Naturally Immortalised Mouse Embryonic Fibroblast Lines Support Human Embryonic Stem Cell Growth Mavi Camarasa, 1 Daniel Brison, 1 Susan J. Kimber, 1 and Alan H. Handyside 2 Abstract Human embryonic stem cell (hESC) growth is dependent on various factors released by feeder cells. Some of them have already been elucidated, although much research is still needed to understand the biology of stem cell maintenance in culture. Traditionally, primary mouse embryonic fibroblasts (PMEFs) have been used as feeder layers, and both murine and human fibroblast cell lines have been shown to support pluripotency and self- renewal of hESC. Here we report the derivation of three new mouse embryonic fibroblast cell lines, MEFLU-M, MEFLU-T, and MEFLU-TB, with different properties regarding growth and support for undifferentiated hESCs. MEFLU-TB is able to support continuous growth of the newly derived Man-1, as well as H1, HUES-1, HUES-7, HUES-8, and HUES-9 human embryonic stem cell lines. After more than 50 passages and doublings, MEFLU-TB feeders compare to early passage primary mouse embryonic fibroblasts in their ability to support undifferen- tiated hESC growth. Our results contradict a previous paradigm that PMEFs tend to lose their capacity to support proliferation of hESCs with increasing passages, and show that the MEFLU-TB mouse embryonic fibroblast cell line and its conditioned medium have the potential to support the maintenance of hESC lines. Also, our results clearly show that spontaneous immortalization of primary fibroblasts can be achieved in culture without any chemical addition or genetic modification. Introduction P luripotent human embryonic stem cells (hESCs) promise to revolutionize the future of regenerative medi- cine through the development of cell and tissue replacement therapies. hESCs were first isolated in 1994 (Bongso et al., 1994) while the first continuous culture of a hES line was published in 1998 (Thompson et al., 1998). Our understanding of hESC bi- ology is still incomplete, but this is changing quickly as an increasing number of groups enrol in hESC research. Never- theless, much work is required if we are to realize their full therapeutic potential. One area in which there is much interest is that of cell culture protocols to obtain purer and safer pop- ulations of functionally normal undifferentiated hESC and hESC-derived cell types, useful in clinical applications (Mallon et al., 2006). Traditionally, primary mouse embryonic fibroblasts (PMEFs) have been used as feeder layers, and hESCs are still routinely cultured on mouse embryonic fibroblasts (MEFs). Cell lines such as the STO line derived from mouse SIM embryonic fi- broblasts and feeders from human origin were soon added to the list (Amit et al., 2003; Cheng et al., 2003; Inzunza et al., 2005; Park et al., 2004; Richards et al., 2002; 2003; Saxena et al., 2008; Xu et al., 2004) of feeder cells for hESCs growth. Genetically modified PMEF lines have been used successfully (Choo et al., 2006) to support hESC lines as well. In all cases the safety of genetically modified cells in a clinical context will have to be evaluated. The generation of immortal, nongenetically modi- fied feeder cell lines is relevant, offering a more stable culture to support hESCs growth, with a lower workload. To the best of our knowledge, no report to date has included nonmodified MEFs older than four to six passages for successful maintenance of hES cells as undifferentiated, pluripotent self-renewing pop- ulations. Here we show that fibroblast cell lines can be derived from fresh PMEFs, and that both they and the medium they have conditioned can effectively support growth of undifferentiated hESCs. These lines will constitute very useful models for the elucidation of the factors secreted by (Chin et al., 2007; Diecke et al., 2008; Greber et al., 2007) or present in feeder layers (Peiffer et al., 2008) that support pluripotent human stem cell growth. Materials and Methods Derivation of MEFLU-M, MEFLU-T, and MEFLU-TB lines Mouse embryonic fibroblasts were isolated from one pregnant MF-1 mouse at 12.5 days of gestation. Briefly, the 1 North West Embryonic Stem Cell Centre, Faculty of life Sciences, University of Manchester, Manchester, United Kingdom. 2 The London Bridge Fertility, Gynaecology and Genetics Centre, London Bridge, London, United Kingdom. CLONING AND STEM CELLS Volume 11, Number 3, 2009 ª Mary Ann Liebert, Inc. DOI: 10.1089=clo.2008.0082 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Naturally Immortalised Mouse Embryonic Fibroblast LinesSupport Human Embryonic Stem Cell Growth
Mavi Camarasa,1 Daniel Brison,1 Susan J. Kimber,1 and Alan H. Handyside2
Abstract
Human embryonic stem cell (hESC) growth is dependent on various factors released by feeder cells. Some ofthem have already been elucidated, although much research is still needed to understand the biology of stem cellmaintenance in culture. Traditionally, primary mouse embryonic fibroblasts (PMEFs) have been used as feederlayers, and both murine and human fibroblast cell lines have been shown to support pluripotency and self-renewal of hESC. Here we report the derivation of three new mouse embryonic fibroblast cell lines, MEFLU-M,MEFLU-T, and MEFLU-TB, with different properties regarding growth and support for undifferentiated hESCs.MEFLU-TB is able to support continuous growth of the newly derived Man-1, as well as H1, HUES-1, HUES-7,HUES-8, and HUES-9 human embryonic stem cell lines. After more than 50 passages and doublings, MEFLU-TBfeeders compare to early passage primary mouse embryonic fibroblasts in their ability to support undifferen-tiated hESC growth. Our results contradict a previous paradigm that PMEFs tend to lose their capacity tosupport proliferation of hESCs with increasing passages, and show that the MEFLU-TB mouse embryonicfibroblast cell line and its conditioned medium have the potential to support the maintenance of hESC lines.Also, our results clearly show that spontaneous immortalization of primary fibroblasts can be achieved inculture without any chemical addition or genetic modification.
Introduction
Pluripotent human embryonic stem cells (hESCs)promise to revolutionize the future of regenerative medi-
cine through the development of cell and tissue replacementtherapies. hESCs were first isolated in 1994 (Bongso et al., 1994)while the first continuous culture of a hES line was published in1998 (Thompson et al., 1998). Our understanding of hESC bi-ology is still incomplete, but this is changing quickly as anincreasing number of groups enrol in hESC research. Never-theless, much work is required if we are to realize their fulltherapeutic potential. One area in which there is much interestis that of cell culture protocols to obtain purer and safer pop-ulations of functionally normal undifferentiated hESC andhESC-derived cell types, useful in clinical applications (Mallonet al., 2006).
Traditionally, primary mouse embryonic fibroblasts (PMEFs)have been used as feeder layers, and hESCs are still routinelycultured on mouse embryonic fibroblasts (MEFs). Cell linessuch as the STO line derived from mouse SIM embryonic fi-broblasts and feeders from human origin were soon added tothe list (Amit et al., 2003; Cheng et al., 2003; Inzunza et al., 2005;Park et al., 2004; Richards et al., 2002; 2003; Saxena et al., 2008;Xu et al., 2004) of feeder cells for hESCs growth. Genetically
modified PMEF lines have been used successfully (Choo et al.,2006) to support hESC lines as well. In all cases the safety ofgenetically modified cells in a clinical context will have to beevaluated. The generation of immortal, nongenetically modi-fied feeder cell lines is relevant, offering a more stable culture tosupport hESCs growth, with a lower workload. To the best ofour knowledge, no report to date has included nonmodifiedMEFs older than four to six passages for successful maintenanceof hES cells as undifferentiated, pluripotent self-renewing pop-ulations. Here we show that fibroblast cell lines can be derivedfrom fresh PMEFs, and that both they and the medium they haveconditioned can effectively support growth of undifferentiatedhESCs. These lines will constitute very useful models for theelucidation of the factors secreted by (Chin et al., 2007; Dieckeet al., 2008; Greber et al., 2007) or present in feeder layers (Peifferet al., 2008) that support pluripotent human stem cell growth.
Materials and Methods
Derivation of MEFLU-M, MEFLU-T,and MEFLU-TB lines
Mouse embryonic fibroblasts were isolated from onepregnant MF-1 mouse at 12.5 days of gestation. Briefly, the
1North West Embryonic Stem Cell Centre, Faculty of life Sciences, University of Manchester, Manchester, United Kingdom.2The London Bridge Fertility, Gynaecology and Genetics Centre, London Bridge, London, United Kingdom.
CLONING AND STEM CELLSVolume 11, Number 3, 2009ª Mary Ann Liebert, Inc.DOI: 10.1089=clo.2008.0082
1

mouse was sacrificed by cervical dislocation and uterine hornswere dissected out and rinsed in PBS containing 0.5 U=mLpenicillin and 0.5mg=mL of streptomycin. Each embryo wasseparated from its placenta and surrounding membranes anddecapitated. Their body walls were isolated and minced withscissors, before 0.05% trypsin and 0.02% EDTA (Lonza Wo-kingham, UK, http:==www.lonza.com) and modified HanksBalanced Salt Solution (HBSS, Lonza) was added to incubatethem at 378C for 30 min. A crude cell isolate composed oftissue pieces, cell clumps, and single-cell suspension waswashed, centrifugued, and resuspended in complete DMEM,consisting of high glucose DMEM supplemented with 10%fetal bovine serum (FBS), 1.5 g=L sodium bicarbonate, 2 mML-Glutamine and 1� nonessential aminoacids (NEAA) (allfrom Lonza), and cultured in the same medium as describeduntil confluence. Three different subcultures were set up fromthe initial isolate by separating adherent monolayers (Fig. 1A)from tissue clumps (Fig. 1B) and single-cell suspensions (Fig.1C) by centrifugation, and were cultured in separate flasks asdescribed. All three subcultures yielded adherent monolayersof cells with fibroblastic cell morphology after continuouspassages performed by trypsinization. They were bulked upseparatedly to passage 2 and mostly frozen to prepare pri-mary MEF feeder layers at passage 4 after successive thawingand expansion. Further passages of aliquots of the three sep-arate subcultures were undertaken using trypsin-EDTA(0.05% trypsin 1:250 and 0.02% EDTA, from Lonza) every 4 to6 days, until steady growth was established. Cultures weresplit for 20 consecutive passages prior to calculating thepopulation doubling curve. The three resulting fibroblastic cell
lines were assigned as MEFLU-M (Fig. 1A), MEFLU-T (Fig.1B), and MEFLU-TB (Fig. 1C).
Feeder layer preparation and feeder-free culture
PMEFs at passage 4 were treated with 10mg=mLmitomycin-C (Sigma M4287, Sigma-Aldrich, Dorset, www.sigmaaldrich.com) in complete DMEM for 6 h, trypsinized,and seeded on 0.1% gelatin (Sigma G1890)-coated tissue cul-ture dishes a day before the hESCs were plated at a density of5–7�104 cells=cm2 onto six-well plates (Corning Surrey,www.corning.com). Alternatively, 4�106 cells were frozen pervial using a slow-freezing method, in 90% FBS and 10%DMSO (Sigma D2650), and thawed onto six-well gelatinizedplates to prepare feeder layers as described before. For theMEFLUs lines, treatment was as described for PMEFs, andpassage number was the only variable parameter. Feederswere used no later than 48 h after plating, and discarded after8–10 days of use as support for stem cell growth. Feeder-freecultures were set up using Matrigel� (BD Biosciences Oxford,http:==www.bdeurope.com) according to manufacturer pro-tocol, as follows: tissue culture plates were coated with 1:30dilution of Matrigel� in cold KO-DMEM (Invitrogen, Carls-bad, CA, www. Invitrogen.com) medium, and left for 2 h atroom temperature (RT) before use. After washing in KO-DMEM, medium conditioned by either PMEFs and MEFLUscultures was added and equilibrated in the CO2 incubatorprior to hESCs seeding.
HES cell culture and embryoid body formation
The newly derived hESC line Man-1 at the North WestEmbryonic Stem Cell Centre (Camarasa et al, in preparation),H1 from WiCell, and HUES-1, HUES-7, HUES-8, and HUES-9from Harvard University, were maintained as pluripotentcolonies on PMEFs feeder layers in knockout Dulbecco’smodified Eagle’s Medium (KO-DMEM, Invitrogen), and=orDMEM=F12 (Lonza) media. Supplementation in each casewas with 10% or 20% knockout serum replacement (KO-SR),(Invitrogen), 4 or 8 ng=mL basic fibroblast growth factor(bFGF) (Invitrogen), 2 mM L-glutamine, 1% NEAA (Lonza),and 0.1 mM b-mercaptoethanol (Sigma). Cells were passagedeither mechanically or with 200 U collagenase IV (Invitrogen)per well in six-well plates. To demonstrate their pluripotency,a basic protocol of in vitro differentiation via embryoid bodyformation was followed: hESC colonies were mechanicallysplit and cultured in bacteriological Petri dishes in suspensionfor 8 to 10 days in DMEM high glucose supplemented with10% fetal bovine serum, 2 mM L-glutamine, and 1% NEAA(all from Lonza), during which embryoid body formation wasfollowed. When cystic morphologies appeared, embryoidbodies were harvested and gently triturated by pipetting.They were plated onto serum-coated glass coverslips, andsubsequently cultured in DMEM high glucose supplementedwith 10% FBS for 25 to 32 days, after which immunofluores-cence assays were performed as described below to detectdifferentiation markers.
Mycoplasma determination
All cultures were checked routinely for mycoplasma infec-tion with either the Immu-Mark inmunofluorescence kit (MPBiomedicals Cambridge, http:==www.mpbio.com) according
FIG. 1. Diagram representing the derivation of the threemouse embryonic fibroblasts cell lines MEFLU-M, MEFLU-T,and MEFLU-TB. From initial pooling and digestion of 12embryos on day 12.5 of development, the whole culture wasdivided in three fractions: cells forming an adherent mono-layer (A), tissue clumps (B), and suspended cells (C). In ev-ery case the subculture produced a monolayer of fibroblasts,which were subsequently cultured separately to generate thethree sublines subjected to study.
2 CAMARASA ET AL.
to the manufacturer instructions, or DNA staining using DAPI(Sigma D9564).
Immunofluorescence detection of stem celland pluripotency markers
Primary antibodies to human NANOG, OCT-4, TRA-1-60,TRA-1-81, SSEA-3, and SSEA-4 (R&D Systems, Abingdon,http:==www.rndsystems.com) were localized using AMCA(Millipore, Livingston, http:==www.millipore.com), FITCand TRITC (R&D Systems) conjugated secondary antibodies.Briefly, cells were seeded on feeder layers or Matrigel matrixwith conditioned medium in 48-well plates (Corning, Dorset,http:==www.corning.com), and cultured for 2 to 6 days.Afterward, cells were washed three times in PBS (Lonza) andfixed in 4% buffered paraformaldehyde (PFA, Sigma) for
15 min at RT. Washing buffer with 0.2% Tween-20 (Sigma)was used after fixation, and blocked with 10% FBS, 0.2%Tween-20 and 0.2% gelatin from cold water fish skin (Sigma)for 2 h in a humid chamber under gentle agitation. Primaryantibodies were diluted in the same blocking buffer and in-cubated overnight at 48C. Three washes were performedduring 5 min each afterward, and secondary antibodies wereincubated for 2 h at RT. Images were acquired using a Zeissinverted microscope equipped with a camera and processedusing AxioVision software (Carl Zeiss, Hertfordshire, www.zeiss.com=micro).
Primary antibodies to markers of each of the three germlayers AFP (alpha-phetoprotein) and Pax-6 (endoderm andendoderm=neurectoderm, respectively), ASMA (alpha-smoothmuscle actin), CD31 and aggrecan (mesoderm), and beta-tubulin III and Musashi-1 (ectoderm), were detected using
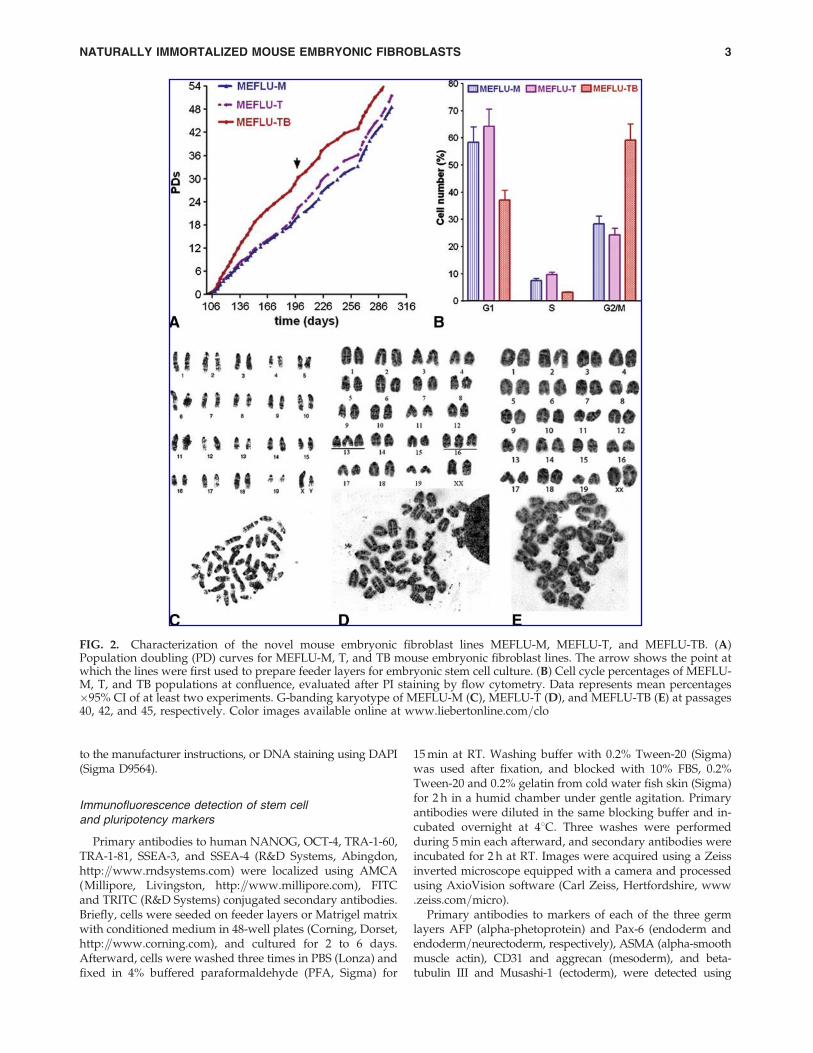
FIG. 2. Characterization of the novel mouse embryonic fibroblast lines MEFLU-M, MEFLU-T, and MEFLU-TB. (A)Population doubling (PD) curves for MEFLU-M, T, and TB mouse embryonic fibroblast lines. The arrow shows the point atwhich the lines were first used to prepare feeder layers for embryonic stem cell culture. (B) Cell cycle percentages of MEFLU-M, T, and TB populations at confluence, evaluated after PI staining by flow cytometry. Data represents mean percentages�95% CI of at least two experiments. G-banding karyotype of MEFLU-M (C), MEFLU-T (D), and MEFLU-TB (E) at passages40, 42, and 45, respectively. Color images available online at www.liebertonline.com=clo
NATURALLY IMMORTALIZED MOUSE EMBRYONIC FIBROBLASTS 3

appropriate FITC and TRITC conjugated secondary antibodies(all from R&D Systems).
AP detection
Alkaline phosphatase (AP) activity was measured using APmagenta kit (Sigma), according to manufacturer instructions.Briefly, the cells were fixed with buffered paraformaldehyde(PFA) solution for 15 min, and stained with AP staining so-lution in the dark for at least 45 min. Bright-field images wereacquired using an Olympus CKX41 inverted microscopeequipped with a C5060 color digital camera.
G-banding
A standard optimized G-banding technique (Henegariuet al., 2001) was used to karyotype MEFs MEFLU and hesclines, with slight modifications. Briefly, actively proliferatingcells were treated with colcemid (1 mg=mL, Sigma) for 3 h,trypsinized, washed in PBS, and incubated in an hypotonicPBS (74.8� 5 mOsm=kg H2O) before incubating in Carnoy’sfixative (cold methanol=acetic acid 3:1) (BDH AnalAR�,VWR, Poole, http:==uk.vwr.com). Extracted nuclei werespread in standard or 5-mm diameter multiwell Teflon-coated slides (Electron Microscopy Sciences, Hatfield, PA,http:==www.emsdiasum.com=microscopy) and dried rapidlyat 858C. After a minimum ‘‘aging’’ period of 24 h at 558C,spreads were treated with 0.025% trypsin for 60 sec, beforebeing incubated with freshly diluted Giemsa stain in Gurr’sbuffer (Sigma) for 30 min. Slides were mounted in mineral oilor with DEPEX, and pictures were taken on a bright-fieldmicroscope under an immersion 100� objective. Thirty se-lected metaphases were counted, and 10 of them werealigned to evaluate the banding pattern in each case.
Cell Cycle=DNA profile analysis
MEFLU cells in single-cell suspensions of approximately1.0�106 cells=mL in PBS were fixed in 70% ethanol on ice for1.5 h and incubated with 500 mg=mL RNAse A (Sigma) for30 min on ice. Cells were then incubated with 25 mg=mL ofpropidium iodide in PBS for 30 min at RT, and analyzed in aFACScan Flow Cytometer (Beckton Dickinson, Oxford, http:==www.bdeurope.com) with associated software. At least 10,000events were collected for each sample. Unstained controlswere included in the experiments, which were performed induplicate.
Results
Derivation of MEFLU-M, MEFLU-T,and MEFLU-TB lines
Fresh PMEFs were isolated from pooled 12.5 dpc embryosfrom a single pregnant MF-1 mouse and cultured in absenceof antibiotics from first passage (considering explanted cells aspassage 0) and maintained routinely in exponential growth.Pooled MEFs were expanded up to passage 2 and frozen, sofeeder layers were prepared from thawed MEFs, inactivatedat passage 4 in each case. In parallel, three sublines were de-rived from the first primary culture, which were namedMEFLU-M, MEFLU-T, and MEFLU-TB, as they were originallyseeded from the first fibroblast monolayer (M), a secondary
digestion from tissue clumps formed at first digestion (T),and secondary attached fibroblasts from suspension cultureisolated from the same first culture (TB) (see Fig. 1). Theirdifferent growth properties as population doublings for allthree sublines from passage 20 and onwards are shown inFigure 2A. MEFLU-TB grew faster from the beginning of theestablishment of the MEF sublines, with an average increaseof 47.2� 3.1 percentage difference in doublings with respect toMEFLU-M during the whole evaluation period. Feeder layerswere prepared from all three sublines from passage 40 on-wards (denoted by arrow). The establishment of the contin-uous culture for all three sublines surpassed replicative crisiseasily (cellular senescence was compensated by cell growth atall times in all cultures), and all seemed to show a weak orabsent contact inhibition from the beginning of their culture.Cell cycle profile in a MEFLU-TB apparently confluent popu-lation showed no G1 enrichment, and a considerably increasedpopulation in late S and G2=M phases in exponentiallygrowing cultures, consistent with the absence of a cell cycleblockade (G1 arrest) typical of those at high density (Camarasaet al., 2005). Representative data of cell cycle analysis in ap-parently confluent populations of all three sublines MEFLU-M,MEFLU-T, and MEFLU-TB showed 59% of cells in G2=Mphase in MEFLU-TB, which is more than twice that expectedfor a confluent population, as seen in MEFLU-M and -T pop-ulations, whose G2=M phase at confluence was of 28 and 24%,respectively (Fig. 2B). MEFLU-TB cells did not show contactinhibition, so we explored the possibility of embryonic stemcell growth support by cell lines showing different growthproperties. Despite continuous growth shown by all threesublines, and the uncommon cell cycle profile shown byMEFLU-TB, it retained a normal karyotype along with ME-FLU-M (see Fig. 2) after more than 30 passages, and supportedhESC growth when used as feeder layers in standard stem cellculture conditions and as conditioned medium in feeder-freecultures (Fig. 3 and Supplemental Fig. 1; see online supple-mentary material at www.liebertonline.com). In contrast,MEFLU-T was trisomic for chromosomes 13 and 16, and wasless consistent in its support of undifferentiated hESc growth,both in feeder and feeder-free cultures with their CM (Sup-plementary Fig. 1).
The morphology of MEFs within each culture did not changeduring the derivation process. Nevertheless, feeders preparedunder the same conditions from MEFs and MEFLUs weremorphologically different: the processes formed by MEFLUcells were longer and thinner than those observed inPMEFs during their use as feeder layers (see SupplementaryFig. 2).
Characterization of HESC lines grown on feedersderived from MEFLU fibroblast lines
Stem cell cultures were maintained routinely on primaryMEFs for more than 4 years on PMEFS and characterizedusing immunofluorescence for the typical stem cell markersNanog, Oct-4, TRA-1-60, TRA-1-80, SSEA-3, SSEA-4, andalkaline phosphatase (AP). Stem cell marker analyses wereperformed on Man-1 at passage 8, and on H1, HUES-1,HUES-7, HUES-8, and HUES-9 after 5 passages in our lab-oratory (at passages 20–35), and were checked routinelythereafter. H1 and HUES lines showed intense stain for TRA-
4 CAMARASA ET AL.
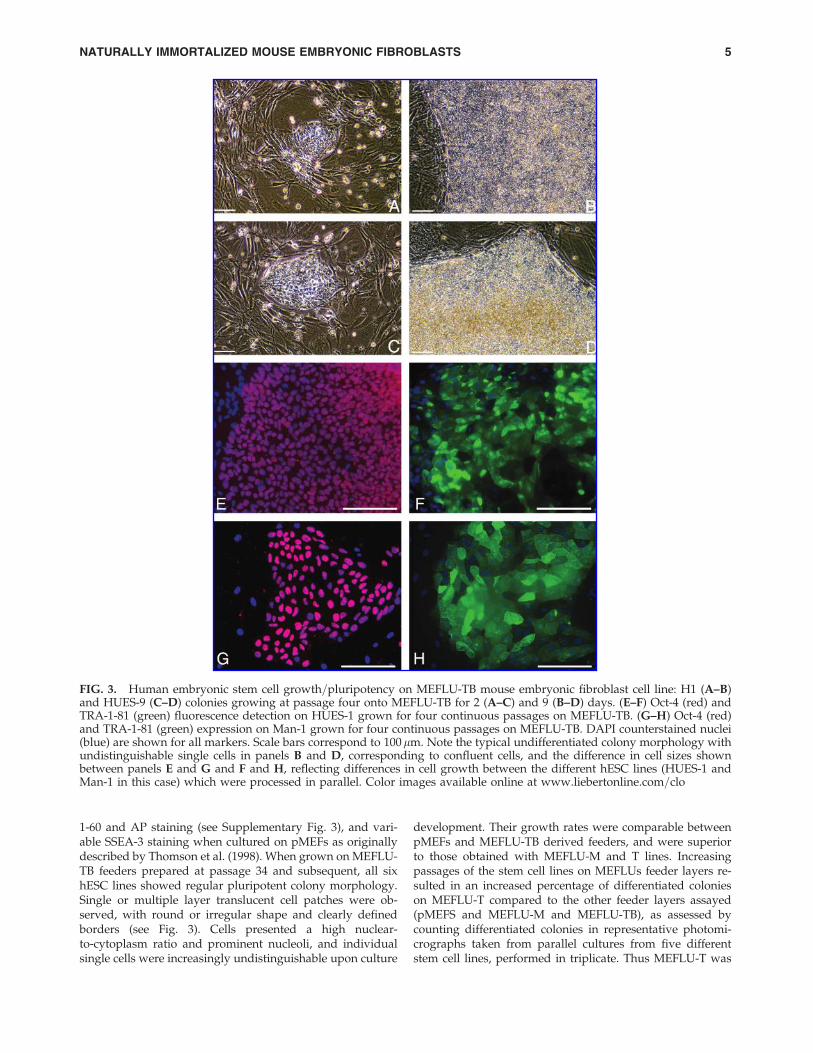
1-60 and AP staining (see Supplementary Fig. 3), and vari-able SSEA-3 staining when cultured on pMEFs as originallydescribed by Thomson et al. (1998). When grown on MEFLU-TB feeders prepared at passage 34 and subsequent, all sixhESC lines showed regular pluripotent colony morphology.Single or multiple layer translucent cell patches were ob-served, with round or irregular shape and clearly definedborders (see Fig. 3). Cells presented a high nuclear-to-cytoplasm ratio and prominent nucleoli, and individualsingle cells were increasingly undistinguishable upon culture
development. Their growth rates were comparable betweenpMEFs and MEFLU-TB derived feeders, and were superiorto those obtained with MEFLU-M and T lines. Increasingpassages of the stem cell lines on MEFLUs feeder layers re-sulted in an increased percentage of differentiated colonieson MEFLU-T compared to the other feeder layers assayed(pMEFS and MEFLU-M and MEFLU-TB), as assessed bycounting differentiated colonies in representative photomi-crographs taken from parallel cultures from five differentstem cell lines, performed in triplicate. Thus MEFLU-T was
FIG. 3. Human embryonic stem cell growth=pluripotency on MEFLU-TB mouse embryonic fibroblast cell line: H1 (A–B)and HUES-9 (C–D) colonies growing at passage four onto MEFLU-TB for 2 (A–C) and 9 (B–D) days. (E–F) Oct-4 (red) andTRA-1-81 (green) fluorescence detection on HUES-1 grown for four continuous passages on MEFLU-TB. (G–H) Oct-4 (red)and TRA-1-81 (green) expression on Man-1 grown for four continuous passages on MEFLU-TB. DAPI counterstained nuclei(blue) are shown for all markers. Scale bars correspond to 100mm. Note the typical undifferentiated colony morphology withundistinguishable single cells in panels B and D, corresponding to confluent cells, and the difference in cell sizes shownbetween panels E and G and F and H, reflecting differences in cell growth between the different hESC lines (HUES-1 andMan-1 in this case) which were processed in parallel. Color images available online at www.liebertonline.com=clo
NATURALLY IMMORTALIZED MOUSE EMBRYONIC FIBROBLASTS 5
nonsupportive, followed by MEFLU-M and MEFLU-TB. Thelatter was equivalent to primary MEF cultures in ability tosupport pluripotent stem cell growth of all six lines assayed.All six lines were maintained for a year in culture on ME-FLU-TB feeders (see Table 1 for detail on minimum numberof passages performed), were successfully frozen–thawed,and characterised as described below.
Pluripotent stem cell growth support of MEFLU-TB wasconfirmed by immunostaining for the typical stem cellmarkers. OCT-4 and TRA-1-81 (see Fig. 3), NANOG andhTERT (data not shown) were detected in HUES-1 and Man-1 after four consecutive passages on MEFLU-TB feeders(passages 41–44), and colony morphology and growth ratiowere maintained in all six lines tested during more than ayear.
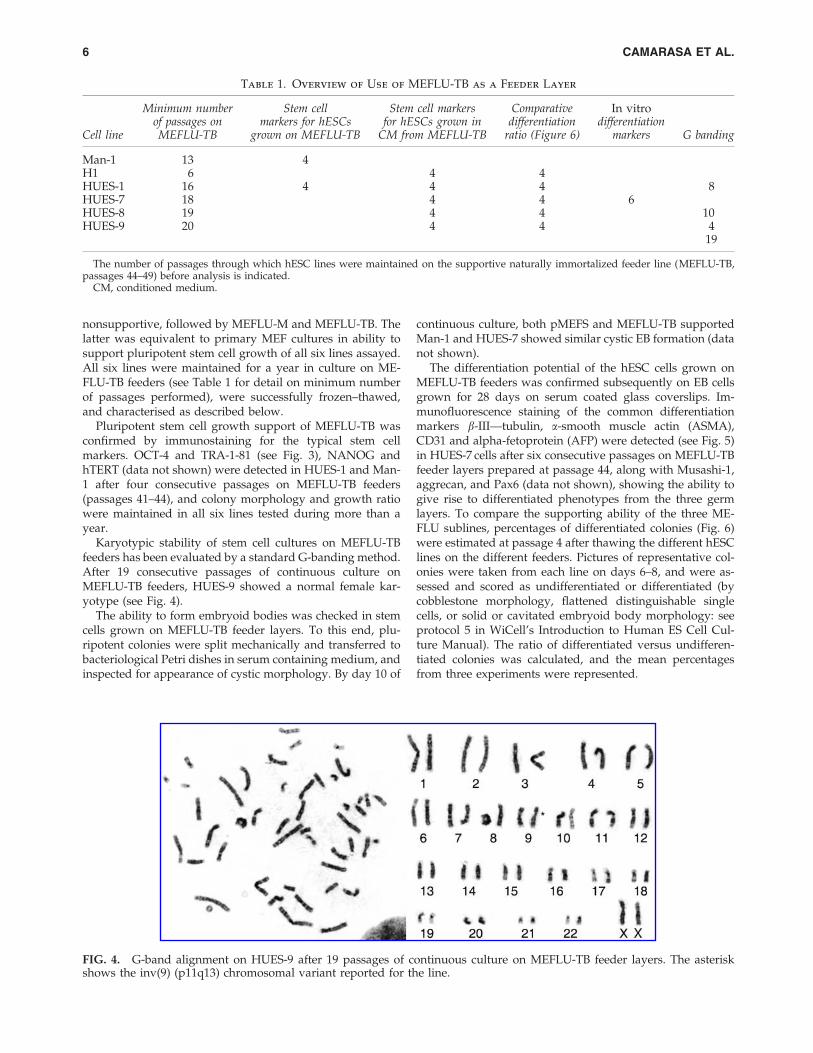
Karyotypic stability of stem cell cultures on MEFLU-TBfeeders has been evaluated by a standard G-banding method.After 19 consecutive passages of continuous culture onMEFLU-TB feeders, HUES-9 showed a normal female kar-yotype (see Fig. 4).
The ability to form embryoid bodies was checked in stemcells grown on MEFLU-TB feeder layers. To this end, plu-ripotent colonies were split mechanically and transferred tobacteriological Petri dishes in serum containing medium, andinspected for appearance of cystic morphology. By day 10 of
continuous culture, both pMEFS and MEFLU-TB supportedMan-1 and HUES-7 showed similar cystic EB formation (datanot shown).
The differentiation potential of the hESC cells grown onMEFLU-TB feeders was confirmed subsequently on EB cellsgrown for 28 days on serum coated glass coverslips. Im-munofluorescence staining of the common differentiationmarkers b-III—tubulin, a-smooth muscle actin (ASMA),CD31 and alpha-fetoprotein (AFP) were detected (see Fig. 5)in HUES-7 cells after six consecutive passages on MEFLU-TBfeeder layers prepared at passage 44, along with Musashi-1,aggrecan, and Pax6 (data not shown), showing the ability togive rise to differentiated phenotypes from the three germlayers. To compare the supporting ability of the three ME-FLU sublines, percentages of differentiated colonies (Fig. 6)were estimated at passage 4 after thawing the different hESClines on the different feeders. Pictures of representative col-onies were taken from each line on days 6–8, and were as-sessed and scored as undifferentiated or differentiated (bycobblestone morphology, flattened distinguishable singlecells, or solid or cavitated embryoid body morphology: seeprotocol 5 in WiCell’s Introduction to Human ES Cell Cul-ture Manual). The ratio of differentiated versus undifferen-tiated colonies was calculated, and the mean percentagesfrom three experiments were represented.
FIG. 4. G-band alignment on HUES-9 after 19 passages of continuous culture on MEFLU-TB feeder layers. The asteriskshows the inv(9) (p11q13) chromosomal variant reported for the line.
Table 1. Overview of Use of MEFLU-TB as a Feeder Layer
Cell line
Minimum numberof passages onMEFLU-TB
Stem cellmarkers for hESCs
grown on MEFLU-TB
Stem cell markersfor hESCs grown in
CM from MEFLU-TB
Comparativedifferentiation
ratio (Figure 6)
In vitrodifferentiation
markers G banding
Man-1 13 4H1 6 4 4HUES-1 16 4 4 4 8HUES-7 18 4 4 6HUES-8 19 4 4 10HUES-9 20 4 4 4
19
The number of passages through which hESC lines were maintained on the supportive naturally immortalized feeder line (MEFLU-TB,passages 44–49) before analysis is indicated.
CM, conditioned medium.
6 CAMARASA ET AL.
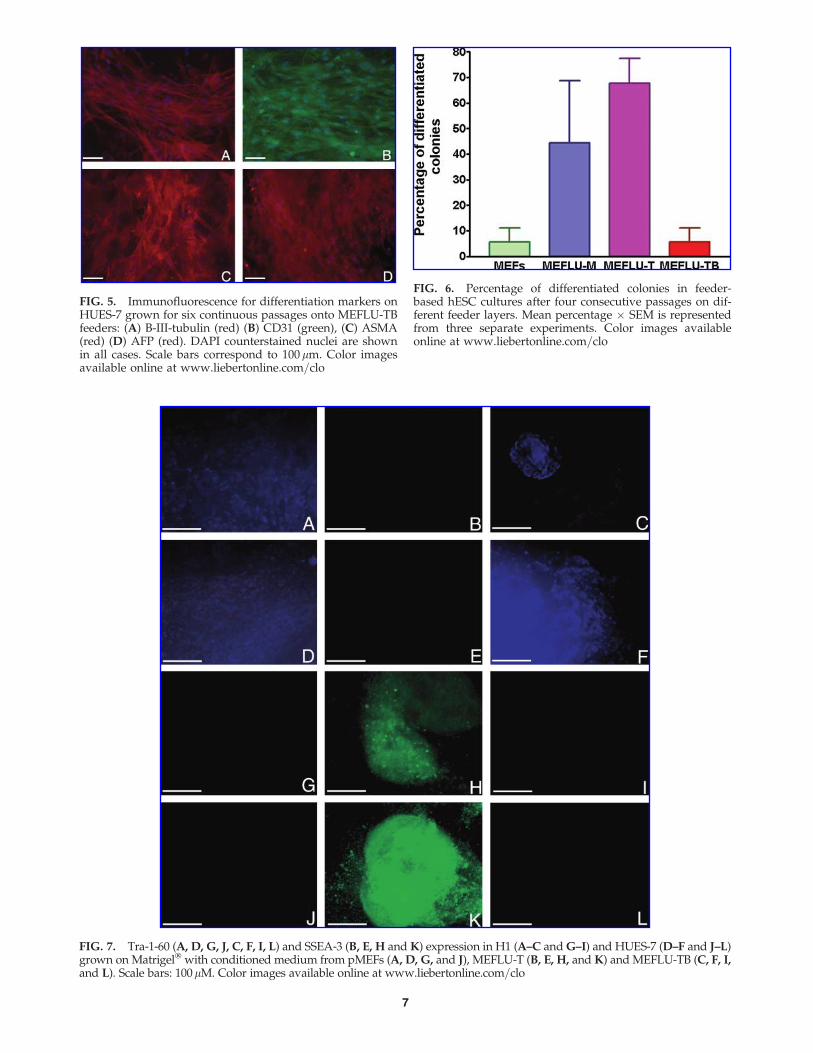
FIG. 5. Immunofluorescence for differentiation markers onHUES-7 grown for six continuous passages onto MEFLU-TBfeeders: (A) B-III-tubulin (red) (B) CD31 (green), (C) ASMA(red) (D) AFP (red). DAPI counterstained nuclei are shownin all cases. Scale bars correspond to 100 mm. Color imagesavailable online at www.liebertonline.com=clo
FIG. 6. Percentage of differentiated colonies in feeder-based hESC cultures after four consecutive passages on dif-ferent feeder layers. Mean percentage � SEM is representedfrom three separate experiments. Color images availableonline at www.liebertonline.com=clo
FIG. 7. Tra-1-60 (A, D, G, J, C, F, I, L) and SSEA-3 (B, E, H and K) expression in H1 (A–C and G–I) and HUES-7 (D–F and J–L)grown on Matrigel� with conditioned medium from pMEFs (A, D, G, and J), MEFLU-T (B, E, H, and K) and MEFLU-TB (C, F, I,and L). Scale bars: 100mM. Color images available online at www.liebertonline.com=clo
7

MEFLU conditioned media (CM) supportsundifferentiated growth of hESCs
Preliminary experiments in which cultures were split fromfresh PMEFs to MEFLU sublines (at passages 30–50) inclassical feeder-based culture systems, resulted in failure ofMEFLU-T to sustain pluripotent hESC growth, with coloniesgrowing in height without expansion soon after the begin-ning of each culture, and signs of differentiation (increasedcytoplasm=nucleus ratio), whereas MEFLU-TB and MEFLU-M continuously supported undifferentiated cell growth aswell as fresh PMEFs for more than four passages. Whenfrozen stocks of hESC colonies, enzymatically separatedfrom feeders, were used for the experiments with MEFLUssublines, all sublines initially sustained pluripotent colonyformation, but differences in pluripotent cell growth anddifferentiation were observed across successive passages.Three separate experiments were conducted with three dif-ferent passages of five of the six hESC lines studied (H1,HUES-1, HUES-7, HUES-8, and HUES-9) for four consecu-tive passages in each case on all four feeder layers (pMEFS,MEFLU-M, MEFLU-T, and MEFLU-TB), resulting in undif-ferentiated cell and colony morphologies for all hESC lineson all three MEFLU lines.
To further address the issue of differences in pluripotentstem cell growth support (ability) between the four differentfeeder layers (primary MEFS as reference and the threeMEFLU sublines), conditioned media (CM) were preparedfrom all three sublines, and assayed in parallel with CM fromprimary MEFs in hESC cultures on Matrigel� as described inthe Methods section. Colonies maintained growth and un-differentiated morphology during at least four consecutivepassages in this feeder-free system (performed in triplicateusing different passages of five hESC lines in each case,ranging from passage 16 to 38). When conditioned mediafrom the new MEFLUs lines were used in these feeder-freecultures, pluripotent colony growth was also demonstrated,although changes were observed in pluripotency markersexpression (see Fig. 7). For MEFLU-M and MEFLU-TB re-sults were the same as described for fresh PMEFs, with hESCstaining strongly for TRA-1-60 in all five hESC lines assayed,but not for SSEA-3. In contrast, when MEFLU-T conditionedmedium was used on Matrigel� cultures, TRA-1-60 was notdetectable and SSEA-3 (green fluorescence in Fig. 7 in theonline version of the manuscript) was consistently strong inall hESC lines.
Discussion
Scientists and clinicians are now pursuing the applica-bility of human embryonic stem cells in regenerative med-icine, and in generating patient-specific stem cell lines. Thederivation of embryonic stem cell lines from embryosbearing monogenic disorders is also seen as a goal withgreat promise in the advancement of our understanding ofsuch conditions. In the era of successful hESC lines, culturewith human feeders, or in feeder-free cultures, it seemsplausible that the embryonic origin of the feeders may beimportant to support of the pluripotent cell state, as re-ported previously (Richards et al., 2003). Moreover, cell linesfrom adult human skin have been used successfully tosupport hESC culture (Richards et al., 2003), and a systemhas been optimized to use fibroblasts derived from hESCs as
feeders for other hESC lines (Stojkovic et al., 2005; Wanget al., 2005; Yoo et al., 2005). Previous limitations of thetechniques for the culture of human stem cells, such as theuse of only embryonic nonhuman feeders, have alreadybeen overcome by the scientific community. Recently, ahigh-passage human foreskin fibroblast line was used tosupport hESC growth in xeno-free conditions (Meng et al.,2008). As we show in this report, primary MEFs do not needto be newly derived and as generally assumed used onlyuntil fifth to sixth passage to prepare feeder layers or con-ditioned medium for stem cell growth support. In this re-port we have shown that novel mouse embryonic fibroblastcell lines cultured in serum-containing media are a valuabletool to substitute for primary cultures in the preparation offeeder layers for human embryonic stem cell culture. Ourthree new lines, MEFLU-M, MEFLU-T, and MEFLU-TB,have been characterized in terms of their support of plu-ripotent stem cell growth of six different human embryonicstem cell lines, of three different origins, namely, H1 fromThe University of Wisconsin (Thompson et al., 1998), four ofthe HUES lines from Harvard University (Cowan et al.,2004), and one of the new lines derived in-house at theNorth West Embryonic Stem Cell Centre, Man-1. All hESClines were maintained and expanded for at least 6 months inculture on MEFLU-TB feeder cells at passages 44–49, andevery experiment described was performed using at leastone hESC line. No difference was observed in the extent ofdifferentiation for hESCs grown on primary MEFs andMEFLU-TBs, even at high passage number. It is importantto note that MEFLU-T, which supported pluripotent statuspoorly compared to fresh PMEFs, was found to promotechanges in pluripotent marker expression in hESCs when itsconditioned medium was used in a feeder-free culture sys-tem. In our hands, H1 cell line does not express SSEA-3 andis strongly positive for TRA-1-60 as initially described byThomson et al. (1998). Also, the HUES lines tested negativefor SSEA-3 expression in regular or feeder-free cultures ofPMEFs, but were strongly positive in the colonies formedin MEFLU-T-based cultures after 3 days of growth. As thepercentage of differentiated hES cells increased with pas-sages in MEFLU-T supported cultures (both with feedersand in feeder-free cultures with conditioned medium), thisobserved change in SSEA-3 expression could be reflecting anearly response to a change in suboptimal culture conditions.
We have characterized hESC cultured for between 4 and19 passages on MEFLU lines, including thawing vials di-rectly on the new feeder lines instead of switching after asplit, thus avoiding the carryover effect from donor feederlayers and growth factors. Thus, the comparison betweenPMEFs and MEFLUs has been direct, as duplicate vials havebeen used in the experiments and approximately the samenumbers of cells were plated onto different feeder layers.When cells pooled from a split were used to plate on ME-FLUs to compare them, differences in growth were less clear,possibly because the culture retained factors from previousfresh PMEFs feeders, known to support long-term self-renewal in hES cells. Thawing stem cell lines on MEFLUswas repeated three times to rule out artifactual results. Ineach case MEFLU-TB was supportive and indistinguishablefrom primary MEFs, as evaluated by cell growth (as colonynumbers) and percentage of differentiated cells, MEFLU-Twas increasingly less-supportive over several passages and
8 CAMARASA ET AL.

effectively unusable for continuous hESc support, andMEFLU-M was somewhat less supportive than PMEFs andMEFLU-TB. The MEFLU-TB subline has been successfullysplit routinely to prepare feeder layers, and all six stem celllines assayed have reached more than 20 consecutive pas-sages on several late passages of the MEFLU-TB line (44 to50). It is now clear that cell lines generated from MEFs canretain the characteristics of primary MEFs, providing aconvenient, long-lived and lasting supply of feeder cells forhuman embryonic stem cell culture. More importantly, theability of the MEFLU-TB conditioned medium to supportpluripotent hESC growth makes the system a true substitutefor primary MEFs.
Interestingly, the MEFLU cell line with chromosomal ab-normalities (MEFLU-T) is the least-supportive one. AlthoughMEFLU-TB had similar potency to MEFS for hESC support,another chromosomally normal line (MEFLU-M) offeredvariable results regarding support of pluripotency, implyingthat every feeder line generated would need to be validated.In future work it will be very interesting and informative todecipher which factors are involved in the loss of pluripotenthESc support. In the elucidation of which factors are released=expressed by feeder cells to allow human pluripotent cellgrowth, the study of the putative genetic differences betweenMEFLU-M, MEFLU-T, and MEFLU-TB at the subchromo-some level may shed light on both the factors that supportstemness and those that cause immortalization. Genotypiccharacterization of all three lines and primary cultures is inprogress for this purpose. It seems though that a chromo-somally normal cell line with immortal growth propertiesprovides adequate long-term support for pluripotent hESCgrowth, and as such, will be a valuable hESC culture tool. Itwill be essential to avoid the need for repeatedly generatingpMEFs for hESC culture support if hESC propagation movesto an industrial scale. Alternative development of feeder-freeculture systems for hESC is being carried out by us (Baxteret al., in press) and others (Cheon et al., 2006; Klimanskayaet al., 2005; Ludwig et al., 2006; Vallier et al., 2005) as sci-entists pursue the complete avoidance of feeder cell culture,and protocols are being optimized so that universal systemsfor efficient and reproducible maintenance of hESCs can bedelivered.
In summary, we have pioneered the spontaneous im-mortalization of mouse embryonic fibroblast lines andproved that such cell lines are true substitutes for primaryMEFs in human embryonic stem cell culture technology.Ongoing research will analyze the mechanisms of the im-mortalization, and dissect the characteristics of and compo-nents released by MEF feeder layers that are required tosupport hESC renewal and pluripotency.
Acknowledgments
The authors want to greatly thank the technical helpprovided by Mrs Beverly Merry at the Faculty of BiologicalSciences at the University of Leeds, expert advice in kar-yotyping provided by Mrs Fiona Small, at the CytogeneticsRegional Unit at St Mary’s Hospital in Manchester, andcritical reading of the manuscript by Dr. Rachel Oldershaw.This work was supported by a grant from the North WestDevelopment Agency to D.R.B. and S.J.K. and from a grantfrom the MRC to A.H.
Author Disclosure Statement
All authors declare they have no competing financialinterests.
References
Amit, M., Margulets, V., Segev, H., et al. (2003). Human feederlayers for human embryonic stem cells. Biol. Reprod. 68, 2150–2156.
Bongso, A., Fong, C.Y., Ng, S.C., et al. (1994). Isolation andculture of inner cell mass cells from human blastocysts. Hum.Reprod. 9, 2110–2117.
Camarasa, M.V., Castro-Galache, M.D., Carrasco-Garcia, E., et al.(2005). Differentiation and drug resistance relationships inleukemia cells. J. Cell Biochem. 94, 98–108.
Cheng, L., Hammond, H., Ye, Z., et al. (2003). Human adultmarrow cells support prolonged expansion of human em-bryonic stem cells in culture. Stem Cells 21, 131–142.
Cheon, S.H., Kim, S.J., Jo, J.Y., et al. (2006). Defined feeder-freeculture system of human embryonic stem cells. Biol. Reprod.74, 611–618.
Chin, A.C., Fong, W.J., Goh, L.T., et al. (2007). Identification ofproteins from feeder conditioned medium that support humanembryonic stem cells. J Biotechnol. 30, 320–328.
Choo, A., Padmanamhan, J., Chin, A., et al. (2006). Immortalizedfeeders for the scale-up of human embryonic stem cellsin feeder and feeder-free conditions. J. Biotechnol. 122, 130–141.
Cowan, C.A., Klimanskaya, I., Mcmahon, J., et al. (2004). Deri-vation of embryonic stem-cell lines from human blastocysts.N. Engl. J. Med. 350, 1353–1356.
Diecke, S., Quiroga-Negreira, A., Redmer, T., et al. (2008). FGF2signaling in mouse embryonic fibroblasts is crucial for self-renewal of embryonic stem cells. Cells Tissues Organs 188,52–61.
Greber, B., Lehrach, H., and Adjaye, J. (2007). Fibroblast growthfactor 2 modulates transforming growth factor beta signalingin mouse embryonic fibroblasts and human ESCs (hESCs) tosupport hESC self-renewal. Stem Cells. 25, 455–464.
Henegariu, O., Heerema, N.A., Wright, L.L., et al. (2001). Im-provements in cytogenetic slide preparation, controlled chro-mosome spreading, chemical aging and gradual denaturing.Cytometry 43, 101–109.
Inzunza, J., Gertow, K., Stromberg, M.A., et al. (2005). Deriva-tion of human embryonic stem cell lines in serum replacementmedium using postnatal human fibroblasts as feeder cells.Stem Cells 23, 544–549.
Klimanskaya, I., Chung, Y., Meisner, L., et al. (2005). Humanembryonic stem cells derived without feeder cells. Lancet 365,1636–1641.
Ludwig, T.E., Levenstein, M.E., Jones, J.M., et al. (2006). Deri-vation of human embryonic stem cells in defined conditions.Nat. Biotechnol. 24, 185–187.
Mallon, B.S., Park, K.Y., Chen, K.G., et al. (2006). Toward xeno-free culture of human embryonic stem cells. Int. J. Biochem.Cell Biol. 38, 1063–1075.
Meng, G., Liu, S., Krawetz, R., et al. (2008). A novel method forgenerating xeno-free human feeder cells for human embryonicstem cell culture. Stem Cells Dev. 17, 413–422.
Park, S.P., Lee, Y.J., Lee, K.S., et al. (2004). Establishment ofhuman embryonic stem cell lines from frozen-thawed blasto-cysts using STO cell feeder layers. Hum. Reprod. 19, 676–684.
Peiffer, I., Barbet, R., Zhou, Y.P., et al. (2008). Use of xenofreematrices and molecularly-defined media to control human
NATURALLY IMMORTALIZED MOUSE EMBRYONIC FIBROBLASTS 9

embryonic stem cell pluripotency, effect of low physiologicalTGF-beta concentrations. Stem Cells Dev. 17, 519–533.
Richards, M. Fong, C.Y., Chan, W.K., et al. (2002). Humanfeeders support prolonged undifferentiated growth of humaninner cell masses and embryonic stem cells. Nat. Biotechnol.20, 933–936.
Richards, M., Tan, S., Fong, C.Y., et al. (2003). Comparativeevaluation of various human feeders for prolonged undiffer-entiated growth of human embryonic stem cells. Stem Cells21, 546–556.
Saxena, S., Hanwate, M., Deb, K., et al. (2008). FGF2 secretinghuman fibroblast feeder cells, a novel culture system for hu-man embryonic stem cells. Mol. Reprod. Dev. 75, 1523–1532.
Stojkovic, P., Lako, M., Stewart, R., et al. (2005). An autogeneicfeeder cell system that efficiently supports growth of undif-ferentiated human embryonic stem cells. Stem Cells 23, 306–314.
Thompson, J.A., Itskovitz-Eldor, J., Shapiro, S.S., et al. (1998).Embryonic stem cell lines derived from human blastocysts.Science 282, 1145–1147.
Vallier, L., Alexander, M., and Pedersen, R.A. (2005). Activin=nodal and FGF pathways cooperate to maintain pluripotency
of human embryonic stem cells. J. Cell. Sci. 118(Pt 19), 4495–4509.
Wang, Q., Fang, Z.F., Jin, F., et al. (2005). Derivation andgrowing human embryonic stem cells on feeders derived fromthemselves. Stem Cells 23, 1221–1227.
Xu, C., Jiang, J., Sottile, V., et al. (2004). Immortalized fibroblast-like cells derived from human embryonic stem cells supportundifferentiated cell growth. Stem Cells 22, 972–980.
Yoo, S.J., Yoon, B.S., Kim, J.M., et al. (2005). Efficient culturesystem for human embryonic stem cells using autologoushuman embryonic stem cell-derived feeder cells. Exp. Mol.Med. 37, 399–407.
Address correspondence to:Mavi Camarasa
North West Embryonic Stem Cell CentreFaculty of life Sciences
University of Manchester46 Grafton Street
M13 9NT Manchester, UK
E-mail: [email protected]
10 CAMARASA ET AL.
Related Documents











