTPALRLRFamide are, ex-
cept for one amino acid, identical to the previously
identified peptides GYNRSFLRFamide (Callinectes
sapidus) and DGRTPALRLRFamide (P. monodon),
respectively. For one peptide the MS/MS spectrum
could be read as EPL DHVFLRFamide or PELDHV
FLRFamide, the feature that a proline residue in posi-
tion 2 protects the N-terminus from general aminopep-
tidases [41] favors the first sequence. Anyway, the C-
terminal (-DHVFLRFamide) is completely identical to
the C-terminal of myosuppressins of other, non-crusta-
cean arthropods including cockroaches, flies, and
grasshoppers [42].
Orcokinins
Orcokinin, NFDEIDRSGFGFN, was the first iden-
tified member (isolated from 1200 abdominal nerve
cords of the crayfish O. limosus) of a peptide family
specific for crustaceans [34]. Activity studies showed thatorcokinins are potent hindgut-stimulating factors [43].
Immunocytochemical studies showed the presence of
orcokinin in all parts of the nervous system [44]. To

Fig. 5. Collision induced dissociation spectra of two de novo sequenced members of the RFamide neuropeptide family. The upper panel showing the
fragmentation of PE(EP)LDHVFLRFamide and the lower panel showing the fragmentation of APQRNFLRFamide. a-type, b-type, y-type, and z-
type fragment ions are indicated in the tables. The theoretical fragment ion masses found in the spectrum are indicated in bold. Mass differences
between expected and observed fragment ions are indicated by d.
J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544 541
date, in total six different orcokinin isoforms are iden-
tified in several crustacean species namely orcokinin,
[Ser9]-orcokinin, [Ala13]-orcokinin, [Val13]-orcokinin
[43], [Thr8–His13]-orcokinin, FDAFTTGFGHS [45],
and [Ala8–Ala13]-orcokinin [46]. Each crustacean species
contains more than one orcokinin isoform. Variations in
isoforms (inter and intra species) are related to point
mutations at the DNA level. In C. borealis we foundbiochemical evidence for the presence of orcokinin (F33,
m/z: 1186.5, Fig. 2), [Ala13]-orcokinin (F36, m/z: 1473.7),
and [Val13]-orcokinin (F38, m/z: 1502.8, Fig. 2). Two
additional orcokinin isoforms, [Ser9–Val13]-orcokinin
(m/z: 766.91, Fig. 6 lower panel) and NFDEIDRSGFA
(m/z: 635.90, Fig. 6 upper panel), were sequenced de
novo. The latter is a shorter peptide than the “classical”
orcokinins, with the amino acids at position 11 and 12
from the original peptide apparantly deleted. These re-
sults are summarized in Table 1.
Protein fragments
Besides peptides, a number of fragments were se-
quenced as well (KIFEPLR, YKIFEPL, YKIFEPLR,
YKIFEPLRS, YKIFEPLRSN, and YKIFEPLRSNL).
These fragments were all present as high intenstity peaks
in the MALDI-TOF spectra (Fig. 2). NCBI BLAST
search revealed that they are fragments of cryptocyanin.
Cryptocyanin was isolated and characterized in Cancer
magister from hepatopancreas tissue [47]. The concen-tration of cryptocyanin in the hemolymph is closely
coordinated with the molt cycle and peaks in late pre-
molt. It is not clear why these fragments appear in a

Fig. 6. Collision induced dissociation spectra of two de novo sequenced orcokinin like neuropeptides. The upper panel showing the fragmentation of
NFDEIDRSGFA and the lower panel showing the fragmentation of NFDEIDRSSFGFV. a-type, b-type, y-type, and z-type fragment ions are
indicated in the tables. The theoretical fragment ion masses found in the spectrum are indicated in bold. Mass differences between expected and
observed fragment ions are indicated by d.
542 J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544
brain–thoracic ganglion extract. This could be due to
contamination of the brain–thoracic ganglion extract
with hemolymph.
Several crustacean neuropeptides within the mass
range of 750–3000 Da were not seen in our peptidomicapproach (ion peaks corresponding to proctolin and
the enkephalins, below 750 Da, were also actively
searched for but not found). This could be attributed
to the following factors: they are not expressed in the
brain–thoracic ganglion (RPCH and PDH), their con-
centration may be below the sensitivity of the used
instrumental setup (CCAP), and not all peptides are
extracted or ionize with the same efficiency. Severalpeptide ion peaks of low intensity that were fragmented
could not be sequenced because of the low quality and
signal intensity of the obtained daughter ions. Never-
theless, the combined use of HPLC fractionation and
mass spectrometric analysis by MALDI-TOF MS and
Q-TOF MS proves to be an elegant tool to analyse the
peptidome of the crustacean nervous system. Starting
from a relative small amount of tissue we were able to
identify 28 peptides, members of four different peptidefamilies, of which eight were sequenced de novo. The
ability to survey an animal for putative neuropeptides
is important for improving our knowledge on the ex-
pression of these signalling molecules within an or-
ganism. A disadvantage, inherent to the mass
spectrometric methods used, is the inability to distin-
guish unequivocally between certain amino acids such
as Gln/Lys and Ile/leu. Therefore, the given amino acidsequences in this study were based on obtained se-
quence data combined with sequences resulting from
NCBI BLAST searches, minimizing errors against Gln/
Lys and Ile/Leu.

J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544 543
Acknowledgments
We thank the members from the laboratory of Michael P. Nus-
baum (Dept. of Neuroscience, University of Pennsylvania school of
Medicine) for providing the C. borealis tissue used in this experiment.
This project was sponsored by the Flemish Science Foundation (Fonds
voor Wetenschappelijk Onderzoek-Vlaanderen, FWO, G.0187.00 and
G.0175.02), and a grant from the National Insitute of Neurological
Disorders and Stroke (NIH NS29436) to M.P.N. J.H. benefits from a
scholarship from the FWO.
References
[1] M.D. Adams et al., The genome sequence of Drosophila melano-
gaster, Science 287 (2000) 2185–2195.
[2] A.N. Starratt, B.E. Brown, Structure of the pentapeptide proct-
olin, a proposed neurotransmitter in insects, Life Sci. 17 (1975)
1253–1256.
[3] G. Baggerman, A. Cerstiaens, A. De Loof, L. Schoofs, Peptido-
mics of the larval Drosophila melanogaster central nervous system,
J. Biol. Chem. 277 (2002) 40368–40374.
[4] M. Yamashita, J.B. Fenn, Negative ion production with the
electrospray ion source, J. Phys. Chem. 88 (1984) 4671–4675.
[5] M. Karas, F. Hillenkamp, Laser desorption ionisation of proteins
with molecular masses exceeding 10 000 Daltons, Anal. Chem. 60
(1988) 1299–1301.
[6] D. Veelaert, G. Baggerman, R. Derua, E. Waelkens, T. Meeusen,
G. Van de Water, A. De Loof, L. Schoofs, Identification of a new
tachykinin from the midgut of the desert locust, Schistocerca
gregaria, by ESI-Qq-oa-TOF mass spectrometry, Biochem. Bio-
phys. Res. Commun. 266 (1999) 237–242.
[7] E. Clynen, G. Baggerman, D. Veelaert, A. Cerstiaens, D. Van der
Horst, L. Harthoorn, R. Derua, E. Waelkens, A. De Loof, L.
Schoofs, Peptidomics of the pars intercerebralis–corpus cardiacum
complex of the migratory locust, Locusta migratoria, Eur.
J. Biochem. 268 (2001) 1929–1939.
[8] E. Clynen, J. Huybrechts, A. De Loof, L. Schoofs, Mass
spectrometric analysis of the perisympathetic organs in locusts:
identification of novel periviscerokinins, Biochem. Biophys. Res.
Commun. 300 (2003) 422–428.
[9] R. Predel, Peptidergic neurohemal system of an insect: mass
spectrometric morphology, J. Comp. Neurol. 436 (2001) 363–
375.
[10] L. Li, R.W. Garden, E.V. Romanova, J.V. Sweedler, In situ
sequencing of peptides from biological tissues and single cells
using MALDI-PSD/CID analysis, Anal. Chem. 71 (1999) 5451–
5458.
[11] S.S. Rubakhin, R.W. Garden, R.R. Fuller, J.V. Sweedler,
Measuring the peptides in individual organelles with mass
spectrometry, Nat. Biotechnol. 18 (2000) 172–175.
[12] J. Stangier, C. Hilbich, K. Beyreuther, R. Keller, Unusual
cardioactive peptide (CCAP) from pericardial organs of the shore
crab, Carcinus maenas, Proc. Natl. Acad. Sci. USA 84 (1987) 575–
580.
[13] H. Dircksen, Distribution and physiology of the crustacean
cardioactive peptide in arthropods, in: K.G. Davey, R.E. Peter,
S.S. Tobe (Eds.), Perspectives in Comparative Endocrinology,
National Research Council of Canada, Ottawa, 1994, pp. 138–
148.
[14] H. Dircksen, R. Keller, Immunocytochemical localization of
CCAP, a novel crustacean cardioactive peptide, in the nervous
system of the shore crab, Carcinus maenas L., Cell Tissue Res. 254
(1988) 347–360.
[15] T.L. Schwarz, G.M.H. Lee, K.K. Siwicki, D.G. Standaert, E.A.
Kravitz, Proctolin in the lobster: the distribution, release and
chemical characterization of a likely neurohormone, J. Neurosci. 4
(1984) 1300–1311.
[16] J. Stangier, H. Dircksen, R. Keller, Identification and immu-
nocytochemical localization of proctolin in pericardial organs
of the shore crab, Carcinus maenas, Peptides 7 (1986) 67–72.
[17] A.J. Mercier, A.B. Lange, V. TeBrugge, I. Orchard, Evidence for
proctolin-like and RFamide-like neuropeptides associated with
the hindgut of the crayfish Procambarus clarkii, Can. J. Zool. 75
(1997) 1208–1225.
[18] P. Fernlund, L. Josefsson, Crustacean color change hormone:
amino acid sequence and chemical synthesis, Science 177 (1972)
173–175.
[19] G. Gaus, L.H. Kleinholz, G. Kegel, R. Keller, Isolation and
characterization of red-pigment-concentrating hormone (RPCH)
from six crustacean species, J. Comp. Physiol. B 160 (1990) 373–
379.
[20] P. Fernlund, Structure of a light-adapting hormone from the
shrimp Pandalus borealis, Biochim. Biophys. Acta 439 (1976) 17–
25.
[21] H. Duve, A.H. Johnsen, J.L. Maestro, A.G. Scott, P.P. Jaros, A.
Thorpe, Isolation and indentification of multiple neuropeptides of
the allatostatin superfamily in the shore crab Carcinus maenas,
Eur. J. Biochem. 250 (1997) 727–734.
[22] H. Duve, A.H. Johnsen, A.G. Scott, A. Thorpe, Allatostatins of
the tiger prawn, Penaeus monodon (Crustacea: Penaeidea), Pep-
tides 23 (2002) 1039–1051.
[23] H. Dircksen, P. Skiebe, B. Abel, H. Agricola, K. Buchner, J.E.
Muren, D.R. N€aassel, Structure, distribution, and biological
activity of novel members of the allatostatin family in the crayfish
Orconectes limosus, Peptides 20 (1999) 695–712.
[24] W.G. Bendena, B.C. Donly, S.S. Tobe, Allatostatins: a growing
family of neuropeptides with structural and functional diversity,
Ann. N. Y. Acad. Sci. 897 (1999) 311–329.
[25] B.A. Trimmer, L.A. Kobierski, E.A. Kravitz, Purification and
characterization of FMRFamide-like immunoreactive substances
from the lobster nervous system: isolation and sequence analysis
of two closely related peptides, J. Comp. Neurol. 266 (1987) 16–
26.
[26] K.G. Krajniak, The identification and stucture-activity relations
of a cardioactive FMRFamide-related peptide from the blue crab
Callinectes sapidus, Peptides 12 (1991) 1295–1302.
[27] A.J. Mercier, I. Orchard, V. TeBrugge, M. Skerret, Isolation of
two FMRFamide-related peptides from crayfish pericardial
organs, Peptides 14 (1993) 137–143.
[28] P. Sithigorngul, W. Saraithonkum, S. Jaideechoey, S. Longy-
ant, W. Sithigorngul, Novel FMRFamide-like neuropeptides
from the eyestalk of the giant freshwater prawn Macrobrach-
ium rosenbergii, Comp. Biochem. Physiol. B 120 (1998) 587–
595.
[29] P. Sithigorngul, W. Saraithonkum, S. Longyant, N. Panchan, W.
Sithigorngul, A. Petsom, Three more FMRF-amide-like neuro-
peptide sequences from the giant freshwater prawn Macrobrach-
ium rosenbergii, Peptides 22 (2001) 191–197.
[30] P. Sithigorngul, J. Pupuem, C. Krungkasem, S. Longyant, P.
Chaivisuthangkura, W. Sithigorngul, A. Petsom, Seven novel
FMRFamide-like neuropeptide sequences from the eyestalk of the
giant tiger prawn Penaeus monodon, Comp. Biochem. Physiol. B
131 (2002) 325–337.
[31] I. Orchard, A.B. Lange, W.G. Bendena, FMRFamide-related
peptides: a multifunctional family of structurally related neuro-
peptides in insects, Adv. Insect Physiol. 28 (2001) 267–329.
[32] P. Torfs, J. Nieto, D. Veelaert, D. Boon, G. Van de Water, E.
Waelkens, R. Derua, J. Calderon, A. De Loof, L. Schoofs, The
kinin peptide family in invertebrates, Ann. N. Y. Acad. Sci. 897
(1999) 361–373.
[33] J. Nieto, D. Veelaert, R. Derua, E. Waelkens, A. Cerstiaens, G.
Coast, B. Devreese, J. Van Beeumen, J. Calderon, A. De Loof, L.

544 J. Huybrechts et al. / Biochemical and Biophysical Research Communications 308 (2003) 535–544
Schoofs, Identification of one tachykinin- and two kinin-related
peptides in the brain of the white shrimp, Penaeus vannamei,
Biochem. Biophys. Res. Commun. 248 (1998) 406–411.
[34] J. Stangier, C. Hilbich, S. Burdzik, R. Keller, Orcokinin: a novel
myotropic peptide from the nervous system of the crayfish,
Orconectes limosus, Peptides 13 (1992) 859–864.
[35] D. Bungart, C. Hilbich, H. Dircksen, R. Keller, Occurrence of
analogs of the myotropic neuropeptide orcokinin in the shore crab
Carcinus maenas: evidence for a novel neuropeptide family,
Peptides 16 (1995) 67–72.
[36] H. Dircksen, S. Burdzik, A. Sauter, R. Keller, Two orcokinins and
the novel octapeptide orcomyotropin in the hindgut of the crayfish
Orconectes limosus: identified myostimulatory neuropeptides orig-
inating together in neurones of the terminal abdominal ganglion,
J. Exp. Biol. 203 (2000) 2807–2818.
[37] L. Schoofs, G.M. Holman, T.K. Hayes, A. Tips, R.J. Nachman,
F. Vandesande, A. De Loof, Isolation, identification and synthesis
of locustamyotropin (Lom-MT), a novel biologically active insect
peptide, Peptides 11 (1989) 427–433.
[38] D.A. Price, M.J. Greenberg, Structure of a molluscan cardioex-
citatory neuropeptide, Science 197 (1977) 670–671.
[39] J.M. Weimann, E. Marder, B. Evans, R.L. Calabrese, The effects
of SDRNFLRFamide and TNRNFLRFamide on the motor
patterns of the stomatogastric ganglion of the crab Cancer
borealis, J. Exp. Biol. 181 (1993) 1–26.
[40] A.J. Mercier, R. Friedrich, M. Boldt, Physiological functions of
FMRFamide-like peptides (FLPs) in crustaceans, Microsc. Res.
Tech. 60 (2003) 313–324.
[41] R.E. Isaac, R.J. Siviter, P. Stancombe, D. Coates, D. Shirras,
Conserved roles for peptidases in the processing of invertebrate
neuropeptides, Biochem. Soc. Trans. 28 (2000) 460–464.
[42] L. Schoofs, G.M. Holman, L. Paemen, D. Veelaert, M. Ame-
linckx, A. De Loof, Isolation, identification, and synthesis of
PDVDHFLRFamide (SchistoFLRFamide) in Locusta migratoria
and its association with the male accessory glands, the salivary
glands, the heart, and the oviduct, Peptides 14 (1993) 409–421.
[43] D. Bungart, G. Kegel, S. Burdzik, R. Keller, Structure–activity
relationships of the crustacean myotropic neuropeptide orcokin,
Peptides 16 (1994) 199–204.
[44] D. Bungart, H. Dircksen, R. Keller, Quantitative-determination
and distribution of the myotropic neuropeptide orcokinin in the
nervous-system of astacidean crustaceans, Peptides 15 (1994) 393–
400.
[45] Y. Yasuda-Kamatani, A. Yasuda, Identification of orcokinin
gene-related peptides in the brain of the crayfish Procambarus
clarkii by the combination of MALDI-TOF and on line capillary
HPLC/Q-TOF mass spectrometries and molecular cloning, Gen.
Comp. Endocrinol. 118 (2000) 161–172.
[46] P. Skiebe, M. Dreger, M. Meseke, J.F. Evers, F. Hucho,
Identification of orcokinins in single neurons in the stomatogastric
nervous system of the crayfish, Cherax destructor, J. Comp.
Neurol. 444 (2002) 245–259.
[47] N.B. Terwilliger, L. Dangott, M. Ryan, Cryptocyanin, a crusta-
cean molting protein: evolutionary link with arthropod hemocy-
anins and insect hexamerins, Proc. Natl. Acad. Sci. USA 96 (1999)
2013–2018.
Related Documents











