Multiple determinants are involved in HIV coreceptor use as demonstrated by CCR4/CCL22 interaction in peripheral blood mononuclear cells (PBMCs) Lokesh Agrawal, Zainab Vanhorn-Ali, and Ghalib Alkhatib Department of Microbiology and Immunology and Walther Oncology Center, Indiana University School of Medicine; and The Walther Cancer Institute, Indianapolis, IN Abstract: Although a number of chemokine re- ceptors display coreceptor activities in vitro, chemokine receptor 5 (CCR5) and CXC chemo- kine receptor 4 (CXCR4) remain the major co- receptors used by the human immunodeficiency virus type 1 (HIV-1). In this study, we used an envelope-mediated fusion assay to demonstrate low CCR4 coreceptor activity with some primary HIV-1 and simian immunodeficiency virus-1 (mac316) isolates in vitro. The coreceptor activ- ity was sensitive to CCR4-specific antibodies and to the CCR4-specific chemokine ligand macro- phage-derived chemokine (MDC)/chemokine li- gand 22 (CCL22). Treatment of peripheral blood mononuclear cells (PBMCs; which express high levels of CCR4) with CCL22 caused down-mod- ulation of endogenous CCR4 but had no signifi- cant effect on HIV-1 entry, suggesting that CCR4 may not be used as an entry coreceptor. Despite expression of other minor coreceptors on PBMCs, CCR5 and CXCR4 are preferentially used by HIV-1 isolates, as shown by chemokine- inhibition data. To determine the factors in- volved in this selective use, we analyzed CCR4 coreceptor activity and compared it with CCR5 use in PBMCs. We used a quantitative fluores- cence-activated cell-sorting assay to estimate the numbers of CCR4 and CCR5 antibody-binding sites (ABS) on PBMCs. Although CCR4 was found on a higher percentage of CD4() cells, CCR5 ABS was twofold greater than CCR4 ABS on CD4( ) cells. Confocal microscopy revealed strong cell-surface CD4/CCR5 but weak CD4/ CCR4 colocalization in PBMCs. Binding studies demonstrated that soluble gp120 had greater af- finity to CCR5 than CCR4. The results suggested that coreceptor density, colocalization with CD4, and affinity of the viral gp120 to the core- ceptor may determine preferential coreceptor use by HIV-1. J. Leukoc. Biol. 72: 1063–1074; 2002. Key Words: (MDC) CCRA coreceptors HIV-1 PBMCs mac- rophage-derived chemokine INTRODUCTION Chemotactic cytokines (chemokines) are proinflammatory me- diators that recruit various cell types to the inflammatory sites (reviewed in refs. [1, 2]). The CC chemokines macrophage- inflamatory protein-1 (MIP-1)/chemokine ligand 3 (CCL3), MIP-1/CCL4, and regulated on activation, normal T ex- pressed and secreted (RANTES)/CCL5, as well as the CXC chemokine stromal cell-derived factor-1 (SDF-1)/CXC chemo- kine ligand 12 (CXCL12), have all been shown to suppress human immunodeficiency virus type 1 (HIV-1) replication of R5 and X4 isolates [3–5]. Macrophage-derived chemokine (MDC)/CCL22, a member of the CC chemokine family, is produced and secreted by monocytes, dendritic cells [6 – 8], and natural killer cells [9]. The relevance of CCL22 to HIV-1 entry was suggested by Pal et al. [10], who demonstrated a broad antiviral activity by high-pressure liquid chromatogra- phy (HPLC)-purified fractions from supernatants of CD8()T lymphocytes. The anti-HIV-1 activity of CCL22 has been con- troversial [11, 12]; however, recent studies have confirmed the antiviral effects of CCL22 against X4 and R5 HIV-1 infection of phytohemagglutinin (PHA)-activated PBMCs [7, 13] or R5 infection of primary macrophages [14]. The chemokine recep- tor 4 (CCR4) is the only known functional receptor for CCL22 [15], and CCR4 mRNA is expressed predominantly in the thymus, spleen, and peripheral blood leukocytes, including T cells, basophils, monocytes [16], macrophages, and platelets [17, 18]. The T cell-directed CC chemokine thymus and acti- vation-regulated chemokine (TARC/CCL17) is also a specific biological ligand for CCR4 [19]. The CXC chemokine receptor 4 (CXCR4) and the CCR5 are the major coreceptors used by X4 and R5 HIV-1 isolates to gain entry into susceptible cells (reviewed in ref. [20]). Addi- tional coreceptors including CCR4 have been demonstrated to support HIV-1 entry in vitro; however, their contribution to HIV disease in vivo remains unknown. Previous studies de- scribed CCR4 as an entry coreceptor [21–24], and others have not observed coreceptor activity for CCR4 [25–27]. These Correspondence: Ghalib Alkhatib, Department of Microbiology and Immu- nology and Walther Oncology Center, Indiana University School of Medicine, 635 Barnhill Drive, Room 420, Indianapolis, IN 46202. E-mail: [email protected] Received April 25, 2002; revised May 29, 2002; accepted May 30, 2002. Journal of Leukocyte Biology Volume 72, November 2002 1063

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Multiple determinants are involved in HIV coreceptor use asdemonstrated by CCR4/CCL22 interaction in peripheralblood mononuclear cells (PBMCs)
Lokesh Agrawal, Zainab Vanhorn-Ali, and Ghalib Alkhatib
Department of Microbiology and Immunology and Walther Oncology Center, Indiana University School of Medicine;and The Walther Cancer Institute, Indianapolis, IN
Abstract: Although a number of chemokine re-ceptors display coreceptor activities in vitro,chemokine receptor 5 (CCR5) and CXC chemo-kine receptor 4 (CXCR4) remain the major co-receptors used by the human immunodeficiencyvirus type 1 (HIV-1). In this study, we used anenvelope-mediated fusion assay to demonstratelow CCR4 coreceptor activity with some primaryHIV-1 and simian immunodeficiency virus-1(mac316) isolates in vitro. The coreceptor activ-ity was sensitive to CCR4-specific antibodies andto the CCR4-specific chemokine ligand macro-phage-derived chemokine (MDC)/chemokine li-gand 22 (CCL22). Treatment of peripheral bloodmononuclear cells (PBMCs; which express highlevels of CCR4) with CCL22 caused down-mod-ulation of endogenous CCR4 but had no signifi-cant effect on HIV-1 entry, suggesting that CCR4may not be used as an entry coreceptor. Despiteexpression of other minor coreceptors onPBMCs, CCR5 and CXCR4 are preferentiallyused by HIV-1 isolates, as shown by chemokine-inhibition data. To determine the factors in-volved in this selective use, we analyzed CCR4coreceptor activity and compared it with CCR5use in PBMCs. We used a quantitative fluores-cence-activated cell-sorting assay to estimate thenumbers of CCR4 and CCR5 antibody-bindingsites (ABS) on PBMCs. Although CCR4 was foundon a higher percentage of CD4(�) cells, CCR5ABS was twofold greater than CCR4 ABS onCD4(�) cells. Confocal microscopy revealedstrong cell-surface CD4/CCR5 but weak CD4/CCR4 colocalization in PBMCs. Binding studiesdemonstrated that soluble gp120 had greater af-finity to CCR5 than CCR4. The results suggestedthat coreceptor density, colocalization withCD4, and affinity of the viral gp120 to the core-ceptor may determine preferential coreceptoruse by HIV-1. J. Leukoc. Biol. 72: 1063–1074;2002.
Key Words: (MDC) � CCRA � coreceptors � HIV-1 � PBMCs � mac-rophage-derived chemokine
INTRODUCTION
Chemotactic cytokines (chemokines) are proinflammatory me-diators that recruit various cell types to the inflammatory sites(reviewed in refs. [1, 2]). The CC chemokines macrophage-inflamatory protein-1� (MIP-1�)/chemokine ligand 3 (CCL3),MIP-1�/CCL4, and regulated on activation, normal T ex-pressed and secreted (RANTES)/CCL5, as well as the CXCchemokine stromal cell-derived factor-1 (SDF-1)/CXC chemo-kine ligand 12 (CXCL12), have all been shown to suppresshuman immunodeficiency virus type 1 (HIV-1) replication ofR5 and X4 isolates [3–5]. Macrophage-derived chemokine(MDC)/CCL22, a member of the CC chemokine family, isproduced and secreted by monocytes, dendritic cells [6–8],and natural killer cells [9]. The relevance of CCL22 to HIV-1entry was suggested by Pal et al. [10], who demonstrated abroad antiviral activity by high-pressure liquid chromatogra-phy (HPLC)-purified fractions from supernatants of CD8(�) Tlymphocytes. The anti-HIV-1 activity of CCL22 has been con-troversial [11, 12]; however, recent studies have confirmed theantiviral effects of CCL22 against X4 and R5 HIV-1 infectionof phytohemagglutinin (PHA)-activated PBMCs [7, 13] or R5infection of primary macrophages [14]. The chemokine recep-tor 4 (CCR4) is the only known functional receptor for CCL22[15], and CCR4 mRNA is expressed predominantly in thethymus, spleen, and peripheral blood leukocytes, including Tcells, basophils, monocytes [16], macrophages, and platelets[17, 18]. The T cell-directed CC chemokine thymus and acti-vation-regulated chemokine (TARC/CCL17) is also a specificbiological ligand for CCR4 [19].
The CXC chemokine receptor 4 (CXCR4) and the CCR5 arethe major coreceptors used by X4 and R5 HIV-1 isolates togain entry into susceptible cells (reviewed in ref. [20]). Addi-tional coreceptors including CCR4 have been demonstrated tosupport HIV-1 entry in vitro; however, their contribution toHIV disease in vivo remains unknown. Previous studies de-scribed CCR4 as an entry coreceptor [21–24], and others havenot observed coreceptor activity for CCR4 [25–27]. These
Correspondence: Ghalib Alkhatib, Department of Microbiology and Immu-nology and Walther Oncology Center, Indiana University School of Medicine,635 Barnhill Drive, Room 420, Indianapolis, IN 46202. E-mail:[email protected]
Received April 25, 2002; revised May 29, 2002; accepted May 30, 2002.
Journal of Leukocyte Biology Volume 72, November 2002 1063

negative results probably resulted from poor transfection effi-ciency of CCR4 and inability to verify its surface expression.The recent development of monoclonal antibodies (mAb)against CCR4 made it possible to detect its abundant expres-sion on PBMCs [28]. HIV-1 uses the major coreceptors forentry into PBMCs; however, the factors contributing to thisselective coreceptor use are not entirely understood. The pref-erential use of the major coreceptors was revealed by chemo-kine-inhibition experiments previously reported by severalgroups [29]. In this study, we used MDC/CCL22 as well asCCR4-specific antibodies to examine whether CCR4 is used asan entry coreceptor in PBMCs in vitro.
Our previous studies demonstrated that CCL4, a chemokineligand specific to CCR5, did not result in complete inhibitionof envelope (Env) fusion [30], suggesting expression of otherR5 coreceptors on primary cells. To examine the role of alter-native coreceptors, functional cDNA screening was performedto isolate these putative coreceptors. We used a nonbiasedcDNA screening strategy previously used to isolate CXCR4[31] to isolate a cDNA clone encoding CCR4. The isolatedcDNA showed coreceptor activity when challenged with HIV-1Envs derived from primary isolates and the macrophage-tropicsimian immunodeficiency virus (SIV)-mac316 Env. CCR4-me-diated fusion activity was more sensitive to the truncated formof CCL22(3–69) than native CCL22(1–69), suggesting a morepotent binding of the truncated form to CCR4. Despite the highlevels of CCR4 expression on PBMCs, CCL22- and CCR4-specific antibodies had no significant effect on viral entry.However, a significant, donor-dependent CCL22 antiviral ac-tivity was consistently obtained in HIV-1 infectivity assays.The results argued against the use of CCR4 as a coreceptor forentry into PBMCs. Therefore, we wished to determine thefactors that contribute to the preferential use of CCR5 as acoreceptor by HIV-1 isolates taking advantage of the highlevels of cell-surface CCR4 on PBMCs [28].
We used a quantitative fluorescence-activated cell-sorting(QFACS) assay to measure antibody-binding sties (ABS) ofCCR4, CCR5, and CXCR4 on PBMCs isolated from six differ-ent donors. Our data showed that high levels of CCR4 detectedon PBMCs from all donors tested did not correlate with theobserved CCR4 coreceptor activity and its inhibition by CCL22in transfected murine cells. Confocal microscopy studies dem-onstrated efficient colocalization of CCR5 and CD4 in allPBMC samples tested compared with weak CCR4/CD4 colo-calization. Binding studies using radiolabeled gp120 revealedthat gp120 had greater affinity to CCR5 than CCR4. The datasuggested that coreceptor density, colocalization with CD4, andgp120 affinity for CCR5 might contribute to the selectiveHIV-1 coreceptor use.
MATERIALS AND METHODS
Cells and viruses
HeLa cells and NIH 3T3 cells were purchased from the American TypeCulture Collection (Manassas, VA). The 3T3.T4 cell line was obtained from theAIDS Research and Reference Reagent Program, Division of AIDS, NIAID,NIH (Rockville, MD). HeLa and 3T3.T4 cell lines were cultured in Dulbecco’smodified Eagle’s medium (Quality Biologicals, Gaithersburg, MD) containing
10% fetal bovine serum (FBS; HyClone, Logan, UT), 2 mM L-glutamine, andantibiotics. Recombinant vaccinia virus stocks were prepared by standardprocedures [32]. The recombinant viruses used vCB-21R(pT7-LacZ), vTF7-3(T7 polymerase), vCB-3(CD4), vCB-16(Unc), vCB-43(BaL), vCB-41(LAV), vCB-28(JR-FL), vCB-39(ADA), vCB-74(SIVmac239), and vCB-75(SIVmac316) andwere previously described [30, 33].
PBMCs from normal donors were activated with PHA (10 �g/ml; SigmaChemical Co., St. Louis, MO) and recombinant human interleukin-2 (rhIL-2;100 U/ml; NIH AIDS Reagent Program) for 3 days. PBMCs were depleted ofCD8(�) T cells by positive selection of cytotoxic cells by microbeads coatedwith antibodies to CD8 (Mitenyi Biotec, Auburn, CA). CD8(�) T cells weresubsequently used in the fusion assay or in HIV-1 infection assays.
Chemokines, antibodies, and soluble proteins
All chemokines including CCL22(1–69) and CCL22(3–69) were purchasedfrom PeproTech (Rocky Hill, NJ). CCL22(3–69) is a truncated form ofCCL22(1–69) in which the first two amino acids are deleted. Polyclonalantibodies to CCR4 were raised in rabbits using a synthetic peptide corre-sponding to the first 15 amino acids of the N-terminus of CCR4. mAb to CCR4were initially obtained from Dr. Lijun Wu (Millennium Inc., Cambridge, MA)and were subsequently purchased from PharMingen (San Diego, CA). Fluo-rescein isothiocyanate (FITC)-conjugated anti-CD4 and phycoerythrin (PE)-conjugated CXCR4 and CCR5 were purchased from R&D Systems (Minneap-olis, MN). The isotype-matched antibody controls, immunoglobulin G (IgG)2a-PE, IgG1-PE, and IgG1-FITC, were all purchased from PharMingen. Solublegp120 was obtained from the laboratory of Dr. Chris Broder. Soluble CD4 wasobtained from the laboratory of Dr. Edward A. Berger (donated to Edward A.Berger by S. Johnson, Pharmacia Upjohn, Kalamazoo, MI).
Macrophage cDNA library, receptor expressionplasmids, and sequencing
The human macrophage cDNA library (purchased from Invitrogen, San Diego,CA) was prepared in pCDNA3.1 using a unidirectional cloning strategy underthe T-7 and cytomegalovirus (CMV) promoters. The cDNA library was used ina functional screening protocol to isolate a cDNA clone that conferred Env-fusion competence to murine cells. The screening method has been previouslydescribed [31]. The isolated cDNA was used to transfect (using DOTAP)NIH-3T3 cells, and expression of the cDNA clones was activated by infectionwith vTF7-3 (T7 RNA polymerase). The transfected cells were then infectedwith vCB-3 (CD4) to allow expression of the CD4 surface antigen. EffectorHeLa cells were coinfected with vCB21R(lacZ) and vCB-43 (Ba-L), vEV-1(89.6), or vCB-16 (Unc). Fusion activity, as a result of mixing the two cellpopulations, was detected by staining the fused cells with X-gal and countingblue cells. It is interesting that one of the cDNA clones gave a positive fusionsignal with primary 89.6 Env but not with the R5 Ba-L. This cDNA clone wasdesignated CCRX. Nucleotide sequencing of the cDNA fragment encodingCCRX indicated that it is identical to CCR4. The nucleotide sequence wasconfirmed by double-stranded DNA sequencing using the Sequenase DNAsequencing kit, Version 2.0 (USB, Cleveland, OH). The cDNA fragmentencoding CCR4 was purified and cloned into pSC59 [34] under the control ofa synthetic early/late vaccinia virus promoter and subsequently used to trans-fect 3T3.CD4 cells to determine its Env-mediated fusion specificity.
Env-mediated fusion and single-roundinfection assays
Coreceptor activity was determined by a vaccinia-based reporter gene assayquantitating Env-mediated cell fusion [35]. The cell fusion assay involves theanalysis of fusion between two distinct cell populations, one expressing CD4(endogenous or encoded by a recombinant virus) and the other expressing theindicated HIV-1 Env glycoprotein encoded by a recombinant vaccinia virus.Cell fusion is scored by a reporter gene activation assay in which the cytoplasmof one cell population expressing vaccinia virus-encoded T7 RNA polymeraseand the cytoplasm of the other expressing lacZ gene linked to the T7 promoter;cell fusion activates �-galactosidase. The cDNA fragments encoding the openreading frames of chemokine receptor CCR4 or CCR5 cloned in pSC59 [30]were used in subsequent fusion assays. Plasmids encoding CCR5 or CCR4were transfected into NIH-3T3 cells using DOTAP lipofection (BoehringerMannheim, Indianapolis, IN). After 4 h incubation with DOTAP at 37°C, cells
1064 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org
were infected with vTF7-3 (encoding T7 RNA polymerase under the control ofthe vaccinia virus early/late synthetic promoter) and vCB-3 (encoding humanCD4) at a multiplicity of infection of 10 pfu/cell for each virus. Control cellswere transfected with the pCDNA3.1 or pSC59 vector alone. Effector HeLacells were infected with vCB-21R (encoding �-galactosidase under control ofthe T7 promoter) and recombinant vaccinia viruses encoding one of thefollowing HIV-1 Envs: vCB-16 (uncleaved mutant of IIIB Env as a negativecontrol), vCB-43 (encoding Ba-L), vCB-39 (encoding ADA), vCB-28 (encodingJR-FL), vCB-41 (encoding LAV), vSC60 (encoding IIIB), 89.6, vCB-74 (en-coding SIV-mac239), or vCB-75 (encoding SIV-mac316). All cell populations(effectors and targets) were incubated overnight at 31°C to allow expression ofthe vaccinia-encoded proteins and were then washed and resuspended inEarle’s modified Eagle’s medium with 2.5% FBS. Duplicate samples contain-ing 105 target cells expressing CD4 and 105 effector cells (expressing theindicated Env) were mixed in a 96-well microtiter plate and incubated at 37°Cfor 2 h. �-Galactosidase activity, produced as a result of cell fusion activitybetween partner cells, was quantitated by colorimetric assay of detergent-treated cell lysates [34].
HIV-1 pseudotyped viral stocks were prepared as previously described[36]. Briefly, the HIV-1 genome used in this protocol includes the lucif-erase gene under the regulation of the SV40 promoter replacing the HIV-1Env gene (HIV�Luc�Env�). Producer 293T cells were transfected withHIV�Luc�Env� and another plasmid DNA containing the R5 ADAHIV-1 Env. After 60 –72 h, pseudotyped viral particles were harvestedfrom the culture supernatants and titered by measuring the amount of p24gag viral antigen.
To analyze the effect of chemokines on Env fusion activity, the CC chemo-kines CCL22(1–69), CCL22(3–69), CCL17, or CCL5 were added individuallyto target cells and incubated for 30–45 min at 37°C before mixing withEnv-expressing HeLa cells or infection with the pseudotyped virus.
Productive HIV-1 infection assays
CD8(�) T cells were depleted from PBMCs that had been stimulated withPHA � rIL-2 for 3 days. Total PBMCs and CD8-depleted PBMC were used inthe infectivity assay. Infection was performed in a 96-well plate (105 cells/well). Cells were incubated with chemokines CCL22(3–69) or CCL2 for 1 h,inoculated with ADA (105.5 IU/ml) or IIIB (104.5 IU/ml) for 3 h, and washedafter virus adsorption, and fresh medium containing chemokines was added.Production of p24 was measured over the course of 12 days using a p24enzyme-linked immunosorbent assay kit purchased from NCI-FCDRC (Fred-erick, MD). Monocyte chemoattractant protein-1 (MCP-1; R&D Systems) wasused as negative control. CCL4 and CCL5 (PeproTech) and azidothymidine(AZT; 1 �M) were used as controls for inhibition of productive infection.Cell-containing aliquots were taken every 3 days and replaced with freshmedium containing fresh chemokines.
Cell-surface expression and QFACS
Human PBMCs stimulated with PHA � IL-2 were washed twice in FACSbuffer (Hanks’ balanced saline solution supplemented with 0.5% FBS and0.02% sodium azide) and were resuspended in 100 �l FACS buffer at 107/ml.Cells were then incubated with a 1:200 dilution of mAb raised against CCR4[28] at 4°C for 30 min. Cells were then washed twice, resuspended in 100 �lice-cold FACS buffer in the presence of PE-conjugated antimouse IgG (Phar-Mingen), and incubated at 4°C for 30 min. Finally, cells were washed twice,resuspended in 500 �l ice-cold FACS buffer, and analyzed in a FACScancytometer (Becton-Dickinson, San Jose, CA). QFACS was performed by con-verting the mean channel fluorescence into ABS by using a standardizedmicrobeads kit purchased from Sigma Chemical Co. For quantitative analysisof ABS, cells were reacted with FITC-conjugated (anti-CD4) or PE-conjugated(anti-CCR4, anti-CCR5, and anti-CXCR4) antibodies. The conjugated anti-bodies were added at saturating amounts to 105 microbeads, and the meanfluorescence intensity (MFI) of the stained receptor was converted into ABS bycoupling with the standard regression (once generated). ABS in each experi-ment was calculated as: ABS value of stained sample minus ABS valueobtained with staining the cells with the isotype-matched control.
Confocal microscopy
PHA � IL-2-activated PBMCs were stained with an anti-CCR4-PE or anti-CCR5-PE mAb followed by anti-CD4-FITC in suspension. The cells were then
fixed in 2% paraformaldehyde, and colocalization was performed on a glass-bottom microwell Petri dish (MatTek, Ashland, MA). Data were collected by ascanning confocal system (Bio-Rad, Hercules, CA) with a krypton/argon laserconfigured on an Axiovert microscope (Ziess, Oberkochen, Germany). Imageswere obtained with a 60� oil immersion objective lens.
Soluble gp120 binding assay
NIH-3T3 cells (2�106) were transfected with pcDNA3 vector, pcDNA3/CCR5,or pcDNA3/CCR4. Expression of CCR5 or CCR4 was activated by infectionwith vTF7-3 (vaccinia encoding T7 RNA polymerase). The cells were incu-bated in duplicates with 0.25 nM I125-labeled gp120 (89.6) at a specificactivity of 2200 Ci/mmol plus varying concentrations of unlabeled gp120 and100 nM sCD4 (NIH AIDS Reagent Program) in 20 �l binding buffer (50 mMHEPES with 0.5% bovine serum albumin, 1 mM CaCl2, and MgCl2, pH 7.2).After 1 h incubation at room temperature, cells were washed with bindingbuffer containing 0.5 M sodium chloride, and cell pellet-associated countswere measured in a � counter.
RESULTS
CCR4 coreceptor activity in transfectedmurine cells
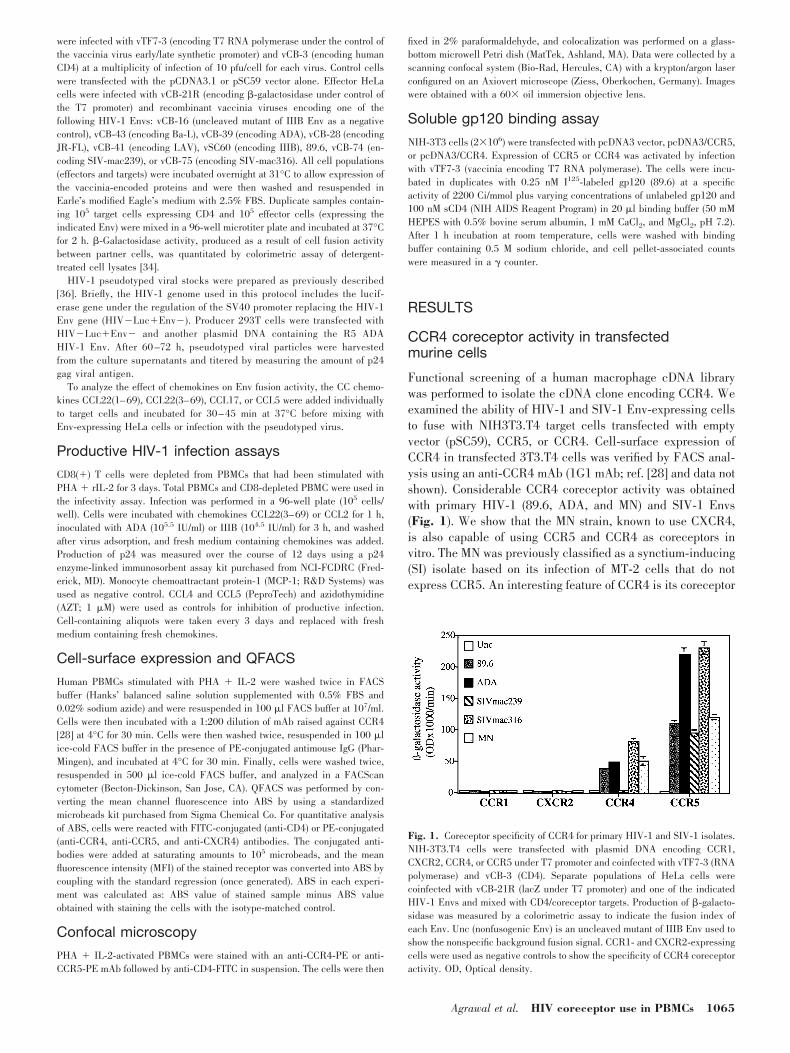
Functional screening of a human macrophage cDNA librarywas performed to isolate the cDNA clone encoding CCR4. Weexamined the ability of HIV-1 and SIV-1 Env-expressing cellsto fuse with NIH3T3.T4 target cells transfected with emptyvector (pSC59), CCR5, or CCR4. Cell-surface expression ofCCR4 in transfected 3T3.T4 cells was verified by FACS anal-ysis using an anti-CCR4 mAb (1G1 mAb; ref. [28] and data notshown). Considerable CCR4 coreceptor activity was obtainedwith primary HIV-1 (89.6, ADA, and MN) and SIV-1 Envs(Fig. 1). We show that the MN strain, known to use CXCR4,is also capable of using CCR5 and CCR4 as coreceptors invitro. The MN was previously classified as a synctium-inducing(SI) isolate based on its infection of MT-2 cells that do notexpress CCR5. An interesting feature of CCR4 is its coreceptor
Fig. 1. Coreceptor specificity of CCR4 for primary HIV-1 and SIV-1 isolates.NIH-3T3.T4 cells were transfected with plasmid DNA encoding CCR1,CXCR2, CCR4, or CCR5 under T7 promoter and coinfected with vTF7-3 (RNApolymerase) and vCB-3 (CD4). Separate populations of HeLa cells werecoinfected with vCB-21R (lacZ under T7 promoter) and one of the indicatedHIV-1 Envs and mixed with CD4/coreceptor targets. Production of �-galacto-sidase was measured by a colorimetric assay to indicate the fusion index ofeach Env. Unc (nonfusogenic Env) is an uncleaved mutant of IIIB Env used toshow the nonspecific background fusion signal. CCR1- and CXCR2-expressingcells were used as negative controls to show the specificity of CCR4 coreceptoractivity. OD, Optical density.
Agrawal et al. HIV coreceptor use in PBMCs 1065
activity with the macrophage-tropic SIV-mac316 and not withthe T cell line-tropic SIV-mac239, which reached 50% of theactivity obtained with CCR5 (Fig. 1). These experiments dem-onstrated functional HIV-1 coreceptor activity of the isolatedCCR4 cDNA clone in transfected murine cells.
Chemokine inhibition of CCR4 coreceptor activity
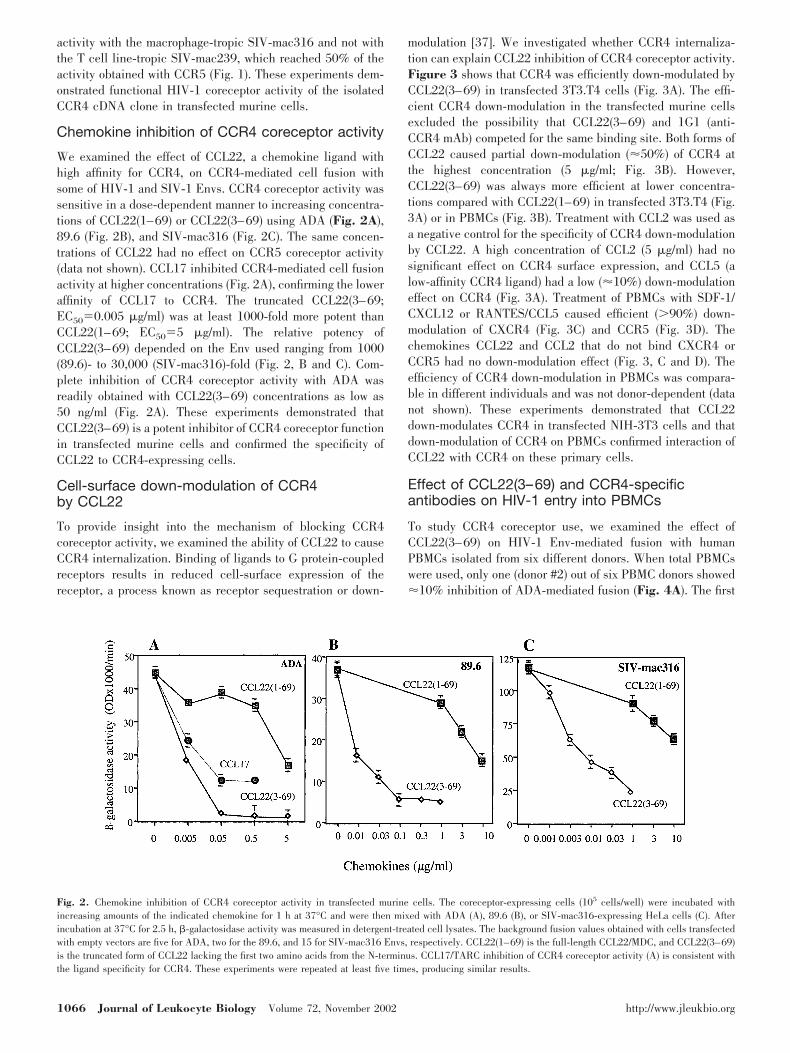
We examined the effect of CCL22, a chemokine ligand withhigh affinity for CCR4, on CCR4-mediated cell fusion withsome of HIV-1 and SIV-1 Envs. CCR4 coreceptor activity wassensitive in a dose-dependent manner to increasing concentra-tions of CCL22(1–69) or CCL22(3–69) using ADA (Fig. 2A),89.6 (Fig. 2B), and SIV-mac316 (Fig. 2C). The same concen-trations of CCL22 had no effect on CCR5 coreceptor activity(data not shown). CCL17 inhibited CCR4-mediated cell fusionactivity at higher concentrations (Fig. 2A), confirming the loweraffinity of CCL17 to CCR4. The truncated CCL22(3–69;EC50�0.005 �g/ml) was at least 1000-fold more potent thanCCL22(1–69; EC50�5 �g/ml). The relative potency ofCCL22(3–69) depended on the Env used ranging from 1000(89.6)- to 30,000 (SIV-mac316)-fold (Fig. 2, B and C). Com-plete inhibition of CCR4 coreceptor activity with ADA wasreadily obtained with CCL22(3–69) concentrations as low as50 ng/ml (Fig. 2A). These experiments demonstrated thatCCL22(3–69) is a potent inhibitor of CCR4 coreceptor functionin transfected murine cells and confirmed the specificity ofCCL22 to CCR4-expressing cells.
Cell-surface down-modulation of CCR4by CCL22
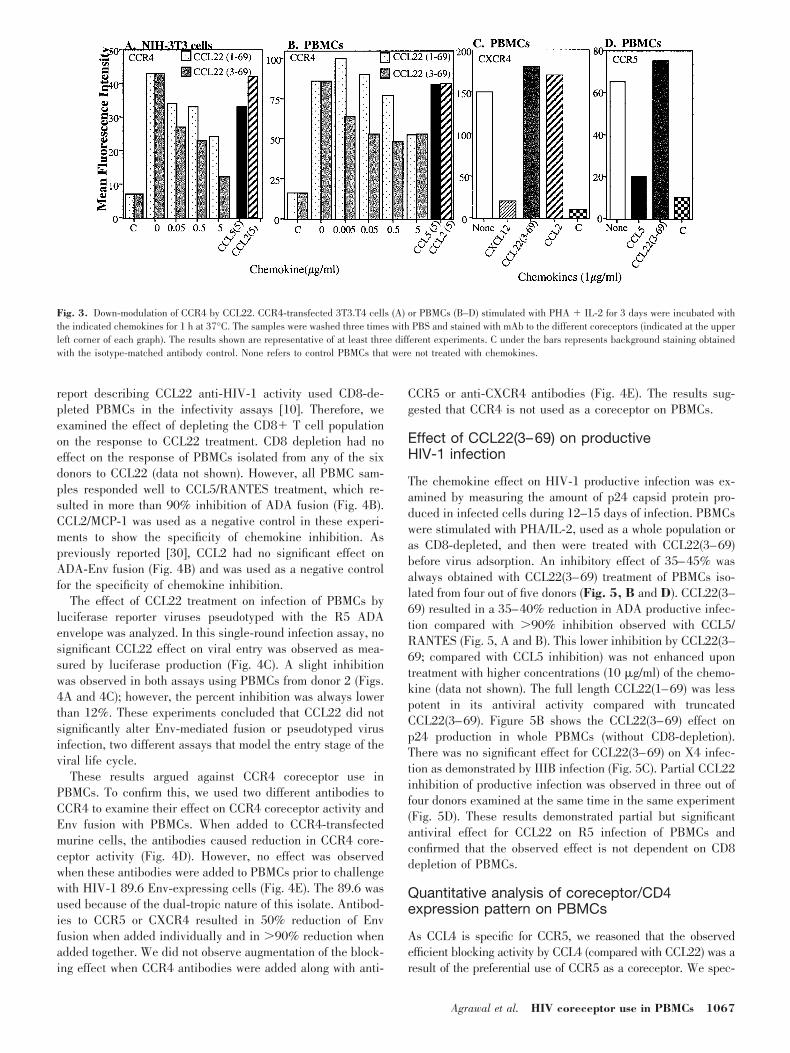
To provide insight into the mechanism of blocking CCR4coreceptor activity, we examined the ability of CCL22 to causeCCR4 internalization. Binding of ligands to G protein-coupledreceptors results in reduced cell-surface expression of thereceptor, a process known as receptor sequestration or down-
modulation [37]. We investigated whether CCR4 internaliza-tion can explain CCL22 inhibition of CCR4 coreceptor activity.Figure 3 shows that CCR4 was efficiently down-modulated byCCL22(3–69) in transfected 3T3.T4 cells (Fig. 3A). The effi-cient CCR4 down-modulation in the transfected murine cellsexcluded the possibility that CCL22(3–69) and 1G1 (anti-CCR4 mAb) competed for the same binding site. Both forms ofCCL22 caused partial down-modulation (50%) of CCR4 atthe highest concentration (5 �g/ml; Fig. 3B). However,CCL22(3–69) was always more efficient at lower concentra-tions compared with CCL22(1–69) in transfected 3T3.T4 (Fig.3A) or in PBMCs (Fig. 3B). Treatment with CCL2 was used asa negative control for the specificity of CCR4 down-modulationby CCL22. A high concentration of CCL2 (5 �g/ml) had nosignificant effect on CCR4 surface expression, and CCL5 (alow-affinity CCR4 ligand) had a low (10%) down-modulationeffect on CCR4 (Fig. 3A). Treatment of PBMCs with SDF-1/CXCL12 or RANTES/CCL5 caused efficient (90%) down-modulation of CXCR4 (Fig. 3C) and CCR5 (Fig. 3D). Thechemokines CCL22 and CCL2 that do not bind CXCR4 orCCR5 had no down-modulation effect (Fig. 3, C and D). Theefficiency of CCR4 down-modulation in PBMCs was compara-ble in different individuals and was not donor-dependent (datanot shown). These experiments demonstrated that CCL22down-modulates CCR4 in transfected NIH-3T3 cells and thatdown-modulation of CCR4 on PBMCs confirmed interaction ofCCL22 with CCR4 on these primary cells.
Effect of CCL22(3–69) and CCR4-specificantibodies on HIV-1 entry into PBMCs
To study CCR4 coreceptor use, we examined the effect ofCCL22(3–69) on HIV-1 Env-mediated fusion with humanPBMCs isolated from six different donors. When total PBMCswere used, only one (donor #2) out of six PBMC donors showed10% inhibition of ADA-mediated fusion (Fig. 4A). The first
Fig. 2. Chemokine inhibition of CCR4 coreceptor activity in transfected murine cells. The coreceptor-expressing cells (105 cells/well) were incubated withincreasing amounts of the indicated chemokine for 1 h at 37°C and were then mixed with ADA (A), 89.6 (B), or SIV-mac316-expressing HeLa cells (C). Afterincubation at 37°C for 2.5 h, �-galactosidase activity was measured in detergent-treated cell lysates. The background fusion values obtained with cells transfectedwith empty vectors are five for ADA, two for the 89.6, and 15 for SIV-mac316 Envs, respectively. CCL22(1–69) is the full-length CCL22/MDC, and CCL22(3–69)is the truncated form of CCL22 lacking the first two amino acids from the N-terminus. CCL17/TARC inhibition of CCR4 coreceptor activity (A) is consistent withthe ligand specificity for CCR4. These experiments were repeated at least five times, producing similar results.
1066 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org
report describing CCL22 anti-HIV-1 activity used CD8-de-pleted PBMCs in the infectivity assays [10]. Therefore, weexamined the effect of depleting the CD8� T cell populationon the response to CCL22 treatment. CD8 depletion had noeffect on the response of PBMCs isolated from any of the sixdonors to CCL22 (data not shown). However, all PBMC sam-ples responded well to CCL5/RANTES treatment, which re-sulted in more than 90% inhibition of ADA fusion (Fig. 4B).CCL2/MCP-1 was used as a negative control in these experi-ments to show the specificity of chemokine inhibition. Aspreviously reported [30], CCL2 had no significant effect onADA-Env fusion (Fig. 4B) and was used as a negative controlfor the specificity of chemokine inhibition.
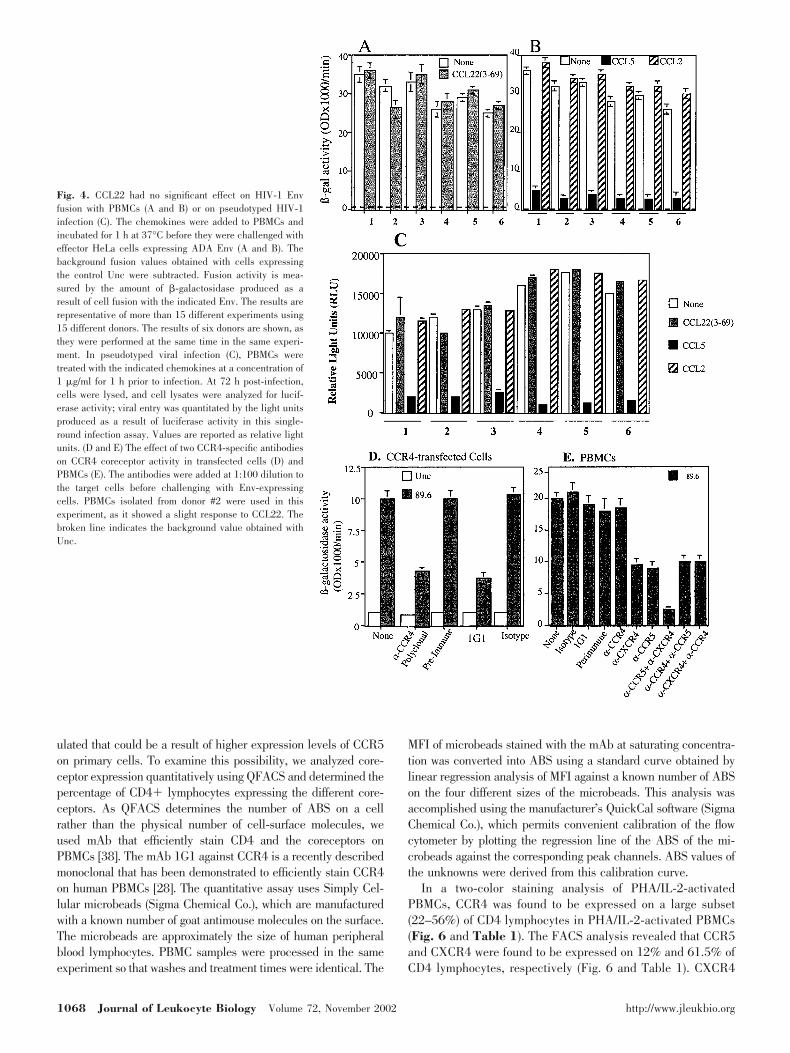
The effect of CCL22 treatment on infection of PBMCs byluciferase reporter viruses pseudotyped with the R5 ADAenvelope was analyzed. In this single-round infection assay, nosignificant CCL22 effect on viral entry was observed as mea-sured by luciferase production (Fig. 4C). A slight inhibitionwas observed in both assays using PBMCs from donor 2 (Figs.4A and 4C); however, the percent inhibition was always lowerthan 12%. These experiments concluded that CCL22 did notsignificantly alter Env-mediated fusion or pseudotyped virusinfection, two different assays that model the entry stage of theviral life cycle.
These results argued against CCR4 coreceptor use inPBMCs. To confirm this, we used two different antibodies toCCR4 to examine their effect on CCR4 coreceptor activity andEnv fusion with PBMCs. When added to CCR4-transfectedmurine cells, the antibodies caused reduction in CCR4 core-ceptor activity (Fig. 4D). However, no effect was observedwhen these antibodies were added to PBMCs prior to challengewith HIV-1 89.6 Env-expressing cells (Fig. 4E). The 89.6 wasused because of the dual-tropic nature of this isolate. Antibod-ies to CCR5 or CXCR4 resulted in 50% reduction of Envfusion when added individually and in 90% reduction whenadded together. We did not observe augmentation of the block-ing effect when CCR4 antibodies were added along with anti-
CCR5 or anti-CXCR4 antibodies (Fig. 4E). The results sug-gested that CCR4 is not used as a coreceptor on PBMCs.
Effect of CCL22(3–69) on productiveHIV-1 infection
The chemokine effect on HIV-1 productive infection was ex-amined by measuring the amount of p24 capsid protein pro-duced in infected cells during 12–15 days of infection. PBMCswere stimulated with PHA/IL-2, used as a whole population oras CD8-depleted, and then were treated with CCL22(3–69)before virus adsorption. An inhibitory effect of 35–45% wasalways obtained with CCL22(3–69) treatment of PBMCs iso-lated from four out of five donors (Fig. 5, B and D). CCL22(3–69) resulted in a 35–40% reduction in ADA productive infec-tion compared with 90% inhibition observed with CCL5/RANTES (Fig. 5, A and B). This lower inhibition by CCL22(3–69; compared with CCL5 inhibition) was not enhanced upontreatment with higher concentrations (10 �g/ml) of the chemo-kine (data not shown). The full length CCL22(1–69) was lesspotent in its antiviral activity compared with truncatedCCL22(3–69). Figure 5B shows the CCL22(3–69) effect onp24 production in whole PBMCs (without CD8-depletion).There was no significant effect for CCL22(3–69) on X4 infec-tion as demonstrated by IIIB infection (Fig. 5C). Partial CCL22inhibition of productive infection was observed in three out offour donors examined at the same time in the same experiment(Fig. 5D). These results demonstrated partial but significantantiviral effect for CCL22 on R5 infection of PBMCs andconfirmed that the observed effect is not dependent on CD8depletion of PBMCs.
Quantitative analysis of coreceptor/CD4expression pattern on PBMCs
As CCL4 is specific for CCR5, we reasoned that the observedefficient blocking activity by CCL4 (compared with CCL22) was aresult of the preferential use of CCR5 as a coreceptor. We spec-
Fig. 3. Down-modulation of CCR4 by CCL22. CCR4-transfected 3T3.T4 cells (A) or PBMCs (B–D) stimulated with PHA � IL-2 for 3 days were incubated withthe indicated chemokines for 1 h at 37°C. The samples were washed three times with PBS and stained with mAb to the different coreceptors (indicated at the upperleft corner of each graph). The results shown are representative of at least three different experiments. C under the bars represents background staining obtainedwith the isotype-matched antibody control. None refers to control PBMCs that were not treated with chemokines.
Agrawal et al. HIV coreceptor use in PBMCs 1067
ulated that could be a result of higher expression levels of CCR5on primary cells. To examine this possibility, we analyzed core-ceptor expression quantitatively using QFACS and determined thepercentage of CD4� lymphocytes expressing the different core-ceptors. As QFACS determines the number of ABS on a cellrather than the physical number of cell-surface molecules, weused mAb that efficiently stain CD4 and the coreceptors onPBMCs [38]. The mAb 1G1 against CCR4 is a recently describedmonoclonal that has been demonstrated to efficiently stain CCR4on human PBMCs [28]. The quantitative assay uses Simply Cel-lular microbeads (Sigma Chemical Co.), which are manufacturedwith a known number of goat antimouse molecules on the surface.The microbeads are approximately the size of human peripheralblood lymphocytes. PBMC samples were processed in the sameexperiment so that washes and treatment times were identical. The
MFI of microbeads stained with the mAb at saturating concentra-tion was converted into ABS using a standard curve obtained bylinear regression analysis of MFI against a known number of ABSon the four different sizes of the microbeads. This analysis wasaccomplished using the manufacturer’s QuickCal software (SigmaChemical Co.), which permits convenient calibration of the flowcytometer by plotting the regression line of the ABS of the mi-crobeads against the corresponding peak channels. ABS values ofthe unknowns were derived from this calibration curve.
In a two-color staining analysis of PHA/IL-2-activatedPBMCs, CCR4 was found to be expressed on a large subset(22–56%) of CD4 lymphocytes in PHA/IL-2-activated PBMCs(Fig. 6 and Table 1). The FACS analysis revealed that CCR5and CXCR4 were found to be expressed on 12% and 61.5% ofCD4 lymphocytes, respectively (Fig. 6 and Table 1). CXCR4
Fig. 4. CCL22 had no significant effect on HIV-1 Envfusion with PBMCs (A and B) or on pseudotyped HIV-1infection (C). The chemokines were added to PBMCs andincubated for 1 h at 37°C before they were challenged witheffector HeLa cells expressing ADA Env (A and B). Thebackground fusion values obtained with cells expressingthe control Unc were subtracted. Fusion activity is mea-sured by the amount of �-galactosidase produced as aresult of cell fusion with the indicated Env. The results arerepresentative of more than 15 different experiments using15 different donors. The results of six donors are shown, asthey were performed at the same time in the same experi-ment. In pseudotyped viral infection (C), PBMCs weretreated with the indicated chemokines at a concentration of1 �g/ml for 1 h prior to infection. At 72 h post-infection,cells were lysed, and cell lysates were analyzed for lucif-erase activity; viral entry was quantitated by the light unitsproduced as a result of luciferase activity in this single-round infection assay. Values are reported as relative lightunits. (D and E) The effect of two CCR4-specific antibodieson CCR4 coreceptor activity in transfected cells (D) andPBMCs (E). The antibodies were added at 1:100 dilution tothe target cells before challenging with Env-expressingcells. PBMCs isolated from donor #2 were used in thisexperiment, as it showed a slight response to CCL22. Thebroken line indicates the background value obtained withUnc.
1068 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org
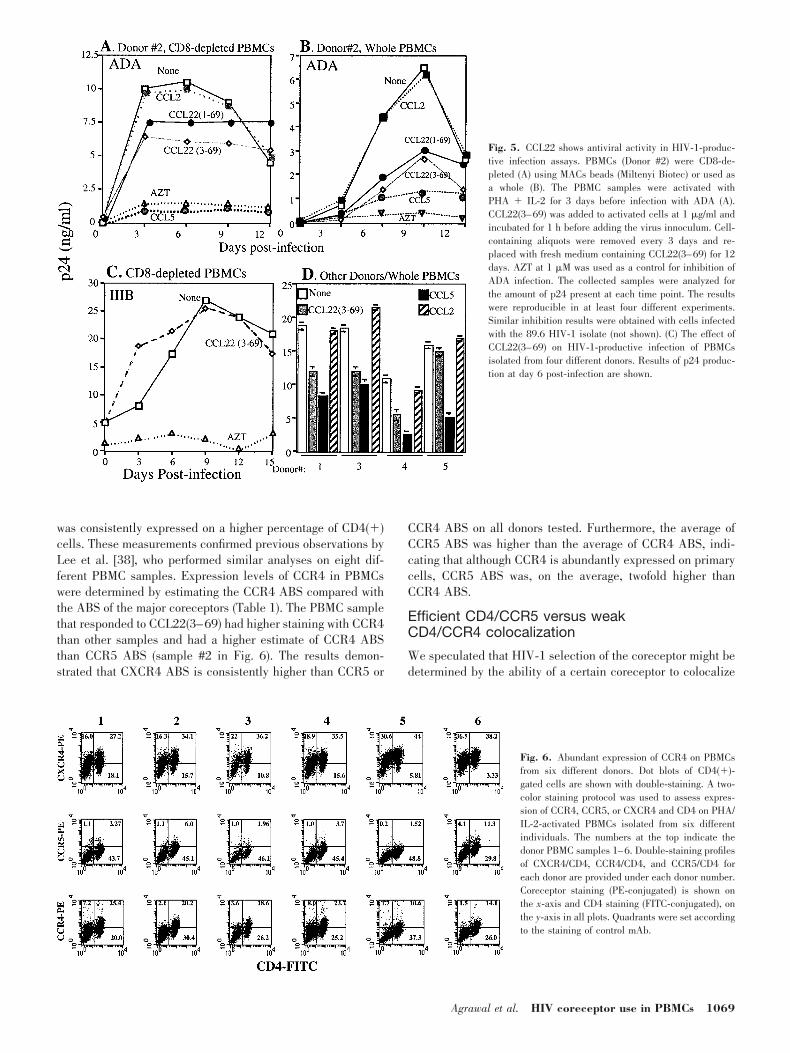
was consistently expressed on a higher percentage of CD4(�)cells. These measurements confirmed previous observations byLee et al. [38], who performed similar analyses on eight dif-ferent PBMC samples. Expression levels of CCR4 in PBMCswere determined by estimating the CCR4 ABS compared withthe ABS of the major coreceptors (Table 1). The PBMC samplethat responded to CCL22(3–69) had higher staining with CCR4than other samples and had a higher estimate of CCR4 ABSthan CCR5 ABS (sample #2 in Fig. 6). The results demon-strated that CXCR4 ABS is consistently higher than CCR5 or
CCR4 ABS on all donors tested. Furthermore, the average ofCCR5 ABS was higher than the average of CCR4 ABS, indi-cating that although CCR4 is abundantly expressed on primarycells, CCR5 ABS was, on the average, twofold higher thanCCR4 ABS.
Efficient CD4/CCR5 versus weakCD4/CCR4 colocalization
We speculated that HIV-1 selection of the coreceptor might bedetermined by the ability of a certain coreceptor to colocalize
Fig. 5. CCL22 shows antiviral activity in HIV-1-produc-tive infection assays. PBMCs (Donor #2) were CD8-de-pleted (A) using MACs beads (Miltenyi Biotec) or used asa whole (B). The PBMC samples were activated withPHA � IL-2 for 3 days before infection with ADA (A).CCL22(3–69) was added to activated cells at 1 �g/ml andincubated for 1 h before adding the virus innoculum. Cell-containing aliquots were removed every 3 days and re-placed with fresh medium containing CCL22(3–69) for 12days. AZT at 1 �M was used as a control for inhibition ofADA infection. The collected samples were analyzed forthe amount of p24 present at each time point. The resultswere reproducible in at least four different experiments.Similar inhibition results were obtained with cells infectedwith the 89.6 HIV-1 isolate (not shown). (C) The effect ofCCL22(3–69) on HIV-1-productive infection of PBMCsisolated from four different donors. Results of p24 produc-tion at day 6 post-infection are shown.
Fig. 6. Abundant expression of CCR4 on PBMCsfrom six different donors. Dot blots of CD4(�)-gated cells are shown with double-staining. A two-color staining protocol was used to assess expres-sion of CCR4, CCR5, or CXCR4 and CD4 on PHA/IL-2-activated PBMCs isolated from six differentindividuals. The numbers at the top indicate thedonor PBMC samples 1–6. Double-staining profilesof CXCR4/CD4, CCR4/CD4, and CCR5/CD4 foreach donor are provided under each donor number.Coreceptor staining (PE-conjugated) is shown onthe x-axis and CD4 staining (FITC-conjugated), onthe y-axis in all plots. Quadrants were set accordingto the staining of control mAb.
Agrawal et al. HIV coreceptor use in PBMCs 1069

with CD4. The viral Env may preferentially use coreceptorsfound in close proximity with CD4. Evidence for CD4/CCR5colocalization and physical association has been previouslydemonstrated [39]; however, colocalization of CD4 with theother minor coreceptors has never been investigated. We an-alyzed CCR4 colocalization with CD4 by confocal microscopy,where colocalization of two proteins can be demonstrated. Toidentify fluorescence colocalization, correlation maps were cal-culated by using a local statistical method. We consistentlyobserved weaker colocalization of CD4 and CCR4 in all PBMCsamples tested. Figure 7A shows a representative experiment.Colocalization of CCR5 and CD4 proteins was always moreefficient, as demonstrated by the strong yellow (red-green co-localization) staining (Fig. 7A). To determine whether CD4/CCR4 can colocalize in transfected murine cells, we performedthe same confocal analysis on NIH-3T3 cells transiently ex-pressing CCR5 or CCR4. The results indicated that CCR5/CD4colocalization was pronounced and much stronger than CCR4/CD4 colocalization in murine 3T3 cells transiently expressingthe coreceptors (Fig. 7B). We reproduced these colocalizationresults using other mAb to human CD4 (OKT4, Ortho Diag-nostic Systems, Raritan, NJ; T4-4 from NIH AIDS ReagentProgram). The data suggested that colocalization with CD4 isan important factor that may contribute to the preferential useof CCR5 on PBMCs.
HIV-1 gp120 has higher affinity to CCR5
We used a technique based on previously published bindingexperiments using soluble gp120, in which CD4 was shown tosignificantly enhance the gp120 coreceptor-binding interaction[40–46]. Interaction of HIV-1 gp120 subunit with coreceptor-expressing cells was analyzed by comparing gp120 bindingwith CCR5 versus CCR4-expressing murine cells. NIH-3T3cells were transfected with empty vector pCDNA3/CCR5 orpCDNA3/CCR4. Expression of the coreceptor was activated byinfection with vTF7-3 (encoding T7 RNA polymerase). Theability of the purified gp120 subunit of HIV-1 Env to competewith I125-labeled gp120 derived from the primary 89.6 Env wasstudied in the presence of soluble CD4. Our choice of thesoluble gp120 subunit was based on the fact that CCR4-expressing cells showed detectable coreceptor activity with the89.6 Env (Fig. 1). Radiolabeled gp120 bound to CCR5- orCCR4-expressing murine cells was competed out with coldgp120. The results demonstrated that radiolabeled gp120bound 104 times more efficiently to CCR5-expressing cells(Fig. 8A). This high signal of bound I125–gp120 was competedout with 1 nM cold gp120 added to CCR5-expressing cells.Despite the much lower signal of bound I125–gp120 on CCR4-expressing cells, a much higher concentration was needed tocompete out 50% of the bound radioactivity (Fig. 8B), whereas
TABLE 1. Comparison of CXCR4, CCR4, CCR5, and CD4 ABS on PBMCs
PBMCs
Mean positive, % of104 cells analyzed(range, n � 6)
Median ABS/cell(range, n � 6)
Median *% CD4(�)/coreceptor(�)(range, n � 6) Median ABS on CD4� cells
CCR4 22.75% (11–34%) 15,745 (7491–33,252) 40.4% (22–56%) CD4�CCR4� 16,802 (12,748–22,655)CCR5 12% (6.3–23%) 35,410 (13,606–66,603) 10.05% (3–27.3%) CD4�CCR5� 33,812 (18,903–52,472)CXCR4 61.5% (40–78%) 46,265 (28,783–85,674) 75.9% (60–92%) CD4�CXCR4� 59,578 (42,777–79,070)CD4 46.6% (42.2–50.6%) 125,662 (108,546–149,743) NA 118,283 (90,043–1,454,320)
Percent-positive gate is defined as the histogram gate giving �2% positive cells using an isotype control. The corresponding ABS values were calculated basedon the gated positive population only. The ABS values obtained with the isotype control were subtracted. * Calculations are based on Figure 6 showing doublestaining of coreceptor and CD4.
Fig. 7. Confocal microscopic analysis of coreceptor/CD4 colocalization. Activated PBMCs (A) or transfected NIH-3T3 cells (B) were stained with ananti-CCR4-PE or anti-CCR5-PE mAb followed by FITC-coupled anti-CD4 in suspension. The cells were then fixed in 2% paraformaldehyde, and colocalizationwas performed on a glass-bottom microwell petri dish (MatTek). Data were gathered by a scanning confocal system (Bio-Rad) with a krypton/argon laser configuredon an Axiovert microscope (Ziess).
1070 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org
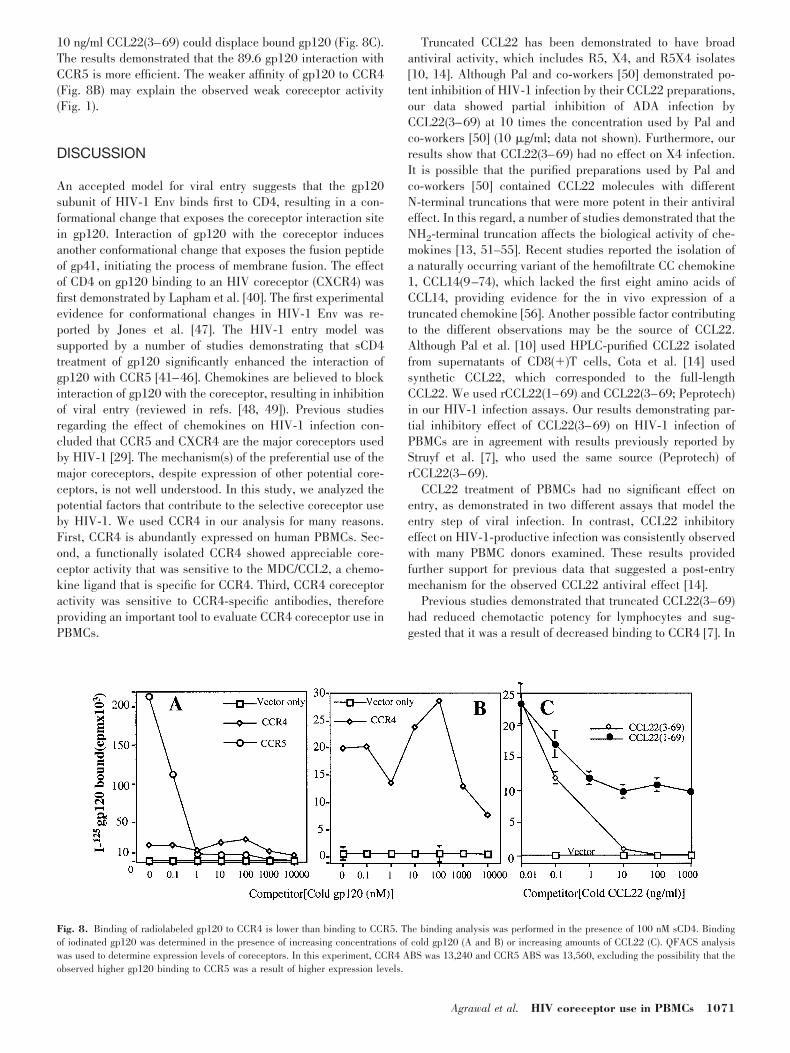
10 ng/ml CCL22(3–69) could displace bound gp120 (Fig. 8C).The results demonstrated that the 89.6 gp120 interaction withCCR5 is more efficient. The weaker affinity of gp120 to CCR4(Fig. 8B) may explain the observed weak coreceptor activity(Fig. 1).
DISCUSSION
An accepted model for viral entry suggests that the gp120subunit of HIV-1 Env binds first to CD4, resulting in a con-formational change that exposes the coreceptor interaction sitein gp120. Interaction of gp120 with the coreceptor inducesanother conformational change that exposes the fusion peptideof gp41, initiating the process of membrane fusion. The effectof CD4 on gp120 binding to an HIV coreceptor (CXCR4) wasfirst demonstrated by Lapham et al. [40]. The first experimentalevidence for conformational changes in HIV-1 Env was re-ported by Jones et al. [47]. The HIV-1 entry model wassupported by a number of studies demonstrating that sCD4treatment of gp120 significantly enhanced the interaction ofgp120 with CCR5 [41–46]. Chemokines are believed to blockinteraction of gp120 with the coreceptor, resulting in inhibitionof viral entry (reviewed in refs. [48, 49]). Previous studiesregarding the effect of chemokines on HIV-1 infection con-cluded that CCR5 and CXCR4 are the major coreceptors usedby HIV-1 [29]. The mechanism(s) of the preferential use of themajor coreceptors, despite expression of other potential core-ceptors, is not well understood. In this study, we analyzed thepotential factors that contribute to the selective coreceptor useby HIV-1. We used CCR4 in our analysis for many reasons.First, CCR4 is abundantly expressed on human PBMCs. Sec-ond, a functionally isolated CCR4 showed appreciable core-ceptor activity that was sensitive to the MDC/CCL2, a chemo-kine ligand that is specific for CCR4. Third, CCR4 coreceptoractivity was sensitive to CCR4-specific antibodies, thereforeproviding an important tool to evaluate CCR4 coreceptor use inPBMCs.
Truncated CCL22 has been demonstrated to have broadantiviral activity, which includes R5, X4, and R5X4 isolates[10, 14]. Although Pal and co-workers [50] demonstrated po-tent inhibition of HIV-1 infection by their CCL22 preparations,our data showed partial inhibition of ADA infection byCCL22(3–69) at 10 times the concentration used by Pal andco-workers [50] (10 �g/ml; data not shown). Furthermore, ourresults show that CCL22(3–69) had no effect on X4 infection.It is possible that the purified preparations used by Pal andco-workers [50] contained CCL22 molecules with differentN-terminal truncations that were more potent in their antiviraleffect. In this regard, a number of studies demonstrated that theNH2-terminal truncation affects the biological activity of che-mokines [13, 51–55]. Recent studies reported the isolation ofa naturally occurring variant of the hemofiltrate CC chemokine1, CCL14(9–74), which lacked the first eight amino acids ofCCL14, providing evidence for the in vivo expression of atruncated chemokine [56]. Another possible factor contributingto the different observations may be the source of CCL22.Although Pal et al. [10] used HPLC-purified CCL22 isolatedfrom supernatants of CD8(�)T cells, Cota et al. [14] usedsynthetic CCL22, which corresponded to the full-lengthCCL22. We used rCCL22(1–69) and CCL22(3–69; Peprotech)in our HIV-1 infection assays. Our results demonstrating par-tial inhibitory effect of CCL22(3–69) on HIV-1 infection ofPBMCs are in agreement with results previously reported byStruyf et al. [7], who used the same source (Peprotech) ofrCCL22(3–69).
CCL22 treatment of PBMCs had no significant effect onentry, as demonstrated in two different assays that model theentry step of viral infection. In contrast, CCL22 inhibitoryeffect on HIV-1-productive infection was consistently observedwith many PBMC donors examined. These results providedfurther support for previous data that suggested a post-entrymechanism for the observed CCL22 antiviral effect [14].
Previous studies demonstrated that truncated CCL22(3–69)had reduced chemotactic potency for lymphocytes and sug-gested that it was a result of decreased binding to CCR4 [7]. In
Fig. 8. Binding of radiolabeled gp120 to CCR4 is lower than binding to CCR5. The binding analysis was performed in the presence of 100 nM sCD4. Bindingof iodinated gp120 was determined in the presence of increasing concentrations of cold gp120 (A and B) or increasing amounts of CCL22 (C). QFACS analysiswas used to determine expression levels of coreceptors. In this experiment, CCR4 ABS was 13,240 and CCR5 ABS was 13,560, excluding the possibility that theobserved higher gp120 binding to CCR5 was a result of higher expression levels.
Agrawal et al. HIV coreceptor use in PBMCs 1071

contrast, our analysis revealed that CCL22(3–69) was a morepotent inhibitor of CCR4 coreceptor activity, which is consis-tent with increased binding. This discrepancy may be ex-plained by the different assay systems used to evaluate CCL22binding to CCR4. Our assay measures HIV-1 Env-mediatedfusion activity that is dependent on Env binding to rCCR4expressed on murine cells. Blocking this interaction by CCL22resulted in the reduction of cell-fusion activity. In contrast,Struyf et al. [7] demonstrated reduced chemotactic activity forthe truncated CCL22(3–69) using the HUT-78 T cell line(endogenous CCR4) in chemotaxis assay.
To understand the mechanism of coreceptor use, we com-pared CCR4 and CCR5 in terms of their densities on PBMCs,their abilities to colocalize with CD4, and their affinities togp120. We demonstrated that CCL4, a chemokine ligand spe-cific for CCR5, caused 90% blocking activity of HIV-1 entry,indicating the efficient use of CCR5 as a coreceptor. In con-trast, CCL22/MDC had no significant effect on HIV-1 entryinto PBMCs, implicating that CCR4 is not used as an entrycoreceptor. Although CCR4 coreceptor activity was sensitive toCCR4-specific antibodies, treatment of PBMCs with these an-tibodies had no effect on HIV-1-Env fusion, suggesting thatCCR4 is not used as a coreceptor.
Quantitative FACS analysis revealed that although CCR4was expressed on a higher percentage of CD4� lymphocytes,the CCR5 ABS was on the average twofold higher than CCR4ABS. Unless there is a threshold effect, such a difference in thesurface concentrations may not contribute to the dramaticdifference in their coreceptor function. Because the CCR4surface concentration is still relatively high, and it is knownthat at such concentrations CCR5 is a very efficient mediator ofHIV entry, it is unlikely that the difference in the surfaceconcentrations can be a major factor.
Consistent with the published data in the literature [39], weshowed that CCR5 colocalizes with CD4 on activated PBMCs.However, we were unable to show significant CCR4/CD4 co-localization in any of the PBMC samples tested. It is possiblethat inability to associate with CD4 may explain why CCR4 isnot efficiently used by HIV-1. The results are in agreementwith previous literature that demonstrated strong CCR5/CD4colocalization [39] in a HeLa-CD4 cell line transiently express-ing CCR5 and in human macrophages [57]. Although Xiao etal. [39] were able to demonstrate that CD4/CCR5 colocaliza-tion translated into physical association by coimmunoprecipi-tation of CD4 and CCR5, Singer et al. [57] reported thatCD4/CCR5 colocalization in human macrophages and T lym-phocytes did not translate into physical association. Usingimmunogold electron microscopy (IEM), Singer et al. [57]showed that CCR5 and CD4 were clustered and closely ap-posed on micovilli of human macrophages and T cells. Thisdiscrepancy may be explained by the different experimentalprocedures used by Xiao et al. [39] and Singer et al. [57]. It iswell known that IEM is not an appropriate technique to dem-onstrate protein-protein interaction in cases when the proteinconformation is very sensitive to interactions with antibodiesconjugated with gold particles. In particular, chemokine recep-tors have a relatively small extracellular portion (only severalnm in diameter), which is comparable in size with the anti-body-binding region, and are very conformationally flexible.
Therefore, IEM can be used to detect single molecules ofchemokine receptors but appears to be inappropriate to detecttheir interactions with other molecules unless special con-structs are used, e.g., chimeric molecules, etc. In the study bySinger et al. [57], there was no positive control; e.g., in thepresence of gp120, CD4 and CCR5 must associate and bedetected. Singer et al. [57] had to use 13B8.2 CD4 mAb, as itwas the only anti-CD4 antibody that intensely stained CD4-expressing HeLa cells following fixation. Xiao et al. [39] andwe used similar CD4 mAb (OKT4 or T4-4), which gave similarCD4/CCR5 colocalization results using live cells.
Our analysis of gp120 binding to CCR4 or CCR5 indicatedthat the gp120 affinity to the coreceptor might play a criticalrole in coreceptor use. We found that gp120 affinity to CCR5was 104 times higher than its affinity to CCR4. These resultsargue against the use of CCR4 as an entry coreceptor inPBMCs. The weak affinity of gp120 to CCR4 correlated withthe observed weak coreceptor activity of CCR4 in transfectedmurine cells. Despite its lower affinity to CCR4, our resultsdemonstrated for the first time that gp120 binding to CCR4 canoccur in cells expressing CCR4 alone. Therefore, the failure touse CCR4 as a coreceptor in PBMCs may be explained by thepresence of higher affinity coreceptors, coreceptor density, andcolocalization with CD4. We believe that in the case of CCR4versus CCR5, affinity to gp120 and colocalization with CD4seem to determine the selective HIV coreceptor use in PBMCs.
Future studies should extend the analysis described in thispaper to include the other minor coreceptors. Such studies willsignificantly contribute to our knowledge about HIV coreceptoruse and the importance of the minor coreceptors in HIVdisease.
ACKNOWLEDGMENTS
This research is supported by a grant from the AmericanFoundation for AIDS Research (amfAR grant #02709-28-RG).The following reagents were obtained through the AIDS Re-search and Reference Reagent Program, Division of AIDS,NIAID, NIH (Rockville, MD): rhIL-2 from Dr. Maurice Gately,Hoffman-La Roch, Inc. (Nutley, NJ); VP1174 (recombinantvaccinia encoding MN envelope) from Virogenetics Corp.(Troy, NY); HIV-1 89.6 from Dr. Ronald Collman, Universityof Pennsylvania (Philadelphia); and HIV-1 ADA from Dr.Howard Gendelman, University of Nebraska Medical Center(Omaha). The authors thank Dr. Lujin Wu for providing 1G1mAb against human CCR4; Edyta Vieth for excellent technicalassistance; Arun Srivastiva and Johnny He for their commentson the manuscript; and Dr. Rebecca Chan for bleeding in-formed consent blood donors (Indiana University, Indianapolis,IRB Study #9812-05).
REFERENCES
1. Lukacs, N. W., Tekkanat, K. K. (2000) Role of chemokines in asthmaticairway inflammation. Immunol. Rev. 177, 21–30.
2. Baggiolini, M. (2000) Reflections on chemokines. Immunol. Rev. 177,5–7.
1072 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org

3. Cocchi, F., DeVico, A. L., Garzinodemo, A., Arya, S. K., Gallo, R. C.,Lusso, P. (1995) Identification of RANTES, MIP-1�, and MIP-1� as themajor HIV-suppressive factors produced by CD8(�) T cells. Science 270,1811–1815.
4. Bleul, C. C., Farzan, M., Choe, H., Parolin, C., Clark-Lewis, I., Sodroski,J., Springer, T. A. (1996) The lymphocyte chemoattractant SDF-1 is aligand for LESTR/fusin and blocks HIV-1 entry. Nature 382, 829–833.
5. Oberlin, E., Amara, A., Bachelerie, F., Bessia, C., Virelizier, J. L.,Arenzana-Seisdedos, F., Schwartz, O., Heard, J. M., Clark-Lewis, I.,Legler, D. F., Loetscher, M., Baggiolini, M., Moser, B. (1996) The CXCchemokine SDF-1 is the ligand for LESTR/fusin and prevents infection byT-cell-line-adapted HIV-1. Nature 382, 833–835.
6. Godiska, R., Chantry, D., Raport, C. J., Sozzani, S., Allavena, P., Leviten,D., Mantovani, A., Gray, P. W. (1997) Human macrophage-derived che-mokine (MDC), a novel chemoattractant for monocytes, monocyte-deriveddendritic cells, and natural killer cells. J. Exp. Med. 185, 1595–1604.
7. Struyf, S., Proost, P., Sozzani, S., Mantovani, A., Wuyts, A., De Clercq, E.,Schols, D., Van Damme, J. (1998) Enhanced anti-HIV-1 activity andaltered chemotactic potency of NH2-terminally processed macrophage-derived chemokine (MDC) imply an additional MDC receptor. J. Immunol.161, 2672–2675.
8. Tang, H. L., Cyster, J. G. (1999) Chemokine up-regulation and activatedT cell attraction by maturing dendritic cells. Science 284, 819–822.
9. Inngjerdingen, M., Damaj, B., Maghazachi, A. A. (2000) Human NK cellsexpress CC chemokine receptors 4 and 8 and respond to thymus andactivation-regulated chemokine, macrophage-derived chemokine, andI-309. J. Immunol. 164, 4048–4054.
10. Pal, R., Garzino-Demo, A., Markham, P. D., Burns, J., Brown, M., Gallo,R. C., DeVico, A. L. (1997) Inhibition of HIV-1 infection by the beta-chemokine MDC. Science 278, 695–698.
11. Arenzana-Seisdedos, F., Amara, A., Thomas, D., Virelizier, J. L., Baleux,F., Clark-Lewis, I., Legler, D. F., Moser, B., Baggiolini, M. (1998) �-Che-mokine MDC and HIV-1 infection. Science 281, 478–478a.
12. Lee, B., Rucker, J., Doms, R., Tsang, M., Hu, X., Dietz, M., Bailer, R.,Montaner, L., Gerard, C., Sullivan, N., Sodroski, J., Stantchev, T., Broder,C. (1998) �-Chemokine MDC and HIV-1 infection. Science 281, 487–487a.
13. Proost, P., Struyf, S., Schols, D., Opdenakker, G., Sozzani, S., Allavena, P.,Mantovani, A., Augustyns, K., Bal, G., Haemers, A., Lambeir, A. M.,Scharpe, S., Van Damme, J., De Meester, I. (1999) Truncation of macro-phage-derived chemokine by CD26/dipeptidyl-peptidase IV beyond itspredicted cleavage site affects chemotactic activity and CC chemokinereceptor 4 interaction. J. Biol. Chem. 274, 3988–3993.
14. Cota, M., Mengozzi, M., Vicenzi, E., Panina-Bordignon, P., Sinigaglia, F.,Transidico, P., Sozzani, S., Mantovani, A., Poli, G. (2000) Selectiveinhibition of HIV replication in primary macrophages but not T lympho-cytes by macrophage-derived chemokine. Proc. Natl. Acad. Sci. USA 97,9162–9167.
15. Imai, T., Chantry, D., Raport, C. J., Wood, C. L., Nishimura, M., Godiska,R., Yoshie, O., Gray, P. W. (1998) Macrophage-derived chemokine is afunctional ligand for the CC chemokine receptor 4. J. Biol. Chem. 273,1764–1768.
16. Power, C. A., Meyer, A., Nemeth, K., Bacon, K. B., Hoogewerf, A. J.,Proudfoot, A. E., Wells, T. N. (1995) Molecular cloning and functionalexpression of a novel CC chemokine receptor cDNA from a humanbasophilic cell line. J. Biol. Chem. 270, 19495–19500.
17. Power, C. A., Clemetson, J. M., Clemetson, K. J., Wells, T. N. (1995)Chemokine and chemokine receptor mRNA expression in human plate-lets. Cytokine 7, 479–482.
18. Kowalska, M. A., Ratajczak, M. Z., Majka, M., Jin, J., Kunapuli, S., Brass,L., Poncz, M. (2000) Stromal cell-derived factor-1 and macrophage-derived chemokine: 2 chemokines that activate platelets. Blood 96, 50–57.
19. Imai, T., Baba, M., Nishimura, M., Kakizaki, M., Takagi, S., Yoshie, O.(1997) The T cell-directed CC chemokine TARC is a highly specificbiological ligand for CC chemokine receptor 4. J. Biol. Chem. 272,15036–15042.
20. Berger, E. A., Murphy, P. M., Farber, J. M. (1999) Chemokine receptorsas HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu.Rev. Immunol. 17, 657–700.
21. Bogers, W. M., Koornstra, W. H., Dubbes, R. H., ten Haaft, P. J.,Verstrepen, B. E., Jhagjhoorsingh, S. S., Haaksma, A. G., Niphuis, H.,Laman, J. D., Norley, S., Schuitemaker, H., Goudsmit, J., Hunsmann, G.,Heeney, J. L., Wigzell, H. (1998) Characteristics of primary infection of aEuropean human immunodeficiency virus type 1 clade B isolate in chim-panzees. J. Gen. Virol. 79, 2895–2903.
22. Bron, R., Klasse, P. J., Wilkinson, D., Clapham, P. R., Pelchen-Matthews,A., Power, C., Wells, T. N., Kim, J., Peiper, S. C., Hoxie, J. A., Marsh, M.
(1997) Promiscuous use of CC and CXC chemokine receptors in cell-to-cell fusion mediated by a human immunodeficiency virus type 2 envelopeprotein. J. Virol. 71, 8405–8415.
23. McKnight, A., Dittmar, M. T., Moniz-Periera, J., Ariyoshi, K., Reeves,J. D., Hibbitts, S., Whitby, D., Aarons, E., Proudfoot, A. E., Whittle, H.,Clapham, P. R. (1998) A broad range of chemokine receptors are used byprimary isolates of human immunodeficiency virus type 2 as coreceptorswith CD4. J. Virol. 72, 4065–4071.
24. Owen, S. M., Ellenberger, D., Rayfield, M., Wiktor, S., Michel, P., Grieco,M. H., Gao, F., Hahn, B. H., Lal, R. B. (1998) Genetically divergentstrains of human immunodeficiency virus type 2 use multiple coreceptorsfor viral entry. J. Virol. 72, 5425–5432.
25. Deng, H., Liu, R., Ellmeier, W., Choe, S., Unutmaz, D., Burkhart, M., DiMarzio, P., Marmon, S., Sutton, R. E., Hill, C. M., Davis, C. B., Peiper,S. C., Schall, T. J., Littman, D. R., Landau, N. R. (1996) Identification ofa major co-receptor for primary isolates of HIV-1. Nature 381, 661–666.
26. Dragic, T., Litwin, V., Allaway, G. P., Martin, S. R., Huang, Y. X.,Nagashima, K. A., Cayanan, C., Maddon, P. J., Koup, R. A., Moore, J. P.,Paxton, W. A. (1996) HIV-1 entry into CD4(�) cells is mediated by thechemokine receptor CC-CKR-5. Nature 381, 667–673.
27. He, J., Chen, Y., Farzan, M., Choe, H., Ohagen, A., Gartner, S., Busciglio,J., Yang, X., Hofmann, W., Newman, W., Mackay, C. R., Sodroski, J.,Gabuzda, D. (1997) CCR3 and CCR5 are co-receptors for HIV-1 infectionof microglia. Nature 385, 645–649.
28. Andrew, D. P., Ruffing, N., Kim, C. H., Miao, W., Heath, H., Li, Y.,Murphy, K., Campbell, J. J., Butcher, E. C., Wu, L. (2001) C-C chemokinereceptor 4 expression defines a major subset of circulating nonintestinalmemory T cells of both Th1 and Th2 potential. J. Immunol. 166, 103–111.
29. Agrawal, L., Alkhatib, G. (2001) Chemokine receptors: emerging oppor-tunities for new anti-HIV therapies. Expert Opin. Ther. Targets 5, 303–325.
30. Alkhatib, G., Combadiere, C., Broder, C. C., Feng, Y., Kennedy, P. E.,Murphy, P. M., Berger, E. A. (1996) CC CKR5: a RANTES, MIP-1alpha,MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1.Science 272, 1955–1958.
31. Feng, Y., Broder, C. C., Kennedy, P. E., Berger, E. A. (1996) HIV-1 entrycofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272, 872–877.
32. Broder, C. C., Earl, P. L. (1999) Recombinant vaccinia viruses. Design,generation, and isolation. Mol. Biotechnol. 13, 223–245.
33. Alkhatib, G., Liao, F., Berger, E. A., Farber, J. M., Peden, K. W. (1997)A new SIV co-receptor, STRL33. Nature 388, 238.
34. Alkhatib, G., Broder, C. C., Berger, E. A. (1996) Cell type-specific fusioncofactors determine human immunodeficiency virus type 1 tropism forT-cell lines versus primary macrophages. J. Virol. 70, 5487–5494.
35. Nussbaum, O., Broder, C. C., Berger, E. A. (1994) Fusogenic mechanismsof enveloped-virus glycoproteins analyzed by a novel recombinant vac-cinia virus-based assay quantitating cell fusion-dependent reporter geneactivation. J. Virol. 68, 5411–5422.
36. He, J., Landau, N. R. (1995) Use of a novel human immunodeficiencyvirus type 1 reporter virus expressing human placental alkaline phospha-tase to detect an alternative viral receptor. J. Virol. 69, 4587–4592.
37. Sabroe, I., Williams, T. J., Hebert, C. A., Collins, P. D. (1997) Chemoat-tractant cross-desensitization of the human neutrophil IL-8 receptor in-volves receptor internalization and differential receptor subtype regula-tion. J. Immunol. 158, 1361–1369.
38. Lee, B., Sharron, M., Montaner, L. J., Weissman, D., Doms, R. W. (1999)Quantification of CD4, CCR5, and CXCR4 levels on lymphocyte subsets,dendritic cells, and differentially conditioned monocyte-derived macro-phages. Proc. Natl. Acad. Sci. USA 96, 5215–5220.
39. Xiao, X., Wu, L., Stantchev, T. S., Feng, Y. R., Ugolini, S., Chen, H.,Shen, Z., Riley, J. L., Broder, C. C., Sattentau, Q. J., Dimitrov, D. S. (1999)Constitutive cell surface association between CD4 and CCR5. Proc. Natl.Acad. Sci. USA 96, 7496–7501.
40. Lapham, C. K., Ouyang, J., Chandrasekhar, B., Nguyen, N. Y., Dimitrov,D. S., Golding, H. (1996) Evidence for cell-surface association betweenfusin and the CD4-gp120 complex in human cell lines. Science 274,602–605.
41. Bandres, J. C., Wang, Q. F., O’Leary, J., Baleaux, F., Amara, A., Hoxie,J. A., Zolla-Pazner, S., Gorny, M. K. (1998) Human immunodeficiencyvirus (HIV) envelope binds to CXCR4 independently of CD4, and bindingcan be enhanced by interaction with soluble CD4 or by HIV envelopedeglycosylation. J. Virol. 72, 2500–2504.
42. Hill, C. M., Deng, H., Unutmaz, D., Kewalramani, V. N., Bastiani, L.,Gorny, M. K., Zolla-Pazner, S., Littman, D. R. (1997) Envelope glyco-proteins from human immunodeficiency virus types 1 and 2 and simianimmunodeficiency virus can use human CCR5 as a coreceptor for viral
Agrawal et al. HIV coreceptor use in PBMCs 1073

entry and make direct CD4-dependent interactions with this chemokinereceptor. J. Virol. 71, 6296–6304.
43. Hoffman, T. L., LaBranche, C. C., Zhang, W., Canziani, G., Robinson, J.,Chaiken, I., Hoxie, J. A., Doms, R. W. (1999) Stable exposure of thecoreceptor-binding site in a CD4-independent HIV-1 envelope protein.Proc. Natl. Acad. Sci. USA 96, 6359–6364.
44. Edinger, A. L., Blanpain, C., Kunstman, K. J., Wolinsky, S. M., Parmen-tier, M., Doms, R. W. (1999) Functional dissection of CCR5 coreceptorfunction through the use of CD4-independent simian immunodeficiencyvirus strains. J. Virol. 73, 4062–4073.
45. Trkola, A., Dragic, T., Arthos, J., Binley, J. M., Olson, W. C., Allaway,G. P., Cheng-Mayer, C., Robinson, J., Maddon, P. J., Moore, J. P. (1996)CD4-dependent, antibody-sensitive interactions between HIV-1 and itsco- receptor CCR-5. Nature 384, 184–187.
46. Wu, L., Gerard, N. P., Wyatt, R., Choe, H., Parolin, C., Ruffing, N.,Borsetti, A., Cardoso, A. A., Desjardin, E., Newman, W., Gerard, C.,Sodroski, J. (1996) CD4-induced interaction of primary HIV-1 gp120glycoproteins with the chemokine receptor CCR-5. Nature 384, 179–183.
47. Jones, P. L., Korte, T., Blumenthal, R. (1998) Conformational changes incell surface HIV-1 envelope glycoproteins are triggered by cooperationbetween cell surface CD4 and co-receptors. J. Biol. Chem. 273, 404–409.
48. Proudfoot, A. E., Power, C. A., Wells, T. N. (2000) The strategy ofblocking the chemokine system to combat disease. Immunol. Rev. 177,246–256.
49. Simmons, G., Reeves, J. D., Hibbitts, S., Stine, J. T., Gray, P. W.,Proudfoot, A. E., Clapham, P. R. (2000) Co-receptor use by HIV andinhibition of HIV infection by chemokine receptor ligands. Immunol. Rev.177, 112–126.
50. DeVico, A., Pal, R., Markham, P., Garzino-Demo, A., Gallo, R. (1998)�-Chemokine MDC and HIV-1 Infection. Science 281, 487a.
51. Struyf, S., Proost, P., Schols, D., De Clercq, E., Opdenakker, G., Lenaerts,J. P., Detheux, M., Parmentier, M., De Meester, I., Scharpe, S., VanDamme, J. (1999) CD26/dipeptidyl-peptidase IV down-regulates the eo-
sinophil chemotactic potency, but not the anti-HIV activity of humaneotaxin by affecting its interaction with CC chemokine receptor 3. J. Im-munol. 162, 4903–4909.
52. Struyf, S., De Meester, I., Scharpe, S., Lenaerts, J. P., Menten, P., Wang,J. M., Proost, P., Van Damme, J. (1998) Natural truncation of RANTESabolishes signaling through the CC chemokine receptors CCR1 and CCR3,impairs its chemotactic potency and generates a CC chemokine inhibitor.Eur. J. Immunol. 28, 1262–1271.
53. Proost, P., Struyf, S., Schols, D., Durinx, C., Wuyts, A., Lenaerts, J. P., DeClercq, E., De Meester, I., Van Damme, J. (1998) Processing by CD26/dipeptidyl-peptidase IV reduces the chemotactic and anti-HIV-1 activityof stromal-cell-derived factor-1alpha. FEBS Lett. 432, 73–76.
54. Proudfoot, A. E., Buser, R., Borlat, F., Alouani, S., Soler, D., Offord, R. E.,Schroder, J. M., Power, C. A., Wells, T. N. (1999) Amino-terminallymodified RANTES analogues demonstrate differential effects on RANTESreceptors. J. Biol. Chem. 274, 32478–32485.
55. Proost, P., Menten, P., Struyf, S., Schutyser, E., De Meester, I., VanDamme, J. (2000) Cleavage by CD26/dipeptidyl peptidase IV converts thechemokine LD78beta into a most efficient monocyte attractant and CCR1agonist. Blood 96, 1674–1680.
56. Detheux, M., Standker, L., Vakili, J., Munch, J., Forssmann, U.,Adermann, K., Pohlmann, S., Vassart, G., Kirchhoff, F., Parmentier,M., Forssmann, W. G. (2000) Natural proteolytic processing of hemo-filtrate CC chemokine 1 generates a potent CC chemokine receptor(CCR)1 and CCR5 agonist with anti-HIV properties. J. Exp. Med. 192,1501–1508.
57. Singer, I. I., Scott, S., Kawka, D. W., Chin, J., Daugherty, B. L., De-Martino, J. A., DiSalvo, J., Gould, S. L., Lineberger, J. E., Malkowitz, L.,Miller, M. D., Mitnaul, L., Siciliano, S. J., Staruch, M. J., Williams, H. R.,Zweerink, H. J., Springer, M. S. (2001) CCR5, CXCR4, and CD4 areclustered and closely apposed on microvilli of human macrophages and Tcells. J. Virol. 75, 3779–3790.
1074 Journal of Leukocyte Biology Volume 72, November 2002 http://www.jleukbio.org
Related Documents
![Perception of Damaged Self in Plants1[OPEN]...INSENSITIVE1 (BRI1)-ASSOCIATED KINASE1 (BAK1) coreceptor-dependentmanner(Table1;Gullyetal.,2019). We explore potential roles of DAMPs](https://static.cupdf.com/doc/110x72/60d06382ad3bd13b7900cff8/perception-of-damaged-self-in-plants1open-insensitive1-bri1-associated-kinase1.jpg)










