Netrin1 is required for neural and glial precursor migrations into the olfactory bulb Janne Hakanen a , Sébastien Duprat b,1 , Marjo Salminen a, ⁎ a Department of Veterinary Biosciences, University of Helsinki, Finland b Developmental Biology Program, Institute of Biotechnology, University of Helsinki, Finland abstract article info Article history: Received for publication 15 December 2010 Revised 23 March 2011 Accepted 15 April 2011 Available online 23 April 2011 Keywords: Interneuron Migration Netrin1 Oligodendrocyte Rostral migratory stream Olfactory bulb Netrin1 (NTN1) deficiency in mouse brain causes defects in axon guidance and cell migration during embryonic development. Here we show that NTN1 is required for olfactory bulb (OB) development at late embryogenesis and at early postnatal stages to facilitate the accumulation of proper numbers of granular and glomerular neuron subtypes and oligodendrocytes into the OB. In addition to the analysis of Ntn1-/- mice we made tissue and neurosphere cultures to clarify the role of NTN1 in the anterior forebrain. We propose that a subset of neural progenitors/precursors requires NTN1 to efficiently enter the rostral migratory stream to migrate into the OB. The analysis of postnatal Ntn1-/- OBs revealed a reduction of specific types of interneurons which have been shown to originate from particular subregions of the lateral ventricle walls. Based on Ntn1 expression in ventral parts of the ventricle walls, we observed a decrease in the mainly ventrally derived type II interneurons that express calcium-binding proteins calretinin and calbindin. Instead, no change in the numbers of dorsally derived tyrosine hydroxylase expressing interneurons was detected. In addition to the specific reduction of type II interneurons, our results indicate that NTN1 is required for oligodendroglial migration into the OB. Furthermore, we characterised the Ntn1 expressing subpopulation of neurosphere-forming cells from embryonic and adult brain as multipotent and self-renewing. However, NTN1 is dispensable for the proliferation of neurosphere forming progenitor cells and for their differentiation. © 2011 Elsevier Inc. All rights reserved. Introduction Neurogenesis, gliogenesis and precursor migration are fundamen- tal processes during central nervous system (CNS) development. Olfactory bulb (OB) development involves two waves of neurogenesis and migration. The first wave starts around E12–13 in mouse when projection neuron precursors and some interneuron precursor sub- types from the anterior part of the telencephalic ventricular zone (VZ) migrate out radially (Batista-Brito et al., 2008; Hinds, 1968). During the second wave starting around E14 and continuing postnatally, most interneuron precursors arrive to the OB from the telencephalic subventricular zone (SVZ) tangentially via the rostral migratory stream (RMS) (Lledo et al., 2008; Luskin, 1998; Pencea and Luskin, 2003). The OB gliogenesis is less well understood. The first astrocytes arrive in the OB around birth and at least some of them use tangential migration from the SVZ through the RMS (Law et al., 1999). Oligodendrocyte progenitor cells (OPCs) are heterogeneous and have multiple origins in the developing forebrain (Chong and Chan, 2010; Le Bras et al., 2005; Spassky et al., 2001; Woodruff et al., 2001). One population expressing the myelin proteolipid protein 1 (Plp1)- driven transgene is detected in the OB already at E14.5, whilst a platelet-derived growth factor receptor α (Pdgfra)-expressing popu- lation appears at E16.5 (Spassky et al., 2001). As with the in- terneurons, the number of OB glial cells increases strongly postnatally. Neural precursor migration from the telencephalic germinal regions to the OB is a highly regulated process influenced by attractive or repulsive cues as well as cell adhesion and extracellular matrix molecules (de Castro and Bribian, 2005; Ghashghaei et al., 2007). Also OPCs migrate long distances before differentiating into myelinating cells surrounding the axons (de Castro and Bribian, 2005; Jarjour and Kennedy, 2004; Miller, 2002; Qi et al., 2002; Tsai and Miller, 2002) but the molecules required for their migration especially to the OB have remained unknown. Netrins comprise a conserved family of laminin-related secreted molecules with members originally implicated in axon guidance and neuronal migration. Netrin1 (NTN1) is the best-characterised mem- ber of the family and in CNS it signals mainly through transmembrane receptors belonging to the Deleted in Colorectal Cancer (DCC) and Unc-5 homolog (UNC5) families (Barallobre et al., 2005; Hedgecock and Norris, 1997; Yurchenco and Wadsworth, 2004). Recently NTN1- integrin interactions have also been shown to guide neuronal migration into the cortex (Stanco et al., 2009). Here we have studied the role of NTN1 in OB development using prenatal and postnatal Ntn1-/- hypomorphic mice as well as tissue Developmental Biology 355 (2011) 101–114 ⁎ Corresponding author at: Department of Veterinary Biosciences, University of Helsinki, Agnes Sjöbergin katu 2, 00790 Helsinki, Finland. Fax: + 358 9 19157033. E-mail addresses: janne.hakanen@helsinki.fi (J. Hakanen), [email protected] (S. Duprat), marjo.salminen@helsinki.fi (M. Salminen). 1 Present address: CECS/I-STEM, Genopole Campus 1, 5 rue Henri Desbruères, 91030 EVRY Cedex. 0012-1606/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2011.04.016 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Developmental Biology 355 (2011) 101–114
Contents lists available at ScienceDirect
Developmental Biology
j ourna l homepage: www.e lsev ie r.com/deve lopmenta lb io logy
Netrin1 is required for neural and glial precursor migrations into the olfactory bulb
Janne Hakanen a, Sébastien Duprat b,1, Marjo Salminen a,⁎a Department of Veterinary Biosciences, University of Helsinki, Finlandb Developmental Biology Program, Institute of Biotechnology, University of Helsinki, Finland
⁎ Corresponding author at: Department of VeterinHelsinki, Agnes Sjöbergin katu 2, 00790 Helsinki, Finlan
E-mail addresses: [email protected] (J. Haka(S. Duprat), [email protected] (M. Salminen).
1 Present address: CECS/I-STEM, Genopole Campus 1,EVRY Cedex.
0012-1606/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.ydbio.2011.04.016
a b s t r a c t
a r t i c l e i n f oArticle history:Received for publication 15 December 2010Revised 23 March 2011Accepted 15 April 2011Available online 23 April 2011
Keywords:InterneuronMigrationNetrin1OligodendrocyteRostral migratory streamOlfactory bulb
Netrin1 (NTN1) deficiency in mouse brain causes defects in axon guidance and cell migration duringembryonic development. Here we show that NTN1 is required for olfactory bulb (OB) development at lateembryogenesis and at early postnatal stages to facilitate the accumulation of proper numbers of granular andglomerular neuron subtypes and oligodendrocytes into the OB. In addition to the analysis of Ntn1−/− micewemade tissue and neurosphere cultures to clarify the role of NTN1 in the anterior forebrain.We propose thata subset of neural progenitors/precursors requires NTN1 to efficiently enter the rostral migratory stream tomigrate into the OB. The analysis of postnatal Ntn1−/− OBs revealed a reduction of specific types ofinterneurons which have been shown to originate from particular subregions of the lateral ventricle walls.Based on Ntn1 expression in ventral parts of the ventricle walls, we observed a decrease in the mainlyventrally derived type II interneurons that express calcium-binding proteins calretinin and calbindin. Instead,no change in the numbers of dorsally derived tyrosine hydroxylase expressing interneurons was detected. Inaddition to the specific reduction of type II interneurons, our results indicate that NTN1 is required foroligodendroglial migration into the OB. Furthermore, we characterised the Ntn1 expressing subpopulation ofneurosphere-forming cells from embryonic and adult brain asmultipotent and self-renewing. However, NTN1is dispensable for the proliferation of neurosphere forming progenitor cells and for their differentiation.
ary Biosciences, University ofd. Fax: +358 9 19157033.nen), [email protected]
5 rue Henri Desbruères, 91030
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Neurogenesis, gliogenesis and precursor migration are fundamen-tal processes during central nervous system (CNS) development.Olfactory bulb (OB) development involves two waves of neurogenesisand migration. The first wave starts around E12–13 in mouse whenprojection neuron precursors and some interneuron precursor sub-types from the anterior part of the telencephalic ventricular zone (VZ)migrate out radially (Batista-Brito et al., 2008; Hinds, 1968). Duringthe second wave starting around E14 and continuing postnatally,most interneuron precursors arrive to the OB from the telencephalicsubventricular zone (SVZ) tangentially via the rostral migratorystream (RMS) (Lledo et al., 2008; Luskin, 1998; Pencea and Luskin,2003).
The OB gliogenesis is less well understood. The first astrocytesarrive in the OB around birth and at least some of them use tangentialmigration from the SVZ through the RMS (Law et al., 1999).Oligodendrocyte progenitor cells (OPCs) are heterogeneous andhave multiple origins in the developing forebrain (Chong and Chan,
2010; Le Bras et al., 2005; Spassky et al., 2001; Woodruff et al., 2001).One population expressing the myelin proteolipid protein 1 (Plp1)-driven transgene is detected in the OB already at E14.5, whilst aplatelet-derived growth factor receptor α (Pdgfra)-expressing popu-lation appears at E16.5 (Spassky et al., 2001). As with the in-terneurons, the number of OB glial cells increases strongly postnatally.
Neural precursor migration from the telencephalic germinalregions to the OB is a highly regulated process influenced by attractiveor repulsive cues as well as cell adhesion and extracellular matrixmolecules (de Castro and Bribian, 2005; Ghashghaei et al., 2007). AlsoOPCs migrate long distances before differentiating into myelinatingcells surrounding the axons (de Castro and Bribian, 2005; Jarjour andKennedy, 2004; Miller, 2002; Qi et al., 2002; Tsai andMiller, 2002) butthe molecules required for their migration especially to the OB haveremained unknown.
Netrins comprise a conserved family of laminin-related secretedmolecules with members originally implicated in axon guidance andneuronal migration. Netrin1 (NTN1) is the best-characterised mem-ber of the family and in CNS it signals mainly through transmembranereceptors belonging to the Deleted in Colorectal Cancer (DCC) andUnc-5 homolog (UNC5) families (Barallobre et al., 2005; Hedgecockand Norris, 1997; Yurchenco and Wadsworth, 2004). Recently NTN1-integrin interactions have also been shown to guide neuronalmigration into the cortex (Stanco et al., 2009).
Here we have studied the role of NTN1 in OB development usingprenatal and postnatal Ntn1−/− hypomorphic mice as well as tissue

102 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
and cell culture assays. We show that NTN1 is required for efficientmigration of a subpopulation of CNS precursors from the lateralventricle walls into the RMS and OB. In line with this, the Ntn1−/−OBs were reduced in size and showed a clear reduction inoligodendrocytes in addition to a significant decrease in the numbersof specific types of ventrally derived interneurons.
Material and methods
Mouse strains
The generation and genotyping of mice carrying a gene trapinsertion in Ntn1 locus in C57Bl6 background have been des-cribed before (Salminen et al., 2000). To generate Ntn1−/− animals,Ntn1+/− mice from C57Bl6 and ICR backgrounds were crossed andalso pure C57Bl6 background was used for some of the analyses. TheNtn1mice carry an IRESβgeo cassette, which produces a fusion proteincontaining both β-galactosidase (β-gal) and neomycin phosphotrans-ferase (NeoR) activities (Friedrich and Soriano, 1991) conferring thusGeneticin resistance to Ntn1 expressing cells. All mouse work hasbeen approved by the University of Helsinki ethical review board.
Neurosphere cultures
Embryonic forebrains, the lateral walls of the adult forebrainventricles and spinal cord were isolated and cultured as described inJohansson et al. (1999). The number of primary neurospheres fromfour separate cultures/genotype isolated from E18.5 brains wascalculated after 7–10 days of culture. To generate secondary ortertiary spheres from adult brain or spinal cord, neurospheres weredissociatedmechanically with a pipette after 7–10 days of culture. Theresulting single-cell suspension was centrifuged down and plated infresh medium.
To select Geneticin resistant neurospheres, 1.5 mg/ml of Geneticin(Invitrogen) was added into the neurosphere medium. This concen-tration was tested and it kills all cells that do not contain at least onecopy of IRESβgeo in the Ntn1 locus (data not shown). Geneticin wasomitted from differentiating sphere cultures.
For differentiation assays, neurospheres were grown for twoweeks on poly-L-ornithine (Sigma) coated glass coverslips in neuro-sphere medium where growth factors were replaced by 5% foetal calfserum (HyClone). Immunostaining of 4% paraformaldehyde (PFA)fixed differentiated neurosphere derived cells was performed withantibodies against O4 1:80 and GFAP 1:1000 (Chemicon), TUJ1 1:250(Biosite). The number of differentiated cells was calculated from atleast 10 fields/slide, each field having at least 40 cells. Four slides/genotype and at least 3 animals/genotype were used for calculationsfor both selected and non-selected spheres.
For clonal cultures, neurospheres were dissociated and in average4–5 cells were placed into each well of a 96-well plate. The number offormed spheres was calculated after 7 days.
RNA isolation and RT-PCR
Total RNA extraction from neurospheres was performed witheither RNAeasy microkit or minikit (Qiagen). cDNA was obtainedwith Superscript III enzyme using random hexamer primers(Invitrogen). PCR amplification was performed in 35 cycles withDynazyme II (Finnzymes) and following settings: 45 s at 94 °C, 60 sat 55 °C and 75 s at 72 °C; followed by 60 s at 94 °C, 60 s at 55 °C and75 s at 72 °C. The used primers were the following: Mash1 forward:cgtcctctccggaactgat, reverse: tcctgcttccaaagtccatt; Noggin forward:aaggatctgaacgagacg, reverse: gcaggaacacttacactcg; Bmp4 forward:ctcccaagaatcatggactg, reverse: aaagcagagctctcactggt; Musashi1 for-ward: gtttcggcttcgtcactttca, reverse: atgtcctcgctctcaaacgtg; Nestinforward: gtggcctctgggatgatg, reverse: ttgaccttcctccccctc; Ntn1 for-
ward: aagcctatcacccaccggaag, reverse: gcatcaagattcctgtggcg, Olig2forward: ccacgtcttccaccaagaaa, reverse: caccagtcgcttcatctcct; Pax6forward: agttcttcgcaacctggcta, reverse: tgaagctgctgctgatagga; Gadphforward: accacagtccatgccatcac, reverse: tccaccaccctgttgctgta.
TUNEL assay, X-gal staining, RNA in situ hybridisation andimmunohistochemistry
To analyse programmed cell death, 5 μm thick paraffin sectionsfrom Ntn1−/− and control littermate Pn1 mice were subjected toTUNEL analysis with the ApopTag Fluorescein In Situ ApoptosisDetection Kit (Chemicon) according to the manufacturer's instruc-tions and as described in Cecconi et al. (2004). The average proportionof TUNEL positive cells from all cells in the 300×300 μm areaindicated in Suppl. Fig. 5 was calculated as below.
For X-gal staining, brains were cut in two halves and the fixedbrain tissues were stained as in Hogan et al. (1994). The stainedsamples were blocked in 0.5% gelatin/30% albumin/20% sucrose/2%glutaraldehyde (all from Sigma) and 100–200 μm sections were cutwith a vibratome. The vibratome sections were cleared with glycerolbefore photography.
The radioactive RNA in situ hybridisation assays on 5–10 μm serialparaffin sections from E14.5–18.5 embryos and Pn6 mouse brainswere performed as before (Salminen et al., 2000) and the imagesweretreated as in Lilleväli et al. (2006). The preparation of antisense andsense probes were performed as described in Salminen et al. (2000).No specific signals were detected with any of the sense probes (datanot shown).
For immunohistochemistry, either paraffin embedded or frozenbrain sections were used. For paraffin sections antigen retrieval wasperformed after deparaffination with Antigen unmasking solution(Vector). Sections were blocked for 1 h at room temperature with 10%goat serum and 3% triton X-100 in PBS, except for OLIG2 where horseserum was used, followed by incubation with primary antibodyovernight at +4 °C. Antibody dilutions: CNP 1:100, calR 1:200, NG21:1000, PSA-NCAM 1:500 and GFAP 1:500 (Chemicon), calB 1:500(Swant), OLIG2 1:800 (Neuromics), S-100β 1:500 (Sigma), PDGFRA1:100 (Cell Signaling), NeuN 1:150 and TH 1:500 (Millipore), β-gal1:1000 (MP Biomedicals, Inc), and TUJ1 1:500 (Biosite). Secondaryantibodies were Alexa Fluor 1:500 (Molecular Probes), except for NG2and PDGFRA for which detection was performed with UltravisionDetection System (Thermo Scientific) combined with TSA PlusFluorescence System (PerkinElmer). NG2/β-gal double-labelling wasperformed using CARD amplification method (Hunyady et al., 1996).The nuclei in the samples were visualised using bis-benzimide(Hoechst No. 33258, Sigma).
The stained sections were mounted in Vectashield (Vector) andphotographed with an Olympus DP70 CCD-camera attached to anOlympus AX70 microscope. Confocal stacks were collected with aLeica TCS SP3 confocal system. Images and colocalisation analysis wasmade with a Bitplane Imaris software (Bitplane Inc.).
Calculation of cell numbers in the olfactory bulb and RMS
Calculations of cell numbers were performed on 5–10 μm thickserial paraffin sections prepared from Ntn1−/− and control wildtype littermates (n=3, at least 3 sections/sample, if not otherwisestated, corresponding sections from Ntn1−/− and controls wereused for comparison). Student's t-test was used for all statisticalanalyses. Significant differences between populations are indicatedwith P values: Pb0.05*, Pb0.01** or Pb0.001***.
To calculate the numbers of calR+, calB+ or TH+ cells in theglomeruli at Pn16 and Pn1, sections were stained with correspondingantibodies and Hoechst. The calculation was performed on 14glomeruli/OB section. Every second glomerulus was used to avoid

103J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
overlapping cell counting of adjacent glomeruli. The control andmutant sections were matched for calculation.
The total number of β-gal+ cells and proportions of double-labelled β-gal+ and NG2+, PSA-NCAM+, TUJ1+ or GFAP+ cells in thesagittal cryostat 10 μm sections through the RMS of heterozygous Pn6mice (n=2) was counted from 4 to 5 confocal images/each antibody.Colocalisation was confirmed by following individual cells throughthe confocal stacks.
Forebrain explants
Cultures of forebrain explants were in general performed aspreviously described (Hack et al., 2002). Briefly, brains from E18.5embryos were dissected out and placed in ice-cold serum-freemedium (Invitrogen) after which the dorsal part was removed. Theremaining anterior forebrain was cut into three pieces as shown inFig. 7A. The explants were mixed with Matrigel (Beckton Dickinson)and placed in 24-well plates. After polymerisation for 10 min, sampleswere overlaid with 500 μl of serum-free medium containing B-27supplement (Invitrogen). Explant cultures were maintained 48 h in ahumidified, 5% CO2, 37 °C incubator. After incubation, the explantswere X-gal stained as above. To measure migration distances, imagesof explants were collected using inverted light microscope (Leica DMIRB) and analysed using Image-Pro Plus 5.1 software (MediaCybernetics, Inc.). Migration distances were measured from a totalof 6 explants/genotype.
Results
Ntn1 is expressed in VZ and RMS during brain development and in theependymal cells of the adult CNS
We verified Ntn1 expression in developing and adult mouse brainby RNA in situ hybridisation and X-gal staining or anti-β-galimmunohistochemistry, where we took advantage of a mouse linecontaining a lacZ insertion in Ntn1 locus (Salminen et al., 2000).
During brain development at embryonic day (E)14.5, Ntn1 mRNAwas detected in the VZ of the lateral wall of the ventral telencephalonin medial and lateral ganglionic eminences (MGE and LGE, respec-tively) as well as in striatum and septum. Very little or no expressioncould be detected in the SVZ (Fig. 1A). Also the ventralmost portion ofthe septal wall VZ expressed Ntn1 (data not shown). Prior to birth, atE18.5, a similar Ntn1 mRNA distribution pattern was observed withhighest levels confined to the VZ of the lateral wall of the forebrainventricles (Figs. 1B,C) excluding however the dorsalmost part (arrowin Fig. 1B). Expression was also detected in a rostral region continuousto the Ntn1 expression in the VZ, therefore referred to as the rostralextension of the VZ (arrow in Fig. 1C). This region represents also themost proximal part of the RMS. In the developing OB itself, very weakexpression was detected in the granular cell layer at E18.5 (Fig. 1C).
In general, Ntn1-driven lacZ expression co-localised with Ntn1mRNA during development (Figs. 1C,D and data not shown).We had acloser look at the presence of β-gal positive cells in the proximal RMSof Ntn1+/− mice with immunohistochemistry and Hoechst-counter-staining (Figs. 1E,F). β-gal positive cells extended from the VZ into theRMS (until the white arrowhead in Fig. 1E), but populated only theventral half of the nuclei-dense horizontal RMS (Fig. 1F).
In postnatal day 6 (Pn6) brain, Ntn1/lacZwas strongly expressed inventral forebrain areas including the striatum and septum and as inembryos, strongest expression was observed along the walls of thelateral ventricle. Strong β-gal activity was detected also along thewhole horizontal RMS and in the OB granular cell layer (Fig. 1G).The β-gal protein was detected mainly ventrally and in the middle ofthe RMS throughout its length but was missing in the most dorsalparts (Fig. 1J). Also at Pn6, Ntn1 mRNA distribution coincided withβ-gal (Figs. 1G,H) except for the RMSwhere β-gal protein distribution
(Fig. 1J) was wider than Ntn1 mRNA distribution and the few Ntn1mRNA expressing cells detected were restricted to the ventral-mostborder (arrow in Figs. 1H,K). This difference could be due to therelatively longer stability of the β-gal protein compared toNtn1mRNAin the cells migrating out from the VZ. In the OB layers, Ntn1 mRNAwas detected only in the granular cells at Pn6 (Fig. 1I) whereas noexpressionwas detected at any of the stages analysed in themitral celllayer (Fig. 1 and data not shown) in contrast to the observations ofMurase and Horwitz (2002).
In adult brain, strongest Ntn1/lacZ expression could be detected inthe lateral walls of the lateral ventricles and in the ventralmost part ofthe medial wall as during development (Fig. 1M and data not shown).In the ventricle walls the expression was confined to one continuouslayer of cells next to the ventricle lumen (Fig. 1M) corresponding tothe ependymal cells. Ntn1/lacZ expression could also be detected inthe ependymal layer of more caudal brain regions and even in thespinal cord (Fig. 1L). To confirm the ependymal expression of Ntn1/lacZ we double-stained adult brain cryosections with β-gal (red) andan ependymal marker S-100β (green) antibodies (Figs. 1N,O). Themerged picture (Fig. 1P) shows a clear co-expression of these markers(yellow).
Taken together, our results suggest that Ntn1-expressing cells inthe VZ and/or its rostral extension produce precursor cells thatmigrate into the OB along the ventral part of the RMS duringembryonic and early postnatal development.
Characterisation of Ntn1/lacZ-expressing cells in RMS
The postnatal RMS contains a heterogeneous population ofmigrating cells including precursors for both neurons and glia ontheir way to the OB (Aguirre and Gallo, 2004; Gritti et al., 2002). Inorder to characterise the Ntn1/lacZ-positive cell population in RMS(Fig. 2A) we performed double-immunohistochemistry with β-galantibody together with antibodies against specific markers detectedin the postnatal RMS. GFAP is a marker for astrocytes and SVZprogenitor cells and GFAP+ cells have also been detected in postnatalRMS (Law et al., 1999; Pencea and Luskin, 2003). PSA-NCAM and classIIIb-tubulin (TUJ1) are markers for migrating neural precursors(Chazal et al., 2000; Rousselot et al., 1995) whilst NG2 is expressedin OPCs and interneuron precursors migrating through the RMS(Aguirre and Gallo, 2004).
In our assay, a proportion of the β-gal+ cells were also GFAP+,PSA-NCAM+, TUJ1+ or NG2+ (Figs. 2B–E and Table 1) suggesting thatthe Ntn1-expressing cells in the ventral VZ of the forebrain produce aheterogeneous subset of precursor cells into the RMS that contributeto both neural and glial populations in the OB. Highest proportionof Ntn1-expressing cells were neural precursors. Interestingly, likeβ-gal+ cells, also the NG2+ cells located mainly in the ventral portionof the RMS (Fig. 2E). However, a proportion of the β-gal+ cells did notexpress any of the tested markers (data not shown) suggesting thatthey may be more immature cells such as proliferating progenitorsthat have also been detected in the RMS (Coskun and Luskin, 2002).
Ntn1-expressing cells from adult brain and spinal cord are multipotentand self-renewing
Proliferating cells can be isolated from neurogenic brain regions ofembryonic and adult rodents and grown as floating neurospheres. WeverifiedwhetherNtn1was expressed in cultured neurospheres, in linewith its expression in the embryonic germinal zone and in one of thecell types included in the adult neural stem cell niche, the ependymalcell. We established neurosphere cultures from forebrains of E12.5,E14.5 and E18.5 wild-type embryos as well as from adult spinal cordsand the dissected lateral ventricle walls of adult brains. Ntn1expression was detected in all proliferating neurosphere culturestogether with the stem/progenitor cell markers (Suppl. Figs. 1A,B).
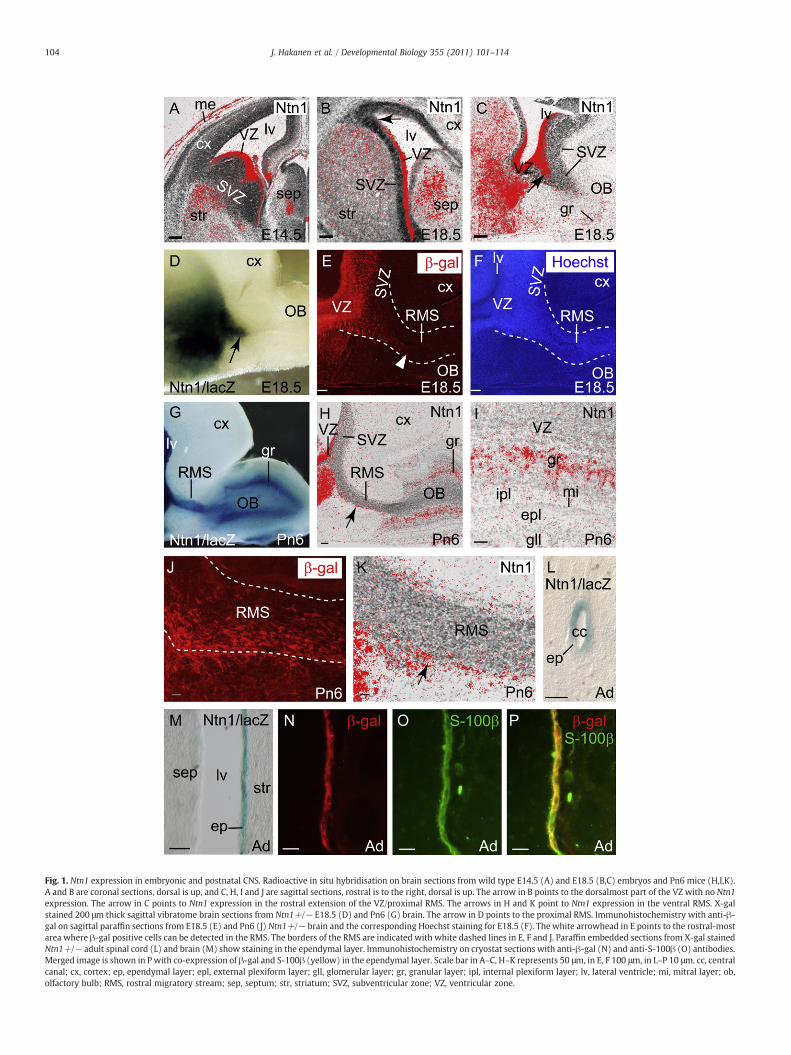
Fig. 1. Ntn1 expression in embryonic and postnatal CNS. Radioactive in situ hybridisation on brain sections from wild type E14.5 (A) and E18.5 (B,C) embryos and Pn6 mice (H,I,K).A and B are coronal sections, dorsal is up, and C, H, I and J are sagittal sections, rostral is to the right, dorsal is up. The arrow in B points to the dorsalmost part of the VZ with no Ntn1expression. The arrow in C points to Ntn1 expression in the rostral extension of the VZ/proximal RMS. The arrows in H and K point to Ntn1 expression in the ventral RMS. X-galstained 200 μm thick sagittal vibratome brain sections from Ntn1+/− E18.5 (D) and Pn6 (G) brain. The arrow in D points to the proximal RMS. Immunohistochemistry with anti-β-gal on sagittal paraffin sections from E18.5 (E) and Pn6 (J) Ntn1+/− brain and the corresponding Hoechst staining for E18.5 (F). The white arrowhead in E points to the rostral-mostarea where β-gal positive cells can be detected in the RMS. The borders of the RMS are indicated with white dashed lines in E, F and J. Paraffin embedded sections from X-gal stainedNtn1+/− adult spinal cord (L) and brain (M) show staining in the ependymal layer. Immunohistochemistry on cryostat sections with anti-β-gal (N) and anti-S-100β (O) antibodies.Merged image is shown in P with co-expression of β-gal and S-100β (yellow) in the ependymal layer. Scale bar in A–C, H–K represents 50 μm, in E, F 100 μm, in L–P 10 μm. cc, centralcanal; cx, cortex; ep, ependymal layer; epl, external plexiform layer; gll, glomerular layer; gr, granular layer; ipl, internal plexiform layer; lv, lateral ventricle; mi, mitral layer; ob,olfactory bulb; RMS, rostral migratory stream; sep, septum; str, striatum; SVZ, subventricular zone; VZ, ventricular zone.
104 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
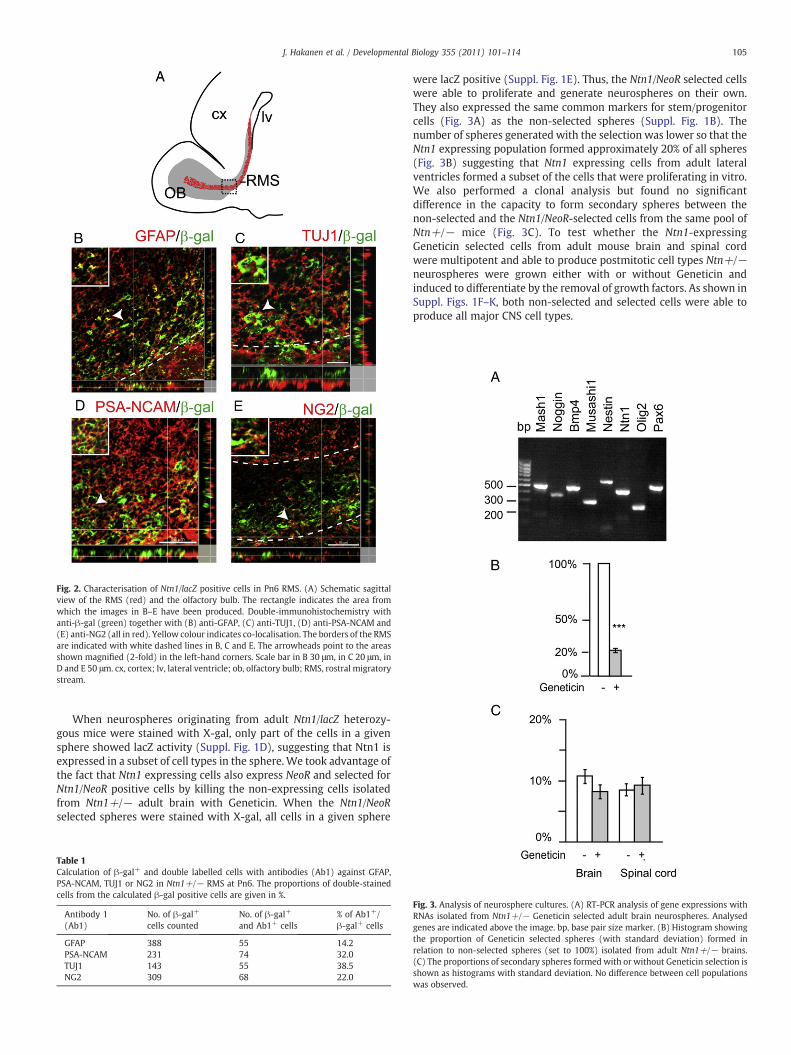
Fig. 2. Characterisation of Ntn1/lacZ positive cells in Pn6 RMS. (A) Schematic sagittalview of the RMS (red) and the olfactory bulb. The rectangle indicates the area fromwhich the images in B–E have been produced. Double-immunohistochemistry withanti-β-gal (green) together with (B) anti-GFAP, (C) anti-TUJ1, (D) anti-PSA-NCAM and(E) anti-NG2 (all in red). Yellow colour indicates co-localisation. The borders of the RMSare indicated with white dashed lines in B, C and E. The arrowheads point to the areasshown magnified (2-fold) in the left-hand corners. Scale bar in B 30 μm, in C 20 μm, inD and E 50 μm. cx, cortex; lv, lateral ventricle; ob, olfactory bulb; RMS, rostral migratorystream.
105J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
When neurospheres originating from adult Ntn1/lacZ heterozy-gous mice were stained with X-gal, only part of the cells in a givensphere showed lacZ activity (Suppl. Fig. 1D), suggesting that Ntn1 isexpressed in a subset of cell types in the sphere. We took advantage ofthe fact that Ntn1 expressing cells also express NeoR and selected forNtn1/NeoR positive cells by killing the non-expressing cells isolatedfrom Ntn1+/− adult brain with Geneticin. When the Ntn1/NeoRselected spheres were stained with X-gal, all cells in a given sphere
Table 1Calculation of β-gal+ and double labelled cells with antibodies (Ab1) against GFAP,PSA-NCAM, TUJ1 or NG2 in Ntn1+/− RMS at Pn6. The proportions of double-stainedcells from the calculated β-gal positive cells are given in %.
Antibody 1(Ab1)
No. of β-gal+
cells countedNo. of β-gal+
and Ab1+ cells% of Ab1+/β-gal+ cells
GFAP 388 55 14.2PSA-NCAM 231 74 32.0TUJ1 143 55 38.5NG2 309 68 22.0
were lacZ positive (Suppl. Fig. 1E). Thus, the Ntn1/NeoR selected cellswere able to proliferate and generate neurospheres on their own.They also expressed the same common markers for stem/progenitorcells (Fig. 3A) as the non-selected spheres (Suppl. Fig. 1B). Thenumber of spheres generated with the selection was lower so that theNtn1 expressing population formed approximately 20% of all spheres(Fig. 3B) suggesting that Ntn1 expressing cells from adult lateralventricles formed a subset of the cells that were proliferating in vitro.We also performed a clonal analysis but found no significantdifference in the capacity to form secondary spheres between thenon-selected and the Ntn1/NeoR-selected cells from the same pool ofNtn+/− mice (Fig. 3C). To test whether the Ntn1-expressingGeneticin selected cells from adult mouse brain and spinal cordwere multipotent and able to produce postmitotic cell types Ntn+/−neurospheres were grown either with or without Geneticin andinduced to differentiate by the removal of growth factors. As shown inSuppl. Figs. 1F–K, both non-selected and selected cells were able toproduce all major CNS cell types.
Fig. 3. Analysis of neurosphere cultures. (A) RT-PCR analysis of gene expressions withRNAs isolated from Ntn1+/− Geneticin selected adult brain neurospheres. Analysedgenes are indicated above the image. bp, base pair size marker. (B) Histogram showingthe proportion of Geneticin selected spheres (with standard deviation) formed inrelation to non-selected spheres (set to 100%) isolated from adult Ntn1+/− brains.(C) The proportions of secondary spheres formed with or without Geneticin selection isshown as histograms with standard deviation. No difference between cell populationswas observed.
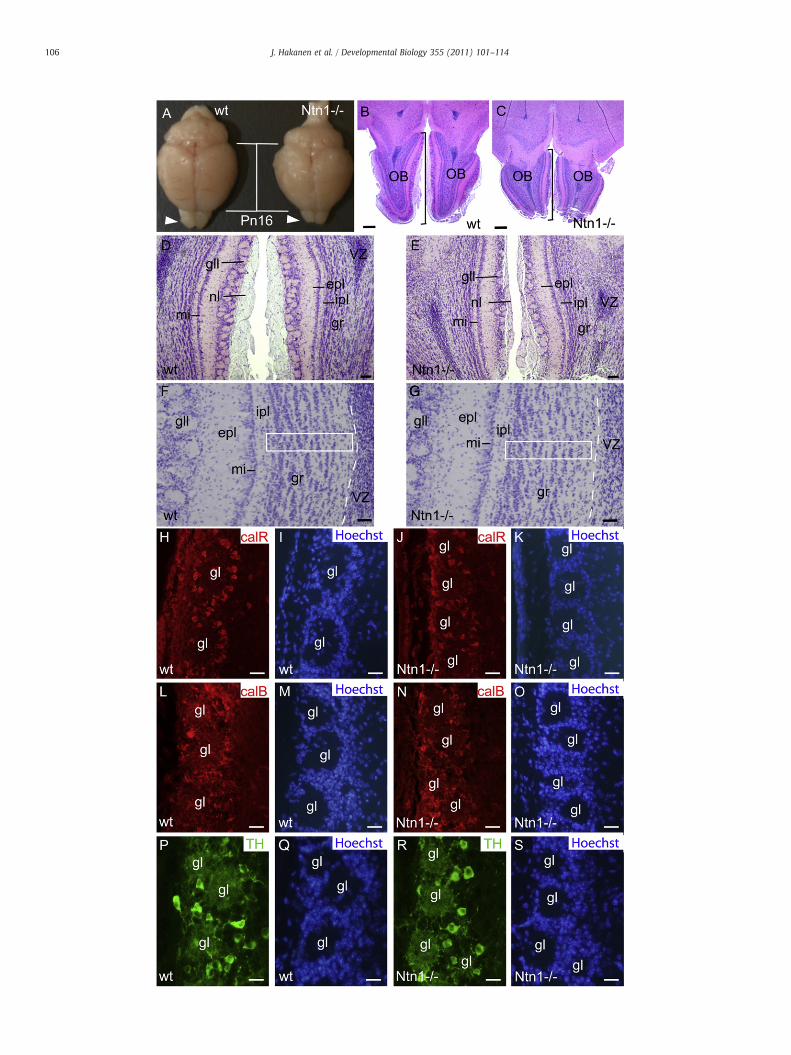
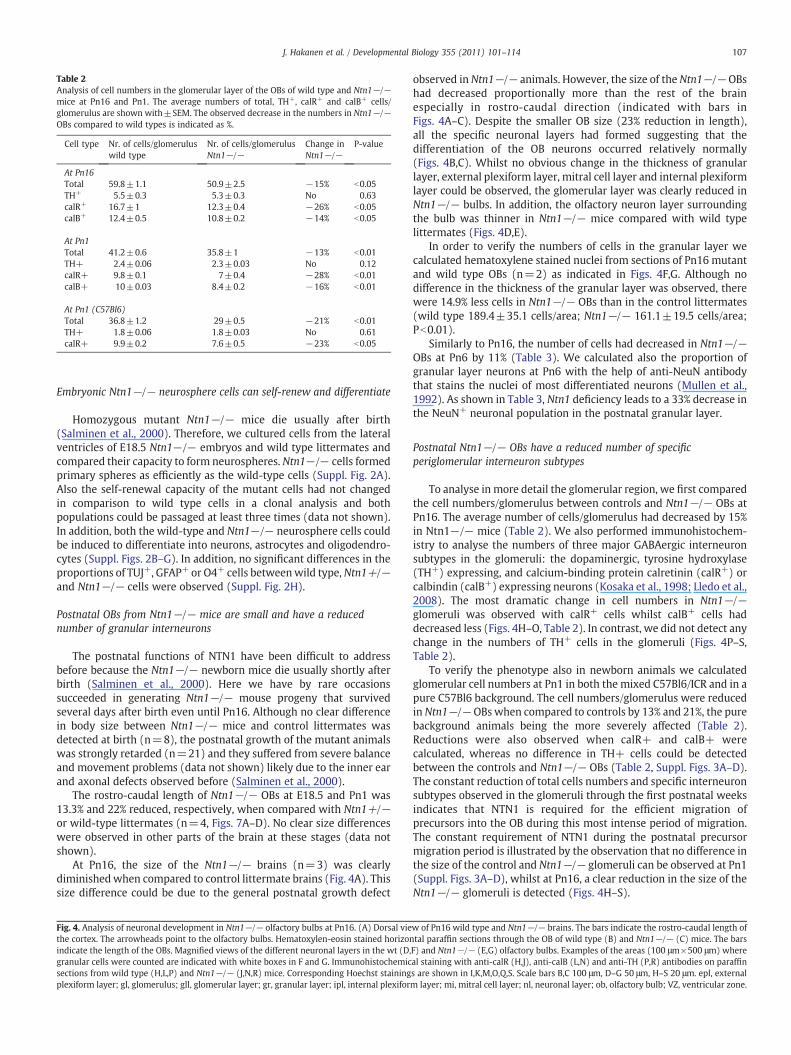
Table 2Analysis of cell numbers in the glomerular layer of the OBs of wild type and Ntn1−/−mice at Pn16 and Pn1. The average numbers of total, TH+, calR+ and calB+ cells/glomerulus are shown with±SEM. The observed decrease in the numbers in Ntn1−/−OBs compared to wild types is indicated as %.
Cell type Nr. of cells/glomeruluswild type
Nr. of cells/glomerulusNtn1−/−
Change inNtn1−/−
P-value
At Pn16Total 59.8±1.1 50.9±2.5 −15% b0.05TH+ 5.5±0.3 5.3±0.3 No 0.63calR+ 16.7±1 12.3±0.4 −26% b0.05calB+ 12.4±0.5 10.8±0.2 −14% b0.05
At Pn1Total 41.2±0.6 35.8±1 −13% b0.01TH+ 2.4±0.06 2.3±0.03 No 0.12calR+ 9.8±0.1 7±0.4 −28% b0.01calB+ 10±0.03 8.4±0.2 −16% b0.01
At Pn1 (C57Bl6)Total 36.8±1.2 29±0.5 −21% b0.01TH+ 1.8±0.06 1.8±0.03 No 0.61calR+ 9.9±0.2 7.6±0.5 −23% b0.05
107J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
Embryonic Ntn1−/− neurosphere cells can self-renew and differentiate
Homozygous mutant Ntn1−/− mice die usually after birth(Salminen et al., 2000). Therefore, we cultured cells from the lateralventricles of E18.5 Ntn1−/− embryos and wild type littermates andcompared their capacity to form neurospheres. Ntn1−/− cells formedprimary spheres as efficiently as the wild-type cells (Suppl. Fig. 2A).Also the self-renewal capacity of the mutant cells had not changedin comparison to wild type cells in a clonal analysis and bothpopulations could be passaged at least three times (data not shown).In addition, both the wild-type and Ntn1−/− neurosphere cells couldbe induced to differentiate into neurons, astrocytes and oligodendro-cytes (Suppl. Figs. 2B–G). In addition, no significant differences in theproportions of TUJ+, GFAP+ or O4+ cells betweenwild type,Ntn1+/−and Ntn1−/− cells were observed (Suppl. Fig. 2H).
Postnatal OBs from Ntn1−/− mice are small and have a reducednumber of granular interneurons
The postnatal functions of NTN1 have been difficult to addressbefore because the Ntn1−/− newborn mice die usually shortly afterbirth (Salminen et al., 2000). Here we have by rare occasionssucceeded in generating Ntn1−/− mouse progeny that survivedseveral days after birth even until Pn16. Although no clear differencein body size between Ntn1−/− mice and control littermates wasdetected at birth (n=8), the postnatal growth of the mutant animalswas strongly retarded (n=21) and they suffered from severe balanceand movement problems (data not shown) likely due to the inner earand axonal defects observed before (Salminen et al., 2000).
The rostro-caudal length of Ntn1−/− OBs at E18.5 and Pn1 was13.3% and 22% reduced, respectively, when compared with Ntn1+/−or wild-type littermates (n=4, Figs. 7A–D). No clear size differenceswere observed in other parts of the brain at these stages (data notshown).
At Pn16, the size of the Ntn1−/− brains (n=3) was clearlydiminishedwhen compared to control littermate brains (Fig. 4A). Thissize difference could be due to the general postnatal growth defect
Fig. 4. Analysis of neuronal development in Ntn1−/− olfactory bulbs at Pn16. (A) Dorsal viethe cortex. The arrowheads point to the olfactory bulbs. Hematoxylen-eosin stained horizonindicate the length of the OBs. Magnified views of the different neuronal layers in the wt (D,granular cells were counted are indicated with white boxes in F and G. Immunohistochemicsections from wild type (H,L,P) and Ntn1−/− (J,N,R) mice. Corresponding Hoechst stainingplexiform layer; gl, glomerulus; gll, glomerular layer; gr, granular layer; ipl, internal plexifor
observed in Ntn1−/− animals. However, the size of the Ntn1−/− OBshad decreased proportionally more than the rest of the brainespecially in rostro-caudal direction (indicated with bars inFigs. 4A–C). Despite the smaller OB size (23% reduction in length),all the specific neuronal layers had formed suggesting that thedifferentiation of the OB neurons occurred relatively normally(Figs. 4B,C). Whilst no obvious change in the thickness of granularlayer, external plexiform layer, mitral cell layer and internal plexiformlayer could be observed, the glomerular layer was clearly reduced inNtn1−/− bulbs. In addition, the olfactory neuron layer surroundingthe bulb was thinner in Ntn1−/− mice compared with wild typelittermates (Figs. 4D,E).
In order to verify the numbers of cells in the granular layer wecalculated hematoxylene stained nuclei from sections of Pn16 mutantand wild type OBs (n=2) as indicated in Figs. 4F,G. Although nodifference in the thickness of the granular layer was observed, therewere 14.9% less cells in Ntn1−/− OBs than in the control littermates(wild type 189.4±35.1 cells/area; Ntn1−/− 161.1±19.5 cells/area;Pb0.01).
Similarly to Pn16, the number of cells had decreased in Ntn1−/−OBs at Pn6 by 11% (Table 3). We calculated also the proportion ofgranular layer neurons at Pn6 with the help of anti-NeuN antibodythat stains the nuclei of most differentiated neurons (Mullen et al.,1992). As shown in Table 3, Ntn1 deficiency leads to a 33% decrease inthe NeuN+ neuronal population in the postnatal granular layer.
Postnatal Ntn1−/− OBs have a reduced number of specificperiglomerular interneuron subtypes
To analyse in more detail the glomerular region, we first comparedthe cell numbers/glomerulus between controls and Ntn1−/− OBs atPn16. The average number of cells/glomerulus had decreased by 15%in Ntn1−/− mice (Table 2). We also performed immunohistochem-istry to analyse the numbers of three major GABAergic interneuronsubtypes in the glomeruli: the dopaminergic, tyrosine hydroxylase(TH+) expressing, and calcium-binding protein calretinin (calR+) orcalbindin (calB+) expressing neurons (Kosaka et al., 1998; Lledo et al.,2008). The most dramatic change in cell numbers in Ntn1−/−glomeruli was observed with calR+ cells whilst calB+ cells haddecreased less (Figs. 4H–O, Table 2). In contrast, we did not detect anychange in the numbers of TH+ cells in the glomeruli (Figs. 4P–S,Table 2).
To verify the phenotype also in newborn animals we calculatedglomerular cell numbers at Pn1 in both the mixed C57Bl6/ICR and in apure C57Bl6 background. The cell numbers/glomerulus were reducedin Ntn1−/− OBs when compared to controls by 13% and 21%, the purebackground animals being the more severely affected (Table 2).Reductions were also observed when calR+ and calB+ werecalculated, whereas no difference in TH+ cells could be detectedbetween the controls and Ntn1−/− OBs (Table 2, Suppl. Figs. 3A–D).The constant reduction of total cells numbers and specific interneuronsubtypes observed in the glomeruli through the first postnatal weeksindicates that NTN1 is required for the efficient migration ofprecursors into the OB during this most intense period of migration.The constant requirement of NTN1 during the postnatal precursormigration period is illustrated by the observation that no difference inthe size of the control and Ntn1−/− glomeruli can be observed at Pn1(Suppl. Figs. 3A–D), whilst at Pn16, a clear reduction in the size of theNtn1−/− glomeruli is detected (Figs. 4H–S).
w of Pn16 wild type and Ntn1−/− brains. The bars indicate the rostro-caudal length oftal paraffin sections through the OB of wild type (B) and Ntn1−/− (C) mice. The barsF) and Ntn1−/− (E,G) olfactory bulbs. Examples of the areas (100 μm×500 μm) whereal staining with anti-calR (H,J), anti-calB (L,N) and anti-TH (P,R) antibodies on paraffins are shown in I,K,M,O,Q,S. Scale bars B,C 100 μm, D–G 50 μm, H–S 20 μm. epl, externalm layer; mi, mitral cell layer; nl, neuronal layer; ob, olfactory bulb; VZ, ventricular zone.
108 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
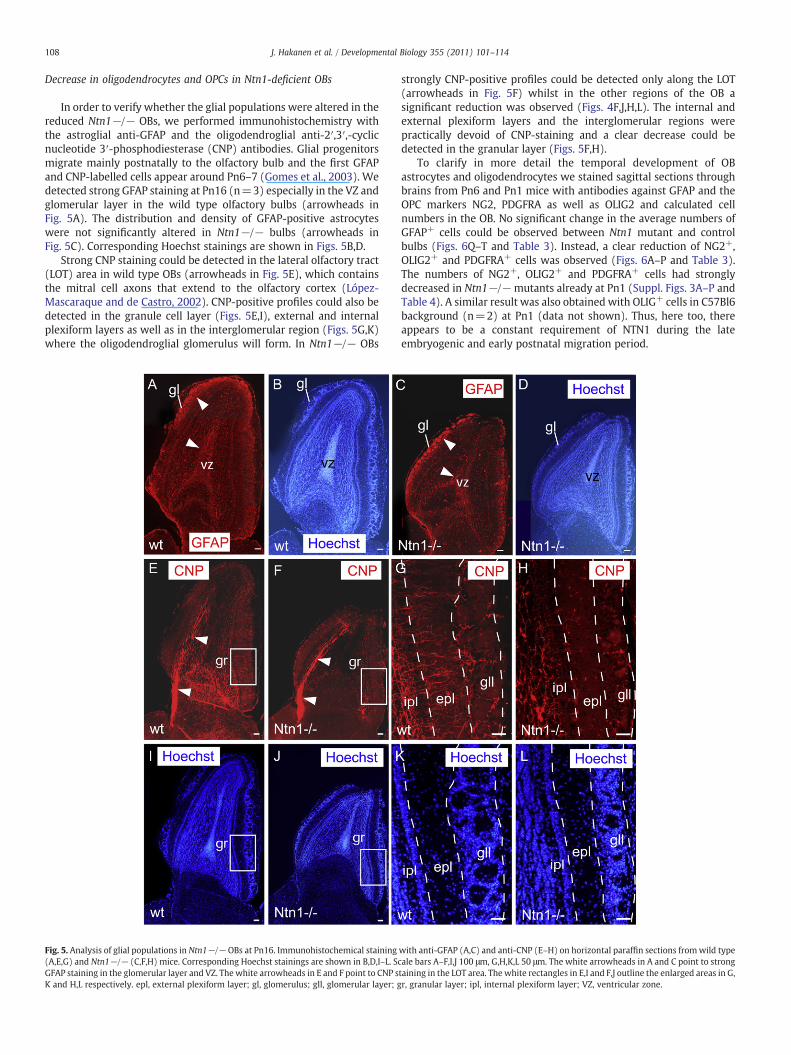
Decrease in oligodendrocytes and OPCs in Ntn1-deficient OBs
In order to verify whether the glial populations were altered in thereduced Ntn1−/− OBs, we performed immunohistochemistry withthe astroglial anti-GFAP and the oligodendroglial anti-2′,3′,-cyclicnucleotide 3′-phosphodiesterase (CNP) antibodies. Glial progenitorsmigrate mainly postnatally to the olfactory bulb and the first GFAPand CNP-labelled cells appear around Pn6–7 (Gomes et al., 2003). Wedetected strong GFAP staining at Pn16 (n=3) especially in the VZ andglomerular layer in the wild type olfactory bulbs (arrowheads inFig. 5A). The distribution and density of GFAP-positive astrocyteswere not significantly altered in Ntn1−/− bulbs (arrowheads inFig. 5C). Corresponding Hoechst stainings are shown in Figs. 5B,D.
Strong CNP staining could be detected in the lateral olfactory tract(LOT) area in wild type OBs (arrowheads in Fig. 5E), which containsthe mitral cell axons that extend to the olfactory cortex (López-Mascaraque and de Castro, 2002). CNP-positive profiles could also bedetected in the granule cell layer (Figs. 5E,I), external and internalplexiform layers as well as in the interglomerular region (Figs. 5G,K)where the oligodendroglial glomerulus will form. In Ntn1−/− OBs
Fig. 5. Analysis of glial populations in Ntn1−/− OBs at Pn16. Immunohistochemical staining(A,E,G) and Ntn1−/− (C,F,H) mice. Corresponding Hoechst stainings are shown in B,D,I–L. ScGFAP staining in the glomerular layer and VZ. The white arrowheads in E and F point to CNP sK and H,L respectively. epl, external plexiform layer; gl, glomerulus; gll, glomerular layer; g
strongly CNP-positive profiles could be detected only along the LOT(arrowheads in Fig. 5F) whilst in the other regions of the OB asignificant reduction was observed (Figs. 4F,J,H,L). The internal andexternal plexiform layers and the interglomerular regions werepractically devoid of CNP-staining and a clear decrease could bedetected in the granular layer (Figs. 5F,H).
To clarify in more detail the temporal development of OBastrocytes and oligodendrocytes we stained sagittal sections throughbrains from Pn6 and Pn1 mice with antibodies against GFAP and theOPC markers NG2, PDGFRA as well as OLIG2 and calculated cellnumbers in the OB. No significant change in the average numbers ofGFAP+ cells could be observed between Ntn1 mutant and controlbulbs (Figs. 6Q–T and Table 3). Instead, a clear reduction of NG2+,OLIG2+ and PDGFRA+ cells was observed (Figs. 6A–P and Table 3).The numbers of NG2+, OLIG2+ and PDGFRA+ cells had stronglydecreased in Ntn1−/− mutants already at Pn1 (Suppl. Figs. 3A–P andTable 4). A similar result was also obtained with OLIG+ cells in C57Bl6background (n=2) at Pn1 (data not shown). Thus, here too, thereappears to be a constant requirement of NTN1 during the lateembryogenic and early postnatal migration period.
with anti-GFAP (A,C) and anti-CNP (E–H) on horizontal paraffin sections fromwild typeale bars A–F,I,J 100 μm, G,H,K,L 50 μm. The white arrowheads in A and C point to strongtaining in the LOT area. The white rectangles in E,I and F,J outline the enlarged areas in G,r, granular layer; ipl, internal plexiform layer; VZ, ventricular zone.
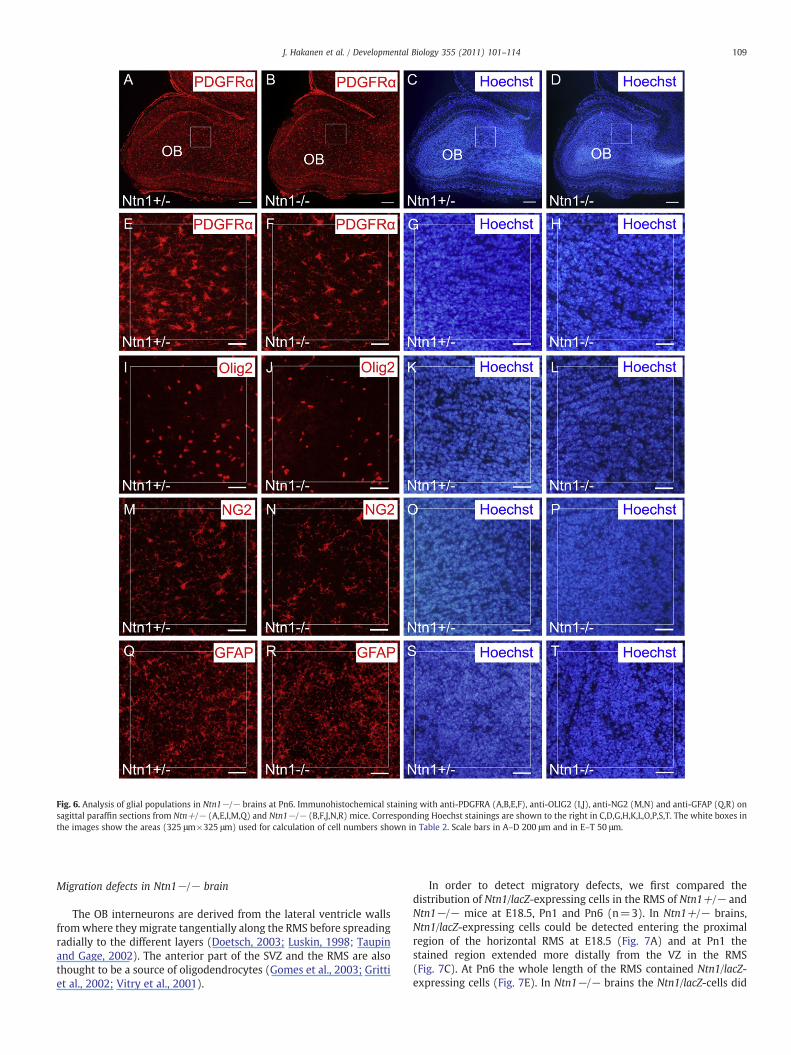
Fig. 6. Analysis of glial populations in Ntn1−/− brains at Pn6. Immunohistochemical staining with anti-PDGFRA (A,B,E,F), anti-OLIG2 (I,J), anti-NG2 (M,N) and anti-GFAP (Q,R) onsagittal paraffin sections from Ntn+/− (A,E,I,M,Q) and Ntn1−/− (B,F,J,N,R) mice. Corresponding Hoechst stainings are shown to the right in C,D,G,H,K,L,O,P,S,T. The white boxes inthe images show the areas (325 μm×325 μm) used for calculation of cell numbers shown in Table 2. Scale bars in A–D 200 μm and in E–T 50 μm.
109J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
Migration defects in Ntn1−/− brain
The OB interneurons are derived from the lateral ventricle wallsfromwhere theymigrate tangentially along the RMS before spreadingradially to the different layers (Doetsch, 2003; Luskin, 1998; Taupinand Gage, 2002). The anterior part of the SVZ and the RMS are alsothought to be a source of oligodendrocytes (Gomes et al., 2003; Grittiet al., 2002; Vitry et al., 2001).
In order to detect migratory defects, we first compared thedistribution of Ntn1/lacZ-expressing cells in the RMS of Ntn1+/− andNtn1−/− mice at E18.5, Pn1 and Pn6 (n=3). In Ntn1+/− brains,Ntn1/lacZ-expressing cells could be detected entering the proximalregion of the horizontal RMS at E18.5 (Fig. 7A) and at Pn1 thestained region extended more distally from the VZ in the RMS(Fig. 7C). At Pn6 the whole length of the RMS contained Ntn1/lacZ-expressing cells (Fig. 7E). In Ntn1−/− brains the Ntn1/lacZ-cells did
Table 3Analysis of cell numbers in OBs of wild type and Ntn1−/− mice at Pn6. The averagenumbers of total granular cells, granular NeuN+, NG2+, OLIG2+, PDGFRA+ and GFAP+
cells/area (325 μm×325 μm, for NeuN+ 100 μm×100 μm) are shown with ±SEM andthe decrease in the numbers observed in Ntn1−/− compared to wild types is indicatedas %.
Cell type at Pn6 Nr. of cells/areaNtn1+/−
Nr. of cells/areaNtn1−/−
Change inNtn1−/−
P-value
Granular total 1230.9±35.8 1094.7±29.1 −11% b0.05Granular NeuN+ 55.2±1.3 37±1.2 −33% b0.001NG2+ 66.7±1.3 43.1±2.3 −35% b0.001OLIG2+ 47.7±2.5 33.4±2.7 −30% b0.05PDGFRA+ 58±2.4 43.4±1.4 −25% b0.01GFAP+ 66.2±5.1 61±5.7 No 0.53
110 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
not reach as far distally from the VZ as in the controls at all stagesanalysed (Figs. 7B,D,F). In contrast, especially seen at Pn1, the Ntn1/lacZ-cells in Ntn1−/− brains were spread in a more broad area in theproximal part of the horizontal RMS close to the VZ (indicated witharrowheads in Fig. 7D) compared to the cells in Ntn1+/− brains(arrowheads in Fig. 7C). At Pn6 the β-gal-stained cells did not reachthe OB as in the controls (arrowheads in Figs. 7E,F). Thus, withoutsufficient amounts of NTN1, the Ntn1/lacZ-expressing cells accumu-lated in the proximal RMS and their migration further towards the OBappeared inefficient.
To test the migratory behaviour of the CNS precursors in theanterior forebrain we cultured VZ/SVZ-RMS/OB explants from wildtype, Ntn1+/− and Ntn1−/− E18.5 embryos and stained them withX-gal to visualiseNtn1/lacZ-expressing cells. Three parts from the areawere cut out for explants as indicated in Figs. 7A,B with dashed whitelines and pieces a, b and c were cultured. C-explants from allgenotypes showed considerable out-migration as networks or chains(Figs. 7G–L and data not shown) whilst a- and b-explants did notshow significant outgrowth (data not shown). No difference betweenthe out-migration from wild type and Ntn1+/− explants could beobserved (data not shown).
When the average lengths of 8 outgrown chains/explant from 6explants/genotype were measured in Ntn1+/− and Ntn1−/−samples, no statistically significant difference in their length wasobserved (student's t-test P=0.2; Figs. 7G–I). Instead, a significant24% reduction in the migration distance of the Ntn1/lacZ-expressingcells from the explants was observed between Ntn1+/− andNtn1−/− cultures (Pb0.05*; Figs. 7J–L). These observations suggestthat NTN1 is required for the migration of CNS precursors out of theexplants.
To analyse in more detail the brain region where the Ntn1/lacZexpressing cells accumulated inNtn1−/−mice, we also calculated thecell numbers in three areas in the proximal RMS region at Pn1(indicated as white boxes a, b and c in Figs. 8A,B). In the ventral part ofthe proximal RMS area (in box c) of the Ntn1−/− samples, more cellswere present than in the control samples. Instead, the cell numberswere significantly reduced in more distal and dorsal areas of the RMS
Table 4Analysis of oligodendroglial cell numbers in OBs of wild type and Ntn1−/−mice at Pn1.The average numbers of total, NG2+, OLIG2+ and PDGFRA+ cells/area(200 μm×200 μm) are shown with±SEM and the decrease in the numbers observedin Ntn1−/− compared to wild types is indicated as %.
Cell typeat Pn1
Nr. of cells/areaNtn1+/−
Nr. of cells/areaNtn1−/−
Change inNtn1−/−
P-value
Total 674.6±7.3 607.1±1.2 −10% b0.001NG2+ 35.6±1.9 24±1.2 −33% b0.01OLIG2+ 18.6±0.3 12.2±1.1 −34% b0.01PDGFRA+ 27.3±0.7 19.1±1.4 −30% b0.01
(boxes a and b, respectively) of the Ntn1−/− animals compared tothe controls (Fig. 8C) further demonstrating that a reduced number ofprecursor cells succeeded in migrating distally into the horizontalRMS in Ntn1−/− mice.
In order to verify whether the Ntn1−/− cells that remainectopically in the proximal area of the horizontal RMS undergoexcess of programmed cell death we performed TUNEL analyses onsections prepared from the anterior forebrain region at Pn1. Wealso calculated the proportion of TUNEL positive cells from all cells(nuclei) in the proximal RMS area. No statistically significantdifference could be observed in the average numbers of TUNELpositive cells (8.3±2.0 cells/area in Ntn1+/− and 8.4±1.4 cells/areainNtn1−/−) or in the proportions of TUNEL positive cells (0.45±0.1%in Ntn1+/− and 0.45±0.01% in Ntn1−/−) between the samples(P=0.9; Suppl. Figs. 5A–D). This suggests that the cells thataccumulate in the proximal RMS do not immediately die throughapoptosis.
Discussion
Ntn1-expressing cells from embryonic VZ and adult ependyma formmultipotent neurospheres
The regulated production of neural and glial precursor cells in thegerminal regions is essential for CNS development. In embryonic andneonatal mice, the main germinal zone, the VZ, contains proliferatingneural stem cells. During the first weeks of life these cells disappeargradually and give rise to the adult ventricular wall cell typesincluding the non-proliferative ependymal cells and the GFAP+
astrocyte-type cells that touch the ventricle and are thought to bethe postnatal neural stem cells (Spassky et al., 2005; Tramontin et al.,2003). A second germinal zone, the SVZ, develops later duringembryonic stages beneath the lateral ventricle VZ and participates inthe expansion of the forebrain (Conover and Allen, 2002). Duringembryonic forebrain development Ntn1 is strongly expressed in theventral parts of the VZ whereas very little, if any expression can bedetected in the adjacent proliferative area, the SVZ (this work andHamasaki et al., 2001; Kawasaki et al., 2006; Serafini et al., 1996). Inthe neurogenic niches of adult brain and spinal cord, Ntn1 expressionis conserved exclusively in the ependymal cells.
Ntn1 expression could be detected also in both embryonic andadult neurosphere cultures. By selecting proliferating neurospheresaccording to Ntn1 expression we showed that this subset of cells wasmultipotent and self-renewing on its own. However, we show thatNTN1 protein function is not critical for the formation of neurospheresor for the self-renewal and differentiation of neural stem/progenitorcells in culture.
NTN1 is required for the efficient migration of OB progenitors/precursorsinto the RMS
The migration of neural and glial precursors to their correctlocations for the final differentiation and innervations has to bestrictly controlled in a temporal and spatial manner and migratorydefects may be involved in diseases such as mental retardation,epilepsy, multiple sclerosis or glial cancers (de Castro and Bribian,2005; Marin and Rubenstein, 2003). Interestingly, long-range attrac-tive signals from OB seem not to be required for the directedmigration of precursors since at least in adult rodents, neuralprecursors are able to enter the RMS and move forward also in thecomplete absence of OB (Jankovski et al., 1998; Kirschenbaum et al.,1999). Instead, a combination of SLIT-mediated repulsion togetherwith motility increasing factors such as astrocyte-derived migration-inducing activity (MIA), PSA-NCAM and Eph/ephrin are thought to bethe main forces behind precursor migration (Ghashghaei et al., 2007;Marin and Rubenstein, 2003). However, rather mild or no defects
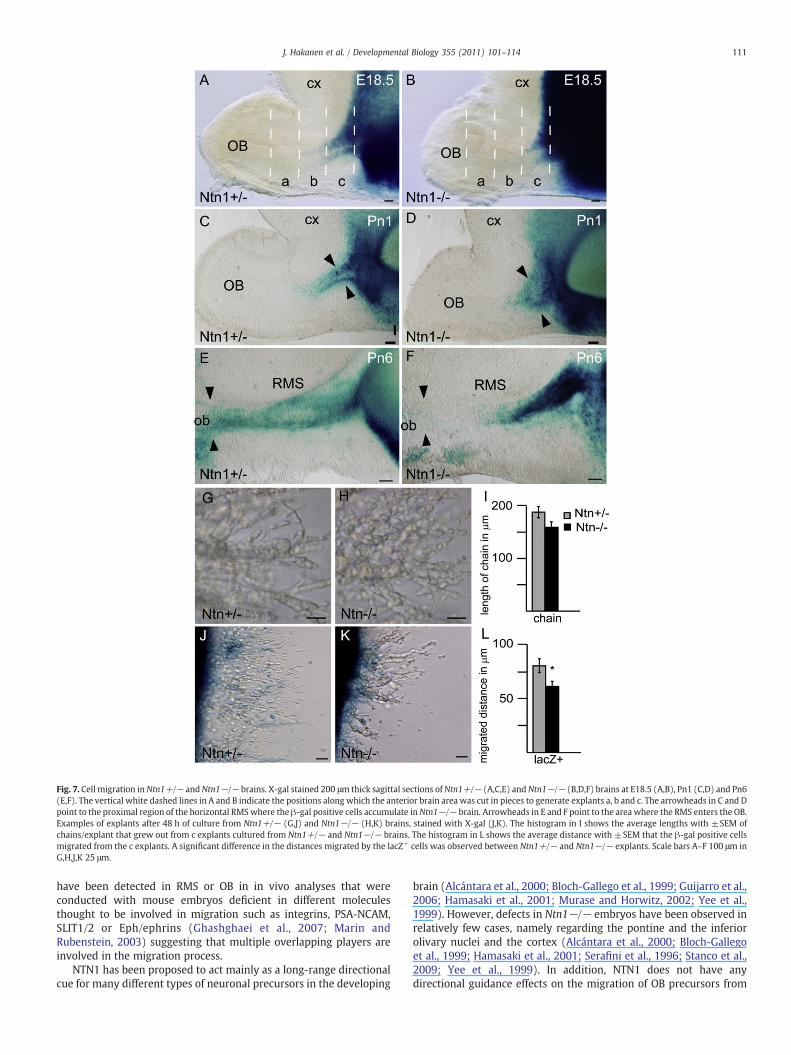
Fig. 7. Cell migration inNtn1+/− andNtn1−/− brains. X-gal stained 200 μm thick sagittal sections ofNtn1+/− (A,C,E) andNtn1−/− (B,D,F) brains at E18.5 (A,B), Pn1 (C,D) and Pn6(E,F). The vertical white dashed lines in A and B indicate the positions along which the anterior brain area was cut in pieces to generate explants a, b and c. The arrowheads in C and Dpoint to the proximal region of the horizontal RMSwhere the β-gal positive cells accumulate inNtn1−/− brain. Arrowheads in E and F point to the area where the RMS enters the OB.Examples of explants after 48 h of culture from Ntn1+/− (G,J) and Ntn1−/− (H,K) brains, stained with X-gal (J,K). The histogram in I shows the average lengths with ±SEM ofchains/explant that grew out from c explants cultured from Ntn1+/− and Ntn1−/− brains. The histogram in L shows the average distance with ±SEM that the β-gal positive cellsmigrated from the c explants. A significant difference in the distances migrated by the lacZ+ cells was observed between Ntn1+/− and Ntn1−/− explants. Scale bars A–F 100 μm inG,H,J,K 25 μm.
111J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
have been detected in RMS or OB in in vivo analyses that wereconducted with mouse embryos deficient in different moleculesthought to be involved in migration such as integrins, PSA-NCAM,SLIT1/2 or Eph/ephrins (Ghashghaei et al., 2007; Marin andRubenstein, 2003) suggesting that multiple overlapping players areinvolved in the migration process.
NTN1 has been proposed to act mainly as a long-range directionalcue for many different types of neuronal precursors in the developing
brain (Alcántara et al., 2000; Bloch-Gallego et al., 1999; Guijarro et al.,2006; Hamasaki et al., 2001; Murase and Horwitz, 2002; Yee et al.,1999). However, defects in Ntn1−/− embryos have been observed inrelatively few cases, namely regarding the pontine and the inferiorolivary nuclei and the cortex (Alcántara et al., 2000; Bloch-Gallegoet al., 1999; Hamasaki et al., 2001; Serafini et al., 1996; Stanco et al.,2009; Yee et al., 1999). In addition, NTN1 does not have anydirectional guidance effects on the migration of OB precursors from

Fig. 8. Calculation of cell numbers in the proximal RMS at Pn1. Cell numbers werecalculated in Hoechst stained paraffin sections from Ntn1+/− (A) and Ntn1−/−(B) mice. Three regions (150 μm×150 μm) were used for analysis indicated with boxesand marked with a, b and c. (C) The average number of cells/section in areas a, b and care shown in histograms with ±SEM. Scale bars 50 μm.
112 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
anterior brain explants (Hu and Rutishauser, 1996; Mason et al.,2001).
We propose here a novel role for NTN1 in the developing forebrainin the initiation of tangential migration of Ntn1-expressing OBprecursors for neurons and oligodendrocytes into the RMS. Ntn1 isexpressed in the VZ and its rostral extension and without NTN1 thesecells accumulate in the rostral extension/proximal RMS regionresulting in fewer cells in the more distal RMS at early postnatalstages when compared to controls. Thus, the progress of Ntn1/lacZcells further into the RMS is inefficient in Ntn1−/−mice. Similarly, inNtn1−/− explant cultures a specific migration defect of the Ntn1/lacZexpressing cells was observed when compared to Ntn1+/− explantswhereas the overall migration activity had not changed in theseexplants.
The inefficient entrance of the Ntn1/lacZ expressing cells into theRMS may be due to deficient detachment/release of these cells fromthe VZ/rostral extension area. A similar problem has been describedbefore in the embryonic Ntn1−/− spinal cord where the detachmentand/or dispersal of the OPCs from the Ntn1 expressing ventral VZwas inefficient (Jarjour et al., 2003; Tsai et al., 2006). Moreover,NTN1 has a local release function also in inner ear epitheliumwhere the normally Ntn1-expressing cells cannot detach from theunderlying basement membrane in Ntn1−/− embryos (Matilainenet al., 2007; Salminen et al., 2000). Similar “short-range” functionsfor NTN1 have been reported in various tissues where NTN1 mediatescell adhesion/migration and organ morphogenesis (Baker et al.,2006). Thus, in addition to its traditional role as a diffusible long-range guidance molecule, NTN1 may function in CNS also in avery local cell-autonomous or autocrine manner, influencing themovement/detachment/release of specific cells that produce it.Interestingly, an autocrine function of NTN1 has been proposed topromote also oligodendrocyte maturation (Rajasekharan et al., 2009).Furthermore, a cell-autonomous function has been suggested forSLIT1 in migrating neuroblasts in addition to its long-range chemor-epulsive function (Nguyen-Ba-Charvet et al., 2004) suggesting thatmany secreted guidance molecules may adopt various functionsduring development.
Reduction of granular cells and calR+ and calB+ glomerularinterneurons in Ntn1−/− OB
The OB interneuron precursors are born in the walls of theforebrain lateral ventricles fromwhere they migrate through the RMSto generate granular and periglomerular cells (Doetsch, 2003; Hinds,1968; Luskin, 1993; Taupin and Gage, 2002). Interestingly, there areparticular subregions in the lateral ventricle wall which contribute tospecific interneuron subtypes (De Marchis et al., 2007; Kohwi et al.,2007; Merkle et al., 2007; Stenman et al., 2003; Toresson andCampbell, 2001; Ventura and Goldman, 2007; Waclaw et al., 2006).The dorsolateral wall (dLGE) produces mainly superficial granule cellsas well as TH+ periglomerular type I neurons (Kohwi et al., 2005;Kohwi et al., 2007; Lledo et al., 2008; Long et al., 2003; Yun et al.,2003) whilst the ventrolateral wall (vLGE and MGE) gives rise tomainly deep granule cells and calB+ periglomerular neurons(Toresson and Campbell, 2001; Yun et al., 2003). The calR+
periglomerular neurons are not produced by the lateral wall, butinstead originate from dorsal and ventral subregions, including thepallium (cortical) wall, themedial wall (septum) and the RMS (Kohwiet al., 2007; Lledo et al., 2008).
According to the strong expression of Ntn1 in the ventralsubregions of the lateral ventricle walls as well as in the ventralseptum wall and in the proximal RMS, we observed a significantdecrease in the numbers of deep granule cells as well as in calR+ andcalB+ glomerular cells in the Ntn1−/− OB. In contrast, the number ofthe exclusively dorsally derived TH+ cells had not changed. Theseobservations suggest that Ntn1-expressing cells in the ventral VZ/proximal RMS produce specific types of neural precursors into the OBand that in the absence of NTN1 these cells are not able to populatethe OB efficiently.
Reduction of a subpopulation of oligodendrocytes in Ntn1−/− OBs
The OPCs are heterogeneous and have multiple origins inmammalian forebrain. Most OPCs originate in the MGE and somealso in the LGE and anterior entopeduncular area (Jarjour andKennedy, 2004; Kessaris et al., 2006; Spassky et al., 2001). Very littleis known of how the OPCs populate the OB during development andwhichmechanisms control this process. The OB oligodendrocytesmayrepresent two distinct lineages with different origins: the early Plp1-expressing lineage and the later, Pdgfra- and Olig1/2-expressinglineage which is thought to originate in the MGE (Spassky et al.,2001). In addition, the RMS may also contain additional “intrinsic”glial progenitors during embryogenesis (Gritti et al., 2002; Reid et al.,1999; Spassky et al., 2001).
Ntn1 is strongly expressed in the VZ of MGE and vLGE but itsexpression is rapidly downregulated in cells that leave the VZincluding the OPCs, but is induced again later in mature oligoden-drocytes in adult CNS (Manitt et al., 2001; Rajasekharan et al., 2009).We did not detect Ntn1mRNA or Ntn1/lacZ expression in the OB VZ atE14.5 (data not shown) and therefore, Ntn1 is not likely to contributeto the Plp1 expressing early lineage. Instead, Ntn1 expression can bedetected in the rostral extension of the ventricular VZ at E18.5 and atPn6 Ntn1 expression is detected in NG2+ cells migrating in the RMSand thus, Ntn1 could contribute to the later oligodendrocytic lineage.At Pn16, a large portion of the OB oligodendrocytes weremissing fromthe Ntn1−/− outer and granular layers. Instead, no clear change wasobserved in the population myelinating the LOT. These observationsmay indicate that the early OPCs could be the main lineagecontributing to LOT myelination whereas the Ntn1-expressing later-appearing lineage would mainly contribute to myelinate the granularand the outer layers of the OB.
In contrast to the oligodendroglial lineage, no significant decreasein the numbers of GFAP+ astrocytes was detected in Ntn1−/− OBs.A similar observation has been made earlier concerning the spinal

113J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
cord (Tsai et al., 2006). Thus, although Ntn1/lacZ is expressed in someGFAP+ cells in the RMS it may be that most OB astrocytes are derivedfrom progenitors that do not express Ntn1, residing for example in theOB itself (Fukushima et al., 2002) or in the posterior SVZ (Law et al.,1999).
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.ydbio.2011.04.016.
Acknowledgments
This study was funded by the Finnish Academy, the Sigrid JuséliusFoundation, by the Finnish Cultural Foundation fellowships to S.D andJ.H and by the Ella and Georg Ehrnrooth Foundation to J.H. We aregrateful to Kirmo Wartiovaara, Juha Partanen and Jyrki Kukkonen fordiscussions and critical reading of the manuscript, to Raija Savolainenfor expert technical assistance, and to Kimmo Tanhuanpää and MikaMolin for help with the confocal analyses.
References
Aguirre,A.,Gallo,V., 2004. Postnatal neurogenesisandgliogenesis in theolfactorybulb fromNG2-expressing progenitors of the subventricular zone. J. Neurosci. 24, 10530–10541.
Alcántara, S., Ruiz, M., De Castro, F., Soriano, E., Sotelo, C., 2000. Netrin 1 acts as anattractive or as a repulsive cue for distinct migrating neurons during thedevelopment of the cerebellar system. Development 127, 1359–1372.
Baker, K.A., Moore, S.W., Jarjour, A.A., Kennedy, T.E., 2006. When a diffusible axonguidance cue stops diffusing: roles for netrins in adhesion and morphogenesis.Curr. Opin. Neurobiol. 16, 529–534.
Barallobre, M.J., Pascual, M., Del Rio, J.A., Soriano, E., 2005. The Netrin family of guidancefactors: emphasis on Netrin-1 signalling. Brain Res. Brain Res. Rev. 49, 22–47.
Batista-Brito, R., Close, J., Machold, R., Fishell, G., 2008. The distinct temporal origins ofolfactory bulb interneuron subtypes. J. Neurosci. 28, 3966–3975.
Bloch-Gallego, E., Ezan, F., Tessier-Lavigne, M., Sotelo, C., 1999. Floor plate and Netrin-1are involved in the migration and survival of inferior olivary neurons. J. Neurosci.19, 4407–4420.
Cecconi, F., Roth, K., Dolgov, O., Munarriz, E., Anokhin, K., Gruss, P., Salminen, M., 2004.Apaf1 dependent programmed cell death is required for inner ear morphogenesisand growth. Development 131, 2125–2135.
Chazal, G., Durbec, P., Jankovski, A., Rougon, G., Cremer, H., 2000. Consequences ofneural cell adhesion molecule deficiency on cell migration in the rostral migratorystream of the mouse. J. Neurosci. 20, 1446–1457.
Chong, S.Y.C., Chan, J.R., 2010. Tapping into the glial reservoir: cells committed toremaining uncommitted. J. Cell Biol. 188, 305–312.
Conover, J.C., Allen, R.L., 2002. The subventricular zone: new molecular and cellulardevelopments. Cell. Mol. Life Sci. 59, 2128–2135.
Coskun, V., Luskin, M.B., 2002. Intrinsic and extrinsic regulation of the proliferation anddifferentiation of cells in the rodent rostral migratory stream. J. Neurosci. Res. 69,795–802.
de Castro, F., Bribian, A., 2005. The molecular orchestra of the migration ofoligodendrocyte precursors during development. Brain Res. Rev. 49, 227–241.
De Marchis, S., Bovetti, S., Carletti, B., Hsieh, Y.C., Garzotto, D., Peretto, P., Fasolo, A.,Puche, A.C., Rossi, F., 2007. Generation of distinct types of periglomerular olfactorybulb interneurons during development and in adult mice: implication for intrinsicproperties of the subventricular zone progenitor population. J. Neurosci. 27,657–664.
Doetsch, F., 2003. The glial identity of neural stem cells. Nat. Neurosci. 6, 1127–1134.Friedrich, G., Soriano, P., 1991. Promoter traps in embryonic stem cells: a genetic screen
to identify and mutate developmental genes in mice. Genes Dev. 5, 1513–1523.Fukushima, N., Yokouchi, K., Kawagishi, K., Moriizumi, T., 2002. Differential neurogen-
esis and gliogenesis by local and migrating neural stem cells in the olfactory bulb.Neurosci. Res. 44, 467–473.
Ghashghaei, H.T., Lai, C., Anton, E.S., 2007. Neuronal migration in the adult brain: are wethere yet? Nat. Rev. 8, 141–151.
Gomes, S.S., Carvalho, S.L., Santiago, M.F., Lopez, L.B., Barradas, P.C., Cavalcante, L.A.,2003. Expression of 2′,3′-cyclic nucleotide 3′-phosphodiesterase (CNPase) in thedeveloping olfactory bulb and subventricular zone rostral extension. J. Neurosci.Res. 73, 471–480.
Gritti, A., Bonfanti, L., Doetsch, F., Caille, I., Alvarez-Buylla, A., Lim, D.A., Galli, R., GarciaVerdugo, J.M., Herrera, D.G., Vescovi, A.L., 2002. Multipotent neural stem cellsreside into the rostral extension and olfactory bulb of adult rodents. J. Neurosci. 22,437–445.
Guijarro, P., Simó, S., Pascual, M., Abasolo, I., Del Rio, J.A., Soriano, E., 2006. Netrin1exerts a chemorepulsive effect on migrating cerebellar interneurons in a Dcc-independent way. Mol. Cell. Neurosci. 33, 389–400.
Hack, I., Bancila, M., Loulier, K., Carroll, P., Cremer, H., 2002. Reelin is a detachmentsignal in tangential chain-migration during postnatal neurogenesis. Nat. Neurosci.5, 939–945.
Hamasaki, T., Goto, S., Nishikawa, S., Ushio, Y., 2001. A role of Netrin-1 in the formationof the subcortical structure striatum: repulsive action on the migration of late-bornstriatal neurons. J. Neurosci. 21, 4272–4280.
Hedgecock, E.M., Norris, C.R., 1997. Netrins evoke mixed reactions in motile cells.Trends Genet. 13, 251–253.
Hinds, J.W., 1968. Autoradiographic study of histogenesis in the mouse olfactory bulb. I.Time of origin of neurons and glia. J. Comp. Neurol. 134, 287–304.
Hogan, B., Beddington, R., Constantini, F., Lacy, E., 1994. Manipulating the MouseEmbryo, second ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor.
Hu, H., Rutishauser, U., 1996. A septum-derived chemorepulsive factor for migratingolfactory interneuron precursors. Neuron 16, 933–940.
Hunyady, B., Krempels, K., Harta, G., Mezey, E., 1996. Immunohistochemical signalamplification by catalyzed reporter deposition and its application in doubleimmunostaining. J. Histochem. Cytochem. 44, 1353–1362.
Jankovski, A., Garcia, C., Soriano, E., Sotelo, C., 1998. Proliferation, migration anddifferentiation of neuronal progenitor cells in the adult mouse subventricular zonesurgically separated from its olfactory bulb. Eur. J. Neurosci. 10, 3853–3868.
Jarjour, A.A., Kennedy, T.E., 2004. Oligodendrocyte precursors on the move: mecha-nisms directing migration. Neuroscientist 10, 99–105.
Jarjour, A.A., Manitt, C., Moore, S.W., Thompson, K.M., Yuh, S.J., Kennedy, T.E., 2003.Netrin-1 is a chemorepellent for oligodendrocyte precursor cells in the embryonicspinal cord. J. Neurosci. 23, 3735–3744.
Johansson, C.B., Momma, S., Clarke, D.L., Risling, M., Lendahl, U., Frisen, J., 1999.Identification of a neural stem cell in the adult mammalian central nervous system.Cell 96, 25–34.
Kawasaki, T., Ito, K., Hirata, T., 2006. Netrin 1 regulates ventral tangential migration ofguidepost neurons in the lateral olfactory tract. Development 133, 845–853.
Kessaris, N., Fogarty, M., Iannarelli, P., Grist, M., Wegner, M., Richardson, W.D., 2006.Competing waves of oligodendrocytes in the forebrain and postnatal elimination ofan embryonic lineage. Nat. Neurosci. 9, 173–179.
Kirschenbaum, B., Doetsch, F., Lois, C., Alvarez-Buylla, A., 1999. Adult subventricularzone neuronal precursors continue to proliferate and migrate in the absence of theolfactory bulb. J. Neurosci. 19, 2171–2180.
Kohwi, M., Osumi, N., Rubestein, J.L.R., Alvarez-Buylla, A., 2005. Pax6 is required formaking specific subpopulations of granule and periglomerular neurons in theolfactory bulb. J. Neurosci. 25, 6997–7003.
Kohwi, M., Petryniak, M.A., Long, J.E., Ekker, M., Obata, K., Yanagawa, Y., Rubenstein,J.L.R., Alvarez-Buylla, A., 2007. A subpopulation of olfactory bulb GABAergicinterneurons is derived from Emx1- and Dlx5/6-expressing progenitors. J. Neurosci.27, 6878–6891.
Kosaka, K., Toida, K., Aika, Y., Kosaka, T., 1998. How simple is the organisation of theolfactory glomerulus? The heterogeneity of the so-called periglomerular cells.Neurosci. Res. 30, 101–110.
Law, A.K., Pencea, V., Buck, C.R., Luskin, M.B., 1999. Neurogenesis and neuronalmigration in the neonatal rat forebrain anterior subventricular zone do not requireGFAP-positive astrocytes. Dev. Biol. 216, 622–634.
Le Bras, B., Chatzopoulou, E., Heydon, K., Martinez, S., Ikenaka, K., Prestoz, L., Spassky, N.,Zalc, B., Thomas, J.L., 2005. Oligodendrocyte development in the embryonic brain:the contribution of the plp lineage. Int. J. Dev. Biol. 49, 209–220.
Lilleväli, K., Haugas, M., Matilainen, T., Pussinen, C., Karis, A., Salminen, M., 2006. Gata3is required for early morphogenesis and Fgf10 expression during otic development.Mech. Dev. 123, 415–429.
Lledo, P.M., Merkle, F.T., Alvarez-Buylla, A., 2008. Origin and function of olfactory bulbinterneuron diversity. Trends Neurosci. 31, 392–400.
Long, J.E., Garel, S., Depew, M.J., Tobet, S., Rubenstein, J.L.R., 2003. DLX5 regulatesdevelopment of peripheral and central components of the olfactory system.J. Neurosci. 23, 568–578.
López-Mascaraque, L., de Castro, F., 2002. The olfactory bulb as an independentdevelopmental domain. Cell Death Differ. 9, 1279–1286.
Luskin, M.B., 1993. Restricted proliferation and migration of postnatally generatedneurons derived from the forebrain subventricular zone. Neuron 11, 173–189.
Luskin, M.B., 1998. Neuroblasts of the postnatal mammalian forebrain: their phenotypeand fate. J. Neurobiol. 36, 221–233.
Manitt, C., Colicos, M.A., Thompson, K.M., Rousselle, E., Peterson, A.C., Kennedy, T.E.,2001. Widespread expression of Netrin-1 by neurons and oligodendrocytes in theadult mammalian spinal cord. J. Neurosci. 21, 3911–3922.
Marin, O., Rubenstein, J.L.R., 2003. Cell migration in the forebrain. Annu. Rev. Neurosci.26, 441–483.
Mason, H.A., Ito, S., Corfas, G., 2001. Extracellular signals that regulate the tangentialmigration of olfactory bulb neuronal precursors: Inducers, inhibitors, and re-pellents. J. Neurosci. 21, 7654–7663.
Matilainen, T., Haugas, M., Kreidberg, J.A., Salminen, M., 2007. Analysis of Netrin1receptors during inner ear development. Int. J. Dev. Biol. 51, 409–413.
Merkle, F.T., Mirzadeh, Z., Alvarez-Buylla, A., 2007. Mosaic organisation of neural stemcells in the adult brain. Science 317, 381–384.
Miller, R.H., 2002. Regulation of oligodendrocyte development in the vertebrate CNS.Progr. Neurobiol. 67, 451–467.
Mullen, R.J., Buck, C.R., Smith, A.M., 1992. NeuN, a neuronal specific nuclear protein invertebrates. Development 116, 201–211.
Murase, S., Horwitz, A.F., 2002. Deleted in colorectal carcinoma and differentiallyexpressed integrins mediate the directional migration of neural precursors in therostral migratory stream. J. Neurosci. 22, 3568–3579.
Nguyen-Ba-Charvet, K.T., Picard-Riera, N., Tessier-Lavigne, M., Baron-Van Evercooren,A., Sotelo, C., Chédotal, A., 2004. Multiple roles for slits in the control of cellmigration in the rostral migratory stream. J. Neurosci. 24, 1497–1506.
Pencea, V., Luskin, M.B., 2003. Prenatal development of the rodent rostral migratorystream. J. Comp. Neurol. 463, 402–418.
Qi, Y., Stapp, D., Qiu, M., 2002. Origin andmolecular specification of oligodendrocytes inthe telencephalon. Trends Neurosci. 25, 223–225.

114 J. Hakanen et al. / Developmental Biology 355 (2011) 101–114
Rajasekharan, S., Baker, K.A., Horn, K.E., Jarjour, A.A., Antel, J.P., Kennedy, T.E., 2009.Netrin 1 and Dcc regulate oligodendrocyte process branching and membraneextension via Fyn and RhoA. Development 136, 415–426.
Reid, C.B., Liang, I., Walsh, C.A., 1999. Clonal mixing, clonal restriction, and specification ofcell types in the developing rat olfactory bulb. J. Comp. Neurol. 403, 106–118.
Rousselot, P., Lois, C., Alvarez-Buylla, A., 1995. Embryonic (PSA) N-CAM reveals chainsof migrating neuroblasts between the lateral ventricle and the olfactory bulb ofadult mice. J. Comp. Neurol. 351, 51–61.
Salminen, M., Meyer, B.I., Bober, E., Gruss, P., 2000. Netrin 1 is required for semicircularcanal formation in the mouse inner ear. Development 127, 13–22.
Serafini, T., Colamarino, S.A., Leonardo, E.D., Wang, H., Beddington, R., Skarnes, W.C.,Tessier-Lavigne, M., 1996. Netrin-1 is required for commissural axon guidance inthe developing vertebrate nervous system. Cell 87, 1001–1014.
Spassky, N., Heydon, K., Mangatal, A., Jankovski, A., Olivier, C., Queraud-Lesaux, F.,Goujet-Zalc, C., Thomas, J.L., Zalc, B., 2001. Sonic hedgehog-dependent emergenceof oligodendrocytes in the telencephalon: evidence for a source of oligodendro-cytes in the olfactory bulb that is independent of PDGFRα signaling. Development128, 4993–5004.
Spassky, N., Merkle, F.T., Flames, N., Tramontin, A.D., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 2005. Adult ependymal cells are postmitotic and are derived from radialglial cells during embryogenesis. J. Neurosci. 25, 10–18.
Stanco, A., Szekeres, C., Patel, N., Rao, S., Campbell, K., Kreidberg, J.A., Polleux, F., Anton, E.S.,2009. Netrin-1-α3β1 integrin interactions regulate the migration of interneuronsthrough the cortical marginal zone. Proc. Natl. Acad. Sci. U. S. A. 106, 7595–7600.
Stenman, J., Toresson, H., Campbell, K., 2003. Identification of two distinct progenitorpopulations in the lateral ganglionic eminence: implications for striatal andolfactory bulb neurogenesis. J. Neurosci. 23, 167–174.
Taupin, P., Gage, F.H., 2002. Adult neurogenesis and neural stem cells of the centralnervous system in mammals. J. Neurosci. Res. 69, 745–749.
Toresson, H., Campbell, K., 2001. A role for Gsh1 in the developing striatum andolfactory bulb of Gsh2 mutant mice. Development 128, 4769–4780.
Tramontin, A.D., Garcia-Verdugo, J.M., Lim, D.A., Alvarez-Buylla, A., 2003. Postnataldevelopment of radial glia and the ventricular zone (VZ): a continuum of the neuralstem cell compartment. Cereb. Cortex 13, 580–587.
Tsai, H., Miller, R.H., 2002. Glial cell migration directed by axon guidance cues. TrendsNeurosci. 25, 173–175.
Tsai, H., Macklin, W.B., Miller, R.H., 2006. Netrin-1 is required for the normaldevelopment of spinal cord oligodendrocytes. J. Neurosci. 26, 1913–1922.
Ventura, R.E., Goldman, J.E., 2007. Dorsal radial glia generate olfactory bulbinterneurons in the postnatal murine brain. J. Neurosci. 27, 4297–4302.
Vitry, S., Avellana-Adalid, V., Lachapelle, F., Baron-Van Evercooren, A., 2001. Migrationand multipotentiality of PSA-NCAM+neural precursors transplanted in thedeveloping brain. Mol. Cell. Neurosci. 17, 983–1000.
Waclaw, R.R., Allen, Z.J.I.I., Bell, S.M., Erdèlyi, F., Szabó, G., Potter, S.S., Campbell, K., 2006.The zinc finger transcription factor Sp8 regulates the generation and diversity ofolfactory bulb interneurons. Neuron 49, 503–516.
Woodruff, R.H., Tekki-Kessaris, N., Stiles, C.D., Rowitch, D.H., Richardson, W.D., 2001.Oligodendrocyte development in the spinal cord and telencephalon: commonthemes and new perspectives. Int. J. Dev. Neurosci. 19, 379–385.
Yee, K.T., Simon, H.H., Tessier-Lavigne, M., O'Leary, D.D.M., 1999. Extension of longleading processes and neuronal migration in the mammalian brain directed by thechemoattractant Netrin-1. Neuron 24, 607–622.
Yun, K., Garel, S., Fischman, S., Rubenstein, J.L.R., 2003. Patterning of the lateralganglionic eminence by the Gsh1 and Gsh2 homeobox genes regulates striatal andolfactory bulb histogenesis and the growth of axons through the basal ganglia. J.Comp. Neurol. 461, 151–165.
Yurchenco, P.D., Wadsworth, W.G., 2004. Assembly and tissue functions of earlyembryonic laminins and netrins. Curr. Opin. Cell Biol. 16, 572–579.
Related Documents











