Neuroscience Letters www.sciencedirect.com Available online at Volume 416 No. 2, 12 April 2007 (Cited in Biological Abstracts (BIOSIS), Chemical Abstracts, Elsevier BIOBASE/Current Awareness in Biological Sciences, Current Contents/Life Sciences, EMBASE/Excerpta Medica, Index Medicus, PASCAL M and Science Citation Index). Also covered in the abstract and citation database SCOPUS ® .Full text available in ScienceDirect ® . CONTENTS Auditory stimulation affects renal sympathetic nerve activity and blood pressure in rats Takuo Nakamura, Mamoru Tanida, Akira Niijima, Hiroshi Hibino, Jiao Shen, Katsuya Nagai (Japan) 107 Case–control association study of the 2,3-cyclic nucleotide 3-phosphodiesterase (CNP) gene and schizophrenia in the Han Chinese population Fulei Tang, Mei Qu, Lifang Wang, Yan Ruan, Tianlan Lu, Handi Zhang, Zhonghua Liu, Weihua Yue, Dai Zhang (PR China) 113 Differential after-effects of bimanual activity on mirror movements Alistair N. Vardy, Andreas Daffertshofer, Arne Ridderikhoff, Peter J. Beek (The Netherlands) 117 The peripheral role of group I metabotropic glutamate receptors on nociceptive behaviors in rats with knee joint inflammation Kyu Sang Lee, Junesun Kim, Young Wook Yoon, Min-Goo Lee, Seung Kil Hong, Hee Chul Han (South Korea) 123 Fos-like immunoreactivity in rat dorsal raphe nuclei induced by alkaloid extract of Mitragyna speciosa Ekkasit Kumarnsit, Uraporn Vongvatcharanon, Niwat Keawpradub, Pranom Intasaro (Thailand) 128 Extensive training in a maze task reduces neurogenesis in the adult rat dentate gyrus probably as a result of stress E. Aztiria, G. Capodieci, L. Arancio, G. Leanza (Italy) 133 Neural coupling between the upper and lower limbs in humans Maria Knikou (USA) 138 mHGTD-P mediates hypoxic neuronal cell death via the release of apoptosis-inducing factor Young-Eun Cho, Jeong-Hun Ko, Yong-Jun Kim, Ji-Hye Yim, Su-Mi Kim, Jae-Hoon Park (Republic of Korea) 144 Intracerebroventricular injection of losartan inhibits angiotensin II-sensitive neurons via GABA inputs in the anterior hypothalamic area of rats Yukihiko Hagiwara, Takao Kubo (Japan) 150 Nocistatin and its derivatives antagonize the impairment of short-term acquisition induced by nociceptin Eugene Hern C. Liu, Tat-Leang Lee, Yuji Nishiuchi, Terutoshi Kimura, Shinro Tachibana (Singapore, Japan) 155 Ethanol withdrawal posttranslationally decreases the activity of cytochrome c oxidase in an estrogen reversible manner Marianna E. Jung, Rajnee Agarwal, James W. Simpkins (USA) 160 The neurobehavioral benefit conferred by a single systemic administration of 8-OH-DPAT after brain trauma is confined to a nar- row therapeutic window Jeffrey P. Cheng, Haris A. Aslam, Ann N. Hoffman, Ross D. Zafonte, Anthony E. Kline (USA) 165 SB203580 enhances interleukin-1 receptor antagonist gene expression in IFN--stimulated BV2 microglial cells through a com- posite nuclear factor-B/PU.1 binding site Jin-Sun Park, Soo-Hyun Jung, Hyemyung Seo, Hee-Sun Kim (South Korea) 169 Morphological changes in the trigemino-rubral pathway in dystrophic (mdx) mice Magali Luci Pinto, Heloisa Helena Vieira Olyntho Tokunaga, Caden Souccar, Guus H.M. Schoorlemmer, Rita de Cássia Ribeiro da Silva Lapa (Brazil) 175 ABCA1 polymorphisms and Alzheimer’s disease Fabienne Wavrant-De Vrièze, Danielle Compton, Meridith Womick, Sampath Arepalli, Omanma Adighibe, Ling Li, Jordi Pérez- Tur, John Hardy (USA, Spain) 180 Feeding and systemic D-amphetamine increase extracellular acetylcholine in the medial thalamus: A possible reward enabling func- tion P. Rada, L. Hernandez, B.G. Hoebel (Venezuela, USA) 184 Bradycardia elicited by microinjections of nociceptin/orphanin FQ into the intermediolateral cell column at T1–T2 in the rat Vineet C. Chitravanshi, Hreday N. Sapru (USA) 188 In vivo effects of leptin on autonomic nerve activity and lipolysis in rats Jiao Shen, Mamoru Tanida, Akira Niijima, Katsuya Nagai (Japan) 193 Central administration of glucagon suppresses food intake in chicks Kazuhisa Honda, Hiroshi Kamisoyama, Noboru Saito, Yohei Kurose, Kunio Sugahara, Shin Hasegawa (Japan) 198 GABA-immunoreactive photoreceptors in the retina of an anuran, Pelobates fuscus Dávid A. Schäffer, Robert Gábriel (Hungary) 202 Apoptosis induces neuronal apolipoprotein-E synthesis and localization in apoptotic bodies David A. Elliott, Woojin S. Kim, David A. Jans, Brett Garner (Australia) 206

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN 0304-3940
EL
SEV
IER
NE
UR
OS
CIE
NC
E L
ET
TE
RS
Vol. 416/2 (2007) 107–210
416/2NeuroscienceLetters
www.sciencedirect.com
Available online at
Volume 416 No. 2, 12 April 2007(Cited in Biological Abstracts (BIOSIS), Chemical Abstracts, Elsevier BIOBASE/Current Awareness in Biological Sciences, Current Contents/Life
Sciences, EMBASE/Excerpta Medica, Index Medicus, PASCAL M and Science Citation Index). Also covered in the abstract and citation database SCOPUS ®.Full text available in ScienceDirect®.
CONTENTS
Auditory stimulation affects renal sympathetic nerve activity and blood pressure in ratsTakuo Nakamura, Mamoru Tanida, Akira Niijima, Hiroshi Hibino, Jiao Shen, Katsuya Nagai (Japan) 107
Case–control association study of the 2�,3�-cyclic nucleotide 3�-phosphodiesterase (CNP) gene and schizophrenia in the HanChinese population
Fulei Tang, Mei Qu, Lifang Wang, Yan Ruan, Tianlan Lu, Handi Zhang, Zhonghua Liu, Weihua Yue, Dai Zhang (PR China) 113Differential after-effects of bimanual activity on mirror movements
Alistair N. Vardy, Andreas Daffertshofer, Arne Ridderikhoff, Peter J. Beek (The Netherlands) 117The peripheral role of group I metabotropic glutamate receptors on nociceptive behaviors in rats with knee joint inflammation
Kyu Sang Lee, Junesun Kim, Young Wook Yoon, Min-Goo Lee, Seung Kil Hong, Hee Chul Han (South Korea) 123Fos-like immunoreactivity in rat dorsal raphe nuclei induced by alkaloid extract of Mitragyna speciosa
Ekkasit Kumarnsit, Uraporn Vongvatcharanon, Niwat Keawpradub, Pranom Intasaro (Thailand) 128Extensive training in a maze task reduces neurogenesis in the adult rat dentate gyrus probably as a result of stress
E. Aztiria, G. Capodieci, L. Arancio, G. Leanza (Italy) 133Neural coupling between the upper and lower limbs in humans
Maria Knikou (USA) 138mHGTD-P mediates hypoxic neuronal cell death via the release of apoptosis-inducing factor
Young-Eun Cho, Jeong-Hun Ko, Yong-Jun Kim, Ji-Hye Yim, Su-Mi Kim, Jae-Hoon Park (Republic of Korea) 144Intracerebroventricular injection of losartan inhibits angiotensin II-sensitive neurons via GABA inputs in the anterior hypothalamicarea of rats
Yukihiko Hagiwara, Takao Kubo (Japan) 150Nocistatin and its derivatives antagonize the impairment of short-term acquisition induced by nociceptin
Eugene Hern C. Liu, Tat-Leang Lee, Yuji Nishiuchi, Terutoshi Kimura, Shinro Tachibana (Singapore, Japan) 155Ethanol withdrawal posttranslationally decreases the activity of cytochrome c oxidase in an estrogen reversible manner
Marianna E. Jung, Rajnee Agarwal, James W. Simpkins (USA) 160The neurobehavioral benefit conferred by a single systemic administration of 8-OH-DPAT after brain trauma is confined to a nar-row therapeutic window
Jeffrey P. Cheng, Haris A. Aslam, Ann N. Hoffman, Ross D. Zafonte, Anthony E. Kline (USA) 165SB203580 enhances interleukin-1 receptor antagonist gene expression in IFN-�-stimulated BV2 microglial cells through a com-posite nuclear factor-�B/PU.1 binding site
Jin-Sun Park, Soo-Hyun Jung, Hyemyung Seo, Hee-Sun Kim (South Korea) 169Morphological changes in the trigemino-rubral pathway in dystrophic (mdx) mice
Magali Luci Pinto, Heloisa Helena Vieira Olyntho Tokunaga, Caden Souccar, Guus H.M. Schoorlemmer, Rita de Cássia Ribeiroda Silva Lapa (Brazil) 175
ABCA1 polymorphisms and Alzheimer’s diseaseFabienne Wavrant-De Vrièze, Danielle Compton, Meridith Womick, Sampath Arepalli, Omanma Adighibe, Ling Li, Jordi Pérez-Tur, John Hardy (USA, Spain) 180
Feeding and systemic D-amphetamine increase extracellular acetylcholine in the medial thalamus: A possible reward enabling func-tion
P. Rada, L. Hernandez, B.G. Hoebel (Venezuela, USA) 184Bradycardia elicited by microinjections of nociceptin/orphanin FQ into the intermediolateral cell column at T1–T2 in the rat
Vineet C. Chitravanshi, Hreday N. Sapru (USA) 188In vivo effects of leptin on autonomic nerve activity and lipolysis in rats
Jiao Shen, Mamoru Tanida, Akira Niijima, Katsuya Nagai (Japan) 193Central administration of glucagon suppresses food intake in chicks
Kazuhisa Honda, Hiroshi Kamisoyama, Noboru Saito, Yohei Kurose, Kunio Sugahara, Shin Hasegawa (Japan) 198GABA-immunoreactive photoreceptors in the retina of an anuran, Pelobates fuscus
Dávid A. Schäffer, Robert Gábriel (Hungary) 202Apoptosis induces neuronal apolipoprotein-E synthesis and localization in apoptotic bodies
David A. Elliott, Woojin S. Kim, David A. Jans, Brett Garner (Australia) 206
Volume 416 No. 2, 12 April 2007
NSL OFC 416(2).qxd:OUTER COVER 3/20/07 8:27 AM Page 1
MARIA
Highlight
MARIA
Highlight

A
aamofIMtr©
K
Lea[otcar[
moira[
DI
m
0d
Neuroscience Letters 416 (2007) 138–143
Neural coupling between the upper and lower limbs in humans
Maria Knikou a,b,∗a Department of Physical Therapy & Neuroscience, The Graduate Center, City University of New York, Staten Island, NY 10314, USA
b Rehabilitation Institute of Chicago & Department of Physical Medicine & Rehabilitation, Northwestern University, Chicago, IL 60611, USA
Received 8 January 2007; received in revised form 27 January 2007; accepted 31 January 2007
bstract
The aim of this study was to establish the effects of active sinusoidal ipsilateral and contralateral upper limb flexion, extension, abduction, anddduction with elbows extended on the right soleus H-reflex with subjects seated and standing. Reflex effects were also established when bothrms moved synchronously in a reciprocal pattern with elbows flexed in seated and standing subjects. Sinusoidal arm movements were timed to aetronome and performed at 0.2 Hz. Soleus H-reflexes were elicited only once (every 4 s) in every movement cycle of the upper limbs. Position
f arms, and activity of shoulder muscles were recorded through twin-axis goniometers and surface electromyography (EMG), respectively. Weound that in seated subjects, regardless the direction of the active movement or the upper limb being moved, the soleus H-reflex was depressed.n standing subjects, a reflex depression was observed during extension, abduction, and adduction of the ipsilateral and contralateral upper limbs.
uscles were active during arm flexion and abduction in all directions of arm movement with subjects either seated or standing. It is suggestedhat arm movement might be incorporated in the rehabilitation training of people with a supraspinal or spinal cord lesion, since it can benefit motorecovery by decreasing spinal reflex excitability of the legs in these patients.
2007 Elsevier Ireland Ltd. All rights reserved.
n; Pos
tru
Aaraarwwsr
eywords: Arm movement; H-reflex; Crossed interlimb connections; Inhibitio
ong propriospinal neurons (LPNs) originate in the cervical (C)nlargement (C3-C5), travel in the ventral and lateral funiculi,nd terminate in the ventral horn of the lumbosacral enlargement1,2,12,17]. These neurons are involved in the neural couplingf arms and legs during rhythmical movements [13], and con-ribute to limb coordination during locomotion. LPNs couple theervical and lumbar enlargements in lower vertebrates [18,20]nd in humans [19], through connections with alpha motoneu-ons and inhibitory interneurons projecting to motoneurons13].
Coordination of the arms and legs during rhythmic move-ents is evident in humans [8,21]. Cutaneous and H-reflexes
f the upper limb muscles during arm cycling are modulatedn a phase and task dependent manner [22,24], with cutaneous
eflex reversals to be present [23], corresponding to the phasend task dependency of the leg reflexes during human walking4,26]. These findings suggest of similar control mechanisms in∗ Correspondence address: City University of New York, The Graduate Center,epartment of Physical Therapy, 2800 Victory Blvd., Bldg. 5N-207, Staten
sland, NY 10314, USA. Tel.: +1 718 982 3316; fax: +1 718 982 2984.E-mail addresses: [email protected],
saumpTidt
304-3940/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.neulet.2007.01.072
ture
he reflex modulation of upper and lower limbs, which might beepresented by a system of two coupled oscillators like the onenderlying quadruped locomotion [7,21].
Changes in static positioning of the shoulder modulate thechilles, quadriceps, and biceps femoris tendon reflexes as well
s the soleus H-reflex [5,6]. The effects were expressed in aeciprocal pattern, e.g. ipsilateral arm flexion and contralateralrm extension facilitated the Achilles, the quadriceps tendonnd soleus H-reflexes and depressed the biceps femoris tendoneflex. The modulation pattern of these reflexes was reversedhen the position of the ipsilateral and contralateral shoulderas switched. In contrast, in seated subjects the soleus H-reflex
ize is decreased during bilateral arm cycling when compared toeflexes recorded with the arms held statically in flexion or exten-ion [9]. Although it is apparent that interlimb reflex connectionsre present in humans and affect motor output promoting humanpright posture and balance, it is currently not known if activeovement of both upper limbs in various planes at different body
ostures affect differently lower limb spinal reflex excitability.
hus, the objectives of this study were to establish whether activepsilateral and/or contralateral upper limb movement in differentirections and active movement of both arms in a reciprocal pat-ern (mimicking the movement pattern observed during human

e Lett
wh
(ttwdaad
itmsmwwtce
maebwmte
ljmtaaarwtTdwo
dMmbatItrH
tm2rectatdadws
toemceTsmim
tMwrmtsctdam
ssiwfltcn
iaI
M. Knikou / Neuroscienc
alking) affect the soleus H-reflex size in seated and standinguman subjects.
Fourteen subjects (six men and eight women) aged 23–4129 ± 5) years participated in the study. The experimental pro-ocol was approved by the Institutional Review Board (IRB) ofhe College of Staten Island (IRB No. 48-N-06), all proceduresere conducted in accordance with the ethical standards laidown in the 1964 Declaration of Helsinki, and each subject gavewritten consent before testing. None of the subjects reported
ny neurological deficit, low back pain, or other musculoskeletalisorder.
In all subjects, the H-reflex was elicited and recorded accord-ng to procedures described previously in detail [14,15]. Briefly,he H-reflex was elicited on the ascending part of the recruit-
ent curve by stimulation of the posterior tibial nerve using aingle pulse of 1 ms, the reflexes were 15–30% of the maxi-al M-wave (Mmax), and the corresponding M-waves (whichere monitored online) were below 5% of the Mmax. H-reflexesere recorded from the right soleus muscle via a single differen-
ial electrode (Delsys Inc., Boston, USA) having established theorrect site of stimulation by using a stainless steel monopolarlectrode as a probe (see more details in Refs. [14,15]).
The ipsilateral and the contralateral active upper limb move-ent included flexion/extension (0–90◦) in the sagittal plane,
nd abduction/adduction (0–90◦) in the frontal plane withlbows extended. In addition, alternated flexion/extension ofoth upper limbs in a reciprocal pattern with elbows flexedas tested. The frequency of the movement was set by a digitaletronome and was equivalent to 0.2 Hz. The subject matched
he sound of the metronome with the movement of the arm, whileach movement cycle lasted 4 s.
Soleus H-reflexes during ipsilateral and contralateral upperimb movement (conditioned reflexes) were recorded with sub-ects seated and standing, and were elicited only once in every
ovement cycle of the upper limbs. The stimulus to the pos-erior tibial nerve was sent at the beginning of the sinusoidalctive arm movement (see top traces of Fig. 3). In each subject,t least three control reflexes for each testing condition (seatednd standing) were randomly recorded with the conditionedeflexes. For every control and conditioned reflex 15 sweepsere recorded. M-wave amplitudes were continually monitored
o ensure stability in the stimulation and recording procedures.he M-wave size was used for screening conditioned H-reflexesuring active movement of the upper limbs. Experimental dataere rejected when a significant difference between the M-wavef the conditioned and the control reflexes was established.
Surface electromyograms (EMG) were recorded via singleifferential electrodes (Bagnoli System, Delsys Inc., Boston,A) from the ipsilateral and contralateral anterior deltoid (AD),iddle deltoid (MD), posterior deltoid (PD), triceps (Tric), and
iceps brachii (Bic) muscles following standard procedures. Therm position during each sinusoidal movement was recordedhrough twin-axis goniometers (World Precision Instruments
nc., FL, USA). All signals were amplified and band-pass fil-ered (10–500 Hz) before being sampled at 2 kHz (1401 plusunning Spike 2 software, Cambridge Electronics Design Ltd.,ertfordshire, England, UK).asir
ers 416 (2007) 138–143 139
The soleus M-wave and H-reflex of the control and condi-ioned reflexes were full-wave rectified and their sizes were
easured as the area under the respective waveforms (Spike, CED Ltd., UK) [15]. For each subject, the conditionedeflexes (all 15 sweeps) recorded during active movement ofither upper limb in all directions were expressed as a per-entage of the mean size of the control reflex collected withhe subject seated. A one-way analysis of variance (ANOVA)nd post hoc Bonferroni tests were applied to the experimen-al data sets for each subject to establish statistically significantifferences between the control and the conditioned H-reflexesnd across the conditioning trials. This analysis was also con-ucted for conditioned reflexes recorded with subjects standing,hile in this case the control reflex was recorded with subjects
tanding.The mean size of each subject’s conditioned H-reflex was
hen grouped based on the upper limb being moved (ipsilateralr contralateral) and the direction of the movement (flexion,xtension, abduction, and adduction). An ANOVA for repeatedeasures (2 × 4) was applied to the data sets to determine
hanges in the size of the conditioned reflexes across differ-nt experimental conditions investigated for the subject group.his analysis was performed separately for tests conducted withubjects seated and standing. Lastly, an ANOVA for repeatedeasures (2 × 4) was also applied to the conditioned reflexes
n order to establish whether the conditioned H-reflexes wereodulated differently with subjects seated and standing.For each subject, the M-waves of the control and condi-
ioned H-reflexes were expressed as a percentage of the maximal-wave. A one-way ANOVA with post hoc Bonferroni testsas used to test for differences between the M-waves of the
eflexes recorded under control conditions and during upper limbovement. When significant differences were encountered the
rial was rejected. Each EMG recording was full-wave rectified,moothed, and averaged over the 15 consecutive arm movementycles. Then, the average EMG signal for each muscle was plot-ed as a function of time. In all tests, statistically significantifferences were established at 95% of confidence level. Resultsre presented as mean values along with the standard error ofean (S.E.M.).Ipsilateral and contralateral upper limb movement induced
ignificant changes in the magnitude of the soleus H-reflex withubjects seated. Representative examples of these effects arellustrated for two subjects in Fig. 1. The right soleus H-reflexas significantly depressed when either upper limb moved inexion, extension, abduction, and adduction. The reduction in
he H-reflex occurred without any apparent changes in M-waveharacteristics demonstrating that the stimulus conditions wereot altered.
The population data for the conditioned H-reflex followingpsilateral and contralateral upper limb movement in the frontalnd sagittal planes with subjects seated are illustrated in Fig. 2A.psilateral arm flexion and extension as well as abduction and
dduction with elbows extended resulted in a significant depres-ion of the soleus H-reflex across subjects with the reflex reach-ng overall amplitudes that ranged from 80 to 85% of controleflex values (P < 0.05). Similarly, the soleus H-reflex was signif-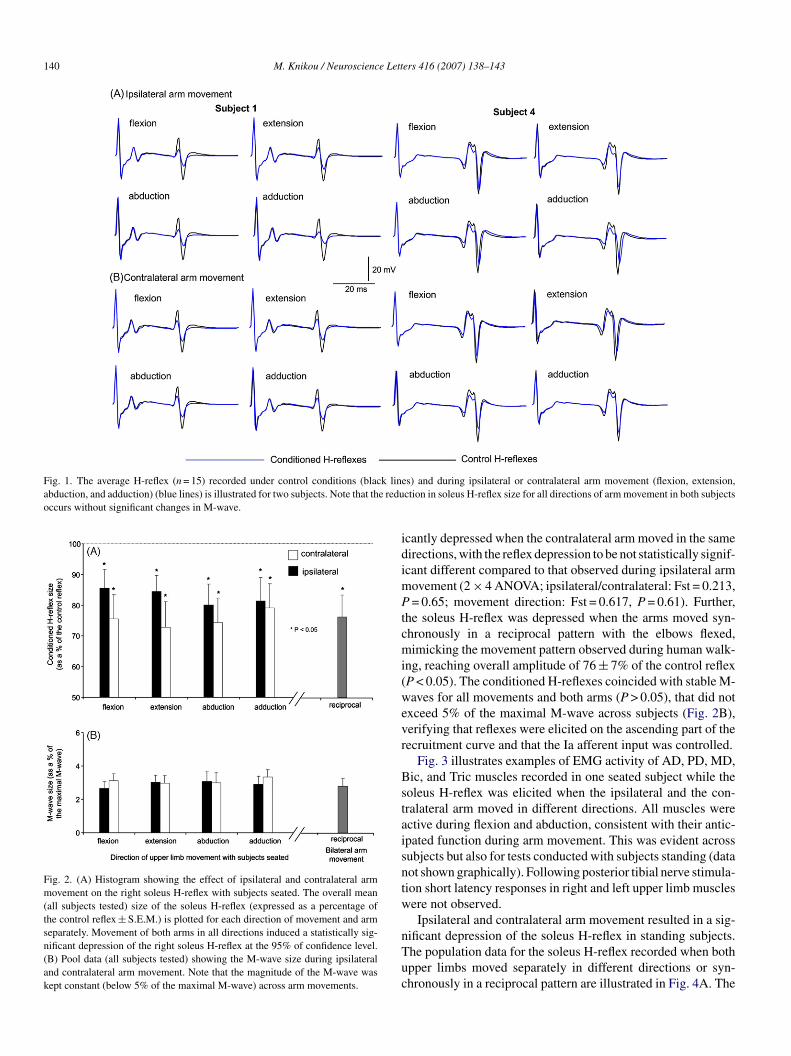
140 M. Knikou / Neuroscience Letters 416 (2007) 138–143
Fig. 1. The average H-reflex (n = 15) recorded under control conditions (black linabduction, and adduction) (blue lines) is illustrated for two subjects. Note that the reduoccurs without significant changes in M-wave.
Fig. 2. (A) Histogram showing the effect of ipsilateral and contralateral armmovement on the right soleus H-reflex with subjects seated. The overall mean(all subjects tested) size of the soleus H-reflex (expressed as a percentage ofthe control reflex ± S.E.M.) is plotted for each direction of movement and armseparately. Movement of both arms in all directions induced a statistically sig-nificant depression of the right soleus H-reflex at the 95% of confidence level.(B) Pool data (all subjects tested) showing the M-wave size during ipsilateraland contralateral arm movement. Note that the magnitude of the M-wave waskept constant (below 5% of the maximal M-wave) across arm movements.
idimPtcmi(wevr
Bstaisntw
nTuc
es) and during ipsilateral or contralateral arm movement (flexion, extension,ction in soleus H-reflex size for all directions of arm movement in both subjects
cantly depressed when the contralateral arm moved in the sameirections, with the reflex depression to be not statistically signif-cant different compared to that observed during ipsilateral arm
ovement (2 × 4 ANOVA; ipsilateral/contralateral: Fst = 0.213,= 0.65; movement direction: Fst = 0.617, P = 0.61). Further,
he soleus H-reflex was depressed when the arms moved syn-hronously in a reciprocal pattern with the elbows flexed,imicking the movement pattern observed during human walk-
ng, reaching overall amplitude of 76 ± 7% of the control reflexP < 0.05). The conditioned H-reflexes coincided with stable M-aves for all movements and both arms (P > 0.05), that did not
xceed 5% of the maximal M-wave across subjects (Fig. 2B),erifying that reflexes were elicited on the ascending part of theecruitment curve and that the Ia afferent input was controlled.
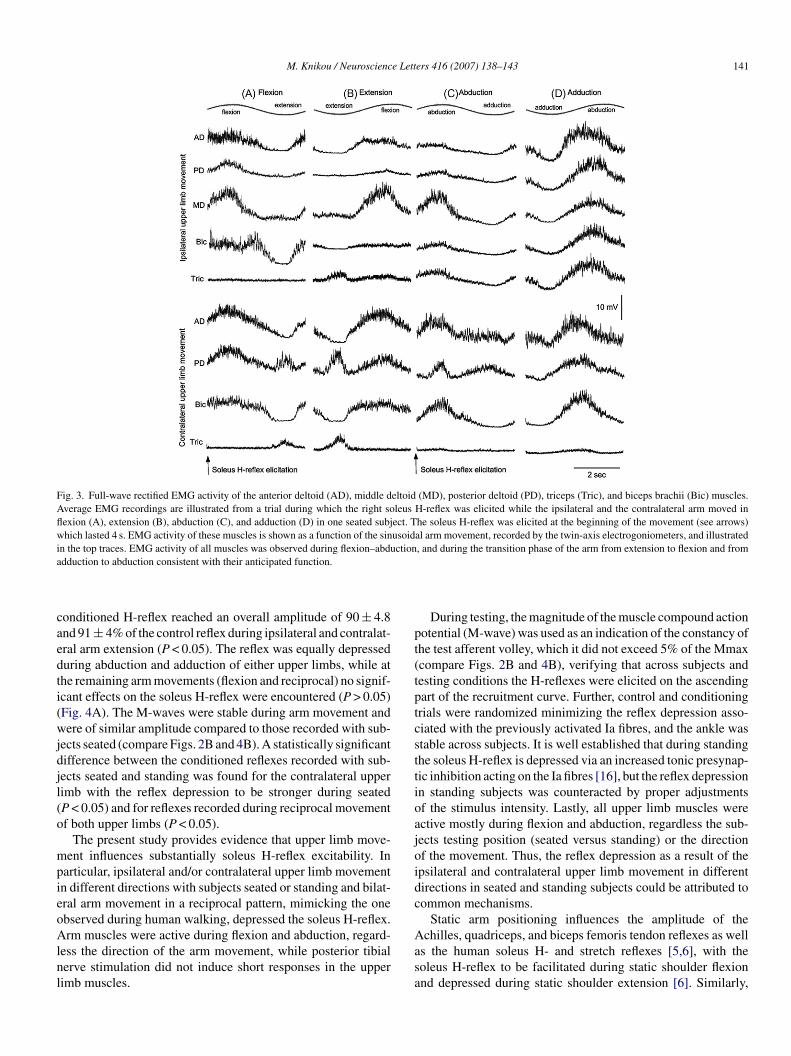
Fig. 3 illustrates examples of EMG activity of AD, PD, MD,ic, and Tric muscles recorded in one seated subject while the
oleus H-reflex was elicited when the ipsilateral and the con-ralateral arm moved in different directions. All muscles werective during flexion and abduction, consistent with their antic-pated function during arm movement. This was evident acrossubjects but also for tests conducted with subjects standing (dataot shown graphically). Following posterior tibial nerve stimula-ion short latency responses in right and left upper limb musclesere not observed.Ipsilateral and contralateral arm movement resulted in a sig-
ificant depression of the soleus H-reflex in standing subjects.he population data for the soleus H-reflex recorded when bothpper limbs moved separately in different directions or syn-hronously in a reciprocal pattern are illustrated in Fig. 4A. The
M. Knikou / Neuroscience Letters 416 (2007) 138–143 141
Fig. 3. Full-wave rectified EMG activity of the anterior deltoid (AD), middle deltoid (MD), posterior deltoid (PD), triceps (Tric), and biceps brachii (Bic) muscles.Average EMG recordings are illustrated from a trial during which the right soleus H-reflex was elicited while the ipsilateral and the contralateral arm moved inflexion (A), extension (B), abduction (C), and adduction (D) in one seated subject. The soleus H-reflex was elicited at the beginning of the movement (see arrows)which lasted 4 s. EMG activity of these muscles is shown as a function of the sinusoidal arm movement, recorded by the twin-axis electrogoniometers, and illustratedin the top traces. EMG activity of all muscles was observed during flexion–abduction, and during the transition phase of the arm from extension to flexion and froma
caedti(wjdjl(o
mpieoAlnl
pt(tptcsttioajoidc
dduction to abduction consistent with their anticipated function.
onditioned H-reflex reached an overall amplitude of 90 ± 4.8nd 91 ± 4% of the control reflex during ipsilateral and contralat-ral arm extension (P < 0.05). The reflex was equally depresseduring abduction and adduction of either upper limbs, while athe remaining arm movements (flexion and reciprocal) no signif-cant effects on the soleus H-reflex were encountered (P > 0.05)Fig. 4A). The M-waves were stable during arm movement andere of similar amplitude compared to those recorded with sub-
ects seated (compare Figs. 2B and 4B). A statistically significantifference between the conditioned reflexes recorded with sub-ects seated and standing was found for the contralateral upperimb with the reflex depression to be stronger during seatedP < 0.05) and for reflexes recorded during reciprocal movementf both upper limbs (P < 0.05).
The present study provides evidence that upper limb move-ent influences substantially soleus H-reflex excitability. In
articular, ipsilateral and/or contralateral upper limb movementn different directions with subjects seated or standing and bilat-ral arm movement in a reciprocal pattern, mimicking the onebserved during human walking, depressed the soleus H-reflex.
rm muscles were active during flexion and abduction, regard-ess the direction of the arm movement, while posterior tibialerve stimulation did not induce short responses in the upperimb muscles.
Aasa
During testing, the magnitude of the muscle compound actionotential (M-wave) was used as an indication of the constancy ofhe test afferent volley, which it did not exceed 5% of the Mmaxcompare Figs. 2B and 4B), verifying that across subjects andesting conditions the H-reflexes were elicited on the ascendingart of the recruitment curve. Further, control and conditioningrials were randomized minimizing the reflex depression asso-iated with the previously activated Ia fibres, and the ankle wastable across subjects. It is well established that during standinghe soleus H-reflex is depressed via an increased tonic presynap-ic inhibition acting on the Ia fibres [16], but the reflex depressionn standing subjects was counteracted by proper adjustmentsf the stimulus intensity. Lastly, all upper limb muscles werective mostly during flexion and abduction, regardless the sub-ects testing position (seated versus standing) or the directionf the movement. Thus, the reflex depression as a result of thepsilateral and contralateral upper limb movement in differentirections in seated and standing subjects could be attributed toommon mechanisms.
Static arm positioning influences the amplitude of the
chilles, quadriceps, and biceps femoris tendon reflexes as wells the human soleus H- and stretch reflexes [5,6], with theoleus H-reflex to be facilitated during static shoulder flexionnd depressed during static shoulder extension [6]. Similarly,

142 M. Knikou / Neuroscience Lett
Fig. 4. (A) Histogram showing the effect of ipsilateral and contralateral armmovement on the right soleus H-reflex with subjects standing. The overall mean(all subjects tested) magnitude of the soleus H-reflex (expressed as a percentageof the control reflex ± S.E.M.) is plotted for each direction of movement and armseparately. Movement of both arms induced a statistically significant depressionof the right soleus H-reflex at the 95% of confidence level when the arm moved inextension, abduction, and adduction. (B) Pool data (all subjects tested) showingthe average M-wave size during ipsilateral and contralateral arm movement.Nmw
shcmla
mothdiaoo
ltactS
asmibat[mmwt
eersbwcbr
A
v
R
[
[11] F. Ginanneschi, F. Dominici, A. Biasella, F. Gelli, A. Rossi, Changes in
ote that the magnitude of the M-wave was kept constant (below 5% of theaximal M-wave) across arm movements, and similar to the M-waves recordedith subjects seated.
tatic changes in arm positioning influence the reflex gain ofand and wrist muscles, attributed partly to changes at a corti-al level [10,11], while rhythmic ankle dorsi flexion/extensionodulate the flexor carpi radialis H-reflex in the resting upper
imbs [3]. It is thus apparent that upper and lower limbs interactnd modulate motor output depending on the task.
The soleus H-reflex was depressed during upper limb move-ent in the frontal and sagittal planes with subjects either seated
r standing. This is in line with the postulated reduction ofhe soleus H-reflex size during arm cycling using a customizedydraulic arm ergometer [9], with reflex depression to be presenturing arm flexion or extension. At this point we should note thatn this study subjects moved actively their upper limbs withoutny force constraints at the wrist, while arms moved separatelyr synchronously with subjects seated and standing in more thanne plane.
The H-reflex conditioned by common peroneal nerve stimu-ation at a long conditioning test interval (a conditioning stimulushat allows one to assess indirectly the presynaptic inhibition
cting on soleus Ia afferent terminals) was reduced during armycling compared to that observed in static trials [9], suggestinghat the reflex modulation occurs at a pre-motoneuronal level.imilarly, Jendrassik maneuver modulates soleus H-reflex by[
ers 416 (2007) 138–143
ffecting the level of presynaptic inhibition [25]. These studiesuggest that the soleus H-reflex modulation during arm move-ent might be associated with changes in the level of presynaptic
nhibition acting on Ia afferent terminals. Such changes mighte driven by propriospinal pathways which couple the cervicalnd lumbosacral enlargements subserving interlimb coordina-ion during locomotion. These pathways are evident in animals17,18] and humans [19]. Thus, muscle and joint afferentsediating the position of the upper limbs might affect alphaotoneuronal excitability of the legs through propriospinal path-ays that influence the level of monosynaptic Ia excitation of
he lower limbs at a presynaptic level.This study has demonstrated that ipsilateral or contralat-
ral upper limb movement in different directions with elbowsxtended and bilateral arm movement with elbows flexed in aeciprocal pattern depressed the soleus H-reflex in seated andtanding subjects. It is suggested that upper limb movement cane employed so to decrease spinal reflex excitability of peopleith a lesion to the central nervous system (e.g. stroke or spinal
ord injury), while arm movement involvement during the reha-ilitation training might benefit locomotion and sensorimotorecovery in these patients.
cknowledgement
The present study was partially supported by the City Uni-ersity of New York Research Foundation.
eferences
[1] B. Alstermark, A. Lundberg, M. Pinter, S. Sasaki, Subpopulations andfunctions of long C3-C5 propriospinal neurones, Brain Res. 404 (1987)395–400.
[2] B. Alstermark, A. Lundberg, M. Pinter, S. Sasaki, Long C3-C5 pro-priospinal neurones in the cat, Brain Res. 404 (1987) 382–388.
[3] F. Baldissera, P. Cavallari, L. Leocani, Cyclic modulation of the H-reflexin a wrist flexor during rhythmic flexion–extension movements of the ipsi-lateral foot, Exp. Brain Res. 118 (1998) 427–430.
[4] C. Capaday, R.B. Stein, Amplitude modulation of the soleus H-reflex inthe human during walking and standing, J. Neurosci. 6 (1986) 1308–1313.
[5] P.J. Delwaide, C. Figiel, C. Richelle, Influence de la position du membresuperieur sur l’ excitabilite de l’arc soleaire, Electromyogr. Clin. Neuro-physiol. 13 (1973) 515–523.
[6] P.J. Delwaide, C. Figiel, C. Richelle, Effects of postural changes of theupper limb on reflex transmission in the lower limb: cervicolumbar reflexinteractions in man, J. Neurol. Neurosurg. Psychiatry 40 (1977) 616–621.
[7] V. Dietz, Do human bipeds use quadrupedal coordination, Trends Neurosci.25 (2002) 462–467.
[8] V. Dietz, R. Muller, G. Colombo, Locomotor activity in spinal man: sig-nificance of afferent input from joint and load receptors, Brain 125 (2002)2626–2634.
[9] A. Frigon, D.F. Collins, E.P. Zehr, Effect of rhythmic arm movement onreflexes in the legs: modulation of soleus H-reflexes and somatosensoryconditioning, J. Neurophysiol. 91 (2004) 1516–1523.
10] F. Ginanneschi, F. Del Santo, F. Dominici, F. Gelli, R. Mazzocchio, A.Rossi, Changes in corticomotor excitability of hand muscles in relation tostatic shoulder positions, Exp. Brain Res. 161 (2005) 374–382.
corticomotor excitability of forearm muscles in relation to static shoulderpositions, Brain Res. 1073–1074 (2006) 332–338.
12] B.M. Giovanelli, H.G. Kuypers, Propriospinal fibers interconnecting thespinal enlargements in the cat, Brain Res. 14 (1969) 321–330.

e Lett
[
[
[
[
[
[
[
[
[
[
[
[
M. Knikou / Neuroscienc
13] E. Jankowska, A. Lundberg, W.J. Roberts, D.A. Stuart, A long propriospinalsystem with direct effects on motoneurones and on interneurones in the catlumbosacral cord, Exp. Brain Res. 21 (1974) 169–174.
14] M. Knikou, B.A. Conway, Modulation of soleus H-reflexes following ipsi-lateral mechanical loading of the sole of the foot in normal and completespinal cord injured humans, Neurosci. Lett. 303 (2001) 107–110.
15] M. Knikou, C. Taglianetti, On the methods employed to record andmeasure the human soleus H-reflex, Somatosens. Mot. Res. 23 (2006)55–62.
16] D.M. Koceja, M.H. Trimble, D.R. Earles, Inhibition of the soleus H-reflexin standing man, Brain Res. 629 (1993) 155–158.
17] S. Miller, R. van Berkum, J. van der Burg, F.G. van der Meche, Interlimbcoordination in stepping in the cat, J. Physiol. 230 (1973) 30–31.
18] S. Miller, J. van der Burg, F.G.A. van der Meche, Coordination of move-ments of the hindlimbs and forelimbs in different forms of locomotion innormal and decerebrate cats, Brain Res. 91 (1975) 217–257.
19] P.W. Nathan, M. Smith, P. Deacon, Vestibulospinal, reticulospinal anddescending propriospinal nerve fibres in man, Brain 119 (1996) 1809–1833.
[
[
ers 416 (2007) 138–143 143
20] C.S. Sherrington, E.E. Laslett, Observations on some spinal reflexes andthe interconnections of spinal segments, J. Physiol. 29 (1903) 58–96.
21] T. Wannier, C. Bastiaanse, G. Colombo, V. Dietz, Arm to leg coordinationduring walking, creeping and swimming activities, Exp. Brain Res. 141(2001) 375–379.
22] E.P. Zehr, D.F. Collins, A. Frigon, N. Hoogenboom, Neural control ofrhythmic human arm movement: phase dependence and task modulation ofHoffmann reflexes in forearm muscles, J. Neurophysiol. 89 (2003) 12–21.
23] E.P. Zehr, C. Haridas, Modulation of cutaneous reflexes in arm muscles dur-ing walking: further evidence of similar control mechanisms for rhythmichuman arm and leg movement, Exp. Brain Res. 149 (2003) 260–266.
24] E.P. Zehr, A. Kido, Neural control of rhythmic, cyclical human armmovement: task dependency, nerve specificity and phase modulation of
cutaneous reflexes, J. Physiol. 537 (2001) 1033–1045.25] E.P. Zehr, R.B. Stein, Interaction of the Jendrassik maneuver with segmen-tal presynaptic inhibition, Exp. Brain Res. 124 (1999) 474–480.
26] E.P. Zehr, R.B. Stein, T. Komiyama, Function of sural nerve reflexes duringhuman walking, J. Physiol. 507 (1997) 305–314.
Related Documents











