73 Movement Models from Sports Provide Representative Task Constraints for Studying Adaptive Behavior in Human Movement Systems Keith Davids 1 , Chris Button 1 , Duarte Araújo 2 , Ian Renshaw 3 , Robert Hristovski 4 1 School of Physical Education, University of Otago, New Zealand 2 Faculty of Human Kinetics, Technical University of Lisbon, Portugal 3 Division of Sport and Recreation, Faculty of Health, Auckland University of Technology, New Zealand 4 Faculty of Physical Culture, University of St Cyril and Methodius, Republic of Macedonia Researchers studying adaptive behavior in human movement systems have traditionally employed simplified, laboratory-based movement models in an effort to conserve experimental rigor. Brunswi- kian psychology raises questions over the representativeness of many of these popular experimental models for studying how movements are coordinated with events, objects, and surfaces of dynamic environments. In this article we argue that sports provide rich ecological constraints for representative task design in modeling the complex interactions of human performers with their environments. Adopting a functionalist perspective enriched by ideas from ecological psychology and nonlinear dynamics, we consider data from exemplar movement models in basketball and boxing to support this contention. We show that this preference for movement models from sports, although not completely novel, has accelerated over recent years, mainly due to the theoretical re-emphasis on studying the interaction of individual and task constraints. The implications of using such applied models of move- ment behavior in studying the design of natural and artificial systems are also discussed. Keywords movement coordination and control · constraints · representative design · perception and action · degrees of freedom 1 Introduction As Seth and Edelman (2004) have argued, in order to understand adaptive behavior, it has become increasingly clear that the interactions between complex organisms, task constraints, and environments need to be studied. For this reason, sub-disciplines such as robotics and artificial intelligence are drawing inspiration from research on how biological movement systems interact with complex environments. Traditional models of robotic design, based on “sense, model, plan, act” prin- ciples, are being complemented by “behavior-based con- trol” approaches with the goal of engineering machines to operate in unpredictable and challenging environ- ments, such as during space exploration or playing in robocup soccer competitions (Mataric, 1998). Alterna- Copyright © 2006 International Society for Adaptive Behavior (2006), Vol 14(1): 73–95. [1059–7123(200612) 14:1; 73–95; 063050] Figures 5–7 appear in color online: http://adb.sagepub.com Correspondence to: Keith Davids, School of Physical Education, University of Otago, 46 Union Street West, Dunedin, New Zealand. E-mail: [email protected] Tel.:+6434798994 Fax: +6434798309

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

73
Movement Models from Sports Provide
Representative Task Constraints for Studying
Adaptive Behavior in Human Movement Systems
Keith Davids1, Chris Button1, Duarte Araújo2, Ian Renshaw3, Robert Hristovski41School of Physical Education, University of Otago, New Zealand2Faculty of Human Kinetics, Technical University of Lisbon, Portugal3Division of Sport and Recreation, Faculty of Health, Auckland University of Technology, New Zealand4Faculty of Physical Culture, University of St Cyril and Methodius, Republic of Macedonia
Researchers studying adaptive behavior in human movement systems have traditionally employedsimplified, laboratory-based movement models in an effort to conserve experimental rigor. Brunswi-
kian psychology raises questions over the representativeness of many of these popular experimental
models for studying how movements are coordinated with events, objects, and surfaces of dynamicenvironments. In this article we argue that sports provide rich ecological constraints for representative
task design in modeling the complex interactions of human performers with their environments.
Adopting a functionalist perspective enriched by ideas from ecological psychology and nonlineardynamics, we consider data from exemplar movement models in basketball and boxing to support this
contention. We show that this preference for movement models from sports, although not completely
novel, has accelerated over recent years, mainly due to the theoretical re-emphasis on studying theinteraction of individual and task constraints. The implications of using such applied models of move-
ment behavior in studying the design of natural and artificial systems are also discussed.
Keywords movement coordination and control · constraints · representative design · perception and
action · degrees of freedom
1 Introduction
As Seth and Edelman (2004) have argued, in order tounderstand adaptive behavior, it has become increasinglyclear that the interactions between complex organisms,task constraints, and environments need to be studied.For this reason, sub-disciplines such as robotics andartificial intelligence are drawing inspiration from
research on how biological movement systems interactwith complex environments. Traditional models ofrobotic design, based on “sense, model, plan, act” prin-ciples, are being complemented by “behavior-based con-trol” approaches with the goal of engineering machinesto operate in unpredictable and challenging environ-ments, such as during space exploration or playing inrobocup soccer competitions (Mataric, 1998). Alterna-
Copyright © 2006 International Society for Adaptive Behavior(2006), Vol 14(1): 73–95.[1059–7123(200612) 14:1; 73–95; 063050]Figures 5–7 appear in color online: http://adb.sagepub.com
Correspondence to: Keith Davids, School of Physical Education, University of Otago, 46 Union Street West, Dunedin, New Zealand. E-mail: [email protected] Tel.:+6434798994 Fax: +6434798309

74 Adaptive Behavior 14(1)
tive design principles, including “situated robotics,”are informed by research on how biological movementsystems search for and pick up information to supportsophisticated interactions with their environments (e.g.,Di Paolo, 2002; Paine & Tani, 2005).
However, as Robert Rosen (1978) has proposed,the question of design in complex biological systems isof a different order of magnitude compared to machineengineering. Unlike artificial devices, there are non-trivial differences between biological structure andfunction and the challenge of capturing adaptive func-tional behaviors makes designing a complex biologicalsystem from a multitude of individual components animpossible task. According to Mikulecky (2000) evolu-tionary design principles target functional componentsof organisms, which are not permanent static featuresof biological systems but dynamically constrained bycontext. This view is harmonious with some “behavior-based” robotic implementations that have been foundedon ecological principles in which an agent and envi-ronment are viewed as an inseparable system studiedat an ecological scale of analysis. In “ecological robot-ics,” adaptive behavior is specified by information inthe environment which does not need to be representedinternally in explicit detail (Fajen, Warren, Temizer, &Kaelbling, 2003), and there have been attempts to foregoa central planner for behaviors and to map sensory sys-tems directly to actuators (Duchon & Warren, 2002).
1.1 The Role of Movement Models from Sport in Studying Adaptive Behavior
The problem of understanding how robots can adapttheir behaviors to interact with dynamically changingconditions has parallels in the study of how humansassemble functional, goal-directed movement behaviorssuch as prehension, locomotion and interceptive actions.In this position paper we discuss how the study ofhuman movement systems can provide meaningfultheoretical insights into how behaviors can be adaptedto relevant constraints on action during interactionswith complex environments. In particular, we notehow, increasingly, the experimental models used bymany psychologists and movement scientists to studyadaptive behavior of complex movement systems arebeing provided by a rich range of sports and physicalactivities (e.g., Davids, Savelsbergh, Bennett, & Vander Kamp, 2002). Accordingly, we explore the propo-sition that sports provide an important performance
context with relevant experimental movement modelsfor studying a wide range of constraints on human move-ment systems. We argue that movement models insports readily lend themselves to the “representativedesign” of tasks for studying key processes in adaptivebehavior such as perception, decision-making, inten-tionality and action.
Although technological advances have also impactedon the selection of movement models for studyingcoordination and control, complex movement modelsfrom sports are gaining in popularity because of thepowerful influence of a theoretical framework sug-gesting that movement coordination is best viewed asadaptive, emergent behavior which self-organizes underconstraints, particularly informational and cognitiveconstraints. Preference by movement scientists and psy-chologists for more complex experimental models fromsports has accelerated rapidly in recent years and insightsinto the adaptive nature of movement behavior havebeen provided by activities such as playground swinging(Post, Peper, & Beek, 2003), pole-balancing (DeGuz-man, 2004), hula-hooping (Balasubramaniam & Tur-vey, 2003), sit-ups (Cordo & Gurfinkel, 2004), basketballfree-throw shooting (Oudejans, et al., 1996), javelinand discus throwing (Schöllhorn, 2003), rowing (Daf-fertshofer, Huys, & Beek, 2004; Shuttleworth, 2004),sailing (Araújo, Davids, & Serpa, 2005), cascade jug-gling (Post, Daffertshofer, & Beek, 2000; Huys, Daf-fertshofer, & Beek, 2004; Haibach, Daniels, & Newell,2004), pistol shooting (Scholz, Schoner, & Latash, 2000),volleyball serving (Handford, 2002), long jumpingrun-ups (Montagne, Cornus, Glize, Quaine, & Lau-rent, 2000), ball catching and hitting (Bennett, van derKamp, Savelsbergh, & Davids, 2000; Scott, van derKamp, Savelsbergh, Oudejans, & Davids, 2004), soccerjuggling (Tlili, Mottet, Dupuy, & Pavis, 2004), basket-ball ball bouncing (Broderick, Pavis & Newell, 2000),ball-racket bouncing (Sternad & Dijkstra, 2004), golfdriving (Knight, 2004), pedalo paddle boating (Chen,Liu, Mayer-Kress, & Newell, 2005), and front crawlswimming (Satkunskiene, Schega, Kunze, Birzinyte,& Daly, 2005).
Traditionally, selection of movement models toinvestigate motor system functioning has been biasedaway from dynamic, multijoint, functional actions prev-alent in sports because of the reductionist view thatexperimental rigor could be better maintained in labo-ratory studies of simple movements. Experimentalmovement models created in laboratory settings tend

Davids et al. Representative Task Design and Movement 75
to involve fewer motor system degrees of freedomthan sports movements, have a limited range of scalingfor key variables such as spatial and temporal con-straints, include a lower level of intentional constrainton behavior and typically have a reduced amount ofperceptual information needed to regulate action (New-ell, 1989; Davids, Kingsbury, Bennett, & Handford,2001; Wulf & Shea, 2002). The distinction betweenexperimental rigor and field research is now being rec-ognized as a false dichotomy, at least in part due to theinfluence of a functionalist theoretical paradigm onmovement coordination, dominated by ecological psy-chology (e.g., Brunswik, 1956; Gibson, 1979) andnonlinear dynamics (e.g., Jirsa & Kelso, 2004). Thistheoretical paradigm has been characterized as a “con-straints-based framework” emphasizing the study ofmovement behavior as an emergent property of thecontinuous interactions of a biological movement sys-tem and its environment (Newell, 1986; Davids, Araújo,Shuttleworth, & Button, 2003). Whilst it is clearlypossible to explain adaptive movements from a varietyof theoretical standpoints, including representationalaccounts (e.g., Rieger, Knoblich, & Prinz, 2005), aconstraints-based perspective is a popular biologi-cally inspired approach which is harmonious with thetenets of “situated” and “ecological” robotics and theiremphasis on enhancing our understanding of the con-straints of natural law on cognition (Duchon & War-ren, 2002).
1.2 Representative Task Design: Insights of Egon Brunswik
Egon Brunswik’s (1956) theoretical insights in eco-logical psychology on the adaptive nature of humanpsychological processes also provide a case for rejectingthe view of laboratory tasks as diametrically opposedto field based research designs for studying humanmovement systems. The issue of task constraints andtheir effect on behavior has often been mistakenly pre-sented in the psychology literature as an issue of dif-ferences in ecological validity (misconceived asexperimental design realism) involving contrived, lab-oratory tasks versus natural tasks. Brunswik (1956)showed how this interpretation of scientific method-ology was a false dichotomy, based on a misunder-standing of the concept of task “representativeness.”He defended the view that perceptual variables shouldbe sampled from an organism’s natural environment
so as to be representative of the typical environmen-tal stimulation to which it had adapted and whichformed the focus of an experimenter’s generalisation.Brunswik’s (1956) contribution to understanding offunctional movement behavior has been immense inmany respects including: (i) promoting idiographic anal-yses of how individuals solve motor problems (i.e.,examining unique functional adaptations of individu-als to the environment); and (ii), stressing design ofspecialized experimental contexts with a high level ofrepresentativeness of a biological movement system’sniche habitat. The generalization of findings outsideof specific experimental contexts is still recognized asproblematic in many different areas of psychology,including educational, social, developmental and exper-imental fields. These issues are also significant inunderstanding how artificial and biological systemsadapt to dynamic environments.
To support our theoretical analysis, we highlightextant data from our program of research in the sportsof basketball and boxing. In these experiments westudied adaptation of human movement systems toother individuals and important objects in the environ-ment during tasks such as dribbling a ball and hitting atarget, respectively (for recent examples see Araújo etal., 2005b; Hristovski, Davids, & Araújo, in press).We start by briefly overviewing key concepts from non-linear dynamics and ecological psychology for study-ing movement coordination and control. In particular,we discuss in more detail the idea of task representa-tiveness, defined earlier, which provides a strongrationale for selection and design of experimental taskconstraints based on sport movements.
1.3 The Theoretical Impetus of Nonlinear Dynamics and Ecological Psychology
Prominent ideas in nonlinear dynamics and ecologicalpsychology have been integrated into a constraints-based framework for developing understanding of howmovements are coordinated with respect to complexand dynamical environments (for detailed overviewssee Araújo, Davids, Bennett, Button, & Chapman, 2004;Davids et al., 2001; Williams, Davids, & Williams,1999). A functionalist perspective in both areas hascharacterized biological movement systems as com-plex, adaptive systems. A key focus concerns how themany interacting system components, or motor sys-tem degrees of freedom, are coordinated and control-

76 Adaptive Behavior 14(1)
led during goal-directed movements (see Bernstein,1967; Berthouze & Lungarella, 2004). Research hasrevealed that patterns emerge between parts of dynami-cal movement systems through processes of self-organ-ization ubiquitous to physical and biological systemsin nature (for reviews see Davids et al., 2003; Guerin& Kunkle, 2004). It has been revealed how dynamicalmovement systems exploit surrounding constraints toshape the functional, self-sustaining patterns of behav-ior that emerge in specific performance contexts. Indynamical movement systems, information plays animportant role in constraining the number of biome-chanical degrees of freedom of the motor system thatthe central nervous system (CNS) needs to regulateduring functional behavior. Rather than controllingeach degree of freedom separately during goal-directedmovement, the CNS exploits self-organization in move-ment systems to form temporarily-assembled musclecomplexes called coordinative structures. Coordinativestructures are functional synergies that emerge betweenparts of the body used to achieve movement goalssuch as locomotion, reaching and grasping, and hitting.Adaptive behavior in dynamical movement systemshas been studied by focusing on “phase transitions”(movements of system components into a differentmacro-state of organization). Phase transitions betweenpatterns of coordination in biological movement sys-tems are considered highly relevant for study becausethey exemplify the emergent nature of behavior.
Ecological psychologists have also addressed theproblem of how perceptual information guides actionsin natural environments, by emphasizing how biologi-cal movement systems are surrounded by huge banksof energy (e.g., light reflected from objects and surfacesor emitted sound waves from an event or another indi-vidual). These energy arrays are highly structured andcan act as information constraints to support decisionmaking, planning, and organization of action. Gibson(e.g., 1979) argued that specific movements of a per-former and/or environmental objects to be acted uponled to the pick up of these key information sources foradapting actions. Movements cause changes to energyflows that provide specific information to organismson the properties of a dynamic environment leading tolawful relations between a kinetic property of a move-ment and the kinematic property of the surroundingperceptual flow. Because flow patterns are specific toparticular environmental properties, they can act asinvariant information variables to be picked by indi-
vidual performers to constrain their actions within spe-cific contexts.
Ecological constraints regulate movement coordi-nation and it has become evident that task constraintsinteract with informational constraints, sometimesoverriding the influence of important structural con-straints in human movement systems. For example, anumber of studies of rhythmical finger movements havereported that neuroanatomical constraints of musclesinvolved in coordination have a strong influence onstability observed (for reviews see Carson, 2004; Car-son & Kelso, 2004). In these studies the basic move-ment model required participants to flex and extend anindex finger either synchronous or syncopated withacoustic information provided by a metronome. Fingerflexions synchronous with the acoustic beat werefound to be more stable than extension movementssyncopated with the beat. It was proposed that evolu-tionary constraints on the neuroanatomical system hada profound influence on differential stability of flexorscompared to extensors (Carson, 2004). Flexor musclesoriginated phylogenetically as anti-gravity musclesand have evolved to produce relatively stronger forcesfrom similar numbers of motor units. However, theseeffects of neuroanatomical constraints can be overrid-den by the addition of informational constraints so thatneither flexion nor extension phases of the rhythmicoscillatory finger movements are more stable. In onestudy a small padded plastic stop was inserted into thetypical rhythmic finger movement experimental proto-col to interrupt the trajectory of participants’ indexfingers as they were rhythmically syncopated or syn-chronized to a metronomic beat. The stop providedadditional haptic information to constrain flexion andextension movements (Kelso, Fink, Delaplain, & Car-son, 2001). Data suggested that afferent informationfrom contact of the finger with the haptic stop deviceoverrode the neuroanatomical design constraints of themetacarpophalangeal joints during rhythmical fingerflexion and extension movements. These findings showhow constraints interact during many natural activitiesand underline why movement models from sportscould provide rich task vehicles for studying adaptivebehaviors, exemplifying how coordinative structuresare soft-assembled and tuned to prevailing task condi-tions (Kugler & Turvey, 1987).
The constraints-led perspective has considerableimplications for understanding variability, traditionallythought of as noise or error in biological movement

Davids et al. Representative Task Design and Movement 77
systems. Rather, instances of variability in movementpatterns, exemplified by fluctuations in stability, maymore accurately reflect flexible and adaptive motorsystem behavior during exploration of performancecontexts. This theoretical position is in line with recentattempts to decompose observed movement systemvariability into separate components which are func-tional and stochastic (e.g., Riley & Turvey, 2002; But-ton, McLeod, Sanders, & Coleman, 2003; Rosengren,Savelsbergh, & Van der Kamp, 2003; Müller & Ster-nad, 2004). This paradox between stability and varia-bility explains why skilled individuals are capable ofboth persistence and change in motor output duringperformance (Davids et al., 2003). The radical impli-cation of these ideas is that perceptions, memories,intentions, decisions or actions may be conceived of asemergent, self-organizing, macroscopic patterns formedas individuals adapt to the ecological constraints oftheir environments (see van Rooij, Bongers, & Hase-lager, 2002; Williams et al., 1999). These ideas havesome parallels in the theoretical views of Egon Brunswik(1956) who viewed psychological processes as adapta-tions to complex environments, effectively openingthe window on to movement models from sports andexemplifying why they have gained such rapid popu-larity in recent years.
1.4 Task Representativeness and a Constraints-Led Perspective
Brunswik (1956) drew attention to some major con-cepts—representative design and intra-ecological cor-relation—which can be understood with reference tothe emergence of movement coordination and controlunder constraints. Representativeness refers to thegeneralization of task constraints in specific researchsettings to performance constraints outside the experi-ment. Brunswik (1956) noted the need for congruencebetween experimental conditions for controlled studyof a particular aspect of behavior and the environmen-tal conditions of performance. The principle of taskrepresentativeness is predicated on the accurate sam-pling of environmental conditions for the design of aresearch study on a specific characteristic of behavior.Brunswik (1956) also suggested that, to be representa-tive, environmental constraints of a “to-be-studiedbehavior” need to interact, as they do in many con-texts of performance. He labeled this feature of exper-imental design ecological intercorrelation. His ideas
on sampling environmental conditions pre-empted thepopularization of the current strategy of focusing onthe relationship or intercorrelations between key vari-ables in a behavioral task. The more extensive thesample of constraints, the more representative was theexperimental task design. As in other areas of ecologi-cal psychology, Brunswik’s (1956) attention to theconcept of task representativeness was focused onenhancing the study of the organismic behavior rela-tive to environmental constraints. The critical role ofexperimental task constraints, exemplified in the ear-lier discussion on interaction with flexion/extensionphases of finger movements, points to the significanceof the principle of ecological intercorrelation in taskrepresentativeness.
In this way, Brunswik (1956) contributed the strongconceptualization of representative task design to thetraditional dichotomization of empirical work as “lab-oratory versus field-based.” Clearly, understanding ofthe interaction between key organismic, task, and envi-ronmental constraints for the emergence of adaptivebehavior provides a powerful theoretical frameworkfor designing representative tasks, regardless of whetherthey are located in a laboratory or field setting.
So, what are the key principles of a constraints-led perspective which could inform the design of rep-resentative tasks? How are these principles highlightedin sports tasks? First, a constraints-led perspectiveemphasizes understanding of the ecological constraintson behavior and rejects any attempt to provide unilat-eral explanations of mind, body, and environment. Asin the implementation of ecological robotics a sys-tems-based relationship between an organism and itsenvironment is advocated. An important implicationof this theoretical commitment for empirical researchon processes of perception, decision-making, and action,is that representative task designs are needed to exam-ine the role of ecological constraints during active taskexploration, i.e., functional adaptive behaviors need tobe investigated at the level of the organism–environ-ment relationship. Second, the primacy of perceptionis recognized in a functionalist view of knowledge.Task design needs to ensure that perceptual systemsare allowed to function as they were designed in orderto pick up information for movement. Use of movementmodels from sports is a powerful method for achiev-ing this goal of research. Third, the adaptation oforganisms to their environments needs to be explainedthrough the complex interactions between perception,

78 Adaptive Behavior 14(1)
intentions, and actions. Fourth, information and move-ment become functionally coupled to support adaptivebehaviors, an important principle for designing repre-sentative tasks for experimentation and training.
These principles receive support from recenttrends in ecological psychology aimed at exploring theintegration of ideas from two founding figures: EgonBrunswik and James J. Gibson (Vicente, 2003; Davids,Bennett, & Newell, 2006; Araújo, Davids, & Passos,2005a). A broad ecological psychology begins by ana-lyzing the ecological constraints on behavior, beforeaddressing the organism’s adaptation (Vicente, 2003).The main aim of a broader ecological psychology is tosample environmental variables in range and distribu-tion to be “representative of a carefully defined set ofconditions” (Brunswik, 1956, page 53). Gibson (1979)demonstrated that a serious analysis of perception asan adaptive process requires a simultaneous analysisof the environment and the information that it affordsor invites in the form of actions. An object affordswhat it does because it has certain specific properties,which are not intrinsic to that object. On the contrary,object properties are taken with reference to agentswho will perceive information about the object orintend to use it for action. To realize these affordanceproperties requires that an agent regulates its activityaccording to information concerning both the objectand itself. Many of the affordances that an agent usesin adapting to its ecology require extensive practiceand learning to be perceived or to be used. Represent-ative experimental design is likely to vary, based onparticipant, task, and environmental constraints (Kugler,Kelso, & Turvey, 1980; Newell, 1986). For this reasonthe constraints-based framework provides a way ofmeasuring the degree of adaptation between an organ-ism’s behavior and the structure of its environment.The interaction of key task and environment constraintsprovides a referent for evaluating the fitness of behav-ior in individual performers. The empirical datareviewed in this paper demonstrate that relatively sim-ple changes in task constraints can provide powerfulinsights into the adaptive coordinative structures thatemerge as individuals find functional coordinationsolutions. They also show that sports abound with avariety of unique task constraints and that analysis ofmovement coordination solutions after manipulatingtask constraints provide particularly useful windowson the role of specific intentions and perceptual infor-mation sources in constraining movements.
1.5 Representativeness of Task Constraints and Research on Perception and Action
An integration of Gibsonian and Brunswikian ideas ina broader ecological psychology also alerts us to theidea that in order to be representative, an experimentaltask should be set up in such a way that the actor canperceive in order to act, but also can act in order toperceive (Gibson, 1979). Gibson (1957) deplored thedesign of experimental contexts that were convenientfor investigators, but relatively alien for participantswho typically use exploratory actions to seek and useperceptual information to guide goal-directed behavior.This is necessary because actions by which cognitionis expressed require that information about environ-mental facts be referential to the energy for behavingwith respect to those facts (Turvey & Shaw, 1995).A clear finding in the literature on expertise and inanticipation and decision making in sport is thatunrepresentative experimental task constraints tend toundermine the expert’s performance advantage incomparison with less skilled performers (Abernethy,Thomas, & Thomas, 1993). There are a number of rea-sons for this observation in the literature, with perhapsthe most important relating to experimental designswhich restrict access of experts to key informationsources which are normally available during perform-ance (Williams et al., 1999). For example, Oudejans,Michaels, Bakker, & Dolné, (1996) found that expertbaseball outfielders could only be distinguished fromnon-baseball players when the experimental task ofcatching allowed them to act on ball-flight informa-tion, and not just react to the sudden appearance ofball-flight information. Additionally, experimental taskconstraints designed to elicit movement responsesrather than verbal responses are better able to distin-guish expert performers from novices in sport (Wil-liams et al., 1999).
To exemplify these arguments, in this section wediscuss research on dynamic interceptive actions illus-trating how subtle alterations in task constraints canlead to adaptive behavior by participants and underlin-ing how sports tasks can enhance representativenessin studying adaptive movement behaviors. There are anumber of key differences between traditional labora-tory methodologies for studying processes of percep-tion and movement, and the task constraints of naturalinterceptive movements such as catching, batting orrunning towards a target area (Davids et al., 2002).

Davids et al. Representative Task Design and Movement 79
Some traditional experimental contexts for studyingperception have been characterized by “informational-minimalism” because of a reductionist belief that hold-ing a huge number of variables constant could allowinvestigators to produce laws of perception (Harris &Jenkin, 1998). For example, an inordinate emphasison use of simulated motion has led to the popularity ofcoincident timing tasks in which participants have totypically predict when a (suddenly occluded) object/image will arrive at a designated target point in space.The task constraints of motion prediction paradigmsemphasize the significance of perceptual anticipationprocesses in studying how predictive visual timing proc-esses are used for interception. The assumption is thatparticipants can “represent” part of the approach tra-jectory of an object image in short term memory topredict its time to arrival at a target location (e.g., seeTodd, 1981). This traditional emphasis in designingstudies to investigate predictive visual processes fittedwell with theories of hierarchically organized percep-tual and movement systems (see Poulton, 1957; Wil-liams et al., 1999). From this theoretical standpoint,the performer is highly dependent on accurate percep-tion of spatio-temporal characteristics of projectilemotion in order to program the movement of an effec-tor towards a predicted future contact point.
However, it has been argued that this type of designmay not be representative of many natural interceptiveactions which emphasize receptor anticipation proc-esses (Poulton, 1957; Davids et al., 2001). Under theconstraints of natural interceptive tasks like ball catch-ing or hitting, participants are usually able to continu-ously regulate movements by viewing the ball until itarrives at the location of the hands or the bat. They donot need to perceptually construct the ball’s trajectoryfrom earlier remembered information from ball flight.This view of the task constraints of natural inter-ceptive actions fits well with the concept of informa-tion–movement coupling in ecological psychology,promoting a strategy of prospective movement con-trol, rather than predictive visual timing (Beek, Jacobs,Daffertshoffer, & Huys, 2003b). Prospection exempli-fies a heterarchical view of the performer, and a highlyintegrated, functional relationship between movementand perceptual systems (for other examples of heterar-chical control models see Hill & Raab, 2005). In eco-logical psychology, it involves a close and continuouscoupling of movement and perceptual systems based onthe relationship between the instantaneous states of
the performer and environment during task perform-ance (Montagne, Laurent, Durey, & Bootsma, 1999).
1.6 Coupling Perception and Action in Experiments on Interceptive Actions
The different constraints of motion prediction and nat-ural interceptive tasks have signalled the existence ofdifferent control mechanisms for successful comple-tion of each type of task, exemplifying the relevanceof Brunswik’s (1956) concept of task representative-ness. For example, slower velocities are typically usedin perceptual anticipation tasks (>1 s) which mightafford the perceptual construction of the stimulus tra-jectory for prediction of future contact points. In mostnatural interceptive actions, however, movement exe-cution times are usually more brief [e.g., 300 ms forcatching balls traveling at 10 m/s velocity with onehand (Alderson, Sully, & Sully, 1974)], facilitating thedevelopment of strategies of continuous regulation orperception-movement coupling (Tresilian, 1995).
Extant data on movement outcome variability alsosuggest that performers find the task constraints of nat-ural interceptive actions more functionally relevantthan psychophysically based experiments (in line withWoodworth, 1938). For example, in some studiesinvolving computer simulations, participants underes-timated time to arrival of a stimulus object at a desig-nated location point on monitor screens. The amountof underestimation increased with increasing time toarrival (e.g., see data from Schiff & Detwiler, 1979;Schiff & Oldak, 1990). Tresilian (1994) has calculatedthat the average underestimate of reported time toarrival in these tasks is around 60% of actual time toarrival at the point of execution of the button press,with the SD (standard deviation) of the estimates being50%. These error margins and levels of outcome vari-ability suggest that participants do not find these taskconstraints functionally relevant. In contrast, data ontiming behavior in adults and infants suggest that thelarge amount of variability in estimated time to arrivalis often not observed in performance of natural taskssuch as fast interceptive actions (see section below onspatio-temporal constraints of interceptive move-ments documenting the results of several studies ofinterceptive actions in sport). Tresilian (1995) high-lighted the fact that “... the variability (standard deviationof response times) of responses in CA [coincidenceanticipation] tasks is some five or six times greater

80 Adaptive Behavior 14(1)
than that observed in IAs [interceptive actions] per-formed under the same stimulus conditions.…”
This observation demonstrates Bootsma’s (1989)criticism of the “unprincipled” way in which processesof perception and movement have sometimes beenstudied separately. Bootsma (1989) examined effects onperformance of de-coupling information and move-ment under three different types of task constraints.Participants in his study were presented with squashballs dropped through a plastic tube (length = 50 cm,diameter = 4.3 cm) from a height of 270 cm above atable surface. They attempted to intercept the ball inone of three different conditions: (i) using their ownarm and a bat; (ii) pressing a button to release a 55-cmmechanical arm to hit the ball just before it landed onthe table top; and (iii) pressing a button when the ballwas judged to be level with the table-top surface (equiv-alent to motion prediction task constraints).
These conditions represented a successive degra-dation in information–movement coupling, and percent-age accuracy data on hitting performance under natural-arm task constraints (82.4%) and artificial-arm con-straints (49.5%) were significantly different. Further,movement times were more variable for the artificial-arm task constraints (14.6 ms) than the natural-arm taskconstraints (5.0 ms). These findings supported the eco-logical notion of information as “specificational” inregulating behavior under the natural task constraintsof interceptive actions. Variability in timing initiationof striking movements was lower under natural-armconstraints (SD = 16.3 ms) compared to artificial armtask constraints (SD = 34.0 ms). When only perceptualjudgements of interception points were required byparticipants, variability was significantly higher (SD =61.8 ms) than under both arm conditions.
These findings implied that structuring experimen-tal tasks to keep key sources of perceptual informationand actions together seems to be a principle for design-ing representative tasks to study interceptive actions.Interestingly, these findings cannot be attributed to tran-sient effects due to lack of familiarity with task con-straints by participants. Even after a 100-trial learningperiod (including 20 familiarization trials), althoughmean movement initiation time (MIT) in the motionprediction conditions was higher than the natural-armconditions, these observed differences were not statis-tically significant (Davids et al., 2001). In line withBootsma’s (1989) original findings, statistically sig-nificant differences were obtained for the variability
of MIT (operationalized as the SD around the mean)between the two conditions. Variability of MIT wasgreater in the motion prediction conditions comparedto the natural-arm condition.
Bootsma’s (1989) data illustrated the effects of de-coupling perception and movement in studies of inter-ceptive actions and highlight the significance of sportstasks as one important means of designing representa-tive tasks for studying adaptive behavior. When thenumber of trials in the perceptual judgement (i.e., CA)task was quintupled, performance was still significantlybetter in the natural-arm striking condition, as evidencedby variability of movement initiation time. It seemsthat information-movement coupling is a robust prin-ciple of constraints-led perspective, and there is somemerit in the proposal of Bootsma (1989) for recognizing“a movement-dependent use of perceptual information,emphasising the inseparability [of perception and move-ment]” (Bootsma, 1989, page 498). The task constraintsof striking a ball set up the saliency of various sourcesof perceptual information involved in timing the move-ment (Davids et al., 2001). Under these specific task andinformational constraints, the performer develops acoupling of movement to relevant sources of perceptualinformation. As evident from the data from the motionprediction task, the developed coupling cannot easily betransferred to different task constraints (Bootsma, 1989),highlighting the importance of task representativenessfor studying adaptive movement behavior.
To summarize, these findings indicate the extensiveinsights into adaptive behavior that might be availablefrom studying movement models under the enrichedecological constraints found in sports. The constraints-led perspective signals the need to carefully distin-guish variability in movement organization, a healthysign of adaptive behavior in biological movement sys-tems, from variability in movement output which isless functional (Davids, Shuttleworth, Button, Renshaw,& Glazier, 2004). The latter is often caused by the useof task constraints which are unrepresentative, as datareported from studies of hitting actions have shown.Compelling arguments exist for empirical work to ade-quately reflect “enriched action environments” basedon a broad range of task constraints, to ensure thatmovement model selection does not occur to confirm apriori theorizing (Newell, 1991; Cordo & Gurfinkel,2004). In the following section we show how thesports of basketball and boxing provide a rich back-drop for the study of emergent functional behaviors

Davids et al. Representative Task Design and Movement 81
because of the many different movement models avail-able for research.
2 Movement Models in Basketball and Boxing: Windows on Adaptive Movement Behavior
2.1 Dribbling in Basketball
Spontaneous movement variability can play an impor-tant role in allowing performers to create differentsolutions to fit different situations. In a dynamic sportsuch as basketball, one can get a good impression ofhow skilled players utilize a range of coordination pat-terns to achieve individual and team goals. Araújo et al.(2002, 2004) recently examined the agent–agent inter-actions in a dyad formed by competing players in bas-ketball dribbling. In spite of the enormous variabilityand complexity of team ball game situations, theoreti-cal and experimental evidence points to the existenceof patterned interpersonal interactions (Schmidt, Carello,& Turvey, 1990). In physical terms, these processes areexpressed in the emergence of organized structures inphase-space describing the interpersonal interactions ina dribbling dyad. Interpersonal coordination can bestudied in the emergence and regulation of coordi-nated states through inherently perceptual processes,based on the dynamics between individuals in a dyador group (Kugler & Turvey, 1987).
From the theoretical perspective of nonlineardynamics, the behavior of interacting players could beinterpreted as an emergent process resulting from thespatio-temporal relations established during game sit-uations. Two individuals in a dribbling dyad may beconsidered as a single system with dyadic synergy(Schmidt et al., 1990). A dyadic synergy can show non-linear properties, namely entrainment and sustainedperiodic behavior, and specific modes of interpersonalcoordination emerge from contextual, personal, and taskconstraints. Team ball sports can be described as aseries of sub-goals, each constraining the coordina-tion of different players to different extents (Schmidt,O’Brien, & Sysko, 1999). In basketball, the interac-tion of a dribbler and a defender in a one-on-one situa-tion can result in a relatively stable interactive dynamicstructure, since the defender may counteract any move-ment towards the basket by an attacker. According tocoaching knowledge it is this stable balance between
attackers and defenders that characterizes the one-on-one situation in basketball (e.g, Bain, Hayes, & Quance,1978). In this classic game situation the attacker needsto de-stabilize or perturb the stable state of this momen-tary dyad. If this system is successfully de-stabilized,the attacker can dribble past the defender towards thebasket. This de-stabilization corresponds to a symme-try-breaking process where the previous stable inter-personal state transits to a new dynamic state (Kugler& Turvey, 1987). This transition from one orderedstate where the defender has the advantage, to anotherwhere the attacker has the advantage (after dribblingpast the defender) provides an alternative, biologicallyinspired explanation, to the information processingapproach (e.g., Schmidt & Lee, 2005). In dynamicenvironments there is typically not one stimulus to setthe player going (such as when a sprinter reacts to thestarter’s gun), but a constant flux of stimulation avail-able to be picked up from the environment in support ofaction (Reed, 1996; Whiting, 1991). Therefore, althoughpossible, an explanation for a particular aspect of acomplex dyad interaction, such as a fake move by oneplayer, cannot easily be explained by relationshipslike Hick’s law (the relation between the number ofstimulus–response alternatives and reaction time), stim-ulus–response compatibility, or the psychologicalrefractory period. Also, the fact that both players ininteraction exhibit symmetry indicates that one playeris not merely responding to the other, but there is acoupling effect as an emergent property, as predictedby the dynamical systems approach, a more parsimo-nious explanation. A dynamical systems analysis of thejust-described transition process (i.e., coupling anddecoupling of the dyad) needs to begin with a measureof order in the stable interpersonal pattern formed bythe position of the attacker and defender with respectto the ball and the basket.
2.2 A Description of a Methodology for Investigating Interpersonal Dynamics in Team Games
Analysis of coaching literature reveals that an orderparameter (i.e., a collective variable that synthesizesthe relevant coordinated parts of the system as a whole)to describe the organization of an attacker–defendersystem could be the distance between the basket andthe dyad (i.e., medium point of the distance betweenthe attacker and defender) during a 1 v 1 sub-phase of

82 Adaptive Behavior 14(1)
the game (e.g., Bain et al., 1978). A specific controlparameter could be the interpersonal distance betweenattacker and defender. Araújo, Davids, Sainhas, &Fernandes, (2002) examined whether the distancefrom the attacker–defender dyad to the basket wouldbecome less stable (i.e., not maintaining a similar dis-tance from the dyad to the basket) until some criticalvalue of interpersonal distance was reached. This inves-tigation considered whether changes in interpersonaldistance were associated with dribbling success byattackers. A specific issue of interest was whether theattacker–defender dyad became more frequently de-stabilized at critical values of interpersonal distance.Ten players (regional level, 15–16 year-old males) par-ticipated in the experiment after recommendation bytheir coaches and informed consent. They were pairedto form 5 dyads. Each dyad started in the free throwline, with the other members of both teams placed onfield based on the “attack system 1:2:2” (see Figure 1).
The horizontal plane trajectory (two-dimensional)of the mass center of each player in the dyad wasrecorded by one digital camera. Body mass center wascalculated with 6 markers (2 ankles, 2 hips, and 2shoulders). Task instructions were for the attacker toscore and the defender to prevent a score, within therules of basketball. The eight other players started toparticipate in the play five seconds after the beginningof the task (temporal task constraints), after having beingplaced in valid positions based on typical basketballmatch strategy (spatial task constraints). The actionsequence started when the defender passed the ball tothe attacker, after a signal from experimenters. Theaction sequence was completed when the attacker shotthe ball at the basket or when the defender took it away
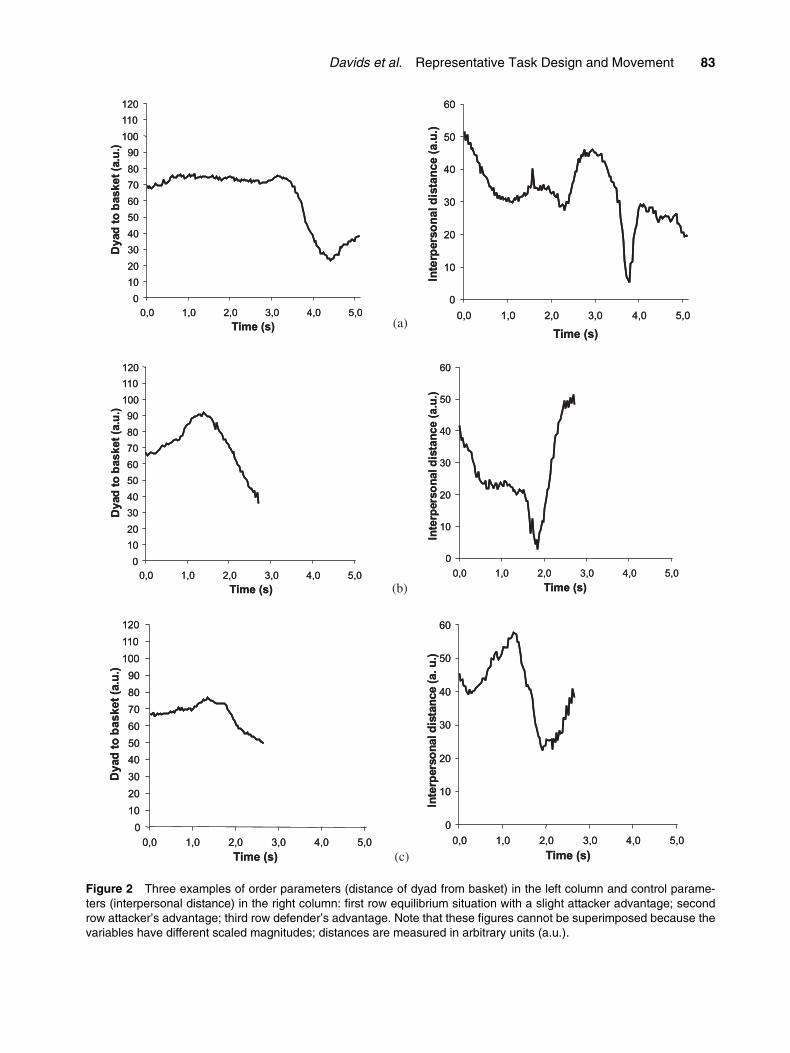
from the attacker. From Figures 2a and 3a, it can beobserved that during the initial part of the dyadic entrain-ment there is a stable state of the order parameter, fol-lowed by an abrupt change in the state of the systemdue to an attacker’s success in de-stabilizing the dyad.
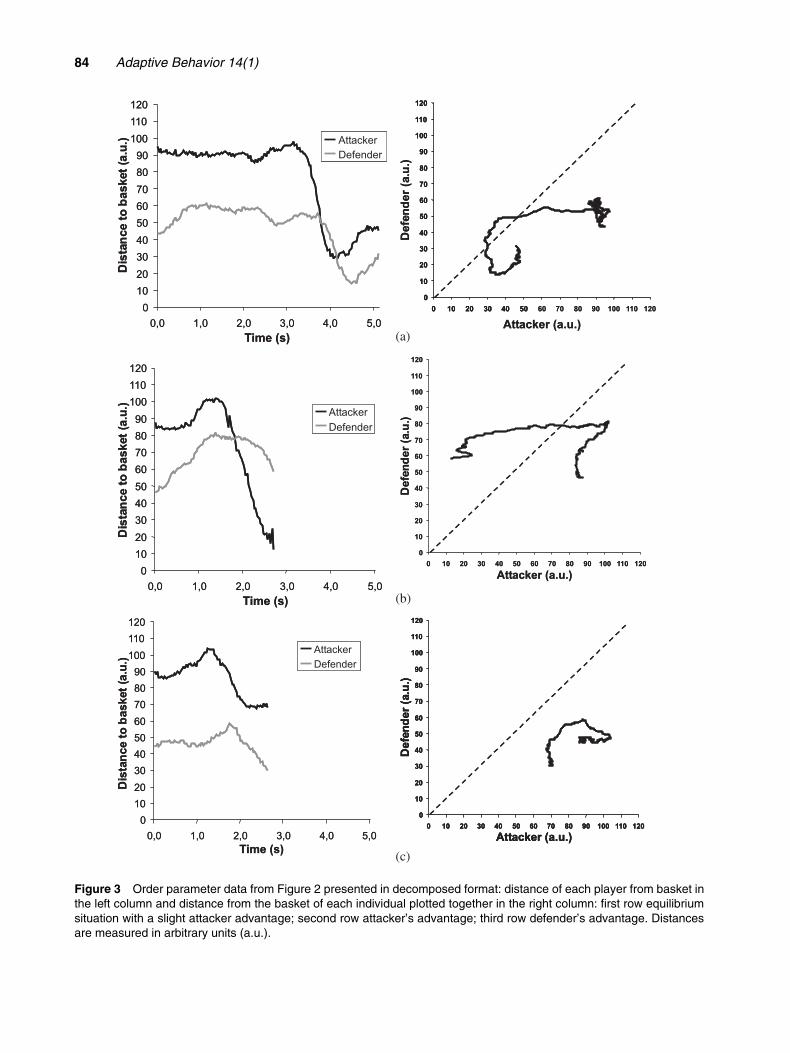
In interpersonal equilibrium, one stable state ofthe dyadic system is maintained during the whole situ-ation, as in Figures 2c and 3c, where the defender hasmanaged to constantly counter-balance the symmetry-breaking actions of the attacker. On the other hand,when the attacker is very successful there is almost noequilibrium. Figures 2b and 3b show that the transi-tion to a new state started immediately after thebeginning of the interaction. The transition is seen inFigure 3b when the initial order characterized by adefender’s advantage transits to a new order charac-terized by the attacker’s advantage. Interestingly, thesymmetry was broken exactly during the shortest val-ues of the control parameter: i.e., interpersonal dis-tance (right side of the figure). This is the transitionphase from dyad equilibrium to a new system order(attacker’s supremacy). In general, the attacker–defendersystem exhibited initial symmetry (showed in Fig-ures 2a and c), which was broken during transition toa new state at a critical value of the control parameter(as shown in Figures 2a and b). Further insights wereprovided when we decomposed our collective variable(distance from the medium point of the dyad to thebasket) showing each player’s distance to the basket.In Figure 3 we present the same situations as depictedin Figure 2 and we can clearly observe the nature ofsymmetry-breaking processes (Figures 3a and b), andhow symmetry was maintained (graph c).
The attacker–defender system exhibited initial sym-metry, which was broken during transition to a newstate at a certain value of the control parameter. Inother words, the attacker was trying to dribble past thedefender, but the defender was attempting to maintainthe initial steady state. The attacker increased the vari-ability of dribbling actions in order to force the emer-gence of a system transition (decision when to “go”).Suddenly (when the symmetry was broken), the deci-sion emerged in the “intending–perceiving–actingcycle.” Considering the characteristics of the figuresshowing the distance from the basket to the attackerand defender plotted together, it is possible to con-sider that the positions above its diagonal representattacker supremacy, and the positions of the systembelow the diagonal represent defender supremacy. In
Figure 1 A schematic diagram illustrating the startingposition of a typical basketball dyad (the attacker withthe ball is represented by the dark circle), based on the“attack system 1:2:2.” X = defenders; O = attackers.
Davids et al. Representative Task Design and Movement 83
Figure 2 Three examples of order parameters (distance of dyad from basket) in the left column and control parame-ters (interpersonal distance) in the right column: first row equilibrium situation with a slight attacker advantage; secondrow attacker’s advantage; third row defender’s advantage. Note that these figures cannot be superimposed because thevariables have different scaled magnitudes; distances are measured in arbitrary units (a.u.).
84 Adaptive Behavior 14(1)
Figure 3 Order parameter data from Figure 2 presented in decomposed format: distance of each player from basket inthe left column and distance from the basket of each individual plotted together in the right column: first row equilibriumsituation with a slight attacker advantage; second row attacker’s advantage; third row defender’s advantage. Distancesare measured in arbitrary units (a.u.).

Davids et al. Representative Task Design and Movement 85
summary, the data suggest that it is possible to inter-pret the dynamics of player interactions in dribbling asemergent properties under constraints.
2.3 Affordance Regulated Hitting Behavior in a Boxing Training Task
The nature of agent–agent and agent–environmentinteractions (Guerin & Kunkle, 2004), are useful forunderstanding how creation, maintenance, and decayof key constraints affects the interactions of perceptual,cognitive, and action sub-systems during sport perform-ance (Araújo et al., 2005b; Chow et al., 2006). Hand-striking movements in martial arts such as boxing arerepresentative movement models in this respect because(i) they exemplify classes of short lived, multi-jointinterceptive action patterns formed from the abun-dance of upper limb motor system degrees of freedom,which are assembled or dissolved depending on per-formance constraints, and (ii) they represent time-var-ying intra- and inter-limb coordinative states with respectto more or less dynamic targets such as opponents ortraining equipment such as speed bags, hand pads, andheavy bags (Turvey, 1990).
Diversity of handstriking actions plays a crucialrole in boxing since it increases uncertainty for oppo-nents and constrains them to depend on probabilisticperceptual judgments for decision making, rather thanknowledge of opponent actions. Game theory predictsthat unpredictability of an organism’s actions is a requi-site feature of efficient adaptive behavior in competitiveenvironments and many organisms show indetermi-nate behavior when they find it to have survival value(Glimcher, 2005). The diversity of actions is based onmotor equivalence, namely the ability of different actionpatterns to achieve the same goal, suggesting that theseproperties of the organisms evolved as a consequenceof their adaptive value. More generally, this ability ofaction systems can be viewed as a specific case of theevolutionary stabilized degeneracy inherent to biolog-ical systems (Edelman & Gally, 2001).
The possibility of exploiting motor equivalence,and consequently the diversity of actions, depends onthe variables to which boxers’ perceptual systems aresensitive. Warren (1984) reported that young adultsranging in stature could, by visual inspection alone,determine which in a series of stairs varying in riserheights afforded bipedal climbing. At a critical point,around 0.88 of leg length, climbers were unable to
ascend the stair bipedally and a sudden change inbehavior occurred: Climbers ascended quadrupedally.The sensitivity of climbers’ perceptual system to thisaffordance (i.e., climbability) controlled the behavio-ral transition between the two climbing modes. In simi-lar vein, reaching towards a target involves detection ofaffordances for a specific mode of action and requiresthe pick-up of critical reach distances. These variablesare specifically scaled to each individual, aroundwhich distinct coordination modes for reaching maybe available (Choi & Mark, 2004).
Striking modes of action in martial arts differ fromother kinds of reaching modes in a number of biody-namic parameters. In the martial arts, movements suchas reaching to hit a target require performers to pick upbody-scaled information (e.g., arm length) to judgewhether a target is reachable with a specific action (e.g.,an uppercut or jab) at a particular distance. Additionally,it is possible that the perception of some biodynamicparameters (Choi & Mark, 2004) (e.g., fist–target energyof collision) might also contribute to constraining strik-ing activity. Therefore, it is theoretically likely thataffordances of different hitting actions emerge in martialarts such as boxing, constrained by the scaling of athletepositioning and arm segment dimensions with referenceto a target, as well as by their perceived efficiency.
In ecological psychology, perception has beendefined as the detection and use of information (Gib-son, 1979), and an important experimental approach inresearching interceptive actions has been to manipu-late properties of environmental information to observechanges in movement behavior, as evident in move-ment kinematics. The assumption is that observedchanges to movement kinematics are likely due to theexperimental manipulations of perceptual variables,which act as information specifying or constrainingactions such as hitting (Beek, Dessing, Peper, & Bullock,2003a). Since perception is specific to the environmen-tal properties uniquely constraining each performancesituation, it follows that changing the informationalconstraints during performance can influence emergentaction patterns (Beek et al., 2003b).
2.4 A Description of a Methodology for Studying Emergent Decision Making During Boxing Practice
Hitting a heavybag is a key task in the basic trainingof boxers. By exploring their own perceptual–motor

86 Adaptive Behavior 14(1)
landscape, boxers can discover areas where their strik-ing diversity is maximal and differences between theefficiency characteristics of their strokes are minimal,which is of utmost importance for adaptation of boxersto stylistic differences in opponents. Hristovski, Davids,and Araújo (in press) examined how the interaction ofthe repetitive handstriking task, the gradually changingperformer–target distance, and efficiency informationconstrained the decision–action behavior in boxers.Specifically, they were interested in the dynamicalinteractions between the task and gradual changes inperceptual information from the task solution space ofthe performers, and effects on the decision–actiondiversity of the performers. It was predicted that theinitial activation of an action, the probability of occur-rence, and the dissolution of actions would exhibitdependence on the distance to the target and on per-ceived striking efficiency by novice boxers. Boxersaged 21–23 years were studied as they selected appro-priate action patterns to ensure shots of efficient colli-sion magnitude to a black leather hanging heavybagfixed to the wall with its bottom 95 cm from the floor.
Figure 4A shows how a distance of one meter fromthe target was calibrated in 10 equal segments of 10 cmso that each boxer could perform 60 shots at each dis-tance. These task constraints were formulated to allowperformers to exploit both efficiency and diversity ofboxing actions. Six classical boxing actions could beperformed during the task: right jab, left jab, right hook,left hook, right uppercut, and left uppercut. A conven-tional procedure of a gradual change of the hypothe-sized control parameter (the participant–target distance)was used to observe the distances at which new actionsemerged as well as the changes of the associated per-ceived efficiencies (Kelso, 1995).
Before starting the activity, participants were strin-gently reminded not to overbalance. This instructionalconstraint is particularly emphasized in the advice ofexpert boxers (Walker, 2003). The choice of distancebetween the lower limbs in the parallel stance and thetiming between the shots in the sequence was left tothe participants. Participants were also free to applyuppercut strokes to any location of the target. Thismethodology allowed Hristovski et al. (in press) to
Figure 4 (a), The apparatus for the investigation seen from above. (b), Schematic representation of the jab and hooktrajectories and the dominant projection axes of these action modes. (c), As (b) for the uppercuts.

Davids et al. Representative Task Design and Movement 87
plot the distances of the earliest activation of upper-cuts, since due to the specific patterning of this strokefrom larger distances, performers could only efficientlystrike the lower or middle part of the heavybag. Eachboxer was asked about their perceptions of stroke effi-ciency for each shot selected after they had finishedeach sequence of activity. Perceived efficiency wasscaled by the performers on a 6 point (0–5) continuousscale with 0 reflecting absence of a stroke and 5 signi-fying a maximally efficient stroke. Modes of performer–target co-ordination patterns were operationally definedas directions of upper limb movements with respect tothe central visual line connecting the participant andthe target facing him. This procedure allowed a nota-tional strategy to be used in classifying shot frequencyas a measure of decision-making stability in the dynamiccontext of boxing. For example, right jabs have adominant projection parallel to the central visual axis(z axis) and were numerically classified as 180 degrees.Left jabs, providing a mirror image of right jabs, wereclassified as –180 deg (see Figure 4b). Notation ofhooks was considered to have projections along thesagittal (z) and horizontal (x) axes, with right hooksbeing classified as 90xz deg. and their mirror image lefthooks as –90xz deg. (see also Figure 4b). Furthermore,upper cut handstrikes were recorded as projections onthe sagittal (z) axis, with right and left uppercuts vary-ing along the vertical (y) axis at 90yz deg. and –90yz
deg. respectively (see Figure 4c).For the purposes of the analysis the following
measures were calculated: (i) Relative scaled distancesD determined as a ratio between the physical distanceof participants from the target and their arm length; (ii)the absolute probabilities of occurrence of action pat-terns (relative frequencies) p(k(D)) = k(D)/n, wherek(D) is the number of occurrences of a certain type ofstroke at distance D and n is the constant length of thesequence (n = 60) for each distance from the target.This variable measured the degree of attraction of theaction modes; (iii) the mean group probabilities of eachof the three general types of boxing shots (jabs, hooks,and uppercuts) for each distance P(D); (iv) meangroup perceived striking efficiencies for each generalaction mode (jabs, hooks, and uppercuts) at scaled dis-tance D, E(D); (v) unpredictability and diversity ofstriking activity by the participants assessed by the
Shannon entropy H(p(D)) = ; (j = 6),
i.e. as average information over all outcomes in
the sequence; and symmetry measures: S(p(D)) =
, where N is the number of combi-
nations of class 2 for a set of 6 elements (6 types ofstrokes), and |∆p(D)|i is the absolute value of the dif-ferences between the probabilities of occurrence ofeach action pattern. In this study, for 6 possible typesof strokes, there were N = 15 probability differencesto calculate for each sequence per participant.
Bifurcation points of the action modes were deter-mined by analyzing the behavior of the first derivativeof the probability of occurrence of action patternsP (D) with respect to scaled distance to target (Sugakov,1998) in order to find the points of discontinuity (non-analyticity) of the performers’ movement patterns. Spe-cifically, the bifurcations of the first-time emergenceof action modes were identified by analyzing thebehavior of the left and right derivatives, i.e., P (D–)and P (D+) respectively, in the vicinity of pointswhere the first nonzero values of Pi(D) were obtained.In similar vein, for detecting the action mode dissolu-tion bifurcations analytical treatments of the behaviorof P (D) in the vicinity of points where Pi(D) switchedto Pi(D) = const. = 0 were conducted.
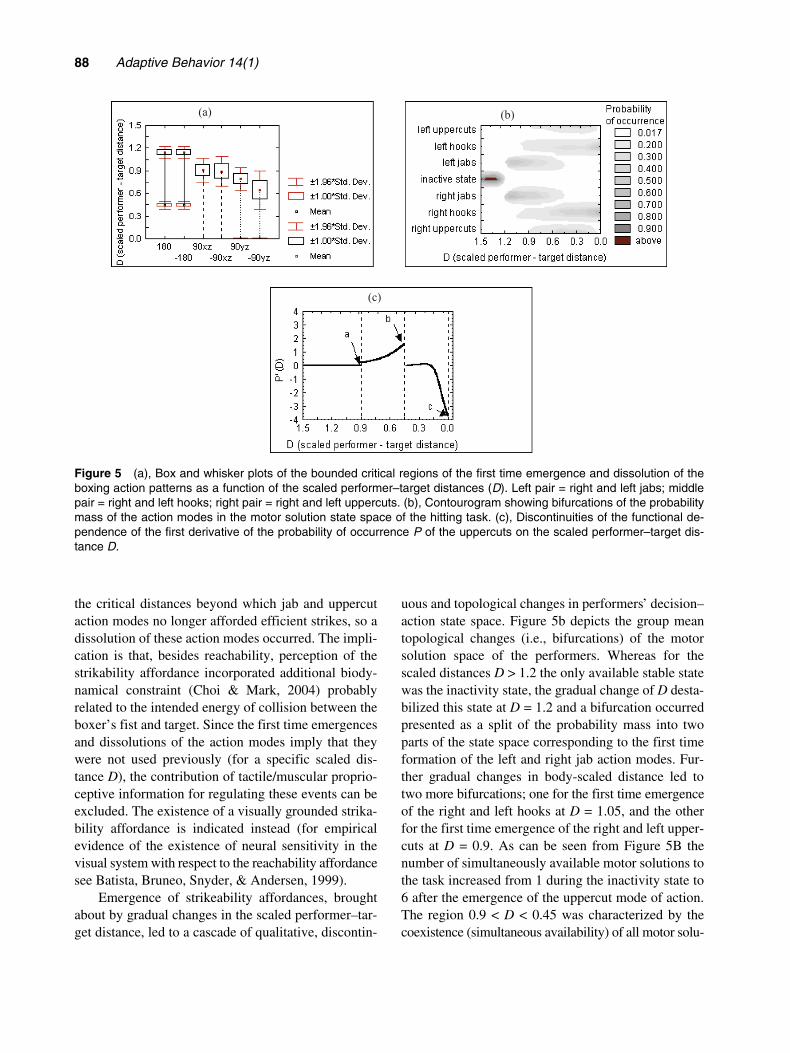
Results showed several characteristics of boxingbehavior. First, significant differences between thebounded regions of scaled performer–target distancesat which specific action modes emerged or weredissoluted (i.e., ceased to exist) for the first time wereindicative of the existence of affordances specificto the boxing action patterns used by performers (Fig-ure 5a).
Data showed that the performers used thesebounded regions of scaled distance as contextual infor-mation specifying a change in the number of possiblemotor solutions to the hitting task. The perceived con-textual information was body-scaled distance informa-tion (Konczak, 1990; Konczak, Meeuwsen, & Cress,1992) and depended on the interaction of the environ-mental constraints (absolute distance from the target)and morphological constraints of performers (effectivelength of upper limbs specific to each of the actionmodes).
The data also suggested that strikability is notmerely a specific case of the reachability affordance,as indicated by dissolution events of jab and uppercutmodes of action. At both these critical points of disso-lution the target was reachable, but jabs and uppercutsceased to exist. It appears that boxers could perceive
pi D( )ln pi D( )i=1
j
∑
11N---- ∆p D( ) i
i=1
N
∑–
i′
i′i′
i′
88 Adaptive Behavior 14(1)
the critical distances beyond which jab and uppercutaction modes no longer afforded efficient strikes, so adissolution of these action modes occurred. The impli-cation is that, besides reachability, perception of thestrikability affordance incorporated additional biody-namical constraint (Choi & Mark, 2004) probablyrelated to the intended energy of collision between theboxer’s fist and target. Since the first time emergencesand dissolutions of the action modes imply that theywere not used previously (for a specific scaled dis-tance D), the contribution of tactile/muscular proprio-ceptive information for regulating these events can beexcluded. The existence of a visually grounded strika-bility affordance is indicated instead (for empiricalevidence of the existence of neural sensitivity in thevisual system with respect to the reachability affordancesee Batista, Bruneo, Snyder, & Andersen, 1999).
Emergence of strikeability affordances, broughtabout by gradual changes in the scaled performer–tar-get distance, led to a cascade of qualitative, discontin-
uous and topological changes in performers’ decision–action state space. Figure 5b depicts the group meantopological changes (i.e., bifurcations) of the motorsolution space of the performers. Whereas for thescaled distances D > 1.2 the only available stable statewas the inactivity state, the gradual change of D desta-bilized this state at D = 1.2 and a bifurcation occurredpresented as a split of the probability mass into twoparts of the state space corresponding to the first timeformation of the left and right jab action modes. Fur-ther gradual changes in body-scaled distance led totwo more bifurcations; one for the first time emergenceof the right and left hooks at D = 1.05, and the otherfor the first time emergence of the right and left upper-cuts at D = 0.9. As can be seen from Figure 5B thenumber of simultaneously available motor solutions tothe task increased from 1 during the inactivity state to6 after the emergence of the uppercut mode of action.The region 0.9 < D < 0.45 was characterized by thecoexistence (simultaneous availability) of all motor solu-
Figure 5 (a), Box and whisker plots of the bounded critical regions of the first time emergence and dissolution of theboxing action patterns as a function of the scaled performer–target distances (D). Left pair = right and left jabs; middlepair = right and left hooks; right pair = right and left uppercuts. (b), Contourogram showing bifurcations of the probabilitymass of the action modes in the motor solution state space of the hitting task. (c), Discontinuities of the functional de-pendence of the first derivative of the probability of occurrence P of the uppercuts on the scaled performer–target dis-tance D.

Davids et al. Representative Task Design and Movement 89
tions to the hitting task with different degrees ofattraction as estimated by their probability of occur-rence. This is also a region of the full exploitation ofthe capacity for motor equivalence by performers. Thecascade of topological changes in state space charac-terized by a reduction in the number of co-existentstates of action modes started with the dissolutionbifurcation of jab actions around D = 0.45 and endedwith the dissolution bifurcation of uppercut actions atD = 0.00 when only right and left hook action modesremained available. Hence, the perception of affordancesat the critical points of the body-scaled distances ledto drastic changes in the motor solution manifold of theperformers.
Figure 5c depicts the functional discontinuities ofthe degree of attraction of the uppercut action patternas measured by the probability of occurrence. Theextant discontinuities show that, at critical points, theperceived strikeability affordances led to non-analyti-cal behavior of the action system of performers whereit switched into different forms of dependence on thescaled distance D. Hence, the bifurcation cascade eventsin the motor solution space and the discontinuouschanges in behavior of participants at critical points ofthe scaled distance point strongly to extant affordance-regulated, macroscopic nonlinear dynamics of theirperceptual–action systems.
Furthermore, this investigation showed that box-ing action modes (i.e., jabs, hooks, and uppercuts) wereconcurrent to and competing with each other as revealedby the values and signs of partial correlations betweenthe scaled distance dependent perceived efficienciesEi(D) and the scaled distance dependent probabilitiesof occurrence of particular action modes Pi(D). Thestrong and positive partial correlations between thejab, hook, and uppercut action modes and their per-ceived efficiencies (0.968, p = 0.004; 0.949, p = 0.001,and 0.962, p = 0.0005, respectively) showed that per-ceived efficiencies enhanced the degree of attractive-ness of those action modes. In contrast to this finding,strong negative partial correlations between the per-ceived efficiencies of concurrent action modes and theprobabilities of occurrence of other action modes, suchas that between the perceived efficiency of hooks andthe probability of occurrence of jabs (–0.959, p =0.0007) as well as other highly negative associations,highlight the suppressing influences of concurrent effi-ciencies on the degree of attractiveness of other actions.In other words, the perceived efficiency of a particular
action mode tended to have inhibitory influence on thedegree of attractiveness of other concurrent modes andsimultaneously an excitatory influence on the associ-ated action mode. These scaled distance-dependentreciprocal enhancing and suppressing influences of theperceived efficiencies shaped the competition effectsbetween the concurrent modes of action.
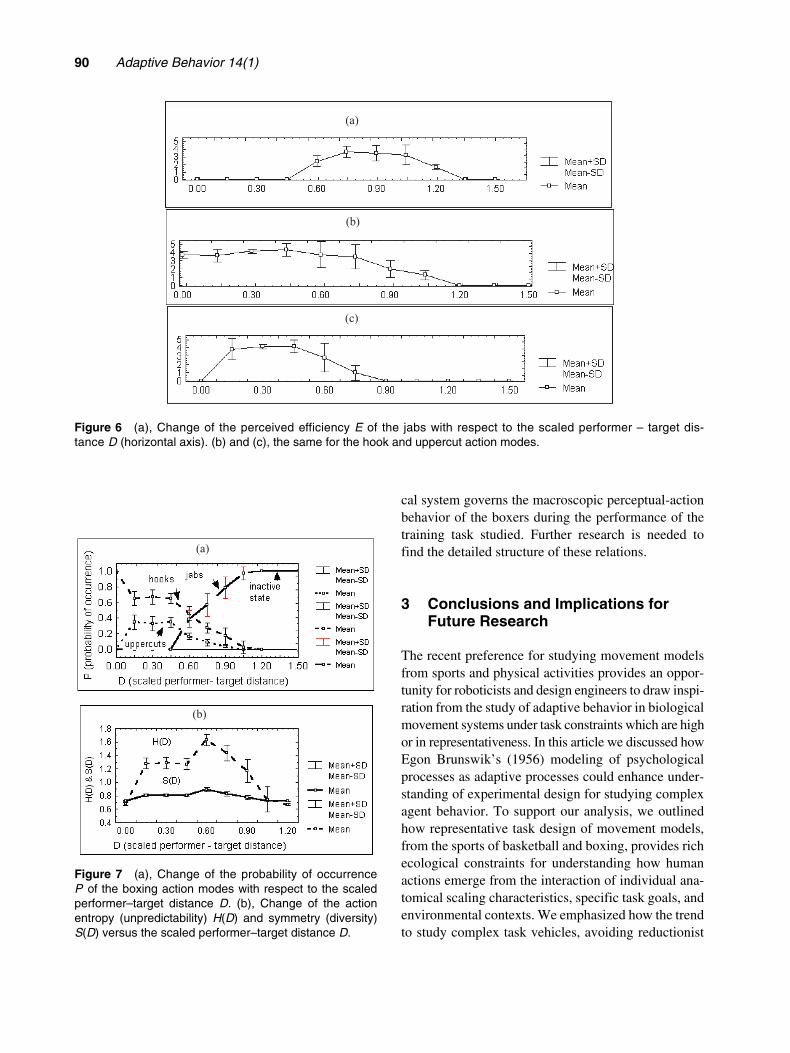
Perceived efficiencies Ei(D) also showed a body-scaled performer–target discontinuous, i.e., nonlinear,distance dependence (see Figures 6a–c).
This finding is corroborated by a change in thefunctional dependence of Ei(D) at the points whenEi(D) = 0 Ei(D) 0 and at the points when Ei(D) 0
Ei(D) = 0. Hence, the occurrence of action modesat each scaled participant–target distance was a netemergent result of the selective pressure and competitionbetween these interacting enhancing and suppressingcontextual constraints. The contextual dependence ofthe attractiveness of the action modes was representedby a clearly visible distance-dependent polarized struc-ture of the action mode degree of attraction (Figure 7a),where for scaled distances 1.2 > D > 0.6 jab actionsdominated over hooks and uppercuts, and for 0.6 > D >0.00 hook and uppercut actions dominated over jabs.
Figure 7b depicts the scaled performer–target dis-tance-dependent changes in the action entropy (unpre-dictability) and symmetry (diversity) measures. Theirmaxima lie in the region where the dominance of jabaction modes transits to dominance of the hook anduppercut action modes: i.e., In the region of their min-imum probability (degree of attraction) difference whichis located around D = 0.6. This region of a maximaldynamic competition of action modes is thus the opti-mal region of exploitation of the capacity for motorequivalence in performers. The exploitation of motorequivalence, and consequently the diversity and unpre-dictability of actions, shows a clear contextual depend-ence. Performers were able to discover the body-scaled perceptual context that afforded them full useof the largest set of available motor system degree offreedom configurations to enhance the diversity andunpredictability of actions which are key determinantsof adaptive behavior in boxing. The possibility ofexploitation of this one (goal) to many (action modes)mapping depended on the context created by the inter-action of the perceptual information such as body-scaled performer–target distances and the perceivedefficiencies of actions. In other words, this findingsuggests that a basic P = P(E(D)) nonlinear dynami-
→ ≠ ≠→
90 Adaptive Behavior 14(1)
cal system governs the macroscopic perceptual-actionbehavior of the boxers during the performance of thetraining task studied. Further research is needed tofind the detailed structure of these relations.
3 Conclusions and Implications for Future Research
The recent preference for studying movement modelsfrom sports and physical activities provides an oppor-tunity for roboticists and design engineers to draw inspi-ration from the study of adaptive behavior in biologicalmovement systems under task constraints which are highor in representativeness. In this article we discussed howEgon Brunswik’s (1956) modeling of psychologicalprocesses as adaptive processes could enhance under-standing of experimental design for studying complexagent behavior. To support our analysis, we outlinedhow representative task design of movement models,from the sports of basketball and boxing, provides richecological constraints for understanding how humanactions emerge from the interaction of individual ana-tomical scaling characteristics, specific task goals, andenvironmental contexts. We emphasized how the trendto study complex task vehicles, avoiding reductionist
Figure 6 (a), Change of the perceived efficiency E of the jabs with respect to the scaled performer – target dis-tance D (horizontal axis). (b) and (c), the same for the hook and uppercut action modes.
Figure 7 (a), Change of the probability of occurrenceP of the boxing action modes with respect to the scaledperformer–target distance D. (b), Change of the actionentropy (unpredictability) H(D) and symmetry (diversity)S(D) versus the scaled performer–target distance D.

Davids et al. Representative Task Design and Movement 91
(or biased) approaches, in research on human move-ment behavior has coincided with advances in theorizingfrom ecological psychology and dynamical systemstheory combined in a constraints-led perspective. Aconstraints-led view of movement coordination andcontrol provides the theoretical impetus for consider-ing task representative design because it highlights theneed to adopt a systems perspective in viewing bio-logical and artificial agents as systems composed ofmany interacting subsystems. A systems perspectiveencourages the view that the appropriate scale ofanalysis for studying design of biological and artificialsystems is at the level of the agent–environment inter-action. From a systems perspective, there is a need tounderstand the emergent couplings that can occurbetween components of the environment and to studythe close link between biological and artificial move-ment systems and their environments (particularly theirsensory and movement subsystems and the surround-ing energy sources).
We argued that movement models from sportsprovide excellent vehicles for studying the basis ofbehavior-based robotic implementations in complexenvironments. Cross-disciplinary fertilization couldbe supported by the study of coordination withinand between different sub-systems of biological andartificial movement systems as well as coordinationbetween agents and the rich range of task constraintsfound in complex environments (Jirsa & Kelso, 2004).The use of movement models from sport yield excel-lent insights into agent–agent and agent–environmentinteractions, particularly from the perspective of “fat-agent” modeling which includes reference to constraintson behaviors like memory, perception, cognitions, andactions (Guerin & Kunkle, 2004). The data describedin the example movement models from basketball andboxing revealed (i) the nature of the perceptual infor-mation used to constrain action; and (ii) the inextrica-ble relationship between information, cognitions, andmovements in specific performance contexts. This the-oretical and empirical emphasis on how system con-straints interact with task constraints is signaling afresh perspective on the role of variability in facilitatingadaptation of agents to dynamic task environments.Analysis of behavior at the level of agent–environmentinteractions is revealing coordination modes as anexample of adaptive, emergent phenomena which canbe functionally varied to meet the challenge of per-forming in dynamic contexts.
The emergence of specific modes of behavior pro-vide insights into the strong influence of task constraints,suggesting the need to carefully consider the nature oftasks used in design laboratories. Designers and engi-neers of robotic and artificial devices could try tomaintain the important perception–action links towardsspecific goals that characterize performance competi-tion or match play in sports contexts as faithfully aspossible. The idea is to design a robot or an artificialdevice attending to the nature of the external informa-tion that should be acted upon in order to achieve acertain goal. By acting upon particular contextual infor-mation the agent should be able to adapt its behavioraccordingly. This means that behavior is not tailoredin advance (internally), but that it is regulated ongo-ingly by environmental information. The data dis-cussed in this position paper suggest that researchersin robotics, artificial intelligence, and machine designengineering can promote understanding of adaptivebehavior of smart agents by continuing to adopt move-ment models from sports in the future.
References
Abernethy, B., Thomas, K. T., & Thomas, J. T. (1993). Strate-gies for improving understanding of motor expertise [ormistakes we have made & things we have learned!]. In J.L. Starkes & F. Allard (Eds.), Cognitive issues in motorexpertise (pp. 317–356). Amsterdam, Netherlands: Else-vier Science.
Alderson, G. J. K., Sully, D. J., & Sully, H. G. (1974). An oper-ational analysis of one-handed catching task using highspeed photography. Journal of Motor Behavior, 6, 217–226.
Araújo, D., Davids. K., Sainhas, J., & Fernandes, O. (2002).Emergent decision-making in sport: A constraints-ledapproach. In L. Toussaint & P. Boulinguez (Eds.) Interna-tional congress “movement, attention & perception” (p. 77).Poitiers, France; Université do Poitiers.
Araújo, D., Davids, K., Bennett, S., Button, C., & Chapman, G.(2004). Emergence of Sport Skills under Constraints. InA. M. Williams & N. J. Hodges (Eds.), Skill acquisition insport: Research, theory and practice (pp. 409–434). Lon-don: Routledge, Taylor & Francis.
Araújo, D., Davids, K., & Passos, P. (2005a). Operating in atheoretical twilight: Towards a stronger representativeexperimental design: Comment on “Rogers, S., Kadar, E.,& Costall, A. (2005). Gaze patterns in the visual control ofstraight-road driving and braking as a function of speedand expertise”. Ecological Psychology, 17, 19–38.

92 Adaptive Behavior 14(1)
Araújo, D., Davids, K., & Serpa, S. (2005b). An ecologicalapproach to expertise effects in decision-making in a sim-ulated sailing regatta. Psychology of Sport and Exercise, 6,671–692.
Bain, B., Hayes, D., & Quance, A. (1978). Coaching certifica-tion manual—Level two. Toronto, ON: Canada Basketball.
Balasubramaniam, R., & Turvey, M. T. (2004). Coordinationmodes in the multisegmental dynamics of hula hooping.Biological Cybernetics, 90, 176–190.
Batista, A. P., Bruneo, C. A., Snyder, L. H., & Andersen, R. A.(1999). Reach plans in eye—centered coordinates. Science,285, 257–260.
Beek, P. J., Dessing, J. C., Peper, C. E., & Bullock, D. (2003a).Modelling the control of interceptive actions. Philosophi-cal Transactions of the Royal Society of London B, 358,1511–1523.
Beek, P. J., Jacobs, D. M., Daffertshoffer, A., & Huys, R.(2003b). Expert performance in sport: Views from the jointperspectives of ecological psychology and dynamical sys-tems theory. In J. L. Starkes & K. A. Ericcson (Eds.), Expertperformance in sports: Advances in research on sportexpertise (pp. 321–344). Champaign, IL: Human Kinetics.
Bennett, S. J., van der Kamp, J., Savelsbergh, G. J. P., & Dav-ids, K. (2000). Discriminating the role of binocular infor-mation in the timing of a one-hand catch: The effects oftelestereoscopic viewing and ball size. ExperimentalBrain Research, 135, 341–347.
Bernstein, N. (1967). The co-ordination and regulation ofmovements. Oxford: Pergamon.
Berthouze, L. & Lungarella, M. (2004). Motor skill acquisitionunder environmental perturbations: On the necessity ofalternate freezing and freeing degrees of freedom. Adap-tive Behavior, 12, 47–64.
Bootsma, R. J. (1989). Accuracy of perceptual processes sub-serving different perception-action systems. QuarterlyJournal of Experimental Psychology, 41A, 489–500.
Broderick, M. P., Pavis, B., & Newell, K. M. (2000). Assess-ment of the adiabatic transformability hypothesis in a ball-bouncing task. Biological Cybernetics, 82, 433–442.
Brunswik, E. (1956). Perception and the representative designof psychology experiments (2nd ed.). Berkeley, CA: Uni-versity of California Press.
Button, C., McLeod, M., Sanders, R., & Coleman, S. (2003).Examining movement variability in the basketball free-throw action at different skill levels. Research Quarterlyfor Exercise and Sport, 74, 257–269.
Carson, R. G. (2004). Governing coordination: Why do mus-cles matter? In V. K. Jirsa & J. A. S. Kelso (Eds.), Coordi-nation Dynamics: Issues and Trends (pp.141–154). NewYork: Springer.
Carson, R. G., & Kelso, J. A. S. (2004). Governing coordina-tion: Behavioral principles and neural correlates. Experi-mental Brain Research, 154, 267–274.
Chen, H. H., Liu, Y.-T., Mayer-Kress, G., & Newell, K. M.(2005). Learning the pedalo locomotion task. Journal ofMotor Behaviour, 37, 247–256.
Choi, H. J., & Mark, L. S. (2004). Scaling affordances forhuman reach actions. Human Movement Science, 23, 785–806.
Chow, J.-Y., Davids, K., Button, C., Shuttleworth, R., Ren-shaw, I., & Araújo, D. (2006). Nonlinear pedagogy: Aconstraints-led framework for understanding emergenceof game play and movement skills. Nonlinear Dynamics,Psychology & the Life Sciences, 10, 71–103.
Cordo, P. J., & Gurfinkel, V. S. (2004). Motor coordination canbe fully understood only by studying complex move-ments. Progress in Brain Research, 143, 29–38.
Daffertshofer, A., Huys, R., & Beek, P. J. (2004). Dynamicalcoupling between locomotion and respiration. BiologicalCybernetics, 90, 157–164.
Davids, K., Kingsbury, D., Bennett, S. J., & Handford, C.(2001). Information–movement coupling: Implications forthe organisation of research and practice during acquisi-tion of self-paced extrinsic timing skills. Journal of SportsSciences, 19, 117–127.
Davids, K., Savelsbergh, G., Bennett, S. J., & Van Der Kamp,J. (Eds.) (2002). Interceptive Actions in Sport. London:Routledge, Taylor & Francis.
Davids, K., Araújo, D., Shuttleworth, R., & Button, C. (2003).Acquiring skill in sport: A constraints-led perspective. Inter-national Journal of Computer Sciences in Sport, 2, 31–39.
Davids, K., Shuttleworth, R., Button, C., Renshaw, I., & Gla-zier, P. (2004). “Essential noise”—enhancing variabilityof informational constraints benefits movement control: Acomment on Waddington and Adams (2003). British Jour-nal of Sports Medicine, 38, 601–605.
Davids, K., Bennett, S. J., & Newell, K. M. (Eds.) (2006). Move-ment system variability. Champaign, IL: Human Kinetics.
DeGuzman, G. C. (2004). Using visual information in func-tional stabilization: Pole balancing example. In V. K. Jirsa& J. A. S. Kelso (Eds.), Coordination dynamics: Issuesand trends (pp. 91–101). New York: Springer.
Di Paolo, E. (2002). Spike-timing dependent plasticity forevolved robots. Adaptive Behavior, 10, 243–263.
Duchon, A. P., & Warren, W. H. (2002). A visual equalizationstrategy for locomotor control: Of honeybees, robots andhumans. Psychological Science, 13, 272–278.
Edelman, G. M., & Gally, J. A. (2001). Degeneracy and com-plexity in biological systems. Proceedings of the NationalAcademy of Sciences of the USA, 98, 13763–13768.
Fajen, B. R., Warren, W. H., Temizer, S., & Kaelbling, L. P.(2003). A dynamical model of visually-guided steering,obstacle avoidance and route selection. InternationalJournal of Computer Vision, 54, 13–34.
Gibson, J. J. (1957). Survival in a world of probable objects.Review of Perception and the representative design of

Davids et al. Representative Task Design and Movement 93
psychological experiments by E. Brunswik, ContemporaryPsychology, 2, 33–35.
Gibson, J. J. (1979). The ecological approach to visual percep-tion. Boston: Houghton Mifflin.
Glimcher, P. W. (2005). Indeterminacy of brain and behavior.Annual Review of Psychology, 56, 25–56.
Guerin, S., & Kunkle, D. (2004). Emergence of constraint inself-organized systems. Nonlinear Dynamics, Psychologyand Life Sciences, 8, 131–146.
Haibach, P. S., Daniels, G. L., & Newell, K. M. (2004). Coor-dination changes in the early stages of learning to cascadejuggle. Human Movement Science, 23, 185–206.
Handford, C. (2002). Strategy and practice for acquiring timingin discrete, self-paced interceptive actions. In K. Davids,G. J. P. Savelsbergh, S. J. Bennett & J. van Der Kamp(Eds.), Interceptive actions in sport (pp.288–300). Lon-don: Routledge, Taylor and Francis.
Harris, L. R., & Jenkin, M. (1998). Vision and action. Cam-bridge, UK: Cambridge University Press.
Heft, H. (2001). Ecological psychology in context: James Gibson,Roger Barker, and the legacy of William James’ radicalempiricism. Mahwah, NJ: Lawrence Erlbaum Associ-ates.
Hill, H., & Raab, M. (2005). Spatio-temporal analysis of acomplex visuo-motor tracking task with brain electricalrelated potentials. Human Movement Science, 23, 238–251.
Hristovski, R., Davids, K., & Araújo, D. (in press). Affordance-controlled bifurcations of action patterns in martial arts.Nonlinear Dynamics, Psychology and the Life Sciences.
Huys, R., Daffertshofer, A., & Beck, P. J. (2004), Emergenceof sport skills under constraint. In A. M. Willinams &N. J. Hodges (Eds.), Skill acquisition in sport: Research,theory and practice (pp. 351–373). London: Routledge,Taylor & Francis.
Jirsa, V. K., & Kelso, J. A. S. (Eds.) (2004). Coordination dynam-ics: Issues and trends. New York: Springer.
Kelso, J. A. S. (1995). Dynamic patterns. The self-organizationof brain and behavior. Cambridge, MA: MIT Press.
Kelso, J. A. S., Fink, P. W., Delaplain, C. R., & Carson, R. G.(2001). Haptic information stabilizes and destabilizes coor-dination dynamics. Proceedings of the Royal Society ofLondon B, 268, 1207–1213.
Knight, C. A. (2004). Neuromotor issues in the learning andcontrol of golf skill. Research Quarterly for Exercise andSport, 75, 9–15.
Konczak, J. (1990). Toward an ecological theory of motor devel-opment: The relevance of the Gibsonian approach tovision for motor development research. In J. Clark & J.Hymphrey (Eds), Advances in Motor DevelopmentResearch: Vol. 3 (pp. 3–17). New York: AMS Press.
Konczak, J., Meeuwsen, H., & Cress, M. (1992). Changingaffordances in stair climbing: The perception of maximumclimbability in young and older adults. Journal of Experi-
mental Psychology: Human Perception and Performance,18, 691–697.
Kugler, P. N., & Turvey, M. T. (1987). Information, naturallaw and the self-assembly of rhythmical movement. Hills-dale, NJ: Lawrence Erlbaum Associates.
Kugler, P. N., Kelso, J. A. S., & Turvey, M. T., (1980). On theconcept of coordinated structures as dissipative structures.I : Theoretical lines of convergence. In G. E. Stelmach &J. Requin (Eds.), Tutorials in motor behavior (pp. 3–45).Amsterdam: North-Holland.
Mataric, M. (1998). Behavior-based robotics as a tool for syn-thesis of artificial behavior and analysis of natural behav-ior. Trends in Cognitive Science, 2, 82–87.
Mikulecky, D. C. (2000). Robert Rosen: The well-posed questionand its answer—Why are organisms different from machines.Systems Research and Behavioral Science, 17, 419–432.
Montagne, G., Cornus, S., Glize, D., Quaine, F., & Laurent, M.(2000). A perception–action coupling type of control inlong jumping. Journal of Motor Behavior, 32, 37–43.
Montagne, G., Laurent, M., Durey, A., & Bootsma, R. J.(1999). Movement reversals in ball catching. Experimen-tal Brain Research, 129, 87–92.
Müller, H., & Sternad, D. (2004). Decomposition of variabilityin the execution of goal-oriented tasks: Three componentsof skill improvement. Journal of Experimental Psychol-ogy: Human Perception and Performance, 30, 212–233.
Newell, K. M. (1986). Constraints on the development of coor-dination. In M. G. Wade & H. T. A. Whiting (Eds.), Motordevelopment in children. Aspects of coordination and con-trol (pp. 341–360). Dordrecht: Martinus Nijhoff.
Newell, K. M. (1989). On task and theory specificity. Journalof Motor Behavior, 21, 92–96.
Newell, K. M. (1991). Motor skill acquisition. Annual Reviewof Psychology, 42, 213–237.
Oudejans, R. R. D., Michaels, C. F., Bakker, F. C., & Dolné,M. A. (1996). The relevance of catching in perceivingaffordances: Perception of catchableness of fly balls.Journal of Experimental Psychology: Human Perceptionand Performance, 22, 879–891.
Paine, R. W., & Tani, J. (2005). How hierarchical control self-organizes in artificial adaptive systems. Adaptive Behav-ior, 13, 211–225.
Post, A. A., Daffertshofer, A., & Beek, P. J. (2000). Principalcomponent analysis in three-ball cascade juggling. Biolog-ical Cybernetics, 82, 143–152.
Post, A. A., Peper, C. E., & Beek, P. J. (2003). Effects of visualinformation and task constraints on intersegmental coordi-nation in playground swinging. Journal of Motor Behav-ior, 35, 64–78.
Poulton, E. C. (1957). On prediction in skilled movements.Psychological Bulletin, 54, 467–478.
Reed, E. S. (1996). Encountering the world: Toward an ecolog-ical psychology. Oxford: Oxford University Press.

94 Adaptive Behavior 14(1)
Rieger, M., Knoblich, G., & Prinz, W. (2005). Compensationfor and adaptation to changes in the environment. Experi-mental Brain Research, 163, 487–502.
Riley, M. A., & Turvey, M. T. (2002). Variability and determin-ism in motor behavior. Journal of Motor Behavior, 34, 99–125.
Rosen, R. (1978). Fundamentals of measurement and represen-tation of natural systems. New York: North-Holland
Rosengren, K. S., Savelsbergh, G., & Van der Kamp, J. (2003).Development and learning: A TASC-based perspective ofthe acquisition of perceptual–motor behaviors. InfantBehavior and Development, 26, 473–494.
van Rooij, I., Bongers, R. M., & Haselager, W. F. G. (2002). Anon-representational approach to imagined action. Cogni-tive Science, 26, 345–375.
Satkunskiene, D., Schega, L., Kunze, K., Birzinyte, K., & Daly,D. (2005). Coordination on arm movements during crawlstroke in elite swimmers with a loco-motor disability. HumanMovement Science, 24, 54–65.
Schiff, W., & Detwiler, M. L. (1979). Information used in judg-ing impending collision. Perception, 8, 647–658.
Schiff, W., & Oldak, R. (1990). Accuracy of judging time-to-arrival: Effects of modality, trajectory, and gender. Jour-nal of Experimental Psychology: Human Perception andPerformance, 16, 303–316.
Schmidt, R. A., & Lee, T. D. (2005). Motor control and learn-ing: A behavioral emphasis (3rd ed.). Champaign, IL:Human Kinetics.
Schmidt, R. C., Carello, C., & Turvey, M. T. (1990). Phasetransitions and critical fluctuations in the visual coordina-tion of rhythmic movements between people. Journal ofExperimental Psychology: Human Perception and Per-formance, 16, 227–247.
Schmidt, R. C., O’Brien, B., & Sysko, R. (1999). Self-organi-zation of between-persons cooperative tasks and possibleapplications to sport. International Journal of Sport Psy-chology, 30, 558–579.
Schöllhorn, W. (2003). Coordination dynamics and its conse-quences on sports. International Journal of Computer Sci-ence in Sport, 2, 40–46.
Scholz, J. P., Schoner, G., & Latash, M. L. (2000). Identifyingthe control structure of multijoint coordination duringshooting. Experimental Brain Research, 135, 382–404.
Scott, M. A., van der Kamp, J., Savelsbergh, G. J. P., Oudejans,R. R. D., & Davids, K. (2004). Object rotation effects onthe timing of a hitting action. Research Quarterly forExercise and Sport, 75, 130–137.
Seth, A. K., & Edelman, G. M. (2004). Environment andbehavior influence the complexity of evolved neural net-works. Adaptive Behavior 12, 5–20.
Shuttleworth, R. (2004). Differential effects of varying atten-tional-focus through instruction in acquisition of move-ment coordination in rowing. Paper presented at the 31stAustralasian Experimental Psychology Conference, April2004, Dunedin, New Zealand.
Sternad, D., & Dijkstra, T. M. H. (2004), Dynamical stability inthe acquisition and performance of rhythmic ball manipu-lation: Theoretical insights with a clinical slant. Journal ofClinical Neurophysiology, 21, 215–227.
Sugakov, V. I. (1998). Lectures in synergetics. Singapore:World Scientific.
Tlili, M., Mottet, D., Dupuy, M.-A., & Pavis, B. (2004). Stabil-ity and phase locking in human soccer juggling, Neuro-science Letters, 360, 45–48.
Todd, J. T. (1981). Visual information about moving objects.Journal of Experimental Psychology: Human Perceptionand Performance, 7, 795–810.
Tresilian, J. R. (1994). Approximate information sources andperceptual variables in interceptive timing. Journal of Exper-imental Psychology: Human Perception and Performance,20, 154–173.
Tresilian, J. R. (1995). Perceptual and cognitive processes intime-to-contact estimation: Analysis of prediction–motionand relative judgement tasks. Perception and Psychophys-ics, 57, 231–245.
Turvey, M. T. (1990). Co-ordination. American Psychologist,45, 938–953.
Turvey, M. T., & Shaw, R. E. (1995). Toward an ecologicalphysics and a physical psychology. In R. L. Solso & D. W.,Massaro (Eds.), The science of the mind: 2001 and beyond(pp. 144–169). New York: Oxford University Press.
Vicente, J. K. (2003). Beyond the lens model and direct percep-tion: Toward a broader ecological psychology. EcologicalPsychology, 15, 241–267.
Walker, J. (2003). Self-defence techniques and tactics. Cham-paign, IL: Human Kinetics.
Warren, W. H. Jr. (1984). Perceiving affordances: Visual guid-ance of stairclimbing. Journal of Experimental Psychol-ogy: Human Perception and Performance, 10, 683–703.
Whiting, H. T. A. (1991). Action is not reaction! A reply toMcLeod and Jenkins. International Journal of Sport Psy-chology, 22, 296–303.
Williams, A. M., Davids, K., & Williams, J. G. (1999). Visualperception and action in sport. London: Routledge, Taylor& Francis.
Woodworth, R. S. (1938). Experimental Psychology. New York:Holt.
Wulf, G., & Shea, C. (2002). Principles derived from the studyof simple skills do not generalize to complex skill learn-ing. Psychonomic Bulletin and Review, 9, 185–211.

Davids et al. Representative Task Design and Movement 95
About the AuthorsKeith Davids is Professor and Dean of the School of Physical Education at the Univer-sity of Otago, NZ. His research interests are founded on the theoretical frameworks ofecological psychology and dynamical systems theory applied to the study of movementcoordination and control in sports. A particular interest concerns the role of constraints inmotor learning and the implications for the development of a nonlinear pedagogy. FromFebruary 2006 he will begin an appointment as Professor and Head of the School of HumanMovement Studies at the Queensland University of Technology in Brisbane, Australia.
Chris Button is Director of the Human Performance Centre in the School of PhysicalEducation at the University of Otago in NZ. He is a Senior Lecturer in motor behavior andresearch methods. Dr Button’s research interests include movement coordination andinterception skills, skill learning, and coaching. He has published his research in severalinternational peer-reviewed journals, and a summary can be found in the textbook,Dynamics of skill acquisition (2007, Human Kinetics). Dr Button has negotiated joint ven-ture agreements and contracts with the New Zealand Academy of Sport in the Central andSouth Island regions. He is also currently responsible for several commercial contractswith sports clothing and equipment companies to test their products (names protected byconfidentiality agreements). Address: School of Physical Education, University of Otago,46 Union St West, Dunedin, NZ. E-mail: [email protected]
Duarte Araújo is Assistant Professor at the Faculty of Human Kinetics at Technical Uni-versity of Lisbon in Portugal. His research involves the study of expert decision making insport, which led to him receiving a young researcher award at the European College ofSport Science in 2001 and at the Association des Chercheurs en Activités Physiques etSportives (ACAPS) in 2002. He was elected as a member of the managing council of theInternational Society of Sport Psychology in 2001, and has been Editor of the BulletinBoard of the International Journal of Sport (and Exercise) Psychology. He is a founder ofthe European Network of Young Specialists in Sport Psychology, being now its past coor-dinator. He has been a performance consultant for the Portuguese Sailing Federation since1995. Address: Laboratorio de Psicologia do Desporto, Faculdade de Motricidade Humana,Universidade Técnica de Lisboa, Portugal. E-mail: [email protected]
Ian Renshaw worked as a Lecturer in the Centre for Sport and Exercise Sciences at Shef-field Hallam University, UK, between 2000 and 2003. He is currently a Senior ResearchLecturer in the New Zealand Institute of Sport and Recreation Research at the AucklandUniversity of Technology, where he is in charge of the Coaching Stream. Ian’s currentresearch interests include the visual regulation of gait in sporting tasks, perceptual train-ing and the development of skill through nonlinear pedagogy. Address: Division of Sportand Recreations, Auckland University of Technology, Private Bag 92006, Auckland 1020,New Zealand. E-mail: [email protected]
Robert Hristovski is currently an Associate Professor at the Faculty of Physical Culturein the University of St. Cyril and Methodius in the city of Skopje, Republic of Macedonia.He was first appointed to the University in 1989 where he now teaches classes in the The-ory and Methodology of Sport Training. He obtained his Master of Science degree (Octo-ber 1994) and Ph.D. (July 1997) at the Faculty of Physical Culture and has researchinterests in methods of analysis used in nonlinear dynamics, particularly as applied tomodeling of human movements and adaptation to the training process. Address: Facultyof Physical Culture, University of St.Cyril and Methodius, Zeleznicka BB, Skopje 1000,Republic of Macedonia. E-mail: [email protected]
Related Documents











