Oecologia (2009) 160:675–685 DOI 10.1007/s00442-009-1337-6 123 PLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER Nectar replenishment and pollen receipt interact in their eVects on seed production of Penstemon roseus Juan Francisco Ornelas · Carlos Lara Received: 23 April 2008 / Accepted: 20 March 2009 / Published online: 18 April 2009 Springer-Verlag 2009 Abstract Resource supply and pollen delivery are often thought to equally limit seed production in animal-polli- nated plants. At equilibrium, plants should show no response to experimental pollen supplementation because resources limit seed set above the current level of pollen attraction, while experimental reduction in pollen deposi- tion below the equilibrium level would reduce seed set. The predicted equilibrium may be disrupted, however, if plants expend additional energy to replenish removed nectar. We investigated the combined eVects of nectar removal and pollen delivery on female reproductive success of Penste- mon roseus (Plantaginaceae), a hummingbird-pollinated plant that replenishes removed nectar. We Wrst documented that the frequency of experimental nectar removal was cor- related with total nectar secretion; and increased frequency of nectar removal resulted in increased female reproductive costs to the plant. Trade-oVs between investing resources in nectar and investing resources in seeds were then investi- gated in two contrasting natural populations by removing nectar from Xowers at increasing frequencies while simul- taneously hand-pollinating Xowers with increasing amounts of pollen. Seed set was lowest at low levels of pollen depo- sition, highest at medium-sized pollen loads, and intermedi- ate when pollen loads were highest. At both sites, the frequency of nectar removal and pollen deposition had an interactive eVect on seed production, in that intermediate levels of nectar removal result in the absolute highest seed set, but only at intermediate pollen loads. At high pollen loads, seed set was higher following little to no nectar removal, and at low pollen loads, all rates of nectar removal aVected fecundity equally. Seed mass responded to nectar removal and pollination diVerently than did seed set. High levels of nectar removal and pollen delivery both lowered seed mass, with little interaction between main eVects. Our Wndings are among the Wrst to demonstrate that nectar replenishment costs and pollination intensity jointly aVect seed production. This conXict between nectar replenish- ment costs and pollen-limiting factors results in trade-oVs between pollinator attraction and seed provisioning. Thus, resource allocation towards nectar production should more often be considered in future studies of pollen limitation. Keywords Haig–Westoby model · Nectar replenishment · Pollen limitation · Penstemon · Mexico Introduction Theoretical models predict that seed set is equally limited by both pollen receipt and resource supply (Haig and West- oby 1988; Burd 2008). At equilibrium, pollen supplementa- tion experiments should not enhance seed production because resources limit seed set above the current level of pollen attraction, while experimental reduction in pollen receipt below the equilibrium level would reduce seed set (Ashman et al. 2004). Despite these predictions, many studies Communicated by Rebecca Irwin. J. F. Ornelas (&) Departamento de Biología Evolutiva, Instituto de Ecología, AC, Km 2.5 Carretera Antigua a Coatepec No. 351, Congregación El Haya, Apdo. Postal 63, 91070 Xalapa, Veracruz, Mexico e-mail: [email protected] C. Lara Laboratorio de Ecología de la Conducta, Centro Tlaxcala de Biología de la Conducta UAT-UNAM, Km 1.5 Carretera Tlaxcala-Puebla s/n, Colonia Xicohténcatl, Apdo. Postal 262, 90070 Tlaxcala, Tlaxcala, Mexico

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Oecologia (2009) 160:675–685DOI 10.1007/s00442-009-1337-6
123
PLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER
Nectar replenishment and pollen receipt interact in their eVects on seed production of Penstemon roseus
Juan Francisco Ornelas · Carlos Lara
Received: 23 April 2008 / Accepted: 20 March 2009 / Published online: 18 April 2009! Springer-Verlag 2009
Abstract Resource supply and pollen delivery are oftenthought to equally limit seed production in animal-polli-nated plants. At equilibrium, plants should show noresponse to experimental pollen supplementation becauseresources limit seed set above the current level of pollenattraction, while experimental reduction in pollen deposi-tion below the equilibrium level would reduce seed set. Thepredicted equilibrium may be disrupted, however, if plantsexpend additional energy to replenish removed nectar. Weinvestigated the combined eVects of nectar removal andpollen delivery on female reproductive success of Penste-mon roseus (Plantaginaceae), a hummingbird-pollinatedplant that replenishes removed nectar. We Wrst documentedthat the frequency of experimental nectar removal was cor-related with total nectar secretion; and increased frequencyof nectar removal resulted in increased female reproductivecosts to the plant. Trade-oVs between investing resources innectar and investing resources in seeds were then investi-gated in two contrasting natural populations by removingnectar from Xowers at increasing frequencies while simul-taneously hand-pollinating Xowers with increasing amounts
of pollen. Seed set was lowest at low levels of pollen depo-sition, highest at medium-sized pollen loads, and intermedi-ate when pollen loads were highest. At both sites, thefrequency of nectar removal and pollen deposition had aninteractive eVect on seed production, in that intermediatelevels of nectar removal result in the absolute highest seedset, but only at intermediate pollen loads. At high pollenloads, seed set was higher following little to no nectarremoval, and at low pollen loads, all rates of nectar removalaVected fecundity equally. Seed mass responded to nectarremoval and pollination diVerently than did seed set. Highlevels of nectar removal and pollen delivery both loweredseed mass, with little interaction between main eVects. OurWndings are among the Wrst to demonstrate that nectarreplenishment costs and pollination intensity jointly aVectseed production. This conXict between nectar replenish-ment costs and pollen-limiting factors results in trade-oVsbetween pollinator attraction and seed provisioning. Thus,resource allocation towards nectar production should moreoften be considered in future studies of pollen limitation.
Keywords Haig–Westoby model · Nectar replenishment · Pollen limitation · Penstemon · Mexico
Introduction
Theoretical models predict that seed set is equally limitedby both pollen receipt and resource supply (Haig and West-oby 1988; Burd 2008). At equilibrium, pollen supplementa-tion experiments should not enhance seed productionbecause resources limit seed set above the current level ofpollen attraction, while experimental reduction in pollenreceipt below the equilibrium level would reduce seed set(Ashman et al. 2004). Despite these predictions, many studies
Communicated by Rebecca Irwin.
J. F. Ornelas (&)Departamento de Biología Evolutiva, Instituto de Ecología, AC, Km 2.5 Carretera Antigua a Coatepec No. 351, Congregación El Haya, Apdo. Postal 63, 91070 Xalapa, Veracruz, Mexicoe-mail: [email protected]
C. LaraLaboratorio de Ecología de la Conducta, Centro Tlaxcala de Biología de la Conducta UAT-UNAM, Km 1.5 Carretera Tlaxcala-Puebla s/n, Colonia Xicohténcatl, Apdo. Postal 262, 90070 Tlaxcala, Tlaxcala, Mexico

676 Oecologia (2009) 160:675–685
123
have detected pollen limitation in natural populations, i.e.,increased seed production with increasing pollen receipt(e.g., Campbell 1985; Galen 1985; Johnston 1991; Camp-bell and Halama 1993), and others have detected resourcelimitation through experimental additions of water or nutri-ents (reviewed in Stephenson 1981; Zimmerman 1983).Extensive reviews of pollen-limitation studies reveal thatHaig and Westoby’s model rarely Wts, that is, pollen sup-plementation often increases (Burd 1994; Larson and Bar-rett 2000; Ashman et al. 2004; Knight et al. 2005) andsometimes decreases (Young and Young 1992) fruit or seedproduction. Such responses to supplementation suggest thatmost populations are not at the equilibrium level purport-edly favored by natural selection. Departures from equilib-rium were also predicted by Haig and Westoby (1988), andcan be produced under stochastic environmental variation,so that pollen supplementation would yield increased seedset even if the population is, on average, at the expectedequilibrium (Burd 2008). For instance, recent ecologicalchanges in the pollination or resource environment such ashabitat fragmentation, or decrease in population size, orloss of native pollinators (reviewed in Ashman et al. 2004),may produce stochastic variation among Xowers in pollenreceipt and eventually may produce populations to divergefrom Haig and Westoby equilibrium (Burd 1995). Demon-strations of how stochastic pollination and resource envi-ronments inXuence this equilibrium are nevertheless scarce.
The interaction between the eVects of adding pollen andresources on seed production is widely accepted (e.g.,McCall and Primack 1987; Ehrlén 1992; Lawrence 1993;Mattila and Kuitunen 2000; Totland 2001); however,resources that contribute to attracting and rewarding pollina-tors could additionally inXuence pollination levels indirectly(e.g., Zimmerman and Pyke 1988a, b; Mitchell and Waser1992). Traditional pollen supplementation experiments oftenincompletely describe the pollen-seed-set curve posed byHaig and Westoby (1988), because at least one importantindirect cost of seed production is excluded. Plants thatreceive experimental pollen supplementation do not pay thecost of attracting [and rewarding] additional pollinationunder natural conditions and therefore have more resourcesavailable for seed maturation (Ashman et al. 2004). Thus, tocorrectly evaluate whether a population is at its equilibriumvalue for traits inXuencing pollinator service (Ashman et al.2004), the additional costs of replenishing removed nectarmust also be measured. Here we examined the predictedequilibrium of the Haig and Westoby graphical model byremoving nectar from Xowers at increasing frequencieswhile simultaneously hand-pollinating Xowers with increas-ing amounts of pollen in a factorial design.
The purpose of our study was to examine the combinedeVects of nectar removal and pollen delivery on the femalereproductive success of Penstemon roseus, a hummingbird-
pollinated plant species that replenishes removed nectar. Ifnectar production costs interact in their eVects with pollenreceipt, then increases in seed production with increasingpollen receipt should be limited by the amount of resourcesdevoted to nectar production costs. Because the cost ofnectar replenishment is not always maternally expensive(Ordano and Ornelas 2005), we undertook two Weld experi-ments. In the Wrst, we measured the direct cost of nectarreplenishment in terms of seed production. In the secondexperiment, we manipulated the frequency of nectarremoval and pollination intensity to assess trade-oVs withseed production. The experiments were aimed at addressingthe following speciWc questions:
1. Does increased nectar removal result in an increasedsubsequent nectar production by individual Xowers?
2. Does experimentally increased nectar replenishmentaVect seed production?
3. Are there trade-oVs between investing resources in nec-tar replenishment and investing resources in developingseeds (is pollen limitation of seed set aVected by theamount of resources devoted to nectar production costs)?
Materials and methods
System description
Study species
Penstemon roseus (Cerv. ex Sweet) G. Don (Plantagina-ceae) is a perennial, endemic to Mexico, commonly found inWr, oak and cloud forests (2,200–3,900 m above sea level;Calderón and Rzedowski 2001). Individuals of P. roseus(0.4–1.2 m high) bear ten to 20 paniculate inXorescences,each with two to four pendant Xowers from terminal branch-ing stems opening per day, and ca. 80 Xoral buds asynchro-nously reach the Xower stage during the blooming season(3 months), which extends from July to December for theregion; there is large overlap between fruit production of theWrst Xowers and anthesis of the later Xowers. The self-com-patible, hermaphroditic, magenta tubular Xowers (ca. 28 mmcorolla length) are protandrous, with 1–2 days in the malephase (staminate) then 1–2 days in the female phase (pistil-late) (Lara and Ornelas 2008). In our samples, ovules perovary average 182 § 0.8 SE (n = 60 Xowers). Althoughmanually outcrossed Xowers produce twice as many seedsas when manually self-pollinated (Lara and Ornelas 2008),outcrossed hand-pollination yields half seed set in P. roseus,which suggests limitation on the resources to mature seedsor that stigmas on these outcrossed pollinations were pro-vided with insuYcient and/or low-quality pollen. In ourstudy area, P. roseus Xowers are visited by bumblebees and

Oecologia (2009) 160:675–685 677
123
butterXies, and most frequently by eight hummingbird spe-cies (Lara 2006; Lara and Ornelas 2008). The most frequenthummingbird is the resident Hylocharis leucotis (ca. 50% ofXoral visits). Unlike most Penstemon species, P. roseusreceives low levels of pollen receipt (10–30 pollen grainsper stigma) and pollen removal (20–30% of pollen pre-sented), and a long time elapses before a hummingbird visit(100–150 min), suggesting low pollinator traYc and pollenshortages in this species. Daily nectar secretion rates rangefrom 0.3 to 0.6 mg of sugar per Xower in P. roseus. Basedon nectar accumulation measures, P. roseus Xowers produce0.63 § 0.063 !l of Xuid and 0.202 § 0.007 mg of sugarafter 24 h of accumulation (mean § SE, n = 30 Xowers,three Xowers from each of ten plants). In 48 h, Xowers accu-mulate twice as much nectar (1.497 § 0.083 !l and0.299 § 0.008 mg, n = 30) and, as in 48 h, produce aboutthe same amount of nectar after 72 (1.33 § 0.066 !l and0.301 § 0.008 mg, n = 30) and 120 h of accumulation(1.242 § 0.076 !l and 0.218 § 0.005 mg, n = 30). Thesedata indicate that little evaporation and/or resorption occursin this species. Flowers respond to repeated nectar removalby producing more nectar (higher volume and more totalsugar); however, our previous nectar-removal study did notinclude control Xowers emptied of nectar once at the end oftheir life span (Lara and Ornelas 2008).
Study sites
Fieldwork was carried out with two populations ofP. roseus from central Mexico during the 2006 and 2007Xowering periods (July–November):
1. Parque Nacional La Malinche (19°14!N, 98°58!W;2,900 m above sea level) is a protected area (45,711 ha)of pine, oak and sacred Wr forest, 80 km from TlaxcalaCity, Tlaxcala, Mexico. Mean annual precipitation is850 mm and mean annual temperature is 13°C (López-Domínguez and Acosta 2005).
2. Acuitlapilco (19°17!N, 98°14!W; 2,200 m above sealevel) is a heavily disturbed area with forested rem-nants located 3 km from the city of Tlaxcala. Meanannual precipitation is 700 mm and mean annual tem-perature is 16°C (Ritter et al. 2004).
These localities represent two extremes of the environmen-tal gradient where P. roseus lives.
Experimental design
Experiment 1: eVects of nectar replenishment on seed production
In this experiment we examined the eVects of repeated nec-tar removal on total nectar production and its eVects on
seed production of P. roseus Xowers. In July 2006, werandomly selected 30 P. roseus plants at La Malinche.InXorescences with fully developed buds were baggedthe evening before nectar measurements. Then, recentlyopened Xowers per plant were subjected to one of the fol-lowing treatments: one removal, nectar was removed onceat the end of the Xowers’ lifetime in which the Xowers werein day 2 of the pistillate phase (1); two removals, nectarwas removed once a day (at 0900 hours) over 2 days inwhich the Xowers were in the staminate phase (2S); tworemovals, nectar was removed once a day (at 0900 hours)over 2 days in which the Xowers were in the pistillate phase(2P); four removals, nectar was removed once a day (at0900 hours) over 4 days in which the Xowers were 2 daysin the staminate phase and 2 days in the pistillate phase (4);four removals, nectar was removed twice a day (at 0900and 1300 hours) over 2 days in which the Xowers were inthe staminate phase (4S); four removals, nectar wasremoved twice a day (at 0900 and 1300 hours) over 2 daysin which the Xowers were in the pistillate phase (4P); eightremovals, nectar was removed twice a day (at 0900 and1300 hours) over 4 days in which the Xowers were 2 daysin the staminate phase and 2 days in the pistillate phase (8);no removal, plants were exposed for 4 days to natural levelsof nectar removal (control, unbagged Xowers) (0). Nectarvolume per Xower was measured using graduated micropi-pettes (5 !l) and a ruler, sugar concentration (percentagesucrose) with a pocket refractometer (American Optical10431, BuValo, N.Y.; range concentration 1–50°, BRIXunits), and the amount of sugar produced (milligrams) cal-culated according to Bolten et al. (1979). Nectar wasextracted carefully without removing the Xowers from theplants. Flowers remained bagged between probes.
Bagged Xowers subjected to experimental nectar removalwere manually outcrossed at day 1 of their pistillate phase(period of peak stigma receptivity; Lara and Ornelas 2008).Hand pollinations were achieved by smearing one antherfrom randomly selected plant donors onto the receptivevirgin stigma. Flowers remained bagged after pollination.Pollen grains deposited on individual stigmas by hand-pollination were not counted, but half the ovules are fertil-ized when smearing one anther on the stigma (Lara andOrnelas 2008). The control for the eVects of micropipetteinsertion (plugged micropipettes are inserted into the Xowerbut nectar is not removed; see Ordano and Ornelas 2005),was not included in this study because we previouslyshowed that our manipulations did not injure Xowers andno signiWcant eVects of micropipette insertion on nectar orseed production occurred in P. roseus (Lara and Ornelas2008). The treatment in which a Xower is emptied once atthe end of its life is an attempt to determine whether or nottotal nectar production in all other treatments applied (rang-ing from 2 to 8 in this experiment) was greater, due to

678 Oecologia (2009) 160:675–685
123
replenishment, than if a Xower was not visited at all(Castellanos et al. 2002; Ordano and Ornelas 2004). None-theless, potential problems with this treatment due to evap-oration and/or resorption may occur in P. roseus Xowers(see also Búrquez and Corbet 1991); nectar secretion isdynamically modiWed by the plant to ensure visits by poll-inators and/or to reallocate resources especially in post-fer-tilization development of ovules (Nepi and Stpiczynska2007). The natural unbagged treatment acts as a control forseed production, although the mechanisms aVecting seedproduction in this treatment are unclear and may be due toresource allocation or pollen load, or both. Fruit capsuleswere harvested in October and seeds per capsule weighedand counted under a microscope. We used 30 Xowers pertreatment, a total of 210 Xowers for nectar measurementsand 240 Xowers for seed production.
To evaluate the eVects of replenishment after repeatedremoval on seed production, we used nested ANOVAs withtype III sum of squares on nectar volume, sugar production,seed number, and total seed mass. Nectar volume and totalsugar produced and seed number and total seed mass herewere intercorrelated response variables (volume vs. sugar,r = 0.87, n = 210, P = 0.0001; seed number vs. total seedmass, r = 0.31, n = 240, P = 0.0001). Therefore, we Wrstperformed nested multivariate ANOVAs (MANOVAs)incorporating these response variables. In the model nectar-removal treatment was a Wxed factor and plant factor wasnested within treatment. Using a MANOVA followed byunivariate ANOVAs, as described above, will reduce theprobability of inXating the type I error rate. Nectar data andtotal seed mass were log10(x + 1) transformed and seednumber data were transformed using a square-root transfor-mation before statistical analyses to achieve normality.
Experiment 2: the combined eVects of nectar removal and pollen deposition on seed production
To simulate nectar removal and pollen deposition, wesimultaneously outcrossed hand-pollinated Xowers addingdiVerent amounts of pollen (pollination intensity) to Xowersof P. roseus, which were diVerently emptied of nectar (fre-quency of nectar removal). In August 2007, an outcrossedhand-pollination experiment was conducted with 60 ran-domly selected plants, 30 at La Malinche (12–14 August)and 30 at Acuitlapilco (16–18 August). We followed thesame general protocol for exclusion of Xoral visitors, as inexperiment 1. However, Xowers at Acuitlapilco had short-ened lifetimes (3 days), 1 day in each sexual phase and1 day with sexual phases intermingled. So, we adjusted ourexperimental protocol there by reducing the overallremoval frequency. Nectar and pollination treatments wereassigned to each of 30 plants per site in a replicated crosseddesign where each level of the nectar-removal treatment
was combined with each level of pollen addition (4 £ 3 =12 treatment combinations applied to ten Xowers each,n = 120 Xowers). Frequency of nectar removal included thefollowing treatment levels: (1) one removal, nectar wasremoved once (at 0900 hours) at the end of Xower’s life-span, i.e., the Xower was in day 2 of the pistillate phase(low frequency); (2) three or four removals, nectar wasremoved once a day (at 0900 hours) over 3 or 4 days(medium frequency); or (3) six or eight removals, nectarwas removed twice a day (at 0900 and 1300 hours) over 3or 4 days in which the Xower was 1–2 days in the staminatephase and 1–2 days in the pistillate phase (high frequency).Hummingbirds visiting P. roseus surely drain Xowers com-pletely as Xowers with 5–10 !l of nectar are completelydrained by a hummingbird in 5-s foraging bouts (n = 100Xowers; unpublished data).
Repeatedly emptied Xowers in a given plant were out-crossed hand-pollinated early in the morning at day 1 of thepistillate phase by smearing: (1) one anther from one plantdonor (low frequency), (2) two anthers from two plantdonors (medium frequency), or (3) four anthers also fromtwo plant donors (high frequency) onto virgin stigmas. Dis-tances between randomly selected plant donors and recipi-ent plants ranged from 20 to 50 m. Because pollendeposition in P. roseus wild plants may diVer from that ofthe applied treatments, we quantiWed how much pollen wasactually applied on stigmas. The application of one anthersmeared on a stigma results in 140 § 7 SE (n = 5) pollengrains. When two or four anthers were smeared on stigmas,the number of pollen grains applied increases up to257 § 11 (n = 5) and 847 § 59 (n = 5), respectively. Like-wise, the number of pollen grains deposited on stigmas is32 § 12 (n = 5) when Xowers are probed once by Hylo-charis leucotis and 148 § 26 (n = 5) when probed by Selas-phorus platycercus. When Xowers are probed 2–4 times thenumber of pollen grains deposited on stigmas increases upto 130 § 20 (n = 5) and 396 § 27 (n = 5) if visited byH. leucotis, and 272 § 98 (n = 5) and 868 § 148 (n = 5) ifvisited by S. platycercus. These data also indicate that ourpollination manipulations simulate natural variation in pol-len receipt. An unbagged Xower per plant (n = 30 Xowers)was also exposed to natural levels of pollination and nectarremoval. Again, the natural unbagged treatment acts as acontrol for seed production. Because plants can reallocateresources among Xowers (Zimmerman and Pyke 1988a), anunbagged open-pollinated control was also applied to oneXower on each of ten additional non-treated plants per site(Zimmerman 1988). Then, our experimental set-up usesone control on the treated plant and one on the control non-treated plant. Unbagged Xowers from treated and non-treated plants were exposed for 4 days to natural levels ofpollination and nectar removal. The mechanisms aVectingseed production in the natural unbagged treatment are

Oecologia (2009) 160:675–685 679
123
unclear and may be due to resource allocation or pollenload, or both. To avoid this problem, an additional treat-ment was added on treated plants, in which Xowers werepollinated then bagged, with no nectar removal. This addi-tional treatment was included to disassociate the eVects ofnectar removal from those of pollination intensity. Experi-mental Xowers were re-bagged after pollinations andremained bagged until fruit production. Fruit capsules wereharvested 6 weeks later and seeds per capsule wereweighed and counted under a microscope. We used a totalof 160 Xowers and 40 plants per site for seed production,including the control treatment on additional non-treatedplants.
The eVects of replenishment after repeated removal andpollination addition, and their interaction on seed produc-tion were tested in factorial ANOVAs with type III sum ofsquares on seed number and total seed mass. We again Wrstperformed two-way MANOVAs incorporating these tworesponse variables to explicitly account for the possiblecorrelation between the dependent variables, followed bytwo-way ANOVAs as described above. We used separateanalyses for each population because frequency of nectarremoval was adjusted due to the site diVerences in Xowerlongevity. Seed production of Xowers exposed to naturalpollination was then contrasted with that from experimentalXowers using post hoc mean comparisons (Tukey’s honestsigniWcant diVerence methods, ! = 0.05). Total seed masswas log10(x + 1) transformed and seed number data weretransformed using a square-root transformation beforestatistical analyses to achieve normality.
Results
Experiment 1: eVects of nectar replenishment on seed production
Repeated nectar removal aVected subsequent nectarproduction positively in P. roseus. A MANOVA showedthat nectar-removal treatment signiWcantly inXuenced totalnectar production for both dependent variables (volumeand amount of sugar, Wilks’ " = 0.026, F12,348 = 0.015,P = 0.0001). Plant individuals (nested within treatments)were not signiWcantly heterogeneous for the two dependentvariables (Wilks’ " = 0.732, F56,348 = 1.044, P = 0.397).Because the MANOVA was signiWcant, we then followedwith univariate ANOVAs. Total volume produced variedsigniWcantly with number of removals, increasing withremoval intensity (F6,175 = 222.12, P = 0.0001). The cumu-lative nectar volume produced by Xowers with the highestremoval frequency (8 times) was about 6 times as high asthat produced by the control (Xowers emptied once). Flow-ers emptied of nectar 2 times, 4 times or 8 times produced
signiWcantly more nectar than Xowers emptied once at theend of their lives (Fig. 1a). Total sugar production also var-ied signiWcantly with frequency of nectar removal(F6,175 = 582.02, P = 0.0001), increasing 2.8 times more inXowers with highest nectar removal than in the control.However, Xowers emptied 4 times (once a day) produced asmuch sugar as the control Xowers emptied once (Fig. 1b).No signiWcant diVerences (P > 0.01) in nectar replenish-ment were observed between sexual phases in Xowers emp-tied 2 or 4 times (Fig. 1).
Seed production was negatively aVected with repeatednectar removal. The MANOVA results showed that nectar-removal treatment signiWcantly inXuenced total seed pro-duction for both dependent variables (seed number andtotal seed mass, Wilks’ " = 0.023, F14,398 = 156.86,P = 0.0001). Again, plant individuals (nested within treat-ment) were not signiWcantly heterogeneous for the twodependent variables (Wilks’ " = 0.726, F64,398 = 1.078,P = 0.329). Because the MANOVA was signiWcant, wethen followed with univariate ANOVAs. Both seed numberand total seed mass varied signiWcantly with number ofremovals, decreasing with removal intensity (seed number,F7,232 = 363.34, P = 0.0001; total seed mass, F7,232 = 67.51,P = 0.0001). Fruit capsules from manually cross-pollinated,Xowers emptied 8 or 4 times during the pistillate phase pro-duced ca. 20 fewer seeds than the control Xowers emptiedonce or other treatment levels (Fig. 1c). Fewer seeds wereproduced when nectar-removal frequency increased duringthe pistillate phase compared to when nectar was repeatedlyremoved during the staminate phase (treatments 4P and 4S;Fig. 1c). Higher seed production among Xowers exposed tonatural visitation indicates that our hand-pollination levels(smearing one anther) were not enough to ensure fertiliza-tion of all ovules or that our manipulations had direct nega-tive eVects on seed production. Lastly, total seed massdecreased ca. one-third when more than one removal wasapplied per day throughout a Xower’s lifetime (Fig. 1d).
Experiment 2: the combined eVects of nectar removal and pollen deposition on seed production
The results from two-way MANOVAs showed thatfrequency of nectar removal and pollination intensity inXu-enced seed production in both sites (nectar-removal eVect,La Malinche, Wilks’ " = 0.291, F6,290 = 41.28, P = 0.0001,Acuitlapilco, Wilks’ " = 0.319, F6,290 = 37.19, P = 0.0001;pollination eVect, La Malinche, Wilks’ " = 0.100, F4,290 =0.02, P = 0.0001, Acuitlapilco, Wilks’ " = 0.201, F4,290 =89.02, P = 0.0001). Furthermore, the nectar removal £ pol-lination intensity interaction was signiWcant for La Malin-che (Wilks’ " = 0.608, F12,290 = 6.81, P = 0.0001) andAcuitlapilco (Wilks’ " = 0.822, F12,290 = 2.49, P = 0.004).Two-way ANOVAs showed that seed number in P. roseus

680 Oecologia (2009) 160:675–685
123
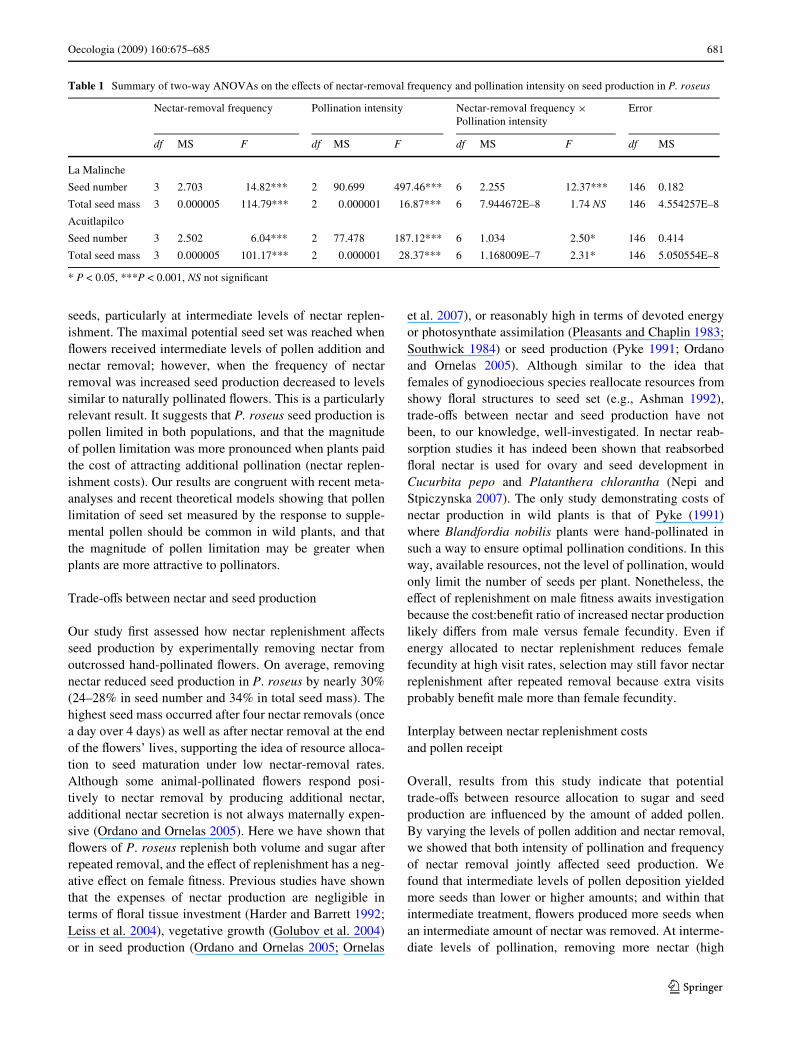
was signiWcantly aVected by frequency of nectar removaland pollination intensity, and their interactions were alsosigniWcant in both sites (Table 1). Flowers that receivedintermediate levels of pollen addition and nectar removalproduced more seeds than those that received low, high, ornatural levels of pollination and nectar removal in both sites(Fig. 2). Hand-pollinated Xowers with intermediate pollina-tion levels produced signiWcantly more seeds than thosewith lower or higher pollination intensities in both sites,and signiWcantly more than those under natural circum-stances (controls in treated and non-treated plants). Whennectar removal was intermediate in frequency (three orfour removals, once a day over 3 or 4 days), Xowers pro-duced signiWcantly more seeds than those that receivedother treatment levels, including those with no nectarremoval and pollinated. Again, when hand-pollinationswere increased, seed production decreased signiWcantly anddecreases were signiWcantly higher when nectar-removalfrequencies were higher (six to eight removals, twice a dayover 3 or 4 days; Fig. 2). Interestingly, Xowers with no nec-tar removal and higher pollination levels produced signiW-cantly fewer seeds than those with intermediate pollinationlevels. However, these Xowers produced signiWcantly moreseeds than those with higher nectar-removal frequencies,suggesting an independent negative eVect of pollinationintensity (Fig. 2).
For total seed mass, hand-pollinations with mixed pollenloads yielded diVerent results. Total seed mass is moreinformative, since that takes into account both diVerencesin the number and the weight (quality) of individual seeds.Two-way ANOVAs showed that total seed mass inP. roseus was signiWcantly aVected by frequency of nectarremoval and pollination intensity, and their interactionswere only signiWcant at Acuitlapilco (Table 1). A signiW-cant increase in total seed mass was observed in hand-polli-nated Xowers with low and intermediate pollination levelscompared to those with higher pollination intensities inboth sites, and to those exposed to natural levels of pollina-tion and nectar removal (Fig. 3). Total seed mass signiW-cantly decreased in Xowers with higher nectar-removalfrequency (six to eight removals, twice a day over 3 or4 days), regardless of pollination intensity.
Discussion
Our Wndings suggest that pollination intensity and nectarreplenishment interact in their eVects on seed production.By manipulating rates of nectar replenishment and patternsof pollen receipt, we found evidence for a trade-oV betweeninvesting resources in nectar and investing resources in
Fig. 1 EVects of nectar removal in Penstemon roseus. a Nectar pro-duction (volume), b sugar production, c seed number, and d total seedmass. Data are means § 1 SE of untransformed values. n = 30 plants(30 Xowers/treatment) and a total of 210 Xowers for nectar measure-ments and 240 Xowers for seed production. DiVerent letters indicatediVerences between groups based on Tukey’s honest signiWcant diVer-ence (HSD) methods (P < 0.05). 1 Flowers emptied once at the end oftheir lives, 2S Xowers emptied once a day over 2 days of the staminatephase, 2P Xowers emptied once a day over 2 days of the pistillatephase, 4 Xowers emptied once a day over 4 days, 4S Xowers emptiedtwice a day over 2 days of the staminate phase, 4P Xowers emptiedtwice a day over 2 days of the pistillate phase, 8 Xowers emptied twicea day over 4 days, O unbagged Xowers exposed to natural levels ofpollination and nectar removal
Oecologia (2009) 160:675–685 681
123
seeds, particularly at intermediate levels of nectar replen-ishment. The maximal potential seed set was reached whenXowers received intermediate levels of pollen addition andnectar removal; however, when the frequency of nectarremoval was increased seed production decreased to levelssimilar to naturally pollinated Xowers. This is a particularlyrelevant result. It suggests that P. roseus seed production ispollen limited in both populations, and that the magnitudeof pollen limitation was more pronounced when plants paidthe cost of attracting additional pollination (nectar replen-ishment costs). Our results are congruent with recent meta-analyses and recent theoretical models showing that pollenlimitation of seed set measured by the response to supple-mental pollen should be common in wild plants, and thatthe magnitude of pollen limitation may be greater whenplants are more attractive to pollinators.
Trade-oVs between nectar and seed production
Our study Wrst assessed how nectar replenishment aVectsseed production by experimentally removing nectar fromoutcrossed hand-pollinated Xowers. On average, removingnectar reduced seed production in P. roseus by nearly 30%(24–28% in seed number and 34% in total seed mass). Thehighest seed mass occurred after four nectar removals (oncea day over 4 days) as well as after nectar removal at the endof the Xowers’ lives, supporting the idea of resource alloca-tion to seed maturation under low nectar-removal rates.Although some animal-pollinated Xowers respond posi-tively to nectar removal by producing additional nectar,additional nectar secretion is not always maternally expen-sive (Ordano and Ornelas 2005). Here we have shown thatXowers of P. roseus replenish both volume and sugar afterrepeated removal, and the eVect of replenishment has a neg-ative eVect on female Wtness. Previous studies have shownthat the expenses of nectar production are negligible interms of Xoral tissue investment (Harder and Barrett 1992;Leiss et al. 2004), vegetative growth (Golubov et al. 2004)or in seed production (Ordano and Ornelas 2005; Ornelas
et al. 2007), or reasonably high in terms of devoted energyor photosynthate assimilation (Pleasants and Chaplin 1983;Southwick 1984) or seed production (Pyke 1991; Ordanoand Ornelas 2005). Although similar to the idea thatfemales of gynodioecious species reallocate resources fromshowy Xoral structures to seed set (e.g., Ashman 1992),trade-oVs between nectar and seed production have notbeen, to our knowledge, well-investigated. In nectar reab-sorption studies it has indeed been shown that reabsorbedXoral nectar is used for ovary and seed development inCucurbita pepo and Platanthera chlorantha (Nepi andStpiczynska 2007). The only study demonstrating costs ofnectar production in wild plants is that of Pyke (1991)where Blandfordia nobilis plants were hand-pollinated insuch a way to ensure optimal pollination conditions. In thisway, available resources, not the level of pollination, wouldonly limit the number of seeds per plant. Nonetheless, theeVect of replenishment on male Wtness awaits investigationbecause the cost:beneWt ratio of increased nectar productionlikely diVers from male versus female fecundity. Even ifenergy allocated to nectar replenishment reduces femalefecundity at high visit rates, selection may still favor nectarreplenishment after repeated removal because extra visitsprobably beneWt male more than female fecundity.
Interplay between nectar replenishment costs and pollen receipt
Overall, results from this study indicate that potentialtrade-oVs between resource allocation to sugar and seedproduction are inXuenced by the amount of added pollen.By varying the levels of pollen addition and nectar removal,we showed that both intensity of pollination and frequencyof nectar removal jointly aVected seed production. Wefound that intermediate levels of pollen deposition yieldedmore seeds than lower or higher amounts; and within thatintermediate treatment, Xowers produced more seeds whenan intermediate amount of nectar was removed. At interme-diate levels of pollination, removing more nectar (high
Table 1 Summary of two-way ANOVAs on the eVects of nectar-removal frequency and pollination intensity on seed production in P. roseus
* P < 0.05, ***P < 0.001, NS not signiWcant
Nectar-removal frequency Pollination intensity Nectar-removal frequency £Pollination intensity
Error
df MS F df MS F df MS F df MS
La Malinche
Seed number 3 2.703 14.82*** 2 90.699 497.46*** 6 2.255 12.37*** 146 0.182
Total seed mass 3 0.000005 114.79*** 2 0.000001 16.87*** 6 7.944672E–8 1.74 NS 146 4.554257E–8
Acuitlapilco
Seed number 3 2.502 6.04*** 2 77.478 187.12*** 6 1.034 2.50* 146 0.414
Total seed mass 3 0.000005 101.17*** 2 0.000001 28.37*** 6 1.168009E–7 2.31* 146 5.050554E–8

682 Oecologia (2009) 160:675–685
123
frequency) reduced seed production in P. roseus on averageby nearly 30% at both sites. When nectar removal wasincreased there was a detrimental eVect in terms of seedproduction, i.e., an increase in pollen deposition increasedseed production to a point where nectar replenishmentbecame presumably costly for the maturation of additionalseeds. The control of no nectar removal and diVerent polli-nation intensities helped to untangle the eVect of nectar
removal and the eVect of pollination intensity; seed produc-tion did not increase linearly by increasing pollinationintensity without nectar removal, which conWrmed the inde-pendent eVect of pollination intensity.
Our earlier hand-pollination studies investigating thecosts of nectar production (Ordano and Ornelas 2005; Orn-elas et al. 2007; Lara and Ornelas 2008) have not controlledand/or varied the amount of pollen received, and so havenot determined whether or not the excess of pollen deposi-tion reduces seed production when plants are forced toreplenish nectar. It is likely that excessive pollen deposi-tion, removal of previously deposited pollen, and/or exces-sive nectar removal interfered with maximal seedproduction in P. roseus. A negative eVect of overpollina-tion (e.g., stigma clogging and pollen interference) isunlikely because the unbagged Xowers did not diVer fromthose treated with the higher pollination intensity andnectar-removal frequency. Normally, an excess of pollenwould lead to higher seed set unless the deposited pollenhas a genetic eVect (i.e., inbreeding). Also, success of hand-pollination experiments may have overestimated the eVectsof pollen deposition on seed production by applying pure
Fig. 2 Combined eVects of nectar removal and pollen deposition onseed number for wild plants at a La Malinche and b Acuitlapilco. Dataare means § 1 SE across fruits of untransformed values. Nectar-re-moval frequency: Xowers emptied once at the end of their lives (Low),Xowers emptied once a day over 3 or 4 days (Medium), Xowers emp-tied twice a day over 3 or 4 days (High), bagged Xowers with no nectarremoval and pollinated (No). Repeatedly emptied Xowers were out-crossed hand-pollinated with mixed pollen loads. Pollination intensity:stigmas smeared once with pollen from one anther (Low), stigmassmeared once with pollen from two anthers (Medium), stigmassmeared once with pollen from four anthers (High). Unbagged Xowersfrom the same treated plants and from additional non-treated plantswere exposed for 4 days to natural levels of pollination and nectarremoval. DiVerent letters indicate diVerences between groups based onTukey’s HSD methods (P < 0.05). n = 40 plants (30 treated and tennon-treated) and 160 Xowers at each site (ten bagged Xowers per treat-ment combination, 30 unbagged Xowers from treated plants, tenunbagged Xowers from additional non-treated plants)
Fig. 3 Combined eVects of nectar removal and pollen deposition ontotal seed mass for wild plants at a La Malinche and b Acuitlapilco. Fortreatments and abbreviations, see Fig. 2

Oecologia (2009) 160:675–685 683
123
outcross pollen (see Ashman et al. 2004), and potentiallyincreased the intensity of pollen competition when pollenwas delivered simultaneously. In the two natural popula-tions of P. roseus, opportunities for pollen competitionseemed very limited because low pollen deposition rateshad been observed (ca. 15–30 pollen grains per stigma over3 days; Lara and Ornelas 2008). The use of one or severalanthers from the same plant donor to conduct outcrosstreatments per Xower presents the possibility of incompati-bility in speciWc crosses. However, when the combinedeVects of nectar removal and pollen deposition on seed pro-duction were investigated, pollen donation was done withpollen mixtures from two diVerent randomly selected focalplants (intermediate and high pollination treatments). Thus,inbreeding or other sources of incompatibility can be dis-carded in this study, except in the low pollination treatmentwhere quality (one anther from one plant donor) and quan-tity of pollen may be confounded. Nonetheless, our resultshighlight the need to further study pollen performance andpollen competition under natural conditions where stigmassurely end up with mixed pollen loads composed of self andoutcross pollen (Lara and Ornelas 2008). Given that Xowersare not injured by micropipette insertion and no signiWcanteVects of micropipette insertion on nectar or seed produc-tion were detected (Lara and Ornelas 2008), the strongestdrop-oVs in seed set at high nectar-removal levels suggeststhat pollen augmentation and nectar removal are having asynergistic eVect on seed set. Nonetheless, damage to thestigma as an alternative mechanism may be driving the eVect.
Natural nectar-removal rates as well as pollen depositionby bumblebees and hummingbirds in P. roseus wild plantsboth depend on pollinator visit rates, pollen carrying capac-ities, and preferences, and may diVer from those of theapplied treatments. Some studies have shown a peak inseed production at intermediate levels of pollinator visita-tion, with a decrease in seed production at higher visitationlevels (Búrquez et al. 1987; Young 1988; Herre 1990) thatmay result in the removal of previously deposited pollen onthe stigma (i.e., Gori 1983). We minimized this variation inforaging behavior by excluding pollinators from theseexperiments and assumed that our treatment levels closelymatched natural hummingbird visits. Also, the averagenumbers of seeds per fruit produced under natural (100–120) and controlled conditions (140–160) were lower thanthe number of ovules per Xower to fertilize (ca. 180 ovulesper Xower in both sites). Furthermore, low rates of pollenremoval may be common for hummingbird-visited Xowersbecause of short probe duration and lack of movementwithin Xowers while probing (Mitchell and Waser 1992).Therefore, unless stigmas end up heavily loaded with selfversus outcross pollen, the low seed production and pollendeposition rates observed under natural conditions suggestpollen limitation in P. roseus.
Departures from the Haig–Westoby equilibrium
The Haig and Westoby’s (1988) graphical model predictsthat seed set is equally limited by both pollen receipt andresource supply. At equilibrium, plants should show noresponse to pollen addition, because resources limit seed setabove the current level of pollen attraction, while experi-mental reduction in pollen receipt below the equilibriumlevel would reduce seed set (Ashman et al. 2004). Despitethese predictions, later meta-analyses of pollen supplemen-tation experiments showed that the magnitude of pollenlimitation of seed set is overestimated (Knight et al. 2006)but common and taxonomically widespread in plants (Burd1994; Larson and Barrett 2000; Ashman et al. 2004; Knightet al. 2005). In a provocative review, Ashman et al. (2004)propose two possible explanations for the incongruencebetween theoretical predictions and experimental results:that wild populations are generally not in equilibrium, orthat they are in an equilibrium that is determined by a sto-chastic rather than a constant pollination environment. Theresults of our second experiment are congruent with Ash-man’s et al. (2004) second explanation that both popula-tions of P. roseus are in an equilibrium determined by astochastic pollination environment.
Stochastic variation around pollen receipt can be gener-ated by the spatial proximity of Xowering individuals, poll-inators’ foraging behavior, composition of mixed stigmaticpollen loads, abundance of eVective pollinators, loss ofnative pollinators, and habitat fragmentation (reviewed inAshman et al. 2004; Burd 2008), and may produce popula-tions which diverge from Haig–Westoby equilibrium.Departures from equilibrium were brieXy explored by Haigand Westoby (1988), and theoretically examined by Burd(2008) incorporating stochastic environmental variation inthe resource and fertilization functions that describe theHaig–Westoby graphical model. Burd’s (2008) modiWedmodel makes two general predictions: pollen limitation ofseed set measured by the response to supplemental pollenshould be common in wild plants, and the magnitude ofpollen limitation may be greater when plants are moreattractive to pollinators. The Wrst prediction accords withrecent meta-analyses of pollen limitation. Our experimentalresults are congruent with both predictions because themagnitude of pollen limitation in P. roseus was greaterwhen plants paid nectar replenishment costs (i.e., moreattractive to pollinators). The same predictions would applyfor plants with constant nectar production rates, becauseplant access to environmental resources could inXuence notonly nectar and seed production, but also other Xoral traitsthat are important for pollinator attraction.
The fertilization and resource availability functions rep-resented in the Haig-Westoby graphical model are Wxed inboth ecological and evolutionary time, and there is no

684 Oecologia (2009) 160:675–685
123
indication of how the mechanistic representation of thesefunctions might evolve. Ashman et al. (2004) extendedtheir conceptual framework incorporating costly reproduc-tive strategies that enhance female fertility. Their formal-ized model illustrates how life history aspects may shapethese functions, and shows how pollen limitation mayresult in the evolution of these functions. If P. roseus Xow-ers are chronically pollen limited, nectar replenishmentcould evolve in a population as a reproductive strategy toincrease pollinator visitation and pollen deposition ratesuntil an equilibrium is reached at which female Wtness islimited simultaneously by both pollen and resource acquisi-tion. Our comparison between pristine and disturbed habi-tats (i.e., diVerent mean resource availability) might help usto understand the causes and consequences of pollen limita-tion for P. roseus. Plant populations with low resourceavailability and low pollinator traYc should experiencestronger trade-oVs between allocation to nectar productionand post-pollination reproductive eVort than those withmore favorable resource conditions. Surprisingly, Burd’s(2008) model predicts that pollen limitation might increaseas plants become more attractive to pollinators in favorableresource conditions. Our population data might support thisprediction. In the most pristine population, La Malinche,Xowers are more frequently visited and have a higher prob-ability to be visited by hummingbirds, and have a more spe-cies-rich pool of hummingbirds compared to Acuitlapilco,the most disturbed population (Lara and Ornelas 2008).Accordingly, rates of removal and deposition of pollen, andnectar replenishment rates, were about twofold in plants atLa Malinche (Lara and Ornelas 2008). When Xowers werepollen supplemented, pollen limitation of seed set increasedin plants at La Malinche compared to those at Acuitlapilco,suggesting higher pollen limitation in the population withmore favorable resource conditions.
Theoretical models consider whole-plant ovule fertiliza-tion and seed set as the appropriate level at which to con-sider maternal Wtness. When using supplemental pollinationas a measure of the pollination environment, the immediateincrease in fruit or seed production is potentially con-founded with plastic responses to additional pollen becauseplants can reallocate resources among Xowers (reviewed inKnight et al. 2006). To solve the problem of resource real-location, researchers typically apply hand-pollinations tothe whole-plant level assuming that all Xowers in a planthave equal access to nutrients and are connected to oneanother. However, the modular nature of Xowering repro-duction should interact with selective factors that cause sto-chastic variation around pollen receipt and resourceavailability. That is, nutrients are not a Wxed pool availableto all Xowers, both spatial and temporal variation inresource availability occur, which means that diVerentXowers have diVerent amounts of energy available for fruit
production (reviewed in Wesselingh 2007). Also, pollendeposition is variable from one Xower to another (Stephenson1981; Burd 1995). This will cause pollen limitation of seedproduction in some Xowers, while in other Xowers it is lim-ited by resource availability (Wesselingh 2007). Ignoringthese inherent diVerences between Xowers can lead to spu-rious results in the study of pollen limitation. Given within-plant resource allocation, we applied nectar-removal andpollination treatments to the level of a Xower includingunbagged open-pollinated controls applied to Xowers onadditional non-treated plants (Zimmerman 1988). By doingso, we assumed that not all Xowers are equal, there beingvariation in natural and experimental pollen deposition andresource availability depending on environmental condi-tions (e.g., amount of nectar replenished) and phenologicalstage, and that resources (e.g., carbon based assimilates), donot freely move around all Xowers, but are restricted inspace and time in integrated physiological units (see alsoCasper and Niesenbaum 1993).
In conclusion, our study showed that frequency of nectarremoval and pollen addition had an interactive eVect ontotal seeds produced. The demonstration that nectarremoval (hence production costs) and pollen receipt jointlyaVect seed set in P. roseus will help to stimulate more com-parative study of both factors in other plant species, partic-ularly in other species in a genus that has become a modelsystem for pollination ecologists. Thus, resource allocationtowards nectar production, together with several other fac-tors (e.g., belowground resources), cannot be overlookedand should more often be considered in future pollen limi-tation studies.
Acknowledgments We are grateful to R. E. Irwin, P. Wilson, J.Walters, A. Hernández, M. Ordano, and two anonymous reviewers forinsightful comments on previous versions of the manuscript, L. E.Núñez Rosas for help in Weldwork, and staV from the Estación Cientí-Wca La Malinche, UAT-UNAM for logistical support. This work hasbeen partially funded by the Departamento de Biología Evolutiva,Instituto de Ecología, AC (reference 902-12-563). We declare that allexperiments presented in this paper comply with the current laws ofMexico.
References
Ashman TL (1992) Indirect costs of seed production between andwithin seasons in a gynodioecious species. Oecologia 92:266–272
Ashman TL, Knight TM, Steets JA, Amarasekare P, Burd M,Campbell DR, Dudash MR, Johnston MO, Mazer SJ, Mitchell RJ,Morgan MT, Wilson WG (2004) Pollen limitation of plant repro-duction: ecological and evolutionary causes and consequences.Ecology 85:2408–2421
Bolten AB, Feinsinger P, Baker HG, Baker I (1979) On the calculationof sugar concentration in Xower nectar. Oecologia 41:301–304
Burd M (1994) Bateman’s principle and plant reproduction: the role ofpollen limitation in fruit and seed set. Bot Rev 60:83–139

Oecologia (2009) 160:675–685 685
123
Burd M (1995) Ovule packaging in stochastic pollination and fertiliza-tion environments. Evolution 49:100–109
Burd M (2008) The Haig-Westoby model revisited. Am Nat 171:400–404
Búrquez A, Corbet SA (1991) Do Xowers reabsorb nectar? Funct Ecol5:369–379
Búrquez A, Sarukhán J, Pedroza AL (1987) Floral biology of a primaryrain forest palm, Astrocaryum mexicanum Liebm. Biol J Linn Soc31:161–174
Calderón G, Rzedowski J (2001) Flora fanerogámica del Valle deMéxico, 2nd edn. Instituto de Ecología, AC, Xalapa
Campbell DR (1985) Pollinator sharing and seed set of Stellariapubera: competition for pollination. Ecology 66:544–553
Campbell DR, Halama KJ (1993) Resource and pollen limitations tolifetime seed production in a natural plant population. Ecology74:1043–1051
Casper BB, Niesenbaum RA (1993) Pollen versus resource limitationof seed production: a reconsideration. Curr Sci 65:210–214
Castellanos MC, Wilson P, Thomson JD (2002) Dynamic nectarreplenishment in Xowers of Penstemon (Scrophulariaceae). Am JBot 89:111–118
Ehrlén J (1992) Proximate limits to seed production in a herbaceousperennial legume, Lathyrus vernus. Ecology 73:1820–1831
Galen C (1985) Regulation of seed-set in Polemonium viscosum: Xoralscents, pollination, and resources. Ecology 66:792–797
Golubov J, Mandujano MC, Montaña C, López-Portillo J, Eguiarte LE(2004) The demographic costs of nectar production in the desertperennial Prosopis glandulosa (Mimosoideae): a modularapproach. Plant Ecol 170:267–275
Gori DF (1983) Post-pollination phenomena and adaptive Xoral chang-es. In: Jones CE, Little RJ (eds) Handbook of experimental polli-nation biology. Van Nostrand Reinhold, New York, pp 31–45
Haig D, Westoby M (1988) On limits of seed production. Am Nat131:757–759
Harder ID, Barrett SCH (1992) The energy cost of bee pollination forPontederia cordata (Pontederiaceae). Funct Ecol 6:226–233
Herre EA (1990) Coevolution of reproductive characteristics in 12 spe-cies of New World Wgs and their pollinator wasps. Experientia45:637–647
Johnston MO (1991) Pollen limitation of female reproduction in Lobe-lia cardinalis and L. siphilitica. Ecology 72:1500–1503
Knight TM, Steets JA, Varmosi JC, Mazer SJ, Burd M, Campbell DR,Dudash MR, Johnston MO, Mitchell RJ, Ashman TL (2005) Pol-len limitation of plant reproduction: patterns and process. AnnuRev Ecol Evol Syst 36:467–497
Knight TM, Steets JA, Ashman TL (2006) A quantitative synthesis ofpollen supplementation experiments highlights the contributionof resource reallocation to estimates of pollen limitation. AmJ Bot 93:271–277
Lara C (2006) Temporal dynamics of Xower use by hummingbirds ina highland temperate forest in Mexico. Ecoscience 13:23–29
Lara C, Ornelas JF (2008) Pollination ecology of Penstemon roseus(Plantaginaceae), an endemic perennial shifted toward humming-bird specialization? Plant Syst Evol 271:223–237
Larson BMH, Barrett SCH (2000) A comparative analysis of pollenlimitation in Xowering plants. Biol J Linn Soc 69:503–520
Lawrence WS (1993) Resource and pollen limitation: plant size-dependent reproductive patterns in Physalis longifolia. Am Nat141:296–313
Leiss KA, Vrieling K, Klinkhamer PGL (2004) Heritability of nectarproduction in Echium vulgare. Heredity 92:446–451
López-Domínguez J, Acosta R (2005) Descripción del Parque Nacion-al La Malinche. In: Fernández J, López-Domínguez J (eds) Bio-diversidad del Parque Nacional La Malinche, Tlaxcala.Coordinación General de Ecología del Gobierno del Estado deTlaxcala, México, pp 73–100
Mattila E, Kuitunen MT (2000) Nutrient versus pollination limitationin Platanthera bifolia and Dactylorhyza incarnata (Orchidaceae).Oikos 89:360–366
McCall C, Primack RB (1987) Resources limit the fecundity of threewoodland herbs. Oecologia 71:431–435
Mitchell RJ, Waser NM (1992) Adaptive signiWcance of Ipomopsisaggregata nectar production: pollination success of single Xow-ers. Ecology 73:633–638
Nepi M, Stpiczyjska M (2007) Nectar resorption and translocation inCucurbita pepo L. and Platanthera chlorantha Custer (Rchb.).Plant Biol 9:93–100
Ordano M, Ornelas JF (2004) Generous-like Xowers: nectar produc-tion in two epiphytic bromeliads and a meta-analysis of removaleVects. Oecologia 140:495–505
Ordano M, Ornelas JF (2005) The cost of nectar replenishment in twoepiphytic bromeliads. J Trop Ecol 21:541–547
Ornelas JF, Ordano M, Lara C (2007) Nectar removal eVects on seedproduction in Moussonia deppeana (Gesneriaceae), a humming-bird-pollinated shrub. Ecoscience 14:117–123
Pleasants JM, Chaplin SJ (1983) Nectar production rates of Asclepiasquadrifolia: causes and consequences of individual variation.Oecologia 59:232–238
Pyke GH (1991) What does it cost a plant to produce Xoral nectar?Nature 350:59
Ritter W, Jáuregui E, Gúzman S, Estrada A, Muñoz H, Súarez J,Corona MC (2004) Ecological and agricultural productivityindices and their dynamics in a sub-humid/semi-arid region fromcentral Mexico. J Arid Environ 59:753–769
Southwick EE (1984) Photosynthate allocation to Xoral nectar: aneglected energy investment. Ecology 65:1775–1779
Stephenson AG (1981) Flower and fruit abortion: proximate causesand ultimate functions. Annu Rev Ecol Syst 12:253–279
Totland Ø (2001) Environment-dependent pollen limitation and selec-tion on Xoral traits in an alpine species. Ecology 82:2233–2244
Wesselingh RA (2007) Pollen limitation meets resource allocation:towards a comprehensive methodology. New Phytol 174:26–37
Young HJ (1988) DiVerential importance of beetle species pollinatingDieVenbachia longispatha (Araceae). Ecology 69:832–844
Young HJ, Young TP (1992) Alternative outcomes of natural andexperimental high pollen loads. Ecology 73:639–647
Zimmerman M (1983) Plant reproduction and optimal foraging: exper-imental nectar manipulation in Delphinium nelsonii. Oikos41:57–63
Zimmerman M (1988) Nectar production, Xowering phenology, andstrategies for pollination. In: Lovett Doust J, Lovett Doust L (eds)Plant reproductive ecology. Oxford University Press, Oxford,pp 157–178
Zimmerman M, Pyke GH (1988a) Reproduction in Polemonium:assessing the factors limiting seed set. Am Nat 131:723–738
Zimmerman M, Pyke GH (1988b) Experimental manipulations ofPolemonium foliosissimum: eVects on subsequent nectar produc-tion, seed production and growth. J Ecol 76:777–789
Related Documents











