ARTICLES MT5-MMP regulates adult neural stem cell functional quiescence through the cleavage of N-cadherin Eva Porlan 1,2,3 , Beatriz Martí-Prado 1,2 , José Manuel Morante-Redolat 1,2 , Antonella Consiglio 4,5 , Ana C. Delgado 1,2 , Robert Kypta 6 , Carlos López-Otín 7 , Martina Kirstein 1,2 and Isabel Fariñas 1,2,8 The identification of mechanisms that maintain stem cell niche architecture and homeostasis is fundamental to our understanding of tissue renewal and repair. Cell adhesion is a well-characterized mechanism for developmental morphogenetic processes, but its contribution to the dynamic regulation of adult mammalian stem cell niches is still poorly defined. We show that N-cadherin-mediated anchorage of neural stem cells (NSCs) to ependymocytes in the adult murine subependymal zone modulates their quiescence. We further identify MT5-MMP as a membrane-type metalloproteinase responsible for the shedding of the N-cadherin ectodomain in this niche. MT5-MMP is co-expressed with N-cadherin in adult NSCs and ependymocytes and, whereas MT5-MMP-mediated cleavage of N-cadherin is dispensable for the regulation of NSC generation and identity, it is required for proper activation of NSCs under physiological and regenerative conditions. Our results indicate that the proliferative status of stem cells can be dynamically modulated by regulated cleavage of cell adhesion molecules. Mammalian stem cell niches are still poorly characterized owing to the changes required for continuous production of new cells and to the complexity of the interactions between stem cells and their neighbours. In the adult mouse subependymal zone (SEZ), astroglial/radial glial-derived neural stem cells (NSCs) continually generate neuroblasts and oligodendroblasts, through transit- amplifying progenitor (TAP) cells 1,2 . Glial fibrillary acidic protein (GFAP) + NSCs (called B1 cells) express nestin, transcription factor sex determining region Y-box containing gene 2 (Sox2) and inhibitor of DNA binding 1 (Id1) and seem to cycle slowly 1,3–5 . These ‘activated’ B1 cells with relatively long cell cycles may be coexisting with more quiescent/dormant B1 cells, as in the bone marrow 6 . Elimination of dividing cells, such as TAP cells and neuroblasts, by exposure to anti-mitotic β-cytosine-arabinofuranoside (AraC) revealed that surviving B cells are capable of niche reconstitution 7 . Interestingly, a proliferative subpopulation of GFAP + bromodeoxyuridine (BrdU)- label retaining cells (LRCs) exhibiting high levels of the epidermal growth factor receptor (EGFR) were also eliminated and reappeared with the first dividing cells 8 . GFAP + EGFR + cells isolated from the SEZ are efficient in neurosphere formation 8 , and high levels of Notch activity in GFAP + Sox2 + BrdU-LRCs result in EGFR expression and correlate with self-renewal in vitro 9 . These data suggest the existence of a subpopulation of NSCs in a reserved state that can fully regenerate the niche and a pool of primed, albeit slowly cycling, EGF-responsive NSCs with broad developmental potential. However, there is little knowledge about the regulation of cell quiescence/activation by niche interactions, especially when it comes to adhesive interactions. The importance of calcium-dependent homophilic cell–cell adhesion proteins of the cadherin family in morphogenesis is well established 10,11 . Cadherins are also likely candidates to mediate attachment of stem cells to their niche 12 . In Drosophila, cadherin- mediated adhesion but not signalling is necessary for the recruitment of stem cells to their microenvironment 13 . However, the implication of N-cadherin (cadherin 2, Cdh2) in the anchorage to bone marrow and quiescence of mammalian hematopoietic stem cells has been a matter of intense debate 14–16 . Classical E- and N-cadherins are more highly expressed in the brain, and their presence in neurogenic niches has already been described 17–20 . Although the loss of E-cadherin (cadherin 1, Cdh1) results in increased proliferation of subependymal TAP cells 17,21 , the function of N-cadherin in adult neurogenic niches has not been investigated. Here, we find that N-cadherin-mediated adhesion to ependymocytes contributes to the quiescence of B cells in the SEZ niche. We also show that the membrane-type metalloproteinase MT5-MMP (also known as Mmp24), which can 1 Centro de Investigaciones Biomédicas en Red sobre Enfermedades Neurodegenerativas (CIBERNED), 28049 Madrid, Spain. 2 Departamento de Biología Celular, Universidad de Valencia, 46100 Valencia, Spain. 3 Cell Division and Cancer Group, Spanish National Cancer Research Centre (CNIO), 28029 Madrid, Spain. 4 Institut de Biomedicina de la Universitat de Barcelona, (IBUB), 08028 Barcelona, Spain. 5 Department of Biomedical Sciences and Biotechnologies, University of Brescia and National Institute of Neuroscience, 25123 Brescia, Italy. 6 Cic BioGUNE, Parque Tecnológico de Vizcaya, 48160 Derio, Spain. 7 Departamento de Bioquímica y Biología Molecular, Instituto Universitario de Oncología (IUOPA), Universidad de Oviedo, 33006 Oviedo, Spain. 8 Correspondence should be addressed to I.F. (e-mail: [email protected]) Received 19 November 2012; accepted 20 May 2014; published online 22 June 2014; DOI: 10.1038/ncb2993 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 1 © 2014 Macmillan Publishers Limited. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ART ICLES
MT5-MMP regulates adult neural stem cell functionalquiescence through the cleavage of N-cadherinEva Porlan1,2,3, Beatriz Martí-Prado1,2, José Manuel Morante-Redolat1,2, Antonella Consiglio4,5, Ana C. Delgado1,2,Robert Kypta6, Carlos López-Otín7, Martina Kirstein1,2 and Isabel Fariñas1,2,8
The identification of mechanisms that maintain stem cell niche architecture and homeostasis is fundamental to our understandingof tissue renewal and repair. Cell adhesion is a well-characterized mechanism for developmental morphogenetic processes, but itscontribution to the dynamic regulation of adult mammalian stem cell niches is still poorly defined. We show thatN-cadherin-mediated anchorage of neural stem cells (NSCs) to ependymocytes in the adult murine subependymal zone modulatestheir quiescence. We further identify MT5-MMP as a membrane-type metalloproteinase responsible for the shedding of theN-cadherin ectodomain in this niche. MT5-MMP is co-expressed with N-cadherin in adult NSCs and ependymocytes and, whereasMT5-MMP-mediated cleavage of N-cadherin is dispensable for the regulation of NSC generation and identity, it is required forproper activation of NSCs under physiological and regenerative conditions. Our results indicate that the proliferative status ofstem cells can be dynamically modulated by regulated cleavage of cell adhesion molecules.
Mammalian stem cell niches are still poorly characterized owingto the changes required for continuous production of new cellsand to the complexity of the interactions between stem cells andtheir neighbours. In the adult mouse subependymal zone (SEZ),astroglial/radial glial-derived neural stem cells (NSCs) continuallygenerate neuroblasts and oligodendroblasts, through transit-amplifying progenitor (TAP) cells1,2. Glial fibrillary acidic protein(GFAP)+ NSCs (called B1 cells) express nestin, transcription factorsex determining region Y-box containing gene 2 (Sox2) and inhibitorof DNA binding 1 (Id1) and seem to cycle slowly1,3–5. These ‘activated’B1 cells with relatively long cell cycles may be coexisting with morequiescent/dormant B1 cells, as in the bone marrow6. Eliminationof dividing cells, such as TAP cells and neuroblasts, by exposureto anti-mitotic β-cytosine-arabinofuranoside (AraC) revealed thatsurviving B cells are capable of niche reconstitution7. Interestingly, aproliferative subpopulation of GFAP+ bromodeoxyuridine (BrdU)-label retaining cells (LRCs) exhibiting high levels of the epidermalgrowth factor receptor (EGFR) were also eliminated and reappearedwith the first dividing cells8. GFAP+EGFR+ cells isolated from theSEZ are efficient in neurosphere formation8, and high levels of Notchactivity in GFAP+Sox2+ BrdU-LRCs result in EGFR expression andcorrelate with self-renewal in vitro9. These data suggest the existence
of a subpopulation of NSCs in a reserved state that can fully regeneratethe niche and a pool of primed, albeit slowly cycling, EGF-responsiveNSCs with broad developmental potential. However, there is littleknowledge about the regulation of cell quiescence/activation by nicheinteractions, especially when it comes to adhesive interactions.
The importance of calcium-dependent homophilic cell–celladhesion proteins of the cadherin family in morphogenesis is wellestablished10,11. Cadherins are also likely candidates to mediateattachment of stem cells to their niche12. In Drosophila, cadherin-mediated adhesion but not signalling is necessary for the recruitmentof stem cells to their microenvironment13. However, the implicationof N-cadherin (cadherin 2, Cdh2) in the anchorage to bone marrowand quiescence of mammalian hematopoietic stem cells has been amatter of intense debate14–16. Classical E- and N-cadherins are morehighly expressed in the brain, and their presence in neurogenic nicheshas already been described17–20. Although the loss of E-cadherin(cadherin 1, Cdh1) results in increased proliferation of subependymalTAP cells17,21, the function of N-cadherin in adult neurogenic nicheshas not been investigated. Here, we find that N-cadherin-mediatedadhesion to ependymocytes contributes to the quiescence of Bcells in the SEZ niche. We also show that the membrane-typemetalloproteinase MT5-MMP (also known as Mmp24), which can
1Centro de Investigaciones Biomédicas en Red sobre Enfermedades Neurodegenerativas (CIBERNED), 28049 Madrid, Spain. 2Departamento de Biología Celular,Universidad de Valencia, 46100 Valencia, Spain. 3Cell Division and Cancer Group, Spanish National Cancer Research Centre (CNIO), 28029 Madrid, Spain. 4Institutde Biomedicina de la Universitat de Barcelona, (IBUB), 08028 Barcelona, Spain. 5Department of Biomedical Sciences and Biotechnologies, University of Brescia andNational Institute of Neuroscience, 25123 Brescia, Italy. 6Cic BioGUNE, Parque Tecnológico de Vizcaya, 48160 Derio, Spain. 7Departamento de Bioquímica y BiologíaMolecular, Instituto Universitario de Oncología (IUOPA), Universidad de Oviedo, 33006 Oviedo, Spain.8Correspondence should be addressed to I.F. (e-mail: [email protected])
Received 19 November 2012; accepted 20 May 2014; published online 22 June 2014; DOI: 10.1038/ncb2993
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 1
© 2014 Macmillan Publishers Limited. All rights reserved.
ART ICLES
c Ncad GFAP Merge
d GFAP EC-Ncad DAPI
∗
z+2 z+3
∗
z+1
e
∗ee
e
e
ze GFAP γ-tubulin GFAP γ-tubulin Ncad
b DCX NcadDAPI DCX Ncad
a S100BNcad Merge
SEZ
S100BDAPI
Ep
endym
al layer
f GFAP EC-Ncad
e
e e
e
e
e
e
e e
e
e
Inset
Ncad-L
929
L929
Ecad-L
929
g
0.0
0.5
2.0
1.5
1.0
∗∗∗ ∗∗∗NS
Rel
ativ
e nu
mb
er o
f G
FAP
+ c
ells
per
fiel
d
h
IgGEC-Ncad
Rel
ativ
e nu
mb
er o
f ce
lls p
er fi
eld
0.0
0.5
1.5
1.0
0.008∗∗
0.044∗
GFAP+ –S
100B
–
GFAP– –S
100B
+
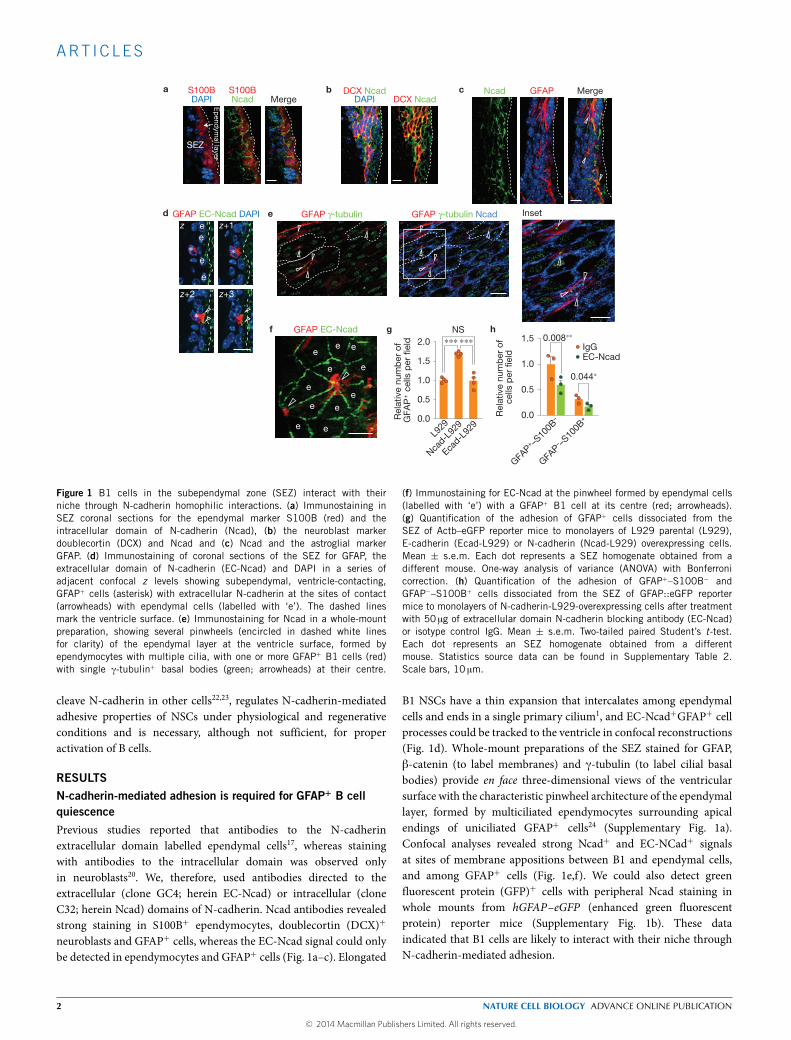
Figure 1 B1 cells in the subependymal zone (SEZ) interact with theirniche through N-cadherin homophilic interactions. (a) Immunostaining inSEZ coronal sections for the ependymal marker S100B (red) and theintracellular domain of N-cadherin (Ncad), (b) the neuroblast markerdoublecortin (DCX) and Ncad and (c) Ncad and the astroglial markerGFAP. (d) Immunostaining of coronal sections of the SEZ for GFAP, theextracellular domain of N-cadherin (EC-Ncad) and DAPI in a series ofadjacent confocal z levels showing subependymal, ventricle-contacting,GFAP+ cells (asterisk) with extracellular N-cadherin at the sites of contact(arrowheads) with ependymal cells (labelled with ‘e’). The dashed linesmark the ventricle surface. (e) Immunostaining for Ncad in a whole-mountpreparation, showing several pinwheels (encircled in dashed white linesfor clarity) of the ependymal layer at the ventricle surface, formed byependymocytes with multiple cilia, with one or more GFAP+ B1 cells (red)with single γ-tubulin+ basal bodies (green; arrowheads) at their centre.
(f) Immunostaining for EC-Ncad at the pinwheel formed by ependymal cells(labelled with ‘e’) with a GFAP+ B1 cell at its centre (red; arrowheads).(g) Quantification of the adhesion of GFAP+ cells dissociated from theSEZ of Actb–eGFP reporter mice to monolayers of L929 parental (L929),E-cadherin (Ecad-L929) or N-cadherin (Ncad-L929) overexpressing cells.Mean ± s.e.m. Each dot represents a SEZ homogenate obtained from adifferent mouse. One-way analysis of variance (ANOVA) with Bonferronicorrection. (h) Quantification of the adhesion of GFAP+–S100B− andGFAP−–S100B+ cells dissociated from the SEZ of GFAP::eGFP reportermice to monolayers of N-cadherin-L929-overexpressing cells after treatmentwith 50 µg of extracellular domain N-cadherin blocking antibody (EC-Ncad)or isotype control IgG. Mean ± s.e.m. Two-tailed paired Student’s t-test.Each dot represents an SEZ homogenate obtained from a differentmouse. Statistics source data can be found in Supplementary Table 2.Scale bars, 10 µm.
cleave N-cadherin in other cells22,23, regulates N-cadherin-mediatedadhesive properties of NSCs under physiological and regenerativeconditions and is necessary, although not sufficient, for properactivation of B cells.
RESULTSN-cadherin-mediated adhesion is required for GFAP+ B cellquiescencePrevious studies reported that antibodies to the N-cadherinextracellular domain labelled ependymal cells17, whereas stainingwith antibodies to the intracellular domain was observed onlyin neuroblasts20. We, therefore, used antibodies directed to theextracellular (clone GC4; herein EC-Ncad) or intracellular (cloneC32; herein Ncad) domains of N-cadherin. Ncad antibodies revealedstrong staining in S100B+ ependymocytes, doublecortin (DCX)+
neuroblasts and GFAP+ cells, whereas the EC-Ncad signal could onlybe detected in ependymocytes and GFAP+ cells (Fig. 1a–c). Elongated
B1 NSCs have a thin expansion that intercalates among ependymalcells and ends in a single primary cilium1, and EC-Ncad+GFAP+ cellprocesses could be tracked to the ventricle in confocal reconstructions(Fig. 1d). Whole-mount preparations of the SEZ stained for GFAP,β-catenin (to label membranes) and γ-tubulin (to label cilial basalbodies) provide en face three-dimensional views of the ventricularsurface with the characteristic pinwheel architecture of the ependymallayer, formed by multiciliated ependymocytes surrounding apicalendings of uniciliated GFAP+ cells24 (Supplementary Fig. 1a).Confocal analyses revealed strong Ncad+ and EC-NCad+ signalsat sites of membrane appositions between B1 and ependymal cells,and among GFAP+ cells (Fig. 1e,f). We could also detect greenfluorescent protein (GFP)+ cells with peripheral Ncad staining inwhole mounts from hGFAP–eGFP (enhanced green fluorescentprotein) reporter mice (Supplementary Fig. 1b). These dataindicated that B1 cells are likely to interact with their niche throughN-cadherin-mediated adhesion.
2 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION
© 2014 Macmillan Publishers Limited. All rights reserved.
ART ICLES
b
0 5 1510
0.0 1.50
Cdh2floxed
GFAP(Cre);Cdh2Δ
0.001∗∗
0.027∗
0.75
Percentage of total cells
Percentage of total cells
c
60 d
ays
pos
t su
rger
y
cc
str
vSEZ
cc
str
v
DAPI GFP
Lent–GFP striatal delivery
SEZ
GFA
P+
Ki6
7+G
FAP
+
d
0 10 20
0 2
Lent–GFP Lent–GFP–DN-CDH
0.005∗∗
4
0.024∗
Percentage of total cells
Percentage of total cells
GFA
P+
Ki6
7+G
FAP
+
Lent–GFP intravent. deliverye
str
cc
vSE
Z
15 d
ays
pos
t su
rger
y cc
str
SE
Z
DAPI GFP
v
Percentage of total cells
0 10 20
0.042∗
0.005∗∗
0 2 4
Lent–GFP Lent–GFP–DN-CDH
f
Percentage of total cells
GFA
P+
Ki6
7+
GFA
P+
Ki6
7 G
FAP
DA
PI
a Cdh2floxed GFAP(Cre);Cdh2Δ
0
50
100
250
LRC
-Brd
U+ c
ells
mm
–2
Corpuscallosum
0.048∗
0
500
1,0001,000
4,000
2,000
SEZ0
45
90
0.016∗
Glomeruli(OB)
0.014∗
Lent–GFP Lent–GFP–DN-CDH
g
Infection
21
days
Euthanasia
BrdU
10
days
Intr
aven
tric
ular
del
iver
y
100
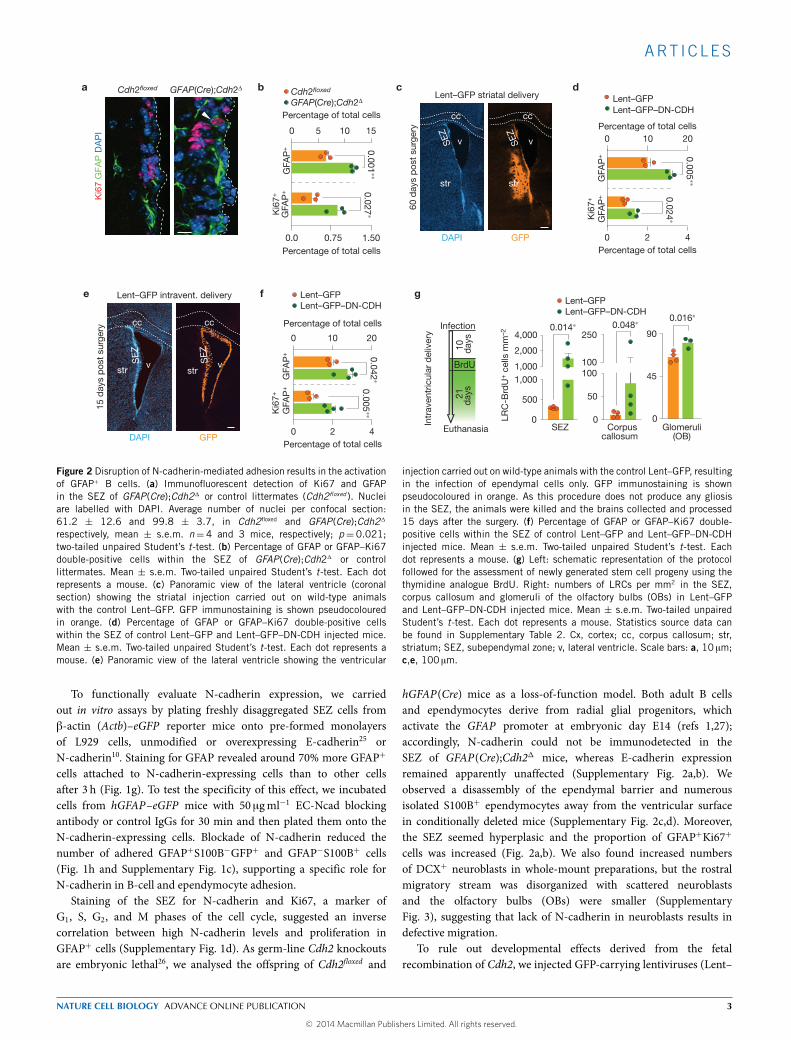
Figure 2 Disruption of N-cadherin-mediated adhesion results in the activationof GFAP+ B cells. (a) Immunofluorescent detection of Ki67 and GFAPin the SEZ of GFAP(Cre);Cdh21 or control littermates (Cdh2floxed ). Nucleiare labelled with DAPI. Average number of nuclei per confocal section:61.2 ± 12.6 and 99.8 ± 3.7, in Cdh2floxed and GFAP(Cre);Cdh21
respectively, mean ± s.e.m. n= 4 and 3 mice, respectively; p= 0.021;two-tailed unpaired Student’s t-test. (b) Percentage of GFAP or GFAP–Ki67double-positive cells within the SEZ of GFAP(Cre);Cdh21 or controllittermates. Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Each dotrepresents a mouse. (c) Panoramic view of the lateral ventricle (coronalsection) showing the striatal injection carried out on wild-type animalswith the control Lent–GFP. GFP immunostaining is shown pseudocolouredin orange. (d) Percentage of GFAP or GFAP–Ki67 double-positive cellswithin the SEZ of control Lent–GFP and Lent–GFP–DN-CDH injected mice.Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Each dot represents amouse. (e) Panoramic view of the lateral ventricle showing the ventricular
injection carried out on wild-type animals with the control Lent–GFP, resultingin the infection of ependymal cells only. GFP immunostaining is shownpseudocoloured in orange. As this procedure does not produce any gliosisin the SEZ, the animals were killed and the brains collected and processed15 days after the surgery. (f) Percentage of GFAP or GFAP–Ki67 double-positive cells within the SEZ of control Lent–GFP and Lent–GFP–DN-CDHinjected mice. Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Eachdot represents a mouse. (g) Left: schematic representation of the protocolfollowed for the assessment of newly generated stem cell progeny using thethymidine analogue BrdU. Right: numbers of LRCs per mm2 in the SEZ,corpus callosum and glomeruli of the olfactory bulbs (OBs) in Lent–GFPand Lent–GFP–DN-CDH injected mice. Mean ± s.e.m. Two-tailed unpairedStudent’s t-test. Each dot represents a mouse. Statistics source data canbe found in Supplementary Table 2. Cx, cortex; cc, corpus callosum; str,striatum; SEZ, subependymal zone; v, lateral ventricle. Scale bars: a, 10 µm;c,e, 100 µm.
To functionally evaluate N-cadherin expression, we carriedout in vitro assays by plating freshly disaggregated SEZ cells fromβ-actin (Actb)–eGFP reporter mice onto pre-formed monolayersof L929 cells, unmodified or overexpressing E-cadherin25 orN-cadherin10. Staining for GFAP revealed around 70% more GFAP+
cells attached to N-cadherin-expressing cells than to other cellsafter 3 h (Fig. 1g). To test the specificity of this effect, we incubatedcells from hGFAP–eGFP mice with 50 µgml−1 EC-Ncad blockingantibody or control IgGs for 30 min and then plated them onto theN-cadherin-expressing cells. Blockade of N-cadherin reduced thenumber of adhered GFAP+S100B−GFP+ and GFAP−S100B+ cells(Fig. 1h and Supplementary Fig. 1c), supporting a specific role forN-cadherin in B-cell and ependymocyte adhesion.
Staining of the SEZ for N-cadherin and Ki67, a marker ofG1, S, G2, and M phases of the cell cycle, suggested an inversecorrelation between high N-cadherin levels and proliferation inGFAP+ cells (Supplementary Fig. 1d). As germ-line Cdh2 knockoutsare embryonic lethal26, we analysed the offspring of Cdh2floxed and
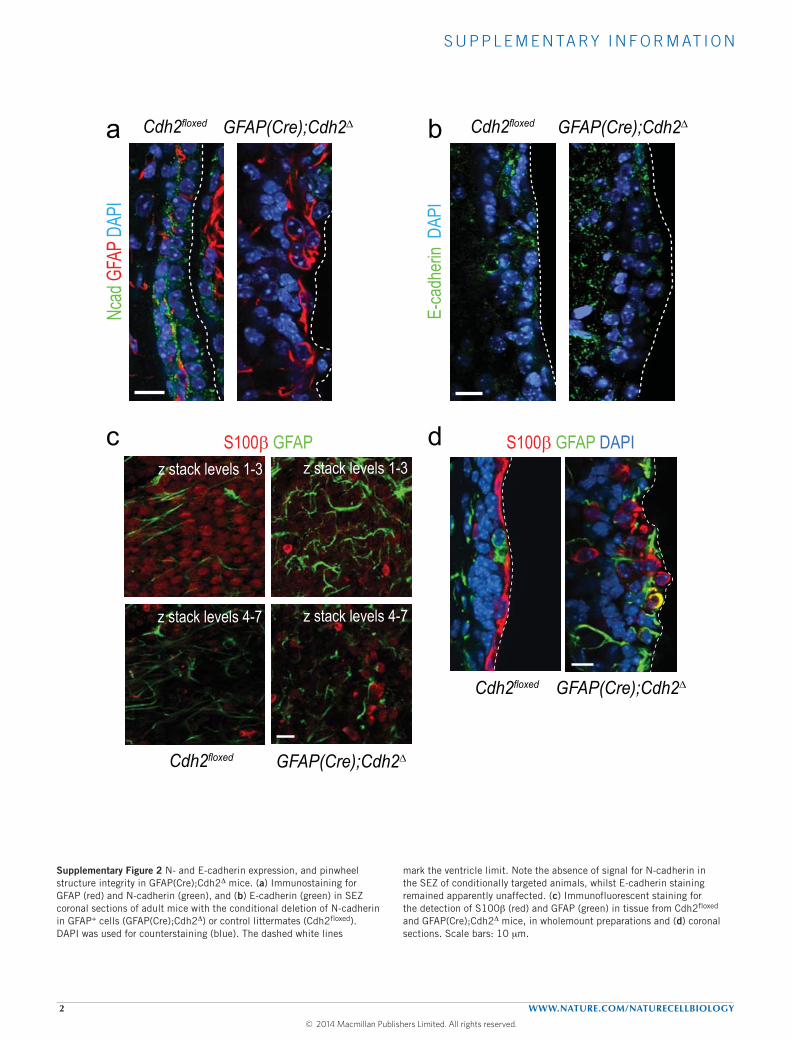
hGFAP(Cre) mice as a loss-of-function model. Both adult B cellsand ependymocytes derive from radial glial progenitors, whichactivate the GFAP promoter at embryonic day E14 (refs 1,27);accordingly, N-cadherin could not be immunodetected in theSEZ of GFAP(Cre);Cdh21 mice, whereas E-cadherin expressionremained apparently unaffected (Supplementary Fig. 2a,b). Weobserved a disassembly of the ependymal barrier and numerousisolated S100B+ ependymocytes away from the ventricular surfacein conditionally deleted mice (Supplementary Fig. 2c,d). Moreover,the SEZ seemed hyperplasic and the proportion of GFAP+Ki67+
cells was increased (Fig. 2a,b). We also found increased numbersof DCX+ neuroblasts in whole-mount preparations, but the rostralmigratory stream was disorganized with scattered neuroblastsand the olfactory bulbs (OBs) were smaller (SupplementaryFig. 3), suggesting that lack of N-cadherin in neuroblasts results indefective migration.
To rule out developmental effects derived from the fetalrecombination of Cdh2, we injected GFP-carrying lentiviruses (Lent–
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 3
© 2014 Macmillan Publishers Limited. All rights reserved.
ART ICLES
GFP; ref. 28) or Lent–GFP–DN-CDH, encoding an extracellularly-truncated version of E-cadherin29, which interfereswith the expressionof all cadherins30, into the SEZ (ref. 28). Lent–GFP–DN-CDH-injectedmice exhibited higher proportions of GFAP+ cells and GFAP+Ki67+
cells (Fig. 2c,d), indicating again that loss of N-cadherin promotesB-cell proliferation. We next designed a procedure to interfere withN-cadherin function only in ependymal cells. Injection of Lent–GFP–DN-CDH into the lateral ventricle resulted in the transduction ofmostependymocytes, a noticeable reduction in EC-N-cadherin stainingin transduced S100B−GFP+ cells, and disassembly of the ependymallayer (Fig. 2e and Supplementary Fig. 4a–d). Although we foundhardly any GFP+GFAP+ cells21, transduction of ependymocytes withLent–GFP–DN-CDH, leading to disruption of the homotypic N-cadherin-mediated adhesion, resulted again in increased proportionsof GFAP+ and GFAP+Ki67+ cells (Fig. 2f and SupplementaryFig. 4b,d,e). In Lent–GFP–DN-CDH-injected mice treated with BrdU10 days after the ventricular delivery and 21 days before euthanasia,we found increased numbers of BrdU+ LRCs in the SEZ, corpuscallosum and OB glomerular layer (Fig. 2g). Thus, perturbation ofN-cadherin in the SEZ results in higher numbers of proliferating Bcells and enhanced neurogenic/oligodendrogenic output, suggestingthat N-cadherin-mediated adhesion in this niche may be partof the mechanism that controls quiescence of GFAP+ B cells.Interestingly, we observed newly generated S100B+ ependymocytesin Lent–GFP–DN-CDH-injected animals (Supplementary Fig. 4f–h),suggesting potential repair mechanisms when the ependymal barrieris compromised.
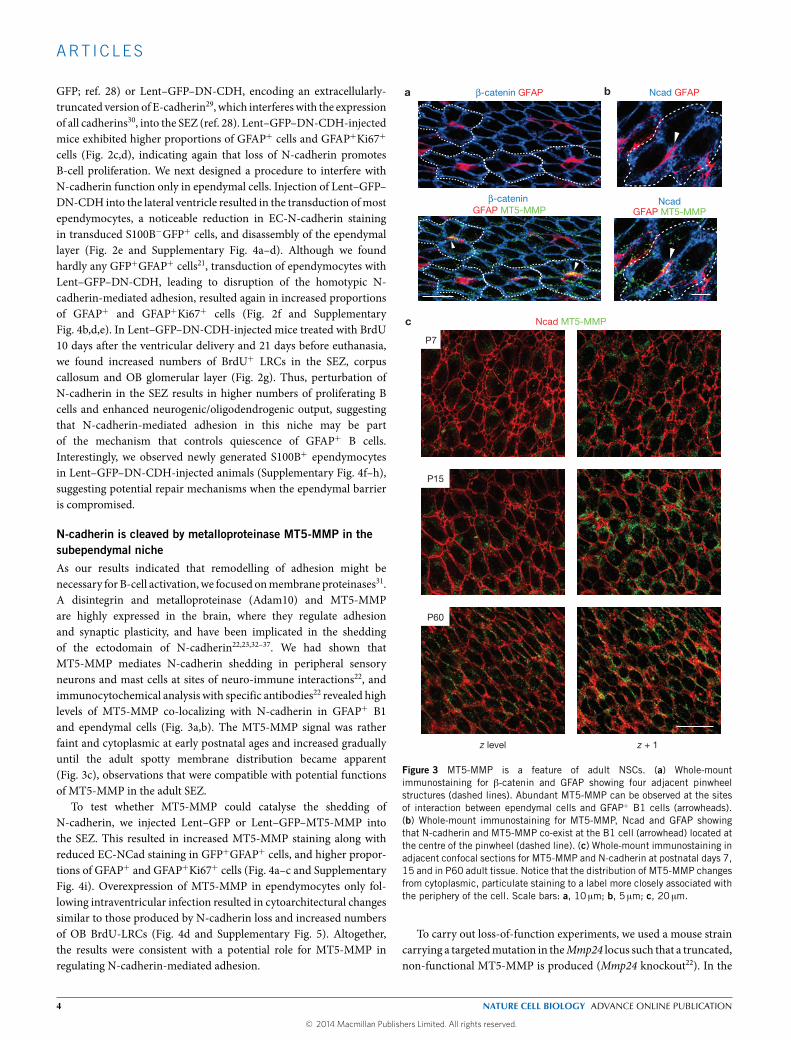
N-cadherin is cleaved by metalloproteinase MT5-MMP in thesubependymal nicheAs our results indicated that remodelling of adhesion might benecessary for B-cell activation,we focused onmembrane proteinases31.A disintegrin and metalloproteinase (Adam10) and MT5-MMPare highly expressed in the brain, where they regulate adhesionand synaptic plasticity, and have been implicated in the sheddingof the ectodomain of N-cadherin22,23,32–37. We had shown thatMT5-MMP mediates N-cadherin shedding in peripheral sensoryneurons and mast cells at sites of neuro-immune interactions22, andimmunocytochemical analysis with specific antibodies22 revealed highlevels of MT5-MMP co-localizing with N-cadherin in GFAP+ B1and ependymal cells (Fig. 3a,b). The MT5-MMP signal was ratherfaint and cytoplasmic at early postnatal ages and increased graduallyuntil the adult spotty membrane distribution became apparent(Fig. 3c), observations that were compatible with potential functionsof MT5-MMP in the adult SEZ.
To test whether MT5-MMP could catalyse the shedding ofN-cadherin, we injected Lent–GFP or Lent–GFP–MT5-MMP intothe SEZ. This resulted in increased MT5-MMP staining along withreduced EC-NCad staining in GFP+GFAP+ cells, and higher propor-tions of GFAP+ and GFAP+Ki67+ cells (Fig. 4a–c and SupplementaryFig. 4i). Overexpression of MT5-MMP in ependymocytes only fol-lowing intraventricular infection resulted in cytoarchitectural changessimilar to those produced by N-cadherin loss and increased numbersof OB BrdU-LRCs (Fig. 4d and Supplementary Fig. 5). Altogether,the results were consistent with a potential role for MT5-MMP inregulating N-cadherin-mediated adhesion.
a β-catenin GFAP
NcadGFAP MT5-MMP
Ncad GFAPb
c Ncad MT5-MMP
z level z + 1
P7
P15
P60
β-catenin GFAP MT5-MMP
Figure 3 MT5-MMP is a feature of adult NSCs. (a) Whole-mountimmunostaining for β-catenin and GFAP showing four adjacent pinwheelstructures (dashed lines). Abundant MT5-MMP can be observed at the sitesof interaction between ependymal cells and GFAP+ B1 cells (arrowheads).(b) Whole-mount immunostaining for MT5-MMP, Ncad and GFAP showingthat N-cadherin and MT5-MMP co-exist at the B1 cell (arrowhead) located atthe centre of the pinwheel (dashed line). (c) Whole-mount immunostaining inadjacent confocal sections for MT5-MMP and N-cadherin at postnatal days 7,15 and in P60 adult tissue. Notice that the distribution of MT5-MMP changesfrom cytoplasmic, particulate staining to a label more closely associated withthe periphery of the cell. Scale bars: a, 10 µm; b, 5 µm; c, 20 µm.
To carry out loss-of-function experiments, we used a mouse straincarrying a targetedmutation in theMmp24 locus such that a truncated,non-functional MT5-MMP is produced (Mmp24 knockout22). In the
4 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION
© 2014 Macmillan Publishers Limited. All rights reserved.

ART ICLES
Lent
–GFP
–MT5
-MM
P
b EC-Ncad GFAP EC-Ncad GFP
Lent
–GFP
a
cc
str
cx
cc
str
v
cx
SE
Z
DAPI
SE
Z
GFP
v
Lent
–GFP
str
iata
l del
iver
y
60 days Lent–GFP Lent–GFP–MT5-MMP
c
Per
cent
age
of t
otal
cel
ls
0
10
25
0.0
1.0
2.0
15
20
5
0.014∗ 0.009∗∗
GFAP+
Ki67+GFAP+
0
45
90
90
135
1800.018∗
LRC
-Brd
U+ c
ells
mm
–2
Lent–GFPLent–GFP–MT5-MMP
d
Infection
21 day
s
Euthanasia
BrdU
10
day
s
Intr
aven
tric
ular
del
iver
y
Glomeruli(OB)
Figure 4 MT5-MMP activity sheds N-cadherin ectodomain in vivo andmodulates NSC proliferation. (a) Intrastriatal delivery of Lent–GFP orLent–GFP–MT5-MMP. Upper panel: general view of the infected area. Lowerpanel: confocal micrographs for GFP immunostaining (pseudocoloured inorange) showing the infected region after 60 days. (b) Immunostainingfor extracellular N-cadherin (EC-Ncad), GFAP and GFP (pseudocoloured) incontrol (Lent–GFP) or Lent–GFP–MT5-MMP transduced SEZs. Arrowheadspoint to GFAP–EC-Ncad–GFP triple-positive cells, and empty arrowheadspoint to GFAP–GFP double-positive, EC-Ncad-negative cells. Nuclei arecounterstained with DAPI, and the dashed white lines mark the ventricle limit.(c) Percentages of GFAP-positive and GFAP–Ki67 double-positive cells within
the SEZ of wild-type mice injected with Lent–GFP or Lent–GFP–MT5-MMP.Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Each dot represents amouse. (d) Left: schematic representation of the experimental design followedfor the evaluation of the number of LRCs, BrdU-positive newborn neurons, inthe olfactory bulb (OB) after the expression of MT5-MMP in the ependymallayer of the lateral ventricle of wild-type mice. Right: number of BrdU-positiveLRCs scored in the glomeruli of wild-type mice infected in the lateral ventriclewith Lent–GFP or Lent–GFP–MT5-MMP. Mean ± s.e.m. Two-tailed unpairedStudent’s t-test. Each dot represents a mouse. Statistics source data can befound in Supplementary Table 2. Str, striatum; cx, cortex; v, lateral ventricle;cc, corpus callosum. Scale bars: a, 100 µm; b, 10 µm.
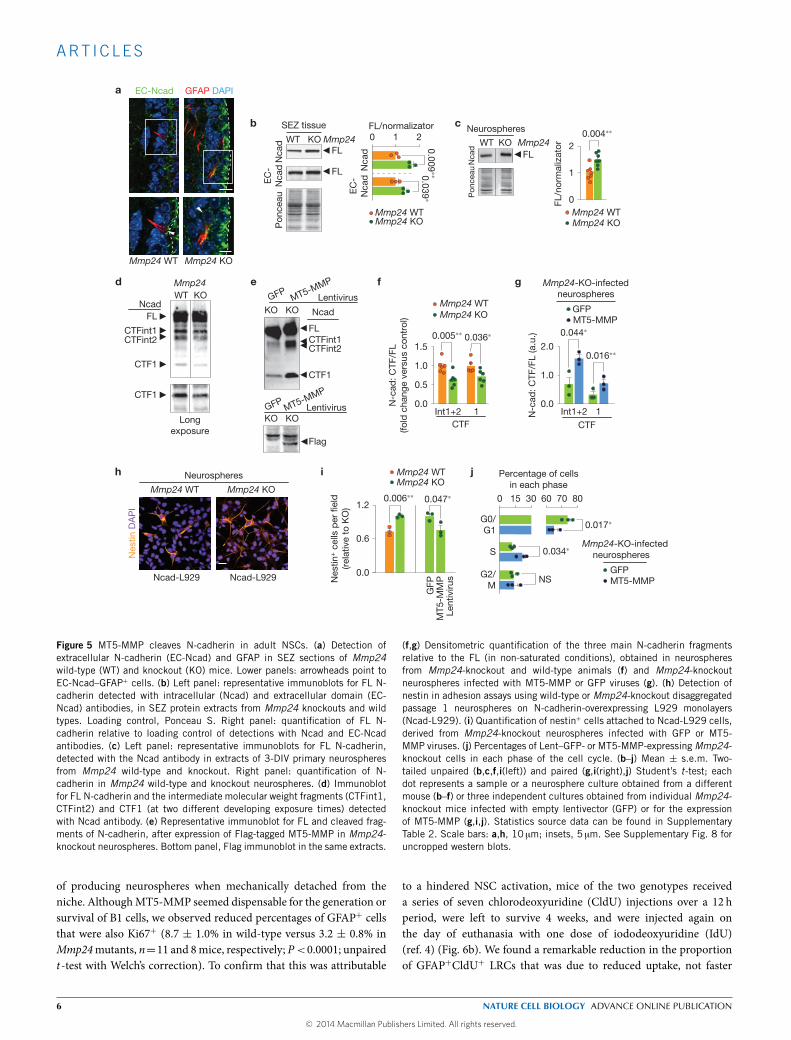
mutants, we could observe a more intense and homogeneous signalfor extracellular N-cadherin in the SEZ and increased levels of full-length (FL)N-cadherin in SEZ and neurosphere homogenates withoutchanges in Cdh2messenger RNA levels (Fig. 5a–c and SupplementaryFig. 6a). Classical cadherins found at adherens junctions are linkedto a ring-like actin cytoskeleton by the β-catenin, α-catenin andplakoglobin proteins38. Because β-catenin can be at the junctions orfree in the cytoplasm in response to Wnt (ref. 38), N-cadherin can actas a trap for β-catenin35, and we found increased levels of β-cateninin Mmp24-knockout neurospheres (Supplementary Fig. 6b), furthersuggesting a deficient processing in the absence of MT5-MMP.
Ectodomain cleavage of N-cadherin leads to the generation ofa soluble 90 kDa amino-terminal soluble fragment and a 45 kDamembrane-bound carboxy-terminal fragment 1 (CTF1), which isfurther cleaved by γ-secretase into a 35 kDa CTF2 (refs 22,23). Inmutant neurospheres we found reduced levels of the CTF1 fragmentand of two other fragments of higher molecular weight (CTFint1 and2) that could be restored to normal levels following lentiviral deliveryof MT5-MMP (Fig. 5d–g). In contrast to reported effects of Adam10on the regulated cleavage of the Notch receptor, which also requiresregulated proteolysis31, modulation of the levels of MT5-MMP inneurospheres did not alter the level of Notch intracellular fragments(Supplementary Fig. 6c,d).
Because cells lacking MT5-MMP exhibited higher levels of FLN-cadherin, we next evaluated their adhesive potential. Nestin+ cellsfrom the mutant SEZ adhered more efficiently than wild types to N-cadherin-L929 monolayers. This increase could be reduced by ectopicexpression of MT5-MMP or by pre-incubation with blocking EC-Ncad antibodies (Fig. 5h,i). Interestingly, mutant neurospheres thathad been transduced withMT5-MMP contained lower proportions ofcells in G0/G1 together with increased percentages of cells in S phase(Fig. 5j). These results demonstrated thatMT5-MMP is a physiologicalregulator of the shedding of N-cadherin ectodomain and of B-celladhesion, and suggested again that N-cadherin processing regulatesthe cycling of NSCs.
Functional quiescence in the SEZ is regulated by MT5-MMPThe SEZ of Mmp24-knockout mice exhibited an apparently normaldistribution of B1 cells in whole mounts (Fig. 6a) and containednormal proportions of GFAP+ cells (relative to 4′,6-diamidino-2-phenylindole (DAPI): 7.94 ± 1.01%, n=10, versus a wild-type valueof 8.17 ± 0.85%, n=13; SEZs of the two genotypes did not differ insize) and GFAP+–Id1high cells (50.7± 2.05%, n=5, versus a wild-typevalue of 49.9 ± 5.43%, n= 6). Moreover, Mmp24-null mice yieldedthe same number of neurospheres as wild-type mice (SupplementaryFig. 6e), indicating that mutant GFAP+ B cells were still capable
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 5
© 2014 Macmillan Publishers Limited. All rights reserved.
ART ICLES
a EC-Ncad GFAP DAPI
Mmp24 WT Mmp24 KO
GFP
MT5
-MM
P
i
Nes
tin+ c
ells
per
fiel
d(r
elat
ive
to K
O)
0.006∗∗ 0.047∗
0.0
0.6
1.2
Lent
iviru
s
Mmp24 WTMmp24 KO
h
Ncad-L929
Nes
tin D
AP
I
Neurospheres
Mmp24 WT Mmp24 KO
Ncad-L929
f
1.5
0.0
0.5
1.0
0.036∗0.005∗∗
Int1+2
Mmp24 WTMmp24 KO
CTF1
N-c
ad: C
TF/F
L(fo
ld c
hang
e ve
rsus
con
trol
)
0.016∗∗
0.044∗
N-c
ad: C
TF/F
L (a
.u.)
2.0
0.0
1.0
g
Int1+2 1
GFPMT5-MMP
Mmp24-KO-infectedneurospheres
CTF
j
G0/G1
S
0 60
G2/M
80
Percentage of cellsin each phase
0.017∗
0.034∗
NS
30 7015
Mmp24-KO-infectedneurospheres
GFPMT5-MMP
d Mmp24WT
Longexposure
CTF1
Ncad
CTFint1CTFint2
CTF1
FL
KO
b
FL
FL
SEZ tissue
Nca
dE
C-
Nca
d
Mmp24WT KO
Pon
ceau
FL/normalizator0 1 2
Nca
dE
C-
Nca
d0.009
∗∗0.039∗
Mmp24 WTMmp24 KO
c
Pon
ceau
FL
Nca
d 2
0
1
0.004∗∗Neurospheres
Mmp24WT
FL/n
orm
aliz
ator
Mmp24 WTMmp24 KO
KO
e
FL
Flag
CTF1
GFPMT5-MMP
CTFint2CTFint1
KO KOLentivirus
GFPMT5-MMP
LentivirusKO KO
Ncad
Figure 5 MT5-MMP cleaves N-cadherin in adult NSCs. (a) Detection ofextracellular N-cadherin (EC-Ncad) and GFAP in SEZ sections of Mmp24wild-type (WT) and knockout (KO) mice. Lower panels: arrowheads point toEC-Ncad–GFAP+ cells. (b) Left panel: representative immunoblots for FL N-cadherin detected with intracellular (Ncad) and extracellular domain (EC-Ncad) antibodies, in SEZ protein extracts from Mmp24 knockouts and wildtypes. Loading control, Ponceau S. Right panel: quantification of FL N-cadherin relative to loading control of detections with Ncad and EC-Ncadantibodies. (c) Left panel: representative immunoblots for FL N-cadherin,detected with the Ncad antibody in extracts of 3-DIV primary neurospheresfrom Mmp24 wild-type and knockout. Right panel: quantification of N-cadherin in Mmp24 wild-type and knockout neurospheres. (d) Immunoblotfor FL N-cadherin and the intermediate molecular weight fragments (CTFint1,CTFint2) and CTF1 (at two different developing exposure times) detectedwith Ncad antibody. (e) Representative immunoblot for FL and cleaved frag-ments of N-cadherin, after expression of Flag-tagged MT5-MMP in Mmp24-knockout neurospheres. Bottom panel, Flag immunoblot in the same extracts.
(f,g) Densitometric quantification of the three main N-cadherin fragmentsrelative to the FL (in non-saturated conditions), obtained in neurospheresfrom Mmp24-knockout and wild-type animals (f) and Mmp24-knockoutneurospheres infected with MT5-MMP or GFP viruses (g). (h) Detection ofnestin in adhesion assays using wild-type or Mmp24-knockout disaggregatedpassage 1 neurospheres on N-cadherin-overexpressing L929 monolayers(Ncad-L929). (i) Quantification of nestin+ cells attached to Ncad-L929 cells,derived from Mmp24-knockout neurospheres infected with GFP or MT5-MMP viruses. (j) Percentages of Lent–GFP- or MT5-MMP-expressingMmp24-knockout cells in each phase of the cell cycle. (b–j) Mean ± s.e.m. Two-tailed unpaired (b,c,f,i(left)) and paired (g,i(right),j) Student’s t-test; eachdot represents a sample or a neurosphere culture obtained from a differentmouse (b–f) or three independent cultures obtained from individual Mmp24-knockout mice infected with empty lentivector (GFP) or for the expressionof MT5-MMP (g,i,j). Statistics source data can be found in SupplementaryTable 2. Scale bars: a,h, 10 µm; insets, 5 µm. See Supplementary Fig. 8 foruncropped western blots.
of producing neurospheres when mechanically detached from theniche. AlthoughMT5-MMP seemed dispensable for the generation orsurvival of B1 cells, we observed reduced percentages of GFAP+ cellsthat were also Ki67+ (8.7 ± 1.0% in wild-type versus 3.2 ± 0.8% inMmp24mutants, n=11 and 8mice, respectively; P<0.0001; unpairedt-test with Welch’s correction). To confirm that this was attributable
to a hindered NSC activation, mice of the two genotypes receiveda series of seven chlorodeoxyuridine (CldU) injections over a 12 hperiod, were left to survive 4 weeks, and were injected again onthe day of euthanasia with one dose of iododeoxyuridine (IdU)(ref. 4) (Fig. 6b). We found a remarkable reduction in the proportionof GFAP+CldU+ LRCs that was due to reduced uptake, not faster
6 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION
© 2014 Macmillan Publishers Limited. All rights reserved.

ART ICLES
0
6
CldU+-LRC
Per
cent
age
ofG
FAP
+ c
ells
c
3
0.005∗∗
Mmp24 WTMmp24 KO
Per
cent
age
of
Cld
U+LR
C
0
30
60
d
IdU+
0.034∗
Mmp24 WTMmp24 KO
h
0
20
Ki67+
Per
cent
age
ofG
FAP
+ c
ells
10
<0.001∗∗∗<0.001∗∗∗
Naive lgG EC-Ncad
Mmp24 WTMmp24 KO
Ki6
7 M
CM
2 G
FAP
DA
PI
e Mmp24 WT Mmp24 KO
b
Cld
U Id
U G
FAP
DA
PI
IdU CldUIdUGFAP CldU GFAP
1 h
IdUCldU
12 h
28 days Eut
h.Mmp24WT
Mmp24KO
a Mmp24 WT Mmp24 KO
GFA
P γ
-tub
ulin
Mm
p24
KO
GFA
P γ
-tub
ulin
GFA
P γ
-tub
ulin
Mm
p24
WT
Percentage of GFAP+
MCM2+ cells0 10050
f
Ki67+ 0.003∗∗
Mmp24 WTMmp24 KO
Merge Merge
Ki67 MCM2 GFAPMmp24 WT Mmp24 KO
Lateral ventricles
Osmoticmini- pump
glgG/EC-Nad
intraventricular infusion
Figure 6 MT5-MMP activity is necessary for the activation of adultNSCs. (a) Z -stack confocal micrographs of whole-mount immunostainingfor GFAP and γ-tubulin. Pinwheel structures are encircled within whitedashed lines for clarity. Insets (lower panels) show higher-magnificationexamples of pinwheels at the ventricle level in Mmp24 knockouts and wild-type controls. Arrowheads point at B cells. (b) Upper panel: schematicrepresentation of the thymidine-analogue administration protocol followedto label slow-dividing (CldU-positive), fast-dividing (IdU-positive) and slow-dividing cells re-entering cell cycle (double-positive cells). Immunostainingfor GFAP (pseudocoloured), CldU-positive LRCs (green) and IdU-positivehighly proliferative cells (red) in the SEZ of wild-type and Mmp24-knockoutanimals. Lower panels are higher-magnification images of the area comprisedby the white rectangle in the upper left panel. (c,d) Percentages of CldU-LRCswithin the GFAP+ population (c) and of IdU-positive cells within the CldU-LRC population (d) in Mmp24-knockout and wild-type animals. Mean ±s.e.m. Two-tailed unpaired Student’s t-test. Dots represent independent mice.
(e) Immunostaining for Ki67, MCM2 and GFAP in the SEZ of wild-type andMmp24-knockout animals. Bottom panels are higher-magnification imagesof the areas comprised by the white rectangles in the top panels. The whitearrowhead indicates a triple-positive cell, and the empty arrowhead pointsto a GFAP–MCM2 double-positive cell, negative for Ki67. (f) Percentagesof Ki67-positive cells within the population of GFAP–MCM2 double-positivecells. Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Dots representindependent mice. (g) Schematic representation of the experimental settingfor the 7 day intraventricular infusion of the blocking antibody for theextracellular domain of N-cadherin (EC-Ncad) or isotype control IgG.(h) Percentages of Ki67-positive cells within the GFAP population in the SEZof naive wild-type and Mmp24-knockout, and of intraventricularly infusedMmp24-knockout mice. Mean ± s.e.m. One-way ANOVA with Bonferronicorrection. Dots represent independent mice. Statistics source data canbe found in Supplementary Table 2. DAPI was used for counterstaining.Scale bars: a, 15 µm; b,e, 10 µm; insets in a,e, 5 µm.
dilution of the nucleotide, as fewer CldU+ LRCs incorporatedIdU in mutant mice (Fig. 6c,d). Reduced uptake was not theresult of premature exit from the cell cycle and terminal astrocyticdifferentiation, as we found similar percentages of CldU+ LRCsexpressing the marker of differentiated astrocytes S100B (ref. 39)(Supplementary Fig. 6f,g). Minichromosome maintenance proteinMCM2 is one licensing factor of the pre-replication complex presentin all replication-competent cells, including quiescent ones4.We foundreduced percentages of GFAP+ cells that were MCM2+ (7.2 ± 0.8%in wild type versus 4.6 ± 0.6% in Mmp24 knockout, n=4; P=0.031,two-tailed unpaired t-test) in the absence ofMT5-MMP (Fig. 6e), and,although over 90% of the GFAP+MCM2+ cells were also Ki67+ inwild-type mice, the percentage of triple-positive cells was reduced inthe mutants (Fig. 6e,f). As MCM2 is expressed throughout the cell
cycle whereas Ki67 is predominantly expressed in late G1, S, G2 andM phases, more GFAP+ B cells seem to be retained in G0/early G1
in the absence of MT5-MMP. These data suggested that MT5-MMPdefective B cells progress more slowly through the cell cycle.
To evaluate the involvement of N-cadherin in the Mmp24phenotype, we infused the EC-Ncad blocking antibodies into thelateral ventricle of mutant mice using osmotic mini-pumps. We foundthat the reduction in GFAP+Ki67+ cells was rescued by EC-Ncad,but not non-relevant antibody infusion (Fig. 6g,h), indicating thatincreased B-cell quiescence in the absence of a functional MT5-MMPwas the result of increased N-cadherin-mediated attachment.
To test MT5-MMP role during regeneration, we infused AraCinto the surface of the brain for 6 days. This ablates rapidly dividingTAP cells and neuroblasts, but spares ependymocytes and the most
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 7
© 2014 Macmillan Publishers Limited. All rights reserved.

ART ICLES
quiescent GFAP+ B cells, which, on withdrawal of the drug, becomeactivated and enter the cell cycle within the next 12–24 h, leadingto rapid regeneration of the SEZ (refs 5,7,8). Immediately afterpump removal (time t = 0 h), surviving GFAP+ B cells showedmore conspicuous and homogeneous N-cadherin staining, but thesignal became dimmer after 12 and 24 h (Supplementary Fig. 7a,b),suggesting again an inverse correlation between the levels of FLN-cadherin and quiescence in B cells. In addition, although mostGFAP+Ki67+ cells had been eliminated by the infusion in wild-typeanimals (decreased from 11.1 ± 1.4 in saline to 2.4 ± 0.7 in AraC-infused mice), most of the Ki67+GFAP+ cells in mutant animals hadsurvived the treatment (Fig. 7a,b), another indication that activated Bcells in MT5-MMP-deficient mice progress more slowly through thecell cycle, potentially by lengthening G1.
Quantitation ofGFAP+ cells that were alsoKi67+ at different times,relative to those present at t = 0, showed that normal repopulationin wild-type animals involves an initial sixfold boost in GFAP+ cellproliferation at t=12 h, that decreases at 24 and 72 h by approximatelytwo- to threefold. In Mmp24-knockout mice, however, the initialactivation was less than half, and, even though it increased slightlyat 24 and 72 h, never reached the wild-type peak activation (Fig. 7c).In agreement with this result, we observed reduced generation ofMash1+ TAP cells inMmp24mutants (Fig. 7d,e), suggesting deficientactivation of quiescent B cells and slower progression through thecell cycle. In conclusion, our data indicate that N-cadherin-mediatedattachment of B cells to their niche modulates their functionalquiescence and that MT5-MMP is part of the mechanism.
DISCUSSIONAdult NSCs lie in an epithelial stem cell niche: their apical domaininteracts with ependymocytes through adhesive junctions17 and theirbasal process adheres to irrigating vasculature through α6β1 integrin-mediated binding to laminin19. This anchorage seems necessaryfor NSC lodging and quiescence19. Moreover, Id transcriptionfactors, which regulate NSC stemness-associated expression, alsomodulate residency in the vascular extracellular matrix niche throughrepression of the gene Rap1GAP (ref. 40). Therefore, molecularprograms involved in stemness regulation are likely to control bothstem cell intrinsic properties and restricted positioning. Recentevidence supports an emerging role also for cell–cell adhesiveinteractions in the organization and modulation of mammalianstem cell reservoirs, although the players involved are still a matterof investigation and debate12,16. Our results demonstrate that N-cadherin-dependent homotypic interactions with ependymal cellsregulate NSC quiescence. More importantly, we provide evidencethat MT5-MMP is required for the proteolytic processing of N-cadherin and the cycling of B1 cells under physiological andregenerative conditions.
Detection of N-cadherin has been controversial in certaintissues, casting doubt on the specificity of some antibodies16. Weobserve staining of ependymocytes and B1 cells with two differentantibodies, in agreement with other reports17. Our studies reveal thatextracellularly directed antibodies do not provide a detectable signalfor N-cadherin in neuroblasts, whereas high levels of N-cadherin inneuroblasts are detected with an antibody directed to intracellularepitopes18,20. This may be related to antigen accessibility and/or
epitope modification, but migratory defects in the rostral migratorystream when N-cadherin is eliminated indicates a functional rolein neuroblasts. Reduction of cadherin function with the dominant-negative construct in B cells could be interpreted as the result ofinterference with E-cadherin17. However, we obtained similar resultsinCdh2 conditional mutants, and it has been reported that E-cadherinloss results in increased proliferative activity of TAP cells, withoutapparent changes in LRCs (ref. 17).
MT5-MMP is a transmembrane protein with an N-terminalextracellular region containing the catalytic domain, a singletransmembrane domain and a cytosolic short C-terminal tail requiredfor its localization36,41,42. Its expression is highly regulated duringbrain development and remains elevated in the adult hippocampus,OB and cerebellum32. Although these regions exhibit continuedsynaptic plasticity, and MT5-MMP seems to be implicated, they alsoexhibit postnatal neurogenesis. Our genetic data indicate that MT5-MMP is necessary, but not sufficient, for N-cadherin ectodomainshedding in the SEZ. Although the lack of this metalloproteinaseresults in defective N-cadherin processing and reduction of B-cellproliferation, OB neurogenesis takes place. Moreover, regenerationfollowing treatment with AraC is delayed, but not abrogated, by thelack of MT5-MMP, suggesting the involvement of other enzymes,such as Adam10. Nevertheless, Adam10 has also been associated withproteolytic activation of neuregulin and Notch31,35,43, whereas wesee no effect of MT5-MMP in the generation of Notch intracellularfragments in the SEZ, suggesting that these metalloproteinases differin their selectivity of action and may regulate different processes.
It is unclear how proteolytic processing and cell detachment couldregulate the cycling of the NSCs. It is possible that perturbation oftissue integrity leads to hyperproliferation, as we observed generationof new ependymal cells, something that is never observed in youngmice unless the ependymal layer is mildly lesioned44. In addition,as B1 cells seem to have restricted access to activating moleculespresent in the cerebrospinal fluid through the selective localizationof receptors at the cilial membrane, we cannot formally discardincreased exposure of B1 cells to pro-mitogenic molecules45. However,lack of MT5-MMP does not result in tissue disorganization, butproduces the opposite result and decreases proliferation, suggestingthat N-cadherin-mediated adhesion could dynamically regulate theactivation of B cells. This could be the result of a permissivecontribution of N-cadherin cleavage to cell cycling or a signallingeffect. Notch, amyloid precursor protein and Erb-B4 undergoregulated cleavage, and signalling mechanisms associated with theirintracellular fragments have been elucidated31. Overexpressed CTF2translocates to the nucleus and stimulate transcription of cyclin D1andβ-catenin genes in pre-delaminating neural crest cells46.Moreover,the N-cadherin intracellular fragment can apparently induce thedegradation of the transcriptional co-activator cyclic AMP responseelement binding protein (CREB; refs 34,47). Alternatively, N-cadherincleavage could contribute to signalling through the release of β-catenin38. Cytoplasmic β-catenin is known to influence the balancebetween proliferation and differentiation of fetal and adult neuralprecursor cells, probably in response to Wnt signalling48. Adulttransgenic mice that express a stabilized form of β-catenin exhibit anenlarged SEZ, owing to the proliferation of Mash1+ TAP cells andinhibition of their differentiation into neuroblasts47,49. The potential
8 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION
© 2014 Macmillan Publishers Limited. All rights reserved.
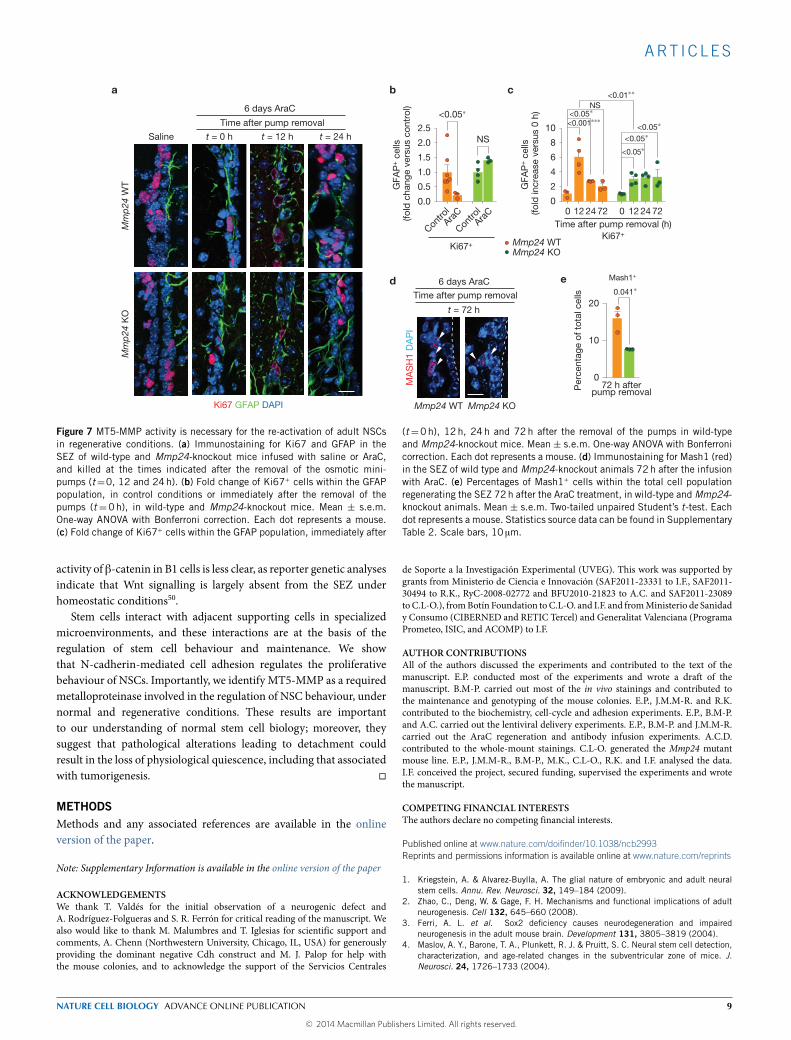
ART ICLES
0 12 24 0 12 2472 72
GFA
P+ c
ells
(fol
d in
crea
se v
ersu
s 0
h)
10
4
8
6
2
<0.05∗
<0.05∗
<0.001∗∗∗
NS
Time after pump removal (h)
c
0
<0.05∗
<0.05∗
<0.01∗∗
GFA
P+ c
ells
(fol
d c
hang
e ve
rsus
con
trol
)
Contro
l
2.5
2.0
1.5
1.0
0.5
0.0
<0.05∗
NS
Contro
lAra
CAra
C
Mmp24 WTMmp24 KO
Mm
p24
WT
Mm
p24
KO
Ki67 GFAP DAPI
6 days AraC
Saline t = 12 ht = 0 h
Time after pump removal
t = 24 h
t = 72 h
MA
SH
1 D
AP
IMmp24 WT Mmp24 KO
6 days AraC
Time after pump removal
0
20
10
Per
cent
age
of t
otal
cel
ls 0.041∗
Mash1+
72 h afterpump removal
e
ba
d
Ki67+Ki67+
Figure 7 MT5-MMP activity is necessary for the re-activation of adult NSCsin regenerative conditions. (a) Immunostaining for Ki67 and GFAP in theSEZ of wild-type and Mmp24-knockout mice infused with saline or AraC,and killed at the times indicated after the removal of the osmotic mini-pumps (t=0, 12 and 24h). (b) Fold change of Ki67+ cells within the GFAPpopulation, in control conditions or immediately after the removal of thepumps (t =0h), in wild-type and Mmp24-knockout mice. Mean ± s.e.m.One-way ANOVA with Bonferroni correction. Each dot represents a mouse.(c) Fold change of Ki67+ cells within the GFAP population, immediately after
(t=0h), 12 h, 24h and 72h after the removal of the pumps in wild-typeand Mmp24-knockout mice. Mean ± s.e.m. One-way ANOVA with Bonferronicorrection. Each dot represents a mouse. (d) Immunostaining for Mash1 (red)in the SEZ of wild type and Mmp24-knockout animals 72h after the infusionwith AraC. (e) Percentages of Mash1+ cells within the total cell populationregenerating the SEZ 72h after the AraC treatment, in wild-type andMmp24-knockout animals. Mean ± s.e.m. Two-tailed unpaired Student’s t-test. Eachdot represents a mouse. Statistics source data can be found in SupplementaryTable 2. Scale bars, 10 µm.
activity of β-catenin in B1 cells is less clear, as reporter genetic analysesindicate that Wnt signalling is largely absent from the SEZ underhomeostatic conditions50.
Stem cells interact with adjacent supporting cells in specializedmicroenvironments, and these interactions are at the basis of theregulation of stem cell behaviour and maintenance. We showthat N-cadherin-mediated cell adhesion regulates the proliferativebehaviour of NSCs. Importantly, we identify MT5-MMP as a requiredmetalloproteinase involved in the regulation of NSC behaviour, undernormal and regenerative conditions. These results are importantto our understanding of normal stem cell biology; moreover, theysuggest that pathological alterations leading to detachment couldresult in the loss of physiological quiescence, including that associatedwith tumorigenesis. �
METHODSMethods and any associated references are available in the onlineversion of the paper.
Note: Supplementary Information is available in the online version of the paper
ACKNOWLEDGEMENTSWe thank T. Valdés for the initial observation of a neurogenic defect andA. Rodríguez-Folgueras and S. R. Ferrón for critical reading of the manuscript. Wealso would like to thank M. Malumbres and T. Iglesias for scientific support andcomments, A. Chenn (Northwestern University, Chicago, IL, USA) for generouslyproviding the dominant negative Cdh construct and M. J. Palop for help withthe mouse colonies, and to acknowledge the support of the Servicios Centrales
de Soporte a la Investigación Experimental (UVEG). This work was supported bygrants from Ministerio de Ciencia e Innovación (SAF2011-23331 to I.F., SAF2011-30494 to R.K., RyC-2008-02772 and BFU2010-21823 to A.C. and SAF2011-23089toC.L-O.), fromBotín Foundation toC.L-O. and I.F. and fromMinisterio de Sanidady Consumo (CIBERNED and RETIC Tercel) and Generalitat Valenciana (ProgramaPrometeo, ISIC, and ACOMP) to I.F.
AUTHOR CONTRIBUTIONSAll of the authors discussed the experiments and contributed to the text of themanuscript. E.P. conducted most of the experiments and wrote a draft of themanuscript. B.M-P. carried out most of the in vivo stainings and contributed tothe maintenance and genotyping of the mouse colonies. E.P., J.M.M-R. and R.K.contributed to the biochemistry, cell-cycle and adhesion experiments. E.P., B.M-P.and A.C. carried out the lentiviral delivery experiments. E.P., B.M-P. and J.M.M-R.carried out the AraC regeneration and antibody infusion experiments. A.C.D.contributed to the whole-mount stainings. C.L-O. generated the Mmp24 mutantmouse line. E.P., J.M.M-R., B.M-P., M.K., C.L-O., R.K. and I.F. analysed the data.I.F. conceived the project, secured funding, supervised the experiments and wrotethe manuscript.
COMPETING FINANCIAL INTERESTSThe authors declare no competing financial interests.
Published online at www.nature.com/doifinder/10.1038/ncb2993Reprints and permissions information is available online at www.nature.com/reprints
1. Kriegstein, A. & Alvarez-Buylla, A. The glial nature of embryonic and adult neuralstem cells. Annu. Rev. Neurosci. 32, 149–184 (2009).
2. Zhao, C., Deng, W. & Gage, F. H. Mechanisms and functional implications of adultneurogenesis. Cell 132, 645–660 (2008).
3. Ferri, A. L. et al. Sox2 deficiency causes neurodegeneration and impairedneurogenesis in the adult mouse brain. Development 131, 3805–3819 (2004).
4. Maslov, A. Y., Barone, T. A., Plunkett, R. J. & Pruitt, S. C. Neural stem cell detection,characterization, and age-related changes in the subventricular zone of mice. J.Neurosci. 24, 1726–1733 (2004).
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 9
© 2014 Macmillan Publishers Limited. All rights reserved.

ART ICLES
5. Nam, H. S. & Benezra, R. High levels of Id1 expression define B1 type adult neuralstem cells. Cell Stem Cell 5, 515–526 (2009).
6. Wilson, A. et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell 135, 1118–1129 (2008).
7. Doetsch, F., Garcia-Verdugo, J. M. & Alvarez-Buylla, A. Regeneration of agerminal layer in the adult mammalian brain. Proc. Natl Acad. Sci. USA 96,11619–11624 (1999).
8. Pastrana, E., Cheng, L. C. & Doetsch, F. Simultaneous prospective purification ofadult subventricular zone neural stem cells and their progeny. Proc. Natl Acad. Sci.USA 106, 6387–6392 (2009).
9. Andreu-Agullo, C., Morante-Redolat, J. M., Delgado, A. C. & Farinas, I. Vascular nichefactor PEDF modulates Notch-dependent stemness in the adult subependymal zone.Nat. Neurosci. 12, 1514–1523 (2009).
10. Miyatani, S. et al. Neural cadherin: Role in selective cell–cell adhesion. Science245, 631–635 (1989).
11. Takeichi, M. Morphogenetic roles of classic cadherins. Curr. Opin. Cell Biol. 7,619–627 (1995).
12. Marthiens, V., Kazanis, I., Moss, L., Long, K. & Ffrench-Constant, C. Adhesionmolecules in the stem cell niche–more than just staying in shape? J. Cell Sci. 123,1613–1622 (2010).
13. Gonzalez-Reyes, A. Stem cells, niches and cadherins: A view from Drosophila. J. CellSci. 116, 949–954 (2003).
14. Kiel, M. J., Acar, M., Radice, G. L. & Morrison, S. J. Hematopoietic stem cellsdo not depend on N-cadherin to regulate their maintenance. Cell Stem Cell 4,170–179 (2009).
15. Kiel, M. J., Radice, G. L. & Morrison, S. J. Lack of evidence that hematopoietic stemcells depend on N-cadherin-mediated adhesion to osteoblasts for their maintenance.Cell Stem Cell 1, 204–217 (2007).
16. Li, P. & Zon, L. I. Resolving the controversy about N-cadherin and hematopoieticstem cells. Cell Stem Cell 6, 199–202 (2010).
17. Karpowicz, P. et al. E-Cadherin regulates neural stem cell self-renewal. J. Neurosci.29, 3885–3896 (2009).
18. Seki, T., Namba, T., Mochizuki, H. & Onodera, M. Clustering, migration, and neuriteformation of neural precursor cells in the adult rat hippocampus. J. Comp. Neurol.502, 275–290 (2007).
19. Shen, Q. et al. Adult SVZ stem cells lie in a vascular niche: A quantitative analysisof niche cell–cell interactions. Cell Stem Cell 3, 289–300 (2008).
20. Yagita, Y. et al. N-cadherin mediates interaction between precursor cells in thesubventricular zone and regulates further differentiation. J. Neurosci. Res. 87,3331–3342 (2009).
21. Barnabe-Heider, F. et al. Genetic manipulation of adult mouse neurogenic niches byin vivo electroporation. Nat. Methods 5, 189–196 (2008).
22. Folgueras, A. R. et al. Metalloproteinase MT5-MMP is an essential modulator ofneuro-immune interactions in thermal pain stimulation. Proc. Natl Acad. Sci. USA106, 16451–16456 (2009).
23. Monea, S., Jordan, B. A., Srivastava, S., DeSouza, S. & Ziff, E. B. Membranelocalization of membrane type 5 matrix metalloproteinase by AMPA receptor bindingprotein and cleavage of cadherins. J. Neurosci. 26, 2300–2312 (2006).
24. Mirzadeh, Z., Merkle, F. T., Soriano-Navarro, M., Garcia-Verdugo, J. M. & Alvarez-Buylla, A. Neural stem cells confer unique pinwheel architecture to the ventricularsurface in neurogenic regions of the adult brain. Cell Stem Cell 3, 265–278 (2008).
25. Nose, A., Nagafuchi, A. & Takeichi, M. Expressed recombinant cadherins mediatecell sorting in model systems. Cell 54, 993–1001 (1988).
26. Radice, G.L. et al. Developmental defects in mouse embryos lacking N-cadherin.Dev. Biol. 181, 64–78 (1997).
27. Malatesta, P. et al. Neuronal or glial progeny: Regional differences in radial glia fate.Neuron 37, 751–764 (2003).
28. Consiglio, A. et al. Robust in vivo gene transfer into adult mammalianneural stem cells by lentiviral vectors. Proc. Natl Acad. Sci. USA 101,14835–14840 (2004).
29. Noles, S. R. & Chenn, A. Cadherin inhibition of beta-catenin signaling regulates theproliferation and differentiation of neural precursor cells. Mol. Cell. Neurosci. 35,549–558 (2007).
30. Nieman, M. T., Kim, J. B., Johnson, K. R. & Wheelock, M. J. Mechanism ofextracellular domain-deleted dominant negative cadherins. J. Cell Sci. 112,1621–1632 (1999).
31. Fortini, M. E. Gamma-secretase-mediated proteolysis in cell–surface–receptorsignalling. Nat. Rev. Mol. Cell Biol. 3, 673–684 (2002).
32. Jaworski, D. M. Developmental regulation of membrane type-5 matrixmetalloproteinase (MT5-MMP) expression in the rat nervous system. Brain Res.860, 174–177 (2000).
33. Malinverno, M. et al. Synaptic localization and activity of ADAM10 regulate excitatorysynapses through N-cadherin cleavage. J. Neurosci. 30, 16343–16355 (2010).
34. Marambaud, P. et al. A CBP binding transcriptional repressor produced by thePS1/epsilon-cleavage of N-cadherin is inhibited by PS1 FAD mutations. Cell 114,635–645 (2003).
35. Reiss, K. et al. ADAM10 cleavage of N-cadherin and regulation of cell–cell adhesionand beta-catenin nuclear signalling. Embo J. 24, 742–752 (2005).
36. Sekine-Aizawa, Y. et al. Matrix metalloproteinase (MMP) system in brain:Identification and characterization of brain-specific MMP highly expressed incerebellum. Eur. J. Neurosci. 13, 935–948 (2001).
37. Uemura, K. et al. Characterization of sequential N-cadherin cleavage by ADAM10and PS1. Neurosci. Lett. 402, 278–283 (2006).
38. Heuberger, J. & Birchmeier, W. Interplay of cadherin-mediated cell adhesion andcanonical Wnt signaling. Cold Spring Harb. Perspect. Biol. 2, a002915 (2010).
39. Raponi, E. et al. S100B expression defines a state in which GFAP-expressing cellslose their neural stem cell potential and acquire a more mature developmental stage.Glia 55, 165–177 (2007).
40. Niola, F. et al. Id proteins synchronize stemness and anchorage to the niche of neuralstem cells. Nat. Cell Biol. 14, 477–487 (2012).
41. Llano, E. et al. Identification and characterization of human MT5-MMP, a newmembrane-bound activator of progelatinase a overexpressed in brain tumors. CancerRes. 59, 2570–2576 (1999).
42. Pei, D. Identification and characterization of the fifth membrane-type matrixmetalloproteinase MT5-MMP. J. Biol. Chem. 274, 8925–8932 (1999).
43. Esteve, P. et al. SFRPs act as negative modulators of ADAM10 to regulate retinalneurogenesis. Nat. Neurosci. 14, 562–569 (2011).
44. Luo, J., Shook, B. A., Daniels, S. B. & Conover, J. C. Subventricular zone-mediated ependymal repair in the adult mammalian brain. J. Neurosci. 28,3804–3813 (2008).
45. Falcao, A. M. et al. The path from the choroid plexus to the subventricular zone: Gowith the flow! Front. Cell. Neurosci. 6, 1–8 (2012).
46. Shoval, I., Ludwig, A. & Kalcheim, C. Antagonistic roles of full-length N-cadherin andits soluble BMP cleavage product in neural crest delamination. Development 134,491–501 (2007).
47. Andreyeva, A. et al. C-terminal fragment of N-cadherin accelerates synapsedestabilization by amyloid-beta. Brain 135, 2140–2154 (2012).
48. Zhang, L., Yang, X., Yang, S. & Zhang, J. The Wnt/beta-catenin signaling pathway inthe adult neurogenesis. Eur. J. Neurosci. 33, 1–8 (2011).
49. Adachi, K. et al. Beta-catenin signaling promotes proliferation of progenitor cells inthe adult mouse subventricular zone. Stem Cells 25, 2827–2836 (2007).
50. Piccin, D. & Morshead, C. M. Wnt signaling regulates symmetry of division ofneural stem cells in the adult brain and in response to injury. Stem Cells 29,528–538 (2011).
10 NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION
© 2014 Macmillan Publishers Limited. All rights reserved.

DOI: 10.1038/ncb2993 METHODS
METHODSAnimals and in vivo manipulations. Generation and genotyping of Mmp24mutant mice have been described previously22. B6.129S6(SJL)-Cdh2tm1Glr/J micecarrying a conditional Cdh2 allele (Cdh2floxed) and FVB-Tg(GFAP-cre)25Mes/Jmice expressing Cre recombinase specifically in astrocytes and astroglial-likeNSCs (GFAP(Cre)) were purchased from The Jackson Laboratory and werecrossed to obtain GFAP(Cre);Cdh21 and control Cdh2floxed littermates. Tg(CAG–eGFP)B5Nagy/J and Tg(GFAP–lacZ,eGFP)7Kdmc reporter mice were obtainedfrom The Jackson Laboratory and the MMRRC, respectively. Housing of miceand all experiments were carried out according to European Union 86/609/EECand Spanish RD-1201/2005 guidelines, following protocols approved by thecorresponding local ethics committees. For BrdU and CldU, mice were injectedintraperitoneally with seven consecutive injections (one every 2 h) at 50mg per kgof body weight in 0.09% NaCl (saline) solution and left to survive for 28 days. ForIdU, mice received one single pulse at 50mg per kg of body weight in 20% dimethylsulfoxide in saline and were killed 1 h after the injection. Mice were anaesthetizedusing a veterinary supervised mixture of ketamine and medetomidine and fixedto a stereotactic device (NeuroLab, Leica Biosystems). For cerebral infusions, AraC(2%, Sigma) in saline, or saline alone, was infused onto the surface of the brain ofadult two-month-old Mmp24 mutant or wild-type mice (n= 4, per genotype andsurvival time condition) with osmotic mini-pumps (ALZET, Palo Alto, CA, model1007D; flow rate 0.5 µl h−1) coupled to a brain infusion kit 3 cannula. Stereotacticcoordinates used were (mm from Bregma): mediolateral (ML)=+1.1. After 6 daysof infusion, mice were killed immediately or at the indicated survival times afterpump removal. Efficiency of the ablation of TAP cells and neuroblasts was assessedfor each animal using Ki67 and DCX stainings. One mouse was excluded from thestudy because no efficient ablation was found. No significant differences were foundbetween saline-infused and naive animals. For lentivector injection, 1.5-month-oldC57/BL6 femalemicewere used.Onemicroliter of vector concentrate (∼1×106)wasinjected with a 33G Hamilton syringe (0.2 µl min−1). Stereotactic coordinates usedwere (mm from Bregma): for the SEZ, anterior–posterior (AP) =+0.6, ML =+1.2and dorsoventral (DV) =−3 from the skull surface; for the lateral ventricle, AP =−0.3, ML=+1, DV=−2.6. Mice were killed and brains analysed two months afterinjection in the SEZ or 15 days after injection into the ventricle to allow for transgeneexpression and to minimize confounding factors caused by the injection procedure.For BrdU-LRC assessment, BrdU was injected 10 days after the intraventricularinfections, and mice were allowed to survive for another 21 days before euthanasia.For cerebral infusions of the EC-N-cadherin antibody, osmoticmini-pumps coupledto a brain infusion kit 3 cannula (ALZET, model 1007D; flow rate 0.5 µl h−1) wereloaded with 80 µg of antibody or isotype IgG per mini-pump diluted in saline andimplanted in two month-oldMmp24-knockout mice. Stereotactic coordinates usedwere (mm from Bregma): AP =−0.3, ML =−0.8 and DV =−3 from the skullsurface. After 7 days of infusion, mice were killed. Stereotactic coordinates wereoptimized for each mouse strain in advance.
Immunohistochemistry. Animals were deeply anaesthetized and transcardiallyperfused with 4% paraformaldehyde in 0.1 M PBS and brains processed forvibratome sectioning (Leica VT1,000 vibratome), or fresh tissue was dissectedto obtain whole mounts of the SEZ as described24. Samples were blocked in 5%normal goat serum and 0.1% Triton X-100 in PBS for 1 h and incubated for48 h in blocking buffer with the appropriate primary antibodies: rabbit anti-Ki67(1/100; Abcam); chick anti-GFAP (1/600; Millipore); mouse anti-N-cadherin cloneC32 (N-cad; 1/100; BD); mouse anti-N-cadherin clone GC4 (EC-N-cad; 1/100;Sigma); mouse anti-E-cadherin (1/100; BD); mouse anti-Mash1 (1/100; BD); mouseanti-nestin (1/500; Abcam); rabbit anti-Id1 (1/200; BioCheck); rabbit anti-S100B(1/100; Dako); goat anti-MCM2 (1/100; Santa Cruz Biotechnology); goat anti-DCX(1/200; Santa Cruz Biotechnology); goat anti-GFP (1/500; Rockland); rabbit anti-GFP (1/200; Abcam); rabbit anti-β-catenin (1/100; Cell Signaling); goat anti-γ-tubulin (1/200; Santa Cruz Biotechnology) and goat anti-MT5-MMP (1/100; SantaCruz Biotechnology). For the specific detection of the synthetic nucleotides, a 20min 2N HCl incubation was carried out before the addition of the antibodies;rat anti-BrdU for CldU or BrdU (1/1,000; Abcam), and mouse anti-BrdU for IdU(1/500; BD). Immunofluorescent detections were carried out with Alexa Fluor(Invitrogen) or DyLight (Pierce) conjugated secondary antibodies. For N- andE-cadherin immunodetection, sections were incubated with biotinylated horseanti-mouse antibodies (Vector Laboratories) followed by Cy2- or Cy3-conjugatedstreptavidin (Jackson ImmunoResearch Laboratories). DAPI (1mgml−1) was usedfor counterstaining. Samples were analysed in anOlympus FluoView FV10i confocallaser scanning microscope equipped with 405, 458, 488 and 633 nm lasers. Imageswere acquired and processed using FV10-ASW 2.1 viewer software.
Cell culture, cell transduction, co-culture and cell-cycle analyses. Methodsfor neurosphere culture have been previously described. Briefly, adult SEZs weredissected and disaggregated using 0.9mgml−1 papain, and the obtained cells were
grown in a defined medium51 using 20 ngml−1 murine EGF and 10 ngml−1 hu-man basic fibroblast growth factor (FGF) as mitogens. L929 cells were grown inDMEM supplemented with 1% l-glutamine and 10% fetal bovine serum. 293Tcells were grown in Iscove’s modified Dulbecco’s medium supplemented with 10%fetal bovine serum and 1% l-glutamine. MT5-MMP complementary DNA wasFlag tagged at the C-terminus and subsequently cloned in the lentiviral vector pRRL-SIN-PPThPGK–GFP–WPRE (Addgene plasmid no 12252) using XhoI and SpeIsites. Primer sequences used for cloning MT5-MMP in the lentiviral vector areforward, 5′-ATTCTC GAGAATGCCGAGGAGCGCGGG-3′, and reverse, 5′-GCACT AGTCTACTTGTCATCGTCGT-3′. TheDN-CDH–GFP constructwas a giftfrom A. Chenn29, and was subcloned in the lentivector using XhoI and SpeIsites. Primer sequences used for cloning are forward, 5′-A TAT CTC GAG ACTGGT GTG GGA GCC-3′, and reverse, 5′-CG ACT AGT TTA CTT GTA CAGCTC GTC-3′. High-titre VSV-pseudotyped lentiviral vector stocks were producedin 293T cells by transient transfection using Lipofectamine 2000 (Invitrogen) ofthe transfer vectors: pRRL-SIN-PPThPGK–GFP–WPRE, pRRL-SIN-PPThPGK–GFP–WPRE-DN-CDH–GFP and pRRL-SIN-PPThPGK–GFP–WPRE–MT5-MMP(herein Lent–GFP, Lent–GFP–DN-CDH and Lent–GFP–MT5-MMP), the late-generation packaging construct pMDL, the rev construct pRSV-rev and the VSVenvelope-expressing construct pMD2.G, and purified by ultracentrifugation as pre-viously described28. Expression titres, determined on 293T cells by fluorescence-activated cell sorter analysis (EPICS, Coulter), were 9×106 TU µl−1 for Lent–GFP,3.5× 107 TU µl−1 for Lent–GFP–DN-CDH and 3.2× 106 TU µl−1 for Lent–GFP–MT5-MMP. For adhesion experiments, either parental or stable L929 clones over-expressing E- or N-cadherin were seeded on glass coverslips and left to reachconfluence for 24–48 h. Freshly disaggregated cells were obtained from the SEZ ofat least three adult mice of each of the appropriate genotypes; 1.25×104 cells cm−2were seeded on the pre-formedmonolayers andwere left to attach for 3 h, after whichcultures were extensively washed to remove unattached cells and cultured in neuralstem cell medium supplementedwith FGF (ref. 51) for 10–12 h. Cells were then fixedand stained for analysis. For the adhesion rescue experiments, neurospheres fromMmp24-knockout mutant mice were obtained and infected with the concentratedlentiviral particles for the overexpression of MT5-MMP, or empty Lent–PGK–GFP,using 1× 106 TU µl−1 in the presence of 4 µgml−1 polybrene, FACS-sorted usinga MoFlo XDP cytometer (Beckman Coulter) and subjected to the latter protocol.Cells were pre-incubated for 1 h with 50 µgml−1 of the blocking antibody against theextracellular domain of N-cadherin or isotype control immunoglobulins before plat-ing. A representative sample was photographed and the number of attached nestin+cells was scored in each condition. For cell-cycle analysis, FACS-sorted lentiviral-transduced neurospheres were collected by centrifugation, disaggregated usingAccutase (Sigma) and fixed and permeabilized before incubation with 40 µgml−1propidium iodide and 40 µgml−1 ribonuclease. Samples of each condition wereanalysed for DNA content with a FACSCanto II (Becton Dickinson), and cell-cycleprofiles and phase percentages were determined using FlowJo software.
Immunoblot. Cell and tissue extracts were lysed in SDS buffer (25mM Tris-HClat pH 7.4, 1mM EDTA, 1% SDS, 10mM sodium pyrophosphate, 20mM β-glycerolphosphate, 2mM sodium orthovanadate) and a protease inhibitor cocktail (Roche).Protein concentration was determined using a commercial colorimetric assay (BCAprotein assay kit, Pierce) and 30–50 µg of protein were resolved by 10% SDS–PAGEor 4–15% Mini-PROTEAN TGX gels (Bio-Rad) and transferred to nitrocellulosemembranes (Protran, Sigma). Membranes were blocked in PBS containing 3% BSAor 5% skimmed milk, and 0.025% Tween-20, and incubated with mouse anti-N-cadherin (BD; Sigma, both 1:1,000), β-actin (1:5,000; Sigma) or Flag (clone M2,1:1,000; Sigma) and rabbit β-catenin (Cell Signaling; 1:1,000) anti-NICD (Abcam;1:1,000) antibodies, followed by appropriate peroxidase-conjugated anti-mouse oranti-rabbit IgGs (1:5,000; Dako). Signal was obtained by enhanced chemolumines-cence (Western Lightning, Perkin Elmer). Autoradiographic films were scanned andthe bands were analysed by densitometry using ImageJ software (NIH).
Statistical analyses.Analyses of significant differences betweenmeanswere carriedout using the unpaired or paired two-tailed Student t-test or one-way ANOVAwith Bonferroni correction when appropriate. When comparisons were carried outwith relative values (normalized values and percentages), data were first normalizedby using a log or arcsin transformation, respectively. No statistical method wasused to predetermine sample size. Sample sizes were based on previously publishedexperiments by us and others, and on our own unpublished data. The experimentswere not randomized. The investigators were not blinded to allocation duringexperiments or outcome assessment. Data are always presented as the mean ±standard error of the mean (s.e.m.), and the number of experiments carried out withindependent cultures/animals (n) is shown as dots in the graphs.
51. Ferron, S. R. et al. A combined ex/in vivo assay to detect effects of exogenouslyadded factors in neural stem cells. Nat. Protocol. 2, 849–859 (2007).
NATURE CELL BIOLOGY
© 2014 Macmillan Publishers Limited. All rights reserved.

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
DOI: 10.1038/ncb2993
projection
b
z z+1
z+2 z+3
aNcad GFPGFAP γ-tubulinGFAPγ-tubulinβ-catenin
GFAP γ-tubulin
c GFP
GFP
S100β GFPS100β DAPI
Supplementary Figure 1
z-1 z+1
Merg
eNcad
GFA
P Ki
67 D
API
z level
zd
Ncad
Ki67
DAP
I
inset
inset
Supplementary Figure 1 Adult neural stem cells express functional N-cadherin. (a) Immunofluorescent detection of GFAP (red), b-catenin (blue) and g-tubulin (green) in the wall of the lateral ventricle of wild type mice. (b) Wholemount staining for N-cadherin (red) and GFP (green) in GFAP-eGFP mice. Note that the GFP+ cell shown is also positive for N-cadherin (white arrowheads). Stainings at specific confocal levels are shown (z). Lower panel; higher magnification of z-stack projection showing a GFP+, B cell. (c) Immunofluorescent detection of the ependymal marker S100b (red), GFP (green) and DAPI (blue) in preparations from co-culture experiments using SEZ homogenates from GFAP-eGFP mice on N-cadherin
overexpressing L929 cells. (d) Immunofluorescent detection of GFAP (green), Ki67 (cyan) and N-cadherin (red) in the SEZ of wild type mice. Shown is the staining at a specific confocal level (z). Right set of smaller panels show a higher magnification of the area outlined by the white line in the left panel, at the specified confocal levels. Non-proliferative GFAP+ cells show higher levels of N-cadherin staining (full arrowheads) whilst activated to proliferate, Ki67-GFAP double positive cells, have lowered N-cadherin levels (empty arrowheads). Nuclei are counterstained with DAPI (blue), and the dashed white lines mark the ventricle limit. Scale bars: (a, c, d) 5 mm; (b) 10 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.
S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
Ncad
GFA
P DAP
Ia b
E-ca
dher
in D
API
S100β GFAP DAPIz stack levels 1-3 z stack levels 1-3
z stack levels 4-7 z stack levels 4-7
S100β GFAPc d
Cdh2floxed GFAP(Cre);Cdh2Δ Cdh2floxed GFAP(Cre);Cdh2Δ
Cdh2floxed GFAP(Cre);Cdh2Δ
Cdh2floxed GFAP(Cre);Cdh2Δ
Supplementary Figure 2
Supplementary Figure 2 N- and E-cadherin expression, and pinwheel structure integrity in GFAP(Cre);Cdh2D mice. (a) Immunostaining for GFAP (red) and N-cadherin (green), and (b) E-cadherin (green) in SEZ coronal sections of adult mice with the conditional deletion of N-cadherin in GFAP+ cells (GFAP(Cre);Cdh2D) or control littermates (Cdh2floxed). DAPI was used for counterstaining (blue). The dashed white lines
mark the ventricle limit. Note the absence of signal for N-cadherin in the SEZ of conditionally targeted animals, whilst E-cadherin staining remained apparently unaffected. (c) Immunofluorescent staining for the detection of S100b (red) and GFAP (green) in tissue from Cdh2floxed and GFAP(Cre);Cdh2D mice, in wholemount preparations and (d) coronal sections. Scale bars: 10 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
1
2 4
3 5 7
6 8
ba
rostral medial caudal
12
34
5 6 7 8
OB DCX
DCX
1
2 4
3 5 7
6 8
c
Cdh2
floxe
d
GFA
P(Cr
e);C
dh2Δ
Cdh2floxed GFAP(Cre);Cdh2Δ
Cdh2
floxe
dG
FAP(
Cre)
;Cdh
2Δ
Supplementary Figure 3
Supplementary Figure 3 Neurogenic input in GFAP(Cre);Cdh2D mice. (a) Macroscopic images of Cdh2floxed and GFAP(Cre);Cdh2D brains. Note the overall increased brain size, and the reduced olfactory bulbs (dashed black lines). (b) Confocal reconstructions of wholemount stainings for the
neuroblast marker DCX (gray) in Cdh2floxed and GFAP(Cre);Cdh2D mice. (c) Magnified images showing the chain-like structures of the migrating neuroblasts (DCX, orange) at the positions indicated in the schematic representation. Scale bars: (a) 2500 mm; (b) 200 mm; (c) 50 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.

S U P P L E M E N TA RY I N F O R M AT I O N
4 WWW.NATURE.COM/NATURECELLBIOLOGY
Supplementary Figure 4
a
Lent-
GFP-
DN-C
DHLe
nt-GF
P
GFAP Ki67 DAPI GFP DAPI
S100β GFP EC-NCad GFP
Lent-
GFP-
DN-C
DHLe
nt-GF
P
*
*
*
*
Intraventricular delivery
c
eb
d
** **
***
**
*GFP
S100
β LR
C DA
PI
Lent-G
FP-MT5-MMP
Lent-G
FP-DN-CDH
Lent-G
FP
f g hIntraventricular delivery
GFP MT5-MMPGFPDAPI
MT5-MMP GFP MT5-MMP
Lent-
GFP-
MT5-
MMP
Lent-
GFP
Striatal delivery
str
str
lv
lv
i
GFP
S100β GFP EC-NCad GFP
Supplementary Figure 4 In vivo localized loss and gain of function assays. (a,c) Immunofluorescent detection of GFP (green) in wholemounts from intraventricularly infected mice with control vector (Lent-GFP) and dominant-negative cadherin (Lent-GFP-DN-CDH). Infection with Lent-GFP-DN-CDH provokes the detachment of ependymocytes from the ventricle wall. (b,d) Detection of S100b (blue), GFP (green) and EC-Ncadherin (red) in wholemounts of the infected ventricles. Note that the discrete membrane localization of EC-Ncadherin (arrowheads) in GFP+ (asterisks) and non-infected ependymal cells shown in control infections (b) is disrupted in Lent-GFP-DN-CDH infected cells (d). (e) GFAP (green) and Ki67 (red) inmunodetection in the SEZ of DN-CDH intraventricularly infected mice.
GFP is shown pseudocolored in orange and nuclei are counterstained with DAPI (blue). (f-h) Immunofluorescent detection of S100b (green) and BrdU (red) in coronal sections from infected mice using control (Lent-GFP, f), dominant-negative cadherin (Lent-GFP-DN-CDH, g) and MT5-MMP (Lent-GFP-MT5-MMP, h) carrying lentiviruses. BrdU-label retaining cells (LRC)-S100b double positive cells are pointed at with white arrowheads, while LRC+ cells negative for S100b are pointed at with empty arrowheads. (i) Staining for GFP (green) and MT5-MMP (red) in coronal sections from control and MT5-MMP infected striata, showing the overexpression of the metalloproteinase in infected cells. Scale bars: (a, c) 200 mm; (b-e) 10 mm; (f-h) 15 mm; (i) 200 mm, 10 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 5
GFAP γtub βcat GFP GFAP γtub βcat GFP GFAP γtub βcat c d
fe
h
Lent-
MT5-
MMP
Lent-
DN-C
DHLe
nt-GF
P
Lent-GFP-MT5-MMP a S100β GFP EC-NCad GFP
Intraventricular delivery
g
b
Supplementary Figure 5
GFP
Supplementary Figure 5. In vivo localized loss and gain of function provokes the disruption of pinwheel-structures at the surface of the ventricle. (a) Immunofluorescent detection of GFP in wholemounts from MT5-MMP infected mice. Note that infection with Lent-GFP-MT5-MMP also provokes the detachment of ependymocytes from the ventricle wall, however the effect is milder than with Lent-GFP-DN-CDH (Supplementary Figure 4c). (b) Detection of S100b (blue), GFP (green) and EC-Ncadherin (red) in wholemounts of the infected ventricles. Note that the discrete membrane localization of EC-Ncadherin in GFP+ and non-infected
(empty arrowheads) ependymal cells is disrupted in Lent-GFP-MT5-MMP infected cells (white arrowheads). Parallel control stainings can be seen in Supplementary Figure 4b. (c-h) Wholemount staining with anti-GFAP (cyan), b-catenin and g-tubulin (red) and GFP (green) to assess pinwheel structures (white dashed lines and arrows) in infected ventricles. (c,e,g: general view; d,f,h: detailed images). We were able to find some pinwheels in MT5-MMP-infected ventricles (white arrow), in contrast to DN-CDH infected areas. Scale bars: (a,b) 200 mm; 50 mm; 10 mm. (c,e,g) 10 mm. (d, f, h) 5 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.
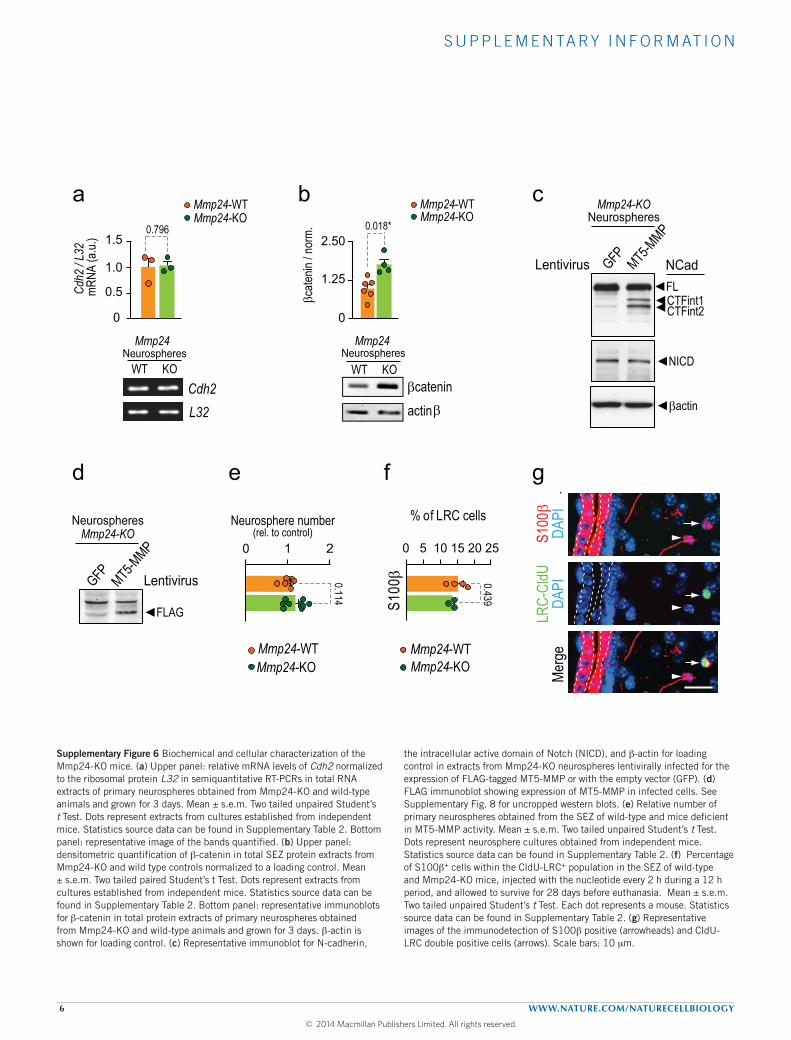
S U P P L E M E N TA RY I N F O R M AT I O N
6 WWW.NATURE.COM/NATURECELLBIOLOGY
Supplementary Figure 6
Cdh2
/ L3
2 mR
NA (a
.u.) 1.5
0
1.0
0.5
0.796
a
Cdh2 L32
NeurospheresMmp24
WT KO
Mmp24-WTMmp24-KO
β
b
Mmp24
actinβcatenin
2.50
0
1.25
βcate
nin / n
orm.
0.018*
NeurospheresWT KO
Mmp24-WTMmp24-KO
S100
β
% of LRC cells
0 10 25
f
Me
rge
S100
βDA
PILR
C-Cl
dUDA
PI
g
Neurosphere number(rel. to control)
0 21
e
20155
0.439
0.114
Mmp24-WTMmp24-KO
Mmp24-WTMmp24-KO
◄FL
◄NICD
◄FLAG
◄βactin
◄CTFint1CTFint2◄
c
d
Lentivirus GFP MT5-M
MP
NeurospheresMmp24-KO
Lentivirus GFP MT5-M
MP
NeurospheresMmp24-KO
NCad
◄
◄
◄
Supplementary Figure 6 Biochemical and cellular characterization of the Mmp24-KO mice. (a) Upper panel: relative mRNA levels of Cdh2 normalized to the ribosomal protein L32 in semiquantitative RT-PCRs in total RNA extracts of primary neurospheres obtained from Mmp24-KO and wild-type animals and grown for 3 days. Mean ± s.e.m. Two tailed unpaired Student’s t Test. Dots represent extracts from cultures established from independent mice. Statistics source data can be found in Supplementary Table 2. Bottom panel: representative image of the bands quantified. (b) Upper panel: densitometric quantification of b-catenin in total SEZ protein extracts from Mmp24-KO and wild type controls normalized to a loading control. Mean ± s.e.m. Two tailed paired Student’s t Test. Dots represent extracts from cultures established from independent mice. Statistics source data can be found in Supplementary Table 2. Bottom panel: representative immunoblots for b-catenin in total protein extracts of primary neurospheres obtained from Mmp24-KO and wild-type animals and grown for 3 days. b-actin is shown for loading control. (c) Representative immunoblot for N-cadherin,
the intracellular active domain of Notch (NICD), and b-actin for loading control in extracts from Mmp24-KO neurospheres lentivirally infected for the expression of FLAG-tagged MT5-MMP or with the empty vector (GFP). (d) FLAG immunoblot showing expression of MT5-MMP in infected cells. See Supplementary Fig. 8 for uncropped western blots. (e) Relative number of primary neurospheres obtained from the SEZ of wild-type and mice deficient in MT5-MMP activity. Mean ± s.e.m. Two tailed unpaired Student’s t Test. Dots represent neurosphere cultures obtained from independent mice. Statistics source data can be found in Supplementary Table 2. (f) Percentage of S100b+ cells within the CldU-LRC+ population in the SEZ of wild-type and Mmp24-KO mice, injected with the nucleotide every 2 h during a 12 h period, and allowed to survive for 28 days before euthanasia. Mean ± s.e.m. Two tailed unpaired Student’s t Test. Each dot represents a mouse. Statistics source data can be found in Supplementary Table 2. (g) Representative images of the immunodetection of S100b positive (arrowheads) and CldU-LRC double positive cells (arrows). Scale bars: 10 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.
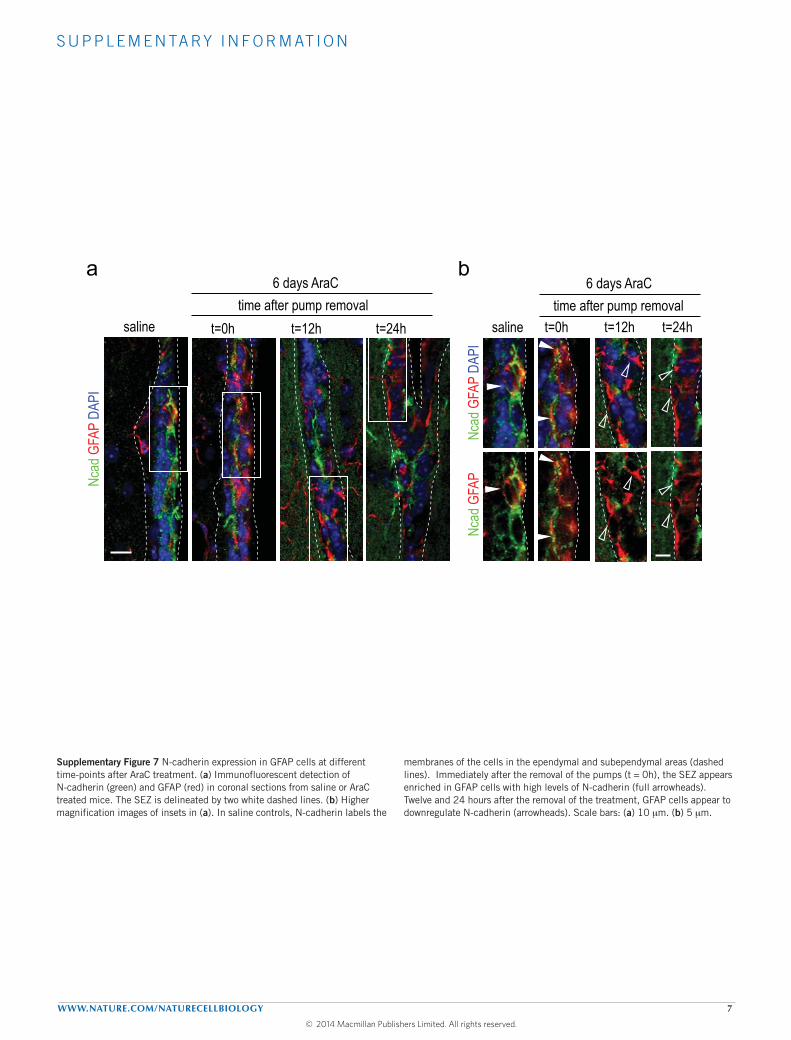
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 7
6 days AraC
saline t=12ht=0htime after pump removal
Ncad
GFA
P DA
PI
Ncad
GFA
P Nc
ad G
FAP
DAPI
t=24h
ba6 days AraC
saline t=12ht=0htime after pump removal
t=24h
Supplementary Figure 7
Supplementary Figure 7 N-cadherin expression in GFAP cells at different time-points after AraC treatment. (a) Immunofluorescent detection of N-cadherin (green) and GFAP (red) in coronal sections from saline or AraC treated mice. The SEZ is delineated by two white dashed lines. (b) Higher magnification images of insets in (a). In saline controls, N-cadherin labels the
membranes of the cells in the ependymal and subependymal areas (dashed lines). Immediately after the removal of the pumps (t = 0h), the SEZ appears enriched in GFAP cells with high levels of N-cadherin (full arrowheads). Twelve and 24 hours after the removal of the treatment, GFAP cells appear to downregulate N-cadherin (arrowheads). Scale bars: (a) 10 mm. (b) 5 mm.
© 2014 Macmillan Publishers Limited. All rights reserved.

S U P P L E M E N TA RY I N F O R M AT I O N
8 WWW.NATURE.COM/NATURECELLBIOLOGY
IB: Ncad
755037
100150
kDa
75kDa
100150
75kDa
IB: NICD
37kDa
50
IB: βactin
IB: Ncad
GFP MT5-MMP
75
50kDa
IB: FLAG
IB: FLAG
75
50kDa
GFP MT5-MMP
SEZ tissue
Ponceau
IB: Ncad
IB: EC-Ncad75kDa
100150
75100150
kDa
75100150
kDa
5037
Culture #1 Culture #2
GFP
MT5-
MMP
GFP
MT5-
MMP
Culture #1 Culture #2
GFP
MT5-
MMP
GFP
MT5-
MMP
Culture #1
Culture #2
75150
kDa
5037
Ponceau
IB: Ncad
Mmp24 Neurospheres
75150
kDa
5037
Cdh2L32
Neurospheres Mmp24
NTC
NTC
Neurospheres
MW
100bp
Mmp24
IB: βactin
IB: βcatenin
75150
kDa
5037
kDa
Neurospheres
Supplementary Figure 8
WT WT WT WT WTKO KO KO KOKO
WT WT WT KO KO KO WT WT WT KO KO KO
Mmp24WT WT KO KO
Mmp24a b
i
h
c
d f
IB: Ncad (long exposure)
IB: Ncad
75
50
100150
37kDa
75
50
100150
37kDa
Neurospheres
WT WT WT KO KO KO
Mmp24
NeurospheresMmp24-KO
g
e
NeurospheresMmp24-KO
NeurospheresMmp24-KO
Supplementary Figure 8 Uncropped western blots and DNA agarose gels. (a,b) Immunoblots (IB) and ponceau-s staining in Fig. 5b and Fig. 5c. (c,d) Immunoblots in Fig. 5e-d. (f-i) Immunoblots and agarose gel in Supplementary Fig. 6a,b,c,d. MW: molecular weight marker; NTC: no template control.
© 2014 Macmillan Publishers Limited. All rights reserved.
Related Documents











