This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
www.elsevier.com/locate/brainres
Available online at www.sciencedirect.com
Research Report
Multiple effects of copper on NMDA receptor currents
Carla Marchettia,n, Irena Baranowska-Bosiackab, Paola Gavazzoa
aInstitute of Biophysics, National Research Council, Genova, ItalybDepartment of Biochemistry and Medical Chemistry, Pomeranian Medical University, Szczecin, Poland
a r t i c l e i n f o
Article history:
Accepted 16 October 2013
Available online 23 October 2013
Keywords:
Copper
GluN1/GluN2A receptor
GluN1/GluN2B receptor
Cerebellar granule cells
Nickel
Redox site
a b s t r a c t
Copper (Cu) is an essential metal present in the human brain and released from synaptic
vesicles following neuronal depolarization. Cu is known to reduce the NMDA receptor (NR)
current with IC50E20 mM. We have studied the effect of Cu on the NR current in cultured
neonatal rat cerebellum granule cells (CGC) and in transiently transfected HEK293 cells
(HEK), expressing either GluN1/GLUN2A or GluN1/GluN2B receptors. In CGCs, Cu causes a
potentiation of the NR current at concentrations o30 mM (EC50¼4.6 mM) and a block at
higher concentrations (IC50¼24 mM). In Fura2 loaded CGCs, Cu (r30 mM) caused an increase
of NMDA-driven calcium influx. This facilitating effect was prevented by pre-treatment
with the reducing agent DTT. Cu also caused an increase of the NR current in GluN1/
GluN2A receptors (EC50¼2 mM) and a block at higher concentrations (IC50¼26 mM). Both
facilitation and inhibition were independent of voltage. The effect of Cu was quantitatively
similar in GluN1/GluN2B receptors, which were potentiated by 10 mM and inhibited by
100 mM Cu. Potentiation was absent in mutants deleted of their entire amino terminal
domain (ATD) of the protein, suggesting an involvement of this region in the interaction.
These results indicate that Cu can facilitate the NR current at lower concentrations than
those required for blocking it; this effect can have consequences on the activity of the
metal at synaptic and extrasynaptic sites.
& 2013 Elsevier B.V. All rights reserved.
1. Introduction
Glutamate is the major excitatory neurotransmitter in thevertebrate central nervous system. It acts pre- and post-synaptically through different classes of ionotropic and meta-botropic receptors, which work in concert to mediate manyforms of synaptic plasticity and, as a consequence, high brainfunctions, such as memory and spatial learning, cognition andbehavior (Traynelis et al., 2010). On the other hand, excessive
activation of ionotropic glutamate receptors causes a degen-eration called excitotoxicity, involved in neurological diseases,stroke, head trauma and epilepsy. Excitotoxicity is mainlycaused by elevated influx of Ca through the Ca-permeableN-methyl-D-aspartate (NMDA) subtype of glutamate receptor(NR) (Choi, 1992). NR is a ligand-gated ionic channel endowedwith unique properties: it requires two agonists, glutamateand glycine, to activate and its permeability is modulated byMg in a voltage-dependent manner. NRs are most commonly
0006-8993/$ - see front matter & 2013 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.brainres.2013.10.029
Abbreviations: NMDA, N-methyl-D-aspartate; NR, NMDA receptor; GluN1, NMDA receptor subunit 1; GluN2A, NMDA receptor
subunit 2A; GluN2B, NMDA receptor subunit 2B; HEPES, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; DTT, Dithiothreitol;
DTNB, 5,5′-dithiobis-(2-nitrobenzoic acid).nCorrespondence to: Istituto di Biofisica, Consiglio Nazionale delle Ricerche, via De Marini 6, 16149 Genova, Italy. Fax: þ39 010 6475500.E-mail address: [email protected] (C. Marchetti).
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1
Author's personal copy
assembled from two types of subunits, the essential GluN1and at least one, but more frequently two types of GluN2 (A,B,C,D) and contain a number of distinct recognition sites forendogenous and exogenous ligands, including metal ionsand redox active reagents, which are able to modulate theirfunctions (Paoletti et al., 2013).
Copper (Cu) is an essential transition metal that is requiredas a cofactor for the correct functioning of vital enzymes andplays major roles in brain metabolism (Linder and Hazegh-Azam, 1996). Its deficiency or deregulation is part of theetiology of very serious inherited diseases (Lutsenko andPetris, 2003), while high level of Cu can be toxic due to theproduction of free radicals (Valko et al., 2005; Pal et al., 2013).Physiologically, Cu is present in both oxidized (Cu2þ) andreduced (Cuþ) state and can bind proteins in both states,preferentially through thiol groups. Cu is abundant in thevertebrate brain where its extracellular concentration wasreported from less than 1 to a few mM (Gaier et al., 2013;Mathie et al., 2006; Stys et al., 2012). It is released duringneuronal activity (Hartter and Barnea, 1988; Kardos et al., 1989;Hopt et al., 2003) and its levels in the synaptic cleft can rise toas high as a few hundred micromolar (White and Cappai, 2003;Millhauser, 2007). Previous studies reported Cu-mediated inhi-bition of neurotransmitter-activated channels, including GABA(Xie et al., 1994; Trombley and Shepherd, 1996) and glutamate(NMDA and AMPA, Trombley and Shepherd, 1996; Weiser and
Wienrich, 1996) receptor channels, as well as voltage-gated ionchannels (Horning and Trombley, 2001). In particular, Cu is anestablished NR antagonist (Trombley and Shepherd, 1996;Vlachova et al., 1996; Weiser and Wienrich, 1996) and inhibitsNR-dependent LTP in hippocampal slices (Doreulee et al., 1997;Leiva et al., 2009; Salazar-Weber and Smith, 2011).
The role of Cu in human CNS disease is still poorlyunderstood, but a strict connection between Cu and NRactivity has been postulated and described in several studies(Stys et al., 2012). This connection was evident in the criticalrole of Cu in the pathogenesis of Alzheimer disease (AD,Hung et al., 2010) and in the link between copper homeostasisand NR activity (Schlief and Gitlin, 2006). Other recent worksshowed that NR activity is modulated by cellular prionprotein (PrPC) in a Cu-dependent manner (You et al., 2012)and this interaction may be important not only in scrapiedisease, but also in AD, epilepsy, pain and depression (Styset al., 2012). All these studies underscore the link between Cuand NR receptor activity and stress the importance of char-acterizing the interaction of this crucial metal and thisprotein. In this work, we have undertaken research tocharacterize Cu effect at the molecular level by studying theaction of Cu on native neuronal and recombinant NR chan-nels. Our experiments provided evidence of an additionaleffect of Cu on NR channels: at relatively low concentrations,Cu can enhance the activity of NR channels, both in primary
Cu (µM)1 10 100 1000
I Cu/
I con
0.0
0.5
1.0
1.5
2.01 Cu
10 Cu
100 Cu
5 s
100pA
NMDA
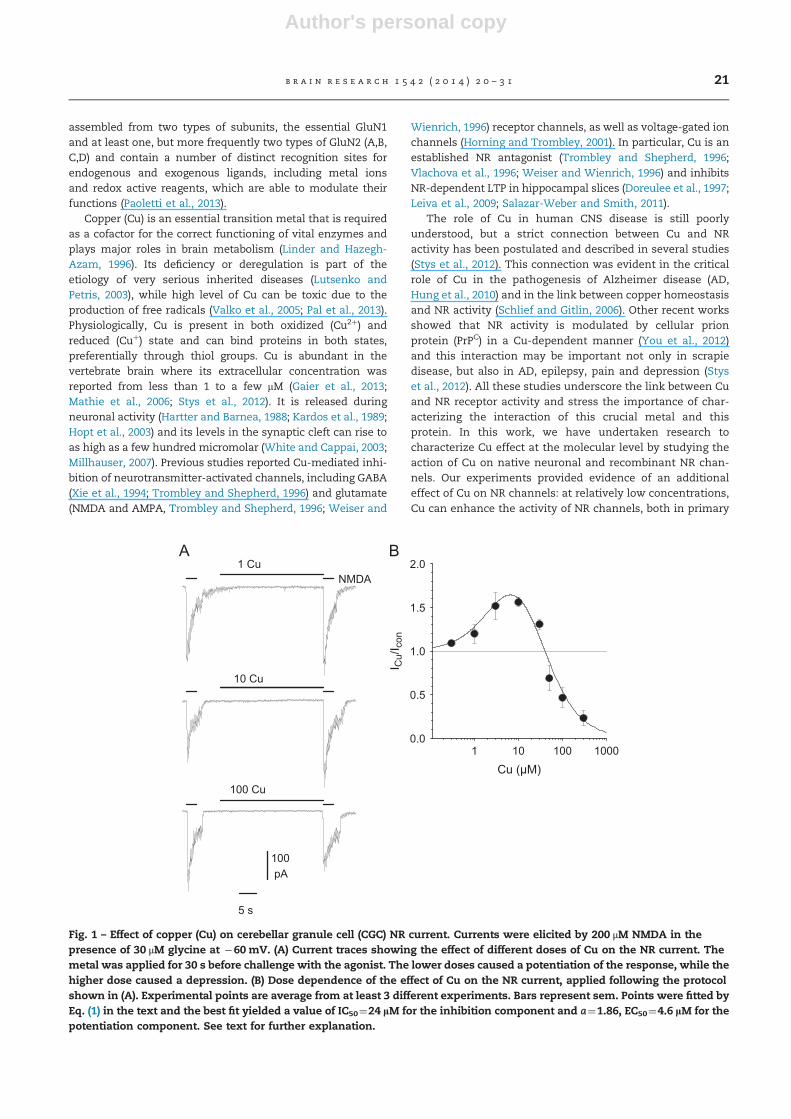
Fig. 1 – Effect of copper (Cu) on cerebellar granule cell (CGC) NR current. Currents were elicited by 200 μM NMDA in thepresence of 30 μM glycine at �60 mV. (A) Current traces showing the effect of different doses of Cu on the NR current. Themetal was applied for 30 s before challenge with the agonist. The lower doses caused a potentiation of the response, while thehigher dose caused a depression. (B) Dose dependence of the effect of Cu on the NR current, applied following the protocolshown in (A). Experimental points are average from at least 3 different experiments. Bars represent sem. Points were fitted byEq. (1) in the text and the best fit yielded a value of IC50¼24 lM for the inhibition component and a¼1.86, EC50¼4.6 lM for thepotentiation component. See text for further explanation.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 21
Author's personal copy
neurons and in transiently transfected cells expressing eitherGluN1/GluN2A or GluN1/GluN2B receptor.
2. Results
2.1. Effect of Cu on NMDA currents in cerebellar granulecells
In neonatal rat cerebellar granule cells (CGC), we studied theaction of Cu on the NR current, measured in whole-cellvoltage-clamp and elicited by 200 mM NMDA in the presenceof 30 mM glycine and 0 Mg. The aim was to characterize thealready reported antagonist effect of the metal ion Cu2þ,which we expect to be the predominant ionic copper speciesin solution. However, it was evident in early experiments thatCu also exerted a potentiating activity on the NR current andthis observation prompted us to study this effect. Cu wasapplied for different time courses immediately before, but notduring challenge with NMDA. This protocol was adopted tominimize chelating effects (Vlachova et al., 1996). With a 30 streatment, the current was increased by 1–30 mM Cu, while itwas reduced at higher concentrations (Fig. 1). The dosedependence could be approximated by the product of aninhibitory component and a stimulatory component, withHill slope fixed to 1 (Marchetti and Gavazzo, 2003), i.e. with
the equation:
ICu=Icon ¼ 1=½1þ ð½Cu�=IC50Þ� � ð1þ a=ð½1þ ðEC50=½Cu�ÞÞ ð1Þ
where ICu/Icon is the ratio between the current in thepresence of Cu and the current in control, [Cu] is the concen-tration of added Cu, IC50 is the Cu dose which provokes a 50%block of the NMDA current, a is the maximum potentiation andEC50 is the dose that causes 50% of the maximum potentiation.The best fit yielded a value of IC50¼24 mM for the inhibitioncomponent and a¼1.86, EC50¼4.6 mM for the potentiationcomponent (Fig. 1B).
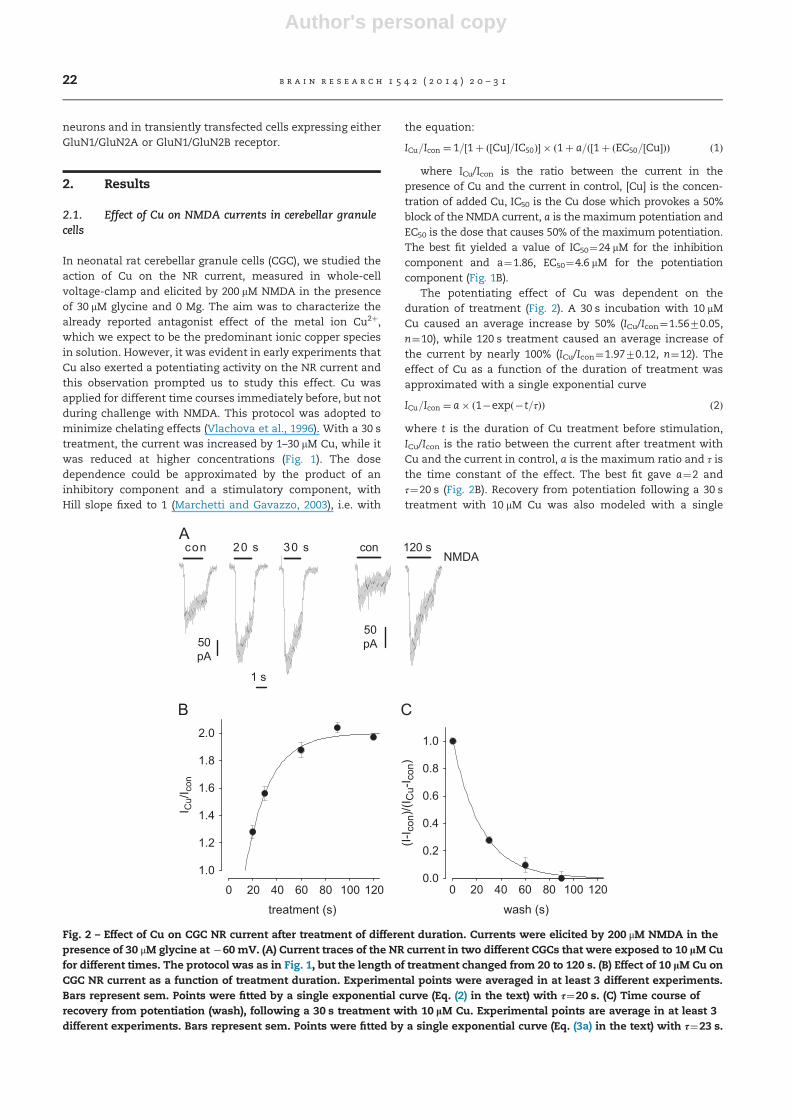
The potentiating effect of Cu was dependent on theduration of treatment (Fig. 2). A 30 s incubation with 10 mMCu caused an average increase by 50% (ICu/Icon¼1.5670.05,n¼10), while 120 s treatment caused an average increase ofthe current by nearly 100% (ICu/Icon¼1.9770.12, n¼12). Theeffect of Cu as a function of the duration of treatment wasapproximated with a single exponential curve
ICu=Icon ¼ a� ð1�expð�t=τÞÞ ð2Þ
where t is the duration of Cu treatment before stimulation,ICu/Icon is the ratio between the current after treatment withCu and the current in control, a is the maximum ratio and τ isthe time constant of the effect. The best fit gave a¼2 andτ¼20 s (Fig. 2B). Recovery from potentiation following a 30 streatment with 10 mM Cu was also modeled with a single
con 120 s
50pA
1 s
50pA
con 20 s 30 sNMDA
treatment (s)0 20 40 60 80 100 120
I Cu/
I con
1.0
1.2
1.4
1.6
1.8
2.0
wash (s)
0 20 40 60 80 100 120
(I-I c
on)/(
I Cu-
I con
)
0.0
0.2
0.4
0.6
0.8
1.0
Fig. 2 – Effect of Cu on CGC NR current after treatment of different duration. Currents were elicited by 200 μM NMDA in thepresence of 30 μM glycine at �60 mV. (A) Current traces of the NR current in two different CGCs that were exposed to 10 lM Cufor different times. The protocol was as in Fig. 1, but the length of treatment changed from 20 to 120 s. (B) Effect of 10 lM Cu onCGC NR current as a function of treatment duration. Experimental points were averaged in at least 3 different experiments.Bars represent sem. Points were fitted by a single exponential curve (Eq. (2) in the text) with τ¼20 s. (C) Time course ofrecovery from potentiation (wash), following a 30 s treatment with 10 lM Cu. Experimental points are average in at least 3different experiments. Bars represent sem. Points were fitted by a single exponential curve (Eq. (3a) in the text) with τ¼23 s.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 122

Author's personal copy
exponential curve
ðI� IconÞ=ðICu� IconÞ ¼ expð�t=τÞ ð3aÞ
where t is the duration of wash, Icon is the current in theabsence of Cu (before treatment and after complete recovery),ICu is the current in the presence of Cu and τ is the timeconstant of the recovery. The best fit gave τ¼23 s (Fig. 2C).
As the potentiation had a relatively slow onset and recovery,we thought it might be mediated by structural modification ofthe protein, such as those caused by oxidation and reduction ofdisulfide bridges at the redox modulatory site (Aizenman et al.,1990; Choi and Lipton, 2000). Treatment (2 min) with thereducing agent dithiothreitol (DTT, 1 mM) always caused anincrease of CGC NR current (IDTT/Icon¼1.8870.10; n¼8), whichrecovered to control value only after prolonged wash (see Fig. 3).In particular, a 20 s wash was never effective in reversing theeffect of DTT. So, Cu was applied immediately after DTTtreatment and, in this condition, a 20 s application of 10 mMCu never caused any increase in the current, but a moderatedecrease (ICu/Icon¼0.8370.09, n¼5). After a prolonged wash ofDTT, the potentiating effect of Cu was restored (Fig. 3).
2.2. Effect of Cu on NMDA-driven Ca influx in Fura2loaded cerebellar granule cells
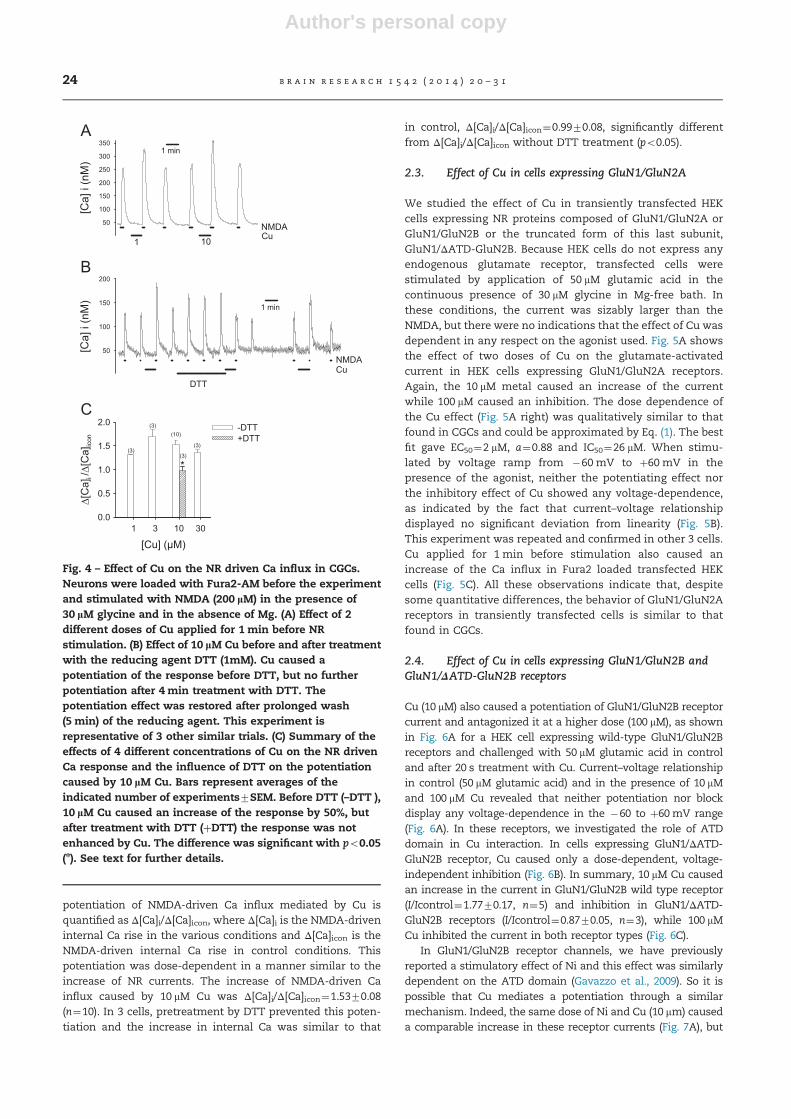
In Fura2-loaded CGCs, challenge with 200 mM NMDA in theabsence of Mg always caused a rise of internal Ca, from basallevel of 4074 nM to 230713 nM (n¼40). The internal Ca levelrecovered to the basal level in approximately 2 min. When Cuwas applied for 1 min before the stimulus with NMDA, thesubsequent response was potentiated in a dose-dependentmanner and recovered to the initial value after wash of themetal (Fig. 4A). The reducing agent DTT (1 mM, Fig. 4B) alsocaused potentiation of Ca influx, while the oxidizing agentDTNB (0.5 mM, not shown) caused a depression of the sub-sequent response, when applied before the stimulus. Whenneurons were subjected to prolonged treatment with DTT(4min in Fig. 4B), a subsequent application of Cu (10 mM,1 min) did not cause any increase, but rather a slight decreaseof the response with respect to control. Similar to the observa-tion on the currents (Fig. 3), the potentiating effect of Cu wasrestored after prolonged (45 min) wash of DTT. These resultsare summarized in the graph of Fig. 4C. In this graph, the
10 Cu
control
after DTTtreatment
wash DTT
NMDA
-DTT +DTT
I/Ico
n
0.0
0.5
1.0
1.5
50pA
3 s
*
t (min)0 5 10 15 20 25
I/Ico
n
0.0
0.5
1.0
1.5
2.0
2.5 DTT
Cu
Fig. 3 – Effect of treatment with the reducing agent DTT on Cu potentiation of CGC NR current. Currents were elicited by 200 μMNMDA in the presence of 30 μM glycine at �60 mV. (A) Effect of Cu on CGC NR current before and after DTT treatment andwash. The upper trace (control) was recorded with the same condition as Fig. 1A, but treatment with Cu lasted 20 s. Cu causedan enhancement that recovered after wash of the metal. Then the cell was incubated for 2 min with 1 mM DTT. The middletrace (after DTT treatment) shows the effect of Cu immediately after the treatment with DTT. The current was increased byDTT, but Cu did not cause any further increase. Prolonged (5 min) wash restored the current to the initial value. The lowertrace (wash DTT) shows the potentiating effect of Cu after wash of DTT. (B) Time course of the effect of DTT and Cu in 3 cells.Application of DTT caused a current increase, which persisted as long as the reducing agent was present. Then DTT waswashed and Cu (10 lM ) was applied for 20 s, but did not caused any increase. The potentiating effect of Cu was restored afterprolonged wash (5 min) of DTT. (C) Summary of the influence of DTT on Cu potentiation in CGC NR current. Bars representaverage of 5 experiments7SEM. Before DTT (–DTT), 10 lM Cu caused an increase of the current by 35%, while no potentiationwas present after treatment with DTT (þDTT). nindicates a significant difference with po0.05.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 23
Author's personal copy
potentiation of NMDA-driven Ca influx mediated by Cu isquantified as Δ[Ca]i/Δ[Ca]icon, where Δ[Ca]i is the NMDA-driveninternal Ca rise in the various conditions and Δ[Ca]icon is theNMDA-driven internal Ca rise in control conditions. Thispotentiation was dose-dependent in a manner similar to theincrease of NR currents. The increase of NMDA-driven Cainflux caused by 10 mM Cu was Δ[Ca]i/Δ[Ca]icon¼1.5370.08(n¼10). In 3 cells, pretreatment by DTT prevented this poten-tiation and the increase in internal Ca was similar to that
in control, Δ[Ca]i/Δ[Ca]icon¼0.9970.08, significantly differentfrom Δ[Ca]i/Δ[Ca]icon without DTT treatment (po0.05).
2.3. Effect of Cu in cells expressing GluN1/GluN2A
We studied the effect of Cu in transiently transfected HEKcells expressing NR proteins composed of GluN1/GluN2A orGluN1/GluN2B or the truncated form of this last subunit,GluN1/ΔATD-GluN2B. Because HEK cells do not express anyendogenous glutamate receptor, transfected cells werestimulated by application of 50 mM glutamic acid in thecontinuous presence of 30 mM glycine in Mg-free bath. Inthese conditions, the current was sizably larger than theNMDA, but there were no indications that the effect of Cu wasdependent in any respect on the agonist used. Fig. 5A showsthe effect of two doses of Cu on the glutamate-activatedcurrent in HEK cells expressing GluN1/GluN2A receptors.Again, the 10 mM metal caused an increase of the currentwhile 100 mM caused an inhibition. The dose dependence ofthe Cu effect (Fig. 5A right) was qualitatively similar to thatfound in CGCs and could be approximated by Eq. (1). The bestfit gave EC50¼2 mM, a¼0.88 and IC50¼26 mM. When stimu-lated by voltage ramp from �60 mV to þ60 mV in thepresence of the agonist, neither the potentiating effect northe inhibitory effect of Cu showed any voltage-dependence,as indicated by the fact that current–voltage relationshipdisplayed no significant deviation from linearity (Fig. 5B).This experiment was repeated and confirmed in other 3 cells.Cu applied for 1 min before stimulation also caused anincrease of the Ca influx in Fura2 loaded transfected HEKcells (Fig. 5C). All these observations indicate that, despitesome quantitative differences, the behavior of GluN1/GluN2Areceptors in transiently transfected cells is similar to thatfound in CGCs.
2.4. Effect of Cu in cells expressing GluN1/GluN2B andGluN1/ΔATD-GluN2B receptors
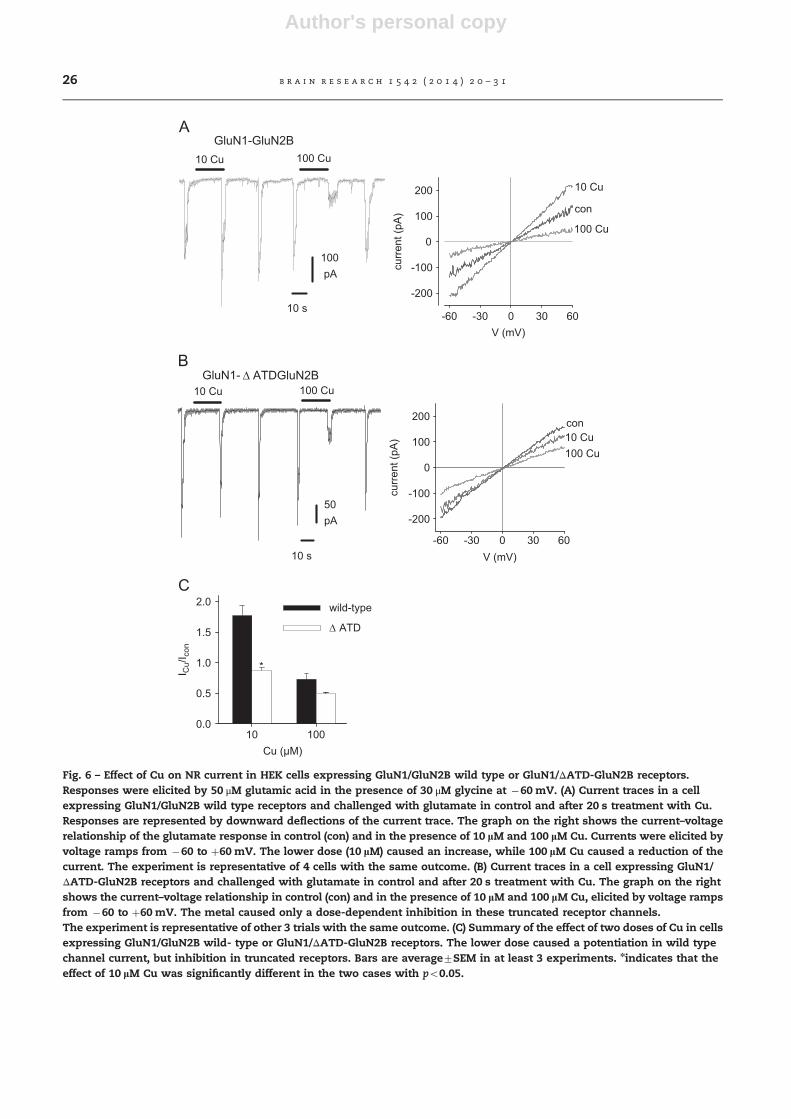
Cu (10 mM) also caused a potentiation of GluN1/GluN2B receptorcurrent and antagonized it at a higher dose (100 mM), as shownin Fig. 6A for a HEK cell expressing wild-type GluN1/GluN2Breceptors and challenged with 50 mM glutamic acid in controland after 20 s treatment with Cu. Current–voltage relationshipin control (50 mM glutamic acid) and in the presence of 10 mMand 100 mM Cu revealed that neither potentiation nor blockdisplay any voltage-dependence in the �60 to þ60mV range(Fig. 6A). In these receptors, we investigated the role of ATDdomain in Cu interaction. In cells expressing GluN1/ΔATD-GluN2B receptor, Cu caused only a dose-dependent, voltage-independent inhibition (Fig. 6B). In summary, 10 mM Cu causedan increase in the current in GluN1/GluN2B wild type receptor(I/Icontrol¼1.7770.17, n¼5) and inhibition in GluN1/ΔATD-GluN2B receptors (I/Icontrol¼0.8770.05, n¼3), while 100 mMCu inhibited the current in both receptor types (Fig. 6C).
In GluN1/GluN2B receptor channels, we have previouslyreported a stimulatory effect of Ni and this effect was similarlydependent on the ATD domain (Gavazzo et al., 2009). So it ispossible that Cu mediates a potentiation through a similarmechanism. Indeed, the same dose of Ni and Cu (10 mm) causeda comparable increase in these receptor currents (Fig. 7A), but
[Ca]
i (n
M)
50
100
150
200
250
300
3501 min
101Cu NMDA
[Ca]
i (n
M)
50
100
150
200
NMDA Cu
1 min
DTT
[Cu] (µM)31
Δ[C
a]i /Δ
[Ca]
icon
0.0
0.5
1.0
1.5
2.0 -DTT+DTT
*(3)
(3)(10)
(3)
(3)
3010
Fig. 4 – Effect of Cu on the NR driven Ca influx in CGCs.Neurons were loaded with Fura2-AM before the experimentand stimulated with NMDA (200 lM) in the presence of30 lM glycine and in the absence of Mg. (A) Effect of 2different doses of Cu applied for 1 min before NRstimulation. (B) Effect of 10 lM Cu before and after treatmentwith the reducing agent DTT (1mM). Cu caused apotentiation of the response before DTT, but no furtherpotentiation after 4 min treatment with DTT. Thepotentiation effect was restored after prolonged wash(5 min) of the reducing agent. This experiment isrepresentative of 3 other similar trials. (C) Summary of theeffects of 4 different concentrations of Cu on the NR drivenCa response and the influence of DTT on the potentiationcaused by 10 lM Cu. Bars represent averages of theindicated number of experiments7SEM. Before DTT (–DTT ),10 lM Cu caused an increase of the response by 50%, butafter treatment with DTT (þDTT) the response was notenhanced by Cu. The difference was significant with po0.05(n). See text for further details.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 124

Author's personal copy
the time course of the effects was different. Cu potentiation ofGluN1/GluN2B receptor current increased with the duration oftreatment and was larger for longer incubations (Fig. 7B). Thepotentiation caused by Ni (10 mM ) was independent of theduration of treatment and already maximum for incubation410 s (Fig. 7C).
Recovery from potentiation followed a single exponentialtime course after Ni and was modeled with a single expo-nential curve
ðI� IconÞ=ðIm� IconÞ ¼ expð�t=τÞ ð3bÞ
where Icon is the current before treatment and after completerecovery, Im is the current in the presence of the metal (Ni inthis case) and τ is the time constant of the recovery. The bestfit gave τ¼15 s (Fig. 7D, filled circles).
In contrast, recovery from potentiation after Cu treatmenthad a double exponential time course and it was modeledwith the sum of two exponential curves:
ðI� IconÞ=ðIm� IconÞ ¼ a1expð�t=τ1Þ þ a2expð�t=τ2Þ ð3cÞ
where Icon is the current before treatment and after completerecovery, Im is the current in the presence of the metal (Cu inthis case), a1 and a2 are the relative amplitudes, and τ1 and τ2the time constants of the two components. The best fita1¼0.48, a2¼0.52, τ1¼11 s and τ2¼ 165 se (Fig. 7D, emptycircles). So the shortest time constant was similar for the twometals, but the time course of recovery from Cu potentiationcontained an additional, slower component.
3. Discussion
This work describes distinct effects of Cu on native andrecombinant NR channels, activated by saturating doses ofboth agonists (glycine and glutamate or NMDA). Cu has beenpreviously reported to inhibit NR channels with IC50 close to
GluN1-GluN2A
100pA
10 sCu (µM)
1
I Cu/
I con
0.0
0.5
1.0
1.5
100 Cu10 Cu
100010010
V (mV)-60
curr
ent (
pA)
-400
-200
0
200 con
10 Cu
100 Cu[C
a]i (
nM)
50
100
150
20030 s
glutcon 6040200-20-40 washCu
Fig. 5 – Effect of Cu on the NR current in HEK cells expressing GluN1/GluN2A receptors. (A) Current traces showing currentfacilitation following treatments with 10 lM and block after 100 lM Cu. Responses to 50 μM glutamic acid were elicited by 2 sperfusion in the presence of 30 μM glycine at �60 mV and are represented by downward deflections of the current trace.The graph on the right shows the dose-dependence of the effect of Cu, applied with the same protocol. Experimental pointsare average from at least 3 different experiments. Bars represent sem. Points were fitted by Eq. (1) (see Fig. 1 and text) and thebest fit yielded a value of EC50¼2 lM, a¼0.88 for the potentiation component and IC50¼26 lM for the inhibition component.(B) Current–voltage relationship in control (50 lM glutamic acid) and in the presence of 10 lM and 100 lM Cu. Currents wereelicited by voltage ramps from �60 to þ60 mV. The lower dose (10 lM) caused an increase, while 100 lM Cu caused areduction of the current in the whole voltage range. This experiment was repeated for 3 additional cells with the sameoutcome. (C) Glutamate-driven Ca influx in HEK cells expressing GluN1/GluN2A receptors loaded with Fura2 (left), after 1 mintreatment with 10 lM Cu (middle) and after wash of the metal (right). Ca influx was induced by 50 lM glutamic acid (glut) inthe presence of 30 lM glycine.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 25
Author's personal copy
GluN1-GluN2B
100pA
10 s
100 Cu10 Cu
V (mV)-60
curr
ent (
pA)
-200
-100
0
100
200
con
10 Cu
100 Cu
60300-30
GluN1- Δ ATDGluN2B10 Cu 100 Cu
50pA
10 s V (mV)-60
curr
ent (
pA)
-200
-100
0
100
200 con10 Cu100 Cu
Cu (µM)10
I Cu/
I con
0.0
0.5
1.0
1.5
2.0 wild-type
Δ ATD
*
60300-30
100
Fig. 6 – Effect of Cu on NR current in HEK cells expressing GluN1/GluN2B wild type or GluN1/ΔATD-GluN2B receptors.Responses were elicited by 50 μM glutamic acid in the presence of 30 μM glycine at �60 mV. (A) Current traces in a cellexpressing GluN1/GluN2B wild type receptors and challenged with glutamate in control and after 20 s treatment with Cu.Responses are represented by downward deflections of the current trace. The graph on the right shows the current–voltagerelationship of the glutamate response in control (con) and in the presence of 10 lM and 100 lM Cu. Currents were elicited byvoltage ramps from �60 to þ60 mV. The lower dose (10 lM) caused an increase, while 100 lM Cu caused a reduction of thecurrent. The experiment is representative of 4 cells with the same outcome. (B) Current traces in a cell expressing GluN1/ΔATD-GluN2B receptors and challenged with glutamate in control and after 20 s treatment with Cu. The graph on the rightshows the current–voltage relationship in control (con) and in the presence of 10 lM and 100 lM Cu, elicited by voltage rampsfrom �60 to þ60 mV. The metal caused only a dose-dependent inhibition in these truncated receptor channels.The experiment is representative of other 3 trials with the same outcome. (C) Summary of the effect of two doses of Cu in cellsexpressing GluN1/GluN2B wild- type or GluN1/ΔATD-GluN2B receptors. The lower dose caused a potentiation in wild typechannel current, but inhibition in truncated receptors. Bars are average7SEM in at least 3 experiments. nindicates that theeffect of 10 lM Cu was significantly different in the two cases with po0.05.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 126

Author's personal copy
20 mM, but the range of values reported in the different studieswas quite wide (0.27–30 mM; Trombley and Shepherd, 1996;Vlachova et al., 1996) and the molecular mechanism under-lying inhibition of NR activity is largely unknown. The presentwork was undertaken to investigate and characterize Cuinhibition, but we soon acquired evidence that Cu canmodulate the NR current also in a positive direction andcause facilitation, although at doses 430 mM the blockingeffect always prevails. The potentiation effect was evidentin whole-cell clamp recordings of NR membrane currents incultured CGCs, as well as in HEK293 cells that had beentransiently transfected with GluN1/GluN2A or GluN1/GluN2B.The potentiation of the response was also observed in mea-surements of internal Ca in Fura2-loaded cells, in both CGCsand transiently transfected cells expressing GluN1/GluN2A
channels. Thus the facilitation causes an enhanced elevationof the intracellular Ca level.
To study the facilitating effect of Cu, we used a protocol(see Experimental procedures section) that minimized com-plex formation of the metal with glutamate or NMDA andconsequent loss of effect at high agonist concentration(Vlachova et al., 1996). The observations that (i) Cu does notneed to be present during challenge with the agonist, and (ii)the facilitating effect is independent of voltage (Figs. 5 and 6)suggest that Cu is acting through a site external to theconducting pathway and accessible when the NR channel isclosed. Therefore it appears that, similar to other divalentcations such as Mg (Paoletti et al., 1995), Zn (Paoletti et al.,1997) and Ni (Gavazzo et al., 2009), Cu is an allosteric mod-ulator of the NR.
treatment (s)0 20 40 60 80 100 120
I m/I c
on
0.0
0.5
1.0
1.5
2.0
wash (s)0 60 120 180
(I-I co
n)/(I
m-I c
on)
0.0
0.2
0.4
0.6
0.8
1.0CuNi
10 Cu 10 Ni
100pA
10 s
200pA
5 s
10 Cu
10 Cu
30 s
CuNi
Fig. 7 – Effect of Cu and Ni on the NR current in HEK cells expressing GluN1/GluN2B receptors. Responses were elicited by50 μM glutamic acid in the presence of 30 μM glycine at �60 mV and are represented by downward deflections of the currenttrace. (A) Effect of 30 s treatment with 10 lM Cu or 10 lM Ni. The effects are very similar, but recovery is faster after Ni washout than after Cu. (B) Current traces showing current facilitation following treatments with 10 lM Cu for 20 and 60 s. Note thetwo different time calibration bars. (C) Effect of Cu (empty circles) and Ni (filled circles) in GluN1/GluN2B NR current as afunction of duration of treatment. Im is the current modified by either metal (Cu, empty circles or Ni, filled circles) and Icon isthe current before treatment. Points are averages of 3 to 7 experiments and bars represent sem. The dotted line is the best fitcurve of Fig. 2B (CGC) and it is shown for visual comparison. The facilitating effect of Ni did not change with the duration oftreatment (horizontal line). (D) Time course of recovery from Cu and Ni potentiation. Im is the current modified by either metal(Cu, empty circles or Ni, filled circles) and Icon is the current before treatment and after complete recovery. Points are averagesof 4 experiments, bars represent sem. Points were best fitted with Eq. (3b) for Ni and Eq. (3c) for Cu. The recovery from Ni effect(filled circles) had a single exponential time course with τ¼15 s. The recovery from the Cu effect (empty circles) displayed adouble exponential time course with a fast (τ1¼11 s) and a slower component (τ2¼165 s) of very similar amplitude (a1¼0.48and a2¼0.52).
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 27

Author's personal copy
The effect of Cu on CGC NR currents was modeled by theproduct of an inhibitory component and a stimulatory com-ponent, as we previously described for Ni (Marchetti andGavazzo, 2003). The value of IC50 for the inhibition compo-nent (24 mM) was close to the value reported previously forthe inhibition of NR channel in olfactory bulb neurons(Trombley and Shepherd, 1996). This model appeared suitableto describe properly the effect of Cu also in GluN1/GluN2Aexpressing cells (Fig. 5A). However, in the transfected chan-nels, the maximum potentiation was half (a¼0.88) than innative neuron receptors (a¼1.86) and ED50 was lower (2 mM intransfected cells and 4.6 mM in native neurons), probablybecause of differences in the intracellular environments.
The effect of Cu on NR channels differs from that of otherdivalent metals in several respects. Different from Mg, butalso from Zn and Ni, Cu never exerted a voltage-dependentblocking effect, an observation that was reported previously(Vlachova et al., 1996) and places it among the metals notinteracting with a site inside the pore. Although the poten-tiating effect of Cu is reminiscent of the effect of Ni, whichhas also multiple effects on NR channels (Marchetti andGavazzo, 2003), including a facilitation induced by pretreat-ment (Gavazzo et al., 2006), the potentiating effect of Ni waslimited to receptors containing the GluN2B subunit, whileCu potentiated the activity of both GluN2B-containing andGluN2A-containing receptors. As a consequence, in CGCs Niexerted a prominent facilitating action only in the first daysin vitro, when these neurons express GluN2B-containing recep-tors (Gavazzo et al., 2006), while Cu facilitation of CGC NRcurrents was independent of the days in vitro.
The potentiating effect of Cu in GluN2B-containing recep-tors is qualitatively comparable to that described for Ni inthese receptors, and possibly occurs through a binding sitelocated in the ATD because, similar to Ni (Gavazzo et al.,2009), Cu does not facilitate the NR current in the truncatedform ΔATDGluN2B receptor. However, the effects of the twometals differ in kinetics, because Ni was quite fast in onsetand recovery, while Cu effect was slower to develop and torecover (Fig. 7C and D).
The voltage-independent inhibitory effect of Cu hasbeen described previously (Trombley and Shepherd, 1996;Vlachova et al., 1996; Weiser and Wienrich, 1996), althoughthe mechanism of action is still debatable. As the inhibitionwas present also in the truncated GluN2B subunit, ΔATD-GluN2B, the site of interaction that mediates this inhibitionmust be located outside the ATD. The existence of anadditional divalent cation binding site responsible for specificheavy metal interaction located in an area different from ATDhas been postulated previously for Zn (Fayyazuddin et al.,2000) and for Ni (Gavazzo et al., 2009).
Another explanation for the mechanism of Cu-drivenvoltage-independent inhibition has been proposed. BecauseCu can function as electron acceptor promoting the reactionof nitric oxide with thiols, release of this metal could functionas a molecular switch to control extracellular S-nitrosation ofthe NR (Schlief et al., 2005; Schlief et al., 2006), a post-translational mechanism critical for modulating receptorfunction (Lipton et al., 2002). This regulation would dependon a couple of cysteines on the GluN1 subunit and thereforebe independent on GluN2 ATD. However, a role of this
regulation has been interpreted by others as an artifact(Hopper et al., 2004).
Interpretation of the facilitation effect is even more com-plex. An involvement of the redox sites is suggested by theobservation that Cu-mediated facilitation is abolished imme-diately after treatment with reducing agents (Fig. 3 and 4).Redox reagents modulate NRs, acting at extracellular sitesto increase or decrease macroscopic currents and Ca flux.Sulfhydryl reducing agents, such as dithiothreitol (DTT),enhance, whereas oxidizing agents decrease, NR currents(Aizenman et al., 1989; Brimecombe et al., 1999). Experimentswere undertaken to examine the involvement of Cu in thisinteraction and the effect of Cu in the different redox state ofthe protein and DTT was chosen as a reducing agent. DTT hasthe disadvantage to chelate divalent cations (Krezel et al.,2001), but the chelating and the reducing effects could beresolved by their time course, because the former was readilywashed out with the fast perfusion system used in this study(10 mL/min, Marchetti and Gavazzo, 2003), while the latterpersisted longer. DTT was never used in the presence of themetal and the effect of Cu was tested immediately after DTT,once assessed that the facilitating action of DTT was notreversed by short (20 to 60 s) wash. In this conditions, thetreatment with Cu did not modify the NR current in anyrespect (Fig. 3), indicating that in the presence of DTT inducedfacilitation, Cu was ineffective. Similar results were obtainedin Ca influx measurements, where long incubation with DTTrequired several minutes to be washed and the effect of Cuimmediately after was mainly inhibitory (Fig. 4). However,these observations are not sufficient to postulate a directeffect of Cu on cystein disulfide bridges because (i) the mostlikely oxidation form of Cu in our conditions is Cu2þ that isexpected to function as electron acceptor, facilitating oxidationof intramolecular disulfide linkages and not the opposite,although a possibility for Cu to break disulfides was postulatedin a different system (Kirakosyan et al., 2008); (ii) determinationof accessible SH-group by Elmann's reagent (DTNB, Riddles et al.,1979), did not put in evidence disulfide bridge breakage caused bytreatment with Cu (Baranowska-Bosiacka, personal observation);(iii) pre-treatment with DTNB reagent caused a decrease of thecurrent, but did not enhance Cu effect (not shown). Therefore,the influence of Cu on the redox site appears an indirectone, with the reduced form of the channel less sensitive to Cupotentiation.
In GluN2A, the facilitating effect of Cu may result from
competition at the high affinity Zn binding site, with Cu
removing Zn from its site and eliminating the tonic inhibi-
tion. In this respect, we have preliminary evidence that the
potentiation induced by Cu is absent in the presence of Zn,
both in CGC and in GluN2A-containing receptors expressed
in HEK cells (Marchetti C., unpublished observation). In
the model proposed by Gielen et al. (2009), the GluN2-ATD
alternates between open and closed cleft conformations, the
latter favoring pore closure. While Zn binding causes the cleft
to close, Cu may bind in the ATD, displacing Zn from its
binding site, favoring the open cleft conformation and open-
ing of the pore. Competition at the Zn high affinity site was
observed also with lead (Pb) in GluN2A-containing receptors,
but not in GluN2B-containing receptors (Gavazzo et al., 2008).
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 128

Author's personal copy
However, in the case of Pb, the metal inhibited the current,favoring the closed cleft conformation, as Zn does.
Physiological implications of the potentiating effect of Cu aredifficult to foresee and so far we have no evidence of any directeffect on neuron survival or implication in NMDA excitoxicity.Experimental data have suggested that ATP7A-mediatedrelease of synaptic copper protects neurons from excitotoxicityby downregulating their activity (Schlief et al., 2006) and Cuchelation exacerbates NMDA-mediated excitotoxic cell death inprimary hippocampal neurons, whereas the addition of a highconcentration of Cu is protective and results in a significantdecrease in cytoplasmic Ca levels after NR activation (Schliefand Gitlin, 2006). However, these effects were caused by a largeCu concentration (200 mM) that always mediates a predominantinhibition. Our data show that Cu acts as a enhancer of NRactivity in a narrow range of concentration (o30 mM), a rangeclose to the extracellular physiological concentration value.In addition the effect may depend on the oxidation state ofcysteine thiols (Lipton et al., 2002), because our data indicatethat the reduced form of the channel is not subjected toCu potentiation. By this effect, Cu may trigger excitability orexacerbate neuronal injury depending on the cellular state.At synaptic level, Cu has been reported to induce biphasicchanges in neurotransmission (Peters et al., 2011) and to inhibitboth NR-dependent (Doreulee et al., 1997) and NR-independent(Salazar-Weber and Smith, 2011) LTP.
In conclusion, beside the antagonistic effect, we haveshown that Cu is able to mediate a stimulatory effect onnative and recombinant NR channels. It is possible thatdifferent sites in the different NRs are involved in thismodulation by Cu. This mechanism can play a role in Cumediated synaptic function (Gaier et al., 2013), as well as inCu-mediated neurotoxicity (Valko et al., 2005; Pal et al., 2013).Regardless of a specific action of Cu on neuronal function,this effect may be of interest to unravel allosteric modulationof NRs and to model specific binding sites of therapeuticpotential.
4. Experimental procedure
4.1. Neuronal cultures
Cerebellar granule cells (CGC) were prepared from 8-day-oldSprague Dawley rats as previously described ( Marchetti et al.,1995; Gavazzo et al., 2006). Cells were seeded on poly-L-lysinecoated glass coverslips and treated with 10 mM cytosinearabinoside from day 1 to minimize proliferation of non-neuronal cells. Cultures were maintained in Basal Eagle'sculture medium, containing 10% fetal calf serum, 100 mg/mlgentamycin and 25 mM KCl, in a humidified atmosphere with5% CO2, 95% air at 37 1C. Experiments were performed incultures between 5 and 16 days in vitro.
4.2. Functional expression of NR channels in HEK293 cells
DNA clones for mammalian cell expression, encoding the ratGluN1 (GenBank number X63255), GluN2A (GenBank AF142377),and GluN2B (Gen-Bank U11419) were a generous gift of StefanoVicini (Georgetown University, Washington, USA). The splice
variant of GluN1 subunit used in this study contains both Csegments, but is devoid of the N segment or exon 5. The ATD-deleted GluN2B subunit was kindly provided by Pierre Paoletti(CNRS, Paris) and is referred to as ΔATD-GluN2B (Gavazzo et al.,2009; Rachline et al., 2005). Human CD8 clone were kindly givenby J. Barhanin (CNRS UPR 411, Valbonne, France). Expressionvectors were pRC for GluN1, pcDNA1.1 for GluN2A, pRK-5 forGluN2B and ΔATD-GluN2B.
HEK 293 cells (CRL 1573; ATC, Rockville, MA, USA) werecultured as in previous work (Marchetti and Gavazzo, 2003)and transiently transfected by incubation with Effectenetransfection reagent (Qiagen, Valencia, CA, USA) and a 2 mgDNA mixture containing 0.8 mg GluN1, 1 mg GluN2A or GluN2Band 0.2 mg CD8. After 6–7 h of incubation, cells were washed,detached and seeded on poly-L-lysine coated glass cover slipsat a concentration of 2.3�105 cells/ml. They were maintainedin DMEM/F12 medium supplemented with 10% fetal bovineserum and 100 mM (7)-2-amino-5-phosphopentanoic acid(D-AP5), a NR antagonist. NR channel expression was obtained18–36 h after wash of the DNA-reagent mixture. Successfullytransfected cells were identified by a brief incubation with CD8antibody coated polystyrene beads (Dynabeads M-450, DynalBiotech, Oslo, Norway).
4.3. Electrophysiology
Total membrane currents were measured in whole-cellclamp configuration as previously described (Marchetti andGavazzo, 2003; Gavazzo et al., 2006). Electrodes were pulledfrom Clark borosilicate glass capillaries and had a resistanceof 5–8 MΩ when filled with the intracellular solution (seebelow). Currents and membrane potential were recorded byan Axopatch amplifier (Molecular Devices, Sunnyvale, CA,USA). Voltage stimulation and data acquisition were per-formed by a PC through a Digidata 1440A interface andPclamp-10 software (Molecular Devices). Capacitance transi-ents were minimized by analog compensation and the valueobtained by this compensation was taken as an estimate ofthe cell capacitance, when needed. Traces were sampled at5 kHz and low-pass filtered at 2 kHz.
Cells were continuously superfused by gravity flow(10ml/min) and all modifiers were applied by bath solutionexchange, with exchange time of approximately 400ms(Marchetti and Gavazzo, 2003). The bath solution contained(in mM) NaCl 140, KCl 5.4, CaCl2 1.0, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) 10, glucose 10. The pHwas adjusted at 7.4 with NaOH and 30 mM glycine was added inall solutions. The pipette solution contained (inmM): CsCl 50, CsF80, EGTA 11, CaCl2 1, MgCl2 1, HEPES 10. The pH was adjusted at7.3 with Trizma base. Cells were voltage-clamped at a potentialof �60mV or stimulated by voltage ramps (from �60 toþ60mV; 1mV/ms). Cells were challenged several times withthe agonist before testing the modifiers. Cu was prepared freshby dilution from a 100mM stock solution of CuCl2.
4.4. Ca influx by fluorescence measurements
The influx of Ca through NR channels was measured by theintracellular fluorescence Ca chelator Fura2 (Marchetti et al.,1995; Mazzolini et al., 2001). Cells were incubated with 10 μm
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 29

Author's personal copy
Fura-2-acetoxymethyl ester (Fura-2AM, Invitrogen) in a phy-siological saline containing 1 mM Mg for 30–45 min at 37 1Cand then washed with the same solution. The coverslips werethen mounted on the stage of an inverted microscope (ZeissIM35, Germany) and cells were viewed through a X40 quartzobjective. [Ca]i measurements and calibrations were performedas described previously (Marchetti et al., 1995; Mazzolini et al.,2001). Cells were challenged by agonists in the absence of Mg(same external solution as in whole-cell recording experiments).
4.5. Data analysis
Traces were analyzed by Pclamp 10 software (Clampfit,Molecular Devices) and by Sigma Plot (SPSS Science, Chicago,IL, USA) software. Data are shown as mean7SEM. Statisticalsignificance was evaluated by paired t-test (In Stat, GraphPadSoftware, San Diego, CA, USA). The difference between twoconditions was considered significant if po0.05.
Acknowledgments
We are grateful to dr Elena Gatta for help in preparingcerebellar granule neuron cultures, to Francesca Quartino,Paolo Guastavino, Alessandro Barbin and Damiano Magliozzifor technical assistance and help.
r e f e r e n c e s
Aizenman, E., Lipton, S.A., Loring, R.H., 1989. Selectivemodulation of NMDA responses by reduction and oxidation.Neuron 2, 1257–1263.
Aizenman, E., Hartnett, K.A., Reynolds, I.J., 1990. Oxygen freeradicals regulate NMDA receptor function via a redoxmodulatory site. Neuron 5, 841–846.
Brimecombe, J.C., Potthoff, W.K., Aizenman, E., 1999. A criticalrole of the N-methyl-D-aspartate (NMDA) receptor subunit(NR) 2A in the expression of redox sensitivity of NR1/NR2Arecombinant NMDA receptors. J. Pharmacol. Exp. Ther. 291,785–792.
Choi, D.W., 1992. Excitotoxic cell death. J. Neurobiol. 23,1261–1276.
Choi, Y.B., Lipton, S.A., 2000. Redox modulation of the NMDAreceptor. Cell. Mol. Life Sci. 57, 1535–1541.
Doreulee, N., Yanovsky, Y., Haas, H.L., 1997. Suppression of long-term potentiation in hippocampal slices by copper.Hippocampus 7, 666–669.
Fayyazuddin, A., Villarroel, A., Le Goff, A., Lerma, J., Neyton, J.,2000. Four residues of the extracellular N-terminal domain ofthe NR2A subunit control high-affinity Zn2þ binding to NMDAreceptors. Neuron 25, 683–694.
Gaier, E.D., Eipper, B.A., Mains, R.E., 2013. Copper signaling in themammalian nervous system: synaptic effects. J. Neurosci. Res.91, 2–19.
Gavazzo, P., Mazzolini, M., Tedesco, M., Marchetti, C., 2006. Nickeldifferentially affects NMDA receptor channels in developingcultured rat neurons. Brain Res. 1078, 71–79.
Gavazzo, P., Zanardi, I., Baranowska-Bosiacka, I., Marchetti, C.,2008. Molecular determinants of Pb2þ interaction with NMDAreceptor channels. Neurochem. Int. 52, 329–337.
Gavazzo, P., Guida, P., Zanardi, I., Marchetti, C., 2009. Moleculardeterminants of multiple effects of nickel on NMDA receptorchannels. Neurotox Res. 15, 38–48.
Gielen, M., Siegler Retchless, B., Mony, L., Johnson, J.W., Paoletti, P.,2009. Mechanism of differential control of NMDA receptoractivity by NR2 subunits. Nature 459, 703–707.
Hartter, D.E., Barnea, A., 1988. Evidence for release of copper inthe brain: depolarization-induced release of newly taken-up67copper. Synapse 2, 412–415.
Hopper, R., Lancaster, B., Garthwaite, J., 2004. On the regulation ofNMDA receptors by nitric oxide. Eur. J. Neurosci. 19, 1675–1682.
Hopt, A., Korte, S., Fink, H., Panne, U., Niessner, R., Jahn, R.,Kretzschmar, H., Herms, J., 2003. Methods for studyingsynaptosomal copper release. J. Neurosci. Methods 128,159–172.
Horning, M.S., Trombley, P.Q., 2001. Zinc and copper influenceexcitability of rat olfactory bulb neurons by multiplemechanisms. J. Neurophysiol. 86, 1652–1660.
Hung, Y.H., Bush, A.I., Cherny, R.A., 2010. Copper in the brain andAlzheimer's disease. J. Biol. Inorg. Chem. 15, 61–76.
Kardos, J., Kovacs, I., Hajos, F., Kalman, M., Simonyi, M., 1989.Nerve endings from rat brain tissue release copper upondepolarization. A possible role in regulating neuronalexcitability. Neurosci. Lett. 103, 139–144.
Kirakosyan, G., Trchounian, K., Vardanyan, Z., Trchounian, A.,2008. Copper (II) ions affect Escherichia coli membranevesicles' SH-groups and a disulfide-dithiol interchangebetween membrane proteins. Cell. Biochem. Biophys. 51,45–50.
Krezel, A., Lesniak, W., Jezowska-Bojczuk, M., Mlynarz, P., Brasun, J.,Kozlowski, H., Bal, W., 2001. Coordination of heavy metals bydithiothreitol, a commonly used thiol group protectant. J. Inorg.Biochem. 84, 77–88.
Leiva, J., Palestini, M., Infante, C., Goldschmidt, A., Motles, E.,2009. Copper suppresses hippocampus LTP in the rat, but doesnot alter learning or memory in the morris water maze. BrainRes. 1256, 69–75.
Linder, M.C., Hazegh-Azam, M., 1996. Copper biochemistry andmolecular biology. Am. J. Clin. Nutr. 63, 797S–811S.
Lipton, S.A., Choi, Y.B., Takahashi, H., Zhang, D., Li, W., Godzik, A.,Bankston, L.A., 2002. Cysteine regulation of protein function—asexemplified by NMDA-receptor modulation. Trends Neurosci. 25,474–480.
Lutsenko, S., Petris, N.J., 2003. Function and regulation of themammalian copper-transporting ATPases: Insights frombiochemical and cell biological approaches. J. Membr. Biol.191, 1–12.
Marchetti, C., Amico, C., Usai, C., 1995. Functionalcharacterization of the effect of nimodipine on the calciumcurrent in rat cerebellar granule cells. J. Neurophysiol. 73,1169–1180.
Marchetti, C., Gavazzo, P., 2003. Subunit-dependent effects ofnickel on NMDA receptor channels. Brain Res. Mol. Brain Res.117, 139–144.
Mathie, A., Sutton, G.L., Clarke, C.E., Veale, E.L., 2006. Zinc andcopper: pharmacological probes and endogenous modulatorsof neuronal excitability. Pharmacol. Ther. 111, 567–583.
Mazzolini, M., Traverso, S., Marchetti, C., 2001. Multiple pathwaysof Pb(2þ) permeation in rat cerebellar granule neurones. J.Neurochem. 79, 407–416.
Millhauser, G.L., 2007. Copper and the prion protein: methods,structures, function, and disease. Annu. Rev. Phys. Chem. 58,299–320.
Pal, A., Badyal, R.K., Vasishta, R.K., Attri, S.V., Thapa, B.R., Prasad, R.,2013. Biochemical, histological, and memory impairment effectsof chronic copper toxicity: a model for non-wilsonian braincopper toxicosis in wistar rat. Biol. Trace Elem. Res. 153, 257–268.
Paoletti, P., Neyton, J., Ascher, P., 1995. Glycine-independent andsubunit-specific potentiation of NMDA responses byextracellular Mg2þ. Neuron 15, 1109–1120.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 130

Author's personal copy
Paoletti, P., Ascher, P., Neyton, J., 1997. High-affinity zincinhibition of NMDA NR1–NR2A receptors. J. Neurosci. 17,5711–5725.
Paoletti, P., Bellone, C., Zhou, Q., 2013. NMDA receptor subunitdiversity: impact on receptor properties, synaptic plasticityand disease. Nat. Rev. Neurosci. 14, 383–400.
Peters, C., Munoz, B., Sepulveda, F.J., Urrutia, J., Quiroz, M., Luza, S.,De Ferrari, G.V., Aguayo, L.G., Opazo, C., 2011. Biphasic effectsof copper on neurotransmission in rat hippocampal neurons.J. Neurochem. 119, 78–88.
Rachline, J., Perin-Dureau, F., Le Goff, A., Neyton, J., Paoletti, P.,2005. The micromolar zinc-binding domain on the NMDAreceptor subunit NR2B. J. Neurosci. 25, 308–317.
Riddles, P.W., Blakeley, R.L., Zerner, B., 1979. Ellman's reagent:5,5′-dithiobis(2-nitrobenzoic acid) – a reexamination. Anal.Biochem. 94, 75–81.
Salazar-Weber, N.L., Smith, J.P., 2011. Copper Inhibits NMDAReceptor-Independent LTP and modulates the paired-pulseratio after LTP in mouse hippocampal slices. Int. J. AlzheimersDis. 2011, 864753.
Schlief, M.L., Craig, A.M., Gitlin, J.D., 2005. NMDA receptoractivation mediates copper homeostasis in hippocampalneurons. J. Neurosci. 25, 239–246.
Schlief, M.L., Gitlin, J.D., 2006. Copper homeostasis in the CNS: anovel link between the NMDA receptor and copperhomeostasis in the hippocampus. Mol. Neurobiol. 33, 81–90.
Schlief, M.L., West, T., Craig, A.M., Holtzman, D.M., Gitlin, J.D.,2006. Role of the Menkes copper-transporting ATPase inNMDA receptor-mediated neuronal toxicity. Proc. Natl. Acad.Sci. USA 103, 14919–14924.
Stys, P.K., You, H.T., Zamponi, G.W., 2012. Copper-dependentregulation of NMDA receptors by cellular prion protein:implications for neurodegenerative disorders. J. Physiol. 590,1357–1368.
Traynelis, S.F., Wollmuth, L.P., McBain, C.J., Menniti, F.S.,Vance, K.M., Ogden, K.K., Hansen, K.B., Yuan, H., Myers, S.J.,Dingledine, R., 2010. Glutamate receptor ion channels:structure, regulation, and function. Pharmacol. Rev. 62,405–496.
Trombley, P.Q., Shepherd, G.M., 1996. Differential modulation byzinc and copper of amino acid receptors from rat olfactorybulb neurons. J. Neurophysiol. 76, 2536–2546.
Valko, M., Morris, H., Cronin, M.T.D., 2005. Metals, toxicity andoxidative stress. Curr. Med. Chem. 12, 1161–1208.
Vlachova, V., Zemkova, H., Vyklicky Jr., L., 1996. Coppermodulation of NMDA responses in mouse and rat culturedhippocampal neurons. Eur. J. Neurosci. 8, 2257–2264.
Weiser, T., Wienrich, M., 1996. The effects of copper ions onglutamate receptors in cultured rat cortical neurons. BrainRes. 742, 211–218.
White, A.R., Cappai, R., 2003. Neurotoxicity from glutathionedepletion is dependent on extracellular trace copper.J. Neurosci. Res. 71, 889–897.
Xie, X., Hider, R.C., Smart, T.G., 1994. Modulation of GABA-mediated synaptic transmission by endogenous zinc in theimmature rat hippocampus in vitro. J. Physiol. 478 (1), 75–86.
You, H., Tsutsui, S., Hameed, S., Kannanayakal, T.J., Chen, L.,Xia, P., Engbers, J.D., Lipton, S.A., Stys, P.K., Zamponi, G.W.,2012. Abeta neurotoxicity depends on interactions betweencopper ions, prion protein, and N-methyl-D-aspartatereceptors. Proc. Natl. Acad. Sci. USA 109, 1737–1742.
b r a i n r e s e a r c h 1 5 4 2 ( 2 0 1 4 ) 2 0 – 3 1 31
Related Documents











