Necrosis, sarcolemmal damage and apoptotic events in myofibers rejected by CD8+ lymphocytes: Observations in nonhuman primates Daniel Skuk ⇑ , Jacques P. Tremblay Neurosciences Division – Human Genetics, CHUQ Research Center – CHUL, Quebec, QC, Canada Received 28 December 2011; received in revised form 29 March 2012; accepted 14 May 2012 Abstract To detect the mechanisms of death in allogeneic myofibers rejected by the immune system, myoblasts were allotransplanted in muscles of macaques immunosuppressed with tacrolimus. Immunosuppression was stopped 1 month later to induce a massive rejection of allogeneic myofibers. Grafted sites were biopsied at 2-week intervals and analyzed by histology. The loss of allogeneic myofibers was rapid and concomitant with an intense infiltration of CD8+ lymphocytes. Several necrotic myofibers were observed in the lymphocyte accumulations by intracellular complement immunodetection. Dystrophin and spectrin immunodetection showed sarcolemmal damage in myofibers surrounded and invaded by CD8+ lymphocytes. Active caspase-3 was immunodetected in some myofibers surrounded by CD8+ lymphocytes. This is the first evidence that the collapse of myofibers attacked by T lymphocytes occurs by necrosis possibly due to damage of the sarcolemma. Caspase 3 is activated at least in some myofibers, but there was no evidence of a complete classical process of apoptosis. Ó 2012 Elsevier B.V. All rights reserved. Keywords: Acute rejection; Apoptosis; CD8+ lymphocytes; Cell transplantation; Myofibers; Necrosis; Nonhuman primates 1. Introduction Recent trials of myoblast allotransplantation conducted by our research team in patients with Duchenne muscular dystrophy [1–3] obtained better results in terms of engraft- ment than in previous clinical trials (see reference [4] for a table compiling these results). However, when we had to diagnose whether or not there was an ongoing acute rejec- tion, we faced a lack of information on acute rejection in this context. We found only some studies in mice [5–7], from which was not possible to extrapolate clear guidelines for the diagnosis of rejection in clinical situations. We have been using nonhuman primates from the genus Macaca to analyze the transplantation of myogenic cells in a model more relevant to humans [8,9]. The close phyloge- netic relationship between humans and macaques (both are primates of the parvorder Catarrhini) provides optimal immunological similarities to preclinical transplantation research [10–12]. Unlike mice, macaques share with humans important transplantation immune parameters, including conserved homology in the genes that encode the major histocompatibility complex and the T-cell recep- tor [13–16]. Importantly, rejection of organ allografts in macaques is driven by the same immune elements as in humans, and shows comparable histological features and cadence [12]. We have recently defined the main histologi- cal characteristics of acute rejection of allogeneic myofibers in macaques [17]. This acute rejection is driven essentially by T lymphocytes that focally accumulate to surround and invade myofibers. However, the mechanism of 0960-8966/$ - see front matter Ó 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.nmd.2012.05.005 ⇑ Corresponding author. Address: Unite ´ de recherche en Ge ´ne ´tique humaine, Centre Hospitalier de l’Universite ´ Laval, 2705 boulevard Laurier, Que ´bec, QC, Canada G1V 4G2. Tel.: +1 418 654 2186; fax: +1 418 654 2207. E-mail address: [email protected] (D. Skuk). www.elsevier.com/locate/nmd Available online at www.sciencedirect.com Neuromuscular Disorders 22 (2012) 997–1005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
www.elsevier.com/locate/nmd
Neuromuscular Disorders 22 (2012) 997–1005
Necrosis, sarcolemmal damage and apoptotic eventsin myofibers rejected by CD8+ lymphocytes: Observations
in nonhuman primates
Daniel Skuk ⇑, Jacques P. Tremblay
Neurosciences Division – Human Genetics, CHUQ Research Center – CHUL, Quebec, QC, Canada
Received 28 December 2011; received in revised form 29 March 2012; accepted 14 May 2012
Abstract
To detect the mechanisms of death in allogeneic myofibers rejected by the immune system, myoblasts were allotransplanted in musclesof macaques immunosuppressed with tacrolimus. Immunosuppression was stopped 1 month later to induce a massive rejection ofallogeneic myofibers. Grafted sites were biopsied at 2-week intervals and analyzed by histology. The loss of allogeneic myofibers wasrapid and concomitant with an intense infiltration of CD8+ lymphocytes. Several necrotic myofibers were observed in the lymphocyteaccumulations by intracellular complement immunodetection. Dystrophin and spectrin immunodetection showed sarcolemmal damagein myofibers surrounded and invaded by CD8+ lymphocytes. Active caspase-3 was immunodetected in some myofibers surrounded byCD8+ lymphocytes. This is the first evidence that the collapse of myofibers attacked by T lymphocytes occurs by necrosis possibly due todamage of the sarcolemma. Caspase 3 is activated at least in some myofibers, but there was no evidence of a complete classical process ofapoptosis.� 2012 Elsevier B.V. All rights reserved.
Keywords: Acute rejection; Apoptosis; CD8+ lymphocytes; Cell transplantation; Myofibers; Necrosis; Nonhuman primates
1. Introduction
Recent trials of myoblast allotransplantation conductedby our research team in patients with Duchenne musculardystrophy [1–3] obtained better results in terms of engraft-ment than in previous clinical trials (see reference [4] for atable compiling these results). However, when we had todiagnose whether or not there was an ongoing acute rejec-tion, we faced a lack of information on acute rejection inthis context. We found only some studies in mice [5–7],from which was not possible to extrapolate clear guidelinesfor the diagnosis of rejection in clinical situations.
0960-8966/$ - see front matter � 2012 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.nmd.2012.05.005
⇑ Corresponding author. Address: Unite de recherche en Genetiquehumaine, Centre Hospitalier de l’Universite Laval, 2705 boulevardLaurier, Quebec, QC, Canada G1V 4G2. Tel.: +1 418 654 2186; fax:+1 418 654 2207.
E-mail address: [email protected] (D. Skuk).
We have been using nonhuman primates from the genusMacaca to analyze the transplantation of myogenic cells ina model more relevant to humans [8,9]. The close phyloge-netic relationship between humans and macaques (both areprimates of the parvorder Catarrhini) provides optimalimmunological similarities to preclinical transplantationresearch [10–12]. Unlike mice, macaques share withhumans important transplantation immune parameters,including conserved homology in the genes that encodethe major histocompatibility complex and the T-cell recep-tor [13–16]. Importantly, rejection of organ allografts inmacaques is driven by the same immune elements as inhumans, and shows comparable histological features andcadence [12]. We have recently defined the main histologi-cal characteristics of acute rejection of allogeneic myofibersin macaques [17]. This acute rejection is driven essentiallyby T lymphocytes that focally accumulate to surroundand invade myofibers. However, the mechanism of

998 D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005
myofiber death by the immune cells is yet to be determined.Although apoptosis is considered the main mechanism ofT-lymphocyte cytotoxicity in mononuclear cells [18–20],we previously failed to detect myofiber apoptosis inmacaques in which immunosuppression was not optimalto control acute rejection of allogeneic myofibers. Neithermyofiber necrosis or apoptosis was observed in those cases.
Trying to delve into this subject, the present study wascarried out with the hypothesis that the detection ofnecrosis or apoptosis was elusive because the collapse ofthe rejected myofibers is very rapid once started, so macro-phages eliminate dead myofibers rapidly. Therefore, toincrease the chances of detecting the mechanism of myofi-ber death in acute rejection, we wanted to produce a mas-sive and rapid attack of T lymphocytes against myofibersexpressing allogeneic epitopes, trying to capture themoment of this attack in a muscle sample.
2. Material and methods
2.1. Animals
Four male cynomolgus monkeys (Macaca fascicularis),4–5 years old, were used as recipients of myoblast allo-transplantation. For transplantation and biopsies, theywere kept under general anesthesia with isofluorane (1.5–2% in oxygen) after induction with intramuscular ketamine(10 mg/kg) and glycopyrrolate (0.05 mg/kg). Buprenor-phine (0.01 mg/kg b.i.d. for 3 days) was given as postoper-ative analgesia. Because of the many muscle samples taken,euthanasia was performed at the end of each experiment byintravenous administration of an overdose of pentobarbital(120 mg/kg) after intramuscular ketamine anesthesia(15 mg/kg). The Laval University Animal Care Committeeauthorized these procedures.
2.2. Cell transplantation
The cell cultures were obtained from a skeletal musclebiopsy done previously in another cynomolgus monkey.This biopsy was minced with fine scissors into fragments ofless than 1 mm3 and then dissociated with 0.2% collagenase(Sigma, St-Louis, MO, USA) in Hank’s balanced saline solu-tion (HBSS) for 1 h, followed by another dissociation with0.125% trypsin (Gibco, Grand Island, NY, USA) in HBSSfor 45 min. The isolated cells were subcultured in vitro inMCDB-120 culture medium [21], with 15% fetal bovineserum (FBS) (Hyclone, Logan, UT, USA), 10 ng/ml of basicfibroblast growth factor (Feldan, St-Laurent, QC, Canada),0.5 mg/ml of bovine serum albumin (Sigma), 0.39 lg/ml ofdexamethasone (Sigma) and 5 lg/ml of human insulin(Sigma). The cells were infected 2 times in vitro with a repli-cation-defective retroviral vector LNPOZC7 (gift from Dr.Constance Cepko, Harvard University, Boston, MA,USA) encoding a LacZ reporter gene and a neomycin-resis-tance gene. The transduced cells were selected twice with600 lg/ml Geneticin (Invitrogen, Burlington, ON, Canada)
within 4 days, proliferated until confluence, and frozen forstorage in liquid nitrogen. A sample of cells to be frozenwas analyzed to determine the percentage of CD56+ cellsas an indicator of the percentage of myoblasts in the cultures.They were incubated with a phycoerythrin-coupled anti-CD56 antibody (Beckman Coulter, Fullerton, CA, USA)and analyzed by flow cytometry. Ninety nine percent of cellswere CD56+. Additional details on the cell culture protocolhave been published previously [8].
For transplantation, cells were thawed, proliferated dur-ing 1 or 2 passages in culture, detached from the flasks using0.1% trypsin in HBSS and washed 3 times with HBSS. Cellswere resuspended in HBSS for transplantation. Intramuscu-lar cell transplantation was performed percutaneously byparallel equidistant injections (1 mm between each other)perpendicular to the muscle surface and forming a squareof approximately 1 cm2. Cell injections were performed witha 250 ll syringe (Hamilton, Reno, NV, USA) attached to aPB600-1 repeating dispenser (Hamilton) and using 27-gaugeneedles of 0.5 inches [22]. The amounts of cells delivered percm3 of muscle are shown in Table 1 (these amounts variedwith the total cells produced at the time of transplantation).The cell implantation involved the penetration of the needleto the maximum depth and the delivery of 5 ll of cell sus-pension during the withdrawal of the needle. An OpSitetransparent sterile dressing with a 5-mm grid (Smith &Nephew, Hull, United Kingdom) was adhered to the skinto see the pattern of needle penetrations. To identify theinjected muscle sites at the time of biopsy, two stitches ofinert non-absorbable polypropylene 4.0 suture (Prolene,Ethicon Inc, Somerville, NJ, USA) were placed at �5 mmfrom both sides of the sites. The muscles used in the exper-iment were the left and right biceps brachii, quadratus femo-ris and gastrocnemius. Up to 2 sites of 1 cm2 were graftedper biceps brachii and gastrocnemius, and up to 3 sites of1 cm2 per quadratus femoris. These sites were separatedby at least 1 cm. Additional details on the cell transplanta-tion protocol have been published previously [8].
2.3. Immunosuppression
An intramuscular formulation of tacrolimus (a generousgift from Astellas Pharma Inc., Osaka, Japan) was admin-istered once daily for immunosuppression, beginning 5–7 days before transplantation. Blood samples were takenat different intervals and analyzed on an IMx tacrolimusII kit for micro-particle enzyme immunoassay (Abbott,Wiesbaden, Germany). The dose of immunosuppressantwas adjusted to obtain optimal tacrolimus blood levelsfor myoblast transplantation in monkeys (>50 lg/L) dur-ing the first month post-transplantation, and then thetacrolimus administration was stopped.
2.4. Sampling
Biopsies were performed in one of the transplanted sitesin each monkey 1 month after transplantation and then

Table 1Percentages of b-Gal+ myofibers in the 7 muscle biopsies performed per monkey in the myoblast-grafted sites, at 2-weeks intervals after tacrolimuswithdrawal.
Monkey Cells grafted per cm3 Weeks after tacrolimus withdrawal
0 2 4 6 8 10 12
Percentage of b-Gal+ myofibers in the biopsies (%)
1 28 � 106 27.8 28.2 33 32.6 5.3 0 02 23.8 � 106 27.2 20.5 21.1 6.2 0 0 03 23.5 � 106 10.7 14.6 11.8 3.8 0 0 04 18.1 � 106 35 31.3 31.8 19.1 0 0 0
D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005 999
every 2 weeks in one of the other transplanted sites (rotat-ing muscles) until resolution of rejection. Preliminaryexperiments gave an idea of the optimal frequency ofbiopsies. Muscle biopsies were performed by open surgery,sampling approximately 1 g of muscle between the twostitches of polypropylene suture placed on either side ofthe cell-grafted site. Biopsies were mounted in embeddingmedium and snap frozen in liquid nitrogen. Serial sectionsof 10–15 lm were made in a cryostat.
2.5. Histological techniques
Sections were stained routinely with hematoxylin-eosin(H&E). For histochemical detection of b-galactosidase(b-Gal), the muscle sections were fixed 3 min in 0.25%glutaraldehyde, rinsed with phosphate buffered saline(PBS), incubated 24 h at room temperature in a solutionof 0.4 mg/ml X-gal (5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside) (Boehringer Mannheim, Vienna, Aus-tria) containing 1 mM MgCl2, 3 mM Ke3F(CN)6�3 mMKe4F(CN)6�3H20 in PBS, and mounted in 1:1 glycerin-jelly.
For immunohistology, the following antibodies (Ab)were used: mouse anti-human CD8 monoclonal Ab (BDBiosciences, Mississauga, ON, Canada), rabbit anti-human/sheep CD8 polyclonal Ab (Abcam, Cambridge,MA, USA), mouse anti-human CD163 monoclonal Ab(BD Biosciences), mouse anti-human/baboon C5b-9monoclonal Ab (Abcam), rabbit anti-human/mouse cas-pase 3 active polyclonal Ab (R&D Systems, Minneapolis,MN, USA), mouse anti-human dystrophin (C-terminus)monoclonal Ab (Vector Laboratories, Burlingame, CA,USA) and mouse anti-human spectrin monoclonal Ab(NCL-SPEC-1, Leica Biosystems Newcastle Ltd, Newcas-tle upon Tyne, United Kingdom).
For immunohistology, nonspecific binding was blockedby a 30-min incubation with 10% FBS in PBS. Sectionswere incubated 1 h with the primary Ab at the concentra-tions recommended by the manufacturers. Then followeda 30-min incubation with a biotinylated anti-mouse Ab(1/150, Dako, Copenhagen, Denmark) or a biotinylatedanti-rabbit Ab (1/150, Dako) followed in both cases by a30-min incubation with streptavidin-Cy3 (1/700, Sigma).For double immunostaining, an anti-mouse IgG Abconjugated to Alexa Fluor 488 (1:300; Molecular Probes,Eugene, OR, USA) was also used as second Ab. Antibodies
and streptavidin were diluted in PBS, pH 7.4, containing1% FBS. Incubations were performed at room tempera-ture. Sections of tissues from macaques were used as posi-tive controls and to verify the reactivity of the Ab withcynomolgus monkeys.
2.6. Microscopic analysis
The muscle sections were analyzed using an Axiophotmicroscope with epifluorescence and bright field optics(Zeiss, Oberkochen, Germany). Pictures were taken withan A650 IS digital camera (Canon, Tokyo, Japan). Selectedmuscle sections with fluorescent immunostaining were ana-lyzed using an Olympus IX70 microscope with epifluores-cent optics (Olympus, Tokyo, Japan) with a FluoView300 confocal microscopy scanning unit (Olympus) usinglasers of 488 nm and 543 nm. Confocal microscopy imageswere taken with FluoView 300 software (Olympus).
3. Results
Tacrolimus blood levels gradually decreased afterimmunosuppression withdrawal, and were not detectedafter 5–9 weeks. Biopsies taken 1 month after transplanta-tion showed abundant b-Gal+ myofibers (Table 1). Abun-dant b-Gal+ myofibers were also observed up to 4 or6 weeks after tacrolimus withdrawal (Table 1). An exampleof these biopsies is shown in Fig. 1A. A drastic decrease inthe amount of b-Gal+ myofibers was observed 6 or8 weeks after tacrolimus withdrawal, preceding the absenceof b-Gal+ myofibers in consecutive biopsies (Table 1,Fig. 1B and C). Therefore, the graft was lost in less than4 weeks (time between the last biopsy with large amountsof b-Gal+ myofibers and the first biopsy with no b-Gal+myofibers). This occurred when tacrolimus blood levels fellbelow 10 ng/ml.
Simultaneously with the abrupt loss of Gal+ myofibers,there were dense focal accumulations of lymphocytes inH&E stained sections (Fig. 1E). Lymphocyte accumula-tions were absent from biopsies taken 1 month after trans-plantation (at the time of tacrolimus withdrawal) and inbiopsies performed 2 and 4 weeks later (Fig. 1D), duringwhich tacrolimus blood levels decreased but were stillabove 10 ng/ml. This prominent lymphocyte infiltrationwas observed 6 or 8 weeks after tacrolimus withdrawal
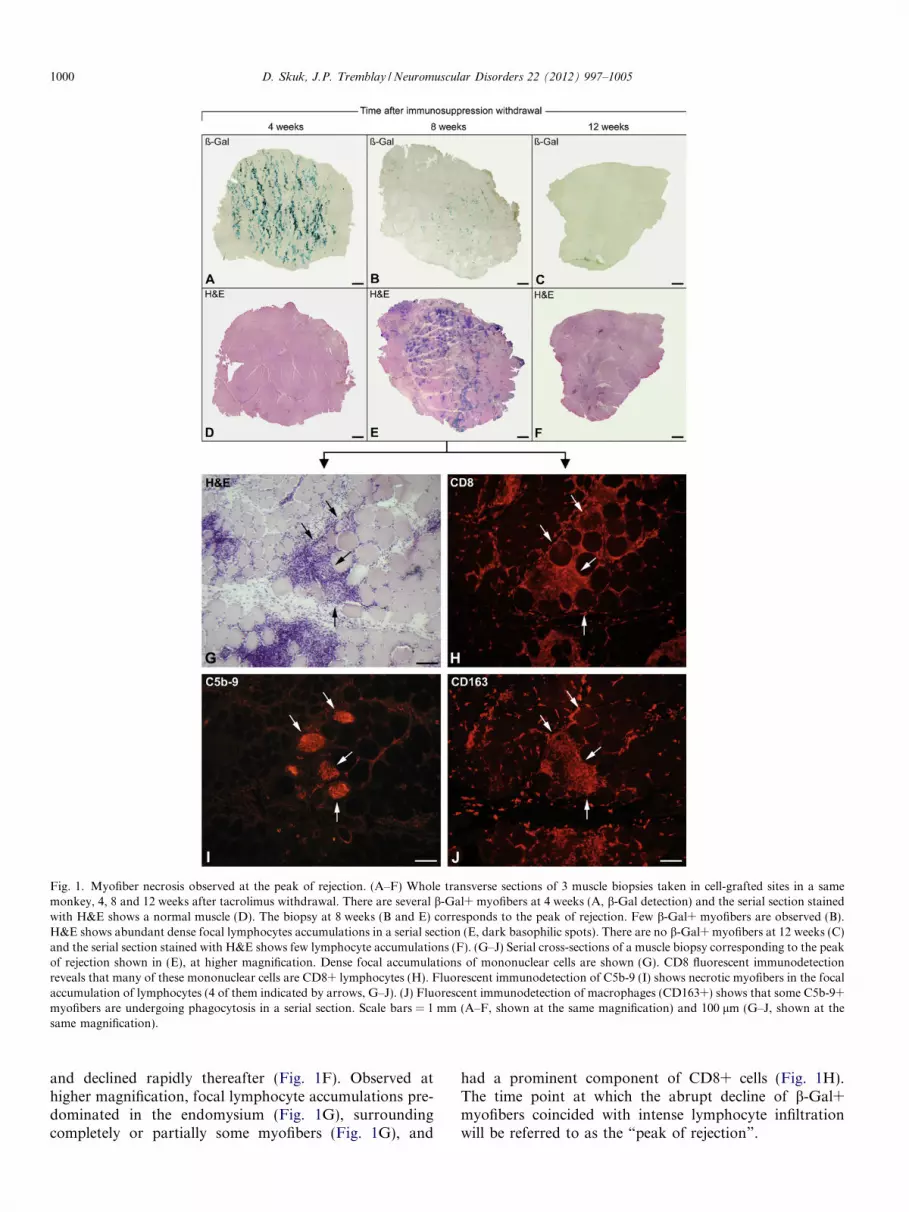
Fig. 1. Myofiber necrosis observed at the peak of rejection. (A–F) Whole transverse sections of 3 muscle biopsies taken in cell-grafted sites in a samemonkey, 4, 8 and 12 weeks after tacrolimus withdrawal. There are several b-Gal+ myofibers at 4 weeks (A, b-Gal detection) and the serial section stainedwith H&E shows a normal muscle (D). The biopsy at 8 weeks (B and E) corresponds to the peak of rejection. Few b-Gal+ myofibers are observed (B).H&E shows abundant dense focal lymphocytes accumulations in a serial section (E, dark basophilic spots). There are no b-Gal+ myofibers at 12 weeks (C)and the serial section stained with H&E shows few lymphocyte accumulations (F). (G–J) Serial cross-sections of a muscle biopsy corresponding to the peakof rejection shown in (E), at higher magnification. Dense focal accumulations of mononuclear cells are shown (G). CD8 fluorescent immunodetectionreveals that many of these mononuclear cells are CD8+ lymphocytes (H). Fluorescent immunodetection of C5b-9 (I) shows necrotic myofibers in the focalaccumulation of lymphocytes (4 of them indicated by arrows, G–J). (J) Fluorescent immunodetection of macrophages (CD163+) shows that some C5b-9+myofibers are undergoing phagocytosis in a serial section. Scale bars = 1 mm (A–F, shown at the same magnification) and 100 lm (G–J, shown at thesame magnification).
1000 D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005
and declined rapidly thereafter (Fig. 1F). Observed athigher magnification, focal lymphocyte accumulations pre-dominated in the endomysium (Fig. 1G), surroundingcompletely or partially some myofibers (Fig. 1G), and
had a prominent component of CD8+ cells (Fig. 1H).The time point at which the abrupt decline of b-Gal+myofibers coincided with intense lymphocyte infiltrationwill be referred to as the “peak of rejection”.

Fig. 2. C5b-9 immunodetection in a transverse section of a muscle biopsy taken in a myoblast-grafted site at the peak of rejection, using confocalmicroscopy to show the intracellular localization of the complement membrane attack complex. Double labeling of C5b-9 (A and C, green fluorescence)and CD8+ lymphocytes (B and C, red fluorescence) was carried out. The arrow indicates a myofiber with intracellular deposition of complementmembrane attack complex, surrounded by CD8+ lymphocytes. Scale bars = 50 lm.
D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005 1001
Several myofibers with sarcoplasmic C5b-9 labeling (asign of necrosis) were observed in muscle biopsies corre-sponding to the peak of rejection (Fig. 1I, Fig. 2). Myofiberswith intracellular C5b-9 labeling were not observed in mus-cle biopsies taken before this peak, and were rare in biopsiestaken after this peak, even if there were some lymphocyteaccumulations after the peak of rejection. Myofibers withsarcoplasmic C5b-9 labeling were surrounded totally orpartially, and sometimes invaded, by CD8+ lymphocytes(Fig. 1H and I, Fig. 2). Most of them were also invadedby macrophages (CD163+ cells) in serial sections, evidenceof phagocytosis of necrotic debris (Fig. 1J).
In several myofibers invaded by CD8+ lymphocytes, theimmunodetection of dystrophin and spectrin showed dele-tions of these sarcolemma associated proteins at sites ofinvasion (Fig. 3). No intracellular C5b-9 was observed inmost myofibers with focal sarcolemmal deletions. In C5b-9+ myofibers, dystrophin and spectrin were either detectedweakly and with large deletions or were not detected, whichshowed a total or near total collapse of these sarcolemma-associated proteins in necrotic myofibers.
Active caspase 3 was detected in the sarcoplasm of somemyofibers. These myofibers were surrounded and eveninvaded by CD8+ lymphocytes (Fig. 4A–C). Most myofi-bers surrounded and even invaded by CD8+ lymphocytes,however, were not positive for active caspase-3. Myofiberspositive for active caspase-3 were associated with signifi-cant sarcolemmal damage, as evidenced by dystrophinimmunodetection in serial sections (Fig. 4D–F). Myofibersexpressing active caspase 3 were not fragmented into apop-totic bodies.
Muscle biopsies taken 2 weeks after the peak of rejectionbiopsy showed no b-Gal+ myofibers and had a lower infil-tration of lymphocytes. Conversely, they showed abundantmyofibers with classical histological features of recent orongoing regeneration: small diameter, sarcoplasmic baso-philia and central nuclei (Fig. 5).
4. Discussion
The present study sought to reveal the mechanisms ofdeath of the allogeneic myofibers attacked by the immunesystem during acute rejection by concentrating in a shortperiod of time an intense attack of T lymphocytes againstmyofibers. To achieve this, intramuscular engraftment ofallogeneic myoblasts was obtained under optimal tacroli-mus immunosuppression and tacrolimus was withdrawnafter a month. Tacrolimus blood levels then decreasedgradually until they were unable to control acute rejection.Once triggered, the rejection of the allogeneic myofiberswas massive and rapid as expected, allowing detection ina muscle sample (the biopsy corresponding to the peak ofrejection), which probably escaped in a more insidiousrejection: myofibers attacked by CD8+ lymphocytesshowed apoptotic events and necrosis. Furthermore, itwas possible to observe in another muscle sample (thebiopsy after the peak of rejection), massive myofiber regen-eration, reflecting the recent massive myofiber destruction.
That the lymphocyte attack and the myofiber reactionswere caused by acute rejection and were not related to mus-cle damage produced by the transplantation procedure isvalidated by the fact that muscle histology was normal1 month after transplantation. Both the immune cell attackand the myofiber reactions were observed only at the peakof rejection, associated with tacrolimus blood levels fallingbelow 10 ng/ml.
4.1. Myofiber necrosis
Necrosis occurs when a cell undergoes irreversible dam-age resulting in metabolic impairments that lead to death.The early loss of membrane integrity (observed in the pres-ent study in myofibers attacked by CD8+ lymphocytes) is aclassical feature of necrosis. By contrast, membrane integ-rity is preserved in apoptosis. The loss of membrane

Fig. 3. Damage of the sarcolemma in myofibers attacked by T lymphocytes. The images correspond to fluorescent co-immunodetection of sarcolemmal-associated proteins (red) and CD8 (green) in transverse sections of muscle biopsies taken at myoblast-grafted sites at the peak of rejection, analyzed byconfocal microscopy. (A) An asterisk indicates a myofiber with a large deletion of dystrophin (arrowheads), corresponding to a region hit by CD8+lymphocytes (B and C, arrowheads). (D–F) Two myofibers with different images of CD8+ lymphocyte invasion are shown. In myofiber 1, CD8+lymphocytes penetrate the myofiber through a region where the absence of dystrophin evidences membrane damage (arrowhead). Myofiber 2 exhibits a“tunneling” pattern of lymphocyte invasion (arrow). Since lymphocytes penetrate from a site out of the section, the dystrophin + sarcolemma of thismyofiber is quite preserved in the image. (G) Asterisks indicate 2 myofibers with different degrees of deletion of the spectrin-labeled sarcolemma. Thesemyofibers have different degrees of invasion by CD8+ lymphocytes at the sites of spectrin deletion (I, arrowheads). Scale bars = 50 lm.
1002 D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005
integrity leads to the intracellular influx of extracellularmolecules and ions, including complement proteins. Com-plement is activated after penetration into myofibers, prob-ably by exposure to intracellular organelles, alteredbiological membranes or proteolysis [23]. Membraneattack complex is assembled as a final step of complementactivation, binding to surfaces within the myofiber [23].The quaternary configuration of membrane attack complex
has neoantigens that are not present in precursor mole-cules, such as C5b-9. Therefore, sarcoplasmic C5b-9 immu-nodetection (as done in the present study) allows easyvisualization of myofiber necrosis [23].
In the present study there was evidence of a direct linkbetween myofiber necrosis (i.e., myofibers with intracellu-lar C5b-9 deposition) and CD8+ lymphocyte attack. Thislink was chronological (necrotic myofibers were
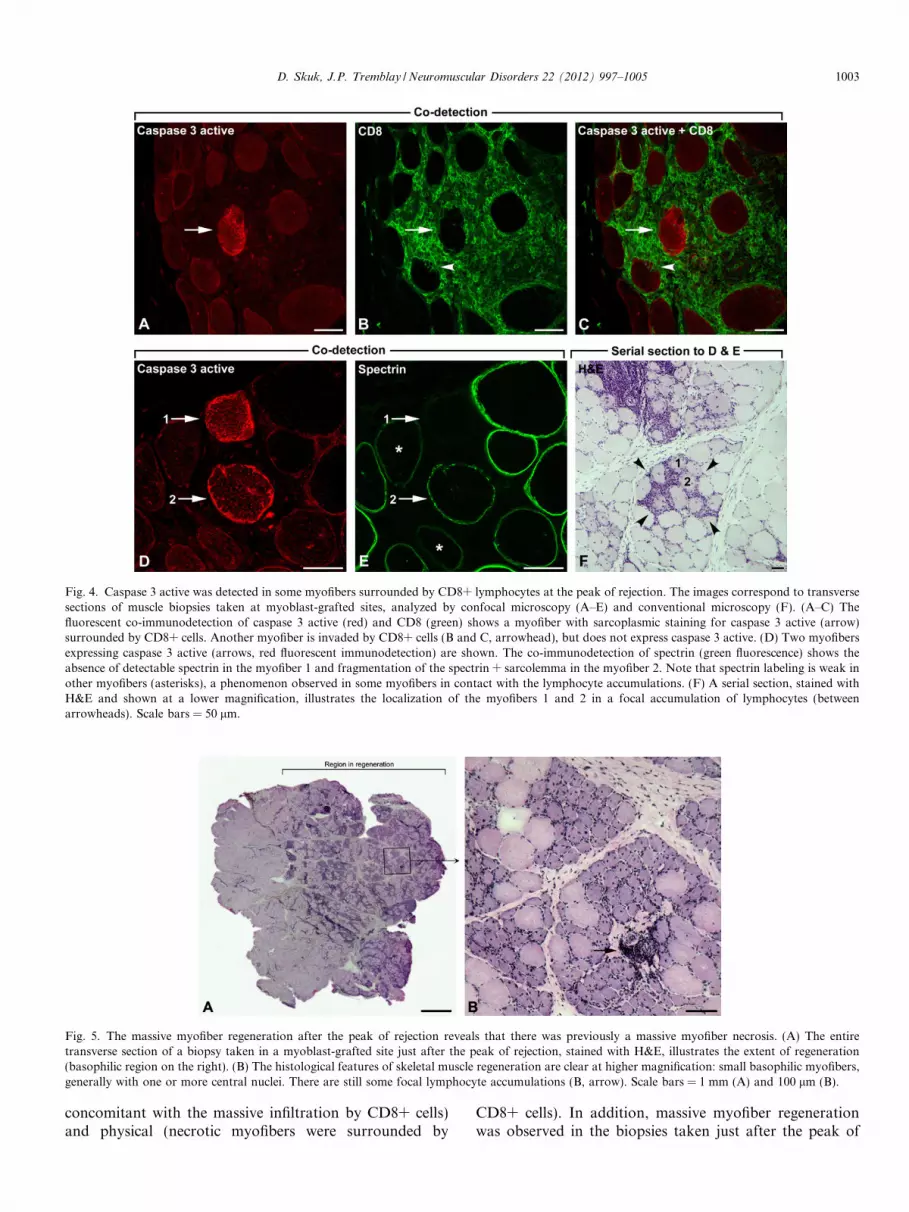
Fig. 4. Caspase 3 active was detected in some myofibers surrounded by CD8+ lymphocytes at the peak of rejection. The images correspond to transversesections of muscle biopsies taken at myoblast-grafted sites, analyzed by confocal microscopy (A–E) and conventional microscopy (F). (A–C) Thefluorescent co-immunodetection of caspase 3 active (red) and CD8 (green) shows a myofiber with sarcoplasmic staining for caspase 3 active (arrow)surrounded by CD8+ cells. Another myofiber is invaded by CD8+ cells (B and C, arrowhead), but does not express caspase 3 active. (D) Two myofibersexpressing caspase 3 active (arrows, red fluorescent immunodetection) are shown. The co-immunodetection of spectrin (green fluorescence) shows theabsence of detectable spectrin in the myofiber 1 and fragmentation of the spectrin + sarcolemma in the myofiber 2. Note that spectrin labeling is weak inother myofibers (asterisks), a phenomenon observed in some myofibers in contact with the lymphocyte accumulations. (F) A serial section, stained withH&E and shown at a lower magnification, illustrates the localization of the myofibers 1 and 2 in a focal accumulation of lymphocytes (betweenarrowheads). Scale bars = 50 lm.
Fig. 5. The massive myofiber regeneration after the peak of rejection reveals that there was previously a massive myofiber necrosis. (A) The entiretransverse section of a biopsy taken in a myoblast-grafted site just after the peak of rejection, stained with H&E, illustrates the extent of regeneration(basophilic region on the right). (B) The histological features of skeletal muscle regeneration are clear at higher magnification: small basophilic myofibers,generally with one or more central nuclei. There are still some focal lymphocyte accumulations (B, arrow). Scale bars = 1 mm (A) and 100 lm (B).
D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005 1003
concomitant with the massive infiltration by CD8+ cells)and physical (necrotic myofibers were surrounded by
CD8+ cells). In addition, massive myofiber regenerationwas observed in the biopsies taken just after the peak of

1004 D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005
rejection. Regenerating myofibers were evident in H&Estained sections by classical histological features: smalldiameter, sarcoplasmic basophilia and central nuclei [24].Since myofiber regeneration reflects recent myofiber necro-sis, this observation supports the fact that a massive necro-sis occurred in the rejected myofibers.
The only available human results to compare with thisobservation are those in autoimmune myopathies inwhich the pathogenesis is also a direct cytotoxicity ofCD8+ lymphocytes against myofibers, i.e., polymyositisand inclusion body myositis (IBM) [25]. As acute rejectionin the present study, the predominant histological featurein polymyositis and IBM is the presence of focal accumu-lations of lymphocytes in the endomysium, which par-tially or completely surround and invade non-necroticmyofibers [26–29]. There are necrotic myofibers in bothdiseases, although the link with the lymphocyte attack isnot so clear because the predominant invading cells inthese myofibers are macrophages, and CD8+ lymphocytesconstitute only 14.5% of the mononuclear cells associatedwith them [30]. Regarding sarcolemmal damage, it isinteresting to remark that ultrastructural studies in poly-myositis and sIBM showed that CD8+ lymphocytes sendprocesses that traverse the basal lamina and focally com-press the myofibers, but without causing sarcolemmaldeletions [31]. The present study did not analyze the integ-rity of the sarcolemma by electron microscopy, but thedeletion of dystrophin and spectrin is evidence that thesarcolemma was not spared by the lymphocyte attack inthe present context of acute rejection.
4.2. Apoptotic events
Induction of apoptosis in their targets is considered themain mechanism of CD8+ lymphocyte cytotoxicity inmononuclear cells [18–20]. Consequently, the authorwanted to search apoptotic events in myofibers attackedby CD8+ lymphocytes. Immunodetection of the activeform of caspases is an efficient method to demonstrateapoptosis in tissue sections [32]. Caspases are intracellularproteases that orderly dissembled cells during apoptosisand their activation provides a common biochemical basisfor apoptosis [33]. Active caspase 3 is most commonly usedto detect apoptotic events in myofibers and it was reportedin several myopathies (for a review see reference [34]).
In the present study, active caspase 3 was detected inseveral myofibers surrounded by CD8+ lymphocytes dur-ing the peak of rejection. This would indicate that thesemyofibers received a pro-apoptotic signal from the sur-rounding lymphocytes. In almost all cases, CD8+ lympho-cytes induce apoptosis of their targets by the perforin–granzyme and the ligand-and-Fas pathways [18–20]. Inhuman skeletal muscle, there is evidence that the perfo-rin–granzyme pathway may play a cytotoxic role on myof-ibers in polymyositis and sIBM [35,36], whereas Fas-mediated mechanism may not be involved [37,38]. Thus,activation of caspase 3 during myofiber rejection could be
also produced by a perforin–granzyme attack of CD8+lymphocytes. Further studies are needed to confirm that.
Does the expression of active caspase 3 indicate that themyofibers were also rejected by apoptosis? Probably not, ifone takes into account both the rest of the present observa-tions and the previous studies on apoptosis. In mononu-clear cells, apoptosis is clear when active caspase 3 isdetected during the fragmentation of the cell into apoptoticbodies, while active caspase 3 expression in morphologi-cally preserved cells is considered only a putative pre-apop-totic stage [39]. In the present study, active caspase 3 wasdetected in myofibers with preserved profile and size, thuscorresponding rather to a putative pre-apoptotic stage.Given that there were no myofibers fragmented into apop-totic bodies, we can assume that caspase 3 activation didnot lead to a classical apoptotic process. In addition, it iscontroversial whether and how myofibers develop apopto-sis [34]. Unlike mononuclear cells, large sincytia containinghundreds of nuclei are not a proper ground for the devel-opment of a classical process of apoptosis [38,25]. On theother hand, activation of caspase 3 was frequently associ-ated with significant sarcolemmal damage in the presentstudy. This early loss of cell membrane integrity is a classi-cal feature of necrosis but not of apoptosis. Therefore,since the deletions of the sarcolemma at the sites of lym-phocyte invasion may cause necrosis and that there wasevidence of necrosis as a terminal phase of myofiber death,the author hypothesizes that necrosis ultimately prevails onan aborted process of apoptosis. It is noteworthy that thepossibility of an apoptotic process preceding myofibernecrosis was previously suggested in dystrophic mice [40].Finally, again comparing with polymyositis and sIBM,most studies found no evidence of myofiber apoptosis inthese entities [34].
5. Conclusion
This study shows a chronological and topographicallink between the attack of CD8+ lymphocytes against allo-geneic myofibers and myofiber necrosis, suggesting thatthis could be the ultimate mechanism of cell death in thiscontext. It also shows that the lymphocyte attack producessarcolemmal damage and, to a lesser extent, induces activa-tion of caspase 3 in myofibers (although there was no evi-dence of complete apoptosis). Therefore, the removal ofallogeneic myofibers by acute rejection appears to followa different path than the classical pathway described inthe removal of allogeneic mononuclear cells.
Acknowledgments
The author wishes to express his gratitude to Ms.Marlyne Goulet for excellent technical work in cell culture,immunohistochemistry and follow-up of immunosuppres-sion in monkeys.

D. Skuk, J.P. Tremblay / Neuromuscular Disorders 22 (2012) 997–1005 1005
This work was supported by a grant of the Jesse’s Jour-ney Foundation for Gene and Cell Therapy of Canada toDr. Daniel Skuk.
References
[1] Skuk D, Roy B, Goulet M, et al. Dystrophin expression inmyofibers of Duchenne muscular dystrophy patients followingintramuscular injections of normal myogenic cells. Mol Ther2004;9:475–82.
[2] Skuk D, Goulet M, Roy B, et al. Dystrophin expression inmuscles of Duchenne muscular dystrophy patients after high-density injections of normal myogenic cells. J Neuropathol ExpNeurol 2006;65:371–86.
[3] Skuk D, Goulet M, Roy B, et al. First test of a “high-densityinjection” protocol for myogenic cell transplantation throughoutlarge volumes of muscles in a Duchenne muscular dystrophy patient:eighteen months follow-up. Neuromuscul Disord 2007;17:38–46.
[4] Skuk D. Myoblast transplantation for inherited myopathies: a clinicalapproach. Expert Opin Biol Ther 2004;4:1871–85.
[5] Guerette B, Asselin I, Vilquin JT, Roy R, Tremblay JP. Lymphocyteinfiltration following allo- and xenomyoblast transplantation in mdxmice. Muscle Nerve 1995;18:39–51.
[6] Irintchev A, Zweyer M, Wernig A. Cellular and molecular reactionsin mouse muscles after myoblast implantation. J Neurocytol1995;24:319–31.
[7] Wernig A, Irintchev A. “Bystander” damage of host muscle caused byimplantation of MHC- compatible myogenic cells. J Neurol Sci1995;130:190–6.
[8] Skuk D, Goulet M, Paradis M, Tremblay JP. Myoblast transplan-tation: techniques in nonhuman primates as a bridge to clinical trials.In: Soto-Gutierrez A, Navarro-Alvarez N, Fox IJ, editors. Methodsin bioengineering: cell transplantation. Boston: Artech House; 2011.p. 219–36.
[9] Skuk D, Tremblay JP. Intramuscular cell transplantation as apotential treatment of myopathies: clinical and preclinical relevantdata. Expert Opin Biol Ther 2011;11:359–74.
[10] Rose SM, Blustein N, Rotrosen D. Recommendations of the expertpanel on ethical issues in clinical trials of transplant tolerance.National Institute of Allergy and Infectious Diseases of the NationalInstitutes of Health. Transplantation 1998;66:1123–5.
[11] Kirk AD. Transplantation tolerance: a look at the nonhuman primateliterature in the light of modern tolerance theories. Crit Rev Immunol1999;19:349–88.
[12] Kirk AD. Crossing the bridge: large animal models in translationaltransplantation research. Immunol Rev 2003;196:176–96.
[13] Levinson G, Hughes AL, Letvin NL. Sequence and diversity of rhesusmonkey T-cell receptor beta chain genes. Immunogenetics1992;35:75–88.
[14] Slierendregt BL, van Noort JT, Bakas RM, Otting N, Jonker M,Bontrop RE. Evolutionary stability of transspecies major histocom-patibility complex class II DRB lineages in humans and rhesusmonkeys. Hum Immunol 1992;35:29–39.
[15] Geluk A, Elferink DG, Slierendregt BL, et al. Evolutionary conser-vation of major histocompatibility complex–DR/peptide/T cellinteractions in primates. J Exp Med 1993;177:979–87.
[16] Jaeger EE, Bontrop RE, Lanchbury JS. Structure, diversity, andevolution of the T-cell receptor VB gene repertoire in primates.Immunogenetics 1994;40:184–91.
[17] Skuk D. Acute rejection of myofibers in nonhuman primates: Keyhistopathologic features. J Neuropathol Exp Neurol2012;71:398–412.
[18] Berke G. The CTL’s kiss of death. Cell 1995;81:9–12.[19] Graubert TA, Ley TJ. How do lymphocytes kill tumor cells? Clin
Cancer Res 1996;2:785–9.
[20] Shresta S, Pham CT, Thomas DA, Graubert TA, Ley TJ. How docytotoxic lymphocytes kill their targets? Curr Opin Immunol1998;10:581–7.
[21] Ham RG, St. Clair JA, Webster C, Blau HM. Improved media fornormal human muscle satellite cells: serum-free clonal growth andenhanced growth with low serum. In Vitro Cell Dev Biol1988;24:833–44.
[22] Skuk D, Goulet M, Tremblay JP. Use of repeating dispensers toincrease the efficiency of the intramuscular myogenic cell injectionprocedure. Cell Transplant 2006;15:659–63.
[23] Engel AG, Biesecker G. Complement activation in muscle fibernecrosis: demonstration of the membrane attack complex of comple-ment in necrotic fibers. Ann Neurol 1982;12:289–96.
[24] Cullen MJ, Mastaglia FL. Pathological reactions of skeletal muscle.In: Mastaglia FL, Walton J, editors. Skeletal muscle pathology. Lon-don: Churchill Livingstone; 1982. p. 88–139.
[25] Hohlfeld R, Engel AG, Goebels N, Behrens L. Cellular immunemechanisms in inflammatory myopathies. Curr Opin Rheumatol1997;9:520–6.
[26] Arahata K, Engel AG. Monoclonal antibody analysis of mononu-clear cells in myopathies. I: Quantitation of subsets according todiagnosis and sites of accumulation and demonstration and counts ofmuscle fibers invaded by T cells. Ann Neurol 1984;16:193–208.
[27] Engel AG, Arahata K. Mononuclear cells in myopathies: quantita-tion of functionally distinct subsets, recognition of antigen-specificcell-mediated cytotoxicity in some diseases, and implications for thepathogenesis of the different inflammatory myopathies. Hum Pathol1986;17:704–21.
[28] Arahata K, Engel AG. Monoclonal antibody analysis of mononu-clear cells in myopathies. IV: Cell-mediated cytotoxicity and musclefiber necrosis. Ann Neurol 1988;23:168–73.
[29] Dalakas MC. Muscle biopsy findings in inflammatory myopathies.Rheum Dis Clin North Am 2002;28:779–98.
[30] Engel AG, Arahata K. Monoclonal antibody analysis of mononu-clear cells in myopathies. II: Phenotypes of autoinvasive cells inpolymyositis and inclusion body myositis. Ann Neurol1984;16:209–15.
[31] Arahata K, Engel AG. Monoclonal antibody analysis of mononu-clear cells in myopathies. III: Immunoelectron microscopy aspects ofcell-mediated muscle fiber injury. Ann Neurol 1986;19:112–25.
[32] Stadelmann C, Lassmann H. Detection of apoptosis in tissue sections.Cell Tissue Res 2000;301:19–31.
[33] Samali A, Zhivotovsky B, Jones D, Nagata S, Orrenius S. Apoptosis:cell death defined by caspase activation. Cell Death Differ1999;6:495–6.
[34] Tews DS. Muscle-fiber apoptosis in neuromuscular diseases. MuscleNerve 2005;32:443–58.
[35] Dalakas MC. Pathophysiology of inflammatory and autoimmunemyopathies. Presse Med 2011;40:e237–47.
[36] Dalakas MC. Review: an update on inflammatory and autoimmunemyopathies. Neuropathol Appl Neurobiol 2011;37:226–42.
[37] Schneider C, Gold R, Dalakas MC, et al. MHC class I-mediatedcytotoxicity does not induce apoptosis in muscle fibers nor ininflammatory T cells: studies in patients with polymyositis, dermat-omyositis, and inclusion body myositis. J Neuropathol Exp Neurol1996;55:1205–9.
[38] Behrens L, Bender A, Johnson MA, Hohlfeld R. Cytotoxic mecha-nisms in inflammatory myopathies. Co-expression of Fas andprotective Bcl-2 in muscle fibres and inflammatory cells. Brain1997;120(Pt 6):929–38.
[39] Eckle VS, Buchmann A, Bursch W, Schulte-Hermann R, Schwarz M.Immunohistochemical detection of activated caspases in apoptotichepatocytes in rat liver. Toxicol Pathol 2004;32:9–15.
[40] Tidball JG, Albrecht DE, Lokensgard BE, Spencer MJ. Apoptosisprecedes necrosis of dystrophin-deficient muscle. J Cell Sci1995;108(Pt 6):2197–204.
Related Documents











