Chemico-Biological Interactions 184 (2010) 474–483 Contents lists available at ScienceDirect Chemico-Biological Interactions journal homepage: www.elsevier.com/locate/chembioint Neurogenic-committed human pre-adipocytes express CYP1A isoforms Chiara Scanarotti a,∗ , Anna Maria Bassi a , Mariafrancesca Catalano a , Chiara Guida b , Rosella Coradeghini c , Carla Falugi d , Mariagrazia Aluigi d , Pierluigi Santi e , Edoardo Raposio e a Cell Culture Lab, DIMES, University of Genoa, Via L.B. Alberti 2, 16132 Genoa, Italy b Tissue Engineering Lab, Plastic Surgery Division, DICMI, University of Genoa, Via De Toni 14, 16132 Genoa, Italy c Nanobiosciences Unit, JRC - European Commission Institute for Health and Consumer Protection, Via E. Fermi 1, 21020 Ispra (VA), Italy d Biology Department, DIBIO, University of Genoa, Viale Benedetto XV 5, 16132 Genoa, Italy e National Cancer Institute, IST, University of Genoa, L.go R. Benzi 10, 16132 Genoa, Italy article info Article history: Received 23 July 2009 Received in revised form 23 December 2009 Accepted 5 January 2010 Available online 18 January 2010 Keywords: Neurogenic pre-adipocytes Cytochrome P450 1A1 Cholinergic system ECOD EROD -Naphthoflavone abstract Stem cell models offer an opportunity both for therapeutic use and for the assessment of alternative in vitro models. Human lipoaspirate is a source of adult stem cells (pre-adipocytes), which are able to differentiate into various phenotypes, such as neurogenic lineage. Here, we analyse the suitability of these in vitro models in screening exogenous compounds, such as environmental pollutants, that may affect adipose cells and neurogenic development. To evaluate neurogenic differentiation, we analysed expression of cholinergic system and acetylcholinesterase immunoreactivity. Heterocyclic derivatives of polycyclic aromatic hydrocarbons (PAHs) are often significant components of environmental con- taminants. As they contain inducers of cytochrome P450 1A1 (CYP1A1), we explored the activity of CYP1A1-related enzymes, i.e. 7-ethoxycoumarin- and 7-ethoxyresorufin-O-deethylase (ECOD and EROD) in both cell systems in basal conditions and after exposure to non-cytotoxic doses of -naphthoflavone (BNF), a well-known PAH-type inducer. Both cell models showed basal and inducible levels of ECOD. Analysis of CYP1A1 protein expression and EROD-related enzyme activity confirmed the inducibility of the CYP1A1 isoform by BNF. These results demonstrate that mesenchymal adult stem cells can constitute innovative models. We therefore propose the use of pre-adipocytes and their neurogenic derivates to evaluate the cytotoxic/biological effects of unintended exposure to contaminants. © 2010 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Fat is an active and dynamic tissue that can easily be har- vested in large quantities from several regions of the human body. It is composed of various cell types, including adipocytes, fibroblasts, smooth-muscle cells, endothelial cells and adipogenic progenitor cells called pre-adipocytes (PA) [1,2]. PA possess a char- acteristic pathway of multipotent adult stem cells and are able to differentiate in vitro into adipo-, osteo-, chondro-, myo- and neurogenic lineages under specific culture conditions [3–7]. The primary source of pre-adipocytes is human liposuctioned fat aspi- Abbreviations: BNF, beta-naphthoflavone; ECOD, 7-ethoxycoumarin-O- deethylase; EROD, 7-ethoxyresorufin-O-deethylase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PAH, polycyclic aromatic hydrocarbon; CYP1A1, cytochrome P450 1A1 isoform. ∗ Corresponding author. Tel.: +39 010 3538823 fax: +39 010 3538836. E-mail addresses: chiara [email protected] (C. Scanarotti), [email protected] (A.M. Bassi), [email protected] (M. Catalano), [email protected] (C. Guida), r [email protected] (R. Coradeghini), [email protected] (C. Falugi), [email protected] (M. Aluigi), [email protected] (P. Santi), [email protected] (E. Raposio). rates and, in particular, processed lipoaspirate cells obtained from the washing and digestion of lipoaspirate extracellular matrix [4,6]. Pre-adipocytes express many CD marker antigens that are similar to those observed on human mesenchymal stem cells, such as CD105, CD90, CD71 and CD44 [8–10]. Moreover, they are capable of secret- ing angiogenesis-related cytokines, including vascular endothelial growth factor, granulocyte/macrophage colony stimulating factor, stromal-derived factor-1alpha and hepatocyte growth factor [11]. When pre-adipocytes are cultivated in very low-serum and serum- free conditions, they maintain their ability to differentiate into adipo-, chondro- and osteogenic lineages in vitro, and the expres- sion of HLA, CD133, CD184 and CD31 is comparable to that seen in cells cultured in 10% serum [12]. Consequently autologous pre- adipocytes that are cultured in vitro may be used in human clinical applications when they are cultivated in an animal-product-free environment; moreover, they have the potential to be used in tissue repair and regeneration [13]. Many recent literature data suggest that neural disorders may begin early in life, and that air pollutants, drugs and other xeno- biotics play a crucial role in this process [14,15]. Therefore we propose the use of human multipotent adult stem cells derived from fat tissue and their neurogenic derivates as an alternative 0009-2797/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.cbi.2010.01.009

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemico-Biological Interactions 184 (2010) 474–483
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journa l homepage: www.e lsev ier .com/ locate /chembio int
Neurogenic-committed human pre-adipocytes express CYP1A isoforms
Chiara Scanarotti a,∗, Anna Maria Bassi a, Mariafrancesca Catalanoa, Chiara Guidab, Rosella Coradeghini c,Carla Falugid, Mariagrazia Aluigid, Pierluigi Santi e, Edoardo Raposioe
a Cell Culture Lab, DIMES, University of Genoa, Via L.B. Alberti 2, 16132 Genoa, Italyb Tissue Engineering Lab, Plastic Surgery Division, DICMI, University of Genoa, Via De Toni 14, 16132 Genoa, Italyc Nanobiosciences Unit, JRC - European Commission Institute for Health and Consumer Protection, Via E. Fermi 1, 21020 Ispra (VA), Italyd Biology Department, DIBIO, University of Genoa, Viale Benedetto XV 5, 16132 Genoa, Italye National Cancer Institute, IST, University of Genoa, L.go R. Benzi 10, 16132 Genoa, Italy
a r t i c l e i n f o
Article history:Received 23 July 2009Received in revised form23 December 2009Accepted 5 January 2010Available online 18 January 2010
Keywords:Neurogenic pre-adipocytesCytochrome P450 1A1Cholinergic systemECODEROD
a b s t r a c t
Stem cell models offer an opportunity both for therapeutic use and for the assessment of alternativein vitro models. Human lipoaspirate is a source of adult stem cells (pre-adipocytes), which are able todifferentiate into various phenotypes, such as neurogenic lineage. Here, we analyse the suitability ofthese in vitro models in screening exogenous compounds, such as environmental pollutants, that mayaffect adipose cells and neurogenic development. To evaluate neurogenic differentiation, we analysedexpression of cholinergic system and acetylcholinesterase immunoreactivity. Heterocyclic derivativesof polycyclic aromatic hydrocarbons (PAHs) are often significant components of environmental con-taminants. As they contain inducers of cytochrome P450 1A1 (CYP1A1), we explored the activity ofCYP1A1-related enzymes, i.e. 7-ethoxycoumarin- and 7-ethoxyresorufin-O-deethylase (ECOD and EROD)in both cell systems in basal conditions and after exposure to non-cytotoxic doses of �-naphthoflavone(BNF), a well-known PAH-type inducer. Both cell models showed basal and inducible levels of ECOD.Analysis of CYP1A1 protein expression and EROD-related enzyme activity confirmed the inducibility of
�-Naphthoflavone the CYP1A1 isoform by BNF. These results demonstrate that mesenchymal adult stem cells can constituteinnovative models. We therefore propose the use of pre-adipocytes and their neurogenic derivates to
ologic
1
vbfipatnp
d3c
acf(
0d
evaluate the cytotoxic/bi
. Introduction
Fat is an active and dynamic tissue that can easily be har-ested in large quantities from several regions of the humanody. It is composed of various cell types, including adipocytes,broblasts, smooth-muscle cells, endothelial cells and adipogenicrogenitor cells called pre-adipocytes (PA) [1,2]. PA possess a char-
cteristic pathway of multipotent adult stem cells and are ableo differentiate in vitro into adipo-, osteo-, chondro-, myo- andeurogenic lineages under specific culture conditions [3–7]. Therimary source of pre-adipocytes is human liposuctioned fat aspi-Abbreviations: BNF, beta-naphthoflavone; ECOD, 7-ethoxycoumarin-O-eethylase; EROD, 7-ethoxyresorufin-O-deethylase; GAPDH, glyceraldehyde-phosphate dehydrogenase; PAH, polycyclic aromatic hydrocarbon; CYP1A1,ytochrome P450 1A1 isoform.∗ Corresponding author. Tel.: +39 010 3538823 fax: +39 010 3538836.
E-mail addresses: chiara [email protected] (C. Scanarotti),[email protected] (A.M. Bassi), [email protected] (M. Catalano),[email protected] (C. Guida), r [email protected] (R. Coradeghini),[email protected] (C. Falugi), [email protected] (M. Aluigi), [email protected]. Santi), [email protected] (E. Raposio).
009-2797/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.cbi.2010.01.009
al effects of unintended exposure to contaminants.© 2010 Elsevier Ireland Ltd. All rights reserved.
rates and, in particular, processed lipoaspirate cells obtained fromthe washing and digestion of lipoaspirate extracellular matrix [4,6].Pre-adipocytes express many CD marker antigens that are similar tothose observed on human mesenchymal stem cells, such as CD105,CD90, CD71 and CD44 [8–10]. Moreover, they are capable of secret-ing angiogenesis-related cytokines, including vascular endothelialgrowth factor, granulocyte/macrophage colony stimulating factor,stromal-derived factor-1alpha and hepatocyte growth factor [11].When pre-adipocytes are cultivated in very low-serum and serum-free conditions, they maintain their ability to differentiate intoadipo-, chondro- and osteogenic lineages in vitro, and the expres-sion of HLA, CD133, CD184 and CD31 is comparable to that seenin cells cultured in 10% serum [12]. Consequently autologous pre-adipocytes that are cultured in vitro may be used in human clinicalapplications when they are cultivated in an animal-product-freeenvironment; moreover, they have the potential to be used in tissuerepair and regeneration [13].
Many recent literature data suggest that neural disorders maybegin early in life, and that air pollutants, drugs and other xeno-biotics play a crucial role in this process [14,15]. Therefore wepropose the use of human multipotent adult stem cells derivedfrom fat tissue and their neurogenic derivates as an alternative

gical
ite
cmddsiinnnswts
atmteTittaaif
rAt
mh
utotogid
2
2
sIcriAiCfcc
C. Scanarotti et al. / Chemico-Biolo
n vitro procedure that may be converted into safe toxicologicalests for human beings. To this end, specific endpoints must bestablished in order to provide predictive biomarkers of risk.
The aim of the present study was therefore to assess bioethicallyompatible predictive testing to evaluate the effects of environ-ental pollutants (i.e. polycyclic hydrocarbons, PAH), foodstuffs,
rugs, etc. on in vitro cultures of pre-adipocytes and neurogenicerivates. As described in our previous papers [4,16] and in othertudies [3,17], lipoaspirate cells have the ability to differentiaten vitro towards neurogenic and myogenic lineages. In particular,n neurogenic derivate cells, we demonstrated the expression ofeuron-specific enolase, an early neuronal progenitor marker, butot of glial fibrillary acidic protein, Tau and MAP2, which are matureeuron and glial markers. To ensure that our in vitro model couldimulate the environment of the neural system reasonably well,e had to verify the expression of other markers of neural func-
ion, such as molecules belonging to neurotransmitter (cholinergic)ystems.
Moreover, since the lack of a detoxification system placessevere restriction on in vitro testing, it is also very impor-
ant to demonstrate the expression and modulation of xenobioticetabolizing enzymes in this model. As is well known, exposure
o hydrophobic xenobiotics results in the induction of phase Inzymes, such as the cytochrome P450 (CYP) enzymatic system.he CYP1A isoform specifically metabolizes PAHs to carcinogenicntermediates. CYP1A induction is used as a biomarker of exposureo toxic substances [18,19]. The vulnerability of the nervous sys-em to neurotoxins, and in particular to PAHs, has to be taken intoccount. As PAHs are lipophilic, they may accumulate not only indipose tissue, but also at several other sites, such as in the brain;ndeed, recent evidence has indicated that they can alter neuralunction in fishes [20,21].
Moreover, PAHs are also known to bind the aryl hydrocarboneceptor (AhR), and several recent studies have reported a role ofhR in regulating the cell cycle, cell proliferation and apoptosis,
hus suggesting a promoter role of AhR in carcinogenesis [22,23].Taken together, the available data seem to suggest that PAHs
ay be implicated in affecting neural development and function inumans, too.
In order to ascertain whether our in vitro models could beseful in investigating the bioactivation potential of environmen-al contaminants, we analysed the expression and modulationf CYP1A in pre-adipocytes and their neurogenic derivate cul-ures by exposure to �-naphthoflavone (BNF), a classic inducerf several AhR battery genes, enclosed Cyp1A1, 1A2, and 1B1enes. We also evaluated specific CYP4501A-related activities,.e. 7-ethoxycoumarin-O-deethylase (ECOD), 7-ethoxyresorufin-O-eethylase (EROD), and CYP1A1 protein expression.
. Materials and methods
.1. Chemicals
All cell culture products, including standard medium and humanerum, were purchased from Euroclone (Life Sciences Division,taly). Supplemented compounds, including collagenase, were pur-hased from Sigma Chemical Co. (St. Louis, MO, USA). Bio-Radeagent protein assay kit, reagents and apparatus for westernmmunoblot were obtained from Bio-Rad (Hercules, CA, USA).nti-CYP1A1, anti-GAPDH and anti-acetylcholinesterase antibod-
es were obtained from Santa Cruz Biotechnology Inc. (Santa Cruz,A, USA); anti-choline acetyltransferase antibody was obtained
rom Chemicon (Millipore Corporate Billerica, MA, USA). All otherhemicals and solvents were of analytical grade and were fromommon commercial sources.
Interactions 184 (2010) 474–483 475
2.2. Lipoaspirate cell processing
Human adipose tissues were obtained by means of lipo-suction procedures from six healthy female donors who werenon-smokers. Our Ethics Committee ensured that the procedurecomplied with national and European bioethical laws regard-ing the criteria of anonymity, informed consent, safety etc. Theliposuctioned fat aspirates were processed to obtain the lipoaspi-rate cell population, as described by Zuk et al. [7] with minormodifications [4,16]. Briefly, after washing in phosphate bufferedsaline (PBS), the lipoaspirates underwent extracellular matrixdigestion with collagenase (2 mg/ml PBS) and were centrifugedat 1200 × g for 10 min. To remove contaminating erythrocytes,the pellet was incubated with 160 mM ammonium chloride for10 min at room temperature. The pre-adipocytes thus obtainedwere subsequently resuspended in control medium, Dulbecco’sModified Eagle’s Medium (DMEM), 10% human serum, 2 mMglutamine, penicillin (100 UI/ml)–streptomycin (50 �g/ml) andamphotericin B (2.5 �g/ml), and set up for neurogenic differenti-ation.
2.3. Neurogenic differentiation procedure
Pre-adipocytes were subjected to a neurogenic differentiationprocedure, as previously described [3,4,16]. Briefly, cellular sam-ples were grown in neurogenic medium: DMEM with 10% humanserum, 2 mM glutamine, penicillin (100 UI/ml)–streptomycin(50 �g/ml), amphotericin B (2.5 �g/ml), 5 �g/ml insulin, 200 �Mindomethacin, and 0.5 mM isobutylmethylxanthine. In parallel,control cultures were maintained in standard culture conditions.Culture medium was replaced with fresh medium twice a week.To assess the stage of neurogenic development, both controlcells and induced cells were observed daily under the micro-scope (Nikon, Eclipse TS100), before specific neural markers wereanalysed. All differentiation procedures were performed for 2weeks.
2.4. FACS analysis
Pre-adipocytes in the initial stage of culture were har-vested at about 30% confluence and incubated with fluores-cein isothiocyanate (FITC)-, phycoerythein (PE)-, and peridinin–chlorophyll–protein complex (PerCP)-conjugated anti-CD 14, anti-CD44, anti-CD90, anti-CD105 and anti-CD45 (BD Pharmingen, SanJose, CA). Anti-IgG isotype control antibodies were used. Labelledcells were resuspended in FACS fix solution (PBS containing 0.1%(v/v) formalin and 2% (w/v) glucose) and stored at 4 ◦C until analy-sis. Data on 10,000 cell events were acquired and analysed by meansof a FACS Calibur instrument (Becton–Dickinson, San Jose, CA, USA)and CellQuest and Paint-a-gate software.
2.5. Immunocytochemistry analysis of molecules related toneurotransmitter systems
After neural induction, pre-adipocyte and neurogenic derivatecultures were grown on poly-d-lysine-coated slides (Bio-Optica,Milan, Italy) for 24 h (1 × 106 cells/slide). Differentiation towards aneurogenic lineage was checked by monitoring cholinergic expres-sion by means of the indirect immunofluorescence method. To thisend, we utilised molecules related to this neurotransmitter systemthat are commonly used to reveal excitatory neural commitment
[24,25]. The molecules related to the cholinergic system were:AChE = acetylcholinesterase, EC: 3.1.1.7, the lytic enzyme of acetyl-choline (ACh); and ChAT = choline acetyltransferase, EC: 2.3.1.6,the biosynthetic enzyme of ACh. Slides were washed twice in PBSand fixed in 4% paraformaldehyde in PBS for 30 min. They were
4 ogical
t2aA1wsflinCflLbgwWTiewstSis
2
matama
2
raa(e
2
(Acift
2
rtapan
76 C. Scanarotti et al. / Chemico-Biol
hen washed in 1% Triton X-100 in PBS for 10 min, 0.02% Tween-0 in PBS for 5 min and 0.02% Tween-20/1% BSA (bovine serumlbumin) in PBS for 5 min. Primary monoclonal anti-mammalianChE and anti-ChAT antibodies were incubated at a dilution of:100 in 3% BSA in PBS at 37 ◦C for 45 min. Slides were thenashed in 0.02% Tween-20/1% BSA in PBS for 5 min. Anti-mouse
econdary antibody (obtained from Cappel, Italy) conjugated withuoresceine isothiocyanate (FITC) was diluted 1:400 in 3% BSA
n PBS and incubated at 37 ◦C for 45 min. After thorough rinsing,uclei were stained by adding 1 �g/ml of propidium iodide (Bio-hemika, Milan, Italy). The cells were then observed under theuorescence microscope (Leica DMRB epifluorescence microscope,eica Microsystems AG Wetzlar, Bensheim, Germany). After incu-ation, the slides were rinsed thoroughly in PBS, mounted withelvatol [26] and observed under a Leitz microscope equippedith fluorescence apparatus for FITC epifluorescence (� = 470 nm).e also analysed the histochemical localisation of AChE activity.
he histochemical AChE reaction, which was aimed at detect-ng AChE enzyme activity, was carried out both in vitro and inlectrophoresis gels, as described earlier [27]. Specificity controlsere performed by pre-exposing the cells to 10−4 M BW284c51, a
pecific inhibitor of nerve AChE, by pre-incubation in medium con-aining 10−4 M tetra-isomethylpyrophosphoramide (iso-OMPA,igma), in order to inhibit pseudocholinesterase activities andsolate the “nerve” AChE activity, and by omitting the sub-trate.
.6. CYP450 inducer treatments
A stock solution of BNF in DMSO was added to serum-freeedium (final solvent concentration <0.2%). Twenty-four hours
fter seeding, cultures were exposed for the last 24, 48 and 72 ho BNF, in the range of concentration from 0.1 to 10 �M. Untreatednd solvent-control cultures were prepared by adding serum-freeedium, alone or supplemented with the same amount of DMSO
s the experimental medium.
.7. Cell viability assessment
Cell viability was assessed by means of two cytotoxicity tests:eduction of tetrazolium salt by mitochondrial enzymes (MTT)nd neutral red uptake (NRU). For these tests, pre-adipocytesnd their neurogenic derivates were seeded on 96-well plates5 × 103 cells/well) 24 h before treatment and the cultures werexposed to each treatment for 24, 48 and 72 h.
.8. MTT assay
MTT was dissolved in PBS (5 mg/ml) and added to the cells10 �l/ml medium) according to the method of Mosmann [28].fter incubation for 3 h at 37 ◦C, a solution of 1N hydrogenhloride–isopropanol (1:25, v/v) was added to each well and mixedn order to dissolve the dark-blue formazan crystals that hadormed. After being kept for a few minutes at room temperature,he plates were read at 570 nm.
.9. NRU assay
After BNF treatment, cell monolayers were exposed to a neutral
ed dye-containing medium (50 �g/ml) and processed accordingo the method of Borenfreund et al. [29]. After incubation for 3 ht 37 ◦C, cells were washed twice in PBS and fixed according to therocedure described by Riddell et al. [30]. The plates were then leftt room temperature for 10 min and the absorbance of the releasedeutral red dye was read at 550 nm.Interactions 184 (2010) 474–483
2.10. Drug metabolism enzyme activities
Twenty-four hours before treatment, pre-adipocyte andneurogenic derivate cultures were seeded on 24-well plates(8 × 104 cell/well) and exposed to BNF for 24 and 48 h. The CYP1A-, 2A-, 2B-, 2C-, 2E- and 3A-dependent 7-ethoxycoumarin-O-deethylase (ECOD) and CYP1A1-dependent 7-ethoxyresorufin-O-deethylase (EROD) activities were determined spectrofluo-rimetrically in living cells, after exposure to their specificsubstrates (7-ethoxycoumarin and 7-ethoxyresorufin, respec-tively). Intact cells were incubated in 24-well plates with150 �M 7-ethoxycoumarin and 8 �M 7-ethoxyresorufin plus10 �M dicumarol [31,32]. Resorufin fluorescence was measuredby using a Perkin-Elmer LS-5 (Beaconsfield, UK) spectrofluorom-eter with excitation and emission wavelengths of 365 and 456 nmfor ECOD and 530 and 585 nm for EROD. The pmoles of 7-hydroxycoumarin and resorufin produced per mg of cell protein perminute were evaluated by comparison with a standard solution.
2.11. Western immunoblotting
Cellular lysates were analysed as described elsewhere [33];30 �g proteins/lane were resolved on SDS-PAGE, transferred toa Hybond ECL membrane and processed by using an ECL + Pluskit Amersham (GE Healthcare Europe GmbH, Germany). TheCYP1A1 protein was detected by means of its specific antibodyand subsequently by means of secondary antibody conjugates tohorseradish peroxidase. Finally, the bands were detected by meansof ECL reagents according to the manufacturer’s protocol. Westernimmunoblots were stripped and re-probed for the glyceraldehyde3-phosphate dehydrogenase (GAPDH) protein by means of a poly-clonal antibody, in order to confirm loading equivalency. The bandswere quantified by Quantity One software in a Gel-Doc system(BioRad), and the densitometry data obtained were subjected tostatistical analysis. The density of the control was set at 100 andthe treated samples were measured as percentages of the controlvalue.
2.12. Protein determination
Total cellular protein content was estimated on cell lysateobtained after exposure of the monolayer to 0.1 M NaOH at 37 ◦Cfor 1 h. The samples were then processed according to Bradford [34]by means of a Bio-Rad assay kit, with bovine serum albumin beingused as standard.
2.13. Statistical analysis
All experiments were performed in triplicate on the lipoaspi-rates from each donor. Results are presented as the arithmeticmean ± DS. The data regarding cytotoxicity and immunoblottingwere subjected to one-way analysis of ANOVA variance for multi-ple comparisons, followed by Dunnett’s test, by means of the Instatand Prism software package (GraphPad Software, Inc). The data ondrug metabolism enzyme activities were statistically analysed bymeans of STATA/SE9TM software (StataCorp LP, College Station, TX,USA). In this case, the statistical analysis was carried out by usinga t-test to evaluate the difference between the two cell groups. Ap-value of less than 5% was considered significant.
3. Results
3.1. Immunochemical analysis
In vitro, pre-adipocytes possess a fibroblast-like morphology;during the first steps of neurogenic differentiation, cell morphology
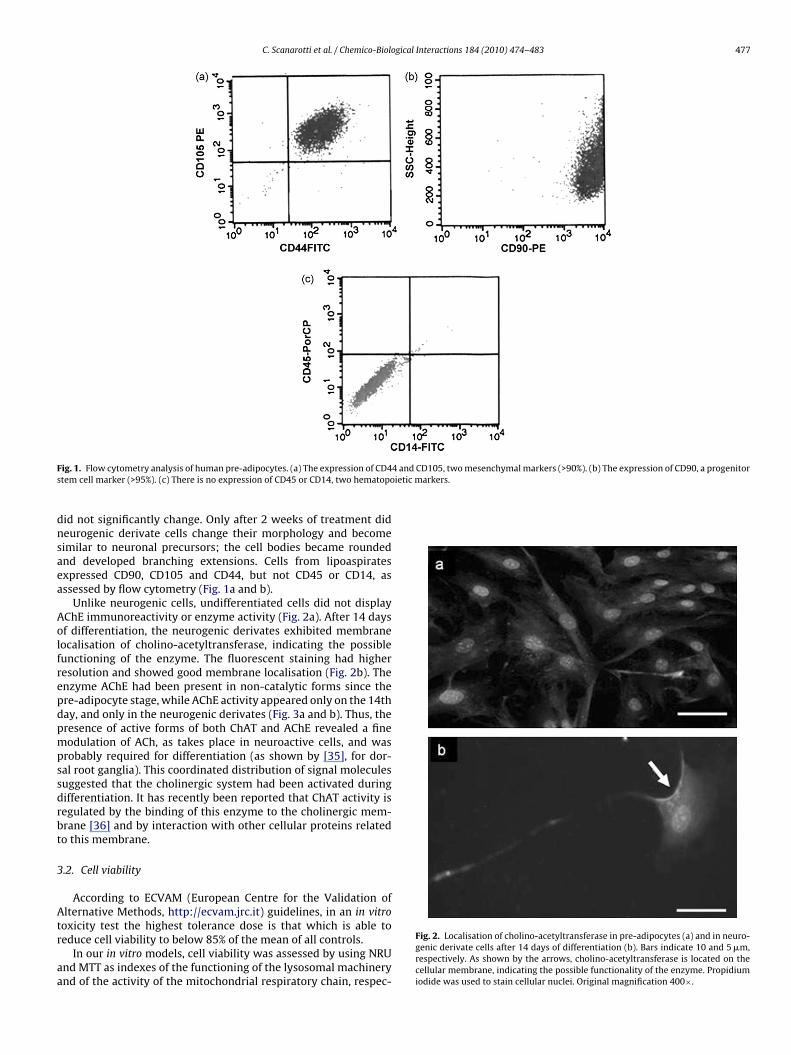
C. Scanarotti et al. / Chemico-Biological Interactions 184 (2010) 474–483 477
F 4 and CD105, two mesenchymal markers (>90%). (b) The expression of CD90, a progenitors ietic markers.
dnsaea
Aolfrepdpmpssdrbt
3
Atr
aa
ig. 1. Flow cytometry analysis of human pre-adipocytes. (a) The expression of CD4tem cell marker (>95%). (c) There is no expression of CD45 or CD14, two hematopo
id not significantly change. Only after 2 weeks of treatment dideurogenic derivate cells change their morphology and becomeimilar to neuronal precursors; the cell bodies became roundednd developed branching extensions. Cells from lipoaspiratesxpressed CD90, CD105 and CD44, but not CD45 or CD14, asssessed by flow cytometry (Fig. 1a and b).
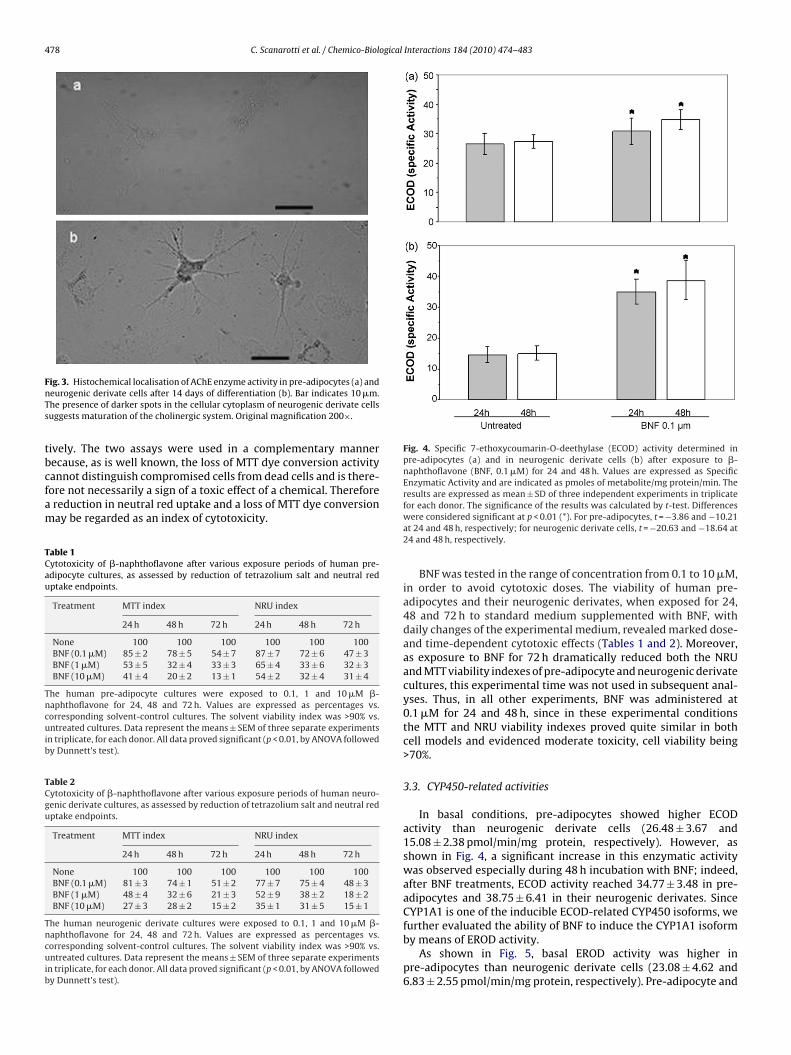
Unlike neurogenic cells, undifferentiated cells did not displayChE immunoreactivity or enzyme activity (Fig. 2a). After 14 daysf differentiation, the neurogenic derivates exhibited membraneocalisation of cholino-acetyltransferase, indicating the possibleunctioning of the enzyme. The fluorescent staining had higheresolution and showed good membrane localisation (Fig. 2b). Thenzyme AChE had been present in non-catalytic forms since there-adipocyte stage, while AChE activity appeared only on the 14thay, and only in the neurogenic derivates (Fig. 3a and b). Thus, theresence of active forms of both ChAT and AChE revealed a fineodulation of ACh, as takes place in neuroactive cells, and was
robably required for differentiation (as shown by [35], for dor-al root ganglia). This coordinated distribution of signal moleculesuggested that the cholinergic system had been activated duringifferentiation. It has recently been reported that ChAT activity isegulated by the binding of this enzyme to the cholinergic mem-rane [36] and by interaction with other cellular proteins relatedo this membrane.
.2. Cell viability
According to ECVAM (European Centre for the Validation oflternative Methods, http://ecvam.jrc.it) guidelines, in an in vitro
oxicity test the highest tolerance dose is that which is able toeduce cell viability to below 85% of the mean of all controls.
In our in vitro models, cell viability was assessed by using NRUnd MTT as indexes of the functioning of the lysosomal machinerynd of the activity of the mitochondrial respiratory chain, respec-
Fig. 2. Localisation of cholino-acetyltransferase in pre-adipocytes (a) and in neuro-genic derivate cells after 14 days of differentiation (b). Bars indicate 10 and 5 �m,respectively. As shown by the arrows, cholino-acetyltransferase is located on thecellular membrane, indicating the possible functionality of the enzyme. Propidiumiodide was used to stain cellular nuclei. Original magnification 400×.
478 C. Scanarotti et al. / Chemico-Biological Interactions 184 (2010) 474–483
FnTs
tbcfam
TCau
Tncuib
TCgu
Tncuib
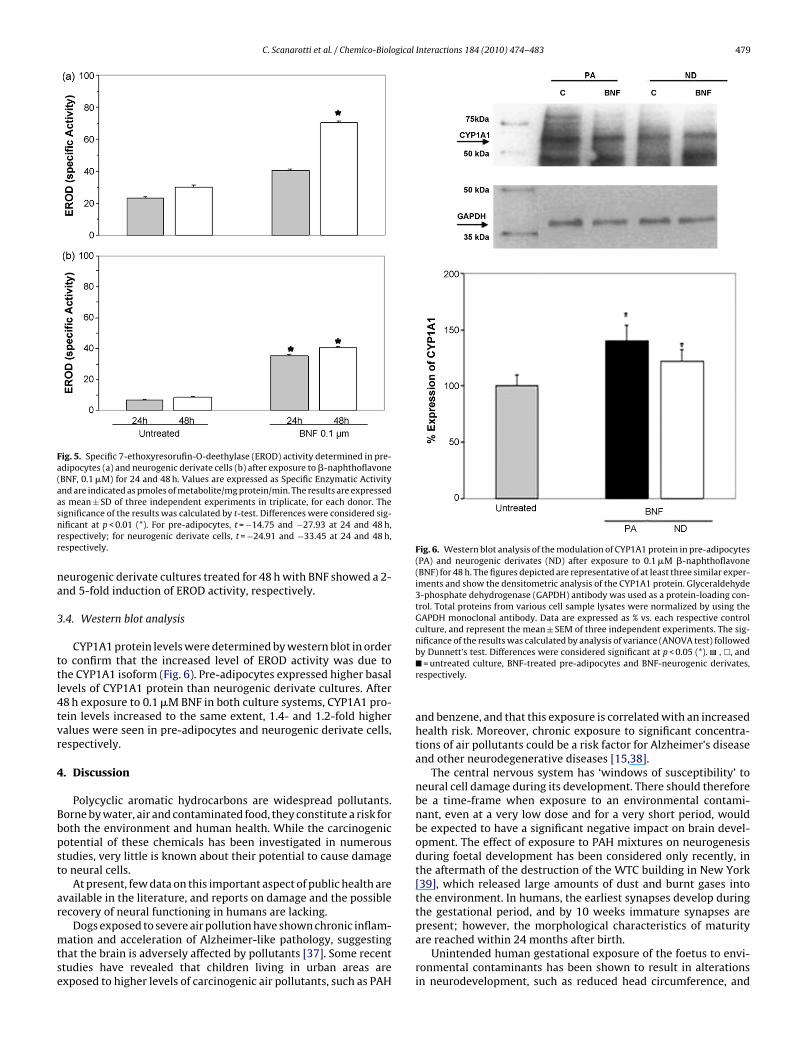
Fig. 4. Specific 7-ethoxycoumarin-O-deethylase (ECOD) activity determined inpre-adipocytes (a) and in neurogenic derivate cells (b) after exposure to �-
ig. 3. Histochemical localisation of AChE enzyme activity in pre-adipocytes (a) andeurogenic derivate cells after 14 days of differentiation (b). Bar indicates 10 �m.he presence of darker spots in the cellular cytoplasm of neurogenic derivate cellsuggests maturation of the cholinergic system. Original magnification 200×.
ively. The two assays were used in a complementary mannerecause, as is well known, the loss of MTT dye conversion activity
annot distinguish compromised cells from dead cells and is there-ore not necessarily a sign of a toxic effect of a chemical. Thereforereduction in neutral red uptake and a loss of MTT dye conversionay be regarded as an index of cytotoxicity.able 1ytotoxicity of �-naphthoflavone after various exposure periods of human pre-dipocyte cultures, as assessed by reduction of tetrazolium salt and neutral redptake endpoints.
Treatment MTT index NRU index
24 h 48 h 72 h 24 h 48 h 72 h
None 100 100 100 100 100 100BNF (0.1 �M) 85 ± 2 78 ± 5 54 ± 7 87 ± 7 72 ± 6 47 ± 3BNF (1 �M) 53 ± 5 32 ± 4 33 ± 3 65 ± 4 33 ± 6 32 ± 3BNF (10 �M) 41 ± 4 20 ± 2 13 ± 1 54 ± 2 32 ± 4 31 ± 4
he human pre-adipocyte cultures were exposed to 0.1, 1 and 10 �M �-aphthoflavone for 24, 48 and 72 h. Values are expressed as percentages vs.orresponding solvent-control cultures. The solvent viability index was >90% vs.ntreated cultures. Data represent the means ± SEM of three separate experiments
n triplicate, for each donor. All data proved significant (p < 0.01, by ANOVA followedy Dunnett’s test).
able 2ytotoxicity of �-naphthoflavone after various exposure periods of human neuro-enic derivate cultures, as assessed by reduction of tetrazolium salt and neutral redptake endpoints.
Treatment MTT index NRU index
24 h 48 h 72 h 24 h 48 h 72 h
None 100 100 100 100 100 100BNF (0.1 �M) 81 ± 3 74 ± 1 51 ± 2 77 ± 7 75 ± 4 48 ± 3BNF (1 �M) 48 ± 4 32 ± 6 21 ± 3 52 ± 9 38 ± 2 18 ± 2BNF (10 �M) 27 ± 3 28 ± 2 15 ± 2 35 ± 1 31 ± 5 15 ± 1
he human neurogenic derivate cultures were exposed to 0.1, 1 and 10 �M �-aphthoflavone for 24, 48 and 72 h. Values are expressed as percentages vs.orresponding solvent-control cultures. The solvent viability index was >90% vs.ntreated cultures. Data represent the means ± SEM of three separate experiments
n triplicate, for each donor. All data proved significant (p < 0.01, by ANOVA followedy Dunnett’s test).
naphthoflavone (BNF, 0.1 �M) for 24 and 48 h. Values are expressed as SpecificEnzymatic Activity and are indicated as pmoles of metabolite/mg protein/min. Theresults are expressed as mean ± SD of three independent experiments in triplicate
for each donor. The significance of the results was calculated by t-test. Differenceswere considered significant at p < 0.01 (*). For pre-adipocytes, t = −3.86 and −10.21at 24 and 48 h, respectively; for neurogenic derivate cells, t = −20.63 and −18.64 at24 and 48 h, respectively.BNF was tested in the range of concentration from 0.1 to 10 �M,in order to avoid cytotoxic doses. The viability of human pre-adipocytes and their neurogenic derivates, when exposed for 24,48 and 72 h to standard medium supplemented with BNF, withdaily changes of the experimental medium, revealed marked dose-and time-dependent cytotoxic effects (Tables 1 and 2). Moreover,as exposure to BNF for 72 h dramatically reduced both the NRUand MTT viability indexes of pre-adipocyte and neurogenic derivatecultures, this experimental time was not used in subsequent anal-yses. Thus, in all other experiments, BNF was administered at0.1 �M for 24 and 48 h, since in these experimental conditionsthe MTT and NRU viability indexes proved quite similar in bothcell models and evidenced moderate toxicity, cell viability being>70%.
3.3. CYP450-related activities
In basal conditions, pre-adipocytes showed higher ECODactivity than neurogenic derivate cells (26.48 ± 3.67 and15.08 ± 2.38 pmol/min/mg protein, respectively). However, asshown in Fig. 4, a significant increase in this enzymatic activitywas observed especially during 48 h incubation with BNF; indeed,after BNF treatments, ECOD activity reached 34.77 ± 3.48 in pre-adipocytes and 38.75 ± 6.41 in their neurogenic derivates. SinceCYP1A1 is one of the inducible ECOD-related CYP450 isoforms, we
further evaluated the ability of BNF to induce the CYP1A1 isoformby means of EROD activity.As shown in Fig. 5, basal EROD activity was higher inpre-adipocytes than neurogenic derivate cells (23.08 ± 4.62 and6.83 ± 2.55 pmol/min/mg protein, respectively). Pre-adipocyte and
C. Scanarotti et al. / Chemico-Biological Interactions 184 (2010) 474–483 479
Fig. 5. Specific 7-ethoxyresorufin-O-deethylase (EROD) activity determined in pre-adipocytes (a) and neurogenic derivate cells (b) after exposure to �-naphthoflavone(BNF, 0.1 �M) for 24 and 48 h. Values are expressed as Specific Enzymatic Activityand are indicated as pmoles of metabolite/mg protein/min. The results are expressedas mean ± SD of three independent experiments in triplicate, for each donor. Thesnrr
na
3
ttl4tvr
4
Bbpst
ar
mtse
Fig. 6. Western blot analysis of the modulation of CYP1A1 protein in pre-adipocytes(PA) and neurogenic derivates (ND) after exposure to 0.1 �M �-naphthoflavone(BNF) for 48 h. The figures depicted are representative of at least three similar exper-iments and show the densitometric analysis of the CYP1A1 protein. Glyceraldehyde3-phosphate dehydrogenase (GAPDH) antibody was used as a protein-loading con-trol. Total proteins from various cell sample lysates were normalized by using theGAPDH monoclonal antibody. Data are expressed as % vs. each respective controlculture, and represent the mean ± SEM of three independent experiments. The sig-nificance of the results was calculated by analysis of variance (ANOVA test) followed
ignificance of the results was calculated by t-test. Differences were considered sig-ificant at p < 0.01 (*). For pre-adipocytes, t = −14.75 and −27.93 at 24 and 48 h,espectively; for neurogenic derivate cells, t = −24.91 and −33.45 at 24 and 48 h,espectively.
eurogenic derivate cultures treated for 48 h with BNF showed a 2-nd 5-fold induction of EROD activity, respectively.
.4. Western blot analysis
CYP1A1 protein levels were determined by western blot in ordero confirm that the increased level of EROD activity was due tohe CYP1A1 isoform (Fig. 6). Pre-adipocytes expressed higher basalevels of CYP1A1 protein than neurogenic derivate cultures. After8 h exposure to 0.1 �M BNF in both culture systems, CYP1A1 pro-ein levels increased to the same extent, 1.4- and 1.2-fold higheralues were seen in pre-adipocytes and neurogenic derivate cells,espectively.
. Discussion
Polycyclic aromatic hydrocarbons are widespread pollutants.orne by water, air and contaminated food, they constitute a risk foroth the environment and human health. While the carcinogenicotential of these chemicals has been investigated in numeroustudies, very little is known about their potential to cause damageo neural cells.
At present, few data on this important aspect of public health arevailable in the literature, and reports on damage and the possibleecovery of neural functioning in humans are lacking.
Dogs exposed to severe air pollution have shown chronic inflam-ation and acceleration of Alzheimer-like pathology, suggesting
hat the brain is adversely affected by pollutants [37]. Some recenttudies have revealed that children living in urban areas arexposed to higher levels of carcinogenic air pollutants, such as PAH
by Dunnett’s test. Differences were considered significant at p < 0.05 (*). , �, and�= untreated culture, BNF-treated pre-adipocytes and BNF-neurogenic derivates,respectively.
and benzene, and that this exposure is correlated with an increasedhealth risk. Moreover, chronic exposure to significant concentra-tions of air pollutants could be a risk factor for Alzheimer’s diseaseand other neurodegenerative diseases [15,38].
The central nervous system has ‘windows of susceptibility’ toneural cell damage during its development. There should thereforebe a time-frame when exposure to an environmental contami-nant, even at a very low dose and for a very short period, wouldbe expected to have a significant negative impact on brain devel-opment. The effect of exposure to PAH mixtures on neurogenesisduring foetal development has been considered only recently, inthe aftermath of the destruction of the WTC building in New York[39], which released large amounts of dust and burnt gases intothe environment. In humans, the earliest synapses develop duringthe gestational period, and by 10 weeks immature synapses arepresent; however, the morphological characteristics of maturity
are reached within 24 months after birth.Unintended human gestational exposure of the foetus to envi-ronmental contaminants has been shown to result in alterationsin neurodevelopment, such as reduced head circumference, and

4 ogical
niIfe
b(bthic
nsda
nmtx[oesflsdav
tiotapretkcaTtiTiteiepal
idprrte
80 C. Scanarotti et al. / Chemico-Biol
eurobehavioral deficits, such as poorer outcome on selective test-ng of cognitive and neuromotor functioning in offspring [40].t therefore seems that nervous system malformations can ariserom alterations in neurogenesis subsequent to environmental PAHxposure.
Several studies in rats have evidenced neurotoxicity inducedy gestational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxinTCDD) or Benzo(a)pyrene (BP). Exposure to TCDD and B(a)P haseen seen to down-regulate early developmental glutamate recep-or subunit expression and function. Gestational exposure to B(a)Pas been found to reduce the capacity for learning in progeny by
nducing cortical axonal alterations, and to impair somatic sensoryortex functions [41–44].
Transplacental exposure to PAHs affects the outcome of preg-ancy not only in laboratory animals but also in humans. Indeed,uch exposure has been associated with the risk of pretermelivery. The metabolites of PAHs can therefore be regarded asborticidal agents [45].
Another important question concerns the bioaccumulation of aumber of highly lipophilic environmental contaminants or phar-aceutical drugs, such as TCDD, in white adipose tissue. The risk of
his phenomenon is related to the long half-lives of these lipophilicenobiotics – in the order of months or years – in the body tissue46–50]. Although the liver is the main organ involved in metab-lizing drugs, the expression and induction of drug-metabolizingnzymes by xenobiotics is also often observed in extrahepatic tis-ues. It has been reported that rodent adipose tissue expressesunctional AhR and NF-E2-related factor 2 (Nrf2) [51,52]. Theipophilic xenobiotics, including BNF, activate AhR and Nrf2 tran-cription factors to enhance the transcription of genes encodingetoxifying enzymes. Thus, lipophilic xenobiotics accumulated indipose tissue may exert as yet unknown effects through the acti-ation of these transcription factors.
Many literature data indicate that variations in the susceptibilityo chemical-induced mutagenic or carcinogenic activity in differentndividuals may partly be explained by differences in the activityf enzymes responsible for the metabolic activation or detoxifica-ion of these toxicants. CYP1A1 is a key enzyme in the metabolicctivation of polycyclic aromatic hydrocarbons [53,54] and alsolays a role in the pathogenesis of adverse effects by generatingeactive intermediates, i.e. free radicals, which increase the toxicffects of active metabolites. Although the hepatic and pulmonaryoxicity of PAH has been extensively characterized, very little isnown about the potential of these chemicals to damage neuralells. Monitoring the effects of environmental pollution, especiallyir pollution, on transgenic mice has not proved successful [55].o date, research on aquatic species has yielded some evidencehat PAH and BNF exert toxic effects on brain functions by affect-ng dopaminergic, noradrenergic and serotonergic systems [20,21].hese studies on the effects of BNF and other PAH-like compoundsn animal species other than mammals could contribute to iden-ifying some pathways or biomarkers for evaluating the risk ofnvironmental pollution in humans. An innovative and reliablen vitro model is needed in order to evaluate the possible toxicffects of a chemical compound on the human nervous system. Weropose human mesenchymal adult stem cells from fat tissue asmodel, since these cells can differentiate towards a neurogenic
ineage in appropriate conditions.In our study, an initial observation of cell morphology was made
n order to ensure that the pre-adipocyte samples from the sixonors were sufficiently similar, and to exclude those samples that
roved too different from the others. Thus, all experimental dataeported refer to three of the donors. Moreover, a cytofluorimet-ic analysis was performed in the early pre-adipocyte populationo confirm the mesenchymal origin of these cells: pre-adipocytesxpressed CD44 (95%), CD90 (99%) and CD105 (96%). The expres-Interactions 184 (2010) 474–483
sion of CD44 and CD105 and the absence of CD45 and CD14 indicatethat isolated cells possess typical mesenchymal features, includingtheir capacity for differentiation. As shown in this research, neu-rogenic derivate cultures seem to maintain several differentiationmarkers of neurogenic lineage. The membrane localisation of ChATseems to suggest that our neurogenic cells were functioning and theperformance of this enzyme has been reported to be regulated byits binding to the plasma membrane [36]. The enzyme is not activewhen it is inside the cytoplasm, while it is active when it is localisedon the cholinergic membrane. At this differentiative stage, AChE ispresent in non-catalytic forms, during the first culture stages, andbegins to acquire catalytic activity (also demonstrated by Aluigi etal. [16]). This coordinated distribution of signal molecules suggestsmaturation of the cholinergic system, and indicates a commitmentto neural differentiation. We used these signal molecules as mark-ers to identify the early steps of differentiation, in order to relatethe detoxification ability of these pre-adipocytes to their ultimatefate as neural cells.
The commercial human neural progenitor line, ReNcell VM,which was derived from 10-week embryonic midbrain tissue,exhibits a dopaminergic neural phenotype [56] and is considered asource of differentiated neurons for the study of neurological dis-eases. Our in vitro model of neurogenic-committed cells derivedfrom pre-adipocytes could, like adult stem cells, be useful in inves-tigating the neurotoxic potential of a xenobiotic on biochemical andfunctional markers during the differentiation and development ofthe neurogenic phenotype.
The neurotoxic potential of a chemical in vivo is dependentupon metabolic parameters, such as biotransformation, lipid-aqueous partitioning, and distribution. Furthermore, certain classesof compounds must be activated by cytochrome P450-dependentmonooxygenase enzymes, and therefore require a metabolic-activating system. As some chemical compounds can modulateCYP expression, an in vitro culture system may partly satisfy theserequirements. The present study is the first to demonstrate thatpre-adipocytes and their neurogenic derivates display basal activ-ity of ECOD and EROD, and that this latter is expressed at lowerlevels in neurogenic derivate cells than in pre-adipocytes. Bothenzymatic activities were increased by a classical PAH-like inducer:BNF-treated neurogenic cells showed higher levels of ECOD activ-ity than pre-adipocytes, and the increase in induced activity wasmore marked (2.6- and 1.3-fold, respectively). EROD activity andimmunoblotting analysis confirmed the presence and inducibilityof the specific CYP1A1 protein.
An interesting finding is the striking sensitivity of pre-adipocytes and neurogenic derivates to the classical CYP450-inducer BNF, as shown by cytotoxicity indexes. Little evidence ofthe toxicity of BNF or other CYP450 inducers on mammalian cellsis available in the literature. In our previous study [57] on theeffects of �-hexachlorocyclohexane in primary cultures of mouse,rat and human hepatocytes and in three hepatoma cell lines, wereported that human HepG2 and mouse Hepa1c1c7 hepatoma cellswere much more sensitive to the cytotoxic activity of methyl-cholanthrene, the well-known CYP1A1 inducer, than the rat FaOhepatoma cell line. However, recent papers have reported thatexposure of human endothelial cells to 1 �M BNF for 48 h increasedEROD activity and caused significant DNA damage, and that BNFadministration to rats, in an in vivo hepatocarcinogenesis model,exacerbated oxidative damage and lipid peroxidation [58,59].
The toxicity of BNF has been investigated in several fish models.In primary cultures of rainbow trout hepatocytes exposed to high
doses of BNF, cytotoxic effects were seen to increase as EROD activ-ity declined [60]. Moreover, in other studies on fishes, erythrocyticnuclear abnormalities and hepatic damage after 6-day exposure to0.9 �M BNF were reported, while at a lower concentration (0.1 �M)an increase in hepatic transaminase was observed. In addition,
gical
ei
scSoiatlmao
0Asar5acrio[
mAiPibtt
oieebthxrtc
bpetp
5
tvns
pp
[
[
[
[
[
C. Scanarotti et al. / Chemico-Biolo
xposure to BNF, alone or in association with PAH heterocycles,nduced marked embryotoxicity [61,62].
We first investigated ECOD activity because this is related toeveral CYP isoforms, including CYP1A1; this provided some indi-ation of the bioactivating potential of both our in vitro models.ubsequently, after finding evidence of ECOD induction, we focusedur research on CYP1A-dependent-EROD activity and, more specif-cally, on the analysis of CYP1A1 protein expression, in order topply our models to the investigation of the carcinogenic activi-ies of PAH-related compounds. Obviously, the protein expressionevel of an enzyme does not totally reflect its activity; indeed, enzy-
atic activity could be increased because of higher reaction speednd/or induced transcription of the gene, or because of the numberf protein molecules.
In our study, pre-adipocytes and neurogenic cells exposed to.1 �M BNF for 48 h displayed a viability index of about 70%.lthough this BNF dose was lower than those reported in othertem cell models [63], we observed a significant induction of ECODnd EROD in both cell cultures. This induction was greater in neu-ogenic cells, especially with regard to EROD, which exhibited a-fold increase after BNF treatment. Flavonoids may act as AhRgonists, and the AhR ligand-dependent activation of AhR inducesell-cycle arrest [22]. In our study, the toxicity of BNF could beelated to the stem-like phenotype of our model, a possibility whichs in line with some literature data on DNA-adducts and apoptosisbserved in rat liver epithelial stem-like cells after exposure to PAH64].
Toxic effects of PAHs are involved in the disruption of lipidetabolism as well as in immunotoxicity [65]. Moreover, severalhR-regulated genes have recently been linked to PAH-induced
mmunotoxicity, teratogenicity and metabolic syndrome [66].AHs have been seen to suppress NR1H4, a nuclear receptornvolved in the expression of genes critical to the homeostasis ofile acid and cholesterol [67–70]. These effects could suggest thathe PAHs reduced expression of NR1H4, via AhR, causes the disrup-ion of lipid metabolism.
With regard to these observations, another relevant result ofur study was the demonstration of the inducibility of CYP4501An human pre-adipocytes. The evidence of drug-metabolizingnzymes in adipose tissue highlights the potential risk of unknownffects of exposure to xenobiotics [51,52]. In other words, theioaccumulation of lipophilic compounds and the biotransforma-ion capability of adipocytes may have important implications foruman health, even in the absence of direct intensive exposure toenobiotics. This issue deserves further study in order to assess theeal risk to human health of the systemic uptake of environmen-al toxicants, and to investigate their metabolic activation, whichould lead to the release of reactive intermediates.
Considering that the toxicological effects of aromatic hydrocar-ons on humans result from the absorption and metabolism ofollutants, we propose the use of our two alternative in vitro mod-ls – pre-adipocytes and neurogenic derivate cells – to evaluatehe cytotoxic/biological effects of environmental chemical com-ounds.
. Conclusions
The main point that emerged from the present study was thathe analyses performed confirmed the usefulness of our models inerifying an increase in CYP1A protein and its enzymatic activity in
eurogenic differentiating cells and in pre-adipocytes, as in adulttem cell models.Other relevant findings can be summarised as follows: (1) asre-adipocytes can be obtained in large numbers from liposuctionrocedures, they constitute a source of human adult mesenchy-
[
Interactions 184 (2010) 474–483 481
mal stem cells that does not raise ethical issues; (2) since thesehuman adult mesenchymal stem cells can be differentiated intoseveral different cell types, our innovative models may be useful inanalysing, in the same donor, the effects of xenobiotics on severalcell-specific biological functions; (3) as pre-adipocytes constitute areservoir of lipophilic compounds, such as PAH, this in vitro modelcould be used to analyse the in vivo effects of unintended exposureto contaminants; (4) we also suggest using neurogenic derivativesfrom pre-adipocytes to study the first steps of morphological andfunctional foetal–embryonic development.
Further studies using genomic, proteomic and metabonomictechnologies will be required in order to test the ability of bothin vitro models to identify novel biomarkers of the toxicity of PAHand other xenobiotics/pollutants.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgements
The authors gratefully acknowledge the skilful help of Mr.Luca Mastrobuono and Dr. Bernard Patrick; Dr. Marina Sartini,researcher of the Health Science Department of the University ofGenova, for statistical analysis; Prof. Roberto Fiocca for his hospi-tality and assistance in preparing this work.
References
[1] A.J. Katz, R. Llull, M.H. Hedrick, J.W. Furtrell, Emerging approaches to the tissueengineering of fat, Clin. Plast. Surg. 26 (1999) 587–603.
[2] J.M. Gimble, A.J. Katz, B.A. Bunnell, Adipose-derived stem cells for regenerativemedicine, Circ. Res. 100 (2007) 1249–1260.
[3] P.H. Ashjian, A.S. Elbarbary, B. Edmonds, D. DeUgarte, M. Zhu, P.A. Zuk, H.P.Lorenz, P. Benhaim, M.H. Hedrick, In vitro differentiation of human processedlipoaspirate cells into early neural progenitors, Plast. Reconstr. Surg. 111 (2003)1922–1931.
[4] E. Raposio, C. Guida, I. Baldelli, F. Benvenuto, M. Curto, L. Paleari, F. Filippi,R. Fiocca, G. Robello, P.L. Santi, Characterization and induction of human pre-adipocytes, Toxicol. In Vitro 21 (2007) 330–334.
[5] B.M. Strem, K.C. Hicok, M. Zhu, I. Wulur, Z. Alfonso, R.E. Schreiber, J.K. Fraser,M.H. Hedrick, Multipotential differentiation of adipose tissue-derived stemcells, Keio J. Med. 54 (2005) 132–141.
[6] P.A. Zuk, M. Zhu, P. Ashjian, D.A. De Ugarte, J. Huang, H. Mizuno, Z.C. Alfonso,J.K. Fraser, P. Benhaim, M.H. Hedrick, Human adipose tissue is a source of mul-tipotent stem cells, Mol. Biol. Cell 13 (2002) 4279–4295.
[7] P.A. Zuk, M. Zhu, H. Mizuno, J. Huang, J.W. Futrell, A.J. Katz, P. Benhaim, H.P.Lorenz, M.H. Hedrick, Multilineage cells from human adipose tissue: implica-tion for cell-based therapies, Tissue Eng. 7 (2001) 211–228.
[8] G. Astori, F. Vignati, S. Bardelli, M. Tubio, M. Gola, V. Albertini, F. Bambi, G. Scali,D. Castelli, V. Rasini, G. Soldati, T. Moccetti, In vitro and multicolor phenotypiccharacterization of cell subpopulations identified in fresh human adipose tissuestromal vascular fraction and in the derived mesenchymal stem cells, Transl.Med. 5 (2007) 55.
[9] D.A. De Ugarte, K. Morizono, A. Elbarbary, Z. Alfonso, P.A. Zuk, M. Zhu, J.L.Dragoo, P. Ashjian, B. Thomas, P. Benhaim, I. Chen, J. Fraser, M.H. Hedrick, Com-parison of multi-lineage cells from human adipose tissue and bone marrow,Cells Tissues Organs 174 (2003) 101–109.
10] S. Gronthos, D.M. Franklin, H.A. Leddy, P.G. Robey, R.W. Storms, J.M. Gimble,Surface protein characterization of human adipose tissue-derived stromal cells,J. Cell. Physiol. 189 (2001) 54–63.
11] E. Meliga, B.M. Strem, H.J. Duckers, P.W. Serruys, Adipose-derived cells, CellTransplant. 16 (2007) 963–970.
12] A.M. Parker, H. Shang, M. Khurgel, A.J. Katz, Low serum and serum-free cultureof multipotent human adipose stem cells, Cytotherapy 9 (2007) 637–646.
13] H. Mizuno, H. Hyakusoku, Mesengenic potential and future clinical perspec-tive of human processed lipoaspirate cells, J. Nippon Med. School 70 (2003)300–306.
14] L. Calderón-Garciduenas, B. Azzarelli, H. Acuna, R. Garcia, T.M. Gambling, N.Osnaya, S. Monroy, M.R. Del Tizapantzi, J.L. Carson, A. Villarreal-Calderon, B.
Rewcastle, Air pollution and brain damage, Toxicol. Pathol. 30 (2002) 373–389.15] L. Calderón-Garciduenas, M. Franco-Lira, R. Torres-Jardón, C. Henriquez-Roldán, G. Barragán-Mejía, G. Valencia-Salazar, A. González-Maciel, R.Reynoso-Robles, R. Villarreal-Calderón, W. Reed, Pediatric respiratory and sys-temic effects of chronic air pollution exposure: nose, lung, heart, and brainpathology, Toxicol. Pathol. 35 (2007) 154–162.

4 ogical
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
82 C. Scanarotti et al. / Chemico-Biol
16] M.G. Aluigi, R. Coradeghini, C. Guida, C. Scanarotti, A.M. Bassi, C. Falugi, P.Santi, E. Raposio, Pre-adipocytes commitment to neurogenesis. 1. Preliminarylocalisation of cholinergic molecules, Cell Biol. Int. 33 (2009) 594–601.
17] K.M. Safford, K.C. Hicok, S.D. Safford, Y.D. Halvorsen, W.O. Wilkison, J.M. Gimble,H.E. Rice, Neurogenic differentiation of murine and human adipose-derivedstromal cells, Biochem. Biophys. Res. Commun. 294 (2002) 371–379.
18] E. Arinc, A. Sen, A. Bozcaarmutlu, Cytochrome P4501A and associated mixed-function oxidase in fish as a biomarker for toxic carcinogenic polluntants in theaquatic environment, Pure Appl. Chem. 72 (2000) 958–994.
19] T. Shimada, K. Inoune, Y. Suzuki, T. Kawai, E. Azuna, T. Nakajima, M. Shindo,K. Kurose, A. Sugie, Y. Yamagishi, Y. Fujii-Kuriyama, M. Hashimoto, Arylhy-drocarbon receptor-dependent induction of liver and lung cytochromes P4501A1, 1A2 and 1B1 by polycyclic aromatic hydrocarbons and polychlorinatedbiphenyls in genetically engineered C57BL/6J mice, Carcinogenesis 23 (2002)1199–1207.
20] N. Aluru, M.M. Vijayan, Brain transcriptomics in response to beta-naphthoflavone treatment in rainbow trout: the role of aryl hydrocarbonreceptor signaling, Aquat. Toxicol. 87 (2008) 1–12.
21] M. Gesto, A. Tintos, J.L. Soengas, J.M. Míguez, beta-Naphthoflavone andbenzo(a)pyrene alter dopaminergic, noradrenergic, and serotonergic systemsin brain and pituitary of rainbow trout (Oncorhynchus mykiss), Ecotoxicol.Environ. Saf. 72 (2009) 191–198.
22] A. Puga, J. Marlowe, S. Barnes, C.Y. Chang, A. Maier, Z. Tan, J.K. Kerzee, X. Chang,M. Strobeck, E.S. Knudsen, Role of the aryl hydrocarbon receptor in cell cycleregulation, Toxicology 181–182 (2002) 171–177.
23] M. Schwarz, A. Buchmann, S. Stinchcombe, A. Kalkuhl, K. Bock, Ah receptor lig-ands and tumor promotion: survival of neoplastic cells, Toxicol. Lett. 112–113(2000) 69–77.
24] M.G. Aluigi, C. Angelini, C. Falugi, R. Fossa, P. Genever, L. Gallus, P.G. Layer,G. Prestipino, Z. Rakonczay, M. Sgro, H. Thielecke, S. Trombino, Interactionbetween organophosphate compounds and cholinergic functions during devel-opment, Chem. Biol. Interact. 157–158 (2005) 305–316.
25] M. Fukui, N. Nakamichi, M. Yoneyama, S. Ozawa, S. Fujimori, Y. Takahata, N.Nakamura, H. Taniura, Y. Yoneda, Modulation of cellular proliferation and dif-ferentiation through GABA(B) receptors expressed by undifferentiated neuralprogenitor cells isolated from fetal mouse brain, J. Cell. Physiol. 216 (2008)507–519.
26] J.R. Davis, R.J. Penny, Improved fluorescence method for identifying sex chro-matin in formalin-fixed tissue, Am. J. Clin. Pathol. 75 (1981) 731–733.
27] A. Miganti, C. Falugi, An epithelial localization of acethylcholinesterase in theascidian Ciona intestinalis embryos and larve, Acta Embryol. Morphol. Exp. 1(1980) 143–155.
28] T. Mosman, Rapid colorimetric assay for cellular growth and survival: appli-cation to proliferation and cytotoxicity assays, J. Immunol. Methods 65 (1983)55–63.
29] E. Borenfreund, H. Babik, N. Martin-Alguacil, Comparison of two in vitro cyto-toxicity assays-the neutral red (NR) and tetrazolium MTT tests, Toxicol. In Vitro2 (1988) 1–6.
30] R.J. Riddell, R.H. Clothier, M. Ball, An evaluation of three in vitro cytotoxicityassays, Food Chem. Toxicol. 24 (1986) 469–471.
31] M.T. Donato, J.V. Castell, M.J. Gomez-Lechon, Cytochrome P450 activities inpure and co-cultured rat hepatocytes. Effects of model inducers, In Vitro Cell.Dev. Biol. Anim. 30 (1994) 825–832.
32] X. Wen, U.K. Walle, T. Walle, 5,7-Dimethoxyflavone downregulates CYP1A1expression and benzo[a]pyrene-induced DNA binding in Hep G2 cells, Carcino-genesis 26 (2005) 803–809.
33] A.M. Bassi, S. Ledda, S. Penco, S. Menini, G. Muzio, R.A. Canuto, M. Ferro,Changes of CYP1A1, GST and ALDH3 enzymes in hepatoma cell lines under-going enhanced lipid peroxidation, Free Radic. Biol. Med. 29 (2000) 1186–1196.
34] M.M. Bradford, A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein–dye binding, Anal.Biochem. 72 (1976) 248–254.
35] S. Biagioni, A.M. Tata, C. Agrati, F. Cianfrani, G. Augusti-Tocco, Modulation ofcholinergic marker expression by nerve growth factor in dorsal root ganglia, J.Neurosci. Res. 62 (2000) 591–599.
36] T. Dobransky, R.J. Rylett, Functional regulation of choline acetyltransferase byphosphorylation, Neurochem. Res. 28 (2003) 537–542.
37] L. Calderón-Garciduenas, W. Reed, R.R. Maronpot, C. Henríquez-Roldán, R.Delgado-Chavez, A. Calderón-Garciduenas, I. Dragustinovis, M. Franco-Lira, M.Aragón-Flores, A.C. Solt, M. Altenburg, R. Torres-Jardón, J.A. Swenberg, Braininflammation and Alzheimer’s-like pathology in individuals exposed to severeair pollution, Toxicol. Pathol. 32 (2004) 650–658.
38] P.L. Grover, Pathways involved in the metabolism and activation of polycyclichydrocarbons, Xenobiotica 16 (1986) 915–931.
39] F.P. Perera, D. Tang, V. Rauh, K. Lester, W.Y. Tsai, Y.H. Tu, L. Weiss, L. Hoepner,J. King, G. Del Priore, S.A. Lederman, Relationships among polycyclic aromatichydrocarbon–DNA adducts, proximity to the World Trade Center, and effectson fetal growth, Environ. Health Perspect. 113 (2005) 1062–1067.
40] F.P. Perera, V. Rauh, W.-Y. Tsai, P. Kinney, D. Camann, D. Barr, T. Bernert,
R. Garfinkel, Y.-H. Tu, D. Diaz, J. Dietrich, R. Whyatt, Effects of transplacen-tal exposure to environmental pollutants on birth outcomes in a multiethnicpopulation, Environ. Health Perspect. 111 (2003) 201–205.41] D.D. Wormley, A. Ramesh, D.B. Hood, Environmental contaminant-mixtureeffects on CNS development, plasticity and behavior, Toxicol. Appl. Pharm. 197(2004) 49–65.
[
Interactions 184 (2010) 474–483
42] D.B. Hood, L. Woods, L. Brown, S. Johnson, F.F. Ebner, Gestational 2,3,7,8-tetrachlorodibenzo-p-dioxin exposure effects on sensory cortex function,Neurotoxicology 27 (2006) 1032–1042.
43] L.A. Brown, H. Khousbouei, J.S. Goodwin, C.V. Irvin-Wilson, A. Ramesh, L.Sheng, M.M. McCallister, G.C. Jiang, M. Aschner, D.B. Hood, Down-regulationof early ionotrophic glutamate receptor subunit developmental expression asa mechanism for observed plasticity deficits following gestational exposure tobenzo(a)pyrene, Neurotoxicology 28 (2007) 965–978.
44] M.M. McCallister, M. Maguire, A. Ramesh, Q. Aimin, S. Liu, H. Khoshbouei, M.Aschner, F.F. Ebner, D.B. Hood, Prenatal exposure to benzo(a)pyrene impairslater-life cortical neuronal function, Neurotoxicology 29 (2008) 846–854.
45] G. Huel, J. Godin, N. Frery, F. Girard, T. Moreau, C. Nessman, Arylhydrocarbonhydroxylase activity in human placenta and threatened preterm delivery, J.Expo. Anal. Environ. Epidemiol. 3 (1993) 149–187.
46] M.H. Bickel, The role of adipose tissue in the distribution and storage of drug,Prog. Drug Res. 28 (1984) 273–303.
47] R. Latini, G. Tognoni, R.E. Kates, Clinical pharmacokinetics of amiodarone, Clin.Pharmacokinet. 9 (1984) 136–156.
48] S. Mühlebach, P.A. Wyss, M.H. Bickel, Comparative adipose tissue kinetics ofthiopental, DDE and 2,4,5,2′ ,4′ ,5′-hexachlorobiphenyl in the rat, Xenobiotica15 (1985) 485–491.
49] D. Flesch-Janys, H. Becher, P. Gurn, D. Jung, J. Konietzko, A. Manz, O. Päpke,Elimination of polychlorinated dibenzo-p-dioxins and dibenzofurans in occu-pationally exposed persons, J. Toxicol. Environ. Health 47 (1996) 363–378.
50] J.E. Michalek, J.L. Pirkle, S.P. Caudill, R.C. Tripathi, D.G. Patterson Jr., L.L. Need-ham, Pharmacokinetics of TCDD in veterans of Operation Ranch Hand: 10-yearfollow-up, J. Toxicol. Environ. Health 47 (1996) 209–220.
51] K. Yoshinari, T. Sato, N. Okino, J. Sugatani, M. Miwa, Expression and inductionof cytochromes p450 in rat white adipose tissue, J. Pharmacol. Exp. Ther. 311(2004) 147–154.
52] K. Yoshinari, N. Okino, T. Sato, J. Sugatani, M. Miwa, Induction of detoxifyingenzymes in rodent white adipose tissue by aryl hydrocarbon receptor agonistsand antioxidants, Drug Metab. Dispos. 34 (2006) 1081–1089.
53] D.R. Thakker, H. Yagi, W. Levin, A.W. Wood, A.H. Conney, D.M. Jerina, Polycyclicaromatic hydrocarbons: metabolic activation to ultimate carcinogens, in: M.W.Anders (Ed.), Bioactovation of Foreign Compounds, Academic Press, London,1985, pp. 177–242.
54] A. Kasai, N. Hiramatsu, K. Hayakawa, J. Yao, M. Kitamura, Direct, continuousmonitoring of air pollution by transgenic sensor mice responsive to halo-genated and polycyclic aromatic hydrocarbons, Environ. Health Perspect. 116(2008) 349–354.
55] M. Ferro, A.M. Bassi, D. Adamo, F. Mattioli, L. Robbiano, G. Nanni, Studies onalpha-hexachlorocyclohexane cytotoxicity, genotoxicity and cytochrome P450induction in primary hepatocytes and hepatoma cell lines from rodents andhumans, ATLA, Altern. Lab. Anim. 25 (1997) 139–152.
56] R. Donato, E.A. Miljan, S.J. Hines, S. Aouabdi, K. Pollock, S. Patel, F.A. Edwards,J.D. Sinden, Differential development of neuronal physiological responsivenessin two human neural stem cell lines, BMC Neurosci. 8 (2007) 36.
57] A. Annas, E. Brittebo, B. Hellman, Evaluation of benzo(a)pyrene-induced DNAdamage in human endothelial cells using alkaline single cell gel electrophoresis,Mutat. Res. 471 (2000) 145–155.
58] Y. Dewa, J. Nishimura, M. Muguruma, M. Jin, Y. Saegusa, T. Okamura, M.Tasaki, T. Umemura, K. Mitsumori, beta-Naphthoflavone enhances oxida-tive stress responses and the induction of preneoplastic lesions in adiethylnitrosamine-initiated hepatocarcinogenesis model in partially hepate-ctomized rats, Toxicology 244 (2008) 179–189.
59] M. Pesonen, T. Andersson, Characterization and induction of xenobiotic metab-olizing enzyme activities in a primary culture of rainbow trout hepatocytes,Xenobiotica 21 (1991) 461–471.
60] M. Pacheco, M.A. Santos, Biotransformation, genotoxic, and histopathologicaleffects of environmental contaminants in European eel (Anguilla anguilla L.),Ecotoxicol. Environ. Saf. 53 (2002) 331–347.
61] D.M. Wassenberg, A.L. Nerlinger, L.P. Battle, R.T. Di Giulio, Effects of the poly-cyclic aromatic hydrocarbon heterocycles, carbazole and dibenzothiophene,on in vivo and in vitro CYP1A activity and polycyclic aromatic hydrocarbon-derived embryonic deformities, Environ. Toxicol. Chem. 24 (2005) 2526–2532.
62] T. Miki, T. Lehman, H. Cai, D.B. Stolz, S.C. Strom, Stem cell characteristics ofamniotic epithelial cells, Stem Cells 23 (2005) 1549–1559.
63] Z. Andrysík, J. Vondrácek, M. Machala, P. Krcmár, L. Svihálková-Sindlerová, A.Kranz, C. Weiss, D. Faust, A. Kozubík, C. Dietrich, The aryl hydrocarbon receptor-dependent deregulation of cell cycle control induced by polycyclic aromatichydrocarbons in rat liver epithelial cells, Mutat. Res. 615 (2007) 87–97.
64] K. Chramostová, J. Vondrácek, L. Sindlerová, B. Vojtesek, A. Kozubík, M. Machala,Polycyclic aromatic hydrocarbons modulate cell proliferation in rat hep-atic epithelial stem-like WB-F344 cells, Toxicol. Appl. Pharmacol. 196 (2004)136–148.
65] K.P. Miller, K.S. Ramos, Impact of cellular metabolism on the biological effectsof benzo[a]pyrene and related hydrocarbons, Drug Metab. Rev. 33 (1999) 1–35.
66] S. Iwano, et al., Identification of AhR-regulated genes involved in PAH-
induced immunotoxicity using a highly-sensitive DNA chip, 3D-GeneTMHuman Immunity and Metabolic Syndrome 9k, Toxicol. In Vitro (2009),doi:10.1016/j.tiv.2009.08.030.67] M. Makishima, A.Y. Okamoto, J.J. Repa, H. Tu, R.M. Learned, A. Luk, M.V. Hull,K.D. Lustig, D.J. Mangelsdorf, B. Shan, Identification of a nuclear receptor forbile acids, Science 284 (1999) 1362–1365.

[
C. Scanarotti et al. / Chemico-Biological
68] D.J. Parks, S.G. Blanchard, R.K. Bledsoe, G. Chandra, T.G. Consler, S.A. Kliewer,J.B. Stimmel, T.M. Willson, A.M. Zavacki, D.D. Moore, J.M. Lehmann, Bile acids:natural ligands for an orphan nuclear receptor, Science 284 (1999) 1365–1368.
[
[
Interactions 184 (2010) 474–483 483
69] H. Wang, J. Chen, K. Hollister, L.C. Sowers, B.M. Forman, Endogenous bile acidsare ligands for the nuclear receptor FXR/BAR, Mol. Cell 3 (1999) 543–553.
70] H. Tu, A.Y. Okamoto, B. Shan, FXR, a bile acid receptor and biological sensor,Trends Cardiovasc. Med. 10 (2000) 30–35.
Related Documents











