6 Neuro-Immune Associative Learning M.-B. Niemi . G. Pacheco‐Lo ´pez . H. Engler . C. Riether . R. Doenlen . M. Schedlowski 1 Introduction ...................................................................................... 124 1.1 Cross Talk between Central Nervous System and Immune System ............................... 124 1.2 Classical Conditioning .............................................................................. 125 1.3 Neuroimmune Associative Learning ................................................................ 125 1.4 Historical Overview ................................................................................. 126 2 Evoking Neuroimmune Associative Learning Responses ........................................ 127 3 Conceptual Framework for Neuroimmune Associative Learning ............................... 129 3.1 Association Phase ................................................................................... 129 3.2 Recall Phase ......................................................................................... 129 4 Principles of Neuroimmune Associative Learning ............................................... 131 4.1 General Learning Rules of Neuroimmune Associative Learning .................................. 131 4.2 Features of Neuroimmune Associative Learning ................................................... 132 5 Mechanisms of Neuroimmune Associative Learning ............................................ 133 5.1 Neurobiology of Association and Recall ........................................................... 133 5.2 Peripheral Mediation of the Conditioned Effects on Immunity .................................. 137 6 Clinical Implications ............................................................................. 139 6.1 Animal Models ...................................................................................... 139 6.2 Human Studies ...................................................................................... 139 7 Methodological Considerations for Neuroimmune Associative Learning Experiments ......... 141 8 Closing Remarks .................................................................................. 142 # 2008 Springer ScienceþBusiness Media, LLC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

6 Neuro-Immune AssociativeLearning
M.-B. Niemi . G. Pacheco‐Lopez . H. Engler . C. Riether . R. Doenlen . M. Schedlowski
1
# 200
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124
1.1
Cross Talk between Central Nervous System and Immune System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1241.2
Classical Conditioning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1251.3
Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1251.4
Historical Overview . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1262
Evoking Neuroimmune Associative Learning Responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1273
Conceptual Framework for Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1293.1
Association Phase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1293.2
Recall Phase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1294
Principles of Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1314.1
General Learning Rules of Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1314.2
Features of Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1325
Mechanisms of Neuroimmune Associative Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1335.1
Neurobiology of Association and Recall . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1335.2
Peripheral Mediation of the Conditioned Effects on Immunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1376
Clinical Implications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1396.1
Animal Models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1396.2
Human Studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1397
Methodological Considerations for Neuroimmune Associative Learning Experiments . . . . . . . . . 1418
Closing Remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1428 Springer ScienceþBusiness Media, LLC.

124 6 Neuro-immune associative learning
Abstract: Neuroimmune associative learning constitutes the concomitant presentation of a neutral stimu-
lus (conditioned stimulus, CS) and an immunomodulating agent (unconditioned stimulus, US). Repre-
sentation of the CS alone is able to activate the centrally stored engram often resulting in changes in
immune parameters that mimic those formed by the US (conditioned immune response). This experimen-
tal model enables to study and dissect the afferent (at association) as well as efferent (at recall) pathway of
brain–immune communication. In this chapter, we will introduce this behavioral conditioning paradigm,
review the available literature focusing on features of the conditioned and unconditioned stimuli, and the
conditioned responses, the so far knownmechanisms steering the conditioned response, and the underlying
learning principles. Moreover, a theoretical framework modeling the pathways as well as the guidelines how
to design neuroimmune associative learning experiments will be provided. Finally, the clinical feasibility of
this behavioral approach will be discussed as a supplement to standard therapies with the aim of optimizing
individual healing conditions.
List of Abbreviations: CRs, conditioned responses; CTA, conditioned taste avoidance; SRBCs, sheep red
blood cells; 6‐OHDA, 6‐hydroxydopamine
1 Introduction
Clinical observations and experimental evidence demonstrate the intensive bidirectional communication
between the central nervous system (CNS) and the immune system (for review, see Elmquist et al., 1997;
Straub and Schedlowski, 2002; Tracey, 2002; Dantzer, 2004; Glaser and Kiecolt‐Glaser, 2005). We termed the
process of sensing and encoding immune inputs to the CNS, and further associating themwith the memory
traces of exteroceptive clues as neuroimmune associative learning. Retrieving such engrams may result in a
complex repertory of physiological responses affecting neurobehavioral, endocrine, as well as immune
parameters. Furthermore, neuroimmune associative learning paradigms can be employed to experimentally
study the principles by which the immune and the nervous system exchange information.
1.1 Cross Talk between Central Nervous System and Immune System
One put a lymphocyte into a culture dish, added an antigen, and out came an antibody. So who needed a
nervous system? (Spector, 1996).
The paradigm of total independence of the immune system has been dismissed by experimental
evidence accumulating mainly during the last three decades. Here are only some relevant issues related to
the present topic enlisted.
A whole series of neuropeptides, neurotransmitters, and neuroendocrine hormones are endogenously
produced by immune cells (for review, see Blalock, 1994; Tayebati et al., 2002; Warthan et al., 2002) and
many cytokines are found to be produced and have significant biological activity in the central and
peripheral nervous system (Blalock, 2005).
1. Stimulation or silencing of distinct brain areas affects immune functioning by different mechanism (for
a detailed review, see Meisel et al., 2005; Wrona, 2006; Ziemssen and Kern, 2007).
2. There is evidence for rich neural connections with lymphoid tissue (Steinman, 2004; Straub, 2004), and
receptors for neurotransmitters are also present on lymphocytes.
Immune‐to‐brain pathway: On the afferent pathway, peripheral immunological changes are signaled to the
CNS, and different ascending pathways (neural and humoral) have been identified. The vagus nerve
provides the major neural pathway identified to date. The initial chemosensory transduction events
occur in immune cells, which respond to specific chemical components expressed by dangerous microorgan-
isms. These immune chemosensory cells release mediators, such as cytokines, to activate neural elements,
including primary afferent neurons of the vagal sensory ganglia. Primary afferent activation initiates
local reflexes (e.g., cardiovascular and gastrointestinal) that support host defense (Goehler et al., 2000).

Neuro-immune associative learning 6 125
This neural afferent pathway is complemented by a humoral afferent pathway that involves cytokines or
other immunotransmitters transported in the blood or produced at the level of the circumventricular
organs. It is possible that such immunotransmitters cross the blood–brain barrier or originate a secondwave of
cytokines produced in the brain parenchyma (Banks, 2006). Depending on their source, these locally produced
cytokines can either activate neurons that project to specific brain areas or diffuse by volume transmission into
the brain parenchyma to reach their targets. Activation of neurons by cytokines can be direct or indirect. The
way the neural pathway of transmission interacts with the humoral pathway remains to be elucidated;
however, it has been proposed that each pathway may engage in different conditions (e.g., localized vs.
systemic infections), and thus codify different information (Dantzer et al., 2000).
Brain‐to‐immune pathway: There are three main pathways: the hypothalamo–pituitary–adrenal (HPA)
axis (humoral), the sympathetic–adrenal–medullary axis and the parasympathetic nervous system includ-
ing the vagus nerve (both neural). Activation of the HPA axis results in the production of glucocorticoid
hormones and catecholamines (Meisel et al., 2005), regulating cytokine balance (for review, see Ziemssen
and Kern, 2007) and vice versa. The sympathetic nervous system regulates immunity by innervation of
lymphoid organs and the release of noradrenaline, and a hormonal component that regulates immunity
systemically through the release of adrenaline from the medulla of the adrenal glands (Sternberg, 2006).
Coupling of the sympathetic nervous system and the HPA axis leads in the spleen to stronger effects
through activation of b‐adrenoceptors and glucocorticoid receptors (Straub, 2004).
In summary, the CNS and immune system intensively and extensively interact, sharing pathways,
messengers, and their receptors.
1.2 Classical Conditioning
Since the CNS has the capability to alter the activity of the immune system as well as of many other organs/
systems, it can be hypothesized that behavioral approaches such as classical conditioning may be able to
modulate immune functions. Any paradigm of associative learning or classical conditioning comprises two
basic phases: association and recall. During association phase, a neutral stimulus (to become a conditioned
stimulus: CS) is paired with an unconditioned stimulus (US), which elicits vigorous responding (uncondi-
tioned response, UR) (Domjan, 2005). Contingent pairing(s) of the CS and the US leads to the establish-
ment of a temporal/causal association of both stimuli, stored within the CNS. This learned association can
then be identified during recall by the emergence of new responses to the CS, which can be now presented in
the absence of the US. These new responses are termed conditioned responses (CRs) and they usually
mimic the URs.
1.3 Neuroimmune Associative Learning
During an association trial, a gustatory/olfactory stimulus as a CS (e.g., saccharin solution; taste, chocolate
milk; flavor, or camphor; odor) precedes the administration of an immunomodulating agent, that is, US
(e.g., cyclosporine A, cyclophosphamide, polyinosinic: polycytidylic acid or antigens). Paired administra-
tion of these stimuli leads to a CS–US association. During recall, the CS is able to produce some effects,
formerly ascribed just to the immunomodulating compound (US) (> Figure 6-1). This phenomenon
implies that the total effects of a drug are composed of pharmacological effects per se plus potential CRs
(Pacheco‐Lopez et al., 2006). Neuroimmune associative learning has been reported employing visual and/or
auditory stimuli such as a light or a tone as CS (MacQueen et al., 1989; Palermo‐Neto and Guimaraes, 2000;
Irie et al., 2002; Costa‐Pinto et al., 2005), or touch stimuli such as scratching or heating of the skin as CS
(Metal’nikov and Chorine, 1926; Nicolau and Antinescu‐Dimitriu, 1929). However, the naturalistic relation
of postprandial immunotoxicological consequences facilitates neuroimmune associative learning that
employs gustatory/olfactory clues as CS. It contributes to the feasibility, duration, and magnitude of the
association. Therefore, it is not surprising that stimuli causing gastrointestinal irritation get more easily

. Figure 6-1
Neuroimmune associative learning. During association, animals are exposed to a conditioned stimulus (CS)
paired with an immunomodulating unconditioned stimulus (US). During recall, the CS is presented without the
US, evoking the formerly learned association. This is often accompanied by avoidance behavior and a complex
physiological response that may also affect peripheral immune functions
126 6 Neuro-immune associative learning
associated with the postprandial taste/odor memories (e.g., after saccharin intake) than with an acoustic or
visual stimulus, since under natural circumstances the former are more likely to become associated
(Domjan et al., 2004).
1.4 Historical Overview
The first documented cases of behavioral conditioned immune effects are of anecdotal character and date
back to the late nineteenth century. Mackenzie (1886) reported a patient who was suffering from a severe
coryza at the sight of an artificial rose, which the presence of natural roses invariably produced in her case.
Others reported similar findings in that environmental stimuli that have been associated with an allergen in
the past provoke allergic symptoms in sensitive patients by a picture of a hay field (Hill, 1930, cited in Ader
and Cohen, 1992).

Neuro-immune associative learning 6 127
The first scientifical reports of classically conditioned immune functions arose from Russian scientific
contemporaries of I. P. Pavlov. Lukyanenko (1961) cites an observation of Makukahin (1911) and a report
by Voronov and Riskin (1925) as possibly the first ones to demonstrate ‘‘conditioned leukocytic reactions.’’
However, according to Luk’yanenko they could not be interpreted properly and were not carried forward.
In the 1920s, Metal’nikov and his colleagues at the Pasteur Institute in Paris, France systematically
documented that an immune response could be elicited without the presence of an antigen but just by
evoking a previous experience, that is, CS exposure. After scratching or heating the skin of guinea pigs with
a warmed metallic plate as CS, the animals received intraperitoneal (i.p.) injections of various bacteria‐derived compounds such as small doses of either Bacillus anthrax, a Staphylococcus filtrate, tapioca
emulsion, or Vibrio cholera as US. After association phase and a delay to allow the return to baseline levels
was completed, the CS alone yielded significant and rapid influx of polymorphonuclear leukocytes into the
peritoneal cavity (Metal’nikov and Chorine, 1926, 1928). As a proof of the biological relevance of the
elicited conditioned response on the immune system, it was reported that conditioned animals survived to
lethal doses of V. cholera or Streptococcus if they were previously behaviorally evoked. Follow‐up experi-
ments replicated and extended these findings (Nicolau and Antinescu‐Dimitriu, 1929; Ostrovskaya, 1929).
Additional evidence occurred largely provided by A. O. Dolin in the Soviet Union and his fellows Krylov,
Flerov, and Luk’yanenko. Both specific and nonspecific immune reactions and both immunosuppression
and immunoenhancement due to conditioning were achieved in mice, rats, guinea pigs, rabbits, dogs, oxen,
monkeys and also in humans (Dolin and Krylov, 1952; Doroshkevich, 1954; Vygodchikov, 1955). At the
same time, but less recognized, reports from Romania showed a conditioned increase in phagocytic activity
of blood polymorphonuclear cells in dogs (Benetato, 1955; Baciu et al., 1965).
In addition, a series of reports published in Switzerland documented conditioned asthma‐likeresponse by using an auditory CS in guinea pigs (Noelpp and Noelpp‐Eschenhagen, 1951a–c, 1952a–c).Such findings were replicated by Ottenberg et al. (1958), and extended to humans (Dekker et al., 1957;
Turnbull, 1962).
In 1975, Ader and Cohen published their seminal work in which they constructed the term behaviorally
conditioned immunosuppression, which retrospectively set the stage for the field of psychoneuroimmunology.
During that time, R. Ader was working on extinction of conditioned taste aversion employing saccharin
taste as CS paired with a drug that induced significant visceral malaise. He realized that some rats of the
conditioned group died during the course of a series of extinctions trials (i.e., CS representation). Moreover,
those animals had received the largest of three different amounts of saccharin and displayed the most
pronounced conditioned taste avoidance (CTA) behavior (Ader, 1974). Knowing the additional immuno-
suppressive properties of the drug employed as US, cyclophosphamide: CY, Ader and his colleague,
immunologist N. Cohen, tested the hypothesis that the increased mortality on the conditioned animals
during extinction phase was related to a compromised immune status resulted from a conditioned
immunosuppressive response. This hypothesis was systematically assessed and confirmed showing that
rats receiving paired administration of saccharin and CY not only displayed a strong CTA but also a reduced
production of antibody titers on challenge with sheep red blood cells (SRBC) (Ader and Cohen, 1975). This
initial finding was verified by independent laboratories basically under the same conditions (i.e., Rogers
et al., 1976; Wayner et al., 1978), and further extended and elaborated (Ader et al., 1982).
2 Evoking Neuroimmune Associative Learning Responses
As a synopsis, >Table 6-1 is provided summarizing several neuroimmune associative learning protocols in
which immune functions are evaluated after recall phase.
Different antigens have been applied as US eliciting conditioned immunoenhancement effects, for
example, T‐dependent antigens such as ovalbumin (Chen et al., 2004; Huang et al., 2004), keyhole limpet
hemocyanin (Ader et al., 1993), T‐independent antigens such as lipopolysaccharides (Bull et al., 1991), or
superantigens such as staphylococcal enterotoxin (Pacheco‐Lopez et al., 2004), as well as viral synthetic
patterns such as poly I:C (Solvason et al., 1993; Coussons‐Read et al., 1994; Demissie et al., 1995; Hsueh
et al., 1995, 1999, 2002; Kuo et al., 2001; Chao et al., 2005). On the other hand, immunosuppressive drugs
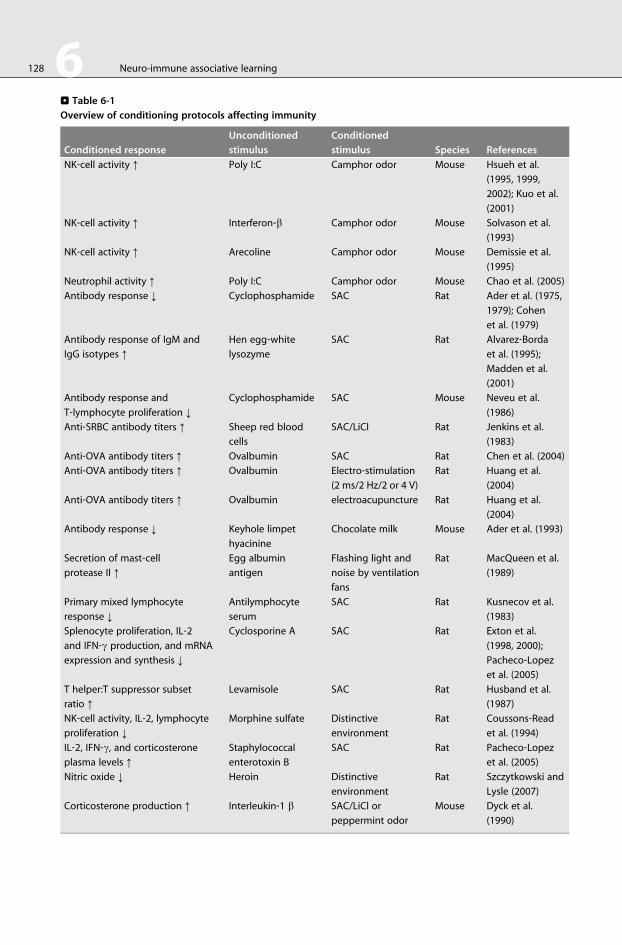
. Table 6-1
Overview of conditioning protocols affecting immunity
Conditioned response
Unconditioned
stimulus
Conditioned
stimulus Species References
NK‐cell activity ↑ Poly I:C Camphor odor Mouse Hsueh et al.
(1995, 1999,
2002); Kuo et al.
(2001)
NK‐cell activity ↑ Interferon‐b Camphor odor Mouse Solvason et al.
(1993)
NK‐cell activity ↑ Arecoline Camphor odor Mouse Demissie et al.
(1995)
Neutrophil activity ↑ Poly I:C Camphor odor Mouse Chao et al. (2005)
Antibody response ↓ Cyclophosphamide SAC Rat Ader et al. (1975,
1979); Cohen
et al. (1979)
Antibody response of IgM and
IgG isotypes ↑
Hen egg‐whitelysozyme
SAC Rat Alvarez‐Bordaet al. (1995);
Madden et al.
(2001)
Antibody response and
T‐lymphocyte proliferation ↓
Cyclophosphamide SAC Mouse Neveu et al.
(1986)
Anti‐SRBC antibody titers ↑ Sheep red blood
cells
SAC/LiCl Rat Jenkins et al.
(1983)
Anti‐OVA antibody titers ↑ Ovalbumin SAC Rat Chen et al. (2004)
Anti‐OVA antibody titers ↑ Ovalbumin Electro‐stimulation
(2 ms/2 Hz/2 or 4 V)
Rat Huang et al.
(2004)
Anti‐OVA antibody titers ↑ Ovalbumin electroacupuncture Rat Huang et al.
(2004)
Antibody response ↓ Keyhole limpet
hyacinine
Chocolate milk Mouse Ader et al. (1993)
Secretion of mast‐cellprotease II ↑
Egg albumin
antigen
Flashing light and
noise by ventilation
fans
Rat MacQueen et al.
(1989)
Primary mixed lymphocyte
response ↓
Antilymphocyte
serum
SAC Rat Kusnecov et al.
(1983)
Splenocyte proliferation, IL‐2and IFN‐g production, and mRNA
expression and synthesis ↓
Cyclosporine A SAC Rat Exton et al.
(1998, 2000);
Pacheco‐Lopezet al. (2005)
T helper:T suppressor subset
ratio ↑
Levamisole SAC Rat Husband et al.
(1987)
NK‐cell activity, IL‐2, lymphocyte
proliferation ↓
Morphine sulfate Distinctive
environment
Rat Coussons‐Readet al. (1994)
IL‐2, IFN‐g, and corticosterone
plasma levels ↑
Staphylococcal
enterotoxin B
SAC Rat Pacheco‐Lopezet al. (2005)
Nitric oxide ↓ Heroin Distinctive
environment
Rat Szczytkowski and
Lysle (2007)
Corticosterone production ↑ Interleukin‐1 b SAC/LiCl or
peppermint odor
Mouse Dyck et al.
(1990)
128 6 Neuro-immune associative learning

Neuro-immune associative learning 6 129
like cyclophosphamide (CY) and cyclosporine A (CsA) have been successfully employed as US, resulting in
conditioned immunosuppressive effects when associated with an appropriate CS such as taste/flavor (Ader
and Cohen, 1975; Exton et al., 2001).
Morphine sulfate as US paired with a distinctive environment as CS served to achieve a condi-
tioned suppression of NK‐cell activity, IL‐2 production, and lymphocyte proliferation (Coussons‐Readet al., 1994).
3 Conceptual Framework for Neuroimmune Associative Learning
The following section aims to provide a framework which takes basic assumptions about stimuli and sites of
action within neuroimmune associative learning into account. Within classical conditioning theory, a
prerequisite for its occurrence is that the CNS must sense both the CS (changes in the external environ-
ment) and the US (changes in the internal environment), where the signals are then processed, associated,
consolidated and recalled at evocation time (Eikelboom and Stewart, 1982; Bovbjerg, 2003; Pacheco‐Lopezet al., 2006, 2007b). Accordingly, the CNS must initiate both the UR and the CR. Translated to neuroim-
mune associative learning, this implies that only immune changes that are detected by the brain can serve as
US, in turn, only immune changes that are executed by the brain should then be called CR. However, the
nature of the US and the CR in most of the neuroimmune associative learning protocols so far reported is
not evident (> Figure 6-2).
3.1 Association Phase
During association phase, there are two possible USs that may be detected by the brain. A directly perceived
US, which is defined as a stimulus that itself is recognized by the CNS. The second kind is an indirectly
perceived US, which is defined to be signaled by intermediary molecules that are then indeed detected by the
CNS. In that case those molecules are the genuine US and the applied drug is a sham US, or indirectly
perceived US. This has important implications, for example, pairing of a CS with a compound that uncondi-
tionally decreases body temperature as US could result in a conditioned hypothermic response; however,
conditioned hyperthermia is also possible, that is, ‘‘paradoxically conditioned effects’’ or ‘‘counterconditioning
effects’’ (e.g., Bull et al., 1991). The proposed framework predicts such possibilities also for conditioned immune
responses and provides a tentative explanation. As Eikelboom and Stewart (1982) proposed, it is necessary to
correctly determine the nature of the US and UR to predict the direction of the CR.
There are two possible afferent pathways for any US, regardless whether directly or indirectly perceived –
a humoral and a neural pathway (see > Section 1.1). Within the neural pathway, the US information, on
detection, may be translated into neural activity. This sensing process requires immunoceptive capacities of
the CNS (Goehler et al., 2000; Blalock, 2005).
The humoral afferent pathway is required more often for indirectly perceived USs that induce molecules
that reach the brain via the blood stream as well as for USs that are not detected locally by the immune
system. In addition, if an indirectly perceived US affects several cell types, all of the involved molecules
become candidates for serving as a genuine US to be detected by the CNS. This implies a more complex,
longer, and therefore maybe slower signaling process for indirectly perceived US compared with directly
perceived US. Therefore, it can be assumed that the CNSmay take longer to respond to an indirectly perceived
US than to a directly perceived US.
3.2 Recall Phase
The CR represents the ultimate proof that an association has formerly taken place. There are two possible
pathways by which immune functions can be modulated by the CNS: the humoral efferent pathway and the
neural efferent pathway (see > Section 1.1). During recall phase, humoral efferent pathway may affect

. Figure 6-2
Theoretical framework for neuroimmune associative learning. At association, the conditioned stimulus (CS) can
potentially be associated with two possible unconditioned stimuli (US). The US directly detected by the CNS is
defined as a directly perceived US. The stimulus that needs one or more intermediary molecules released
by another system to be detected by the CNS is termed indirectly perceived US. Any US, indirectly or directly
perceived, has two possible afferent pathways to the CNS: a neural afferent pathway and a humoral efferent
pathway. At recall, the CNS canmodulate immune function via these twopathways. Thehumoral efferent pathway
may imply changes in neuroendocrine mediators that directly or indirectly modify the immune response. The
neural efferent pathway is supported by direct innervations of primary and secondary lymphoid organs
130 6 Neuro-immune associative learning
neurohormones that in turn affect immune responses. These peripheral effects are diffuse and long‐lastingas any neuroendocrine responses. Direct innervation of primary and secondary lymphoid organs (Elenkov
et al., 2000; Mignini et al., 2003; Tracey, 2007) may be part of the neural efferent pathway. Since several
immune parameters such as T‐cell differentiation (Sanders and Kohm, 2002a; Sanders and Straub, 2002b),
hematopoiesis (Miyan et al., 1998; Artico et al., 2002), T‐ and B‐cell activity (Downing and Miyan,
2000), NK‐cell activity (Katafuchi et al., 1993; Hori et al., 1995), and inflammatory responses (Czura and
Tracey, 2005; Pavlov and Tracey, 2005) are affected by neural activity, and they may also be subjected to be
affected by neuroimmune associative learning protocols. It may also be the case that besides the analyzed
and reported parameters there are still others not yet identified that are controlled or initiated by the
conditioning procedure. The picture becomes more complex considering that the immune system under-
takes sensitization (memory) and habituation (tolerance) processes, basically independent from the CNS.
In addition, it is known that several immune functions underlie circadian rhythms (Buijs et al., 2006).
These conditions complicate the prediction of the final nature and magnitude of the CR, often requesting
experimental trials. In this regard, some immune parameters modulated at recall may be the bizarre
reflection of neural activity that cannot be explained by orthodox learning and memory rules. For instance,
one such scenario would be that the delay between two recall trials is not long enough for certain immune
functions to return to baseline levels. Such successive recall trials may yield an additive effect in these and
related immune parameters rather than an extinction, which may occur in the neural correlates that elicited
these conditioned immune effects. In summary, immune responses can be affected recalling neuroimmune
engrams, but this does not necessarily imply that such immune responses were behaviorally conditioned.

Neuro-immune associative learning 6 131
4 Principles of Neuroimmune Associative Learning
The following section introduces findings of neuroimmune associative learning experiments and discusses
them with regard to rules of orthodox learning and memory theory that are commonly accepted to apply
when nonimmune USs are employed. This is insofar of interest as the conditioned taste aversion paradigm,
which is employed in the majority of neuroimmune associative learning experiments, has certain unique
features. It has been reported that odor–immune association can be established under long interstimulus
intervals (e.g., up to 24 h) (Hsueh et al., 1992). Taste–visceral associations are weak with interstimulus
delays longer than 4 h (Hiramoto et al., 1992; Solvason et al., 1992). Taste/odor–immune association could
occur after only a single CS–US association, whereas visual/auditory/touch–immune association requires
further reinforcement trials (Domjan, 2005). Therefore, given the CNS–immune communication complex-
ity, it seems worth to take a closer look at rules applying to a given neuroimmune associative learning.
4.1 General Learning Rules of Neuroimmune Associative Learning
Intensive training by increasing the number of CS–US trials during association strengthens the conditioned
response. This is a commonly agreed principle in diverse conditioning paradigms and was first described by
Pavlov (1927) and also applying to neuroimmune associative learning (Espinosa et al., 2004). Although one
learning trial conditioning (e.g., pairing antigenic challenge with saccharin, Alvarez‐Borda et al., 1995;
Madden et al., 2001) has been reported many times, it is not a phenomenon general to all USs employed in
neuroimmune associative learning protocols. Therefore, a paradigm including several association trials may
produce more reliable CRs (Espinosa et al., 2004). In this regard, it has been reported that the magnitude of
the conditioned effects on the immunity is larger after intensive learning (Niemi et al., 2007).
Extinction is defined as the reduction in magnitude and duration of the conditioned response as a
consequence of unreinforced trials (i.e., CS alone) (Szczytkowski and Lysle, 2007). The extinction rate was
directly related to the volume of CS presented on the association trial (Ader, 1974). This principle has been
proven to apply to neuroimmune associative learning in many other studies (e.g., Bovbjerg et al., 1984;
Lysle et al., 1988). In contrast, a recent finding indicates that evoking a consolidated taste–CsA engram
several times resulted in a stronger conditioned immunosuppression (Niemi et al., 2007). Similar heterodox
findings have been reported on an analog taste–immunosuppression engram resulting from pairing
saccharin and cyclophosphamide. Here, the immunosuppression was also more pronounced after several
CS unreinforced exposures (Ader and Cohen, 1975; Rogers et al., 1976; Wayner et al., 1978). Possible
explanations for such peculiar results are due to an insufficient delay between recall trials (too‐near trials),in that immune functions do not have enough time to return to baseline levels, an additive effect may result
in a cumulative immunosuppression. An alternative hypothesis is that the conditioned immunosuppressive
state occurring at first recall is sensed by the CNS and acts as a reinforcement to induce reconsolidation
(Berman and Dudai, 2001; Eisenberg et al., 2003; Dudai, 2006) of the taste–immunosuppression engram
and hereby working as an additional association trial.
Regarding the phenomenon of passive forgetting (the retention rate after a delay between association
and recall phases), Markovic et al. (1988) found excellent retention of CS–US association 8 weeks after
association. Here, rats were sensitized with ovalbumin injections; afterward, conditioning comprised
saccharin as CS and ovalbumin as US. No extinction occurred during a 6‐day test period in terms of the
CTA behavior.
Contingency is defined as the occurrence of CS and US and is important for achieving associative
learning. Ader and Cohen (1982) demonstrated this in showing that a partial reinforcement protocol
significantly reduced the conditioned immunosuppressive effect. Apparently, contingency in neuroimmune
associative learning adheres also to common learning principles.
Backward association would be the case if the US precedes the occurrence of the CS. Odor–poly I:C
backward conditioning resulted in conditioned increased NK‐cell activity (Solvason et al., 1992). This
should not be possible for directly (fast) perceived US (see > Section 3), whereas an indirectly perceived
US, which takes longer to be sensed by the CNS, is more likely to occur in parallel with sensory and

132 6 Neuro-immune associative learning
encoding steps of the CS, and therefore may become associated. In this case, a backward conditioned group
may help to delineate the nature of a given US or to discover the genuine US, respectively. An alternative
approach would be to systematically vary the CS–US interstimulus interval. In addition, if the UR kinetic is
known, the determination of the time point of the most pronounced CR enables extrapolation to the
genuine US that mediates the relevant information.
Latent inhibition, the CS preexposure may retard or diminish CS–US engram, thus reducing the
strength of the CR at recall. Such learning interference phenomena have been documented also in
neuroimmune associative learning (e.g., saccharin–ovalbumin Chen et al., 2004). However, the naturalistic
relation among stimuli should be considered to estimate the relevance of such a phenomenon in a given
conditioning protocol.
4.2 Features of Neuroimmune Associative Learning
Neuroimmune associative learning across life span: The strength of neuroimmune associative learning seems
to be age‐dependent. For instance Spector et al. (1994) report that old mice (24 months old) were able to
display a conditioned enhancement of NK activity in an odor–poly I:C conditioning protocol, but young
(3 months old) mice showed a stronger conditioned response. This reflects very much the unconditioned
response, which is detectable, although weaker in aged mice. This assumption is in line with reports
demonstrating a diminished conditioned immunosuppression in aged animals (Gorczynski, 1987b, 1991).
Two main factors could be the basis of aging effects on neuroimmune associative learning. Learning and
memory deficits are a common characteristic of CNS senescence (Nomura and Hori, 1996). However it
should also be considered that innervations to the immune organs change across life span, reducing
significantly during elderly stages. As has been reviewed (> Section 3.2), such neuroimmune efferent
neural pathways may be essential to induce the conditioned effect on immunity. Thus, it cannot be assumed
that a given conditioning protocol could be extrapolated to any stage of life.
Gender effects: Few systematic attempts have been undertaken to elucidate the role of gender or estruous
cycle on neuroimmune associative learning. Spector et al. (1994) reported conditioned increases in NK‐cellactivity in both male and female mice. However, it should be taken into account that a significant effect of
gonadal hormones on the neural (Kritzer et al., 2007) as well as on immune processes (Cutolo et al., 2006)
has been documented. Thus, it may not be surprising to find gender effects in a given neuroimmune
associative learning protocol.
Compensatory or paradoxical conditioned effects: These effects on immune responses have been reported
after evocation of a given neuroimmune associative learning. For example, Gorczynski and Kennedy (1984)
varied the time of day at which the initial association trial began and found that conditioned suppression
was developed by taste cues paired with CYduring the light portion of the diurnal cycle, whereas association
that began during the dark portion of the diurnal cycle resulted in either no CR or a conditioned
immunoenhancement. The authors hypothesized that the background level of neuroendocrine hormones
is critical to the direction of the CR to the taste cue paired with CY. However, Siegel et al. (1987) posited an
alternative hypothesis, indicating that the observation that most conditioning studies of regulatory
responses use stimuli other than taste cues suggest that the immunosuppression results in evoking a
taste–CY association may be unique to the use of taste cues. Other cues, inadvertently present in the
association regimen may control CRs different than those CRs elicited by taste cues (e.g., cue to conse-
quence specificity, Garcia and Koelling, 1967b).
Consciousness: An interesting finding is that association and recall phases can occur in anesthetized
subjects. Hsueh et al. (1992) reported that mice associate CS (camphor odor) and US (poly I:C) under
anesthesia. Second, if conditioned consciously, they could recall under anesthesia, even when CS/US
interstimulus interval was separated by 1–2 days. Since this unusual long interval differed from conscious
learning where the organism seeks information using a logical perceptual relation among events, the
authors reasoned that CS/US learning must be taken place unconsciously by different rules.
Immune history: Different immune histories among subjects may result in a divergent response to the
same immune stimulus. This yields a different immune reaction signaled to the CNS that may become

Neuro-immune associative learning 6 133
associated with a CS. Therefore, at recall phase, the conditioned response may vary in magnitude or even in
direction depending on the initial immune reaction. Evidence supporting this hypothesis comes from a
study showing that in an anaphylaxis model, physiological responses to first and second antigen exposure
differs drastically: CTA response (Djuric et al., 1988; Markovic et al., 1988), immunological, neural, and
behavioral responses (Costa‐Pinto et al., 2005). For instance, a recent study documented a highly significant
difference in lipopolysaccharide‐induced CTA behavior depending on the individual immune history (i.e.,
tolerant vs. naıve) (Pacheco‐Lopez et al., 2007a). Drug tolerance is another phenomenon that may affect
neuroimmune associative learning. Repeated treatment with a drug may reduce its specific effect on the
organism, a phenomenon termed drug tolerance. Therefore, if used as US with several CS–US pairings, the
signaling of this drug may alter with each administration. If so, this yields to different information possibly
becoming associated with the CS (e.g., Dyck et al., 1987). The first pairing may involve a different US signal
than the second, where it may be less pronounced.
Another key finding comes from experiments of odor–poly I:C conditioning in which several immune
responses (e.g., NK cell, neutrophil, and cytotoxic T cell) are affected on recall. However, a systematic
analysis led to the conclusion that the enhanced immune responses after recall depended on the immuno-
logical history of the particular subject (Demissie et al., 1997, 2000).
Species differences: Differences among species in a given conditioning paradigm do also exist. Although
conditioned immunosuppression in a saccharin–CsA conditioning paradigm is apparent in all three
species, it is reported that mice did not develop CTA behavior (Niemi et al., 2006), but rats do whereas
humans have reported a reduced palatability of the conditioned taste (Goebel et al., 2002). Dissociation of
CTA behavior and conditioned immunosuppression was found here in mice (e.g., Bovbjerg et al., 1984).
CS/US administration route: The delivery route of CS and US may affect the conditioned response. For
instance, it was found that the mode of administration of saccharin (CS) and ovalbumin (US) significantly
affects the conditioned behavioral response. The most effective mode to induce a conditioned taste aversion
behavior was CS periorally and US intraperitoneally. Strikingly, this mode yielded the mildest symptoms of
anaphylactic shock (UR) compared with CS intravenously/US intravenously mode and CS periorally/US
intravenously (weakest CTA response) (Markovic et al., 1988). Such results are in concordance with the
naturalistic relation of postprandial immunotoxicological consequences. Although according to established
associative learning rules, the extent of the UR determines the extent of the CR (Domjan, 2005); the
magnitude of the CR is often smaller than the UR. However, in the earlier case changing the route of
administration led to violation of this law.
Neuroimmune engram specificity: Solvason et al. (1991) demonstrated that a given CS creates specific
and independent neuroimmune engrams. Two odors (camphor and citronella oil) were paired with poly I:C
as a US. Mice were able to discriminate between the two CS when reexposed to either one of them in the
sense that they showed a CR to that CS that had been paired with the US during association phase, but not
to the other one.
Different CS served successfully in different conditioning protocols. Among the reported data are environ-
ments (Coussons‐Read et al., 1994; Szczytkowski and Lysle, 2007); visual (MacQueen et al., 1989) and auditory
(Harris and Fitzgerald, 1989;MacQueen et al., 1989); stimuli or such complex procedures like surgical sham skin
grafts (Gorczynski et al., 1982) (see also >Table 6‐1 and >Table 6‐2). In general, it can be assumed that
novelty, intensity, distinctiveness, and uniqueness are relevant characteristics of an effective CS.
5 Mechanisms of Neuroimmune Associative Learning
5.1 Neurobiology of Association and Recall
The neural network involved in taste–visceral associative learning includes mainly sensory and hedonic
pathways (Sewards and Sewards, 2002; Sewards, 2004). Among the involved brain structures are consistently
the nucleus tractus solitary, parabracchial nucleus, medial thalamus, amygdala, and insular cortex (Yamamoto
et al., 1994). In particular, the insular cortex subserves the association, retrieval, retention, and extinction of
taste–visceral memories (Nerad et al., 1996; Bermudez‐Rattoni et al., 1997, 2004; Pacheco‐Lopez et al., 2007b),
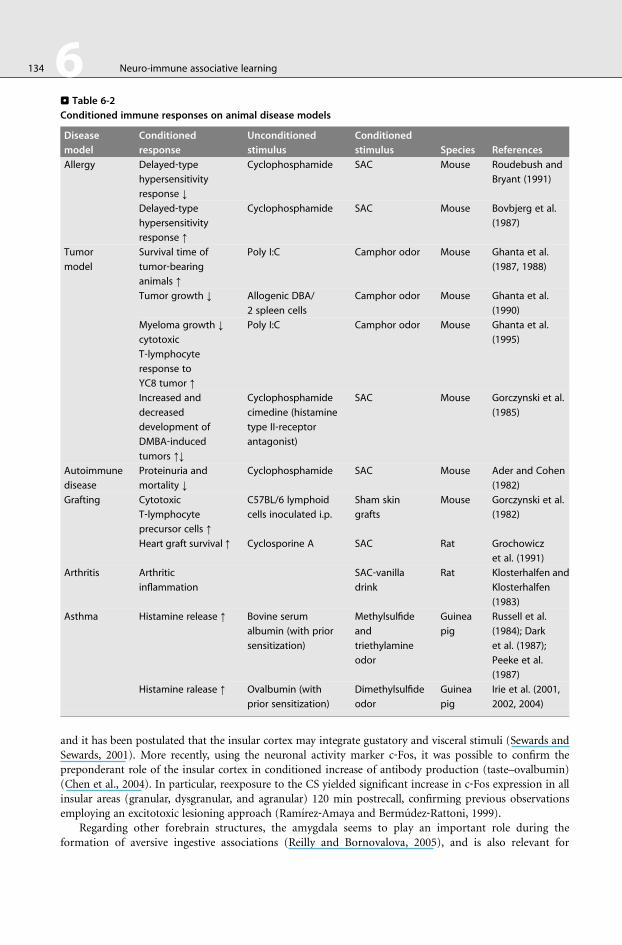
. Table 6-2
Conditioned immune responses on animal disease models
Disease
model
Conditioned
response
Unconditioned
stimulus
Conditioned
stimulus Species References
Allergy Delayed‐typehypersensitivity
response ↓
Cyclophosphamide SAC Mouse Roudebush and
Bryant (1991)
Delayed‐typehypersensitivity
response ↑
Cyclophosphamide SAC Mouse Bovbjerg et al.
(1987)
Tumor
model
Survival time of
tumor‐bearinganimals ↑
Poly I:C Camphor odor Mouse Ghanta et al.
(1987, 1988)
Tumor growth ↓ Allogenic DBA/
2 spleen cells
Camphor odor Mouse Ghanta et al.
(1990)
Myeloma growth ↓
cytotoxic
T‐lymphocyte
response to
YC8 tumor ↑
Poly I:C Camphor odor Mouse Ghanta et al.
(1995)
Increased and
decreased
development of
DMBA‐inducedtumors ↑↓
Cyclophosphamide
cimedine (histamine
type II‐receptorantagonist)
SAC Mouse Gorczynski et al.
(1985)
Autoimmune
disease
Proteinuria and
mortality ↓
Cyclophosphamide SAC Mouse Ader and Cohen
(1982)
Grafting Cytotoxic
T‐lymphocyte
precursor cells ↑
C57BL/6 lymphoid
cells inoculated i.p.
Sham skin
grafts
Mouse Gorczynski et al.
(1982)
Heart graft survival ↑ Cyclosporine A SAC Rat Grochowicz
et al. (1991)
Arthritis Arthritic
inflammation
SAC‐vanilladrink
Rat Klosterhalfen and
Klosterhalfen
(1983)
Asthma Histamine release ↑ Bovine serum
albumin (with prior
sensitization)
Methylsulfide
and
triethylamine
odor
Guinea
pig
Russell et al.
(1984); Dark
et al. (1987);
Peeke et al.
(1987)
Histamine ralease ↑ Ovalbumin (with
prior sensitization)
Dimethylsulfide
odor
Guinea
pig
Irie et al. (2001,
2002, 2004)
134 6 Neuro-immune associative learning
and it has been postulated that the insular cortex may integrate gustatory and visceral stimuli (Sewards and
Sewards, 2001). More recently, using the neuronal activity marker c‐Fos, it was possible to confirm the
preponderant role of the insular cortex in conditioned increase of antibody production (taste–ovalbumin)
(Chen et al., 2004). In particular, reexposure to the CS yielded significant increase in c‐Fos expression in all
insular areas (granular, dysgranular, and agranular) 120 min postrecall, confirming previous observations
employing an excitotoxic lesioning approach (Ramırez‐Amaya and Bermudez‐Rattoni, 1999).Regarding other forebrain structures, the amygdala seems to play an important role during the
formation of aversive ingestive associations (Reilly and Bornovalova, 2005), and is also relevant for
Neuro-immune associative learning 6 135
limbic–autonomic interaction (Swanson and Petrovich, 1998). A series of reports has indicated that the
insular cortex and the amygdala are key structures in conditioned immunosuppression after evoking taste–
CY and odor–CY associations (Ramırez‐Amaya et al., 1996, 1998).
It has also been proposed that the ventromedial hypothalamic nucleus, widely recognized as a satiety
center (Vettor et al., 2002), is intimately associated with sympathetic facilitation in peripheral tissues
(Saito et al., 1989), including modulation of peripheral immune reactivity (Okamoto et al., 1996). In
agreement with previous reports employing a taste–CY engram (Ramırez‐Amaya et al., 1996, 1998, 1999),
the neural substrates involved in the immunosuppression resulting in evoking a taste–CsA association in
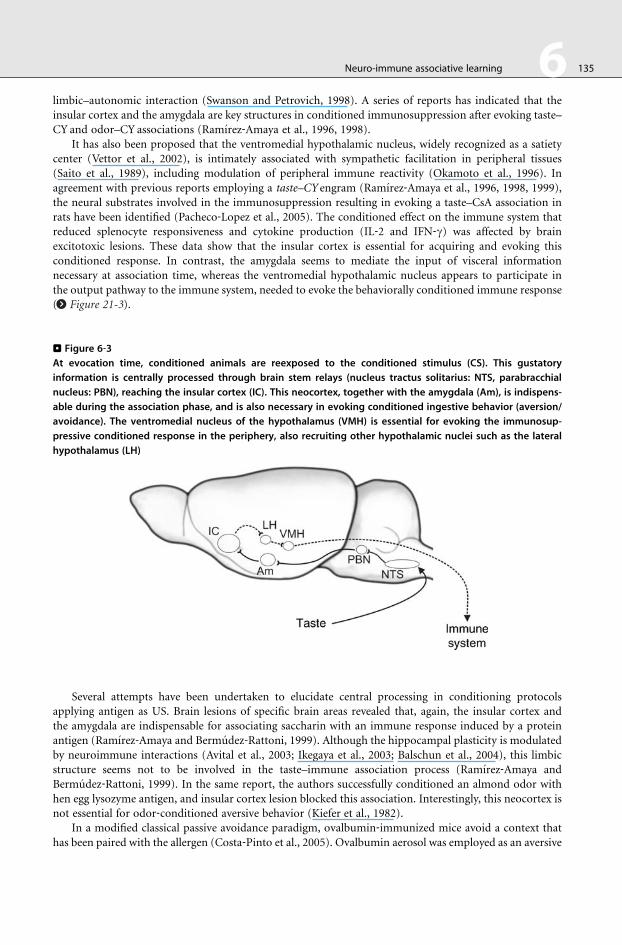
rats have been identified (Pacheco‐Lopez et al., 2005). The conditioned effect on the immune system that
reduced splenocyte responsiveness and cytokine production (IL‐2 and IFN‐g) was affected by brain
excitotoxic lesions. These data show that the insular cortex is essential for acquiring and evoking this
conditioned response. In contrast, the amygdala seems to mediate the input of visceral information
necessary at association time, whereas the ventromedial hypothalamic nucleus appears to participate in
the output pathway to the immune system, needed
(> Figure 21-3 ).
to evoke the behaviorally conditioned immune response
. Figure 6‐3At evocation time, conditioned animals are reexposed to the conditioned stimulus (CS). This gustatory
information is centrally processed through brain stem relays (nucleus tractus solitarius: NTS, parabracchial
nucleus: PBN), reaching the insular cortex (IC). This neocortex, together with the amygdala (Am), is indispens-
able during the association phase, and is also necessary in evoking conditioned ingestive behavior (aversion/
avoidance). The ventromedial nucleus of the hypothalamus (VMH) is essential for evoking the immunosup-
pressive conditioned response in the periphery, also recruiting other hypothalamic nuclei such as the lateral
hypothalamus (LH)
Several attempts have been undertaken to elucidate central processing in conditioning protocols
applying antigen as US. Brain lesions of specific brain areas revealed that, again, the insular cortex and
the amygdala are indispensable for associating saccharin with an immune response induced by a protein
antigen (Ramırez‐Amaya and Bermudez‐Rattoni, 1999). Although the hippocampal plasticity is modulated
by neuroimmune interactions (Avital et al., 2003; Ikegaya et al., 2003; Balschun et al., 2004), this limbic
structure seems not to be involved in the taste–immune association process (Ramırez‐Amaya and
Bermudez‐Rattoni, 1999). In the same report, the authors successfully conditioned an almond odor with
hen egg lysozyme antigen, and insular cortex lesion blocked this association. Interestingly, this neocortex is
not essential for odor‐conditioned aversive behavior (Kiefer et al., 1982).
In a modified classical passive avoidance paradigm, ovalbumin‐immunized mice avoid a context that
has been paired with the allergen (Costa‐Pinto et al., 2005). Ovalbumin aerosol was employed as an aversive

136 6 Neuro-immune associative learning
US, the CS was a usually preferred dark compartment of a box. After association phase, animals avoided
this compartment and spent more time in the bright (usually aversive) compartment of the box. The
expression of c‐Fos expression was tracked after airway ovalbumin challenge and it was found to be
enhanced in the hypothalamic paraventricular nucleus and the central nucleus of the amygdala in
sensitized mice. Therefore, it is likely that the immediate hypersensitivity reaction in allergic asthma,
which is characterized by IgE‐mediated mast‐cell deregulation with concomitant histamine release, plays
a major role in the reported brain activity (Costa‐Pinto et al., 2005). In addition, these structures are
commonly linked to emotional behavioral patterns that are also involved in conditioned taste aversion
(Bermudez‐Rattoni, 2004).Regarding neural mechanisms behind the odor–poly I:C conditioning paradigm, it has been reported
that IFN‐b is the major input to the CNS during association phase. It can replace poly I:C as US to acquire a
conditioned increase in NK‐cell activity; interestingly, this is not the case for IFN‐a (Solvason et al., 1988).
This association can be achieved by intravenous (i.v.) administration of IFN‐b at a dose of 10,000 IU, but
not at 1,000 IU. Further, if IFN‐b was directly delivered into the CNS via the cisterna magna, a smaller dose
(100 IU) sufficed to be associated with the CS. In turn, the CR was blocked by injecting anti‐IFN‐b(100 neutralizing units) into the cisterna magna 24 h before association (Solvason et al., 1993). The afferent
pathway for association yielding conditioned increased NK‐cell activity seems to be a different one to that of
the conditioned fever in the same associative learning conditioning protocol (Rogers et al., 1992). The i.p.
injection of sodium carbonate blocked CS–US association for enhancement of NK‐cell activity, but leftconditioned fever response unaffected. Conversely, indomethacin treatment which is known to prevent
prostaglandin synthesis blocked the conditioned fever response but not the conditioned NK‐cell response.Using a pharmacological approach, the neurochemical features of the conditioned effect enhancing NK‐cellactivity in rodents (odor–poly I:C association) have been described in detail by one group. Administering
lidocaine centrally blocked both association and recall of the CR (Rogers et al., 1994a) and the CNS sensory
processes of the CS, but not of the US. Similarly, peripheral and central treatment with monosodium
glutamate (Ghanta et al., 1994) or sodium carbonate (Rogers et al., 1994b) apparently blocks association,
but not CS perception. Moreover, the hypothalamic arcuate nucleus is necessary for association, but not for
recall of the CR (Ghanta et al., 1994). Recall in this paradigm seems to be mediated by central opioid
pathways; peripheral injection of the opioid receptor antagonist quaternary naltrexone (not penetrating the
CNS) does not affect recall of the CR. Naltrexone given before association did not prevent the conditioned
effect (Solvason et al., 1989), indicating independency of central opioid pathways.
Central catecholamines seem to be essential, and glutamate – but not GABA – is also required at recall
stage (Hsueh et al., 1999; Kuo et al., 2001). In particular, reserpine treatment before recall, which
unspecifically depletes central and peripheral catecholamine contents, blocked the CR (Hiramoto et al.,
1990). More recently, it has been shown that a‐ and b‐adrenoceptor antagonists or dopamine (DA)‐1 and
DA‐2 receptor antagonists given shortly before recall also blocked the CR (Hsueh et al., 1999).
Furthermore, it has been demonstrated that cholinergic, as well as serotonergic, central systems are
required in triggering conditioned NK‐cell response (Hsueh et al., 2002). At association, acetylcholine is
believed to act through nicotinic, M2‐, and M3‐muscarinic receptors, whereas at recall M1‐, M2‐ and M3‐muscarinic receptors have been identified to be crucially involved. In both association and recall phase,
serotonin acts through 5‐HT1 and 5‐HT2 receptors to affect the CR.
Neurotransmitter contents in certain brain areas during recall stage have been analyzed, revealing a
significantly higher norepinephrine content in the cerebellum and dopamine content in the striatum and
hippocampus in conditioned animals compared with controls (Hsueh et al., 1999). Interestingly, glutamate
contents at recall in this paradigm in the same brain areas did not differ between groups (Kuo et al., 2001).
In an identical conditioning protocol (camphor odor, poly I:C) with enhanced neutrophil activity as CR
instead of NK‐cell activity (Chao et al., 2005) measured levels of tyrosine hydroxylase in several brain areas
24 h after recall to localize action sites of catecholamines. Conditioned animals displayed significantly more
tyrosine hydroxylase expressing neurons in the hypothalamus, cortex, and locus coeruleus compared with
control animals. But this is unlikely to be related to a neural memory process since a 24 h memory trace is
rather unusual.

Neuro-immune associative learning 6 137
In addition to classical neurotransmitters, cytokines have been demonstrated to play an important role
within the CNS, modulating neuronal and glial function in nonpathological settings such as learning and
memory processes (Balschun et al., 2004; Dantzer, 2004; Tonelli et al., 2005). Specifically, proinflammatory
cytokines, such as IL‐1, IL‐6, and TNF‐a, have been shown to modulate spatial learning tasks, as well as
long‐term potentiation phenomena (Gibertini, 1996; Schneider et al., 1998; Fiore et al., 2000; Banks et al.,
2001; Matsumoto et al., 2001, 2002; Rachal Pugh et al., 2001; Lynch, 2002). In this sense, it can be assumed
that cytokines may be a significant factor in the associative processes occurring during behavioral condi-
tioning of immune functions. Apart from these neuromodulatory properties, proinflammatory cytokines
seem to play an important part in the afferent pathway between the immune system and the CNS
(Besedovsky and del Rey, 1996; Turnbull and Rivier, 1999; Dantzer, 2004). Therefore, it can be hypothesized
that central cytokines may act as mediators in the brain during an ‘‘immune‐sensing’’ phase in the
association phase. These hypotheses are supported by observations that (1) receptors for these proinflam-
matory cytokines are expressed in the CNS (Szelenyi, 2001; Sredni‐Kenigsbuch, 2002), (2) peripheral
immune changes affect central cytokine production and cytokine receptor expression in the brain (Pitossi
et al., 1997; Del Rey et al., 2000), and (3) cytokines can act as unconditioned stimuli to induce conditioned
taste aversion/avoidance (Tazi et al., 1988; Dyck et al., 1990; Janz et al., 1991; Hiramoto et al., 1993).
The underlying mechanisms when using SEB as US are completely unknown. However, systemic IL‐2administration has been found to modify central monoamine activity (Lacosta et al., 2000). Similarly,
striatum catecholamine concentrations followed a dose–response curve in reaction to increased peripheral
SEB immunization (Pacheco‐Lopez et al., 2004). Therefore, association may involve T‐cell‐derived cyto-
kines like IL‐2 signaling to the CNS.
5.2 Peripheral Mediation of the Conditioned Effects on Immunity
The available data suggest that the effects of conditioning could be mediated by a preferential effect on
T cells. Conditioned suppression of lymphoproliferative responses in rats and mice, for example, has been
observed in response to T‐cell mitogens but not (or less reliable) in response to B‐cell mitogens (Neveu
et al., 1986; Kusnecov et al., 1988; Lysle et al., 1990, 1991). Immune adoptive transfer experiments also
suggest that conditioning may be mediated by T‐cell changes (Gorczynski, 1987a). Splenocytes from
conditioned or experimentally naıve animals were transferred into irradiated naıve or conditioned animals
that were or were not subsequently reexposed to the CS. The observed increases or decreases in the
antibody‐forming cell response to SRBC depended on the donor cells and the conditioning treatment
experienced by the recipient. The separate transfer of enriched Tand B cells into naıve or conditioned animals
suggested that conditioning effects were attributable to the adoptively transferred T cells (Gorczynski, 1991).
However, the specificity of whether conditioning can modulate the antibody response to different types of
antigens has not been resolved.
With respect to adrenocortical influences, it is reasonable to hypothesize that conditioned alterations in
immunologic reactivity could be mediated by conditioned neuroendocrine changes. In experiments in
which humoral immune responses were assessed in conditioned animals, the recall of a taste–LiCl
association did not affect antibody responses to SRBC, in contrast to the immunosuppression that was
observed after evoking the taste–CY engram (Ader et al., 1979). This finding indicates that an immuno-
suppressive drug was needed to cause immunosuppression during association phase, thus supporting a
taste–immunosuppression engram interpretation.
In follow‐up experiments, circulating levels of adrenocortical steroids were artificially elevated by
exogenous administration of corticosterone at the time of antigen injection to mimic the stress response
occurring in animals reexposed to the taste paired with sickness (Ader et al., 1979). Elevated glucocorticoid
levels did not significantly lower antibody titers. These data would appear, a priori, to exclude the possibility
of a stress‐mediated phenomenon in the genesis of conditioned immunosuppression.
However, experimental data, in which the delayed‐type hypersensitivity (DTH) response was used as an
index of T‐cell function, support the hypothesis that changes in the immune function of animals subjected

138 6 Neuro-immune associative learning
to conditioned taste aversion might be better viewed as a secondary consequence of the psychological
conflict affecting thirsty animals that are exposed to a taste solution previously associated with sickness
(Kelley and Dantzer, 1988). According to this hypothesis, the immunosuppressive status after evocation
phase is a consequence of the conflict of the strong motivation to drink in fluid‐deprived animals against
the aversive memories associated to the CS. Accordingly, it should be possible to induce immunosuppres-
sion in conditioned animals even when no immunosuppressive drug is used during association phase (as
US). After a taste–LiCl association, conditioned animals showed a strong conditioned taste aversion, but
also an immunosuppressive status in the T‐cell function only when a forced choice test was implemented,
but not in a two‐bottle preference procedure which eliminates the psychological conflict of thirst versus
aversive memories (Kelley et al., 1984, 1985). In contrast to this stress hypothesis, a serum corticosterone
time–course study performed to examine the possible involvement of glucocorticoids in conditioned
immunosuppression of the DTH response has been published (Roudebush and Bryant, 1991). Animals
were sacrificed 30, 60, 90, 120 min and 24 h after evoking a taste–CY engram. No significant differences in
serum corticosterone levels were detected between nonconditioned controls and the conditioned groups at
any time point. Supporting this line of thinking, several other research groups have consistently reported that
the suppression of splenic T‐cell proliferation was independent of the stressor‐induced increase in adrenocor-
tical activity (Mormede et al., 1988; Lysle et al., 1990; Exton et al., 1998).
Importantly, both conditioned and stressor‐induced alterations in immune and nonspecific defense
responses have been attributed to the action of central and peripheral catecholamines. For instance, it has
been reported that both chlorpromazine and amitriptyline, both centrally acting, abolish the immunosup-
pressive status elicited by recalling a taste–CYengram (Gorczynski and Holmes, 1989). Furthermore, it has
also been reported that the b‐adrenergic antagonist, propranolol, blocked the immunosuppressive effects of
a conditioning stress paradigm (Lysle et al., 1992). Supporting the involvement of peripheral catechola-
mines, nadolol (a/b‐adrenergic antagonist that does not cross the blood–brain barrier) blocked the electric
shock‐induced suppression of splenic, but not peripheral, blood lymphocyte proliferation following
mitogenic stimulation ex vivo (Cunnick et al., 1990). In this regard, we have previously revealed that the
immunosuppressed status after evoking a taste–CsA engram is not related to the activation of the HPA axis
and is merely mediated by the neural innervation of the spleen, via noradrenaline–b‐adrenoceptors‐dependent mechanisms (Exton et al., 1999, 2002; Xie et al., 2002). It has been demonstrated that splenocyte
reactivity is modulated in part by tonic inhibition from the splenic nerve (Okamoto et al., 1996) and
sympathetic splenic innervation seems to be under the central control of the ventromedial hypothalamus
(Katafuchi et al., 1993, 1994). While electrical stimulation of the ventromedial hypothalamus has been
found to arouse sympathetic activity (Saito et al., 1989), the lateral hypothalamus seemed to do the
opposite (Bernardis and Bellinger, 1993). Thus, the lateral hypothalamus may exert immunoenhancing
properties (Wrona and Trojniar, 2003) in part by antagonizing the ventromedial hypothalamus, and thereby
reducing sympathetic tone inhibition. Importantly, such hypothalamic regulation of sympathetic activity
seems to be modulated by the insular cortex (Allen et al., 1991; Cechetto and Chen, 1992; Oppenheimer
et al., 1992; Butcher and Cechetto, 1998).
Concerning the conditioned enhancement of NK‐cell activity, several mechanisms have already been
elucidated. On the neural efferent pathway, sympathetic innervation of the spleen seems not to be responsi-
ble for mediating the conditioned response. Despite peripheral sympathectomy using 6‐hydroxydopamine
(6‐OHDA) between association and recall, the CR still occurred (Hiramoto et al., 1990). However, splenic
denervation should give final evidence. Interestingly, applying the same conditioning protocol, but measur-
ing neutrophil activity as UR and CR, respectively, peripheral sympathectomy before recall completely
abrogated the CR (Chao et al., 2005). On the humoral efferent pathway, elevated plasma adrenocorticotropic
hormone (ACTH) levels and splenic IFN‐a expression were measured in conditioned animals at recall time
(Hsueh et al., 1994b); no effect was found for b‐endorphin concentrations. Peripheral administration of the
synthetic glucocorticoid dexamethasone blocked recall, but not association of the CR, assumingly by
negative feedback inhibition of HPA axis activity (Hsueh et al., 1994a). Dexamethasone treatment before
recall resulted in elevated neutrophil activity in conditioned animals compared with controls, indicating that
this specific CR is independent of HPA activity (Chao et al., 2005).

Neuro-immune associative learning 6 139
6 Clinical Implications
6.1 Animal Models
>Table 6‐2 summarizes conditioned effects affecting critical immune responses in animal disease models
with potential clinical application.
The DTH response refers to an overreaction produced by the immune system given a presensitized
immune state of the host. A conditioned decrease (Roudebush and Bryant, 1991) and increase (Bovbjerg
et al., 1987) of this response has been observed by pairing saccharin with CY. Similarly, Exton et al. (2000)
reported a conditioned suppression of the contact hypersensitivity reaction by pairing saccharin with CsA.
Conditioning protocols attested a significant impact on tumor‐related models. For instance, tumor
growth was demonstrated to be enhanced as well as delayed by applying different US (CY and cimedine, a
histamine type II receptor antagonist) (Gorczynski et al., 1985). Moreover, the survival time of tumor‐bearing mice has been prolonged in conditioned animals (Ghanta et al., 1988).
Evoking a taste–CY engram, Ader and Cohen (1982) demonstrated a conditioned retardation in
proteinuria and mortality in New Zealand hybrid mice which are prone to develop an autoimmune disease
resembling human lupus erythematosus. Most interestingly, in a follow‐up study they could show that
the lupus‐prone MRL‐lpr/lpr mice displayed a weaker CTA compared with congenic controls. The
authors interpreted that the lupus‐prone animals seek to regain homeostasis by consuming the tasting
solution to achieve the therapeutic immunosuppressive status elicited by evoking the taste–CY engram
(Grota et al., 1987).
Another example of conditioning effects on immunity is documented by grafting experiments. Gorczynski
et al. (1982) performed skin grafting as CS in combination with i.p. injected lymphoid cells of another
mouse strain as US. Reexposing the conditioned animals to the sham‐grafting procedure, they showed
an increase in cytotoxic T‐lymphocyte precursor cells specific for alloantigens on the grafted tissue.
In addition, Grochowicz et al. (1991) reported a conditioned prolongation of the survival time of heart
allografts in rats evoking a saccharin–CsA engram. In follow‐up experiments, Exton and colleagues
(1999) demonstrated a significant prolongation of the survival of transplanted hearts, including long‐term survival (>100 days) of transplants in 20% of the animals that were conditioned and additionally
subtherapeutically treated with CsA. In addition, employing taste–CsA association Klosterhalfen and
Klosterhalfen (1983, 1990) extended the conditioned immunosuppression to an arthritis model. Before
induction of adjuvant arthritis, rats were dosed with cyclophosphamide (US) after presenting a distinctive
saccharin/vanilla solution (CS). Conditioned rats showed no external signs of a proliferation of inflamma-
tion, whereas approximately half of the animals in the control groups developed small lesions.
A series of studies dealt with conditioning of asthma‐like symptoms, anaphylactic shock (Noelpp
and Noelpp‐Eschenhagen, 1951c; Djuric et al., 1988; Palermo‐Neto and Guimaraes, 2000), or histamine
release, respectively (Dark et al., 1987; Irie et al., 2001, 2002, 2004) that have been already reviewed
(> Sections 1.3, 1.4, and 4.2).
6.2 Human Studies
This section summarizes the few, but promising research reports of conditioned effects modulating immune
responses in human subjects, indicating potential therapeutic outcomes of this behavioral approach.
A Japanese study from the early 1960s reported a conditioned dermatitis response in adult male subjects
elicited by evoking a specific association (CS: blue solution topically applied also contains 2% raw extract
Rhus vernicifera: US) (Ikemi et al., 1962). A case report of asthmatic patients suffering from skin sensitivities
to house‐dust extract and grass pollen shows a conditioned effect. The subjects were exposed to these
allergens by inhalation (Dekker et al., 1957). After a series of conditioning trials, they experienced allergic
attacks after inhalation of the neutral solvent used to deliver the allergens. This work showed not only fast
conditioning of the asthmatic attack (CR), but also tenacious retention, that is, lack of extinction. Similarly,

140 6 Neuro-immune associative learning
presenting a novel‐tasting and novel‐appearing drink as CS with house‐dust mite allergen as US to patients
with allergic rhinitis yielded an associative learning (Gauci et al., 1994). In addition, pairing an olfactory cue
as CS with nasal challenge with seasonal grass allergens to investigate conditioning of the immediate
hypersensitivity reaction in hay fever (seasonal allergic rhinitis) produced conditioned increase of histamine
release and decrease nasal airflow at recall (Barrett et al., 2000). Furthermore, there was evidence of
extinction and a follow‐up experiment showed a more pronounced conditioned effect after three associa-
tion trials compared with one, indicating a reinforcement effect. An elevated mast‐cell tryptase in mucosa
was observed when an intranasal saline application was given simultaneously with the CS at recall. Another
type of allergic reaction, the DTH response, was tested in healthy volunteers who received five monthly
tuberculin skin tests (Smith and McDaniel, 1983). In this conditioning protocol, both tuberculin (US) and
saline were injected; the latter was taken from a green vial (CS�) and the former was drawn from a red vial
(CSþ). On the test day, the color labeling of the substances was reversed. Although the saline injections did
not induce a skin reaction (erythema and induration), the severity of the symptoms was significantly
blunted in all the subjects tested when the tuberculin was drawn from the green vial (i.e., conditioned
compensatory effect), where subjects expected their reactions to be negative. However, a similar protocol
using various allergens (e.g., mite dust, fur) taken from colored vials did not result in conditioned
modulation of skin reactions in the subjects tested (Booth et al., 1995).
Another series of experiments investigated the role of conditioning in the context of cancer treatment
and chemotherapy (Bovbjerg, 2003). Some chemotherapeutic agents such as CY have immunosuppressive
effects. For the patient, often the treatment visits to the hospital become aversive and act as CS–US
association trials pairing whereby a variety of features such as white coat, distinct smell when entering
the building, the clinic itself, its smell, the clinician’s voice, and so on may act as distinctive salient stimuli
(CS) that are contingently paired with the chemotherapy (US) (e.g., CY). Immune function was assessed in
cancer patients in the hospital before chemotherapy and compared with assessments conducted at home.
Proliferative responses to T‐cell mitogens were lower for cells isolated from blood samples taken in the
hospital (i.e., after recall) than for home samples (Bovbjerg et al., 1990). These results were replicated in
ovarian cancer patients (Lekander et al., 1995) and pediatric cancer patients receiving chemotherapy (Stock-
horst et al., 2000). In addition, chemotherapy patients often develop conditioned/anticipatory nausea (Andry-
kowski, 1988; Bovbjerg et al., 1990; Morrow et al., 1991; Matteson et al., 2002), anxiety (Jacobsen et al., 1993;
DiLorenzo et al., 1995), and fatigue (Bovbjerg et al., 2005) responses to reminders of chemotherapy. To
resolve such severe consequence of chemotherapy treatment, one may use some of the learning principles
that have been shown to apply to neuroimmune associative learning (> Section 4), for example, latent
inhibition to prevent such undesirable conditioning effects. Several visits to the clinic where the chemo-
therapy will take place should be recommended before starting the chemotherapy. Another possible
behavioral prophylactic therapy may be accomplished before chemotherapy, a specific novel stimulus
that could be administered under the physician’s control, for example, a novel and distinctive tasting/
flavored/colored/sparkling solution. Such artificial stimulus will be categorized and associated to negative
chemotherapy hedonic values; however, it may prevent the development of possible associations to other
high‐protein/caloric food input (Scalera, 2002).
Regarding conditioning of cellular immune parameters, one group assessed the conditionability of
augmentation of NK‐cell numbers and their lytic activity in healthy subjects. Although a conditioned
response was evoked after pairing a given taste with subcutaneous administered adrenaline (Buske‐Kirschbaum et al., 1992, 1994), these effects could not be replicated (Kirschbaum et al., 1992). In multiple
sclerosis patients, four monthly CY infusions (US) were paired with an anise‐flavored syrup (CS) (Giang
et al., 1996). Long‐term treatment with CY decreases blood leukocyte numbers, which often leads to
leukopenia. After a long period such as 6 months of administering the placebo infusion paired with the
drink, 8 out of 10 patients showed a conditioned reduction in peripheral leukocytes numbers. In addition,
by pairing subcutaneously interferon‐g injections (US) with a distinctively flavored drink (CS), it was
possible to induce an elevation of neopterin and quinolinic acid serum levels after evoking such an
association in healthy volunteers (Longo et al., 1999). However, it has been hypothesized that more than
a single associative learning trial pairing a distinctive taste (CS) with interferon‐b injections (US) is

Neuro-immune associative learning 6 141
necessary to produce immune‐conditioned effects (Goebel et al., 2005). This view is supported by experi-
mental data for healthy male volunteers where the immunosuppressive drug cyclosporine A (US) was
paired four times with a distinctively flavored/colored solution (CS) (Goebel et al., 2002), inducing taste–
immune associative learning. After association, the drink (CS) alone induced conditioned inhibition of
ex vivo cytokine (IL‐2 and IFN‐g), mRNA expression, and cytokine release, as well as of the proliferative
responsiveness of human peripheral blood lymphocytes, similar to the CsA effect. In addition, a study with
patients suffering from allergic house‐dust mite rhinitis received pairing of a novel‐tasting drink (CS) and
the H1‐receptor antagonist, desloratadine (US) on five consecutive days. On reexposure, conditioned
patients displayed a decreased basophil activation, assessed in a skin prick test, and subjective symptom
score to a degree that was similar to drug treated control group (Goebel et al., 2007).
In summary, the reported results imply that introduction of behavioral conditioning to supplement
standard pharmacotherapeutic treatment may enable the physician to maintain some desired physiological
state or diminish pathological states. The outlook will be of reducing undesired side effects and maximizing
the effects of pharmacological therapies with a lower cumulative amount of active drug than is currently
used (Ader and Cohen, 1985).
7 Methodological Considerations for Neuroimmune AssociativeLearning Experiments
The following methodological considerations may improve neuroimmune associative learning establish-
ment in new settings as well as its further experimental development and clinical translation.
Ader (2003) suggested the following experimental groups to be included in conditioning protocols,
testing its impact on immune parameters:
Conditioned group: It constitutes the primary experimental group. After CS–US pairing during associa-
tion, this group is reexposed to the CS during recall.
Conditioned not reexposed group: This group is identically treated as conditioned group during associa-
tion phase, and it is not reexposed to the CS during recall phase. It serves as a control for any direct or
indirect immunomodulating effects of the association procedure per se, as well as for possible residual
effects of the US at time of recall.
Unconditioned response group: This group is also identically treated to conditioned group during
association, and receives additionally US reexposure during recall without the CS. This is the pharmaco-
logical control that defines the magnitude and direction of the UR, and also controls for immunological and
other assays being performed to gather the data.
Latent inhibition group: This group is preexposed to the CS before submitted to the entire conditioning
procedure identical to conditioned group. Here, a reduced CR is expected since preexposure should weaken
the strength of association.
Noncontingent conditioned group: This group receives the same number and amount of stimuli as
conditioned group, but CS and US are not timely paired. During recall, this group is also reexposed to the
CS. It controls for nonassociative factors, and certifies the neutrality of the CS in terms of immunological
effects.
Placebo group: This group is exposed to the CS at times as conditioned group is, but never receives the
US; instead, an immunologically neutral stimulus such as saline is administered as placebo. It controls for
residual CS effects and possible procedural artifacts like handling, injections, and so on.
The difficulty for the investigator lies not so much in inducing conditioned immune alterations, but in
employing the proper controls, both immunological and psychological, to demonstrate the predictability
and reliability of such effects, and consequently, the controllability. Therefore, the more control groups one
employs, the better the data are to interpret. Although it is a matter of costs and in the case of human
studies, in particular such involving patients, an ethical issue.
Some considerations about the nature of both the US and CS are provided in the following paragraphs,
complementing previous elaborations (Spector, 1987).

142 6 Neuro-immune associative learning
The unconditioned stimulus should be known in detail in terms of immunologic sequel, and the sequel
must be reproducible. This means that applying a drug with an attenuating effect with subsequent
administration is useless for more than one association trial, since different immunological statuses
would be paired with the CS. In addition, the dose level must be reproducible. For ethical reasons, the
drug should have minimal side effects when experimenting with humans.
The conditioned stimulus should be easily perceived by each subject. This means that it should be novel
and distinct. This is not trivial, since the experimenter needs to find a stimulus that is new to a diverse
population that encountered numerous tastes, odors, colors, and so on in daily routine. Note also
that for instance about 30% of the human population may be anosmic to camphor odor (Spector, 1987).
The CS must also be continuously perceived by each subject without attenuation on repetition, similarly
to the US. It must be immunologically neutral and harmless. The conditioning procedure must be carried
out in a constant environment, since its cues have been proven to be part of the CS. The conditioning
procedures often inadvertently manipulate additional stimulus components that may not be identified
explicitly (e.g., the deodorant or voice of the experimenter, the smell of the room, the sound of a church
bell nearby, etc.). A lack of attention to environmental components of the CS may make conditioned effects
in the immune system appear to be small, although they may just be masked by the presence of environ-
mental cues. In addition, it should be taken into account that the development of the CR depends on
the modality of the CS; aversions produced by association with drug‐induced visceral illness are
often specific to gustatory or olfactory components of the CS (Garcia and Koelling, 1967a). The procedure
should be carried out at the same time of the day, since most immunological parameters underlie circadian
rhythm.
Finally, some issues should be solved to standardize a given conditioning protocol with a clinical
perspective. For instance, it is unknown how long the conditioned response lasts. Is reconditioning
possible (to maintain the conditioned response in a longer perspective, extinction must be avoided)?
When does forgetting set in? Are side effects of the US also conditioned? How controllable is the
conditioned response in a population that in contrast to a laboratory animal comprises differing immune
and psychological history/status? In addition, other factors such as age, gender, and cultural background
must be taken into consideration by health practitioners implementing behavioral conditioning as a
supplement therapy.
8 Closing Remarks
Neuroimmune associative learning is one of the most fascinating examples which uncover the multiple
interactions among the nervous, endocrine, and immune systems. Contributions to this line of research are
coming from various fields of expertise, from the molecular to the behavioral level. This behavioral model is
unique in elucidating the CNS–immune interactions, because it unites the afferent (during association
phase) and efferent (during evocation phase) pathway between these systems in one model, but with the
experimental possibility to dissect such interactions. The potential clinical application to use CRs affecting
immunity as supplementary treatment in a therapeutic setting is a further argument to promote the
understanding of the principles behind neuroimmune associative learning.
References
Ader R. 1974. Letter: Behaviorially conditioned immunosup-
pression. Psychosom Med 36: 183-184.
AderR. 2003. Conditioned immunomodulation:Researchneeds
and directions. Brain Behav Immun 17 (Suppl 1): S51-S57.
Ader R, Cohen N. 1975. Behaviorally conditioned immuno-
suppression. Psychosom Med 37: 333-340.
Ader R, Cohen N. 1982. Behaviorally conditioned immuno-
suppression and murine systemic lupus erythematosus.
Science 215: 1534-1536.
Ader R, Cohen N. 1985. CNS-immune system interac-
tions: Conditioning phenomena. Beh Brain Sci 8:
379-394.

Neuro-immune associative learning 6 143
Ader R, Cohen N. 1992. Conditioned immunopharmacologic
effects on cell‐mediated immunity. Int J Immunopharma-
col 14: 323-327.
Ader R, Cohen N, Bovbjerg D. 1982. Conditioned suppression
of humoral immunity in the rat. J Comp Physiol Psychol
96: 517-521.
Ader R, Cohen N, Grota LJ. 1979. Adrenal involvement
in conditioned immunosuppression. Int J Immunophar-
macol 1: 141-145.
Ader R, Kelly K, Moynihan JA, Grota LJ, Cohen N. 1993.
Conditioned enhancement of antibody production using
antigen as the unconditioned stimulus. Brain Behav
Immun 7: 334-343.
Allen GV, Saper CB, Hurley KM, Cechetto DF. 1991. Organi-
zation of visceral and limbic connections in the insular
cortex of the rat. J Comp Neurol 311: 1-16.
Alvarez‐Borda B, Ramirez‐Amaya V, Perez‐Montfort R,
Bermudez‐Rattoni F. 1995. Enhancement of antibody pro-
duction by a learning paradigm. Neurobiol Learn Mem 64:
103-105.
Andrykowski MA. 1988. Defining anticipatory nausea and
vomiting: Differences among cancer chemotherapy patients
who report pretreatment nausea. J Behav Med 11: 59-69.
ArticoM, Bosco S, Cavallotti C, Agostinelli E, Giuliani‐Piccari G,
et al. 2002. Noradrenergic and cholinergic innervation of
the bone marrow. Int J Mol Med 10: 77-80.
Avital A, Goshen I, Kamsler A, Segal M, Iverfeldt K, et al. 2003.
Impaired interleukin‐1 signaling is associated with deficits
in hippocampal memory processes and neural plasticity.
Hippocampus 13: 826-834.
Baciu I, Sotuz V, Stoica N, Raducanu N. 1965. Sur les centres
nerveux regulateurs de l’erythropoiese. Rev Roum Physiol
2: 123-133.
Balschun D, Wetzel W, Del Rey A, Pitossi F, Schneider H, et al.
2004. Interleukin‐6: A cytokine to forget. FASEB J 18:
1788-1790.
Banks WA. 2006. The blood–brain barrier in psychoneuroim-
munology. Neurol clin 24: 413-419.
Banks WA, Farr SA, La Scola ME, Morley JE. 2001. Intrave-
nous human interleukin‐1alpha impairs memory proces-
sing in mice: Dependence on blood–brain barrier transport
into posterior division of the septum. J Pharmacol Exp
Ther 299: 536-541.
Barrett JE, King MG, Pang G. 2000. Conditioning rhinitis in
allergic humans. Ann N YAcad Sci 917: 853-859.
Benetato G. 1955. [Central nervous mechanism of the leuko-
cytic and phagocytic reaction]. J Physiol (Paris) 47:
391-403.
Berman DE, Dudai Y. 2001. Memory extinction, learning
anew, and learning the new: Dissociations in the molec-
ular machinery of learning in cortex. Science 291:
2417-2419.
Bermudez‐Rattoni F. 2004. Molecular mechanisms of taste‐
recognition memory. Nat Rev Neurosci 5: 209-217.
Bermudez‐Rattoni F, Introini‐Collison I, Coleman‐Mesches K,
McGaugh JL. 1997. Insular cortex and amygdala lesi-
ons induced after aversive training impair retention:
Effects of degree of training. Neurobiol Learn Mem 67:
57-63.
Bermudez‐Rattoni F, Ramirez‐Lugo L, Gutierrez R, MirandaMI.
2004. Molecular signals into the insular cortex and amyg-
dala during aversive gustatory memory formation. Cell
Mol Neurobiol 24: 25-36.
Bernardis LL, Bellinger LL. 1993. The lateral hypothalamic
area revisited: Neuroanatomy, body weight regulation,
neuroendocrinology and metabolism. Neurosci Biobehav
Rev 17: 141-193.
Besedovsky HO, del Rey A. 1996. Immune‐neuro‐endocrine
interactions: Facts and hypotheses. Endocr Rev 17: 64-102.
Blalock JE. 1994. The syntax of immune–neuroendocrine
communication. Immunol Today 15: 504-511.
Blalock JE. 2005. The immune system as the sixth sense.
J Intern Med 257: 126-138.
Booth RJ, Petrie KJ, Brook RJ. 1995. Conditioning allergic
skin responses in humans: A controlled trial. Psychosom
Med 57: 492-495.
Bovbjerg DH. 2003. Conditioning, cancer, and immune regu-
lation. Brain Behav Immun 17 (Suppl 1): S58-S61.
Bovbjerg D, Ader R, Cohen N. 1984. Acquisition and extinc-
tion of conditioned suppression of a graft‐vs‐host response
in the rat. J Immunol 132: 111-113.
Bovbjerg D, Cohen N, Ader R. 1987. Behaviorally conditioned
enhancement of delayed‐type hypersensitivity in the
mouse. Brain Behav Immun 1: 64-71.
Bovbjerg DH, Montgomery GH, Raptis G. 2005. Evidence for
classically conditioned fatigue responses in patients receiv-
ing chemotherapy treatment for breast cancer. J Behav Med
28: 231-237.
Bovbjerg DH, Redd WH, Maier LA, Holland JC, Lesko LM,
et al. 1990. Anticipatory immune suppression and nausea
in women receiving cyclic chemotherapy for ovarian can-
cer. J Consult Clin Psychol 58: 153-157.
Buijs RM, Scheer FA, Kreier F, Yi C, Bos N, et al. 2006.
Organization of circadian functions: Interaction with the
body. Prog Brain Res 153: 341-360.
Bull DF, Brown R, King MG, Husband AJ. 1991. Modulation
of body temperature through taste aversion conditioning.
Physiol Behav 49: 1229-1233.
Buske‐Kirschbaum A, Kirschbaum C, Stierle H, Jabaij L,
Hellhammer D. 1994. Conditioned manipulation of natu-
ral killer (NK) cells in humans using a discriminative
learning protocol. Biol Psychol 38: 143-155.
Buske‐Kirschbaum A, Kirschbaum C, Stierle H, Lehnert H,
Hellhammer D. 1992. Conditioned increase of natural killer

144 6 Neuro-immune associative learning
cell activity (NKCA) in humans. Psychosom Med 54:
123-132.
Butcher KS, Cechetto DF. 1998. Neurotransmission in
the medulla mediating insular cortical and lateral
hypothalamic sympathetic responses. Can J Physiol
Pharmacol 76: 737-746.
Cechetto DF, Chen SJ. 1992. Hypothalamic and cortical sym-
pathetic responses relay in the medulla of the rat. Am
J Physiol 263: 544-552.
Chao HJ, Hsu YC, Yuan HP, Jiang HS, Hsueh CM.
2005. The conditioned enhancement of neutrophil activ-
ity is catecholamine dependent. J Neuroimmunol 158:
159-169.
Chen J, Lin W, Wang W, Shao F, Yang J, et al. 2004. Enhance-
ment of antibody production and expression of c‐Fos in the
insular cortex in response to a conditioned stimulus after
a single‐trial learning paradigm. Behav Brain Res 154:
557-565.
Cohen N, Ader R, Green N, Bovbjerg D. 1979. Conditioned
suppression of a thymus-independent antibody response.
Psychosom Med 41: 487-91.
Costa‐Pinto FA, Basso AS, Britto LR, Malucelli BE, Russo M.
2005. Avoidance behavior and neural correlates of allergen
exposure in a murine model of asthma. Brain Behav
Immun 19: 52-60.
Coussons‐Read ME, Dykstra LA, Lysle DT. 1994. Pavlovian
conditioning of morphine‐induced alterations of immune
status: Evidence for opioid receptor involvement. J Neu-
roimmunol 55: 135-142.
Cunnick JE, Lysle DT, Kucinski BJ, Rabin BS. 1990. Evidence
that shock‐induced immune suppression is mediated by
adrenal hormones and peripheral beta‐adrenergic recep-
tors. Pharmacol Biochem Behav 36: 645-651.
Cutolo M, Capellino S, Sulli A, Serioli B, Secchi ME, et al.
2006. Estrogens and autoimmune diseases. Ann N Y Acad
Sci 1089: 538-547.
Czura CJ, Tracey KJ. 2005. Autonomic neural regulation of
immunity. J Intern Med 257: 156-166.
Dantzer R. 2004. Cytokine‐induced sickness behaviour: A
neuroimmune response to activation of innate immunity.
Eur J Pharmacol 500: 399-411.
Dantzer R, Konsman JP, Bluthe RM, Kelley KW. 2000. Neural
and humoral pathways of communication from the im-
mune system to the brain: Parallel or convergent? Auton
Neurosci 85: 60-65.
Dark K, Peeke HV, Ellman G, Salfi M. 1987. Behaviorally
conditioned histamine release. Prior stress and condition-
ability and extinction of the response. Ann N Y Acad Sci
496: 578-582.
Dekker E, Pelser HE, Groen J. 1957. Conditioning as a cause
of asthmatic attacks; a laboratory study. J Psychosom Res 2:
97-108.
Del Rey A, Randolf A, Pitossi F, Rogausch H, Besedovsky HO.
2000. Not all peripheral immune stimuli that activate
the HPA axis induce proinflammatory cytokine gene
expression in the hypothalamus. Ann N Y Acad Sci 917:
169-174.
Demissie S, Ghanta VK, Hiramoto NS, Hiramoto RN. 2000.
NK cell and CTL activities can be raised via conditioning
of the CNS with unrelated unconditioned stimuli. Int
J Neurosci 103: 79-89.
Demissie S, Rogers CF, Hiramoto NS, Ghanta VK,
Hiramoto RN. 1995. Arecoline a muscarinic cholinergic
agent conditions central pathways that modulate natural
killer cell activity. J Neuroimmunol 59: 57-63.
DiLorenzo TA, Jacobsen PB, Bovbjerg DH, Chang H,
Hudis CA, et al. 1995. Sources of anticipatory emotional
distress in women receiving chemotherapy for breast
cancer. Ann Oncol 6: 705-711.
Djuric VJ, Markovic BM, Lazarevic M, Jankovic BD. 1988.
Anaphylactic shock‐induced conditioned taste aversion. II.
Correlation between taste aversion and indicators of ana-
phylactic shock. Brain Behav Immun 2: 24-31.
Dolin AO, Krylov VN. 1952. [Role of the cerebral cortex in
immunologic reactions of the organism]. Zh Vyssh Nerv
Deiat Im I P Pavlova 2: 547-560.
Domjan M. 2005. Pavlovian conditioning: A functional per-
spective. Annu Rev Psychol 56: 179-206.
Domjan M, Cusato B, Krause M. 2004. Learning with arbi-
trary versus ecological conditioned stimuli: Evidence from
sexual conditioning. Psychon Bull Rev 11: 232-246.
Doroshkevich AA. 1954. [Effect of conditioned stimuli on
formation of immunological reactions]. Zh Vyssh Nerv
Deiat Im I P Pavlova 4: 108-115.
Downing JE, Miyan JA. 2000. Neural immunoregulation:
Emerging roles for nerves in immune homeostasis and
disease. Immunol Today 21: 281-289.
Dudai Y. 2006. Reconsolidation: The advantage of being refo-
cused. Curr Opin Neurobiol 16: 174-178.
Dyck DG, Driedger SM, Nemeth R, Osachuk TA, Greenberg
AH. 1987. Conditioned tolerance to drug‐induced (poly I:
C) natural killer cell activation: Effects of drug‐dosage
and context‐specificity parameters. Brain Behav Immun 1:
251-266.
Dyck DG, Janz L, Osachuk TA, Falk J, Labinsky J, et al. 1990.
The Pavlovian conditioning of IL‐1‐induced glucocorticoid
secretion. Brain Behav Immun 4: 93-104.
Eikelboom R, Stewart J. 1982. Conditioning of drug‐induced
physiological responses. Psychol Rev 89: 507-528.
Eisenberg M, Kobilo T, Berman DE, Dudai Y. 2003. Stability
of retrieved memory: Inverse correlation with trace domi-
nance. Science 301: 1102-1104.
Elenkov IJ, Wilder RL, Chrousos GP, Vizi ES. 2000. The
sympathetic nerve – an integrative interface between two

Neuro-immune associative learning 6 145
supersystems: The brain and the immune system. Pharma-
col Rev 52: 595-638.
Elmquist JK, Scammell TE, Saper CB. 1997. Mechanisms of
CNS response to systemic immune challenge: The febrile
response. Trends Neurosci 20: 565-570.
Espinosa E, Calderas T, Flores‐Mucino O, Perez‐Garcia G,
Vazquez‐Camacho AC, et al. 2004. Enhancement of anti-
body response by one‐trial conditioning: Contrasting results
using different antigens. Brain Behav Immun 18: 76-80.
Exton MS, Elfers A, Jeong WY, Bull DF, Westermann J, et al.
2000. Conditioned suppression of contact sensitivity is
independent of sympathetic splenic innervation. Am
J Physiol 279: 1310-1315.
Exton MS, Gierse C, Meier B, Mosen M, Xie Y, et al. 2002.
Behaviorally conditioned immunosuppression in the rat
is regulated via noradrenaline and beta‐adrenoceptors.
J Neuroimmunol 131: 21-30.
Exton MS, Herklotz J, Westermann J, Schedlowski M. 2001.
Conditioning in the rat: An in vivo model to investigate the
molecular mechanisms and clinical implications of brain–
immune communication. Immunol Rev 184: 226-235.
Exton MS, Schult M, Donath S, Strubel T, Bode U, et al. 1999.
Conditioned immunosuppression makes subtherapeutic
cyclosporin effective via splenic innervation. Am J Physiol
276: 1710-1717.
Exton MS, von Horsten S, Schult M, Voge J, Strubel T, et al.
1998. Behaviorally conditioned immunosuppression using
cyclosporine A: Central nervous system reduces IL‐2
production via splenic innervation. J Neuroimmunol 88:
182-191.
Fiore M, Angelucci F, Alleva E, Branchi I, Probert L, et al.
2000. Learning performances, brain NGF distribution and
NPY levels in transgenic mice expressing TNF‐alpha. Behav
Brain Res 112: 165-175.
Garcia J, Koelling RA. 1967a. A comparison of aversions
induced by X‐rays, toxins, and drugs in the rat. Radiat
Res (Suppl 7): 439–450.
Garcia J, Koelling RA. 1967b. The relationship of cue to
consequence in avoidance learning. Psychon Sci 4: 123-124.
Gauci M, Husband AJ, Saxarra H, King MG. 1994. Pavlovian
conditioning of nasal tryptase release in human subjects
with allergic rhinitis. Physiol Behav 55: 823-825.
Ghanta V, Hiramoto RN, Solvason B, Spector NH. 1987.
Influence of conditioned natural immunity on tumor
growth. Ann N YAcad Sci 496: 637-646.
Ghanta VK, Hiramoto NS, Solvason HB, Soong SJ, Hiramoto
RN. 1990. Conditioning: A new approach to immunother-
apy. Cancer Res 50: 4295-4299.
Ghanta VK, Hiramoto NS, Soong SJ, Hiramoto RN. 1995.
Conditioning of the secondary cytotoxic T-lymphocyte
response to YC8 tumor. Pharmacol Biochem Behav 50:
399-403.
Ghanta VK, Miura T, Hiramoto NS, Hiramoto RN. 1988.
Augmentation of natural immunity and regulation of
tumor growth by conditioning. Ann N Y Acad Sci 521:
29-42.
Ghanta VK, Rogers CF, Hsueh CM, Demissie S, Lorden JF,
et al. 1994. Role of arcuate nucleus of the hypothalamus in
the acquisition of association memory between the CS and
US. J Neuroimmunol 50: 109-114.
Giang DW, Goodman AD, Schiffer RB, Mattson DH,
Petrie M, et al. 1996. Conditioning of cyclophosphamide‐
induced leukopenia in humans. J Neuropsychiatry Clin
Neurosci 8: 194-201.
Gibertini M. 1996. IL1 beta impairs relational but not proce-
dural rodent learning in a water maze task. Adv Exp Med
Biol 402: 207-217.
Glaser R, Kiecolt‐Glaser JK. 2005. Stress‐induced immune
dysfunction: Implications for health. Nat Rev Immunol 5:
243-251.
Goebel MU, Hubell D, Kou W, Janssen OE, Katsarava Z, et al.
2005. Behavioral conditioning with interferon beta‐1a in
humans. Physiol Behav 84: 807-814.
Goebel MU, Meykadeh N, Kou W, Schedlowski M, Hengge
UR. 2007. Behavioral conditioning of antihistamine effects
in patients with allergic rhinitis. Psychother Psychosom (in
press).
Goebel MU, Trebst AE, Steiner J, Xie YF, Exton MS, et al.
2002. Behavioral conditioning of immunosuppression is
possible in humans. FASEB J 16: 1869-1873.
Goehler LE, Gaykema RP, HansenMK, Anderson K, Maier SF,
et al. 2000. Vagal immune‐to‐brain communication: A
visceral chemosensory pathway. Auton Neurosci 85:
49-59.
Gorczynski RM. 1987a. Analysis of lymphocytes in, and host
environment of, mice showing conditioned immuno-
suppression to cyclophosphamide. Brain Behav Immun 1:
21-35.
Gorczynski RM. 1987b. Diversity in the lymphocyte recogni-
tion repertoire is altered during ageing. Biomed Pharmac-
other 41: 124-126.
Gorczynski RM. 1991. Toward an understanding of the
mechanisms of classical conditioning of antibody
responses. J Gerontol 46: P152-P156.
Gorczynski RM, Holmes W. 1989. Neuroleptic and anti‐
depressant drug treatment abolishes conditioned immuno-
suppression in mice. Brain Behav Immun 3: 312-319.
Gorczynski RM, Kennedy M. 1984. Associative learning and
regulation of immune responses. Prog Neuropsychophar-
macol Biol Psychiatry 8: 593–600.
Gorczynski RM, Kennedy M, Ciampi A. 1985. Cimetidine
reverses tumor growth enhancement of plasmacytoma
tumors in mice demonstrating conditioned immunosup-
pression. J Immunol 134: 4261-4266.

146 6 Neuro-immune associative learning
Gorczynski RM, Macrae S, Kennedy M. 1982. Conditioned
immune response associated with allogeneic skin grafts in
mice. J Immunol 129: 704-709.
Gorczynski RM, Macrae S, Kennedy M. 1984. Factors
involved in the classical conditioning of antibody
responses in mice. Breakdown in Human Adaptation to
Stress: Towards a multidisciplinary Approach. Ballieux R,
Fielding J, L’Abbatte A, editors. Boston: Kluwer‐Nijhoff;
pp. 704-712.
Grochowicz PM, Schedlowski M, Husband AJ, King MG,
Hibberd AD, et al. 1991. Behavioral conditioning prolongs
heart allograft survival in rats. Brain Behav Immun 5:
349-356.
Grota LJ, Ader R, Cohen N. 1987. Taste aversion learning in
autoimmune Mrl‐lpr/lpr and Mrlþ/þ mice. Brain Behav
Immun 1: 238-250.
Harris GC, Fitzgerald RD. 1989. Impaired learning of
classically conditioned bradycardia in rats following
fourth ventricle administration of D‐Ala2‐methionine‐
enkephalinamide. Behav Neurosci 103: 77-83.
Hill LE. 1930. Philosophy of a Biologist. London: Arnold.
Hiramoto R, Ghanta V, Lorden J, Solvason H, Soong S‐J, et al.
1992. Conditioning of enhanced natural killer cell activity:
Effects of changing inter‐stimulus intervals and evidence
for long‐delayed learning. Prog Neuroendocrinol 5: 13-20.
Hiramoto R, Ghanta V, Solvason B, Lorden J, Hsueh CM,
et al. 1993. Identification of specific pathways of communi-
cation between the CNS and NK cell system. Life Sci 53:
527-540.
Hiramoto R, Solvason B, Ghanta V, Lorden J, Hiramoto N.
1990. Effect of reserpine on retention of the conditioned
NK cell response. Pharmacol Biochem Behav 36: 51-56.
Hori T, Katafuchi T, Take S, Shimizu N, Niijima A. 1995. The
autonomic nervous system as a communication channel
between the brain and the immune system. Neuroimmu-
nomodulation 2: 203-215.
Hsueh CM, Chen SF, Ghanta VK, Hiramoto RN. 1995.
Expression of the conditioned NK cell activity is beta‐
endorphin dependent. Brain Res 678: 76-82.
Hsueh CM, Chen SF, Lin RJ, Chao HJ. 2002. Cholinergic and
serotonergic activities are required in triggering con-
ditioned NK cell response. J Neuroimmunol 123: 102-111.
Hsueh C, Kuo J, Chen S, Huang H, Cheng F, et al. 1999.
Involvement of catecholamines in recall of the conditioned
NK cell response. J Neuroimmunol 94: 172-181.
Hsueh CM, Lorden JF, Hiramoto RN, Ghanta VK. 1992.
Acquisition of enhanced natural killer cell activity under
anesthesia. Life Sci 50: 2067-2074.
Hsueh CM, Rogers C, Hiramoto RN, Ghanta VK. 1994a.
Effect of dexamethasone on conditioned enhancement of
natural killer cell activity. Neuroimmunomodulation 1:
370-376.
Hsueh CM, Tyring SK, Hiramoto RN, Ghanta VK. 1994b.
Efferent signal(s) responsible for the conditioned augmen-
tation of natural killer cell activity. Neuroimmunomodula-
tion 1: 74-81.
Huang JX, Lin WJ, Chen J. 2004. Antibody response can be
conditioned using electroacupuncture as conditioned stim-
ulus. Neuroreport 15: 1475-1478.
Husband AJ, King MG, Brown R. 1987. Behaviourally
conditioned modification of T cell subset ratios in rats.
Immunol Lett 14: 91-94.
Ikegaya Y, Delcroix I, Iwakura Y, Matsuki N, Nishiyama N.
2003. Interleukin‐1beta abrogates long‐term depression of
hippocampal CA1 synaptic transmission. Synapse 47: 54-57.
Ikemi Y, Kusano T, Fukamachi K. 1962. Psychogenic factors in
headache. Int J Neurol 3: 368-378.
Irie M, Maeda M, Nagata S. 2001. Can conditioned histamine
release occur under urethane anesthesia in guinea pigs?
Physiol Behav 72: 567-573.
Irie M, Nagata S, Endo Y. 2002. Fasting stress exacerbates
classical conditioned histamine release in guinea pigs. Life
Sci 72: 689-698.
Irie M, Nagata S, Endo Y. 2004. Diazepam attenuates
conditioned histamine release in guinea pigs. Int J Psycho-
physiol 51: 231-238.
Jacobsen PB, Bovbjerg DH, Redd WH. 1993. Anticipatory
anxiety in women receiving chemotherapy for breast can-
cer. Health Psychol 12: 469-475.
Janz LJ, Brown R, Zuo L, Falk J, Greenberg AH, et al. 1991.
Conditioned taste aversion but not adrenal activity devel-
ops to ICV administration of interleukin‐1 in rats. Physiol
Behav 49: 691-694.
Jenkins P, Chadwick R, Nevin J. 1983. Classically conditioned
enhancement of antibody production. Bull Psychonom Soc
21: 485-487.
Katafuchi T, Ichijo T, Take S, Hori T. 1993. Hypothalamic
modulation of splenic natural killer cell activity in rats.
J Physiol 471: 209-221.
Katafuchi T, Okada E, Take S, Hori T. 1994. The biphasic
changes in splenic natural killer cell activity following
ventromedial hypothalamic lesions in rats. Brain Res 652:
164-168.
Kelley KW, Dantzer R. 1988. The importance of conditioning
in conditioned immunosuppression. Int J Neurosci 39:
289-297.
Kelley KW, Dantzer R, Mormede P, Salmon H, Aynaud JM.
1984. [Induction of immunosuppression by dietary aver-
sion acquired in the absence of immunosuppressive treat-
ment]. C R Acad Sci III 299: 123-126.
Kelley KW, Dantzer R, Mormede P, Salmon H, Aynaud JM.
1985. Conditioned taste aversion suppresses induction of
delayed‐type hypersensitivity immune reactions. Physiol
Behav 34: 189-193.

Neuro-immune associative learning 6 147
Kiefer SW, Rusiniak KW, Garcia J. 1982. Flavor‐illness aver-
sions: Gustatory neocortex ablations disrupt taste but not
taste‐potentiated odor cues. J Comp Physiol Psychol 96:
540-548.
Kirschbaum C, Jabaaij L, Buske‐Kirschbaum A, Hennig J,
Blom M, et al. 1992. Conditioning of drug‐induced immu-
nomodulation in human volunteers: A European collabo-
rative study. Br J Clin Psychol 31(4): 459-472.
Klosterhalfen W, Klosterhalfen S. 1983. Pavlovian condition-
ing of immunosuppression modifies adjuvant arthritis in
rats. Behav Neurosci 97: 663-666.
Klosterhalfen S, Klosterhalfen W. 1990. Conditioned cyclo-
sporine effects but not conditioned taste aversion in immu-
nized rats. Behav Neurosci 104: 716-724.
Kritzer MF, Brewer A, Montalmant F, Davenport M,
Robinson JK. 2007. Effects of gonadectomy on performance
in operant tasks measuring prefrontal cortical function in
adult male rats. Horm Behav 51: 183-194.
Kuo J, Chen S, Huang H, Yang C, Tsai P, et al. 2001. The
involvement of glutamate in recall of the conditioned NK
cell response. J Neuroimmunol 118: 245-255.
Kusnecov AW, Husband AJ, King MG. 1988. Behaviorally
conditioned suppression of mitogen‐induced proliferation
and immunoglobulin production: Effect of time span be-
tween conditioning and reexposure to the conditioning
stimulus. Brain Behav Immun 2: 198-211.
Kusnecov AW, Sivyer M, King MG, Husband AJ, Cripps AW,
Clancy RL. 1983. Behaviorally conditioned suppression
of the immune response by antilymphocyte serum.
J immunol 130: 2117-2120.
Lacosta S, Merali Z, Anisman H. 2000. Central monoamine
activity following acute and repeated systemic interleukin‐2
administration. Neuroimmunomodulation 8: 83-90.
Lekander M, Furst CJ, Rotstein S, Blomgren H, Fredrikson M.
1995. Anticipatory immune changes in women treated
with chemotherapy for ovarian cancer. Int J Behav Med 2:
1-12.
Longo DL, Duffey PL, KoppWC, Heyes MP, AlvordWG, et al.
1999. Conditioned immune response to interferon‐gamma
in humans. Clin Immunol 90: 173-181.
Lukyanenko VI. 1961. The functional structure of the immu-
nogenetic process and its nervous regulation. Folia Biol
(Praha) 7: 379-389.
Lynch MA. 2002. Interleukin‐1 beta exerts a myriad of effects
in the brain and in particular in the hippocampus: Analysis
of some of these actions. Vitam Horm 64: 185-219.
Lysle DT, Cunnick JE, Fowler H, Rabin BS. 1988. Pavlovian
conditioning of shock‐induced suppression of lymphocyte
reactivity: Acquisition, extinction, and preexposure effects.
Life Sci 42: 2185-2194.
Lysle DT, Cunnick JE, Maslonek KA. 1991. Pharmacological
manipulation of immune alterations induced by an
aversive conditioned stimulus: Evidence for a beta‐adren-
ergic receptor‐mediated Pavlovian conditioning process.
Behav Neurosci 105: 443-449.
Lysle DT, Cunnick JE, Rabin BS. 1990. Stressor‐induced alter-
ation of lymphocyte proliferation in mice: Evidence for
enhancement of mitogenic responsiveness. Brain Behav
Immun 4: 269-277.
Lysle DT, Luecken LJ, Maslonek KA. 1992. Suppression of the
development of adjuvant arthritis by a conditioned aversive
stimulus. Brain Behav Immun 6: 64-73.
Mackenzie JN. 1886. The production of the so‐called ‘‘rose
cold’’ by means of an artificial rose. Am JMed Sci 91: 45-47.
MacQueen G, Marshall J, Perdue M, Siegel S, Bienenstock J.
1989. Pavlovian conditioning of rat mucosal mast cells to
secrete rat mast cell protease II. Science 243: 83-85.
Madden KS, Boehm GW, Lee SC, Grota LJ, Cohen N, et al.
2001. One‐trial conditioning of the antibody response to
hen egg lysozyme in rats. J Neuroimmunol 113: 236-239.
Makukahin I. 1911. O Leykocitoze. Dissert. St. Petersburg.
Markovic BM, Djuric VJ, Lazarevic M, Jankovic BD. 1988.
Anaphylactic shock‐induced conditioned taste aversion. I.
Demonstration of the phenomenon by means of three
modes of CS–US presentation. Brain Behav Immun 2:
11-23.
Matsumoto Y, Yoshida M, Watanabe S, Yamamoto T. 2001.
Involvement of cholinergic and glutamatergic functions in
working memory impairment induced by interleukin‐1beta
in rats. Eur J Pharmacol 430: 283-288.
Matsumoto Y, Watanabe S, Suh YH, Yamamoto T. 2002.
Effects of intrahippocampal CT105, a carboxyl terminal
fragment of beta‐amyloid precursor protein, alone/with
inflammatory cytokines on working memory in rats. J
Neurochem 82: 234-239.
Matteson S, Roscoe J, Hickok J, Morrow GR. 2002. The role of
behavioral conditioning in the development of nausea. Am
J Obstet Gynecol 186: S239-S243.
Meisel C, Schwab JM, Prass K, Meisel A, Dirnagl U. 2005.
Central nervous system injury‐induced immune deficiency
syndrome. Nat Rev Neurosci 6: 775-786.
Metal’nikov S, Chorine V. 1926. Role des reflexes condition-
nels dans l’immunite. Ann L’Inst Pasteur 40: 893-900.
Metal’nikov S, Chorine V. 1928. Role des reflexes condition-
nels dans la formation des anticorps. C R Soc Biol 102:
133-134.
Mignini F, Streccioni V, Amenta F. 2003. Autonomic innerva-
tion of immune organs and neuroimmune modulation.
Auton Autacoid Pharmacol 23: 1-25.
Miyan JA, Broome CS, Whetton AD. 1998. Neural regulation
of bone marrow. Blood 92: 2971-2973.
Mormede P, Dantzer R, Michaud B, Kelley KW, Le Moal M.
1988. Influence of stressor predictability and behavioral
control on lymphocyte reactivity, antibody responses

148 6 Neuro-immune associative learning
and neuroendocrine activation in rats. Physiol Behav 43:
577-583.
Morrow GR, Lindke J, Black PM. 1991. Anticipatory nausea
development in cancer patients: Replication and extension
of a learning model. Br J Psychol 82: 61-72.
Nerad L, Ramirez‐Amaya V, Ormsby CE, Bermudez‐Rattoni
F. 1996. Differential effects of anterior and posterior
insular cortex lesions on the acquisition of conditioned
taste aversion and spatial learning. Neurobiol Learn Mem
66: 44-50.
Neveu PJ, Dantzer R, Le Moal M. 1986. Behaviorally
conditioned suppression of mitogen‐induced lymphopro-
liferation and antibody production in mice. Neurosci Lett
65: 293-298.
Nicolau I, Antinescu‐Dimitriu O. 1929. Roles des reflexes
conditionnels dans, la formation des anticorps. I Compt
Rend d I Soc Biol 102: 144-145.
Niemi M‐B, Haerting M, Kou W, del Rey A, Besedovsky HO,
Schedlowski M, Pacheco-Lopez G. 2007. Taste‐immuno-
suppression engram: Reinforcement and extinction.
J Neuroimmunol (in press).
Niemi MB, Pacheco‐Lopez G, Kou W, Harting M, del Rey A,
et al. 2006. Murine taste‐immune associative learning.
Brain Behav Immun 20: 527-531.
Noelpp B, Noelpp‐Eschenhagen I. 1951a. [Experimental
bronchial asthma in guinea pigs, Part I. Methods for objec-
tive registration of asthma attacks]. Int Arch Allergy Appl
Immunol 2: 308-320.
Noelpp B, Noelpp‐Eschenhagen I. 1951b. [Experimental
bronchial asthma in guinea pigs. Part II. The role of
conditioned reflexes in the pathogenesis of bronchial
asthma]. Int Arch Allergy Appl Immunol 2: 321-329.
Noelpp B, Noelpp‐Eschenhagen I. 1951c. [Role of
conditioned reflex in bronchial asthma; experimental
investigation on the pathogenesis of bronchial asthma].
Helv Med Acta 18: 142-158.
Noelpp B, Noelpp‐Eschenhagen I. 1952a. [Experimental
bronchial asthma in the guinea pig. IV. Experimental
asthma in the guinea pig as an experimental model]. Int
Arch Allergy Appl Immunol 3: 207-217.
Noelpp B, Noelpp‐Eschenhagen I. 1952b. [Experimental
bronchial asthma of guinea‐pigs. III. Studies on the sig-
nificance of conditioned reflexes; ability to develop
conditioned reflexes and their duration under stress]. Int
Arch Allergy Appl Immunol 3: 108-136.
Noelpp B, Noelpp‐Eschenhagen I. 1952c. [The experimental
bronchial asthma of the guinea pig. V. Experimental patho-
physiological studies]. Int Arch Allergy Appl Immunol 3:
302-323.
Nomura M, Hori K. 1996. The aging effects on learning and
memory. Neurosci Res 25: 14-14(1).
Okamoto S, Ibaraki K, Hayashi S, Saito M. 1996. Ventrome-
dial hypothalamus suppresses splenic lymphocyte activity
through sympathetic innervation. Brain Res 739: 308-313.
Oppenheimer SM, Saleh T, Cechetto DF. 1992. Lateral hypo-
thalamic area neurotransmission and neuromodulation of
the specific cardiac effects of insular cortex stimulation.
Brain Res 581: 133-142.
Ostrovskaya OA. 1929. The conditioned reflex and the immu-
nologic reaction. J Exp Biol Med 12: 174-182.
Ottenberg P, Stein M, Lewis J, Hamilton C. 1958. Learned
asthma in the guinea pig. Psychosom Med 20: 395-400.
Pacheco‐Lopez G, Niemi MB, Engler H, Engler A, Riether C,
et al. 2007a. Weaken taste‐LPS association during endotox-
in tolerance. Physiol Behav (in press).
Pacheco‐Lopez G, Niemi MB, Engler H, Schedlowski M.
2007b. Neuro‐immune associative learning. Neural Plastic-
ity and Memory. From Genes to Brain Imaging, First
edition. Bermudez‐Rattoni F, editor. Boca Raton: CRC
Press; pp: 285-310.
Pacheco‐Lopez G, Niemi MB, Engler H, Schedlowski M.
2006. Conditioned enhancement of immune responses.
Ader R, Felten DL, Cohen N, editors. Psychoneuroimmu-
nology, Fourth edition. Academic Press; San Diego:
pp. 631-660.
Pacheco‐Lopez G, Niemi MB, Kou W, Harting M, Del Rey A,
et al. 2004. Behavioural endocrine immune‐conditioned
response is induced by taste and superantigen pairing.
Neuroscience 129: 555-562.
Pacheco‐Lopez G, Niemi MB, Kou W, Harting M, Fandrey J,
et al. 2005. Neural substrates for behaviorally conditioned
immunosuppression in the rat. J Neurosci 25: 2330-2337.
Palermo‐Neto J, Guimaraes RK. 2000. Pavlovian conditioning
of lung anaphylactic response in rats. Life Sci 68: 611-623.
Pavlov IP. 1927. Conditioned Reflexes. London: Oxford
University Press.
Pavlov VA, Tracey KJ. 2005. The cholinergic anti‐inflammato-
ry pathway. Brain Behav Immun 19: 493-499.
Peeke HV, Dark K, Ellman G, McCurry C, Salfi M. 1987a.
Prior stress and behaviorally conditioned histamine release.
Physiol Behav 39: 89-93.
Peeke HV, Ellman G, Dark K, Salfi M, Reus VI. 1987b. Corti-
sol and behaviorally conditioned histamine release. Ann N
YAcad Sci 496: 583-587.
Pitossi F, del Rey A, Kabiersch A, Besedovsky H. 1997. Induc-
tion of cytokine transcripts in the central nervous system
and pituitary following peripheral administration of endo-
toxin to mice. J Neurosci Res 48: 287-298.
Rachal Pugh C, Fleshner M, Watkins LR, Maier SF, Rudy JW.
2001. The immune system and memory consolidation: A
role for the cytokine IL‐1beta. Neurosci Biobehav Rev 25:
29-41.

Neuro-immune associative learning 6 149
Ramırez‐Amaya V, Alvarez‐Borda B, Bermudez‐Rattoni F.
1998. Differential effects of NMDA‐induced lesions into
the insular cortex and amygdala on the acquisition and
evocation of conditioned immunosuppression. Brain
Behav Immun 12: 149-160.
Ramırez‐Amaya V, Alvarez‐Borda B, Ormsby C, Martınez R,
Perez‐Montfort R, et al. 1996. Insular cortex lesions impair
the acquisition of conditioned immunosuppression. Brain
Behav Immun 10: 103-114.
Ramırez‐Amaya V, Bermudez‐Rattoni F. 1999. Conditioned
enhancement of antibody production is disrupted by insu-
lar cortex and amygdala but not hippocampal lesions. Brain
Behav Immun 13: 46-60.
Reilly S, Bornovalova MA. 2005. Conditioned taste aversion
and amygdala lesions in the rat: A critical review. Neurosci
Biobehav Rev 29: 1067-1088.
Rogers CF, Ghanta VK, Demissie S, Hiramoto N, Hiramoto R.
1994a. Lidocaine interrupts the conditioned natural killer
cell response by interfering with the conditioned stimulus.
Neuroimmunomodulation 1: 278-283.
Rogers C, Ghanta V, Demissie S, Hiramoto N, Hiramoto R.
1994b. Sodium carbonate prevents NK cell conditioning by
interfering with the US signal. Int J Neurosci 77: 277-286.
Rogers C, Ghanta V, Hsueh CM, Hiramoto N, Hiramoto R.
1992. Indomethacin and sodium carbonate effects on
conditioned fever and NK cell activity. Pharmacol Biochem
Behav 43: 417-422.
Rogers MP, Reich P, Strom TB, Carpenter CB. 1976. Behav-
iorally conditioned immunosuppression: Replication of a
recent study. Psychosom Med 38: 447-451.
Roudebush RE, Bryant HU. 1991. Conditioned immunosup-
pression of a murine delayed type hypersensitivity re-
sponse: Dissociation from corticosterone elevation. Brain
Behav Immun 5: 308-317.
Saito M, Minokoshi Y, Shimazu T. 1989. Accelerated norepi-
nephrine turnover in peripheral tissues after ventromedial
hypothalamic stimulation in rats. Brain Res 481: 298-303.
Sanders VM, Kohm AP. 2002a. Sympathetic nervous system
interaction with the immune system. Int Rev Neurobiol 52:
17-41.
Sanders VM, Straub RH. 2002b. Norepinephrine, the beta‐
adrenergic receptor, and immunity. Brain Behav Immun
16: 290-332.
Scalera G. 2002. Effects of conditioned food aversions on
nutritional behavior in humans. Nutr Neurosci 5: 159-188.
Schneider H, Pitossi F, Balschun D, Wagner A, del Rey A, et al.
1998. A neuromodulatory role of interleukin‐1beta
in the hippocampus. Proc Natl Acad Sci USA 95:
7778-7783.
Sewards TV. 2004. Dual separate pathways for sensory and
hedonic aspects of taste. Brain Res Bull 62: 271-283.
Sewards TV, Sewards MA. 2001. Cortical association areas
in the gustatory system. Neurosci Biobehav Rev 25:
395-407.
Sewards TV, Sewards M. 2002. Separate, parallel sensory and
hedonic pathways in the mammalian somatosensory sys-
tem. Brain Res Bull 58: 243-260.
Siegel S, Krank M, Hinson R. 1987. Anticipation of pharma-
cological and nonpharmacological events: Classical condi-
tioning and addictive behavior. J Drug Issues 17: 83-110.
Smith GR, McDaniel SM. 1983. Psychologically mediated
effect on the delayed hypersensitivity reaction to tuberculin
in humans. Psychosom Med 45: 65-70.
Solvason HB, Ghanta VK, Hiramoto RN. 1988. Conditioned
augmentation of natural killer cell activity. Independence
from nociceptive effects and dependence on interferon‐
beta. J Immunol 140: 661-665.
Solvason HB, Ghanta VK, Hiramoto RN. 1993. The identity
of the unconditioned stimulus to the central nervous sys-
tem is interferon‐beta. J Neuroimmunol 45: 75-81.
Solvason HB, Ghanta VK, Lorden JF, Soong SJ, Hiramoto RN.
1991. A behavioral augmentation of natural immunity:
Odor specificity supports a Pavlovian conditioning model.
Int J Neurosci 61: 277-288.
Solvason HB, Ghanta VK, Soong SJ, Rogers CF, Hsueh CM,
et al. 1992. A simple, single, trial‐learning paradigm for
conditioned increase in natural killer cell activity. Proc
Soc Exp Biol Med 199: 199-203.
Solvason HB, Hiramoto RN, Ghanta VK. 1989. Naltrexone
blocks the expression of the conditioned elevation of natu-
ral killer cell activity in BALB/c mice. Brain Behav Immun
3: 247-262.
Spector NH. 1987. Old and new strategies in the conditioning
of immune responses. Ann N YAcad Sci 496: 522-531.
Spector NH. 1996. Neuroimmunomodulation: A brief
review. Can conditioning of natural killer cell activity
reverse cancer and/or aging? Regul Toxicol Pharmacol 24:
32-38.
Spector NH, Provinciali M, di Stefano G, Muzzioli M,
Bulian D, et al. 1994. Immune enhancement by condition-
ing of senescent mice. Comparison of old and young mice
in learning ability and in ability to increase natural
killer cell activity and other host defense reactions in re-
sponse to a conditioned stimulus. Ann N Y Acad Sci 741:
283-291.
Sredni‐Kenigsbuch D. 2002. TH1/TH2 cytokines in the cen-
tral nervous system. Int J Neurosci 112: 665-703.
Steinman L. 2004. Elaborate interactions between the
immune and nervous systems. Nature Immunol 5: 575-581.
Sternberg EM. 2006. Neural regulation of innate immunity:
A coordinated nonspecific host response to pathogens.
Nature Rev Immunol 6: 318-328.

150 6 Neuro-immune associative learning
Stockhorst U, Spennes‐Saleh S, Korholz D, Gobel U,
Schneider ME, et al. 2000. Anticipatory symptoms and
anticipatory immune responses in pediatric cancer patients
receiving chemotherapy: Features of a classically conditioned
response? Brain Behav Immun 14: 198-218.
Straub RH. 2004. Complexity of the bi‐directional neuroim-
mune junction in the spleen. Trends Pharmacol Sci 25:
640-646.
Straub RH, Schedlowski M. 2002. Immunology and multi-
modal system interactions in health and disease. Trends
Immunol 23: 118-120.
Swanson LW, Petrovich GD. 1998. What is the amygdala?
Trends Neurosci 21: 323-331.
Szczytkowski JL, Lysle DT. 2007. Conditioned effects of
heroin on the expression of inducible nitric oxide synthase
in the rat are susceptible to extinction and latent inhibition.
Psychopharmacology 191: 879-889.
Szelenyi J. 2001. Cytokines and the central nervous system.
Brain Res Bull 54: 329-338.
Tayebati SK, El‐Assouad D, Ricci A, Amenta F. 2002. Immu-
nochemical and immunocytochemical characterization of
cholinergic markers in human peripheral blood lympho-
cytes. J Neuroimmunol 132: 147-155.
Tazi A, Dantzer R, Crestani F, Le Moal M. 1988. Interleukin‐1
induces conditioned taste aversion in rats: A possible
explanation for its pituitary–adrenal stimulating activity.
Brain Res 473: 369-371.
Tonelli LH, Postolache TT, Sternberg EM. 2005. Inflammato-
ry genes and neural activity: Involvement of immune
genes in synaptic function and behavior. Front Biosci 10:
675-680.
Tracey KJ. 2002. The inflammatory reflex. Nature 420:
853-859.
Tracey KJ. 2007. Physiology and immunology of the choliner-
gic antiinflammatory pathway. J Clin Investig 117: 289-296.
Turnbull JW. 1962. Asthma conceived as a learned response.
J Psychosom Res 6: 59-70.
Turnbull AV, Rivier CL. 1999. Regulation of the hypothalamic–
pituitary–adrenal axis by cytokines: Actions andmechanisms
of action. Physiol Rev 79: 1-71.
Vettor R, Fabris R, Pagano C, Federspil G. 2002. Neuroendo-
crine regulation of eating behavior. J Endocrinol Invest 25:
836-854.
Voronov A, Riskin I. 1925. Russkija Medizina, III, 3/2.
Vygodchikov GV. 1955. [Certain controversial problems in
immunity]. Zh Mikrobiol Epidemiol Immunobiol 1: 5-14.
Warthan MD, Freeman JG, Loesser KE, Lewis CW, Hong M,
et al. 2002. Phenylethanolamine N‐methyl transferase
xpression in mouse thymus and spleen. Brain Behav
Immun 16: 493-499.
Wayner EA, Flannery GR, Singer G. 1978. Effects of taste
aversion conditioning on the primary antibody response
to sheep red blood cells and Brucella abortus in the albino
rat. Physiol Behav 21: 995-1000.
Wrona D. 2006. Neural–immune interactions: An integrative
view of the bidirectional relationship between the brain
and immune systems. J Neuroimmunol 172: 38-58.
Wrona D, Trojniar W. 2003. Chronic electrical stimulation of
the lateral hypothalamus increases natural killer cell cyto-
toxicity in rats. J Neuroimmunol 141: 20-29.
Xie Y, Frede S, Harnish MJ, Exton MS, Schedlowski M. 2002.
Beta‐adrenoceptor‐induced inhibition of rat splenocyte
proliferation: Cytokine gene transcription as the target of
action. Immunobiology 206: 345-353.
Yamamoto T, Shimura T, Sako N, Yasoshima Y, Sakai N. 1994.
Neural substrates for conditioned taste aversion in the rat.
Behav Brain Res 65: 123-137.
Ziemssen T, Kern S. 2007. Psychoneuroimmunology – cross‐
talk between the immune and nervous systems. J Neurol
254 (Suppl 2): 8-11.
Related Documents











