Abstract Retinal degeneration represents a huge burden of blind- ing disease, and currently there are no effective treat- ments that reverse the most common causes of neural retinal degeneration. Stem cell biology has the potential to significantly ease this burden, not only through the development of disease models of retinal degeneration but also in the manufacture of a replacement for the neural retinal tissue. This review summarizes the major advancements in the last decade in the field of neural retinal regeneration with an emphasis on the differen- tiation of embryonic and induced pluripotent stem cells into cells with retinal and specifically photoreceptor characteristics. © 2014 S. Karger AG, Basel Diseases of the retina are responsible for the ma- jority of irreversible blindness worldwide. Age- related macular degeneration (AMD) is the most common cause of blindness in the over 55 age group [1, 2]. It is a complex multifactorial disease with a hereditary component that results in chronic inflammation and subsequent deteriora- tion of the supporting cell layer of the retina the retinal pigment epithelium (RPE) in the central retina. The gradual loss of RPE leads to degenera- tion of the fragile (fig. 1), highly metabolic photo- receptor (PR) cell layer. Neither the RPE nor the cells of the PR layer divide in the adult human, and hence the gradual loss of central vision in AMD patients is irreversible. There is currently no cure for AMD. Stargardt disease (SGTD) is the most common inherited retinal degeneration oc- curring in 1:10,000 live births [3]. The most com- mon mutation is in the ABCA4 gene, which codes for a transmembrane transporter protein that is exclusively expressed in PR cells [4]. Defects in ABCA4 lead to the accumulation of unwanted proteins and the retinal toxin lipofuscin, which damages the PR and RPE. There are currently no established treatments for Stargardt disease. Reti- nitis pigmentosa (RP) is a heterogeneous group of diseases that predominantly affect the rod PR sys- tem. The many gene mutations associated with RP result in a broad phenotype, but in many cas- es, severe vision loss substantially affects quality Casaroli-Marano RP, Zarbin MA (eds): Cell-Based Therapy for Retinal Degenerative Disease. Dev Ophthalmol. Basel, Karger, 2014, vol 53, pp 97–110 (DOI: 10.1159/000357363) Neural Retinal Regeneration with Pluripotent Stem Cells Conor M. Ramsden a, b · Michael B. Powner a · Amanda-Jayne F. Carr a · Matthew J.K. Smart a · Lyndon da Cruz a, b · Peter J. Coffey a, c a The London Project to Cure Blindness, Division of ORBIT, Institute of Ophthalmology, University College London, b NIHR Biomedical Research Centre at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology, London, UK; c Center for Stem Cell Biology and Engineering, NRI, UC Santa Barbara, Santa Barbara, Calif., USA Downloaded by: UCL 144.82.134.189 - 4/25/2014 3:34:51 PM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AbstractRetinal degeneration represents a huge burden of blind-ing disease and currently there are no effective treat-ments that reverse the most common causes of neural retinal degeneration Stem cell biology has the potential to significantly ease this burden not only through the development of disease models of retinal degeneration but also in the manufacture of a replacement for the neural retinal tissue This review summarizes the major advancements in the last decade in the field of neural retinal regeneration with an emphasis on the differen-tiation of embryonic and induced pluripotent stem cells into cells with retinal and specifically photoreceptor characteristics copy 2014 S Karger AG Basel
Diseases of the retina are responsible for the ma-jority of irreversible blindness worldwide Age-related macular degeneration (AMD) is the most common cause of blindness in the over 55 age group [1 2] It is a complex multifactorial disease with a hereditary component that results in chronic inflammation and subsequent deteriora-
tion of the supporting cell layer of the retina the retinal pigment epithelium (RPE) in the central retina The gradual loss of RPE leads to degenera-tion of the fragile (fig 1) highly metabolic photo-receptor (PR) cell layer Neither the RPE nor the cells of the PR layer divide in the adult human and hence the gradual loss of central vision in AMD patients is irreversible There is currently no cure for AMD Stargardt disease (SGTD) is the most common inherited retinal degeneration oc-curring in 1 10000 live births [3] The most com-mon mutation is in the ABCA4 gene which codes for a transmembrane transporter protein that is exclusively expressed in PR cells [4] Defects in ABCA4 lead to the accumulation of unwanted proteins and the retinal toxin lipofuscin which damages the PR and RPE There are currently no established treatments for Stargardt disease Reti-nitis pigmentosa (RP) is a heterogeneous group of diseases that predominantly affect the rod PR sys-tem The many gene mutations associated with RP result in a broad phenotype but in many cas-es severe vision loss substantially affects quality
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363)
Neural Retinal Regeneration with Pluripotent Stem Cells
Conor M Ramsden
a b Michael B Powner
a Amanda-Jayne F Carr
a Matthew JK Smart
a Lyndon da Cruz
a b Peter J Coffey
a c
a The London Project to Cure Blindness Division of ORBIT Institute of Ophthalmology University College
London b NIHR Biomedical Research Centre at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology London UK c Center for Stem Cell Biology and Engineering NRI UC Santa Barbara Santa Barbara Calif USA
Dow
nloa
ded
by
UC
L
144
821
341
89 -
42
520
14 3
34
51 P
M
98 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
of life by the end of the third or fourth decade [5] It affects 1 4000 live births and the most com-monly affected gene is rhodopsin
Clearly there is a need to develop therapies to fully reverse these devastating diseases and a re-generative medicine approach could provide a so-lution Stem cells could be used to either directly replace damaged tissue or rescue cells through their ability to secrete soluble factors that act in a paracrine fashion [6] or by fusion with the failing cells [7] Additionally the development of disease models using induced pluripotent stem cells (IPSC) from patients with identified gene defects in SGTD and RP will enable a greater understand-ing of the underlying molecular pathology of these diseases and facilitate drug discovery and the development of gene therapy
There are several trials currently listed in the World Health Organization clinical trials register which employ a number of stem cell types to treat a variety of retinal diseases [8ndash14] Early results have been published of a trial using human embry-onic stem cell (HESC)-derived RPE cells injected
into the subretinal space as a suspension in one patient with dry AMD and another with SGTD The patient with SGTD improved from hand mo-tions vision to 20800 in the treated eye with no change in hand motions vision in the fellow eye While there were no significant gains in vision for the AMD patient at 4 months there have been no significant adverse effects reported so far [8]
The first question that these trials will answer concerns the safety of retinal stem cell transplan-tation and as result many of the trial subjects have very advanced disease It is therefore unlike-ly that attempting to replace just the RPE [8] or using the paracrine restorative properties of stem cells [10] will reverse pathology in patients with advanced retinal disease where PR cell death and loss of vision are already apparent For these ad-vanced cases the replacement of PR cells will also be required The survival of the output neurons of the eye the retinal ganglion cells in advanced dis-ease [15] suggests that the replacement of PR cells may be a viable option for restoring vision in ret-inal degeneration
BMPh
BM
RPERPE
OSOS
ONL
INLINL
GCLGCGC
BPBP
RRCC
GCL
ONL
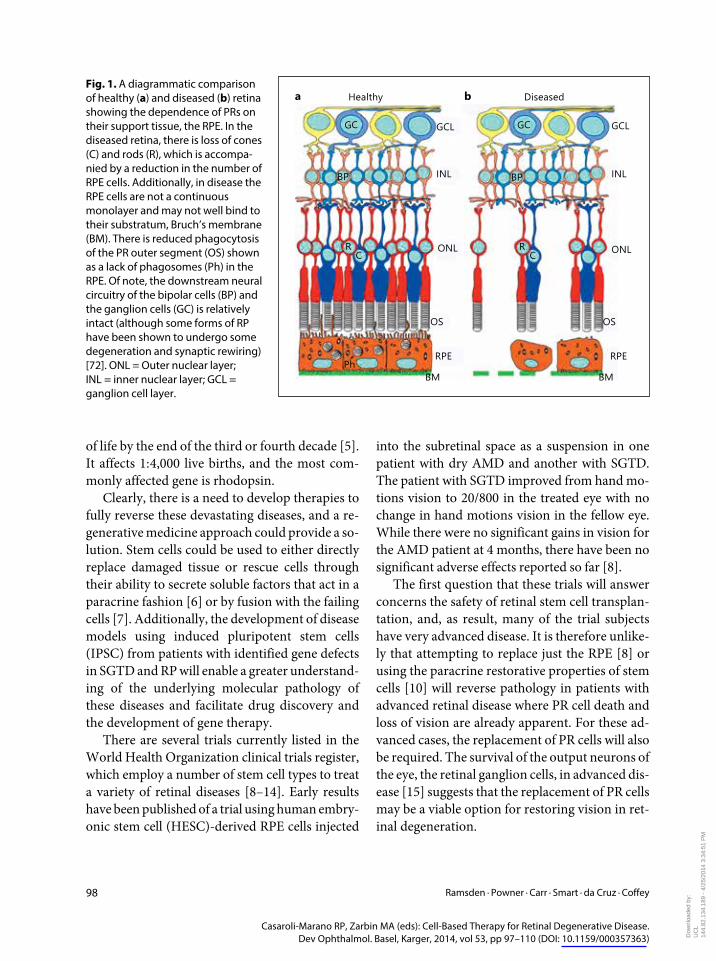
Healthya b DiseasedFig 1 A diagrammatic comparison of healthy (a) and diseased (b) retina showing the dependence of PRs on their support tissue the RPE In the diseased retina there is loss of cones (C) and rods (R) which is accompa-nied by a reduction in the number of RPE cells Additionally in disease the RPE cells are not a continuous monolayer and may not well bind to their substratum Bruchrsquos membrane (BM) There is reduced phagocytosis of the PR outer segment (OS) shown as a lack of phagosomes (Ph) in the RPE Of note the downstream neural circuitry of the bipolar cells (BP) and the ganglion cells (GC) is relatively intact (although some forms of RP have been shown to undergo some degeneration and synaptic rewiring) [72] ONL = Outer nuclear layer INL = inner nuclear layer GCL = ganglion cell layer
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 99
The eye is an ideal organ in which to investi-gate the potential of stem cell therapy because it is surgically and diagnostically accessible many of its diseases are well characterized and it is nones-sential to life Importantly there is a degree of im-mune privilege in the eye conferred by the blood retinal barrier and transplanted cells can be di-rectly monitored in the eye using noninvasive ophthalmic instruments Furthermore there is a wealth of surgical knowledge in dealing with the eye bolstered by the development of experimen-tal techniques and tools in attempts to treat AMD surgically over the last 20 years [16 17]
Retinal Histogenesis and Anatomy
In order to better understand the methods to gen-erate neural retina described herein a brief intro-duction to ocular embryology is included to con-textualize the many exogenous factors used to drive the retinal phenotype and the markers that allow identification of differing retinal cells It should be noted that none of these pathways act in isolation and there is considerable cross talk and activation and antagonism that can reverse depending on the concentration and hence dis-tance between adjacent groups of developing cells
Cell division in the hours following fertiliza-tion is driven by growth factors which in turn switch on and off genes leading to growth and dif-ferentiation of embryonic cells These ontogenet-ically early cells of the inner cell mass at the blas-tocyst stage of development will express so-called markers of pluripotency such as octamer-bind-ing transcription factor 4 (OCT4) sex-determin-ing region Y-box 2 (SOX2) and the homeobox protein Nanog Bone morphogenetic protein 4 (BMP4) is a member of the transforming growth factor-β family BMP4 induces SOX2 and thus helps keep cells in an undifferentiated dividing state As the embryo develops cells begin the pro-cess of differentiation primarily into the three
germ layers the endoderm mesoderm and ecto-derm The cells of the ectoderm undergo further specialization and migration to form the neuro-ectoderm from which the nervous system and eye subsequently develop
Notch signaling pathway is involved in body axis determination and is also expressed later in the developing mammalian retina [18] and acti-vation of this pathway may be involved in the de-termination of a glial neural support tissue the Muller cells [19] Nodal signaling is involved in left-right determination Sonic hedgehog (Shh) expression marks the line of division in the com-mon eye field and knockouts have cyclopia [20] The wingless-related integration site (Wnt) sig-naling pathway is active at increasing levels in the more posterior neural tissues in the neural plate and since the eye develops from the anterior neu-roectoderm it follows that some antagonism of this pathway during ocular organogenesis is nec-essary [21] Exogenous fibroblast growth factor (FGF) drives cells to form posterior mesoderm and hence forms more caudal tissues [22]
There are both acidic (αFGF) and basic (βFGF) FGF receptors in the optic vesicle [23] which evaginates from the diencephalon at 4 weeksrsquo ges-tation in humans and then subsequently invagi-nates upon itself to form the bilayered optic cup at 6 weeks This bilayered structure is formed of two apex-in epithelial cell layers that produce ex-tracellular matrix proteins and growth factors specific to their cell type and polarity This extra-cellular matrix is important for intercellular com-munication and therefore subsequent develop-ment [24] Both layers of the optic cup express the early eye field markers paired homeobox 6 (PAX6) and the retina and anterior neural fold homeobox (RAX) The outer more caudal layer destined to become the RPE initially expresses microphthalmia-associated transcription factor (MITF) a marker of immature RPE The inner layer is destined to become the neural retina which preferentially expresses the CEH-10 ho-meobox protein (CHX10) These neuroretinal
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
100 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
progenitor cells begin to express the PR-specific transcription factor cone rod homeobox (CRX) which together with neural leucine zipper (NRL) a marker of post mitotic rods and RAX activate a cascade of PR-specific genes These include in-terphotoreceptor retinoid-binding protein (IRBP) which can first be detected by immuno-histochemistry at 9 weeksrsquo gestation Blue light-sensitive cones express S-opsin expressed at 14 weeks and M-opsin and L-opsin shown to be ex-pressed in green and red sensitive cones respec-tively are detected by week-16 [25] (fig 2)
Transplantation Strategies
Following the observation that ocular embryo-logical tissue was able to differentiate into retinal tissue when transplanted into the cerebral tissue of rats [26] and transplanted fetal retina was shown to survive in the anterior chamber of adult rat eyes [27] there has been considerable study of fetal retina as a source of transplant tissue Sub-retinal grafts were shown to survive for longer pe-riods if younger donors were used However do-
nors from the very early embryological period produced less organized grafts following trans-plantation Other challenges encountered with retinal transplants are immune rejection and method of transplant Transplants of cell suspen-sions are less able to survive the host immune re-sponse compared to the transplantation of fetal retinal sheets [28]
Using mice with green fluorescent protein (GFP) at the Nrl locus MacLaren et al [29] were able to purify postmitotic rods for transplant A small percentage of the transplants were shown to integrate into the host retina and improve pupil re-sponse to light in a mouse model of RP and more recently improved visual navigation in a mouse model of congenital stationary night blindness [30] Their experiments showed that the ideal on-togenetic age for immature PR transplant in mice is in the first postnatal week However in contrast to these findings Gust and Reh [31] have found that adult mouse retina can also integrate into host retina and suggest that integration of PR cells is dependent on their survival after dissociation
A pilot study of 8 transplants of human retinal fetal aggregates in immunosuppressed human
RPE65bestrophin
PAX6MITFOTX2
IRBPTULP
S-opsinM-opsinL-opsin
9 weeks 14ndash16 weeksPAX6CHX10
CRX
Progenitorcell
PRprecursor
PRdifferentiation
PAX6MITFOTX2
PAX6RAX
OCT4NANOG
SOX2Mature
RPEImmature
RPE
Maturecone
Immaturecone
Maturerod
ImmaturerodRod
Cone
Optic cup
9 weeks
IRBPTULPNRL
RecoverinRhodopsin
14ndash16 weeks
RPE
Retina
FGF-driveninvaginationBlastocyst Eye field
Opticvesicle
4 weeks
Fig 2 Schematic of the markers in human retinal development from the blastocyst stage to the appearance of PR precursors at 16 weeks with time points
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 101
subjects with RP and AMD in the USA and 14 RP patients in India [32 33] did not show any sig-nificant lasting visual improvement however there was no clear evidence of graft rejection While such studies are informative the issues as-sociated with harvesting fetal tissue for trans-plants suggest that this approach may not be ex-trapolated to treat the huge burden of disease caused by retinal pathology worldwide
With the aim to manufacture cells for neural retinal transplant and thus reduce the require-ment for human fetal tissue Klassen et al [34] were able to expand mouse fetal retina cells in cell culture for up to 60 passages The cells expressed early markers of retinal PRs in vitro Following transplantation into a mouse model of RP the cells migrated to the appropriate layer of the reti-na the outer nuclear layer and began to express later markers of retinal development the PR pro-teins recoverin and rhodopsin The mice were able to better perform tasks in low light levels fol-lowing transplantation In discussing the results the authors stated that this functional rescue may have been due to replacement of the damaged PRs but they could not rule out a neuroprotective capacity for the new cells or the role of the mela-nopsin system
Such a manufacturing approach is a move to-wards a more ethically acceptable and easier to manufacture cellular source Add this benefit to a low rate of immunoreactivity and cheaper costs and this approach represents an ideal cellular source for neural retinal regeneration Many of these conditions could be met by the use of plu-ripotent stem cells both embryonic and induced pluripotent cells
Stem Cells
Stem cells have the ability to differentiate into other cell types and are also capable of indefinite self-replication in their undifferentiated state Stem cells may be classified according to the
range of cell types into which they can differenti-ate For example pluripotent cells have the abil-ity to form all three embryological germ layers Stem cells can also be classified by their source HESC originate from the inner cell mass of the blastocyst of 3- to 5-day-old preimplantation embryos and are pluripotent [35] Fetal stem cells are harvested from fetal tissue and are con-sidered multipotent having already specialized to some degree Adult stem cells have also been characterized in most major organs including the eye [36] however these cells exist at a low fre-quency are difficult to harvest and have limited potency IPSC were first generated from differ-entiated mouse somatic cells by forced expres-sion of four transcription factors Oct4 Sox2 c-Myc and Klf4 [37]
Protocols for Deriving Neural Retina from Pluripotent Stem Cells
In essence many of the protocols to derive neural retinal cells from stem cells utilize a two-step pro-cess (table 1) The first step is to drive the cells toward an embryologically anterior neural deter-mination and from there towards a retinal fate These steps are achieved using known agonists and antagonists of early developmental pathways A majority of protocols use embryoid bodies (EBs) or aggregates of stem cells The intercellular cues within these EBs are thought to promote an environment similar to the developing embryo with cells at the outside producing extracellular matrix which in turn promotes the development of cellular polarity Hence the default lineage in minimal culture conditions is ectoderm and spe-cifically neuroectoderm [38]
Ikeda et al [39] reported the generation of neu-ral retinal precursors from mouse embryonic stem cells (ESC) This was achieved by aggregating the cells into EBs in a suspension culture In order to push the cells to an anterior developmental fate the embryological signaling pathways Wnt and
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72
98 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
of life by the end of the third or fourth decade [5] It affects 1 4000 live births and the most com-monly affected gene is rhodopsin
Clearly there is a need to develop therapies to fully reverse these devastating diseases and a re-generative medicine approach could provide a so-lution Stem cells could be used to either directly replace damaged tissue or rescue cells through their ability to secrete soluble factors that act in a paracrine fashion [6] or by fusion with the failing cells [7] Additionally the development of disease models using induced pluripotent stem cells (IPSC) from patients with identified gene defects in SGTD and RP will enable a greater understand-ing of the underlying molecular pathology of these diseases and facilitate drug discovery and the development of gene therapy
There are several trials currently listed in the World Health Organization clinical trials register which employ a number of stem cell types to treat a variety of retinal diseases [8ndash14] Early results have been published of a trial using human embry-onic stem cell (HESC)-derived RPE cells injected
into the subretinal space as a suspension in one patient with dry AMD and another with SGTD The patient with SGTD improved from hand mo-tions vision to 20800 in the treated eye with no change in hand motions vision in the fellow eye While there were no significant gains in vision for the AMD patient at 4 months there have been no significant adverse effects reported so far [8]
The first question that these trials will answer concerns the safety of retinal stem cell transplan-tation and as result many of the trial subjects have very advanced disease It is therefore unlike-ly that attempting to replace just the RPE [8] or using the paracrine restorative properties of stem cells [10] will reverse pathology in patients with advanced retinal disease where PR cell death and loss of vision are already apparent For these ad-vanced cases the replacement of PR cells will also be required The survival of the output neurons of the eye the retinal ganglion cells in advanced dis-ease [15] suggests that the replacement of PR cells may be a viable option for restoring vision in ret-inal degeneration
BMPh
BM
RPERPE
OSOS
ONL
INLINL
GCLGCGC
BPBP
RRCC
GCL
ONL
Healthya b DiseasedFig 1 A diagrammatic comparison of healthy (a) and diseased (b) retina showing the dependence of PRs on their support tissue the RPE In the diseased retina there is loss of cones (C) and rods (R) which is accompa-nied by a reduction in the number of RPE cells Additionally in disease the RPE cells are not a continuous monolayer and may not well bind to their substratum Bruchrsquos membrane (BM) There is reduced phagocytosis of the PR outer segment (OS) shown as a lack of phagosomes (Ph) in the RPE Of note the downstream neural circuitry of the bipolar cells (BP) and the ganglion cells (GC) is relatively intact (although some forms of RP have been shown to undergo some degeneration and synaptic rewiring) [72] ONL = Outer nuclear layer INL = inner nuclear layer GCL = ganglion cell layer
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 99
The eye is an ideal organ in which to investi-gate the potential of stem cell therapy because it is surgically and diagnostically accessible many of its diseases are well characterized and it is nones-sential to life Importantly there is a degree of im-mune privilege in the eye conferred by the blood retinal barrier and transplanted cells can be di-rectly monitored in the eye using noninvasive ophthalmic instruments Furthermore there is a wealth of surgical knowledge in dealing with the eye bolstered by the development of experimen-tal techniques and tools in attempts to treat AMD surgically over the last 20 years [16 17]
Retinal Histogenesis and Anatomy
In order to better understand the methods to gen-erate neural retina described herein a brief intro-duction to ocular embryology is included to con-textualize the many exogenous factors used to drive the retinal phenotype and the markers that allow identification of differing retinal cells It should be noted that none of these pathways act in isolation and there is considerable cross talk and activation and antagonism that can reverse depending on the concentration and hence dis-tance between adjacent groups of developing cells
Cell division in the hours following fertiliza-tion is driven by growth factors which in turn switch on and off genes leading to growth and dif-ferentiation of embryonic cells These ontogenet-ically early cells of the inner cell mass at the blas-tocyst stage of development will express so-called markers of pluripotency such as octamer-bind-ing transcription factor 4 (OCT4) sex-determin-ing region Y-box 2 (SOX2) and the homeobox protein Nanog Bone morphogenetic protein 4 (BMP4) is a member of the transforming growth factor-β family BMP4 induces SOX2 and thus helps keep cells in an undifferentiated dividing state As the embryo develops cells begin the pro-cess of differentiation primarily into the three
germ layers the endoderm mesoderm and ecto-derm The cells of the ectoderm undergo further specialization and migration to form the neuro-ectoderm from which the nervous system and eye subsequently develop
Notch signaling pathway is involved in body axis determination and is also expressed later in the developing mammalian retina [18] and acti-vation of this pathway may be involved in the de-termination of a glial neural support tissue the Muller cells [19] Nodal signaling is involved in left-right determination Sonic hedgehog (Shh) expression marks the line of division in the com-mon eye field and knockouts have cyclopia [20] The wingless-related integration site (Wnt) sig-naling pathway is active at increasing levels in the more posterior neural tissues in the neural plate and since the eye develops from the anterior neu-roectoderm it follows that some antagonism of this pathway during ocular organogenesis is nec-essary [21] Exogenous fibroblast growth factor (FGF) drives cells to form posterior mesoderm and hence forms more caudal tissues [22]
There are both acidic (αFGF) and basic (βFGF) FGF receptors in the optic vesicle [23] which evaginates from the diencephalon at 4 weeksrsquo ges-tation in humans and then subsequently invagi-nates upon itself to form the bilayered optic cup at 6 weeks This bilayered structure is formed of two apex-in epithelial cell layers that produce ex-tracellular matrix proteins and growth factors specific to their cell type and polarity This extra-cellular matrix is important for intercellular com-munication and therefore subsequent develop-ment [24] Both layers of the optic cup express the early eye field markers paired homeobox 6 (PAX6) and the retina and anterior neural fold homeobox (RAX) The outer more caudal layer destined to become the RPE initially expresses microphthalmia-associated transcription factor (MITF) a marker of immature RPE The inner layer is destined to become the neural retina which preferentially expresses the CEH-10 ho-meobox protein (CHX10) These neuroretinal
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
100 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
progenitor cells begin to express the PR-specific transcription factor cone rod homeobox (CRX) which together with neural leucine zipper (NRL) a marker of post mitotic rods and RAX activate a cascade of PR-specific genes These include in-terphotoreceptor retinoid-binding protein (IRBP) which can first be detected by immuno-histochemistry at 9 weeksrsquo gestation Blue light-sensitive cones express S-opsin expressed at 14 weeks and M-opsin and L-opsin shown to be ex-pressed in green and red sensitive cones respec-tively are detected by week-16 [25] (fig 2)
Transplantation Strategies
Following the observation that ocular embryo-logical tissue was able to differentiate into retinal tissue when transplanted into the cerebral tissue of rats [26] and transplanted fetal retina was shown to survive in the anterior chamber of adult rat eyes [27] there has been considerable study of fetal retina as a source of transplant tissue Sub-retinal grafts were shown to survive for longer pe-riods if younger donors were used However do-
nors from the very early embryological period produced less organized grafts following trans-plantation Other challenges encountered with retinal transplants are immune rejection and method of transplant Transplants of cell suspen-sions are less able to survive the host immune re-sponse compared to the transplantation of fetal retinal sheets [28]
Using mice with green fluorescent protein (GFP) at the Nrl locus MacLaren et al [29] were able to purify postmitotic rods for transplant A small percentage of the transplants were shown to integrate into the host retina and improve pupil re-sponse to light in a mouse model of RP and more recently improved visual navigation in a mouse model of congenital stationary night blindness [30] Their experiments showed that the ideal on-togenetic age for immature PR transplant in mice is in the first postnatal week However in contrast to these findings Gust and Reh [31] have found that adult mouse retina can also integrate into host retina and suggest that integration of PR cells is dependent on their survival after dissociation
A pilot study of 8 transplants of human retinal fetal aggregates in immunosuppressed human
RPE65bestrophin
PAX6MITFOTX2
IRBPTULP
S-opsinM-opsinL-opsin
9 weeks 14ndash16 weeksPAX6CHX10
CRX
Progenitorcell
PRprecursor
PRdifferentiation
PAX6MITFOTX2
PAX6RAX
OCT4NANOG
SOX2Mature
RPEImmature
RPE
Maturecone
Immaturecone
Maturerod
ImmaturerodRod
Cone
Optic cup
9 weeks
IRBPTULPNRL
RecoverinRhodopsin
14ndash16 weeks
RPE
Retina
FGF-driveninvaginationBlastocyst Eye field
Opticvesicle
4 weeks
Fig 2 Schematic of the markers in human retinal development from the blastocyst stage to the appearance of PR precursors at 16 weeks with time points
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 101
subjects with RP and AMD in the USA and 14 RP patients in India [32 33] did not show any sig-nificant lasting visual improvement however there was no clear evidence of graft rejection While such studies are informative the issues as-sociated with harvesting fetal tissue for trans-plants suggest that this approach may not be ex-trapolated to treat the huge burden of disease caused by retinal pathology worldwide
With the aim to manufacture cells for neural retinal transplant and thus reduce the require-ment for human fetal tissue Klassen et al [34] were able to expand mouse fetal retina cells in cell culture for up to 60 passages The cells expressed early markers of retinal PRs in vitro Following transplantation into a mouse model of RP the cells migrated to the appropriate layer of the reti-na the outer nuclear layer and began to express later markers of retinal development the PR pro-teins recoverin and rhodopsin The mice were able to better perform tasks in low light levels fol-lowing transplantation In discussing the results the authors stated that this functional rescue may have been due to replacement of the damaged PRs but they could not rule out a neuroprotective capacity for the new cells or the role of the mela-nopsin system
Such a manufacturing approach is a move to-wards a more ethically acceptable and easier to manufacture cellular source Add this benefit to a low rate of immunoreactivity and cheaper costs and this approach represents an ideal cellular source for neural retinal regeneration Many of these conditions could be met by the use of plu-ripotent stem cells both embryonic and induced pluripotent cells
Stem Cells
Stem cells have the ability to differentiate into other cell types and are also capable of indefinite self-replication in their undifferentiated state Stem cells may be classified according to the
range of cell types into which they can differenti-ate For example pluripotent cells have the abil-ity to form all three embryological germ layers Stem cells can also be classified by their source HESC originate from the inner cell mass of the blastocyst of 3- to 5-day-old preimplantation embryos and are pluripotent [35] Fetal stem cells are harvested from fetal tissue and are con-sidered multipotent having already specialized to some degree Adult stem cells have also been characterized in most major organs including the eye [36] however these cells exist at a low fre-quency are difficult to harvest and have limited potency IPSC were first generated from differ-entiated mouse somatic cells by forced expres-sion of four transcription factors Oct4 Sox2 c-Myc and Klf4 [37]
Protocols for Deriving Neural Retina from Pluripotent Stem Cells
In essence many of the protocols to derive neural retinal cells from stem cells utilize a two-step pro-cess (table 1) The first step is to drive the cells toward an embryologically anterior neural deter-mination and from there towards a retinal fate These steps are achieved using known agonists and antagonists of early developmental pathways A majority of protocols use embryoid bodies (EBs) or aggregates of stem cells The intercellular cues within these EBs are thought to promote an environment similar to the developing embryo with cells at the outside producing extracellular matrix which in turn promotes the development of cellular polarity Hence the default lineage in minimal culture conditions is ectoderm and spe-cifically neuroectoderm [38]
Ikeda et al [39] reported the generation of neu-ral retinal precursors from mouse embryonic stem cells (ESC) This was achieved by aggregating the cells into EBs in a suspension culture In order to push the cells to an anterior developmental fate the embryological signaling pathways Wnt and
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 99
The eye is an ideal organ in which to investi-gate the potential of stem cell therapy because it is surgically and diagnostically accessible many of its diseases are well characterized and it is nones-sential to life Importantly there is a degree of im-mune privilege in the eye conferred by the blood retinal barrier and transplanted cells can be di-rectly monitored in the eye using noninvasive ophthalmic instruments Furthermore there is a wealth of surgical knowledge in dealing with the eye bolstered by the development of experimen-tal techniques and tools in attempts to treat AMD surgically over the last 20 years [16 17]
Retinal Histogenesis and Anatomy
In order to better understand the methods to gen-erate neural retina described herein a brief intro-duction to ocular embryology is included to con-textualize the many exogenous factors used to drive the retinal phenotype and the markers that allow identification of differing retinal cells It should be noted that none of these pathways act in isolation and there is considerable cross talk and activation and antagonism that can reverse depending on the concentration and hence dis-tance between adjacent groups of developing cells
Cell division in the hours following fertiliza-tion is driven by growth factors which in turn switch on and off genes leading to growth and dif-ferentiation of embryonic cells These ontogenet-ically early cells of the inner cell mass at the blas-tocyst stage of development will express so-called markers of pluripotency such as octamer-bind-ing transcription factor 4 (OCT4) sex-determin-ing region Y-box 2 (SOX2) and the homeobox protein Nanog Bone morphogenetic protein 4 (BMP4) is a member of the transforming growth factor-β family BMP4 induces SOX2 and thus helps keep cells in an undifferentiated dividing state As the embryo develops cells begin the pro-cess of differentiation primarily into the three
germ layers the endoderm mesoderm and ecto-derm The cells of the ectoderm undergo further specialization and migration to form the neuro-ectoderm from which the nervous system and eye subsequently develop
Notch signaling pathway is involved in body axis determination and is also expressed later in the developing mammalian retina [18] and acti-vation of this pathway may be involved in the de-termination of a glial neural support tissue the Muller cells [19] Nodal signaling is involved in left-right determination Sonic hedgehog (Shh) expression marks the line of division in the com-mon eye field and knockouts have cyclopia [20] The wingless-related integration site (Wnt) sig-naling pathway is active at increasing levels in the more posterior neural tissues in the neural plate and since the eye develops from the anterior neu-roectoderm it follows that some antagonism of this pathway during ocular organogenesis is nec-essary [21] Exogenous fibroblast growth factor (FGF) drives cells to form posterior mesoderm and hence forms more caudal tissues [22]
There are both acidic (αFGF) and basic (βFGF) FGF receptors in the optic vesicle [23] which evaginates from the diencephalon at 4 weeksrsquo ges-tation in humans and then subsequently invagi-nates upon itself to form the bilayered optic cup at 6 weeks This bilayered structure is formed of two apex-in epithelial cell layers that produce ex-tracellular matrix proteins and growth factors specific to their cell type and polarity This extra-cellular matrix is important for intercellular com-munication and therefore subsequent develop-ment [24] Both layers of the optic cup express the early eye field markers paired homeobox 6 (PAX6) and the retina and anterior neural fold homeobox (RAX) The outer more caudal layer destined to become the RPE initially expresses microphthalmia-associated transcription factor (MITF) a marker of immature RPE The inner layer is destined to become the neural retina which preferentially expresses the CEH-10 ho-meobox protein (CHX10) These neuroretinal
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
100 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
progenitor cells begin to express the PR-specific transcription factor cone rod homeobox (CRX) which together with neural leucine zipper (NRL) a marker of post mitotic rods and RAX activate a cascade of PR-specific genes These include in-terphotoreceptor retinoid-binding protein (IRBP) which can first be detected by immuno-histochemistry at 9 weeksrsquo gestation Blue light-sensitive cones express S-opsin expressed at 14 weeks and M-opsin and L-opsin shown to be ex-pressed in green and red sensitive cones respec-tively are detected by week-16 [25] (fig 2)
Transplantation Strategies
Following the observation that ocular embryo-logical tissue was able to differentiate into retinal tissue when transplanted into the cerebral tissue of rats [26] and transplanted fetal retina was shown to survive in the anterior chamber of adult rat eyes [27] there has been considerable study of fetal retina as a source of transplant tissue Sub-retinal grafts were shown to survive for longer pe-riods if younger donors were used However do-
nors from the very early embryological period produced less organized grafts following trans-plantation Other challenges encountered with retinal transplants are immune rejection and method of transplant Transplants of cell suspen-sions are less able to survive the host immune re-sponse compared to the transplantation of fetal retinal sheets [28]
Using mice with green fluorescent protein (GFP) at the Nrl locus MacLaren et al [29] were able to purify postmitotic rods for transplant A small percentage of the transplants were shown to integrate into the host retina and improve pupil re-sponse to light in a mouse model of RP and more recently improved visual navigation in a mouse model of congenital stationary night blindness [30] Their experiments showed that the ideal on-togenetic age for immature PR transplant in mice is in the first postnatal week However in contrast to these findings Gust and Reh [31] have found that adult mouse retina can also integrate into host retina and suggest that integration of PR cells is dependent on their survival after dissociation
A pilot study of 8 transplants of human retinal fetal aggregates in immunosuppressed human
RPE65bestrophin
PAX6MITFOTX2
IRBPTULP
S-opsinM-opsinL-opsin
9 weeks 14ndash16 weeksPAX6CHX10
CRX
Progenitorcell
PRprecursor
PRdifferentiation
PAX6MITFOTX2
PAX6RAX
OCT4NANOG
SOX2Mature
RPEImmature
RPE
Maturecone
Immaturecone
Maturerod
ImmaturerodRod
Cone
Optic cup
9 weeks
IRBPTULPNRL
RecoverinRhodopsin
14ndash16 weeks
RPE
Retina
FGF-driveninvaginationBlastocyst Eye field
Opticvesicle
4 weeks
Fig 2 Schematic of the markers in human retinal development from the blastocyst stage to the appearance of PR precursors at 16 weeks with time points
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 101
subjects with RP and AMD in the USA and 14 RP patients in India [32 33] did not show any sig-nificant lasting visual improvement however there was no clear evidence of graft rejection While such studies are informative the issues as-sociated with harvesting fetal tissue for trans-plants suggest that this approach may not be ex-trapolated to treat the huge burden of disease caused by retinal pathology worldwide
With the aim to manufacture cells for neural retinal transplant and thus reduce the require-ment for human fetal tissue Klassen et al [34] were able to expand mouse fetal retina cells in cell culture for up to 60 passages The cells expressed early markers of retinal PRs in vitro Following transplantation into a mouse model of RP the cells migrated to the appropriate layer of the reti-na the outer nuclear layer and began to express later markers of retinal development the PR pro-teins recoverin and rhodopsin The mice were able to better perform tasks in low light levels fol-lowing transplantation In discussing the results the authors stated that this functional rescue may have been due to replacement of the damaged PRs but they could not rule out a neuroprotective capacity for the new cells or the role of the mela-nopsin system
Such a manufacturing approach is a move to-wards a more ethically acceptable and easier to manufacture cellular source Add this benefit to a low rate of immunoreactivity and cheaper costs and this approach represents an ideal cellular source for neural retinal regeneration Many of these conditions could be met by the use of plu-ripotent stem cells both embryonic and induced pluripotent cells
Stem Cells
Stem cells have the ability to differentiate into other cell types and are also capable of indefinite self-replication in their undifferentiated state Stem cells may be classified according to the
range of cell types into which they can differenti-ate For example pluripotent cells have the abil-ity to form all three embryological germ layers Stem cells can also be classified by their source HESC originate from the inner cell mass of the blastocyst of 3- to 5-day-old preimplantation embryos and are pluripotent [35] Fetal stem cells are harvested from fetal tissue and are con-sidered multipotent having already specialized to some degree Adult stem cells have also been characterized in most major organs including the eye [36] however these cells exist at a low fre-quency are difficult to harvest and have limited potency IPSC were first generated from differ-entiated mouse somatic cells by forced expres-sion of four transcription factors Oct4 Sox2 c-Myc and Klf4 [37]
Protocols for Deriving Neural Retina from Pluripotent Stem Cells
In essence many of the protocols to derive neural retinal cells from stem cells utilize a two-step pro-cess (table 1) The first step is to drive the cells toward an embryologically anterior neural deter-mination and from there towards a retinal fate These steps are achieved using known agonists and antagonists of early developmental pathways A majority of protocols use embryoid bodies (EBs) or aggregates of stem cells The intercellular cues within these EBs are thought to promote an environment similar to the developing embryo with cells at the outside producing extracellular matrix which in turn promotes the development of cellular polarity Hence the default lineage in minimal culture conditions is ectoderm and spe-cifically neuroectoderm [38]
Ikeda et al [39] reported the generation of neu-ral retinal precursors from mouse embryonic stem cells (ESC) This was achieved by aggregating the cells into EBs in a suspension culture In order to push the cells to an anterior developmental fate the embryological signaling pathways Wnt and
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72
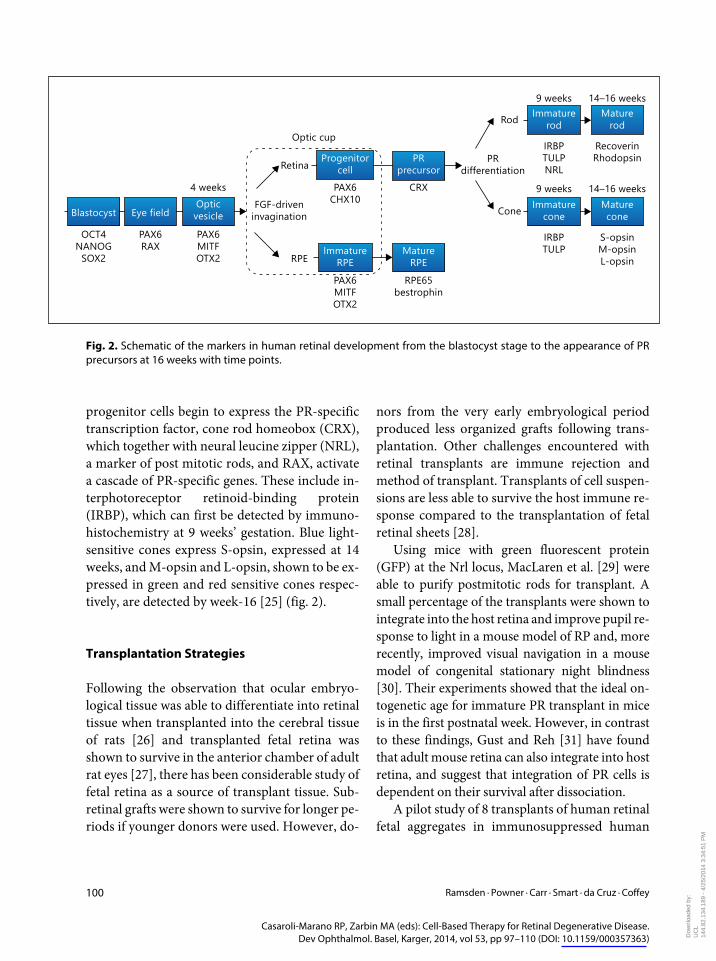
100 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
progenitor cells begin to express the PR-specific transcription factor cone rod homeobox (CRX) which together with neural leucine zipper (NRL) a marker of post mitotic rods and RAX activate a cascade of PR-specific genes These include in-terphotoreceptor retinoid-binding protein (IRBP) which can first be detected by immuno-histochemistry at 9 weeksrsquo gestation Blue light-sensitive cones express S-opsin expressed at 14 weeks and M-opsin and L-opsin shown to be ex-pressed in green and red sensitive cones respec-tively are detected by week-16 [25] (fig 2)
Transplantation Strategies
Following the observation that ocular embryo-logical tissue was able to differentiate into retinal tissue when transplanted into the cerebral tissue of rats [26] and transplanted fetal retina was shown to survive in the anterior chamber of adult rat eyes [27] there has been considerable study of fetal retina as a source of transplant tissue Sub-retinal grafts were shown to survive for longer pe-riods if younger donors were used However do-
nors from the very early embryological period produced less organized grafts following trans-plantation Other challenges encountered with retinal transplants are immune rejection and method of transplant Transplants of cell suspen-sions are less able to survive the host immune re-sponse compared to the transplantation of fetal retinal sheets [28]
Using mice with green fluorescent protein (GFP) at the Nrl locus MacLaren et al [29] were able to purify postmitotic rods for transplant A small percentage of the transplants were shown to integrate into the host retina and improve pupil re-sponse to light in a mouse model of RP and more recently improved visual navigation in a mouse model of congenital stationary night blindness [30] Their experiments showed that the ideal on-togenetic age for immature PR transplant in mice is in the first postnatal week However in contrast to these findings Gust and Reh [31] have found that adult mouse retina can also integrate into host retina and suggest that integration of PR cells is dependent on their survival after dissociation
A pilot study of 8 transplants of human retinal fetal aggregates in immunosuppressed human
RPE65bestrophin
PAX6MITFOTX2
IRBPTULP
S-opsinM-opsinL-opsin
9 weeks 14ndash16 weeksPAX6CHX10
CRX
Progenitorcell
PRprecursor
PRdifferentiation
PAX6MITFOTX2
PAX6RAX
OCT4NANOG
SOX2Mature
RPEImmature
RPE
Maturecone
Immaturecone
Maturerod
ImmaturerodRod
Cone
Optic cup
9 weeks
IRBPTULPNRL
RecoverinRhodopsin
14ndash16 weeks
RPE
Retina
FGF-driveninvaginationBlastocyst Eye field
Opticvesicle
4 weeks
Fig 2 Schematic of the markers in human retinal development from the blastocyst stage to the appearance of PR precursors at 16 weeks with time points
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 101
subjects with RP and AMD in the USA and 14 RP patients in India [32 33] did not show any sig-nificant lasting visual improvement however there was no clear evidence of graft rejection While such studies are informative the issues as-sociated with harvesting fetal tissue for trans-plants suggest that this approach may not be ex-trapolated to treat the huge burden of disease caused by retinal pathology worldwide
With the aim to manufacture cells for neural retinal transplant and thus reduce the require-ment for human fetal tissue Klassen et al [34] were able to expand mouse fetal retina cells in cell culture for up to 60 passages The cells expressed early markers of retinal PRs in vitro Following transplantation into a mouse model of RP the cells migrated to the appropriate layer of the reti-na the outer nuclear layer and began to express later markers of retinal development the PR pro-teins recoverin and rhodopsin The mice were able to better perform tasks in low light levels fol-lowing transplantation In discussing the results the authors stated that this functional rescue may have been due to replacement of the damaged PRs but they could not rule out a neuroprotective capacity for the new cells or the role of the mela-nopsin system
Such a manufacturing approach is a move to-wards a more ethically acceptable and easier to manufacture cellular source Add this benefit to a low rate of immunoreactivity and cheaper costs and this approach represents an ideal cellular source for neural retinal regeneration Many of these conditions could be met by the use of plu-ripotent stem cells both embryonic and induced pluripotent cells
Stem Cells
Stem cells have the ability to differentiate into other cell types and are also capable of indefinite self-replication in their undifferentiated state Stem cells may be classified according to the
range of cell types into which they can differenti-ate For example pluripotent cells have the abil-ity to form all three embryological germ layers Stem cells can also be classified by their source HESC originate from the inner cell mass of the blastocyst of 3- to 5-day-old preimplantation embryos and are pluripotent [35] Fetal stem cells are harvested from fetal tissue and are con-sidered multipotent having already specialized to some degree Adult stem cells have also been characterized in most major organs including the eye [36] however these cells exist at a low fre-quency are difficult to harvest and have limited potency IPSC were first generated from differ-entiated mouse somatic cells by forced expres-sion of four transcription factors Oct4 Sox2 c-Myc and Klf4 [37]
Protocols for Deriving Neural Retina from Pluripotent Stem Cells
In essence many of the protocols to derive neural retinal cells from stem cells utilize a two-step pro-cess (table 1) The first step is to drive the cells toward an embryologically anterior neural deter-mination and from there towards a retinal fate These steps are achieved using known agonists and antagonists of early developmental pathways A majority of protocols use embryoid bodies (EBs) or aggregates of stem cells The intercellular cues within these EBs are thought to promote an environment similar to the developing embryo with cells at the outside producing extracellular matrix which in turn promotes the development of cellular polarity Hence the default lineage in minimal culture conditions is ectoderm and spe-cifically neuroectoderm [38]
Ikeda et al [39] reported the generation of neu-ral retinal precursors from mouse embryonic stem cells (ESC) This was achieved by aggregating the cells into EBs in a suspension culture In order to push the cells to an anterior developmental fate the embryological signaling pathways Wnt and
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 101
subjects with RP and AMD in the USA and 14 RP patients in India [32 33] did not show any sig-nificant lasting visual improvement however there was no clear evidence of graft rejection While such studies are informative the issues as-sociated with harvesting fetal tissue for trans-plants suggest that this approach may not be ex-trapolated to treat the huge burden of disease caused by retinal pathology worldwide
With the aim to manufacture cells for neural retinal transplant and thus reduce the require-ment for human fetal tissue Klassen et al [34] were able to expand mouse fetal retina cells in cell culture for up to 60 passages The cells expressed early markers of retinal PRs in vitro Following transplantation into a mouse model of RP the cells migrated to the appropriate layer of the reti-na the outer nuclear layer and began to express later markers of retinal development the PR pro-teins recoverin and rhodopsin The mice were able to better perform tasks in low light levels fol-lowing transplantation In discussing the results the authors stated that this functional rescue may have been due to replacement of the damaged PRs but they could not rule out a neuroprotective capacity for the new cells or the role of the mela-nopsin system
Such a manufacturing approach is a move to-wards a more ethically acceptable and easier to manufacture cellular source Add this benefit to a low rate of immunoreactivity and cheaper costs and this approach represents an ideal cellular source for neural retinal regeneration Many of these conditions could be met by the use of plu-ripotent stem cells both embryonic and induced pluripotent cells
Stem Cells
Stem cells have the ability to differentiate into other cell types and are also capable of indefinite self-replication in their undifferentiated state Stem cells may be classified according to the
range of cell types into which they can differenti-ate For example pluripotent cells have the abil-ity to form all three embryological germ layers Stem cells can also be classified by their source HESC originate from the inner cell mass of the blastocyst of 3- to 5-day-old preimplantation embryos and are pluripotent [35] Fetal stem cells are harvested from fetal tissue and are con-sidered multipotent having already specialized to some degree Adult stem cells have also been characterized in most major organs including the eye [36] however these cells exist at a low fre-quency are difficult to harvest and have limited potency IPSC were first generated from differ-entiated mouse somatic cells by forced expres-sion of four transcription factors Oct4 Sox2 c-Myc and Klf4 [37]
Protocols for Deriving Neural Retina from Pluripotent Stem Cells
In essence many of the protocols to derive neural retinal cells from stem cells utilize a two-step pro-cess (table 1) The first step is to drive the cells toward an embryologically anterior neural deter-mination and from there towards a retinal fate These steps are achieved using known agonists and antagonists of early developmental pathways A majority of protocols use embryoid bodies (EBs) or aggregates of stem cells The intercellular cues within these EBs are thought to promote an environment similar to the developing embryo with cells at the outside producing extracellular matrix which in turn promotes the development of cellular polarity Hence the default lineage in minimal culture conditions is ectoderm and spe-cifically neuroectoderm [38]
Ikeda et al [39] reported the generation of neu-ral retinal precursors from mouse embryonic stem cells (ESC) This was achieved by aggregating the cells into EBs in a suspension culture In order to push the cells to an anterior developmental fate the embryological signaling pathways Wnt and
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

102 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Nodal were antagonized with respective antago-nists Dickkopf (Dkk1) and left-right determina-tion factor (Lefty) Serum was omitted until day 3 The EBs were then moved to an adherent culture with the addition of Activin a member of the transforming growth factor-β superfamily The cultures showed significant increase in expression of retinal progenitor markers Rax and Pax6 com-pared to controls On day-12 6 of the colonies and 05 of the cells were positive for Crx an ear-ly marker of PR development Crx and orthoden-ticle homeobox 2 (Otx2) expression co-localized consistent with PR development in vivo Follow-ing this step these differentiated ESC derived cells were co-cultured for 12 days with retinal explants from embryonic mouse eyes Some of these ESC-
derived cells became positive for the PR markers rhodopsin and recoverin
The same research group generated retinal precursors from HESC using a similar protocol but with longer time period reflecting the relative difference in gestation period between the two species [40] After day 120 retinoic acid taurine and N-2 supplement (see box 1) were added to the culture medium for a further 30 days At day 120 11 of cells were CRX positive and rhodopsin was observed in 9 of cells at day 200 Using an almost identical protocol Hirami et al [41] de-rived retinal precursors from mouse and human IPSC In the human cells 14 of colonies were CRX positive at day 80 and 26 showed staining for recoverin on day 120
Table 1 Summary of protocols used to derive PR-like cells from pluripotent stem cells
Species Celltype
Conditions Crx+cells
Crx+colonies
Day First author and year
Mouse ESC EBAd D L Ac 05 6 12 Ikeda 2005 [39]Human ESC EBAd D IGF1 B27 No N2 βFGF 12 24 Lamba 2006 [43]Human ESC EBAd D L RA Ta 11 120 Osakada 2008 [40]Mouse IPSC EBAd D L RA Ta na na na Hirami 2009 [41]Human IPSC EBAd D L RA Ta 14 80 Hirami 2009 [41]Human ESC and IPSC EBAd CKI7 SB431542 RA Ta + 100 Osakada 2009 [66]Human IPSC Ad Ma D IGF1 No B27 N2 11 60 Lamba 2009 [44]Human ESC Ad No B27 EGF βFGF 01 28 Banin 2006 [45]Mouse IPSC EBAd N2 B27 Fi No βFGF and retinal explant 23 50 Parameswaran 2010 [46]Human ESC AgAd ndashβFGF N2 B27 19 80 Meyer 2009 [47]Human IPSC AgAd ndashβFGF N2 B27 14 80 Meyer 2009 [47]Human ESC AgAd ndashβFGF No D N2 B27 56 80 Meyer 2011 [48]Human ESC and IPSC EBAd B27 N2 No D IGF1 βFGF Shh T3
RA Ta AcA16 45 Mellough 2012 [52]
Mouse ESC EB Ma N2 RA Ta + 18 Eiraku 2011 [53]Human ESC EB wntndash wnt+ SAG N2 RA 15 126 Nakano 2012 [55]Human ESC Ad Ma N2 B27 RA Ta βFGF αFGF Shh 60 10 Boucherie 2013 [57]Human IPSC AgAd ndashβFGF N2 B27 + 70 Phillips 2012 [49]Mouse IPSC EBAd B27 N2 No IGF1 D βFGF αFGF DAPT 33 33 Tucker 2011 [50]Human ESC Ad B27 N2 No D IGF1 FGF9 βFGF na na Hambright 2012 [51]Mouse ESC EBAd D L βFGF αFGF DAPT Ac Shh RA Ta 75 28 Garita-Hernandez 2013
[56]
Ad = Adherent culture D = Dkk1 L = Lefty Ac = Activin B27 = B-27 supplement No = Noggin N2 = N-2 supplement ndashβFGF = without βFGF RA = retinoic acid Ta = taurine Ma = Matrigel EGF = epidermal growth factor Fi = fibronectin T3 = 33prime5-triiodo-L-thyronine DAPT = Notch pathway inhibitor + = present but no given na = not available
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 103
Incorporating a previous observation that in-sulin-like growth factor 1 (IGF1) was able to in-duce eye field formation in Xenopus embryos [42] Lamba et al [43] generated retinal progeni-tors using HESC in EB serum-free culture with the addition of DKK1 IGF1 Noggin (an antago-nist of BMP4) and B-27 supplement (see box 1) for 3 days Following this step the EBs were trans-ferred to adherent cultures and factors N-2 and βFGF were added for a further 3 weeks These cul-ture conditions allowed a majority of cells to ex-press PAX6 12 of cells expressed CRX and 6 expressed NRL a postmitotic marker of rod PR development These cells were later transplanted into the eyes of an animal model of Leber con-genital amaurosis which has the Crx gene knocked out The mice with successful trans-plants had some electrical activity restored to their retinae the amplitude of which was propor-tional the number of cells that had integrated [44]
The same group was able to derive cells that express markers of PRs using human IPSC In-stead of making EBs the cells were allowed to form clumps and transferred immediately to ad-
herent culture on MatrigelTM a mixture of growth factors and basement membrane proteins (in-cluding collagen IV and laminin) derived from mouse sarcoma cells Retinal induction was driv-en by the addition of IGF1 Noggin and Dkk1 for 3 weeks and then the culture was maintained for several months with N-2 and B-27 supplement At month 2 11 of cells expressed CRX of which 30 were NRL positive While there was some evidence of markers of more differentiated PRs such as recoverin and rhodopsin they were at low levels (lt1) This IPSC-derived retinal line was then infected with a lentivirus containing GFP under the control of PR-specific promoter of IRBP Cells expressing GFP were then sorted by fluorescence to purify a PR population for trans-plantation to the subretinal space of wild-type mice in which a small subset of these survived and expressed PR markers such as recoverin and rhodopsin
Banin et al [45] derived neural precursors from HESC using adherent culture conditions with serum Noggin was added for the first 8 days and then removed for 5 days Darker colony patches were excised and transferred to serum-
Box 1 Cell culture supplementation
Many cell culture protocols supplement basic culture media with fetal bovine serum (FBS) of which albumin is a major component FBS is preferred to adult bovine serum as it has a lower concentration of antibodies and higher concentration of growth factors Major drawbacks to using FBS in cell culture arise from the fact that not all its of constituents can be identified and those that could be are often not quantified Additionally there is variability between batches This inconsistency makes it difficult to ascertain what effect exogenous factors added by an investigator to cell culture may have
In order to overcome some of these difficulties various supplements with known additives are used in place of FBS N-2 supplement which contains insulin transferrin putrescine selenite and progesterone was first defined in 1978 [70] The addition of just these five factors allowed proliferation of rat neuroblastoma cells at a rate equivalent to the addition of 10 FBS B-27 supplement was first described as a useful supplement for the culture of rat hippocampal neurons [71] B-27 comprises biotin α-tocopherol retinol albumin catalase insulin transferrin superoxide dismutase corticosterone galactose ethanolamine glutathione carnitine linoleic acid linolenic acid progesterone putrescine selenite and tri-iodo-L-thyronine
N-2 and B-27 overcome much of the inconsistency of using serum supplementation however they contain animal-derived albumin and as such are not ideal for use in cells for human transplantation For this application chemically defined media which do not contain any animal derivatives may be preferable
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

104 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
free adherent culture with B-27 βFGF and epi-dermal growth factor thereafter These cells ex-pressed markers of immature and mature neu-rons While many of the cells expressed PAX6 CRX was only detected in lt01 of cells The cells were subsequently transplanted to neonatal and adult rat eyes and were shown to survive in 50 of cases In some cases the cells were shown to migrate into the inner plexiform layer and poten-tially formed synapses There was histological ev-idence of NRL expression and the cone-specific marker blue cone opsin in the HESC-derived transplanted cells
Parameswaran et al [46] were able to generate PR-like cells from mouse IPSC using an EB pro-tocol with supplementation of N-2 B-27 insulin fibronectin and Noggin for 10 days The EBs were subsequently dissociated and allowed to ad-here to laminin-coated plates for 25 days Finally the cells were co-cultured with explants from postnatal mouse retina for 10 days with just βFGF and N-2 added to the culture medium At day-50 immunocytochemistry revealed that cells ex-pressed both Crx (23) and Nrl (15) Following intravitreal transplantation there was some evi-dence of rhodopsin expression a later marker of PRs and no teratoma formation was observed
Using HESC and human IPSC Meyer et al [47] derived retina-like cells using floating aggre-gates of cells initially cultured without βFGF fol-lowed by the addition of N-2 and heparin for neu-ral induction These aggregates were then allowed to adhere to laminin-coated plates and were cul-tured with B-27 supplement At day 80 19 of HESC-derived colonies and 14 of IPSC colonies were positive for CRX of which almost half ex-pressed markers of more mature PRs such as re-coverin
The same group refined their protocol with the addition of Noggin and Dkk1 in the first days of the protocol With this modification they were able to increase the amount of CRX-positive cells to 56 by day 80 [48] Within the colonies cells that expressed recoverin appeared to segregate
apart from cells expressing brain-specific homeo-box 3 (BRN3) a marker of ganglion cells Fur-thermore the cells positive for recoverin had an electrophysiological profile consistent with PRs The same group was also able to generate PR-like cells using a human IPSC line derived from circu-lating T cells using a similar protocol [49] These retina-like cells formed basic laminar structures akin to the developing retina in vivo and ex-pressed CRX recoverin rhodopsin and S-opsin
Tucker et al [50] derived PR-like cells using mouse ESC and EBs in serum-free media with B-27 N-2 Noggin IGF1 Dkk1 βFGF αFGF and N-[N-(35-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) In 33 days they produced colonies in which 33 of cells were positive for Crx Expression of recoverin and rhodopsin was also present Calcium imag-ing studies showed a physiological pattern simi-lar to normal PR cells The cells were transplant-ed into the subretinal space of RP disease model mice where they migrated to the outer nuclear layer and rescued some retinal electric activity In a separate experiment cells that expressed mark-ers of less differentiation were injected into the eyes of immune-deficient mice and were shown to form teratomata
Hambright et al [51] generated PR-like cells from HESC cells on adherent culture with B-27 N-2 Noggin Dkk1 IGF1 FGF9 and βFGF While the cells were positive for the early eye field markers PAX6 and RAX there was no expression of recoverin Following subretinal transplanta-tion into mouse eyes some expression of recov-erin was noted after 3 months Of interest they noted no rejection of these xenotransplants de-spite not using any immunosuppression
Using an EB system Mellough et al [52] com-pared the effects of three different culture condi-tions with EBs derived from human IPSC and HESC that were changed to adherent culture on day 30 The experiment ran over 60 days Firstly they used the comprehensive collection of B-27 and N-2 supplements Noggin Dkk1 IGF1
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 105
βFGF Shh 33prime5-triiodo-L-thyronine retinoic acid taurine and activin A in serum-free basal culture medium These cultures were compared with EBs grown in basal medium without any fac-tors and with EBs grown in basal medium supple-mented only with B-27 and N-2 Flow cytometry revealed that 16 of cells expressed CRX with all the factors in the medium at day 45 There was a peak of expression of the L-opsin at day 45 (41 of cells) but this was reduced by day 60 (13 of cells) Interestingly the basal medium with just the N-2 and B-27 supplements was also able to generate cells with PR qualities with only slightly lower efficiency In an attempt to tease out which factors had the greatest effect in retinal determi-nation the EBs were grown in single isolated fac-tor conditions They determined that B-27 N-2 Dkk1 and Noggin have important roles in gen-eration of PR-like cells and that there is an endog-enous upregulation of transcription factors in-volved in neural retinal determination in stem cells exposed to minimal culture conditions
This endogenous ability of stem cell lines to develop into neural retina was elegantly illustrat-ed by Eiraku et al [53] and Eiraku and Sasai [54] who used mouse ESCs in EBs grown in culture medium containing Matrigel With just the addi-tion of N-2 and taurine the EBs spontaneously form vesicles that express Rax that undergo an in-vagination process quite like the formation of the optic cup in vivo By seeding HESC at a higher number to form larger EBs over a longer period of time the same group generated optic vesicle-like structures by firstly blocking the Wnt path-way and subsequent agonism of both the Wnt and hedgehog pathways [55] under high-oxygen con-ditions Vesicles expressing high levels of RAX were excised from the EBs and cultured with N-2 taurine and retinoic acid and were shown to form a multilayered tissue similar to the in vivo devel-oping eye cup that expresses CRX NRL recov-erin and BRN3 CRX was expressed in 15 of cells at day 126 The expression of CRX and re-coverin was increased to 40 by the addition of
DAPT an inhibitor of the Notch pathway In ad-dition to neural retinal precursors these optic cup-like structures also express markers of RPE such as MITF in the external pigmented cell lay-er next to the putative PR layer which mirrors the anatomy in vivo These stem cell-derived eye cups are likely to be very useful in generating disease models of outer retinal degeneration to study the close interaction between the PRs and RPE In or-der to produce these eyecup-like cells high oxy-gen conditions were used
In contrast to the high oxygen conditions re-quired for PR generation by the Sasai laboratory another group found that relative hypoxia in-creases the efficiency of PR-like cell generation from mouse ESC cultured as EBs and plated out after exposure to Dkk1 Lefty serum Activin A DAPT αFGF βFGF Shh retinoic acid and tau-rine resulting in 75 expression of Crx The rea-son proposed for the high yield of Crx cells is that the relative hypoxic conditions are similar to the oxygen tension in utero [56] Subretinal injection of these cells in wild-type mice showed continued expression of rhodopsin migration throughout the layers of the retina and no tera-toma formation
An accelerated method has been devised to generate PR-like cells from HESC using an adher-ent culture with Matrigel N-2 B-27 taurine ret-inoic acid FGF and Shh that promotes 60 of the cells to express CRX at day 10 and NRL at day 28 [57] This approach further highlights the im-portant role that the extracellular matrix plays in promoting neural retinal generation
While there has been success in generating cells that express markers consistent with PR cells the biggest obstacle yet to overcome is the generation of cells with full PR morphology in-cluding the outer segments and the ability to re-spond to light After that another hurdle is to ensure that upon transplantation they can form meaningful synaptic connections with their complex downstream circuitry of the bipolar amacrine horizontal and ganglion cells
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

106 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
Disease Modeling
IPSC technology is a powerful method with which to develop disease models Not only is this tech-nology useful in enabling better pathological characterization of the molecular and histological basis of diseases it will allow more targeted drug development and gene therapy An IPSC line was developed from a patient with gyrate atrophy a retinal degeneration that is caused by a single gene defect in the gene for the enzyme ornithine aminotransferase (OAT) These cells were in-duced to a retinal fate using Dkk1 Noggin B-27 and N-2 supplements Enzyme assay showed low levels of OAT activity in patient-derived IPSC cells compared to controls The addition of in-creasing levels of vitamin B6 which is thought to be an OAT cofactor increased OAT activity in the IPSC line In a further experiment a function-al OAT gene was delivered to the gyrate atrophy IPSC line using a bacterial artificial chromosome This genetically modified IPSC line was then ex-posed to the retinal differentiation protocol and assessed for OAT activity which was shown to be at normal levels [48]
Using a modified version of the protocol orig-inally developed by Jin et al [58] IPSC lines from 5 patients with RP covering differing mutations in three causative genes of late-onset RP RP1 PRPH2 and RHO and one causative of early on-set RP RP9 were differentiated into retinal phe-notypes At day 120 there was no significant dif-ference in rod cell differentiation between the RP1 PRPH2 and RHO cell lines as measured by expression of rhodopsin In contrast the RP9 line showed a much lower level of rhodopsin expres-sion which is in keeping with the relative severity of the disease phenotypes The RP9 line was also observed to have higher levels of oxidative stress at day 100 The RHO cell line displayed an abnor-mal accumulation of rhodopsin in the cytoplasm This was suggested to be because of abnormal un-folding of the protein consistent with the pre-sumed pathological mechanism of this subtype of
RP In the final part of the report the effects of differing levels of α-tocopherol ascorbic acid and β-carotene on the IPSC-derived RP lines at day 120 for 7 days were assessed α-Tocopherol increased the levels of rhodopsin-positive cells in the RP9 IPSC line but had no effect on the others These experiments suggest that IPSC cells may be a powerful tool to investigate the pathogenesis of human retinal diseases originating in PR cells
Future Perspectives Transplantation of Stem Cell-Derived Neural Retina in Humans
There are several phase 1 and 2 clinical trials un-derway that address various retinal pathologies with different sources of stem cells and disparate surgical approaches [59] Not only are the stem cell sources different their mode of action varies as well The first question these trials must answer is how safe stem cell therapy is in ocular therapy None of the animal transplantation studies de-tailed above showed a significant rate of teratoma formation upon transplantation of stem cells that are differentiated Conversely less differentiated stem cells that continue to express markers of plu-ripotency do form teratomata upon retinal trans-plantation [50 60]
The next question to answer is regarding the efficacy of a cellular therapy The current trials are using cell suspensions to either replace the RPE in AMD or rescue some retinal function through the paracrine release of growth factors While these approaches might be of value in milder forms of disease it is unlikely that they will show much functional improvement in severe pathology where PR cells and vision are lost For such pa-tients replacement of the neural retina will likely be necessary In many diseases of the neural reti-na and PRs there is a gradual decline of function that is attributed to the attritional effects of the buildup of toxic metabolites or damage to struc-tures pertinent to cell survival such as Bruchrsquos membrane [61] These factors need to be consid-
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 107
ered when developing strategies for cell replace-ment so that graft function is not compromised As such many groups developing potential stem cell therapy for AMD are investigating the use of a lsquopatch graftrsquo of HESC-RPE as a monolayer ad-hered to a membrane support [14 62] Trans-plantation of cells on a pre-prepared therapeutic patch will not only provide the structural support required for the survival of the RPE cells trans-plantation of cells as a monolayer will ensure that cells are fully differentiated polarized and func-tional This patch graft system may also serve as an ideal approach with which to transplant both the RPE and PR layers required for those with more advanced retinal degeneration
Small Molecule Induced Pluripotent Stem Cell Reprogramming and Retinal Induction
While further results are eagerly awaited from current and future studies of stem cells for retinal repair there have been extensive regulatory hur-dles to overcome when translating potential ther-apeutics from the laboratory to clinic Any meth-od to minimize this delay by avoiding the use of animal-derived factors and potentially oncogenic protocols is greatly welcomed
The use of IPSC-derived therapeutics heralds additional challenges associated with the original production of cells using viral reprogramming and in particular the use of the proto-oncogene c-Myc for the induction of pluripotency More re-cently IPSC have also been generated using dif-ferent delivery systems such as nonintegrative vi-ral vectors [63] and direct RNA [64] and protein transfer [65] Furthermore the use of small mol-ecules to replace the transcription factors origi-nally used may be preferable as they avoid the issues associated with transformation of the cells
With the consideration that one of the primary purposes of generating neural retinal cells from stem cells is for transplantation Osakada et al [66] analyzed the ability of small molecules to
drive neural retinal differentiation Dkk1 and Lefty the primary drivers of retinal differentia-tion in their previous protocol are produced in animals or bacteria The use of new animal prod-ucts in clinical trials adds a further layer of ethical and regulatory constraint to an already complex process Using the small molecules CK1ndash7 and SD-431542 in place of Dkk1 and Lefty in HESC and IPSC culture they were able to show down-regulation of genes that determine pluripotency Moreover these cells then began to express many of the proteins typical of PR precursors such as PAX6 RAX CRX and rhodopsin in appropriate sequence over 120 days circumventing the need for differentiation of retinal cells with animal de-rived products
Immunological Considerations of Stem Cell Therapy in Retinal Disease
Patient-specific IPSC therapy is theoretically less immunogenic than ESC therapy However inves-tigations by Zhao et al [67] have shown that un-differentiated IPSC injected into mice may be more immunogenic that ESC A potential expla-nation for this increased immune reaction is that the IPSC cells are constitutively expressing tran-scription factors that would otherwise be down-regulated However further research suggests that differentiated IPSC are less immunogenic [68] Other drawbacks of patient-specific IPSC include the cost and time delay as current proto-cols take weeks to months to derive stem cells and then differentiate to the desired tissue type An-other potential application of IPSC is to develop a bank of disparate human leucocyte antigen (HLA) types of undifferentiated cells ready to be directed towards the required tissue for HLA-matched pa-tients [69]
Prior to human transplantation a question to be addressed is the need for immunosuppression The retina has a degree of immune privilege in health conferred by the blood retinal barrier This
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

108 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
privilege is often compromised in disease states and would need to be breached to transplant cells to the neural retina Therefore immunosuppres-sion would likely be warranted and all its side ef-fects managed appropriately
Conclusion
In conclusion there is a huge need for therapies that reverse neural retinal degeneration Several protocols have been developed in recent years to generate cells with a neural retinal phenotype us-ing HESC and IPSC with varying efficiencies There are many similarities between the proto-cols such as the use of cellular aggregates and EBs and the modulation of the Wnt Shh Nodal and Notch pathways to name but a few Major hurdles to overcome include the need to generate cells
with full PR morphology deciding which trans-plantation method to adopt and deciphering what ontogenetic stage of developing PR will best integrate into the adult retina It will be exciting to see which of these protocols is first to make it to the clinical arena
Acknowledgements
This work was supported by funding from The London Project to Cure Blindness The Medical Research Council UK The Californian Institute of Regenerative Medicine Fight for Sight UK The Lincy Foundation The Macular Society and the National Institute for Health Research Biomedical Research Centre based at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology The views expressed are those of the au-thors and not necessarily those of the NHS the NIHR or the Department of Health
References
1 Jager RD Mieler WF Miller JW Age-related macular degeneration N Engl J Med 2008 358 2606ndash2617
2 de Jong PTVM Age-related macular de-generation N Engl J Med 2006 355
1474ndash1485 3 Walia S Fishman GA Natural history
of phenotypic changes in Stargardt mac-ular dystrophy Ophthalmic Genet 2009
30 63ndash68 4 Allikmets R Singh N Sun H Shroyer
NF Hutchinson A Chidambaram A et al A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mu-tated in recessive Stargardt macular dys-trophy Nat Genet 1997 15 236ndash246
5 Hartong DT Berson EL Dryja TP Reti-nitis pigmentosa Lancet 2006 368
1795ndash1809 6 Baglio SR Pegtel DM Baldini N Mes-
enchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy Front Physiol 2012 3
359 7 Ying Q-L Nichols J Evans EP Smith
AG Changing potency by spontaneous fusion Nature 2002 416 545ndash548
8 Schwartz SD Hubschman J-P Heilwell G Franco-Cardenas V Pan CK Ostrick RM et al Embryonic stem cell trials for macular degeneration a preliminary report Lancet 2012 379 713ndash720
9 McGill TJ Cottam B Lu B Wang S Gir-man S Tian C et al Transplantation of human central nervous system stem cells ndash neuroprotection in retinal degen-eration Eur J Neurosci 2012 35 468ndash477
10 Lund RD Wang S Lu B Girman S Holmes T Sauveacute Y et al Cells Isolated from umbilical cord tissue rescue photo-receptors and visual functions in a ro-dent model of retinal disease Stem Cells 2009 25 602ndash611
11 Siqueira RC Messias A Voltarelli JC Scott IU Jorge R Intravitreal injection of autologous bone marrow-derived mononuclear cells for hereditary retinal dystrophy a phase I trial Retina 2011
31 1207ndash1214
12 Park SS Caballero S Bauer G Shibata B Roth A Fitzgerald PG et al Long-term effects of intravitreal injection of GMP-grade bone-marrow-derived CD34+ cells in NOD-SCID mice with acute isch-emia-reperfusion injury Invest Oph-thalmol Vis Sci 2012 53 986ndash994
13 Kumar A Raj M Mochi TB Mohanty S Seth T Azad R Assessment of central retinal function after autologous bone marrow derived intravitreal stem cells injection in patients with retinitis pig-mentosa using multifocal ERG a pilot study World J Retin Vitreous 2012 2
5ndash13 14 Carr A-J Vugler AA Hikita ST Law-
rence JM Gias C Chen LL et al Protec-tive effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat PLoS One 2009 4e8152
15 Medeiros NE Curcio CA Preservation of ganglion cell layer neurons in age-related macular degeneration Invest Ophthalmol Vis Sci 2001 42 795ndash803
16 da Cruz L Chen FK Ahmado A Green-wood J Coffey P RPE transplantation and its role in retinal disease Prog Retin Eye Res 2007 26 598ndash635
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

Neural Retinal Regeneration with Pluripotent Stem Cells 109
17 Chen FK Patel PJ Uppal GS Rubin GS Coffey PJ Aylward GW et al A compar-ison of macular translocation with patch graft in neovascular age-related macular degeneration Invest Ophthalmol Vis Sci 2009 50 1848ndash1855
18 Bao ZZ Cepko CL The expression and function of Notch pathway genes in the developing rat eye J Neurosci 1997 17
1425ndash143419 Jadhav AP Roesch K Cepko CL Devel-
opment and neurogenic potential of Muumlller glial cells in the vertebrate retina Progr Retin Eye Res 2009 28 249ndash262
20 Chiang C Litingtung Y Lee E Young KE Corden JL Westphal H et al Cyclo-pia and defective axial patterning in mice lacking Sonic hedgehog gene func-tion Nature 1996 383 407ndash413
21 Fuhrmann S Wnt signaling in eye or-ganogenesis Organogenesis 2008 4 60ndash67
22 Tripathi BJ Tripathi RC Livingston AM Borisuth NS The role of growth factors in the embryogenesis and differ-entiation of the eye Am J Anat 1991
192 442ndash47123 Zhang SS-M Fu X-Y Barnstable CJ
Molecular aspects of vertebrate retinal development Mol Neurobiol 2002 26
137ndash15224 Hausman RE Ocular extracellular ma-
trices in development Prog Retin Eye Res 2007 26 162ndash188
25 OrsquoBrien KMB Schulte D Hendrickson AE Expression of photoreceptor-associ-ated molecules during human fetal eye development Mol Vis 2003 9 401ndash409
26 Tansley K The development of the rat eye in graft J Exp Biol 1946 22 221ndash224
27 Royo PE Quay WB Retinal transplanta-tion from fetal to maternal mammalian eye Growth 1959 23 313ndash336
28 Seiler MJ Aramant RB Cell replacement and visual restoration by retinal sheet transplants Prog Retin Eye Res 2012 31
661ndash68729 MacLaren RE Pearson RA MacNeil A
Douglas RH Salt TE Akimoto M et al Retinal repair by transplantation of pho-toreceptor precursors Nature 2006 444
203ndash20730 Pearson RA Barber AC Rizzi M Hip-
pert C Xue T West EL et al Restora-tion of vision after transplantation of photoreceptors Nature 2012 485 99ndash103
31 Gust J Reh TA Adult donor rod photo-receptors integrate into the mature mouse retina Invest Ophthalmol Vis Sci 2011 52 5266ndash5272
32 Humayun MS de Juan E del Cerro M Dagnelie G Radner W Sadda SR et al Human neural retinal transplantation Invest Ophthalmol Vis Sci 2000 41
3100ndash310633 Das T del Cerro M Jalali S Rao VS
Gullapalli VK Little C et al The trans-plantation of human fetal neuroretinal cells in advanced retinitis pigmentosa patients results of a long-term safety study Exp Neurol 1999 157 58ndash68
34 Klassen HJ Ng TF Kurimoto Y Kirov I Shatos M Coffey P et al Multipotent retinal progenitors express developmen-tal markers differentiate into retinal neurons and preserve light-mediated behavior Invest Ophthalmol Vis Sci 2004 45 4167ndash4173
35 Thomson JA Embryonic stem cell lines derived from human blastocysts Sci-ence 1998 282 1145ndash1147
36 Singhal S Bhatia B Jayaram H Becker S Jones MF Cottrill PB et al Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation Stem Cells Transl Med 2012 1 188ndash199
37 Takahashi K Yamanaka S Induction of pluripotent stem cells from mouse em-bryonic and adult fibroblast cultures by defined factors Cell 2006 126 663ndash676
38 Ying Q-L Smith AG Defined conditions for neural commitment and differentia-tion Meth Enzymol 2003 365 327ndash341
39 Ikeda H Osakada F Watanabe K Mizuseki K Haraguchi T Miyoshi H et al Generation of Rx+Pax6+ neural reti-nal precursors from embryonic stem cells Proc Natl Acad Sci USA 2005 102
11331ndash1133640 Osakada F Ikeda H Mandai M Wataya
T Watanabe K Yoshimura N et al To-ward the generation of rod and cone photoreceptors from mouse monkey and human embryonic stem cells Nat Biotechnol 2008 26 215ndash224
41 Hirami Y Osakada F Takahashi K Oki-ta K Yamanaka S Ikeda H et al Gen-eration of retinal cells from mouse and human induced pluripotent stem cells Neurosci Lett 2009 458 126ndash131
42 Pera EM Wessely O Li SY De Robertis EM Neural and head induction by insu-lin-like growth factor signals Dev Cell 2001 1 655ndash665
43 Lamba DA Karl MO Ware CB Reh TA Efficient generation of retinal progenitor cells from human embryonic stem cells Proc Natl Acad Sci USA 2006 103
12769ndash1277444 Lamba DA Gust J Reh TA Transplan-
tation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice Cell Stem Cell 2009 4 73ndash79
45 Banin E Obolensky A Idelson M Hemo I Reinhardtz E Pikarsky E et al Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells Stem Cells 2006
24 246ndash25746 Parameswaran S Balasubramanian S
Babai N Qiu F Eudy JD Thoreson WB et al Induced pluripotent stem cells generate both retinal ganglion cells and photoreceptors therapeutic implica-tions in degenerative changes in glau-coma and age-related macular degen-eration Stem Cells 2010 28 695ndash703
47 Meyer JS Shearer RL Capowski EE Wright LS Wallace KA McMillan EL et al Modeling early retinal development with human embryonic and induced pluripotent stem cells Proc Natl Acad Sci USA 2009 106 16698ndash16703
48 Meyer JS Howden SE Wallace KA Ver-hoeven AD Wright LS Capowski EE et al Optic vesicle-like structures derived from human pluripotent stem cells fa-cilitate a customized approach to retinal disease treatment Stem Cells 2011 29
1206ndash121849 Phillips MJ Wallace KA Dickerson SJ
Miller MJ Verhoeven AD Martin JM et al Blood-derived human iPS cells gener-ate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses Invest Ophthalmol Vis Sci 2012 53 2007ndash2019
50 Tucker BA Park I-H Qi SD Klassen HJ Jiang C Yao J et al Transplantation of adult mouse iPS cell-derived photore-ceptor precursors restores retinal struc-ture and function in degenerative mice PLoS One 2011 6e18992
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72

110 Ramsden middot Powner middot Carr middot Smart middot da Cruz middot Coffey
51 Hambright D Park K-Y Brooks M McKay R Swaroop A Nasonkin I Long-term survival and differentiation of reti-nal neurons derived from human em-bryonic stem cell lines in un-immunosuppressed mouse retina Mol Vis 2012 18 920
52 Mellough CB Sernagor E Moreno-Gi-meno I Steel DHW Lako M Efficient stage-specific differentiation of human pluripotent stem cells toward retinal photoreceptor cells Stem Cells 2012 30
673ndash68653 Eiraku M Takata N Ishibashi H Kawa-
da M Sakakura E Okuda S et al Self-organizing optic-cup morphogenesis in three-dimensional culture Nature 2011
472 51ndash5654 Eiraku M Sasai Y Mouse embryonic
stem cell culture for generation of three-dimensional retinal and cortical tissues Nat Protoc 2012 7 69ndash79
55 Nakano T Ando S Takata N Kawada M Muguruma K Sekiguchi K et al Self-formation of optic cups and stor-able stratified neural retina from human ESCs Cell Stem Cell 2012 10 771ndash785
56 Garita-Hernaacutendez M Diaz-Corrales F Lukovic D Guede IG Lloret AD Saacuten-chez MLV et al Hypoxia increases the yield of photoreceptors differentiating from mouse embryonic stem cells and improves the modeling of retinogenesis in vitro Stem Cells 2013 31 966ndash978
57 Boucherie C Mukherjee S Henckaerts E Thrasher AJ Sowden JC Ali RR Brief report self-organizing neuroepithelium from human pluripotent stem cells fa-cilitates derivation of photoreceptors Stem Cells 2013 31 408ndash414
58 Ramsden CM Powner MB Carr A-JF Smart MJK da Cruz L Coffey PJ Stem cells in retinal regeneration past pres-ent and future Development 2013 140
2576ndash2585 59 Jin Z-B Okamoto S Osakada F Homma
K Assawachananont J Hirami Y et al Modeling retinal degeneration using patient-specific induced pluripotent stem cells PLoS One 2011 6e17084
60 Arnhold S Klein H Semkova I Addicks K Schraermeyer U Neurally selected embryonic stem cells induce tumor for-mation after long-term survival follow-ing engraftment into the subretinal space Invest Ophthalmol Vis Sci 2004
45 4251ndash425561 Gullapalli VK Sugino IK Van Patten Y
Shah S Zarbin MA Impaired RPE sur-vival on aged submacular human Bruchrsquos membrane Exp Eye Res 2005
80 235ndash24862 Vugler A Carr A-J Lawrence J Chen
LL Burrell K Wright A et al Elucidat-ing the phenomenon of HESC-derived RPE anatomy of cell genesis expansion and retinal transplantation Exp Neurol 2008 214 347ndash361
63 Fusaki N Ban H Nishiyama A Saeki K Hasegawa M Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai vi-rus an RNA virus that does not inte-grate into the host genome Proc Jpn Acad Ser B Phys Biol Sci 2009 85 348ndash362
64 Warren L Manos PD Ahfeldt T Loh Y-H Li H Lau F et al Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA Cell Stem Cell 2010 7 618ndash630
65 Kim D Kim C-H Moon J-I Chung Y-G Chang M-Y Han B-S et al Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins Cell Stem Cell 2009 4 472ndash476
66 Osakada F Jin Z-B Hirami Y Ikeda H Danjyo T Watanabe K et al In vitro differentiation of retinal cells from hu-man pluripotent stem cells by small-molecule induction J Cell Sci 2009 122
3169ndash317967 Zhao T Zhang Z-N Rong Z Xu Y Im-
munogenicity of induced pluripotent stem cells Nature 2011 474 212ndash215
68 Guha P Morgan JW Mostoslavsky G Rodrigues NP Boyd AS Lack of im-mune response to differentiated cells derived from syngeneic induced plurip-otent stem cells Cell Stem Cell 2013 12
407ndash41269 Zimmermann A Preynat-Seauve O
Tiercy J-M Krause K-H Villard J Hap-lotype-based banking of human pluripo-tent stem cells for transplantation po-tential and limitations Stem Cells Dev 2012 21 2364ndash2373
70 Bottenstein JE Skaper SD Varon SS Sato GH Selective survival of neurons from chick embryo sensory ganglionic dissociates utilizing serum-free supple-mented medium Exp Cell Res 1980 125
183ndash19071 Brewer GJ Torricelli JR Evege EK Price
PJ Optimized survival of hippocampal neurons in B27-supplemented Neuro-basal a new serum-free medium combi-nation J Neurosci Res 1993 35 567ndash576
72 Phillips MJ Otteson DC Sherry DM Progression of neuronal and synaptic remodeling in the rd10 mouse model of retinitis pigmentosa J Comp Neurol 2010 518 2071ndash2089
Conor M RamsdenThe London Project to Cure Blindness Division of ORBITInstitute of Ophthalmology University College London11ndash43 Bath StreetLondon EC1V 9EL (UK)E-Mail conorramsden09uclacuk
Casaroli-Marano RP Zarbin MA (eds) Cell-Based Therapy for Retinal Degenerative DiseaseDev Ophthalmol Basel Karger 2014 vol 53 pp 97ndash110 (DOI 101159000357363) D
ownl
oade
d by
U
CL
14
482
134
189
- 4
25
2014
33
451
PM
- CitRef_1
- CitRef_2
- CitRef_3
- CitRef_4
- CitRef_5
- CitRef_6
- CitRef_7
- CitRef_9
- CitRef_8
- CitRef_10
- CitRef_11
- CitRef_12
- CitRef_14
- CitRef_15
- CitRef_16
- CitRef_17
- CitRef_18
- CitRef_19
- CitRef_20
- CitRef_21
- CitRef_22
- CitRef_23
- CitRef_24
- CitRef_25
- CitRef_26
- CitRef_27
- CitRef_28
- CitRef_29
- CitRef_30
- CitRef_31
- CitRef_32
- CitRef_33
- CitRef_34
- CitRef_35
- CitRef_36
- CitRef_37
- CitRef_38
- CitRef_39
- CitRef_40
- CitRef_41
- CitRef_42
- CitRef_43
- CitRef_44
- CitRef_45
- CitRef_46
- CitRef_47
- CitRef_48
- CitRef_49
- CitRef_50
- CitRef_51
- CitRef_52
- CitRef_53
- CitRef_54
- CitRef_55
- CitRef_56
- CitRef_57
- CitRef_58
- CitRef_59
- CitRef_60
- CitRef_61
- CitRef_62
- CitRef_63
- CitRef_64
- CitRef_65
- CitRef_66
- CitRef_67
- CitRef_68
- CitRef_69
- CitRef_70
- CitRef_71
- CitRef_72
Related Documents











