Multiple pools of PtdIns 4-phosphate detected using the pleckstrin homology domain of Osh2p Anjana Roy and Timothy P Levine Division of Cell Biology, Institute of Ophthalmology, University College London, Bath Street, London EC1V 9EL Address correspondence to Tim Levine: phone (+44)-20 7608 4027/8, fax (+44)-20 7608 4034 email [email protected] Running Title: A PH domain that binds multiple pools of PtdIns(4)P JBC Papers in Press. Published on July 22, 2004 as Manuscript M401583200 Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on February 13, 2018 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Multiple pools of PtdIns 4-phosphate detected using the
pleckstrin homology domain of Osh2p
Anjana Roy and Timothy P Levine
Division of Cell Biology, Institute of Ophthalmology, University College London, Bath
Street, London EC1V 9EL
Address correspondence to Tim Levine:
phone (+44)-20 7608 4027/8, fax (+44)-20 7608 4034
email [email protected]
Running Title:
A PH domain that binds multiple pools of PtdIns(4)P
JBC Papers in Press. Published on July 22, 2004 as Manuscript M401583200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
2
SUMMARY
Phosphatidylinositol (PtdIns) phosphate (PtdInsP) lipids are used as intracellular
signposts for the recruitment and activation of peripheral membrane proteins. While
the distribution of most PtdInsPs is restricted to a single organelle, PtdIns(4)P is
unique in that it exists in several discrete pools, and so proteins that bind
PtdIns(4)P must use extra receptors to achieve a restricted localization. Here we
compare the two highly related pleckstrin homology (PH) domains from Osh1p and
Osh2p, yeast homologues of oxysterol binding protein (OSBP), that target
membranes using PtdIns(4)P, and in vitro bind both PtdIns(4)P and PtdIns(4,5)P2.
We show that Golgi targeting is specified by an additional site on PHOsh1, that lies on
a face of the domain not previously known to interact with receptors. In contrast,
PHOsh2 does not have a demonstrable second site, and targets multiple pools of
PtdInsPs, each dependent on a different PtdIns 4-kinase. This lack of a second site
in PHOsh2 allows it to be used as an unbiased reporter for altered distribution of
4–phosphorylated PtdIns. For example, in cells with excess PtdIns(4)P caused by
inactivation of the phosphatase Sac1p, PHOsh2 indicates that PtdIns(4)P accumulates
on the plasma membrane, while other Golgi-targeted PH domains fail to detect this
change.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
3
INTRODUCTION
Membrane recruitment of peripheral membrane proteins is determined by one or
more targeting domains that bind other proteins or membrane lipids. An important
class of membrane lipid used to recruit proteins are the PtdInsPs, phosphates being
added at positions 3, 4 or 5 of the inositol sugar in any one of 7 combinations. Most
PtdInsPs have quite restricted distributions, and can be considered as intracellular
signposts or signals that recruit proteins to specific compartments (1). These
distributions are created by the targeting of lipid modifying enzymes. For example,
PtdIns(3)P is largely restricted to endosomes: it is synthesized by a PtdIns 3-kinase
lying on the trans-Golgi network to endosome pathway, and it is prevented from
reaching other compartments by degradation in the vacuole/lysosome (2).
PtdIns(4)P differs from other PtdInsPs as it performs multiple roles. Not only is it a
signpost for protein recruitment, but it is also an essential intermediate in the
production of PtdIns(4,5)P2. Further, PtdIns(4)P is synthesized by multiple,
separate PtdIns 4-kinases and is therefore likely to have multiple intracellular pools.
There are two families of PtdIns 4-kinases: types II and III (type I are now known
to be PtdIns 3-kinases (3)). In mammalian cells, the predominant PtdIns 4-kinases
are type II, but have only recently been cloned and are still partially characterized
(4-7). In addition, there are two mammalian type III kinases: α and β (8). Analysis
of the relative importance of the different kinases in membrane traffic has been
carried out in the model organism S. cerevisiae, which also has type II and III
kinases, but with a different emphasis, since here the type III kinases are dominant
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
4
(9). The yeast type III α PtdIns 4-kinase homologue, Stt4p, is an essential protein
that has been localized to the plasma membrane (10), and has been shown to
synthesize the bulk of PtdIns(4)P for cellular requirements. This pool acts as a
precursor for PtdIns(4,5)P2 (11), which is perhaps exclusively on the plasma
membrane (12,13), and which is made by a single PtdIns(4)P 5-kinase, Mss4p, that
is active on the plasma membrane (14,15). However, there are important roles for
PtdIns(4)P beyond being an intermediate in PtdIns(4,5)P2 production, in particular
in budding from the late Golgi. This is the location of Pik1p, another essential PtdIns
4-kinase that is homologous to the human type III β kinase (16,17). A number of
peripheral Golgi proteins are now known to be recruited by PtdIns(4)P. Pleckstrin
homology (PH) domains from three families of lipid binding proteins target the Golgi
(12,13,18-21). In addition, clathrin adaptor AP1 complex was also shown to
interact with PtdIns(4)P on the Golgi (22). Yeast also harbor a third PtdIns 4-
kinase, Lsb6p, which is in the type II family, and has no known function (23).
The question posed by the presence of these different PtdIns 4-kinases is how
might proteins detect one specific pool of PtdIns(4)P among the multiple organelles
that have active PtdIns 4-kinases? One plausible explanation supported by previous
observations with the PH domain of OSBP is that proteins cannot target a specific
compartment by virtue of PtdIns(4)P alone, but achieve restricted targeting by
binding a membrane lipid in combination with other membrane receptors (12,24).
OSBP is a peripheral membrane protein that targets the late Golgi solely by virtue
of a PH domain that binds to PtdIns(4)P and PtdIns(4,5)P2 with equal affinity
(12,18). In addition, PHOSBP binds a Golgi-restricted receptor dependent on the
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
5
small GTPase Arf1p (12), and has recently been shown to bind weakly to ARF1
(21).
Here, we focus on two PH domains from yeast homologues of OSBP: Osh1 and
Osh2. While PHOsh1 targets Golgi membranes, PHOsh2, a closely related domain (71%
identical and 91% homologous) targets membranes weakly (25). We have
identified the site within PHOsh1 that binds a second membrane receptor, and
developed a tool based on PHOsh2 that lacks the second site and so detects pools of
4-phosphorylated PtdIns in an unbiased manner. Using PHOsh2, we demonstrate the
physical separation of the two pools of PtdIns(4)P dependent on Stt4p and Pik1p,
on the plasma membrane and Golgi respectively. We then compare PHOsh2 to other
PH domains that bind PtdIns(4)P and PtdIns(4,5)P2 as a probe for excess
PtdIns(4)P in ∆sac1 cells.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
6
Experimental Procedures
Plasmids
PH domains were defined as follows (alternative gene names together with
accession numbers given in brackets, followed by starting and ending residues,
identified using single letter amino acid code): Osh1p (YAR042W/YAR044W,
AY241177) E280-R384; Osh2p (YDL019C, NP_010265) long form: 256P-424P;
short form: S286-K391; phospholipase C δ1 (PLCδ1), ceramide transfer protein
(CERT, previously called Goodpasture’s antigen binding protein – GPBP) and
phosphoinositol 4-phosphate adaptor protein 1 (FAPP1) as before (12). For
chimeras between Osh1p and Osh2p PH domains, 4 restriction sites that do not
alter the translated sequence were introduced at roughly equal spacing in both PH
domains, to produce 5 segments that shared the same boundary residues in both
two domains. Plasmids containing PH domains were based on either pRS406
(integrating at URA3) or pRS416 (CEN URA3) and contained the constitutive portion
of the PHO5 promoter, followed by GFP, a linker with the Myc epitope
(GSMEQKLISEEDLRS), then the PH domain and a carboxy-terminal extension of
XNS (where X is encoded by the last two nucleotides from the source DNA and a
guanine) as previously (12,24,25). Plasmids are listed in Table 1.
Insert Table 1 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
7
Yeast strains For localization in wild-type yeast, plasmids were routinely
transformed into strain RS453B. Strains harboring the temperature-sensitive alleles
pik1-83, stt4-4 and mss4-2 together with their wild type controls and ∆vps34 were
as described previously (12). A ∆sac1 strain (TLY218) was derived from SEY6210
by replacing SAC1 with the heterologous marker gene S. pombe HIS5 using the
PCR method (26), and deletion of SAC1 was confirmed by PCR and inositol
auxotrophy.
Liposome centrifugation assays His6-tagged fluorescent PH domains were
expressed in bacteria and purified as previously (12). Small unilamellar liposomes
were prepared, incubated with GFP-PH domain and removed from suspension by
centrifugation as described previously (12). The GFP content remaining in the
supernatant was measured in a LS50 spectrophotometer (excitation 485 nm,
emission 515 nm, slit widths 5 nm). Phosphoinositides were assumed to be equally
distributed between the two leaflets of the liposome bilayer.
Cell imaging GFP-PH domain fusion proteins were visualized in yeast cells growing
at mid-log phase (OD 600 nm = 1) and in COS cells as previously (12,24). To label
yeast with FM4-64, the dye was added at final concentration of 30 µM 15 minutes
prior to imaging endosomal/late Golgi compartments.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
8
RESULTS
Identification of the second site in PHOsh1 for Golgi targeting. PHOsh1 targets
Golgi membranes in yeast (Fig. 1A) and mammalian fibroblasts (24,25), while
PHOsh2 shows barely discernible membrane targeting in yeast (Fig. 1B) and
fibroblasts (data not shown). This differs from a recent study showing stronger
Golgi targeting by monomeric PHOsh2, although in that case a basic patch adjacent
to the core PH domain sequence was also included, which may alter targeting (27).
The portion of primary sequence responsible for the difference in targeting between
PHOsh1 and PHOsh2 was mapped using a series of chimeras in which portions of the
Osh1p sequence were exchanged for the homologous Osh2p sequence. For this
purpose, the PH domain (105 amino acids in Osh1p) was divided into five
approximately equal segments, and chimeras made that contained mixtures of
Osh1p and Osh2p sequence (see Materials and Methods). Substitution of the N-
terminal three fifths of PHOsh1 with the equivalent region of PHOsh2 had no effect on
targeting (Fig. 1C), despite the almost universal location of the binding site in PH
domains for PtdInsPs within this N-terminal region (28,29). In contrast, inclusion of
the next (i.e. fourth) segment of PHOsh2 inhibited membrane localization (Fig. 1D).
The same requirement for the fourth segment of PHOsh1 was seen for Golgi targeting
in mammalian COS cell fibroblasts (data not show).
Insert Figure 1 about here
Insert Table 2 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
9
Within the critical fourth segment of 22 residues, PHOsh1 differs from PHOsh2 at 8
positions. By further dissection of this segment, a single residue, histidine at
position 79 in PHOsh1 compared to arginine at position 80 in PHOsh2, was shown to be
critical for targeting (Table 2). To confirm the role of this single residue, arginine
was introduced at position 79 of PHOsh1, in which it strongly inhibited Golgi targeting
(Fig. 1E). Introduction of histidine at the same site in PHOsh2 induced a striking gain
of Golgi targeting (Fig. 1F).
Modeling of the sequence of PHOsh1 on known PH domain structures (28,29) shows
that this residue lies in the middle of the β7 strand. In all known structures, this
residue points away from the core of the domain on a side of the molecule at some
distance from the membrane interface, assuming that this is defined by the site
that binds PtdInsPs (Fig. 2). These results therefore indicate that the second site is
unlikely to interact with a membrane lipid receptor, but is more likely to be a
protein.
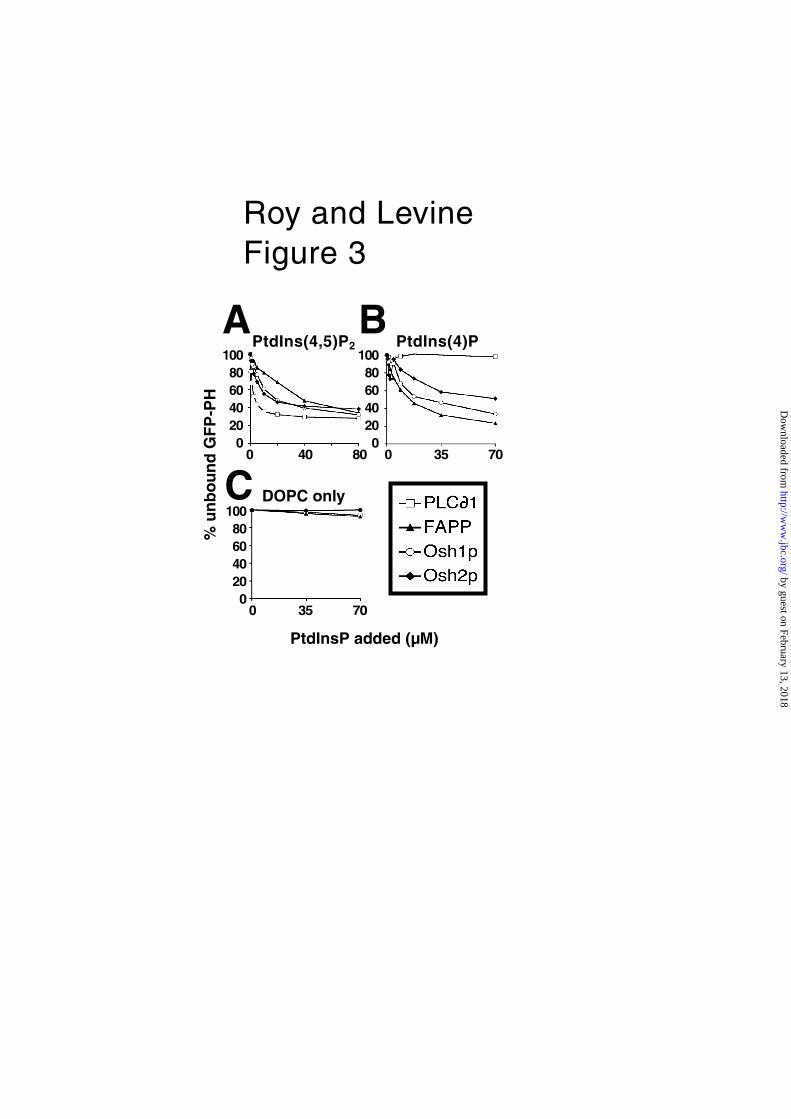
PHOsh2 binds both PtdIns(4)P and PtdIns(4,5)P2. If PHOsh2 is fully capable of
membrane targeting, but just lacks a second site, it should bind PtdInsPs similarly
to PHOsh1. To test this, we compared the binding of GFP-tagged PH domains to
liposomes containing 2.5% PtdIns(4)P or PtdIns(4,5)P2 (12) (Fig. 3). As controls,
PHPlcδ1 showed its well-documented absolute preference for PtdIns(4,5)P2 over
PtdIns(4)P, and PHFAPP1 bound both lipids, with a slight preference for PtdIns(4)P
(Fig. 3). PHOsh1 and PHOsh2 bound PtdIns(4,5)P2 with almost identical affinity, greater
Insert Figure 2 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
10
than that of PHFAPP1 (Fig. 3A); PHOsh1 bound PtdIns(4)P more strongly than PHOsh2
(approximately 2-fold more) (Fig. 3B). Overall, these experiments show that PHOsh2
binds PtdIns(4)P and PtdIns(4,5)P2 with affinity similar to other PH domains that
target membranes efficiently, despite its very weak membrane targeting in vivo.
A PHOsh2 dimer targets two pools of 4 phosphorylated PtdInsPs in growing
cells. The above data show that PHOsh2 binds 4-phosphorylated PtdIns, but it differs
from closely related PH domains by the absence of a second site. Therefore, we
reasoned that if a reporter construct could be made to target membranes
dependent purely on PHOsh2, it would reveal the overall distribution of PtdIns(4)P
and PtdIns(4,5)P2. To achieve membrane localization, we expressed a tandem
dimer of PHOsh2, as has been done for PHPLCδ1 and FYVE domains previously. The
presence of two membrane binding sites vastly increases avidity without altering
specificity (12,30). GFP-PHOsh2-dimer showed tight dual localisation to punctate
internal structures and the plasma membrane, with little cytoplasmic background
(Fig. 4A). GFP-PHOsh2-dimer expressed from either a relatively weak promoter
(OSH1), or a rapidly inducible promoter (GAL1) produced the same dual localisation
(data not shown), indicating that it is independent of duration and level of
expression. Although the distribution of GFP-PHOsh2-dimer became diffuse after
fixation (data not shown), partial co-localisation was found in live cells with punctae
stained by FM4-64, which traverses the endocytic pathway and also enters late
Golgi compartments prior to being sorted back to the cell surface (Fig. 4B) (31).
Therefore, we conclude that the punctate structures are late Golgi and/or early
Insert Figure 3 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
11
endosomes. In addition, the dual targeting of PHOsh2 implies either that it interacts
with two different ligands, or alternatively that a single ligand is present in two
sites. An indication that both localisations of GFP-PHOsh2-dimer are PtdInsP-
dependent was gained using a dimer in which two conserved positive residues of
both PH domain sequences contained mutations that are known to prevent PtdInsP
binding site of the PH domains (12), which had an entirely diffuse localisation (Fig.
4C).
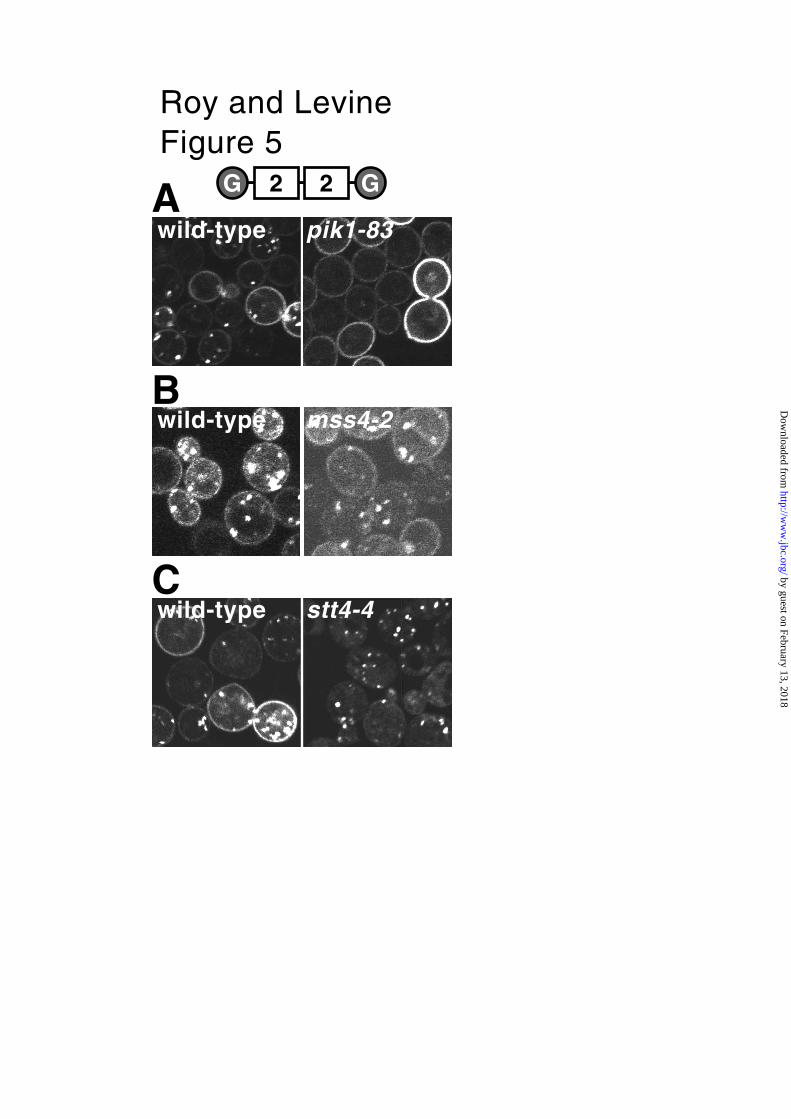
To test whether Pik1p plays a role in the localisation of PHOsh2, as it does for PHOSBP
(12), GFP-PHOsh2-dimer was expressed in a strain carrying a conditional pik1 allele.
Shift to the non-permissive temperature caused a rapid and almost complete loss of
internal punctate localisation in the mutant cells, with only a slight effect seen in
wild-type cells undergoing the same temperature shift (Fig. 5A). By comparison,
PHOSBP is only incompletely delocalized under these conditions (12). Therefore,
PtdIns(4)P produced by Pik1p is likely to be the Golgi ligand for PHOsh2. Next, we
sought to identify the plasma membrane ligand for PHOsh2. Initially we looked at the
role of PtdIns(4,5)P2 by expressing GFP-PHOsh2-dimer in cells carrying the mss4-2ts
allele. Little effect on the plasma membrane localisation was detected at the non-
permissive temperature under conditions where PtdIns(4,5)P2 levels are known to
fall (Fig. 5B) (12). This result does not rule out a role for PtdIns(4,5)P2 at the
plasma membrane, but indicates that there must be an additional ligand present
when PtdIns(4,5)P2 production is blocked.
Insert Figure 4 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
12
We next tested whether the other essential PtdIns(4)-kinase, Stt4p, was
responsible for synthesising a pool of PtdInsP on the plasma membrane that is
recognised by PHOsh2. In temperature sensitive strains carrying the mutant allele
stt4-4ts, the localisation of PHOsh2-dimer was altered, with much reduced plasma
membrane localization even at the permissive temperature (data not shown).
Plasma membrane targeting was almost completely lost at the non-permissive
temperature, while punctate localisation was preserved (Fig. 5C, 37˚C), and these
effects were not seen in wild-type controls (Fig. 5C), indicating that Stt4p
synthesizes the 4-phosphorylated PtdIns that PHOsh2 binds on the plasma
membrane. We also examined the role of the single PtdIns 3-kinase in yeast,
VPS34, since it is possible that PHOsh2 binds either PtdIns(3)P or PtdIns(3,5)P2.
However, the localisation of GFP-PHOsh2-dimer was unaffected in ∆vps34 cells (data
not shown). The combination of this result and the complete loss of punctate
localization with pik1 mutation indicates that the punctate localization of PHOsh2 is
not mediated by PtdIns(3)P. Overall, these results show that each of the dual
localisations of this PH domain is sensitive to inactivation of a different essential
PtdIns 4-kinase, demonstrating that PHOsh2 interacts with two separate pools of 4-
phosphorylated PtdIns.
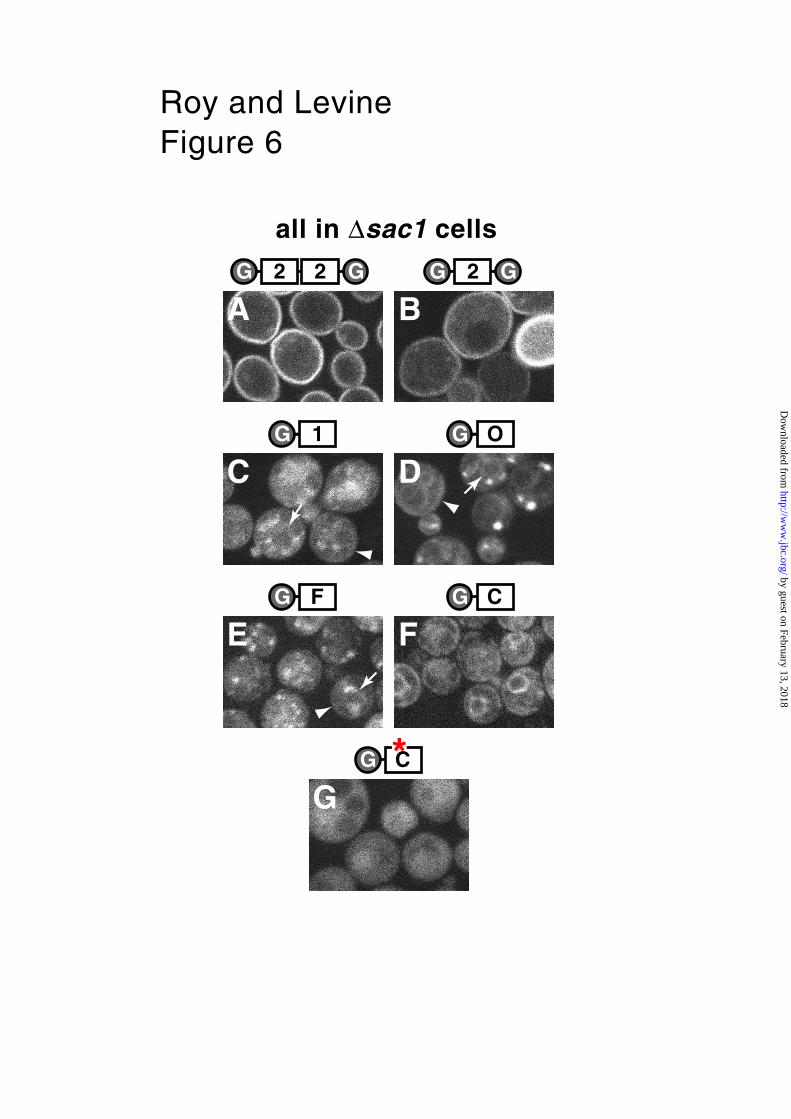
PHOsh2 as a reporter for altered PtdIns(4)P levels in ∆sac1 cells.
Insert Figure 5 about hereInsert Figure 5 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
13
The results above indicate that PHOsh2 is an unbiased reporter of PtdIns(4)P and
PtdIns(4,5)P2, so we next examined how its distribution was affected by a well-
characterised mutation that alters PtdInsP levels. Sac1p is one of five PtdInsP
phosphatases in yeast, and its inactivation leads to a large increase in cellular
PtdIns(4)P, up to ten-fold, together with a fall in PtdIns(4,5)P2 (16,32). To
determine the effect of these changes in 4-phospholrylated PtdIns, we expressed
tagged PH domains in a ∆sac1 strain. The GFP-PHOsh2-dimer showed much
increased plasma membrane targeting, and an apparent decrease in punctate
localization (Fig. 6A, compare with Fig. 4A). This change might be caused by either
a reduction in ligand concentration on Golgi/endosomes or an increase in ligand on
the plasma membrane. To distinguish between these possibilities, we expressed
monomeric GFP-PHOsh2 in the ∆sac1 strain, and found that a single copy of PHOsh2
now localized appreciably to the plasma membrane (Fig. 6B, compare with Fig. 1B).
This result indicates that the ∆sac1 mutation leads to an increase in PtdIns(4)P at
the plasma membrane, which is likely, from the results above, to be synthesized by
Stt4p. Therefore, our findings agree with previous results that the excess
PtdIns(4)P that accumulates in ∆sac1 cells is synthesized by Stt4p rather than by
Pik1p (32).
We next examined the effect of the ∆sac1 mutation on other PH domains that bind
PtdIns(4)P. GFP-PHOsh1 did not target the enlarged pool of PtdIns(4)P at the plasma
membrane, but remained largely punctate (Fig. 6C). This is further evidence that
PHOsh1 does not target membranes solely because they are rich in PtdIns(4)P. We
Insert Figure 6 about here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
14
next examined the effect of ∆sac1 on the distribution of other Golgi-targeted PH
domains, using sequences from OSBP, FAPP1 and ceramide transfer protein (CERT,
previously called Goodpasture’s antigen binding protein – GPBP). Like PHOsh1, these
target the Golgi by interacting with both PtsIns(4)P and a second site (12). The
distribution of GFP-PHOSBP and GFP-PHFAPP1 remained punctate, as seen for GFP-
PHOsh1 (Fig. 6D and E). However, GFP-PHCERT was notably different, being targeted
to the nuclear envelope and to peripheral patches, characteristic of the yeast ER
(Fig. 6F). To check whether this targeting was caused by binding to PtdInsPs, we
next expressed a mutated version of this construct, in which the critical basic
residue for PtdInsP binding was mutated to an arginine. GFP-PHCERT(R43E) was
entirely diffuse, with no ER or punctate localization (Fig. 6G). On close inspection,
all three of PHOsh1, PHOSBP and PHFAPP1showed much weaker targeting to the nuclear
envelope and periphery (Fig. 6). These results suggest that some PtdIns(4)P
accumulates on the ER in ∆sac1 cells, and that this pool is specifically targeted by
PHCERT, possibly because of a unique second site interaction.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
15
Discussion
PH domains are sequences usually 100–120 amino acids in length that have a
characteristic fold of a seven-stranded β sandwich closed off at one side by an
amphipathic alpha helix (28,29,33). PH domains have been found to interact with a
variety of different ligands, using sites that are somewhat conserved for each
purpose. The alpha helix can interact with WD40 proteins including Gβγ (34), and
differing parts of the alpha helix of PH domains in Guanine nucleotide exchange
factors bind the neighbouring catalytic Dbl homology domain (35). Finally, the best
characterized interaction of PH domains arises from the formation of a pocket by
loops β1-β2 and β3-β4 to bind PtdInsPs. Lipid specificity is determined by a
combination of basic residues lining the pocket and a generally positive electrostatic
charge on this face of the domain. While a few PH domains, in particular those that
bind PtdIns(3,4,5)P3, show high affinity binding for just a single PtdInsP, most PH
domains, including PHOSBP and related sequences, bind PtdInsPs relatively
promiscuously (28,36-38). This lack of specificity need not preclude specific overall
targeting if the domain also interacts with a second receptor (12).
We compared two highly related PH domains with different Golgi targeting. Both PH
domains bound PtdInsPs with moderate strength, similar to the recent findings of
Lemmon and co-workers (38). Indeed, out of the 33 PH domains in the entire yeast
genome, these two are among only seven that demonstrably bind PtdInsPs at all
(38). Given a similar binding affinity for PtdInsPs, what determines the difference in
targeting? This was mapped to the third residue of strand β7, which in PHOsh1 (tight
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
16
binding) consists of RWHLKG, compared to RWRLKG in PHOsh2 (weak binding). The
histidine is conserved in Osh1p homologues in humans (OSBP, ORP4 and ORP9),
flies, worms, plants and in other fungi (C. albicans, A. nidulans but not S. pombe –
serine, or M. Grisea and N. crassa – threonine), whereas the arginine is unique to
Osh2p in Saccharomyces. Different PH domain structures all indicate that this
residue lies at the bottom of a shallow groove on a face of the domain adjacent to
but not overlapping with the PtdInsP binding site (Fig. 2) (28,29,37). The finding
that this histidine can specify Golgi localization identifies this side of PH domains as
a site in this sub-family of PH domains that binds a second ligand. Given the
location of the site away from the membrane, it would appear that the ligand is a
protein, rather than a membrane lipid. Following our initial studies where we found
that this second site depends on Arf1p in yeast (12), De Matteis and co-workers
have now demonstrated a direct interaction between both PHFAPP1 and PHOSBP with
ARF1 in mammalian cells (the equivalent of yeast Arf1p) (21). On this basis, we
therefore predict that ARF1/Arf1p interacts with histidine 79 in PHOsh1 and PHOSBP
(12).
Although Golgi targeting can be achieved by combining two binding interactions,
without a second site monomeric PHOsh2 does not target the Golgi. This reduced
affinity for the Golgi may be crucial to targeting of full-length Osh2p, which has
diverged in multiple ways from Osh1p (25). Because PHOsh2 interacts only with
PtdInsPs, we have been able to develop a tool based on dimeric PHOsh2 that detects
PtdIns(4)P and PtdIns(4,5)P2 alone (Fig. 3). Binding to other PtdInsPs was shown to
be irrelevant by the lack of a role for Vps34p. The dual localization of GFP-PHOsh2-
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
17
dimer in log-phase cells was consistent with previous studies of the two essential
PtdIns 4-kinases (9), and their localizations (10,17). While previous studies have
shown that Pik1p produces PtdIns(4)P at the Golgi (12,13), this is the first
demonstration that Stt4p synthesizes PtdIns(4)P on the plasma membrane. Since
the Stt4-dependent pools of PtdIns(4)P and PtdIns(4,5)P2 are of roughly equivalent
size (11), it is difficult to determine which lipid is being detected by the GFP-PHOsh2-
dimer, even using cells in which Mss4p can be inactivated.
If GFP-PHOsh2-dimer is indeed a probe for 4-phosphorylated PtdIns, its distribution
should change in response to changes in PtdInsP metabolism. We used cells in
which the SAC1 gene was disrupted, since in the absence of this PtdInsP
phosphatase cells accumulate up to ten times the normal levels of PtdIns(4)P, while
PtdIns(4,5)P2 decreases by 50% (16,32). Results with PHOsh2 in these cells showed
that excess PtdIns(4)P accumulated mainly on the plasma membrane. However,
other PH domains that bind PtdIns(4)P, such as those from Osh1p, OSBP and FAPP1
remained largely localized to the Golgi in ∆sac1 cells, similar to the recent findings
of Lemmon and co-workers (38). This is further evidence that PH domains that
target the Golgi as monomers localize using a second receptor in addition to
PtdInsP. Interestingly, PHCERT behaved differently in cells carrying ∆sac1 compared
to the other sequences tested in targeting the ER in a PtdInsP-dependent manner.
This suggests that PHCERT recognizes a second Golgi receptor that is relocated to the
ER in a ∆sac1 strain. Since this receptor differs from that recognized by PHOsh1p,
PHOSBP and PHFAPP1, it may be that PHCERT does not bind ARF1. ER relocalization of a
Golgi protein in ∆sac1 cells has also been reported for the short yeast OSBP
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
18
homologue Kes1p, which also binds PtdIns(4)P and PtdIns(4,5)P2 (39). CERT was
recently shown to transfer ceramide in mammalian cells from its site of synthesis
(the ER) to the site of its conversion to complex sphingolipid (late Golgi) (19), a
process which also occurs in yeast (40). Identical targeting by PHCERT and Kes1p
suggests the possibility that they both access a sub-domain of the late Golgi that is
specialized for non-vesicular ceramide import.
It is clear from all studies of PH domains targeted to Golgi membranes that they
bind PtdIns(4)P. However, these PH domains can also show fairly indiscriminate
binding to a broad range of PtdInsPs (12,24,36,38), i.e. there is no intrinsic
specificity for PtdIns(4)P. This is sometimes over-looked (41) because different
techniques for measuring protein-lipid interaction yield differing results. By binding
protein to lipids immobilized on nitrocellulose (so-called “Fat Western”) originally
showed that PHFAPP1 specifically binds PtdIns(4)P (20), however this specificity was
not seen with an amended technique (38). Although it is not yet known if other
proteins that target PtdIns(4)P on the Golgi are equally promiscuous (22), it is
tempting to speculate why such lack of specificity exists for PtdIns(4)P binding
proteins. PtdIns(4)P exists in multiple discrete pools, not only in yeast, but also in
mammalian cells where multiple organelles can recruit PtdIns 4-kinases (4-
8,22,42). Thus, proteins that target PtdIns(4)P can only achieve a narrow
distribution among the multiple pools of PtdIns(4)P by combining this interaction
with others. Therefore, there may be very little evolutionary disadvantage to
binding the combination of PtdIns(4)P and PtdIns(4,5)P2, and it is possible that a
truly PtdIns(4)P-specific binding protein may not exist.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
19
In summary, PHOsh2 appears to detect two independent pools of 4-phosphorylated
PtdIns without the bias introduced by a second site that is otherwise found in PH
domains that bind PtdIns(4)P. The potential use of the probe is complicated by its
interaction with another lipid, PtdIns(4,5)P2. Now that the micro-heterogeneity of
all intracellular organelles has become evident (43-45), it will become increasingly
important to study how multiple targeting signals integrate to specify a protein’s
overall distribution.
Acknowledgements We would like to acknowledge the support of Sean Munro, in
whose lab this work was initiated, and who has given much useful advice. We would
also like to thank Scott Emr for providing yeast strains and communicating results
prior to publication, and Anjon Audhya, Olga Perisic, Howard Riezman, Steve Moss
and Chris Loewen for helpful discussions. Funded by the Wellcome Trust (Grant no.
060537) and Fight For Sight.
Abbreviations:
CERT – ceramide transfer protein, GPBP - Goodpasture’s antigen binding protein,
NLS - nuclear localization signal, OSBP - oxysterol binding protein, PH – pleckstrin
homology, PtdIns – phosphatidylinositol, PtdInsP - phosphatidylinositol phosphate
(PtdInsP), FAPP1 - phosphoinositol 4-phosphate adaptor protein 1, PLCδ1 -
phospholipase C δ1.
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
20
References
1. Insall, R. H., and Weiner, O. D. (2001) Dev Cell 1, 743-747
2. Wurmser, A. E., and Emr, S. D. (1998) Embo J 17, 4930-4942
3. Whitman, M., Downes, C. P., Keeler, M., Keller, T., and Cantley, L. (1988)
Nature 332, 644-646
4. Minogue, S., Anderson, J. S., Waugh, M. G., dos Santos, M., Corless, S.,
Cramer, R., and Hsuan, J. J. (2001) J Biol Chem 276, 16635-16640
5. Barylko, B., Gerber, S. H., Binns, D. D., Grichine, N., Khvotchev, M., Sudhof,
T. C., and Albanesi, J. P. (2001) J Biol Chem 276, 7705-7708
6. Balla, A., Tuymetova, G., Barshishat, M., Geiszt, M., and Balla, T. (2002) J Biol
Chem 277, 20041-20050
7. Wei, Y. J., Sun, H. Q., Yamamoto, M., Wlodarski, P., Kunii, K., Martinez, M.,
Barylko, B., Albanesi, J. P., and Yin, H. L. (2002) J Biol Chem 277, 46586-
46593
8. Wong, K., Meyers dd, R., and Cantley, L. C. (1997) J Biol Chem 272, 13236-
13241
9. Audhya, A., Foti, M., and Emr, S. D. (2000) Mol Biol Cell 11, 2673-2689
10. Audhya, A., and Emr, S. D. (2002) Dev Cell 2, 593-605
11. Perera, N. M., Michell, R. H., and Dove, S. K. (2004) J Biol Chem 279, 5216-
5226
12. Levine, T. P., and Munro, S. (2002) Curr Biol 12, 695-704
13. Stefan, C. J., Audhya, A., and Emr, S. D. (2002) Mol Biol Cell 13, 542-557
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
21
14. Homma, K., Terui, S., Minemura, M., Qadota, H., Anraku, Y., Kanaho, Y., and
Ohya, Y. (1998) J Biol Chem 273, 15779-15786
15. Audhya, A., and Emr, S. D. (2003) Embo J 22, 4223-4236
16. Hama, H., Schnieders, E. A., Thorner, J., Takemoto, J. Y., and DeWald, D. B.
(1999) J Biol Chem 274, 34294-34300
17. Walch-Solimena, C., and Novick, P. (1999) Nat Cell Biol 1, 523-525
18. Ridgway, N. D., Dawson, P. A., Ho, Y. K., Brown, M. S., and Goldstein, J. L.
(1992) J Cell Biol 116, 307-319
19. Hanada, K., Kumagai, K., Yasuda, S., Miura, Y., Kawano, M., Fukasawa, M.,
and Nishijima, M. (2003) Nature 426, 803-809
20. Dowler, S., Currie, R. A., Campbell, D. G., Deak, M., Kular, G., Downes, C. P.,
and Alessi, D. R. (2000) Biochem J 351, 19-31
21. Godi, A., Di Campli, A., Konstantakopoulos, A., Di Tullio, G., Alessi, D. R.,
Kular, G. S., Daniele, T., Marra, P., Lucocq, J. M., and De Matteis, M. A.
(2004) Nat Cell Biol 6, 393-404
22. Wang, Y. J., Wang, J., Sun, H. Q., Martinez, M., Sun, Y. X., Macia, E.,
Kirchhausen, T., Albanesi, J. P., Roth, M. G., and Yin, H. L. (2003) Cell 114,
299-310
23. Han, G. S., Audhya, A., Markley, D. J., Emr, S. D., and Carman, G. M. (2002)
J Biol Chem 277, 47709-47718
24. Levine, T. P., and Munro, S. (1998) Curr Biol 8, 729-739
25. Levine, T. P., and Munro, S. (2001) Mol Biol Cell 12, 1633-1644
26. Wach, A., Brachat, A., Alberti-Segui, C., Rebischung, C., and Philippsen, P.
(1997) Yeast 13, 1065-1075
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
22
27. Nagel, W., Schilcher, P., Zeitlmann, L., and Kolanus, W. (1998) Mol Biol Cell 9,
1981-1994
28. Ferguson, K. M., Kavran, J. M., Sankaran, V. G., Fournier, E., Isakoff, S. J.,
Skolnik, E. Y., and Lemmon, M. A. (2000) Mol Cell 6, 373-384
29. Lemmon, M. A. (2003) Traffic 4, 201-213
30. Gillooly, D. J., Morrow, I. C., Lindsay, M., Gould, R., Bryant, N. J., Gaullier, J.
M., Parton, R. G., and Stenmark, H. (2000) Embo J 19, 4577-4588
31. Wiederkehr, A., Avaro, S., Prescianotto-Baschong, C., Haguenauer-Tsapis, R.,
and Riezman, H. (2000) J Cell Biol 149, 397-410
32. Foti, M., Audhya, A., and Emr, S. D. (2001) Mol Biol Cell 12, 2396-2411
33. Musacchio, A., Gibson, T., Rice, P., Thompson, J., and Saraste, M. (1993)
Trends Biochem Sci 18, 343-348
34. Fushman, D., Najmabadi-Haske, T., Cahill, S., Zheng, J., LeVine, H., 3rd, and
Cowburn, D. (1998) J Biol Chem 273, 2835-2843
35. Rossman, K. L., Worthylake, D. K., Snyder, J. T., Siderovski, D. P., Campbell,
S. L., and Sondek, J. (2002) Embo J 21, 1315-1326
36. Rameh, L. E., Arvidsson, A., Carraway, K. L., 3rd, Couvillon, A. D., Rathbun,
G., Crompton, A., VanRenterghem, B., Czech, M. P., Ravichandran, K. S.,
Burakoff, S. J., Wang, D. S., Chen, C. S., and Cantley, L. C. (1997) J Biol
Chem 272, 22059-22066
37. Saxena, A., Morozov, P., Frank, D., Musalo, R., Lemmon, M. A., Skolnik, E. Y.,
and Tycko, B. (2002) J Biol Chem 277, 49935-49944
38. Yu, J. W., Mendrola, J. M., Audhya, A., Singh, S., Keleti, D., DeWald, D. B.,
Murray, D., Emr, S. D., and Lemmon, M. A. (2004) Mol Cell 13, 677-688
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
23
39. Li, X., Rivas, M. P., Fang, M., Marchena, J., Mehrotra, B., Chaudhary, A., Feng,
L., Prestwich, G. D., and Bankaitis, V. A. (2002) J Cell Biol 157, 63-77
40. Funato, K., and Riezman, H. (2001) J Cell Biol 155, 949-959
41. De Matteis, M. A., and Godi, A. (2004) Nat Cell Biol 6, 487-492
42. Waugh, M. G., Minogue, S., Anderson, J. S., Balinger, A., Blumenkrantz, D.,
Calnan, D. P., Cramer, R., and Hsuan, J. J. (2003) Biochem J 373, 57-63
43. de Renzis, S., Sonnichsen, B., and Zerial, M. (2002) Nat Cell Biol 4, 124-133
44. Collins, T. J., and Bootman, M. D. (2003) J Exp Biol 206, 1993-2000
45. Staehelin, L. A. (1997) Plant J 11, 1151-1165
46. Thomas, C. C., Deak, M., Alessi, D. R., and van Aalten, D. M. (2002) Curr Biol
12, 1256-1262
47. Schneiter, R., Brugger, B., Sandhoff, R., Zellnig, G., Leber, A., Lampl, M.,
Athenstaedt, K., Hrastnik, C., Eder, S., Daum, G., Paltauf, F., Wieland, F. T.,
and Kohlwein, S. D. (1999) J Cell Biol 146, 741-754
48. Sikorski, R. S., and Hieter, P. (1989) Genetics 122, 19-27
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
24
Figure Legends:
Figure 1. Identification of second site within PHOsh1 that determines Golgi targeting.
PHOsh1, PHOsh2 and the indicated chimeras of these two homologous sequences were
expressed as GFP fusion proteins as indicated in RS453, a wild-type yeast strain,
and images taken of cells growing in log-phase. The different PH domains were: A.
PHOsh1 (105 residues); B. PHOsh2 (106 residues); C. residues 1-61 of PHOsh2 followed
by residues 61-105 of PHOsh1; D. PHOsh1 with residues 61-82 replaced with 62-83 of
PHOsh2. The region of 61-82 near the C-terminus of PHOsh1 was critical for Golgi
targeting. Fine dissection of this region then indicated that a single residue is
critical (see Table 2). E. PHOsh1 with mutation H79R; F. PHOsh2 with mutation R80H. ,
Substitution of arginine into PHOsh1 inhibited targeting, whereas histidine 79 brought
about Golgi targeting when inserted into PHOsh2. Plasmids used in A-F were pTL331,
pTL502, pTL503, pTL504, pTL508 and pTL509 respectively.
Fig. 2. Separation of the site that determines Golgi localization from the site that
binds PtdIns.
Ribbon diagram of the structure of PH domain from the kinase Akt (PHAkt) together
with Ins(1,3,4,5)-tetrakisphosphate ligand, the soluble headgroup of PtdIns(4,5)P2
(red and grey) drawn by CHIMERA software using known co-ordinates (46).
Features indicated are: loop β1-β2 (1), and loop β3-β4 (2) that encompass the
binding site of the ligand, leading to the predicted orientation of the domain with
respect to membrane lipids, which have been drawn to approximate scale (PH
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
25
domain diameter = approx 4 nm, Golgi membrane thickness = 7.3 nm (47),
phospholipid diameter 0.7 nm) ; and T77 of PHAkt (blue, arrowed), the residue
equivalent to H79 of PHOsh1 and R80 of PHOsh2, positioned on a side of the domain at
one side away from the known ligand binding site. Out of 14 solved PH domain
structures, the predicted secondary structure of the core β sheets and α helix in
PHOsh1 is most similar to PHAkt. The extra helix (asterisk) in loop β3-β4 of PHAkt is a
unique feature absent from PHOsh1/2 that is unlikely to affect the core structure.
Fig. 3. Binding of PHOsh2 to PtdIns(4)P and PtdIns(4,5)P2 in vitro.
GFP-tagged PH domains from PLCδ1, FAPP1, Osh1p and Osh2p (final concentration
0.5–0.8 µM) were incubated with increasing amounts of small unilamellar liposomes
made up from DOPC containing A. 2.5% PtdIns(4,5)P2 B. 2.5% PtdIns(4)P, or C.
neither (DOPC alone). Binding was detected as the decrease in the proportion of
GFP fluorescence remaining in the supernatant after centrifugation to remove
liposomes and bound GFP-PH domain. PHPlcδ1 specifically bound PtdIns(4,5)P2, and
the other PH domains bound both this and PtdIns(4)P, PHOsh2 alone showing a slight
preference for PtdIns(4,5)P2. None of the PH domains interacted with DOPC alone.
Fig. 4. Dual localisation of PHOsh2 dimer.
A. PHOsh2 was expressed as a tandemly repeated dimer of PHOsh2 extended
approximately 30 amino acids at each end (total of 169 amino acids) bracketed by
GFP (using pTL511). The dimer showed a dual localisation: at the plasma
membrane particularly enriched in small buds; and in numerous punctate structures
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
26
often seen at the bud-neck in large budded cells, which is the site of secretion
(arrows). The same dual localization was seen with another dimeric PHOsh2 construct
with only a single GFP, and much narrower definition of PHOsh2 (106 residues,
pTL512, data not shown).
B. Cells expressing GFP-PHOsh2-dimer as in A, co-stained with the endocytic tracer
FM4-64 (15 mins uptake), which has entered late Golgi compartments. In the
bottom panel, each image has been falsely coloured: GFP in green, FM4-64 in
magenta, with co-localization indicated in white. The very brightest punctae all
showed good co-localization, as do a proportion of the less bright punctae
(arrowhead). Using these settings, cells containing single markers showed complete
separation of the fluorophores (data not shown).
C. GFP-PHOsh2 dimer as in A, but both PH domains of the dimer carry mutations in
two basic residues implicated in PtdInsP binding (K307E R309E, pTL513). The
construct was entirely cytoplasmic, indicating loss of both of the localisations of
wild-type PHOsh2.
Fig. 5. Different PtdIns 4-kinases synthesize the two pools of PtdIns(4)P.
All images are of the GFP-PHOsh2-dimer expressed (from pTL511) in the indicated
mutant strains and the corresponding wild-type parental strains, grown to mid-log
phase at the permissive temperature (25˚C) and shifted to a strongly non-
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
27
permissive temperature (39˚C) for 15 minutes prior to imaging. A. Inactivation of
Pik1p completely inhibited the punctate localization of PHOsh2. B. Targeting to both
sites was unaffected by inactivation of Mss4p, indicating no requirement for
PtdIns(4,5)P2 for localization. C. Plasma membrane localisation was lost selectively
on inactivation of Stt4p, but not in the wild-type control. Plasma membrane
targeting was also reduced in the stt4-4 mutant strain at a temperature permissive
for growth (25˚C, data not shown).
Fig. 6. PHOsh2 and other Golgi-targeted PH domains reveal an altered distribution of
PtdIns(4)P in cells lacking Sac1p.
Images of GFP-tagged PH domains expressed in a ∆sac1 strain, grown to mid-log
phase at 30˚C. A. GFP-PHOsh2-dimer, B. GFP-PHOsh2-monomer, C. GFP-PHOsh1, D.
GFP-PHOSBP, E. GFP-PHFAPP, F. GFP-PHCERT and G. GFP-PHCERT* carrying the mutation
R43E . The constructs used are indicated diagrammatically above each image (2–2,
2, 1, O, F, C, C*), and were expressed from pTL511, pTL342, pTL331, pTL332,
pTL334, pTL333 and pTL514 respectively. Deletion of ∆sac1 shifted dimeric PHOsh2
from punctate to plasma membrane targeting. Monomeric PHOsh2 showed enhanced
targeting to the plasma membrane (compare with Fig 1A). In comparison, PHOsh1,
PHOSBP and PHFAPP1 targeted punctae predominantly in ∆sac1 cells. In addition, these
sequences showed minor targeting to the nuclear envelope (arrows) and cell
periphery (arrowheads). PHCERT was unique in targeting the ER (nuclear envelope
and peripheral patches). This was dependent on binding to PtdInsPs, as is was not
abolished by mutating the critical basic residue in PHCERT (R43) to an acid (E).
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
28
Table 1. Plasmids used.
pRS406 Integrates at URA3 ref (48)
pRS416 CEN URA3 (low copy) ref (48)
pTL331 pRS406 PHO5 prom., GFP, myc, PHOsh1 (105 aa = 280–122 aa) ref (25)
pTL332 pRS406 PHO5 prom., GFP, myc, PHOSBP (99 aa = 87-185 aa) ref (12)
pTL333 pRS406 PHO5 prom., GFP, myc, PHCERT (=GPBP) (122 aa = 1-122 aa) ref (12)
pTL334 pRS406 PHO5 prom., GFP, myc, PHFAPP1 (98 aa = 1-98 aa) ref (12)
pTL342 pRS416 PHO5 prom., GFP, PHOsh2 (256–424 aa), GFP ref (25)
pTL502 pRS406 PHO5 prom., GFP, myc, PHOsh2 (106 aa = 286-391 aa)
391K391
this study
pTL503 pRS406 PHO5 prom., GFP, myc, PHOsh2 (aa 1-61) – PHOsh1 (aa 61-105) this study
pTL504 pRS406 PHO5 prom., GFP, myc, PHOsh1 (aa 61-82 62-83 of PHOsh2) this study
pTL505 pRS406 PHO5 prom., GFP, myc, PHOsh1 (aa 61-70 62-71 of PHOsh2) this study
pTL506 pRS406 PHO5 prom., GFP, myc, PHOsh1 (aa 71-82 72-83 of PHOsh2) this study
pTL507 pRS406 PHO5 prom., GFP, myc, PHOsh1 (aa 71-77 72-78 of PHOsh2) this study
pTL508 pRS406 PHO5 prom., GFP, myc, PHOsh1 as pTL331 (with mutation H79R) this study
pTL509 pRS406 PHO5 prom., GFP, myc, PHOsh2 as pTL502 (with mutation R80H) this study
pTL510 pTrcHisA His6, GFP, myc, PHOsh2 (106 aa)
391K391
this study
pTL511 pRS416 PHO5 prom., GFP, PHOsh2 (169 aa = 256-424 aa) – VNSKL linker –
PHOsh2 repeat - GFP
391K391
this study
pTL512 pRS406 PHO5 prom., GFP, PHOsh2
(106 aa) – KGAGAGAGS linker – PHOsh2 repeat
this study
pTL513 pTL511 carrying mutations K307E R309E in both copies of PHOsh2 this study
pTL514 pTL333 carrying mutation R43E in PHCERT this study
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine, 2004
29
Table 2. Analysis of the section of PHOsh1 that determines Golgi targeting.
GFP-tagged PHOsh1 constructs which varied only in the sequences in the fourth out of
five segments of PHOsh1 (residues 61-82) were scored for their punctate localization.
Introduction of the Osh2p sequence (construct 1) made 8 changes and rendered
membrane targeting only weakly detectable (w). While the N-terminal 3 variant
residues had little effect on targeting (construct 2), the C-terminal 5 variant
residues were responsible for the majority of targeting (construct 3). Introduction
of the first 4 of these 5 changes had no effect on targeting (construct 4), indicating
an important role for the last variant residue: H79 in PHOsh1 (R80 in PHOsh2).
construct plasmid Sequence from residues 61-82
in PHOsh1
# changes
cf Osh1p
punctae
Osh1p pTL331 SEKLKFEIIGGNNGVIRWHLKG 0 ++
1 pTL504 SEKLNFELLGGITGTTRWRLKG 8 w
2 pTL505 SEKLNFELLGGNNGVIRWHLKG 3 +
3 pTL506 SEKLKFEIIGGITGTTRWRLKG 5 w
4 pTL507 SEKLKFEIIGGITGTTRWHLKG 4 ++
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

A B
C
E
D
FR80HH79R
Roy and Levine Figure 1
G 2G 1
G 1 G 2
G G
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine Figure 2
*1 2
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from
7035
B
0
C100
60
200
80
40
70350
PtdInsP added (µM)
Roy and Levine Figure 3
0 8040
A100
60
200
80
40
% u
nb
ou
nd
GF
P-P
H
PtdIns(4,5)P2 PtdIns(4)P
DOPC only
100
60
200
80
40
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roy and Levine Figure 4
A BG 2 G2 GFP
FM4-64
mergeG G
C2* *2
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from
Roy and Levine Figure 5
A
B
C
pik1-83wild-type
stt4-4wild-type
mss4-2wild-type
G 2 G2
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from
F
Roy and Levine Figure 6
G 2 G2 G 2 G
all in ∆sac1 cells
G OG 1
G CG F
G *C
A B
C
E
G
D
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Anjana Roy and Timothy P. Levineof Osh2p
Multiple pools of PtdIns 4-phosphate detected using the pleckstrin homology domain
published online July 22, 2004J. Biol. Chem.
10.1074/jbc.M401583200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 13, 2018http://w
ww
.jbc.org/D
ownloaded from
Related Documents











