Indian Journal of Experimental Biology Vol. 52, August 2014, pp. 763-772 Nano gold conjugation, anti-arthritic potential and toxicity studies of snake Naja kaouthia (Lesson, 1831) venom protein toxin NKCT1 in male albino rats and mice Partha Pratim Saha 1 , Tanmoy Bhowmik 1 , Anjan Kumar Dasgupta 2 & Antony Gomes 1 * 1 Laboratory of Toxinology & Experimental Pharmacodynamics Department of Physiology, University of Calcutta, 92 A P C Road, Kolkata 700 009, India 2 Department of Biochemistry, University of Calcutta 35 Ballygunge Circular Road, Kolkata 700 019, India Received 20 January 2014; revised 8 May 2014 Nanoscience and Nanotechnology have found their way in the fields of pharmacology and medicine. The conjugation of drug to nanoparticles combines the properties of both. In this study, gold nanoparticle (GNP) was conjugated with NKCT1, a cytotoxic protein toxin from Indian cobra venom for evaluation of anti-arthritic activity and toxicity in experimental animal models. GNP conjugated NKCT1 (GNP-NKCT1) synthesized by NaBH 4 reduction method was stable at room temperature (25±2 °C), pH 7.2. Hydrodynamic size of GNP-NKCT1 was 68–122 nm. Arthritis was developed by Freund's complete adjuvant induction in male albino rats and treatment was done with NKCT1/GNP-NKCT1/standard drug. The paw/ankle swelling, urinary markers, serum markers and cytokines were changed significantly in arthritic control rats which were restored after GNP-NKCT1 treatment. Acute toxicity study revealed that GNP conjugation increased the minimum lethal dose value of NKCT1 and partially reduced the NKCT1 induced increase of the serum biochemical tissue injury markers. Histopathological study showed partial restoration of toxic effect in kidney tissue after GNP conjugation. Normal lymphocyte count in culture was in the order of GNP-NKCT1>NKCT1>Indomethacine treatment. The present study confirmed that GNP conjugation increased the antiarthritic activity and decreased toxicity profile of NKCT1. Keywords: Gold nanoparticle, Gold nanoparticle conjugation, Naja kaouthia, NKCT1, Rheumatoid arthritis, Snake venom, Toxicity study Application of snake venom toxins in health and diseases have been mentioned in folk-traditional medicine 1 . Snake venom toxins showed activity against various experimental pathology/animal studies such as cancer 2,3 , stroke 4 , neural trauma, alzheimer's disease, parkinson's disease 5 . Anti-arthritic activity of Indian monocellate cobra (Naja kaouthia) venom on adjuvant induced arthritis has been reported 6 . Chen and Robinson 7 , isolated cobrotoxin from snake Naja naja atra, which possessed anti-nociceptive effect in mice. Pretreatment of cobra venom factor delayed the acute inflammation followed by induction of chronic arthritis in rat model 8 . Treatment with small dose of cobra venom factor delayed the onset of adjuvant arthritis and inhibited the maximum inflammatory response 8 . One of the major drawbacks of most of the snake venom toxins that they failed to cross the barrier of safety line due to their toxicity and thereby hindered the process of drug development. In order to overcome the toxicity of these potential toxins, several attempts have been made which include liposome encapsulation, silica coating and nanoparticle tagging 9,10 . Toxins have been conjugated with nanoparticles to increase their functional efficacy, reduction of toxicity and targeted drug delivery 11,12 . Snake venom from Walterinessia aegyptia (WEV) was combined with silica nanoparticles, decreased the expression of Bcl-2 and enhanced the activation of caspase-3 in breast cancer cell line (MDAMB-231). Nano-particle along with WEV also showed significant reduction in actin polymerization and cytoskeletal rearrangement 13 . Melittin (a component of bee venom) loaded nanoparticles have the ability to inhibit infectivity by CXCR4 and CCR5 tropic HIV-1 strains 14 . The foundation of nanotechnology research is based on the size and shape of the nanoparticles, where distinct optical, electronic, or magnetic properties can be tuned during chemical synthesis. ________________ *Correspondent author Telephone: 91-33-23508386/6387/6396/1397 (Extn: 229) Fax: 91-33-2351-9755/2241-3288 E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Indian Journal of Experimental Biology
Vol. 52, August 2014, pp. 763-772
Nano gold conjugation, anti-arthritic potential and toxicity studies of snake Naja
kaouthia (Lesson, 1831) venom protein toxin NKCT1 in male albino rats and mice
Partha Pratim Saha1, Tanmoy Bhowmik
1, Anjan Kumar Dasgupta
2 & Antony Gomes
1*
1Laboratory of Toxinology & Experimental Pharmacodynamics
Department of Physiology, University of Calcutta, 92 A P C Road, Kolkata 700 009, India 2Department of Biochemistry, University of Calcutta
35 Ballygunge Circular Road, Kolkata 700 019, India
Received 20 January 2014; revised 8 May 2014
Nanoscience and Nanotechnology have found their way in the fields of pharmacology and medicine. The conjugation
of drug to nanoparticles combines the properties of both. In this study, gold nanoparticle (GNP) was conjugated with
NKCT1, a cytotoxic protein toxin from Indian cobra venom for evaluation of anti-arthritic activity and toxicity in
experimental animal models. GNP conjugated NKCT1 (GNP-NKCT1) synthesized by NaBH4 reduction method was stable
at room temperature (25±2 °C), pH 7.2. Hydrodynamic size of GNP-NKCT1 was 68–122 nm. Arthritis was developed by
Freund's complete adjuvant induction in male albino rats and treatment was done with NKCT1/GNP-NKCT1/standard drug.
The paw/ankle swelling, urinary markers, serum markers and cytokines were changed significantly in arthritic control rats
which were restored after GNP-NKCT1 treatment. Acute toxicity study revealed that GNP conjugation increased the
minimum lethal dose value of NKCT1 and partially reduced the NKCT1 induced increase of the serum biochemical tissue
injury markers. Histopathological study showed partial restoration of toxic effect in kidney tissue after GNP conjugation.
Normal lymphocyte count in culture was in the order of GNP-NKCT1>NKCT1>Indomethacine treatment. The present
study confirmed that GNP conjugation increased the antiarthritic activity and decreased toxicity profile of NKCT1.
Keywords: Gold nanoparticle, Gold nanoparticle conjugation, Naja kaouthia, NKCT1, Rheumatoid arthritis, Snake venom,
Toxicity study
Application of snake venom toxins in health and
diseases have been mentioned in folk-traditional
medicine1. Snake venom toxins showed activity against
various experimental pathology/animal studies such as
cancer2,3
, stroke4, neural trauma, alzheimer's disease,
parkinson's disease5. Anti-arthritic activity of Indian
monocellate cobra (Naja kaouthia) venom on adjuvant
induced arthritis has been reported6. Chen and
Robinson7, isolated cobrotoxin from snake Naja naja
atra, which possessed anti-nociceptive effect in mice.
Pretreatment of cobra venom factor delayed the acute
inflammation followed by induction of chronic arthritis
in rat model8. Treatment with small dose of cobra venom
factor delayed the onset of adjuvant arthritis and
inhibited the maximum inflammatory response8.
One of the major drawbacks of most of the snake
venom toxins that they failed to cross the barrier of
safety line due to their toxicity and thereby hindered
the process of drug development. In order to
overcome the toxicity of these potential toxins,
several attempts have been made which include
liposome encapsulation, silica coating and
nanoparticle tagging9,10
. Toxins have been conjugated
with nanoparticles to increase their functional
efficacy, reduction of toxicity and targeted drug
delivery11,12
. Snake venom from Walterinessia
aegyptia (WEV) was combined with silica
nanoparticles, decreased the expression of Bcl-2 and
enhanced the activation of caspase-3 in breast cancer
cell line (MDAMB-231). Nano-particle along with
WEV also showed significant reduction in actin
polymerization and cytoskeletal rearrangement13
.
Melittin (a component of bee venom) loaded
nanoparticles have the ability to inhibit infectivity by
CXCR4 and CCR5 tropic HIV-1 strains14
.
The foundation of nanotechnology research is
based on the size and shape of the nanoparticles,
where distinct optical, electronic, or magnetic
properties can be tuned during chemical synthesis.
________________
*Correspondent author
Telephone: 91-33-23508386/6387/6396/1397 (Extn: 229)
Fax: 91-33-2351-9755/2241-3288
E-mail: [email protected]

INDIAN J EXP BIOL, AUGUST 2014
764
There is an enormous interest in exploiting gold
nanoparticals (GNP) in various biomedical
applications since their size scale is similar to that of
biological molecules like protein, DNA etc. Over the
past two decades, their colours and unique
physicochemical properties have also attracted
attention due to their historical importance and current
applications in biotechnology and material science.
The use of GNP as biomarkers in biology has been
rapidly increased due to high scattering efficiencies,
immune to photo bleaching, and non-cytotoxic, which
provides advantage over semiconductor
nanoparticles15
. Functionalizing gold nanoparticles
with proteins could promote efficient binding,
clearance, and biocompatibility16
. Gold nanoparticle
conjugation is a dynamic domain of drug designing in
biomedical research and one of the most important
emerging areas in modern toxicology for effective
targeted delivery, reducing toxicity and increasing
efficacy. In the present study, a snake venom protein
toxin NKCT1, isolated from Indian monocellate
cobra, Naja kaouthia Lesson 1831, venom,
conjugated with gold nanoparticle and its anti-arthritic
property as well as toxicity profile have been
evaluated in experimental animal model.
Materials and Methods
Materials and reagents—Chemicals and reagents
used were of analytic grade. The following chemicals
were used: acetyl acetone reagent, acrylamide,
ammonium persulphate, bisacrylamide, disodium
hydrogen phosphate, cupric sulphate, ethanol, folin
ciocalteu reagent, glycine, indomethacin,
mercaptoethanol, methanol, para dimethyl amino
benzaldehyde, paraffin, P-nitrophenyl phosphate,
poly(ethylene)glycol, TRIS, sodium borohydride,
sodium lauryl sulphate were of SRL (India); BSA,
carboxymethyl cellulose, coomassie brilliant blue,
Freund’s complete adjuvant, HAuCL4 and histopaque
were from Sigma (USA); bromophenol blue, CK-MB
kit, diastix-reagent strips, LDH, methyline blue,
osteomol, potassium dihydrogen phosphate, SDS,
SGOT, SGPT kit, sodium potassium tartarate,
TEMED, urea kit and, xylene were from Merck
(India); eosin, formaldehyde and hematoxylin were
from Qualigen (India); ELISA kit - CINC1, TNF-α,
IL-1β, IL-6, CINC-1, IL-10 and VEGF kit were from
R & D (USA); Drabkin solution of Cogent (India) and
tris buffer were from Spectrochem (India).
Collection of snake venom—Lyophilized Naja
kaouthia snake venom (NKV) was purchased from
Calcutta Snake Park, Kolkata, India. Venom
concentration was expressed in terms of dry
weight/protein equivalent17
.
Purification of NKCT1—NKCT1 was purified from NKV by Ion-exchange column chromatography
and HPLC18
. The fraction was desalted and
concentrated by centricon (Millipore MWCO 3 k).
Purified NKCT1 was checked for homogeneity by
HPLC and SDS-PAGE gel electrophoresis19
.
Synthesis and characterization of GNP-NKCT1—
GNP was prepared by sodium borohydrate reduction
method20
with modification. Using poly
ethyleneglycol (PEG), NKCT1 conjugation and
characterization was done as reported earlier10
.
Experimental animals—Wistar male albino rats
(120±10 g) and Swiss albino male mice (20±2 g) were
procured from the approved animal breeders and
housed in standard polypropylene cages at controlled
temperature (25±2 °C), with light conditions (12 h light
and dark cycle) and 65±5% RH. The animals were
provided with pellet diet, green vegetables, gram and
water ad libitum. The experiments were conducted
according to the departmental animal ethics
committee for the purpose of control and supervision
of experiments on animals. All animal experiments
were approved by the Institutional Animal Ethics
Committee (AEC Ref: 820/04/ac/CPC SEA.2010).
Antiarthritic activity—Development of experimental
rheumatoid arthritis: Experimental rheumatoid arthritis
(RA) was developed by injecting (sub-plantar) 0.1 mL
emulsion of FCA in olive oil (1:1 v/v) at a
concentration of 0.25 mg heat killed Mycobacterium
tuberculosis/mL of emulsion into the right hind footpad
of male rats on day 021
.
Treatment schedule: Rats were divided into
following 6 groups of 6 each: Gr. 1: sham control;
Gr. 2: arthritis control, Gr. 3: standard (indomethacin,
0.25 mg.100g-1
x 5 days alternately, po), Gr. 4:
NKCT1treated (2µg.100g-1
x 14 days, ip), Gr. 5:
GNP-NKCT1 treated (2µg.100g-1
x 14 days, ip),
Gr. 6: GNP treated (200 µL of 48µM GNP.100g-1 x 14
days, ip). Treatment was started on day 3 after FCA
induction. The rats in the control group received the
same volume of vehicle. Urine was collected on day 14.
On day 16 blood was collected and serum was separated
for the analysis of biochemical parameters. Right hind
paw and ankle were collected for histological studies.
Physical parameters: On day 0, 2, 5, 10 and 15,
paw and ankle swellings of all animals were measured
with digital caliper (Mitutoyo, Japan). Paw and ankle

SAHA et al.: NANOGOLD CONJUGATED PROTEIN TOXIN & ANTIARTHRITIC POTENTIAL
765
diameter measured just prior to arthritis induction
were used as the control diameter (day 0).
Biochemical analysis of urine and serum: Urinary
hydroxyproline22
and glucosamine23
were measured
spectrophotometrically (Analab-180, USA). Serum
alkaline (ACP) and acid phosphatase (ALP) were
measured by biochemical methods24
. The measurements
of serum pro and anti-inflammatory interleukins
(TNF-α, IL-1β, IL-6, CINC1, IL-10, VEGF) were done
by ELISA kit, using ELISA reader (BioTek, ELx800).
Histopathological studies of joints— Joints were
fixed in 10% buffered formalin for 24 h, decalcified
in osteomol for 4-5 days, dehydrated in graded
alcohol, cleared in xylol and embedded in paraffin
wax (56–58 °C). Sections (5 µm thick) were cut with
rotary microtome (Weswox Optik, India), stained
with haematoxylin-eosin, and observed under bright
field microscope (Motic BA 450, Germany) and
photography were captured with Motic software
(Motic Images Plus 2.0 software).
Acute toxicity studies—Determination of minimum
lethal dose: NKCT1 and GNP-NKCT1 minimum
lethal dose (MLD) was determined in Swiss albino
male mice. Different dose (1, 1.5, 2, 2.5, 2.75, 3 and
3.5 mg.kg-1
) of NKCT1 and GNP-NKCT1were
administered (sc) to different animal groups and
mortality were recorded up to 24 h of observation.
The minimum dose that killed 100% animals in 24 h
was considered as the minimum lethal dose (MLD).
Acute toxicity treatment schedule: Swiss albino
male mice (20±2 g) were divided into following 3
groups of 6 each: Gr. 1: Sham control; Gr. 2: NKCT1
treated (2 µg.100g-1
, sc); Gr. 3: GNP-NKCT1 treated
(2 µg.100g-1
, sc). The toxic manifestation, mortality,
food and water intake, fecal consistency of animals
were monitored up to 24 h and qualitative urine
parameters were recorded by Diastix- Reagent Strips
(Merck, India) after 24 h.
Hematology and biochemical parameters: Blood
was collected from all the animals at 8 and 24 h by
cardiac puncture and in heparin vials. Total WBC,
RBC count and Hb% were estimated. Serum glutamate
oxaloacetate transaminase (SGOT), serum glutamate-
pyruvate transaminase (SGPT), urea, creatinine,
creatine kinase (CK) MB isoenzyme (CK-MB), lactate
dehydrogenase (LDH) were estimated after 24 h,
using biochemical kits (Ecoline, Merck, India).
Histopathology of kidney: Kidney tissue was
collected after 24 h of treatment and fixed with 10%
buffer formalin for 24 h. The organs were dehydrated
in graded alcohol, cleared in xylol and embedded in
paraffin wax (56–58 °C). Sections (5 µm thick) were
cut with rotary microtome (Weswox Optik, India),
stained with haematoxylin-eosin, and observed under
bright field microscope (Motic BA 450, Germany).
Photographs were captured with Motic software
(Motic Images Plus 2.0 software) to observe
microscopic changes in the organs.
Human lymphocytes collection and toxicity study:
Blood was collected in heparinized vial from the
healthy adults (aged 24-30 years) after informed
consent. The experiment was approved by the
Institutional human ethics committee (Ref. No. IHEC/
AG/HUM/P17/12). Lymphocytes were collected from
whole blood using histopaque after centrifugation at
1200 rpm for 5 min. They were cultured in sterile
RPMI 1640 media. Cells were grown in a CO2
incubator at 37 °C with 5% CO2 in humidified
condition. Normal lymphocytes (1x 106) were seeded
on 96 well and treated with GNP-NKCT1/NKCT1
(2 µg.mL-1
) of cell lines with respect to standard drug
Indomethacine (250 µg.mL-1
). The cell growth
inhibition studies were done using 0.2% trypan blue
with direct count under light microscope (Olympus,
Tokyo) using haemocytometer chamber.
Statistical analysis—Data were expressed as
mean±SE (n=6). The repeated measure analysis of
variance (ANOVA) was used to determine significant
differences between groups p<0.05 was considered to
be statistically significant.
Results Purification of NKCT1—Naja kaouthia venom was
applied on CM-cellulose column equilibrated with
phosphate buffer (pH 7.2). Protein fraction eluted by
0.5 M NaCl was collected and further purified by
HPLC using Protein Pak 60 column. A single sharp
peak with retention time 22 min was collected.
SDS-molecular weight of the protein was found to be
6.76 kDa and designated as NKCT1.
Synthesis and characterization of GNP-NKCT1—
Preparation of GNP-NKCT1 through sodium
borohydrate reduction method by using NKCT1,
GNP, PEG, resulted a light purple coloured colloidal
solution, stable at room temperature (25±2 °C) and
pH 7.2. Hydrodynamic size of GNP-NKCT1 was
determined by DLS was 68–122 nm with average of
92 nm10
. TEM image analysis confirmed the actual
size of GNP as 5-25 nm, where GNPs were arranged
within the protein (NKCT1) core (Fig. 1).

INDIAN J EXP BIOL, AUGUST 2014
766
Anti-arthritic activity of GNP-NKCT1
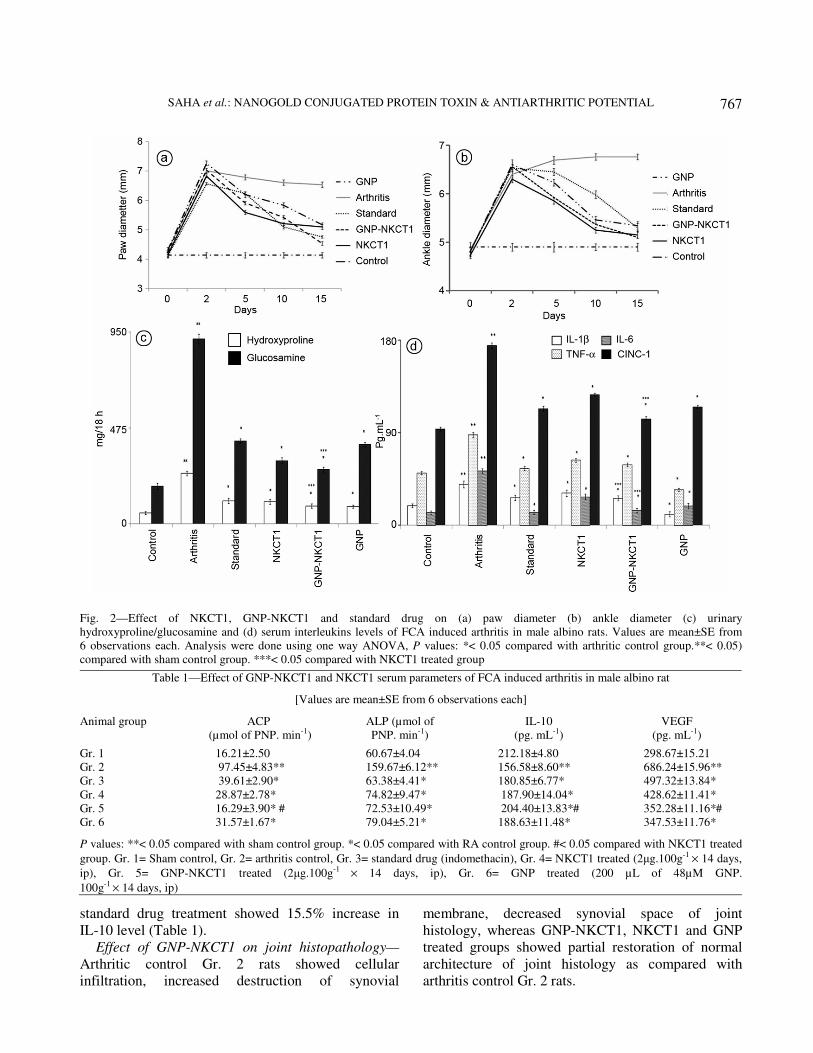
Physical parameters—Induction of RA resulted
significant increase of the paw and ankle diameter in
respect to sham control rats Gr. 1 rats (4.14±0.18 mm
and 4.91±0.13 mm). GNP-NKCT1 treatment
significantly decreased paw and ankle diameter up to
4.54±0.17 mm and 5.12±0.09 mm respectively;
NKCT1 treatment significantly decreased paw and
ankle diameter up to 5.17±0.12 mm and 5.15±0.16 mm
respectively; GNP treatment significantly decreased
paw and ankle diameter up to 5.23±0.22 mm and
5.33±0.13 mm respectively as compared to arthritic
control Gr. 2 rats (6.56±0.24 mm and 6.76±0.21 mm)
after day 15, whereas standard drug treatment
significantly decreased paw and ankle diameter up to
4.76±0.13 mm and 5.3±0.12 mm. (Fig. 2a and b).
Effect of GNP-NKCT1 on urinary parameters—
GNP-NKCT1 treatment significantly decreased
hydroxyproline (64.67%) and glucosamine (70.57%)
levels; NKCT1 treatment significantly decreased
hydroxyproline (56.18%) and glucosamine (65.87%)
levels; GNP significantly decreased hydroxyproline
(65.93%) and glucosamine (57.06%) levels as
compared with arthritic control Gr. 2 rats, whereas
standard drug treatment significantly decreased
54.32% hydroxyproline and 55.26% glucosamine
level (Fig. 2c).
Effect of GNP-NKCT1 on serum parameters—
GNP-NKCT1 treatment significantly decreased ACP
(83.28%) and ALP (54.57%) levels; NKCT1
treatment significantly decreased ACP (70.37%) and
ALP (53.14%) levels; GNP treatment significantly
decreased ACP (67.6%) and ALP (50.49%) levels as
compared to arthritic control Gr. 2 rats, whereas
standard drug treatment significantly decreased
59.35% ACP and 60.01% ALP level (Table 1).
Effect of GNP-NKCT1 on serum cytokines—GNP-
NKCT1 treatment significantly decreased TNF-α
(33.12%), IL-1β (34.06%), IL-6 (71.79%), CINC1
(40.76%) and VEGF (48.66%) level; NKCT1
treatment significantly decreased TNF-α (28.23%),
IL-1β (21.97%), IL-6 (47.39%), CINC1 (27.29%) and
VEGF (29.69%) level; GNP treatment significantly
decreased TNF-α (21.54%), IL-1β (19.18%), IL-6
(51.32%), CINC1 (34.13%) and VEGF (49.35%)
level as compared to arthritic control Gr. 2 rats,
whereas standard drug treatment significantly
decreased 37.17% TNF-α, 32.47% IL-1β, 75.23%
IL-6, 35.18% CINC1 and 27.53% VEGF level
(Fig. 2d, Table 1).
GNP-NKCT1, NKCT1 and GNP treatment
significantly increased serum anti-inflammatory
cytokine IL-10 level (30.54, 20 and 20.46%) as
compared with arthritis control Gr. 2 rats, whereas
Fig. 1—Transmission electron microscopy and DLS study of
(a) only GNP, size 2–8 nm with average size of 4 nm (b) GNP-
NKCT1 where GNP were arranged within protein core, size was
of 68-122 nm with an average of 92 nm
SAHA et al.: NANOGOLD CONJUGATED PROTEIN TOXIN & ANTIARTHRITIC POTENTIAL
767
standard drug treatment showed 15.5% increase in
IL-10 level (Table 1).
Effect of GNP-NKCT1 on joint histopathology—
Arthritic control Gr. 2 rats showed cellular
infiltration, increased destruction of synovial
membrane, decreased synovial space of joint
histology, whereas GNP-NKCT1, NKCT1 and GNP
treated groups showed partial restoration of normal
architecture of joint histology as compared with
arthritis control Gr. 2 rats.
Table 1—Effect of GNP-NKCT1 and NKCT1 serum parameters of FCA induced arthritis in male albino rat
[Values are mean±SE from 6 observations each]
Animal group ACP
(µmol of PNP. min-1)
ALP (µmol of
PNP. min-1)
IL-10
(pg. mL-1)
VEGF
(pg. mL-1)
Gr. 1 16.21±2.50 60.67±4.04 212.18±4.80 298.67±15.21
Gr. 2 97.45±4.83** 159.67±6.12** 156.58±8.60** 686.24±15.96**
Gr. 3 39.61±2.90* 63.38±4.41* 180.85±6.77* 497.32±13.84*
Gr. 4 28.87±2.78* 74.82±9.47* 187.90±14.04* 428.62±11.41*
Gr. 5 16.29±3.90* # 72.53±10.49* 204.40±13.83*# 352.28±11.16*#
Gr. 6 31.57±1.67* 79.04±5.21* 188.63±11.48* 347.53±11.76*
P values: **< 0.05 compared with sham control group. *< 0.05 compared with RA control group. #< 0.05 compared with NKCT1 treated
group. Gr. 1= Sham control, Gr. 2= arthritis control, Gr. 3= standard drug (indomethacin), Gr. 4= NKCT1 treated (2µg.100g-1 × 14 days,
ip), Gr. 5= GNP-NKCT1 treated (2µg.100g-1 × 14 days, ip), Gr. 6= GNP treated (200 µL of 48µM GNP.
100g-1 × 14 days, ip)
Fig. 2—Effect of NKCT1, GNP-NKCT1 and standard drug on (a) paw diameter (b) ankle diameter (c) urinary
hydroxyproline/glucosamine and (d) serum interleukins levels of FCA induced arthritis in male albino rats. Values are mean±SE from
6 observations each. Analysis were done using one way ANOVA, P values: *< 0.05 compared with arthritic control group.**< 0.05)
compared with sham control group. ***< 0.05 compared with NKCT1 treated group

INDIAN J EXP BIOL, AUGUST 2014
768
Acute toxicity of GNP-NKCT1 Determination of minimum lethal dose (MLD)—
The minimum lethal dose (MLD) of GNP-NKCT1
and NKCT1 was found to be 3 and 1.5 mg.k
-1 (sc)
respectively in Swiss albino male mice.
No animal mortality was recorded after 24 h of
treatment. Fecal consistency, food and water intake
were normal in all three groups. After 24 h of
treatment, urine strip test revealed no change in
urinary glucose, bilirubin, ketone, specific gravity,
blood, pH, urobilinogen in GNP-NKCT1 and NKCT1
treated mice. However, NKCT1 treated Gr. 2 mice
urine showed presence of protein (+++) as compared
with GNP-NKCT1 treated Gr. 3 mice urine protein
(+) (Fig. 3a).
Effect on hematological parameters—A significant
increase in WBC count was observed in NKCT1
(80.21%) and GNP-NKCT1 (40.6%) treated mice,
when compared with sham control Gr. 1 mice. No
significant change in total RBC count and Hb% was
observed in NKCT1 and GNP-NKCT1 treated group
of mice as compared to sham control Gr. 1 mice.
Serum biochemical parameters—There was a
significant increase in serum creatinine, urea, CK-MB
and LDH (136.1, 154.9, 75.14 and 51.2%) due to
NKCT1 treated Gr. 2 mice as compared with sham
control Gr. 1 mice. GNP-NKCT1 treatment also
significantly increased serum urea, CK-MB and LDH
levels (46.2, 34.28 and 16.6%) as compared with sham
control Gr. 1 mice. No significant change in serum
creatinine level was observed after GNP-NKCT1
treatment. (Table 2).
Histopathological changes—NKCT1 treated
kidney histology showed partial tubular and
glomerular necrosis and increased capsular space.
Whereas GNP-NKC1 treatment showed less tubular
and glomerular necrotic effect as compared with
NKCT1 treated kidney tissue (Fig. 3b).
Toxicity studies on human lymphocytes—NKCT1
(2 µg.mL-1
), GNP-NKCT1 (2 µg.mL-1
) treatment
decreased the lymphocyte count (28-30 and 16-18%)
as compared with the standard drug Indomethacine
(44-46%).
Table 2—Effect of GNP-NKCT1 and NKCT1 on serum biochemical
parameters of male albino mice.
[Values are mean±SE from 6 observations each]
Animal
group
Creatinine
( mg.dL-1
)
Urea
( mg.dL-1
)
LDH
(U.L-1
)
CK-MB
(IU.L-1
)
Gr. 1 0.83±0.06 28.04±2.01 149.37±7.56 10.67±1.14
Gr. 2 1.96±0.08* 71.48±5.02* 224.14±10.24* 18.71±6.54*
Gr. 3 0.98±0.03*# 40.76±1.48*# 173.83±7.67*# 14.21±2.78*#
P values: *< 0.05 when compared with sham control group. #< 0.05
when compared with NKCT1 treated group. Gr. 1= sham control,
Gr. 2= NKCT1 treated mice, Gr. 3= GNP-NKCT1 treated mice.
Fig. 3— (a): Effect of GNP-NKCT1 and NKCT1 on male albino
mice urinary protein, analyzed by Multistix urine strip b(i), Blank
strip; b(ii), absence of protein in control mice urine;
b(iii), presence of protein (+) in GNP-NKCT1 treated mice urine;
b(iv), presence of protein (+++) in NKCT1 treated mice urine.
(b): Histopathological changes induced by of GNP-NKCT1 and
NKCT1 on male albino mice kidney. b(i), control mice kidney;
b(ii), GNP-NKCT1 treated mice kidney; b(iii and iv), NKCT1
treated mice kidney. Arrows show the glomerular necrosis and
increased capsular space, indicates the nephro-toxicity of NKCT1.

SAHA et al.: NANOGOLD CONJUGATED PROTEIN TOXIN & ANTIARTHRITIC POTENTIAL
769
Discussion In this study, a cytotoxic snake venom protein
toxin NKCT1 was tagged with gold nanoparticles
with a poly ethylene glycol (PEG) ligand protecting
layer. It had been demonstrated that PEG ligands were
very much effective in preventing nonspecific binding
of peptides, proteins and enzymes to the gold
nanoparticle surfaces25
. PEG had been extensively
used as a biomaterial because of its low toxicity, low
immunogenicity26
. PEG coated GNP with functional
carboxyl groups on their surface was formed and
allowed for further controlled conjugation with
NKCT1 that avoided peptide-peptide interaction and
aggregation within the nanoparticles. It was
previously reported that, GNP up to 100 nm size was
easily taken inside the cells and trapped within the
vesicles in cytoplasm27
. The hydrodynamic size of
GNP-NKCT1 obtained by DLS was 68-122 nm with
average of 92 nm, confirmed the effective size of
GNP-NKCT1 for cellular uptake. During the
assessment of anti-rheumatic activity, GNP-NKCT1
(total 28 µg of NKCT1 and 26.5 µg of GNP) was
administrated per rat during the treatment period. Paw
and ankle swelling of FCA induced rats were
significantly reduced after GNP-NKCT1 treatment as
compared with the arthritic control Gr. 2 rats. It was
reported that infiltrated pro-inflammatory cells like
neutrophils attach to the cartilage and invade the
cartilage matrix by its lysosomal enzymes and
oxygen-derived reactive species28
. It was very likely
that the inhibition of hind paw and ankle swellings by
GNP-NKCT1 treatment was due to inhibition of pro-
inflammatory cells infiltration and reducing tissue
destruction. It was further supported by the reduction
of chemo attractant CINC-1 level by GNP-NKCT1
treatment in arthritic rats.
Collagen and cartilage matrix glycoproteins having
high content of hydroxyproline and glucosamine.
Catabolism of insoluble collagen and cartilage matrix
glycoproteins is the major source of urinary
hydroxyproline and glucosamine29
that increases or
decreases in a number of clinical conditions30
including Paget's disease of bone, carcinoma with
metastasis in bone, osteoporosis, arthritis etc31,32,3
.
Increased activities of matrix metalloproteinease
enzymes and collagen degradation were responsible
for the elevated levels of hydroxyproline in arthritic
rat urine33
. Significant restoration of urinary
hydroxyproline and glucosamine by GNP-NKCT1
treatment indicated that GNP-NKCT1 provided
condroprotection by inhibiting the degradation of
collagen and cartilage matrix. Acid and alkaline
phosphatases (ACP and ALP) activity is a measure of
lysosomal integrity. In adjuvant-induced arthritis and
inflammatiory disorders, an increase of ACP and ALP
level was observed due to their release from the
degraded cell lysosomes in the area of inflammation,
which facilitated the degradation of extra cellular
matrix of synovium3. GNP-NKCT1 treatment
significantly decreased the ACP and ALP levels in
FCA induced rats, which indicated that GNP-NKCT1
possess the potential to restore the damages of
lysosomal membrane integrity.
In animal models, inhibition of TNF-α result in
decreased inflammation, while inhibition of IL-1β
effectively prevented cartilage destruction34
.
Similarly, IL-6-deficient mice did not develop bone
erosion35
. Therapeutic inhibition of IL-6 may reduce
generalized osteoporosis, joint inflammation and local
joint destruction in RA36
. The direct involvement of
TNF-α and other pro-inflammatory cytokines in the
development and maintenance of RA suggested that
blocking of their expression in the synovial tissue
could be beneficial for therapy. TNF-α and other
cytokine synthesis is known to be regulated at both
the transcriptional and post transcriptional level. It
was observed that GNP-NKCT1 treatment
significantly reduced the pro-inflammatory cytokines
like TNF-α, IL-1β, IL-6 in respect to arthritic control
rats. Synthesis of those cytokines were regulated in
the transcription level by transcription factor NF-kβ in
pathogenesis of cancer and inflammatory disorders37
.
It was observed that GNP-NKCT1 inhibited the NF-
kβ activation in cancer cells (Gomes et al,
unpublished data). Therefore, antiarthritic property of
the GNP-NKCT1 may attributed to its’ indirect mode
of action, mainly by blocking the upstream regulation
of NF-kβ signaling pathway and synthesis of pro-
inflammatory cytokines. CINC-1, a pro-inflammatory
cytokine promotes recruitment of neutrophil in the
affected joints and helps to progress inflammation38
. It
was also observed that GNP-NKCT1 treatment
antagonized the action of CINC-1. An increased
production of IL-10 by non-T cells was reported in
patients with RA39,40
. There was an initial burst of
pro-inflammatory cytokines (TNF-α, IL1, IL6)
followed by a rise in IL10 synthesis since IL10
production requires both the synthesis of TNF-α and
IL141
. It was found that GNP-NKCT1 treatment
significantly increased the anti-inflammatory cytokine

INDIAN J EXP BIOL, AUGUST 2014
770
IL-10 level in arthritic rats. It may contribute to
diminish T cell mediate inflammation and increased
antibody production in RA.
Beside the onset of inflammation, angiogenesis is a
very important event that provides the vascular
network, necessary to sustain synovial and immune
cell proliferation in RA. VEGF, the key regulatory
molecule of angiogenesis, induced the proliferation
and migration of vascular endothelial cells during the
progression of new blood vessels formation in RA42
.
It was observed that GNP-NKCT1 restored the serum
VEGF level, which was significantly increased in
arthritic rat. Significant reduction of serum VEGF
level by GNP-NKCT1 treatment in arthritic rats
indicate it’s antiangiogenic property, that may
attributed due to the presence of GNP, because GNP
treatment itself significantly reduced the VEGF level
in arthritic rats. Joint histology showed efficacy of
GNP-NKCT1 for partial restoration of synovial space
and the normal joint architecture.
Debnath et al.18
showed that NKCT1 shared
toxicity along with the anticancer activity and the
toxicity being one of the major drawback of this
molecule. One of the major target of the present
study, how to minimize the toxicity of NKCT1?
Whether nanogold conjugation would be able to alter
the toxicity of NKCT1? The toxicity data on GNP-
NKCT1 clearly showed that the toxicity of NKCT1
was reduced due to nanogold conjugation as observed
by the reduced lethal action in mice. In the acute
toxicity studies, several biochemical markers of major
organs (heart, kidney) were increased due to NKCT1
treatment, which were significantly low in GNP-
NKCT1 treated animals. NKCT1 was nephrotoxic in
nature as showed by the presence of protein (+++) in
the urine of NKCT1 treated animals. Low amount of
protein (+) in the urine of GNP-NKCT1 treated
animals confirmed that the nephrotoxic activity of
NKCT1 was minimized by nanogold conjugation thus
making the molecule less toxic. Histopathological
observation of the kidney also confirmed the above
findings.
NKCT1 induced myotoxicity markers (CK-MB,
LDH) were significantly low in GNP-NKCT1 treated
mice, thus the myotoxic action of NKCT1 was also
reduced by nanogold conjugation. In vitro culture of
lymphocyte showed that nanogold conjugated
NKCT1 was less toxic than the NKCT1 and
indomethacine. It is likely that gold conjugation
masked the protein part of NKCT1 responsible for the
toxicity manifestation or inactivated the toxic moiety
of NKCT1 by certain chemical bonding. Tagging of
GNPs with protein/peptide prolongs the circulation
time, thus offering an increased probability of
accumulation in the abnormal cells and targeted
binding to sites of angiogenesis or infection12
. The
delivery of conjugated protein/peptide is small and
decreased in nonspecific targeting attributable to the
use of GNPs. Moreover, pegylation of GNPs
prevented their interaction and uptake by normal
lymphocytic cell or endothelial cell14,10
. Fortuitously
extra loaded GNPs that do not reach their targets are
accumulated and deactivated in the liver12
, while
thermodynamically favorable GNP-NKCT1 (neutral
GNP charged with cationic protein) reach their targets
and engulfed by enhanced phagocytotic abnormal
cells that leads to cell death.
In the present study it was observed that, urinary
serum biochemical markers, pro-inflammatory
cytokines levels were significantly reduced after
GNP-NKCT1 and NKCT1 treatment. However the
levels of those arthritic markers (hydroxiproline,
glucosamine, ACP, IL-1B, IL-6, CINC1, VEGF) were
significantly low in GNP conjugated NKCT1 treated
rats than that of unconjugated NKCT1 treated rats.
GNP-NKCT1 significantly increased in anti-
inflammatory cytokines IL-10 levels than that of
unconjugated NKCT1 treatment. Furthermore present
study also showed that GNP conjugation reduced the
toxicity profile of NKCT1 along with increasing it’s
antiarthritic efficiency. Serum level of tissue injury
markers (creatinine, urea, LDH, CK-MB) were found
to be significantly low in GNP-NKCT1 treated mice
than that of NKCT1 treated mice group. Recently it
had been reported that GNP conjugation reduces the
cardio, neuro and nephrotoxic limitations of the
NKCT143
. Therefore GNP conjugation not only
increased the antiarthritic property of NKCT1 but also
beneficial for the reduction of the toxicity. Further
detail studies on this area are warranted for
understanding the mechanism of reduction of toxicity
and antiarthritic potential of GNP-NKCT1 for
biomedical application in the near future.
Conclusion This study confirmed a successful conjugation of
gold nanoparticle with NKCT1, a cytotoxin from
Naja kaouthia snake venom which showed increased
antiarthritic potential and lower toxicity profile
through in vivo and in vitro experiments. Further

SAHA et al.: NANOGOLD CONJUGATED PROTEIN TOXIN & ANTIARTHRITIC POTENTIAL
771
studies on the molecular mechanism of anti-arthritic
activities of GNP-NKCT1 and its toxicity are in
progress.
Conflict of interest
No conflict of interest exist among the authors.
Acknowledgement This research work was sponsored by Department
of Biotechnology, Govt of India, New Delhi
(Ref. no. BT/PR14811/NNT/28/500/2010).
Reference 1 Pal S K, Gomes A, Dasgupta S C & Gomes A, Snake venom
as therapeutic agents: From toxins to drug development,
Indian J Exp Biol, 40 (2002) 1353.
2 Das T, Bhattacharya S, Haldar B, Biswas A, Dasgupta S,
Gomes A & Gomes A, Cytotoxic and antioxidant property of
a purified fraction (NN-32) of Indian Naja naja venom on
Ehrlich ascites carcinoma in BALB/c mice, Toxicon, 57
(2011) 1065.
3 Gomes A, Bhattacharjee P, Mishra R, Biswas A K,
Dasgupta S C & Giri B, Anticancer potential of animal
venoms and toxins, Indian J Exp Biol, 48 (2010) 93.
4 Sherman D G, Atkinson R P, Chippendale T, Levin K A,
Ng K, Futrell N, Hsu C Y & Levy D E, Intravenous Ancrod
for treatment of acute ischemic stroke: the STAT study: A
randomized controlled trial Stroke Treatment with Ancrod
Trial, JAMA, 283 (2000) 2461.
5 Koh D C I, Armugam A & Jeyaseelan K, Snake venom
components and their applications in biomedicine, CMLS, 63
(2006) 3030.
6 Gomes A, Bhattacharya S, Chakraborty M, Bhattacharjee P
& Mishra R, Anti-arthritic activity of Indian monocellate
cobra (Naja kaouthia) venom on adjuvant induced arthritis,
Toxicon, 55 (2010) 670.
7 Chen R & Robinson S E, The effect of cholinergic
manipulations on the analgesic response to cobrotoxin in
mice, Life Sci, 47 (1990) 1949.
8 Kourounakis L K, Nelson R A & Kupusta M A, The effect of
a cobra venom factor on complement and adjuvant induced
disease in rats, Arthritis Rheum, 16 (1973) 71.
9 Freitas T V & Frézard F, Encapsulation of native crotoxin in
liposomes: A safe approach for the production of antivenom
and vaccination against Crotalus durissus terrificus venom,
Toxicon, 35(1) (1997) 91
10 Bhowmik T, Saha P P, Dasgupta A & Gomes A,
Antileukemic potential of PEGylated Gold nanoparticle
conjugated with protein toxin (NKCT1) isolated from Indian
cobra (Naja kaouthia) venom, Cancer Nano, 4 (2013) 39
11 Biswas A, Gomes A, Sengupta J, Datta P, Singha S,
Dasgupta A K & Gomes A, Nanoparticle-conjugated animal
venom-toxins and their possible therapeutic potential,
J Venom Res, 3 (2012) 15.
12 Soman N R, Baldwin S L, Hu G, Marsh J N, Lanza G M,
Heuser J E, Arbeit J M, Wickline S A & Schlesinger P H,
Molecularly targeted nanocarriers deliver the cytolytic
peptide melittin specifically to tumor cells in mice, reducing
tumor growth, J Clin Invest, 119 (2009) 2830.
13 Al-Sadoon M K, Abdel-Maksoud M A, Rabah D M &
Badr G, Induction of apoptosis and growth arrest in human
breast carcinoma cells by a snake (Walterinnesia aegyptia)
venom combined with silica nanoparticles: crosstalk between
Bcl2 and caspase 3, Cell Physiol Biochem, 30 (2012) 653.
14 Hood J L, Jallouk A P, Campbell N, Ratner L & Wickline S A
Cytolytic nanoparticles attenuate HIV-1 infectivity, Antiviral
Therapy, 18 (2013) 95.
15 Warnasooriya N, Joud F, Bun P, Tessier G, Coppey-Moisan M,
Desbiolles P, Atlan M, Abboud M & Gross M, Imaging gold
nanoparticles in living cell environments using heterodyne
digitalholographic microscopy, Opt Express, 18 (2010) 3264.
16 Dreaden E C, Alkinaly A M, Huang X, Murphy C J &
El-Sayed M A, The golden age: gold nanoparticles for
biomedicine, Chem Soc Rev, 41 (2012) 2740
17 Lowry O H, Rosebrough N J, Farr A L & Randall R J,
Protein measurement with the Folin phenol reagent, J Biol
Chem, 193 (1951) 265.
18 Debnath A, Saha A, Gomes A, Biswas S, Chakraborty P,
Giri B, Biswas A K, Das Gupta S & Gomes A, A lethal
cardiotoxic–cytotoxic protein from the Indian monocellate
cobra (Naja kaouthia) venom, Toxicon, 56 (2010) 569.
19 Laemmli U K, Cleavage of Structural Proteins during the
Assembly of the Head of Bacteriophage T4, Nature, 227
(1970) 680.
20 Samal A K, Sreeprasad T S & Praddep T, Investigation of
role of NaBH4 in the the chemical synthesis of gold
nanorods, J Nanopart Res, 12 (2010) 1777.
21 Newbould B B, Chemotherapy of arthritis induced in rats of
mycobacteria adjuvant, Br J Pharmacol, 21 (1963) 127.
22 Neuman R E & Logan M A, The determination of
hydroxyproline, J Biol Chem, 184 (1950) 299
23 Elson L A & Morgan W T, A colorimetric method for the
determination of glucosamine and chondrosamine, Biochem
J, 27 (1933) 1824.
24 Mitchell R F, Karnovsky M J & Karnovsky M L The
distribution of some granule associated enzymes in guineapig
polymorphonuclear leucocytes, Biochem J, 116 (1970) 207.
25 Harder P, Grunze M, Dahint R, Whitesides G M &
Laibinis P E, Molecular Conformation in Oligo (ethylene
glycol)-Terminated Self-Assembled Monolayers on Gold and
Silver Surfaces Determines Their Ability To Resist Protein
Adsorption, J Phys Chem B, 102 (1998) 426.
26 Zhu B, Eurell T, Gunawan R & Leckband D, Chain-length
dependence of the protein and cell resistance of
oligo(ethylene glycol)-terminated self-assembled monolayers
on gold, J Biomed Mater Res, 56 (2001) 406.
27 Chithrani B D, Ghazani A A & Chan W C W, Determining
the size and shape dependence of gold nanoparticle uptake
into mammalian cells, Nano Lett, 6 (2006) 662.
28 Kowanko I C, Bates E J & Ferrante A, Mechanisms of
human neutrophii mediated cartilage damage in vitro: The
role of lysosomal enzymes, hydrogen peroxide and
hypochlorous acid, Immunol Cell Biol, 67 (1989) 321.
29 Prockop D J, Ebert P S & Shapiro B M, Studies with
porline-3,4,-H3 on the hydroxylation of proline during
collagen synthesis in chick embryos, Arch Biochem Biophys,
106 (1964) 112.
30 Weiss P H & Klein L, The quantitative relationship of
urinary peptide hydroxyproline excretion to collagen
degradation, J Clin Invest, 48 (1969) 1.

INDIAN J EXP BIOL, AUGUST 2014
772
31 Scarpignato C, NSAID-induced intestinal damage: Are
luminal bacteria the therapeutic target? Gut, 57 (2008) 145.
32 Halder S, Dasgupta S, Gomes A, Giri B, Dasgupta S C,
Biswas A, Mishra A & Gomes A, A high molecular weight
protein Bengalin from the Indian black scorpion
(Heterometrus bengalensis C.L. Koch) venom having anti –
osteoporosis activity in female albino rats, Toxicon, 55
(2010) 455.
33 Reddy G K & Dhar S C, Metabolism of collagen in bone of
adjuvant induced arthritic rat, Bone, 10 (1989) 439.
34 Probert L, Plows D, Kontogeorgos G & Kollias G, The type I
interleukin-1 receptor acts in series with tumor necrosis
factor (TNF) to induce arthritis in TNF-transgenic mice, Eur
J Immunol, 25 (1995) 1794.
35 Boe A, Baiocchi M, Carbonatto M, Papoian R &
Serlupi-Crescenzi O, Interleukin 6 knockout mice are
resistant to antigen-induced experimental arthritis,
Eur Cytokine Netw, 11(1999) 1057.
36 Wong P K K, Quinn J M W, Sims N A, Nieuwenhuijze A,
Campbell I K & Wicks I P, Interleukin-6 modulates
production of T lymphocyte–derived cytokines in antigen-
induced arthritis and drives inflammation-induced
osteoclastogenesis, Arthritis Rheum, 54 (2006) 158.
37 Elinav E, Nowarksi R, Thaiss C A, Hu B, Jin C & Flavell A,
Inflammation-induced cancer: Crosstalk between tumours,
immune cells and microorganisms, Nature Rev, 13 (2013)
759.
38 Takano K & Nakagawa H, Contribution of cytokine-induced
neutrophil chemoattractant CINC-2 and CINC-3 to
neutronphil recruitment in lipopolysaccharide-induced
inflammation in rats, Inflammation Res, 50 (2001) 503.
39 Llorente L, Patin Y R, Fior R, Varela J A, Wijdenes J,
Fourrier B M, Galanaud P & Emilie D, In vivo production of
interleukin-10 by non–t cells in rheumatoid arthritis,
sjöugren's syndrome, and systemic lupus erythematosus,
Arthritis Rheum, 37 (1994) 1647.
40 Alanärä T, Karstila K, Moilanen T, Silvennoinen N &
Isomäki P, Expression of IL-10 family cytokines in
rheumatoid arthritis: Elevated levels of IL-19 in the joints,
Scand J Rheumatol, 39 (2010) 118.
41 Clair E W S, Interleukin 10 treatment for rheumatoid
arthritis, Ann Rheum Dis, 58 (1999) 199.
42 Silva L C R, Ortigosa L C M & Benard G, Anti-TNF-α
Agents in the Treatment of Immune-mediated Inflammatory
Diseases: Mechanisms of action and pitfalls, Immunotherapy,
2 (2010) 817.
43 Saha P P, Bhowmik T, Dasgupta A K & Gomes A,
In vivo and in vitro toxicity of nanogold conjugated snake
venom protein toxin GNP-NKCT1, Toxicol Rep, 1(2014)74.
Related Documents











