Neighbouring weeds influence the formation of arbuscular mycorrhiza in grapevine Tomislav Radić & Katarina Hančević & Matevž Likar & Ivan Protega & Marija Jug-Dujaković & Irena Bogdanović Received: 20 December 2011 / Accepted: 3 April 2012 / Published online: 1 May 2012 # Springer Science+Business Media B.V. 2012 Abstract Grapevine (Vitis vinifera L.) and two selected weeds from Mediterranean Croatian vineyards (Plantago lanceolata L. and Tanacetum cinerariifolium (Trevir.) Sch. Bip.) were examined in pot culture experiments, individu- ally or when combined, to see whether multiple hosts influ- enced the formation of the symbiosis with arbuscular mycorrhizal fungi (AMF). The results after six-month period showed that plant identity and density significantly influenced development of mycorrhizal intra- and extraradical mycelium and/or sporulation. Grapevine and T. cinerariifolium individ- ually and in combination resulted in a greater development of arbuscular mycorrhizae in terms of spore production, extra- radical mycelium length and root colonization compared with pots containing P. lanceolata. Herbaceous weed species seemed to promote a different set of dominant AMF, poten- tially providing a wider spectrum of AMF for colonising grapevine roots. This indicates the value of encouraging host plant diversity in vineyards. AMF sequences obtained in this study are the first data reported for soils in Croatia. Keywords Arbuscular mycorrhiza . Grapevine . Weed neighbours . Extraradical mycelium . AMF diversity 1 Introduction The associations between plant hosts and arbuscular mycor- rhizal fungi (AMF) greatly influence the establishment and maintenance of the plant community and ecosystem processes (van der Heijden et al. 1998; Hart et al. 2003). Numerous studies focus on plant responses to arbuscular mycorrhizal fungi (AMF) and the benefits that plants gain from this symbi- osis (Harley and Smith 1983; van der Heijden and Sanders 2003). Nevertheless, the composition and diversity of the plant community can influence the structure of the AMF community (Burrows and Pfleger 2002; Johnson et al. 2003; Hausmann and Hawkes 2010). Thus apart from environmental factors, AMF diversity is dependent on plant species identity and age (e.g. Hetrick and Bloom 1986; McGonigle and Fitter 1990; Genney et al. 2001; Li et al. 2009; Hausmann and Hawkes 2010). In general, plant richness increases AMF diversity (Alguacil et al. 2011). However, this is mostly based on studies of natural habitats where environmental factors interact simul- taneously and cannot be fully assessed and/or controlled. In addition to plant-host specificity, AMF composition is influenced by neighbouring plants that can have beneficial, inhibitory or neutral effects. For example, Mummey et al. (2005) reported that the invasive Centaurea maculosa L. controlled the AMF community composition in surrounding native plants. Plant competition in natural ecosystems may be indirect and depend on the establishment and mainte- nance of mycorrhizal networks and the nutrient exchanges within them (Bever and Schultz 2005). The same also holds true for agroecosystems where crop species interact with weed species. Grapevine (Vitis vinifera L.) is a crop whose Electronic supplementary material The online version of this article (doi:10.1007/s13199-012-0165-3) contains supplementary material, which is available to authorized users. T. Radić (*) : K. Hančević : I. Protega : M. Jug-Dujaković : I. Bogdanović Institute for Adriatic Crops and Karst Reclamation, Put Duilova 11, 21000 Split, Croatia e-mail: [email protected] M. Likar Biotechnical Faculty, Department of Biology, University of Ljubljana, Večna pot 111, Ljubljana, Slovenia Symbiosis (2012) 56:111–120 DOI 10.1007/s13199-012-0165-3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neighbouring weeds influence the formation of arbuscularmycorrhiza in grapevine
Tomislav Radić & Katarina Hančević & Matevž Likar &
Ivan Protega & Marija Jug-Dujaković &
Irena Bogdanović
Received: 20 December 2011 /Accepted: 3 April 2012 /Published online: 1 May 2012# Springer Science+Business Media B.V. 2012
Abstract Grapevine (Vitis vinifera L.) and two selectedweeds from Mediterranean Croatian vineyards (Plantagolanceolata L. and Tanacetum cinerariifolium (Trevir.) Sch.Bip.) were examined in pot culture experiments, individu-ally or when combined, to see whether multiple hosts influ-enced the formation of the symbiosis with arbuscularmycorrhizal fungi (AMF). The results after six-month periodshowed that plant identity and density significantly influenceddevelopment of mycorrhizal intra- and extraradical myceliumand/or sporulation. Grapevine and T. cinerariifolium individ-ually and in combination resulted in a greater development ofarbuscular mycorrhizae in terms of spore production, extra-radical mycelium length and root colonization compared withpots containing P. lanceolata. Herbaceous weed speciesseemed to promote a different set of dominant AMF, poten-tially providing a wider spectrum of AMF for colonisinggrapevine roots. This indicates the value of encouraging hostplant diversity in vineyards. AMF sequences obtained in thisstudy are the first data reported for soils in Croatia.
Keywords Arbuscular mycorrhiza . Grapevine .Weedneighbours . Extraradical mycelium . AMF diversity
1 Introduction
The associations between plant hosts and arbuscular mycor-rhizal fungi (AMF) greatly influence the establishment andmaintenance of the plant community and ecosystem processes(van der Heijden et al. 1998; Hart et al. 2003). Numerousstudies focus on plant responses to arbuscular mycorrhizalfungi (AMF) and the benefits that plants gain from this symbi-osis (Harley and Smith 1983; van der Heijden and Sanders2003). Nevertheless, the composition and diversity of the plantcommunity can influence the structure of the AMF community(Burrows and Pfleger 2002; Johnson et al. 2003; Hausmannand Hawkes 2010). Thus apart from environmental factors,AMF diversity is dependent on plant species identity and age(e.g. Hetrick and Bloom 1986; McGonigle and Fitter 1990;Genney et al. 2001; Li et al. 2009; Hausmann and Hawkes2010). In general, plant richness increases AMF diversity(Alguacil et al. 2011). However, this is mostly based on studiesof natural habitats where environmental factors interact simul-taneously and cannot be fully assessed and/or controlled.
In addition to plant-host specificity, AMF composition isinfluenced by neighbouring plants that can have beneficial,inhibitory or neutral effects. For example, Mummey et al.(2005) reported that the invasive Centaurea maculosa L.controlled the AMF community composition in surroundingnative plants. Plant competition in natural ecosystems maybe indirect and depend on the establishment and mainte-nance of mycorrhizal networks and the nutrient exchangeswithin them (Bever and Schultz 2005). The same also holdstrue for agroecosystems where crop species interact withweed species. Grapevine (Vitis vinifera L.) is a crop whose
Electronic supplementary material The online version of this article(doi:10.1007/s13199-012-0165-3) contains supplementary material,which is available to authorized users.
T. Radić (*) :K. Hančević : I. Protega :M. Jug-Dujaković :I. BogdanovićInstitute for Adriatic Crops and Karst Reclamation,Put Duilova 11,21000 Split, Croatiae-mail: [email protected]
M. LikarBiotechnical Faculty, Department of Biology,University of Ljubljana,Večna pot 111,Ljubljana, Slovenia
Symbiosis (2012) 56:111–120DOI 10.1007/s13199-012-0165-3

establishment and growth is highly dependent on thepresence of AMF (Schreiner 2005). Grapevines have alow root density in comparison to other crops (Mohr1996), and its AMF symbionts play a significant role inits water and mineral acquisition (Abbot and Robson 1984).The formation of grapevine mycorrhizal networks with neigh-bouring plants depends on vineyard soil management practi-ces, especially weed control and cover cropping (Baumgartneret al. 2005).
In the present study, we examined the hypothesis that theplanting density of the host plant (grapevine) and the pres-ence of other neighbouring plants would influence the AMFcolonisation of grapevine roots. More specifically, we exploredwhether: 1) grapevine and the weed species from Croatianvineyards develop arbuscular mycorrhizal symbioses to thesame extent under uniform conditions; 2) different AMF spe-cies associate with the different vineyard weeds; 3) the coex-istence of multiple host plants (grapevine and weeds) in potcultures influenced arbuscular mycorrhizal root colonization ofgrapevine as well as the spread of extraradical hyphae andsporulation of the AMF.
2 Material and methods
2.1 Plant material
Plants selected as hosts for mycorrhizal establishment weregrapevine (Vitis vinifera L. cv. Plavac mali), ribwort plantain(Plantago lanceolata L.), Dalmatian pyrethrum (Tanacetumcinerariifolium (Trevir.) Sch.Bip.), dandelion (Taraxacumofficinale Weber), and white clover (Trifolium repens L.).They were chosen because they are co-occurring weeds invineyards in the Mediterranean region of Croatia.
Grapevine cuttings were collected from a vineyard nearthe town of Vrgorac in south Croatia (43°12’ N, 17°20’ E)and disinfected in 0.4 % solution of fungicide Mikal flash(fosetyl-Al 50 %, folpet 25 %) and 1 % NaOCl, dipped into1,500 ppm Indol Acetic Acid for root growth promotion,and planted into an autoclaved pot mix (50 % vermiculite,35 % 0.3–0.6 mm quartz sand, 15 % <0.3 mm quartz sand).Seeds of P. lanceolata, T. cinerariifolium, T. officinale, andT. repens were disinfected with 0.5 % NaOCl for 30 min,washed with sterile water and germinated in the same potmix as used for grapevine cuttings. Three weeks later theseedlings and the rooted cuttings were transplanted to 2 Lpots (d017 cm) and fertilized with 5 g per pot of slow-solubilizing Osmocote fertilizer pearls of low-phosphorus-content (N:P:K 15:9:15 plus micronutrients). Plants werekept in a growth chamber at a temperature of 25 °C; witha light intensity of 1,000 lx (Flora, Osram, Germany). Theywere periodically illumined (16 h of light and 8 h of darkness)and regularly watered with tap water.
2.2 Inoculum
Grapevine rhizosphere soils were collected from four samplingpoints in a vineyard in south Croatia (43°12’ N, 17°20’ E) atthe stage of bud break. The sampling points were at distancesof approximately 10 m across the diagonal of the rectangularvineyard. The soil was collected at a depth of 0–20 cm. Sam-ples were mixed together and homogenized as much as possi-ble to make a sample of about 10, 000 g. In each 2 L pot 100 gof such soil inoculum was added to the sterilized pot mix, rightbelow the seedlings roots and then the pots were filled to thetop with the same potting mix (prepared as described in sec-tion 2.1.; quartz sand and vermiculite). This way the trapcultures were set up. The pH of the resulting plant growthsubstrate was 8.0. This approach simulated natural con-ditions in the vineyard in terms of the diversity andabundance of AMF species that could interact withany plants grown in the pots.
2.3 Experimental design
The nine combinations of plant hosts (Table 1) were set upin the pots : grapevine–one plant (Gr); grapevine–fourplants (Gr × 4); T. cinerariifolium–one plant (Tc); T. cine-rariifolium–four plants (Tc × 4); P. lanceolata–one plant(Pl); P. lanceolata–four plants (Pl × 4); one grapevine plantwith one T. cinerariifolium plant (Gr + Tc); one grapevineplant with one P. lanceolata plant (Gr + Pl), and one grapevineplant with one P. lanceolata plant, and one T. cinerariifoliumplant (Gr + Tc + Pl). There were nine replicates for each ofthese combinations, i.e. 81 pots in total. Sampling was done 2,4 and 6 months after the inoculation. Each time, three pots ofevery combination were sampled. The pots were sampleddestructively, that is, the entire plant and accompanying sub-strate was removed in order to collect material for the plantand mycorrhizal parameters.
For the determination of the AMF community in the hostplant roots, trap cultures of four plant species (V. vinifera, T.officinale, T. repens and P. lanceolata) were set up as de-scribed in section 2.2. Each plant species was represented bythree pots (one plant per pot). After 6 months their rootswere sampled and processed for assess the AMF community(12 samples in total).
2.4 Mycorrhizal parameters
AMF spores were isolated from 100 g pot substrate samplesby a wet sieving and decanting method, followed by sucrosegradient centrifugation (Gerdeman and Nicolson 1963).They were counted under the stereomicroscope at 50×magnification.
Grapevine and weeds’ roots were examined to determineAMF colonization. After the washing, roots were KOH
112 T. Radić et al.
cleared, acidified, stained with Trypan blue, stored in glyceroland examined under light microscope (Phillips and Hayman1970). Arbuscular, vesicular, hyphae only, and total coloniza-tions were determined according to the magnified intersec-tions method (McGonigle et al. 1990). 150–200 intersectionswere analyzed per each replicate of plant roots.
Hyphal length of extraradical mycelium (ERM) wasassessed using an inserted membrane technique (Balaz andVosatka 2001). One cellulose nitrate membrane filter (MF-Millipore, 47 mm diameter, 0.45 μm pore size) was placedin each pot. Thirty random microscope fields from eachmembrane were recorded using a colour CCD video camera(Axio cam MRc5, Zeiss). Hyphal length was measured usingsoftware AxioVision 4.0 and the results were expressed asERM lengths per 1 cm2 of the inserted membrane.
2.5 Plant parameters
Fresh weight of shoots and roots was measured immediatelyafter washing the roots and excess moisture removal. Shootswere dried in an oven at 70 °C for 48 h to determine dryweight. Fresh root/shoot ratio and dry weight percentagewere calculated.
2.6 DNA extraction from roots, PCR, cloningand sequencing
Plant hosts’ roots were thoroughly rinsed and cut into 1-cmpieces. 150 mg of root pieces per sample were crushed inliquid nitrogen and the procedure for NucleoSpin Plant IIKit (Macherey-Nagel, Duren, Germany) was followed forDNA extraction. For PCR amplifications, the universal eu-karyotic primer NS31 (Simon et al. 1992) was used as a
forward primer and a mixture of AM1 (Helgason et al.1998), AM2 and AM3 (Santos-Gonzalez et al. 2007)was used as the reverse primer. Besides AM1 primer,originally designed for the specific amplification ofGlomeromycota DNA, AM2 and AM3 were used foramplification of DNA from Glomus group B and Glo-mus group C (Diversisporaceae) representatives. Thisprimer combination targets a 550-bp-long region of the18S rRNA gene. Final reaction volume was 50 μL. The PCRthermocycling program was as follows: 94 °C for 10 min,followed by 35 cycles of 94 °C for 1 min, 58 °C for 1 min and72 °C for 2 min. The program ended at 72 °C for 8 min.
After confirming PCR products were the correct size(~550 bp) by electrophoresis on 1 % agarose gels, they werecloned using TOPO TA cloning kit (Invitrogen, San Diego,USA) according to the manufacturer’s instructions. A min-imum of ten positive clones per sample (30 clones per plantspecies, 120 in total) was reamplified using M13 primersand checked by electrophoresis. Overall, 80 clones thatconfirmed to had incorporated DNA fragment of correctsize were purified with NucleoSpin kit (Macherey-Nagel,Duren, Germany) and sequenced (single stranded sequenc-ing) by the Macrogen Company (Korea).
2.7 Phylogenetic analysis
The sequence data have been submitted to the GenBankdatabase for their accession numbers. The sequences weresubjected to a GenBank search to evaluate the taxonomicaffinities of each of the 18S sequences, using the defaultoption of gapped BLAST (Altschul et al. 1997). The sequencealignments were carried out by ClustalX (Larkin et al. 2007),and refined by eye. The dataset subjected to phylogenetic
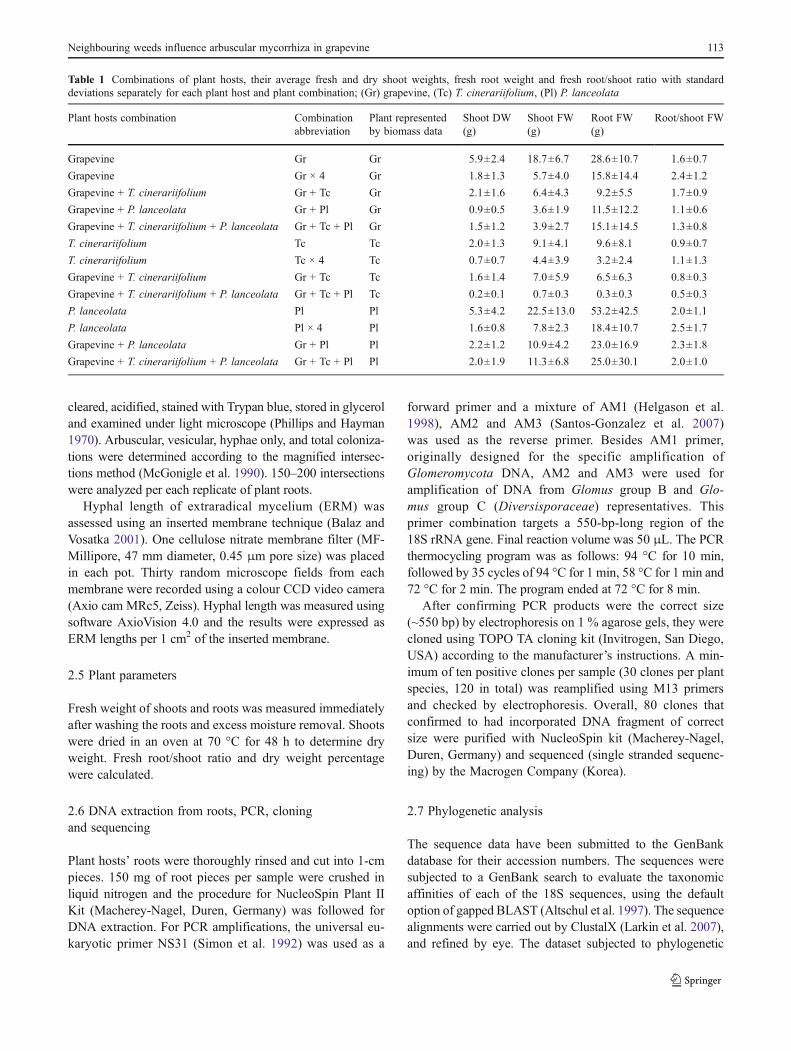
Table 1 Combinations of plant hosts, their average fresh and dry shoot weights, fresh root weight and fresh root/shoot ratio with standarddeviations separately for each plant host and plant combination; (Gr) grapevine, (Tc) T. cinerariifolium, (Pl) P. lanceolata
Plant hosts combination Combinationabbreviation
Plant representedby biomass data
Shoot DW Shoot FW Root FW Root/shoot FW(g) (g) (g)
Grapevine Gr Gr 5.9±2.4 18.7±6.7 28.6±10.7 1.6±0.7
Grapevine Gr × 4 Gr 1.8±1.3 5.7±4.0 15.8±14.4 2.4±1.2
Grapevine + T. cinerariifolium Gr + Tc Gr 2.1±1.6 6.4±4.3 9.2±5.5 1.7±0.9
Grapevine + P. lanceolata Gr + Pl Gr 0.9±0.5 3.6±1.9 11.5±12.2 1.1±0.6
Grapevine + T. cinerariifolium + P. lanceolata Gr + Tc + Pl Gr 1.5±1.2 3.9±2.7 15.1±14.5 1.3±0.8
T. cinerariifolium Tc Tc 2.0±1.3 9.1±4.1 9.6±8.1 0.9±0.7
T. cinerariifolium Tc × 4 Tc 0.7±0.7 4.4±3.9 3.2±2.4 1.1±1.3
Grapevine + T. cinerariifolium Gr + Tc Tc 1.6±1.4 7.0±5.9 6.5±6.3 0.8±0.3
Grapevine + T. cinerariifolium + P. lanceolata Gr + Tc + Pl Tc 0.2±0.1 0.7±0.3 0.3±0.3 0.5±0.3
P. lanceolata Pl Pl 5.3±4.2 22.5±13.0 53.2±42.5 2.0±1.1
P. lanceolata Pl × 4 Pl 1.6±0.8 7.8±2.3 18.4±10.7 2.5±1.7
Grapevine + P. lanceolata Gr + Pl Pl 2.2±1.2 10.9±4.2 23.0±16.9 2.3±1.8
Grapevine + T. cinerariifolium + P. lanceolata Gr + Tc + Pl Pl 2.0±1.9 11.3±6.8 25.0±30.1 2.0±1.0
Neighbouring weeds influence arbuscular mycorrhiza in grapevine 113

analyses was composed of the glomalean sequences obtainedand 34 additional 18S rDNA sequences of glomeromycetes(Glomus group A, Glomus group B, Diversisporaceae, Para-glomaceae, Gigasporaceae, Acaulosporaceae) from Gen-Bank. The sequences of Paraglomus occultum were used asan out-group.
Neighbour-Joining (NJ) and Maximum Likelihood (ML)were used to analyze the aligned sequences. All analyseswere performed in MEGA5 software (Tamura et al. 2011).In both the ML and NJ analyses, the evolutionary modelK80 + Γ (Kimura 1980) of Modeltest 3.7 (Posada andCrandall 1998) was used, selected by hierarchical likelihoodratio tests (hLRTs). Bootstrap values were obtained using200 subsamples for Maximum Likelihood and 1,000 sub-samples for Neighbour Joining.
2.8 Statistical analysis
To test simultaneously for overall plant combination effectsover three harvests during the experiment, we subjected datato repeated-measures analysis of variance (ANOVA, Systat10.2) for one grouping factor (plant combination) and onewithin factor (sampling time). Repeated measures ANOVAfor two grouping factors (plant identity and plant density)and one within factor (sampling time) was used to differentiatedensity and host identity influence over time on mycorrhizalparameters by considering Gr, Gr × 4, Tc, Tc × 4, Pl and Pl × 4treatments. One-way ANOVA with Bonferroni post-hoc testwas used for each sampling separately in order to estimatestatistical significance of differences of mycorrhizal and plantvariables between different treatments. Data were transformedinto natural logarithm values to obtain a normal distributionbefore statistical analyses were performed.
Data on distribution of different AMF 18S rDNA sequencesamong different plant hosts was used for cluster analysis withunweighted pair-group method, with arithmetic average(UPGMA) method in R v2.14.1.
3 Results
3.1 Mycorrhizal parameters
Prior to planting host seedlings and rooted cuttings, sporeabundance in the initial field soil inoculum was assessed.The average abundance was five spores per gram (5.2±0.8,four samples). After the inoculum was added to the pottingsoil mix, the estimated spore abundance was 0.26 spores pergram of pot.
Two months after the experiment was set up, grapevine (Gr)and the combinationGr + Tc exhibited the greatest abundance ofAMF spores. The next two measurements (4 and 6 months)confirmedGr, Tc, andGr +Tc treatments as themost suitable for
spore production (Fig. 1) (P00.008, F02.873), with maximumvalues being about 6 spores per gram. The Pl and Pl × 4 treat-ments had the lowest spore number, on average less than 0.8 pergram. Repeated measures ANOVA integrated different harvestsand confirmed overall significant differences between plantcombinations (P00.004, F05.458) as well as significant timeinfluence (P<0.001, F029.457). However, the interactionbetween plant combinations and time was not significant.
The AMF extraradical mycelium (ERM) (Fig. 2) showedsignificant differences between treatments after four (P00.010,F04.289) and 6 months (P00.043, F02.829), with the highestvalues being (up to 182 cm cm−2) in pots with a single grapevineplant (Gr). The lowest values (17 cm cm−2) were in the Gr + Pltreatment. Generally, ERM differed between plant combinations(F02.863, P00.045) and between three harvests (P<0.001,F025.128). There were significant interaction between thesetwo factors (P00.007, F02.931), which indicates significantchanges with time at least for some plant combinations.
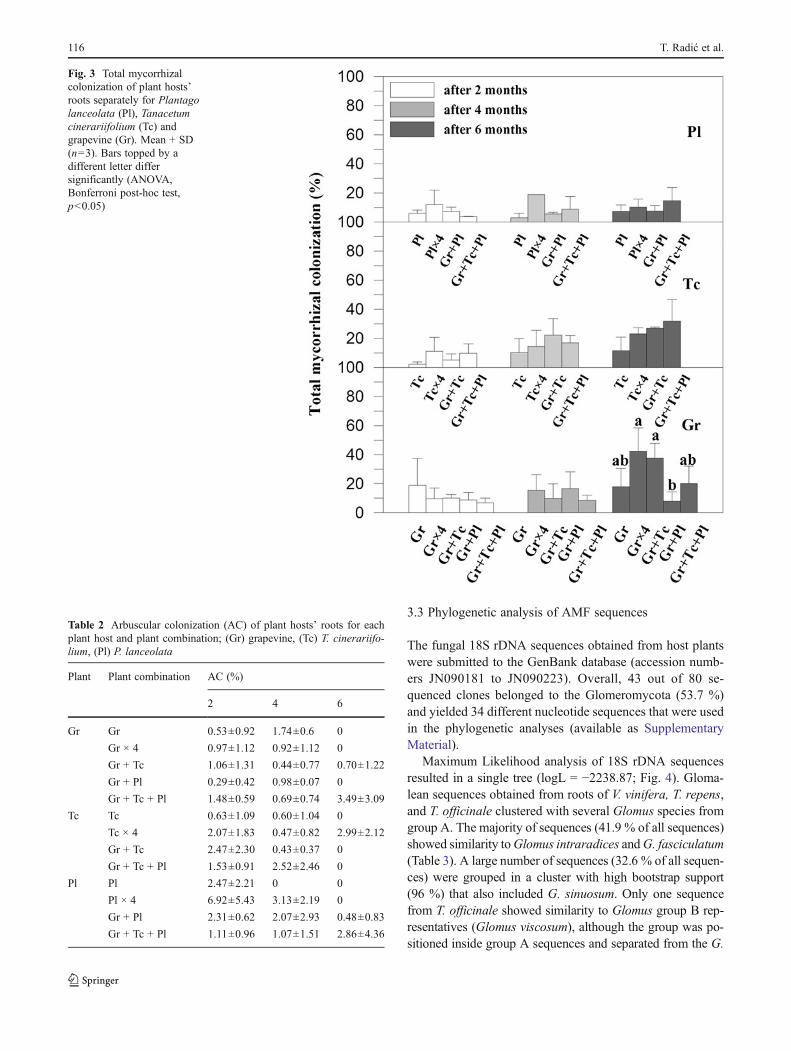
Analyzing the three harvests separately, the percentage ofmycorrhizal colonization (Fig. 3) only differed betweengrapevine combinations (P00.05, F03.442) at 6 months.Gr × 4 and Gr + Tc had the highest colonization (about40 %), significantly greater than Gr + Pl (8 %). However,the overall results only showed significant differences forgrapevine between the three harvests (F03.799, P00.040),and not between the plant combinations. The integratedresults differed between the combinations involving P. lan-ceolata (P00.008, F08.345) and those with T. cinerariifo-lium (P00.009, F08.005), and between the three harvests(P00.003, F08.681).
Arbuscule colonization declined from the first to the thirdsampling. At the end of the experiment, arbuscules wereonly found in grapevine and P. lanceolata when these weregrowing in combinations with other plant species. T. cinerar-iifolium only maintained arbuscules in the Tc × 4 treatment(Table 2). In grapevine and T. cinerariifolium, however, arbus-cular colonization changed between the three harvests(P00.006, F06.641 and P00.007, F06.859, respectively)as did vesicular colonization of T. cinerariifolium (P00.025,F04.682). T. cinerariifoliumwas significantly more colonisedthan P. lanceolata after 4 months (P00.035, F03.785) and6 months (P00.038, F03.616). Throughout the experiment,vesicles were more numerous in grapevine than in T. cinerar-iifolium or P. lanceolata (P00.013, F04.980) irrespective ofthe hosts’ combination in pots, while arbuscules were themostnumerous in P. lanceolata (P00.045).
The density effect was assessed through a comparison ofobserved parameters in pots with one plant versus pots withfour plants. The abundance of spores and ERM lengthshowed no significant difference with planting density.In contrast, significant differences were found betweenone and four plants in terms of total AMF colonization(P00.004, F012.212) as well as for plant × density
114 T. Radić et al.

interaction (P00.028,F04.904). Repeatedmeasures ANOVAshowed significant differences between one and four planttreatments for arbuscular (P00.009, F09.506) and vesicularcolonization (P00.029, F06.153), but there was no plant ×density interaction. Significant differences were also apparentin one and four plant treatments between the first, second andthird sampling (P00.004, F07.011 and P<0.001, F011.036respectively).
3.2 Plant parameters
Average fresh and dry shoot weight, root fresh weight andfresh root/shoot ratios are shown in Table 1 for each of threeplant species and for every plant combination. The greatestshoot biomass was measured in grapevines while the highestroot/shoot ratio was calculated for Plantago lanceolata.Shoot DW for T. cinerariifolium generally differed betweentreatments (P<0.001, F024.102). The same held true for
grapevine (P00.002, F09.577) and P. lanceolata (P00.005,F09.557) but no plant × harvest interaction was found. Rootfresh biomass differed between treatments for T. cinerarii-folium (P<0.001, F047.2009) and P. lanceolata (P00.010,F07.576), but interaction plant × harvest was found only forT. cinerariifolium (P00.032, F03.113). Root/shoot ratiochanged with time (P<0.001) but no significant differencebetween treatments was found for this parameter. Significantdifferences between two contrast densities (one and four) werefound for shoot DW (P<0.001, F013.101) and root FW(P<0.001, F012.704) but not for plant × density interaction.
In order to test whether changes in mycorrhizal parame-ters (spore abundance, ERM, total radical colonization,arbuscules and vesicular colonization) correlated to plantdry weight, root biomass or root/shoot ratio a Pearson cor-relation matrix was analyzed for total data and separately forgroups of data (plant species, plant combination, and monthof sampling). No significant correlations were found.
Fig. 1 AMF spores’ abundancein pot substrates during theexperiment. Mean + SD (n03).Bars topped by a different letterdiffer significantly (ANOVA,Bonferroni post-hoc test,p<0.05)
Fig. 2 AMF extraradicalmycelium (ERM) length in potsubstrates during experiment.Bars topped by a different letterdiffer significantly (ANOVA,Bonferroni post-hoc test,p<0.05)
Neighbouring weeds influence arbuscular mycorrhiza in grapevine 115
3.3 Phylogenetic analysis of AMF sequences
The fungal 18S rDNA sequences obtained from host plantswere submitted to the GenBank database (accession numb-ers JN090181 to JN090223). Overall, 43 out of 80 se-quenced clones belonged to the Glomeromycota (53.7 %)and yielded 34 different nucleotide sequences that were usedin the phylogenetic analyses (available as SupplementaryMaterial).
Maximum Likelihood analysis of 18S rDNA sequencesresulted in a single tree (logL 0 −2238.87; Fig. 4). Gloma-lean sequences obtained from roots of V. vinifera, T. repens,and T. officinale clustered with several Glomus species fromgroup A. The majority of sequences (41.9 % of all sequences)showed similarity toGlomus intraradices andG. fasciculatum(Table 3). A large number of sequences (32.6 % of all sequen-ces) were grouped in a cluster with high bootstrap support(96 %) that also included G. sinuosum. Only one sequencefrom T. officinale showed similarity to Glomus group B rep-resentatives (Glomus viscosum), although the group was po-sitioned inside group A sequences and separated from the G.
Fig. 3 Total mycorrhizalcolonization of plant hosts’roots separately for Plantagolanceolata (Pl), Tanacetumcinerariifolium (Tc) andgrapevine (Gr). Mean + SD(n03). Bars topped by adifferent letter differsignificantly (ANOVA,Bonferroni post-hoc test,p<0.05)
Table 2 Arbuscular colonization (AC) of plant hosts’ roots for eachplant host and plant combination; (Gr) grapevine, (Tc) T. cinerariifo-lium, (Pl) P. lanceolata
Plant Plant combination AC (%)
2 4 6
Gr Gr 0.53±0.92 1.74±0.6 0
Gr × 4 0.97±1.12 0.92±1.12 0
Gr + Tc 1.06±1.31 0.44±0.77 0.70±1.22
Gr + Pl 0.29±0.42 0.98±0.07 0
Gr + Tc + Pl 1.48±0.59 0.69±0.74 3.49±3.09
Tc Tc 0.63±1.09 0.60±1.04 0
Tc × 4 2.07±1.83 0.47±0.82 2.99±2.12
Gr + Tc 2.47±2.30 0.43±0.37 0
Gr + Tc + Pl 1.53±0.91 2.52±2.46 0
Pl Pl 2.47±2.21 0 0
Pl × 4 6.92±5.43 3.13±2.19 0
Gr + Pl 2.31±0.62 2.07±2.93 0.48±0.83
Gr + Tc + Pl 1.11±0.96 1.07±1.51 2.86±4.36
116 T. Radić et al.

lamellosum/G. etunicatum cluster. In addition, severalsequences from P. lanceolata showed similarity to represen-tatives of Diversisporaceae. Two small clusters (unidentifiedgroups 1 and 2) contained sequences only found from ourstudy and could not be assigned to any already identifiedGlomus species.
Cluster analysis of distribution of AMF among differentplant hosts showed the highest similarity between the AMFcommunities of T. officinale and grapevine (Fig. 5a). TheAMF community on the roots of P. lanceolata was the leastsimilar to AMF communities of other host plants examined.Sequences of G. intraradices/G. fasciculatum and G. sinu-osum were observed in three of the plant hosts (Fig. 5b),whereas the majority of other phylogenetic clusters fromFig. 4 was observed in a single plant species.
4 Discussion
Our results confirm the initial hypotheses that host plantidentity and density affect the development and compositionof the AMF community. This has been found previouslywhen different hosts have been examined individually(Tahat et al. 2008). However the studied host plant combi-nations revealed that these were important as well. Thegrapevine and the T. cinerariifolium individually and incombination (Gr + Tc) were very supportive in terms ofmycorrhizal establishment, andwere a significant improvementcompared with P. lanceolata and its combination with grape-vine (Gr + Pl). Our findings indicate that grapevine does notalways have the same facility to interact with AMF which isdependent on its neighbouring plants. We conclude that not allplant interactions are equally favourable for mycorrhizal estab-lishment. In these relationships, the essential and specific char-acteristics of each plant species are important, and diverseallelopathic effects may occur (Mallik and Williams 2005).
Plant root exudates may be involved in suppression orstimulation of AMF root colonization. (Vierheilig et al.2003; Bainard et al. 2009). The influence of root allelo-chemicals was explored in P. lanceolata as a highly mycor-rhizal plant (De Deyn et al. 2009). The authors concludedthat its root exudation (iridoid glycoside) does not reducethe potential benefit of symbioses with AMF although itserves as a defence mechanism. The question remains howsecondary metabolites can influence the colonization of aneighbouring plant, such as grapevine. P. lanceolata mayreduce the AMF colonization in grapevine by being a poorhost. Our results showed that this plant species, when a neigh-bour, reduced the development of grapevine mycorrhizae. Incontrast, T. cinerariifoliumwas a beneficial grapevine neighbourfrom the aspect of AMF development.
Grapevine, T. cinerariifolium and P. lanceolata devel-oped greater shoot biomass when planted alone rather than
Fig. 4 Maximum Likelihood tree of fungal 18S sequences showingthe placement of sequences isolated from Vitis vinifera, Trifoliumrepens, Plantago lanceolata and Taraxacum officinale among identi-fied glomalean fungal species; the groups formed are discussed in themain text; numbers indicate where bootstrap support is above 60 %(200 replicates)
Neighbouring weeds influence arbuscular mycorrhiza in grapevine 117

in higher density or with other species, as already describedfor P. lanceolata (van Ruijven and Berendse 2003). Thisconfirms the disappearance of the positive effects of AMFon plant growth when plants are more crowded (Moora andZobel 1998; Koide and Dickie 2002; Perez and Urcelay2009). Furthermore, it seems that plant density also affectsdevelopment of fungal symbioses. We observed significantlydifferent mycorrhizal colonization in pots where four plants ofthe same species grew together versus pots with only a singleplant. It could be hypothesized that plant density was thedetermining factor. However, plant density was shown to beresponsible for only some of the observed mycorrhizal param-eters. Differing competitive abilities of the selected plantspecies may also have played a role. Those differences could
have constrained the mineral nutrition, photosynthesis andgrowth of particulate plant species, leading to differences insupport of AMF. The role of plant host competition and/orneighbour interaction could not be discerned in the presentstudy.
The resident vegetation may help to maintain high AMfungal population diversity in the vineyards via weed net-works with the grapevines. These networks can affect grape-vine growth or nutrient uptake (Baumgartner et al. 2005;Schreiner and Mihara 2009). Based on our findings it isimportant which weed species are present, as they promoteAMF development to a different degree and have differentdominant AM fungi. We used a primer combination com-posed of a mixture of reverse primers AM1, AM2, and AM3
Table 3 Grouping of sequencesby the nucleotide similarity(97 % similarity was used as thecut-off value) with numbers ofsequences in the individualgroup and the percentage of allobtained sequences; plant host ofthe respective sequences form-ing the individual group andphylogenetic group based on thephylogeny analysis are added
Sequencegroup
No. of sequencesin group
% of allsequence
Phylogenetic group Plant hosts
1 18 41.9 G. intraradices/G. fasciculatum T. repens, T. officinale,V. vinifera
2 14 32.6 G. sinuosum P. lanceolata, T. officinale,V. vinifera
3 4 9.3 Diversisporaceae P. lanceolata
4 3 7.0 unidentified group 1 T. repens, T. officinale
5 2 4.7 unidentified group 2 V. vinifera
6 1 2.3 G. mosseae T. repens
7 1 2.3 G. viscosum T. officinale
Fig. 5 a Unweighted pair-group analysis using arithmetic average(UPGMA) cluster analysis of arbuscular mycorrhial (AM) communi-ties from roots of Vitis vinifera, Trifolium repens, Plantago lanceolataand Taraxacum officinale, as revealed by sequencing of 18S rDNA. bDistribution of the phylogenetic clusters of identified and unidentifiedGlomus species present in the roots of the four host plant species
indicated as the frequency of each fungal species in each plant species.Legend: G.intra./G.fasc-Glomus intraradices/Glomus fasciculatum, G.mos-Glomus mosseae, G.sin-Glomus sinuosum, G.vis-Glomus visco-sum, UG 1-Unidentified Glomus group 1, UG 2-Unidentified Glomusgroup 2, Diversispor-Diversisporaceae
118 T. Radić et al.

(Santos-Gonzalez et al. 2007), since AM1 exhibits somemismatches in the annealing sites with taxa belonging toGlomus groups B and C (Schwarzott et al. 2001). Theseprimers amplified an AMF consortium colonizing the grape-vine and herbaceous weeds roots, composed of various taxawithin Glomus groups A and B (Glomeraceae) and C(Diversisporaceae). We did not detect the presence of fungalsequences belonging to the families Acaulosporaceae, Gig-asporaceae, and Pacisporaceae, which can also be detectedwith this set of primers. The greatest number of AMFsequences (representatives Glomus group A) was found ingrapevine. However, other plant hosts accomodated morediverse AMF communities, as in T. officinale roots whererepresentatives of Glomus groups A and B discovered and inroots of P. lanceolata where fungi of Glomus groups A andC were found. The majority of Glomus A group sequencesshowed similarity to G. sinuosum, G. fasciculatum and G.intraradices, supporting the designation of the latter as acosmopolitan fungus in many ecosystems (Sykorova et al.2007a, b; Öpik et al. 2009). A single sequence belonging toG. mosseae was identified, suggesting rather low presenceof this fungus in the vineyard soil. The total absence of G.mosseae from the grapevine roots in our experiment couldbe due to the composition of the initial inoculum used,although Schreiner and Mihara (2009) also failed to observeany sequences of G. mosseae and Acaulosporaceae ongrapevine roots, despite finding spores of G. mosseae aswell as several Acaulospora species in vineyard soils. Thecommon occurrence of G. mosseae spores in soil, but theabsence of this fungus in root amplicons was also reportedfor agricultural fields (Kjøller and Rosendahl 2001) andmixed grassland communities (Hempel et al. 2007). Fungiwithin the G. mosseae clade are considered to be ruderalspecies, as they are common among early colonisers of treeseedlings (Husband et al. 2002) and greenhouse trap plants(Sykorova et al. 2007a). They are later replaced byother AMF species. In contrast to several members ofGlomus group A that were identified in the roots ofgrapevine and examined herbaceous weeds in our study,the sequences of other Glomus groups were scarce andobserved only in herbaceous weeds. No representativesof either of these Glomus groups were previously de-scribed as colonising grapevine roots. This could be theresult of only limited available evidence on Glomusgroup B and C isolates present in the literature, due tothe extensive use of the primer pair NS31-AM1. Thesequences obtained in this study are the first data onnative AMF species in Croatian soils and only a pre-view of expected AMF biodiversity.
In conclusion, when establishing pot cultures the choiceof plant host or a plant host combination is a factor that cansignificantly alter the extent and characteristics of arbuscularmycorrhizal development. This has important implications
for understanding natural ecosystems, where various plantsintensively communicate and have varying effects on arbus-cular mycorrhizal symbioses. These effects are also presentin the agroecosystems such as vineyards where the choice ofappropriate cover crops may influence mycorrhizal charac-teristics of grapevine.
Acknowledgment This study was financed by Unity throughKnowledge Fund (UKF), Croatia, grant no53, within “My first re-search” program, and Ministry of Science, Education and Sports ofthe Republic of Croatia (091-0910468-0279). The authors thank MScJ. Radić for help in statistical analysis.
References
Abbot LK, Robson AD (1984) The effect of root density, inoculumplacement and the infectivity of inoculum on the development ofvesicular-arbuscular mycorrhizas. New Phytol 97:285–299
Alguacil MM, Torres MP, Torrecillas E, Diaz G, Roldan A (2011) Planttype differently promote the arbuscular mycorrhizal fungi biodi-versity in the rhizosphere after revegetation of a degraded, semi-arid land. Soil Biol Biochem 43:167–173
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Lipman DJ(1997) Gapped BLAST and PSI-BLAST: a new generation ofprotein database search programs. Nucleic Acids Res 25:3389–3402
Bainard LD, Brown PD, Upadhyaya MK (2009) Inhibitory effect oftall hedge mustard (Sisymbrium loeselii) allelochemicals onrangeland plants and arbuscular mycorrhizal fungi. Weed Sci 57(4):386–393
Balaz M, Vosatka M (2001) A novel inserted membrane technique forstudies of mycorrhizal extraradical mycelium. Mycorrhiza11:291–296
Baumgartner K, Smith RF, Bettiga L (2005) Weed control and covercrop management affect mycorrhizal colonization of grapevineroots and arbuscular mycorrhizal fungal spore populations in aCalifornia vineyard. Mycorrhiza 15:111–119
Bever JD, Schultz PA (2005) Mechanisms of mycorrhizal mediation ofplant-plant interactions. In: Dighton J, Oudemans P (eds) Thefungal community, 4th edn. Taylor and Francis, Boca Raton, pp443–460
Burrows RL, Pfleger FL (2002) Arbuscular mycorrhizal fungi respondto increasing plant diversity. Can J Bot 80:120–130
De Deyn GB, Biere A, van der Putten WH, Wagenaar R, KlironomosJN (2009) Chemical defense, mycorrhizal colonization andgrowth responses in Plantago lanceolata L. Oecologia 160:433–442
Genney DR, Hartley SH, Alexander IJ (2001) Arbuscular mycorrhizalcolonization increases with host density in a heathland communi-ty. New Phytol 152:355–363
Gerdeman JW, Nicolson TH (1963) Spore of mycorrhizal endogonespecies extracted from soil by wet sieving and decanting. Trans BrMycol Soc 46:235–244
Harley JL, Smith SE (1983) Mycorrhizal symbiosis. Academic, NewYork
Hart MM, Reader RJ, Klironomos JN (2003) Plant coexistence medi-ated by arbuscular mycorrhizal fungi. Trends Ecol Evol 18(8):418–423
Hausmann NT, Hawkes CV (2010) Order of plant host establishmentalters the composition of arbuscular mycorrhizal communities.Ecology 91(8):2333–2343
Neighbouring weeds influence arbuscular mycorrhiza in grapevine 119

Helgason T, Daniell TJ, Husband R, Fitter AH, Young JPW (1998)Ploughing up the wood-wide web? Nature 394:431–431
Hempel S, Renker C, Buscot F (2007) Differences in the speciescomposition of arbuscular mycorrhizal fungi in spore, root andsoil communities in a grassland ecosystem. Environ Microbiol9:1930–1938
Hetrick BAD, Bloom J (1986) The influence of host plant on produc-tion ability of vesicular-arbuscular mycorrhizal spores. Mycologia78:32–36
Husband R, Herre EA, Turner SL, Gallery R, Young JPW (2002)Molecular diversity of arbuscular mycorrhizal fungi and patternsof host association over time and space in a tropical forest. MolEcol 11:2669–2678
Johnson D, Vandenkoornhuyse P, Leake JR, Gilbert L, Booth RE,Grime JP et al (2003) Plant communities affect arbuscular mycor-rhizal fungal diversity and community composition in grasslandmicrocosm. New Phytol 161:503–515
Kimura M (1980) A simple method for estimating evolutionary rate ofbase substitutions through comparative studies of nucleotidesequences. J Mol Evol 16:111–120
Kjøller R, Rosendahl S (2001) Molecular diversity of Glomalean(arbuscular mycorrhizal) fungi determined as distinct Glomusspecific DNA sequences from roots of field grown peas. MycolRes 105:1027–1032
Koide RT, Dickie IA (2002) Role of mycorrhizal fungi in plant pop-ulations. Plant Soil 244:307–317
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,McWilliam H et al (2007) Clustal W and Clustal X version 2.0.Bioinformatics 23:2947–2948
Li LF, Li T, Zhang Y, Zhao ZW (2009) Molecular diversity of arbus-cular mycorrhizal fungi and their distribution patterns related tohost-plants and habitats in a hot and arid ecosystem, southwestChina. FEMS Microbiol Ecol 71:418–427
Mallik MAB, Williams RD (2005) Allelopathic growth stimulation ofplants and microorganisms. Allelopathy J 16(2):175–198
McGonigle TP, Fitter AH (1990) Ecological specificity of vesicular-arbuscular mycorrhizal associations. Mycol Res 94:120–122
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA (1990)A new method which gives and objective measure of colonizationof roots by vesicular-arbuscular mycorrhizal fungi. New Phytol115:495–501
Mohr HD (1996) Periodicity of root tip growth of vines in the Mosellevalley. Vit Enol Sci 51:83–90
Moora M, Zobel M (1998) Can arbuscular mycorrhiza change theeffect of root competition between conspecific plants of differentages? Can J Bot 76:613–619
Mummey DL, Rillig MC, Holben WE (2005) Neighboring plant influ-ences on arbuscular mycorrhizal fungal community compositionas assessed by T-RFLP analysis. Plant Soil 271:83–90
Öpik M, Metsis M, Daniell TJ, Zobel M, Moora M (2009) Large-scaleparallel 454 sequencing reveals host ecological group specificityof arbuscular mycorrhizal fungi in a boreonemoral forest. NewPhytol 184:424–437
Perez M, Urcelay C (2009) Differential growth response to arbuscularmycorrhizal fungi and plant density in two wild plants belongingto contrasting functional types. Mycorrhiza 19:517–523
Phillips JM, Hayman DS (1970) Improved procedure of clearing rootsand staining parasitic and vesicular-arbuscular mycorrhizal fungifor rapid assessment of infection. Trans Br Mycol Soc 55:159–161
Posada D, Crandall KA (1998) Modeltesst: testing the model of DNAsubstitution. Bioinformatics 14:817–818
Santos-Gonzalez JC, Finlay RD, Tehler A (2007) Seasonal dynamicsor arbuscular mycorrhizal fungal communities in roots in a sem-natural grassland. Appl Environ Microbiol 73:5613–5623
Schreiner RP (2005) Spatial and temporal variation of roots, arbuscularmycorrhizal fungi, and plant and soil nutrients in a mature PinotNoir (Vitis vinifera L.) vineyard in Oregon, USA. Plant Soil276:219–234
Schreiner RP, Mihara KL (2009) The diversity of arbuscular mycor-rhizal fungi amplified from grapevine roots (Vitis vinifera L.) inOregon vineyards is seasonally stable and influenced by soil andvine age. Mycologia 101(5):599–611
Schwarzott D, Walker C, Schuessler A (2001) Glomus, the largestgenus of the arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Mol Phylogenet Evol 21:190–197
Simon L, Lalonde M, Bruns TD (1992) Specific amplification of 18Sfungal ribosomal genes from vesicular-arbuscular endomycorrhi-zal fungi colonizing roots. Appl Environ Microbiol 58:291–295
Sykorova Z, Ineichen K, Wiemken A, Redecker D (2007) The culti-vation bias: different communities of arbuscular mycorrhizal fun-gi detected in roots from the field, from bait plants transplanted tothe field, and from a greenhouse trap experiment. Mycorrhiza18:1–14
Sýkorová Z, Wiemken A, Redecker D (2007) Cooccurring Gentianaverna and Gentiana acaulis and their neighboring plants in twoSwiss upper montane meadows harbor distinct arbuscular mycor-rhizal fungal communities. Appl Environ Microbiol 73:5426–5434
Tahat MM, Kamaruzaman S, Radziah O, Kadir J, Masdek HN (2008)Plant host selectivity for multiplication of Glomus mosseae spore.Int J Bot 4:466–470
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011)MEGA5: molecular evolutionary genetics analysis using maxi-mum likelihood, evolutionary distance, and maximum parsimonymethods. Mol Biol Evol (In press)
van der Heijden MGA, Sanders IR (2003) Mycorrhizal ecology.Springer, Berlin
van der Heijden MGA, Boller T, Wiemken A, Sanders IR (1998)Different arbuscular mycorrhizal fungal species are potentialdeterminants of plant community structure. Ecology 79:2082–2091
van Ruijven J, Berendse F (2003) Positive effects of plant species diver-sity on productivity in absence of legumes. Ecol Lett 6:170–175
Vierheilig H, Lerat S, Piche Y (2003) Systemic inhibition of arbuscularmycorrhiza development by root exudates of cucumber plantscolonised by Glomus mosseae. Mycorrhiza 13:167–170
120 T. Radić et al.
Related Documents











