Research report Neuroanatomical distribution of ARX in brain and its localisation in GABAergic neurons Karine Poirier a,1 , Hilde Van Esch a,1 , Gae ¨lle Friocourt a , Yoann Saillour a , Nadia Bahi a , Ste ´phanie Backer b , Evelyne Souil c , Laetitia Castelnau-Ptakhine a , Cherif Beldjord a , Fiona Francis a , Thierry Bienvenu a , Jamel Chelly a, * a Laboratoire de Ge ´ne ´tique et de Physiopathologie des Retards Mentaux, Institut Cochin. Inserm U567, Universite ´ Paris V. 24, rue du Faubourg Saint Jacques, 75014 Paris, France b De ´veloppement et migration neuronale, Institut Cochin. Inserm U567, Universite ´ Paris V. 24, rue du Faubourg Saint Jacques, 75014 Paris, France c Plateforme de Morphologie et Histologie du petit animal, Institut Cochin, CNRS, INSERM, Universite ´ Paris V. 123, Boulevard Port-Royal, 75014 Paris, France Accepted 11 November 2003 Abstract Recent human genetics approaches identified the Aristaless-related homeobox (ARX) gene as the causative gene in X-linked infantile spasms, Partington syndrome, and non-syndromic mental retardation as well as in forms of lissencephaly with abnormal genitalia. The ARX predicted protein belongs to a large family of homeoproteins and is characterised by a C-terminal Aristaless domain and an octapeptide domain near the N-terminus. In order to learn more about ARX function, we have studied in detail Arx expression in the central nervous system during mouse embryonic development as well as in the adult. During early stages of development, Arx is expressed in a significant proportion of neurons in the cortex, the striatum, the ganglionic eminences and also in the spinal cord. In the adult, expression of Arx is still present and restricted to regions that are known to be rich in GABAergic neurons such as the amygdala and the olfactory bulb. A possible role for Arx in this type of neurons is further reinforced by the expression of Arx in a subset of GABAergic interneurons in young and mature primary cultures of cortical neuronal cells as well as in vivo. Moreover, these data could explain the occurrence of seizures in the great majority of patients with an ARX mutation, due to mislocalisation or dysfunction of GABAergic neurons. We also performed ARX wild-type and mutant over-expression experiments and found that the different ARX mutations tested did not modify the morphology of the cells. Moreover, no abnormal cell death or protein aggregation was observed, hence suggesting that more subtle pathogenic mechanisms are involved. D 2004 Elsevier B.V. All rights reserved. Theme: Disorders of the nervous system Topic: Developmental disorders Keywords: ARX; Mental retardation; Brain development; GABA; Mouse 1. Introduction The human Aristaless-related homeobox gene (ARX) was identified as the causative gene in X-linked infantile spasms (ISSX; West syndrome, MIM308350), in certain families with X-linked syndromic (Partington syndrome, MIM309510) and non-syndromic mental retardation, in- cluding patients with dystonia, and patients with myoclonic epilepsy with spasticity [5,20]. Mutations identified in ARX in these patients included missense mutations, a recurrent in-frame 24-bp duplication predicted to cause an expansion of a polyalanine tract from 12 to 20 alanines and a small in- frame insertion leading to an increase of another polyala- nine tract containing 16 residues. Notably, no nonsense mutations were identified in these cases. More recently, mutations in ARX have been identified in X-linked lissen- cephaly with abnormal genitalia (XLAG, MIM300215) [9]. However, these latter mutations in ARX result predominant- ly in premature termination signals because of frameshifts 0169-328X/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.molbrainres.2003.11.021 * Corresponding author. Tel.: +33-1-44-41-24-81; fax: +33-1-44-21- 24-21. E-mail address: [email protected] (J. Chelly). 1 These authors contirbuted equally to this work. www.elsevier.com/locate/molbrainres Molecular Brain Research 122 (2004) 35 – 46

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/molbrainres
Molecular Brain Research 122 (2004) 35–46
Research report
Neuroanatomical distribution of ARX in brain and its localisation
in GABAergic neurons
Karine Poiriera,1, Hilde Van Escha,1, Gaelle Friocourta, Yoann Sailloura, Nadia Bahia,Stephanie Backerb, Evelyne Souilc, Laetitia Castelnau-Ptakhinea, Cherif Beldjorda,
Fiona Francisa, Thierry Bienvenua, Jamel Chellya,*
aLaboratoire de Genetique et de Physiopathologie des Retards Mentaux, Institut Cochin. Inserm U567, Universite Paris V. 24,
rue du Faubourg Saint Jacques, 75014 Paris, FrancebDeveloppement et migration neuronale, Institut Cochin. Inserm U567, Universite Paris V. 24, rue du Faubourg Saint Jacques, 75014 Paris, France
cPlateforme de Morphologie et Histologie du petit animal, Institut Cochin, CNRS, INSERM, Universite Paris V. 123,
Boulevard Port-Royal, 75014 Paris, France
Accepted 11 November 2003
Abstract
Recent human genetics approaches identified the Aristaless-related homeobox (ARX) gene as the causative gene in X-linked infantile
spasms, Partington syndrome, and non-syndromic mental retardation as well as in forms of lissencephaly with abnormal genitalia. The ARX
predicted protein belongs to a large family of homeoproteins and is characterised by a C-terminal Aristaless domain and an octapeptide
domain near the N-terminus. In order to learn more about ARX function, we have studied in detail Arx expression in the central nervous
system during mouse embryonic development as well as in the adult. During early stages of development, Arx is expressed in a significant
proportion of neurons in the cortex, the striatum, the ganglionic eminences and also in the spinal cord. In the adult, expression of Arx is still
present and restricted to regions that are known to be rich in GABAergic neurons such as the amygdala and the olfactory bulb. A possible role
for Arx in this type of neurons is further reinforced by the expression of Arx in a subset of GABAergic interneurons in young and mature
primary cultures of cortical neuronal cells as well as in vivo. Moreover, these data could explain the occurrence of seizures in the great
majority of patients with an ARX mutation, due to mislocalisation or dysfunction of GABAergic neurons. We also performed ARX wild-type
and mutant over-expression experiments and found that the different ARX mutations tested did not modify the morphology of the cells.
Moreover, no abnormal cell death or protein aggregation was observed, hence suggesting that more subtle pathogenic mechanisms are
involved.
D 2004 Elsevier B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Developmental disorders
Keywords: ARX; Mental retardation; Brain development; GABA; Mouse
1. Introduction cluding patients with dystonia, and patients with myoclonic
The human Aristaless-related homeobox gene (ARX)
was identified as the causative gene in X-linked infantile
spasms (ISSX; West syndrome, MIM308350), in certain
families with X-linked syndromic (Partington syndrome,
MIM309510) and non-syndromic mental retardation, in-
0169-328X/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.molbrainres.2003.11.021
* Corresponding author. Tel.: +33-1-44-41-24-81; fax: +33-1-44-21-
24-21.
E-mail address: [email protected] (J. Chelly).1 These authors contirbuted equally to this work.
epilepsy with spasticity [5,20]. Mutations identified in ARX
in these patients included missense mutations, a recurrent
in-frame 24-bp duplication predicted to cause an expansion
of a polyalanine tract from 12 to 20 alanines and a small in-
frame insertion leading to an increase of another polyala-
nine tract containing 16 residues. Notably, no nonsense
mutations were identified in these cases. More recently,
mutations in ARX have been identified in X-linked lissen-
cephaly with abnormal genitalia (XLAG, MIM300215) [9].
However, these latter mutations in ARX result predominant-
ly in premature termination signals because of frameshifts

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–4636
due to insertions or deletions, or nonsense mutations. In
two cases of XLAG, missense mutations have been iden-
tified (R332H, L343Q) [9]. These two mutations are
located in highly conserved regions of the homeodomain
suggesting dysfunction of DNA binding. The striking
differences in phenotype, which are observed (XLAG vs.
X-linked mental retardation), are likely to be explained by
the consequences of the different ARX mutations on the
stability and/or the function of ARX protein.
The ARX predicted protein belongs to one of the three
largest classes of homeoproteins, the paired (Prd) class.
Within this class, it is member of a specific sub-class of
proteins, which contain a glutamine residue at the critical
position 50 of their homeodomain (Q50), a residue also
found conserved in the Drosophila Aristaless protein (al).
In addition to its paired/Q50 central homeodomain, ARX is
characterised by a 14 amino acid C-terminal Aristaless
domain and by an octapeptide domain located near the
N-terminus designated as the Goosecoid Engrailed Homol-
ogy (GEH) or the eh-1 domain in the Engrailed (En)
homeoprotein [11].
In order to better understand the function of ARX and its
involvement in X-linked mental retardation, we studied Arx
expression in the central nervous system (CNS) during
mouse embryonic development as well as in the adult.
Our results suggest that Arx is likely to play an important
role during embryonic CNS development, as well as in the
adult were ARX expression is still present in the neocortex,
hippocampus, hypothalamus, amygdala and especially in
the olfactory bulb. High expression of Arx in regions which
are known to be rich in GABAergic neurons, and the
expression of Arx in a subset of GABAergic neurons
suggest that mutations in ARX alter GABAergic neuronal
development and/or function, which might contribute to the
seizure phenotype observed in ARX-related disorders.
Over-expression of wild-type and different ARX mutants
involved in certain of these disorders, including a polyala-
nine tract expansion, revealed no dramatic changes in cell
differentiation, morphology and cell death. Moreover, no
abnormal protein aggregation was observed, suggesting that
these mutations result in more subtle cellular changes.
2. Materials and methods
2.1. Expression constructs
The QuikChangek Site-Directed Mutagenesis Kit (Stra-
tagene, La Jolla, CA, USA) was used to introduce the L33P
and the P353L mutations into the mouse Arx cDNA,
subcloned in the pEGFP-C1 vector (Clontech, Palo Alto,
CA, USA). All procedures were performed according to the
manufacturer’s instructions. In the case of the polyalanine
stretch expansion construct (ARX-polyA), we amplified
exon 2 of a patient known to carry the 7 alanine insertion
within the first polyalanine stretch. This exon was subse-
quently subcloned in the mouse Arx cDNA. A final expan-
sion from 16 to 25 alanines within the first polyalanine
stretch of the construct was obtained. All constructs were
confirmed by DNA sequencing (ABI 373, Applied Biosys-
tems, Foster City, CA, USA).
2.2. Antibodies
Two GST-fusion proteins corresponding to amino acids
(aa) 310–396 and aa 496–562 were used to produce
polyclonal antibodies (CovalAb, Oullins, France) against
the Arx homeodomain (anti-ARX-HD) and C-ter domain
(anti-ARX-Cter), respectively. Bleeds were taken after 10
weeks and tested on cell lysates. For Western blot analysis,
anti-Arx antibodies and anti-actin mouse monoclonal anti-
bodies (Chemicon, Temacula, CA, USA) were used at
1:1000. Anti-ARX-antibodies were used at 1:1000 for
immunohistochemistry and 1:100 for immunocytochemis-
try. Anti-a-tubulin (Sigma, 1:2000), anti-h-III tubulin
(BAbCo, Richmond, CA, USA, 1:1000), anti-CaMKII (Sig-
ma, 1:200) and anti-GABA (Sigma, 1:500) mouse mono-
clonal antibodies were also used for immunocytochemistry.
Specific immunostaining using anti-GABA and anti-CaM-
KII mouse monoclonal antibodies was observed only with
rat tissues and primary cultures of neuronal cells.
2.3. GST-fusion proteins production
Two GST-fusion proteins corresponding to the Arx
homeodomain (aa 310–396; GST-HD) and C-ter domain
(aa 496–562; GST-Cter) were produced in B27 bacteria
using IPTG induction. Protein purification was then per-
formed using glutathione Sepharose 4B (Amersham Phar-
macia, Uppsala, Sweden).
2.4. Cell culture and transfection
Animal handling and experiments were performed in
accordance with French ministry of Agriculture recommen-
dations and EEC regulations and guidelines. Adult pregnant
female rats were sacrificed by cervical dislocation and
brains were extracted from day 18 (E18) rat embryos in
phosphate-buffered saline (PBS) containing 0.6% glucose.
After a mechanic dissociation of cortex, living neuronal
cells were counted using trypan blue and cultured in neuro-
basal medium supplemented with 2% B-27 (Invitrogen,
Groningen, The Netherlands) and 100 unit/ml of penicillin
and streptomycin each, until fixation. Neurons were cultured
on glass coverslips or Petri dishes, coated with poly-D-lysine
(Invitrogen) and laminin (Sigma) [4]. Primary glial cultures
were prepared as previously described, from newborn cere-
bral hemispheres [16]. Glial cells were cultured in Dulbec-
co’s modified Eagle’s medium (DMEM, Invitrogen)
supplemented with 4.5 mg/ml glucose, 10% foetal bovine
serum (Invitrogen) and 100 units/ml of penicillin and
streptomycin each. PC12 cells were grown in DMEM

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–46 37
supplemented with glucose (4.5 mg/ml), 5% foetal bovine
serum, 10% horse serum, and 100 units/ml of penicillin and
streptomycin. Plasmid constructs were introduced into PC12
cells (80% confluent) using GenePorter (Gene Therapy
Systems, San Diego, CA, USA) according to the manufac-
turer’s instruction. COS7 cells were cultured in DMEM
supplemented with glucose (4.5 mg/ml), 10% foetal bovine
serum, and 100 units/ml of penicillin and streptomycin each.
Cells were plated at 106 cells per 10-cm culture dishes and
allowed 24 h to adhere before they were transfected using
FuGENE 6 transfection reagent (Roche Molecular Bio-
chemicals, Basel, Switzerland). Expression was allowed
for 24 h before cells were used for immunofluorescence
and biochemical studies. Transfection of PC12 and COS7
cells with all the different constructs was repeated three
times and each time approximately 75 transfected cells were
analysed.
2.5. Immunofluorescence staining and
immunohistochemistry
For immunostaining, PC12 cells were grown on poly-D-
lysine-coated glass coverslips. Immunostaining experiments
using neuronal cells were performed at 5, 10 or 14 days in
culture. COS7, PC12, glial and neuronal cells were fixed in
4% paraformaldehyde (PFA) in 0.1 M PBS (pH 7.4) for 30
min at room temperature. After three washes with 0.1 M
PBS, fixed cells were incubated 1 h in TBST (10 mM Tris–
HCl pH8, 150 mM NaCl, 0.05% Triton X-100) containing
2% goat serum (Dako, Glostrup, Denmark) and then incu-
bated overnight at 4 jC with the primary antibody. The cells
Fig. 1. Specificity of anti-Arx antibody (anti-ARX-HD). (A) Specificity of anti-AR
kDa was observed in E17 rat brain embryo and neonatal brain (NB) extracts. Not
assay. In lanes 1 and 2, equal amounts of adult olfactory bulb protein extracts we
lane 1 was not detected in lane 2 where the antibody was pre-incubated with specifi
transfected COS7 protein extracts. (C) Specificity of anti-ARX-HD tested using
transfected with ARX-GFP, while in lanes 1 and 3 GFP alone was used. The same b
the anti-ARX-HD and the anti-GFP antibodies in lanes 2 and 4.
were washed three times in TBST and incubated for 1 h at
room temperature with affinity purified secondary antibod-
ies (FITC conjugated goat anti rabbit or Texas Red conju-
gated goat anti rabbit antibodies (Jackson Immunoresearch,
West Grove, PA, USA) used at 1:200 dilution. After three
washes, coverslips were mounted in Mowiol and examined
with a Zeiss microscope equipped with epifluorescence
illumination.
Mouse and rat embryos, newborn (P4) and adult brains
were cryoprotected immediately after dissection with 30%
sucrose in phosphate buffer. Embryos were prefixed by
immersion in 2% PFA before being frozen with isopentane
and sectioned (10–15 Am thick). Embryos, newborn and
adult brains sections were then fixed for 20 min (embryos in
2% PFA, newborn and adult brains in 4% PFA) and they
were washed three times with PBS. Immunostaining and
analysis of the sections were performed as described previ-
ously. All immunostaining experiments were performed
with both anti-ARX-antibodies.
2.6. Immunoperoxidase labeling
ARX immunoperoxidase labeling was performed on
free-floating sections of 50 Am obtained from E13.5 mouse
after fixation overnight in 4% paraformaldehyde (PFA).
Endogenous peroxidase activity was blocked by a 30-min
treatment in 0.3% hydrogen peroxidase in phosphate-buff-
ered saline. Sections were pre-incubated in 10% goat serum
and 0.3% Triton X-100 in PBS for an hour and then
incubated with anti-ARX (1/200) in PBS overnight at 4
jC. After three PBS washes, sections were incubated with
X-HD for detecting endogenous Arx. A specific band of approximately 70
e the absence of this protein in cerebellum (Cb). (B) Antibody competition
re loaded on an SDS-PAGE gel. The specific band corresponding to Arx in
c GST-HD fusion protein. Lanes 3 and 4 show the same result obtained with
transfected COS7 cell extracts. Lanes 2 and 4 contain extracts from cells
ands, corresponding to the ARX-GFP fusion protein, were detected by both
K. Poirier et al. / Molecular Brain Research 122 (2004) 35–4638
the secondary antibody for 2 h at room temperature (bio-
tinylated goat anti-rabbit IgG, Vector Laboratories, Burlin-
game, CA). They were then transferred into avidin–biotin–
peroxydase complex (ABC kit, Vector Laboratories) for 90
min at room temperature. Peroxidase enzyme activity was
revealed using 3,3V-diaminobenzidinetetrahydrochloride
(DAB, final concentration 0.005%) as the chromogen and
0.01% hydrogen peroxide as the substrate. The sections
were then mounted onto gelatine-coated slides, dehydrated
and coverslipped.
2.7. Protein extracts and western immunoblot
Proteins from dissected adult mouse brains were extracted
in lysis buffer (50 mM Tris, 150 mMNaCl, 1% NP40 pH8.0)
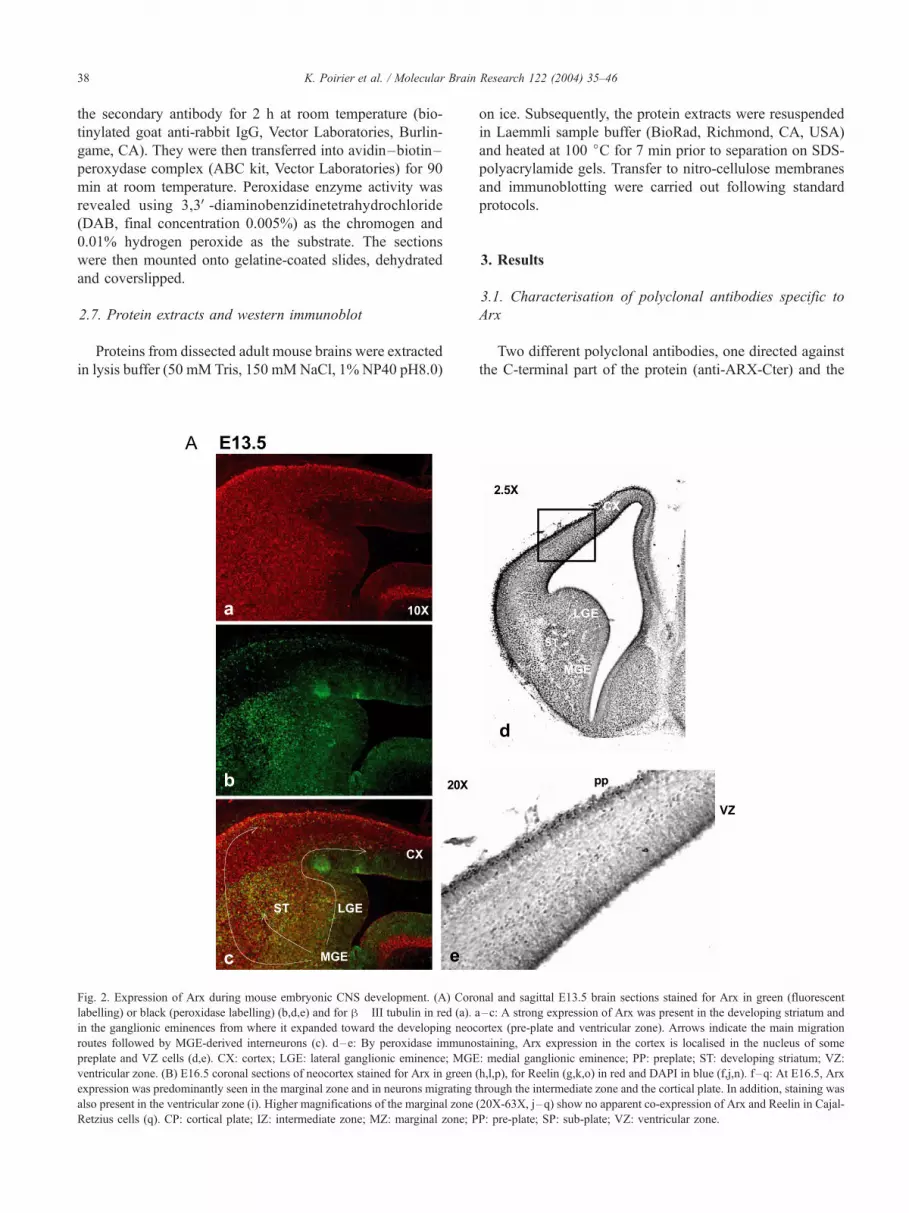
Fig. 2. Expression of Arx during mouse embryonic CNS development. (A) Coro
labelling) or black (peroxidase labelling) (b,d,e) and for h� III tubulin in red (a). a
in the ganglionic eminences from where it expanded toward the developing neoc
routes followed by MGE-derived interneurons (c). d–e: By peroxidase immuno
preplate and VZ cells (d,e). CX: cortex; LGE: lateral ganglionic eminence; MGE
ventricular zone. (B) E16.5 coronal sections of neocortex stained for Arx in green
expression was predominantly seen in the marginal zone and in neurons migrating t
also present in the ventricular zone (i). Higher magnifications of the marginal zone
Retzius cells (q). CP: cortical plate; IZ: intermediate zone; MZ: marginal zone; P
on ice. Subsequently, the protein extracts were resuspended
in Laemmli sample buffer (BioRad, Richmond, CA, USA)
and heated at 100 jC for 7 min prior to separation on SDS-
polyacrylamide gels. Transfer to nitro-cellulose membranes
and immunoblotting were carried out following standard
protocols.
3. Results
3.1. Characterisation of polyclonal antibodies specific to
Arx
Two different polyclonal antibodies, one directed against
the C-terminal part of the protein (anti-ARX-Cter) and the
nal and sagittal E13.5 brain sections stained for Arx in green (fluorescent
–c: A strong expression of Arx was present in the developing striatum and
ortex (pre-plate and ventricular zone). Arrows indicate the main migration
staining, Arx expression in the cortex is localised in the nucleus of some
: medial ganglionic eminence; PP: preplate; ST: developing striatum; VZ:
(h,l,p), for Reelin (g,k,o) in red and DAPI in blue (f,j,n). f–q: At E16.5, Arx
hrough the intermediate zone and the cortical plate. In addition, staining was
(20X-63X, j–q) show no apparent co-expression of Arx and Reelin in Cajal-
P: pre-plate; SP: sub-plate; VZ: ventricular zone.

Fig. 2 (continued).
K. Poirier et al. / Molecular Brain Research 122 (2004) 35–46 39
other against the homeodomain (anti-ARX-HD), were used
to detect the Arx protein. As shown in Fig. 1A, anti-ARX-
HD antibody revealed a band of approximately 70 kDa in
rat neonatal and embryo (E17) brain extracts. In protein
extracts from cerebellum, no protein was detected with these
antibodies. This result is in line with the previously de-
scribed in situ hybridisation and RT-PCR expression studies
that showed an absence of ARX mRNA expression in the
cerebellum [5]. The detected protein is likely to be specific
and corresponds to the native ARX protein, since pre-
immune sera did not produce a signal on the Western blots
and competitive assay of the antibodies with the specific
GST-fusion protein displaced completely the detected band
(Fig. 1B). Moreover, a similar band corresponding to an
ARX-GFP fusion protein obtained by transfection in COS7
cells was recognised by both anti-ARX-HD and anti-GFP
antibodies (Fig. 1C). Similar results were obtained for anti-
ARX-Cter (data not shown). The specificity of these anti-
bodies is further supported by the coherent immunostaining
results obtained with both antibodies. In this study, figures
illustrating the reported results were obtained with the anti-
ARX-HD.
3.2. Arx is predominantly expressed in the marginal zone of
the developing neocortex
Previous RNA in situ analysis showed that Arx can be
detected from day E8 onwards [5]. In this report, we

Fig. 3. Expression of Arx in E13.5 mouse spinal cord and pons. E13.5
transverse sections through mouse cervical spinal cord (b) and pons (a)
stained for Arx by immunoperoxidase technique. At E13.5, detailed
transverse sections showed expression of Arx in the area of the pons (a) and
in the floorplate (a–b). In addition, expression of Arx was seen in the
intermediate region of the spinal cord, extending towards the prospective
dorsal horn. (b) D: dorsal; FP: floorplate; V: ventral.
K. Poirier et al. / Molecular Brain Research 122 (2004) 35–4640
examined the expression of Arx protein in mouse embry-
onic neural tissues including brain and spinal cord by
immunostaining. At E10, a restricted expression of Arx
was observed in the telencephalic vesicles, the ventral
thalamus and in the floor plate (data not shown). At later
stages of development, from E10.5 to E16.5, Arx expres-
sion was observed in telencephalic structures including the
ganglionic eminences, the developing neocortex and the
striatum, in the ventral thalamus (Fig. 2A and B), as well as
in the pons (Fig. 3a) and the medulla oblongata (data not
shown). We did not detect any expression in the rest of the
developing diencephalon neither in the cerebellum (data not
shown). These results are consistent with our previous in
situ hybridisation experiments. At E13.5, a strong expres-
sion of Arx was observed in the developing striatum and in
the intermediate zone of the ganglionic eminences, a region
where interneurons originate [1,10] (Fig. 2A). From this
region, Arx expression expanded toward the developing
neocortex where it was mainly detected in the pre-plate,
characterised by expression of h-III tubulin, a marker of
post-mitotic neurons (Fig. 2A). The number of ARX
positive cells in the pre-plate layer adjacent to the MGE
appears to be much higher than in the preplate
corresponding to a more anterior region of the developing
telencephalon. These results suggest that ARX positive cells
display an anterior–posterior gradient coherent with gradi-
ent of migrating neurons from the MGE to the telenceph-
alon (data not shown). We also observed a staining in the
ventricular zone, suggesting that Arx is expressed in both
dividing and post-mitotic cells (Fig. 2B). The nuclear
localisation of ARX in cells localised in the VZ was
confirmed by peroxidase immunostaining (Fig. 2A, d and
e). At E16.5, Arx expression was predominantly seen in the
marginal zone and in neurons migrating through the inter-
mediate zone and the cortical plate (Fig. 2B). In addition,
staining was still present in the ventricular zone. To further
characterise the type of neurons that expressed Arx in the
marginal zone, we performed co-labelling with Reelin, a
glycoprotein secreted by Cajal-Retzius cells. We did not
observe co-expression of Reelin and Arx in Cajal-Retzius
cells, suggesting that Arx labels another sub-type of early
generated neurons (Fig. 2B).
3.3. Arx expression in the developing spinal cord
We also examined Arx expression in the developing
and adult spinal cord at the cervical level. The expression
of Arx was first seen in the neuroepithelium at E10.5 (data
not shown), confirming our previous in situ data. At this
early stage of development, spinal cord cells are beginning
to differentiate, moving from the ventricular zone to the
intermediate zone during the process of migration. At
E13.5, we observed that Arx was still expressed in the
floorplate and in the area of the pons (Fig. 3). In addition,
an expression was seen in a low proportion of cells in the
mantle layer of the intermediate region and the prospective
dorsal horn (Fig. 3). Analysis at later stages of develop-
ment showed a decreasing number of cells expressing
ARX. In the adult spinal cord, Arx expression is no longer
detected (data not shown).
3.4. Arx expression in the neocortex and in the olfactory
bulb in the adult
We examined the expression of the Arx protein in
adult mouse cortex, striatum, thalamus, colliculi, cerebel-
lum, hippocampus and olfactory bulb by Western blot
analysis using both anti-ARX antibodies. Besides a
strong predominant expression in the olfactory bulb, a
specific band of approximately 70 kDa was present in
the cortex, the striatum, and the thalamus. In the colliculi
and the hippocampus, the Arx protein band was barely
detectable (data not shown). As previously observed by

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–46 41
RNA in situ hybridisation, no expression was detected in
the cerebellum.
To confirm this restricted pattern of Arx expression in the
adult CNS, we performed immunostaining on adult mouse
and rat brain sections. This confirmed that Arx was still
expressed in single cells dispersed in all layers of the
neocortex, in the ventral hypothalamus, in the dentate gyrus
Fig. 4. Expression of Arx in the adult mouse brain. (A) Coronal section through th
Left, a histological section of the mouse brain is given to indicate the different struc
nuclear localisation of ARX in cortical cells is shown with a higher magnification
Granular cell layer, dentate gyrus; PoDG: Polymorph cell layer, dentate gyrus. A
through the olfactory bulb, stained for Arx in green (e) and DAPI in blue (j). Left,
cell layers. Arx expression is evident in glomerular (GL), granule (GR) and mitr
(Fig. 4A) and in a large number of cells in the amygdala
(data not shown) and the olfactory bulb (Fig. 4B). In the
latter, Arx labelled neurons in the granule, glomerular and
mitral cell layers (Fig. 4B). Similar to the results on
embryonic brain, we did not find Arx expression in the
cerebellum. Same results were obtained on adult rat brain
sections (data not shown).
e adult mouse brain, stained for Arx in green (a–d) and DAPI in blue (f– i).
tures where Arx expression was seen. Merged images are shown (k–n). The
(d–n). CX: cortex; DG: dentate gyrus; VH: ventral hypothalamus; GrDG:
ll layers of the neocortex contain Arx positive cells. (B) Coronal section
a histological section of the olfactory bulb is given to indicate the different
al (MI) cell layers.

Brain Research 122 (2004) 35–46
3.5. Arx expression in GABAergic neurons
In order to explore Arx expression in young and matur-
ing neurons, cell extracts from primary cultures of E18 rat
cortical brain neurons grown for 2, 5, 10, 14, 21 and 30 days
(D) were screened with anti-ARX-HD. Expression is al-
ready present in immature neurons at D2 and persisted for
30 days in culture (Fig. 5A). Hence, Arx is detected not only
in young differentiating neurons, but also in mature neurons
K. Poirier et al. / Molecular42
Fig. 5. Arx is predominantly expressed in GABAergic neurons in culture and in
grown for 2, 5, 10, 14, 21 and 30 days, respectively. Arx was found to be expre
double band was detected probably corresponding to post-translation modification
with the protein produced in vitro (data not shown). Anti-actin was used as contro
with a-tubulin (a), CaMKII (e) or GABA (h) in red, DAPI (c) in blue and Arx (b,f
subset of neurons (c versus d). Co-expression was observed in a subset of GA
(C) Coronal sections through the adult rat neocortex showing layers II– IVof the m
red. Merged images (c,g) and higher magnifications (63� , d,h) show that the large
was observed for Arx and CaMKII. The arrow indicates a cell that co-expresse
observed in all parts and layers of the neocortex.
in culture. Neuronal cell cultures of embryonic rat cortex
stained for Arx at D14 of culture, revealed a cell-specific
expression pattern, i.e. only a subset of neuronal cells
express Arx. On the contrary, no Arx expression was
observed in glial cells positive for glial fibrillary acidic
protein (GFAP), a specific marker of this type of cells (data
not shown).
To further characterise the neuronal subset that expressed
Arx, we assessed the expression of calcium-calmodulin-
vivo. (A) Arx expression in primary cultures of cortical rat brain neurons
ssed in both immature and mature neurons in these cultures. In all lanes, a
of the Arx protein or the existence of isoforms. This doublet was also seen
l. (B) Primary neuronal cell cultures of embryonic rat cortex at D14 stained
,i) antibodies in green. Merged images show that Arx is expressed in only a
BAergic neurons (j), while no co-expression was seen with CaMKII (g).
edial cortex, stained for Arx in green (b,f) and GABA (a) and CaMKII (e) in
majority of Arx positive cells co-expresses GABA, while no co-expression
s ARX and GABA. Similar co-expressions of Arx and GABA have been

Fig. 5 (continued).
K. Poirier et al. / Molecular Brain Research 122 (2004) 35–46 43
dependent kinase II (CaMKII) and gamma-aminobutyric acid
(GABA) in young and mature cultures of primary neuronal
cells (cultures of 5, 10 and 14 days). Double immunofluo-
rescent staining using adequate antibodies showed the pres-
ence of dot-like signals for Arx in the nucleus of many cells
that were positive for GABA (Fig. 5B). However, cells
expressing CaMKII, a marker of excitatory glutamatergic
neurons did not express Arx. This pattern of expression was
observed in young as well as in mature cultures. To confirm
this cell-specific expression pattern in vivo, we performed
double-staining experiments on newborn and adult rat brain
sections. GABA/Arx double immunostaining showed that
ARX is expressed in a high proportion of GABAergic cells.
In some brain structures known to be very rich in GABAergic
cells, such as the olfactory bulb or the ventral hypothalamus,
the proportion of ARX/GABA positive cells is very high and
represents approximately 90% of GABAergic neuronal cells
(Fig. 5C). A similar co-expression pattern was observed on
newborn brain sections (data not shown). On the contrary, no
co-expression of Arx and CaMKII was observed on these
sections. These results are in line with those obtained on
primary young and mature cultures of cortical brain neurons.
These data, together with the observed Arx expression pattern
that coincides with known migration routes of interneurons
during embryonic brain development, suggest that Arx is
predominantly expressed in GABAergic neurons and might
be involved in the differentiation or migration or function of
at least a subset of these GABAergic interneurons.
3.6. Phenotypic expression of cells expressing mutated ARX
Different mutations in the ARX gene are involved in
syndromic and non-syndromic forms of mental retardation.
The mutation 98C>T leading to a leucine to proline change
at amino acid 33 of ARX (L33P) is located in the octapep-
tide domain designated as the GEH domain and was
identified in a family with X-linked nonspecific mental
retardation. The mutation 1058C>T is predicted to cause a
change of proline 353 to leucine (P353L) and is one of the
six invariable residues, typical of the paired type homeo-
domain. This mutation has been identified in a rare X-linked
recessive myoclonic epilepsy with spasticity and intellectual
disability. Another mutation is a polyA tract expansion
(insertion of 7 alanines) within the first polyalanine stretch

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–4644
(at position 100–115) of the ARX protein. This type of
mutation has been identified in a family with X-linked
nonspecific mental retardation with aggressive behaviour.
To examine the cellular consequences of these ARX
mutations, COS7 cells and PC12 cells were transfected with
these three different mutant constructs fused to Green
Fluorescent Protein (GFP) as well as with the wild-type
construct. COS7, as well as PC12 cells do not express the
ARX protein endogenously. ARX wild-type and the three
mutant constructs (ARX-L33P, ARX-P353L and ARX-
polyA) were detected exclusively in the nucleus of the cells
(data not shown). No dramatic changes in PC12 cell
morphology before and after nerve growth factor (NGF)
treatment were noted with any of the mutant fusion proteins.
However, we cannot exclude an effect of the GFP-tag on the
function of the different ARX proteins. Interestingly, the
polyA tract expansion mutation did not lead to abnormal
intranuclear protein aggregation 1, 3 and 7 days after
transfection in COS7 cells or PC12 cells. These observa-
tions were confirmed by absence of insoluble aggregates in
the sensitive SDS-PAGE stacking gel performed on day 1
after transfection (data not shown). Moreover, by counting
the number of transfected cells 1, 3 and 7 days after
transfection with wild-type or one of the three mutant
constructs, we observed that the percentages of living cells
expressing ARX mutants were not significantly different
from those expressing wild-type ARX at any time point
examined. These results suggest that the pathogenic effects
of these mutations cause only subtle changes, if any, in these
cell types.
4. Discussion
We analysed the expression of the Arx protein in the
CNS during development and in the adult. Arx was found in
a number of specific brain regions as well as in the
developing spinal cord. During early development of the
telencephalon, expression of Arx is observed in the ventric-
ular zone and in post-mitotic neurons in the pre-plate. As
development emerges, Arx expression is mainly observed in
the marginal zone (MZ) as well as in post-mitotic neurons
that are migrating through the intermediate zone and the
cortical plate towards their final position. However, as Arx
is also expressed in cells of the ventricular zone, we can
suggest that this protein might play not only a role in post-
mitotic and migrating neurons but also in dividing progen-
itor cells. The Arx positive cells present in the MZ are
different from Reelin expressing Cajal-Retzius cells. It is
known that other types of neurons are present in the MZ, but
thus far their identity has not been fully characterised [15].
A strong Arx expression is present in the mouse devel-
oping striatum and in the intermediate zone of the gangli-
onic eminences, a region where GABAergic interneurons
originate [1,10]. Subsequently, these neurons follow a
tangential migratory route to their positions in the develop-
ing cortex. At least three spatially and temporally distinct
migration routes have been identified [12]. Although it
cannot be excluded that the Arx expression observed in
the pre-plate originates from cells produced in the dorsal
part of the cortex, the observed expression pattern seems
most coherent with a flux of migrating cells derived from
the medial ganglionic eminence that contributes to the ARX
positive interneurons. This suggests that Arx plays a role in
development and/or migration of neuronal cells, likely to be
interneurons derived from the ganglionic eminences. These
observations are reinforced by the expression in young
primary cultures of Arx in GABAergic neurons shown in
this study and the findings reported by Ref. [9]. These
authors revealed that deficiency of Arx in the mouse seems
to inhibit migration of GABAergic interneurons from the
medial ganglionic eminence (MGE) to the intermediate
zone, while the migration of GABAergic interneurons from
the MGE to the cortical subventricular zone though the
lateral ganglionic eminence (LGE) is maintained. In con-
clusion, our expression data obtained in late development
(E13.5 to E16.5) are in total agreement with the observed
findings in Arx � /Y mutant mice [9].
Moreover, these Arx expression data in the developing
mouse cortex and the ganglionic eminences are in line with
the expression pattern described during human brain devel-
opment [17].
During mouse spinal cord development, we observed an
early expression of Arx in the neuroepithelium. At E13.5,
we still found an expression in the floorplate, an interme-
diate target for path-finding commissural axons located at
the ventral midline of the developing mouse spinal cord. In
addition, Arx is expressed in the mantle layer of the
intermediate region (IR) and the prospective dorsal horn.
Interestingly, the IR is defined as the area between the dorsal
and ventral horns that contains primarily interneurons. Other
homeobox-containing transcription factors with restricted
domains of expression in post-mitotic cells of the IR have
been previously identified including En-1, Evx-1, Lmx-1
and Dbx-1 [13]. This observation underlies again the
involvement of Arx in the regulation of yet-to-be identified
neuronal process during CNS development.
In the adult, Arx expression is no longer present in the
spinal cord but noted in all cortical layers, the striatum, the
amygdala, the thalamus, the hypothalamus and olfactory
bulb, both by Western blot analysis and by immunohisto-
chemistry. Interestingly, Arx/CAMKII and Arx/GABA dou-
ble immunostaining experiments on adult rat brain sections
and primary cultures of neuronal and glial cells showed an
expression of Arx in a subset of GABA-expressing neurons,
whereas no expression of Arx in CaMKII excitatory neuro-
nal and glial cells was observed. Although it is difficult to
extrapolate from these results on the putative role of Arx in
adult brain, our results suggest that Arx is probably not
involved in intrinsic function of excitatory-type CAMKII-
positive neurons. Although we cannot exclude a role in
early development of this type of neurons, seen in the high

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–46 45
Arx expression present in the germinal ventricular zone. On
the other hand, Arx expression patterns observed in the
developing brain and the expression of Arx in GABAergic
neurons of adult brain and primary cultures suggest that Arx
is likely to be involved in the differentiation, migration and/
or function of at least a subset of GABAergic interneurons.
However, not all brain areas that contain GABAergic
neurons require Arx expression, as is for example the case
in the cerebellum were Arx expression is absent in the
inhibitory Purkinje cells. In the Dlx1/Dlx2 double mutant,
there is a massive reduction of GABAergic interneurons in
the cerebral cortex [1,2]. This is due to defects in the
different migration routes of the neurons originating from
the ganglionic eminences, resulting in reduced numbers of
striatal (GABAergic and cholinergic), olfactory bulb
(GABAergic and dopaminergic) and cortical (GABAergic)
interneurons [18]. However, as Dlx1/Dlx2 mutants still
express GABA in the subcortical telencephalon [1,2], it is
clear that other genes are involved in the control of
GABAergic neuronal development. Arx is thus likely to
fall into this category.
Our results suggest that the Arx protein is expressed in a
large proportion of young developing neurons, with dimi-
nution of expression in the adult, leaving only some
GABAergic neurons continuing to express this protein.
Moreover, these findings could explain the occurrence of
seizures observed in the great majority of patients with an
ARX mutation, due to a reduced number or function of
cortical GABAergic neurons. GABA, the principal inhibi-
tory neurotransmitter in the cerebral cortex, maintains the
inhibitory tone that counterbalances neuronal excitation.
Abnormalities of GABAergic function have already been
associated with epilepsy in humans. Recently, mutations in
the GABAA-receptor g2 were identified not only in patients
with generalised epilepsies and febrile seizures, but also
with severe myoclonic epilepsy in infancy [3,8]. Mutations
have also been identified in GABRA1 encoding the a1 sub-
unit of the GABAA receptor in affected individuals of a
large family with juvenile myoclonic epilepsy [6]. The high
expression of Arx observed in the amygdala is also concor-
dant with the occurrence of seizures. In fact, alterations in
the density of GABAergic interneurons (especially neurons
which contain calbindin and/or neuropeptide Y) in the
basolateral amygdala have been reported in several models
of epilepsy [14,21].
Different mutations have been identified in the ARX gene
and have been shown to be involved in a broad spectrum of
neurological disorders ranging from non-syndromic to syn-
dromic forms of mental retardation and XLAG [5,9,19].
ARX mutations include missense mutations, polyalanine
tract expansions, nonsense and frameshift mutations. To
better understand the clinical heterogeneity associated with
several missense mutations in ARX including the polyala-
nine expansion, we studied the spatial distribution of GFP-
tagged wild-type and ARX mutants in transfected COS7 and
PC12 cells. All tested mutants were localised in the nucleus
and did not affect the morphology of COS7 and PC12 cells
even after nerve growth factor (NGF) treatment. Moreover,
over-expression of wild-type or mutant ARX did not induce
cell death in vitro and no abnormal protein aggregation was
observed by over-expression of the ARX-polyalanine mu-
tant in COS7 and PC12-cells. This observation is in agree-
ment with previous data concerning alanine stretch
expansions in other disease gene products. In fact among
the alanine-expanded proteins involved in human diseases
(HOXA13, HOXD13, CBFA1, ZIC2, PABPN1), mutated
PABPN1 is the only one that is reported to induce the
formation of intranuclear inclusions [7]. A further analysis
of mutant clones in different cell types is required for a
further understanding of molecular and cellular consequen-
ces resulting from the dysfunction of ARX.
Acknowledgements
The authors are grateful to P. Billuart, J. Parnavelas, F.
Fauchereau, M. Mildalgo-Sanchez and E. Bloch-Gallego for
their helpful comments, M.C. Vinet for her technical
assistance, A. Goffinet for providing the Reelin antibodies
and A. Koulakoff for glial cell cultures. This work was
supported by the European Community (QLG2-CT-1999-
00791, QLG3-CT-2002-01810), CNRS, Fondation pour la
recherche Medicale (FRM), Fondation Bettencourt Schuler,
Fondation France Telecom, and Fondation Franc�aise pour laRecherche sur les Epilepsies K.P. is supported by a PhD
fellowship of the Ministere de la Recherche, H.V.E. is a
post-doctoral researcher of the Fund for Scientific Research-
Flanders, Belgium (FWO-Vlaanderen).
References
[1] S.A. Anderson, D.D. Eisenstat, L. Shi, J.L. Rubenstein, Interneuron
migration from basal forebrain to neocortex: dependence on Dlx
genes, Science 278 (1997) 474–476.
[2] S.A. Anderson, M. Qiu, A. Bulfone, D.D. Eisenstat, J. Meneses, R.
Pedersen, J.L. Rubenstein, Mutations of the homeobox genes Dlx-1
and Dlx-2 disrupt the striatal subventricular zone and differentiation
of late born striatal neurons, Neuron 19 (1997) 27–37.
[3] S. Baulac, G. Huberfeld, I. Gourfinkel-An, G. Mitropoulou, A. Bera-
nger, J.F. Prud’homme, M. Baulac, A. Brice, R. Bruzzone, E.
LeGuern, First genetic evidence of GABA(A) receptor dysfunction
in epilepsy: a mutation in the gamma2-subunit gene, Nat. Genet. 28
(2001) 46–48.
[4] Y. Berwald-Netter, N. Martin-Moutot, A. Koulakoff, F. Couraud, Na+-
channel-associated scorpion toxin receptor sites as probes for neuro-
nal evolution in vivo and in vitro, Proc. Natl. Acad. Sci. U. S. A. 78
(1981) 1245–1249.
[5] T. Bienvenu, K. Poirier, G. Friocourt, N. Bahi, D. Beaumont, F.
Fauchereau, L. Ben Jeema, R. Zemni, M.C. Vinet, F. Francis, P.
Couvert, M. Gomot, C. Moraine, H. van Bokhoven, V. Kalscheuer,
S. Frints, J. Gecz, K. Ohzaki, H. Chaabouni, J.P. Fryns, V. Desportes,
C. Beldjord, J. Chelly, ARX, a novel Prd-class-homeobox gene highly
expressed in the telencephalon, is mutated in X-linked mental retar-
dation, Hum. Mol. Genet. 11 (2002) 981–991.
[6] P. Cossette, L. Liu, K. Brisebois, H. Dong, A. Lortie, M. Vanasse,

K. Poirier et al. / Molecular Brain Research 122 (2004) 35–4646
J.M. Saint-Hilaire, L. Carmant, A. Verner, W.Y. Lu, Y.T. Wang, G.A.
Rouleau, Mutation of GABRA1 in an autosomal dominant form of
juvenile myoclonic epilepsy, Nat. Genet. 31 (2002) 184–189.
[7] X. Fan, P. Dion, J. Laganiere, B. Brais, G.A. Rouleau, Oligomeriza-
tion of polyalanine expanded PABPN1 facilitates nuclear protein ag-
gregation that is associated with cell death, Hum. Mol. Genet. 10
(2001) 2341–2351.
[8] L.A. Harkin, D.N. Bowser, L.M. Dibbens, R. Singh, F. Phillips, R.H.
Wallace, M.C. Richards, D.A. Williams, J.C. Mulley, S.F. Berkovic,
I.E. Scheffer, S. Petrou, Truncation of the GABA(A)-receptor gam-
ma2 subunit in a family with generalized epilepsy with febrile seiz-
ures plus, Am. J. Hum. Genet. 70 (2002) 530–536.
[9] K. Kitamura, M. Yanazawa, N. Sugiyama, H. Miura, A. Iizuka-Kogo,
M. Kusaka, K. Omichi, R. Suzuki, Y. Kato-Fukui, K. Kamiirisa, M.
Matsuo, S. Kamijo, M. Kasahara, H. Yoshioka, T. Ogata, T. Fukuda,
I. Kondo, M. Kato, W.B. Dobyns, M. Yokoyama, K. Morohashi,
Mutation of ARX causes abnormal development of forebrain and
testes in mice and X-linked lissencephaly with abnormal genitalia
in humans, Nat. Genet. 32 (2002) 359–369.
[10] A.A. Lavdas, M. Grigoriou, V. Pachnis, J.G. Parnavelas, The medial
ganglionic eminence gives rise to a population of early neurons in the
developing cerebral cortex, J. Neurosci. 19 (1999) 7881–7888.
[11] C. Mailhos, S. Andre, B. Mollereau, A. Goriely, A. Hemmati-Brivan-
lou, C. Desplan, Drosophila Goosecoid requires a conserved hepta-
peptide for repression of paired-class homeoprotein activators, Devel-
opment 125 (1998) 937–947.
[12] O. Marin, J.L. Rubenstein, A long, remarkable journey: tangential
migration in the telencephalon, Nat. Rev., Neurosci. 2 (2001)
780–790.
[13] M.P. Matise, A.L. Joyner, Expression patterns of developmental con-
trol genes in normal and Engrailed-1 mutant mouse spinal cord reveal
early diversity in developing interneurons, J. Neurosci. 17 (1997)
7805–7816.
[14] A.J. McDonald, F. Mascagni, Immunohistochemical characterization
of somatostatin containing interneurons in the rat basolateral amyg-
dala, Brain Res. 943 (2002) 237–244.
[15] G. Meyer, J.M. Soria, J.R. Martinez-Galan, B. Martin-Clemente, A.
Fairen, Different origins and developmental histories of transient
neurons in the marginal zone of the fetal and neonatal rat cortex,
J. Comp. Neurol. 397 (1998) 493–518.
[16] L. Nowak, P. Ascher, Y. Berwald-Netter, Ionic channels in mouse
astrocytes in culture, J. Neurosci. 7 (1987) 101–109.
[17] R. Ohira, Y. Zhang, W. Guo, K. Dipple, S. Shih, J. Doerr, B.
Huang, L. Fu, A. Abukhalil, D. Geschwind, et al, Human ARX
gene: genomic characterization and expression, Mol. Genet.
Metab. 77 (2002) 179.
[18] G. Panganiban, J.L. Rubenstein, Developmental functions of the Dis-
tal-less/Dlx homeobox genes, Development 129 (2002) 4371–4386.
[19] P. Stromme, M.E. Mangelsdorf, I.E. Scheffer, J. Gecz, Infantile
spasms, dystonia, and other X-linked phenotypes caused by mutations
in Aristaless related homeobox gene, ARX, Brain Develop. 24 (2002)
266–268.
[20] P. Stromme, M.E. Mangelsdorf, M.A. Shaw, K.M. Lower, S.M.
Lewis, H. Bruyere, V. Lutcherath, A.K. Gedeon, R.H. Wallace,
I.E. Scheffer, G. Turner, M. Partington, S.G. Frints, J.P. Fryns,
G.R. Sutherland, J.C. Mulley, J. Gecz, Mutations in the human
ortholog of Aristaless cause X-linked mental retardation and epi-
lepsy, Nat. Genet. 30 (2002) 441–445.
[21] J. Tuunanen, T. Halonen, A. Pitkanen, Decrease in somatostatin-im-
munoreactive neurons in the rat amygdaloid complex in a kindling
model of temporal lobe epilepsy, Epilepsy Res. 26 (1997) 315–327.
Related Documents











