This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Neurobiology of AddictionInsight from Neurochemical Imaging
Nina B.L. Urban, MD, MSca,b,�, Diana Martinez, MD
a,b
Drug addiction is a chronically relapsing disorder characterized by:1
1. Compulsion to seek and take the drug2. Loss of control in limiting intake3. Appearance of negative emotional symptoms as well as physical withdrawal
symptoms when the drug cannot be accessed.
The authors have nothing to disclose.a Department of Psychiatry, Division of Substance Abuse, Columbia University, New York, NY10032, USA; b Department of Psychiatry, New York State Psychiatric Institute, 1051 RiversideDrive, Unit 31, New York, NY 10032, USA* Corresponding author. Department of Psychiatry, New York State Psychiatric Institute, 1051Riverside Drive, Unit 31, New York, NY 10032.E-mail address: [email protected]
Psychiatr Clin N Am 35 (2012) 521–541http://dx.doi.org/10.1016/j.psc.2012.03.011 psych.theclinics.com0193-953X/12/$ – see front matter © 2012 Elsevier Inc. All rights reserved.
KEYWORDS
• PET • SPECT • Dopamine • Reward system • Psychostimulants• Neuroimaging
KEY POINTS
• Factors that determine who becomes addicted include genetic, developmental, andenvironmental risk factors.
• PET and SPECT imaging use radioligands that are specific for a given brain receptor,and labeled with a traceable radionuclide, allowing quantification of these receptorsin humans in vivo.
• The mesolimbic and mesocortical dopamine pathways are crucial in drug reward andaddiction. Imaging studies investigate vulnerability to addiction through dopamine-receptor and -transporter binding, as well as exploring dopamine transmission viapharmacologic challenge and depletion studies.
• Both dopamine receptors and dopamine release in areas of the striatum are reducedin cocaine, methamphetamine, heroin, and alcohol dependence, possibly related totreatment refractoriness.
• With the development of new radioligands, neuroimaging exploration of otherneurotransmitter systems and their role in addiction has begun as well.

Author's personal copy
Much of the current neurobiological drug abuse research is focused on understandingthe molecular mechanisms mediating the transition to the loss of behavioral controlover drug-seeking and drug-taking, and the neuroadaptive changes that occur in thebrain. The aim of this article is to summarize the contributions of neurochemicalimaging studies to a better understanding of the neurobiology of addiction.
THE NEUROCIRCUITRY OF REWARD IN ADDICTION
Drugs of abuse mimic or enhance the actions of neurotransmitters, the endogenouschemical messengers in the nervous system, at receptors for these neurotransmitters.Opioids are presumed to be habit-forming because of actions at opiate receptors,and nicotine because of action at nicotinic acetylcholine receptors. Phencyclidineacts at N-methyl-D-aspartate (NMDA) and sigma receptors, and also blocks dopa-mine reuptake.2 �9-Tetrahydrocannabinol (THC) binds to endocannabinoid recep-tors.3,4 Although amphetamine and cocaine do not act directly at dopamine recep-tors, they are reinforcing because they increase the concentration of dopamine at thedopamine receptors of the nucleus accumbens and frontal cortex.5
It has become clear that the acute administration of most drugs of abuse increasesdopamine transmission in the basal ganglia,5 and that dopamine transmission in thisbrain region plays a crucial role in mediating the reinforcing effects of these drugs.6–8
The mesolimbic dopamine pathway is made up of dopaminergic cells in the ventraltegmental area (VTA) projecting into the nucleus accumbens (NAc), located in theventral striatum, and is considered crucial for drug reward.9 The mesostriatal(dopamine cells of the substantia nigra projecting to the dorsal striatum) andmesocortical (dopamine cells of the VTA projecting to the frontal cortex) pathways arealso recognized in contributing to predicting drug reward (anticipation) and addic-tion.10 The time course of dopamine signaling is also a key factor, where the fastesttime course predominantly has a role in reward and attributing value to predictedoutcomes of behavior, while steady activation of dopamine release plays a role inproviding an enabling effect on specific behavior-related systems.11 The mode ofdopamine cell firing (phasic vs tonic) also differently modulates the rewarding,conditioning effects, or drugs (predominantly phasic) versus the changes in executivefunction that occur in addiction (predominantly tonic).12
IMAGING BRAIN RECEPTORS
The studies discussed in this article use positron emission tomography (PET) orsingle-photon emission computed tomography (SPECT) to image specific receptorsand changes in neurotransmitters that occur in addiction. PET and SPECT useagonists and antagonists that are specific for a given brain receptor, and are labeledwith a radionuclide (usually carbon-11 [11C] or fluorine-18 [18F] for PET and iodine-123[123I] for SPECT), which allow quantification of these receptors in human brainimaging studies (for an in-depth review of these imaging techniques, please seeRefs.13,14). Briefly stated, PET and SPECT use radiotracers such that the radionuclideis incorporated into the receptor-specific molecule, which allows detection of theradiotracer with specialized scanners by virtue of its radioactive signal, as it binds tothe receptors in the brain. The main outcome measure used in these imaging studiesis the “binding potential” (BP), which depends on both the density of the receptor andits affinity for the radiotracer. BP is generally measured as either BPND (BP relative tothe free fraction of radiotracer in the nondisplaceable, ie, not receptor specific, braintissue) or BPP (BP relative to the free fraction of radiotracer in the arterial plasma).15
In addition to imaging brain receptors, some radiotracers can be used to image thechange in the levels of neurotransmitters within the brain.
522 Urban & Martinez
Author's personal copy
Pharmacologic Challenges to Measure Dopamine Release
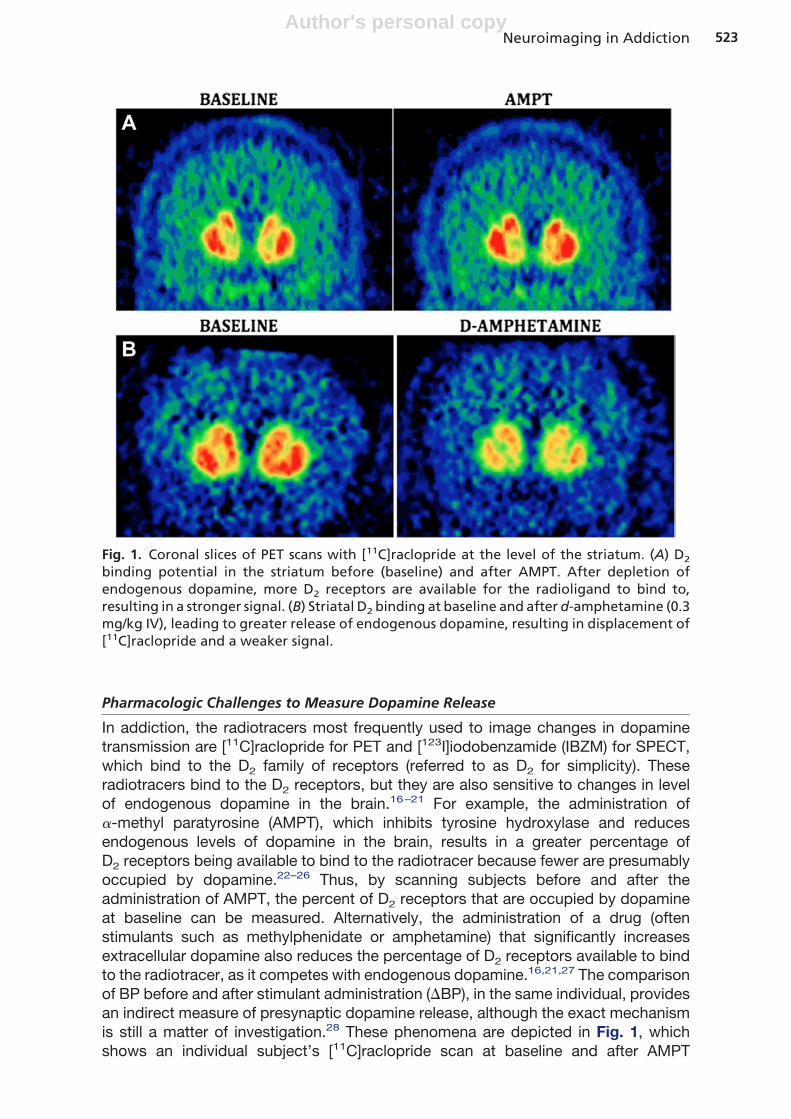
In addiction, the radiotracers most frequently used to image changes in dopaminetransmission are [11C]raclopride for PET and [123I]iodobenzamide (IBZM) for SPECT,which bind to the D2 family of receptors (referred to as D2 for simplicity). Theseradiotracers bind to the D2 receptors, but they are also sensitive to changes in levelof endogenous dopamine in the brain.16–21 For example, the administration of�-methyl paratyrosine (AMPT), which inhibits tyrosine hydroxylase and reducesendogenous levels of dopamine in the brain, results in a greater percentage ofD2 receptors being available to bind to the radiotracer because fewer are presumablyoccupied by dopamine.22–26 Thus, by scanning subjects before and after theadministration of AMPT, the percent of D2 receptors that are occupied by dopamineat baseline can be measured. Alternatively, the administration of a drug (oftenstimulants such as methylphenidate or amphetamine) that significantly increasesextracellular dopamine also reduces the percentage of D2 receptors available to bindto the radiotracer, as it competes with endogenous dopamine.16,21,27 The comparisonof BP before and after stimulant administration (�BP), in the same individual, providesan indirect measure of presynaptic dopamine release, although the exact mechanismis still a matter of investigation.28 These phenomena are depicted in Fig. 1, whichshows an individual subject’s [11C]raclopride scan at baseline and after AMPT
Fig. 1. Coronal slices of PET scans with [11C]raclopride at the level of the striatum. (A) D2
binding potential in the striatum before (baseline) and after AMPT. After depletion ofendogenous dopamine, more D2 receptors are available for the radioligand to bind to,resulting in a stronger signal. (B) Striatal D2 binding at baseline and after d-amphetamine (0.3mg/kg IV), leading to greater release of endogenous dopamine, resulting in displacement of[11C]raclopride and a weaker signal.
523Neuroimaging in Addiction

Author's personal copy
administration (see Fig. 1A, top panels) and another subject before and after theadministration of a psychostimulant (d-amphetamine, 0.3 mg/kg) (see Fig. 1B, bottompanels). The administration of AMPT results in an increase in D2 receptor BP (due toa decrease in endogenous dopamine), whereas the administration of a stimulantreduces [11C]raclopride BP (due to an increase in endogenous dopamine, see Fig. 1B).Using these methods, imaging studies have been able to investigate changes indopamine in addition to the changes in brain receptors that occur in addiction.
STIMULANT ABUSE AND DOPAMINE RECEPTORSCocaine
The drugs of abuse that have been studied most frequently with PET or SPECTimaging are the psychostimulants cocaine and methamphetamine. It was shown inthe early 1990s that cocaine dependence is associated with a decrease in D2 receptorbinding when compared to matched control subjects.29 Subsequent studies haveconsistently shown decreases in D2 receptor BP of 11% to 15% in cocaine-dependent individuals compared to control subjects.30–32 Two PET imaging studiesscanned cocaine-dependent subjects after 3 months of abstinence, and both showedpersistently lower D2 receptor BP with [11C]raclopride.30,33 These findings suggestthat the decrease in D2 receptor BP is either long lasting or not reversible. Importantly,an imaging study in rhesus monkeys exposed to cocaine showed that D2 receptoravailability is decreased by 15% to 20% already after 1 week of cocaine self-administration, and that this decrease can persist even up to 1 year of abstinence,suggesting that the decrease in D2 receptors is long lasting but reversible.34 Only oneimaging study has measured the D1 receptor family in cocaine dependence, and thisstudy showed no difference compared to control subjects.35 Thus, the change indopamine receptor availability associated with cocaine dependence appears to affectprimarily the D2 receptor family.
Methamphetamine
Imaging studies of the D2-receptor in methamphetamine dependence have shownsimilar results. Three studies found a decrease in striatal D2 receptor BP of about 10%to 15% in methamphetamine-dependent subjects compared to controls.36–38 Thesestudies also investigated the behavioral correlates of the decrease in BP. Lee andcolleagues37 showed an inverse relationship between BP and impulsiveness, in whichlower BP was associated with greater impulsivity, which is consistent with an imagingstudy in rodents.39 The study of Volkow and colleagues36 found that D2 receptor BPcorrelates with metabolic rate in the orbitofrontal cortex, which could reflect altera-tions in the dopamine-mediated striatal regulation of orbitofrontal activity via thestriato–thalamo–cortical pathways.
Dopamine Receptors in Other Addictions
The decrease in striatal D2-receptors was first described in cocaine abusers, andinitially it was thought to result from chronic cocaine exposure. However, imagingstudies have now demonstrated that decreased striatal D2 receptor binding occurs ina number of addictions, such as heroin,40 alcohol dependence,41–44 tobacco,45 andeven obesity.46,47 Thus, it is now thought that low D2 receptor binding serves as abiomarker for addictive behavior, and may reflect a propensity to depend onpharmacologic stimulation to experience reward.48,49 Depending on the addictivepotential of each abused substance, only a relatively small percentage of individualsexposed to drugs will become addicted (eg, 15% over 10 years of cocaine use; 10%
524 Urban & Martinez

Author's personal copy
lifetime risk for alcohol dependence50). Factors that determine who becomes ad-dicted include genetic (50% of risk), developmental (the risk is higher in adolescence),and environmental (eg, drug access, stress) risk factors.51
Low Dopamine Receptor Binding May Predispose to Addiction
Imaging studies have also been used to investigate the vulnerability to addiction. Innonhuman primates, social status has been shown to affect dopamine D2 receptor BPin the brain.52 Low status was associated with decreased D2 receptor BP comparedto high social status monkeys, and was also associated with an increased propensityto self-administer cocaine.52 Also in humans, one imaging study showed thatvolunteers with lower social status had lower values for [11C]raclopride BP comparedto subjects of high social status and support.53 Imaging studies have also investi-gated the effect of a family history of addiction and D2 receptors. Two studies haveshown that volunteers with a family history of addiction (cocaine and alcohol), who arenot addicted themselves, have higher D2 receptor binding compared to subjects withno family history.54 Because subjects with a strong family history of addiction wouldbe expected to have a higher risk for dependence, these findings suggest thatincreased D2 receptor BP may be protective.54 Lastly, Volkow and colleagues haveshown that high striatal D2 binding in healthy controls is predictive of an unpleasantreaction to the administration of intravenous methylphenidate, and that low D2
binding was associated with a pleasurable experience.55,56 Thus, taken togetherthese studies indicate that low D2 receptor binding is associated with increased socialstress, increased impulsivity, and an increased positive response to drugs of abuse,factors that have been shown to be risk factors for addiction.
STIMULANT ABUSE AND STRIATAL DOPAMINE TRANSMISSION
As described, PET or SPECT imaging with a D2 antagonist radiotracer and apharmacologic challenge that releases presynaptic dopamine (�BP) can be used toimage changes in the level of endogenous dopamine in the striatum. Brain imagingstudies in humans have shown that drug-induced increases in dopamine in thestriatum are associated with the subjective experience of reward, that is, theexperience of euphoria or feeling “high.”57 In addition, these studies have not beenlimited to the stimulant drugs. In humans, stimulants,58,59 nicotine,60 alcohol,61,62 andmarijuana63 increase dopamine in dorsal and ventral striatum, which is consistentwith animal studies showing that most drugs of abuse increase dopamine releasefrom the presynaptic neurons in the striatum, regardless of their primary mechanismof action.64 In addition, some of these studies have reported that participants whodisplay the greatest dopamine increases with the drug also report the most intense“high” or “euphoria.”57,59
These studies were performed in nonaddicted human volunteers, and support thehypothesis that increased striatal dopamine release is generally associated with thepositive and reinforcing effects of drug of abuse. However, imaging studies in chronicdrug users show that striatal dopamine release is altered in addiction. Volkow andcolleagues,31 using [11C]raclopride displacement to measure dopamine release,previously found that cocaine dependence is associated with blunted dopaminerelease in response to a stimulant challenge (intravenous methylphenidate) comparedto matched controls. In addition, the cocaine-dependent subjects also reported adecrease in the positive effects of the stimulant compared to the control subjects.Using SPECT and an amphetamine challenge (0.3 mg/kg IV), Malison and col-leagues65 performed a similar study in cocaine abusers and controls and reportedonly a 1% change in radiotracer binding in the cocaine abusers compared to a 10%
525Neuroimaging in Addiction

Author's personal copy
decrease in the control subjects. Lastly, Wu and colleagues66 used the levodopaanalogue 6-[18F]-fluoro-L-3,4-dihydroxyphenylalanine (FDOPA), which provides ameasure of presynaptic dopamine activity (ie, dopamine synthesis), to show thatcocaine-dependent subjects who had been abstinent 11 to 30 days had lower uptakecompared to controls. These findings taken together strongly support the hypothesisthat cocaine dependence is associated with a decrease in presynaptic dopaminerelease, measured as presynaptic dopamine release and presynaptic stores ofdopamine in the striatum.
Cocaine Self-Administration
Our group previously performed a similar study using PET to image dopamine releasein cocaine abusers. In this study, non-treatment–seeking cocaine-dependent sub-jects were scanned with [11C]raclopride and an amphetamine challenge to measurepresynaptic dopamine release.67 After the PET scans, the cocaine-dependent sub-jects performed cocaine self-administration sessions to investigate the relationshipbetween dopamine release and a laboratory model of cocaine-seeking behavior.Participants were given five choices between low-dose smoked cocaine and analternative positive reinforcer (money), and the choices were weighted toward themoney. As in previous studies, the cocaine abusers had blunted dopamine releasecompared to the controls subjects. However, this study also showed that, within thecocaine-abusing subjects, low stimulant-induced dopamine release in the ventralstriatum was associated with more choices for cocaine over money, and suggest thatlow dopamine release is associated with compulsive cocaine use.67
STIMULANT ABUSE, DOPAMINE TRANSMISSION, AND RESPONSE TO TREATMENTCocaine
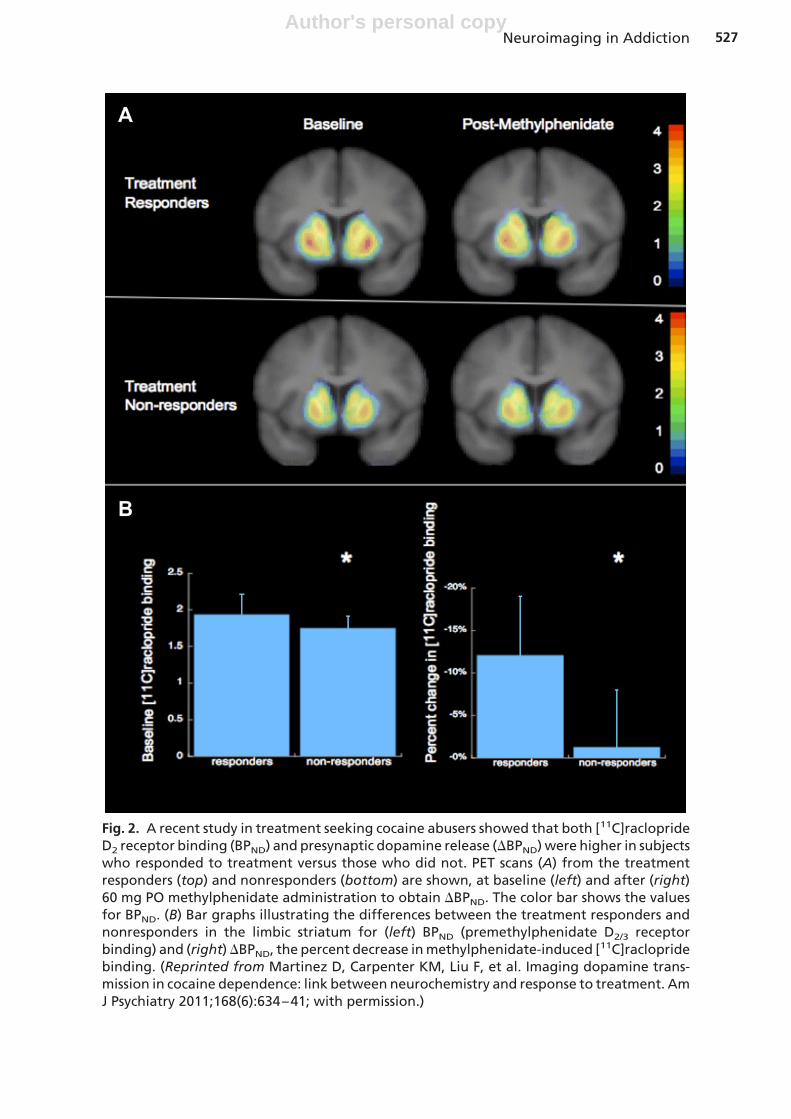
More recently, our group investigated this finding in the clinical setting.33 The goal ofthis study was to investigate whether blunted striatal dopamine transmission pre-dicted response to treatment that uses positive reinforcement to reduce cocaine use.Treatment-seeking cocaine-dependent subjects underwent PET scans using [11C]ra-clopride to image baseline dopamine D2 receptor binding and stimulant-inducedpresynaptic dopamine release. After the scans, the subjects were enrolled in 24weeks of treatment using contingency management combined with communityreinforcement, which is a behavioral treatment that uses monetary vouchers to induceabstinence from cocaine,68,69 and that is similar to the choice presented in thelaboratory in our previous study.67 The results of this study showed that both baselineD2 receptor BP and presynaptic dopamine release (�BP) were lower in the subjectswho relapsed compared to those who were able to respond to treatment (this findingis illustrated in Fig. 2). Importantly, the treatment responders did not differ from thecontrol comparison group in either outcome measure (D2 receptor BP or stimulant-induced �BP), suggesting that this group had intact striatal dopamine transmission.
Methamphetamine
A recent study by Wang and colleagues38 used [11C]raclopride and a stimulantchallenge (oral methylphenidate) to measure dopamine release in methamphetamineabusers and showed that it was blunted compared to controls, as reported previouslyin cocaine abusers. This study also followed the methamphetamine abusers for 9months at an outpatient drug rehabilitation program with an intensive behavioraltreatment and showed that response to treatment was associated with greaterdopamine release. In addition, the methamphetamine abusers who responded to
526 Urban & Martinez
Author's personal copy
Fig. 2. A recent study in treatment seeking cocaine abusers showed that both [11C]racloprideD2 receptor binding (BPND) and presynaptic dopamine release (�BPND) were higher in subjectswho responded to treatment versus those who did not. PET scans (A) from the treatmentresponders (top) and nonresponders (bottom) are shown, at baseline (left) and after (right)60 mg PO methylphenidate administration to obtain �BPND. The color bar shows the valuesfor BPND. (B) Bar graphs illustrating the differences between the treatment responders andnonresponders in the limbic striatum for (left) BPND (premethylphenidate D2/3 receptorbinding) and (right) �BPND, the percent decrease in methylphenidate-induced [11C]raclopridebinding. (Reprinted from Martinez D, Carpenter KM, Liu F, et al. Imaging dopamine trans-mission in cocaine dependence: link between neurochemistry and response to treatment. AmJ Psychiatry 2011;168(6):634–41; with permission.)
527Neuroimaging in Addiction

Author's personal copy
treatment did not differ from the controls in both PET outcome measures. Thesefindings are consistent with our study in cocaine abusers, and suggest that impaireddopamine transmission predicts a failure to respond to intensive behavioral treatmentin stimulant abusers.
Alcohol and Heroin
In addition to stimulant abuse, imaging studies using this technique have shown thatpresynaptic dopamine release is blunted in alcohol and heroin dependence.43,44,70
Thus, the decrease in presynaptic dopamine release measured with PET is notselective for cocaine dependence alone, and, like reduced baseline D2 receptorbinding, it appears to occur across addictions. However, the study by our group inheroin-dependent subjects also used a measure of drug-seeking behavior, andcorrelated the choice to self-administer heroin with the PET outcome measures.70 Inthis study there was no correlation between either outcome measure or the choice toself-administer drug, suggesting that dysregulation in dopamine transmission may besimilar across different addictions, but that the correlation between neurochemistryand drug-seeking behavior appears to vary with the particular drug of abuse.
Imaging Dopamine Transmission in Addiction: Clinical Correlates
Taken together, these studies provide some insight into the alterations in dopaminetransmission that occur in addiction. The findings in the preceding text show that theadministration of a number of drugs of abuse to control subjects produced ameasurable increase in presynaptic dopamine that can be measured with PETimaging. In addition, this increase in endogenous dopamine is consistent with animalstudies showing that most drugs of abuse increase striatal dopamine, which plays acrucial role in their reinforcing effects. However, in addicted individuals, the admin-istration of stimulants that are known to induce presynaptic dopamine release hasbeen shown to have little effect on the presynaptic dopamine neurons. A number ofthese studies show that addicted individuals have not only blunted dopamine releasebut also a reduction in the positive subjective effects of stimulant administration.These findings suggest that addiction is associated with a decrease in dopaminetransmission, and show that addiction extends far beyond simply the “high” thatdrugs induce.
Notably, both studies that have investigated the correlation between dopaminetransmission and treatment response show that the stimulant-dependent subjectswho failed to respond to treatment were those who had the greatest reduction inpresynaptic dopamine release. Importantly, both studies also found that there was nodifference in dopamine transmission in the stimulant-dependent subjects who re-sponded to treatment and the control group. These findings suggest that low striataldopamine transmission may occur in a subset of dependent individuals who are themost refractory to treatment.
Addiction: Compulsive Behavior Perpetuated by Low Dopamine Transmission
The role of dopamine in the striatum is among the most studied phenomena of thebrain. For almost a half-century, it has been shown that striatal dopamine is a crucialcomponent of reward and reward-based learning.71 The NAc, contained within theventral striatum in humans, serves as a hub of the brain’s reward pathways, and playsa central role in selecting adaptive, motivated behavior.72 In animal studies, deficits indopamine signaling in the NAc have been shown to impair operant conditioning,response inhibition, and behavioral flexibility.73 Lesions of the NAc result in a
528 Urban & Martinez

Author's personal copy
profound deficit in the animals’ ability to choose appropriately between two reinforc-ers: they impulsively and consistently choose a lesser reward over a delayedreinforcer of greater value.74 These findings suggest that dopamine signaling is criticalfor making the shift between competing reinforcers. Thus, in the setting of lowdopamine transmission a behavior that is ingrained is carried out, even in thepresence of an alternative reward of greater value. In other words, low dopaminetransmission in addiction appears to be consistent with a habit, rather than playing arole in mediating a “high.”
Treatment of Addiction Through Increasing Dopamine?
This information can then be used in treatment development. Previous clinical trialshave investigated the effect of medications that increase striatal dopamine transmis-sion as treatment for cocaine dependence, and some report success, whereas othersdo not (for review see Ref.75). One reason for this inconsistency may be thatmedications that are known to increase dopamine transmission in some addictedsubjects may have no effect in more refractory patients. Future treatments thatincrease striatal dopamine transmission, particularly in refractory patients, need to beexplored. This could possibly include medications that improve dopamine transmis-sion by increasing presynaptic stores in the striatum.76 Another approach may be toincrease dopamine transmission by targeting other receptor systems, such as the �,glutamate, or acetylcholine receptors (for review see Ref.77,78). In addition, it may benecessary to combine pharmacotherapies that increase striatal dopamine withbehavioral treatments that work to change behavior while dopamine is at work.
Additional Neurocircuitry
Based on this body of literature, the majority of functional imaging studies havefocused on investigating the dopamine receptors and dopamine transmission in thebasal ganglia. There is mounting evidence that additional circuits are engaged in thedevelopment of addiction, including79:
1. Mesolimbic dopamine system2. Ventral striatum3. Ventral striatum/dorsal striatum/thalamus circuits4. Dorsolateral frontal cortex/inferior frontal cortex/hippocampus circuits5. Extended amygdala.
In addition, although dopamine transmission in the nucleus accumbens does play akey role in mediating the positive reinforcement of drugs of abuse,6 other receptorsystem are also clearly affected in the development of addiction.80 At present,radiotracers are available to image the dopamine receptors and transporters, sero-tonin receptors/transporters, �-aminobutyric acid (GABA) and glutamate receptors,opioid receptors, and others.
IMAGING STUDIES OF PSYCHOSTIMULANT ABUSE: BEYOND D2 RECEPTORSDopamine Transporters in Stimulant Abuse
In addition to the dopamine D2 receptors and presynaptic dopamine release, imagingstudies have also investigated other receptors and transporters. One of the mostconsistent findings seen in imaging studies of methamphetamine dependence is adecrease in dopamine transporter (DAT) binding in the striatum. As shown in Table 1,seven published imaging studies in humans have shown a decrease in the DAT inmethamphetamine abusers compared to controls.81–87 In addition, the percent
529Neuroimaging in Addiction
Author's personal copy
decrease in the dopamine transporter appears to correlate with the duration ofabstinence before scanning (less abstinence, greater loss of DAT). These studies areconsistent with imaging studies in nonhuman primates, which also show thatmethamphetamine exposure results in a significant decrease in the DAT, which cantake months to years to recover.88–93 The mechanism behind this reduction intransporter density appears to involve a number of mechanisms, including oxidativestress, activation of transcription factors, DNA damage, excitotoxicity, and variousapoptotic pathways that result in frank injury to the dopamine neurons (for review seeRef.94).
Cocaine
Interestingly, PET studies of the DAT in cocaine dependence show that, unlikemethamphetamine, cocaine has a much less dramatic effect. In a study of cocaineabusers abstinent for 96 hours, Malison and colleagues95 showed a 20% increase inthe DAT compared to matched controls. However, imaging studies performed ateither 5 � 8 days or 42 � 7 days of abstinence show no difference in BP betweenhealthy controls and cocaine abusers.42,96 This suggests that the DAT is elevated invery early abstinence, but does not appear to differ from controls after this time point.Even though cocaine abusers and methamphetamine abusers both show blunteddopamine release, the mechanism mediating this difference likely varies between thetwo drugs of abuse. Thus, whereas the loss of dopamine transmission in metham-phetamine abuse results from injury to the dopamine neurons, blunted dopaminetransmission in cocaine abusers most likely results from other factors, such asdysregulation of the neurotransmitters that regulate dopamine transmission.
Table 1Published studies in human methamphetamine abusers listed in order of least to greatestpercent decrease in DAT reported
StudyNumber ofVolunteers
Abstinence Average(Range) % Decrease DAT
Johanson et al85 MA: 16HC: 18
3.4 y (3 m–18 y) Caudate, 16;putamen, 12
McCann et al86 MA: 6HC: 10
3.3 y (4 m–5 y) Putamen, 25;caudate, 23
McCann et al87 MA: 22HC: 17
2.4 y (8 m– �10 y) Caudate, 23;putamen, 17
Iyo et al84 MA: 11HC: 9
9 ma (0 m–18 m) Ventral striatum, 26; caudate/putamen, 20
Volkow et al83 MA: 15HC: 18
6 m (0.5 m–36 m) Caudate, 28;putamen, 21
Sekine et al81 MA: 11HC: 9
5.6 m (7d–1.5 y) Ventral striatum, 20; caudate/putamen, 20
Chou et al82 MA: 7HC: 7
Days (not specified) Whole striatum, 30
The average abstinence is the average time of abstinence before the PET scan reported in the study.Abstinence is provided as average and range.Abbreviations: d, days; HC, healthy control; m, months; MA, methamphetamine abuser; y, years.
a Estimated average; actual data not provided.
530 Urban & Martinez

Author's personal copy
Serotonin and Opiate Receptors
Although the majority of imaging studies in addiction have focused on the dopaminereceptors and dopamine transmission in the basal ganglia, there is mountingevidence that additional circuits and other neurotransmitters also play a crucial role inthe development of addiction.80 In cocaine dependence, imaging studies have alsoinvestigated alterations in the serotonin transporter and � opioid receptor. Cocainedirectly affects the serotonin system, and altering serotonin transmission in humansubjects affects craving and the subjective effects of cocaine.97–99 However, only oneimaging study has been performed in humans to measure the serotonin transporter,and this study showed that the transporter was increased in the brain stem in cocaineabusers who had been abstinent for 4 to 8 days.100 Three imaging studies havemeasured the � receptor using [11C]carfentanil in cocaine dependence. These studiesshow that cocaine dependence is associated with an increase in � receptor BP in theanterior cingulate, frontal and temporal cortex, caudate, and thalamus compared tocontrols, and remains elevated even after months of abstinence.101,102 This groupalso showed that regional brain � receptor binding in cocaine-dependent subjectswas a significant predictor of time to relapse to cocaine use.103
ALCOHOL DEPENDENCE
Ethanol affects numerous neuronal receptor and channels.104 As described briefly inthe preceding text, ethanol clearly has an effect on dopamine signaling, although theexact mechanism remains uncertain.104 Imaging studies have used the radiotracer[11C]raclopride to investigate whether acute ethanol administration to healthy humansproduces striatal dopamine release that can be measured with PET. Two studies haveshown that acute alcohol administration results in [11C]raclopride displacement61,62
while others have not.105,106 Importantly, a recent study showed that [11C]raclopridedisplacement induced by ethanol varied by gender: men had a much larger effectcompared to women, and percent dopamine release correlated with the positivesubjective effects in men, but not in women.62
Alcoholism Interferes with Dopamine Signaling
However, like stimulant dependence, studies in alcohol-dependent subjects showthat striatal dopamine signaling is impaired once dependence sets in. Several PETand SPECT studies have measured striatal D2 receptor BP in alcohol depen-dence,41,43,44,107–111 and seven of these show that D2 receptor BP is decreasedcompared to that in matched controls (two show nonsignificant decreases). Asdescribed in the preceding text, studies imaging stimulant-induced dopamine releasehave shown this to be blunted in alcohol-dependent subjects compared to con-trols.43,44 Studies using alcohol as a challenge to release dopamine have beenperformed only in healthy controls but not in alcohol-dependent subjects. Otherparameters of dopamine transmission in alcohol dependence have been studied withimaging, overall supporting the hypothesis that striatal dopamine signaling is impairedin alcohol dependence (for review see Ref.112).
Other Neurotransmitter Systems Affected in Alcoholism: Serotonin
The other receptor systems that have been imaged in alcohol dependence include theserotonin, opiate, and GABA receptor systems. Imaging studies of the serotoninsystem have focused on the serotonin transporter, given the important role of theserotonin system in vulnerability to alcohol-induced aggression.113 Overall, the resultsof these studies are mixed in that some studies have reported a decrease in the
531Neuroimaging in Addiction

Author's personal copy
serotonin transporter and other studies have not.114–118 One of the studies alsoinvestigated the serotonin 1A receptor,118 but showed no difference between thealcohol-dependent and control subjects, while another study imaged the serotonin 1Breceptors and found an increase in binding in the ventral striatum of alcohol-dependent subjects compared with controls.119 Given the importance of the sero-tonin system in alcohol dependence, the fact that the findings from these are notconsistent is surprising, particularly given that the imaging studies of dopamine arevery consistent. One possibility is that the alterations in dopamine signaling reportedin alcohol dependence are not specific to alcoholism (see earlier), but belie avulnerability to addiction. In addition, it appears from the imaging studies of theserotonin system in alcoholism that differences in gender, genotype, and aggressionmay explain why there is less uniformity in these results.112,113 Thus, further imagingstudies of the serotonin system in alcohol dependence should be performed tounderstand better how these factors affect serotonin transmission and play a role inthe development of alcoholism.
Opiate Receptors
The other neurotransmitter systems that play a crucial role in alcohol dependenceare the opiate receptors and GABA. Several studies have measured the � receptorin alcohol dependence using the � receptor selective radiotracer [11C]carfentanil.An early study in eight alcohol-dependent men after 4 days of abstinence reporteda decrease in � receptor BP in the prefrontal cortex compared to controls, with nodifferences in other cortical regions.120 However, a subsequent study of 25alcohol-dependent subjects scanned after weeks of abstinence showed thatalcohol dependence was associated with an increase in � receptor BP in theventral striatum compared to the control subjects, with no difference in other brainregions studied.121 These findings are in agreement with another imaging study in20 alcohol-dependent subjects showing significantly higher � receptor BP com-pared to controls in several brain regions including the ventral striatum.122 Inaddition, there was an inverse relationship between [11C]carfentanil BPND andcraving.122
GABA
With respect to the GABA system, imaging studies have focused on measuring theGABAA receptor, given the direct role it plays in ethanol intoxication and withdrawal.Seven PET and SPECT studies have been conducted to look at this receptor inalcohol dependence. Three showed that alcohol dependence is associated with adecrease in the GABAA receptor binding in the frontal cortex, anterior cingulate,cerebellum, and parietal and temporal cortices.123–125 In contrast, two studiesreported no difference in GABAA receptor binding between alcohol-dependentsubjects and healthy controls,126,127 and two studies showed an increase in GABAA
receptor binding in alcohol-dependent subjects compared to controls.128,129 Animportant issue with respect to these discrepancies is not only the number of subjectsincluded (some studies had a very small cohort) but also time of abstinence to thetime of scanning. The study of Staley and colleagues129 imaged early and later inabstinence and showed that radiotracer binding at the GABAA receptor varies withduration of abstinence and likely shifts across the different stages of alcoholdependence (for review see Ref.130).
532 Urban & Martinez

Author's personal copy
CANNABIS DEPENDENCETHC Induces Dopamine Release
Four imaging studies have examined the effect of �9-THC on dopamine release invivo, including an anecdotal case report.63,131–133 Unlike studies using a stimulant toproduce dopamine release in the brain, the studies using THC as a challenge torelease dopamine show very different results. In one study, THC inhalation was foundto reduce [11C]raclopride binding in the ventral striatum and the precommissuraldorsal putamen compared to placebo, consistent with an increase in dopamine levelsin these regions in a small pilot study.63 However, a study using the syntheticcannabinoid dronabinol as a challenge reported no significant difference in THC-induced [11C]raclopride binding (but an increase in psychosis-like symptoms).132 Inaddition, Barkus and colleagues compared intravenous THC to placebo in 11 healthymen using SPECT with [123I]IBZM, and found no significant change in the caudate orputamen.133 Overall, these studies suggest that there may be a small effect of acuteTHC on dopamine release in the human striatum, but that this effect is difficult tomeasure. Given that the increase in striatal dopamine may be slightly larger with astimulant challenge compared to acute THC inhalation,134,135 it is possible that theeffect of chronic THC use on dopamine is present, but not large enough to bemeasurable with these imaging methods.
D2 Receptors Are Not Affected in Cannabis Dependence
As described, the abuse of most drugs of abuse (cocaine, methamphetamine,alcohol, heroin, and nicotine) has been shown to be associated with a reduction in D2
receptor binding. Thus, one would expect the same finding in cannabis abuse.However, imaging studies investigating the dopaminergic system in chronic cannabisusers have not seen any difference in [11C]raclopride BPND in chronic cannabis-dependent subjects compared to controls.134,136,137 Sevy and colleagues 136 re-ported that although cannabis-dependent subjects had lower normalized glucosemetabolism in the right orbitofrontal cortex and bilateral putamen with [18F]FDG, therewas no difference in striatal D2 receptor availability compared to controls. One otherstudy retrospectively evaluated a data set of [11C]raclopride scans of volunteers whohad also reported cannabis use and compared D2 binding to data from a controlsample without any reported use. No difference in D2 binding was found, or anycorrelation with lifetime frequency of use and D2 binding potential.138
Is Dopamine Transmission Affected by Chronic Cannabis Use?
Similarly, imaging studies in addiction to cocaine, methamphetamine, heroin, andalcohol dependence are associated with a reduction in presynaptic dopaminerelease. Only one imaging study to date has evaluated changes in dopamine releasein addition to D2 receptor density in chronic, recently abstinent, cannabis users withthe amphetamine challenge paradigm and [11C]raclopride, and found no difference tohealthy controls.134 Subjects in our sample, however, reported only mild to moderatecannabis use, while meeting Diagnostic and Statistical Manual of Mental Disorders,4th ed. (DSM-IV-TR) criteria for dependence. It is conceivable that more severecannabis use may lead to measurable changes in the dopaminergic system. Inter-estingly, dopamine release in the associative striatum correlated positively with age atonset of the drug use, providing further evidence that the adolescent brain may bemore vulnerable to chronic effects of the drug.
533Neuroimaging in Addiction

Author's personal copy
Endogenous Cannabinoid Receptors
The endocannabinoid system, however, may be more directly affected, and imagingstudies have recently become possible with the development of the inverse agonistradiotracer [18F]FMPEP-d2 [(3R,5R)-5-(3-[18F]Fluoromethoxy-d2)phenyl)-3-((R)-1-phenyl-ethylamino)-1-(4-trifluoromethyl-phenyl)-pyrrolidin-2-one]. A recent study byHirvonen and colleagues138 demonstrated a reversible and regionally selectivedownregulation of brain cannabinoid CB1 receptors in chronic, heavy cannabis users.Downregulation of CB1 receptor correlated with the years of cannabis smoking, waslimited to cortical regions, and returned to normal levels after 1 month of monitoredabstinence from the drug. Downregulation of endocannabinoid receptors may con-tribute to tolerance to cannabis, while the dopaminergic system plays a secondaryrole, but further studies are needed to understand the interplay better.
SUMMARY
Neuroimaging studies have been crucial in understanding changes in the variousneurotransmitter systems implicated in addiction in the living human brain. Predom-inantly reduced striatal dopamine transmission appears to play an important role inpsychostimulant, alcohol and heroin addiction, while addiction to cannabis may bemediated primarily by the endocannabinoid system. However, the study of otherneurotransmitter systems likely involved in addiction, for example glutamate, hasbeen limited by the number and quality of available radiotracers, and data on changesin these systems in the most common addictions are emerging only now. Furtherstudies are needed to understand fully how the interplay of various neurotransmittersystems contributes to addiction and to ultimately help to develop more effectivetreatment approaches.
REFERENCES
1. Koob GF. Hedonic homeostatic dysregulation as a driver of drug-seeking behavior.Drug Discov Today Dis Models 2008;5(4):207–15.
2. Carlezon WA Jr, Wise RA. Rewarding actions of phencyclidine and related drugs innucleus accumbens shell and frontal cortex. J Neurosci 1996;16(9):3112–22.
3. Devane WA, Axelrod J. Enzymatic synthesis of anandamide, an endogenous ligandfor the cannabinoid receptor, by brain membranes. Proc Natl Acad Sci U S A1994;91(14):6698–701.
4. Vogel Z, Barg J, Levy R, et al. Anandamide, a brain endogenous compound,interacts specifically with cannabinoid receptors and inhibits adenylate cyclase. JNeurochem 1993;61(1):352–5.
5. Wise RA, Rompre PP. Brain dopamine and reward. Annu Rev Psychol 1989;40:191–225.
6. Koob GF, Le Moal M. Drug addiction, dysregulation of reward, and allostasis.Neuropsychopharmacology 2001;24(2):97–129.
7. Robinson TE, Berridge KC. Addiction. Annu Rev Psychol 2003;54:25–53.8. Koob GF. Neural mechanisms of drug reinforcement. Ann N Y Acad Sci 1992;654:
171–91.9. Wise RA. The role of reward pathways in the development of drug dependence.
Pharmacol Ther 1987;35(1–2):227–63.10. Wise RA. Roles for nigrostriatal—not just mesocorticolimbic—dopamine in reward
and addiction. Trends Neurosci 2009;32(10):517–24.11. Schultz W. Multiple dopamine functions at different time courses. Annu Rev Neurosci
2007;30:259–88.
534 Urban & Martinez

Author's personal copy
12. Grace AA. The tonic/phasic model of dopamine system regulation and its implica-tions for understanding alcohol and psychostimulant craving. Addiction 2000;95(Suppl 2):S119–28.
13. Carson RE. Parameters estimation in positron emission tomography, in positronemission tomography: principles and applications for the brain and the heart. In:Phelps ME, Mazziotta JC, Schelbert HR, editors. New York: Raven Press; 1986. p.347–90.
14. Slifstein M, Laruelle M. Models and methods for derivation of in vivo neuroreceptorparameters with PET and SPECT reversible radiotracers. Nucl Med Biol 2001;28(5):595–608.
15. Innis RB, Cunningham VJ, Delforge J, et al. Consensus nomenclature for in vivoimaging of reversibly binding radioligands. J Cereb Blood Flow Metab 2007;27(9):1533–9.
16. Breier A, Su TP, Saunders R, et al. Schizophrenia is associated with elevatedamphetamine-induced synaptic dopamine concentrations: evidence from anovel positron emission tomography method. Proc Natl Acad Sci U S A 1997;94(6):2569 –74.
17. Dewey SL, Smith GS, Logan J, et al. Striatal binding of the PET ligand 11C-racloprideis altered by drugs that modify synaptic dopamine levels. Synapse 1993;13(4):350–6.
18. Innis RB, Malison RT, al-Tikriti M, et al. Amphetamine-stimulated dopamine releasecompetes in vivo for [123I]IBZM binding to the D2 receptor in nonhuman primates.Synapse 1992;10(3):177–84.
19. Laruelle M, Iyer RN, al-Tikriti M, et al. Microdialysis and SPECT measurements ofamphetamine-induced dopamine release in nonhuman primates. Synapse 1997;25(1):1–14.
20. Laruelle M, Abi-Dargham A, van Dyck CH, et al. SPECT imaging of striatal dopaminerelease after amphetamine challenge. J Nucl Med 1995;36:1182–90.
21. Volkow ND, Wang G, Fowler RS, et al. Therapeutic doses of oral methylphenidatesignificantly increase extracellular dopamine in the human brain. J Neurosci 2001;21(2):RC121.
22. Martinez D, Greene K, Broft A, et al. Lower level of endogenous dopamine in patientswith cocaine dependence: findings from PET imaging of D(2)/D(3) receptors follow-ing acute dopamine depletion. Am J Psychiatry 2009;166(10):1170–7.
23. Cropley VL, Innis RB, Nathan PJ, et al. Small effect of dopamine release and no effectof dopamine depletion on [18F]fallypride binding in healthy humans. Synapse 2008;62(6):399–408.
24. Riccardi P, Baldwin R, Salomon R, et al. Estimation of baseline dopamine D2receptor occupancy in striatum and extrastriatal regions in humans with positronemission tomography with [18F] fallypride. Biol Psychiatry 2008;63(2):241–4.
25. Verhoeff NP, Hussey D, Lee M, et al. Dopamine depletion results in increasedneostriatal D(2), but not D(1), receptor binding in humans. Mol Psychiatry 2002;7(3):233, 322–8.
26. Laruelle M, D’Souza CD, Baldwin RM, et al. Imaging D2 receptor occupancy byendogenous dopamine in humans. Neuropsychopharmacology 1997;17:162–74.
27. Laruelle M, Al-Tikrity M, Abi-Dargham A, et al. D-amphetamine-induced reduction ofdopamine D2 availability in primates: comparison between [123I]IBF and [123I]IBZM.Soc Neurosci Abstr 1994;20:644.
28. Laruelle M. Imaging synaptic neurotransmission with in vivo binding competitiontechniques: a critical review. J Cereb Blood Flow Metab 2000;20(3):423–51.
535Neuroimaging in Addiction

Author's personal copy
29. Volkow ND, Fowler JS, Wolf AP, et al. Effects of chronic cocaine abuse onpostsynaptic dopamine receptors. Am J Psychiatry 1990;147(6):719–24.
30. Volkow ND, Fowler JS, Wang GJ, et al. Decreased dopamine D2 receptor availabilityis associated with reduced frontal metabolism in cocaine abusers. Synapse 1993;14(2):169–77.
31. Volkow ND, Wang GJ, Fowler JS, et al. Decreased striatal dopaminergic respon-siveness in detoxified cocaine-dependent subjects. Nature 1997;386:830–3.
32. Martinez D, Broft A, Foltin RW, et al. Cocaine dependence and d2 receptoravailability in the functional subdivisions of the striatum: relationship with cocaine-seeking behavior. Neuropsychopharmacology 2004;29(6):1190–202.
33. Martinez D, Carpenter KM, Liu F, et al. Imaging dopamine transmission in cocainedependence: link between neurochemistry and response to treatment. Am J Psy-chiatry 2011;168(6):634–41.
34. Nader MA, Morgan D, Gage HD, et al. PET imaging of dopamine D2 receptors duringchronic cocaine self-administration in monkeys. Nat Neurosci 2006;9(8):1050–6.
35. Martinez D, Slifstein M, Narendran R, et al. Dopamine D1 receptors in cocainedependence measured with PET and the choice to self-administer cocaine. Neuro-psychopharmacology 2009;34(7):1774–82.
36. Volkow ND, Change L, Wang GJ, et al. Low level of brain dopamine D2 receptors inmethamphetamine abusers: association with metabolism in the orbitofrontal cortex.Am J Psychiatry 2001;158(12):2015–21.
37. Lee B, London ED, Poldrack RA, et al. Striatal dopamine d2/d3 receptor availabilityis reduced in methamphetamine dependence and is linked to impulsivity. J Neurosci2009;29(47):14734–40.
38. Wang GJ, Smith L, Volkow ND, et al. Decreased dopamine activity predicts relapsein methamphetamine abusers. Mol Psychiatry 2011. DOI: 10.1038/mp.2011.86.[Epub ahead of print].
39. Dalley JW, Fryer TD, Brichard L, et al. Nucleus accumbens D2/3 receptors predicttrait impulsivity and cocaine reinforcement. Science 2007;315(5816):1267–70.
40. Wang GJ, Volkow ND, Fowler JS, et al. Dopamine D2 receptor availability inopiate-dependent subjects before and after naloxone-precipitated withdrawal. Neu-ropsychopharmacology 1997;16(2):174–82.
41. Hietala J, West C, Swälahti E, et al. Striatal D2 dopamine receptor binding charac-teristics in vivo in patients with alcohol dependence. Psychopharmacology [Berl]1994;116(3):285–90.
42. Volkow ND, Wang GJ, Fowler JS, et al. Decreases in dopamine receptors but not indopamine transporters in alcoholics. Alcohol Clin Exp Res 1996;20(9):1594–8.
43. Martinez D, Gil R, Slifstein M, et al. Alcohol dependence is associated with blunteddopamine transmission in the ventral striatum. Biol Psychiatry 2005;58(10):779–86.
44. Volkow ND, Wang GJ, Telang F, et al. Profound decreases in dopamine release instriatum in detoxified alcoholics: possible orbitofrontal involvement. J Neurosci2007;27(46):12700–6.
45. Fehr C, Yakushev I, Hohmann N, et al. Association of low striatal dopamine d2receptor availability with nicotine dependence similar to that seen with other drugs ofabuse. Am J Psychiatry 2008;165(4):507–14.
46. Wang GJ, Volkow ND, Logan J, et al. Brain dopamine and obesity. Lancet 2001;357(9253):354–7.
47. Volkow ND, Wang GJ, Fowler JS, et al. Overlapping neuronal circuits in addictionand obesity: evidence of systems pathology. Philos Trans R Soc Lond B Biol Sci2008;363(1507):3191–200.
536 Urban & Martinez

Author's personal copy
48. Volkow ND, Fowler JS, Wang GJ. Role of dopamine in drug reinforcement andaddiction in humans: results from imaging studies. Behav Pharmacol 2002;13(5–6):355–66.
49. Melis M, Spiga S, Diana M. The dopamine hypothesis of drug addiction: hypodop-aminergic state. Int Rev Neurobiol 2005;63:101–54.
50. Wagner FA, Anthony JC. From first drug use to drug dependence; developmentalperiods of risk for dependence upon marijuana, cocaine, and alcohol. Neuropsycho-pharmacology 2002;26(4):479–88.
51. Volkow N, Li TK. The neuroscience of addiction. Nat Neurosci 2005;8(11):1429–30.52. Morgan D, Grant KA, Gage HD, et al. Social dominance in monkeys: dopamine D2
receptors and cocaine self-administration. Nat Neurosci 2002;5(2):169–74.53. Martinez D, Orlowska D, Narendran R, et al. Dopamine type 2/3 receptor availability
in the striatum and social status in human volunteers. Biol Psychiatry 2010;67(3):275–8.
54. Volkow ND, Wang GJ, Begleiter H, et al. High levels of dopamine D2 receptors inunaffected members of alcoholic families: possible protective factors. Arch GenPsychiatry 2006;63(9):999–1008.
55. Volkow ND, Wang GJ, Fowler JS, et al. Prediction of reinforcing responses topsychostimulants in humans by brain dopamine D2 receptor levels. Am J Psychiatry1999;156(9):1440–3.
56. Volkow ND, Wang GJ, Fowler JS, et al. Brain DA D2 receptors predict reinforcingeffects of stimulants in humans: replication study. Synapse 2002;46(2):79–82.
57. Volkow ND, Wang GJ, Fowler JS, et al. Relationship between psychostimulant-induced “high” and dopamine transporter occupancy. Proc Natl Acad Sci U S A1996;93(19):10388–92.
58. Volkow ND, Wang GJ, Fowler JS, et al. Reinforcing effects of psychostimulants inhumans are associated with increases in brain dopamine and occupancy of D(2)receptors. J Pharmacol Exp Ther 1999;291(1):409–15.
59. Drevets WC, Gautier C, Price JC, et al. Amphetamine-induced dopamine release inhuman ventral striatum correlates with euphoria. Biol Psychiatry 2001;49(2):81–96.
60. Brody AL, Mandekern MA, Olmstead RE, et al. Ventral striatal dopamine release inresponse to smoking a regular vs a denicotinized cigarette. Neuropsychopharma-cology 2009;34(2):282–9.
61. Boileau I, Assaad JM, Pihl RO, et al. Alcohol promotes dopamine release in thehuman nucleus accumbens. Synapse 2003;49(4):226–31.
62. Urban NB, Kegeles LS, Slifstein M, et al. Sex differences in striatal dopamine releasein young adults after oral alcohol challenge: a positron emission tomography imagingstudy with [11C]raclopride. Biol Psychiatry 2010;68(8):689–96.
63. Bossong MG, van Berckel BN, Boellaard R, et al. Delta 9-tetrahydrocannabinolinduces dopamine release in the human striatum. Neuropsychopharmacology 2009;34(3):759–66.
64. Di Chiara G, Imperato A. Drugs abused by humans preferentially increase synapticdopamine concentrations in the mesolimbic system of freely moving rats. Proc NatlAcad Sci U S A 1988;85(14):5274–8.
65. Malison RT, Mechanic KY, Klummp H, et al. Reduced amphetamine-stimulateddopamine release in cocaine addicts as measured by [123I]IBZM SPECT. J Nucl Med1999;40(5 Suppl):110P.
66. Wu JC, Bell K, Najafi A, et al. Decreasing striatal 6-FDOPA uptake with increasingduration of cocaine withdrawal. Neuropsychopharmacology 1997;17(6):402–9.
537Neuroimaging in Addiction

Author's personal copy
67. Martinez D, Narendran R, Fottin RW, et al. Amphetamine-induced dopamine re-lease: markedly blunted in cocaine dependence and predictive of the choice toself-administer cocaine. Am J Psychiatry 2007;164(4):622–9.
68. Higgins ST, Budney AJ, Bickel WK, et al. Incentives improve outcome inoutpatient behavioral treatment of cocaine dependence. Arch Gen Psychiatry1994;51(7):568 –76.
69. Higgins ST, Sigmon SC, Wong CJ, et al. Community reinforcement therapy forcocaine-dependent outpatients. Arch Gen Psychiatry 2003;60(10):1043–52.
70. Martinez D, Saccone PA, Liu F, et al. Deficits in dopamine D(2) receptors andpresynaptic dopamine in heroin dependence: commonalities and differences withother types of addiction. Biol Psychiatry 2012;71(3):192–8.
71. Wise RA. Addictive drugs and brain stimulation reward. Annu Rev Neurosci 1996;19:319–40.
72. Sesack SR, Grace AA. Cortico-basal ganglia reward network: microcircuitry. Neu-ropsychopharmacology 2010;35(1):27–47.
73. Goto Y, Grace AA. Limbic and cortical information processing in the nucleusaccumbens. Trends Neurosci 2008;31(11):552–8.
74. Cardinal RN, Pennicott DR, Sugathapala CL, et al. Impulsive choice induced in ratsby lesions of the nucleus accumbens core. Science 2001;292(5526):2499–501.
75. Grabowski J, Shearer J, Merrill J, et al. Agonist-like, replacement pharmacotherapyfor stimulant abuse and dependence. Addict Behav 2004;29(7):1439–64.
76. Schmitz JM, Mooney ME, Moeller FG, et al. Levodopa pharmacotherapy for cocainedependence: choosing the optimal behavioral therapy platform. Drug Alcohol De-pend 2008;94(1–3):142–50.
77. Shippenberg TS, Zapata A, Chefer VI. Dynorphin and the pathophysiology of drugaddiction. Pharmacol Ther 2007;116(2):306–21.
78. Lester DB, Rogers TD, Blaha CD. Acetylcholine-dopamine interactions in the patho-physiology and treatment of CNS disorders. CNS Neurosci Ther 2010;16(3):137–62.
79. Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology2010;35(1):217–38.
80. Nestler EJ. Is there a common molecular pathway for addiction? Nat Neurosci2005;8(11):1445–9.
81. Sekine Y, Iyo M, Ouchi Y, et al. Methamphetamine-related psychiatric symptomsand reduced brain dopamine transporters studied with PET. Am J Psychiatry2001;158(8):1206–14.
82. Chou YH, Huang WS, Su TP, et al. Dopamine transporters and cognitive function inmethamphetamine abuser after a short abstinence: a SPECT study. Eur Neuropsy-chopharmacol 2007;17(1):46–52.
83. Volkow ND, Chang L, Wang GJ, et al. Association of dopamine transporter reductionwith psychomotor impairment in methamphetamine abusers. Am J Psychiatry2001;158(3):377–82.
84. Iyo M, Sekine Y, Mori N. Neuromechanism of developing methamphetamine psy-chosis: a neuroimaging study. Ann N Y Acad Sci 2004;1025:288–95.
85. Johanson CE, Frey KA, Lundahl LH, et al. Cognitive function and nigrostriatalmarkers in abstinent methamphetamine abusers. Psychopharmacology [Berl] 2006;185(3):327–38.
86. McCann U, Wong DF, Yokoi F, et al. Reduced striatal dopamine transporterdensity in abstinent methamphetamine and methcathinone users: evidence frompositron emission tomography studies with [11C]WIN-35,428. J Neurosci 1998;18(20):8417–22.
538 Urban & Martinez

Author's personal copy
87. McCann UD, Kuwabara H, Kumar A, et al. Persistent cognitive and dopaminetransporter deficits in abstinent methamphetamine users. Synapse 2008;62(2):91–100.
88. Melega WP, Lacan G, Desalles AA, et al. Long-term methamphetamine-induceddecreases of [(11)C]WIN 35,428 binding in striatum are reduced by GDNF: PETstudies in the vervet monkey. Synapse 2000;35(4):243–9.
89. Hashimoto K, Tsukada H, Nishiyama S, et al. Protective effects of N-acetyl-L-cysteine on the reduction of dopamine transporters in the striatum of mon-keys treated with methamphetamine. Neuropsychopharmacology 2004;29(11):2018 –23.
90. Hashimoto K, Tsukada H, Nishiyama S, et al. Protective effects of minocycline on thereduction of dopamine transporters in the striatum after administration of metham-phetamine: a positron emission tomography study in conscious monkeys. BiolPsychiatry 2007;61(5):577–81.
91. Villemagne V, Yuan J, Wong DF, et al. Brain dopamine neurotoxicity in baboonstreated with doses of methamphetamine comparable to those recreationally abusedby humans: evidence from [11C]WIN-35,428 positron emission tomography studiesand direct in vitro determinations. J Neurosci 1998;18(1):419–27.
92. Harvey DC, Lacan G, Tanious SP, et al. Recovery from methamphetamine inducedlong-term nigrostriatal dopaminergic deficits without substantia nigra cell loss. BrainRes 2000;871(2):259–70.
93. Volkow ND, Chang L, Wang GJ, et al. Loss of dopamine transporters in metham-phetamine abusers recovers with protracted abstinence. J Neurosci 2001;21(23):9414–8.
94. Cadet JL, Krasnova IN. Molecular bases of methamphetamine-induced neurode-generation. Int Rev Neurobiol 2009;88:101–19.
95. Malison RT, Best SE, van Dyck CH, et al. Elevated striatal dopamine transportersduring acute cocaine abstinence as measured by [123I] beta-CIT SPECT. Am JPsychiatry 1998;155(6):832–4.
96. Wang GJ, Volkow ND, Fowler JS, et al. Cocaine abusers do not show loss ofdopamine transporters with age. Life Sci 1997;61(11):1059–65.
97. Aronson SC, Black JE, McDougle CJ, et al. Serotonergic mechanisms of cocaineeffects in humans. Psychopharmacology [Berl] 1995;119(2):179–85.
98. Satel SL, Krystal JH, Delgado PL, et al. Tryptophan depletion and attenuation ofcue-induced craving for cocaine. Am J Psychiatry 1995;152(5):778–83.
99. Walsh SL, Preston KL, Sullivan JT, et al. Fluoxetine alters the effects of intravenouscocaine in humans. J Clin Psychopharmacol 1994;14(6):396–407.
100. Jacobsen LK, Staley JK, Malison RT, et al. Elevated central serotonin transporterbinding availability in acutely abstinent cocaine-dependent patients. Am J Psychiatry2000;157(7):1134–40.
101. Zubieta JK, Gorelick DA, Stauffer R, et al. Increased mu opioid receptor bindingdetected by PET in cocaine-dependent men is associated with cocaine craving. NatMed 1996;2(11):1225–9.
102. Gorelick DA, Kim YK, Bencherif B, et al. Imaging brain mu-opioid receptors inabstinent cocaine users: time course and relation to cocaine craving. Biol Psychiatry2005;57(12):1573–82.
103. Gorelick DA, Kim YK, Bencherif B, et al. Brain mu-opioid receptor binding: relation-ship to relapse to cocaine use after monitored abstinence. Psychopharmacology[Berl] 2008;200(4):475–86.
104. Sulzer D. How addictive drugs disrupt presynaptic dopamine neurotransmission.Neuron 2011;69(4):628–49.
539Neuroimaging in Addiction

Author's personal copy
105. Yoder KK, Constantinescu CC, Kareken DA, et al. Heterogeneous effects of alcoholon dopamine release in the striatum: a PET study. Alcohol Clin Exp Res 2007;31(6):965–73.
106. Salonen I, Hietala J, Laihinen A, et al. A PET study on the acute effect of ethanol onstriatal D2 dopamine receptors with [11C]raclopride in healthy males. Hum Psycho-pharmacol Clin Exp 1997;12(2):145–52.
107. Volkow ND, Wang G-J, Fowler JS, et al. Decreases in dopamine receptors but notin dopamine transporters in alcoholics. J Nucl Med 1996;37:33P.
108. Volkow ND, Wang GJ, Maynard L, et al. Effects of alcohol detoxification ondopamine D2 receptors in alcoholics: a preliminary study. Psychiatry Res 2002;116(3):163–72.
109. Heinz A, Siessmeier T, Wrase J, et al. Correlation between dopamine D(2) receptorsin the ventral striatum and central processing of alcohol cues and craving. Am JPsychiatry 2004;161(10):1783–9.
110. Rominger A, Cumming P, Xiong G, et al. [(18) F]fallypride PET measurement ofstriatal and extrastriatal dopamine D(2/3) receptor availability in recently abstinentalcoholics. Addict Biol 2012;17(2):490–503.
111. Heinz A, Siessmeier T, Wrase J, et al. Correlation of alcohol craving with striataldopamine synthesis capacity and D2/3 receptor availability: a combined [18F]DOPAand [18F]DMFP PET study in detoxified alcoholic patients. Am J Psychiatry 2005;162(8):1515–20.
112. Martinez D, Kim JH, Krystal J, et al. Imaging the neurochemistry of alcohol andsubstance abuse. Neuroimag Clin N Am 2007;17(4):539–55, x.
113. Heinz AJ, Beck A, Meyer-Lindenberg A, et al. Cognitive and neurobiological mech-anisms of alcohol-related aggression. Nat Rev Neurosci 2011;12(7):400–13.
114. Heinz A, Jones DW, Bissette G, et al. Relationship between cortisol andserotonin metabolites and transporters in alcoholism. Pharmacopsychiatry2002;35(4):127–34.
115. Heinz A, Jones DW, Mazzanti C, et al. A relationship between serotonin transportergenotype and in vivo protein expression and alcohol neurotoxicity. Biol Psychiatry2000;47(7):643–9.
116. Szabo Z, Owonikoko T, Peyrot M, et al. Positron emission tomography imaging ofthe serotonin transporter in subjects with a history of alcoholism. Biol Psychiatry2004;55(7):766–71.
117. Brown AK, George DT, Fujita M, et al. PET [11C]DASB imaging of serotonintransporters in patients with alcoholism. Alcohol Clin Exp Res 2007;31(1):28–32.
118. Martinez D, Slifstein M, Gil R, et al. Positron emission tomography imaging of theserotonin transporter and 5-HT(1A) receptor in alcohol dependence. Biol Psychiatry2009;65(2):175–80.
119. Hu J, Henry S, Gallezot JD, et al. Serotonin 1B receptor imaging in alcoholdependence. Biol Psychiatry 2010;67(9):800–3.
120. Bencherif B, Wand GS, McCaul ME, et al. Mu-opioid receptor binding measured by[11C]carfentanil positron emission tomography is related to craving and mood inalcohol dependence. Biol Psychiatry 2004;55(3):255–62.
121. Heinz A, Reimold M, Wrase J, et al. Correlation of stable elevations in striatalmu-opioid receptor availability in detoxified alcoholic patients with alcohol craving: apositron emission tomography study using carbon 11-labeled carfentanil. Arch GenPsychiatry 2005;62(1):57–64.
122. Weerts EM, Wand GS, Kuwabara H, et al. Positron emission tomography imaging ofmu- and delta-opioid receptor binding in alcohol-dependent and healthy controlsubjects. Alcohol Clin Exp Res 2011;35(12):2162–73.
540 Urban & Martinez

Author's personal copy
123. Abi-Dargham A, Krystal JH, Anjilvel S, et al. Alterations of benzodiazepine receptorsin type II alcoholic subjects measured with SPECT and [I-123]iomazenil. Am JPsychiatry 1998;155(11):1550–5.
124. Lingford-Hughes AR, Acton PD, Gacinovic S, et al. Reduced levels of GABA-benzodiazepine receptor in alcohol dependency in the absence of grey matteratrophy. Br J Psychiatry 1998;173:116–22.
125. Gilman S, Koeppe RA, Adams K, et al. Positron emission tomographic studies ofcerebral benzodiazepine-receptor binding in chronic alchoholics. Ann Neurol 1996;40:163–71.
126. Litton JE, Neiman J, Pauli S, et al. PET analysis of [11C]flumazenil binding tobenzodiazepine receptors in chronic alcohol-dependent men and healthy controls.Psychiatry Res 1993;50(1):1–13.
127. Lingford-Hughes AR, Acton PD, Gacinovic S, et al. Levels of gamma-aminobutyricacid-benzodiazepine receptors in abstinent, alcohol-dependent women: preliminaryfindings from an 123I-iomazenil single photon emission tomography study. AlcoholClin Exp Res 2000;24(9):1449–55.
128. Jalan R, Turjanski N, Taylor-Robinson SD, et al. Increased availability of centralbenzodiazepine receptors in patients with chronic hepatic encephalopathy andalcohol related cirrhosis. Gut 2000;46(4):546–52.
129. Staley JK, Gottschalk C, Petrakis IL, et al. Cortical gamma-aminobutyric acid typeA-benzodiazepine receptors in recovery from alcohol dependence: relationship tofeatures of alcohol dependence and cigarette smoking. Arch Gen Psychiatry 2005;62(8):877–88.
130. Krystal JH, Staley J, Mason G, et al. Gamma-aminobutyric acid type A receptors andalcoholism: intoxication, dependence, vulnerability, and treatment. Arch Gen Psy-chiatry 2006;63(9):957–68.
131. Voruganti LN, Slomka P, Zabel P, et al. Cannabis induced dopamine release: anin-vivo SPECT study. Psychiatry Res 2001;107(3):173–7.
132. Stokes PR, Mehta MA, Curran HV, et al. Can recreational doses of THC producesignificant dopamine release in the human striatum? NeuroImage 2009;48(1):186 –90.
133. Barkus E, Morrison PD, Vuletic D, et al. Does intravenous �9–tetrahydrocannabinolincrease dopamine release? A SPET study. J Psychopharmacol 2011;25(11):1462–8.
134. Urban N, Sliftstein M, Thompson JL, et al. Dopamine release in chronic cannabisusers: a [11C]raclopride positron emission tomography study. Biol Psychiatry 2012.[Epub ahead of print].
135. Abi-Dargham A, Kegels LS, Martinez D, et al. Dopamine mediation of positivereinforcing effects of amphetamine in stimulant naive healthy volunteers: results froma large cohort. Eur Neuropsychopharmacol 2003;13(6):459–68.
136. Sevy S, Smith GS, Ma Y, et al. Cerebral glucose metabolism and D2/D3 receptoravailability in young adults with cannabis dependence measured with positronemission tomography. Psychopharmacology [Berl] 2008;197(4):549–56.
137. Stokes PR, Egerton A, Watson B, et al. History of cannabis use is not associated withalterations in striatal dopamine D2/D3 receptor availability. J Psychopharmacol2012;26(1):144–9.
138. Hirvonen J, Goodwin RS, Li CT, et al. Reversible and regionally selective downregu-lation of brain cannabinoid CB(1) receptors in chronic daily cannabis smokers. MolPsychiatry 2011. DOI: 10.1038/mp.2011.82. [Epub ahead of print].
541Neuroimaging in Addiction
Related Documents





![Neurobiology of Addiction presentation [Autosaved]](https://static.cupdf.com/doc/110x72/61df190bd25d6301594124e7/neurobiology-of-addiction-presentation-autosaved.jpg)





