Nasal Fossa of Mouse and Dwarf Lemurs (Primates, Cheirogaleidae) TIMOTHY D. SMITH 1,2 * AND JAMES B. ROSSIE 3 1 School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania 2 Department of Anthropology, University of Pittsburgh, Pittsburgh, Pennsylvania 3 Department of Anthropology, Stony Brook University, Stony Brook, New York ABSTRACT Dimensions of the external midface in mammals are sometimes related to olfactory abilities (e.g., ‘‘olfactory snouts’’ of strepsirrhine pri- mates). This association hinges on the largely unexplored relationship between the protruding midface and internal topography of the nasal fos- sae. Herein, serially sectioned heads of embryonic to adult cheirogaleid primates (mouse and dwarf lemurs) and a comparative sample were stud- ied. To assess the anteroposterior distribution of olfactory epithelium (OE) within the nasal fossa, the surface area of OE and non-OE was measured in two mouse lemurs (one adult, one infant). Prenatally, ethmo- turbinal projections appear in an anteroposterior sequence. Fetal mouse lemurs, tenrecs, voles, and flying lemurs have four ethmoturbinals that project toward the nasal septum. Major distinctions among these mam- mals include the number of turbinals in recesses and the extent of the ol- factory recess. Surface area measurements in the adult mouse lemur reveal that 31% of the entire nasal fossa is lined with OE. The majority is sequestered in a posterior recess (70% OE). Anterior to this space, only 28% of the nasal fossa is lined with OE. Ethmoturbinal I is lined with relatively less OE (35%) compared with more posterior ethmoturbinals (46–57%). Age comparisons support the idea that OE increases less than non-OE between ages. Regionally, results suggest that most growth in surface area occurs in turbinals. But in all ethmoturbinals, surface area of non-OE differs between ages more than that of OE. This study shows that the anterior part of the nasal fossa is mostly nonolfactory in Microcebus murinus. Anat Rec, 291:895–915, 2008. Ó 2008 Wiley-Liss, Inc. Key words: concha; nasal cavity; olfactory; primate; respiratory; turbinal; turbinate Among all vertebrates, mammals have evolved the nasal cavities with the greatest internal complexity. The lateral nasal wall is complicated by projections called turbinals (turbinates, conchae). These invade the nasal airway space and increase surface area for both olfactory and respiratory mucosae (Negus, 1958; Hillenius, 1992, 1994). In morphology, these projections may vary from simple ridges to highly elaborated scrolls. Anatomists first understood the intricate contours of the nasal fos- sae by the study of hemisected skulls (e.g., Zuckerkandl, 1887; Seydel, 1891; Kollmann and Papin, 1925; Cave, 1973), but coronal sections of the rostrum of mammals provided a far clearer picture, because the most median turbinals often overlay other turbinals and recesses (Negus, 1958). The advent of computed tomography now allows nondestructive access to the nasal cavity and the ability to three-dimensionally reconstruct the internal structures and spaces (Rowe et al., 2005; Rossie, 2006; Craven et al., 2007). Such studies reveal the actual space through which inspired air and volatile chemicals travel (Craven et al., 2007). While the available informa- tion on internal nasal morphological adaptations contin- This article contains Supplementary Material available at http://www.interscience.wiley.com/suppmat/1932-8486/suppmat. *Correspondence to: Timothy D. Smith, School of Physical Therapy, Slippery Rock University, Slippery Rock PA, 16057. Fax: 724-738-2113. E-mail: [email protected] Received 23 January 2008; Accepted 4 April 2008 DOI 10.1002/ar.20724 Published online in Wiley InterScience (www.interscience.wiley. com). Ó 2008 WILEY-LISS, INC. THE ANATOMICAL RECORD 291:895–915 (2008)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nasal Fossa of Mouse and DwarfLemurs (Primates, Cheirogaleidae)
TIMOTHY D. SMITH1,2* AND JAMES B. ROSSIE3
1School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania2Department of Anthropology, University of Pittsburgh, Pittsburgh, Pennsylvania3Department of Anthropology, Stony Brook University, Stony Brook, New York
ABSTRACTDimensions of the external midface in mammals are sometimes
related to olfactory abilities (e.g., ‘‘olfactory snouts’’ of strepsirrhine pri-mates). This association hinges on the largely unexplored relationshipbetween the protruding midface and internal topography of the nasal fos-sae. Herein, serially sectioned heads of embryonic to adult cheirogaleidprimates (mouse and dwarf lemurs) and a comparative sample were stud-ied. To assess the anteroposterior distribution of olfactory epithelium(OE) within the nasal fossa, the surface area of OE and non-OE wasmeasured in two mouse lemurs (one adult, one infant). Prenatally, ethmo-turbinal projections appear in an anteroposterior sequence. Fetal mouselemurs, tenrecs, voles, and flying lemurs have four ethmoturbinals thatproject toward the nasal septum. Major distinctions among these mam-mals include the number of turbinals in recesses and the extent of the ol-factory recess. Surface area measurements in the adult mouse lemurreveal that 31% of the entire nasal fossa is lined with OE. The majorityis sequestered in a posterior recess (70% OE). Anterior to this space, only28% of the nasal fossa is lined with OE. Ethmoturbinal I is lined withrelatively less OE (35%) compared with more posterior ethmoturbinals(46–57%). Age comparisons support the idea that OE increases less thannon-OE between ages. Regionally, results suggest that most growth insurface area occurs in turbinals. But in all ethmoturbinals, surface areaof non-OE differs between ages more than that of OE. This study showsthat the anterior part of the nasal fossa is mostly nonolfactory inMicrocebusmurinus. Anat Rec, 291:895–915, 2008. � 2008Wiley-Liss, Inc.
Key words: concha; nasal cavity; olfactory; primate; respiratory;turbinal; turbinate
Among all vertebrates, mammals have evolved thenasal cavities with the greatest internal complexity. Thelateral nasal wall is complicated by projections calledturbinals (turbinates, conchae). These invade the nasalairway space and increase surface area for both olfactoryand respiratory mucosae (Negus, 1958; Hillenius, 1992,1994). In morphology, these projections may vary fromsimple ridges to highly elaborated scrolls. Anatomistsfirst understood the intricate contours of the nasal fos-sae by the study of hemisected skulls (e.g., Zuckerkandl,1887; Seydel, 1891; Kollmann and Papin, 1925; Cave,1973), but coronal sections of the rostrum of mammalsprovided a far clearer picture, because the most medianturbinals often overlay other turbinals and recesses(Negus, 1958). The advent of computed tomography nowallows nondestructive access to the nasal cavity and the
ability to three-dimensionally reconstruct the internalstructures and spaces (Rowe et al., 2005; Rossie, 2006;Craven et al., 2007). Such studies reveal the actualspace through which inspired air and volatile chemicalstravel (Craven et al., 2007). While the available informa-tion on internal nasal morphological adaptations contin-
This article contains Supplementary Material available athttp://www.interscience.wiley.com/suppmat/1932-8486/suppmat.
*Correspondence to: Timothy D. Smith, School of PhysicalTherapy, Slippery Rock University, Slippery Rock PA, 16057.Fax: 724-738-2113. E-mail: [email protected]
Received 23 January 2008; Accepted 4 April 2008
DOI 10.1002/ar.20724Published online in Wiley InterScience (www.interscience.wiley.com).
� 2008 WILEY-LISS, INC.
THE ANATOMICAL RECORD 291:895–915 (2008)

ues to increase, knowledge of the mucosa that lines thesurfaces is far less detailed for most mammals (Smithet al., 2007a,b).In the mammalian nasal cavity, four types of epithelia
have been said to line the internal boundaries. Theseinclude respiratory, olfactory, stratified, and a moreinconsistent ‘‘transitional’’ epithelium (Adams, 1986).These epithelia, combined with underlying elements ofthe lamina propria that support them, are the basis forphysiological properties of internal structures in thenasal region. Mucosal characteristics of the inner con-tours of the nasal fossae (each comprising half of thenasal cavity) are only known in a generalized mannerfor most mammals. In addition, the terminology usedto describe structures varies greatly. These factors arelimitations in the endeavor to understand functionalspecializations and the evolutionary modifications ofthe nasal fossa in different mammalian taxa, particu-larly in groups with highly diverse nasal morphology(Smith et al., 2004; Smith and Rossie, 2006; Smith et al.,2007a).Detailed mucosal ‘‘maps’’ are available for some small-
bodied mammals (Adams, 1972; Clancy et al., 1994). Thepercentage of mucosa found on individual elements (e.g.,turbinals) has been provided for the opossum (Roweet al., 2005) and some bat species (Bhatnagar andKallen, 1975). For larger bodied mammals, internal dis-tribution of nasal mucosa has been generally described(e.g., goats, horses—Kumar et al., 1993, 2000). Quantita-tive data are available for rabbits, cats, and dogs,although without specific associations between osseousstructures and mucosal coverings (see Negus, 1958;Moulton and Biedler, 1967). In sum, a generalized topo-graphic association of internal nasal structures with epi-thelial types can be gleaned from the literature summar-ized above (and see Moore, 1981, for review). The ante-roinferior portions of the nasal fossa, including turbinalstherein, are lined with nonolfactory epithelium (mostlyof respiratory and stratified types). Turbinals of the eth-moid bone and adjacent surfaces of the midline septumbear olfactory epithelium in all mammals described todate. However, descriptions of diverse taxa (ungulates,carnivores, bats, insectivores, primates, and marsupials)indicate that some ethmoidal turbinals are partially cov-ered by respiratory mucosa (Read, 1908; Negus, 1958;Loo, 1974; Bhatnagar and Kallen, 1975; Kumar et al.,1993, 2000; Rowe et al., 2005; Smith et al., 2007b). Thisvariation in the mucosal distribution along ethmoidalturbinals and adjacent structures requires more detailedinvestigation (Smith et al., 2007a,b).Turbinals and other structures that bear nasal mucosa
vary enormously among mammalian taxa (Paulli, 1900;Moore, 1981). The evolutionary history of the lateralnasal wall is primarily inferred from comparative stud-ies, which would indicate that marsupials have theprimitive condition of five ethmoturbinals (Maier,1993a). Moore (1981) suggests that, in carnivores andungulates, the number of turbinals has increased by asplitting of the more posterior turbinals. However,Moore also questions whether the first ethmoturbinal(endoturbinal II in his terminology) is formed by meansof union of two ‘‘originally quite separate endoturbinals’’(1981, p 250). If certain turbinals have fused during evo-lution of the mammalian lateral nasal wall as suggestedby Paulli (1900) and Moore (1981), using the primary
lamina as criteria for nomenclature may be arbitrary.Due to postnatal elaboration of the ethmoidal labyrinth(Maier, 1993a), comparative studies of adult craniaappear to hold little promise for understanding whethersplitting or fusion occurred to produce the varying con-figurations among mammals. Although the sole use ofdevelopmental data to infer homology can be misleading(Hall, 1994), postnatal transformations of the nasal fossain mammals are extensive and lead to divergent adultmorphologies (Witmer, 1995). Thus, an understanding ofthe process by which the turbinals form during late em-bryonic and fetal stages is essential to clarify whetherethmoturbinals are autonomous elements during devel-opment. A neonatal mammalian ‘‘bauplan’’ has beendescribed by Maier (1993a,b), who described several fe-tal strepsirrhines (1993b). At present, however, detailed,cross-sectional ontogenetic studies of the turbinals arelacking.Here, we begin to address the need for further devel-
opmental and histological data on the primate nasalfossa. Species from the family Cheirogaleidae (mouseand dwarf lemurs) are studied. These primates mayhave ecological and behavioral similarities of the earliestprimates (Cartmill, 1974). Each of these species feedsprimarily in a fine-branch milieu to obtain insect and/orplant products. Furthermore, nocturnal cheirogaleidsrely on olfaction as a crucial means of interspecific com-munication (Perret, 1992), and detection of certain fooditems (Siemers et al., 2007). The present study begins toaddress several uncertain aspects of primate nasal anat-omy. First, ontogenetic transformations of the lateralnasal wall are assessed using a cross-sectional embry-onic/fetal sample of Microcebus. A comparative sampleof fetal nonprimate mammals is also studied to deter-mine whether Microcebus spp. have a similar cartilagi-nous template from which the nasal fossa, particularlythe ethmoturbinal complex, develops. A second focus ofthis study is on the distribution of nasal mucosa andstructural associations in postnatal primates. Dimen-sions of the external midface in mammals are sometimesrelated to olfactory abilities, for example, the so-called‘‘olfactory snouts’’ of strepsirrhine primates (Napier andNapier, 1967). Such an association hinges on the largelyunexplored relationship between the protruding midfaceand internal topography of the nasal fossae. The presentstudy provides a detailed account of mucosal distributionover internal osseous structures in a strepsirrhine pri-mate, Microcebus murinus.
MATERIALS AND METHODSSample
The present study describes development and mucosallining of the nasal fossa in three genera from the familyCheirogaleidae, which are small bodied primates thatpossess moderately prognathic midfaces (Fig. 1a). Seri-ally sectioned heads of mouse lemurs (Microcebus spp.;four adults, one neonate, nine embryos and fetuses) anddwarf lemurs (Cheirogaleus medius: two adults, two neo-nates; Mirza coquereli, two neonates) were studied. Afetal sample of two common tenrecs (Tenrec ecaudatus),one rodent (Microtus ochrogaster), and one dermopterid(Cynocephalus volans) were compared with fetal Micro-cebus to assess similarities and distinctions in the carti-laginous template for the nasal capsule.
896 SMITH AND ROSSIE
Ontogenetic primate samples are challenging toobtain, and therefore most specimens were obtainedopportunistically when neonatal (ages 1 to 2 days) or
adult animals died of natural causes in captivity (Table1). However, it must be mentioned that the precise fertil-ization age of ‘‘neonates’’ remains unknown. Some varia-tions may be due to ontogenetic changes within a peri-natal window. Likewise, precise ages for specimens inthe comparative sample remain unknown, although allwere previously estimated to be of fetal (Bhatnagar andWible, 1994; Shimp et al., 2003) or perinatal ages (Smithet al., 2007c). The vole neonate was previously judged tobe of perinatal size based on its crown-rump length(Smith et al., 2007c).Embryonic and fetal Microcebus specimens of the
Bluntschli collection were studied at the American Mu-seum of Natural History (Lozanoff and Diewert, 1989;Shimp et al., 2003). This collection included data cardswith measures such as ‘‘total overall length,’’ which wasroughly equivalent to crown-rump length (CRL) basedon archived photographs of embryos/fetuses. Thearchived photographs revealed details about limb mor-phology (shape, presence of digital rays, etc.). Based onthis information and internal features the Microcebusembryos were assigned Carnegie stages (O’Rahilly andMuller, 1987; Table 1).The assessment of early nasal capsule maturation
below is mostly based on the sample of Microcebusmyoxinus, which comprises a greater number of embry-onic stages. For comparison of nonprimates to primates,fetal specimens that possessed a completely chondrifiedcapsule were selected to assess similarities and differen-ces in the cartilaginous template or bauplan. In additionto images shown in the results section below, serial sec-tional levels can be viewed in primates and nonprimatesat http://www.interscience.wiley.com/jpages/1932-8486/suppmat.These specimens exist in several collections including
those serially sectioned by the first coauthor and col-leagues (Smith et al., 2007b,c), some of which are nowhoused at the Carnegie Museum of Natural History(Table 1). Selected specimens of the Bluntschli collectionand one specimen from Duke University (Bhatnagar andWible, 1994) were used to describe developmentalaspects. Throughout the text, descriptions of the primatenasal structures apply to all species unless otherwiseindicated. Otherwise, full genus and species names arefrequently specified to avoid confusion at the genuslevel.All specimens were previously paraffin-embedded and
serially sectioned (10 to 40 mm). A variety of stainingprocedures were used, including Azan blue, hematoxy-lin-eosin, Gomori trichrome, Mallory’s trichrome, andAlcian blue-periodic acid-Schiff. The details of pre-paration for serial sectioning were previously published(Lozanoff and Diewert, 1989; Shimp et al., 2003; Smithet al., 2004).
Structural Relations and Terminology of Nose
and Nasal Fossa
Cheirogaleids, like most mammals, possess a moisthairless patch of skin that surrounds the nostrils, therhinarium (Fig. 1a). The rhinarium has been related toprimate chemoreception based on the continuity of amidline rhinarial groove with a passage leading to vomero-nasal organ duct (Schilling, 1970; Martin, 1990; Rossieand Smith, 2007). The anatomy of the external nose and
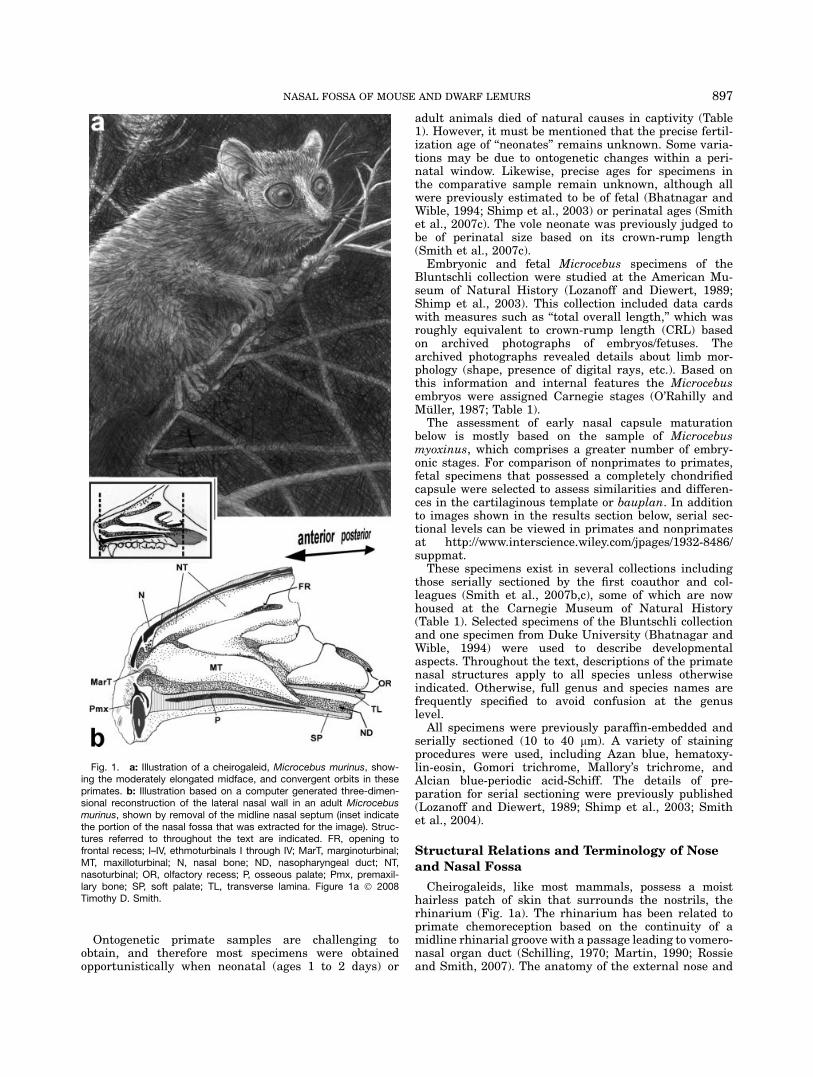
Fig. 1. a: Illustration of a cheirogaleid, Microcebus murinus, show-ing the moderately elongated midface, and convergent orbits in theseprimates. b: Illustration based on a computer generated three-dimen-sional reconstruction of the lateral nasal wall in an adult Microcebusmurinus, shown by removal of the midline nasal septum (inset indicatethe portion of the nasal fossa that was extracted for the image). Struc-tures referred to throughout the text are indicated. FR, opening tofrontal recess; I–IV, ethmoturbinals I through IV; MarT, marginoturbinal;MT, maxilloturbinal; N, nasal bone; ND, nasopharyngeal duct; NT,nasoturbinal; OR, olfactory recess; P, osseous palate; Pmx, premaxil-lary bone; SP, soft palate; TL, transverse lamina. Figure 1a � 2008Timothy D. Smith.
897NASAL FOSSA OF MOUSE AND DWARF LEMURS

the vomeronasal organ in Microcebus has been carefullydescribed in previous publications (Schilling, 1970; Smithet al., 2007a), and will not be considered further.Comparative studies of the turbinals are made diffi-
cult by the numerous schemes of terminology used bydifferent authors. For instance, one system of terminol-ogy identifies the turbinals closest to the septum asendoturbinals, and those positioned more laterally anddeep to the endoturbinals as ectoturbinals (Paulli, 1900;Moore, 1981; or frontoturbinals—Maier, 1993a,b). Otherauthors have dispensed with the term endoturbinal andname the turbinals after the bones from which they pro-ject. These names reflect anatomical articulation inadults: The maxilloturbinal articulates with the maxillaand premaxilla, the nasoturbinal articulates with thenasal bone (and with the more posterior ethmoid to avariable extent), and one or more turbinals arise fromthe ethmoid—the ethmoturbinals. Authors differ in theirtreatment of the nasoturbinal, some referring to it as‘‘endoturbinal I or ethmoturbinal I’’ (Paulli, 1900; Moore,1981; Martin, 1990), whereas other authors consider thenext, more posterior, turbinal as ethmoturbinal I (e.g.,Le Gros Clark, 1959; Maier, 1993a).In part, these differences in terminology vary accord-
ing to fields of study, with human and veterinary anat-omy each possessing its own standardized nomenclature(Warrick and Brookes, 1989; Nomina Anatomica Veteri-naria, 2005). To facilitate a comparison of our findingswith other published reports, Figures 1b and 2 and
Table 2 explicitly assign the terminology used herein tointernal structures of the nose. The major, that is, mostmedially projecting turbinals of the lateral wall of strep-sirrhines present six distinct laminae, schematicallyshown in Figures 1b and 2. In Figure 2, the most dorsaland anterior turbinal (nasoturbinal) is arbitrarilydenoted as ‘‘1,’’ and the remainder are numbered in aposterior direction. The reader is referred to Table 2 fornomenclature relating to these and other elements byvarious authors, and should refer to column 2 of Table 1in regard to the present study. In particular, the readershould note numerous discrepancies exist for elementsof the ethmoid complex (Table 2).
Terminology of Development
We have argued elsewhere in support of a system thatwe believe most accurately reflects the homology ofnasal structures (Rossie, 2003, 2006; Smith and Rossie,2006). Subsequent to the formation of a nasal sac in theembryo, the walls of the nasal fossae first condense asmesenchyme, and then chondrify (Moore, 1981). De Beer(1937) considers this to occur in four regions: the ante-rior cupula, the posterior cupula (also called orbitonasallamina), the paranasal cartilage (which joins the ante-rior and posterior cupulae), and the nasal tectum. Col-lectively, the elements of the lateral wall are referred toas the paries nasi (Fig. 3). The nasal tectum comprisessuperior, laterally projecting processes of the median
TABLE 1. Species, age, size, and source of specimens in the study
Specimen no. Figure(s) Species CRL (mm) stage source
Prenatal/perinatal sampleMiSer. 7 4 Microcebus myoxinus (pygmy mouse lemur) 8 E15 BMiSer. 19 M. myoxinus 9 E16 BMiSer. 1* 4 M. myoxinus 13 E22/23 BMiSer. 24 M. myoxinus 31 F BMiSer. 45* 13 M. myoxinus 32 F BMiSer. 71 Microcebus murinus (gray mouse lemur) 11 E20 BMiSer. 54 5, 6 M. murinus 16 F BMiSer. 55* 6, 8, 12 M. murinus 28.5 F BMiSer. 53 M. murinus 33 F BP886/CM 116661 7, 8, 10, 11 M. murinus 41.4 N DLC/CMP384//CM 116670 Mirza coquereli (Coquerel’s dwarf lemur) 69.4 N DLC/CMP385* 7 Mirza coquereli 60.3 N DLCP623 Cheirogaleus medius (fat-tailed dwarf lemur) 57.2 N DLCP699* 7 C. medius 59.3 N DLCInSer. 20* 12 Tenrec ecaudatus (Madagascar hedgehog) 22 F BInSer. 2* 13 T. ecaudatus 23.5 F BDUCEC804 12 Cynocephalus volans (colugo) 88 F DUVP104 12 Microtus ochrogaster (meadow vole) 25.3 N SRU
Adult sampleSpecimen no. Figure(s) species age sourceP1845 2, 8, 10, 11 Microcebus murinus 4 y DLCP870 M. murinus 7 y DLCP1180/CM 116664 M. murinus 9 y DLC/CMP859/CM 116660 M. murinus DLC/CMP1442 Cheirogaleus medius 5 y DLCCM1 9 C. medius ? CMZ
CRL, crown-rump length; stage: E, Carnegie embryonic stage; F, fetus; N, neonate; source: B, Bluntschli collection; DLC,Duke Lemur Center; CM, Carnegie Museum; CMZ, Cleveland Metroparks zoo DU, Duke University comparative embryol-ogy collection; SRU, Slippery Rock University (collection of first coauthor).*Serial sections (every 10th to 20th section of these specimens may be viewed as movie files at http://www.interscience.wiley.com/jpages/1932-8486/suppmat).
898 SMITH AND ROSSIE

cartilaginous septum and forms the ‘‘roof ’’ of the nasalcapsule. The bilateral paranasal cartilages and orbito-nasal laminae have their own centers of condensation.The tectum, a bilateral projection of the median septalcartilage, is initially continuous with the anterior cupula(together, this is the parietotectal cartilage), but laterforms bridges to the paranasal cartilage and posteriorcupula. As chondrification proceeds, the articulation ofthese centers produces discrete elements. The posterioredge of the anterior cupula overlaps the paranasal carti-
lage internally producing a ridge within the nasalfossa—the semicircular crest. Similarly, the anterioredge of the posterior cupula overlaps the paranasal car-tilages internally. The anterior projecting edge of theposterior cupula forms the first ethmoturbinal. Numer-ous authors consider the next turbinal in the posteriordirection to be the ventral lamina of ethmoturbinal I(Seydel, 1891; Moore, 1981; Maier, 1993a,b; see Table 2for alternate terminologies). The basis for this terminol-ogy is that the two scrolls anchor to a common primarylamina (Dieulafe, 1906; Moore, 1981). More posteriorly,additional ethmoturbinals arise and project mediallyfrom the ventromedial margin of the posterior cupula(de Beer, 1937). Accessory ethmoturbinals arise among(and are hidden by) the ethmoturbinals that reach themedial row. Interturbinals and epiturbinals are exam-ples of these accessory scrolls (see below for defini-tions—Maier, 1993a,b). Conventionally, the entire eth-moturbinal complex of adults is considered to be a deriv-ative of the posterior cupula of the nasal capsule (deBeer, 1937; Smith and Rossie, 2006). The nasoturbinal isconsidered as a distinct element that project anteriorly,but also overlaps the ethmoturbinal complex dorsally (aportion of the adult ethmoid bone forms the posteriorpart of the nasoturbinal—Moore, 1981). The maxillotur-binal forms as a remnant of the inferior part of theparies nasi.Spatially, the cartilaginous wall of the nasal fossae
may be divided into three segments, the pars anterior,pars intermedia, and pars posterior. This is a useful con-struct for discussion because it combines the nasal tec-tum with the lateral elements. Our discussion uses thesedescriptors, but we differ slightly from Moore (1981, p252) who emphasized the use of the posterior projectionof the anterior cupula (crista semicircularis) and the an-terior projection of the posterior cupula (ethmoturbinalI) as ‘‘landmarks’’ separating the three parts. Herein,the three contributions of the paries nasi are empha-sized as defining the tripartite organization (Fig. 3).
Histological Observations and Computerized
Three-Dimensional Reconstructions
Due the small size of the animals and fragile natureof the internal nasal fossa, three-dimensional recon-structions and histological cross-sections were used toascertain the general morphology of the internal nose.An adult and neonatal Microcebus murinus wereselected for computer-based three-dimensional recon-structions and quantification of olfactory mucosa distri-bution. Both exhibited well-preserved epithelia, enablingthe identification of transition points from olfactory tononolfactory mucosa. Observations of Bowman’s glandsin the lamina propria and/or olfactory nerves were madeto initially locate olfactory mucosa. Then, the rows ofround nuclei of olfactory receptor neurons were identi-fied and the transition point to a shorter, ciliated respi-ratory epithelium was identified under high magnifica-tion. The entire contour of the nasal mucosa wasobserved because patches of nonolfactory epitheliumsometimes occurred, interrupting the continuity of olfac-tory mucosa. Examination of adults and neonates wasmade at high magnifications using a Leica photomicro-scope (Leica Microsystems: Wetzlar, Germany) at 3400
Fig. 2. Identification of internal nasal structures and spacesreferred to in the text and Table 2. a: Lateral wall of the nasal fossa ofMicrocebus murinus (septum removed). b: Approximate position ofcross-sectional levels shown in Figures 2c through h (adult Microce-bus murinus). c: Anterior margin of the glandular part of nasoturbinal(1a) and maxilloturbinal (2). d: Section through the anterior part of eth-moturbinal I (3), also showing the anterior limit of the Anterolateralrecess (w) and vomeronasal organ (VNO). e: Section near the anteriorlimit of the orbit (Or) showing the semicircular crest (7). f: Sectionthrough the anterior part of ethmoturbinal II (4), also showing the osse-ous nasoturbinal (1b). The frontomaxillary septum (9) divides the pos-terolateral recess into frontal (x) and maxillary recesses (y) at this level.The frontal recess contains a single frontoturbinal (8) in Microcebusspp (see text). g: Section through the anterior projection of ethmotur-binal III (5), also showing interturbinals (10), the posterior root of eth-moturbinal II, and the posterior, unattached projection of the maxillo-turbinal. Ehtmoturbinal I has fused to the nasoturbinal at this level. h:Section through the olfactory recess (z) which contains ethmoturbinalIV (6). The olfactory recess is separated from the nasopharyngealducts (ND) by the transverse lamina (11). NS, nasal septum; OB, olfac-tory bulb; R, retina. Figures 2a and b are not drawn to scale. Scalebar 5 1 mm.
899NASAL FOSSA OF MOUSE AND DWARF LEMURS

to 3630. Fetal specimens were studied using a Microstarcompound microscope (American Optical).To acquire sections for reconstruction, the serial sec-
tions of each nasal chamber were digitally photographedusing a Leica stereomicroscope at low magnification.Images were transferred to Adobe Photoshop 7.0 andsaved as JPEG files. To reconstruct the nasal fossa oftwo of the fetal specimens from the Bluntschli collection,images were acquired using a digital camera with itslens held against the eyepiece of a Brock monocularmicroscope (Brock Optical, Sarasota, FL). Subsequently,individual files for every 20th (in adults) or 10th section
(in infants) were marked according to fiducial landmarks(specific structures or contours) that delimit the olfac-tory mucosa. At the anterior and posterior ends, inter-vening sections were also marked in adults so that accu-racy was comparable to that used for infants. Themarked digital images were then aligned for reconstruc-tion as described in detail by Smith et al. (2004).Reconstructions of the nasal wall were made using
Scion Image software (release 4.02, NIH). Four speci-mens of M. murinus (two fetal, one neonatal, and oneadult animal) were selected to assess age relatedchanges in the contours of the nasal fossa. To illustrate
TABLE 2. Terminology for structures and spaces of the nasal fossa
Reference number
Terminology
(see Figure 2)Structure name
(this study) Humana Other synonyms
Structures1 nasoturbinal (entire) see below concha nasalis dorsalis (NAVb); endoturbinal I
(Moore, 1981, Fig. 84)1a nasoturbinal,
mucosal partagar nasi pars rostralis (NAV); nasoturbinal (Moore, 1981—p
49); nasoturbinate, anterior crus/cardinal conchaIA (Pedziwiatr; 1972); agar nasi region ofnasoturbinal (Kollmann and Papin, 1925)
1b nasoturbinal,osseous part
semicircular crestand uncinate processare remnants
pars caudalis (NAV); ethmoturbinal I (Martin,1990); nasoturbinate, posterior crus/cardinalconcha IB (Pedziwiatr, 1972); endoturbinal I(Paulli, 1990); nasoturbinal, region of uncinateprocess (Kollmann and Papin, 1925)
2 maxilloturbinal inferior nasal concha concha ventralis (NAV) ; maxilloturbinate; ventralconcha
3 ethmoturbinal I middle nasal concha concha media (NAV); ethmoturbinal II (Martin,1990); endoturbinal II, upper lamella (Paulli,1990; Moore, 1981); cardinal concha II(Pedziwiatr, 1972); endoturbinal I (Allen, 1882)
4 ethmoturbinal II superior nasal concha concha ethmoidalis(NAV); ethmoturbinal III(Martin, 1990); endoturbinal II, lower lamella(Paulli, 1990; Moore, 1981); cardinal concha III(Pedziwiatr, 1972)
5 ethmoturbinal III ? concha ethmoidalis(NAV); ethmoturbinal IV(Martin, 1990); endoturbinal II (Paulli, 1990;Moore, 1981); cardinal concha IV (Pedziwiatr,1972)
6 ethmoturbinal IV ? concha ethmoidalis(NAV); ethmoturbinal V(Martin, 1990); endoturbinal III (Moore, 1981);cardinal concha V (Pedziwiatr, 1972)
7 semicircular crest same continuous with, and part of, structure 1b8 frontoturbinal ethmoid bulla concha frontalis (Pedziwiatr, 1972); ectoturbinal
(Moore, 1981)9 frontomaxillary
septum– lateral root of ethmoturbinal I (Rossie, 2006);
anterior root of ethmoturbinal I (de Beer, 1937);horizontal lamina (Maier, 1993a)
10 interturbinal (afterMaier, 1993b)
ectoturbinal (Paulli, 1900; Moore, 1981); accessoryturbinal (Dieleux, 1906)
11 transverse lamina ossiculum Bertini? lamina terminalis (Kollmann and Papin, 1925;Hill, 1953)
Spacesw lateral recess recessus anterior (de Beer, 1937); maxillary sinus
(Rowe et al., 2005, see Fig. 5 therein)x frontal recess superior maxillary recess (Negus, 1958, p 313)y maxillary recess inferior maxillary recess (Negus, 1958, p 313)z olfactory recess sphenoethmoidal recess ethmoturbinal recess
(Maier, 1993a); cupular recess (van Gilse, 1927);sphenoidal recess (Loo, 1973)
aSee Moore (1981) and Smith and Rossie (2006) for further discussion.bNAV (Nomina Anatomica Veterinaria).
900 SMITH AND ROSSIE

the lateral nasal wall, the septum was manually erasedthroughout the sequence of images using Scion Image.In one fetal specimen, deeper recesses were examined byerasing the anterior projection of ethmoturbinal I.Four reconstructions of the neonatal and adult speci-
men were made. First, the entire nasal fossa and thenasal septum were reconstructed as two separate views.Next, only the olfactory portions of each were recon-structed using the same rotation coordinates in ScionImage. Images generated from these reconstructionswere saved and then superimposed using Adobe Photo-shop software to identify mucosal transitions for picto-rial comparisons (see Smith et al., 2007b, or Rehorekand Smith, 2007, for more details). Next, selected recon-structed images were traced and rendered by hand toclearly emphasize contours while excluding artifactualimperfections of the sectioned tissues (e.g., folds ortears) that distort parts of the image.
Mucosal ‘‘Mapping’’ and Quantitative Methods
To obtain a measurement of olfactory SA in the neona-tal and adult M. murinus, the perimeter of olfactory epi-thelium was measured in each section after calibratingto a digital image of a stage micrometer photographed atthe same magnification. The initial cross-sectional levelused for perimeter measurements was the first sectionin which a completely enclosed nasal fossa was found.
For the posterior end-point, the last section containingthe olfactory recess was used. Below the olfactory recess,the nasopharyngeal meatus continues as bilateral naso-pharyngeal ducts for a variable distance in primates(Smith and Rossie, 2006). These ducts are at least par-tially separated by mucosa alone. Every perimeter meas-ured was recorded in millimeters, and multiplied by thedistance in millimeters to the next section, yielding asegment of olfactory surface area between sections. Allsegmental measurements were then summed andrecorded as total olfactory surface area. The same pro-cess was repeated on the entire perimeter of the nasalfossa to obtain total surface area. Olfactory surface areawas subtracted from total surface area to obtain nonol-factory surface area.
RESULTS
General Structure of the Internal Nose
Three-dimensional reconstructions of the internal to-pography in adults reveal that the internal nose is mostcomplex along the lateral wall, where six distinct muco-sal folds or bulges project to a position adjacent to themidline septum (Figs. 1b, 2a–h). The internal nasal skel-eton passes between the large, well-separated orbits(Fig. 2e–h). In combination, Figure 2 and Table 2 offer adetailed, illustrated list of the terminology used in thisstudy. Only a brief summary of this morphology follows,because some of this anatomy has been previouslyreported in osteological descriptions (Kollmann andPapin, 1925; Hill, 1953; Cave, 1973; Ankel-Simons, 2007).On the medial side of each nasal fossa is the nasal
septum, nearly vertical in its contour except for smallbudges inferiorly in the anterior half (Figs. 2c–1e), corre-sponding to the vomeronasal complex. Also, there is amore posterior projection from the septum, supported bya portion of the vomer. At this level, the vomer projectslaterally to join a medially projecting shelf of the eth-moid bone (Fig. 2h). This is the transverse lamina,which divides a solely respiratory pathway (the naso-pharyngeal duct) from the more dorsal space (olfactoryrecess, in which olfactory mucosa predominates) (Fig.2h; see below). The lateral wall is much more irregular.The most medially projecting elements comprise themaxilloturbinal, nasoturbinal, and four scrolls from theethmoid complex: the ethmoturbinals. The nasoturbinalis not a continuously pronounced ridge from its anteriorto posterior extent (see Fig. 2c–f). However, at both ante-rior and posterior ends, this turbinal corresponds to thelateral extent of the cartilaginous nasal tectum, whichretreats from the nasoturbinal fold anteriorly and ossi-fies posteriorly in the adult (see below for related devel-opmental observations).
Prenatal Development of the Internal Nose
Mesenchymal condensations are the earliest signs ofthe cartilaginous capsule, as shown in the smallest em-bryonic specimen examined (8 mm CRL Microcebusmyoxinus; Fig. 4). The paries nasi of the capsule are con-tinuous with the tectum anteriorly, but they are discon-tinuous posteriorly. A separation first appears at the an-terior limit of the primary choanae, where the pariesnasi is separated from the tectum (Fig. 4a). The tectumitself is continuous with the median condensation for
Fig. 3. Schematic of the organization of the amniote nasal capsule(based on de Beer, 1937), emphasizing the tripartite organizationbased on three regions of the lateral wall (paries nasi). a: Three mes-enchymal condensations contribute to the paries nasi of amniotes: theparietotectal cartilage (= pars anterior, but note tectum is removed),the paranasal cartilages (PnC), and the orbitonasal lamina (OnL).These define the pars anterior, pars intermedia, and pars posterior,respectively. b: During fetal morphogenesis, these segments overlapeach other as indicated. The semicircular crest (SCr) extends posteriorlyand ethmoturbinal I (I) extends anteriorly, each overlapping the parsintermedia. NsC, nasal septal cartilage; II–IV, ethmoturbinals II to IV.
901NASAL FOSSA OF MOUSE AND DWARF LEMURS

the nasal septal cartilage until the level of the parsintermedia (this position is marked by the presence ofthe primitive frontal recess [Fig. 4b]). From this pointposteriorly, there is no distinct condensation of the tec-tum, although the median septal (mesethmoid) conden-sation continues (Fig. 4b,c). Condensations for the parsposterior are indistinct in the 8-mm CRL Microcebus
myoxinus (Fig. 4c), but have commenced in the 9-mmCRL specimen (MiSer. 19). In both 8-mm and 9-mmCRL M. myoxinus, some lateral bridging exists betweenthe pars anterior and pars intermedia. In a larger speci-men (13 mm CRL M. myoxinus, Fig. 4d–f) chondrifica-tion is complete in the nasal septum, paries nasi, andthe nasal tectum. Lesser-developed elements are foundinternally, where the semicircular crest and anterior tipof ethmoturbinal I are represented by condensed mesen-chyme (Fig. 4d). A cartilaginous nasal tectum and pariesnasi are continuous from the level of the pars anterior(not shown) to the intersection of pars intermedia andpars posterior (Fig. 4e). Ethmoturbinals II through IVare incompletely developed. Ethmoturbinal II is repre-sented by a mucosal fold supported by a moderately pro-nounced medial bulge in the paries nasi (Fig. 4e). Eth-moturbinal III exists as condensed mesenchyme in thisspecimen (Fig. 4f), and no trace of Ethmoturbinal IV isobservable.Figure 5 shows a three-dimensional reconstruction
of the nasal capsule and surrounding structures in a16-mm CRL Microcebus murinus. The pars anterior,pars intermedia, and pars posterior, are difficult todelineate externally, except that the pars intermedia isan external bulge of the capsule (Fig. 5a–d). The parsposterior is directly adjacent to the orbits (Fig. 5a). Fig-ure 5e emphasizes the close relationship of the ethmo-turbinals of the pars posterior to the cribriform plate.Divisions between the portions of the tripartite cap-
sule are best visualized internally. Figure 6a displays atransverse view of the nasal capsule in the same fetusas in Figure 5. The capsule appears to be in a moreadvanced state of maturation compared with the speci-men described in Figure 4d–f, because the semicircularcrest and ethmoturbinals II and III have fully cartilagi-nous projections within the mucosal folds. In this speci-men, ethmoturbinal IV is visible, although only as a mu-cosal bulge. Figure 6a also illustrates the intersection ofthe three anteroposterior ‘‘segments’’ of the capsule. Thepars anterior folds inwardly relative to the pars interme-dia, and projects posteriorly as the semicircular crest.Similarly, the pars posterior folds inward relative to thepars intermedia, and projects anteriorly as ethmoturbi-nal I. Note that the semicircular crest and the anteriorfree margin of ethmoturbinal I form the medial wall ofthe lateral recess (Fig. 6a). Larger fetuses, such as the28.5-mm CRL M. murinus, possess a full set of turbinals(Fig. 6b), which all have internal cartilaginous support(Fig. 7a–d). Figure 6b is a drawing based on a computerthree-dimensional reconstruction of the lateral wall ofthis specimen, after the anterior projection of ethmotur-binal I was digitally ‘‘dissected away’’ using Scion Imagesoftware. The intersection of the three parts of the nasalcapsule is mostly revealed. The semicircular crestobscures the anterior limit of the anterolateral recess(i.e., the portion of the lateral recess bordered mediallyby overlap with the pars anterior). More posteriorly inthe lateral recess, the cut edge of the frontomaxillaryseptum (‘‘lateral root’’ of ethmoturbinal I) is seen. Thishorizontal shelf effectively divides the lateral recessinto a frontal recess (superiorly) and maxillary recess(inferiorly). Here, this portion of the lateral recess istermed the posterolateral recess (i.e., the portion of thelateral recess bordered medially by overlap with thepars posterior).
Fig. 4. Embryonic nasal capsules in Microcebus myoxinus at 8-mm CRL (a–c) and 13-mm CRL (d–f), emphasizing the least differenti-ated parts of the capsule in each. Insets in 4a–c identify positions. Inthe 8-mm CRL embryo, the pars anterior is continuous with the sep-tum anteriorly (not shown). a: At the posterior limit of the pars anterior,mesenchyme is mostly loosely organized (light gray) with discontinu-ous condensations visible (dark gray) for the pars anterior (Pa) andseptal cartilage (S). b: The primordial frontal recess (FR) is visible atthe level of the pars intermedia (Pi) which is positioned further laterallyrelative to the pars anterior. c: More posteriorly, the pars intermedia isless distinct (right side of figure) and no distinct condensation for thepars posterior is seen. d: The pars anterior is nearly fully chondrifiedin the 13-mm CRL specimen, except for its posterior projection, thesemicircular crest (sc), which is condensed mesenchyme at its ventrallimits. The free anterior margin of ethmoturbinal I (I) is a mesenchymalcondensation at this level. e: A section near the dorsal root of ethmo-turbinal I. Here, the pars posterior is fused to the medial face of thepars intermedia, as evidenced by small bulges for incipient ethmotur-binals I and II. The tectum (T) is continuous up to this level. The parsposterior (Pp) are separated from the nasal septum posteriorly and arefully chondrified laterally. On the medial side ethmoturbinal III is amesenchymal condensation. E, epithelium; MT, maxillorturbinal; ND,nasopharyngeal duct; NT, nasoturbinal; OR, olfactory recess. Scalebars 5 200 mm in a–c, (insets 0.5 mm), 500 mm in d–f.
902 SMITH AND ROSSIE

Morphology of the Nasal Capsule in Three
Neonatal Cheirogaleids
Neonates of three cheirogaleids (Microcebus murinus,Mirza coquereli, and Cheirogaleus medius) are similaranteriorly, throughout the pars anterior (not shown).The anterolateral recess (the portion of the lateral recessbordered medially by overlap with the pars anterior)similarly has a thick glandular lamina propria ventrallyin all species (Fig. 7a,e,f). A major distinction amongspecies is the number of frontoturbinals in the frontalrecess. Microcebus murinus has one, whereas Mirzacoquereli and C. medius each have two (Fig, 7b,f,i).These differences are also seen in comparisons of adultMicrocebus murinus and C. medius. Frontoturbinal 1,the nasoturbinal, and ethmoturbinal I are all more ro-bust in neonates of Mirza coquereli and C. medius com-
pared with Microcebus murinus. In the neonatal Mirzacoquereli, there is a third, dorsally positioned bulge inthe mucosa and underlying cartilage, suggestive of a ru-dimentary frontoturbinal. All three species are similarthroughout the pars posterior (Figs. 7c,d,g,h,k,l).
Prenatal to Postnatal Transformations of the
Lateral Nasal Wall
Figure 8 shows renderings drawn from three-dimen-sional computer reconstructions of the right side of thenasal fossa in a fetal to adult series of Microcebus muri-nus. Anteriorly, reconstructions begin at the level ofthe marginoturbinal. Posteriorly, the reconstructionsend just anterior to the end of the olfactory recess. The
Fig. 6. a: Horizontal plane through the cartilaginous nasal capsuleof the specimen shown in Figure 5, identifying mucosal/cartilaginousfolds of the nasal fossa at the early fetal period. The tripartite organi-zation of the nasal capsule is emphasized on the left side, where whitelines are drawn between the pars anterior (PA), pars intermedia (PI)and pars posterior (PP). Mucosal folds for the semicircular crest (SC),frontoturbinal (ft), and ethmoturbinals I–IV are shown, although not allhave internal cartilaginous support. FR, frontal recess; NS, nasal sep-tum; PS, presphenoid. b: View of the lateral nasal wall in a 28.5 CRLMicrocebus murinus, illustrating details of the overlapping three partsof the nasal capsule at a later fetal stage. The free anterior protectionsof ethmoturbinals I and II have been ‘‘dissected away’’ to illustratedeeper elements and spaces. ft, frontoturbinal; FR, frontal recess; FS,frontomaxillary septum; MR, maxillary recess; I–IV, ethmoturbinals Ithrough IV; MarT, marginoturbinal; MT, maxilloturbinal; N, nasal bone;ND, nasopharyngeal duct; NT, nasoturbinal; OR, olfactory recess; TL,transverse lamina.
Fig. 5. Computer generated three-dimensional reconstructions ofthe nasal capsule in a 16-mm CRL Microcebus murinus. a: Ventralview illustrating the position of the eyes relative the nasal capsule. Anelement of the nasal floor (solum nasi), the vomeronasal cartilage(VNC), is shown. On the right side of the figure the internal nasal mu-cosa is highlighted. b–d Rotating views of the isolated nasal capsule,the illustrate the pars anterior (PA), pars intermedia (PI, darkened foremphasis), and pars posterior (PP) in ventrolateral (b), dorsolateral (c),and dorsal (d) perspectives. e: The mucosa in d is stripped away onthe right side to emphasize three cartilaginous elements for ethmotur-binals I through III (ethmoturbinal IV is only a mucosal fold. Also, eshows an oblique ventral view with the septum and one side of thenasal cavity removed. The position of ethmoturbinals I through IV rela-tive to the cribriform plate (CP) is indicated. AT, auditory tube; MT,maxilloturbinal, NT, nasoturbinal, OC, otic capsule.
903NASAL FOSSA OF MOUSE AND DWARF LEMURS

median nasal septum is removed to reveal the medialsurface of the lateral wall. Figure 8a shows a reconstruc-tion of a 28.5-mm CRL fetus. In this specimen, all eth-moturbinals are supported internally by cartilage (Fig.7a–d). The nasoturbinal is a pronounced ridge anteriorlyand posteriorly, but is poorly defined in the region of theanterolateral recess (cf. Fig. 12a). Here, the semicircularcrest continues into the posterior part of the nasoturbi-nal. The maxilloturbinal is largest anteriorly, and tapersto a small ridge at the level of ethmoturbinal II. Thefrontal recess is small, with a single prominent fronto-turbinal. All four ethmoturbinals project anteriorly fromtheir ventral attachments to the lateral nasal wall. Eth-
moturbinal IV is proportionally enlarged relative to thesize of the same turbinal in the 16-mm CRL fetus (cf.Fig. 5e), and is the sole visible structure in the olfactoryrecess (demarcated by the floor of this space, the trans-verse lamina, Fig. 8a). Figure 8b shows the same viewin a neonatal Microcebus murinus. The most notable dis-tinction of this specimen compared with the smaller fe-tus is the proportional elongation of ethmoturbinal I. Inthe adult nasal fossa (Fig. 8c; 4-year-old M. murinus),the anterior third of the nasal fossa is spatially packed
Fig. 7. Variations in the developing ethmoidal region of neonatalcheirogaleids. a–l: Neonates of Microcebus murinus (a–d), Mirzacoquereli (e–h), and Cheirogaleus medius (i–l) are similar up to thelevel of the anterolateral recess (AR, top row) and in more posteriorregions of the ethmoturbinal complex (see third and last rows). Amajor distinction among species was the number of frontoturbinals inthe frontal recess. Microcebus murinus has one (b), whereas Mirzacoquereli and C. medius each have two (f,j). In Mirza coquereli, thereis an additional, dorsally positioned bulge in the mucosa and underly-ing cartilage, suggestive of a rudimentary frontoturbinal. f1, frontoturbi-nal 1; f2, frontoturbinal 2; I–IV, ethmoturbinals I through IV; ltp, laminatransversalis posterior; MT, maxilloturbinal; ND, nasopharyngeal duct;NT, nasoturbinal; OR, olfactory recess; SC, semicircular crest; TL,transverse lamina. Scales bars 5 1 mm in a (applies to all images).
Fig. 8. a–c: Fetal to postnatal morphogenesis of the lateral nasalwall in fetal (a, 28.5 mm CRL), neonatal (b), and adult (c) Microcebusmurinus. ft, frontoturbinal. FR, frontal recess; I–IV, ethmoturbinals Ithrough IV; MarT, marginoturbinal; MT, maxilloturbinal; ND, nasopha-ryngeal duct; NT, nasoturbinal; OR, olfactory recess; SC, semicircularcrest; TL, transverse lamina.
904 SMITH AND ROSSIE

with proportionally larger turbinals. The maxilloturbinaland ethmoturbinal I are the major elements in thisregion, while the nasoturbinal remains a small ridge.Relative to the neonatal specimen (Fig. 8b), all ethmo-turbinals are proportionally larger on their anteriormargin, but ethmoturbinals I and IV appear to be themost enlarged. In the adult, ethmoturbinal I projects tooverlap the majority of the maxilloturbinal and alsoobscures all but a small aperture of the frontal recess.Ethmoturbinal IV is elongated in both anterior and pos-terior directions from its dorsal attachment. Relative tofetal and neonatal specimens, the adult maxilloturbinalis enlarged dorsally, to lie closely adjacent to the naso-turbinal and ethmoturbinal I. The maxilloturbinal isalso enlarged posteriorly relative to younger specimens.Note that this turbinal is proportionally elongated tooverlap at least part of all turbinals. In part, this isrelated to a free posterior projection, which tapers to-ward the nasopharyngeal duct (cf. Fig. 2g).
Distribution of Olfactory and Nonolfactory
Mucosae Along the Walls of the Nasal Fossae
Figures 9 to 11 provide a pictorial summary of the dis-tribution of mucosa along the walls of the nasal fossa inC. medius and Microcebus murinus. At the level of themarginoturbinal (Fig. 9a, adult C. medius), mostlystratified epithelia is found but small patches of ciliatedepithelium are found associated with subepithelialglands (Fig. 9b,c). The marginoturbinal is continuousposteriorly with the maxilloturbinal (Fig. 9d). Septaland lateral nasal gland masses are associated with thenasal septum and nasoturbinal, respectively (Fig. 9d).These glands are PAS1/Alcian blue2. At this level(approximately 1.5 mm posterior to Fig. 9a), respiratoryepithelium (ciliated, pseudostratified, or simple colum-nar) covers most of the septum, lateral nasal wall, andportions of the maxilloturbinal. Alcian blue1 goblet cellsare numerous in this epithelium (Fig. 9d). Aside from
Fig. 9. Mucosal characteristics of the nasal fossa in an adult Cheir-ogaleus medius. a–c: At the level of the marginoturbinal (a, MarT), theepithelium is mostly nonciliated simple or stratified epithelium, butsmall patches of respiratory epithelium (cilia, open arrows) can befound lining the masses of the lateral nasal gland (b) or septal glands(c). d: A section at the anterior limit of the maxilloturbinal (MT), show-ing PAS1/alcian blue-negative glands in the lamina propria. Alcianblue 1 glands line the lateral and septal walls of the nasal fossa aswell as most of the concave surface of the MT (insets). e,f: Olfactorymucosa projects most anteriorly along the roof or septum and alongthe dorsal rim of ethmoturbinal I (e), as indicated in f by olfactory epi-thelium (OE) and Bowman’s glands (BG). g,h: The anterolateral recess(AL) is surrounded by a thick, glandular lamina propria, with a ciliated
glandular surface membrane h: enlargement of boxed area in g, show-ing goblet cells (arrows). h,i: The maxilloturbinal has ciliated glandularepithelium on its concave aspect, and a vascular, nonciliated epithe-lium on its convex side (i; vs, venous sinuses). In the posterior half ofthe nasal fossa, proportionally more olfactory mucosa is seen. j,k Theunenclosed and enclosed frontal recess (FR). l: The olfactory recess.Mucosal details are provide for each in m–o. Note the nonolfactory lin-ing on the ventral side of ethmoturbinal IV, where a thin ciliated epithe-lium (CE) is found (o). FS, frontomaxillary septum; f1, frontoturbinal 1;f2, frontoturbinal 2;MR, maxillary recess; NT, nasoturbinal; NLD, naso-lacrimal duct; NPD, nasopalatine duct; NS, nasal septum; OB, olfac-tory bulb; ON, olfactory nerves. Scales bars 5 300 mm in a,d,e,g,j,k,l,30 mm in b,c,f,h,i,m,n,o and insets for d.
905NASAL FOSSA OF MOUSE AND DWARF LEMURS

the floor of the nasal fossa (not shown), which is mostlycovered with stratified epithelium, respiratory mucosacovers most of the anterior third of the nasal fossa.Olfactory epithelium first appears as small patches
along the roof and/or septum of the nasal fossa and thedorsal rim of ethmoturbinal I (Fig. 9e,f; at the level ofthe anterolateral recess). The patch along the roof orseptum extends somewhat more anteriorly than the ol-factory mucosa of ethmoturbinal I, especially at birth(Figs. 10, 11, Microcebus murinus). The first 0.5 to 0.8mm of the anterior projection is entirely nonolfactory inadult Microcebus murinus.The anterolateral recess is mostly filled with glandu-
lar lamina propria, covered with ciliated epithelium withgoblet cells (Fig. 9g,h). A small patch of olfactory epithe-lium extends into the apex of the anterolateral recess,just anterior to the beginning of the frontal recess. Theproportionally extensive anterior distribution of res-piratory mucosa on ethmoturbinal I was previouslydescribed in cheirogaleids (Smith et al., 2007b). The ad-jacent maxilloturbinal is only partly ciliated. Generally,the convex surface of the scroll is nonciliated and highlyvascular, whereas it concave aspects are covered withciliated epithelium with goblet cells (Fig. 9i).
In the posterior half of the nasal fossa, olfactory mu-cosa covers the majority of surfaces of all turbinalsexcluding the maxilloturbinal (Figs. 9j–o; 10a,b). Thefrontal recess of C. medius houses two frontoturbinals(Fig. 9j,k). A portion of all frontoturbinals and ethmotur-binals is lined with at least some nonolfactory mucosa,even within the olfactory recess (Figs. 9l,o).
Olfactory Mucosa Distribution
Table 3 and Figures 10 and 11 provide quantitativeand pictorial accounts, respectively, of the olfactory ver-sus nonolfactory mucosa that lines the nasal fossae inone neonatal (Figs. 10a, 11a) and one adult (Figs. 10b,11b) Microcebus murinus. Calculations of surface area(SA) of olfactory and nonolfactory mucosae, as well asthree-dimensional reconstructions indicate that thenasal fossa is lined with proportionately more olfactorymucosa in the neonatal specimen (38%) compared withthe adult (31%). This difference is more evident in thenasopharyngeal chamber, where the neonatal nasalfossa had approximately 41% olfactory SA, compared the25% in the adult. Based on three-dimensional recon-structions, half or more of the nasal septal mucosa is
Fig. 10. a,b: Mucosal ‘‘maps’’ of the lateral nasal wall in neonatal(a) and adult (b) Microcebus murinus. The turbinals are all emphasizedby shades of gray. Unshaded surfaces or lightly shaded surfaces arenonolfactory. Olfactory parts of the turbinals are emphasized by darkgray. ft, frontoturbinal; FR, frontal recess; I–IV, ethmoturbinals I throughIV; MarT, marginoturbinal; MT, maxilloturbinal; ND, nasopharyngealduct; NT, nasoturbinal; OR, olfactory recess; TL, transverse lamina.
Fig. 11. a,b: Mucosal ‘‘maps’’ of the nasal septum wall in neonatal(a) and adult (b) Microcebus murinus. Unshaded surfaces are nonol-factory. Olfactory parts are shaded. N, nasal bone; ND, nasopharyn-geal duct; NPD, nasopalatine duct; OR, olfactory recess; Pmx, pre-maxilla; SC, septal cartilage; TL, cut edge of transverse lamina; VNC,bulge indicating vomeronasal cartilage.
906 SMITH AND ROSSIE

olfactory. Passing toward the olfactory recess, the extentof olfactory mucosa is greater, ending at the level wherethe vomer projects its bilateral choanal processes(Fig. 11).Olfactory mucosa covers dorsoposterior portions of the
nasoturbinal and ethmoturbinals. The nasoturbinal wasnot quantified separately because it is indistinct for aportion of its anterior half (see Fig. 2e). Three-dimen-sional reconstructions revealed that the most anteriorglandular portion is entirely covered with nonolfactorymucosa, whereas the part posterior to the semicircularcrest is entirely olfactory (Fig. 10). All turbinals possesssome nonolfactory mucosa and a greater proportion ofnonolfactory SA is found in adults (Fig. 10b). Quantita-tive results support this observation (Table 3). Moreover,the maxilloturbinal has the greatest overall adult/neona-tal ratio in SA of the turbinals outside the olfactoryrecess (Table 3).The posterior part of the upper nasal fossa is sepa-
rated from the nasopharyngeal ducts by the transverselamina (posterior 25% of the nasal fossa in neonate andposterior 22% in adult Microcebus murinus); a partialseparation is created anteriorly by the choanal processof the vomer. Only ethmoturbinal IV is completelyenclosed within the olfactory recess. This turbinal hasproportionally the greatest amount of olfactory SA atboth ages and has the greatest overall adult/neonatal ra-tio in SA of all ethmoturbinals (Table 3).The posterolateral recess has olfactory mucosa in the
enclosed frontal recess, and more anteriorly before thefrontomaxillary septum attaches (not shown; cf. Fig.9j,k). The percentage of olfactory SA in the entire lateralrecess is somewhat lower in the adult (43%) comparedwith the neonatal Microcebus murinus (48%). The singlefrontoturbinal of the Microcebus murinus has an adult/neonatal ratio in SA that exceeds ethmoturbinals IIand III.These SA data include some surfaces that would not
be discernable in osseous specimens, such as the firstenclosed cross-sections of the nasal fossa as well as theposterior-most cross sections of the nasopharyngealducts. Each of these regions has some borders formedby soft tissue alone. To generate results that couldbe applied to osseous specimens, surface areas werealso calculated in the adult Microcebus murinus usingosseous landmarks. For this measurement, the startpoint of the nasal fossa is the first section in which theright nasal passageway is completely enclosed by bone.The end point is the last section in which the nasopha-ryngeal ducts are completely divided by the vomer.Using these landmarks, the total mucosal SA of thenasal fossa is 362.1 mm2, of which 113.8 mm2 (31%) isolfactory mucosa.
Comparative Morphology of the Internal Nose
During Late Fetal/Neonatal Development
A comparison of sectioned material from late fetal/neo-natal mammals is provided in Figure 12, including coro-nal sections of a Microcebus murinus (Fig. 12a–e), Cyno-cephalus volans (Fig. 12f–j), Tenrec ecaudatus (Fig. 11k–o), and Microtus ochrogaster (Fig. 12p–t). A basic simi-larity exists in organization of the ethmoturbinal com-plex among these species. Specifically, ethmoturbinal I
projects anteriorly to overlap the pars intermedia, andposteriorly three additional cartilaginous elements (eth-moturbinals II–IV) project to a position adjacent to thenasal septum. In contrast to ethmoturbinal I, which rep-resents an anterior projection of the orbitonasal laminaitself, the posterior three elements each have an attach-ment to the ventral margin of the pars posterior. Ethmo-turbinals III and IV also have more posterior attach-ments to the roof of the capsule in all species, and eth-moturbinal II has a mucosal root near the intersectionof the nasal tectum and ethmoturbinal I. EthmoturbinalI shows subtle variations in contour (Fig. 12b,g,j,q). Allspecies have two frontoturbinals except Microcebus spp.,which possesses a single scroll within the frontal recess(Fig. 12a–c). All possess interturbinals lateral to ethmo-turbinal III (Fig. 12d,i,n,s). Major distinctions existbetween the Microtus ochrogaster and the other three
TABLE 3. Surface areas of structures and boundariesof spaces in the nasal fossa in Microcebus murinus
Structure/ spacefor surface area(SA) measurement
AdultSA (mm2)
NeonatalSA (mm2)
Adult/neonateratio
Total nasal fossa 372.6 65.09 5.7Total OE in nasal fossa 113.8 24.8 4.6Non-OE 258.7 40.3 6.4% OE 30.6 38.1olfactory recess 28.2 8.2 3.4olfactory recess OE 19.6 2.1 9.4% OE 69.5 25.5nasopharyngealchamber total area(incl. recesses)
294.6 55.5 5.3
nasopharyngealchamber OE
73.6 22.7 3.2
%OE 25.0 40.9lateral recess, totalarea
64.2 11.5 5.6
OE in lateral recess 27.3 5.5 4.9%OE 42.5 48.3ET I: total area 40.4 3.2 12.7ET I: OE 14.2 2.4 6.0ET I: Non-OE 26.2 0.8 32.0%OE 35.1 74.2ET II: total area 13.2 1.3 9.9ET II: OE 7.0 1.1 6.6ET II: Non-OE 6.2 0.3 22.4%OE 53.2 79.4ET III: total area 11.0 1.3 8.0ET III: OE 6.1 1.0 6.0ET III: Non-OE 4.6 0.3 14.7%OE 57.3 76.6ET IV: total area 18.3 1.2 15.9ET IV: OE 8.4 08 10.8ET IV: Non-OE 9.9 0.4 26.4%OE 46.0 67.5maxilloturbinal: totalarea
79.1 5.7 13.8
frontoturbinal: totalarea
12.7 1.2 10.7
frontoturbinal: OE 8.1 1.1 7.2frontoturbinal: Non-OE 4.6 0.1 61.9%OE 63.4 93.6
OE, olfactory neuroepithelium.
907NASAL FOSSA OF MOUSE AND DWARF LEMURS

mammals. In Microtus ochrogaster, the transverse lam-ina more extensively overlaps the ethmoturbinal com-plex, encapsulating ethmoturbinals III and IV withinthe olfactory recess (Fig. 12s,t). In Microcebus murinus,
Cynocephalus volans, and Tenrec ecaudatus, only ethmo-turbinal IV is completely enclosed in the olfactory recess(Fig. 12e,j,o). More anterior distinctions include a rela-tively smaller maxilloturbinal in Microtus ochrogaster
Fig. 12. Comparison of sectioned material from late fetal/neonatalmammals, including coronal sections of a 28.5 CRL Microcebus muri-nus (Fig. 6a–e), Cynocephalus sp. (Fig. 6f–j), Tenrec ecaudatus (Fig.6k–o), and Microtus ochrogaster (Fig. 6p–t). In all species, ethmoturbi-nal I (I) projects anteriorly to overlap the pars intermedia (top row; AR,anterolateral recess). In all species, ethmoturbinals II through IV have
ventral roots from the paries nasi (see rows 2 through 5). Differencesin these turbinals mainly pertain to the shape. More significant differ-ences are found in the anteroposterior extent of the transverse lamina(rows 4 and 5). Scale bars 5 1 mm in a–d, 1 mm in f–I, 1 mm in k–n,750 mm in j, 500 mm e,o,9-t.
908 SMITH AND ROSSIE

(not shown, also see Adams, 1972). Additionally, the an-terior part of the nasoturbinal has cartilaginous supportin Microtus ochrogaster (not shown here), while in theother mammals it is only a mucosal, glandular bulge (asin the adult Microcebus murinus, Fig. 2c).Figure 13 shows roughly horizontal sections through
fetal heads of a 32 CRL Microcebus myoxinus (Fig.13a–d) and a 23.5 CRL Tenrec caudatus (Fig. 13e–h).Each series of sections progresses from ventral to dor-sal through the ethmoturbinal complex. The nasal cap-sules are in a similar state of maturation, in that allethmoturbinals have fully developed cartilaginous sup-ports. This view reveals that ethmoturbinal I is a directcontinuation of the orbitonasal lamina (i.e., the lateralwall of the pars posterior) that projects anteriorly, over-lapping the semicircular crest (Fig. 13a–c,e–g). In thisview, the more posterior ethmoturbinals are seen to begrowths from the medial aspect of the orbitonasal lam-ina. Each species can be seen to have two interturbi-nals between ethmoturbinals II and III, although theyare somewhat smaller in Microcebus murinus (Fig.13b,f). Figure 13d,h is near the cribriform plate, a pro-portionally larger frontal recess is seen in Tenrec ecau-
datus (Fig. 13h) compared with Microcebus murinus(Fig. 13d).
DISCUSSION
Functional interpretations of superficial (e.g., midfa-cial prognathism) and minute details (e.g., turbinal sizeand number) of extant and fossil primate noses hinge, inpart, on a detailed knowledge of the inner mucosal lin-ing (Smith et al., 2007a). Recent studies show that evo-lutionary patterns related to chemosensory structurescan be inferred from fossil material (Hillenius, 2000;Bush et al., 2004; Kay et al., 2004; Rossie, 2005).Although evidence is sometime indirect, osteologicalmorphology can yield estimates of the size of soft tissuestructures (Bush et al., 2004; Kay et al., 2004). Observa-tions on the nasal fossa of fossil primates are hamperedby the paucity of well-preserved material. However,some adequate material exists (see Smith and Rossie,2006; Smith et al., 2007a), and more data will likely beuncovered (Kay et al., 2006). The details provided here,on the association between internal osseous nasal struc-
Fig. 13. a–h: Fetal heads sectioned nearly in a horizonatal (palatal)plane, including a 32 CRL Microcebus myoxinus (a–d) and a 23.5 CRLTenrec caudatus (e–h). Each series of sections progresses from ventralto dorsal through the ethmoturbinal complex. The nasal capsules are
in a similar state of maturation, in that all ethmoturbinals have fullydeveloped cartilaginous supports. ft, frontoturbinal; FR, frontal recess;int, interturbinal; MR, maxillary recess; I–IV, ethmoturbinals I throughIV; SC, semicircular crest. Scale bars 5 500 mm.
909NASAL FOSSA OF MOUSE AND DWARF LEMURS

tures and mucosa in two cheirogaleids primates, areintended to inform such future studies.
Ontogeny of the Nasal Template
The prenatal Microcebus sample of the Bluntschli col-lection was opportunistically collected more than 70years ago, and no information on precise ages of theembryos and fetuses can be gleaned. However, this col-lection has been used to demonstrate graded characteris-tics of the solum nasi (Lozanoff and Diewert, 1989) andnasopalatine ducts (Shimp et al., 2003) of specimenswith differing CRL. Because Microcebus myoxinus domi-nates this sample, the bulk of interpretations of earlystages of development are based on this species. Becausethe 13 mm CRL M. myoxinus (MiSer. 1) was previouslyassessed as a Carnegie Stage 22/23 embryo (also seeLozanoff and Diewert, 1989), it can be used to approxi-mate that state of maturation at the end of the embry-onic timeframe. Figure 14a,b is a schematic diagramillustrating the maturation of the tectum and pariesnasi, respectively. The top row is based on observationsof 8- and 9-mm CRL Microcebus myoxinus (Stage 15–16embryos). The middle row represents the stage of matu-ration at the transition from embryonic to fetal periods.The bottom row is based on the fully chondrified capsu-les in fetuses of both M. murinus and M. myoxinus.The tripartite organization of the mammalian paries
nasi, beginning with three separate centers of mesen-chymal condensation, is a common theme of morphogen-esis (de Beer, 1937) with few known exceptions (Zeller,
1987). Because the ensuing maturation and growth ofthese centers varies among higher taxa (Zeller, 1987),early nasal capsule development of the mouse lemur isof interest. The specimens examined in this study sug-gest several spatial trends in development of the nasalcapsule. The nasal tectum develops in an anteroposteriorand mediolateral direction (Fig. 14a, top, middle). Abridging of the tectum to the paries nasi is first appa-rent anteriorly, as the parietotectal cartilage condenses(Fig. 14a, middle), as described for amniotes generally(de Beer, 1937). More posteriorly, it appears that theparanasal cartilages begin to form laterally and thenprogress toward the tectum, based on comparison ofembryos of different sizes. The mesenchymal condensa-tions of the orbitonasal lamina and paranasal cartilagesbegin to overlap relatively early by virtue of the exten-sion of ethmoturbinal I (14a,b, middle row). A schematicview of the full roof is shown in 14a (bottom).In Figure 14b the paries nasi is shown with the tec-
tum removed. The nasal capsules of the smallest Micro-cebus myoxinus suggest that the condensations for theorbitonasal lamina appear after those for the paranasalcartilages. The pars intermedia becomes overlappedinternally by the anterior margin of the par posteriorand the hind edge of the pars anterior by virtue of addi-tions of condensing mesenchyme. Ethmoturbinal I andthe semicircular crest are formed in this manner, bymeans of advancing streams of condensing mesenchyme(Fig. 14b, middle). Ethmoturbinals II to IV form in ananteroposterior sequence, as inferred from prenatalMicrocebus myoxinus and Microcebus murinus (see Figs.4f and 6a, respectively). Specimen MiSer. 54 (Microcebusmurinus) had a slightly more advanced maturation ofthe pars posterior, with a chondrified ethmoturbinal III,but only a mucosal bulge for ethmoturbinal IV.
Ontogeny of the Lateral Wall
Two clear ontogenetic trends are suggested by thecross-sectional sample of fetal, neonatal and adultMicrocebus murinus. First, there is a disproportionateincrease in dimensions of the anterior part of the nasalfossa, with expansion of the maxilloturbinal and the ad-jacent nasal fossa wall (mostly the viscerocranial maxil-lary and nasal bones). Based on reconstructions in Fig-ure 8, ethmoturbinal I may advance in anterior lengthat an even faster pace. This differential growth maybegin in the late fetal period. Three-dimensional ‘‘maps’’of the mucosa and quantitative findings indicate thatthis expansion of surface area involves mostly nonolfac-tory portions (Fig. 10). Whereas this study did not quan-tify respiratory mucosa, the preponderance of respira-tory mucosa on nonolfactory parts of ethmoturbinal I(Smith et al., 2007b) and the mucosa of the maxilloturbi-nal (see below) suggests that the growth in these ante-rior regions augments air-conditioning capacity.The second trend is seen posteriorly. While all ethmo-
turbinals become anteriorly elongated, the first andfourth increase disproportionately. This is evidencedmost plainly by the posterior elongation of ethmoturbi-nal IV, wherein a free projection elongates in tandemwith a posteriorly expanding olfactory recess (seeFig. 8).The quantitative comparisons support these impres-
sions of differential growth, because ethmoturbinals I
Fig. 14. Development of the capsule, organized into three regions(pars), based on prenatal Microcebus specimens. a,b: Schematic dia-gram of changes in the tectum, or roof, of the developing capsule (a)and the paries nasi, or lateral wall (b) are indicated. The top row indi-cates the earliest stage of development based on MiSer. 7 and MiSer.19). The middle row indicates late embryonic/early fetal stages (basedon MiSer. 1 and MiSer. 54). The resultant form is based on largerfetuses and neonates (bottom row). Stippled areas indicate mesenchy-mal condensations. Solid structures indicate chondrified capsule. PtC,parietotectal cartilage; PnC, paranasal cartilages; OnL, orbitonasallamina; NsC, nasal septal cartilage; SCr, semicircular crest; T, tectum;I–IV, ethmoturbinals I to IV.
910 SMITH AND ROSSIE

and IV have the highest of the adult/neonatal specimenratios (see Table 3). If the neonatal specimen is typicalin dimensions for a newborn Microcebus murinus, andthe adult is typical for its age, the differences suggestclear trends in postnatal growth. First, the dispropor-tionate growth of nonolfactory portions of ethmoturbinalI in Microcebus murinus (Smith et al., 2007b) appears tobe part of a trend throughout the entire nasal fossa.There is a decrease in olfactory SA of nearly 10%between neonatal and adult ages. Second, when thenasal fossa is divided into two regions, the olfactoryrecess and the remainder, the changes in SA indicatereverse trends in growth. The olfactory recess increasesits olfactory SA by nearly 45%, while the remainder ofthe nasal fossa decreases by more than 15% in olfactorySA. In the intermediate region of the nasal fossa, quan-titative observations and three-dimensional reconstruc-tions suggest that the anterolateral recess and ethmo-turbinals II and III expand at a rate more commensu-rate with overall nasal fossa growth. These apparentpatterns are interpreted as growth trends with caution,because these represent the first detailed quantificationsof the entire nasal fossa and its components in a pri-mate. Future analyses of other taxa are sorely needed.
Internal, Osseous Nasal Structures and Spaces:
Identification and Physiological Associations
Terminology. The terminology in this report,largely derived from Maier (1993a,b) is specifically use-ful for ontogenetic and evolutionary analyses of themammalian nasal fossa. The most numerous irregular-ities in terminology in Table 2 relate to turbinals. Whilesome terms may be interchangeable with little chance ofconfusion (e.g., frontoturbinal/ectoturbinal), some spe-cific practices have been problematic.First, the use of Maier’s terminology (1993a,b) is advo-
cated with respect to the ‘‘accessory’’ ethmoidal turbi-nals. Frontoturbinals or ectoturbinals are those nestedwithin the frontal recess. Accessory ethmoturbinalsarising more posteriorly include interturbinals and epi-turbinals. The former arise from the medial wall of theorbitonasal lamina, between (and covered medially by)ethmoturbinals. Epiturbinals are accessory scrolls of theethmoturbinals themselves. Developmentally, fronto-turbinals are products of the pars intermedia whileinterturbinals and epiturbinals derive from the pars pos-terior. This system resolves confusion in terms of simpledescriptive anatomy and potential issues regardinghomology. A drawback in the practice of collectively re-ferring to all of these elements as ‘‘ectoturbinals’’ (Paulli,1900) is that they are clearly not serial homologues.Second, it is argued here that the practice of describ-
ing ethmoturbinal I as a bipartite structure (see Table 2)is arbitrary. Ethmoturbinal I is the most anterior medi-ally positioned scroll of the ethmoid bone, and at thesame time, the most anterior limit of the orbitonasallamina. The next ethmoidal scroll in posterior processionis an independent element arising from the ventrome-dial margin of the orbitonasal lamina, as discussed fur-ther below. Historically, this issue has already beenraised, for different reasons. Kollmann and Papin(1925), citing Seydel (1891), also found the practice ofcombining the first two medial scrolls into a compositestructure as problematic. Based on embryology of the
human nasal fossa (‘‘...il y a plus d’ethmoıdaux chezl’Hommme qu’on croyait...’’ —p 5), they argued that thatthe practice of combining these elements is an unneces-sary construct, used to render easier the comparisons ofsome anthropoids to lemurs. Accordingly, Kollmann andPapin considered the series of (ethmo) turbinals as sepa-rate elements (‘‘...suffisamment invidividualisee.’’ —p 5).
Nasoturbinal. The nasoturbinal is among themost problematic internal structures in terms of nomen-clature (see Table 2). This may partially relate to differ-ing preferences among authors, but taxonomic variationmay also contribute. For example, the anterior portionof this turbinal encloses elements of the lateral nasalgland, which create a visible mucosal fold. There is nowell-defined osseous ridge supporting this fold in anyspecies under study (e.g., see Figs. 2c, 9a). However, insome other primates, the nasal bone projects a turbinalcrest inward in the region of this mucosal fold (e.g., lori-soids—Kollmann and Papin, 1925). In at least someinsectivores, an osseous turbinal is adjacent to the lat-eral nasal gland duct (see Fig. 1 in LaRochelle andBaron, 1989). More problematic is the large anteriorlypositioned osseous turbinal in rodents, commonly identi-fied as the nasoturbinal (Adams, 1972; Clancy et al.,1994; Jurcisek et al., 2003), which is directly continuouswith the ossified nasal tectum. The homology of thesestructures with the glandular part of the nasoturbinal, asdefined here, is uncertain. It is beyond the purpose of thisstudy to resolve this issue, and we are following Moore(1981) who used the entire structure defined here as syn-onymous with the nasoturbinal. However, a careful recon-sideration of the variation and development of differentsegments of the nasoturbinal is needed.In cheirogaleids, the nasoturbinal may be traced in de-
velopment by following the lateral margin of the nasaltectum. Anteriorly, the lateral nasal gland mass islocated on the medial margin of the tectum. The tectumhas disappeared from this position in adults (Fig. 2c).Passing posteriorly, the lateral part of the tectum contin-ues as the semicircular crest (see Fig. 7a), which thenbecomes attenuated dorsally (see Fig. 7b). These struc-tures ossify as dorsolateral walls of the ethmoid bone.The mucosal characteristics show that the glandularpart of the nasoturbinal bears respiratory mucosa. Inthe osseous part, the dorsal part of the semicircularcrest and the entire posterior part of the nasoturbinalbear olfactory mucosa. An extension of mucosa from theventral part of the semicircular crest that is unsup-ported by bone (see Figs. 7a, 9g) reveals mucosal surfa-ces that would be difficult to infer from osseous struc-tures alone. This creates extra mucosal surface, and con-tains glands of the lateral recess.
Maxilloturbinal. The maxilloturbinal ossifiesfrom the inferior margin of the paries nasi, as describedpreviously in other mammals (de Beer, 1937; Moore,1981). Observations of the mucosal covering indicatethat the medial convex surface is important for regulat-ing temperature during respiration, based on the pres-ence of numerous venous sinuses. A ciliated epitheliumwith unicellular glands is mostly restricted to the lateralconcave surface, which may be more important for airfiltration.
911NASAL FOSSA OF MOUSE AND DWARF LEMURS

Ethmoid bone. The ethmoid bone ossifies fromportions of all three components of the fetal capsule, thepars anterior (semicircular crest), pars intermedia (wallsof the frontal recess, frontomaxillary septum, frontotur-binal[s]), and the pars posterior (ethmoturbinal complex,posterior nasal tectum, lateral part of transverse lamina).The ethmoturbinal complex is the primary derivative
of the orbitonasal lamina of the pars posterior. Ethmo-turbinal I is a rostrally prolonged extension of the orbi-tonasal lamina of the pars posterior. The remaining eth-moturbinals project ventromedially from the orbitonasallamina. Progressing posteriorly, the lamina transversalisposterior forms the lateral part of the transverse lamina.It resembles the ethmoturbinals as the posterior-mostventromedial projection in the sequence. As such, thelamina transversalis posterior could be the homolog ofthe last ethmoturbinal of metatherians. In fact, the lastethmoturbinal in the opossum forms part of the floor ofthe olfactory recess (see ‘‘ethmoid plate,’’ Fig. 6, in Roweet al., 2005). It is not precisely clear at this time howthe ethmoturbinals of the opossum correspond with thatof Microcebus. A bipartite ‘‘endoturbinal I’’ was describedby Rowe et al. (2005; Fig. 10), corresponding to thesequence assigned as ethmoturbinals I and II here.Although this may indicate six ethmoturbinals in theopossum, the scroll identified as ‘‘endoturbinal III’’ byRowe et al. (2005, Fig. 6) does not appear to occupy themedial row, and may correspond to an interturbinalaccording to the terminology of Table 2. Further compar-ative work is required to establish developmental equiv-alence of ethmoturbinals between metatherian andeutherian mammals.Notwithstanding that all turbinals do derive from the
nasal capsule (a basis for more inclusive terminology perRowe et al., 2005), all ethmoturbinals have two commoncharacteristics. First, all develop from the pars posterior(which also does provide partial origin for the nasoturbi-nal). Second, all of these turbinals are distinctly andwholly part of the adult ethmoid bone. Therefore thereis both a developmental and anatomical basis for refer-ring to the four posterior turbinals in the medial row as‘‘ethmoturbinals.’’Our preference to consider the first two ethmoturbi-
nals as sequentially distinct elements is based on the de-velopmental observation that the so-called ventral lam-ina of ethmoturbinal I is one of three or more invagina-tions from the inner surface of the pars posterior. Thisagrees with Maier’s (1993b) descriptions of individualfetuses of Microcebus and Galago. In Daubentonia, eth-moturbinal I projects farther into the nasal fossa atbirth compared with these species (Maier, 1993b).Maier’s diagram of the ethmoidal labyrinth appears toshow a distinct division of this turbinal into two lami-nae, but without a series of prenatal stages, it is difficultto discern whether this turbinal divides or is at a moreadvanced stage of morphogenesis compared with hisdescriptions of Microcebus and Galago. We argue thatthe mammalian bauplan described by Maier (1993a)similarly shows that the ‘‘ventral lamina of ethmoturbi-nal I’’ is simply the first of a sequence of turbinals thatgrow from the inner face of the pars posterior. Thisargument may seem semantic, but it resolves some prob-lems in comparative morphology. Although the secondethmoidal turbinal does partially merge (postnatally)with the most anterior ethmoidal turbinal and share a
common primary lamina (a distinct lateral root—thefrontomaxillary septum), it also diverges from this lam-ina to anchor into the lateral wall of the nasal fossa byits own distinct root. In cheirogaleids, this separate rootis posterior to its union with the first ethmoturbinal (af-ter the latter merges with the roof of the nasal fossa—Fig. 2g). In rodents, the second ethmoturbinal is distinctanterior to its union with the first ethmoturbinal(Adams, 1972; Clancy et al., 1994).The notion of a bipartite ethmoturbinal I is rendered
questionable by the above information. In summary, thesecond ethmoidal turbinal can be termed ethmoturbinalII based on the following points. First, it is developmen-tally identified as one of three distinct structures on themedial face of the pars posterior. Second, ethmoturbinalI is distinct from ethmoturbinals II–IV because it is thecontinuation of the orbitonasal lamina of the pars poste-rior. Last, although ethmoturbinal II has the mostanterior emergence from the pars posterior, its asso-ciation with the first ethmoturbinal in the sequence isvariable.
Transverse lamina. This plate of bone, a result offusion between the lamina transversalis posterior andthe lateral (choanal) vomerine processes, creates a blind-ended olfactory recess dorsal to the nasopharyngealducts. The arrangement appears to be primitive for ther-ian mammals because the plate of bone is present inmetatherians and numerous eutherians (Negus, 1958),and its presence has been asserted in at least one Trias-sic mammal (Kielan-Jaworowska et al., 2004). The com-parative sample of this study suggests some develop-mental variations. Of particular note is the differinganteroposterior extent of the transverse lamina inMicrotus ochrogaster compared with all others. Therodent exhibits a common condition among mammals. Incats (Negus, 1958) and dogs (Craven et al., 2007), multi-ple ethmoturbinals are enclosed in the olfactory recess.Negus (1958) noted that the transverse lamina (whichhe termed the ‘‘subethmoid shelf ’’) is present in marsu-pials, felids, and canids, among others, but is small inrodents and ungulates. He also considered it absent inprimates, although numerous studies have since contra-dicted this (Maier, 1993b; Smith et al., 2004). The extentof this plate may relate directly to segregation of turbi-nal physiology. Craven et al. (2007) describe ethmoturbi-nals of the dog to be predominantly lined with olfactorymucosa. Furthermore, in canids and felids there is lessoverlap of the maxilloturbinal by the first ethmoturbinalprojection (see Fig. 99 in Negus, 1958; Fig. 3 in Cravenet al., 2007). By comparison, some strepsirrhine prima-tes (e.g., cheirogaleids and galagids—Kollmann andPapin, 1925; Smith et al., 2007b), at least some bats(Bhatnagar and Kallen, 1975), and the opossum (Roweet al., 2005) all have extensive overlap of the maxillotur-binal by ethmoturbinal I. In these animals, the trans-verse lamina is confined to toward the posterior part ofthe ethmoturbinal complex or is absent (in two batsdescribed by Bhatnagar and Kallen, 1975). Furthermore,in at least some of these mammals, a substantial portionof the ethmoturbinal I is lined with nonolfactory epithe-lium (Rowe et al., 2005; Smith et al., 2007b). This sug-gests that a more collaborative turbinal physiologyexists for some mammals, leaving an interesting unan-swered question of the plesiomorphic condition.
912 SMITH AND ROSSIE

Previous studies suggest that the ethmoturbinal com-plex is overlapped to a variable degree by the transverselamina in different strepsirrhines. Loo (1973) shows eth-moturbinal IV to be completely overlapped by the trans-verse lamina, whereas ethmoturbinals I–III are anteriorto the olfactory recess (Fig. 8, therein). While this rela-tionship is also described here for cheirogaleids, otherlemuroids differ. Hill describes ‘‘typical lemurs’’ (refer-ring to genera of the family Lemuridae) as having theethmoturbinals ‘‘more completely welded to the laminaterminalis of the ethmoid than in lorisidae or galagi-dae....’’ (1953, p 277). He also reported variations in thisregard among indriids. Clearly, the transverse lamina isnot a reliable structure to denote the limits of the eth-moturbinal complex.
Lateral recess. The nasal fossa provides an excel-lent example of an anatomical region where human vs.nonhuman or mammal vs. nonmammal anatomy is so di-chotomous that certain homologies remain obscure(Witmer, 1995; Smith and Rossie, 2006). In some circum-stances such as this, different schemes of terminologyfor anatomical structures may be partially unavoidable(see Wilson, 2006). However, paranasal recesses andsinuses have become interchanged repeatedly (see Table2 for some details). For instance, Rae and Koppe (2003)lamented the interchange in terminology of ‘‘lateralrecess’’ with spaces relating to the maxillary sinus or itsprimordium.The present study used a novel terminology for the
lateral recess. Specifically, it is proposed that the lateralrecess should be named regionally based on the intersec-tion of elements of the developing nasal capsule. Ossifi-cations or the pars intermedia form the entire lateralwall of the recess. The anterolateral recess is the portionof the lateral recess where the semicircular crest(derived from the pars anterior) forms the medial wall.Posteriorly, the first ethmoturbinal (derivative of thepars posterior) forms the medial border of the posterolat-eral recess. The posterolateral recess is divided furtherinto frontal and maxillary recesses, which are transectedby the horizontally oriented frontomaxillary septum.These terms explicitly relate to contours of the fetal car-tilaginous capsule. In Microcebus, the frontal recessappears first (see Fig. 4b) in keeping with observationson other mammals by Dieulafe (1906). Subsequently, theshape of the lateral recess relates to the formation of thesemicircular crest and differential growth of ethmoturbi-nal I as well as a region-specific outwardly folded pariesnasi. Confusion with terms such as the maxillary orfrontal sinus is specifically avoided, because these spacesform by means of the distinctive process of secondarypneumatization (Rossie, 2006).The frontal recess is primarily related to olfaction, a
primitive association (Negus, 1958). In Mirza coquereliand C. medius this space is more complex by virtue of asecond frontoturbinal. The anterolateral and maxillaryrecesses are predominantly nonolfactory spaces (except asmall strip of olfactory epithelium extending anterior tothe frontal recess). These spaces demonstrate the poten-tial incongruity of mucosa/bone surface areas. The thickglandular lamina propria would appear to impart agreat difference between estimates using osteological asopposed to mucosal boundaries.
Frontoturbinals. Moore (1981) reports that fron-toturbinals (called ectoturbinals by Moore) vary in num-ber from 13–20 in extant taxa. In part, this is becausehe follows Paulli (1900) by including structures referredto here as ‘‘interturbinals.’’ The terminology advocatedhere provides different names for turbinals that do notreach the medial row. The advantage of this practice isthat it partially differentiates these accessory scrolls,which are difficult to identify and determine relation-ships of homology. Interturbinals then, arise from theorbitonasal lamina. ‘‘Frontoturbinals’’ (Maier, 1993a,b)are found in the frontal recess and articulate with thefrontal bone in adults. Moore reported that ‘‘prosimians’’(clearly referring only to strepsirrhines in that actualcontext) possess only one or two frontoturbinals, andcheirogaleids conform to this. Paulli’s work (1900)shows that carnivores and ungulates have as many assevenfold additional frontoturbinals by comparison.Whether the number is reduced in primates as sug-gested by Moore (1981) or the other taxa evolved addi-tional structures awaits further study. Homologies offrontoturbinals are difficult to deduce, and an arbi-trary medial to lateral numbering scheme is used. Itshould not be assumed that frontoturbinal 1 of cheiro-galeids corresponds with frontoturbinal 1 of nonpri-mates. Indeed, this study found no dorsally positionedfrontoturbinals, except a possible rudiment in Mirza.This is consistent with a reduction in number due, inpart, to medial compression secondary to orbital con-vergence.
Olfactory recess. If the lamina transversalis pos-terior is indeed homologous with the last ethmoturbinalof opossum and other marsupials, the nasal fossa ofmetatherians and eutherians differs in more than eth-moturbinal number. The fundamental difference wouldalso include the complexity and extent of the olfactoryrecess. A potential conundrum exists in that this homol-ogy suggests that the transverse lamina is not formedsimilarly among different mammals. However, this mustbe the case broadly speaking, because the lamina over-laps the ethmoturbinal complex to a varying degree.Broader ontogenetic studies of therian mammals arerequired for a definitive answer.The present study indicates that in cheirogaleid pri-
mates, ethmoturbinal IV is the sole medial projectionwithin the olfactory recess. Although the transverselamina itself serves as valid indicator for a space dedi-cated for olfactory function, it cannot be assumed tocorrelate closely with the complexity or size of the eth-moturbinal complex in osteological studies of extant orfossil primates.The measurements of the olfactory recess in this
study begin in the coronal section where it is com-pletely enclosed, a highly replicable definition. Interms of olfactory air-currents, this definition is cer-tainly somewhat arbitrary and too rigid. Nonetheless,three-dimensional mucosal maps of Microcebus illus-trate that the recess, as defined herein, is predomi-nantly an olfactory space, in contrast to the nasopha-ryngeal chamber. As such, it represents potentiallyreliable proxy for the extent of olfactory mucosa. Ofconcern is the unknown extent to which the nasopha-ryngeal part of the nasal fossa shares in the functionacross different primates.
913NASAL FOSSA OF MOUSE AND DWARF LEMURS

Implications for Future Studies
Maier (1993a) emphasized the need for further studieson the morphogenesis of the mammalian nasal capsule.Few authors have studied or discussed prenatal ontog-eny of the nasal fossa since then (but see Lozanoff et al.,1993; Rowe et al., 2005; Rossie, 2003, 2006). Results ofthe present study illustrate that the ethmoturbinal com-plex of Microcebus develops from a cartilaginous tem-plate similar to mammals of diverse other taxa. Thenumber of ethmoturbinals occupying a medial positionin mammals has been reported to range from three tosix (Negus, 1958; Moore, 1981). The mechanism bywhich turbinals vary among mammalian groups remainunanswered to date. For example, do some mammals ac-quire complexity by adding turbinals or splitting exist-ing lamellae (e.g., pigs—Parker, 1874; Hillmann, 1971)?Which turbinals are lost in taxa with reduced complex-ity? Such questions require a broad range of species andages.The observations made to date suggest that possession
of four medial ethmoturbinals is a common theme, andmay have evolved through reduction of the last ethmo-turbinal after divergence from a common therian ances-tor. If stem primates developed a nasal fossae by meansof the same template, it seems likely that reduction ofthe ethmoturbinals is not a defining feature of the order,but occurred as a complex trend across different taxa.However, a reduction of the frontal recess, and the olfac-tory turbinals therein, could be.In terms of the olfactory versus nonolfactory SA, the
present study provides a broader perspective than a sim-ilar previous study on an individual turbinal (Smithet al., 2007b). An age comparison supports the idea thatolfactory epithelium scales differently than other epithe-lia between ages. Along the surfaces of ethmoturbinal I,Smith et al. (2007b) suggested that nonolfactory mucosaincreased to a lesser extent than olfactory mucosa. Thepresent study suggests this observation may be appliedmore broadly across the nasal fossa. In Microcebus muri-nus, total olfactory SA is 3.5-fold greater in adult versusneonate, whereas nonolfactory SA is more than 5.4-foldgreater.These quantitative data are the most detailed to date
in terms of structure-mucosa relationships. Thus, thesefindings can inform future osteological analyses thatattempt to infer physiological attributes of the nasalfossa, such as olfactory SA or air flow dynamics, in liv-ing or extinct primates. It is possible that the data on ol-factory SA of individual elements reflects specific charac-teristics of Microcebus, so interpretations are necessarilygeneralized at present. However, the microanatomicaland quantitative findings in this study offer guidance in theosteological interpretation of extant and fossil primates.The following generalizations can be made at present:
(1) This study shows that the anterior part of the nasalfossa is mostly nonolfactory in M. murinus, and themagnitude of nonolfactory SA increases postnatally.Thus, the term ‘‘olfactory snout’’ may misrepresent theprognathic midface of strepsirrhines. (2) The SA of theethmoturbinal complex must be interpreted withcaution. Restricted functional associations of individualturbinals with specific physiology may apply to somemammals (e.g., rats, mice, and dogs), but at least someprimates do not exhibit this clear division of labor. (3)
Two spaces, the olfactory recess and frontal recess, arereliable proxies for olfactory SA in Microcebus. However,the olfactory recess in cheirogaleids is not coextensivewith the ethmoturbinal complex. Variations in the trans-verse lamina across mammals, especially with regard tothe extent that it encloses ethmoturbinals, remainobscure. (4) Structures such as the glandular portion ofthe nasoturbinal and the mucosal SA of the olfactoryrecess are difficult to infer from osseous contours.
ACKNOWLEDGMENTS
We thank E. Westwig and D. Lunde of the AmericanMuseum of Natural History for access to the Bluntschlicollection of prenatal mammals and J.R. Wible for accessto a fetal colugo. We also thank M.P. Mooney for accessto a stereomicroscope. Lastly, many thanks to S.B.McLaren for caring for a portion of the sample studiedherein, now housed at the Carnegie Museum, Sectionof Mammals. This is Duke Lemur Center publication# 1130.
LITERATURE CITED
Adams DR. 1972. Olfactory and non-olfactory epithelia in the nasalcavity of the mouse, Peromyscus. Am J Anat 133:37–50.
Adams DR. 1986. Transitional epithelial zone of the bovine nasalmucosa. Am J Anat 176:159–170.
Allen H. 1882. On a revision of the ethmoid bone in Mammalia,with special reference to the description of this bone in and of thesense of smell in the Chiroptera. Bull Mus Comp Zool Harvard10:135–164.
Ankel-Simons F. 2007. Primate anatomy: an introduction. NewYork: Academic Press.Bhatnagar KP, Kallen FC. 1975. Quantita-tive observations on the nasal epithelia and olfactory innervationin bats. Acta Anat (Basel) 91:272–282.
Bhatnagar KP, Wible JR. 1994. Observations on the vomeronasalorgan of the colugo Cynocephalus (Mammalia, Dermoptera). ActaAnat (Basel) 151:43–48.
Bush E, Simons E, Allman A. 2004. High resolution CT study of thecranium of a fossil anthropoid primate, Parapithecus grangeri:new insights into the evolutionary history of primate sensory sys-tems. Anat Rec 281A:1083–1087.
Cartmill M. 1974. Rethinking primate origins. Science 184:436–443.Cave AJE. 1973. The primate nasal fossa. Biol J Linn Soc Lond
5:377–387.Clancy AN, Schoenfeld TA, Forbes WB, Macrides F. 1994. The spa-
tial organization of the peripheral olfactory system of the ham-ster. II: Receptor surfaces and odorant passageways within thenasal cavity. Brain Res Bull 34:211–241.
Craven BA, Neuberger T, Paterson EG, Webb AG, Josephson EM,Morrison EE, Settles GS. 2007. Reconstruction and morphometricanalysis of the nasal airway of the dog (Canis familiaris) andimplications regarding olfactory airflow. Anat Rec 290:1325–1340.
de Beer GR. 1937. The development of the vertebrate skull. Chi-cago: Chicago University Press.
Dieulafe L. 1906. Morphology and embryology of the nasal fossae ofvertebrates. Ann Otol Rhinol Laryngol 15:1–584.
Hall BK. 1994. Introduction. In: Hall BK, editor. Homology. Thehierarchical basis of comparative biology. San Diego: AcademicPress. p 1–19.
Hill WCO. 1953. Primates, comparative anatomy and taxonomy. I.Strepsirrhini. Edinburgh: Edinburgh University Press.
Hillenius WJ. 1992. The evolution of nasal turbinates and mamma-lian endothermy. Paleobiology 18:17–29.
Hillenius WJ. 1994. Turbinates in therapsids: evidence for endo-thermy in mammal-like reptiles. Evolution 48:207–229.
Hillenius WJ. 2000. Septomaxilla of nonmammalian synapsids: soft-tissue correlates and a new functional interpretation. J Morphol245:29–50.
914 SMITH AND ROSSIE

Hillmann DJ. 1971. Anatomy of the nasal cavity and paranasalsinuses of the domestic pig (Sus Scrofa Domestica). Ph.D. disser-tation. Iowa State University.
Jurcisek JA, Durbin JE, Kusewitt DF, Bakaletz LO. 2003. Anatomyof the nasal cavity in the chinchilla. Cells Tissues Org 174:136–152.
Kay RF, Rossie JB, Colbert MW, Rowe TB. 2004. Observations ofTremacebus harringtoni (Platyrrhini, early Miocene, Sacanana,Argentina) based on high-resolution X-ray CT- scans. Anat Rec281A:1157–1172.
Kay RF, Rae TC, Koppe T, Colbert MW. 2006. Paranasal sinuspneumatization in the Miocene platyrrhine Homunculus patago-nicus. Am J Phys Anthropol Suppl 42:112.
Kielan-Jaworowska Z, Cifelli R, Luo Z-X. 2004. Mammals from theage of dinosaurs: origins, evolution, and structure. New York:Columbia University Press.
Kollmann M, Papin L. 1925. Etudes sur lemuriens. Anatomie com-paree des fosses nasales et de leurs annexes. Archs Morph ge exp22:1–60.
Kumar P, Kumar S, Singh Y. 1993. Histological studies on the nasalethmoturbinates of goats. Small Rumin Res 11:85–92.
Kumar P, Timoney JF, Southgate HHP, Sheoran AS. 2000. Lightand scanning electron microscopic studies of the nasal turbinatesof the horse. Anat Histol Embryol 29:103–109.
LaRochelle L, Baron G. 1989. Comparative morphology and mor-phometry of the nasal fossae of four species of North Americanshrews (Soricinae). Am J Anat 186:306–314.
Le Gros Clark WE. 1959. The antecedents of man. Edinburgh:Edinburgh University Press.
Loo SK. 1973. A comparative study of the nasal fossa of four nonhu-man primates. Folia Primatol (Basel) 20:410–422.
Loo SK. 1974. Comparative study of the histology of the nasal fossain four primates. Folia Primatol 21:290–303.
Lozanoff S, Diewert VM. 1989. Developmental morphology of the so-lum nasi in the mouse lemur (Microcebus murinus). J Morphol202:409–424.
Lozanoff S, Zingeser MR, Diewert VM. 1993. Computerized model-ling of nasal capsular morphogenesis in prenatal primates. ClinAnat 6:37–47.
Maier W. 1993a. Cranial morphology of the therian common ances-tor, as suggested by adaptations of neonate marsupials. In: SzalayF, Novacek MJ, McKenna MC, editors. Mammal phylogeny. NewYork: Springer-Verlag. p 165–181.
Maier W. 1993b. Zur evolutiven und funktionellen Morphologie desGesichtsschaedels der Primaten. Z Morph Anthrop 79:279–299.
Martin RD. 1990. Primate origins and evolution. A phylogeneticreconstruction. Princeton, NJ: Princeton University Press.
Moore WJ. 1981. The mammalian skull. London: Cambridge Uni-versity Press.
Moulton DG, Beidler LM. 1967. Structure and function of the pe-ripheral olfactory system. Physiol Rev 47:1–51.
Napier JR, Napier PH. 1967. A handbook of living primates. NewYork: Academic Press.
Negus V. 1958. The comparative anatomy and physiology of thenose and paranasal sinuses. Livingston: Edinburgh and London.
Nomina Anatomica Veterinaria. 2005. www.wava-amav.org/nav.htm.O’Rahilly RM Muller F. 1987. Developmental stages in human
embryos. Washington: Carnegie Institute of Washington.Parker WK. 1874. On the structure and development of the skull
in the pig (Sus scrofa). Philos Trans R Soc Lond 164:289–336.Paulli S. 1900. Uber die Pneumaticitat des Schadels bei den Sauge-
thieren. III. Uber die Morphologie des Siebbeins der Pneumatici-tat bei den Insectivoran, Hyracoideen, Chiropteren, Carnivoren,Pinnepedien, Edentates, Rodentiern, Prosimien und Primaten.Gegenb Morpholog Jahrb 28:483–564.
Pedziwiatr ZF. 1972. Das Siebbeinlabyrinth .II. Differenzierung undSystematik der Hauptmuschel bein einigen Gattungen der Sauge-tiere. Anat Anz 131:378–390.
Perret M. 1992. Environmental and social determinants of sexualfunction in the male lesser mouse lemur (Microcebus murinus).Folia Primatol 59:1–25.
Rae TC, Koppe T. 2003. The term ‘‘lateral recess’’ and craniofacialpneumatization in Old World monkeys (Mammalia, Primates,Cercopithecoidea). J Morphol 258:193–199.
Read EA. 1908. A contribution to the knowledge of the olfactoryapparatus in dog, cat and man. Am J Anat 8:17–47.
Rehorek SJ, Smith TD. 2007. Concurrent 3-D visualization of multi-ple microscopic structures. In: Mendez-Vilas A, Diaz J, editors.Modern research and educational topics on microscopy. No. 3. Vol.2. Badojoz, Spain: Formatex. p 917–923.
Rossie JB. 2003. Ontogeny, homology, and phylogenetic significanceof anthropoid paranasal sinuses. New Haven, CN: Yale Univer-sity.
Rossie JB. 2005. Anatomy of the nasal cavity and paranasal sinusesin Aegyptopithecus and early Miocene African catarrhines. Am JPhys Anthropol 126:250–267.
Rossie JB. 2006. Ontogeny and homology of the paranasal sinusesin Platyrrhini (Mammalia: Primates). J Morphol 267:1–40.
Rossie JB, Smith TD. 2007. Ontogeny of the nasolacrimal duct inprimates: functional and phylogenetic implications. J Anat 210:195–208.
Rowe TB, Eiting TP, Macrini TE, Ketcham RA. 2005. Organizationof the olfactory and respiratory skeleton in the nose of the grayshort-tailed opossum Monodelphis domestica. J Mamm Evol 12:303–336.
Schilling A. 1970. L’organe de Jacobson du lemurien malgacheMicrocebus. murinus. Mem Mus d’Hist Nat (Serie A) 61:203–280.
Seydel O. 1891. Uber die Nasenhohle der hoheren Saugethiere unddes Menschen. Morphol Jahrb 18:44–99.
Shimp KL, Bhatnagar KP, Bonar CJ, Smith TD. 2003. Ontogeny ofthe nasopalatine duct in primate. Anat Rec 274A:862–869.
Siemers BM, Goerlitz HR, Robsomanitrandrasana E, Piep M, Ram-anamanjato JB, Rakotondravony D, Ramilijaona O and GanzhornJU. 2007. Sensory basis of food detection in wild Microcebus mur-inus. Int J Primatol 28:291–304.
Smith TD, Rossie JB. 2006. Primate olfaction: anatomy and evolu-tion. In: Brewer W, Castle D, Pantelis C, editors. Olfaction andthe brain: window to the mind. Cambridge: Cambridge UniversityPress. p 135–166.
Smith TD, Bhatnagar KP, Tuladhar P, Burrows AM. 2004. Distribu-tion of olfactory epithelium in the primate nasal cavity: are‘‘microsmia’’ and ‘‘macrosmia’’ valid morphological concepts? AnatRec 281A:1173–1181.
Smith TD, Rossie JB, Bhatnagar KP. 2007a. Evolution of the noseand nasal skeleton in primates. Evol Anthropol 16:132–146.
Smith TD, Bhatnagar KP, Rossie JB, Docherty BA, Burrows AM,Mooney MP, Siegel MI. 2007b. Scaling of the first ethmoturbinalin nocturnal strepsirrhines: olfactory and respiratory surfaces.Anat Rec 290:215–237.
Smith TD, Bhatnagar KP, Dennis JC, Morrison EE, Park TJ. 2007c.Growth deficient vomeronasal organs in the naked mole-rat (Het-erocephalus glaber). Brain Res 1132:78–83.
Van Gilse PHG. 1927. The development of the sphenoidal in manand its homology in mammals. J Anat 61:153–166.
Warrick R, Brookes M. 1989. Nomina Anatomica. Edinburgh:Churchill Livingstone.
Wilson JA. 2006. Anatomical nomenclature of fossil vertebrates: stand-ardized terminology or ‘lingua franca.’ J Vert Paleontol 26:511–518.
Witmer LM. 1995. Homology of facial structures in extant archo-saurs (birds and crocodilians), with special reference to paranasalpneumaticity and nasal conchae. J Morphol 225:269–327.
Zeller U. 1987. Morphogenesis of the mammalian skull with specialreference to Tupaia. In: Kuhn HJ, Zeller U, editors. Morphogene-sis of the mammalian skull. New York: Springer Verlag. p 17–50.
Zuckerkandl E. 1887. Das periphere Geruchsorgan der Saugethiere.Stuttgart: Verlag von Ferdinand Enke.
915NASAL FOSSA OF MOUSE AND DWARF LEMURS
Related Documents











