Mutational analysis and homology modelling of SyrC, the aminoacyltransferase in the biosynthesis of syringomycin Maria Rosaria Fullone a , Alessandro Paiardini a , Dennis C. Gross b , Shi-En Lu c , Alberto Fiore d , Ingeborg Grgurina a, * a Department of Biochemical Sciences ‘‘A. Rossi Fanelli’’, University of Rome ‘‘La Sapienza’’, P.le A. Moro, 5, 00185 Rome, Italy b Department of Plant Pathology and Microbiology, Texas A&M University, 2132 TAMU, College Station, TX, USA c Department of Entomology and Plant Pathology, Mississippi State University, Mississippi State, MS, USA d Dipartimento di Scienza degli Alimenti, Universita ` di Napoli ‘‘Federico II’’, 80055 Portici, Naples, Italy Received 10 September 2007 Available online 4 October 2007 Abstract SyrC, a component of the multienzyme system of syringomycin biosynthesis, has been shown to shuttle Thr/4-Cl-Thr between the thiolation domains SyrB1-T1 and SyrE-T8,9 by transiently linking it to Cys224 in the enzyme active site. We present data on the struc- ture-function relationship in vivo of this protein and an in silico model of its three-dimensional structure. The biosynthetic activity of SyrC was not influenced when either Asp348 or His376 that together with Cys224 form a putative catalytic triad, were replaced with Ala, but it was abolished by the exchange Cys224 with Ser. The presence of the FLAG peptide on either the N- or C-terminus of the protein did not affect activity, whereas the deletion of the first 16 amino acids at the N-terminus or the insertion of Maltose Binding Protein abolished the production of syringomycin. We present the model of the three-dimensional structure of SyrC suggesting a homodimeric structure for the protein and biochemical data that are supportive of this model. Ó 2007 Elsevier Inc. All rights reserved. Keywords: Syringomycin; SyrC; Aminoacyltransferase; Mutagenesis; Structure–activity relationship; Homology modelling Syringomycin (SRE) is a virulence-conferring and anti- fungal lipodepsinonapeptide produced by many strains of the plant pathogenic bacterium Pseudomonas syringae pv. syringae [1]. It is synthesized by a nonribosomal mecha- nism of peptide synthesis (NRPS) [2], whose machinery and mechanism were recently reviewed [3]. The biosyn- thetic cluster of syringomycin [4], composed of three tran- scriptional units, syrB1/syrB2, syrC and syrE, is a prototype of the ‘‘non linear’’ class of NRPS clusters. In fact, the C-terminal residue in SRE, 4-Cl-L-Thr, is formed by enzymes encoded at the 5 0 -end of the cluster, SyrB1 and SyrB2. SyrB1 is a didomain adenylation-thiolation (A-T1) module which activates L-threonine that is then chlorinated by the halogenase SyrB2 [5]. SyrE is an eight-modules-con- taining synthetase involved in the formation of the octa- peptide in SRE. Its C-terminal portion contains a condensation (C9) and a thiolation (T9) domain which sep- arate the SyrE8 module from the thioesterase domain. The gene syrC, located between syrB1/B2 and syrE, encodes an a,b-hydrolase which was shown to mediate the transfer of 4-Cl-L-Thr from the SyrB1-T1 domain to SyrE-T8,9 domains by transthioesterification [6]. SyrC belongs to the growing family of free-standing proteins, operating within some NRPS and mixed biosynthetic pathways, that shuttle intermediates in the course of the construction of a metabolite, as exemplified by CmaE [7], CouN7 [8] and CloN7 [9] in the biosynthesis of coronatine, coumermycin, and clorobiocin, respectively. In the present study we investigated the structure–func- tion relationship of the SyrC protein by a functional com- plementation approach and its three-dimensional structure by in silico homology modelling. 0006-291X/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2007.09.116 * Corresponding author. Fax: +39 0649917566. E-mail address: [email protected] (I. Grgurina). www.elsevier.com/locate/ybbrc Available online at www.sciencedirect.com Biochemical and Biophysical Research Communications 364 (2007) 201–207

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 364 (2007) 201–207
Mutational analysis and homology modelling of SyrC,the aminoacyltransferase in the biosynthesis of syringomycin
Maria Rosaria Fullone a, Alessandro Paiardini a, Dennis C. Gross b, Shi-En Lu c,Alberto Fiore d, Ingeborg Grgurina a,*
a Department of Biochemical Sciences ‘‘A. Rossi Fanelli’’, University of Rome ‘‘La Sapienza’’, P.le A. Moro, 5, 00185 Rome, Italyb Department of Plant Pathology and Microbiology, Texas A&M University, 2132 TAMU, College Station, TX, USA
c Department of Entomology and Plant Pathology, Mississippi State University, Mississippi State, MS, USAd Dipartimento di Scienza degli Alimenti, Universita di Napoli ‘‘Federico II’’, 80055 Portici, Naples, Italy
Received 10 September 2007Available online 4 October 2007
Abstract
SyrC, a component of the multienzyme system of syringomycin biosynthesis, has been shown to shuttle Thr/4-Cl-Thr between thethiolation domains SyrB1-T1 and SyrE-T8,9 by transiently linking it to Cys224 in the enzyme active site. We present data on the struc-ture-function relationship in vivo of this protein and an in silico model of its three-dimensional structure. The biosynthetic activity ofSyrC was not influenced when either Asp348 or His376 that together with Cys224 form a putative catalytic triad, were replaced withAla, but it was abolished by the exchange Cys224 with Ser. The presence of the FLAG peptide on either the N- or C-terminus of theprotein did not affect activity, whereas the deletion of the first 16 amino acids at the N-terminus or the insertion of Maltose BindingProtein abolished the production of syringomycin. We present the model of the three-dimensional structure of SyrC suggesting ahomodimeric structure for the protein and biochemical data that are supportive of this model.� 2007 Elsevier Inc. All rights reserved.
Keywords: Syringomycin; SyrC; Aminoacyltransferase; Mutagenesis; Structure–activity relationship; Homology modelling
Syringomycin (SRE) is a virulence-conferring and anti-fungal lipodepsinonapeptide produced by many strains ofthe plant pathogenic bacterium Pseudomonas syringae pv.syringae [1]. It is synthesized by a nonribosomal mecha-nism of peptide synthesis (NRPS) [2], whose machineryand mechanism were recently reviewed [3]. The biosyn-thetic cluster of syringomycin [4], composed of three tran-scriptional units, syrB1/syrB2, syrC and syrE, is aprototype of the ‘‘non linear’’ class of NRPS clusters. Infact, the C-terminal residue in SRE, 4-Cl-L-Thr, is formedby enzymes encoded at the 5 0-end of the cluster, SyrB1 andSyrB2. SyrB1 is a didomain adenylation-thiolation (A-T1)module which activates L-threonine that is then chlorinatedby the halogenase SyrB2 [5]. SyrE is an eight-modules-con-
0006-291X/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2007.09.116
* Corresponding author. Fax: +39 0649917566.E-mail address: [email protected] (I. Grgurina).
taining synthetase involved in the formation of the octa-peptide in SRE. Its C-terminal portion contains acondensation (C9) and a thiolation (T9) domain which sep-arate the SyrE8 module from the thioesterase domain. Thegene syrC, located between syrB1/B2 and syrE, encodes ana,b-hydrolase which was shown to mediate the transfer of4-Cl-L-Thr from the SyrB1-T1 domain to SyrE-T8,9domains by transthioesterification [6]. SyrC belongs tothe growing family of free-standing proteins, operatingwithin some NRPS and mixed biosynthetic pathways, thatshuttle intermediates in the course of the construction of ametabolite, as exemplified by CmaE [7], CouN7 [8] andCloN7 [9] in the biosynthesis of coronatine, coumermycin,and clorobiocin, respectively.
In the present study we investigated the structure–func-tion relationship of the SyrC protein by a functional com-plementation approach and its three-dimensional structureby in silico homology modelling.

202 M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207
Materials and methods
Growth of microorganisms. Strains of P. s. pv. syringae were grown at25 �C on SRM-C or PDB (potato dextrose broth) medium as described[2,10]. Escherichia coli strains DH10B and BL21(DE3) used for plasmidpropagation and protein expression, were grown in LB liquid medium at37 �C and 25 �C, respectively.
Microbiological assay. Antimicrobial assays were performed withRhodotorula pilimanae as described [10].
Construction of a syrC-recA mutant. To assure stable maintenance ofoverexpression constructs in syrC mutant strain BR334 [11], plasmidpBR325 [12] carrying a 9-kb BamHI fragment of pEMH9 [13], whichcontains a recA gene from P. s. pv. syringae B728a mutated by insertion ofan X fragment, was introduced into cells of strain BR334 for markerexchange mutagenesis. The resulting recA mutant BR334A1 was con-firmed by testing for sensitivity to UV light as described previously [13].
Construction of pKm12 plasmid. pKm12 plasmid was prepared in orderto express syrC in P. s. pv. syringae BR334A1 as an N-terminus free SyrCunder the control of an inducible promoter. A fragment containing the last282 bp of lacI, tac promoter and malE gene was removed from pMEKm12[14] by EcoRV and BamHI digestion. A 600 bps fragment containing thelacI (800–1081) and tac promoter was obtained by PCR amplification (for:5 0-GTTGGTGCGGATATCTCGGTAG-3 0 and rev: 5 0-GGATCCTGTTTCCTGTGTGAAATT-3 0) and was cloned into the resultingplasmid in order to obtain the pKm12 plasmid. The construct was con-firmed by restriction analysis and by sequencing.
Construction of pMEKm12syrC, pKm12syrC, pKm12D16syrC,
pKm12syrC-Cflag, and the pKm12syrC-Nflag. For the preparation of theabove constructs the syrC gene was amplified from the p601D-1R [14]plasmid with the specific primers described below. The PCR reaction wasperformed by using Taq Gold (Perkin Elmer) according to the manufac-turer’s instructions. The amplicon was directly cloned into pGEMT(Promega) vector. The resulting plasmid was digested with BamHI andHindIII and the fragment obtained was cloned into pKm12 or pMEKm12previously digested with BamH1 and HindIII. The verification of theresulting plasmids was determined by sequencing. The oligo pairs used inPCR amplification of each construct are reported below. pKm12D16syrC:
D16syrCBamH1-for: 5 0-ggattcATGCTCCAGGCACCT-3 0 and syrCHin-dIII-rev: 50-aagcttTCAGGCGACAGCGGG-30; pMEK12syrC and pKm12syrC:syrCBamH1-for: 5 0-ggatccATGACTATTTCCTCCG-3 0 and syrCHin-dIII-rev (see above); pKm12syrC-Cflag: syrCBamH1-for (see above) andsyrC-CflagHindIII-rev: 5 0-aagcttCACTTGTCGTCGTCGTCCTTGTAGTCGGCG ACAG-3 0; pKm12syrC-Nflag: syrCBamHI-Nflag for: 5 0-ggatccATGTCCGACTACAAGGACGACGACGACAAGATGACTATTTCCTCC-3 0 and syrCHindIII-rev (see above).
SyrC mutant construction. Site directed mutagenesis was performedusing Stratagene’s QuickChange kit. pGEMT vector carrying syrC gene wasused as a template for mutagenesis of C224A, C224S, D348A, H376A,H358A, H352A, H369A. The mutagenic primers were: C224A for: 5 0-ATGGGCATCGCCGGCGGCGCG-3 0, C224A rev:5 0- ACCGCGCCGCCGGCGATGCC-3 0; C224S for: 5 0-ATGGGCATCTCCGGCGGCGC-3 0,C224S rev: 5 0ACCGCGCCGCCGGAGATG-3 0; D348A for: 5 0-GCTGGCGATGCCGCCTCGACTACGC-3 0, D348A rev: 5 0-GATGTGCGTAGTCGAGGCGGCATCG-3 0; H376A for: 5 0-CAA TGGCAGCGC
CCTGGCGTT-3 0, H376A rev: 5 0-AAAGAACGCCAGGGCGCTGCC-3 0; H352A for: 5 0-ACTCGACTACGGCCATCGGCG-3 0, H352A rev: 5 0-AGCCGCCGATGGCCGTAGTCG-30; H358A for: 5 0-CGGCTCACGGCAATCGCCG-30, H358A rev: 5 0-ACGCGGCGATTGCCCGTGAG-3 0; H369A for: 5 0-ACGCCACGCTGGCCGTAGAGC-3 0 H369A rev: 5 0-TGCGCTCTACGGCCAGCGTGG-3 0. Incorporation of mutation wascontrolled by DNA sequencing. Mutated plasmids were digested withBamHI and HindIII and the fragments obtained were subcloned intopKm12. The resulting constructs were electroporated into P. s. pv. syringae
BR334A1.Complementation experiments. The P. s. pv. syringae strains BR334A1
and BR334A1 transformed with pMEKm12 or pKm12 plasmids con-taining the syrC gene or the relative mutated forms, were grown in the
presence of appropriate antibiotics as described [2]. For the constructswhere the induction of gene transcription was required, IPTG (5 mM) wasadded in the growth phase (OD 0.5–0.8). After 6–8 h of induction withshaking (150 rpm), the content of the flask was transferred into a 1500 mlRoux bottle. After 8 days the mixtures of secondary metabolites wereextracted and analysed as described [2].
SDS–PAGE and Western blotting. Samples for immunoblotting wererun on 12% SDS–PAGE and electrotransferred onto PVDF membranes.After 1 h incubation with anti-FLAG M2 peroxidase conjugated anti-bodies (Sigma), immunoreactive proteins were detected with BM BluePOD Substrate (Roche).
Protein expression and purification. Escherichia coli host cells BL21(DE3)transformed with pKm12syrC-Cflag were grown at 25 �C in LB broth withampicillin (100 lg/ml) to A600 = 0.8. After 3 h of induction with 0.05 mMIPTG, cells from 1 L of culture were harvested by centrifugation, resus-pended in 12 ml of 25mM MOPS, 150 mM NaCl, pH 7.4 and lysed by threepassages through a French press at 12000 psi. Cell extracts were clarified bycentrifugation (40 min at 60,000g). SyrC-CFLAG was purified on anti-FLAG M2 affinity gel (Sigma) according to the manufacturer.
Crosslinking of SyrC-CFLAG. A solution of 1 mM bis(sulfosuccinim-idyl)suberate (BS3) (Pierce) in 25 mM MOPS buffer pH 7.5 was freshlyprepared. A 50 ll volume of purified SyrC-CFLAG (2.2 lM) was incu-bated for 2 h at 4 �C together with 44 lM of BS3. Reaction was stopped bythe addition of 1 ll of Tris–HCl 1.5 M. BS3 and control samples wereanalysed by SDS–PAGE followed by immunoblotting with anti-FLAGM2 peroxidase conjugated antibodies.
Mass spectrometry experiments. MALDI-TOF spectra were recordedas described in [15].
Modelling of the three-dimensional structure of SyrC. The choice of asuitable structural template for SyrC, multiple sequence alignment, modelconstruction and evaluation methods are described in details in the Sup-plementary material section.
Results
In trans functional complementation assays of the syrC-
inactivated mutant P. s. pv. syringae strain BR334A1 with
plasmids containing modified forms of the syrC gene
To evaluate the effects of structural modifications of SyrCon its biosynthetic activity in vivo, we adopted the functionalcomplementation approach. Therefore, we generated a syrC
mutant that was also mutated in recA, called BR334A1, andtransformed it with plasmids carrying modified forms of thesyrC gene, whose ability to restore the production of SRE inBR334A1 in liquid culture was evaluated by antifungalassay, RP-HPLC analysis and, in some cases, by mass spec-trometry. As shown in Table 1, pKm12syrC carrying thesyrC gene under tac promoter, restored the capability toproduce SRE. We first assayed the effects of three differentmodifications at the N-terminus. The strain BR334A1 trans-formed with pKm12D16syrC, the construct expressing aSyrC form lacking the first 16 amino acids that was availablein our laboratory, was unable to restore the SRE produc-tion. For some unknown reason, this strain did not growon SRM-C medium but did grow on PDB. AlthoughPDB medium supports the production of P. s. pv. syringaeB301D LDPs [10], no SRE production was observedfor BR334A1(pKm12D16syrC). The construct pMEKm12-syrC, expressing a SyrC protein N-terminally fused to MBP(Maltose Binding Protein), was also unable to complement

Table 1The production of SRE in P. s. pv. syringae BR334A1 strain transformedwith vectors carrying syrC forms modified at 5 0- or 3 0-end
Plasmids used for the transformation of the BR334A1strain
SREproduction
pKm12syrC +pKm12D16syrC �pMEKm12syrC �pKm12syrC-Nflag +pKm12 syrC-Cflag +
For each culture the presence of SRE was estimated by antifungal assayand RP-HPLC analysis.The levels of SRE production/ml of broth culture were evaluated by RP-HPLC [2] and shown to be in the range of 9.0–9.5 lg/ml in all the pro-ducer strains.
M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207 203
the BR334A1 strain. To verify if a less bulky tag wouldallow a functional complementation, we constructed theplasmid pKm12syrC-Nflag expressing an N-FLAG-taggedSyrC form, carrying an octapeptide at the N-terminus,which was as biosynthetically competent as the wild-typestrain. The same result was obtained in the complementa-tion with the pKm12syrC-Cflag where the sequence encod-ing the FLAG octapeptide is translationally fused to theC-terminus of SyrC, showing that the C-terminal FLAGtag did not influence the biosynthetic functionality of theprotein.
The results of the complementation experiments withplasmids carrying the site directed mutants of the SyrCactive site residues are shown in Table 2. In accordancewith the results obtained in vitro [6], the C224A mutantfailed to produce SRE. The substitution of Cys224 withSer also caused the loss of SRE production. Cys224 is anucleophile component of a catalytic triad containing alsoAsp348 and His376 in the SyrC active site, described in thenext section. However, the site directed mutation into Alaof either Asp348 or His376, did not influence the biosyn-thetic activity of SyrC. In fact, the chromatographic profileof the broth extract showed the presence of SRE, as con-firmed by the mass spectrometry analysis. Before the con-struction of the three-dimensional model, we also mademutants where His352 or His358 or His369 were exchangedwith Ala, in an attempt to identify another His residue
Table 2The production of SRE in P. s. pv. syringae BR334A1 strain transformedwith vectors carrying site-directed syrC mutated forms
Plasmids used for the transformation of the BR334A1strain
SREproduction
pKm12syrC C224A �pKm12syrC C224S �pKm12syrC D348A + (MH+
1225.00)pKm12syrC H376A + (MH+
1225.03)pKm12syrC H358A +pKm12syrC H352A +pKm12syrC H369A �
The levels of SRE were evaluated as reported in Table 1.
which could functionally replace His376. The mutation ofthe first two did not affect the production of SRE, indicat-ing that the activity of the protein was not influenced. Con-versely, the mutant H369A was deficient in SREproduction. As later shown by homology modelling,His369 is located at the contact surface between the twosubunits of the putative SyrC homodimeric structure.
Construction of the three-dimensional homology model of
SyrC
In order to obtain insight into the three-dimensionalstructure of the protein and into the structure of the activesite, a homology model of SyrC was constructed using theknown crystal structure of the meta-cleavage producthydrolase (CumD) [16] from Pseudomonas fluorescens asa structural template. Only the region spanning residues148–402 of the primary structure was modelled, since nosuitable structural template was found for the N-terminaldomain. Given the low sequence identity of �25% betweenthe two enzymes, the construction of the model required adetailed structural analysis, mainly based on the conservedelements of secondary structure that were identifiedthrough a multiple alignment of sequence homologs ofSyrC (Fig. 1). The Ca backbone trace of the model isshown in Fig. 2. As expected, the structure exhibits a typ-ical a/b-hydrolase fold, with the core of each subunit com-posed of a b-sheet, with the strands connected by helices.Excluding the N-terminal region, the enzyme is constitutedby two domains: the main, or core domain (residues 125–250, and 320–403) and the lid domain (residues 251–319).A buried cleft connected to the solvent through a tightchannel and located between the core and the lid domainsis predicted to be the active site of the enzyme. Inside thecleft, a catalytic triad typical of this fold, composed ofCys224, Asp348 and His376, is observed (Fig. 3). Theseresidues, invariant in the multiple alignment of the closesthomologs of SyrC (Fig. 1), exhibit a hydrogen-bondbetween the Od2 atom of Asp348 and Nd1 of His376. SyrCshows a wider entrance to the active site, as compared toCumD, which is due to the different identity of the residuessurrounding the channel. These are mainly hydrophobicresidues: Phe161, Phe266, Met270, Gln284, Phe287,Leu288, Leu377, Phe379 and Phe380. It can be envisagedthat the channel allows the entrance of the phosphopant-etheine arm.
Several residues are conserved between SyrC andCumD, at the subunit interface of the latter. These include:Ala366, Leu368, His369, Val370 and Glu396. Therefore, itis anticipated that, like CumD, SyrC could assume adimeric quaternary structure in solution (Fig. 2).
Cross-linking of purified SyrC-CFLAG
To gain support on the homodimeric structure of SyrCcross-linking experiments were carried out. The syrC wasexpressed as a fusion protein with the FLAG-tag at its

Fig. 1. Multiple alignment of CumD, SyrC and its closest homologs. Amino acid one-letter code is used. Dashes represent insertions and deletions;numbers on the left represent sequence numbering. Invariant positions are boxed in black. SyrC sequence of Pseudomonas syringae pv. syringae B301D-Ris reported in ref [6]. The other sequences are labelled with their NCBI identifiers: 24158682, CumD (GenBank Accession No.: 1IUN_A); 66045845, SyrC[Pseudomonas syringae pv. syringae B728a; YP_235686]; 20150036, MupV [Pseudomonas fluorescens; AAM12938]; 67544687, conserved hypotheticalprotein [Burkholderia vietnamiensis G4; ZP_00422614]; 115360793, conserved hypothetical protein [Burkholderia cepacia AMMD; YP_777930];118698367, conserved hypothetical protein [Burkholderia ambifaria MC40-6; ZP_01556444]; 53722205, hypothetical protein BPSS1177 [Burkholderia
pseudomallei K96243; YP_111190]; 67738857, predicted hydrolases or acyltransferases (a,b-hydrolase superfamily) [Burkholderia pseudomallei 668;ZP_00489479]; 83717113, hypothetical protein BTH_II1230 [Burkholderia thailandensis E264; YP_439426].
204 M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207
C-terminus and purified by immunoaffinity chromatogra-phy (Fig. 4A) Protein–protein cross-linking, using theamine-specific cross-linker BS3 was performed and ana-
lysed by Western blotting. As shown in Fig. 4B, besides a45 kDa band corresponding to SyrC-CFLAG, a new bandat 90 kDa, corresponding in size to a SyrC homodimer,
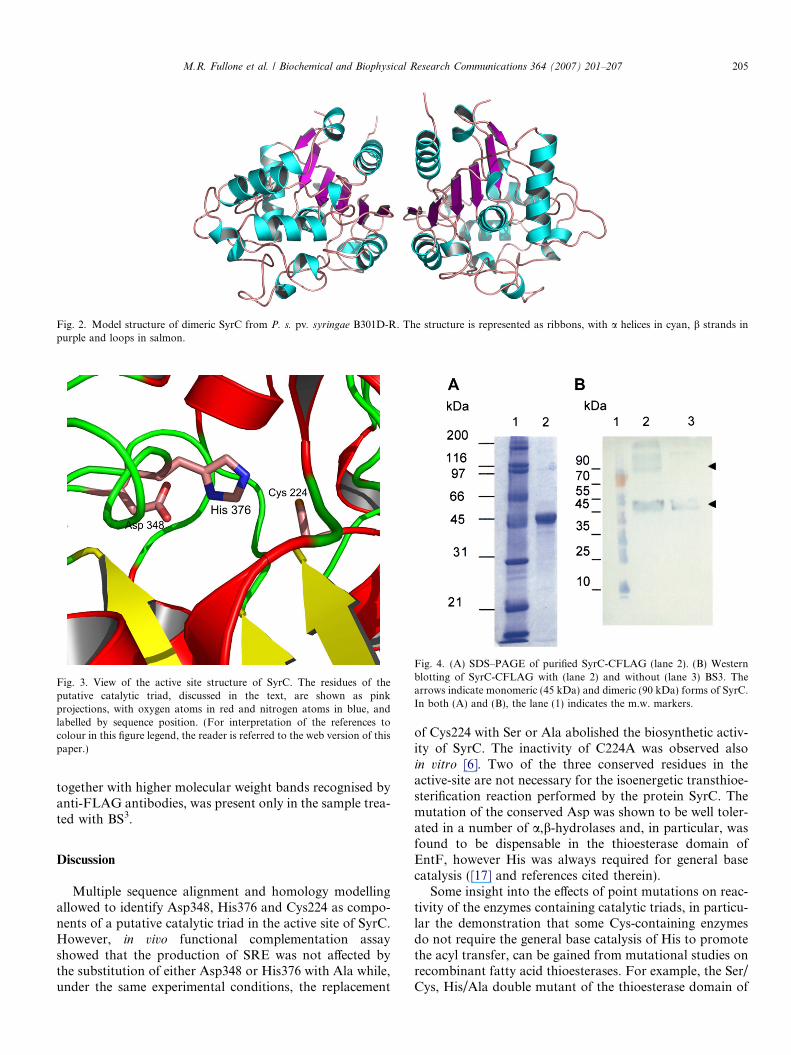
Fig. 2. Model structure of dimeric SyrC from P. s. pv. syringae B301D-R. The structure is represented as ribbons, with a helices in cyan, b strands inpurple and loops in salmon.
Fig. 3. View of the active site structure of SyrC. The residues of theputative catalytic triad, discussed in the text, are shown as pinkprojections, with oxygen atoms in red and nitrogen atoms in blue, andlabelled by sequence position. (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of thispaper.)
Fig. 4. (A) SDS–PAGE of purified SyrC-CFLAG (lane 2). (B) Westernblotting of SyrC-CFLAG with (lane 2) and without (lane 3) BS3. Thearrows indicate monomeric (45 kDa) and dimeric (90 kDa) forms of SyrC.In both (A) and (B), the lane (1) indicates the m.w. markers.
M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207 205
together with higher molecular weight bands recognised byanti-FLAG antibodies, was present only in the sample trea-ted with BS3.
Discussion
Multiple sequence alignment and homology modellingallowed to identify Asp348, His376 and Cys224 as compo-nents of a putative catalytic triad in the active site of SyrC.However, in vivo functional complementation assayshowed that the production of SRE was not affected bythe substitution of either Asp348 or His376 with Ala while,under the same experimental conditions, the replacement
of Cys224 with Ser or Ala abolished the biosynthetic activ-ity of SyrC. The inactivity of C224A was observed alsoin vitro [6]. Two of the three conserved residues in theactive-site are not necessary for the isoenergetic transthioe-sterification reaction performed by the protein SyrC. Themutation of the conserved Asp was shown to be well toler-ated in a number of a,b-hydrolases and, in particular, wasfound to be dispensable in the thioesterase domain ofEntF, however His was always required for general basecatalysis ([17] and references cited therein).
Some insight into the effects of point mutations on reac-tivity of the enzymes containing catalytic triads, in particu-lar the demonstration that some Cys-containing enzymesdo not require the general base catalysis of His to promotethe acyl transfer, can be gained from mutational studies onrecombinant fatty acid thioesterases. For example, the Ser/Cys, His/Ala double mutant of the thioesterase domain of

206 M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207
chicken fatty acid synthase was inactive as a hydrolase, butcould form the acyl-enzyme intermediate [18] and a doublemutation Ser/Cys, His/Arg converted a rat fatty acid thio-esterase into an acyltransferase, efficient in the presence ofthiol acceptors [19]. Regarding the exchange Cys/Ser, theeffects may vary depending on the specific protein, the reac-tion catalysed and complex steric and electrostatic environ-ment of the active site [20]. For example, the activity wasonly modified in a polyhydroxyalkanoate synthase [21]and abolished in arylamine N-acetyltransferases [22]. Itwould be of interest to ascertain if the Cys/Ser exchangein SyrC produces an inactive protein or confers hydrolyticactivity toward T- domain tethered intermediates, whichwould also result in the abrogation of SRE productionin vivo. Other proteins belonging to the a,b-hydrolase super-family that shuttle intermediates between different compo-nents of complex biosynthetic machineries where NRPS ismixed with other pathways, use either Cys or Ser as a nucle-ophile. CmaE involved in the biosynthesis of coronatine [7]is most similar to SyrC in that it utilizes a Cys residue tobind the intermediate and release it to a thiol acceptor ina net transthioesterification. On the other hand, the activesites of CouN7 [8] and CloN7 [9] that donate the acyl inter-mediate to an oxygen nucleophile acceptor in the biosynthe-sis of aminocoumarin antibiotics contain Ser in the activesite, whose role in the covalent binding of the intermediatehas been shown by radiolabelling for CouN7.
The insertion of the FLAG tag either on the N- or theC-terminus of SyrC did not affect its biosynthetic activity.The activity of the N-FLAG-SyrC is in line with thereported in vitro activity of the His6-tagged SyrC [6]. How-ever, both the insertion of a bulky MBP tag and the dele-tion of 16 residues at the N-terminus caused the loss ofbiosynthetic activity which could be due to conformationalmodifications, but could also indicate that the N-terminalportion plays a role in the interaction of SyrC with its pro-tein partners.
The homology modelling suggests a homodimeric struc-ture for SyrC in accordance with the presence of a bandcorresponding to the m.w. of the dimer in the cross-linkingexperiment. Moreover, the inactivation caused by themutation of one of the residues at the interface in the puta-tive homodimer, His369, lends further support of the rele-vance of the dimeric structure for the function of theprotein. A similar example is found in polyhydroxyalkano-ate synthase from Pseudomonas aeruginosa where themutation of Trp398, postulated to play a role in the dimer-isation, inactivated the enzyme [21]. The NRPS systemswere shown to be monomeric [23], with the exception ofVibF, a synthetase implicated in the formation of vibriob-actin [24]. SyrC is not a peptide synthetase, but it is anindispensable part of an NRPS machinery. At present,one can only speculate on the possible functional role ofdimerization of this aminoacyltransferase. Consideringthat one of the advantages of homodimerization is that itenables proteins to bind simultaneously to different proteinpartners [25] one could envisage that it could facilitate the
function of SyrC in transferring 4-chlorothreonine betweentwo T domains. However, more work is needed to clarifythe quaternary structure of SyrC.
Acknowledgments
This work was supported by the SYRTOX project of theProvincia Autonoma di Trento, by a grant from the ItalianMinistero dell 0Istruzione, Universita e Ricerca (MIUR)and Istituto Pasteur-Fondazione Cenci Bolognetti (grantto I.G.).
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.bbrc.2007.09.116.
References
[1] C.L. Bender, F. Alarcoen-Chaidez, D.C. Gross, Pseudomonas syrin-
gae phytotoxins: mode of action, regulation and biosynthesis bypeptide and polyketide synthetases, Microbiol. Mol. Biol. Rev. 63(1999) 266–292.
[2] I. Grgurina, M. Mariotti, Biosynthetic origin of syringomycin andsyringopeptin 22, toxic secondary metabolites of the phytopathogenicbacterium Pseudomonas syringae pv. syringae, FEBS Lett. 462 (1999)151–154.
[3] M.A. Fischbach, C.T. Walsh, Assembly-line enzymology for polyke-tide and nonribosomal antibiotics: logic, machinery and mechanisms,Chem. Rev. 106 (2006) 3468–3496.
[4] E. Guenzi, G. Galli, I. Grgurina, D.C. Gross, G. Grandi, Charac-terization of the syringomycin synthetase gene cluster, J. Biol. Chem.273 (1998) 32857–32863.
[5] F.H. Vaillancourt, J. Yin, C.T. Walsh, SyrB2 in syringomycin Ebiosynthesis is a nonheme Fe (II) a-ketoglutarate and O2-dependent halogenase, Proc. Nat. Acad. Sci. 102 (2005) 10111–10116.
[6] G.M. Singh, F.H. Vaillancourt, J. Yin, C.T. Walsh, Characterizationof SyrC, an aminoacyltransferase shuttling threonyl and chloroth-reonyl residues in the syringomycin biosynthetic assembly line, Chem.Biol. 14 (2007) 31–40.
[7] E.R. Strieter, F.H. Vaillancourt, C.T. Walsh, CmaE: a transferaseshuttling aminoacyl groups between carrier protein domains in thecoronamic acid biosynthetic pathway, Biochemistry 46 (2007) 7549–7557.
[8] C.J. Balibar, S. Garneau-Tsodikova, C.T. Walsh, Covalent CouN7enzyme intermediate for acyl group shuttling in aminocoumarinbiosynthesis, Chem. Biol. 14 (2007) 679–690.
[9] A. Freitag, E. Wemakor, S. M Li, L. Heide, Acyl transfer inclorobiocin biosynthesis: involvement of several proteins in thetransfer of the pyrrole-2-carboxyl moiety to the deoxysugar, Chem-Biochem 6 (2005) 2316–2325.
[10] N.S. Iacobellis, P. Lavermicocca, I. Grgurina, M. Simmaco,A. Ballio, Phytotoxic properties of Pseudomonas syringae pv.syringae toxins, Physiol. Mol. Plant Pathol. 40 (1992) 107–116.
[11] N.B. Quigley, Y.Y. Mo, D.C. Gross, SyrD is required for syringo-mycin production by Pseudomonas syringae pathovar syringae and isrelated to family of binding secretion proteins, Mol. Microbiol. 9(1993) 787–801.
[12] P. Prentki, F. Karch, S. Iida, J. Meyer, The plasmid cloning vectorpBR325 contains a 482 base-pair-long inverted duplication, Gene 14(1981) 289–299.

M.R. Fullone et al. / Biochemical and Biophysical Research Communications 364 (2007) 201–207 207
[13] D.H. Willis, E.M. Hrabak, S.E. Lindow, N.J. Panopoulos, Construc-tion and characterization of Pseudomonas syringae recA mutantstrains, Mol. Plant-Microbe Interact. 1 (1988) 80–86.
[14] S. Lu, B.K. Scholz-Schroeder, D.C. Gross, Construction ofpMEKm12 an expression vector for protein production in Pseudo-
monas syringae, FEMS Microbiol. Lett. 210 (2002) 115–121.[15] S.M. Monti, M. Gallo, R. Ferracane, R.C. Borrelli, A. Ritieni, M.L.
Greco, A. Graniti, V. Fogliano, Analysis of bacterial lipodepsipep-tides by matrix-assisted laser desorption/ionisation time-of-flight andhigh-performance liquid chromatography with electrospray massspectrometry, Rapid Commun. Mass Spectrom. 15 (2001) 623–628.
[16] S. Fushinobu, T. Saku, M. Hidaka, S.-Y. Jun, H. Noiri, H. Yamane,H. Shoun, T. Omori, T. Wakagi, Crystal structures of a meta-cleavage product hydrolase from Pseudomonas fluorescens IP01(CumD) complexed with cleavage products, Protein Sci. 11 (2002)2184–2195.
[17] C.A. Shaw-Reid, N.L. Kelleher, H.C. Losey, A.M. Gehring, C. Berg,C.T. Walsh, Assembly line enzymology by multimodular nonribos-omal peptide synthetases: the thioesterase domain of E. coli EntFcatalyzes both elongation and cyclolactonization, Chem. Biol. 6(1999) 385–400.
[18] M. Pazirandeh, S.S. Chirala, S.J. Wakil, Site-directed mutagenesisstudies on the recombinant thioesterase domain of chicken fatty acidsynthase expressed in Escherichia coli, J. Biol. Chem. 266 (1991)20946–20952.
[19] A. Witkowski, H.E. Witkowska, S. Smith, Reeingeneering thespecificity of a serine active-site enzyme, J. Biol. Chem. 269 (1994)379–383.
[20] M.E. McGrath, M.E. Wilke, J.N. Higaki, C.S. Craik, R.J. Fletterick,Crystal structures of two engineered thiol trypsins, Biochemistry 28(1989) 9264–9270.
[21] A.A. Amara, B.H.A. Rehm, Replacement of the catalytic nucleo-phile cysteine-269 by serine in class II polyhydroxyalkanoatesynthase from Pseudomonas aeruginosa-mediated synthesis of anew polyester: identification of catalytic residues, Biochem. J. 374(2003) 413–421.
[22] J. Sandy, A. Mushtaq, S.J. Holton, P. Schartau, M.E.M. Noble, E.Sim, Investigation of the catalytic triad of arylamine N-acetyltrans-ferases: essential residues required for acetyl transfer to arylamines,Biochem. J. 390 (2005) 115–123.
[23] S. A Sieber, U. Linne, N.J. Hillson, E. Roche, C.T. Walsh, M.A.Marahiel, Evidence for a monomeric structure of nonribosomalpeptide synthetase, Chem. Biol. 9 (2002) 997–1008.
[24] N.J. Hillston, C.J. Balibar, C.T. Walsh, Catalytically inactivecondensation domain C1 is responsibe for dimerization of theVibF subunit of vibriobactin synthetase, Biochemistry 43 (2004)11344–11351.
[25] N.J. Marianayagam, M. Sunde, J.M. Matthews, The power oftwo: protein dimerization in biology, Trends Biochem. Sci. 29 (2004)618–625.
Related Documents











