Neural transplants in patients with Huntington’s disease undergo disease-like neuronal degeneration F. Cicchetti a,b,1 , S. Saporta c,d , R. A. Hauser e , M. Parent f,g , M. Saint-Pierre a , P. R. Sanberg h,i , X. J. Li j , J. R. Parker k , Y. Chu l , E. J. Mufson l , J. H. Kordower l , and T. B. Freeman h,i,1 a Centre de Recherche du CHUL (CHUQ), 2705, Boulevard Laurier, Que ´ bec, QC, Canada G1V 4G2; b Department of Psychiatry/Neuroscience, Universite ´ Laval, Que ´ bec, QC, Canada G1K 7P4; c Department of Pathology and Cell Biology, h Department of Neurosurgery and Brain Repair, i Center of Excellence for Aging and Brain Repair, d University of South Florida, 2 Tampa General Circle, Tampa, FL 33606-3571; e Departments of Neurology; Pharmacology and Experimental Therapeutics, Parkinson’s Disease and Movement Disorders National Parkinson’s Foundation Center of Excellence, University of South Florida, Tampa, FL 33606; f Department of Pathology and Cell Biology; Groupe de recherche sur le syste ` me nerveux central; g Faculty of Medicine; Universite ´ de Montre ´ al, Montreal, QC, Canada; j Departments of Genetics and Neurology, Emory University, Atlanta, GA 30322; k Department of Pathology, University of Louisville Health Sciences Center, 530 South Jackson Street, Louisville, KY 40202; and l Department of Neurological Sciences and Center for Brain Repair, Rush University Medical Center, 1735 West Harrison Street, Chicago, IL 60612 Edited by Solomon H. Snyder, Johns Hopkins University School of Medicine, Baltimore, MD, and approved June 3, 2009 (received for review April 20, 2009) The clinical evaluation of neural transplantation as a potential treatment for Huntington’s disease (HD) was initiated in an at- tempt to replace lost neurons and improve patient outcomes. Two of 3 patients with HD reported here, who underwent neural transplantation containing striatal anlagen in the striatum a de- cade earlier, have demonstrated marginal and transient clinical benefits. Their brains were evaluated immunohistochemically and with electron microscopy for markers of projection neurons and interneurons, inflammatory cells, abnormal huntingtin protein, and host-derived connectivity. Surviving grafts were identified bilaterally in 2 of the subjects and displayed classic striatal projec- tion neurons and interneurons. Genetic markers of HD were not expressed within the graft. Here we report in patients with HD that (i) graft survival is attenuated long-term; (ii) grafts undergo disease-like neuronal degeneration with a preferential loss of projection neurons in comparison to interneurons; (iii) immuno- logically unrelated cells degenerate more rapidly than the patient’s neurons, particularly the projection neuron subtype; (iv) graft survival is attenuated in the caudate in comparison to the putamen in HD; (v) glutamatergic cortical neurons project to transplanted striatal neurons; and (vi) microglial inflammatory changes in the grafts specifically target the neuronal components of the grafts. These results, when combined, raise uncertainty about this poten- tial therapeutic approach for the treatment of HD. However, these observations provide new opportunities to investigate the under- lying mechanisms involved in HD, as well as to explore additional therapeutic paradigms. excitotoxicity inflammation mutant huntingtin glutamate microglia H untington’s disease (HD) is a progressive, untreatable, and fatal neurodegenerative disorder caused by increased CAG repeats in the huntingtin gene (1). The clinical evaluation of neural transplantation as a potential treatment for HD was initiated in an attempt to replace lost neurons and improve patient outcomes (see refs. 2–4). Preclinical rodent and primate experiments demon- strated the feasibility of using embryonic striatal grafts for the treatment of HD. Fetal striatal grafts survive (5), induce behavioral recovery (6, 7), and establish connectivity with the rodent brain (8, 9). Similar results have been reported with xenografts of porcine and human striatal tissue into rodents (2, 9–12). The human striatum is a small enough target to be approached surgically in HD with a realistic goal of achieving conf luent graft integration into the host brain (2, 13, 14). Grafts can be functionally integrated (15–18), although such connectivity may not be necessary (19, 20). Similar behavioral results were reported in primate models of HD (21, 22). The optimal donor age for human striatal graft survival was established in studies using animal models (4, 12, 23, 24). We chose to use embryonic striatal tissue derived from the far lateral portion of the lateral ventricular eminence to optimize striatal-like (P-zone) volume within grafts to greater than 50% (2, 3, 23, 25, 26), which was postulated to correlate with optimal behavioral improvement in rodent models of HD (4, 27–29). Others have used alternative methods of dissection, transplanting the entire ganglionic eminence (4, 30, 31), which includes at least 13 different cell types or nuclei that are not medium spiny neurons of the striatum (23). Several programs initiated neural transplant trials to evaluate the safety, tolerability, and potential efficacy of human striatal trans- plantation for the treatment of HD (26, 30–32). Preliminary, open-label studies of neural transplants in patients with HD have demonstrated approximately 2 to 4 years of modest clinical benefits, followed by progressive clinical deterioration similar to the natural history of the disease (31, 32), although there is 1 anecdotal case report of more meaningful clinical benefit (33). The safety profile for neural transplantation in HD may differ significantly from neural transplantation in Parkinson’s disease (PD) (32, 34). Neu- ritic outgrowth of transplanted medium spiny-projection neurons to appropriate host target areas was postulated to mediate graft- derived behavioral benefits (35), although this afferent connectivity of the graft to the host has never been observed at human autopsy (25, 36). Additionally, recent reports of PD-like Lewy-body inclu- sions within grafts surviving long-term in patients with PD have raised concerns that genetically and immunologically unrelated grafts are susceptible to the disease processes as well (37–39). The brains of 3 patients with HD who received neuronal trans- plants a decade earlier were evaluated at autopsy. Patients were clinically monitored, as described previously (31, 32). Here we report in patients with HD that (i) graft survival is attenuated long-term; (ii) immunologically unrelated grafts degenerate more rapidly than the patient’s brain, particularly the projection neuron subtype; (iii) cortical neurons project to transplanted striatal neu- rons; and (iv) microglial inflammatory changes in the grafts spe- cifically target the neuronal components of the grafts. We also provide evidence that excitotoxicity and inf lammatory mechanisms play a significant role in transplant degeneration and HD as well. Results Clinical Outcomes. Autopsy results from patient 2 (W.G.) of our series (32) have been reported previously (25). The evaluations Author contributions: F.C., P.R.S., and T.B.F. designed research; F.C., S.S., R.A.H., M.P., M.S.-P., J.R.P., Y.C., J.H.K., and T.B.F. performed research; X.J.L. contributed new reagents/ analytic tools; F.C., S.S., M.P., R.A.H., Y.C., J.H.K., E.J.M., and T.B.F. analyzed data; and F.C. and T.B.F. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Freely available online through the PNAS open access option. 1 To whom correspondence should be addressed. E-mail: [email protected] or [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0904239106/DCSupplemental. www.pnas.orgcgidoi10.1073pnas.0904239106 PNAS Early Edition 1 of 6 NEUROSCIENCE AQ: B AQ: C AQ: D AQ: E Fn1 AQ: F AQ: S AQ: T ZSI balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS Cadmus: 8775 Research Article • Neuroscience

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neural transplants in patients with Huntington’sdisease undergo disease-like neuronal degenerationF. Cicchettia,b,1, S. Saportac,d, R. A. Hausere, M. Parentf,g, M. Saint-Pierrea, P. R. Sanbergh,i, X. J. Lij, J. R. Parkerk, Y. Chul,E. J. Mufsonl, J. H. Kordowerl, and T. B. Freemanh,i,1
aCentre de Recherche du CHUL (CHUQ), 2705, Boulevard Laurier, Quebec, QC, Canada G1V 4G2; bDepartment of Psychiatry/Neuroscience, Universite Laval,Quebec, QC, Canada G1K 7P4; cDepartment of Pathology and Cell Biology, hDepartment of Neurosurgery and Brain Repair, iCenter of Excellence for Agingand Brain Repair, dUniversity of South Florida, 2 Tampa General Circle, Tampa, FL 33606-3571; eDepartments of Neurology; Pharmacology and ExperimentalTherapeutics, Parkinson’s Disease and Movement Disorders National Parkinson’s Foundation Center of Excellence, University of South Florida, Tampa,FL 33606; fDepartment of Pathology and Cell Biology; Groupe de recherche sur le systeme nerveux central; gFaculty of Medicine; Universite de Montreal,Montreal, QC, Canada; jDepartments of Genetics and Neurology, Emory University, Atlanta, GA 30322; kDepartment of Pathology, University of LouisvilleHealth Sciences Center, 530 South Jackson Street, Louisville, KY 40202; and lDepartment of Neurological Sciences and Center for Brain Repair, RushUniversity Medical Center, 1735 West Harrison Street, Chicago, IL 60612
Edited by Solomon H. Snyder, Johns Hopkins University School of Medicine, Baltimore, MD, and approved June 3, 2009 (received for review April 20, 2009)
The clinical evaluation of neural transplantation as a potentialtreatment for Huntington’s disease (HD) was initiated in an at-tempt to replace lost neurons and improve patient outcomes. Twoof 3 patients with HD reported here, who underwent neuraltransplantation containing striatal anlagen in the striatum a de-cade earlier, have demonstrated marginal and transient clinicalbenefits. Their brains were evaluated immunohistochemically andwith electron microscopy for markers of projection neurons andinterneurons, inflammatory cells, abnormal huntingtin protein,and host-derived connectivity. Surviving grafts were identifiedbilaterally in 2 of the subjects and displayed classic striatal projec-tion neurons and interneurons. Genetic markers of HD were notexpressed within the graft. Here we report in patients with HD that(i) graft survival is attenuated long-term; (ii) grafts undergodisease-like neuronal degeneration with a preferential loss ofprojection neurons in comparison to interneurons; (iii) immuno-logically unrelated cells degenerate more rapidly than the patient’sneurons, particularly the projection neuron subtype; (iv) graftsurvival is attenuated in the caudate in comparison to the putamenin HD; (v) glutamatergic cortical neurons project to transplantedstriatal neurons; and (vi) microglial inflammatory changes in thegrafts specifically target the neuronal components of the grafts.These results, when combined, raise uncertainty about this poten-tial therapeutic approach for the treatment of HD. However, theseobservations provide new opportunities to investigate the under-lying mechanisms involved in HD, as well as to explore additionaltherapeutic paradigms.
excitotoxicity � inflammation � mutant huntingtin � glutamate � microglia
Huntington’s disease (HD) is a progressive, untreatable, andfatal neurodegenerative disorder caused by increased CAG
repeats in the huntingtin gene (1). The clinical evaluation of neuraltransplantation as a potential treatment for HD was initiated in anattempt to replace lost neurons and improve patient outcomes (seerefs. 2–4). Preclinical rodent and primate experiments demon-strated the feasibility of using embryonic striatal grafts for thetreatment of HD. Fetal striatal grafts survive (5), induce behavioralrecovery (6, 7), and establish connectivity with the rodent brain (8,9). Similar results have been reported with xenografts of porcineand human striatal tissue into rodents (2, 9–12). The humanstriatum is a small enough target to be approached surgically in HDwith a realistic goal of achieving confluent graft integration into thehost brain (2, 13, 14). Grafts can be functionally integrated (15–18),although such connectivity may not be necessary (19, 20). Similarbehavioral results were reported in primate models of HD (21, 22).
The optimal donor age for human striatal graft survival wasestablished in studies using animal models (4, 12, 23, 24). We choseto use embryonic striatal tissue derived from the far lateral portionof the lateral ventricular eminence to optimize striatal-like (P-zone)
volume within grafts to greater than 50% (2, 3, 23, 25, 26), whichwas postulated to correlate with optimal behavioral improvementin rodent models of HD (4, 27–29). Others have used alternativemethods of dissection, transplanting the entire ganglionic eminence(4, 30, 31), which includes at least 13 different cell types or nucleithat are not medium spiny neurons of the striatum (23).
Several programs initiated neural transplant trials to evaluate thesafety, tolerability, and potential efficacy of human striatal trans-plantation for the treatment of HD (26, 30–32). Preliminary,open-label studies of neural transplants in patients with HD havedemonstrated approximately 2 to 4 years of modest clinical benefits,followed by progressive clinical deterioration similar to the naturalhistory of the disease (31, 32), although there is 1 anecdotal casereport of more meaningful clinical benefit (33). The safety profilefor neural transplantation in HD may differ significantly fromneural transplantation in Parkinson’s disease (PD) (32, 34). Neu-ritic outgrowth of transplanted medium spiny-projection neurons toappropriate host target areas was postulated to mediate graft-derived behavioral benefits (35), although this afferent connectivityof the graft to the host has never been observed at human autopsy(25, 36). Additionally, recent reports of PD-like Lewy-body inclu-sions within grafts surviving long-term in patients with PD haveraised concerns that genetically and immunologically unrelatedgrafts are susceptible to the disease processes as well (37–39).
The brains of 3 patients with HD who received neuronal trans-plants a decade earlier were evaluated at autopsy. Patients wereclinically monitored, as described previously (31, 32). Here wereport in patients with HD that (i) graft survival is attenuatedlong-term; (ii) immunologically unrelated grafts degenerate morerapidly than the patient’s brain, particularly the projection neuronsubtype; (iii) cortical neurons project to transplanted striatal neu-rons; and (iv) microglial inflammatory changes in the grafts spe-cifically target the neuronal components of the grafts. We alsoprovide evidence that excitotoxicity and inflammatory mechanismsplay a significant role in transplant degeneration and HD as well.
ResultsClinical Outcomes. Autopsy results from patient 2 (W.G.) of ourseries (32) have been reported previously (25). The evaluations
Author contributions: F.C., P.R.S., and T.B.F. designed research; F.C., S.S., R.A.H., M.P.,M.S.-P., J.R.P., Y.C., J.H.K., and T.B.F. performed research; X.J.L. contributed new reagents/analytic tools; F.C., S.S., M.P., R.A.H., Y.C., J.H.K., E.J.M., and T.B.F. analyzed data; and F.C.and T.B.F. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.
1To whom correspondence should be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0904239106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0904239106 PNAS Early Edition � 1 of 6
NEU
ROSC
IEN
CE
AQ: B
AQ: C
AQ: D
AQ: E
Fn1
AQ: F
AQ: S
AQ: T
ZSI
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
Cadmus: 8775Research Article • Neuroscience
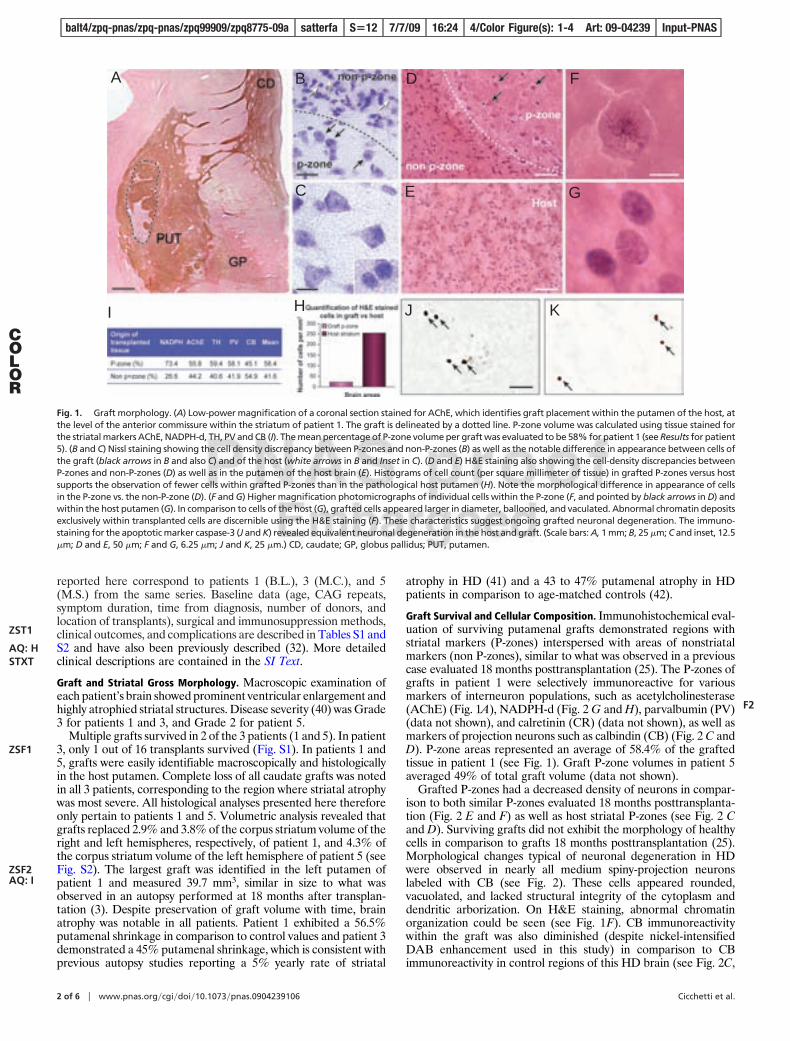
reported here correspond to patients 1 (B.L.), 3 (M.C.), and 5(M.S.) from the same series. Baseline data (age, CAG repeats,symptom duration, time from diagnosis, number of donors, andlocation of transplants), surgical and immunosuppression methods,clinical outcomes, and complications are described in Tables S1 andS2 and have also been previously described (32). More detailedclinical descriptions are contained in the SI Text.
Graft and Striatal Gross Morphology. Macroscopic examination ofeach patient’s brain showed prominent ventricular enlargement andhighly atrophied striatal structures. Disease severity (40) was Grade3 for patients 1 and 3, and Grade 2 for patient 5.
Multiple grafts survived in 2 of the 3 patients (1 and 5). In patient3, only 1 out of 16 transplants survived (Fig. S1). In patients 1 and5, grafts were easily identifiable macroscopically and histologicallyin the host putamen. Complete loss of all caudate grafts was notedin all 3 patients, corresponding to the region where striatal atrophywas most severe. All histological analyses presented here thereforeonly pertain to patients 1 and 5. Volumetric analysis revealed thatgrafts replaced 2.9% and 3.8% of the corpus striatum volume of theright and left hemispheres, respectively, of patient 1, and 4.3% ofthe corpus striatum volume of the left hemisphere of patient 5 (seeFig. S2). The largest graft was identified in the left putamen ofpatient 1 and measured 39.7 mm3, similar in size to what wasobserved in an autopsy performed at 18 months after transplan-tation (3). Despite preservation of graft volume with time, brainatrophy was notable in all patients. Patient 1 exhibited a 56.5%putamenal shrinkage in comparison to control values and patient 3demonstrated a 45% putamenal shrinkage, which is consistent withprevious autopsy studies reporting a 5% yearly rate of striatal
atrophy in HD (41) and a 43 to 47% putamenal atrophy in HDpatients in comparison to age-matched controls (42).
Graft Survival and Cellular Composition. Immunohistochemical eval-uation of surviving putamenal grafts demonstrated regions withstriatal markers (P-zones) interspersed with areas of nonstriatalmarkers (non P-zones), similar to what was observed in a previouscase evaluated 18 months posttransplantation (25). The P-zones ofgrafts in patient 1 were selectively immunoreactive for variousmarkers of interneuron populations, such as acetylcholinesterase(AChE) (Fig. 1A), NADPH-d (Fig. 2 G and H), parvalbumin (PV)(data not shown), and calretinin (CR) (data not shown), as well asmarkers of projection neurons such as calbindin (CB) (Fig. 2 C andD). P-zone areas represented an average of 58.4% of the graftedtissue in patient 1 (see Fig. 1). Graft P-zone volumes in patient 5averaged 49% of total graft volume (data not shown).
Grafted P-zones had a decreased density of neurons in compar-ison to both similar P-zones evaluated 18 months posttransplanta-tion (Fig. 2 E and F) as well as host striatal P-zones (see Fig. 2 Cand D). Surviving grafts did not exhibit the morphology of healthycells in comparison to grafts 18 months posttransplantation (25).Morphological changes typical of neuronal degeneration in HDwere observed in nearly all medium spiny-projection neuronslabeled with CB (see Fig. 2). These cells appeared rounded,vacuolated, and lacked structural integrity of the cytoplasm anddendritic arborization. On H&E staining, abnormal chromatinorganization could be seen (see Fig. 1F). CB immunoreactivitywithin the graft was also diminished (despite nickel-intensifiedDAB enhancement used in this study) in comparison to CBimmunoreactivity in control regions of this HD brain (see Fig. 2C,
A B
C
D
E
F
G
I H J K
Fig. 1. Graft morphology. (A) Low-power magnification of a coronal section stained for AChE, which identifies graft placement within the putamen of the host, atthe level of the anterior commissure within the striatum of patient 1. The graft is delineated by a dotted line. P-zone volume was calculated using tissue stained forthe striatal markers AChE, NADPH-d, TH, PV and CB (I). The mean percentage of P-zone volume per graft was evaluated to be 58% for patient 1 (see Results for patient5). (B and C) Nissl staining showing the cell density discrepancy between P-zones and non-P-zones (B) as well as the notable difference in appearance between cells ofthe graft (black arrows in B and also C) and of the host (white arrows in B and Inset in C). (D and E) H&E staining also showing the cell-density discrepancies betweenP-zones and non-P-zones (D) as well as in the putamen of the host brain (E). Histograms of cell count (per square millimeter of tissue) in grafted P-zones versus hostsupports the observation of fewer cells within grafted P-zones than in the pathological host putamen (H). Note the morphological difference in appearance of cellsin the P-zone vs. the non-P-zone (D). (F and G) Higher magnification photomicrographs of individual cells within the P-zone (F, and pointed by black arrows in D) andwithin the host putamen (G). In comparison to cells of the host (G), grafted cells appeared larger in diameter, ballooned, and vaculated. Abnormal chromatin depositsexclusively within transplanted cells are discernible using the H&E staining (F). These characteristics suggest ongoing grafted neuronal degeneration. The immuno-staining for the apoptotic marker caspase-3 (J and K) revealed equivalent neuronal degeneration in the host and graft. (Scale bars: A, 1 mm; B, 25 �m; C and inset, 12.5�m; D and E, 50 �m; F and G, 6.25 �m; J and K, 25 �m.) CD, caudate; GP, globus pallidus; PUT, putamen.
2 of 6 � www.pnas.org�cgi�doi�10.1073�pnas.0904239106 Cicchetti et al.
ZST1
AQ: HSTXT
ZSF1
ZSF2AQ: I
COLOR
F2
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
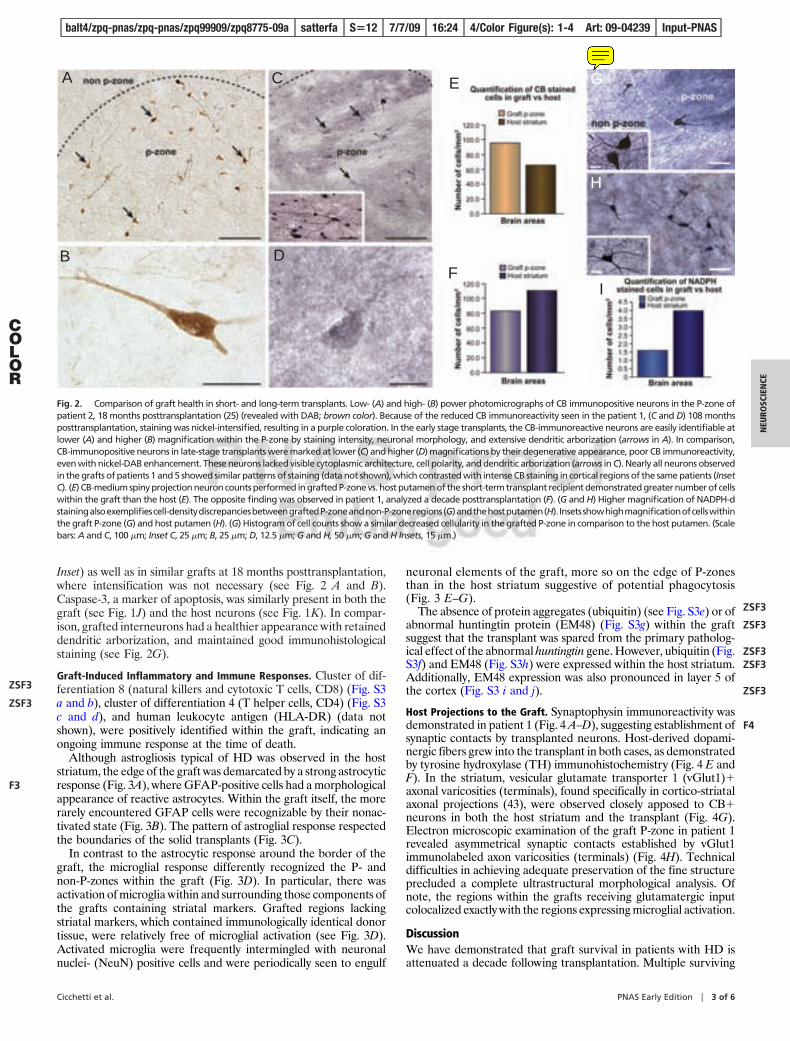
Inset) as well as in similar grafts at 18 months posttransplantation,where intensification was not necessary (see Fig. 2 A and B).Caspase-3, a marker of apoptosis, was similarly present in both thegraft (see Fig. 1J) and the host neurons (see Fig. 1K). In compar-ison, grafted interneurons had a healthier appearance with retaineddendritic arborization, and maintained good immunohistologicalstaining (see Fig. 2G).
Graft-Induced Inflammatory and Immune Responses. Cluster of dif-ferentiation 8 (natural killers and cytotoxic T cells, CD8) (Fig. S3a and b), cluster of differentiation 4 (T helper cells, CD4) (Fig. S3c and d), and human leukocyte antigen (HLA-DR) (data notshown), were positively identified within the graft, indicating anongoing immune response at the time of death.
Although astrogliosis typical of HD was observed in the hoststriatum, the edge of the graft was demarcated by a strong astrocyticresponse (Fig. 3A), where GFAP-positive cells had a morphologicalappearance of reactive astrocytes. Within the graft itself, the morerarely encountered GFAP cells were recognizable by their nonac-tivated state (Fig. 3B). The pattern of astroglial response respectedthe boundaries of the solid transplants (Fig. 3C).
In contrast to the astrocytic response around the border of thegraft, the microglial response differently recognized the P- andnon-P-zones within the graft (Fig. 3D). In particular, there wasactivation of microglia within and surrounding those components ofthe grafts containing striatal markers. Grafted regions lackingstriatal markers, which contained immunologically identical donortissue, were relatively free of microglial activation (see Fig. 3D).Activated microglia were frequently intermingled with neuronalnuclei- (NeuN) positive cells and were periodically seen to engulf
neuronal elements of the graft, more so on the edge of P-zonesthan in the host striatum suggestive of potential phagocytosis(Fig. 3 E–G).
The absence of protein aggregates (ubiquitin) (see Fig. S3e) or ofabnormal huntingtin protein (EM48) (Fig. S3g) within the graftsuggest that the transplant was spared from the primary patholog-ical effect of the abnormal huntingtin gene. However, ubiquitin (Fig.S3f) and EM48 (Fig. S3h) were expressed within the host striatum.Additionally, EM48 expression was also pronounced in layer 5 ofthe cortex (Fig. S3 i and j).
Host Projections to the Graft. Synaptophysin immunoreactivity wasdemonstrated in patient 1 (Fig. 4 A–D), suggesting establishment ofsynaptic contacts by transplanted neurons. Host-derived dopami-nergic fibers grew into the transplant in both cases, as demonstratedby tyrosine hydroxylase (TH) immunohistochemistry (Fig. 4 E andF). In the striatum, vesicular glutamate transporter 1 (vGlut1)�axonal varicosities (terminals), found specifically in cortico-striatalaxonal projections (43), were observed closely apposed to CB�neurons in both the host striatum and the transplant (Fig. 4G).Electron microscopic examination of the graft P-zone in patient 1revealed asymmetrical synaptic contacts established by vGlut1immunolabeled axon varicosities (terminals) (Fig. 4H). Technicaldifficulties in achieving adequate preservation of the fine structureprecluded a complete ultrastructural morphological analysis. Ofnote, the regions within the grafts receiving glutamatergic inputcolocalized exactly with the regions expressing microglial activation.
DiscussionWe have demonstrated that graft survival in patients with HD isattenuated a decade following transplantation. Multiple surviving
A
B
C
D
E
F
G
H
I
Fig. 2. Comparison of graft health in short- and long-term transplants. Low- (A) and high- (B) power photomicrographs of CB immunopositive neurons in the P-zone ofpatient 2, 18 months posttransplantation (25) (revealed with DAB; brown color). Because of the reduced CB immunoreactivity seen in the patient 1, (C and D) 108 monthsposttransplantation, staining was nickel-intensified, resulting in a purple coloration. In the early stage transplants, the CB-immunoreactive neurons are easily identifiable atlower (A) and higher (B) magnification within the P-zone by staining intensity, neuronal morphology, and extensive dendritic arborization (arrows in A). In comparison,CB-immunopositive neurons in late-stage transplants were marked at lower (C) and higher (D) magnifications by their degenerative appearance, poor CB immunoreactivity,even with nickel-DAB enhancement. These neurons lacked visible cytoplasmic architecture, cell polarity, and dendritic arborization (arrows in C). Nearly all neurons observedin the grafts of patients 1 and 5 showed similar patterns of staining (data not shown), which contrasted with intense CB staining in cortical regions of the same patients (InsetC). (E) CB-medium spiny projection neuron counts performed in grafted P-zone vs. host putamen of the short-term transplant recipient demonstrated greater number of cellswithin the graft than the host (E). The opposite finding was observed in patient 1, analyzed a decade posttransplantation (F). (G and H) Higher magnification of NADPH-dstainingalsoexemplifiescell-densitydiscrepanciesbetweengraftedP-zoneandnon-P-zoneregions(G)andthehostputamen(H). Insetsshowhighmagnificationofcellswithinthe graft P-zone (G) and host putamen (H). (G) Histogram of cell counts show a similar decreased cellularity in the grafted P-zone in comparison to the host putamen. (Scalebars: A and C, 100 �m; Inset C, 25 �m; B, 25 �m; D, 12.5 �m; G and H, 50 �m; G and H Insets, 15 �m.)
Cicchetti et al. PNAS Early Edition � 3 of 6
NEU
ROSC
IEN
CE
ZSF3
ZSF3
F3
COLOR
ZSF3
ZSF3
ZSF3ZSF3
ZSF3
F4
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
FC
Note
Is it possible to write the letter "G" in black instead?

grafts were observed in only 2 of our 3 patients. Case 3 (see ref. 32)had only 1 of 16 surviving grafts, which is unusual for any PD or HDtransplant recipient using similar techniques (3, 13, 14, 37, 38).Caudate grafts did not survive in any of these HD patients, similarto what we reported 18 months after transplantation (25). Becausethe caudate displays the greatest neuronal degeneration and astro-gliosis in HD, the current findings suggest that this region is a poortransplant niche in this disease. Long-term putamenal grafts haddecreased cellularity in comparison to those at 18 months post-transplantation. Grafted cells appeared unhealthy, particularlyprojection neurons within the grafts. The expression of intraneu-ronal caspase-3 suggests that striatal cell loss was induced, at leastin part, by apoptosis. These results differ significantly from 18-month and 6-year posttransplantation cases where healthy graftsurvival was reported (25, 36). Neural degeneration at 10 years ofneuronal transplant in HD patients is widespread and potentially
incompatible with any long-term graft-derived benefit. These re-sults differ from similar case reports in patients with PD (37–39).In those cases, 2 to 8% of transplanted cells demonstrated disease-specific markers (Lewy bodies) (37, 39) and 40 to 80% of dopami-nergic neurons and cytoplasmic �-synuclein (38, 39). Despite this,cells were viable, metabolically active (38) and remained differen-tiated with neuritic reinnervation of the host striatum and withlong-term clinical efficacy in individual anecdotal cases (37, 38).
There was diminished cellular viability in all grafted cells at 10years (vs. 18 months) posttransplantation. Importantly, poor cel-lular viability was profound in the grafted striatal projection neu-rons in particular, as opposed to grafted interneurons that stillshowed distinct cellular cytoarchitecture and polarity but with fewerdendritic extensions than was observed at 18 months posttrans-plantation. The preferential degeneration of grafted medium-spinyneurons in comparison to interneurons recapitulates the pattern of
A
B C
D E F
G
Fig. 3. Inflammatory responses to grafts. Both astrocytosis and microgliosis were evaluated and found to be consistent in all 3 hemispheres; representativephotomicrographs of patient 1 are illustrated here. (A) The astrocytic response is significant in the host but minimal within the graft, as investigated by GFAP staining.The edge of the graft is demarcated by a particularly strong astrocytic response, characterized by activated astrocytes. A few nonactivated astrocytes are observablewithin P-zones (B). Abundant GFAP fiber staining is seen within the non-P-zone (C). These nonactivated astrocytes expressed fibers that extended throughout thenon-P-zones but did not significantly invade P-zone. (D) Low-power photomicrograph of double immunostaining for the neuronal nuclear marker NeuN (brown; DABchromogen) and the microglial marker Iba-1 (black; nickel-DAB chromogen) at the interface of a P-zone and non-P-zone and at the interface of the graft and the host.A pronounced microglial response is demonstrated cuffing the edge of the P-zone as well as within the P-zone. Microglial cuffing is seen predominantly at the interfacebetween P-zones and non-P-zones, as opposed to the interface between the non-P-zone portion of the graft and the host. (E) Higher magnification of NeuN/Iba-1staining at the interface graft/host. (F and G) Higher power photomicrographs depicting examples of grafted neurons intermingled with microglial cells. Of note,microglial cells were often observed engulfing neuronal elements, resembling an ongoing phagocytic event. (Scale bars: A, 25 �m; B, 20 �m; C, 100 �m; D, 300 �m;E and F, 25 �m; G, 12.5 �m.)
Fig. 4. Graft connectivity. (A) Low-power photomi-crograph of synaptophysin immunoreactivity insidethe graft (dotted line). The density of synaptophysinreaction product is depicted in (B), in the form of anoptical density plot taken along the line shown in (A).(C and D) High magnification of synaptophysin stain-ing in (C) a P-zone of the graft and (D) in the hoststriatum. (E) Photomicrograph of TH immunostainingof the grafted P-zone (dotted line). (F) Numerous host-derived TH� fibers are observed with seamless pene-tration of the graft border. (G) vGlut1 axon varicosities(black dots; arrows within the neuropile) were foundclosely apposed to cell bodies labeled for CB (brown,arrowheads) in both the host striatum and the trans-plant. (H) Example of a vGlut1-labeled axon varicosity(asterix) within the transplant P-zone exhibiting anasymmetrical synaptic contact (Between small arrows).(Scale bars: C and D, 25 �m; E, 100 �m; F, 25 �m; G, 20�m; H, 500 nm.) av, axon varicosity; sp, spine.
4 of 6 � www.pnas.org�cgi�doi�10.1073�pnas.0904239106 Cicchetti et al.
COLOR
COLOR
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
FC
Note
Abbreviation should be "av" NOT "an".

striatal cell loss seen in HD, suggesting that disease-like cell loss isalso occurring within the genetically unrelated grafts.
Excitotoxicity may play a fundamental role in the degenerativeprocesses seen within these transplants. Glutamatergic axon varicositiesfrom the cerebral cortex were found closely apposed to striatal projec-tion neurons in both the host striatum and the transplants. Within thegraft, they were more abundant in the P-zones that contain the neuronsthat normally receive cortical glutamatergic innervation. Furthermore,microglial infiltration specifically cuffed the P-zones within the grafts,suggesting that microglial activation was responding to specific cuesfound within the striatal P-zone components of the graft. As thenon-P-zone components of the allografts have the same immunologicalsurface antigens as P-zone elements, the selective microglial responseto P-zones in particular is compatible with the hypothesis that thisresponse is mediated by neurotoxicity (44, 45) rather than a subacuteimmune rejection (14). Similar loss of striatal projection neurons is seenfollowing excitotoxicity secondary to glutaric acid in the brains ofpatients with Glutaric Acidemia Type 1, which leads to an HD-likesyndrome in young children (46, 47). In rats, no similar colocalizationof glutamatergic input with P-zone-specific microglial infiltration hasbeen described, suggesting that these pathological findings described inthe study represent a neurotoxic event. However, it is also possible thatthe cortex is making appropriate synapses with the graft.
The lack of EM48 (a marker of the abnormal huntingtin gene) withingrafts, in the presence of robust graft degeneration, indicates that theabnormal HD gene is not required within the striatal neurons to inducedegeneration. Instead, the degeneration of striatal grafts may be aremote consequence of the abnormal gene expressed elsewhere in thebrain. Among other regions, EM48 was located in layer 5 of the cortex,the origin of the vGlut1 innervation of both the striatum and ourtransplants. Normal huntingtin immunolabeling has similarly beenidentified in layer 5 cortico-striatal projection neurons of the normal rat(48). In addition to glutamatergic input to the striatum, cortico-striatalprojections also provide trophic support to striatal neurons (49). Thesedata suggest that grafts degenerate secondary to either excitotoxicglutamatergic inputs from the cortex combined with microglial activa-tion against striatal components of the grafts and to the loss of adequatehost trophic support. The pattern of graft degeneration that so closelyrecapitulates the pattern of striatal degeneration in HD suggests thatfindings in transplantationprovideapossiblemodel for themechanismsof degeneration of the striatum of patients with HD as well.
In the present study, we also observed that the grafts werestrikingly more affected by pathological processes than the hoststriatum, despite the fact that the grafts were young, immunolog-ically unrelated to the patients with HD, and exposed to the diseaseprocess for only a decade. Cortical glutamatergic projections toboth the graft and the host striatum were demonstrated. At theelectron microscopy level, synaptic contacts between glutamatergicaxon varicosities and unlabeled dendrites within the graft were alsoobserved. It is possible that the immune responses to solid grafts inthe absence of immunosuppression (14, 50), combined with therelease of glutamate directly from microglia or glutamatergiccortico-striatal afferents, accelerated the degeneration process ingrafted vs. host striatal neurons, as suggested in animal studies (51).Graft degenerative changes seen here are incompatible with clinicalbenefits after 10 years, vs. PD, where only 2 to 8% of grafted cellshave �-synuclein inclusions (37–39), and clinical benefits lasted for11 to 12 years in those patients (37, 38).
It has been postulated that if striatal transplants are to beeffective for the treatment of HD, they need to prevent localneuronal degeneration within the striatum (26) as well as wide-spread degeneration of cortical afferents projecting to the grafts(2–4, 35). However, meaningful neuritic outgrowth was not dem-onstrated in either the present study or previous studies of graftssurviving 18 months (25) or 6 years posttransplantation (36). Thevolume of striatal loss in our transplant recipients is similar to whathas been described in other patients with HD (42). Therefore, thepresent study failed to demonstrate that embryonic striatal grafts
slowed degeneration of the surrounding striatum via either neuriticoutgrowth or graft-derived neurotrophic expression (35). However,the present study demonstrated that glutamatergic cortico-striatalprojections terminate onto transplanted striatal components of thegrafts. The functional significance of this interaction remains un-known. Rather than the grafts positively influencing the cortex, thepathology within the cortex appeared to induce neuronal degen-eration within the grafts.
Risks associated with neuronal transplantation in this study arenot trivial. Two subjects developed a total of 3 subdural hematomasthat required surgical evacuation, out of a total of 14 hemispherictransplants (32). This was likely because of cerebrospinal fluid lossin the setting of brain atrophy leading to the formation of hygromas,which frequently convert to hematomas. This also led to a targetingerror in the last needle tract in 1 patient. This safety profile differsdramatically from that observed in transplant recipients with PD,where the same surgeon (T.B.F.) had no surgically relevant sub-dural hematomas in 66 consecutive transplant procedures (32).Based on these results, it is our impression that any future surgicaltherapy for HD should intervene in earlier stage patients with lessbrain atrophy, and minimize cerebrospinal fluid loss to diminish therisks of subdural hematomas and targeting errors.
Clinical efficacy has been postulated to require that greater than50% of grafted volume is composed of P-zones (4, 27, 29) thatcontain striatal projection neurons. This anatomical threshold wasachieved in both the present and our previous study (3). However,clinical efficacy in our patients was mild in magnitude, short-lived,subject to the limitations of small open-label trials (placebo effect,as well as potential investigator and subject bias), and did notprevent the unremitting and fatal progression of HD. Similarclinical outcomes have been reported by others (31), despite the factthat their grafts were derived from a much more nonspecificdissection of the striata in their first autopsy report (53). Thesesimilar clinical results, using significantly different dissection meth-ods with a ‘‘subthreshold’’ quantity of P-zones within grafts, sug-gests that clinical benefits, if any, are the result of nonspecific effectsof these grafts (19, 20). The combination of an unfavorable risk-benefit profile, short-term and mild benefits at best, and significantdisease-like graft degeneration makes future trials of fetal-celltransplantation using these techniques potentially unwarranted.Therapeutic strategies (with or without neural grafts) aimed ataltering inflammatory or immune responses, host-derived neuro-toxicity, and neurotrophic support in the brain may prove to bemore promising for the future treatment of HD.
MethodsDonor Tissue Preparation and Transplantation. Methods for tissue preparation,neuronal transplantation, immunosuppression, location of the transplants, num-ber of donors per patient, and clinical genetic as well as radiologic evaluationhave been described previously (3, 23, 25, 32). Solid-tissue transplants measuring0.5 to 1 mm3 were derived from the far lateral portion of the lateral ventriculareminence to optimize the percentage of tissue of striatal origin (23). All patientsreceived transplants into both the caudate and putamen derived from 5 to 8striatal primordia per site. The 3 patients in this series represent patients 1, 3, and5 from our clinical report (32).
Postmortem Tissue Preparation and Histological Evaluation. Postmortem inter-vals were 5, 4, and 5 h for patients 1, 3, and 5 respectively. The brains werebisected, cut serially into1-cmslabs inacoronalplaneand immersed inZamboni’sfixative (25) for 8 days at 4 °C. The brain of patient 5 was collected in the samefashion as patients 1 and 3, but only one-half of the brain was immersed inZamboni’s solution. The other half was immersion-fixed in 4% glutaraldehydeprepared in cacodylate buffer for electron microscopic examination. Followingthe 8-day fixation period, brain slabs were then placed in a 20% sucrose in 0.1 MPB cryoprotectant solution until they sank and were cut frozen into 40-�m thicksections.
Series of adjacent sections were processed for histochemical and immunohis-tochemical (seeSupplementalMethods inSIText forprotocoldetails)analyses forthe visualization of H&E (brain cytoarchitecture) and various striatal subtypes ofinterneurons; NADPH-d (marker for nitric oxide containing striatal interneurons),
Cicchetti et al. PNAS Early Edition � 5 of 6
NEU
ROSC
IEN
CE
AQ: K
AQ: L
AQ: M
STXT
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
FC
Barrer

AChE (enzyme catalyzing hydrolysis of the neurotransmitter acetyl choline incholinergic neurons), PV (Sigma; 1:1,000) or CR (Swant; 1:2,500). The immune/inflammatory response was investigated using GFAP (astrocytes; Dako Canada;1:2,500), CD4 (T helper cells; Serotec; 1:250), CD8 (natural killers and cytotoxic Tcells; Serotec; 1:200), HLA-DR (surface antigen of class II major histocompatibilitycomplex; Serotec; 1:200), while other sections were processed for double immu-nohistochemistry to visualize the microglial response (Iba-1; Wako Chemicals;1:1,000) to grafted neuronal elements (NeuN; Chemicon; 1:2500). Genetic mark-ers of huntingtin were evaluated using EM48 (abnormal huntingtin protein;provided by X.J. Li, Emory University; 1:2,000) and ubiquitin (Calbiochem; 1:250).Finally, host-derived connectivity was assessed by TH (Pel-Freez; 1:1,000), synap-tophysin (Calbiochem; 1:500), double immunostaining for CB (striatal projectionneurons; Sigma; 1:2,500), and vGlut1 (cortico-striatal projections; Sigma; 1:500),and electron microscopy (see Supplemental Methods in SI Text for protocoldetails). Sections intended for electron microscopy were prepared as above forvGlut1 immunohistochemistry, but for single immunostaining were preparedonly using a slightly modified version of previously published protocol (52).
Three-Dimensional Reconstruction, Striatal P-Zone Evaluation and Cell Counts.Three-dimensional reconstruction was performed using the Serial Section Recon-
struction method provided by the Neurolucida software, version 6.0 (Micro-brightfield) on Nissl-stained sections. Volumetric evaluation of graft size (Cava-lieri method) as well as P-zone/non-P-zone areas for patients 1 and 5 wereexplicitly performed using the right hemisphere of both patients 1 and 5. Bothgrafts and P-zones were delineated using Nissl staining and the Tracing Contoursoption in the Stereo Investigator software, version 5.0 (Microbrightfield). Areasfor either P-zones or non-P-zones were calculated using Contour Measurementsoption. Cell counts were performed using the Optical Fractionator probe (StereoInvestigator software) based on 3 different immunostaining markers for NADPH(interneurons), H&E (general cell marker), and CB (projection neurons). Moredetails on the assessment of graft volume, estimation of striatal P-zones, and3-dimensional reconstruction are provided in SI Text.
ACKNOWLEDGMENTS. The authors thank Dr. Peter Gould of the serviced’anatomopathologie et cytologie, Hopital de l’Enfant-Jesus, Quebec, Canadafor pathological diagnosis, and clinical coordinators Deborah Scott and HollyDelgado for their commitment and expertise. This work was supported in part bythe Huntington Society Canada, the Fondation Canadienne pour l’Innovation(F.C.), and Tampa General Healthcare, the International Organization of GlutaricAcidemia (T.B.F.).
1. Huntington’s Disease Collaborative Research Group (1993) A novel gene containing atrinucleotide repeat that is expanded and unstable on Huntington’s disease chromo-somes. Cell 72:971–983.
2. Freeman TB, Hauser RA, Sanberg PR, Saporta S (2000) Neural transplantation for thetreatment of Huntington’s disease. Prog Brain Res 127:405–411.
3. Freeman TB, et al. (2000) Transplantation of human fetal striatal tissue in Huntington’sdisease: rationale for clinical studies. Neural transplantation in neurodegenerativedisease: current status and new directions. Novartis Found Symp 231:129–144.
4. Peschanski M, Cesaro P, Hantraye P (1995) Rationale for intrastriatal grafting of striatalneuroblasts in patients with Huntington’s disease. Neuroscience 68:273–285.
5. Schmidt RH, Bjorklund A, Stenevi U (1981) Intracerebral grafting of dissociated CNStissue suspensions: a new approach for neuronal transplantation to deep brain sites.Brain Res 218:347–356.
6. Deckel AW, Robinson RG, Coyle JT, Sanberg PR (1983) Reversal of long-term locomotorabnormalities in the kainic acid model of Huntington’s disease by day 18 fetal striatalimplants. Eur J Pharmacol 93:287–288.
7. Isacson O, Brundin P, Kelly PA, Gage FH, Bjorklund A (1984) Functional neuronalreplacement by grafted striatal neurones in the ibotenic acid-lesioned rat striatum.Nature 311:458–460.
8. Pritzel M, Isacson O, Brundin P, Wiklund L, Bjorklund A (1986) Afferent and efferentconnections of striatal grafts implanted into the ibotenic acid lesioned neostriatum inadult rats. Exp Brain Res 65:112–126.
9. Wictorin K (1992) Anatomy and connectivity of intrastriatal striatal transplants. ProgNeurobiol 38:611–639.
10. Deacon TW, Pakzaban P, Isacson O (1994) The lateral ganglionic eminence is the originof cells committed to striatal phenotypes: neural transplantation and developmentalevidence. Brain Res 668:211–219.
11. Isacson O, et al. (1995) Transplanted xenogeneic neural cells in neurodegenerativedisease models exhibit remarkable axonal target specificity and distinct growth pat-terns of glial and axonal fibres. Nat Med 1:1189–1194.
12. Wictorin K, Brundin P, Gustavii B, Lindvall O, Bjorklund A (1990) Reformation of longaxon pathways in adult rat central nervous system by human forebrain neuroblasts.Nature 347:556–558.
13. Kordower JH, et al. (1995) Neuropathological evidence of graft survival and striatalreinnervation after the transplantation of fetal mesencephalic tissue in a patient withParkinson’s disease. N Engl J Med 332:1118–1124.
14. Kordower JH, et al. (1997) Fetal grafting for Parkinson’s disease: expression of immunemarkers in two patients with functional fetal nigral implants. Cell Transplant 6:213–219.
15. Campbell K, Wictorin K, Bjorklund A (1995) Neurotransmitter-related gene expressionin intrastriatal striatal transplants–I. Phenotypical characterization of striatal andnon-striatal graft regions. Neuroscience 64:17–33.
16. Campbell K, Wictorin K, Bjorklund A (1995) Neurotransmitter-related gene expressionin intrastriatal striatal transplants–II. Characterization of efferent projecting graftneurons. Neuroscience 64:35–47.
17. Pakzaban P, Deacon TW, Burns LH, Isacson O (1993) Increased proportion of acetyl-cholinesterase-rich zones and improved morphological integration in host striatum offetal grafts derived from the lateral but not the medial ganglionic eminence. Exp BrainRes 97:13–22.
18. Sirinathsinghji DJ, Heavens RP, Torres EM, Dunnett SB (1993) Cholecystokinin-dependentregulation of host dopamine inputs to striatal grafts. Neuroscience 53:651–663.
19. Norman AB, Lehman MN, Sanberg PR (1989) Functional effects of fetal striatal trans-plants. Brain Res Bull 22:163–172.
20. Sanberg PR, Koutouzis TK, Freeman TB, Cahill DW, Norman AB (1993) Behavioral effectsof fetal neural transplants: relevance to Huntington’s disease. Brain Res Bull 32:493–496.
21. Hantraye P, Riche D, Maziere M, Isacson O (1992) Intrastriatal transplantation ofcross-species fetal striatal cells reduces abnormal movements in a primate model ofHuntington disease. Proc Natl Acad Sci USA 89:4187–4191.
22. Isacson O, Riche D, Hantraye P, Sofroniew MV, Maziere M (1989) A primate model ofHuntington’s disease: cross-species implantation of striatal precursor cells to theexcitotoxically lesioned baboon caudate-putamen. Exp Brain Res 75:213–220.
23. Freeman TB, Sanberg PR, Isacson O (1995) Development of the human striatum:implications for fetal striatal transplantation in the treatment of Huntington’s disease.Cell Transplant 4:539–545.
24. Naimi S, Jeny R, Hantraye P, Peschanski M, Riche D (1996) Ontogeny of human striatalDARPP-32 neurons in fetuses and following xenografting to the adult rat brain. ExpNeurol 137:15–25.
25. Freeman TB, et al. (2000) Transplanted fetal striatum in Huntington’s disease: pheno-typic development and lack of pathology. Proc Natl Acad Sci USA 97:13877–13882.
26. Freeman TB, et al. (1998) Human Fetal Tissue Transplantation for the Treatment ofMovement Disorders. (AANS Publications Committee, New York) pp 177–192.
27. Brundin P, Fricker RA, Nakao N (1996) Paucity of P-zones in striatal grafts prohibitcommencement of clinical trials in Huntington’s disease. Neuroscience 71:895–897.
28. Nakao N, Grasbon-Frodl EM, Widner H, Brundin P (1996) DARPP-32-rich zones in graftsof lateral ganglionic eminence govern the extent of functional recovery in skilled pawreaching in an animal model of Huntington’s disease. Neuroscience 74:959–970.
29. Peschanski M, Cesaro P, Hantraye P (1996) What is needed versus what would beinteresting to know before undertaking neural transplantation in patients with Hun-tington’s disease. Neuroscience 71:899–900.
30. Bachoud-Levi A, et al. (2000) Safety and tolerability assessment of intrastriatal neuralallografts in five patients with Huntington’s disease. Exp Neurol 161:194–202.
31. Bachoud-Levi AC, et al. (2006) Effect of fetal neural transplants in patients with Hunting-ton’s disease 6 years after surgery: a long-term follow-up study. Lancet Neurol 5:303–309.
32. Hauser RA, Sandberg PR, Freeman TB, Stoessl AJ (2002) Bilateral human fetal striataltransplantation in Huntington’s disease. Neurology 58:1704, author reply 1704.
33. Reuter I, et al. (2008) Long-term clinical and positron emission tomography outcomeof fetal striatal transplantation in Huntington’s disease. J Neurol Neurosurg Psychiatry79:948–951.
34. Kopyov OV, Jacques S, Lieberman A, Duma CM, Eagle KS (1998) Safety of intrastriatalneurotransplantation for Huntington’s disease patients. Exp Neurol 149:97–108.
35. Watts C, Dunnett SB (2000) Towards a protocol for the preparation and delivery ofstriatal tissue for clinical trials of transplantation in Huntington’s disease. Cell Trans-plant 9:223–234.
36. Keene CD, et al. (2007) Neural transplantation in Huntington disease: long-term graftsin two patients. Neurology 68:2093–2098.
37. Kordower JH, Chu Y, Hauser RA, Freeman TB, Olanow CW (2008) Lewy body-likepathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat Med14:504–506.
38. Kordower JH, Olanow CW, Hauser R, Chu Y, Freeman TB (2008) Transplanted dopa-minergic neurons develop PD pathologic changes: A second case report. MovementDisorders 23:2303–2306.
39. Li JY, et al. (2008) Lewy bodies in grafted neurons in subjects with Parkinson’s diseasesuggest host-to-graft disease propagation. Nat Med 14:501–503.
40. Vonsattel JP, et al. (1985) Neuropathological classification of Huntington’s disease.J Neuropathol Exp Neurol 44:559–577.
41. Henley SM, et al. (2006) Increased rate of whole-brain atrophy over 6 months in earlyHuntington disease. Neurology 67:694–696.
42. Mann DM, Oliver R, Snowden JS (1993) The topographic distribution of brain atrophy inHuntington’s disease and progressive supranuclear palsy. Acta Neuropathol 85:553–559.
43. Raju DV, Smith Y (2005) Differential Localization of Vesicular Glutamate Transporters 1and 2 in the Rat Striatum. (Springer Science and Business Media, New York) pp. 601–610.
44. Biber K, Neumann H, Inoue K, Boddeke HW (2007) Neuronal ‘‘On’’ and ‘‘Off’’ signalscontrol microglia. Trends Neurosci 30:596–602.
45. Conforti L, Adalbert R, Coleman MP (2007) Neuronal death: where does the end begin?Trends Neurosci 30:159–166.
46. Flott-Rahmel B, et al. (1997) Nerve cell lesions caused by 3-hydroxyglutaric acid: a possiblemechanism for neurodegeneration in glutaric acidaemia I. J Inherit Metab Dis 20:387–390.
47. Funk CB, et al. (2005) Neuropathological, biochemical and molecular findings in aglutaric acidemia type 1 cohort. Brain 128:711–722.
48. Fusco FR, et al. (1999) Cellular localization of huntingtin in striatal and cortical neuronsin rats: lack of correlation with neuronal vulnerability in Huntington’s disease. J Neu-rosci 19:1189–1202.
49. Canals JM, et al. (2001) Expression of brain-derived neurotrophic factor in corticalneurons is regulated by striatal target area. J Neurosci 21:117–124.
50. Redmond DJ, Vinuela A, Kordower J, Isacson O (2008) Influence of cell preparation andtarget location on the behavioral recovery after striatal transplantation of fetal dopami-nergic neurons in a primate model of Parkinson’s disease. Neurobiol Dis 29:103–116.
51. Stack EC, et al. (2007) Neuroprotective effects of synaptic modulation in Huntington’sdisease R6/2 mice. J Neurosci 27:12908–12915.
52. Parent M, Descarries L (2008) Acetylcholine innervation of the adult rat thalamus:distribution and ultrastructural features in dorsolateral geniculate, parafascicular, andreticular thalamic nuclei. J Comp Neurol 511:678–691.
53. Capetian P, et al. (2009) Histological findings on fetal striatal grafts in a Huntington’sdisease patient early after transplantation. Neuroscience 160:661–675.
6 of 6 � www.pnas.org�cgi�doi�10.1073�pnas.0904239106 Cicchetti et al.
STXT
AQ: N
STXT
AQ: O
AQ: P
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS

Supporting InformationCicchetti et al. 10.1073/pnas.0904239106SI TextSupplemental Clinical Outcome. Significant cortical atrophy wasnoted on the preoperative MRI scan of patient 1 (B.L.). At theend of surgery, brain shrinkage because of cerebrospinal f luidloss was noted. The surface of the brain was �8 mm from theinner cortex of the skull. No subdural hematoma was noted onthe immediate postoperative MRI scan. However, corticalshrinkage led to targeting error in the last needle tract (in theright caudate) (see Fig. S2). A 2-cm thick asymptomatic subacutesubdural hematoma was noted on the localizing MRI scan donebefore the second operation and was drained uneventfully aftercompletion of the contralateral transplant procedure. Follow-upMRI scans demonstrated complete resolution of the hematoma.The Unified Huntington’s Disease Rating Scale (UHDRS)scores improved from 42 at baseline to 30 to 32 at 3 and 9 monthspostoperatively, but returned to baseline (1) by 18 months aftersurgery. Preoperative gait instability with falling improved for2.5 years, whereupon a wheelchair was required and swallowingdifficulties began to emerge. UHDRS scores stabilized until 3.5years after surgery. Scores then progressively deteriorated to 52,65, and 78 at 4, 6, and 7 years postoperatively. Cognitive functionalso progressively declined with Mini Mental State Evaluation(MMSE) scores worsening from 30 at both baseline and at 21months postsurgery to 14 at 7 years postsurgery. She died at 67years of age of an aspiration pneumonia complicated by amyocardial infarction 9 years postoperatively.
Patient 3 (M.C.) had an asymptomatic 1-cm cortical hemor-rhage after her first operation, as well as a thin (3 mm) subduralhematoma without mass effect. She had postoperative confusionthat resolved in 2 weeks. A 4-mm subdural hematoma was notedafter the second operation, and the bilateral thin subduralhematomas both resolved on subsequent postoperative imagingwithout complications. Her preoperative UHDRS score of 33remained unchanged until 10 to 12 months postsurgery, whenscores improved to 28 and 21, respectively. Her preoperativebalance difficulties and falling (once per month) improvedpostoperatively, and she did not fall for at least 2 years aftersurgery. Symptoms then worsened to a UHDRS score of 37 at20 months after surgery and plateaued there until 3 yearspostoperatively. Cognitive function declined mildly with anMMSE score of 26 at baseline and 24 at 30 months postsurgery.Some depressive symptoms emerged at 2 years after surgery. Thepatient died at the age of 75 of a cardiorespiratory arrest ascomplication of end-stage HD, 10.5 years after transplantation.
Patient 5 (M.S.) was noted to have significant cortical andstriatal atrophy on the preoperative MRI scan. She had 7-mmthick bilateral hygromas postoperatively, which increased to 1 cmafter the second operation. She tripped at home, hitting her head2 weeks after her second operation, with a 1- to 2-min loss ofconsciousness. A CT evaluation demonstrated conversion of herhygromas into subdural hematomas that required surgical drain-age bilaterally. The hematomas were 2.5-cm thick on the left sideand 1.0-cm thick on the right side at the time of surgery. The CTscan 1 month later demonstrated complete resolution of thesubdural hematomas. She never improved back to baseline. HerUHDRS score was 27 before surgery and 31 after drainage of hersubdural hematomas. She deteriorated to a score of 39 by 9months postoperation that remained stable until 2.5 years aftersurgery, when she began a more rapid deterioration to a scoreof 53. By 5 years postoperatively, her UHDRS score was 64.MMSE scores declined from 27 at baseline to 16 at 5 years
postsurgery. She died at the age of 68 of a cardiorespiratoryarrest, secondary to end-stage HD, 9.5 years after surgery.
SI MethodsHistochemistry. Sections were washed 3 times in PBS 0.1M beforehistochemical staining for NADPH-d (marker for nitric oxidecontaining striatal interneurons), AChE (enzyme catalyzinghydrolysis of the neurotransmitter acetyl choline in cholinergicneurons), and H&E (brain cytoarchitecture). For NADPH-dstaining, sections were washed in PBS 0.1M pH 7.4, preincubatedin 0.25% Triton X-100 in PBS for 10 min and transferred in afresh solution of 0.25% Triton X-100, 0.05% of the reduced formof nicotinamide adenine dinucleotide phosphate (b-NADPH;Calbiochem), 0.02% nitro blue tetrazolium (Sigma) in PBS for5 min at room temperature and then at 37 °C for 8 h. Sectionswere rinsed in PBS, mounted on gelatin-coated slides, and keptat 37 °C overnight. They were subsequently dehydrated in as-cending grades of ethanol and coverslipped with DPX mountingmedia (Electron Microscopy Science).
For AChE staining, sections were washed 3 times in distilledwater and incubated in 0.2% acetylthiocholine iodide (Sigma) ina stock solution (copper sulfate, glycine, magnesium chloride,maleic acid in 4% NaOH, 40% sodium sulfate). They were thenrinsed 3 times in 40% sodium sulfate, and incubated in 10%ammonium sulfide for 2 min. After washes in distilled water,sections were counterstained with Kernechtrot red (J.T. Baker)for 1 min and rinsed in tap water. Slices were mounted ongelatin-coated slides, air-dried overnight, dehydrated in ascend-ing grades of ethanol, and cover-slipped with DPX mountingmedia.
For H&E staining, sections were first mounted on gelatin-coated slides, air-dried overnight, and hydrated in 50% ethanol.They were then stained with hematoxylin (Fisher Scientific) for40 sec, washed in running water for 5 min, and placed indifferentiator solution (0.5% pure glacial acetic acid in 95%ethanol) for 1 min. Sections were washed using distilled waterand counterstained with Eosin Y (Sigma) for 40 sec, dehydratedin ethanol and xylene baths (3 � 90% ethanol, 2 � 100% ethanol,2� xylene) and cover-slipped with DPX mounting media.
Immunohistochemistry. Before immunostaining procedures, free-floating sections were washed 3 times in PBS 0.1M pH 7.4 andplaced in 3% peroxide in 0.1M PB for 30 min at room temper-ature. For single immunostaining, sections were subsequentlywashed in PBS and then preincubated for 30 min at roomtemperature in a blocking solution containing, 0.1% TritonX-100 (Sigma) and 5% Normal Goat Serum (NGS, Wisent Inc.)diluted in PBS. Sections were incubated for 24 h at 4 °C in thesame solution to which either anti-GFAP (Dako Canada,;1:2,500) or anti-TH (Pel-Freez; 1:1,000), or for 48 h at 4 °C withanti-PV (Sigma; 1:1,000). Sections were then washed in PBS andincubated for 1 h at room temperature in the blocking solutionto which biotinylated goat anti-rabbit (for GFAP and TH) orbiotinylated goat anti-mouse (for PV) (Vector Laboratories;1:1,500) was added. Following 3 washes in PBS 0.1M, sectionswere placed in a solution of avidin-biotin peroxidase complex(ABC Elite kit, Vector Laboratories) for 1 h at room temper-ature. Antibodies were revealed by placing the sections in Trisbuffer solution containing 0.05% 3.3�-diaminobenzidine tetra-hydrochloride (DAB, Sigma) and 0.1% of 30% hydrogen per-oxidase at room temperature. Reaction was stopped by washingin 0.05M Tris buffer and subsequent PBS washes. Slices were
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 1 of 7
STXT
ZSF2
AQ: U
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
Cadmus: 8775Research Article • Neuroscience

mounted on gelatin-coated slides, air-dried overnight, dehy-drated in ascending grades of ethanol, xylene, and cover-slippedwith DPX mounting media.
Other sections were immunohistochemically processed withnickel-intensification of DAB to enhance the chromogen signal.These sections were treated in a similar manner as describedabove except that the main buffer solution was composed of PBS0.2M pH 7.4 and 1% BSA (Sigma) was added as a blocking agentin the primary and secondary antibodies, as well as in the ABCsolutions. Sections were incubated 48 h at 4 °C with eitheranti-CB (Sigma; 1:2,500), CD4 (Serotec; 1:250), CD8 (Serotec;1:200), HLA-DR (Serotec; 1:200), ubiquitin (Calbiochem;1:250), synaptophysin (Calbiochem; 1:500), EM48 (provided byX.J. Li, Emory University; 1:2,000) or anti-CR (Swant; 1:2,500)and 1 h with biotinylated goat anti-mouse (for CB) or goatanti-rabbit (for CR) (Vector Laboratories; 1:1,500). After incu-bation with ABC, sections were washed twice in acetate imida-zole 0.2M pH 7.2 followed by Ni-DAB solution (dH2O, sodiumacetate 1M pH 7.2, imidazole 0.2M pH 9.2, nickel-sulfate 6.H2O,DAB, H2O2 30%). Immunohistochemical reaction was termi-nated by washes in acetate imidazole 0.2M (pH 7.2) followed by0.2M PBS rinses. Slices were mounted on gelatin-coated slides,air-dried overnight, dehydrated in ascending grades of ethanol,and cover-slipped with DPX mounting media. In these experi-ments, immunohistochemical controls included omission of theprimary or the secondary antibody, which completely abolishedthe immunostaining.
Other sections were processed for double immunohistochem-istry to visualize neuronal nuclei (NeuN) and microglia (Iba-1)or calcium binding protein (CB) and vGlut1. After overnightincubation at 4 °C with an antibody against Iba-1 (Wako Chem-icals; 1:1000) or 48 h at 4 °C with an antibody against vGlut1(Sigma; 1:500), the sections were extensively washed in PBS andincubated for 1 h at room temperature in a PBS solutioncontaining biotinylated goat anti-rabbit IgG (Vector Labs; di-lution 1:1,500), Triton X-100 (0.1%), BSA (1%), and NGS (5%).After further washing in PBS, the sections were placed in asolution containing ABC (Elite kit; Vector Labs) for 1 h at roomtemperature. The bound peroxidase was revealed with nickel-intensified DAB as the chromogen. After immunostaining forIba-1 or vGlut1, the sections were reincubated overnight at 4 °Cwith a NeuN antibody (Chemicon; 1:1,000) or 48h at 4 °C withan antibody against CB (Sigma; 1:2,500). The incubation pro-cedures were the same as above, except that the incubation timein secondary antibody, goat anti-mouse (Vector Labs; 1:1,500),was 2 h and sections were revealed using DAB (Sigma) and0.01% hydrogen peroxide in 0.05 M Tris-imidazole (pH 7.2) atroom temperature. The reaction was stopped after 10 to 15 minby extensive washing in PBS. Controls included omission of
either one of the primary antibodies to exclude cross-reactivityof the secondary antibodies.
Sections intended for electron microscopy were prepared asabove for vGlut1 immunohistochemistry, but for single immu-nostaining only, without Triton X-100 in all solutions and usingDAB as the chromogen. After revelation, these sections wereosmicated, dehydrated in ethanol and propylene oxide, andflat-embedded in Durcapan (Fluka). Rectangular pieces withinthe grafted P-zone were removed from the flat-embeddedvGlut1-immunostained sections, glued to the tip of resin blocks,and sectioned ultrathin (80 nm) with a Reichert Jung ultrami-crotome. These sections were collected on bare 150 mesh coppergrids, stained with lead citrate, and examined with a PhillipsCM100 electron microscope (60 kV, Philips Electronique).
Assessment of Graft Volume and Location. Nissl staining, as de-scribed previously (2), was used to perform the 3-dimensionalreconstruction of transplantation sites using Neurolucida mod-eling software (Microbrightfield) attached to a E800 Nikonmicroscope (Nikon Instruments) (see below). Two distinct setsof calculations were performed in relation to graft volume. Thefirst calculations, which are found in Table 1 of Fig. 1, depict thevolumes of the entire corpus striatum (putamen, caudate and theglobus pallidus). Representative serial sections (1 in 10) werecompared to equivalent sections from the corpus striatum of acontrol brain. The second set of volumetric measurements wasused to evaluate the degree of brain shrinkage of patients 1 and5. For this measurement, serial sections of the putamen of thetransplant recipient were compared to equivalent serial sectionsof the putamen in an age-matched control brain.
Estimation of the Striatal Zones (P-Zones) Within the Grafts. Volu-metric evaluation of graft size (Cavalieri method) as well asP-zone and non-P-zone areas for patients 1 and 5 were explicitlyperformed using the right hemisphere of both patients 1 and 5.Both grafts and P-zones were delineated using Nissl staining andthe Tracing Contours option in the Stereo Investigator software,version 5.0 (Microbrightfield). Areas for either P-zones ornon-P-zones were calculated using Contour Measurements op-tion.
Three-dimensional reconstruction. Three-dimensional reconstruc-tion was performed using the Serial Section Reconstructionmethod provided by the Neurolucida software, version 6.0(Microbrightfield). Briefly, the caudate, putamen, and trans-plant sites were traced using the Tracing Contours function foreach section. Subsequently, each tracing was imported into theNeuroExplorer software, where the drawings were aligned tocomplete the 3-dimensional reconstruction. This procedure al-lowed calculation of structure and graft volumes, which took intoaccount section thickness (40 �m).
1. Vonsattel JP, et al. (1985) Neuropathological classification of Huntington’s disease.J Neuropathol Exp Neurol 44:559–577.
2. Freeman TB, et al. (2000) Transplanted fetal striatum in Huntington’s disease: pheno-typic development and lack of pathology. Proc Natl Acad Sci USA 97:13877–13882.
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 2 of 7
AQ: V
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
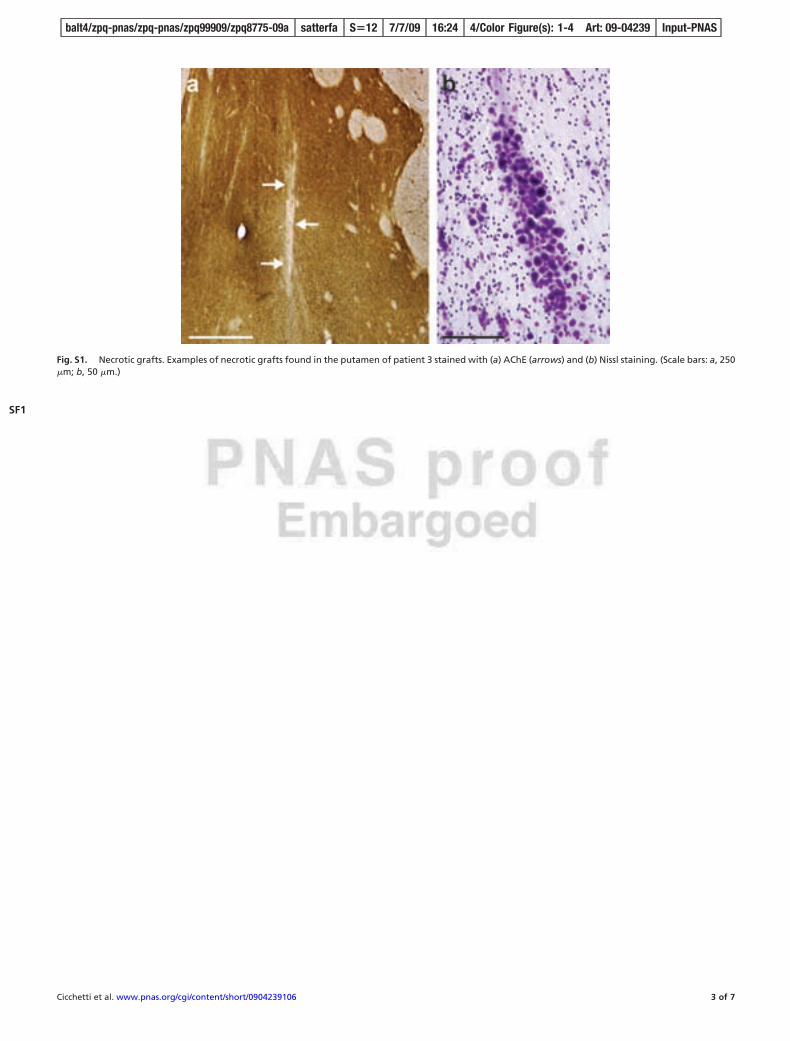
Fig. S1. Necrotic grafts. Examples of necrotic grafts found in the putamen of patient 3 stained with (a) AChE (arrows) and (b) Nissl staining. (Scale bars: a, 250�m; b, 50 �m.)
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 3 of 7
SF1
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
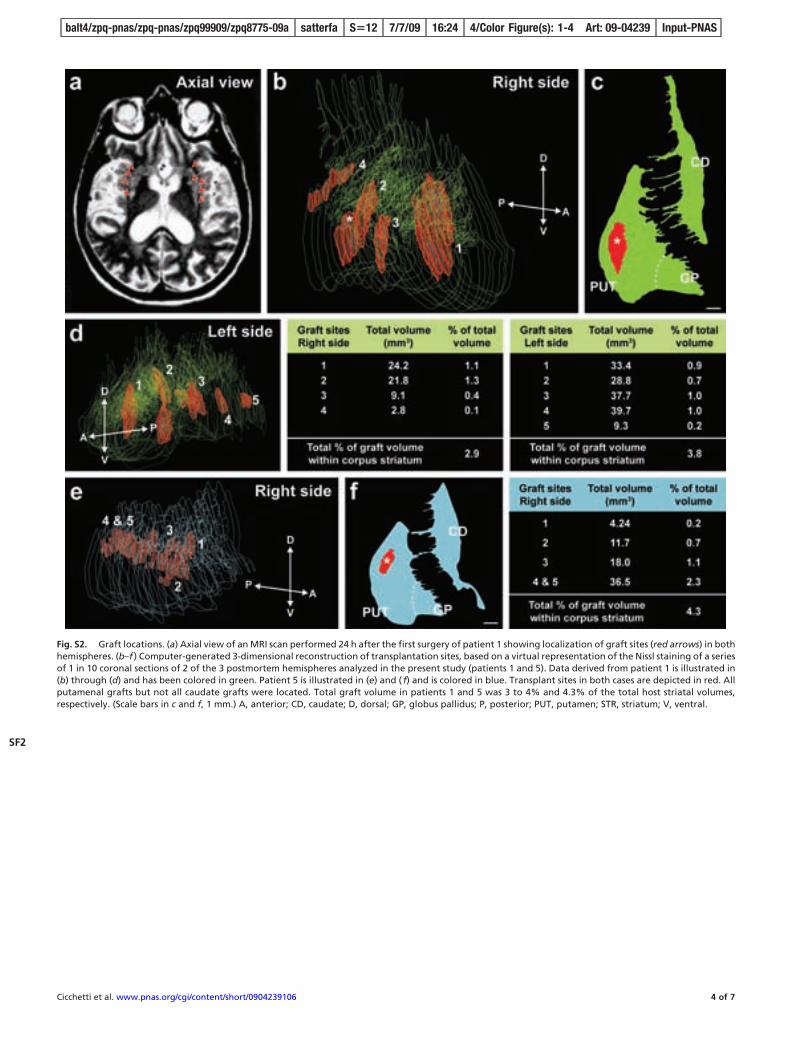
Fig. S2. Graft locations. (a) Axial view of an MRI scan performed 24 h after the first surgery of patient 1 showing localization of graft sites (red arrows) in bothhemispheres. (b–f ) Computer-generated 3-dimensional reconstruction of transplantation sites, based on a virtual representation of the Nissl staining of a seriesof 1 in 10 coronal sections of 2 of the 3 postmortem hemispheres analyzed in the present study (patients 1 and 5). Data derived from patient 1 is illustrated in(b) through (d) and has been colored in green. Patient 5 is illustrated in (e) and ( f) and is colored in blue. Transplant sites in both cases are depicted in red. Allputamenal grafts but not all caudate grafts were located. Total graft volume in patients 1 and 5 was 3 to 4% and 4.3% of the total host striatal volumes,respectively. (Scale bars in c and f, 1 mm.) A, anterior; CD, caudate; D, dorsal; GP, globus pallidus; P, posterior; PUT, putamen; STR, striatum; V, ventral.
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 4 of 7
SF2
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
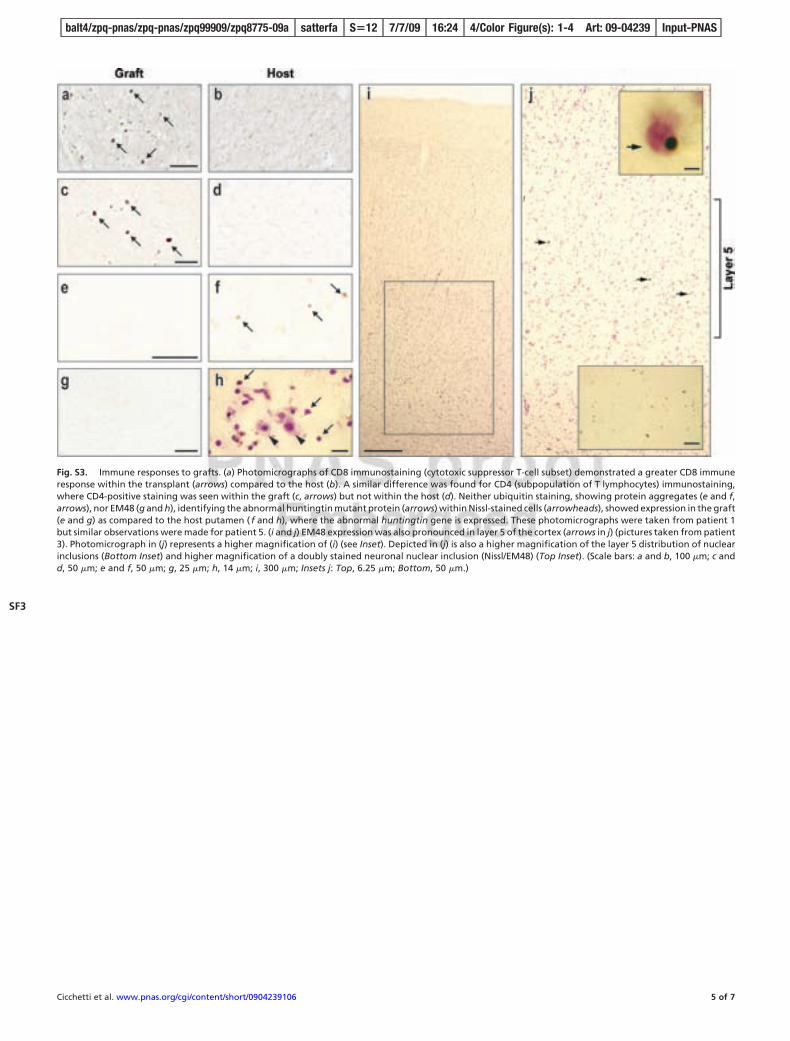
Fig. S3. Immune responses to grafts. (a) Photomicrographs of CD8 immunostaining (cytotoxic suppressor T-cell subset) demonstrated a greater CD8 immuneresponse within the transplant (arrows) compared to the host (b). A similar difference was found for CD4 (subpopulation of T lymphocytes) immunostaining,where CD4-positive staining was seen within the graft (c, arrows) but not within the host (d). Neither ubiquitin staining, showing protein aggregates (e and f,arrows), nor EM48 (g and h), identifying the abnormal huntingtin mutant protein (arrows) within Nissl-stained cells (arrowheads), showed expression in the graft(e and g) as compared to the host putamen ( f and h), where the abnormal huntingtin gene is expressed. These photomicrographs were taken from patient 1but similar observations were made for patient 5. (i and j) EM48 expression was also pronounced in layer 5 of the cortex (arrows in j) (pictures taken from patient3). Photomicrograph in (j) represents a higher magnification of (i) (see Inset). Depicted in (j) is also a higher magnification of the layer 5 distribution of nuclearinclusions (Bottom Inset) and higher magnification of a doubly stained neuronal nuclear inclusion (Nissl/EM48) (Top Inset). (Scale bars: a and b, 100 �m; c andd, 50 �m; e and f, 50 �m; g, 25 �m; h, 14 �m; i, 300 �m; Insets j: Top, 6.25 �m; Bottom, 50 �m.)
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 5 of 7
SF3
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS

Table S1. Patient characteristics
Patient 1 (B.L.) Patient 3 (M.C.) Patient 5 (M.S.)
Gender Female Female FemaleCAG repeats 42 42 42Grades 3 3 2Time from diagnosis (years) 5 12 5Symptom duration (years) 8 17 9Age at transplantation (years) 58 64 59Post-operative latency (years) 9 10.5 9.5Causes of death Aspiration pneumonia
complicated by amyocardial infarction
Cardiorespiratory arrest as a complication of end-stage HD
Immunosuppression CsA 6 mg/kg/day 7 days before first operation continuing for 14 days after second operation, then 2mg/kg/day for 6 months
Number and location oftransplants
1 L caudate 2 L caudate 1 L caudate
4 L putamen 6 L putamen 4 L putamen1 R caudate 2 R caudate 1 R caudate4 R putamen 6 R putamen 5 R putamen
Surgical complications Asymptomatic subacute subduralhematoma after first operation
Asymptomatic corticalhemorrhage and thin subdural
hematoma after first and secondoperations
Bilateral hygromas post-operatively.Conversion of her hygromas intosubdural hematomas requiring
surgical drainage
Data from ref. 1 and subsequent clinical evaluation.
1. Hauser RA, Sandberg PR, Freeman TB, Stoessl AJ (2002) Bilateral human fetal striatal transplantation in Huntington’s disease. Neurology 58:1704, author reply 1704.
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 6 of 7
ST1
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS

Table S2. Clinical outcomes
Patient 1 (B.L.) Patient 3 (M.C.) Patient 5 (M.S.)
UHDRS score pre-operation 42 33 27 (31 after subdural hematoma drainage)UHDRS score post-operation 30 (3 mo) 28 (10 mo) 39 (9 mo)
32 (9 mo) 21 (12 mo) 53 (2.5 yrs)40 (18 mo) 37 (20 mo) 64 (5 yrs)52 (4 yrs)65 (6 yrs)78 (7 yrs)
MMSE score pre-operation 30 26 27MMSE score post-operation 30 (21 mo) 24 (30 mo) 16 (5 yrs)
14 (7 yrs)
MMSE, Mini Mental State Evaluation; UHDRS, Unified Huntington’s Disease Rating Scale.Data from ref. 1 and subsequent clinical evaluation.
1. Hauser RA, Sandberg PR, Freeman TB, Stoessl AJ (2002) Bilateral human fetal striatal transplantation in Huntington’s disease. Neurology 58:1704, author reply 1704.
Cicchetti et al. www.pnas.org/cgi/content/short/0904239106 7 of 7
ST2
balt4/zpq-pnas/zpq-pnas/zpq99909/zpq8775-09a satterfa S�12 7/7/09 16:24 4/Color Figure(s): 1-4 Art: 09-04239 Input-PNAS
Related Documents











