Neuromuscular fatigue during a prolonged intermittent exercise: Application to tennis Olivier Girard a, * , Gregory Lattier a , Nicola A. Maffiuletti b , Jean-Paul Micallef a,c , Gregoire P. Millet d a UPRES – EA 2991, Faculty of Sport Sciences, Montpellier, France b Neuromuscular Research Lab, Schulthess Clinic, Zurich, Switzerland c INSERM ADR 08, Montpellier, France d ASPIRE, Academy for Sport Excellence, Doha, Qatar Received 19 February 2007; received in revised form 24 April 2007; accepted 11 May 2007 Abstract The time course of alteration in neuromuscular function of the knee extensor muscles was characterized during a prolonged intermit- tent exercise. Maximal voluntary contraction (MVC) and surface EMG activity of both vastii were measured during brief interruptions before (T 0 ), during (30, 60, 90, 120, 150 and 180 min: T 30 , T 60 , T 90 , T 120 , T 150 , T 180 ) and 30 min after (T +30 ) a 3 h tennis match in 12 trained players. M-wave and twitch contractile properties were analyzed following single stimuli. Short tetani at 20 Hz and 80 Hz were also applied to six subjects at T 0 and T 180 . Significant reductions in MVC (P < 0.05; 9%) and electromyographic activity normalized to the M wave for both vastii (P < 0.01) occurred with fatigue at T 180 . No significant changes in M-wave duration and amplitude nor in twitch contractile properties were observed. The ratio between the torques evoked by 20 Hz and 80 Hz stimulation declined significantly (P < 0.001; 12%) after exercise. Central activation failure and alterations in excitation–contraction coupling are probable mechanisms contributing to the moderate impairment of the neuromuscular function during prolonged tennis playing. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Intermittent exercise; Maximal voluntary contraction; Muscle activation; M-wave; Low- and high-frequency electrical stimulation 1. Introduction Fatigue is a complex phenomenon that can be described as a time-dependent exercise-induced reduction in the max- imal force generating capacity of a muscle (Gandevia, 2001). Alteration in performance tends to vary across sports that are influenced more or less by factors like decreased muscular power and endurance, decreased motor skill performance, and mental lapses. It is well known that impairment of performance resulting from neuromuscular fatigue differs according to the type of contraction, the muscular group tested and the exercise duration/intensity (Millet and Lepers, 2004). Fatigue can therefore originate from several sites that are usually classified as proximal (central fatigue) and/or distal (peripheral fatigue) to the neuromuscular junction (Fitts, 1994; Gandevia, 2001). Some recent experiments have investigated the origin of strength loss after prolonged continuous exercises such as running, cycling or ski skating (for review, see Millet and Lepers, 2004). For example, Millet et al. (2002) found that 65 km of running induced a 30% decrease in knee exten- sors (KE) maximal voluntary contraction (MVC) that was mainly explained by a decrease in maximal voluntary acti- vation. Place et al. (2004) reported that both KE muscle activation and M-wave characteristics were altered but only toward the final stages during a 5 h running exercise. So, the central activation failure and alterations in muscle action potential transmission were contributing to the impairment of the neuromuscular function observed. 1050-6411/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.jelekin.2007.05.005 * Corresponding author. Tel.: +33 4 67 41 57 49; fax: +33 4 67 41 57 50. E-mail address: [email protected] (O. Girard). Journal of Electromyography and Kinesiology 18 (2008) 1038–1046 www.elsevier.com/locate/jelekin

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Electromyography and Kinesiology 18 (2008) 1038–1046
www.elsevier.com/locate/jelekin
Neuromuscular fatigue during a prolonged intermittentexercise: Application to tennis
Olivier Girard a,*, Gregory Lattier a, Nicola A. Maffiuletti b, Jean-Paul Micallef a,c,Gregoire P. Millet d
a UPRES – EA 2991, Faculty of Sport Sciences, Montpellier, Franceb Neuromuscular Research Lab, Schulthess Clinic, Zurich, Switzerland
c INSERM ADR 08, Montpellier, Franced ASPIRE, Academy for Sport Excellence, Doha, Qatar
Received 19 February 2007; received in revised form 24 April 2007; accepted 11 May 2007
Abstract
The time course of alteration in neuromuscular function of the knee extensor muscles was characterized during a prolonged intermit-tent exercise. Maximal voluntary contraction (MVC) and surface EMG activity of both vastii were measured during brief interruptionsbefore (T0), during (30, 60, 90, 120, 150 and 180 min: T30, T60, T90, T120, T150, T180) and 30 min after (T+30) a 3 h tennis match in 12trained players. M-wave and twitch contractile properties were analyzed following single stimuli. Short tetani at 20 Hz and 80 Hz werealso applied to six subjects at T0 and T180. Significant reductions in MVC (P < 0.05; �9%) and electromyographic activity normalized tothe M wave for both vastii (P < 0.01) occurred with fatigue at T180. No significant changes in M-wave duration and amplitude nor intwitch contractile properties were observed. The ratio between the torques evoked by 20 Hz and 80 Hz stimulation declined significantly(P < 0.001; �12%) after exercise. Central activation failure and alterations in excitation–contraction coupling are probable mechanismscontributing to the moderate impairment of the neuromuscular function during prolonged tennis playing.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Intermittent exercise; Maximal voluntary contraction; Muscle activation; M-wave; Low- and high-frequency electrical stimulation
1. Introduction
Fatigue is a complex phenomenon that can be describedas a time-dependent exercise-induced reduction in the max-imal force generating capacity of a muscle (Gandevia,2001). Alteration in performance tends to vary acrosssports that are influenced more or less by factors likedecreased muscular power and endurance, decreased motorskill performance, and mental lapses. It is well known thatimpairment of performance resulting from neuromuscularfatigue differs according to the type of contraction, themuscular group tested and the exercise duration/intensity(Millet and Lepers, 2004). Fatigue can therefore originate
1050-6411/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jelekin.2007.05.005
* Corresponding author. Tel.: +33 4 67 41 57 49; fax: +33 4 67 41 57 50.E-mail address: [email protected] (O. Girard).
from several sites that are usually classified as proximal(central fatigue) and/or distal (peripheral fatigue) to theneuromuscular junction (Fitts, 1994; Gandevia, 2001).
Some recent experiments have investigated the origin ofstrength loss after prolonged continuous exercises such asrunning, cycling or ski skating (for review, see Millet andLepers, 2004). For example, Millet et al. (2002) found that65 km of running induced a �30% decrease in knee exten-sors (KE) maximal voluntary contraction (MVC) that wasmainly explained by a decrease in maximal voluntary acti-vation. Place et al. (2004) reported that both KE muscleactivation and M-wave characteristics were altered butonly toward the final stages during a 5 h running exercise.So, the central activation failure and alterations in muscleaction potential transmission were contributing tothe impairment of the neuromuscular function observed.

O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046 1039
However, to our knowledge, there is no informationregarding the contribution of central and peripheral factorsto muscle fatigue during a prolonged high-intensity exercisewith a discontinuous load profile such as racquet games.
Tennis can be played over several hours (1–5 h) of briefperiods (5–10 s) of high intensity work (60–75% of maximaloxygen uptake) interspersed with periods (15–20 s) oflighter activity or rest (Fernandez et al., 2006). The inter-mittent nature of this sport is highlighted by the small totalplaying time of 20–30% of total match duration and by thework to rest ratio between 1:1 and 1:4. A strong point ofhaving tennis as a model is that the motivation of the sub-ject remains very high during this intermittent exercisesince it is a dual game. Although a large number of studies(for review, see Fernandez et al., 2006) have focused on thecharacteristics of tennis players during actual match play(e.g., physiological responses, point duration, speed andagility, strength, flexibility) – which provides a great insightinto the energy demands of tennis – less is known, however,on the influence of fatigue on tennis performance (Horneryet al., 2007). It has been recently demonstrated that pro-gressive reductions in MVC and leg stiffness highly corre-lated with increases in perceived exertion and musclesoreness throughout a 3 h tennis match, whereas explosivestrength was maintained and decreased only after thematch (Girard et al., 2006). Nevertheless, to the best ofour knowledge, no previous study has focused on the pos-sible mechanisms responsible for alteration of neuromuscu-lar function induced by this type of activity. In addition,the majority of previous fatigue experiments examinedfunctional impairments induced by prolonged continuousactivity by looking at the difference between before andafter exercise, while little information exists on changesoccurring in the neuromuscular system as exercise pro-gresses (Millet and Lepers, 2004).
The aim of this study was therefore to examine the timecourse of impairment in neural and contractile processesduring prolonged tennis playing. To examine the centraland peripheral components of fatigue, changes in volun-tary and electrically induced (both superimposed and onthe relaxed muscle) contractions of the KE muscles werequantified during interruptions (<10 min) every 30 minduring a 3 h tennis match. Based on previous research oncontinuous exercise (Millet and Lepers, 2004), we hypoth-esized that progressive reduction in muscle activation dur-ing such prolonged intermittent exercise might accompanyperipheral fatigue.
2. Methods
2.1. Subjects
Twelve well-trained male tennis players (mean ± SD age:23.4 ± 1.6 yr; mass: 72.2 ± 4.5 kg; height: 178.5 ± 4.3 cm; bodyfat: 14.1 ± 3.0%) of regional to national level (International tennisnumber ranging from 2 to 4) volunteered to participate in thisstudy after they were informed in detail about the nature of theexperiment and possible risks. The subjects were practicing tennis
regularly for at least 10 yr before the study, and their averageweekly technical/tactical and physical training during the 6months before testing were 4.9 ± 3.2 and 2.5 ± 1.4 h/wk, respec-tively. Written informed consent was given by each subject, andthe local Committee on Human Research gave their approval forthe project.
2.2. Experimental design
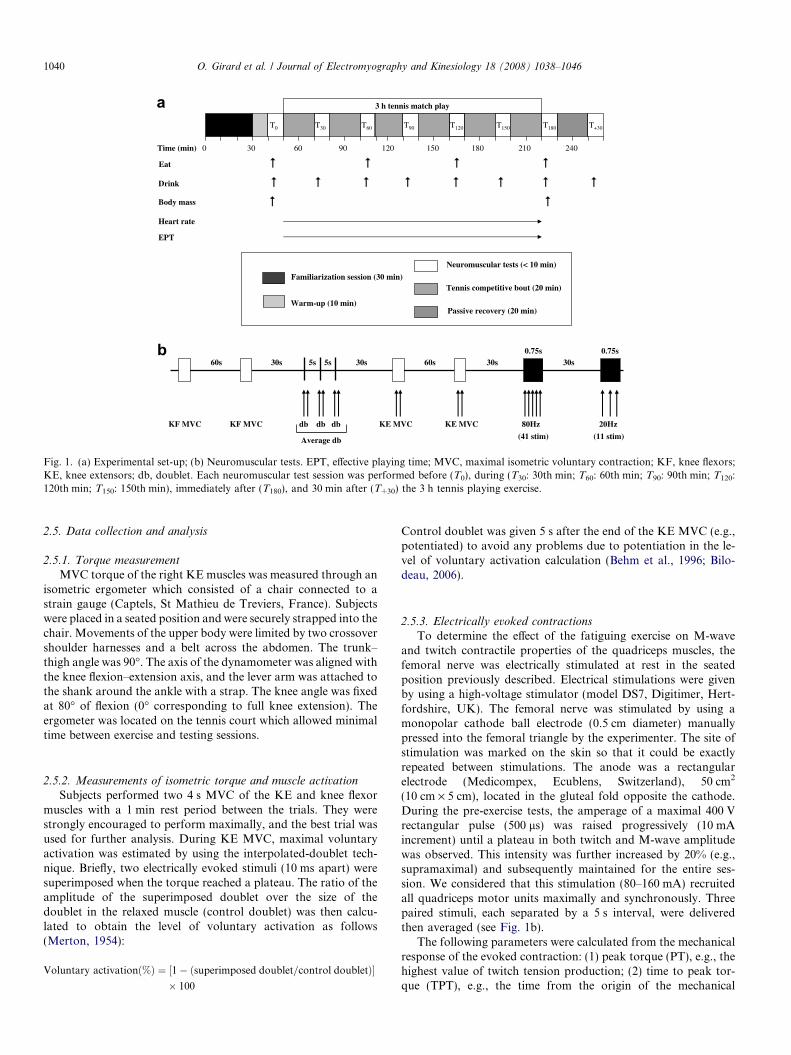
The experimental design is depicted in Fig. 1a. Each subjectwas initially familiarized with several sub-maximal (range 1–30 mA) electrical stimuli and with the isometric ergometer forMVC torque testing. Subjects were requested to perform threeKE MVCs (4 s duration) with a rest interval of �1 min. Foreach subject, the coefficient of variation across trials was calcu-lated. If the maximal torque varied to a large extent (coefficientof variation >5%), three additional KE MVCs were performed.Afterwards, a passive isometric recruitment curve was drawn tocarefully search the optimal stimulus intensity for the maximalM-wave response (see electrically evoked contractions). A stan-dardized warm-up was carried out by each subject before thetesting session. It consisted of 5 min of running at a self-selectedpace (8–10 km/h), followed by 5 min of tennis play. Subsequentto this warm-up protocol, neuromuscular tests including MVCtesting and electrically induced contractions were operated in theorder presented in Fig. 1b. These neuromuscular tests wereconducted before (T0), during (30, 60, 90, 120, and 150 min:T30, T60, T90, T120, T150, respectively), immediately after (T180)and 30 min after (T+30) a 3 h standardized tennis match. Astrict timing regimen allowed maintaining similar conditionsamong repeated measurements. All experiments were performedon a Greenset� tennis court area. The air temperature was 12–15 �C.
2.3. Intermittent exercise
The prolonged fatiguing exercise was a 3 h standardizedcompetitive tennis match (excluding the 10 min warm-up period).The scoring and time characteristics complied with the rules of theInternational Tennis Federation. During tennis match, playerswere therefore allowed to rest for a maximum of 20 s betweenrallies, but no pauses interspersed the games. All subjects playedagainst an opponent of similar standard. Due to the competitivenature of a tennis game, each subject was well motivated to per-form maximally over the total test duration. During the 3 h tennismatch, subjects ingested 250 ml water every 30 min and a 77 kcalenergetic biscuit at the first and second hour of play to preventdehydration and hypoglycaemia. Body mass was measured(Tanita TBF – 300GS, Neuilly-Sur-Seine, France) in all subjectsbefore (1 h after subject’s breakfast) and immediately after thefatiguing exercise.
2.4. Exercise characteristics
Heart rate was recorded continuously and averaged every 5 sduring the match using short range radio telemetry (S610; PolarElectro Oy, Kempele, Finland). A researcher timed the durationof each rally in order to calculate the following mean variables: (1)the duration of rallies (s); and (2) the effective playing time (%),corresponding to the ratio between the sum of duration of ralliesand the total playing time (Girard et al., 2006).
0 30 60 90 120 150 180 210 240
Eat
Drink
Warm-up (10 min)
Tennis competitive bout (20 min)
Passive recovery (20 min)
3 h tennis match play
Heart rate
T30 T60 T90 T120 T150 T180 T+30T0
Time (min)
EPT
Neuromuscular tests (< 10 min)
Body mass
Familiarization session (30 min)
a
b60s 30s 5s 5s 30s 60s 30s 30s
Average db (41 stim) (11 stim)
KF MVC KF MVC db db db KE MVC KE MVC 80Hz 20Hz
0.75s 0.75s
Fig. 1. (a) Experimental set-up; (b) Neuromuscular tests. EPT, effective playing time; MVC, maximal isometric voluntary contraction; KF, knee flexors;KE, knee extensors; db, doublet. Each neuromuscular test session was performed before (T0), during (T30: 30th min; T60: 60th min; T90: 90th min; T120:120th min; T150: 150th min), immediately after (T180), and 30 min after (T+30) the 3 h tennis playing exercise.
1040 O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046
2.5. Data collection and analysis
2.5.1. Torque measurement
MVC torque of the right KE muscles was measured through anisometric ergometer which consisted of a chair connected to astrain gauge (Captels, St Mathieu de Treviers, France). Subjectswere placed in a seated position and were securely strapped into thechair. Movements of the upper body were limited by two crossovershoulder harnesses and a belt across the abdomen. The trunk–thigh angle was 90�. The axis of the dynamometer was aligned withthe knee flexion–extension axis, and the lever arm was attached tothe shank around the ankle with a strap. The knee angle was fixedat 80� of flexion (0� corresponding to full knee extension). Theergometer was located on the tennis court which allowed minimaltime between exercise and testing sessions.
2.5.2. Measurements of isometric torque and muscle activation
Subjects performed two 4 s MVC of the KE and knee flexormuscles with a 1 min rest period between the trials. They werestrongly encouraged to perform maximally, and the best trial wasused for further analysis. During KE MVC, maximal voluntaryactivation was estimated by using the interpolated-doublet tech-nique. Briefly, two electrically evoked stimuli (10 ms apart) weresuperimposed when the torque reached a plateau. The ratio of theamplitude of the superimposed doublet over the size of thedoublet in the relaxed muscle (control doublet) was then calcu-lated to obtain the level of voluntary activation as follows(Merton, 1954):
Voluntary activationð%Þ ¼ ½1� ðsuperimposed doublet=control doubletÞ�� 100
Control doublet was given 5 s after the end of the KE MVC (e.g.,potentiated) to avoid any problems due to potentiation in the le-vel of voluntary activation calculation (Behm et al., 1996; Bilo-deau, 2006).
2.5.3. Electrically evoked contractions
To determine the effect of the fatiguing exercise on M-waveand twitch contractile properties of the quadriceps muscles, thefemoral nerve was electrically stimulated at rest in the seatedposition previously described. Electrical stimulations were givenby using a high-voltage stimulator (model DS7, Digitimer, Hert-fordshire, UK). The femoral nerve was stimulated by using amonopolar cathode ball electrode (0.5 cm diameter) manuallypressed into the femoral triangle by the experimenter. The site ofstimulation was marked on the skin so that it could be exactlyrepeated between stimulations. The anode was a rectangularelectrode (Medicompex, Ecublens, Switzerland), 50 cm2
(10 cm · 5 cm), located in the gluteal fold opposite the cathode.During the pre-exercise tests, the amperage of a maximal 400 Vrectangular pulse (500 ls) was raised progressively (10 mAincrement) until a plateau in both twitch and M-wave amplitudewas observed. This intensity was further increased by 20% (e.g.,supramaximal) and subsequently maintained for the entire ses-sion. We considered that this stimulation (80–160 mA) recruitedall quadriceps motor units maximally and synchronously. Threepaired stimuli, each separated by a 5 s interval, were deliveredthen averaged (see Fig. 1b).
The following parameters were calculated from the mechanicalresponse of the evoked contraction: (1) peak torque (PT), e.g., thehighest value of twitch tension production; (2) time to peak tor-que (TPT), e.g., the time from the origin of the mechanical

O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046 1041
response to PT; and (3) half-relaxation time (HRT), e.g., the timeto obtain half of the decline in twitch maximal torque.
Finally, a 0.75 s high-frequency tetanus (80-Hz) and a 0.75 slow-frequency tetanus (20-Hz) (e.g., 41 and 11 stimuli, respec-tively) were evoked on the relaxed muscle. The peak torque ofboth trains of stimuli was measured (P80 and P20, respectively)and the P20/P80 ratio was calculated. However, since evokedtetanus on relaxed muscles were more painful than doublets andsuperimposed stimulations, only six subjects agreed for thismeasure before and immediately after the 3 h exercise. Tetanicstimulation allowed us to assess muscle contractile function moreaccurately, as a single twitch may not be considered a goodindicator of peripheral fatigue (Millet et al., 2003). For the 80-Hzfrequency stimulation, the torque reached during the tetanuscorresponded to 118% of MVC. Finally, the MVC/P80 ratio, e.g.,another method characterising the extent of muscle activation(Bigland-Ritchie et al., 1986; Millet and Lepers, 2004), wascalculated.
2.6. Surface EMG recordings
Electromyographic (EMG) activity of the vastus lateralis (VL),vastus medialis (VM) and biceps femoris (BF) muscles wasrecorded by means of two pairs of silver chloride circular (inter-distance electrode = 20 mm) surface electrodes (ControleGraphique Medical, Brie-Comte-Robert, France) fixed lengthwiseover the middle of the muscle belly (De Luca, 1997). The referenceelectrode was attached to the left wrist. Low impedance betweenthe two electrodes (<3 kX) was obtained by abrading the skinwith emery paper and cleaning with alcohol. Subjects kept theelectrodes on their skin throughout the duration of the entireexperiment; nevertheless, the position of the electrodes wasmarked because for some subjects electrodes had to be replaceddue to excessive sweating. EMG signals were amplified with abandwidth frequency ranging from 30 to 500 Hz (Biopac MP30,Systems Inc., Santa Barbara, CA). Torque and EMG data werestored (sampling frequency 2000 Hz) with commercially availablesoftware (Acqknowledge 3.6.7, Biopac Systems Inc., Santa Bar-bara, CA).
Peak-to-peak amplitude (PPA) and duration (PPD) of M-waves were determined for VL and VM. During KE and kneeflexors MVCs, the EMG of the VL, VM and BF was analyzedover a 1 s period when the torque had reached a plateau (beforethe superimposed doublet) to calculate the root mean square(RMS). RMS values of both vastii were also normalized to theamplitude of respective M-wave amplitudes (e.g., RMS/PPAratio). A reduction in the MVC RMS without a reduction in PPAmay be interpreted as a central activation failure (Millet andLepers, 2004).
The PT/PPA ratio was also calculated as follows:
PT=PPA ¼ PT=ð½PPAVL þ PPAVM�=2Þ ðLattier et al:; 2004Þ
Finally, antagonist coactivation was quantified as follows. BFRMS during KE MVC was calculated over a 1 s period beforethe superimposed twitch. Then, this value was expressed as a per-centage of maximal BF RMS, previously recorded during kneeflexors MVC (see above).
Fig. 2. Maximal isometric voluntary contraction of knee extensor musclesbefore (T0), during (T30: 30th min; T60: 60th min; T90: 90th min; T120:120th min; T150: 150th min), immediately after (T180), and 30 min after(T+30) the 3 h tennis playing exercise. Values are means ± SE. *P < 0.05;**P < 0.01; ***P < 0.001, significantly different from T0.
2.7. Statistical analysis
All data are expressed as means ± SD (tables) and ± SE (fig-ures). P80, P20 and the P20/P80 ratio measured before and
immediately after exercise were compared with a Student’s pairedt-test (two-tailed). Data recorded during the exercise and recoverywere tested using a one-factor (time) ANOVA for repeated mea-sures. When significant main effects were found, the LSD Fisherpost hoc test was used. Statistical significance was accepted atP < 0.05. The statistical analyses were undertaken by using Sigm-aStat 2.03 software (Jandel Corporation, San Rafael, CA, USA).
3. Results
3.1. Exercise characteristics
After exercise, the body mass loss averaged 1.1 ± 0.7 kg(1.6 ± 1.0% of initial body weight). During the 3 h tennismatch, the mean heart rate was 143.8 ± 7.8 beats/min,which represents a mean intensity of 74.1 ± 4.0% of maxi-mal heart rate (estimated as 220-age). Mean duration ofrallies and effective playing time were 7.0 ± 1.3 s and21.3 ± 3.6%, respectively.
3.2. Isometric torque and muscle activation
Analysis of variance revealed a significant effect of exer-cise upon KE MVC (P < 0.01) (Fig. 2). The mean KEMVC torque decreased gradually during the first 2 h of ten-nis play (�1%, �5%, �6% and �7% at T30, T60, T90, andT120 respectively, when compared to T0) but became signif-icantly lower only after 150 min (�13% at T150; P < 0.001).At the end of exercise, the subjects had lost on average 9%of MVC (P < 0.05) at T180 and did not recover 30 min later(�10%; P < 0.01).
RMS/PPA for both VL and VM muscles were signifi-cantly reduced (P < 0.05) by the exercise as shown inFigs. 3a and b. For the VL muscle, the RMS/PPA decreasebecame significant only after the first hour of exercise(�11% at T30, P = 0.06; and �16%, �18%, �12%, �19%and �15% at T60, T90, T120, T150 and T180 respectively,P < 0.05), when compared to T0. After 30 min of recovery,RMS/PPA for the VL muscle was not significantly lowerthan at rest (�7%; P > 0.05). RMS/PPA for the VM

Fig. 3. Electromyographic activity of the vastus lateralis muscle (a), electromyographic activity of the vastus medialis muscle (b), voluntary activationlevel estimated by the superimposed doublet method (c), and antagonist coactivation (d) before (T0), during (T30: 30th min; T60: 60th min; T90: 90th min;T120: 120th min; T150: 150th min), immediately after (T180), and 30 min after (T+30) the 3 h tennis playing exercise. Values are means ± SE. *P < 0.05;**P < 0.01; ***P < 0.001, significantly different from T0.
1042 O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046
muscle started declining at the 30th min of exercise andremained significantly reduced until the end of exerciseand then at T+30. Fig. 3c shows that voluntary activationgradually decreased – though not significantly so –throughout the exercise from T0 to T150 (83.9 ± 1.7 vs.74.3 ± 4.1%; �11%; P = 0.12) but returned almost to ini-tial values at T180 (81.9 ± 3.1%). Antagonist coactivationdid not change significantly (P > 0.05) during exercise(Fig. 3d). For the six subjects who tolerated the tetani,the MVC/P80 ratio declined significantly from pre- to
Table 1M-wave characteristics before (T0), during (T30: 30th min; T60: 60th min;T90: 90th min; T120: 120th min; T150: 150th min), immediately after (T180),and 30 min after (T+30) the 3 h tennis playing exercise for both vastuslateralis and vastus medialis muscles
Vastus Lateralis Vastus Medialis
PPA (mV) PPD (ms) PPA (mV) PPD (ms)
T0 6.94 ± 1.09 7.90 ± 2.52 7.14 ± 1.79 7.28 ± 1.35T30 6.81 ± 1.38 8.18 ± 2.50 7.28 ± 2.09 7.10 ± 1.31T60 6.76 ± 1.08 7.38 ± 2.20 6.97 ± 2.06 7.34 ± 1.82T90 6.56 ± 0.95 7.69 ± 2.06 6.97 ± 1.97 6.71 ± 1.07T120 6.36 ± 1.68 7.87 ± 2.13 7.19 ± 1.68 6.98 ± 1.27T150 6.48 ± 1.29 7.63 ± 1.71 6.30 ± 2.08 6.60 ± 1.43T180 6.54 ± 1.23 7.77 ± 2.38 6.76 ± 2.11 6.98 ± 0.97T+30 6.06 ± 1.45 7.43 ± 1.52 7.56 ± 2.89 7.13 ± 0.92
Values are means ± SD.PPA, peak to peak amplitude (mV); PPD, peak to peak duration (ms).
post-exercise (from 0.82 ± 0.08 to 0.68 ± 0.12; �16%;P < 0.05).
3.3. M-wave and twitch contractile properties
The characteristics of the M-wave were not altered bythe present exercise (Table 1). In the same way, changesin twitch contractile properties were minimal and insignif-icant during exercise (Table 2). PT tended (P = 0.09) todecrease during exercise, and the reduction at T150 reached
Table 2Twitch contractile properties of the quadriceps muscle before (T0), during(T30: 30th min; T60: 60th min; T90: 90th min; T120: 120th min; T150:150th min), immediately after (T180), and 30 min after (T+30) the 3 h tennisplaying exercise
PT (N m) TPT (ms) HRT (ms)
T0 54.1 ± 8.7 73.3 ± 21.7 89.2 ± 17.1T30 52.6 ± 10.4 61.9 ± 14.9 84.6 ± 19.6T60 51.8 ± 8.9 59.2 ± 9.1 80.7 ± 17.9T90 52.5 ± 7.2 61.8 ± 16.0 77.6 ± 17.2T120 50.1 ± 7.3 58.0 ± 9.4 81.0 ± 17.4T150 48.9 ± 6.5 63.6 ± 11.4 85.9 ± 20.2T180 49.9 ± 6.9 63.4 ± 15.3 80.1 ± 22.7T+30 49.7 ± 6.6 66.1 ± 16.1 82.9 ± 13.7
Values are means ± SD.PT, peak torque; TPT, time to peak torque; HRT, half relaxation time.
91
84 848482
8487
100
50
60
70
80
90
100
T0 T30 T60 T90 T120 T150 T180 T+30
PT/P
PAVL
+VM (%
T 0)
** ** *** **** **
a
150
200
250
300
350
400
450
T0 T180
Torq
ue (N
m)
0
0.2
0.4
0.6
0.8
Rat
io P
20/P
80
P80 P20 P20/P80
**
***b
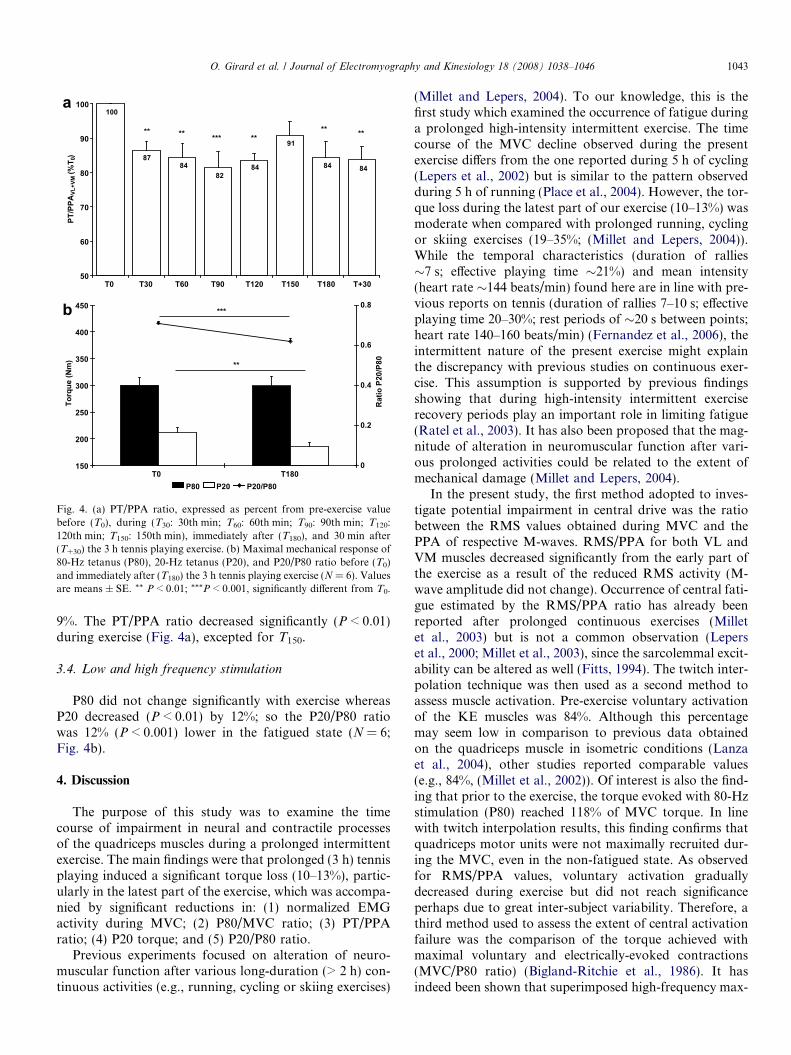
Fig. 4. (a) PT/PPA ratio, expressed as percent from pre-exercise valuebefore (T0), during (T30: 30th min; T60: 60th min; T90: 90th min; T120:120th min; T150: 150th min), immediately after (T180), and 30 min after(T+30) the 3 h tennis playing exercise. (b) Maximal mechanical response of80-Hz tetanus (P80), 20-Hz tetanus (P20), and P20/P80 ratio before (T0)and immediately after (T180) the 3 h tennis playing exercise (N = 6). Valuesare means ± SE. ** P < 0.01; ***P < 0.001, significantly different from T0.
O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046 1043
9%. The PT/PPA ratio decreased significantly (P < 0.01)during exercise (Fig. 4a), excepted for T150.
3.4. Low and high frequency stimulation
P80 did not change significantly with exercise whereasP20 decreased (P < 0.01) by 12%; so the P20/P80 ratiowas 12% (P < 0.001) lower in the fatigued state (N = 6;Fig. 4b).
4. Discussion
The purpose of this study was to examine the timecourse of impairment in neural and contractile processesof the quadriceps muscles during a prolonged intermittentexercise. The main findings were that prolonged (3 h) tennisplaying induced a significant torque loss (10–13%), partic-ularly in the latest part of the exercise, which was accompa-nied by significant reductions in: (1) normalized EMGactivity during MVC; (2) P80/MVC ratio; (3) PT/PPAratio; (4) P20 torque; and (5) P20/P80 ratio.
Previous experiments focused on alteration of neuro-muscular function after various long-duration (> 2 h) con-tinuous activities (e.g., running, cycling or skiing exercises)
(Millet and Lepers, 2004). To our knowledge, this is thefirst study which examined the occurrence of fatigue duringa prolonged high-intensity intermittent exercise. The timecourse of the MVC decline observed during the presentexercise differs from the one reported during 5 h of cycling(Lepers et al., 2002) but is similar to the pattern observedduring 5 h of running (Place et al., 2004). However, the tor-que loss during the latest part of our exercise (10–13%) wasmoderate when compared with prolonged running, cyclingor skiing exercises (19–35%; (Millet and Lepers, 2004)).While the temporal characteristics (duration of rallies�7 s; effective playing time �21%) and mean intensity(heart rate �144 beats/min) found here are in line with pre-vious reports on tennis (duration of rallies 7–10 s; effectiveplaying time 20–30%; rest periods of �20 s between points;heart rate 140–160 beats/min) (Fernandez et al., 2006), theintermittent nature of the present exercise might explainthe discrepancy with previous studies on continuous exer-cise. This assumption is supported by previous findingsshowing that during high-intensity intermittent exerciserecovery periods play an important role in limiting fatigue(Ratel et al., 2003). It has also been proposed that the mag-nitude of alteration in neuromuscular function after vari-ous prolonged activities could be related to the extent ofmechanical damage (Millet and Lepers, 2004).
In the present study, the first method adopted to inves-tigate potential impairment in central drive was the ratiobetween the RMS values obtained during MVC and thePPA of respective M-waves. RMS/PPA for both VL andVM muscles decreased significantly from the early part ofthe exercise as a result of the reduced RMS activity (M-wave amplitude did not change). Occurrence of central fati-gue estimated by the RMS/PPA ratio has already beenreported after prolonged continuous exercises (Milletet al., 2003) but is not a common observation (Leperset al., 2000; Millet et al., 2003), since the sarcolemmal excit-ability can be altered as well (Fitts, 1994). The twitch inter-polation technique was then used as a second method toassess muscle activation. Pre-exercise voluntary activationof the KE muscles was 84%. Although this percentagemay seem low in comparison to previous data obtainedon the quadriceps muscle in isometric conditions (Lanzaet al., 2004), other studies reported comparable values(e.g., 84%, (Millet et al., 2002)). Of interest is also the find-ing that prior to the exercise, the torque evoked with 80-Hzstimulation (P80) reached 118% of MVC torque. In linewith twitch interpolation results, this finding confirms thatquadriceps motor units were not maximally recruited dur-ing the MVC, even in the non-fatigued state. As observedfor RMS/PPA values, voluntary activation graduallydecreased during exercise but did not reach significanceperhaps due to great inter-subject variability. Therefore, athird method used to assess the extent of central activationfailure was the comparison of the torque achieved withmaximal voluntary and electrically-evoked contractions(MVC/P80 ratio) (Bigland-Ritchie et al., 1986). It hasindeed been shown that superimposed high-frequency max-

1044 O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046
imal trains of stimuli may improve the detection of centralactivation failure during isometric knee extension (Milleret al., 1999). Therefore the present reduction in MVC/P80 ratio (�16%) observed after the fatiguing exerciseemphasizes the role of central fatigue in the torquedecrease.
As a whole, the three methods used for estimating mus-cle activation strongly suggest that central components ofmuscle fatigue play a major role in the marked decreaseof force observed during prolonged tennis playing. How-ever, by measuring an activation deficit with these tech-niques, it is not possible to determine if alterations ofneuromuscular activity during such a prolonged exercisecould be due to an impairment in command generated inthe higher cortical structures (supraspinal site) and/or inresponse to afferent inputs from metabolic changes in themuscle (spinal level) (Gandevia, 2001). It has been shownthat the reduction in corticospinal impulses reaching moto-neurons was partly due to an alteration in neurotransmitter(e.g., serotonin, acetylcholine and dopamine) function inthe brain (Davis and Bailey, 1997). Considering that a4 h tennis tournament previously induced a 28% declinein branched-chain amino acids (Struder et al., 1995) –which may impair the brain’s capacity to recruit the opti-mal number of motor units – central fatigue could beexpected after tennis exercise. However, it is not clear ifincreases in neurotransmitter concentrations are causedby the increased production in the peripheral tissues oftheir precursors, which then cross the blood-brain barrier,or whether they result from increased neural activity (Gue-zennec et al., 1998). Nevertheless, change in cortical excit-ability per se is probably not the only cause of centralfatigue. This could also occur at the spinal level fromperipheral reflex inhibition of the a-motoneuron pool viagroup III and IV afferents due to mechanical and biochem-ical changes in exercising muscles (Garland, 1991), disfacil-itation of a-motoneurons by muscle spindle afferents(Bongiovanni and Hagbarth, 1990) and/or recurrent inhi-bition of a-motoneuron (Kukulka et al., 1986). Regardingthis latter point, it is known that antagonist coactivationmay be affected by fatigue: for example, the relativeamount of hamstring coactivation increased duringexhausting isokinetic knee extensions at both slow and fastspeeds (Weir et al., 1998). However, it is noteworthy that inthe present study biceps femoris coactivation wasunchanged with fatigue therefore indicating that this mech-anism was not involved in the isometric MVC torque dec-rement. Nonetheless, further insight on spinal modulationunder fatigue (e.g., 30 km (Millet et al., 2003) or marathonrunning (Avela et al., 1999)) support the view that muscledamage alters the lower limb stiffness regulation throughchange in the afferent input from the muscle (pre-synapticinhibition and/or stretch-reflex disfacilitation) (Komi,2000). These changes in mechanical performance that areassociated with a decreased tolerance to imposed stretchload (Horita et al., 1999) may have in turn reduced thestretch-shortening cycle performance (e.g., powerful stroke
production and rapid movement about the court) in thepresent study (Girard et al., 2006).
The unchanged M-wave duration and amplitude withfatigue is a novel finding, suggesting that a failure in theneuromuscular transmission-propagation of action poten-tials did not occur in this type of prolonged exercise (Fitts,1994). After prolonged continuous exercises, significantreductions in M-wave amplitude have been previouslyobserved (Millet and Lepers, 2004). However, M-wavecharacteristics must be analyzed with caution, since anincrease in extracellular potassium may inhibit the propa-gation of muscle action potentials in sites distal to sarco-lemma, as in T-tubules (West et al., 1996). Furthermore,changes in EMG amplitude may also be confounded bythe presence of cancellation, resulting from the overlappingpositive and negative phases of individual-single motorunit potentials (Farina et al., 2004).
In this study, M-wave amplitude remained unchangedwhereas PT/PPA significantly decreased, thus indicatingthe presence of contractile fatigue. Taken together, thesefindings highlight a failure of excitation–contraction cou-pling, likely related to an impaired sarcoplasmic reticulumactivity (Mador et al., 2000). However, since temporal con-tractile properties (e.g., TPT and HRT) were not signifi-cantly modified during exercise, one may speculate thatcontractile velocity was not affected by fatigue. A decreasein peak twitch torque with insignificant changes in tempo-ral parameters after submaximal exhaustive stretch-short-ening cycle exercise has been associated with changes inthe contractile characteristics of the quadriceps femorismuscle (Strojnik and Komi, 2000).
Low- and high-frequency electrical stimulation allows toassess the signal transfer from the excitation of the sarco-lemma to the binding of intracellular Ca2+ to the contrac-tile proteins (Vøllestad, 1997). In the present study, thewell-preserved torque during 80-Hz stimulation indicatedthat the propagation of the muscle action potential wasnot impaired significantly and that the maximum Ca2+-activated force was not altered. In line with the lack ofchange in M-wave characteristics, this result confirms thatthe present intermittent exercise did not induce any mem-brane excitability failure. The reduced P20/P80 ratio inthe fatigued condition showed a low-frequency fatigue(LFF). LFF, also called long-lasting fatigue, involves bothimpaired excitation–contraction coupling and structuraldamage (Jones, 1996; Metzger and Fitts, 1987). LFFimplies a smaller Ca2+ release from sarcoplasmic reticulum(Russeau and Pinkos, 1990) and/or inhibition of Ca2+
binding to troponin (Blanchard et al., 1984), resulting inless active cross-bridges. This is supported by the fact thatPT/PPA ratio decreased significantly during exercise. Evenif the occurrence of excitation–contraction coupling failurehas been already reported following eccentric (Martinet al., 2005) or plyometric (Strojnik and Komi, 2000) exer-cises, to date, no evidence of knee extensor muscles LFFhas been shown after prolonged running exercise (Milletand Lepers, 2004). Although we did not measure any index

O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046 1045
of muscular damage in the present study, LFF was antici-pated because muscle soreness evaluated using a visualanalogue scale has been shown to increase linearly duringprolonged tennis playing (Girard et al., 2006). One maytherefore speculate that the numerous stretch-shorteningcycle movements (e.g., serves, jumps) and eccentric con-tractions (e.g., flexions, breakages) during the 3 h tennisexercise induced some structural impairment. This assump-tion is supported by previous findings (Thompson et al.,1999) showing that the causes of fatigue when performingprolonged intermittent high-intensity shuttle running withsufficient rest periods are not associated with metabolic fac-tors but most-likely related to muscle damage.
5. Conclusion
This study examined the time course of neural andcontractile alterations of the quadriceps muscles duringprolonged (3 h) tennis playing. Fatigue during suchprolonged high-intensity intermittent exercise inducedprogressive reductions in the maximal voluntary KE torque-generating capability. Central activation failure and altera-tions in excitation–contraction coupling (low-frequencyfatigue) are probably the main mechanisms contributingto the torque loss observed. Nevertheless, further studiesare required to examine the adjustments in the spinal andsupraspinal circuitries during prolonged intermittentactivities.
References
Avela J, Kyrolainen H, Komi PV, Rama D. Reduced reflex sensitivitypersists several days after long-lasting stretch-shortening cycle exercise.J Appl Physiol 1999;86(4):1292–300.
Behm DG, St-Pierre DM, Perez D. Muscle inactivation: assessment ofinterpolated twitch technique. J Appl Physiol 1996;81(5):2267–73.
Bigland-Ritchie B, Furbush F, Woods JJ. Fatigue of intermittentsubmaximal voluntary contractions: central and peripheral factors. JAppl Physiol 1986;61(2):421–9.
Bilodeau M. Central fatigue in continuous and intermittent contractionsof triceps brachii. Muscle Nerve 2006;34(2):205–13.
Blanchard EM, Pan BS, Solaro RJ. The effect of acidic pH on the ATPaseactivity and troponin Ca2+ binding of rabbit skeletal myofilaments. JBiol Chem 1984;259(5):3181–6.
Bongiovanni LG, Hagbarth KE. Tonic vibration reflexes elicited duringfatigue from maximal voluntary contractions in man. J Physiol (Lond)1990;423:1–14.
Davis JM, Bailey SP. Possible mechanisms of central nervous systemfatigue during exercise. Med Sci Sports Exerc 1997;29(1):45–57.
De Luca CJ. The use of surface electromyography in biomechanics. J ApplBiomech 1997;13:135–63.
Farina D, Merletti R, Enoka RM. The extraction of neural strategies fromthe surface EMG. J Appl Physiol 2004;96(4):1495–6.
Fernandez J, Mendez-Villanueva A, Pluim B. Intensity of tennis matchplay. Br J Sports Med 2006;40(5):387–91.
Fitts RH. Cellular mechanisms of muscle fatigue. Physiol Rev1994;74(1):49–94.
Gandevia SC. Spinal and supraspinal factors in human muscle fatigue.Physiol Rev 2001;81(4):1725–89.
Garland SJ. Role of small diameter afferents in reflex inhibition duringhuman fatigue. J Physiol 1991;435:547–58.
Girard O, Lattier G, Micallef JP, Millet GP. Changes in exercisecharacteristics, maximal voluntary contraction, and explosive strengthduring prolonged tennis playing. Br J Sports Med 2006;40(6):521–6.
Guezennec CY, Abdelmalki A, Serrurier B, Merino D, Bigard X,Berthelot M, et al.. Effects of prolonged exercise on brain ammoniaand amino acids. Int J Sports Med 1998;19(5):323–7.
Horita T, Komi PV, Nicol C, Kyrolainen H. Effect of exhausting stretch-shortening cycle exercise on the time course of mechanical behaviour inthe drop jump: possible role of muscle damage. Eur J Appl Physiol1999;79(2):393–403.
Hornery DJ, Farrow D, Mujika I, Young W. Fatigue in tennis.Mechanisms of fatigue and effect on performance. Sports Med2007;37(3):199–212.
International Tennis Federation Rules of Tennis 2006 and 2007. Interna-tional Tennis Federation, London [Updated at 2006 Jan 20; cited 2007Jan 30]. Available from: http://www.itftennis.com/abouttheitf/rules-regs/rules.asp.
Jones DA. High-and low-frequency fatigue revisited. Acta Physiol Scand1996;156(3):265–70.
Komi PV. Stretch-shortening cycle: a powerful model to study normal andfatigued muscle. J Biomech 2000;33(10):1197–206.
Kukulka CG, Moor MA, Russel AG. Changes in human alphamotoneurone excitability during sustained maximum isometric con-tractions. Neurosci Lett 1986;68(3):327–33.
Lanza IR, Russ DW, Kent-Braun JA. Age-related enhancement of fatigueresistance is evident in men during both isometric and dynamic tasks. JAppl Physiol 2004;97(3):967–75.
Lattier G, Millet GY, Martin A, Martin V. Fatigue and recovery afterhigh-intensity exercise Part 1: Neuromuscular fatigue. Int J SportsMed 2004;25(6):450–6.
Lepers R, Hausswirth C, Maffiuletti N, Brisswalter J, van Hoecke J.Evidence of neuromuscular fatigue after prolonged cycling exercise.Med Sci Sports Exerc 2000;32(11):1880–6.
Lepers R, Maffiuletti NA, Rochette L, Brugniaux J, Millet GY.Neuromuscular fatigue during a long-duration cycling exercise. J ApplPhysiol 2002;92(4):1487–93.
Mador MJ, Kufel TJ, Pineda LA, Sharma GK. Diaphragmatic fatigueand high-intensity exercise in patients with chronic obstructivepulmonary disease. Am J Respir Crit Care Med 2000;161(1):118–23.
Martin V, Millet GY, Lattier G, Perrod L. Why does knee extensormuscles torque decrease after eccentric-type exercise?. J Sports MedPhys Fitness 2005;45(2):143–51.
Merton PA. Voluntary strength and failure. J Physiol 1954;123(3):553–64.Metzger JM, Fitts RH. Fatigue from high- and low-frequency muscle
stimulation: contractile and biochemical alterations. J Appl Physiol1987;62(5):2075–82.
Miller M, Downham D, Lexell J. Superimposed single impulse and pulsetrain electrical stimulation: a quantitative assessement during submax-imal isometric knee extension in young healthy men. Muscle Nerve1999;22(8):1038–46.
Millet GY, Lepers R. Alterations of neuromuscular function afterprolonged running, cycling and skiing exercises. Sports Med2004;34(2):105–16.
Millet GY, Lepers R, Maffiuletti NA, Babault N, Martin V, Lattier G.Alterations of neuromuscular function after an ultramarathon. J ApplPhysiol 2002;92(2):486–92.
Millet GY, Martin V, Lattier G, Ballay Y. Mechanisms contributing toknee extensor strength loss after prolonged running exercise. J ApplPhysiol 2003a;94(1):193–8.
Millet GY, Martin V, Maffiuletti NA, Martin A. Neuromuscular fatigueafter a ski skating marathon. Can J Appl Physiol 2003b;28(3):434–45.
Place N, Lepers R, Deley G, Millet GY. Time course of neuromuscularalterations during a prolonged running exercise. Med Sci Sports Exerc2004;36(8):1347–56.
Ratel S, Lazaar N, Williams CA, Bedu M, Duche P. Age differences inhuman skeletal muscle fatigue during high-intensity intermittentexercise. Acta Paediatr 2003;92(11):1248–54.

1046 O. Girard et al. / Journal of Electromyography and Kinesiology 18 (2008) 1038–1046
Russeau E, Pinkos J. pH modulates conducting and gating behaviour ofsingle calcium release channels. Pflugers Arch 1990;415:645–7.
Strojnik V, Komi PV. Fatigue after submaximal intensive stretch-shortening cycle exercise. Med Sci Sports Exerc 2000;32(7):1314–9.
Struder HK, Hollmann W, Duperly J, Weber K. Amino acid metabolismin tennis and its possible influence on the neuroendocrine system. Br JSports Med 1995;29(1):28–30.
Thompson D, Nicholas CW, Williams C. Muscular soreness followingprolonged intermittent high-intensity shuttle running. J Sports Sci1999;17(5):387–95.
Vøllestad NK. Measurement of human muscle fatigue. J Neurosci Meth1997;74(2):219–27.
Weir JP, Keefe DA, Eaton JF, Augustine RT, Tobin DM. Effect of fatigueon hamstring coactivation during isokinetic knee extensions. Eur JAppl Physiol Occup Physiol 1998;78(6):555–9.
West W, Hicks A, McKelvie R, O’Brien J. The relationship betweenplasma potassium, muscle membrane excitability and force followingquadriceps fatigue. Pflugers Arch 1996;432(1):43–9.
Olivier Girard received his Ph.D. in HumanMovement Sciences in December 2006. Duringhis Ph.D., he has studied neuromuscular fati-gue and mechanical alterations in intermittentactivities such as racquet sports. Particularly,he has investigated mechanisms underlyingalterations in muscle function induced by pro-longed tennis playing. His research interestsalso include changes in mechanical and energycost as well as alteration in neural processinduced by different running exercises. Cur-rently he is lecturer at the faculty of sports
science at the University of Montpellier, France.
Gregory Lattier was born in 1975, he receivedhis Ph.D. degree from the University of Bur-gundy in Dijon (France) under the direction ofPr. Guillaume Millet and Dr. Alain Martin in2003. His main research interests are the effectsof neuromuscular electrical stimulation onhuman performance, and the evaluation of theneuromuscular fatigue and recovery processesfollowing various form of physical activity.
Nicola A. Maffiuletti was born in Bergamo,Italy, in 1973. He received the Ph.D. degree inSport Science in 2000 from the University ofBurgundy in Dijon, France. He has been anAssistant Professor in the same University(INSERM U887 Laboratory) since 2001, and iscurrently Director of the NeuromuscularResearch Laboratory at the Schulthess Clinicin Zurich, Switzerland. His current researchinterests include the study of neuromuscularfunction in orthopaedic patients and thephysiology of neuromuscular electrical stimu-
lation. He is a member of the American College of Sports Medicine andthe European College of Sport Science.
Jean-Paul Micallef, engineer, has a Ph.D. inbiomechanics and is professor at the universityof Montpellier. His main research interestsinclude sport biomechanics, rehabilitation,orthopedics and bioengineering. He is alsointerested in the development of innovatingsensors (transducers, strain gauges).
Gregoire Millet is senior physiologist in Aspire,
the Academy for Sport Excellence of Qatar. Hehas been an elite coach in triathlon, moun-tainbike and distance running in several coun-tries. To date, he has published two books and 60 articles in peer-review journals, mainly onthe physiological responses to aerobic, inter-mittent or hypoxic training.
Related Documents









![[Product Monograph Template - Standard] · conventional hemodialysis, sustained -efficiency dialysis (SLED), extended daily dialysis low (EDD), or prolonged intermittent renal replacement](https://static.cupdf.com/doc/110x72/5f08331e7e708231d420d4e3/product-monograph-template-standard-conventional-hemodialysis-sustained-efficiency.jpg)

