Neurochemical, neuroautonomic and neuropharmacological acute effects of sibutramine in healthy subjects Fuad Lechin a,b,c,d, * , Bertha van der Dijs b,c,d , Gerardo Hernandez a,c,d , Beatriz Orozco c,d , Simon Rodriguez c,d , Scarlet Baez b,c a Department of Neurophysiology, Instituto de Medicina Experimental, Faculty of Medicine, Universidad Central de Venezuela, Caracas, Apartado 80.983, Caracas 1080-A, Venezuela b Department of Neurochemistry,, Instituto de Medicina Experimental, Faculty of Medicine, Universidad Central de Venezuela, Caracas, Venezuela c Department of Neuropharmacology, Instituto de Medicina Experimental, Faculty of Medicine, Universidad Central de Venezuela, Caracas, Venezuela d Department of Neuroimmunology, Instituto de Medicina Experimental, Faculty of Medicine, Universidad Central de Venezuela, Caracas, Venezuela Received 20 July 2005; accepted 27 September 2005 Available online 2 November 2005 Abstract Sibutramine is a neuropharmacological drug that exerts central (CNS) and peripheral effects including noradrenaline (NA), and serotonin (5- HT) uptake inhibition. In addition, the drug is able to induce release from DA axons. We measured levels of circulating neurotransmitters in 20 healthy subjects during supine-resting (fasting) state before and after 15 mg of oral sibutramine. Systolic blood pressure (SBP), diastolic blood pressure (DBP), and heart rate (HR) were also monitored. Sibutramine triggered sustained and progressive increase of NA, NA/Ad ratio and DBP. Slight increases of DAwere also registered between the 60 and 240 min periods. The rise in DA tended to fade progressively, reaching basal level at 360 min period. Diastolic blood pressure, but neither SBP nor HR, showed significant increases that correlated positively with NA/Ad ratios. Slight but significant negative correlation was also found between DBP and DA. This correlation tended to fade throughout the trial to show no significance at the 360 min period. Although neither plasma serotonin (f-5HT) nor platelet serotonin (p-5HT) values showed significant variation throughout the trial, the f-5HT/p-5HT ratio showed significant decrease throughout. Significant negative correlation was found between f-5HT/p- 5HT ratio and NA/Ad ratio. Our results indicate that sibutramine stimulates neural sympathetic activity but not adrenal sympathetic activity in healthy individuals. Further, sibutramine lowers parasympathetic activity. The moderate rise in diastolic blood pressure triggered by sibutramine would be associated with CNS-NA enhancement plus parasympathetic inhibition. # 2005 Elsevier Inc. All rights reserved. Keywords: Sibutramine; Plasma noradrenaline; Plasma adrenaline; Plasma dopamine; Blood serotonin; Diastolic blood pressure; Neural sympathetic activity 1. Introduction Sibutramine is a neuropharmacological drug used as an anorexigenic agent and as such it is widely used to treat obesity, insulin resistance and polycystic ovary syndrome (PCOS) (Abolfathi et al., 2004; de Simone et al., 2005; Gaciong and Placha, 2005; Jordan et al., 2005; Karabacak et al., 2004; McMahon et al., 2000). Although it is considered a noradrena- line (NA) + serotonin (5HT) re-uptake inhibitor (Fukagawa and Sakata, 2001; Invernizi and Garattini, 2004), the drug also induces release from DA and 5HT axons (Ukai et al., 2004). Because sibutramine is closely related to amphetamines, it is viewed as a potentially harmful drug able to trigger adverse cardiovascular and neuropsychiatric disorders. The fact that its prolonged administration has provoked enhancement of central DA activity in rodents has been cited as proof that the drug behaves as a DA re-uptake inhibitor (Nakagawa et al., 2001). Although the half-life for sibutramine, M1 and M2 metabolites ranged between 8 and 18 h under fasting conditions, as in our protocol, it has been demonstrated that feeding is able to increase the half-life of the drug (Abolfathi et al., 2004). NeuroToxicology 27 (2006) 184–191 * Corresponding author. Tel.: + 58 212 961 1048; fax: +58 212 961 0172. E-mail address: fl[email protected] (F. Lechin). 0161-813X/$ – see front matter # 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.neuro.2005.09.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neurochemical, neuroautonomic and neuropharmacological
acute effects of sibutramine in healthy subjects
Fuad Lechin a,b,c,d,*, Bertha van der Dijs b,c,d, Gerardo Hernandez a,c,d,Beatriz Orozco c,d, Simon Rodriguez c,d, Scarlet Baez b,c
aDepartment of Neurophysiology, Instituto de Medicina Experimental, Faculty of Medicine, Universidad Central de Venezuela,
Caracas, Apartado 80.983, Caracas 1080-A, VenezuelabDepartment of Neurochemistry,, Instituto de Medicina Experimental, Faculty of Medicine,
Universidad Central de Venezuela, Caracas, VenezuelacDepartment of Neuropharmacology, Instituto de Medicina Experimental, Faculty of Medicine,
Universidad Central de Venezuela, Caracas, VenezueladDepartment of Neuroimmunology, Instituto de Medicina Experimental, Faculty of Medicine,
Universidad Central de Venezuela, Caracas, Venezuela
Received 20 July 2005; accepted 27 September 2005
Available online 2 November 2005
Abstract
Sibutramine is a neuropharmacological drug that exerts central (CNS) and peripheral effects including noradrenaline (NA), and serotonin (5-
HT) uptake inhibition. In addition, the drug is able to induce release from DA axons. We measured levels of circulating neurotransmitters in 20
healthy subjects during supine-resting (fasting) state before and after 15 mg of oral sibutramine. Systolic blood pressure (SBP), diastolic blood
pressure (DBP), and heart rate (HR) were also monitored. Sibutramine triggered sustained and progressive increase of NA, NA/Ad ratio and DBP.
Slight increases of DAwere also registered between the 60 and 240 min periods. The rise in DA tended to fade progressively, reaching basal level at
360 min period. Diastolic blood pressure, but neither SBP nor HR, showed significant increases that correlated positively with NA/Ad ratios. Slight
but significant negative correlation was also found between DBP and DA. This correlation tended to fade throughout the trial to show no
significance at the 360 min period. Although neither plasma serotonin (f-5HT) nor platelet serotonin (p-5HT) values showed significant variation
throughout the trial, the f-5HT/p-5HT ratio showed significant decrease throughout. Significant negative correlation was found between f-5HT/p-
5HT ratio and NA/Ad ratio. Our results indicate that sibutramine stimulates neural sympathetic activity but not adrenal sympathetic activity in
healthy individuals. Further, sibutramine lowers parasympathetic activity. The moderate rise in diastolic blood pressure triggered by sibutramine
would be associated with CNS-NA enhancement plus parasympathetic inhibition.
# 2005 Elsevier Inc. All rights reserved.
Keywords: Sibutramine; Plasma noradrenaline; Plasma adrenaline; Plasma dopamine; Blood serotonin; Diastolic blood pressure; Neural sympathetic activity
NeuroToxicology 27 (2006) 184–191
1. Introduction
Sibutramine is a neuropharmacological drug used as an
anorexigenic agent and as such it is widely used to treat obesity,
insulin resistance and polycystic ovary syndrome (PCOS)
(Abolfathi et al., 2004; de Simone et al., 2005; Gaciong and
Placha, 2005; Jordan et al., 2005; Karabacak et al., 2004;
McMahon et al., 2000). Although it is considered a noradrena-
line (NA) + serotonin (5HT) re-uptake inhibitor (Fukagawa and
* Corresponding author. Tel.: + 58 212 961 1048; fax: +58 212 961 0172.
E-mail address: [email protected] (F. Lechin).
0161-813X/$ – see front matter # 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuro.2005.09.004
Sakata, 2001; Invernizi and Garattini, 2004), the drug also
induces release from DA and 5HT axons (Ukai et al., 2004).
Because sibutramine is closely related to amphetamines, it is
viewed as a potentially harmful drug able to trigger adverse
cardiovascular and neuropsychiatric disorders. The fact that its
prolonged administration has provoked enhancement of central
DA activity in rodents has been cited as proof that the drug
behaves as a DA re-uptake inhibitor (Nakagawa et al., 2001).
Although the half-life for sibutramine, M1 and M2
metabolites ranged between 8 and 18 h under fasting conditions,
as in our protocol, it has been demonstrated that feeding is able to
increase the half-life of the drug (Abolfathi et al., 2004).

F. Lechin et al. / NeuroToxicology 27 (2006) 184–191 185
Littlework has been done to investigate the effects of centrally
acting drugs upon circulating neurotransmitters. Considering our
long experience measuring those parameters we feel that the
assessment of the effects of sibutramine on the latter will add
more information dealing with the neuropharmacological
mechanisms triggered by this drug. Such peripheral drug-
induced effectsmight explainmanyphysiological,metabolic and
clinical features occurring throughout neuropharmacological
treatments. This research, moreover, would help the under-
standing of complex central–peripheral interactions.
Neuropharmacological agents act on one or more CNS
circuits and all changes triggered at those levels provoke
neuroautonomic, neuroendocrine and behavioral effects
according to the subject’s pre-existing physiologic or patho-
physiologic state. Thus, full information about CNS circuitry
and CNS–peripheral interaction is basic to interpreting the
beneficial or adverse effects of all neuropharmacological drugs.
In view of the above, we proposed investigating the effects of
sibutramine, a potentially harmful drug widely used around the
world to treat obesity, and some syndromes linked to it.
2. Subjects and methods
We measured levels of plasma noradrenaline (NA), adrena-
line (Ad), dopamine (DA), free serotonin (f-5HT), tryptophane
(TRP), and platelet–serotonin (p-5HT) before (�30 and 0 min),
and after (60, 90, 120, 180, 240, 300, and 360 min) the oral
administration of 15-mg of sibutramine in 20 healthy
volunteers. We performed similar measurements 1 week earlier
in the same volunteers after administration of placebo. The
group of volunteers comprised 11 men and 9 women whose
ages ranged from 21 to 32 years (mean � S.E. = 25.8 � 2.3).
Informed consent was obtained in writing from all volunteers,
and the procedure was approved by the Ethical Committee of
FUNDAIME. All volunteers were within 10% of ideal body
weight, none had any physical or psychiatric illness. Exclusion
criteria included pregnancy, lactation, smoking, and alcohol
abuse. Volunteers were recumbent during all procedures. A
heparinized venous catheter was inserted into a forearm vein at
least 30 min before the test. We used cold, plastic syringes to
collect blood samples at the times specified above. Sibutramine
15 mg was administered orally after the second blood sample
(0 min) was obtained. Blood samples were obtained for
measuring plasma neurotransmitters and platelet aggregation.
Blood for measuring plasma neurotransmitters was transferred
to plastic tubes, each containing 1 ml of an anti-oxidant
solution (20 mg of EDTA plus 10 mg of sodiummetabisulphite/
ml). The tubes were carefully inverted several times and placed
on ice until centrifugation. To obtain platelet-rich plasma
(PRP), we centrifuged the tubes at 600 rpm at 4 8C for 15 min.
We stored 2 ml of PRP at �70 8C until needed for
determination of p-5HT levels. The remaining blood was
centrifuged again at 7000 rpm. We stored two aliquots of the
supernatant, which was platelet-poor plasma (PPP), at �70 8Cuntil needed for assays of catecholamine and f-5HT. Blood
samples for platelet aggregation were processed immediately.
A physician in constant attendance monitored heart rate and
blood pressure, and noted any symptoms reported by subjects.
2.1. Analytical assays
2.1.1. Neurochemistry
Plasma catecholamine and serotonin samples were mea-
sured in duplicate, and all determinations were made at the
same time. We used reverse phase, ion pair high-pressure liquid
chromatography with electrochemical detection (Davies and
Molyneux, 1982; Eisenhofer et al., 1986; Picard et al., 1985).
2.1.2. Reagents and standards
Noradrenaline, adrenaline, dopamine, serotonin creatinine
sulfate, dihydroxybenzylamine, 5-hydroxy-tryptophane, sodium
octyl sulfate, dibutylamine KH2PO4, citric acid, sodium acetate,
andEDTAwereobtained fromSigma-AldrichCo. (St.Louis,MO,
USA). Acid-washed aluminum oxide and microfilters were
purchased from Bioanalytical Systems Inc. (West Lafayette, IN,
USA).Acetonitrileand2-propanolwereobtainedfromRiedel-de-
Haen AG (Frankfurt, Germany). Glass-distilled water was
deionized and filtered through a Milli-O reagent grade water
system (Millipore, Bedford, MA, USA). Solutions and solvent
were filtered through a 0.2 mmMillipore filter and were vacuum
deaereated. Standard solutions (1 mmol/L) were prepared in
0.1 mol/Lperchloric acidanddiluted to thedesired concentration.
2.1.3. Equipment
Liquid chromatography was performed using Waters 515
pumps (Waters Co., Milford, MA, USA) equipped with 7125
Rheodyne valve injector fitted with a 50 ml sample loop for
detection of catecholamines, and 100 ml sample loop for p- and
f-5HT detection (Rheodyne, Berodine, Berkeley, CA, USA).
For catecholamine assays, a 15 cm � 4 mm i.d. DiscoveryTM
analytical column packed with C18 3 mm particles was used,
fitted with a precolumn filter 0.2 mm (Sigma-Aldrich Co., St.
Louis, MO, USA).
The detection system was a Waters 460 Electrochemical
Detector (Waters Co., Milford, MA, USA). A potential of
0.70 V was applied to the working electrode (glassy carbon)
versus the Ag–AgCl reference electrode. The chromatograms
were registered and quantified using Millennium software
(Waters Assoc., Milford, MA. USA).
2.1.4. Catecholamine assays
These were performed by extraction onto 20 mg of acid-
washed alumina followed by their elution with 200 ml of
0.2 mol/l HClO4 using BAS (Bioanalytical Systems) micro-
filters. The instrument was calibrated with standard plasma.
After incubation with acid-washed aluminum oxide, a plasma
pool free of catecholamines was obtained. This was processed
similarly to plasma samples, but 20 ml of standard solution
containing noradrenaline, adrenaline and dopamine (50 ng/ml
each) was added to l ml of the plasma pool to obtain the
standard plasma. Both standard plasma and sample plasma
were supplemented with 20 ml of internal standard solution
(dihydroxybenzylamine 100 ng/ml). The mobile phase was
F. Lechin et al. / NeuroToxicology 27 (2006) 184–191186
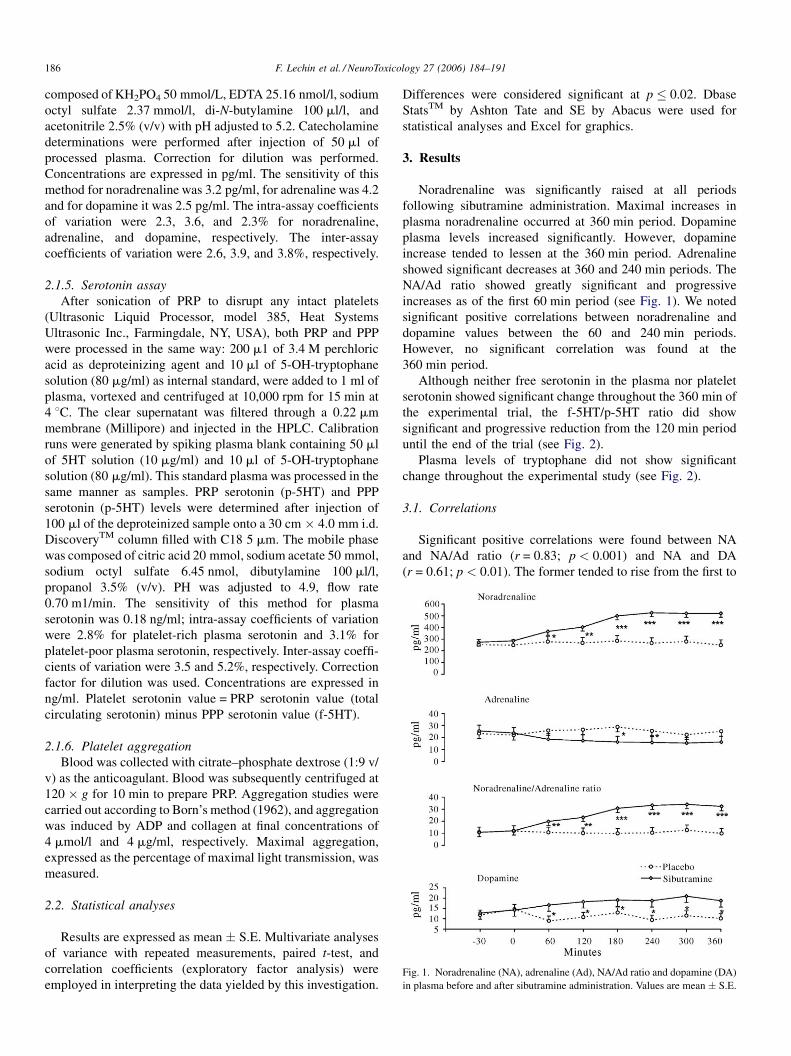
Fig. 1. Noradrenaline (NA), adrenaline (Ad), NA/Ad ratio and dopamine (DA)
in plasma before and after sibutramine administration. Values are mean � S.E.
composed of KH2PO4 50 mmol/L, EDTA 25.16 nmol/l, sodium
octyl sulfate 2.37 mmol/l, di-N-butylamine 100 ml/l, and
acetonitrile 2.5% (v/v) with pH adjusted to 5.2. Catecholamine
determinations were performed after injection of 50 ml of
processed plasma. Correction for dilution was performed.
Concentrations are expressed in pg/ml. The sensitivity of this
method for noradrenaline was 3.2 pg/ml, for adrenaline was 4.2
and for dopamine it was 2.5 pg/ml. The intra-assay coefficients
of variation were 2.3, 3.6, and 2.3% for noradrenaline,
adrenaline, and dopamine, respectively. The inter-assay
coefficients of variation were 2.6, 3.9, and 3.8%, respectively.
2.1.5. Serotonin assay
After sonication of PRP to disrupt any intact platelets
(Ultrasonic Liquid Processor, model 385, Heat Systems
Ultrasonic Inc., Farmingdale, NY, USA), both PRP and PPP
were processed in the same way: 200 m1 of 3.4 M perchloric
acid as deproteinizing agent and 10 ml of 5-OH-tryptophane
solution (80 mg/ml) as internal standard, were added to 1 ml of
plasma, vortexed and centrifuged at 10,000 rpm for 15 min at
4 8C. The clear supernatant was filtered through a 0.22 mm
membrane (Millipore) and injected in the HPLC. Calibration
runs were generated by spiking plasma blank containing 50 ml
of 5HT solution (10 mg/ml) and 10 ml of 5-OH-tryptophane
solution (80 mg/ml). This standard plasma was processed in the
same manner as samples. PRP serotonin (p-5HT) and PPP
serotonin (p-5HT) levels were determined after injection of
100 ml of the deproteinized sample onto a 30 cm � 4.0 mm i.d.
DiscoveryTM column filled with C18 5 mm. The mobile phase
was composed of citric acid 20 mmol, sodium acetate 50 mmol,
sodium octyl sulfate 6.45 nmol, dibutylamine 100 ml/l,
propanol 3.5% (v/v). PH was adjusted to 4.9, flow rate
0.70 m1/min. The sensitivity of this method for plasma
serotonin was 0.18 ng/ml; intra-assay coefficients of variation
were 2.8% for platelet-rich plasma serotonin and 3.1% for
platelet-poor plasma serotonin, respectively. Inter-assay coeffi-
cients of variation were 3.5 and 5.2%, respectively. Correction
factor for dilution was used. Concentrations are expressed in
ng/ml. Platelet serotonin value = PRP serotonin value (total
circulating serotonin) minus PPP serotonin value (f-5HT).
2.1.6. Platelet aggregation
Blood was collected with citrate–phosphate dextrose (1:9 v/
v) as the anticoagulant. Blood was subsequently centrifuged at
120 � g for 10 min to prepare PRP. Aggregation studies were
carried out according to Born’s method (1962), and aggregation
was induced by ADP and collagen at final concentrations of
4 mmol/l and 4 mg/ml, respectively. Maximal aggregation,
expressed as the percentage of maximal light transmission, was
measured.
2.2. Statistical analyses
Results are expressed as mean � S.E. Multivariate analyses
of variance with repeated measurements, paired t-test, and
correlation coefficients (exploratory factor analysis) were
employed in interpreting the data yielded by this investigation.
Differences were considered significant at p � 0.02. Dbase
StatsTM by Ashton Tate and SE by Abacus were used for
statistical analyses and Excel for graphics.
3. Results
Noradrenaline was significantly raised at all periods
following sibutramine administration. Maximal increases in
plasma noradrenaline occurred at 360 min period. Dopamine
plasma levels increased significantly. However, dopamine
increase tended to lessen at the 360 min period. Adrenaline
showed significant decreases at 360 and 240 min periods. The
NA/Ad ratio showed greatly significant and progressive
increases as of the first 60 min period (see Fig. 1). We noted
significant positive correlations between noradrenaline and
dopamine values between the 60 and 240 min periods.
However, no significant correlation was found at the
360 min period.
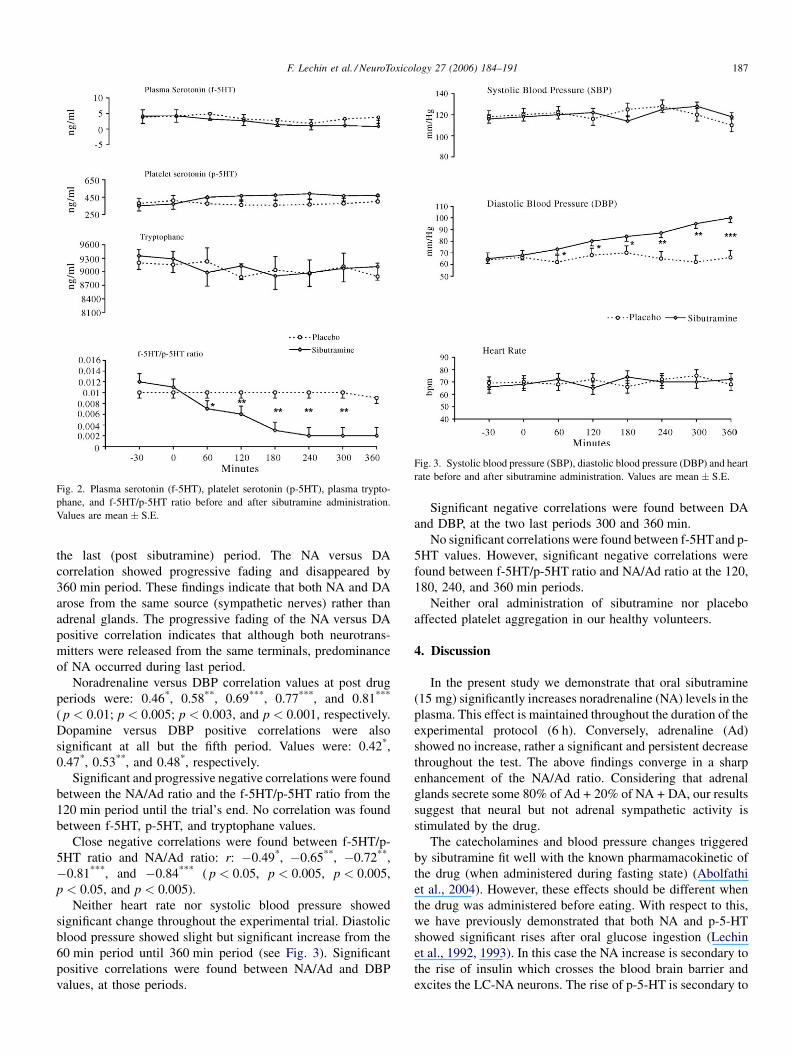
Although neither free serotonin in the plasma nor platelet
serotonin showed significant change throughout the 360 min of
the experimental trial, the f-5HT/p-5HT ratio did show
significant and progressive reduction from the 120 min period
until the end of the trial (see Fig. 2).
Plasma levels of tryptophane did not show significant
change throughout the experimental study (see Fig. 2).
3.1. Correlations
Significant positive correlations were found between NA
and NA/Ad ratio (r = 0.83; p < 0.001) and NA and DA
(r = 0.61; p < 0.01). The former tended to rise from the first to
F. Lechin et al. / NeuroToxicology 27 (2006) 184–191 187
Fig. 2. Plasma serotonin (f-5HT), platelet serotonin (p-5HT), plasma trypto-
phane, and f-5HT/p-5HT ratio before and after sibutramine administration.
Values are mean � S.E.
Fig. 3. Systolic blood pressure (SBP), diastolic blood pressure (DBP) and heart
rate before and after sibutramine administration. Values are mean � S.E.
the last (post sibutramine) period. The NA versus DA
correlation showed progressive fading and disappeared by
360 min period. These findings indicate that both NA and DA
arose from the same source (sympathetic nerves) rather than
adrenal glands. The progressive fading of the NA versus DA
positive correlation indicates that although both neurotrans-
mitters were released from the same terminals, predominance
of NA occurred during last period.
Noradrenaline versus DBP correlation values at post drug
periods were: 0.46*, 0.58**, 0.69***, 0.77***, and 0.81***
( p < 0.01; p < 0.005; p < 0.003, and p < 0.001, respectively.
Dopamine versus DBP positive correlations were also
significant at all but the fifth period. Values were: 0.42*,
0.47*, 0.53**, and 0.48*, respectively.
Significant and progressive negative correlations were found
between the NA/Ad ratio and the f-5HT/p-5HT ratio from the
120 min period until the trial’s end. No correlation was found
between f-5HT, p-5HT, and tryptophane values.
Close negative correlations were found between f-5HT/p-
5HT ratio and NA/Ad ratio: r: �0.49*, �0.65**, �0.72**,
�0.81***, and �0.84*** ( p < 0.05, p < 0.005, p < 0.005,
p < 0.05, and p < 0.005).
Neither heart rate nor systolic blood pressure showed
significant change throughout the experimental trial. Diastolic
blood pressure showed slight but significant increase from the
60 min period until 360 min period (see Fig. 3). Significant
positive correlations were found between NA/Ad and DBP
values, at those periods.
Significant negative correlations were found between DA
and DBP, at the two last periods 300 and 360 min.
No significant correlations were found between f-5HTand p-
5HT values. However, significant negative correlations were
found between f-5HT/p-5HT ratio and NA/Ad ratio at the 120,
180, 240, and 360 min periods.
Neither oral administration of sibutramine nor placebo
affected platelet aggregation in our healthy volunteers.
4. Discussion
In the present study we demonstrate that oral sibutramine
(15 mg) significantly increases noradrenaline (NA) levels in the
plasma. This effect is maintained throughout the duration of the
experimental protocol (6 h). Conversely, adrenaline (Ad)
showed no increase, rather a significant and persistent decrease
throughout the test. The above findings converge in a sharp
enhancement of the NA/Ad ratio. Considering that adrenal
glands secrete some 80% of Ad + 20% of NA + DA, our results
suggest that neural but not adrenal sympathetic activity is
stimulated by the drug.
The catecholamines and blood pressure changes triggered
by sibutramine fit well with the known pharmamacokinetic of
the drug (when administered during fasting state) (Abolfathi
et al., 2004). However, these effects should be different when
the drug was administered before eating. With respect to this,
we have previously demonstrated that both NA and p-5-HT
showed significant rises after oral glucose ingestion (Lechin
et al., 1992, 1993). In this case the NA increase is secondary to
the rise of insulin which crosses the blood brain barrier and
excites the LC-NA neurons. The rise of p-5-HT is secondary to

F. Lechin et al. / NeuroToxicology 27 (2006) 184–191188
the excitation of the enterocromafın cells triggered by glucose
plus the post-prandial parasympathetic drive. This latter
mechanism is absent after the administration of sibutramine.
Small but significant rises of DA were also found after the
drug’s administration. This finding is consistent with the well
known fact that both catecholamines arise from the same source
(sympathetic nerves). The fact that the NA versus DA
significant positive correlation registered in this study
diminished with time is consistent with the progressive
predominance of NA over DA release. It fits well with the
finding that NA/DA ratio tended to rise throughout the 6 h of the
trial. In this regard, it is well known that the DA pool which
exists at sympathetic terminals is released during neural
sympathetic activation (Soares Da Silva, 1986). The DA
released from those terminals acts as a modulatory agent able to
bridle overflowing NA release, by acting at DA2 inhibitory
auto-receptors existing at this level (Artalejo et al., 1985;
Mannelli et al., 1988; Missale et al., 1988; Soares Da Silva,
1986, 1987). Evidently, enhanced release of DA triggered by
the drug was not enough to impede an overwhelming release of
NA, responsible for the diastolic blood pressure (DBP) rise that
we registered in all cases. This phenomenon is the opposite of
that observed in the orthostatic hypotension syndrome when
excessive DA spillover impedes NA release from sympathetic
terminals (Robertson et al., 1979; Van Loon et al., 1979;
Wilffert et al., 1984; Ziegler et al., 1979). The above
observations are reinforced by our findings concerning blood
pressure versus NA, DA, and Ad correlations. Whereas DBP,
but not SBP, showed significant positive correlation versus NA
and DA, no significant correlation was observed between blood
pressure and Ad values.
Birkenfeld et al. (2005) found that resting blood pressure
tends to increase with sibutramine. These findings are similar to
those registered in our study. However, other contradictory
findings in the Birkenfeld et al. (2002, 2005) studies should be
discussed on the basis that peripheral sympathetic activity
depends on two separate branches: neural sympathetic
(sympathetic nerves = 90% NA and 10% DA) and adrenal
sympathetic (adrenal glands = 80% adrenaline + 10% noradre-
naline + 10% dopamine, approximately). These CNS sympa-
thetic drives may function in association or dissociation. Our
experimental design demonstrated that sibutramine triggers
neural but not adrenal sympathetic excitation. For this reason
diastolic but not systolic blood pressure showed significant
increase. These findings are consistent with those shown in our
study demonstrating that sibutramine did not raise either
systolic blood pressure or heart rate, both parameters depending
on beta-adrenergic (adrenal sympathetic) but not alpha-
adrenergic (neural sympathetic) mechanisms. The diastolic
blood pressure rise triggered by the drug showed attenuation
when ingested over several days (Birkenfeld et al., 2002), an
effect which should be attributed to the fact that sibutramine is
an inhibitor of both NA and 5-HT uptake. Obviously, the latter
effect is slower to be triggered than the former. Here, it should
be taken into account that dorsal raphe serotonergic neurons
send inhibitory axons to CNS-NA neurons (Szabo and Blier,
2001). These mechanisms explain why sibutramine-induced
anorectic effects, depending on serotonergic activity, increased
progressively.
At CNS level, the hyper-neural sympathetic activity
provoked by sibutramine in our experimental study fits well
with the postulation that this drug triggers hyperactivity of the
CNS-NA circuitry responsible for neural sympathetic but not
adrenal sympathetic activation. These findings are in accor-
dance with the fact that sibutramine is a central NA-uptake
enhancer drug. With respect to the above, it is well documented
that the two branches of peripheral sympathetic activity can
function in association or dissociation (Lechin et al., 2002b,
2004c; Young et al., 1984). Exhaustive experimental evidence
demonstrates that whereas adrenal sympathetic activity is
directly ruled by the medullary Ad-C1 nuclei, neural
sympathetic release depends on the noradrenergic pontine
nuclei (A5, A6, A7). Whereas the two C1-Ad nuclei send axons
to sympathetic pre-ganglionic neurons located at the cervical
and thoracic intermedio-lateral spinal column, the pontine
noradrenergic nuclei send axons to pre-ganglionic neurons of
the lumbar segment. Axons from the former group innervate the
adrenal glands whereas those of the second group reach the
lumbar sympathetic ganglia (Ader et al., 1979; Commissiong
et al., 1978; Dampney et al., 2003; Folkow and von Euler, 1954;
Gurtu et al., 1984; Hancock and Fougerousse, 1976; Kobayashi
et al., 1975; Nygren and Olson, 1977; Satoh et al., 1977; Smith
and Barrow, 1990; Pearson, 1978). Tracing with retrovirus has
ratified neurophysiological and neuropharmacological research
in this area (Blessing et al., 1981; Ding et al., 1993; Koulu et al.,
1986; Li and Guyenet, 1995; Li et al., 1992; Wessenlingh et al.,
1989). On the other hand, our research work dealing with the
assessment of circulating neurotransmitters before and after
various types of stress tests like orthostasis, exercise (Lechin
et al., 1994, 1995a,b, 1996b), oral glucose tolerance test
(Lechin et al., 1991, 1992, 1993), neuropharmacological drugs
(Lechin et al., 1997, 1985a,b), and others carried out during the
last 25 years, is in line with the clear demonstration that neural
and adrenal sympathetic activities depend on two distinct CNS
circuitries. One of the most important findings stemming from
the above is that diastolic blood pressure is closely and
maximally dependent on the CNS–pontine NA nuclei (A5, A6,
A7) activities, as demonstrated in the experimental model of
essential hypertension (Lewis rats) (Koulu et al., 1986). These
mammals are characterized by a hyperactive pontine NA
nuclei + DBP rise. In addition, they respond to all types of
emotional stimuli with enhancement of the firing of CNS-NA
neurons + diastolic blood pressure rise + increased NA levels at
both cerebro-spinal fluid and blood levels. According to the
foregoing, results emanating from the present investigation
strongly suggest that acute administration of sibutramine
provokes CNS-NA activation, responsible for the peripheral
neural sympathetic release without participation of adrenal
gland secretion. Our findings fit well with the fact that
sibutramine acts as an NA-uptake inhibitor.
The lowered f-5HT/p-5HT ratio provoked by sibutramine in
our experimental trial merits detailed discussion. Most
circulating serotonin is stored in platelets (p-5HT) = 97–
99%. Free serotonin in the plasma (f-5HT) accounts for the

F. Lechin et al. / NeuroToxicology 27 (2006) 184–191 189
remaining 1–3%. The f-5HT/p-5HT ratio increases during
platelet aggregation or platelet activation (Born, 1962). Other
factors able to increase f-5HT include the rise of plasma levels
of both acetylcholine (ACh) (Rausch et al., 1985) and dopamine
(DA) (De Keyser et al., 1988; Ricci et al., 2001). Both
neurotransmitters compete for platelet uptake with serotonin.
Thus, any rise of ACh and DA tends to increase f-5HT. The fact
that we found a reduction of the f-5HT/p-5HT ratio, in our
experimental subjects, despite noting moderate increases of DA
in the plasma, obliges us to postulate that ACh which also
interferes with platelet uptake, would be significantly lowered
after the drug’s administration. Although we cannot directly
measure ACh levels in peripheral blood, we previously
demonstrated a close positive correlation between f-5HT/p-
5HT ratio and peripheral parasympathetic activity in depressed
subjects (Lechin et al., 1995a,b, 1996a) known to present a
hyperparasympathetic profile. In addition, we showed that the
f-5HT/p-5HT ratio correlated positively with parasympathetic
activity occurring during sleep and wake periods (Lechin, 2000;
Lechin and van der Dijs, 2005; Lechin et al., 1992, 1994,
2004a,b) which was reverted by atropine (Lechin, 2000; Lechin
et al., 1994, 1996c). Thus, that ratio seems to be an adequate
index of ACh circulating levels in normal subjects, in absence
of platelet aggregation and other interfering factors. Accord-
ingly we propose that the very low f-5HT/p-5HT ratio observed
in our experimental study reflects a low parasympathetic
activity secondary to the neural sympathetic predominance
triggered by the drug. This postulation is supported by the fact
that the NA pontine nuclei send direct inhibitory axons to the
dorsal motor vagal nuclei (Lechin et al., 2002a). This
postulation is reinforced by the close negative correlations
we found in the present study between f-5HT/p-5HT ratio and
NA/Ad ratio. The low parasympathetic activity is also
consistent with the previously mentioned cardiovascular,
gastrointestinal, psychiatric, and sleep disturbances triggered
by the drug after prolonged administration (Birkenfeld et al.,
2002, 2005; de Simone et al., 2005; Jordan et al., 2005).
Considering that both p- and f-5-HT rises registered after an
oral glucose load (Lechin et al., 1992, 1993) reflects
parasympathetic activity, it is logical to assume that the
lowering of these parameters registered after sibutramine
administration should be associated with hypersympathetic
activity. In addition, it has been shown that the bioavailability of
sibutramine was significantly increased with the administration
of food (Abolfathi et al., 2004).
Sibutramine increases insulin sensitivity and improves
insulin resistance (Wu et al., 2003). With respect to this, it
is a well known fact that insulin crosses the blood brain barrier
and excites CNS-NA neurons. These CNS-NA neurons send
excitatory drive to neural sympathetic nerves which release NA
and DA. In addition, it has been demonstrated that insulin acts
as an NA-uptake inhibitor at the CNS-NA neurons (Boyd et al.,
1986). Thus, this drug behaves as a potentiating factor of the
CNS-induced insulin effect.
The fact that sibutramine decreased f-5HT/p-5HT ratio,
despite the fact that the drug is a serotonin uptake inhibitor (at
both platelets and serotonergic axons), reinforces the postula-
tion that this acute effect is secondary to the reduction of
circulating acetylcholine levels triggered by the enhancement
of neural sympathetic activity. These findings agree with the
postulation of the f-5HT/p-5HT ratio as an index of
parasympathetic activity in absence of platelet aggregation.
We investigated the effects of sibutramine throughout a 6-h
period. We did not prolonged this study because in hour
experience, prolonged supine-resting state tends to induce sleep
and might change the circulating neurotransmitters profile thus,
we would introduce an additional factor able to trigger both
central and peripheral neurochemical changes. However, we
are now investigating the neurotransmitters profile in subjects
who are consuming the drug during prolonged periods.
Summarizing all the above, it is logical to assume that
sibutramine reduces food intake by both acute and chronic
effects. The former would depend on the anti-parasympathetic
mechanisms triggered by the drug, whereas the enhancement of
central serotonergic activity would be responsible for the
chronic anorexigenic effects.
References
Abolfathi Z, Couture J, Vallee F, LeBel M, TanguayM, Masson E. A pilot study
to evaluate the pharmacokinetics of sibutramine in healthy subjects under
fasting and fed conditions. J Pharm Pharmaceut Sci 2004;7(3):345–9.
Ader JP, Postema F, Korf J. Contribution of the locus coeruleus to the adrenergic
innervation of the rat spinal cord: a biochemical study. J Neural Transm
1979;44:159–73.
Artalejo AR, Garcia AG, Montiel C, Sanchez-Garcia P. A dopaminergic
receptor modulates catecgolamine release from the cat adrenal gland. J
Physiol 1985;362:359–68.
Birkenfeld AL, Schroeder C, Boschmann M, Tank J, Franke G, Luft FC, et al.
Paradoxical effect of sibutramine on autonomic cardiovascular regulation.
Circulation 2002;106:2459–65.
Birkenfeld AL, Schroeder C, Pischon T, Tank J, Luft FC, Sharma AM, et al.
Paradoxical effect of sibutramine on autonomic cardiovascular regulation in
obese hypertensive patients. Sibutramine and blood pressure. Clin Auton
Res 2005;15(3):200–6.
Blessing WW, Goodchild AK, Dampney RA, Chalmers JP. Cell groups in the
lower brain stem of the rabbit projecting to the spinal cord, with special
reference to catecholamines-containing neurons.. Brain Res 1981;221:35–55.
Born GVR. Aggregation of blood platelets by adenosine diphosphate and its
reversal. Nature 1962;194:927–9.
Boyd FT, Clarke DW, Raizada MK. Insulin inhibits specific norepinephrine
uptake in neuronal cultures from rat brain. Brain Res 1986;398:1–5.
Commissiong JW, Hellstrom SO, Neff NH. A new projection from locus
coeruleus to the spinal ventral columns: histochemical and biochemical
evidence. Brain Res 1978;148:207–13.
Dampney RA, Horiuchi J, Tagawa T, Fontes MA, Potts PD, Polson JW.
Medullary and supramedullary mechanisms regulating sympathetic vaso-
motor tone. Acta Physiol Scand 2003;177:209–18.
Davies CL, Molyneux SG. Routine determination of plasma catecholamines
using reversed phase ion pair high performance liquid chromatography with
electrochemical detection. J Chromatogr 1982;231:41–51.
De Keyser S, De Waele M, Convents A, Ebinger G, Vauquelin G. Identification
of D1 like dopamine receptors on human blood platelets. Life Sci
1988;42:1797–806.
De Simone G, Romano C, De Caprio C, Contaldo F, Salanitri T, Paparatti UL, et
al. Effects of sibutramine-induced weight loss on cardiovascular system in
obese subjects. Nutr Metab Cardiovasc Dis 2005;15:24–30.
Ding ZQ, Li YW, Wessenlingh SL, Blessing WW. Transneuronal labeling of
neurons in rabbit brain after injection of herpes simplex virus type 1 into the
renal nerve. J Auton Nerv Syst 1993;42:23–31.

F. Lechin et al. / NeuroToxicology 27 (2006) 184–191190
Eisenhofer G, Goldstein DS, Stull R. Simultaneous liquid chromatographic
determination of 3,4-dihydroxyphenylglycol, catecholamines, and 3,4-
dihydroxy-phenylalanine in plasma and their responses to inhibition of
monoamine oxidase. Clin Chem 1986;32:2030–3.
Folkow B, von Euler US. Selective activation of noradrenaline and adrenaline
producing cells in the suprarenal gland of the cat by hypothalamic stimula-
tion. Circul Res 1954;2:191–5.
Fukagawa K, Sakata T. Monoaminergic anorectic agents. Nippon Yakurigaku
Zasshi 2001;118:303–8.
Gaciong Z, Placha G. Efficacy and safety of sibutramine in 2225 subjects with
cardiovascular risk factors: short term, open label, observational study. J
Hum Hypertens 2005;19:737–43.
Gurtu S, Pant KK, Sinha JN, Bhargava KP. An investigation into the mechanism
of cardiovascular responses elicited by electrical stimulation of locus
coeruleus and subcoeruleus in the cat. Brain Res 1984;301:59–64.
Hancock MB, Fougerousse CL. Spinal projections from the nucleus locus
coeruleus and nucleus subcoeruleus in the cat and monkey as demonstrated
by the retrograde transport of horseradish peroxidase. Brain Res Bull
1976;1:229–34.
Invernizi RW, Garattini S. Role of presynaptic alpha2-adrenoceptors in anti-
depressant action: recent findings from microdialysis studies. Prog Neu-
ropsychopharmacol Biol Psychiatry 2004;28:819–27.
Jordan J, Scholze J, Matiba B, Wirth A, Hauner H, Sharma AM. Influence of
sibutramine on blood pressure: evidence from placebo-controlled trials. Int J
Obes 2005;29:509–16.
Karabacak IY, Karabacak O, Toruner FB, Akdemir O, Arslan M. Treatment
effect of sibutramine compared to fluoxetine on leptin levels in polycyctic
ovary disease. Gynecol Endocrinol 2004;19:196–201.
Kobayashi RM, Palkovits M, Jacobwitz DM, Kopin IJ. Biochemical mapping of
the noradrenergic projection from the locus coeruleus. A model for studies
of brain neuronal pathways. Neurology 1975;25:223–33.
Koulu M, Saavedra JM, Niwa M, Linnoila M. Increased catecholamines
metabolism in the locus coeruleus of young spontaneously hypertensive
rats. Brain Res 1986;369:361–4.
Lechin F. Central and plasma 5-HT, vagal tone and airways. Trends Pharmacol
Sci (TiPS) 2000;21:425.
Lechin AE, Varon J, van der Dijs B, Lechin F. Plasma catecholamines and
indoleamines during attacks and remission on severe bronchial asthma:
possible role of stress. Am J Respir Crit Care Med 1994;149:A778.
Lechin F, van der Dijs B. Blood pressure and autonomic nervous system
assessment throughout the sleep cycle in normal adults. Sleep
2005;28(5):645–6.
Lechin F, van der Dijs B, Jakubowicz D, Camero RE, Villa S, Arocha L, et al.
Effects of clonidine on blood pressure, noradrenaline, cortisol, growth
hormone, and prolactin plasma levels in high and low intestinal tone
depressed patients. Neuroendocrinology 1985a;41:156–62.
Lechin F, van der Dijs B, Jakubowicz D, Camero RE, Villa S, Lechin E, et al.
Effects of clonidine on blood pressure, noradrenaline, cortisol, growth
hormone and prolactin plasma levels in low and high intestinal tone
subjects. Neuroendocrinology 1985b;40:253–61.
Lechin F, van der Dijs B, Lechin A, Lechin M, Coll-Garcia E, Jara H, et al.
Doxepin therapy for postprandial symptomatic hypoglycemic patients:
neurochemical, hormonal and metabolic disturbances. Clin Sci
1991;80:373–84.
Lechin F, van der Dijs B, Lechin M, Lechin-Baez S, Jara H, Lechin A, et al.
Effects of an oral glucose load on plasma neurotransmitters in humans:
involvement of REM sleep? Neuropsychobiology 1992;26:4–11.
Lechin F, van der Dijs B, Lechin M, Jara H, Lechin A, Lechin-Baez S, et al.
Plasma neurotransmitters throughout an oral glucose tolerance test in
essential hypertension. Clin Exp Hypertens 1993;15:209–40.
Lechin F, van der Dijs B, Orozco B, Lechin ME, Acosta E, Lechin AE, et al.
Plasma neurotransmitters, blood pressure and heart rate during supine-
resting, orthostasis and moderate exercise conditions in major depressed
patients. Biol Psychiatry 1995a;38:166–73.
Lechin F, van der Dijs B, Orozco B, Lechin AE, Baez S, Lechin ME, et al.
Plasma neurotransmitters, blood pressure and heart rate during supine-
resting, orthostasis and moderate exercise in dysthymic depressed patients.
Biol Psychiatry 1995b;37:884–91.
Lechin F, van der Dijs B, Benaim M. Stress versus depression (Review). Prog
Neuropsychopharmacol Biol Psychiatry 1996a;20:899–950.
Lechin F, van der Dijs B, Orozco B, Lechin AE, Baez S, Lechin ME, et al.
Plasma neurotransmitters, blood pressure and heart during supine-resting,
orthostasis and moderate exercise in severely ill patients: a model of failing
to cope with stress. Psychother Psychosom 1996b;65:129–36.
Lechin F, van der Dijs B, Orozco B, Baez S, Rada I, Leon G, et al. Plasma
neurotransmitters, blood pressure and heart rate during supine-resting,
orthostasis and moderate exercise stress test in healthy humans before
and after parasympathetic blockade with atropine. Res Comm Biol Psychol
Psychiatry 1996c;21:55–72.
Lechin F, van der Dijs B, Jara H, Orozco B, Baez S, Jahn E, et al. Plasma
neurotransmttter profiles of anxiety, phobia and panic disorder patients.
Acute and chronic effects of buspirone. Res Comm Biol Psychol Psychiatry
1997;22:95–110.
Lechin F, van der Dijs B, Lechin ME. Neurocircuitry and neuroautonomic
disorders: Reviews and strategies of therapy. Basel Karger 2002;57–9.
Lechin F, Lechin ME, van der Dijs B. Plasma catecholamines and chronic
congestive heart failure. Circulation 2002b;106(25):222–3.
Lechin F, van der Dijs B, Lechin AE. The autonomic nervous system assessment
throughout thewake-sleep cycle and stress. PsychosomMed 2004a;66:974–
6.
Lechin F, van der Dijs B, Pardey-Maldonado B, Benaim M, Baez S, Orozco B,
Lechin AE. Circulating neurotransmitter profiles during the different wake-
sleep stages in normal subjects. Psychoneuroendocrinology 2004b;29:669–
85.
Lechin F, van der Dijs B, Lechin AE. Neural sympathetic activity in essential
hypertension. Hipertension 2004c;44:e3–4.
Li YW, Ding ZQ, Wessenlingh SL, Blessing WW. Renal and adrenal sympa-
thetic preganglionic neurons in rabbit spinal cord: tracing with herpes
simplex virus.. Brain Res 1992;573:147–52.
Li YW, Guyenet PG. Neuronal excitation by angiotensin II in the rostral
ventrolateral medulla of the rat in vitro. Am J Physiol 1995;268:
R272–7.
Mannelli M, Pupilli C, Fabbri G, Musante R, De Feo ML, Franchi F, et al.
Endogenous dopamine (DA) and DA2 receptors: a mechanism limiting
excessive sympathic-adrenal discharge in humans. J Clin Endocr Metab
1988;66:626–31.
McMahon FG, Fujioka K, Singh BN, Mendel CM, Rowe E, Rolston K, et al.
Efficacy and safety of sibutramine in obese white and African American
patients with hypertension. Arch Int Med 2000;160:2185–91.
Missale C, Memo M, Liberini P, Spano P. Dopamine selectively inhibits
angiotensinII-induced aldosterone secretion by interacting with D2 recep-
tors. J Pharmacol Exp Ther 1988;246:1137–43.
Nakagawa T, Ukai K, Ohyama T, Gomita Y, Okamura H. Effects of sibutrmine
on the central dopaminergic system in rodents. Neurotox Res 2001;3:235–
47.
Nygren LG, Olson L. A new major projection from the locus coeruleus: the
main source of noradrenergic nerve terminals in the ventral and dorsal
columns of spinal cord. Brain Res 1977;132:85–93.
Pearson JA. The coeruleo-spinal pathway: an anatomical, neurochemical and
electrophysiological study. Soc Neurosci Abstr 1978;4:1831.
Picard M, Olichon D, Gombert J. Determination of serotonin in plasma by
liquid chromatography with electrochemical detection. J Chromatogr
1985;341:445–51.
Rausch JL, Janowsky SC, Huey LY. Physostigmine effects on serotonin uptake
in human blood platelets. Eur J Pharmacol 1985;109:91–6.
Ricci A, Bronzetti E, Mannino F, Mignini F, Morosetti C, Tayebati SK, et al.
Dopamine receptors in human platelets. Naunyn Schmiedeberg’s Arch
Pharmacol 2001;109:91–6.
Robertson D, Johnson GA, Robertson RM, Nies AS, Shand DG, Oates JA.
Comparative assessment of stimuli that release neuronal and
adrenomedullary catecholamines in man. Circulation 1979;59:637–
643.
Satoh K, Tohyama M, Yamamoto K, Sukumoto T, Shimuzu N. Noradrenaline
innervation of the spinal cord studied by the horseradish peroxidase method
combined with monoamine oxidase staining. Exp Brain Res 1977;30:175–
86.

F. Lechin et al. / NeuroToxicology 27 (2006) 184–191 191
Smith JK, Barrow KW. Cardiovascular effects of L-glutamate and tetrodotoxin
microinjected into the rostral and caudal ventrolateral medulla in normo-
tensive and spontaneously hypertensive rats. Brain Res 1990;506:1–8.
Soares Da Silva P. Evidence for a non-precursor dopamine pool in noradre-
nergic neurons of the dog mesenteric artery. Naunyn Schmiedeberg’s Arch
Pharmacol 1986;333:219–23.
Soares Da Silva P. A comparison between the pattern of dopamine and
noradrenaline release from sympathetic neurons of the dog mesenteric
artery. Br J Pharmacol 1987;90:91–8.
Szabo ST, Blier P. Functional and pharmacological characterization of the
modulatory role of serotonin on the firing activity of locus coeruleus
norepinephrine neurons. Brain Res 2001;922:9–20.
Ukai K, Nakagawa T, Ohyama T, Nakanishi H. Sibutramine induces potential-
dependent exocytotic release but not carrier-mediated release of dopamine
and 5-hydroxytryptamine. Eur J Pharmacol 2004;484:209–15.
Van Loon GR, Sole MJ, Bain J, Ruse JL. Effects of bromocriptine on plasma
catecholamine in normal men. Neuroendocrinology 1979;28:425–34.
Wessenlingh SL, Li YW, Blessing WW. PNMT-containing neurons in the
rostral medulla oblongata (C1, C3 groups) are transneuronally labeled after
injection of herpes simplex virus type 1 into the adrenal gland. Neurosci Lett
1989;106:99–104.
Wilffert B, Smitt G, de Jonge A, Thoolen MJ, Timmermans P, van Zwieten PA.
Inhibitory dopamine receptors on sympathetic neurons innervating the
cardiovascular system of the pithed rat. Characterization and role in relation
to presynaptic alpha-2-adrenoceptors. Naunyn Schmiedeberg’s Arch Phar-
macol 1984;326:91–8.
Wu J, Lei MX, Chea HL. Serum leptin and insulin resistance in obesity and
effects of sibutramine on them. Hunan Yi Ke DA Xue Xue Bao
2003;28:605–7.
Young JB, Rosa RM, Landsberg L. Dissociation of sympathetic nervous system
and adrenal medullary responses. Am J Physiol 1984;247:E35–40.
Ziegler MG, Lake CR, Williams AC, Teychenne PF, Shoulson I, Steinsland O.
Bromocriptine inhibits norepinephrine release. Clin Pharmacol Ther
1979;25:137–42.
Related Documents











