The Prostate 68:588 ^598 (2008) NeuroendocrineTransdifferentiation Induced by VPAIs Mediated by PPARg Activation and Confers Resistance to AntiblasticTherapy in Prostate Carcinoma Adriano Angelucci, 1 * Paola Muzi, 2 Loredana Cristiano, 1 Danilo Millimaggi, 2 AnnaMaria Cimini, 1 Vincenza Dolo, 2 Roberto Miano, 3 Carlo Vicentini, 4 Maria Paola Ceru `, 1 and Mauro Bologna 1 1 Department of Basic and Applied Biology,University of L’Aquila, L’Aquila, Italy 2 Department of Experimental Medicine,University of L’Aquila, L’Aquila, Italy 3 Department of Urology,University of Tor Vergata, Rome, Italy 4 Department of Surgery,University of L’Aquila, L’Aquila, Italy BACKGROUND. Prostate cancer (PCa) is the most commonly diagnosed cancer in men in the Western Countries. When prostatectomy fails to eradicate the primary tumor, PCa is generally refractory to all therapeutic approaches. Valproic acid (VPA) is a promising anticancer agent recently assigned to the class of histone deacetylase (HDAC) inhibitors. However molecular mechanisms underlying VPA action in PCa cells are largely unknown and further experimental validation to prove its potential application in clinic practice is needed. RESULTS. In our study we show that VPA is a potent inducer of neuro-endocrine transdifferentiation (NET) in androgen receptor null PCa cells, both in vitro and in vivo. NET was an early event detectable through the expression of neuro-endocrine (NE) markers within 72 hr after VPA treatment and it was associated to a reduction in the overall cell proliferation. When we interrupted VPA treatment we observed the recovery in residual cells of the basal proliferation rate both in vitro and in a xenograft model. The NET process was related to Bcl-2 over-expression in non-NE PCa cells and to the activation of PPARg in NE cells. The use of specific PPARg antagonist was able to reduce significantly the expression of NE markers induced by VPA. CONCLUSIONS. Our data indicate that the use of VPA as monotherapy in PCa has to be considered with extreme caution, since it may induce an unfavorable NET. In order to counteract the VPA-induced NET, the inhibition of PPARg may represent a suitable adjuvant treatment strategy and awaits further experimental validation. Prostate 68: 588 – 598, 2008. # 2008 Wiley-Liss, Inc. KEY WORDS: histone deacetylase inhibitors; Bcl-2; secretory neuron-specific enolase; b-tubulin III; cis-platinum INTRODUCTION Although Prostate cancer (PCa) emerges from secretory cells, neuro-endocrine transdifferentiation (NET) has been demonstrated at various extents in virtually all cases of PCa. Since NE cells are thought to be a cell lineage that separates early in development from that of the secretory cells, their origin in PCa is controversial [1]. However NET is a well documented process observed in several PCa cell lines and Abbreviations: PCa, prostate cancer; VPA, valproic acid; HDAC, histone deacetylase; NE, neuro-endocrine; NET, neuro-endocrine transdifferentiation; NSE, secretory neuron-specific enolase; b-tubIII, b-tubulin III; PPARg, peroxisomes proliferators-activated receptor g. *Correspondence to: Dr. Adriano Angelucci, PhD, Department of Experimental Medicine, University of L’Aquila, Via Vetoio, Cop- pito2, 67100 L’Aquila, Italy. E-mail: [email protected] Received 5 June 2007; Accepted 18 October 2007 DOI 10.1002/pros.20708 Published online 20 February 2008 in Wiley InterScience (www.interscience.wiley.com). ȣ 2008 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Prostate 68:588 ^598 (2008)
NeuroendocrineTransdifferentiation InducedbyVPAIsMediatedbyPPARgActivationandConfersResistance
toAntiblasticTherapy in ProstateCarcinomaAdriano Angelucci,1* Paola Muzi,2 Loredana Cristiano,1 Danilo Millimaggi,2
AnnaMaria Cimini,1 Vincenza Dolo,2 Roberto Miano,3 Carlo Vicentini,4
Maria Paola Ceru,1 and Mauro Bologna1
1Departmentof Basicand Applied Biology,Universityof L’Aquila, L’Aquila, Italy2Departmentof ExperimentalMedicine,Universityof L’Aquila, L’Aquila, Italy
3DepartmentofUrology,Universityof TorVergata, Rome, Italy4Departmentof Surgery,Universityof L’Aquila, L’Aquila, Italy
BACKGROUND. Prostate cancer (PCa) is themost commonly diagnosed cancer inmen in theWestern Countries. When prostatectomy fails to eradicate the primary tumor, PCa is generallyrefractory to all therapeutic approaches. Valproic acid (VPA) is a promising anticancer agentrecently assigned to the class of histone deacetylase (HDAC) inhibitors. However molecularmechanisms underlyingVPA action in PCa cells are largely unknown and further experimentalvalidation to prove its potential application in clinic practice is needed.RESULTS. In our study we show that VPA is a potent inducer of neuro-endocrinetransdifferentiation (NET) in androgen receptor null PCa cells, both in vitro and in vivo. NETwas an early event detectable through the expression of neuro-endocrine (NE) markers within72 hr after VPA treatment and it was associated to a reduction in the overall cell proliferation.When we interrupted VPA treatment we observed the recovery in residual cells of the basalproliferation rate both in vitro and in a xenograft model. The NET process was related to Bcl-2over-expression in non-NE PCa cells and to the activation of PPARg in NE cells. The use ofspecific PPARg antagonist was able to reduce significantly the expression of NE markersinduced by VPA.CONCLUSIONS. Our data indicate that the use of VPA as monotherapy in PCa has to beconsidered with extreme caution, since it may induce an unfavorable NET. In order tocounteract the VPA-induced NET, the inhibition of PPARg may represent a suitable adjuvanttreatment strategy and awaits further experimental validation. Prostate 68: 588–598, 2008.# 2008 Wiley-Liss, Inc.
KEY WORDS: histone deacetylase inhibitors; Bcl-2; secretory neuron-specific enolase;b-tubulin III; cis-platinum
INTRODUCTION
Although Prostate cancer (PCa) emerges fromsecretory cells, neuro-endocrine transdifferentiation(NET) has been demonstrated at various extents invirtually all cases of PCa. Since NE cells are thought tobe a cell lineage that separates early in developmentfrom that of the secretory cells, their origin in PCa iscontroversial [1]. However NET is a well documentedprocess observed in several PCa cell lines and
Abbreviations: PCa, prostate cancer; VPA, valproic acid; HDAC,histone deacetylase; NE, neuro-endocrine; NET, neuro-endocrinetransdifferentiation; NSE, secretory neuron-specific enolase; b-tubIII,b-tubulin III; PPARg, peroxisomes proliferators-activated receptor g.*Correspondence to: Dr. Adriano Angelucci, PhD, Department ofExperimental Medicine, University of L’Aquila, Via Vetoio, Cop-pito2, 67100 L’Aquila, Italy. E-mail: [email protected] 5 June 2007; Accepted 18 October 2007DOI 10.1002/pros.20708Published online 20 February 2008 in Wiley InterScience(www.interscience.wiley.com).
� 2008 Wiley-Liss, Inc.

experimental models where it has been associated tothe presence of neuronal-specific markers includingthe secretory neuron-specific enolase (NSE) and cyto-skeleton protein b-tubulin III (b-tubIII). Data on theprognostic impact of NET in PCa are at presentconflicting [2–4]. Studies confirming a prognostic roleof NET have suggested that NE cells are directlyinvolved in the resistance to therapy [5,6]. In fact PCacells presenting NE features are able to escapeapoptosis and at the same time seem to enhance themalignant potential of surrounding non-NE tumorcells. This aspect may be achieved in various waysincluding paracrine release of protective and angio-genic factors [7,8]. Although NET may potentiallyinfluence the outcome of a therapy, to date few dataabout the interaction between NET and drugs areavailable. The best-known association is between anincreased number of NE tumor cells and the androgensuppression therapy [9]. Moreover a potential linkbetween AR inactivation and NET has been describedin vitro suggesting a direct negative role for androgen-depletion therapy in PCa prognosis [10].
Since the demonstration that valproic acid (VPA) is aneffective histonedeacetylase (HDAC) inhibitor [11]manyresearches have focused on its potential role as a validantitumoral drug. The increasing interest about VPA isexplained also by the limited side effects demonstrated inits long clinical use as anticonvulsant drug.We and otherauthors have demonstrated that VPAwhen used at highconcentrations has a significant antiproliferative effect onPCa cells, inducing cell cycle arrest and apoptotic celldeath [12,13]. VPAmediated apoptosis in PCa cell lines isattributable mainly to the downmodulation of Bcl-2 withthe consequent mitochondrial depolarization. However,as usual for HDAC inhibitors, VPA has on tumor cells apleiotropic effect affecting the functioning of severaltransduction pathways and determining a severe mod-ification in tumor phenotype. In particular VPA inducestumor differentiation in neuroblastoma [14] and intransformed cells [11]. The therapeutic impact of thiscomplex action on a multifocal tumor as PCa promptsfurther studies before to individuate the best clinicalapplication.
The peroxisome proliferators-activated receptor g(PPARg) is a hormone nuclear receptor whose tran-scriptional activity controls cell proliferative status. Itsoverexpression has been associated to cell differentia-tion in PCa cells [15]. Recently troglitazone andciglitazone, agonists of PPARg have also been shownto inhibit the proliferation of many cancer cell linesincluding those of PCa [16]. However PPARg activatedtarget genes that mediate antiproliferative effectsremain largely unknown, and several lines of evidenc-es indicate that the present PPARg agonists realizetheir inhibitory effect independently of PPARg activa-
tion [17]. For these reasons the therapeutic strategybased upon the activation of PPARg in tumors iscontroversial. VPA has been shown to be a potentactivator of PPARg [18] but the biological consequencesof this activation have not yet been described.
The aim of the present work was to elucidate in vitroand in vivo the molecular mechanisms underlying thephenotypic changes induced byVPA in PCa cells and toverify their compatibility with a clinical use of VPA asantitumoral drug. Our results indicate that VPA treat-ment induces amassiveNET in residualPCacells andanoverall apoptotic resistance through a late overexpres-sion of Bcl-2. The use of a specific PPARg antagonistwasable to revert some differentiative effects withoutaffecting the VPA cytostatic action. For these reasonswe propose herein the combined use of VPA/PPARgantagonist, once further validated, for a new andeffective therapeutic approach in PCa treatment.
MATERIALSANDMETHODS
Cell Culture and Reagents
The PC3 human prostate cancer cell line wasoriginally obtained by ATCC (Rockville, MD) andwas maintained in DMEM supplemented with 10%fetal bovine serum, glutamine and penicillin-strepto-mycin (Sigma, St. Louis, MI). Primary antibodies anti-NSE was purchased from Cymbus biotechnology(Eastleigh, Hampshire, UK), anti-b-tubIII was pur-chased from Promega (Madison, WI), anti-Bcl-2 (Ab-1)and anti-actin (Ab-1)were purchased fromCalbiochem(Darmstadt, Germany), anti-PPARg was purchasedfrom ABR (Golden, CO). GW9662 is generous gifts ofDr. Timothy M. Willson, GlaxoSmithKline (DiscoveryResearch, Research Triangle Park, NC).
ProliferationAssay
A classical growth curve analysis was used todetermine the inhibition activity of antitumoral drugs.Cells were cultured in DMEM plus 10% FCS, thentrypsinized, counted using a hemocytometer, andadjusted to 1� 104 cells per 50-mm diameter Petri dishin serum-free DMEM. After 24 hr, three dishes weresacrificed for cell counting (time 0) to measure thebaseline cell number. The remaining dishes receivedfresh medium containing drug at the indicated con-centrations. For cell counting, cells were trypsinized,resuspended in 20 ml saline, and counted in a CoulterCounter (LabRecyclers, Gaithersburg, MD). Five inde-pendent counts were made from each dish.
Immunofluorescence
Cells grown on coverslips and treated atdifferent times (2� 104 cells/cm2) were fixed in 4%
The Prostate
Valproate andCell Differentiation 589

paraformaldehyde in PBS for 10 min at room temper-ature and permeabilized in PBS containing 0.1% TritonX-100 for 5 min at RT. Cells were then incubated withprimary antibodies, all diluted in PBS containing 3%BSA according to manufacture’s indications, for 1 hr atRT. After extensive washings with PBS, cells weretreated with fluorescein-labeled IgG secondary anti-body (1:100 in PBS containing 3% BSA) for 30 min atroom temperature. After extensive washings, cellswere mounted with Vectashield mounting mediumand photographed at fluorescence microscope (AXIO-PHOT, Zeiss).
Animals andXenograft
Male CD1 nude mice (Charles River, Milan, Italy)were maintained under the guidelines established byour Institution (University of L’Aquila, Medical Schooland Science andTechnology School BoardRegulations,complying with the Italian government regulationn.116 January 27, 1992 for the use of laboratoryanimals). Before any invasivemanipulation, mice wereanesthesized with a mixture of ketamine (25 mg/ml)/xylazine (5 mg/ml). Xenografts were obtained byinjecting s.c. 1� 106 PC3 cells in 100 ml of 12 mg/mlMatrigel (Becton Dickinson, Franklin Lakes, NJ).Sodium valproate (Sigma) was resuspended in phys-iological solution and administered per os by oralgavage. Tumor growth was monitored daily bymeasuring the average tumor diameter. The volumeof the tumor was expressed in mm3 according to theformula 4/3pr3. At the end point xenografts wereexcised, weighted and processed for histologicalanalysis.
Immunohistochemistry
Subcutaneous tumors excised frommouse were fixedin 4% formaldehyde in 0.1 M phosphate buffer,pH 7.2 and embedded in paraffin. Slide-mounted tissuesections (4-mm thick) were deparaffinized in xylene andhydrated serially in 100%, 95%, and 80% ethanol.Endogenous peroxidases were quenced in 3% H2O2 inPBS for 1 hr and then slides were incubated withantihuman primary antibody for 1 hr at room temper-ature. Sections were washed three times in PBS andantibody binding was revealed using the Sigma fast 3,30-diaminobenzidine tablet set (Sigma, St. Louis, MO).
Western Blot
Total cell lysates were obtained resuspending thecells in buffer containing 1% Triton, 0.1% SDS, 2 mMCaCl2, 100 mg/ml phenylmethyl sulfonyl fluoride.Protein content was determined using the ProteinAssayKit 2 (Bio-Rad Laboratory,Hercules, CA). Eighty
microgram of proteins were electrophoresed in 10%SDS-polyacrylamide gel and then electrotransferred tonitrocellulosemembrane (Schleicher & Schuell, Dassel,Germany), which was then blocked overnight with10 mM Tris–HCl, pH 8.0, 150 mMNaCl, 0.05% Tween-20 (TBS-T) containing 10% non-fat dry milk. Themembrane was then incubated with 1 mg/ml ofprimary antibody in TBS-T and with specific horse-radish peroxidase-conjugated secondary antibodies inTBS-T. Protein bands were visualized using a chem-iluminescent detection system (Amersham Bioscien-ces, Piscataway, NJ).
FluorescenceAnalysis byFluorescence-ActivatedCell Sorting
In order to analyze apoptosis and cell cycle theadherent cells were trypsinized, pooled with theculture supernatant containing the apoptotic cellsalready detached from the dish and centrifuged. Cells(1� 106) were washed in PBS and fixed for 30 min bythe addition of 1 ml 70% ethanol. After 30 min, the cellswere pelleted by centrifugation (720g; 5 min), andrinsed in 1ml ofDNAstaining solution (PBS containing200 mg/ml RNase A, 20 mg/ml propidium iodide and0.1% Triton X-100) and stained by incubation at roomtemperature for 60 min. All cells were then measuredon a FACScan flow cytometer with an argon laser at488 nm for excitation and analyzed using Cell Questsoftware. All flow-cytometric measurements weremade using the same instrument settings, and at least10,000 cells were measured in each sample. Apoptoticcells were detected by a quantifiable peak in sub-G1phase corresponding to the red fluorescent lightemitted by sub-diploid nuclei of cells, and the resultswere expressed as the percentage of death by apoptosisinduced by aparticular treatment. Quantification ofNEmarkers was performed by standard flow cytometryanalysis. Briefly cells were trypsinized, washed insaline buffer and fixed for 10 min at 48C with a 4%buffered formalin solution. Then cells were washedand resuspended at 1� 106 cells/ml in buffer contain-ing 0.1% Triton-X. Cells were incubated with 1 mg/mlprimary antibody at 48C for 1 hr and then withfluorescein isothiocyanate-conjugated secondary anti-body at 48C for 1 hr. Expression levels were evaluatedas fluorescence intensity in the presence of the relevantantibody and calculated as increment respect to back-ground value obtained by analyzing cells incubatedwith fluoreiscinated secondary antibody only (auto-fluorescence).
Statistical Analysis
Results are expressed as means� SD for at leastthree distinct experiments. Demonstration of significant
The Prostate
590 Angelucci et al.
differences among means were performed by Student’st-analysis considering 0.01 the threshold value of P. Allstatistical analyses were performed using Kaleidagraph3.6 (Synergy Software, Reading, PA).
RESULTS
The Effectof VPAontheGrowthof PC3Cells
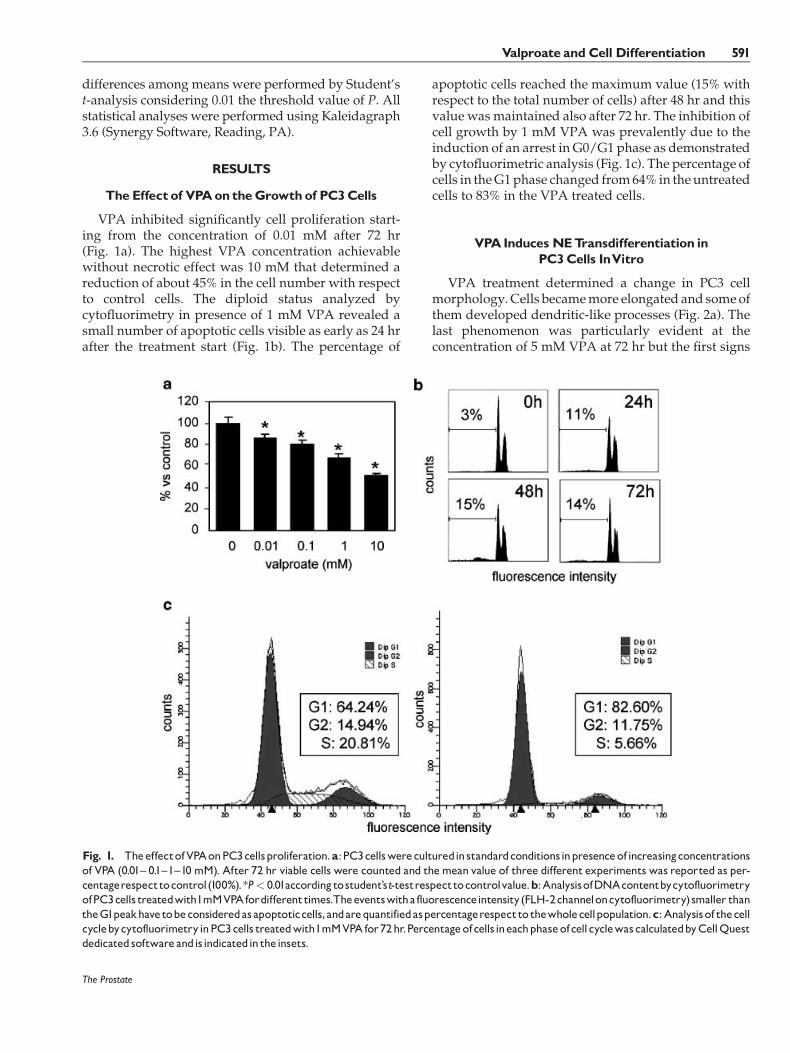
VPA inhibited significantly cell proliferation start-ing from the concentration of 0.01 mM after 72 hr(Fig. 1a). The highest VPA concentration achievablewithout necrotic effect was 10 mM that determined areduction of about 45% in the cell number with respectto control cells. The diploid status analyzed bycytofluorimetry in presence of 1 mM VPA revealed asmall number of apoptotic cells visible as early as 24 hrafter the treatment start (Fig. 1b). The percentage of
apoptotic cells reached the maximum value (15% withrespect to the total number of cells) after 48 hr and thisvalue was maintained also after 72 hr. The inhibition ofcell growth by 1 mM VPA was prevalently due to theinduction of an arrest in G0/G1 phase as demonstratedby cytofluorimetric analysis (Fig. 1c). The percentage ofcells in theG1phase changed from64% in theuntreatedcells to 83% in the VPA treated cells.
VPAInducesNETransdifferentiation inPC3Cells InVitro
VPA treatment determined a change in PC3 cellmorphology.Cells becamemore elongated and someofthem developed dendritic-like processes (Fig. 2a). Thelast phenomenon was particularly evident at theconcentration of 5 mM VPA at 72 hr but the first signs
The Prostate
Fig. 1. TheeffectofVPAonPC3cellsproliferation.a:PC3cellswereculturedinstandardconditionsinpresenceofincreasingconcentrationsof VPA (0.01^ 0.1^1^10 mM). After 72 hr viable cells were counted and themean value of three different experiments was reported as per-centagerespecttocontrol(100%).*P< 0.01accordingtostudent’st-testrespecttocontrolvalue.b:AnalysisofDNAcontentbycytofluorimetryofPC3cells treatedwith1mMVPAfordifferenttimes.Theeventswitha fluorescenceintensity(FLH-2channeloncytofluorimetry)smaller thantheG1peakhave tobeconsideredasapoptoticcells,andarequantifiedaspercentagerespectto thewholecellpopulation.c:Analysisof thecellcyclebycytofluorimetryinPC3cells treatedwith1mMVPAfor72hr.Percentageofcells ineachphaseofcellcyclewascalculatedbyCellQuestdedicatedsoftwareandisindicatedin theinsets.
Valproate andCell Differentiation 591
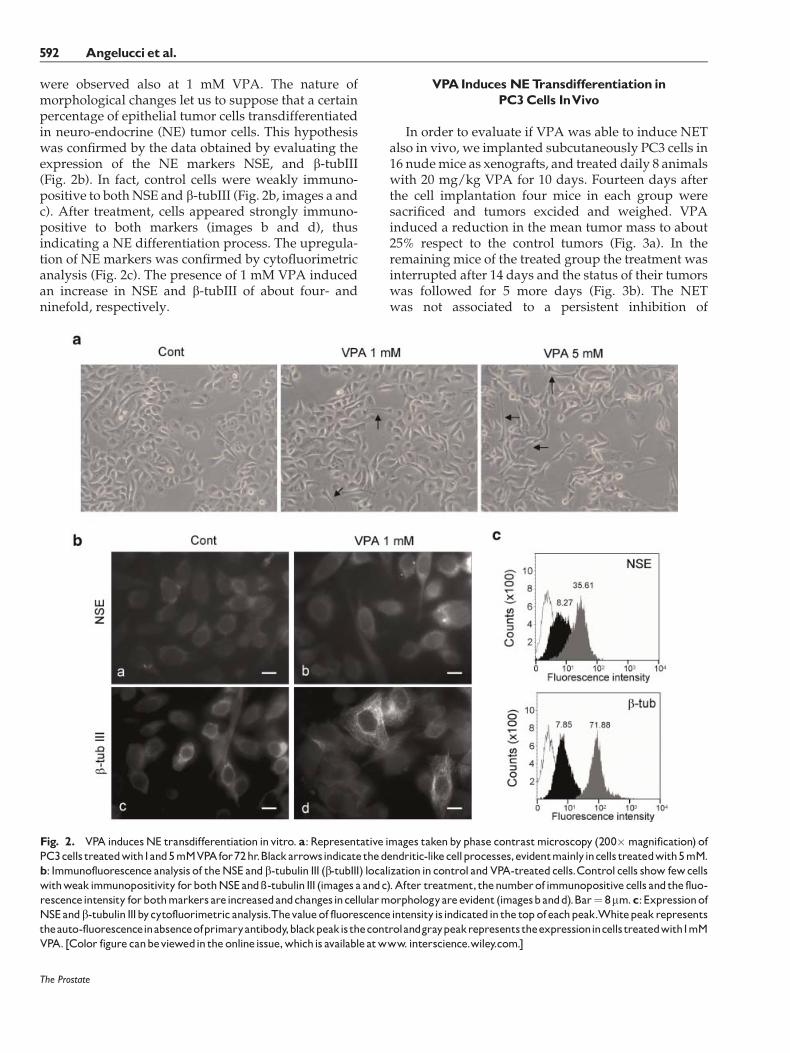
were observed also at 1 mM VPA. The nature ofmorphological changes let us to suppose that a certainpercentage of epithelial tumor cells transdifferentiatedin neuro-endocrine (NE) tumor cells. This hypothesiswas confirmed by the data obtained by evaluating theexpression of the NE markers NSE, and b-tubIII(Fig. 2b). In fact, control cells were weakly immuno-positive to bothNSE and b-tubIII (Fig. 2b, images a andc). After treatment, cells appeared strongly immuno-positive to both markers (images b and d), thusindicating a NE differentiation process. The upregula-tion of NE markers was confirmed by cytofluorimetricanalysis (Fig. 2c). The presence of 1 mM VPA inducedan increase in NSE and b-tubIII of about four- andninefold, respectively.
VPAInducesNETransdifferentiation inPC3Cells InVivo
In order to evaluate if VPA was able to induce NETalso in vivo, we implanted subcutaneously PC3 cells in16 nudemice as xenografts, and treated daily 8 animalswith 20 mg/kg VPA for 10 days. Fourteen days afterthe cell implantation four mice in each group weresacrificed and tumors excided and weighed. VPAinduced a reduction in the mean tumor mass to about25% respect to the control tumors (Fig. 3a). In theremaining mice of the treated group the treatment wasinterrupted after 14 days and the status of their tumorswas followed for 5 more days (Fig. 3b). The NETwas not associated to a persistent inhibition of
The Prostate
Fig. 2. VPA inducesNE transdifferentiation invitro. a:Representative images takenbyphase contrastmicroscopy (200�magnification) ofPC3cells treatedwith1and5mMVPAfor72hr.Blackarrowsindicate thedendritic-likecellprocesses,evidentmainlyincells treatedwith5mM.b: Immunofluorescence analysis of theNSE andb-tubulin III (b-tubIII) localization incontrol andVPA-treatedcells.Control cells show fewcellswithweakimmunopositivity forbothNSEand�-tubulin III (images a andc).After treatment, thenumberof immunopositive cells andthe fluo-rescenceintensity forbothmarkers areincreasedandchangesincellularmorphologyareevident (imagesbandd).Bar¼ 8mm.c:ExpressionofNSEandb-tubulin IIIbycytofluorimetric analysis.Thevalueof fluorescenceintensityisindicatedin the top ofeachpeak.Whitepeakrepresentstheauto-fluorescenceinabsenceofprimaryantibody,blackpeakisthecontrolandgraypeakrepresentstheexpressionincells treatedwith1mMVPA. [Color figurecanbeviewedin theonlineissue,whichis availableatwww. interscience.wiley.com.]
592 Angelucci et al.
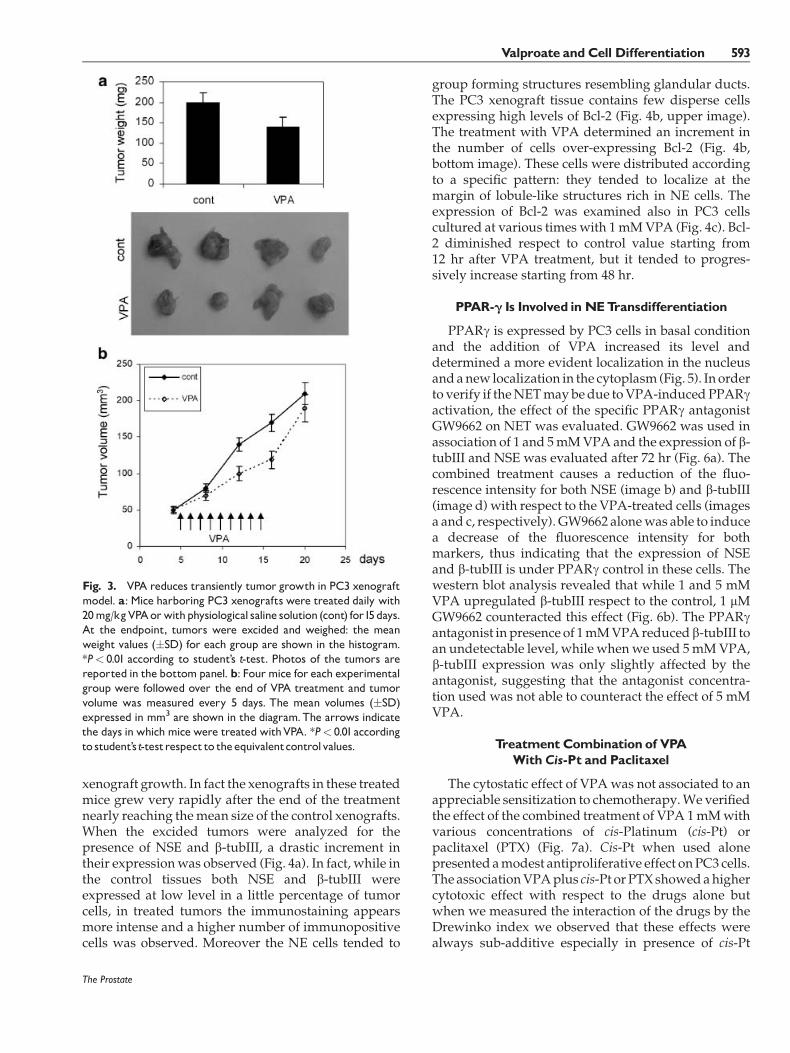
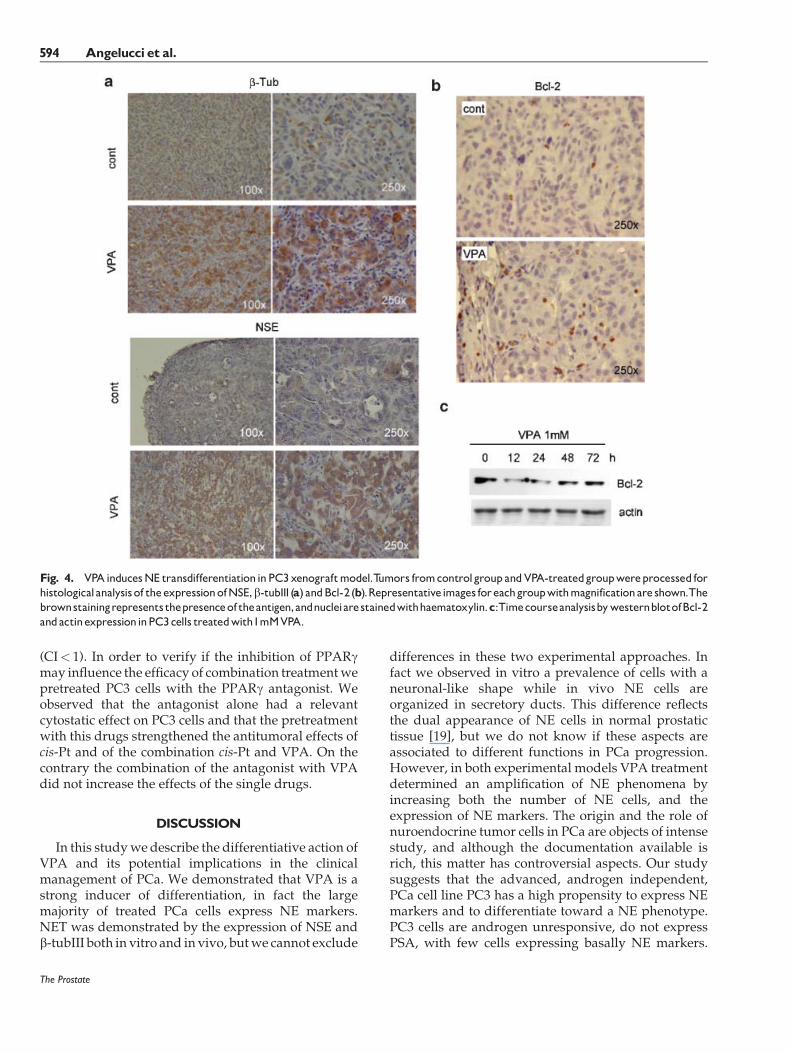
xenograft growth. In fact the xenografts in these treatedmice grew very rapidly after the end of the treatmentnearly reaching themean size of the control xenografts.When the excided tumors were analyzed for thepresence of NSE and b-tubIII, a drastic increment intheir expressionwas observed (Fig. 4a). In fact, while inthe control tissues both NSE and b-tubIII wereexpressed at low level in a little percentage of tumorcells, in treated tumors the immunostaining appearsmore intense and a higher number of immunopositivecells was observed. Moreover the NE cells tended to
group forming structures resembling glandular ducts.The PC3 xenograft tissue contains few disperse cellsexpressing high levels of Bcl-2 (Fig. 4b, upper image).The treatment with VPA determined an increment inthe number of cells over-expressing Bcl-2 (Fig. 4b,bottom image). These cells were distributed accordingto a specific pattern: they tended to localize at themargin of lobule-like structures rich in NE cells. Theexpression of Bcl-2 was examined also in PC3 cellscultured at various times with 1mMVPA (Fig. 4c). Bcl-2 diminished respect to control value starting from12 hr after VPA treatment, but it tended to progres-sively increase starting from 48 hr.
PPAR-g Is Involved inNETransdifferentiation
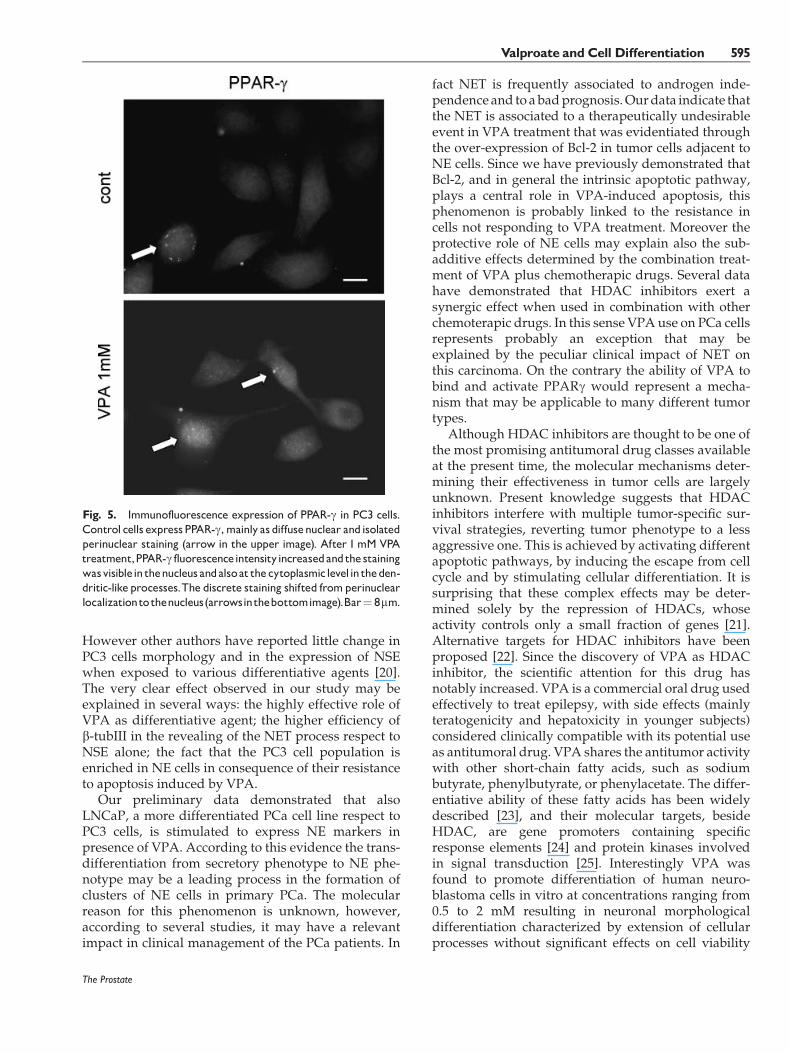
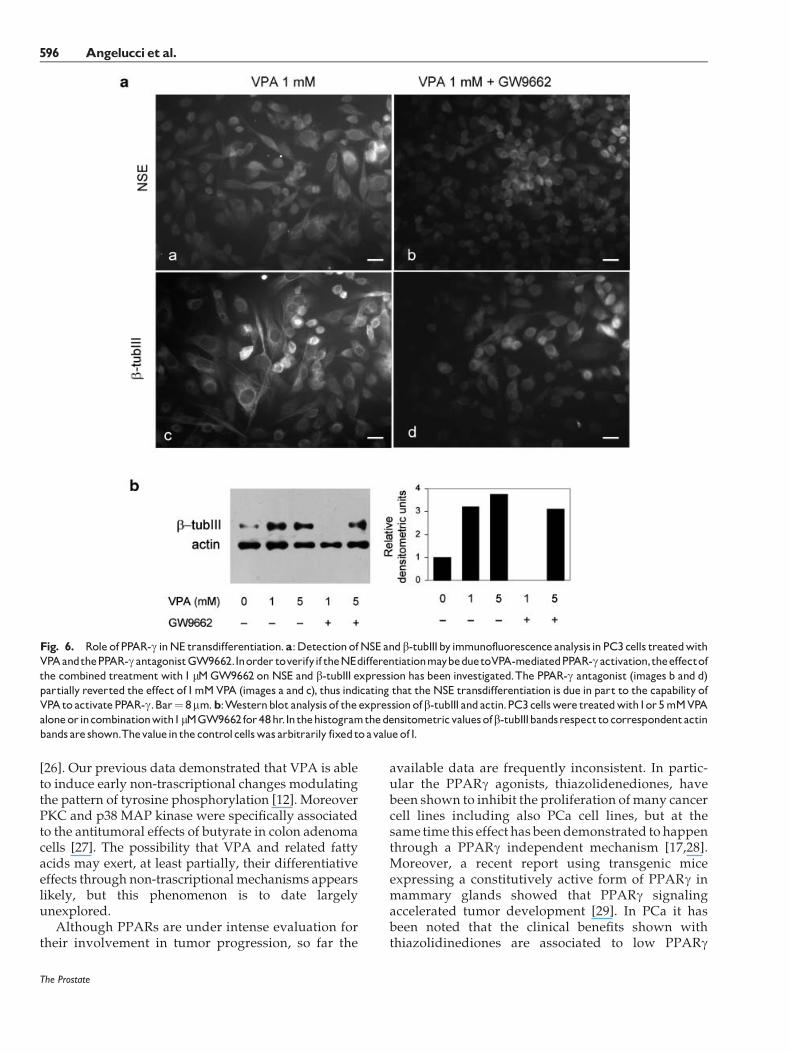
PPARg is expressed by PC3 cells in basal conditionand the addition of VPA increased its level anddetermined a more evident localization in the nucleusandanew localization in the cytoplasm (Fig. 5). In orderto verify if theNETmaybedue toVPA-induced PPARgactivation, the effect of the specific PPARg antagonistGW9662 on NET was evaluated. GW9662 was used inassociation of 1 and 5mMVPAand the expression of b-tubIII and NSE was evaluated after 72 hr (Fig. 6a). Thecombined treatment causes a reduction of the fluo-rescence intensity for both NSE (image b) and b-tubIII(image d) with respect to the VPA-treated cells (imagesa and c, respectively). GW9662 alonewas able to inducea decrease of the fluorescence intensity for bothmarkers, thus indicating that the expression of NSEand b-tubIII is under PPARg control in these cells. Thewestern blot analysis revealed that while 1 and 5 mMVPA upregulated b-tubIII respect to the control, 1 mMGW9662 counteracted this effect (Fig. 6b). The PPARgantagonist in presence of 1mMVPAreducedb-tubIII toan undetectable level, while whenwe used 5mMVPA,b-tubIII expression was only slightly affected by theantagonist, suggesting that the antagonist concentra-tion used was not able to counteract the effect of 5 mMVPA.
TreatmentCombination of VPAWithCis-Pt and Paclitaxel
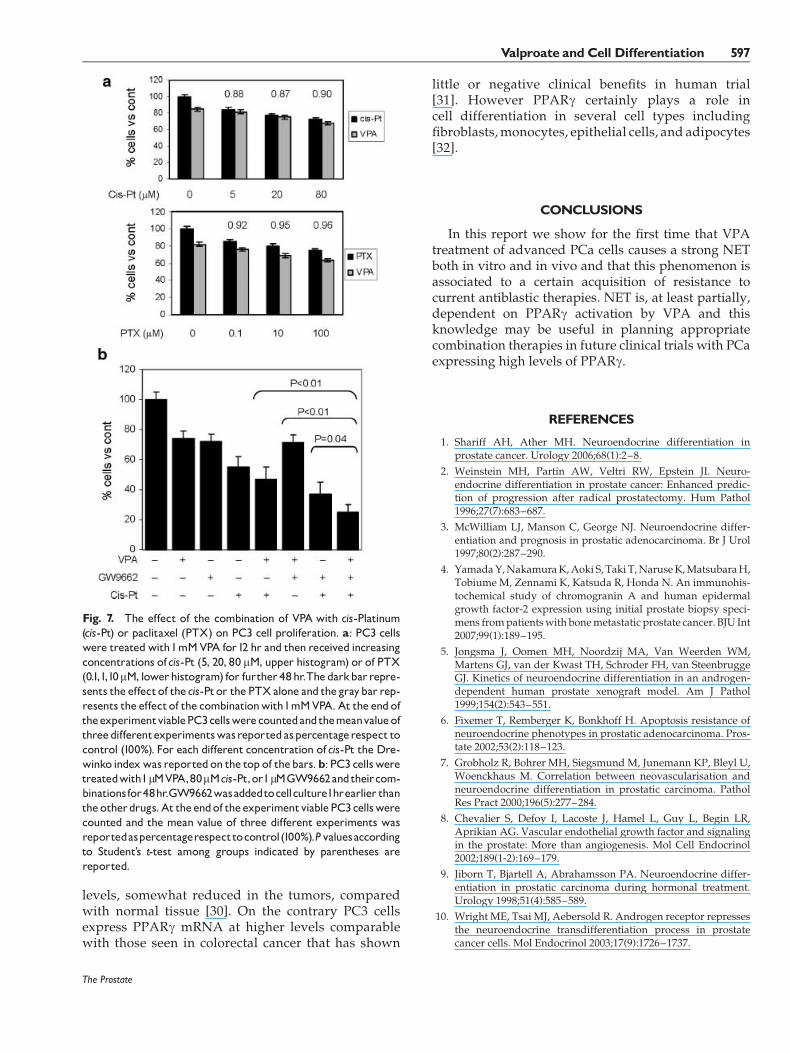
The cytostatic effect of VPAwas not associated to anappreciable sensitization to chemotherapy.We verifiedthe effect of the combined treatment of VPA 1mMwithvarious concentrations of cis-Platinum (cis-Pt) orpaclitaxel (PTX) (Fig. 7a). Cis-Pt when used alonepresented amodest antiproliferative effect onPC3 cells.The associationVPAplus cis-Pt or PTXshowedahighercytotoxic effect with respect to the drugs alone butwhen we measured the interaction of the drugs by theDrewinko index we observed that these effects werealways sub-additive especially in presence of cis-Pt
The Prostate
Fig. 3. VPA reduces transiently tumor growth in PC3 xenograftmodel. a: Mice harboring PC3 xenografts were treated daily with20mg/kgVPAorwithphysiological saline solution (cont) for15days.At the endpoint, tumors were excided and weighed: the meanweight values (�SD) for each group are shown in the histogram.*P< 0.01 according to student’s t-test. Photos of the tumors arereported in the bottompanel. b: Fourmice for each experimentalgroup were followed over the end of VPA treatment and tumorvolume was measured every 5 days. The mean volumes (�SD)expressed in mm3 are shown in the diagram.The arrows indicatethe days in which mice were treatedwithVPA. *P< 0.01accordingto student’st-testrespect to theequivalentcontrolvalues.
Valproate andCell Differentiation 593
(CI< 1). In order to verify if the inhibition of PPARgmay influence the efficacy of combination treatmentwepretreated PC3 cells with the PPARg antagonist. Weobserved that the antagonist alone had a relevantcytostatic effect on PC3 cells and that the pretreatmentwith this drugs strengthened the antitumoral effects ofcis-Pt and of the combination cis-Pt and VPA. On thecontrary the combination of the antagonist with VPAdid not increase the effects of the single drugs.
DISCUSSION
In this studywe describe the differentiative action ofVPA and its potential implications in the clinicalmanagement of PCa. We demonstrated that VPA is astrong inducer of differentiation, in fact the largemajority of treated PCa cells express NE markers.NET was demonstrated by the expression of NSE andb-tubIII both in vitro and in vivo, butwe cannot exclude
differences in these two experimental approaches. Infact we observed in vitro a prevalence of cells with aneuronal-like shape while in vivo NE cells areorganized in secretory ducts. This difference reflectsthe dual appearance of NE cells in normal prostatictissue [19], but we do not know if these aspects areassociated to different functions in PCa progression.However, in both experimental models VPA treatmentdetermined an amplification of NE phenomena byincreasing both the number of NE cells, and theexpression of NE markers. The origin and the role ofnuroendocrine tumor cells in PCa are objects of intensestudy, and although the documentation available isrich, this matter has controversial aspects. Our studysuggests that the advanced, androgen independent,PCa cell line PC3 has a high propensity to express NEmarkers and to differentiate toward a NE phenotype.PC3 cells are androgen unresponsive, do not expressPSA, with few cells expressing basally NE markers.
The Prostate
Fig. 4. VPAinducesNE transdifferentiationinPC3xenograftmodel.Tumors fromcontrolgroupandVPA-treatedgroupwereprocessed forhistologicalanalysisof theexpressionofNSE,b-tubIII (a) andBcl-2 (b).Representativeimages foreachgroupwithmagnificationare shown.Thebrownstainingrepresents thepresenceof theantigen,andnucleiarestainedwithhaematoxylin.c:TimecourseanalysisbywesternblotofBcl-2andactinexpressioninPC3cells treatedwith1mMVPA.
594 Angelucci et al.
However other authors have reported little change inPC3 cells morphology and in the expression of NSEwhen exposed to various differentiative agents [20].The very clear effect observed in our study may beexplained in several ways: the highly effective role ofVPA as differentiative agent; the higher efficiency ofb-tubIII in the revealing of the NET process respect toNSE alone; the fact that the PC3 cell population isenriched in NE cells in consequence of their resistanceto apoptosis induced by VPA.
Our preliminary data demonstrated that alsoLNCaP, a more differentiated PCa cell line respect toPC3 cells, is stimulated to express NE markers inpresence of VPA. According to this evidence the trans-differentiation from secretory phenotype to NE phe-notype may be a leading process in the formation ofclusters of NE cells in primary PCa. The molecularreason for this phenomenon is unknown, however,according to several studies, it may have a relevantimpact in clinical management of the PCa patients. In
fact NET is frequently associated to androgen inde-pendence and to abadprognosis.Ourdata indicate thatthe NET is associated to a therapeutically undesirableevent in VPA treatment that was evidentiated throughthe over-expression of Bcl-2 in tumor cells adjacent toNE cells. Since we have previously demonstrated thatBcl-2, and in general the intrinsic apoptotic pathway,plays a central role in VPA-induced apoptosis, thisphenomenon is probably linked to the resistance incells not responding to VPA treatment. Moreover theprotective role of NE cells may explain also the sub-additive effects determined by the combination treat-ment of VPA plus chemotherapic drugs. Several datahave demonstrated that HDAC inhibitors exert asynergic effect when used in combination with otherchemoterapic drugs. In this sense VPA use on PCa cellsrepresents probably an exception that may beexplained by the peculiar clinical impact of NET onthis carcinoma. On the contrary the ability of VPA tobind and activate PPARg would represent a mecha-nism that may be applicable to many different tumortypes.
Although HDAC inhibitors are thought to be one ofthe most promising antitumoral drug classes availableat the present time, the molecular mechanisms deter-mining their effectiveness in tumor cells are largelyunknown. Present knowledge suggests that HDACinhibitors interfere with multiple tumor-specific sur-vival strategies, reverting tumor phenotype to a lessaggressive one. This is achieved by activating differentapoptotic pathways, by inducing the escape from cellcycle and by stimulating cellular differentiation. It issurprising that these complex effects may be deter-mined solely by the repression of HDACs, whoseactivity controls only a small fraction of genes [21].Alternative targets for HDAC inhibitors have beenproposed [22]. Since the discovery of VPA as HDACinhibitor, the scientific attention for this drug hasnotably increased. VPA is a commercial oral drug usedeffectively to treat epilepsy, with side effects (mainlyteratogenicity and hepatoxicity in younger subjects)considered clinically compatible with its potential useas antitumoral drug. VPA shares the antitumor activitywith other short-chain fatty acids, such as sodiumbutyrate, phenylbutyrate, or phenylacetate. The differ-entiative ability of these fatty acids has been widelydescribed [23], and their molecular targets, besideHDAC, are gene promoters containing specificresponse elements [24] and protein kinases involvedin signal transduction [25]. Interestingly VPA wasfound to promote differentiation of human neuro-blastoma cells in vitro at concentrations ranging from0.5 to 2 mM resulting in neuronal morphologicaldifferentiation characterized by extension of cellularprocesses without significant effects on cell viability
The Prostate
Fig. 5. Immunofluorescence expression of PPAR-g in PC3 cells.Control cells expressPPAR-g,mainly as diffusenuclear andisolatedperinuclear staining (arrow in the upper image). After 1mM VPAtreatment,PPAR-g fluorescenceintensityincreasedandthestainingwasvisiblein thenucleus andalsoat thecytoplasmiclevelintheden-dritic-like processes.The discrete staining shifted fromperinuclearlocalizationtothenucleus(arrowsinthebottomimage).Bar¼ 8mm.
Valproate andCell Differentiation 595
[26]. Our previous data demonstrated that VPA is ableto induce early non-trascriptional changes modulatingthe pattern of tyrosine phosphorylation [12]. MoreoverPKC and p38 MAP kinase were specifically associatedto the antitumoral effects of butyrate in colon adenomacells [27]. The possibility that VPA and related fattyacids may exert, at least partially, their differentiativeeffects through non-trascriptional mechanisms appearslikely, but this phenomenon is to date largelyunexplored.
Although PPARs are under intense evaluation fortheir involvement in tumor progression, so far the
available data are frequently inconsistent. In partic-ular the PPARg agonists, thiazolidenediones, havebeen shown to inhibit the proliferation ofmany cancercell lines including also PCa cell lines, but at thesame time this effect has beendemonstrated to happenthrough a PPARg independent mechanism [17,28].Moreover, a recent report using transgenic miceexpressing a constitutively active form of PPARg inmammary glands showed that PPARg signalingaccelerated tumor development [29]. In PCa it hasbeen noted that the clinical benefits shown withthiazolidinediones are associated to low PPARg
The Prostate
Fig. 6. Role ofPPAR-g inNE transdifferentiation.a:DetectionofNSE andb-tubIIIbyimmunofluorescenceanalysis inPC3cells treatedwithVPAandthePPAR-gantagonistGW9662.Inorder toverifyif theNEdifferentiationmaybeduetoVPA-mediatedPPAR-gactivation,theeffectofthe combined treatmentwith1 mMGW9662 on NSE and b-tubIII expression has been investigated.The PPAR-g antagonist (images b and d)partially reverted the effect of1mMVPA (images a and c), thus indicating that theNSE transdifferentiation is due in part to the capability ofVPAto activatePPAR-g.Bar¼ 8mm.b:Westernblot analysis of theexpressionofb-tubIII andactin.PC3cellswere treatedwith1or5mMVPAaloneorincombinationwith1mMGW9662for48hr.Inthehistogramthedensitometricvaluesofb-tubIIIbandsrespecttocorrespondentactinbandsare shown.Thevaluein thecontrolcellswas arbitrarily fixedtoavalueof1.
596 Angelucci et al.
levels, somewhat reduced in the tumors, comparedwith normal tissue [30]. On the contrary PC3 cellsexpress PPARg mRNA at higher levels comparablewith those seen in colorectal cancer that has shown
little or negative clinical benefits in human trial[31]. However PPARg certainly plays a role incell differentiation in several cell types includingfibroblasts,monocytes, epithelial cells, and adipocytes[32].
CONCLUSIONS
In this report we show for the first time that VPAtreatment of advanced PCa cells causes a strong NETboth in vitro and in vivo and that this phenomenon isassociated to a certain acquisition of resistance tocurrent antiblastic therapies. NET is, at least partially,dependent on PPARg activation by VPA and thisknowledge may be useful in planning appropriatecombination therapies in future clinical trials with PCaexpressing high levels of PPARg.
REFERENCES
1. Shariff AH, Ather MH. Neuroendocrine differentiation inprostate cancer. Urology 2006;68(1):2–8.
2. Weinstein MH, Partin AW, Veltri RW, Epstein JI. Neuro-endocrine differentiation in prostate cancer: Enhanced predic-tion of progression after radical prostatectomy. Hum Pathol1996;27(7):683–687.
3. McWilliam LJ, Manson C, George NJ. Neuroendocrine differ-entiation and prognosis in prostatic adenocarcinoma. Br J Urol1997;80(2):287–290.
4. YamadaY,NakamuraK,Aoki S, Taki T,NaruseK,MatsubaraH,Tobiume M, Zennami K, Katsuda R, Honda N. An immunohis-tochemical study of chromogranin A and human epidermalgrowth factor-2 expression using initial prostate biopsy speci-mens frompatientswith bonemetastatic prostate cancer. BJU Int2007;99(1):189–195.
5. Jongsma J, Oomen MH, Noordzij MA, Van Weerden WM,Martens GJ, van der Kwast TH, Schroder FH, van SteenbruggeGJ. Kinetics of neuroendocrine differentiation in an androgen-dependent human prostate xenograft model. Am J Pathol1999;154(2):543–551.
6. Fixemer T, Remberger K, Bonkhoff H. Apoptosis resistance ofneuroendocrine phenotypes in prostatic adenocarcinoma. Pros-tate 2002;53(2):118–123.
7. Grobholz R, Bohrer MH, Siegsmund M, Junemann KP, Bleyl U,Woenckhaus M. Correlation between neovascularisation andneuroendocrine differentiation in prostatic carcinoma. PatholRes Pract 2000;196(5):277–284.
8. Chevalier S, Defoy I, Lacoste J, Hamel L, Guy L, Begin LR,Aprikian AG. Vascular endothelial growth factor and signalingin the prostate: More than angiogenesis. Mol Cell Endocrinol2002;189(1-2):169–179.
9. Jiborn T, Bjartell A, Abrahamsson PA. Neuroendocrine differ-entiation in prostatic carcinoma during hormonal treatment.Urology 1998;51(4):585–589.
10. Wright ME, Tsai MJ, Aebersold R. Androgen receptor repressesthe neuroendocrine transdifferentiation process in prostatecancer cells. Mol Endocrinol 2003;17(9):1726–1737.
The Prostate
Fig. 7. The effect of the combination of VPA with cis-Platinum(cis-Pt) or paclitaxel (PTX) on PC3 cell proliferation. a: PC3 cellswere treatedwith1mMVPA for12 hr and then received increasingconcentrations ofcis-Pt (5, 20, 80 mM, upper histogram) or of PTX(0.1,1,10mM, lowerhistogram) for further48hr.Thedarkbarrepre-sents the effectof thecis-Ptor the PTXalone and thegraybar rep-resents the effectof the combinationwith1mMVPA.At the endoftheexperimentviablePC3cellswerecountedandthemeanvalueofthreedifferentexperimentswasreportedaspercentagerespect tocontrol (100%). For each different concentration ofcis-Pt the Dre-winko indexwasreportedon the top of thebars.b: PC3 cellsweretreatedwith1mMVPA,80mMcis-Pt,or1mMGW9662andtheircom-binationsfor48hr.GW9662wasaddedtocellculture1hrearlier thantheotherdrugs.At theendof theexperimentviablePC3cellswerecounted and the mean value of three different experiments wasreportedaspercentagerespecttocontrol(100%).Pvaluesaccordingto Student’s t-test among groups indicated by parentheses arereported.
Valproate andCell Differentiation 597

11. Gurvich N, Tsygankova OM, Meinkoth JL, Klein PS. Histonedeacetylase is a target of valproic acid-mediated cellular differ-entiation. Cancer Res 2004;64(3):1079–1086.
12. Angelucci A, Valentini A, Millimaggi D, Gravina GL, Miano R,Dolo V, Vicentini C, Bologna M, Federici G, Bernardini S.Valproic acid induces apoptosis in prostate carcinoma cell linesby activation of multiple death pathways. Anticancer Drugs2006;17(10):1141–1150.
13. Xia Q, Sung J, Chowdhury W, Chen CL, Hoti N, Shabbeer S,Carducci M, Rodriguez R. Chronic administration of valproicacid inhibits prostate cancer cell growth in vitro and in vivo.Cancer Res 2006;66(14):7237–7244.
14. Cinatl J Jr, KotchetkovR, Blaheta R, Driever PH, Vogel JU, CinatlJ. Induction of differentiation and suppression of malignantphenotype of human neuroblastoma BE(2)-C cells by valproicacid: Enhancement by combination with interferon-alpha. IntJ Oncol 2002;20(1):97–106.
15. Kubota T, Koshizuka K, Williamson EA, Asou H, Said JW,Holden S, Miyoshi I, Koeffler HP. Ligand for peroxisomeproliferator-activated receptor gamma (troglitazone) has potentantitumor effect against humanprostate cancer both in vitro andin vivo. Cancer Res 1998;58(15):3344–3352.
16. Koeffler HP. Peroxisome proliferator-activated receptor gammaand cancers. Clin Cancer Res 2003;9(1):1–9.
17. Shiau CW, Yang CC, Kulp SK, Chen KF, Chen CS, Huang JW.Thiazolidenedionesmediate apoptosis in prostate cancer cells inpart through inhibition of Bcl-xL/Bcl-2 functions independentlyof PPARgamma. Cancer Res 2005;65(4):1561–1569.
18. Lampen A, Carlberg C, Nau H. Peroxisome proliferator-activated receptor delta is a specific sensor for teratogenicvalproic acid derivatives. Eur J Pharmacol 2001;431(1):25–33.
19. Vashchenko N, Abrahamsson PA. Neuroendocrine differentia-tion in prostate cancer: Implications for new treatment modal-ities. Eur Urol 2005;47(2):147–155.
20. Zelivianski S, VerniM,MooreC,KondrikovD, TaylorR, LinMF.Multipathways for transdifferentiation of human prostatecancer cells into neuroendocrine-like phenotype. BiochimBiophys Acta 2001;1539(1-2):28–43.
21. Van Lint C, Emiliani S, Verdin E. The expression of a smallfraction of cellular genes is changed in response to histonehyperacetylation. Gene Expr 1996;5(4-5):245–253.
22. Minucci S, Pelicci PG. Histone deacetylase inhibitors and thepromise of epigenetic (andmore) treatments for cancer. Nat RevCancer 2006;6(1):38–51.
23. Newmark HL, Lupton JR, Young CW. Butyrate as a differ-entiating agent: Pharmacokinetics, analogues and current status.Cancer Lett 1994;78(1-3):1–5.
24. Yang J, Kawai Y, Hanson RW, Arinze IJ. Sodium butyrateinduces transcription from the G alpha(i2) gene promoterthrough multiple Sp1 sites in the promoter and by activatingthe MEK-ERK signal transduction pathway. J Biol Chem 2001;276(28):25742–25752.
25. Orchel A, Dzierzewicz Z, Parfiniewicz B, Weglarz L, Wilczok T.Butyrate-induceddifferentiation of colon cancer cells is PKCandJNK dependent. Dig Dis Sci 2005;50(3):490–498.
26. Cinatl J Jr, Cinatl J, ScholzM, Driever PH, Henrich D, KabickovaH, Vogel JU, Doerr HW, Kornhuber B. Antitumor activity ofsodium valproate in cultures of human neuroblastoma cells.Anticancer Drugs 1996;7(7):766–773.
27. McMillan L, Butcher SK, Pongracz J, Lord JM. Opposing effectsof butyrate and bile acids on apoptosis of human colon adenomacells: Differential activation of PKC and MAP kinases. BrJ Cancer 2003;88(5):748–753.
28. Weber SM, Scarim AL, Corbett JA. PPARgamma is not requiredfor the inhibitory actions of PGJ2 on cytokine signaling inpancreatic beta-cells. Am J Physiol Endocrinol Metab2004;286(3):E329–E336.
29. Saez E, Rosenfeld J, Livolsi A, Olson P, Lombardo E, Nelson M,Banayo E, Cardiff RD, Izpisua-Belmonte JC, Evans RM. PPARgamma signaling exacerbates mammary gland tumor develop-ment. Genes Dev 2004;18(5):528–540.
30. Mueller E, Smith M, Sarraf P, Kroll T, Aiyer A, Kaufman DS,Oh W, Demetri G, Figg WD, Zhou XP, Eng C, Spiegelman BM,Kantoff PW. Effects of ligand activation of peroxisome prolifer-ator-activated receptor gamma in human prostate cancer. ProcNatl Acad Sci USA 2000;97(20):10990–10995.
31. Sertznig P, SeifertM, TilgenW,Reichrath J. Present concepts andfuture outlook: Function of peroxisome proliferator-activatedreceptors (PPARs) for pathogenesis, progression, and therapy ofcancer. J Cell Physiol 2007;212(1):1–12.
32. Mangelsdorf DJ, Evans RM. The RXR heterodimers and orphanreceptors. Cell 1995;83(6):841–850.
The Prostate
598 Angelucci et al.
Related Documents











