© 2003 Blackwell Publishing Ltd 403 Parasite Immunology , 2003, 25, 403–412 Blackwell Publishing Ltd. ORIGINAL ARTICLE ANTIBODY RESPONSES TO MSP6 and MSP7 Naturally acquired antibody responses to the components of the Plasmodium falciparum merozoite surface protein 1 complex LINA WANG 1 , LUCY CROUCH 1 , THOMAS L. RICHIE 2 , DOAN HANH NHAN 3 & ROSS L. COPPEL 1 1 Department of Microbiology and the Victoria Bioinformatics Consortium, Monash University, Clayton, Victoria, Australia, 2 Malaria Program, Naval Medical Research Centre, Silver Spring, Maryland, USA and 3 Institute for Malariology, Parasitology and Entomology, Hanoi, Vietnam SUMMARY Merozoite surface protein 6 (MSP6) and 7 (MSP7) of Plas- modium falciparum are peripheral membrane proteins whose cleaved products, MSP6 36 , MSP7 22 and MSP7 19 , are found on the merozoite surface as components of a non-covalently bound complex which also contains four polypeptides derived from merozoite surface protein 1 (MSP1). We have expressed both the precursor regions and the processed mature products of MSP6 and MSP7 in Escherichia coli and showed that these recombinant proteins react with human immune sera. In a set of sera collected from individuals living in malaria- endemic areas of Southern-central Vietnam, antibodies to the mature polypeptides of MSP6 36 and MSP7 22 were detected in 50·6 and 85·6% of the serum samples, whereas antibodies to the precursor regions of MSP6 and MSP7 were detected in only 12·1 and 42·5% of the serum samples, respectively. The predominant subclass of anti-MSP6 antibodies was IgG1, whereas the predominant subclass of anti-MSP7 antibodies was IgG3. In the same set of serum samples, the antibody responses to MSP1 19 are predominantly IgGI, whereas anti- bodies to merozoite surface protein 4 (MSP4) are mainly IgG3. This data is consistent with the proposition that, during malaria infection, variable proteins induce responses that are predominantly of the IgG3 isotype, and conserved proteins induce responses that are predominantly IgG1. The antibodies to MSP6, MSP7 and MSP1 19 all decreased at the time of infection, but increased during the convalescent period. No correlation was observed between the antibodies at the com- mencement of the study and absence of parasitaemia during surveillance in this population. Keywords antibody response, MSP6, MSP7, MSP1, Plas- modium falciparum Abbreviations: ELISA, enzyme-linked immunosorbent assay; MSP, merozoite surface protein INTRODUCTION Effective immunity to Plasmodium falciparum blood-stage infection involves the acquisition of anti-parasite antibodies targeted at various exposed proteins of the parasite, includ- ing surface proteins of the invasive merozoite (1,2). Immuno- globulin (Ig) purified from immune adults can passively transfer protection against P. falciparum infection (3,4). Further, the absence of parasite-specific antibodies to the parasite is associated with a poor clinical prognosis (5). The identity of these target antigens remains to be fully elucid- ated, although several proteins, including merozoite surface protein 1 (MSP1), have been implicated (6). MSP1 is the most-abundant and best-characterized mero- zoite surface component. It is synthesized by the intracellu- lar schizont as a large precursor protein (185–210 kDa) and undergoes proteolytic processing during the maturation of merozoites (7–9). Primary proteolytic processing of the precursor produces four major polypeptide fragments of approximately 83, 28–30, 38 and 42 kDa. These fragments, MSP1 83 , MSP1 30 , MSP1 38 and MSP1 42 , exist as a non- covalently associated complex on the merozoite surface. A second proteolytic processing event cleaves MSP1 42 to yield polypeptides of approximately 33 and 19 kDa (MSP1 33 and MSP1 19 ). MSP1 19 remains anchored to the merozoite membrane and is carried into the invaded erythrocyte (7). MSP1 33 , together with the other polypeptides derived from primary processing, is shed as a complex into the extra- cellular milieu (8). The shed MSP1 complex also contains three polypeptides of 36, 22 and 19 kDa that are the expressed product of two other genes (10,11). The 36-kDa protein is derived from a Correspondence: Ross L. Coppel, Department of Microbiology and the Victoria Bioinformatics Consortium, Monash University, Clayton, Victoria 3800, Australia (e-mail: [email protected]). Received: 20 June 2003 Accepted for publication: 26 August 2003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© 2003 Blackwell Publishing Ltd 403
Parasite Immunology, 2003, 25, 403–412
Blackwell Publishing Ltd.ORIGINAL ARTICLEANTIBODY RESPONSES TO MSP6 and MSP7Naturally acquired antibody responses to the components of the
Plasmodium falciparum merozoite surface protein 1 complex
LINA WANG1, LUCY CROUCH1, THOMAS L. RICHIE2, DOAN HANH NHAN3 & ROSS L. COPPEL1
1Department of Microbiology and the Victoria Bioinformatics Consortium, Monash University, Clayton, Victoria, Australia, 2Malaria Program, Naval Medical Research Centre, Silver Spring, Maryland, USA and 3Institute for Malariology, Parasitology and Entomology, Hanoi, Vietnam
SUMMARY
Merozoite surface protein 6 (MSP6) and 7 (MSP7) of Plas-modium falciparum are peripheral membrane proteins whosecleaved products, MSP636, MSP722 and MSP719, are foundon the merozoite surface as components of a non-covalentlybound complex which also contains four polypeptides derivedfrom merozoite surface protein 1 (MSP1). We have expressedboth the precursor regions and the processed mature productsof MSP6 and MSP7 in Escherichia coli and showed thatthese recombinant proteins react with human immune sera.In a set of sera collected from individuals living in malaria-endemic areas of Southern-central Vietnam, antibodies to themature polypeptides of MSP636 and MSP722 were detected in50·6 and 85·6% of the serum samples, whereas antibodies tothe precursor regions of MSP6 and MSP7 were detected inonly 12·1 and 42·5% of the serum samples, respectively. Thepredominant subclass of anti-MSP6 antibodies was IgG1,whereas the predominant subclass of anti-MSP7 antibodieswas IgG3. In the same set of serum samples, the antibodyresponses to MSP119 are predominantly IgGI, whereas anti-bodies to merozoite surface protein 4 (MSP4) are mainlyIgG3. This data is consistent with the proposition that, duringmalaria infection, variable proteins induce responses that arepredominantly of the IgG3 isotype, and conserved proteinsinduce responses that are predominantly IgG1. The antibodiesto MSP6, MSP7 and MSP119 all decreased at the time ofinfection, but increased during the convalescent period. Nocorrelation was observed between the antibodies at the com-mencement of the study and absence of parasitaemia duringsurveillance in this population.
Keywords antibody response, MSP6, MSP7, MSP1, Plas-modium falciparum
Abbreviations: ELISA, enzyme-linked immunosorbent assay;MSP, merozoite surface protein
INTRODUCTION
Effective immunity to Plasmodium falciparum blood-stageinfection involves the acquisition of anti-parasite antibodiestargeted at various exposed proteins of the parasite, includ-ing surface proteins of the invasive merozoite (1,2). Immuno-globulin (Ig) purified from immune adults can passivelytransfer protection against P. falciparum infection (3,4).Further, the absence of parasite-specific antibodies to theparasite is associated with a poor clinical prognosis (5). Theidentity of these target antigens remains to be fully elucid-ated, although several proteins, including merozoite surfaceprotein 1 (MSP1), have been implicated (6).
MSP1 is the most-abundant and best-characterized mero-zoite surface component. It is synthesized by the intracellu-lar schizont as a large precursor protein (185–210 kDa) andundergoes proteolytic processing during the maturation ofmerozoites (7–9). Primary proteolytic processing of theprecursor produces four major polypeptide fragments ofapproximately 83, 28–30, 38 and 42 kDa. These fragments,MSP183, MSP130, MSP138 and MSP142, exist as a non-covalently associated complex on the merozoite surface. Asecond proteolytic processing event cleaves MSP142 to yieldpolypeptides of approximately 33 and 19 kDa (MSP133 andMSP119). MSP119 remains anchored to the merozoitemembrane and is carried into the invaded erythrocyte (7).MSP133, together with the other polypeptides derived fromprimary processing, is shed as a complex into the extra-cellular milieu (8).
The shed MSP1 complex also contains three polypeptidesof 36, 22 and 19 kDa that are the expressed product of twoother genes (10,11). The 36-kDa protein is derived from a
Correspondence: Ross L. Coppel, Department of Microbiology and the Victoria Bioinformatics Consortium, Monash University, Clayton, Victoria 3800, Australia (e-mail: [email protected]).Received: 20 June 2003 Accepted for publication: 26 August 2003

404 © 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412
L. Wang et al. Parasite Immunology
precursor designated merozoite surface protein 6 (MSP6)(12), while the 22 and 19 kDa proteins are both derivedfrom a precursor designated merozoite surface protein 7(MSP7) (13). Both MSP6 and MSP7 are peripheral mem-brane proteins encoded by genes encoding proteins thatpossess an N-terminal secretion signal, but lack membraneanchor sequences and are stabilized on the merozoite sur-face through protein–protein interactions with MSP1-derived polypeptides (12,13).
MSP1 has been shown to be a major target of humanimmune responses to malaria. Antibodies to MSP119 arefound in most malaria-exposed individuals and have beencorrelated with protection from clinical symptoms of malariain some, but not all, studies (14–18). In an in vitro study,antibodies to MSP119 appear to contribute about 50% of thegrowth inhibitory activity of human immune sera in certainserum samples (6), but its importance as a target for othermechanisms of immunity, such as antibody-dependent cel-lular inhibition, is unknown. Antibodies to other regions ofMSP1 may also play a role in immunity. For example, thepresence of antibodies to block 2 located in MSP183 wasassociated with reduced risk for clinical malaria in childrenin The Gambia (19). Antibodies to MSPl were predomin-antly of the cytophilic subclasses of immunoglobulin G(IgG), indicating that such antibodies may play a role inopsonization and complement mediated lysis of free mero-zoites (15,20,21). There has been no published informationon the prevalence, specificity and isotype distribution ofantibodies induced by infection to the other proteins inthe MSPl complex. In this study, we examined naturallyacquired antibodies to both MSP6 and MSP7 in a group ofindividuals living in the Khanh-Hoa region of high endem-icity of Southern-central Vietnam.
MATERIALS AND METHODS
Construction of recombinant MSP6 and MSP7 proteins
Two fragments (A and B) of MSP6 and MSP7 were ampli-fied from genomic DNA of P. falciparum 3D7 strain byPCR and cloned into pGEX-KG (AMRAD PharmaciaBiotech, Melbourne, Vic, Australia). As shown in Figure 1,MSP6A contains residues 20–160 of MSP6, whereas MSP7Acontains residues 23–176 of MSP7, both of which are notpresent in the mature products. MSP6B contains residues161–371 and spans the region encoding MSP636, whereasMSP7B contains residues 177–351 and spans the regionencoding MSP722. The genes encoding these various frag-ments were sequenced and found to agree completely withthe reported sequences (12,13), with the exception of MSP7Bwhich showed a difference from the reported sequence attwo residues: a mutation from H to Y at residue 241 and amutation from D to G at residue 242. To exclude the possi-bility of cloning- or polymerase-induced artifact, productsof two separate PCR reactions were directly sequencedand the differences were confirmed as being present in thegenomic DNA and representing allelic diversity in MSP7.The recombinant plasmids were transformed into E. colistrain BL21 (Novagen, Milwaukee, WI, USA), the glutathione-S-transferase (GST) fusion proteins were expressed byinduction with isopropyl β--thio-galactopyranoside (Pro-gen Industries Limited, Darra, Qld, Australia) and purifiedby affinity chromatography on glutathione agarose asdescribed previously (22). These fusion proteins are desig-nated GST-MSP6A, GST-MSP6B, GST-MSP7A and GST-MSP7B, their calculated molecular masses are 43, 52, 45and 48 kDa, respectively.
Figure 1 Schematic representation (to scale) of MSP6 and MSP7. Black boxes at the left indicate signal sequences. The shaded boxes indicate the cleaved products of MSP636 and MSP722. The cleavage site in MSP722 to produce MSP719 is indicated by an arrow. Horizontal bars below the schematic indicate the regions that are expressed as GST fusion proteins. The first and last amino acid residues included in each recombinant protein are given.

© 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412 405
Volume 25, Number 8/9, August/September 2003 Antibody responses to MSP6 and MSP7
Recombinant MSP119
MSP119, the carboxyl-terminal fragment of MSP1 thatcontains two EGF-like domains, was expressed as a GSTfusion protein in E. coli. The correct conformation of thisrecombinant protein has been demonstrated by its reactivitywith several biologically important monoclonal antibodies(23).
Human sera
The serum samples examined in this study were collectedfrom residents living in the Khanh-Nam Commune ofKhanh-Hoa Province in Southern-central Vietnam. Threespecies of Plasmodium are endemic to this area. A cross-sectional survey conducted in June 1994 showed bloodsmear positivity rates of 14·4% for P. falciparum, 11·3% forP. vivax and 1·0% for P. malariae. We classified olderchildren and adults living in this area as semi-immune,based on the observation that only half of those with para-sitaemia described symptoms consistent with malaria andthat these symptoms were often mild (e.g. headache). Thestudy subjects and collection of serum samples have beendescribed previously (18). Briefly, at the commencement ofthe study in June 1994, blood samples (T0) were obtainedwith informed consent from 132 volunteers aged from 9 to55 years. These volunteers were radically treated withquinine sulphate, doxycycline hyclate and primaquinephosphate, and then followed up daily by questioning forsymptoms and weekly by obtaining peripheral blood smearsby finger prick, for a period of 6 months. Individuals whodeveloped patent parasitaemia over the course of the studywere treated and blood samples were collected at the timeof treatment (T1) and 28 days later (T28). Of the 112 indi-viduals who completed surveillance, 47 became parasitaemicwith P. falciparum, including six with P. falciparum and P.vivax or P. malariae, 32 became parasitaemic with P. vivax,P. malariae, or both, and 33 did not develop a positive bloodsmear. Overall, infections were detected in about 70% ofthe individuals over the 6-month surveillance period; theirparasitaemia-free intervals (time to infection) were between36 and 156 days. No volunteers had recurrent parasitaemiaduring the 28 days of follow-up after mefloquine treatment.A total of 174 serum samples were chosen in this study toexamine antibody responses to MSP6 and MSP7, includingsamples collected at T0, T1 and T28 from the 47 individualswho acquired P. falciparum parasitaemia and the samplescollected at T0 from the 33 individuals who remained smearnegative.
A second set of sera were obtained from 22 residents ofthe Madang region of Papua New Guinea, all of whom haddocumented exposure to P. falciparum infection (22). Thirty
negative control sera were taken from Australian blooddonors who had no history of exposure to malaria (18).
SDS–PAGE and immunoblotting
Recombinant MSP6 and MSP7 proteins were fractioned bysodium dodecyl sulfate – polyacrylamide gel electrophoresis(SDS–PAGE) and transferred to polyvinylidene difluoride(PVDF) for immunoblotting as previously described (22).
Enzyme-linked immunosorbent assay (ELISA)
The reactivity of human sera with recombinant proteinswas examined by ELISA as previously described (18),except that the optical density (OD) was determined using aLabsystems Multiskan (Labsystems Oy, Helsinki, Finland).Each serum sample was tested in parallel against the GSTfusion proteins and GST alone as a negative control andthe specific OD values were calculated by subtracting thecontrol OD value from the values obtained from the fusionprotein. Positive sera were defined as those that gave anOD value greater than the mean plus 3 standard deviationsof OD values obtained using 30 negative control sera. Fordetection of the Ig isotypes, an isotype-specific ELISA wasperformed as previously described (18).
Data analysis
Statistical analysis was performed using GraphPad Prismsoftware (GraphPad Software Inc., San Diego, CA, USA).The chi-square test was used to compare proportions ofantibody responders in different groups, whereas the Wil-coxon and Mann–Whitney tests were used to compare theantibody levels between groups for paired and unpaireddata, respectively. Spearman’s rank correlation test was usedto correlate antibody reactivity with pairs of individualantigens.
RESULTS
Antigenicity of recombinant MSP6 and MSP7
Both the precursor regions (A) and the mature products (B)of MSP6 and MSP7 were expressed in E. coli as GST fusionproteins. When analysed on SDS–PAGE, GST-MSP6A andGST-MSP7B were present at the expected sizes, whereasGST-MSP6B and GST-MSP7A had higher observed mole-cular masses (Figure 2a). There were also a number ofsmaller bands in each fusion protein, which were partiallydegraded expression products. To assess whether the fusionproteins of MSP6 and MSP7 could be recognized by humanimmune sera, an equal amount of each fusion protein was
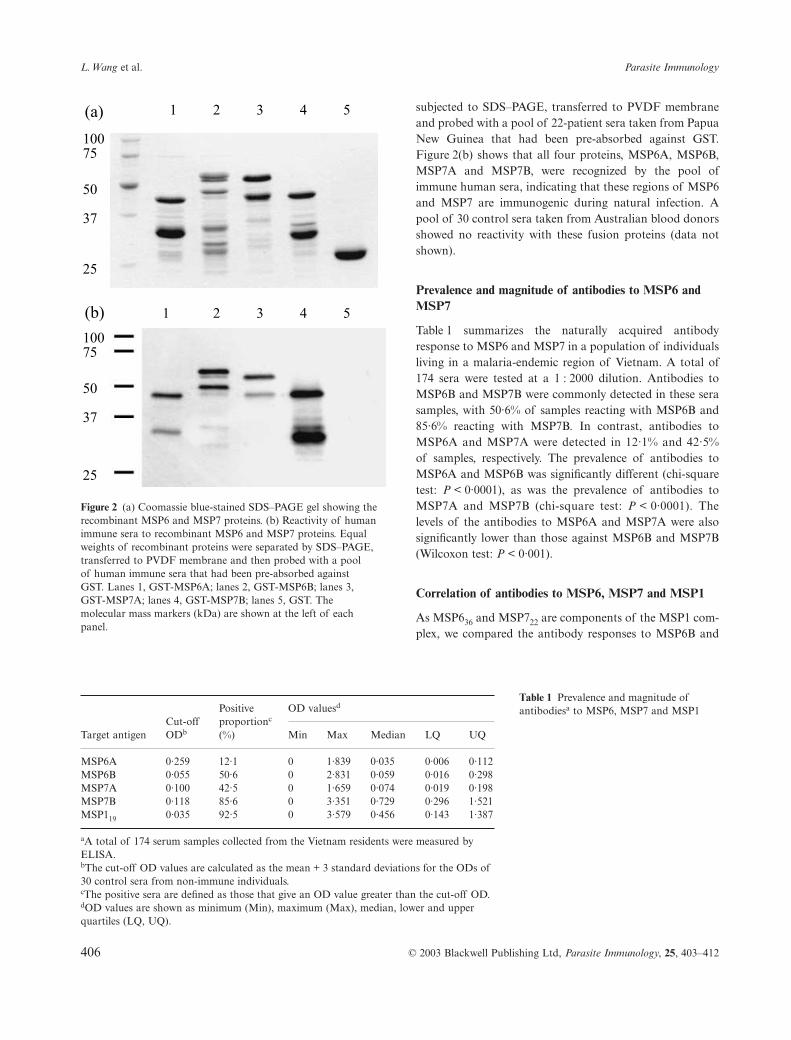
406 © 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412
L. Wang et al. Parasite Immunology
subjected to SDS–PAGE, transferred to PVDF membraneand probed with a pool of 22-patient sera taken from PapuaNew Guinea that had been pre-absorbed against GST.Figure 2(b) shows that all four proteins, MSP6A, MSP6B,MSP7A and MSP7B, were recognized by the pool ofimmune human sera, indicating that these regions of MSP6and MSP7 are immunogenic during natural infection. Apool of 30 control sera taken from Australian blood donorsshowed no reactivity with these fusion proteins (data notshown).
Prevalence and magnitude of antibodies to MSP6 and MSP7
Table 1 summarizes the naturally acquired antibodyresponse to MSP6 and MSP7 in a population of individualsliving in a malaria-endemic region of Vietnam. A total of174 sera were tested at a 1 : 2000 dilution. Antibodies toMSP6B and MSP7B were commonly detected in these serasamples, with 50·6% of samples reacting with MSP6B and85·6% reacting with MSP7B. In contrast, antibodies toMSP6A and MSP7A were detected in 12·1% and 42·5%of samples, respectively. The prevalence of antibodies toMSP6A and MSP6B was significantly different (chi-squaretest: P < 0·0001), as was the prevalence of antibodies toMSP7A and MSP7B (chi-square test: P < 0·0001). Thelevels of the antibodies to MSP6A and MSP7A were alsosignificantly lower than those against MSP6B and MSP7B(Wilcoxon test: P < 0·001).
Correlation of antibodies to MSP6, MSP7 and MSP1
As MSP636 and MSP722 are components of the MSP1 com-plex, we compared the antibody responses to MSP6B and
Cut-offODb
Positiveproportionc
(%)
OD valuesd
Target antigen Min Max Median LQ UQ
MSP6A 0·259 12·1 0 1·839 0·035 0·006 0·112MSP6B 0·055 50·6 0 2·831 0·059 0·016 0·298MSP7A 0·100 42·5 0 1·659 0·074 0·019 0·198MSP7B 0·118 85·6 0 3·351 0·729 0·296 1·521MSP119 0·035 92·5 0 3·579 0·456 0·143 1·387
aA total of 174 serum samples collected from the Vietnam residents were measured by ELISA.bThe cut-off OD values are calculated as the mean + 3 standard deviations for the ODs of 30 control sera from non-immune individuals.cThe positive sera are defined as those that give an OD value greater than the cut-off OD.dOD values are shown as minimum (Min), maximum (Max), median, lower and upper quartiles (LQ, UQ).
Table 1 Prevalence and magnitude of antibodiesa to MSP6, MSP7 and MSP1
Figure 2 (a) Coomassie blue-stained SDS–PAGE gel showing the recombinant MSP6 and MSP7 proteins. (b) Reactivity of human immune sera to recombinant MSP6 and MSP7 proteins. Equal weights of recombinant proteins were separated by SDS–PAGE, transferred to PVDF membrane and then probed with a pool of human immune sera that had been pre-absorbed against GST. Lanes 1, GST-MSP6A; lanes 2, GST-MSP6B; lanes 3, GST-MSP7A; lanes 4, GST-MSP7B; lanes 5, GST. The molecular mass markers (kDa) are shown at the left of each panel.

© 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412 407
Volume 25, Number 8/9, August/September 2003 Antibody responses to MSP6 and MSP7
MSP7B in our study population with those to MSP1. To dothis, we tested the serum samples at a 1 : 2000 dilutionagainst a correctly folded form of MSP119 (23). The anti-bodies to MSP119 were observed at a higher prevalence thanthose to MSP6B (chi-square test: P < 0·0001) or MSP7B (chi-square test: P = 0·025) (Table 1). The level of anti-MSP119
antibodies was also higher than the anti-MSP6B antibodies(Wilcoxon test: P < 0·001), but similar to the anti-MSP7Bantibodies (Wilcoxon test: P = 0·092). Antibodies to MSP6B,MSP7B and MSP119 were positively correlated (Figure 3),suggesting that responses to all three polypeptides werelinked. Of note, 3·8% of the populations did not show react-ivity to any of the three polypeptides in those assays despitemany years of residence in a highly endemic area.
Isotype distribution of the anti-MSP6 and -MSP7 antibodies
The isotype distribution of anti-MSP6 and -MSP7 antibod-ies was measured for MSP6B and MSP7B only, as MSP6Aand MSP7A were relatively poorly immunogenic. As shownin Tables 2 and 3, the anti-MSP6 and -MSP7 antibodieswere mainly of the cytophilic subclasses IgG1 and IgG3.IgG2 and IgG4 antibodies were present at low levels for bothanti-MSP6 and anti-MSP7 antibodies, whereas IgM waspresent at reasonably high levels in some individuals. It isinteresting to note that, within the two cytophilic subclasses,the predominant subclass of anti-MSP6 antibodies was IgG1both in terms of prevalence and level (chi-square test: P <0·0001; Wilcoxon test: P < 0·0001), whereas the predominantsubclass of anti-MSP7 antibodies was IgG3 (chi-squaretest: P < 0·0001; Wilcoxon test: P < 0·0001) (Figure 4a andFigure 4b). In the same set of serum samples, antibodiesto MSP119 were found to be predominantly of the IgG1isotype (Figure 4c). The predominance of IgG1 in antibodiesto MSP6 and MSP119, and the predominance of IgG3 inantibodies to MSP7, were also observed in the 22-patientsera collected in Papua New Guineans (data not shown).
Comparison of antibodies at different time points
The serum samples taken at T0, Tl and T28 from the 47individuals who acquired P. falciparum parasitaemia wereanalysed to compare the change in antibodies to MSP6 andMSP7 during infection and convalescence. The antibodiesto MSP6A, MSP6B, MSP7A, MSP7B and MSP119 had alldecreased at the time of infection, but were boosted duringthe convalescent period (Table 4). The difference in anti-body levels between T0 and Tl was statistically significantfor MSP6A, MSP7A and MSP7B, and the difference inantibody levels between T1 and T28 was statistically signi-ficant for MSP7B and MSP119. At 28 days after infection,
antibodies to MSP6, MSP7 and MSP119 all returned to levelscomparable with those measured at T0.
Lack of correlation between the antibodies and protective immunity
Of the 112 individuals who completed surveillance, weclassified the 47 who acquired P. falciparum parasitaemias
Figure 3 Pairwise correlation between antibody responses (OD) to MSP6, MSP7 and MSP1. (a) MSP6B vs. MSP119; (b) MSP7B vs. MSP119; (c) MSP6B vs. MSP7B. The Spearman’s correlation coefficient (rs) and the associated level of significance (P ) are shown on each graph.

408 © 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412
L. Wang et al. Parasite Immunology
Cut-offODb
Positive proportionc
(%)
OD valuesd
Isotype Min Max Median LQ UQ
IgG1 0·022 54·0 0 1·681 0·031 0·010 0·131IgG2 0·009 35·1 0 0·287 0·004 0·001 0·013IgG3 0·006 50·0 0 1·476 0·004 0 0·030IgG4 0·012 4·6 0 0·097 0 0 0·003IgM 0·087 42·0 0·017 2·896 0·071 0·045 0·133
aA total of 174 serum samples collected from the Vietnam residents were measured by isotype specific ELISA against recombinant MSP6B.bThe cut-off OD values are calculated as the mean + 3 standard deviations for the ODs of 30 control sera from non-immune individuals.cThe positive sera are defined as those that give an OD value greater than the cut-off OD, and the positive proportion represents the proportion of all sera in this study that are positive for the isotype indicated.dOD values are shown as minimum (Min), maximum (Max), median, lower and upper quartiles (LQ, UQ).
Table 2 Isotype-specific antibody responses to MSP6a
Cut-offODb
Positive proportionc
(%)
OD valuesd
Isotype Min Max Median LQ UQ
IgG1 0·057 56·9 0 1·354 0·078 0·029 0·196IgG2 0·038 0·6 0 0·045 0·006 0·003 0·010IgG3 0·011 83·9 0 2·972 0·154 0·036 0·852IgG4 0·016 3·4 0 1·021 0 0 0·003IgM 0·120 20·1 0·014 1·287 0·068 0·049 0·105
aA total of 174 serum samples collected from the Vietnam residents were measured by isotype-specific ELISA against recombinant MSP7B.bThe cut-off OD values are calculated as the mean + 3 standard deviations for the ODs of 30 control sera from non-immune individuals.cThe positive sera are defined as those that give an OD value greater than the cut-off OD, and the positive proportion represents the proportion of all sera in this study that are positive for the isotype indicated.dOD values are shown as minimum (Min), maximum (Max), median, lower and upper quartiles (LQ, UQ).
Table 3 Isotype-specific antibody responses to MSP7a
Table 4 Comparison of antibodies to MSP6, MSP7 and MSP1 in serum samples collected at different time pointsa
Target antigens
Antibody levelsb P valuesc
T0 T1 T28 T0 and T1 T1 and T28 T0 and T28
MSP6A 0·042 (0·008, 0·130) 0·027 (0·005, 0·107) 0·029 (0·005, 0·144) 0·0029 0·2604 0·1536MSP6B 0·065 (0·024, 0·361) 0·030 (0·014, 0·293) 0·074 (0·016, 0·587) 0·0092 0·0060 0·4022MSP7A 0·100 (0·029, 0·251) 0·039 (0·015, 0·206) 0·068 (0·019, 0·220) 0·0034 0·4092 0·0549MSP7B 1·061 (0·335, 1·572) 0·592 (0·280, 1·344) 1·002 (0·374, 1·711) 0·0006 < 0·0001 0·7610MSP119 0·527 (0·110, 1·527) 0·353 (0·140, 1·387) 0·896 (0·287, 2·133) 0·0374 < 0·0001 0·0094
aT0 samples were collected from 47 individuals at the beginning of the survey, T1 and T28 samples were collected from these individuals at the time of treatment for P. falciparum parasitaemia and 28 days after treatment respectively.bAntibody levels are represented as the median (lower quartile, upper quartile) of the OD values.cAntibody levels at two time points are compared using Wilcoxon test; the threshold for significance is 0·0034 after an adjustment is made for the number of comparisons.

© 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412 409
Volume 25, Number 8/9, August/September 2003 Antibody responses to MSP6 and MSP7
during the 6-month surveillance period as susceptible, andthe 33 who did not acquire parasitaemia with any species ofmalaria as potentially protected (18). The 32 individualswho developed parasitaemia with P. vivax or P. malariaewere excluded from this study because of the possibility thatthey were indeed susceptible to P. falciparum, but that thissusceptibility was not revealed because of an interveninginfection with other species. Comparison of the anti-MSP6and -MSP7 antibodies at T0, either for total Ig or individual
Ig isotypes, revealed no significant difference between thesetwo groups (data not shown). Analysis of the individuals inthe susceptible group did not reveal any significant correla-tion between antibody level and time to infection (data notshown). Similarly, no such association was observed in thisstudy population for antibodies to MSP119. Excluding theindividuals from the analysis who were parasitaemic at T0did not change the result.
DISCUSSION
Proteins on the merozoite surface of P. falciparum areimportant vaccine candidates against malaria infection, asthey are exposed to the immune system after merozoiterelease and are potential targets for antibodies that preventmerozoite invasion or lead to opsonization and parasitekilling. Several such proteins, such as MSP1 (14,16,17), mero-zoite surface protein 2 (MSP2) (24–26), merozoite surfaceprotein 3 (MSP3) (27), merozoite surface protein 4 (MSP4)(18), and apical membrane antigen 1 (28), have been examinedin immunoepidemiological studies and shown to be well recog-nized by human immune sera. Here, we show that MSP6and MSP7 are targets of naturally acquired antibodyresponses in people exposed to natural malaria infection.
We found that the precursor regions of MSP6 and MSP7were less strongly recognized by immune sera than their cor-responding mature products. As these precursor regions arenot present on the merozoite surface, nor are shed as solubleproteins (12,13), it is likely that they are degraded shortlyafter being made. This transient existence could tend tominimize exposure and lower overall immunogenicity. Nev-ertheless, the fact that they are immunogenic in a proportionof the study subjects suggests that exposure is taking place.As these regions are not found in free merozoites, it suggeststhat proteins of mature intracellular parasites are involvedin induction of antibody responses. Presumably they arederived from parasites killed by phagocytosis or followingingestion by antigen-presenting cells of parasites killed byother mechanisms. The lack of antibody response to MSP6or MSP7 in some individuals is not due to lack of exposure,as all residents have been exposed to parasites repeatedly(18). In addition, some sera were negative for MSP6 whilebeing highly positive for MSP7 and vice versa, indicatingepitope-specific factors influencing response, among whichmight be genetic restriction of immune responses to distinctepitopes. In general though, individuals tended to respondto multiple antigens and there is an overall positive correla-tion between an individual’s responses to MSP6, MSP7 andMSP1.
The levels of the anti-MSP6 and -MSP7 antibodies in theVietnam residents decreased at the time of P. falciparuminfection, but increased during the convalescent period and
Figure 4 Comparison of IgG subclass levels to MSP6 (a), MSP7 (b) and MSP119 (c). Each point shows antibody levels (in OD value) of IgG1 and IgG3 in a serum sample diluted at 1 : 200 for MSP6, MSP7 and 1 : 500 for MSP119.

410 © 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412
L. Wang et al. Parasite Immunology
were restored to levels comparable with those measured atthe beginning of the survey. This change in antibody levelsis also seen in antibody responses to MSP119 and MSP4(18). This may be a feature of this population, where infec-tions were prematurely terminated by therapy, or reflect thetransmission intensity of that region. No correlation wasobserved in this study population between the presence ofanti-MSP6 and anti-MSP7 antibodies at T0 and the absenceof parasitaemia during surveillance. Similarly, no such asso-ciation was observed for antibodies to MSP119, which is fre-quently reported to be positively associated with protectionfrom high parasitaemias and reduced morbidity (16,17).This suggests that the state of immunity in this study popu-lation may be due to a different set of host factors thanthose responsible for controlling parasitaemias or limitingmorbidity. It would be worthwhile examining the antibodyresponses to MSP6 and MSP7 in a different population forwhich these clinical and parasitological data are available.Alternatively, it could be that most individuals in the studypopulation were susceptible to reinfection with malaria, butthat the number of reinfection was limited by a low entomo-logic inoculation rate.
Anti-MSP6 and anti-MSP7 antibodies were found to bemainly of the cytophilic subclasses IgG1 and IgG3, both ofwhich have been reported to have opsonizing and complex-fixing properties (29). It is interesting to note that, of thetwo cytophilic subclasses, IgG1 is the predominant sub-class of antibodies to MSP6 and IgG3 is the predominantsubclass of antibodies to MSP7. This isotype difference isintriguing, given that MSP6 and MSP7 are in the same pro-tein complex. Imbalance of antibody subclass distributionin antibodies to other P. falciparum blood-stage antigens hasbeen reported. For example, antibody responses to MSP2are mainly of IgG3 subclass (25,26) and responses to MSP119
and rhoptry-associated antigen 1 are mainly of the IgG1subclass (15,20,30). Little is known about the parametersthat regulate human isotype responses and the consequencesof such responses in malaria. A study of the subclass distri-bution of antigen-specific IgG in an in vitro culture systemhas suggested that the intrinsic properties of a protein itselfplay a major role (31). Studies of other systems suggest thatthe cytokine milieu may be critical in determining switchpatterns in naive B cells (32). It is possible that differentmalaria antigens induce different cytokine responses, whichmay influence class switching during the development ofanti-malaria immunity. From the available data, it appearsthat the degree of conservation/polymorphism of an antigenis associated with the IgG subclass distribution in antibod-ies to that antigen. For example, in the same set of serumsamples used in this study, we have found that the antibod-ies to MSP2 and MSP4 are predominantly of IgG 3 subclass(18,33), whereas the antibodies to MSP5 are predominantly
of IgG1 subclass (S. Weisman et al., manuscript in prepara-tion). We have seen the same pattern of isotype bias in natur-ally acquired antibodies to MSP1, MSP2, MSP4, MSP5,MSP6 and MSP7 in a genetically distinct human population(Papua New Guinea) (data not shown). MSP119 is highlyconserved, with only four amino acid variations detected inall laboratory strains and field isolates examined so far(34,35); MSP5 is also a highly conserved protein that lackssequence variation between various isolates (36); and exam-ination of MSP6 in three parasite lines has not detected anysequence variation (12). In contrast, MSP2 is a highly poly-morphic antigen that contains a variable central domain ofrepeats flanked by variable non-repetitive regions (33,37);MSP4 is relatively polymorphic, with more than nine vari-ants detected in a study of 20 isolates (38). Examination ofMSP7 in three parasite lines identified a number of variantresidues (13) and, during construction of MSP7 expressingclones for this study, we detected a number of additionalvariant residues. Interestingly, although MSP1, MSP2,MSP4, MSP5, MSP6 and MSP7 are all merozoite surfaceproteins, antibodies raised to these antigens by parasiteinfection are strongly biased to either IgG1 or IgG3. In par-ticular, MSP4 and MSP5 are encoded by two related genesand have similar structural features (39), but they induceantibodies with distinct isotype distribution. It is possiblethat structural and/or functional constraints may limit theextent of sequence variation, which in turn regulate the iso-type responses. This proposal is supported by the differen-tial patterns of IgG subclass response to two distinct regionsof MSP1: responses to the polymorphic block 2 are predom-inantly IgG3, whereas antibodies to the conserved MSP119 aremainly IgG1 (21). The exact relationship between sequencediversity, protein structure and effect on antigenicity of epitopesremains to be determined for many of these proteins.
ACKNOWLEDGEMENTS
This work was supported by grants from the NationalHealth and Medical Research Council (NHM&RC) of Aus-tralia, the National Institutes of Health (NIH), the HowardHughes Medical Institute International Scholars Programand the UNDP/World Bank/WHO Special Programmefor Research and Training in Tropical Diseases. We thankAnthony Holder for supplying the MSP119 expression con-struct and Philip Hodgkin for useful discussions. The fieldcomponent of this study was conducted in 1994 as a collab-oration between the Institute for Malariology, Parasitologyand Entomology, Hanoi, Vietnam, and US Naval MedicalResearch Unit 2, Jakarta, Indonesia. We thank Le DinhCong, Tran Thi Uyen, Nguyen Dieu Thtiong, Luc NguyenTuyen (IMPE) and Stephen F. Wignall and Andrew L.Corwin (NAMRU-2) for supporting this research.

© 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412 411
Volume 25, Number 8/9, August/September 2003 Antibody responses to MSP6 and MSP7
REFERENCES
1 Anders RF & Saul AJ. Candidate antigens for an asexual bloodstage vaccine against falciparum malaria. In: Good MF, Saul AJ,ed. Molecular Immunological Considerations in Malaria VaccineDevelopment. Boca Raton: CRC Press, 1993, pp. 169–208.
2 Hoffman S, Coppel RL & Chulay J. Vaccines. In: Oaks SC,Mitchell VS, Pearson GW & Carpenter CCJ, eds. Malaria:Obstacles and Opportunities. Washington DC: National AcademyPress, 1991, pp. 169–210.
3 Cohen S, McGregor IA & Carrington SC. Gammaglobulin andacquired immunity to human malaria. Nature 1961; 192: 733–737.
4 Sabchareon A, Burnouf T, Ouattara D et al. Parasitologic andclinical human response to immunoglobulin administration infalciparum malaria. Am J Trop Med Hyg 1991; 45: 297–308.
5 Sarthou JL, Angel G, Aribot G et al. Prognostic value of anti-Plasmodium falciparum-specific immunoglobulin G3, cytokines,and their soluble receptors in West African patients with severemalaria. Infect Immun 1997; 65: 3271–3276.
6 O’Donnell RA, de Koning-Ward TF, Burt RA et al. Antibodiesagainst merozoite surface protein (MSP)-1(19) are a majorcomponent of the invasion-inhibitory response in individualsimmune to malaria. J Exp Med 2001; 193: 1403–1412.
7 Blackman MJ, Heidrich HG, Donachie S, MeBride JS &Holder AA. A single fragment of a malaria merozoite surfaceprotein remains on the parasite during red cell invasion and isthe target of invasion-inhibiting antibodies. J Exp Med 1990;172: 379–382.
8 Blackman MJ & Holder AA. Secondary processing of thePlasmodium falciparum merozoite surface protein-1 (MSP1) bya calcium-dependent membrane-bound serine protease: shed-ding of MSP133 as a noncovalently associated complex withother fragments of the MSP1. Mol Biochem Parasitol 1992; 50:307–315.
9 Blackman MJ, Whittle H & Holder AA. Processing of thePlasmodium falciparum major merozoite surface protein-1:identification of a 33-kilodalton secondary processing productwhich is shed prior to erythrocyte invasion. Mol Biochem Para-sitol 1991; 49: 35–44.
10 Stafford YM, Blackman MJ, Harris A, Shai S, Grainger M &Holder AA. N-tenninal amino acid sequence of the Plasmodiumfalciparum merozoite surface protein-1 polypeptides. Mol BiochemParasitol 1994; 66: 157–160.
11 Stafford WH, Gunder B, Harris A, Heidrich HG, Holder AA &Blackinan MJ. A 22-kDa protein associated with the Plasmodiumfalciparum merozoite surface protein-1 complex. Mol BiochemParasitol 1996; 80: 159–169.
12 Trucco C, Fernandez-Reyes D, Howell S, Stafford WH, Scott-Finnigan U, Grainger M et al. The merozoite surface protein 6gene codes for a 36-kDa protein associated with the Plasmodiumfalciparum merozoite surface protein-1 complex. Mol BiochemParasitol 2001; 112: 91–101.
13 Pachebat JA, Ling IT, Grainger M et al. The 22-kDa compo-nent of the protein complex on the surface of Plasmodium falci-parum merozoites is derived from a larger precursor, merozoitesurface protein 7. Mol Biochem Parasitol 2001; 117: 83–89.
14 al-Yaman F, Genton B, Kramer KJ et al. Assessment of the roleof naturally acquired antibody levels to Plasmodium falciparummerozoite surface protein-1 in protecting Papua New Guineanchildren from malaria morbidity. Am J Trop Med Hyg 1996; 54:443–448.
15 Dodoo D, Theander TG, Kurtzlials JA et al. Levels of antibodyto conserved parts of Plasmodium falciparum merozoite surfaceprotein 1 in Ghanaian children are not associated with protec-tion from clinical malaria. Infect Immun 1999; 67: 2131–2137.
16 Riley EM, Allen SJ, Wheeler JG et al. Naturally acquired cellularand humoral immune responses to the major merozoite surfaceantigen (PfMSP1) of Plasmodium falciparum are associated withreduced malaria morbidity. Parasite Immunol 1992; 14: 321–337.
17 Egan AF, Morris J, Barnish G et al. Clinical immunity to Plas-modium falciparum malaria is associated with serum antibodiesto the 19-kDa C-terminal fragment of the merozoite surfaceantigen, PfMSP-1. J Infect Dis 1996; 173: 765–769.
18 Wang L, Richie TL, Stowers A, Nhan DH & Coppel RL.Naturally acquired antibody responses to Plasmodium falciparummerozoite surface protein 4 in a population living in an area ofendemicity in Vietnam. Infect Immun 2001; 69: 4390–4397.
19 Conway DJ, Cavanagh DR, Tanabe K et al. A principal targetof human immunity to malaria identified by molecular popula-tion genetic and immunological analyses. Nat Med 2000; 6:689–692.
20 Egan AF, Chappel JA, Burghaus PA et al. Serum antibodiesfrom malaria-exposed people recognize conserved epitopesformed by the two epidermal growth factor motifs of MSP1(19),the carboxy-terminal fragment of the major merozoite surfaceprotein of Plasmodium falciparum. Infect Immun 1995; 63: 456–466.
21 Cavanagh DR, Dobano C, Elhassan IM et al. Differentialpatterns of human immunoglobulin G subclass responses todistinct regions of a single protein, the merozoite surface protein1 of Plasmodium falciparum. Infect Immun 2001; 69: 1207–1211.
22 Wang L, Black CG, Marshall VM & Coppel RL. Structural andantigenic properties of merozoite surface protein 4 of Plasmo-dium falciparum. Infect Immun 1999; 67: 2193–2200.
23 Burghaus PA & Holder AA. Expression of the 19-kilodaltoncarboxy-terminal fragment of the Plasmodium falciparum mero-zoite surface protein-1 in Escherichia coli as a correctly foldedprotein. Mol Biochem Parasitol 1994; 64: 165–169.
24 Taylor RR, Allen SJ, Greenwood BM & Riley EM. IgG3 anti-bodies to Plasmodium falciparum merozoite surface protein 2(MSP2) – increasing prevalence with age and association withclinical immunity to malaria. Am J Trop Med Hyg 1998; 58:406–413.
25 Taylor RR, Smith DB, Robinson VJ, McBride JS & Riley EM.Human antibody response to Plasmodium falciparum merozoitesurface protein 2 is serogroup specific and predominantly of theimmunoglobulin G3 subclass. Infect Immun 1995; 63: 4382–4388.
26 Rzepczyk CM, Hale K, Woodroffe N et al. Humoral. immuneresponses of Solomon Islanders to the merozoite surface anti-gen 2 of Plasmodium falciparum show pronounced skewingtowards antibodies of the immunoglobulin G3 subclass. InfectImmun 1997; 65: 1098–1100.
27 Oeuvray C, Bouharoun-Tayoun H, Grass-Masse H et al. Anovel merozoite surface antigen of Plasmodium falciparum(MSP-3) identified by cellular-antibody cooperative mechanismantigenicity and biological activity of antibodies. Mem InstOswaldo Cruz, 1994; 89: 77–80.
28 Thomas AW, Trape JF, Rogier C, Goncalves A, Rosario VE &Narum DL. High prevalence of natural antibodies againstPlasmodium falciparum 83-kilodalton apical membraneantigen (PF83/AMA-1) as detected by capture-enzyme-linked

412 © 2003 Blackwell Publishing Ltd, Parasite Immunology, 25, 403–412
L. Wang et al. Parasite Immunology
immunosorbent assay using full-length baculovirus recombinantPF83/AMA-1. Am J Trop Med Hyg 1994; 51: 730–740.
29 Groux H & Gysin J. Opsonization as an effector mechanism inhuman protection against asexual blood stages of Plasmodiumfalciparum: functional role of IgG subclasses. Res Immunol1990; 141: 529–542.
30 Fonjungo PN, Elhassan IM, Cavanagh DR et al. A longitudinalstudy of human antibody responses to Plasmodium falciparumrhoptry-associated protein 1 in a region of seasonal and unstablemalaria transmission. Infect Immun 1999; 67: 2975–2985.
31 Garraud O, Perraut R, Diouf A, Nambei WS, Tall A, Spiegel Aet al. Regulation of antigen-specific immunoglobulin G sub-classes in response to conserved and polymorphic Plasmodiumfalciparum antigens in an in vitro model. Infect Immun 2002; 70:2820–2827.
32 Garraud O, Nutman TB. The roles of cytokines on human B-cell differentiation into immunoglobulin secreting cells. Bull InstPasteur 1996; 94: 285–309.
33 Weisman S, Wang L, Billman-Jacobe H, Nhan DH, Richie TL& Coppel RL. Antibody responses in patients infected withstarins of Plasmodium falciparum expressing diverse forms ofmerozoite surface protein 2. Infect Immun 2001; 69: 959–967.
34 Miller LH, Roberts T, Shahabuddin M & McCutchan TF.Analysis of sequence diversity in the Plasmodium falciparummerozoite surface protein-1 (MSP-1). Mol Blochem Parasitol1993; 59: 1–14.
35 Kang Y & Long CA. Sequence heterogeneity of the C-terminal,cys-rich region of the merozoite surface protein-1 (MSP-1) infield samples of Plasmodium falciparum. Mol Biochem Parasitol1995; 73: 103–110.
36 Wu T, Black CG, Wang L, Hibbs AR & Coppel RL. Lack ofsequence diversity in the gene encoding merozoite surface pro-tein 5 of Plasmodium falciparum. Mol Biochem Parasitol 1999;103: 243–250.
37 Smythe JA, Coppel RL, Day KP et al. Structural diversity in thePlasmodium falciparum merozoite surface antigen MSA-2. ProcNatl Acad Sci USA 1991; 88: 1751–1755.
38 Wang L, Marshall VM & Coppel RL. Limited polymorphismof the vaccine candidate merozoite surface protein 4 ofPlasmodium falciparum. Mol Biochem Parasitol 2002; 20: 301–303.
39 Marshall VM, Wu T & Coppel RL. Close linkage of threemerozoite surface protein genes on chromosome 2 of Plasmodiumfalciparum. Mol Biochem Parasitol 1998; 94: 13–25.
Related Documents











