THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 19% by The American Society of Biological Chemists, Inc. Vol. 259, No. 21, Issue of November 10, pp. 13541-13549,1984 Printed in U. S. A. Regulation of Light-harvesting Chlorophyll-binding Protein mRNA Accumulation in Chlamydomonas reinhardi POSSIBLE INVOLVEMENT OF CHLOROPHYLL SYNTHESIS PRECURSORS* (Received for publication, March 26, 1984) Udo JohanningmeierS and Stephen H. Howell From the Biology Department, C-016, University of California San Diego, LaJolla, Californin 92093 Light induction of light-harvesting chlorophyll a b - binding protein (LHCP) mRNA accumulation was stud- ied in light-dark synchronized cultures of Chlamydo- monas reinhardi. LHCP mRNA accumulation was pre- vented by the chlorophyll-synthesis inhibitor a,a-di- pyridyl which blocks late steps in the chlorophyll bio- synthetic pathway and leads to the accumulation of the porphyrin intermediate magnesium protoporphyrin methyl ester. LHCP mRNA accumulated normally, however, when chlorophyll synthesis was blocked by inhibitors such as hemin and levulinic acid which in- terfere with early steps in the chlorophyll biosynthesis pathway prior to the formation of magnesium proto- porphyrin methyl ester. Similar effects were observed in the light induction of LHCP mRNA levels in proto- porphyrin IX-accumulating mutants, brc-l and brs-l. These mutants havelow levels of LHCP mRNA when grown under heterotrophic conditions in thedark where they accumulate protoporphyrin IX. However, LHCP mRNA is light-induced in brc-l which synthe- sizes chlorophyll in the light and presumably consumes porphyrin intermediates in doing so. These results sug- gest that the chlorophyll-synthesis intermediates, mag- nesium protoporphyrin methyl ester and its immediate precursors, inhibit by a feedback mechanism the light induction of LHCP mRNA accumulation. Low magne- sium protoporphyrin methyl ester levels permit the light-induced accumulation of LHCP mRNA, whereas high magnesium protoporphyrin methyl ester levels destabilize LHCP mRNA regardless of the illumination conditions. Preliminary experiments show that LHCP mRNA accumulation in C. reinhardi is stimulated by blue light, and not byred light which stimulates LHCP mRNA accumulation in higher plants. In Chlamydomonas reinhardi, as in higher plants, LHCPs’ are encoded by the nuclear genome (Chua and Gillham, 1977), synthesized as precursors on cytoplasmic ribosomes (Hoober, 1970; Hoober and Stegeman, 1976),transported into the chlo- roplast, and inserted into thylakoid membranes (Schmidt et * This work was supported by a grant from the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ Supported by the Deutsche Forschungsgemeinschaft (Jo 123/1- 1). Present address, Department of Chemistry, Box 215, University of Colorado, Boulder, Colorado 80309. The abbreviations used are: LHCP, light-harvesting chlorophyll ab-binding protein; DP, a,a’-dipyridyl; EGTA, ethylene glycol bis(p- aminoethyl ether)-N,N,N’,N’-tetraacetic acid; Et, ethyl: Ac, acetyl. al., 1981) where they serve as apoproteins for the light- harvesting chlorophyll complexes. The biosynthesis and func- tional properties of LHCPs are regulated at many different levels by light. In higher plants, light, acting through phyto- chrome, influences the accumulation of LHCP mRNA (Apel, 1979; Stiekema et al., 1983). Light may also affect the trans- lation of LHCP mRNA (Slovin and Tobin, 1982). Addition- ally, the stable assembly of LHCPs in thylakoid membranes appears to depend on the concomitant light-driven synthesis of chlorophyll (Apel and Kloppstech, 1980; Cuming and Ben- nett, 1981). Finally, the activity of assembled LHCPs may be modulated by light-dependent phosphorylation (Bennett, 1979; Bennett et al., 1980; Allen et al., 1981). In C. reinhurdi, LHCP expression and activity may also be regulated at many different levels. LHCP mRNA can be induced both by light (Shepherd et al., 1983) and heat shock (Hoober et aL, 1982). Also, LHCP is phosphorylated in a light-dependent fashion (Owens and Ohad, 1982). In this paper, we have studied the factors which regulate the accumulation of LHCP mRNA during the light phase in light-dark synchronized C. reinhardi cells. We present evi- dence which implicates both light and porphyrin precursors in chlorophyll biosynthesis as regulators of LHCP mRNA accumulation under these conditions. Evidence for the in- volvement of porphyrin intermediates in the regulation of LHCP mRNA accumulation was obtained through the use of chlorophyll-synthesis inhibitors and protoporphyrin-accu- mulating mutants of C. reinhardi. To examine changes in LHCP mRNA levels, we used the cDNA clone pHs16 as a probe which specifically hybridizes to the 1.2-kb mRNA that encodes the 32-kDa LHCP precursor protein in C. reinhardi (Shepherd et al., 1983). MATERIALS AND METHODS Growth of Cells-A cell-wall-deficient strain of C. reinhardi CW15 (provided by D. R. Davies) was grown photoautotrophically at 20 “C under synchronous conditions (12-h light (L)/12-h dark (D) alternat- ing illumination cycles). Synchronous cells were subjected to at least three light-dark cycles before they were harvested for various exper- iments at cell densities between 5-10 x 10’ cells/ml. C. reinhardi mutants br8-l and brc-l (kindly provided by Dr. W.-Y. Wang) were grown in HSA medium as described by Wang et al. (1974). Cells were treated with hemin (Pfaltz and Bauer, Stamford CN) by adding hemin from a 10 mM stock solution (hemin dissolved in H20 by adding NaOH and finally adjusted to pH 7.5 with NaH,PO,) to a final concentration of 0.1 mM. Before adding hemin to thecells, the pH of the medium was adjusted to pH 7.2 by adding 1 M Tris-HC1 (pH 7.2) to a final concentration of20mM. When levulinic acid was added, the pH of the levulinic acid solution (Sigma) was adjusted to 6.5 before it was added to the liquid culture. a,a-Dipyridyl (Sigma) was added to cultures from a 1 M solution in EtOH. Filters for red and blue light experiments had the following spectral properties. Red filter: A,;, = 0.4 at 665 nm, A 2 1 at 630 and 700 nm; blue filter: Amin = 0.25 at 470 nm, A 2 1 at 460 and 485 nm. 13541

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 19% by The American Society of Biological Chemists, Inc.
Vol. 259, No. 21, Issue of November 10, pp. 13541-13549,1984 Printed in U. S. A .
Regulation of Light-harvesting Chlorophyll-binding Protein mRNA Accumulation in Chlamydomonas reinhardi POSSIBLE INVOLVEMENT OF CHLOROPHYLL SYNTHESIS PRECURSORS*
(Received for publication, March 26, 1984)
Udo JohanningmeierS and Stephen H. Howell From the Biology Department, C-016, University of California San Diego, LaJolla, Californin 92093
Light induction of light-harvesting chlorophyll a b - binding protein (LHCP) mRNA accumulation was stud- ied in light-dark synchronized cultures of Chlamydo- monas reinhardi. LHCP mRNA accumulation was pre- vented by the chlorophyll-synthesis inhibitor a,a-di- pyridyl which blocks late steps in the chlorophyll bio- synthetic pathway and leads to the accumulation of the porphyrin intermediate magnesium protoporphyrin methyl ester. LHCP mRNA accumulated normally, however, when chlorophyll synthesis was blocked by inhibitors such as hemin and levulinic acid which in- terfere with early steps in the chlorophyll biosynthesis pathway prior to the formation of magnesium proto- porphyrin methyl ester. Similar effects were observed in the light induction of LHCP mRNA levels in proto- porphyrin IX-accumulating mutants, brc-l and brs-l. These mutants have low levels of LHCP mRNA when grown under heterotrophic conditions in the dark where they accumulate protoporphyrin IX. However, LHCP mRNA is light-induced in brc-l which synthe- sizes chlorophyll in the light and presumably consumes porphyrin intermediates in doing so. These results sug- gest that the chlorophyll-synthesis intermediates, mag- nesium protoporphyrin methyl ester and its immediate precursors, inhibit by a feedback mechanism the light induction of LHCP mRNA accumulation. Low magne- sium protoporphyrin methyl ester levels permit the light-induced accumulation of LHCP mRNA, whereas high magnesium protoporphyrin methyl ester levels destabilize LHCP mRNA regardless of the illumination conditions. Preliminary experiments show that LHCP mRNA accumulation in C. reinhardi is stimulated by blue light, and not by red light which stimulates LHCP mRNA accumulation in higher plants.
In Chlamydomonas reinhardi, as in higher plants, LHCPs’ are encoded by the nuclear genome (Chua and Gillham, 1977), synthesized as precursors on cytoplasmic ribosomes (Hoober, 1970; Hoober and Stegeman, 1976), transported into the chlo- roplast, and inserted into thylakoid membranes (Schmidt et
* This work was supported by a grant from the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Supported by the Deutsche Forschungsgemeinschaft (Jo 123/1- 1). Present address, Department of Chemistry, Box 215, University of Colorado, Boulder, Colorado 80309.
The abbreviations used are: LHCP, light-harvesting chlorophyll ab-binding protein; DP, a,a’-dipyridyl; EGTA, ethylene glycol bis(p- aminoethyl ether)-N,N,N’,N’-tetraacetic acid; Et, ethyl: Ac, acetyl.
al., 1981) where they serve as apoproteins for the light- harvesting chlorophyll complexes. The biosynthesis and func- tional properties of LHCPs are regulated at many different levels by light. In higher plants, light, acting through phyto- chrome, influences the accumulation of LHCP mRNA (Apel, 1979; Stiekema et al., 1983). Light may also affect the trans- lation of LHCP mRNA (Slovin and Tobin, 1982). Addition- ally, the stable assembly of LHCPs in thylakoid membranes appears to depend on the concomitant light-driven synthesis of chlorophyll (Apel and Kloppstech, 1980; Cuming and Ben- nett, 1981). Finally, the activity of assembled LHCPs may be modulated by light-dependent phosphorylation (Bennett, 1979; Bennett et al., 1980; Allen et al., 1981). In C. reinhurdi, LHCP expression and activity may also be regulated at many different levels. LHCP mRNA can be induced both by light (Shepherd et al., 1983) and heat shock (Hoober et aL, 1982). Also, LHCP is phosphorylated in a light-dependent fashion (Owens and Ohad, 1982).
In this paper, we have studied the factors which regulate the accumulation of LHCP mRNA during the light phase in light-dark synchronized C. reinhardi cells. We present evi- dence which implicates both light and porphyrin precursors in chlorophyll biosynthesis as regulators of LHCP mRNA accumulation under these conditions. Evidence for the in- volvement of porphyrin intermediates in the regulation of LHCP mRNA accumulation was obtained through the use of chlorophyll-synthesis inhibitors and protoporphyrin-accu- mulating mutants of C. reinhardi. To examine changes in LHCP mRNA levels, we used the cDNA clone pHs16 as a probe which specifically hybridizes to the 1.2-kb mRNA that encodes the 32-kDa LHCP precursor protein in C. reinhardi (Shepherd et al., 1983).
MATERIALS AND METHODS
Growth of Cells-A cell-wall-deficient strain of C. reinhardi CW15 (provided by D. R. Davies) was grown photoautotrophically at 20 “C under synchronous conditions (12-h light (L)/12-h dark (D) alternat- ing illumination cycles). Synchronous cells were subjected to at least three light-dark cycles before they were harvested for various exper- iments at cell densities between 5-10 x 10’ cells/ml. C. reinhardi mutants br8-l and brc-l (kindly provided by Dr. W.-Y. Wang) were grown in HSA medium as described by Wang et al. (1974). Cells were treated with hemin (Pfaltz and Bauer, Stamford CN) by adding hemin from a 10 mM stock solution (hemin dissolved in H20 by adding NaOH and finally adjusted to pH 7.5 with NaH,PO,) to a final concentration of 0.1 mM. Before adding hemin to the cells, the pH of the medium was adjusted to pH 7.2 by adding 1 M Tris-HC1 (pH 7.2) to a final concentration of 20 mM. When levulinic acid was added, the pH of the levulinic acid solution (Sigma) was adjusted to 6.5 before it was added to the liquid culture. a,a-Dipyridyl (Sigma) was added to cultures from a 1 M solution in EtOH. Filters for red and blue light experiments had the following spectral properties. Red filter: A,;, = 0.4 at 665 nm, A 2 1 at 630 and 700 nm; blue filter: Amin = 0.25 at 470 nm, A 2 1 at 460 and 485 nm.
13541
13542 Light-harvesting Chlorophyll-binding Protein mRNA Regulation
RNA Extraction-CW15 cells (2 X IO7 in total) were harvested by centrifugation, and the cell pellet was suspended in 100 pl of extrac- tion buffer containing 0.1 M Tris-HCI (pH 8.5), 0.4 M LiCI, 0.01 M EGTA, 0.005 M EDTA. 10 pl of a 20% sodium dodecyl sulfate solution and 2 pl of a proteinase K solution (4 mg/ml, Bethesda Research Laboratories) was added and the sample set on ice for 5-10 min. The resulting cell lysate was extracted twice with phenol, and RNA was precipitated overnight by adding 0.1 volume of 2 M NaOAc (pH 5) and 2.5 volumes of 100% EtOH. The precipitate was washed once with 70% EtOH and finally dissolved in 20 pl of H20.
RNA from walled strains was obtained by suspending cells in extraction buffer as described above and sonicating the suspension for 1 min using a Sonifier Cell Disruptor (Branson Sonic Power Co.) with a microtip (power setting 2) in the presence of diethylpyrocar- bonate a t a final concentration of 2% (v/v). After sonication, sodium dodecyl sulfate was added to a final concentration of 2% (w/v) and the sample set on ice for 5 min. Further extraction steps were performed as described above.
RNA Blots-Total RNA from 4 X lo6 mother cells was subjected to electrophoresis on 1.5% agarose gels containing 10 mM methyl- mercuric hydroxide (Alpha Products, Danvers, MA) according to Bailey and Davidson (1976). RNA was transferred to nitrocellulose paper (BA83, Schleicher and Schuell) and hybridized in dextran- sulfate-containing solutions to 32P-labeled, nick-translated probe DNA (usually -5 X lo6 cpm), according to Wahl et al. (1979). All RNA samples were subjected to electrophoresis on duplicate gels. One gel was used for RNA transfer, the other was stained with ethidium bromide to assess the integrity of the RNA samples. In addition, gels were stained following transfer to check for quantitative RNA transfer to the filter.
RESULTS
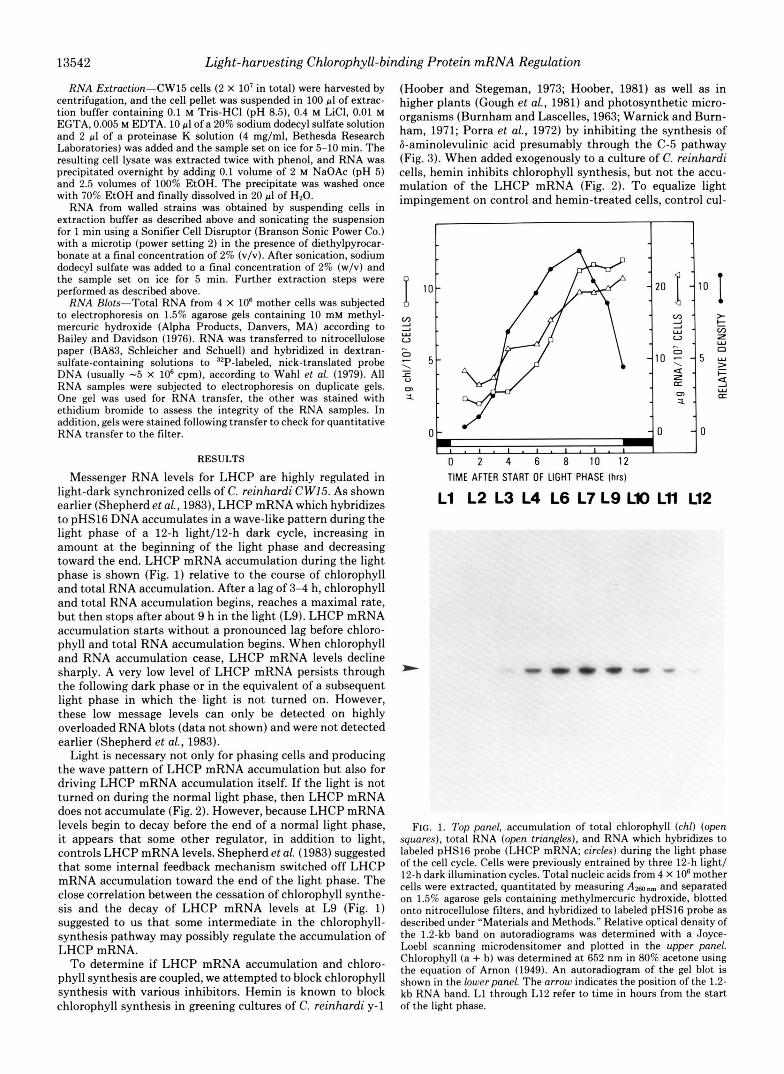
Messenger RNA levels for LHCP are highly regulated in light-dark synchronized cells of C. reinhardi CWI5. As shown earlier (Shepherd et al., 1983), LHCP mRNA which hybridizes to pHs16 DNA accumulates in a wave-like pattern during the light phase of a 12-h light/l2-h dark cycle, increasing in amount at the beginning of the light phase and decreasing toward the end. LHCP mRNA accumulation during the light phase is shown (Fig. 1) relative to the course of chlorophyll and total RNA accumulation. After a lag of 3-4 h, chlorophyll and total RNA accumulation begins, reaches a maximal rate, but then stops after about 9 h in the light (L9). LHCP mRNA accumulation starts without a pronounced lag before chloro- phyll and total RNA accumulation begins. When chlorophyll and RNA accumulation cease, LHCP mRNA levels decline sharply. A very low level of LHCP mRNA persists through the following dark phase or in the equivalent of a subsequent light phase in which the light is not turned on. However, these low message levels can only be detected on highly overloaded RNA blots (data not shown) and were not detected earlier (Shepherd et al., 1983).
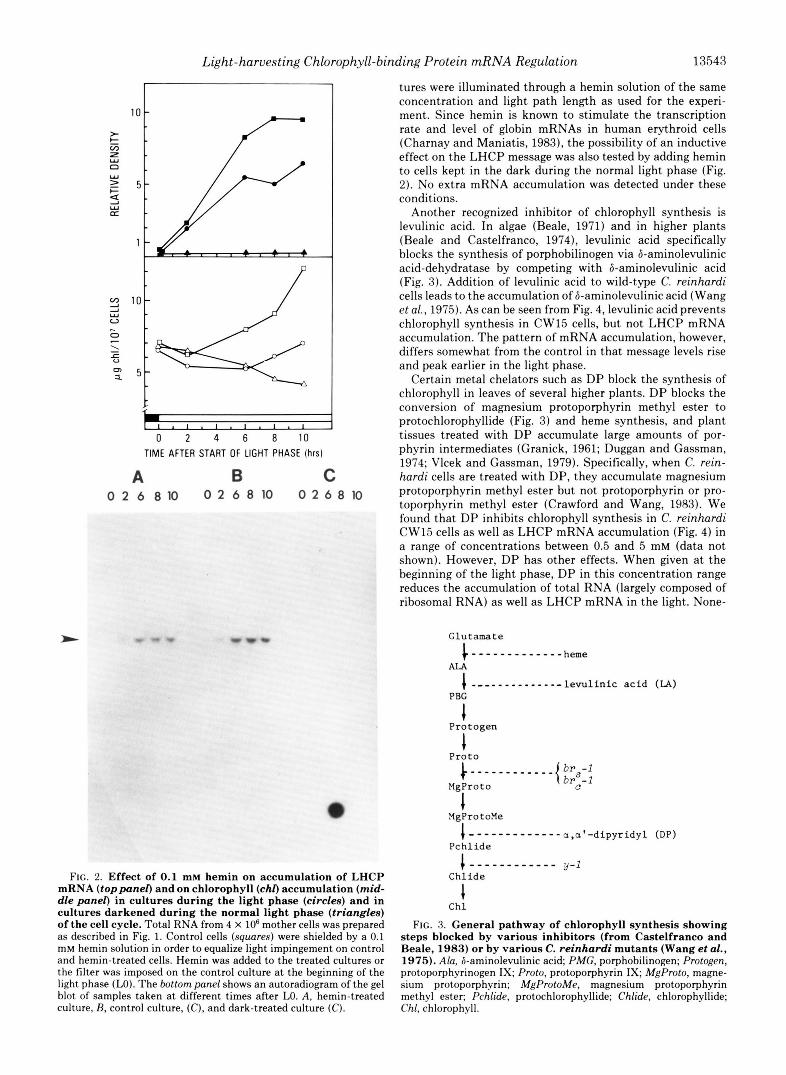
Light is necessary not only for phasing cells and producing the wave pattern of LHCP mRNA accumulation but also for driving LHCP mRNA accumulation itself. If the light is not turned on during the normal light phase, then LHCP mRNA does not accumulate (Fig. 2). However, because LHCP mRNA levels begin to decay before the end of a normal light phase, it appears that some other regulator, in addition to light, controls LHCP mRNA levels. Shepherd et al. (1983) suggested that some internal feedback mechanism switched off LHCP mRNA accumulation toward the end of the light phase. The close correlation between the cessation of chlorophyll synthe- sis and the decay of LHCP mRNA levels at L9 (Fig. 1) suggested to us that some intermediate in the chlorophyll- synthesis pathway may possibly regulate the accumulation of LHCP mRNA.
To determine if LHCP mRNA accumulation and chloro- phyll synthesis are coupled, we attempted to block chlorophyll synthesis with various inhibitors. Hemin is known to block chlorophyll synthesis in greening cultures of C. reinhardi y-1
(Hoober and Stegeman, 1973; Hoober, 1981) as well as in higher plants (Gough et al., 1981) and photosynthetic micro- organisms (Burnham and Lascelles, 1963; Warnick and Burn- ham, 1971; Porra et al., 1972) by inhibiting the synthesis of 8-aminolevulinic acid presumably through the C-5 pathway (Fig. 3). When added exogenously to a culture of C. reinhardi cells, hemin inhibits chlorophyll synthesis, but not the accu- mulation of the LHCP mRNA (Fig. 2). To equalize light impingement on control and hemin-treated cells, control cul-
TIME AFTER START OF LIGHT PHASE (hrs)
L1 L2 L3 L4 L6 L7 L9 LlO Lll L12
F I G . 1. Top panel, accumulation of total chlorophyll (chl) (open sqwres), total RNA (open triangles), and RNA which hybridizes to labeled pHs16 probe (LHCP mRNA; circles) during the light phase of the cell cycle. Cells were previously entrained by three 12-h light/ 12-h dark illumination cycles. Total nucleic acids from 4 X IO6 mother cells were extracted, quantitated by measuring Amnm and separated on 1.5% agarose gels containing methylmercuric hydroxide, blotted onto nitrocellulose filters, and hybridized to labeled pHs16 probe as described under “Materials and Methods.” Relative optical density of the 1.2-kb band on autoradiograms was determined with a Joyce- Loebl scanning microdensitomer and plotted in the upper panel. Chlorophyll (a + b) was determined at 652 nm in 80% acetone using the equation of Arnon (1949). An autoradiogram of the gel blot is shown in the lowerpanel. The arrow indicates the position of the 1.2- kb RNA band. L1 through L12 refer to time in hours from the start of the light phase.
Light-harvesting Chlorophyll-binding Protein m R N A Regulation 13543
t
0 2 4 6 8 1 0 TIME AFTER START OF LIGHT PHASE (hrs)
FIG. 2. Effect of 0.1 mM hemin on accumulation of LHCP mRNA (toppanel) and on chlorophyll (chl) accumulation (mid- dle panel) in cultures during the light phase (circles) and in cultures darkened during the normal light phase (triangles) of the cell cycle. Total RNA from 4 X lo6 mother cells was prepared as described in Fig. 1. Control cells (squures) were shielded by a 0.1 mM hemin solution in order to equalize light impingement on control and hemin-treated cells. Hemin was added to the treated cultures or the filter was imposed on the control culture at the beginning of the light phase (LO). The bottom panel shows an autoradiogram of the gel blot of samples taken at different times after LO. A, hemin-treated culture, R, control culture, (C), and dark-treated culture (C).
tures were illuminated through a hemin solution of the same concentration and light path length as used for the experi- ment. Since hemin is known to stimulate the transcription rate and level of globin mRNAs in human erythroid cells (Charnay and Maniatis, 1983), the possibility of an inductive effect on the LHCP message was also tested by adding hemin to cells kept in the dark during the normal light phase (Fig. 2). No extra mRNA accumulation was detected under these conditions.
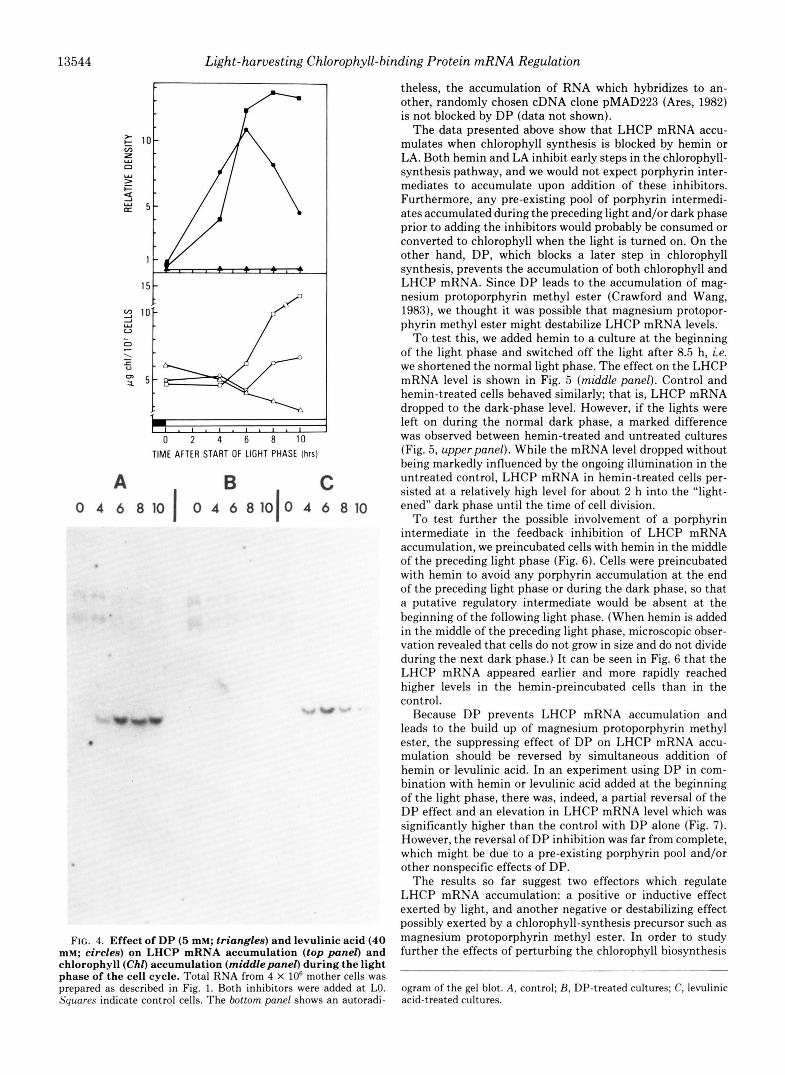
Another recognized inhibitor of chlorophyll synthesis is levulinic acid. In algae (Beale, 1971) and in higher plants (Beale and Castelfranco, 1974), levulinic acid specifically blocks the synthesis of porphobilinogen via 6-aminolevulinic acid-dehydratase by competing with 6-aminolevulinic acid (Fig. 3). Addition of levulinic acid to wild-t,ype C. reinhardi cells leads to the accumulation of b-aminolevulinic acid (Wang et al., 1975). As can be seen from Fig. 4, levulinic acid prevents chlorophyll synthesis in CW15 cells, but not LHCP mRNA accumulation. The pattern of mRNA accumulation, however, differs somewhat from the control in that message levels rise and peak earlier in the light phase.
Certain metal chelators such as DP block the synthesis of chlorophyll in leaves of several higher plants. DP blocks the conversion of magnesium protoporphyrin methyl ester to protochlorophyllide (Fig. 3) and heme synthesis, and plant tissues treated with DP accumulate large amounts of por- phyrin intermediates (Granick, 1961; Duggan and Gassman, 1974; Vlcek and Gassman, 1979). Specifically, when C. rein- hardi cells are treated with DP, they accumulate magnesium protoporphyrin methyl ester but not protoporphyrin or pro- toporphyrin methyl ester (Crawford and Wang, 1983). We found that DP inhibits chlorophyll synthesis in C. reinhardi CW15 cells as well as LHCP mRNA accumulation (Fig. 4) in a range of concentrations between 0.5 and 5 mM (data not shown). However, DP has other effects. When given at the beginning of the light phase, DP in this concentration range reduces the accumulation of total RNA (largely composed of ribosomal RNA) as well as LHCP mRNA in the light. None-
PBG -"-- . . - . . - - - - levul inic acid (LA)
I + +
Protogen
pr;:- - - - - { E$ MgProto
XgProtoMe
1 - - - - - - - - - - - - - a , a ' - d ipy r idy l (DP) Pch l ide + - - - - - - - - - - - -
+ Y-*
Chl ide
Chl
FIG. 3. General pathway of chlorophyll synthesis showing steps blocked by various inhibitors (from Castelfranco and Beale, 1983) or by various C. reinhardi mutants (Wang et al., 1975). A h , 6-aminolevulinic acid; PMG, porphobilinogen; Protogen, protoporphyrinogen IX; Proto, protoporphyrin IX; MgProto, magne- sium protoporphyrin; MgProtoMe, magnesium protoporphyrin methyl ester; Pcchlide, protochlorophyllide; Chlide, chlorophyllide; Chl, chlorophyll.
13544 Light-harvesting Chlorophyll-binding Protein mRNA Regulation
151
L: “
0 2 4 6 8 1 0 TIME AFTER START OF LIGHT PHASE (hrs)
A B C 0 4 6 810 I 0 4 6 81010 4 6 810
e
FIG. 4. Effect of DP (5 mM; triangles) and levulinic acid (40 mM; circles) on LHCP mRNA accumulation (top panel) and chlorophyll (Chl) accumulation (middlepanel) during the light phase of the cell cycle. Total RNA from 4 X lo6 mother cells was DreDared as described in Fig. 1. Both inhibitors were added a t LO.
theless, the accumulation of RNA which hybridizes to an- other, randomly chosen cDNA clone pMAD223 (Ares, 1982) is not blocked by DP (data not shown).
The data presented above show that LHCP mRNA accu- mulates when chlorophyll synthesis is blocked by hemin or LA. Both hemin and LA inhibit early steps in the chlorophyll- synthesis pathway, and we would not expect porphyrin inter- mediates to accumulate upon addition of these inhibitors. Furthermore, any pre-existing pool of porphyrin intermedi- ates accumulated during the preceding light and/or dark phase prior to adding the inhibitors would probably be consumed or converted to chlorophyll when the light is turned on. On the other hand, DP, which blocks a later step in chlorophyll synthesis, prevents the accumulation of both chlorophyll and LHCP mRNA. Since DP leads to the accumulation of mag- nesium protoporphyrin methyl ester (Crawford and Wang, 1983), we thought it was possible that magnesium protopor- phyrin methyl ester might destabilize LHCP mRNA levels.
To test this, we added hemin to a culture at the beginning of the light phase and switched off the light after 8.5 h, i.e. we shortened the normal light phase. The effect on the LHCP mRNA level is shown in Fig. 5 (middle panel). Control and hemin-treated cells behaved similarly; that is, LHCP mRNA dropped to the dark-phase level. However, if the lights were left on during the normal dark phase, a marked difference was observed between hemin-treated and untreated cultures (Fig. 5, upperpanel). While the mRNA level dropped without being markedly influenced by the ongoing illumination in the untreated control, LHCP mRNA in hemin-treated cells per- sisted at a relatively high level for about 2 h into the “light- ened” dark phase until the time of cell division.
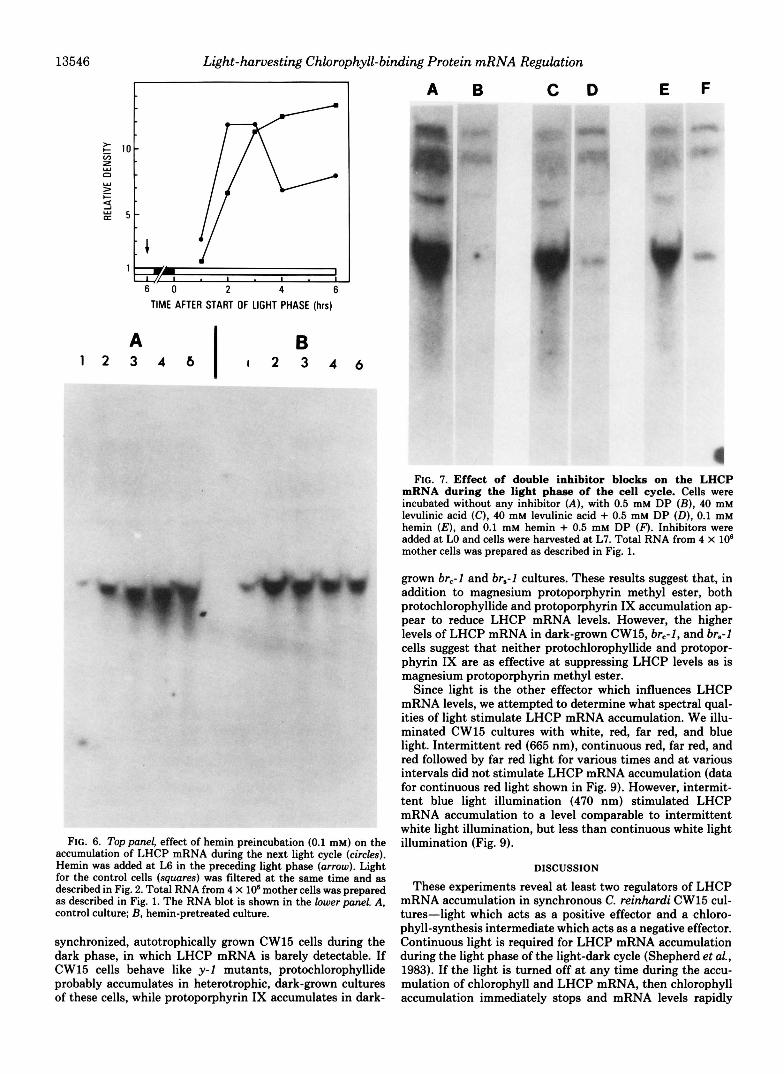
To test further the possible involvement of a porphyrin intermediate in the feedback inhibition of LHCP mRNA accumulation, we preincubated cells with hemin in the middle of the preceding light phase (Fig. 6). Cells were preincubated with hemin to avoid any porphyrin accumulation at the end of the preceding light phase or during the dark phase, so that a putative regulatory intermediate would be absent at the beginning of the following light phase. (When hemin is added in the middle of the preceding light phase, microscopic obser- vation revealed that cells do not grow in size and do not divide during the next dark phase.) It can be seen in Fig. 6 that the LHCP mRNA appeared earlier and more rapidly reached higher levels in the hemin-preincubated cells than in the control.
Because DP prevents LHCP mRNA accumulation and leads to the build up of magnesium protoporphyrin methyl ester, the suppressing effect of D P on LHCP mRNA accu- mulation should be reversed by simultaneous addition of hemin or levulinic acid. In an experiment using DP in com- bination with hemin or levulinic acid added at the beginning of the light phase, there was, indeed, a partial reversal of the DP effect and an elevation in LHCP mRNA level which was significantly higher than the control with DP alone (Fig. 7). However, the reversal of D P inhibition was far from complete, which might be due to a pre-existing porphyrin pool and/or other nonspecific effects of DP.
The results so far suggest two effectors which regulate LHCP mRNA accumulation: a positive or inductive effect exerted by light, and another negative or destabilizing effect possibly exerted by a chlorophyll-synthesis precursor such as magnesium protoporphyrin methyl ester. In order to study further the effects of perturbing the chlorophyll biosynthesis
omam of the gel blot. A. control: B. DP-treated cultures: C. levulinic Squares indicate control cells. The bottom panel shows an autoradi- acid-treated cultures.

t
Light-harvesting Chlorophyll-binding Protein mRNA Regulation 13545
FIG. 5. Effect of extending the light phase by 4 h (toppanel) or shortening the light phase by 3.5 h (middle panel) on the LHCP mRNA accumulation in the presence (circles) and ab- sence (squares) of 0.1 mM hemin added at LO. Total RNA from 4 X 10‘ mother cells was prepared as described in Fig. 1. Light for the control cells was filtered as described in Fig. 2. Autoradiogram of RNA blot of extended light phase (A and B) or of the shortened light phase (C) in the absence ( A ) or presence (B) of hemin.
TIME AFTER START OF LIGHT PHASE fhrr)
pathway on LHCP mRNA accumulation, we made use of two brown mutants of C. reinhurdi, brc-l and brs-l (Wang et al., 1974). Both are known to accumulate protoporphyrin IX and are blocked in the ability to convert protoporphyrin IX into magnesium protoporphyrin in the dark (Fig. 3). However, brc- 1 , but not brs-l cells, are able to carry out this step in the light.
Cells of CW15, brc-l, and brs-l were grown asynchronously under heterotrophic conditions in the dark or mixotrophically in the light. Our CW15 strain is not able to synthesize chlorophyll in the dark, and in that sense behaves as a y-1 mutant which cannot reduce protochlorophyllide in the dark (Sager, 1961). The level of LHCP mRNA in dark-grown CW15 cells, normalized to the same amount of total RNA, is
about 30% of the level found in light-grown cells (Fig. 8). The same difference in light and dark levels of LHCP mRNA was found in brc-l cells, which accumulate protoporphyrin IX and cannot synthesize chlorophyll in the dark but do synthesize chlorophyll in the light. LHCP mRNA levels in br,-l cells do not respond to light (Fig. 8, E and F) and remain at a level comparable to dark-grown brs-l and brc-l (Fig. 80) and CW15 (Fig. 8R) cells. Under the relatively low light intensities used in these experiments, the protoporphyrin IX accumulated in the brs-l mutant is presumably not destroyed even after 24 h in the light, since much higher light intensities (-15,000 lux) are necessary for photodestruction within this time (Wang et al., 1974). LHCP mRNA levels in all dark-grown asynchro- nous cultures (CW15, brc-l, brs-l) were higher than those in
13546 Light-harvesting Chlorophyll-binding Protein mRNA Regulation
1
6 0 2 4 6 I 1
TIME AFTER START OF LIGHT PHASE (hrs)
A 1 2 3 4 6 1 2 3 4 6
B
FIG. 6. Top panel , effect of hemin preincubation (0.1 mM) on the accumulation of LHCP mRNA during the next light cycle (circles). Hemin was added a t L6 in the preceding light phase (arrow). Light for the control cells (squares) was filtered a t the same time and as described in Fig. 2. Total RNA from 4 X lo6 mother cells was prepared as described in Fig. 1. The RNA blot is shown in the lower panel. A, control culture; B, hemin-pretreated culture.
synchronized, autotrophically grown CW15 cells during the dark phase, in which LHCP mRNA is barely detectable. If CW15 cells behave like y-1 mutants, protochlorophyllide probably accumulates in heterotrophic, dark-grown cultures of these cells, while protoporphyrin IX accumulates in dark-
A B C D E F
1 FIG. 7. Effect of double inhibitor blocks on the LHCP
mRNA during the light phase of the cell cycle. Cells were incubated without any inhibitor (A), with 0.5 mM D P (B) , 40 mM levulinic acid (c), 40 mM levulinic acid + 0.5 mM DP (D), 0.1 mM hemin ( E ) , and 0.1 mM hemin + 0.5 mM DP (F). Inhibitors were added at LO and cells were harvested at L7. Total RNA from 4 X lo6 mother cells was prepared as described in Fig. 1.
grown brc-l and br.-l cultures. These results suggest that, in addition to magnesium protoporphyrin methyl ester, both protochlorophyllide and protoporphyrin IX accumulation ap- pear to reduce LHCP mRNA levels. However, the higher levels of LHCP mRNA in dark-grown CW15, brc-l, and br.-1 cells suggest that neither protochlorophyllide and protopor- phyrin IX are as effective at suppressing LHCP levels as is magnesium protoporphyrin methyl ester.
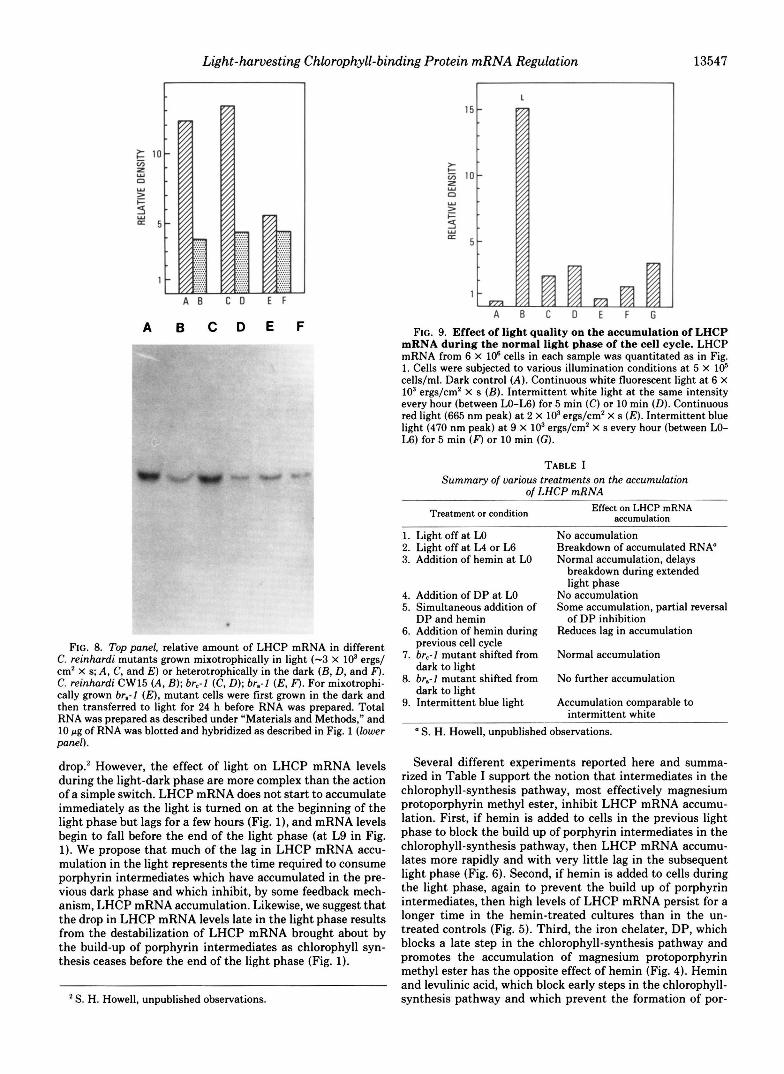
Since light is the other effector which influences LHCP mRNA levels, we attempted to determine what spectral qual- ities of light stimulate LHCP mRNA accumulation. We illu- minated CW15 cultures with white, red, far red, and blue light. Intermittent red (665 nm), continuous red, far red, and red followed by far red light for various times and a t various intervals did not stimulate LHCP mRNA accumulation (data for continuous red light shown in Fig. 9). However, intermit- tent blue light illumination (470 nm) stimulated LHCP mRNA accumulation to a level comparable to intermittent white light illumination, but less than continuous white light illumination (Fig. 9).
DISCUSSION
These experiments reveal at least two regulators of LHCP mRNA accumulation in synchronous C. reinhardi CW15 cul- tures-light which acts as a positive effector and a chloro- phyll-synthesis intermediate which acts as a negative effector. Continuous light is required for LHCP mRNA accumulation during the light phase of the light-dark cycle (Shepherd et al., 1983). If the light is turned off a t any time during the accu- mulation of chlorophyll and LHCP mRNA, then chlorophyll accumulation immediately stops and mRNA levels rapidly
Light-harvesting Chlorophyll-binding Protein mRNA Regulation
A B C D E F . .
FIG. 8. Top pawl , relative amount of LHCP mRNA in different C. reinhrdi mutants grown mixotrophically in light (-3 X lo3 ergs/ cm2 X s; A, C, and E ) or heterotrophically in the dark (B, D, and F). C. reinhrdi CW15 (A, B) ; brc-l (C, D); br.-1 (E , F). For mixotrophi- cally grown br.-l ( E ) , mutant cells were first grown in the dark and then transferred to light for 24 h before RNA was prepared. Total RNA was prepared as described under "Materials and Methods," and 10 pg of RNA was blotted and hybridized as described in Fig. 1 (lower pawl ) .
drop? However, the effect of light on LHCP mRNA levels during the light-dark phase are more complex than the action of a simple switch. LHCP mRNA does not start to accumulate immediately as the light is turned on at the beginning of the light phase but lags for a few hours (Fig. l), and mRNA levels begin to fall before the end of the light phase (at L9 in Fig. 1). We propose that much of the lag in LHCP mRNA accu- mulation in the light represents the time required to consume porphyrin intermediates which have accumulated in the pre- vious dark phase and which inhibit, by some feedback mech- anism, LHCP mRNA accumulation. Likewise, we suggest that the drop in LHCP mRNA levels late in the light phase results from the destabilization of LHCP mRNA brought about by the build-up of porphyrin intermediates as chlorophyll syn- thesis ceases before the end of the light phase (Fig. 1).
S. H. Howell, unpublished observations.
13547
A B C O E F G
FIG. 9. Effect of light quality on the accumulation of LHCP mRNA during the normal light phase of the cell cycle. LHCP mRNA from 6 X lo6 cells in each sample was quantitated as in Fig. 1. Cells were subjected to various illumination conditions at 5 X lo5 cells/ml. Dark control (A) . Continuous white fluorescent light a t 6 X IO3 ergs/cm2 X s (B) . Intermittent white light at the same intensity every hour (between LO-L6) for 5 min (C) or 10 min (D). Continuous red light (665 nm peak) a t 2 X lo3 ergs/cm2 X s (E) . Intermittent blue light (470 nm peak) a t 9 X lo3 ergs/cm2 X s every hour (between LO- L6) for 5 min (F) or 10 min (G).
TABLE I Summary of various treatments on the accumulation
of LHCP mRNA
Treatment or condition Effect on LHCP mRNA accumulation
1. Light off a t LO No accumulation 2. Light off a t L4 or L6 Breakdown of accumulated RNA" 3. Addition of hemin at LO Normal accumulation, delays
breakdown during extended light phase
4. Addition of DP at LO No accumulation 5. Simultaneous addition of Some accumulation, partial reversal
6. Addition of hemin during Reduces lag in accumulation
7. brc-l mutant shifted from Normal accumulation
8. bre-l mutant shifted from No further accumulation
9. Intermittent blue light Accumulation comparable to
DP and hemin of DP inhibition
previous cell cycle
dark to light
dark to light
intermittent white - S. H. Howell, unpublished observations.
Several different experiments reported here and summa- rized in Table I support the notion that intermediates in the chlorophyll-synthesis pathway, most effectively magnesium protoporphyrin methyl ester, inhibit LHCP mRNA accumu- lation. First, if hemin is added to cells in the previous light phase to block the build up of porphyrin intermediates in the chlorophyll-synthesis pathway, then LHCP mRNA accumu- lates more rapidly and with very little lag in the subsequent light phase (Fig. 6). Second, if hemin is added to cells during the light phase, again to prevent the build up of porphyrin intermediates, then high levels of LHCP mRNA persist for a longer time in the hemin-treated cultures than in the un- treated controls (Fig. 5). Third, the iron chelater, DP, which blocks a late step in the chlorophyll-synthesis pathway and promotes the accumulation of magnesium protoporphyrin methyl ester has the opposite effect of hemin (Fig. 4). Hemin and levulinic acid, which block early steps in the chlorophyll- synthesis pathway and which prevent the formation of por-

13548 Light-harvesting Chlorophyll-binding Protein mRNA Regulation
phyrin intermediates, permit LHCP mRNA to accumulate in the light. Fourth, when hemin or levulinic acid are added together with DP, the inhibitory effects of dipyridyl are par- tially overcome (Fig. 7). Most likely this occurs because hemin or levulinic acid block the formation of porphyrin compounds, including magnesium protoporphyrin methyl ester which ac- cumulates in the presence of DP. Fifth, LHCP mRNA is not light-induced in the mutant br8-l, which accumulates proto- porphyrin IX in the dark but cannot convert it to chlorophyll in the light (Fig. 8). On the other hand, LHCP mRNA is light induced in brc-l, which similarly accumulates protoporphyrin IX in the dark but can convert it to chlorophyll in the light.
The effect of light on LHCP mRNA accumulation is com- plex because, in addition to the “direct” effects light may have on LHCP induction, light also drives chlorophyll synthesis, which, as described above, influences LHCP mRNA accu- mulation. Light is needed to photoconvert protochlorophyl- lide to chlorophyllide in C. reinhurdi y-1 mutants (Sager, 1961) and in the absence of light, chlorophyll precursors block the chlorophyll biosynthesis pathway by feedback inhibition (Wang et al., 1974). Thus, in C. reinhardi y-1 mutants and in our CW15 strain which behaves similarly to y - I , the light requirement for chlorophyll synthesis during the light-dark cycle can be attributed to the light requirement to photocon- vert protochlorophyllide to chlorophyllide. However, in syn- chronous wild-type cultures, which should be able to convert protochlorophyllide to chlorophyllide in the dark phase, chlo- rophyll synthesis is restricted to the light phase, and synchro- nous wild-type cultures show the same kinetics of chlorophyll accumulation as our CW15 cultures (see, for example, Schor et al., 1970; Iwanji et al., 1975; Janero and Barrnett, 1981). Unlike higher plants, LHCP mRNA appears to accumulate in response to blue light stimulation in C. reinhardi. The red light receptor which mediates this response in higher plants and Lemna is the pigment phytochrome (Apel, 1979; Stiekema et al., 1983). Either C. reinhurdi has no phytochrome or, if it does, then this response is not mediated by the pigment.
The sort of scheme we suggest for the regulation of LHCP mRNA by chlorophyll-synthesis precursors in C. reinhurdi has certain features in common with that which has been proposed for the regulation of isocytochrome c (CYC-1) gene by heme precursors in yeast. Guarente and Mason (1983) have demonstrated that regulatory elements upstream to the CYC- 1 promoter sense directly or indirectly intracellular levels of heme. In C. reinhurdi we do not know if the LHCP gene itself is influenced by light and chlorophyll-synthesis precursors. Instead, both of these factors may influence the breakdown or turnover of the mRNA and not its transcription. These questions can be addressed when we are able to measure the rates of LHCP mRNA transcript runoff in uitro following various light or inhibitor treatments.
How porphyrin intermediates in chlorophyll synthesis, which are usually thought to be confined to the chloroplast, affect the workings of the nuclear-cytoplasmic genetic system is an interesting question. From our studies, it appears that one of the most effective negative regulators of LHCP mRNA accumulation is magnesium protoporphyrin methyl ester, which has been shown to accumulate in C. reinhardi cells in the presence of DP (Crawford and Wang, 1983). As opposed to all other stroma-soluble chlorophyll precursors, magnesium protoporphyrin and magnesium protoporphyrin methyl ester are probably located in the chloroplast envelope in association with membrane-bound Mg chetalase and extraplastidic S- adenosylmethionine (Crawford et al., 1982; Fuesler and Cas- telfranco, 1983). It is possible that some process such as the importation or processing of the 32-kDa LHCP precursor is
affected by the presence of magnesium protoporphyrin methyl ester in the chloroplast envelope and this, in turn, somehow influences the stability of the LHCP message. In this respect, it is interesting that in other systems, heme compounds affect the importation and processing of certain nuclear-encoded, mitochondrial-protein precursors. In yeast, the organellar up- take of precytochrome c l is dependent on the presence (not the absence) of heme compounds in the mitochondria (Gasser et a i , 1982; Henning et al., 1983). In chick embryo hepatocytes in culture, the importation of cytosolic-synthesized L-amino- levulinate synthase precursor is blocked by added heme (Sri- vastava et aL, 1983).
The hybridization probe we have used in this study recog- nizes the gene encoding the major 32-kDa precursor protein of the LHCP complex. As yet, we do not know how many other genes in C. reinhardi encode the other polypeptides of the light-harvesting complexes. The complexes are composed of several closely related polypeptides ranging in size from about 23-30 kDa (Delepelaire and Chua, 1981; Shepherd et al., 1983). The 32-kDa precursor gives rise to one or more of the major polypeptides in the 23-25-kDa range (Shepherd et al., 1983). In higher plants such as petunia, the genes encoding the major chlorophyll-binding proteins represent a small mul- tigene family (Dunsmuir et al., 1983). The hybridization probe used in this study, pHS16, hybridizes to only a single DNA band on blots of restriction-digested C. reinhardi DNA and is derived largely from the 3’ untranslated portion of the 32- kDa LHCP gene.3
REFERENCES Allen, J . F., Bennett, J., Steinback, K. E., and Arntzen, C. J . (1981)
Apel, K. (1979) Eur. J. Biochem. 97,183-188 Apel, K., and Kloppstech, K. (1980) Planta (Berl.) 150,426-430 Ares, M. (1982) Thesis, University of California, San Diego Arnon, D. J. (1949) Plant Physiol. 24, 1-15 Bailey, J. M., and Davidson, N. (1976) Anal. Biochem. 7 0 , 7 5 4 5 Beale, S. I. (1971) Plant Physiol. 48 , 316-319 Beale, S. I., and Castelfranco, P. A. (1974) Plant Physiol. 5 3 , 291-
Bennett, J. (1979) FEBS Lett. 103,342-344 Bennett, J., Steinback, K. E., and Arntzen, C. J.. (1980) Proc. Natl.
Burnham, B. F., and Lascelles, J. (1963) Biochem. J. 87,462-472 Castelfranco, P. A., and Beale, S. I. (1983) Annu. Reu. Plant PhYsiol.
Charney, P., and Maniatis, T. (1983) Science (Wash. D. C.) 220,
Chua, N.-H., and Gillham, N. W. (1977) J. Cell Bioi. 74,441-452 Crawford, M. S., and Wang, W.-Y. (1983) Plant Physiol. 71.303-306 Crawford. M. S.. Wane. W.-Y., and Jensen. K. G. (1982) Mol. Gen.
Nature (Lord.) 291,25-29
296
Acad. Sci. U. S. A. 77,5253-5257
34,241-278
1281-1283
~~ Y l
Genet. i88,1:6 Cuming, A. C., and Bennett, J. (1981) Eur. J. Biochem. 118, 71-80 Deleaelaire. P., and Chua, N.-H. (1981) J. BWl. Chem. 2 6 6 , 9300-
9307 . .
Duggan, J., and Gassman, M. (1974) Plant Physiol. 53, 206-215 Dunsmuir, P., Smith, S. M., and Bedbrook, J. (1983) J. Mol. Appl.
Fuesler, T. P., and Castelfranco, P. A. (1983) Plant Physiol. 72 , 57 Gasser, S. M., Ohashi, A., Daum, G., Bohni, P. C., Gibson, J., Reid,
G . A,. Yonetani. T.. and Schatz. G . (1982) Proc. Natl. Acad. Sci. U.
Gen. 2, 285-300
S. A.’79, 267-271 Gough, S. P., Girnth, C., and Kannangara, C. G. (1981) in Proceedings
of the 5th International Congress on Photosynthesis (Akoyunoglou,
, ,
G., ed), Vol. V, pp. 107-116, Balaban International Science Serv- ices, Philadelphia, PA
Granick, S. (1961) J . Biol. Chem. 236 , 1168-1172
P. Imbault, U. Johanningmeier, and S. H. Howell, manuscript in preparation.

Light-harvesting Chlorophyll-binding Protein mRNA Regulation 13549
Guarente, L., and Mason, T. (1983) Cell 32, 1279-1286 Hennig, B., Koehler, H., and Neupert, W. (1983) Proc. Natl. Acad.
Hoober, J. K. (1970) J . Biol. Chem. 2 4 5 , 4327-4334 Hoober, J. K. (1981) in Proceedings of the 5th International Congress
on Photosynthesis (Akoyunoglou, G., ed) Vol. V, pp. 859-866, Balaban International Science Services, Philadelphia, PA
Sci. U. S. A. 80,4963-4967
Hoober, J. K., and Stegeman, W. J. (1973) J. Cell Biol. 5 6 , 1-12 Hoober, J. K., and Stegeman, W. J. (1976) J. Cell Biol. 7 0 , 326-337 Hoober, J. K., Marks, D. B., Keller, B. J., and Margulis, M. M. (1982)
Iwanij, V., Chua, N.-H., and Siekevitz, P. (1975) J. Cell Bwl. 64,572-
Janero, D. R., and Barrnett, R. (1981) J. Cell Biol. 9 1 , 126-134 Owens, G. C., and Ohad, I. (1982) J. Cell Biol. 9 3 , 712-718 Porra, R. J., Irving, E. A., and Tennick, A. M. (1972) Arch Biochem.
Sager, R. (1961) Carnegie Zmt. Washington Year Book 60,374-376 Schmidt, G. W., Bartlett, S. G., Grossman, A. R., Cashmore, A. R.,
J. Cell Biol. 95, 552-558
585
Biophys. 148 , 37-43
and Chua, N.-H. (1981) J. Cell Biol. 91,468-478
Sci. U. S. A. 6 6 , 174-180
107
Schor, S., Siekevitz, P., and Palade, G. E. (1970) Proc. Natl. Acad.
Shepherd, H. S., Ledoigt, G., and Howell, S . H. (1983) Cell 32, 99-
Slovin, J. P., and Tobin, E. M. (1982) Planta (Berl.) 154, 465-472 Srivastava, G., Borthwick, I. A., Brooker, J. D., Wallace, J. C., May,
B. K., and Elliot, W. H. (1983) Biochem. Biophys. Res. Commun.
Stiekema, W. J., Wimpee, C. F., Silverthorne, J., and Tobin, E. M.
Vlcek, L. M., and Gassman, M. L. (1979) Plant Physiol. 64,393-397 Wahl, G., Stern, M., and Stark, G. (1979) Proc. Natl. Acad. Sci. U. S.
Wang, W.-Y., Wang, W. L., Boynton, J. E., and Gillham, N. W.
Wang, W., Boynton, J . E., and Gillham, N. W. (1975) Cell 6, 75-84 Warnick, G. R., and Burnham, B. F. (1971) J. Biol. Chem. 246,
117,344-349
(1983) Plant Physiol. 72 , 717-724
A. 76,3683-3687
(1974) J . Cell Biol. 63,806-823
6880-6885
Related Documents











