36 © 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology Veterinary Dermatology 2006, 17, 36–44 BlackwellPublishingLtd Mouse epidermal develo pment: effects of retinoic acid exposure in utero ROSA A. GARCÍA-FERNÁNDEZ*, CLAUDIA PÉ REZ-MARTÍNEZ†, JA VIER ESPINOSA ÁLV AREZ †, ALEX J. DURÁN NAVARRET E† and MARIA J. GARCÍA-I GLE SIAS† *Histology and Pathological Anatomy Section, Department of Animal Medicine and Surgery, F aculty of Vet erinary Science, University of Madrid (UCM), Madrid, Spain, †Histology and Pathological Anatomy Section, Department of Animal Pathology , Animal Medicine, Faculty of Veterinary Science, University of León, León, Spain (Received 20 May 2005; accepted 24 November 2005) Abstract Epidermal morphogenesis was studied in vivo following prenatal exposure to retinoic acid (RA). In pregnant mice, a single oral dose of RA on day 11.5 of gestation failed to induce histological changes in fetal epidermal development except in epidermal thickness. Epidermal thickness increased from 16.5 days post-coitum (dpc) onwards, and temporal and spatial epidermal modifications in keratins K5 and K14 related to proliferativ e activity of keratinocytes were observed. An RA effect on cell proliferation was supported by a statistically significant increase in the number of epidermal S-phase cells, containing BrdU–incorporated DNA in RA- exposed mice compared with nonexposed animal s. The prolonged in utero action of RA on epidermal prolifer- ative activity in fetuses and newborns suggests a long-term RA effect that may play a role on the development and evolution of diseases in adult skin. INTRODUCTION Retinoic acid (RA), the most biologically active natural metabolite of vitamin A, plays a fundamental role in embryogenesis and in the differentia tion of tissues, includ- ing the skin, and is involved in the control of epithelial cell growth. 1 RA is used therapeutically for dermatological 2 and neoplastic diseases, 3 although the clinical usefulness of retinoids is limited by their teratogenic potential. 4–6 Postnatal studies on the effects of retinoids in dermato- logy and oncology have been undertaken in animals 7,8 and humans, 9,10 following topical application or oral administration, but knowledge of the in vivo effects on skin exposed to RA in utero is limited to mouse vibrissal 11 and pelage hair follicle 12 development. The physiological retinoid, RA, media tes in a wide variety of activities such as the proliferation of epithelial cells. 13 It plays a role in the expression of keratin (K) proteins in kera tinocytes, 14 which express different types of RA receptors. 15 These intermediate filaments have therefore been used as tools to investigate cutaneous morphogenesis and differenti- ation. 16 Thus, the present aim was to determine the in vivo effects of prenatal RA exposure on epidermal development by analysing changes in morphology, keratin expression and proliferative activity in Navy Marine Research Institute (NMRI) mice, thereby complementing previous studies on the effects of this retinoid on vibrissal 11 and pelage hair follicles 12 and widening the knowledge of its local, systemic and temporal actions on the skin. METHODS Animals and treatmen t Six- to 8-week-old albino NMRI mice (n = 90), weighing 30–35 g, were obtained from Antibióticos Laboratories S.A. (León, Spain). All were maintained in a cycle of 12 h light and 12 h dark at a controlled temperature (21 ± 1 °C) and relative humidity (55 ± 10%) with free access to food and water. All experiments were carried out following the guidelines of the European Law on the Protection of Animals . 17 To obtain timed pregnancies, individual female mice were placed randomly with individual male mice at 20.00 h and inspected daily thereafter at 09.00 h for the appearance of a vaginal plug, which indicated mating. Appearance of the plug was designated day 0.5 of pregnancy . The pregnant mice were treated by oral administra tion with 30 mg kg −1 body weight of RA (Sigma Chemical Co., St Louis, MO, USA) in corn oil on day 11.5 of gestation. This procedure was carried out in the dark under dim yellow light to retard photodegradation. Two control groups were established at the same gesta- tion time by treating pregnant mice with an identical volume of the corn oil vehicle and by leavin g one group untreated. Five embryos across different litters from each group (RA-exposed, vehicle-exposed and nonexposed) were obtained daily from the mothers post-mortem beginning at gestational day 12.5 until day 18.5. In Correspondence: Dr Rosa Garcia-Fernandez, Histology and Patho- logical Anatomy Section, Department of Animal Pathology, Animal Medicine and Surgery, Faculty of Veterinary Science, University of Madrid (UCM), Madrid, Spain. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 1/9
36 © 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Veterinary Dermatology 2006, 17, 36–44
BlackwellPublishingLtd
Mouse epidermal development: effects ofretinoic acid exposure in utero
ROSA A. GARCÍA-FERNÁNDEZ*, CLAUDIA PÉ REZ-MARTÍNEZ†, JAVIER ESPINOSAÁLVAREZ †, ALEX J. DURÁN NAVARRET E† and MARIA J. GARCÍA-I GLE SIAS†
*Histology and Pathological Anatomy Section, Department of Animal Medicine and Surgery, Faculty ofVeterinary Science, University of Madrid (UCM), Madrid, Spain, †Histology and Pathological Anatomy
Section, Department of Animal Pathology, Animal Medicine, Faculty of Veterinary Science,University of León, León, Spain
(Received 20 May 2005; accepted 24 November 2005)
Abstract Epidermal morphogenesis was studied in vivo following prenatal exposure to retinoic acid (RA). Inpregnant mice, a single oral dose of RA on day 11.5 of gestation failed to induce histological changes in fetalepidermal development except in epidermal thickness. Epidermal thickness increased from 16.5 days post-coitum(dpc) onwards, and temporal and spatial epidermal modifications in keratins K5 and K14 related to proliferative
activity of keratinocytes were observed. An RA effect on cell proliferation was supported by a statisticallysignificant increase in the number of epidermal S-phase cells, containing BrdU–incorporated DNA in RA-exposed mice compared with nonexposed animals. The prolonged in utero action of RA on epidermal prolifer-ative activity in fetuses and newborns suggests a long-term RA effect that may play a role on the developmentand evolution of diseases in adult skin.
INTRODUCTION
Retinoic acid (RA), the most biologically active naturalmetabolite of vitamin A, plays a fundamental role in
embryogenesis and in the differentiation of tissues, includ-ing the skin, and is involved in the control of epithelialcell growth.1
RA is used therapeutically for dermatological2 andneoplastic diseases,3 although the clinical usefulness of retinoids is limited by their teratogenic potential.4–6
Postnatal studies on the effects of retinoids in dermato-logy and oncology have been undertaken in animals7,8
and humans,9,10 following topical application or oraladministration, but knowledge of the in vivo effects onskin exposed to RA in utero is limited to mouse vibrissal11
and pelage hair follicle12 development. The physiological
retinoid, RA, mediates in a wide variety of activities suchas the proliferation of epithelial cells.13 It plays a role inthe expression of keratin (K) proteins in keratinocytes,14
which express different types of RA receptors.15 Theseintermediate filaments have therefore been used as toolsto investigate cutaneous morphogenesis and differenti-ation.16 Thus, the present aim was to determine thein vivo effects of prenatal RA exposure on epidermaldevelopment by analysing changes in morphology,keratin expression and proliferative activity in NavyMarine Research Institute (NMRI) mice, therebycomplementing previous studies on the effects of this
retinoid on vibrissal11 and pelage hair follicles12 andwidening the knowledge of its local, systemic andtemporal actions on the skin.
METHODS
Animals and treatmentSix- to 8-week-old albino NMRI mice (n = 90), weighing30–35 g, were obtained from Antibióticos LaboratoriesS.A. (León, Spain). All were maintained in a cycle of 12 h light and 12 h dark at a controlled temperature(21 ± 1 °C) and relative humidity (55 ± 10%) with freeaccess to food and water. All experiments were carriedout following the guidelines of the European Law on theProtection of Animals.17 To obtain timed pregnancies,
individual female mice were placed randomly withindividual male mice at 20.00 h and inspected dailythereafter at 09.00 h for the appearance of a vaginalplug, which indicated mating. Appearance of the plugwas designated day 0.5 of pregnancy.
The pregnant mice were treated by oral administrationwith 30 mg kg−1 body weight of RA (Sigma ChemicalCo., St Louis, MO, USA) in corn oil on day 11.5 of gestation. This procedure was carried out in the darkunder dim yellow light to retard photodegradation.Two control groups were established at the same gesta-tion time by treating pregnant mice with an identicalvolume of the corn oil vehicle and by leaving one group
untreated. Five embryos across different litters from eachgroup (RA-exposed, vehicle-exposed and nonexposed)were obtained daily from the mothers post-mortembeginning at gestational day 12.5 until day 18.5. In
Correspondence: Dr Rosa Garcia-Fernandez, Histology and Patho-logical Anatomy Section, Department of Animal Pathology, AnimalMedicine and Surgery, Faculty of Veterinary Science, Universityof Madrid (UCM), Madrid, Spain. E-mail: [email protected]

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 2/9
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Mouse epidermal development after RA exposure 37
addition, three to seven pregnant mice from eachgroup were allowed to give birth, and their offspringwere euthanized at birth (0) and at 1 day of age. In eachgroup, five animals from different litters were analysedfor each age.
To assess epidermal cell proliferation, 2 h beforeembryo and offspring collection, all pregnant mice andoffspring were given an intraperitoneal injection of 5-bromo-2′-deoxyuridine (BrdU) (Sigma Chemical Co.)(1 mg per 20 g body weight in 150 µL of saline solution).BrdU is incorporated into newly synthesized strands of DNA of S-phase cells in vivo.
HistologyFetuses and newborn animals were fixed in Bouin’ssolution or in 70% ethanol and processed routinely to
paraffin wax. Sections (3 µm) were cut at three differ-ent levels in each instance: one midsagittal and twoparasagittal (one when an eye was seen and the otherwhen the stomach lumen was observed), and stainedwith haematoxylin and eosin (H&E) and Masson’strichrome stains.
ImmunohistochemistryFrom day 12.5 of gestation to 1 day of age, an immuno-histochemical study was carried out on epidermisfrom the seven body regions cited previously in themorphometric study. Midsagittal sections, mounted on
poly
-lysine-coated slides, were subjected to immuno-histochemical staining using the avidin–biotin–peroxidasecomplex (ABC) method (Peroxidase Standard, Vectastain,ABC Kit, Vector Laboratories, Burlingame, CA, USA).Table 1 shows the working dilution and the character-istics of the primary antibodies used. The immuno-reactivity of masked antigens was restored by treatingsections with a solution of 0.1% trypsin (Sigma ChemicalCo.) for monoclonal mouse anti-BrdU antibody or withheat18 for the other antibodies used. After the sequentialincubations with the primary and secondary anti-bodies and the avidin–biotin–peroxidase complex, theimmunoreactivity was demonstrated using diaminoben-
zidine (DAB, substrate kit for peroxidase, VectorLaboratories) as chromogen to produce a brown stainsuitable for light microscopy, and the nuclei werecounterstained with haematoxylin.
Negative and positive control sections for the primaryand secondary antibodies were included. Adult mouseepidermis was used as positive control for all antikeratinantibodies except anti-K6 and anti-K8. A hyperplasicmouse epidermis and mouse hepatic bile ducts were usedas positive controls for anti-K6 and anti-K8 antibodies,respectively. A mouse previously inoculated with BrdUwas used as positive control for anti-BrdU antibody;for corresponding negative control sections, the primaryantibody was replaced with Tris-buffered saline (TBS).
Morphometric study: epidermal thicknessFrom day 12.5 of gestation to 1 day of age, livingepidermal thickness (i.e. without stratum corneum)was measured in the midsagittal tissue section of eachanimal using an ocular micrometre (PK 12.5 × m) (C.
Richert, Austria) calibrated with a stage micrometre.Ten measures on three different sections in each animalwere recorded in each of seven different body regions:cranium, back, abdomen, thorax, neck, mandible andmuzzle, except in the back where 20 were made.
Epidermal cell kineticsFrom day 12.5 of gestation to 1 day of age, the numberof BrdU-positive nuclei per area (µm2) was counted infive ×400 fields of the epidermis of a midsagittal tissuesection of each animal using an image analysis system( version 3.0, Soft Imaging System, Münster,
Germany). Quantification was undertaken on threedifferent body regions: cranial (frontal bone–atlas length),dorsal (atlas–rump length) and ventral (abdomen– thorax–neck length). Cells that had incorporatedBrdU were identified by the precipitation of a brownsubstance in their nuclei.
Statistical analysisOne-way
19 was used for each age (fetuses andnewborns) to test whether the mean of epidermalthickness and BrdU-positive nuclei per area (µm2) of epidermis differed among the groups examined (RA-exposed, vehicle-exposed and nonexposed). As no
significant difference was found between the meansof the two control groups (vehicle-exposed andnonexposed), the average value of both groups was usedfor comparison with the mean of RA-exposed group.
Table 1. Primary antibodies used for immunohistochemical labelling
Clone Character/species Specificity Dilution† Source
TROMA 1* mo/R K8 ud R. Kemler‡AF 109 po/Rb K1 1 in 2 × 105 BabCO, CA, USAAF 138 po/Rb K5 1 in 105 BabCO, CA, USAMK6 po/Rb K6 1 in 105 BabCO, CA, USA
MK10 po/Rb K10 1 in 4 × 10
5
BabCO, CA, USAAF 64 po/Rb K14 1 in 105 BabCO, CA, USAZBU 30 mo/M S-phase cells 1 in 2 × 102 Zymed Laboratories, San Francisco, CA, USA
mo, monoclonal; po, polyclonal; K, keratin; M, mouse; R, rat; Rb, rabbit; ud, undiluted.*70% ethanol-fixed tissues fetuses.†µL in µL (microlitre in microlitre).‡Dr R. Kemler, Max Planck Institute, Freiburg, Germany.
8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 3/9
38 RA García-Fernández et al .
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Multifactor 19 was carried out to analyse epidermalthickness, using body region as covariate. Differenceswere considered statistically significant at P < 0.05. Datawere expressed as mean ± standard error of the mean(SEM). Analysis was carried out using
version 2.0.
RESULTS
Histogenesis and immunohistochemistryThe main histological features of epidermal develop-ment could be broadly divided into four stages, and thesequence did not differ between control and treatedgroups.
Stage 1. Twelve and a half to 14.5 days post-coitum
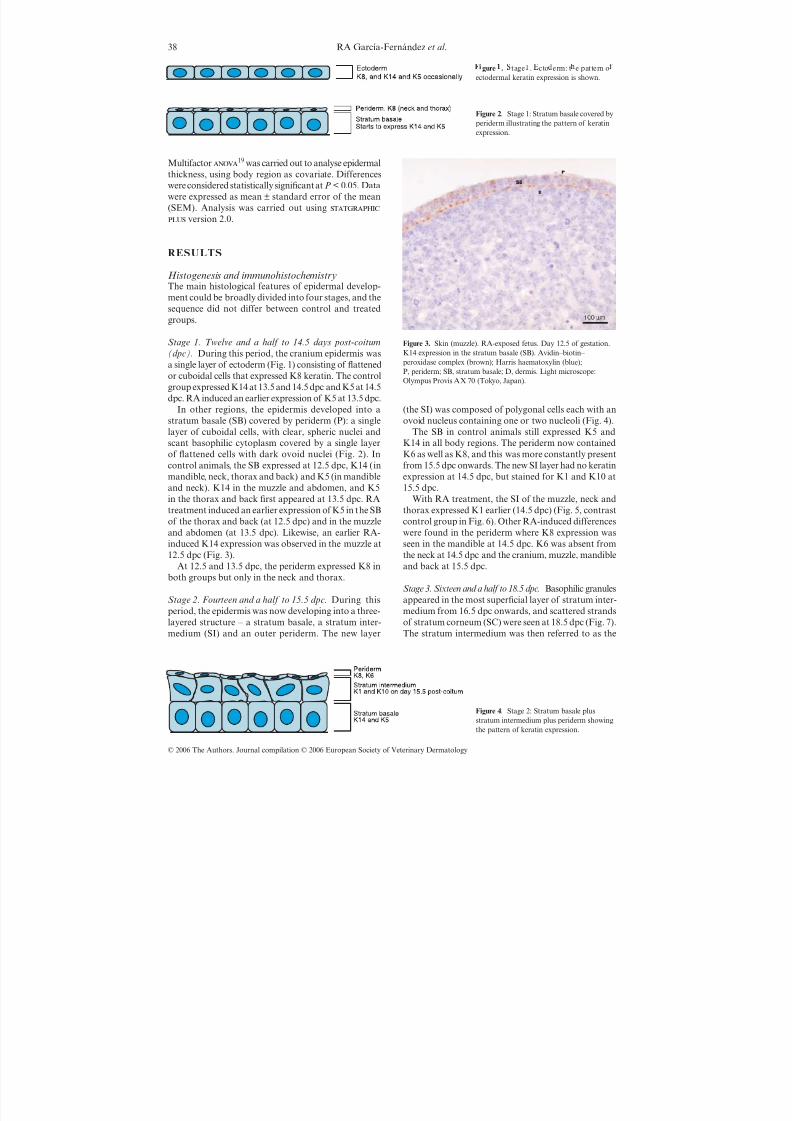
(dpc). During this period, the cranium epidermis wasa single layer of ectoderm (Fig. 1) consisting of flattenedor cuboidal cells that expressed K8 keratin. The controlgroup expressed K14 at 13.5 and 14.5 dpc and K5 at 14.5dpc. RA induced an earlier expression of K5 at 13.5 dpc.
In other regions, the epidermis developed into astratum basale (SB) covered by periderm (P): a singlelayer of cuboidal cells, with clear, spheric nuclei andscant basophilic cytoplasm covered by a single layerof flattened cells with dark ovoid nuclei (Fig. 2). Incontrol animals, the SB expressed at 12.5 dpc, K14 (inmandible, neck, thorax and back) and K5 (in mandibleand neck). K14 in the muzzle and abdomen, and K5in the thorax and back first appeared at 13.5 dpc. RAtreatment induced an earlier expression of K5 in the SBof the thorax and back (at 12.5 dpc) and in the muzzleand abdomen (at 13.5 dpc). Likewise, an earlier RA-induced K14 expression was observed in the muzzle at
12.5 dpc (Fig. 3).At 12.5 and 13.5 dpc, the periderm expressed K8 in
both groups but only in the neck and thorax.
Stage 2. Fourteen and a half to 15.5 dpc. During thisperiod, the epidermis was now developing into a three-layered structure – a stratum basale, a stratum inter-medium (SI) and an outer periderm. The new layer
(the SI) was composed of polygonal cells each with anovoid nucleus containing one or two nucleoli (Fig. 4).
The SB in control animals still expressed K5 andK14 in all body regions. The periderm now containedK6 as well as K8, and this was more constantly presentfrom 15.5 dpc onwards. The new SI layer had no keratinexpression at 14.5 dpc, but stained for K1 and K10 at15.5 dpc.
With RA treatment, the SI of the muzzle, neck andthorax expressed K1 earlier (14.5 dpc) (Fig. 5, contrastcontrol group in Fig. 6). Other RA-induced differenceswere found in the periderm where K8 expression wasseen in the mandible at 14.5 dpc. K6 was absent from
the neck at 14.5 dpc and the cranium, muzzle, mandibleand back at 15.5 dpc.
Stage 3. Sixteen and a half to 18.5 dpc. Basophilic granulesappeared in the most superficial layer of stratum inter-medium from 16.5 dpc onwards, and scattered strandsof stratum corneum (SC) were seen at 18.5 dpc (Fig. 7).The stratum intermedium was then referred to as the
gure . tage . cto erm: t e pattern oectodermal keratin expression is shown.
Figure 2. Stage 1: Stratum basale covered byperiderm illustrating the pattern of keratinexpression.
Figure 4. Stage 2: Stratum basale plusstratum intermedium plus periderm showingthe pattern of keratin expression.
Figure 3. Skin (muzzle). RA-exposed fetus. Day 12.5 of gestation.K14 expression in the stratum basale (SB). Avidin–biotin– peroxidase complex (brown); Harris haematoxylin (blue);P, periderm; SB, stratum basale; D, dermis. Light microscope:Olympus Provis AX 70 (Tokyo, Japan).

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 4/9
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Mouse epidermal development after RA exposure 39
suprabasal layer or stratum suprabasale (SS). Peridermwas only observed when there was no stratum corneum.In controls, the SB continued to express K5 and K14
and the SI K1 and K10. Periderm still contained K8and K6, whereas the stratum corneum always remainedunstained.
Stage 4. Postnatal 0–1 day of age. The postnatal epider-mis consisted of basal and suprabasal layers, where amorphological difference between the stratum spinosumand granulosum was clear, and a stratum corneum waspresent (Fig. 8).
As in the prenatal situation, the SS still expressed K1
and K10 and the SB K5 and K14 in all body regions.Scattered K6 expression in the SS was seen at day 0 of age (neck, thorax and abdomen) and at 1 day of age(cranium, neck, thorax, abdomen and back). There wasno expression in the stratum corneum.
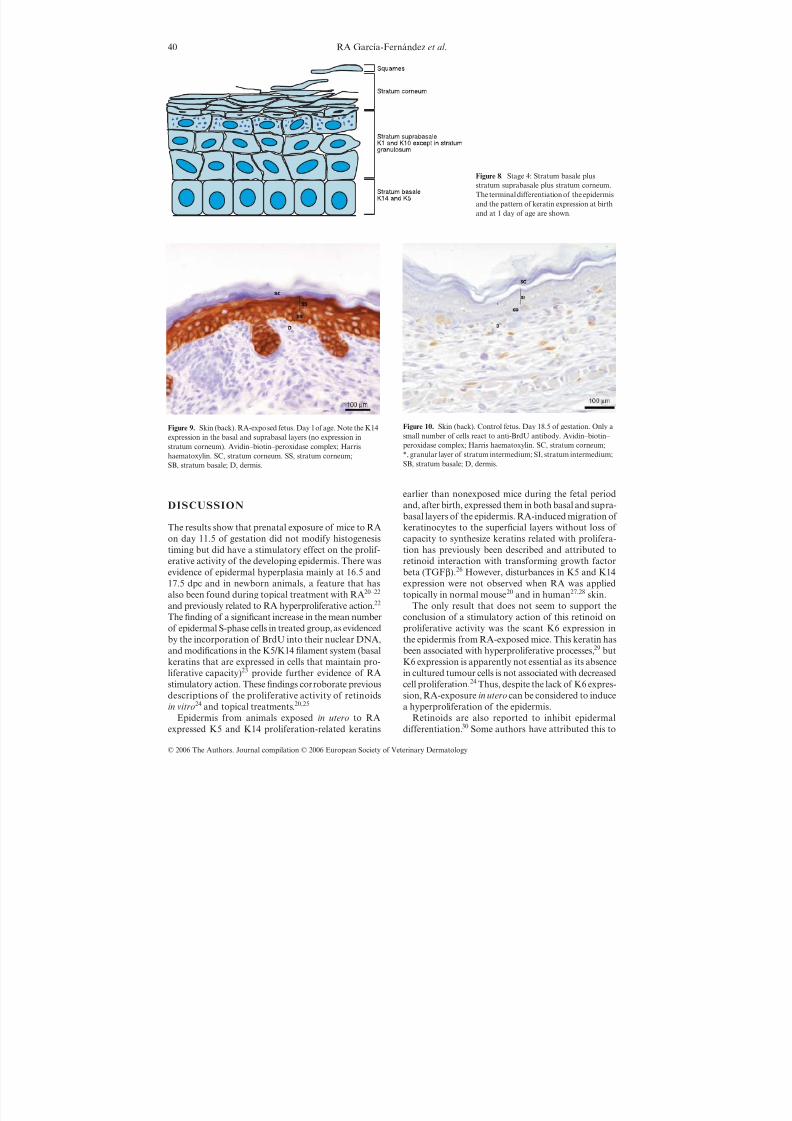
RA treatment caused expression of K14 and K5 inthe suprabasal layer (Fig. 9) in addition to the normalK1 and K10 expression. The first K6-positive suprabasalcells in RA-exposed mice were seen on 1 day of age infewer body regions than in control groups.
In summary, during prenatal development, RAtreatment induced an earlier expression of K5 andK14 than in controls, and these keratins persisted inthe SS after birth. RA treatment also caused an earlierexpression of K1 and decreased K6 expression inthe SS.
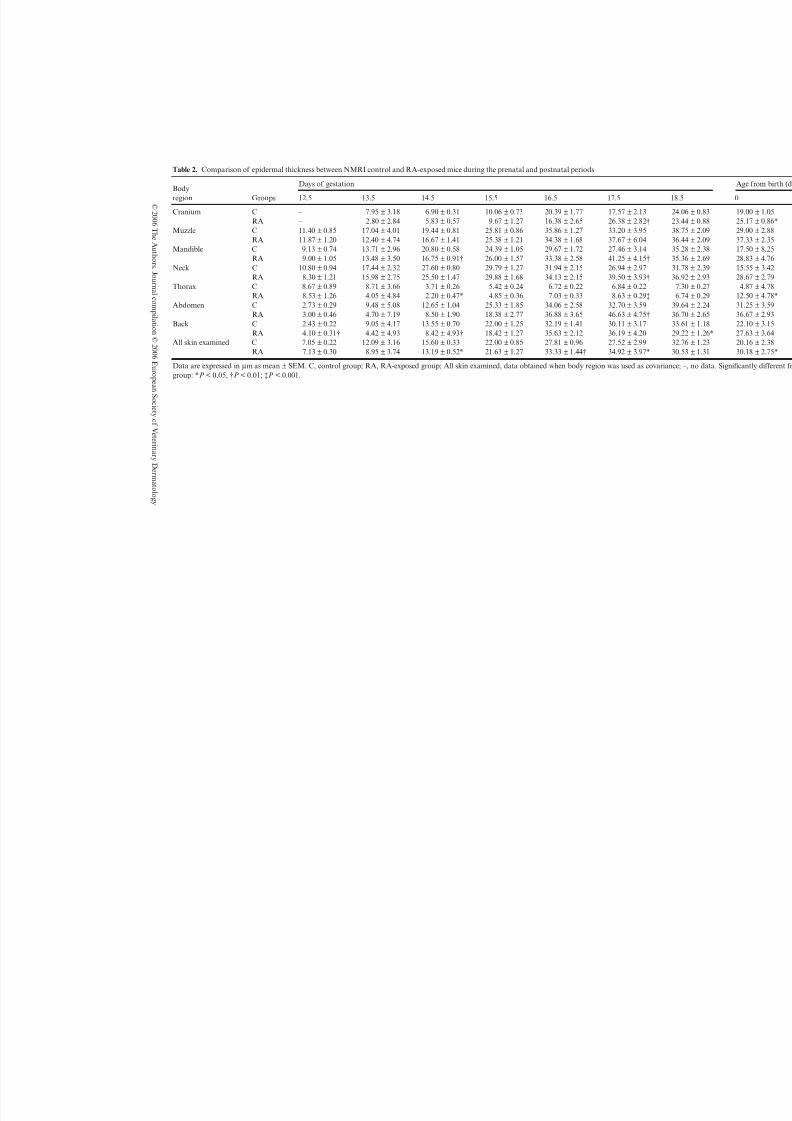
Morphometric study: epidermal thicknessTable 2 shows the mean epidermal thickness in controland RA-exposed groups. One-way for each bodyregion revealed a significant epidermal increase at 12.5dpc (back), 17.5 dpc (cranium, mandible, neck, thoraxand abdomen) and 0 day of age (cranium and thorax),whereas a significant decrease was shown at 14.5 dpc(mandible, thorax and back) and 18.5 dpc (back), both
in RA-exposed animals. When body region was used ascovariate (multifactor), a statistically significantincrease was demonstrated in all skin examined inRA-exposed mice at 16.5 and 17.5 dpc and 0 day of age, whereas a significant decrease was only shown at14.5 dpc.
Epidermal cell kineticsMore BrdU-positive cells were seen in RA-exposedanimals (Figs 10 and 11), demonstrating an increase in S-phase cells and hence indicating proliferative activity.The means of BrdU-positive epidermal cells in RA-
exposed and nonexposed mice were used to comparethe proliferative activity in both groups (Table 3). Astatistically significant increase in proliferative activitywas found in the epidermis from all body regions andin both the prenatal and the postnatal periods follow-ing RA exposure.
Figure 5. Skin (muzzle). RA-exposed fetus. Day 14.5 of gestationshowing K1 expression in the stratum intermedium. Avidin–biotin– peroxidase complex; Harris haematoxylin. P, periderm; SI, stratum
intermedium; SB, stratum basale; D, dermis.
Figure 6. Skin (muzzle). Control fetus. Day 14.5 of gestation.No K1 expression in the stratum intermedium. Avidin–biotin– peroxidase complex; Harris haematoxylin. P, periderm;SI, stratum intermedium; SB, stratum basale; D, dermis.
Figure 7. Stage 3: Appearance of basophilicgranules in the most superficial layer of the
stratum intermedium (stratum granulosum)from 16.5 days post-coitum (dpc) onwardsand scattered strands of stratum corneum at18.5 dpc. The distribution of the layers andthe pattern of keratin expression are shown.
8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 5/9
40 RA García-Fernández et al .
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
DISCUSSION
The results show that prenatal exposure of mice to RAon day 11.5 of gestation did not modify histogenesis
timing but did have a stimulatory effect on the prolif-erative activity of the developing epidermis. There wasevidence of epidermal hyperplasia mainly at 16.5 and17.5 dpc and in newborn animals, a feature that hasalso been found during topical treatment with RA20–22
and previously related to RA hyperproliferative action.22
The finding of a significant increase in the mean numberof epidermal S-phase cells in treated group, as evidencedby the incorporation of BrdU into their nuclear DNA,and modifications in the K5/K14 filament system (basalkeratins that are expressed in cells that maintain pro-liferative capacity)23 provide further evidence of RAstimulatory action. These findings corroborate previous
descriptions of the proliferative activity of retinoidsin vitro24 and topical treatments.20,25
Epidermis from animals exposed in utero to RAexpressed K5 and K14 proliferation-related keratins
earlier than nonexposed mice during the fetal periodand, after birth, expressed them in both basal and supra-basal layers of the epidermis. RA-induced migration of keratinocytes to the superficial layers without loss of capacity to synthesize keratins related with prolifera-
tion has previously been described and attributed toretinoid interaction with transforming growth factorbeta (TGFβ).26 However, disturbances in K5 and K14expression were not observed when RA was appliedtopically in normal mouse20 and in human27,28 skin.
The only result that does not seem to support theconclusion of a stimulatory action of this retinoid onproliferative activity was the scant K6 expression inthe epidermis from RA-exposed mice. This keratin hasbeen associated with hyperproliferative processes,29 butK6 expression is apparently not essential as its absencein cultured tumour cells is not associated with decreasedcell proliferation.24 Thus, despite the lack of K6 expres-
sion, RA-exposure in utero can be considered to inducea hyperproliferation of the epidermis.
Retinoids are also reported to inhibit epidermaldifferentiation.30 Some authors have attributed this to
Figure 8. Stage 4: Stratum basale plusstratum suprabasale plus stratum corneum.The terminal differentiation of the epidermisand the pattern of keratin expression at birthand at 1 day of age are shown.
Figure 9. Skin (back). RA-exposed fetus. Day 1 of age. Note the K14expression in the basal and suprabasal layers (no expression instratum corneum). Avidin–biotin–peroxidase complex; Harrishaematoxylin. SC, stratum corneum. SS, stratum corneum;SB, stratum basale; D, dermis.
Figure 10. Skin (back). Control fetus. Day 18.5 of gestation. Only asmall number of cells react to anti-BrdU antibody. Avidin–biotin– peroxidase complex; Harris haematoxylin. SC, stratum corneum;*, granular layer of stratum intermedium; SI, stratum intermedium;SB, stratum basale; D, dermis.
8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 6/9
©2 0 0 6 T h e A u t h or s . J o ur n al c om pi l a t i on ©2 0 0 6 E ur o p e an S o c i e t y of V e t e r i n ar yD e r m a t ol o g y
Table 2. Comparison of epidermal thickness between NMRI control and RA-exposed mice during the prenatal and postnatal periods
Bodyregion Groups
Days of gestation
12.5 13.5 14.5 15.5 16.5 17.5 1
Cranium C – 7.95 ± 3.18 6.90 ± 0.31 10.06 ± 0.73 20.39 ± 1.77 17.57 ± 2.13 2RA – 2.80 ± 2.84 5.83 ± 0.57 9.67 ± 1.27 16.38 ± 2.65 26.38 ± 2.82† 2
Muzzle C 11.40 ± 0.85 17.04 ± 4.01 19.44 ± 0.81 25.81 ± 0.86 35.86 ± 1.27 33.20 ± 3.95 3RA 11.87 ± 1.20 12.40 ± 4.74 16.67 ± 1.41 25.38 ± 1.21 34.38 ± 1.68 37.67 ± 6.04 3
Mandible C 9.13 ± 0.74 13.71 ± 2.96 20.80 ± 0.58 24.39 ± 1.05 29.67 ± 1.72 27.46 ± 3.14 3RA 9.00 ± 1.05 13.48 ± 3.50 16.75 ± 0.91† 26.00 ± 1.57 33.38 ± 2.58 41.25 ± 4.15† 3
Neck C 10.80 ± 0.94 17.44 ± 2.32 27.60 ± 0.80 29.79 ± 1.27 31.94 ± 2.15 26.94 ± 2.97 3
RA 8.30 ± 1.21 15.98 ± 2.75 25.50 ± 1.47 29.88 ± 1.68 34.13 ± 2.15 39.50 ± 3.93† 3Thorax C 8.67 ± 0.89 8.71 ± 3.66 3.71 ± 0.26 5.42 ± 0.24 6.72 ± 0.22 6.84 ± 0.22
RA 8.53 ± 1.26 4.05 ± 4.84 2.20 ± 0.47* 4.85 ± 0.36 7.03 ± 0.33 8.63 ± 0.29‡Abdomen C 2.73 ± 0.29 9.48 ± 5.08 12.65 ± 1.04 25.33 ± 1.85 34.06 ± 2.58 32.70 ± 3.59 3
RA 3.00 ± 0.46 4.70 ± 7.19 8.50 ± 1.90 18.38 ± 2.77 36.88 ± 3.65 46.63 ± 4.75† 3Back C 2.43 ± 0.22 9.05 ± 4.17 13.55 ± 0.70 22.00 ± 1.25 32.19 ± 1.41 30.11 ± 3.17 3
RA 4.10 ± 0.31† 4.42 ± 4.93 8.42 ± 4.93† 18.42 ± 1.27 35.63 ± 2.12 36.19 ± 4.20 2All skin examined C 7.05 ± 0.22 12.09 ± 3.16 15.60 ± 0.33 22.00 ± 0.85 27.81 ± 0.96 27.52 ± 2.99 3
RA 7.13 ± 0.30 8.95 ± 3.74 13.19 ± 0.52* 21.63 ± 1.27 33.33 ± 1.44† 34.92 ± 3.97* 3
Data are expressed in µm as mean ± SEM. C, control group; RA, RA-exposed group; All skin examined, data obtained when body region was used as covariangroup: *P < 0.05, †P < 0.01; ‡P < 0.001.

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 7/9
42 RA García-Fernández et al .
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
RA inhibition of normal regulation of signalling in thebasal layer, associated with RA-induced K6 expressionin epidermis in vivo.24 In this study, reduced K6 expressionfollowing RA treatment indicates that RA does nothave an inhibitory effect on epidermal differentiation.
Changes in keratinocyte maturation have beendemonstrated in previous in vitro studies with RAinducing a decrease in the expression of K131,32 or K1033
or both.24,28,34 This action has also been observed inin vivo studies after topical RA treatment of adult
epidermis.31,35 However, in this study, RA induced anearlier epidermal K1 expression in fetuses. This indi-cates an earlier RA-induced epidermal differentiationin contrast to a delay in the terminal differentiation
of keratinocytes as described previously for in vitro
treatment.36 They also contrast with the absence of aneffect on epidermal differentiation following in vivo RAtreatment in human skin.28,34
In conclusion, RA administered to pregnant miceinduced, in developing skin from their offspring, anearlier epidermal differentiation. It also stimulated cellproliferation both in the epidermis and in the pelage hair
follicle.12 Such prolonged RA action on skin proliferationraises the question as to whether RA-exposure in utero
could induce a long-term effect on adult skin, therebyplaying a role in development, evolution and prognosisof skin diseases, such as neoplasms, in adult animals.Longer-term studies of animals exposed to RA in utero
are needed to determine the long-term action of thisretinoid on adult mouse skin carcinogenesis.
ACKNOWLEDMENTS
This work was supported by grants from the Junta deCastilla y León (España) (LE04/94). The authors thankDr Miguel Angel Vidal Caballero of the Centro deInvestigaciones Biológicas (Madrid, Spain) for supplyingthe anti-K8 antibody and Dr Juan García Vieitez of theDepartment of Pharmacology, Toxicology and Nursery(University of León, Spain) for advice on RA dosage.
REFERENCES
1. Reynolds NJ, Fisher GJ, Griffiths EM et al. Retinoicacid metabolites exhibit biological activity in humankeratinocytes, mouse melanoma cells and hairless mouseskin in vivo. Journal of Pharmacology and ExperimentalTherapeutics 1993; 266: 1636– 42.
2. Sitzmann JH, Bauer FW, Cunliff WJ et al. In situ
Figure 11. Skin (back). RA-exposed fetus. Day 18.5 of gestation.Many more cells react positively for anti-BrdU than in controlanimals. Avidin–biotin–peroxidase complex; Harris haematoxylin.SC, stratum corneum; *, granular layer of stratum intermedium;SI, stratum intermedium; SB, stratum basale; D, dermis.
Table 3. BrdU-positive nuclei per µm of epidermis from NMRI control and RA-exposed mice during prenatal and postnatal periods
Age (days) Groups
BrdU-positive nuclei per µm2 according to body region
Cranial Dorsal Ventral
12.5 dpc C 1.7 × 10−2 ± 1.6 × 10−2 2.6 × 10−2 ± 1.2 × 10−2 2.6 × 10−2 ± 1.2 × 10−2
RA 8.5 × 10−2 ± 1.5 × 10−2* 8.3 × 10−2 ± 0.6 × 10−2† 14 × 10−2 ± 1.5 × 10−2†13.5 dpc C 2.5 × 10−2 ± 0.5 × 10−2 3.3 × 10−2 ± 0.5 × 10−2 3.5 × 10−2 ± 0.8 × 10−2
RA 4.2 × 10−2 ± 0.4 × 10−2* 7.1 × 10−2 ± 0.4 × 10−2‡ 10 × 10−2 ± 0.7 × 10−2‡14.5 dpc C 2.4 × 10−2 ± 0.3 × 10−2 2.3 × 10−2 ± 0.5 × 10−2 4.7 × 10−2 ± 0.7 × 10−2
RA 6.4 × 10−2 ± 0.4 × 10−2‡ 10.2 × 10−2 ± 0.6 × 10−2‡ 10.4 × 10−2 ± 0.7 × 10−2‡15.5 dpc C 3.2 × 10−2 ± 0.2 × 10−2 4.3 × 10−2 ± 0.3 × 10−2 5.5 × 10−2 ± 0.6 × 10−2
RA 7.0 × 10−2 ± 0.2 × 10−2† 7.9 × 10−2 ± 0.3 × 10−2† 11 × 10−2 ± 0.6 × 10−2†16.5 dpc C 2.2 × 10−2 ± 0.3 × 10−2 3.0 × 10−2 ± 0.3 × 10−2 3.7 × 10−2 ± 0.4 × 10−2
RA 4.1 × 10−2 ± 0.2 × 10−2† 5.1 × 10−2 ± 0.2 × 10−2† 6.2 × 10−2 ± 0.3 × 10−2†17.5 dpc C 1.8 × 10−2 ± 0.2 × 10−2 1.9 × 10−2 ± 0.2 × 10−2 3.3 × 10−2 ± 0.3 × 10−2
RA 3.8 × 10−2 ± 0.2 × 10−2† 4.1 × 10−2 ± 0.2 × 10−2† 6.1 × 10−2 ± 0.3 × 10−2†18.5 dpc C 2.0 × 10−2 ± 0.2 × 10−2 1.9 × 10−2 ± 0.2 × 10−2 3.0 × 10−2 ± 0.3 × 10−2
RA 3.0 × 10−2 ± 0.2 × 10−2* 2.9 × 10−2 ± 0.2 × 10−2† 4.0 × 10−2 ± 0.3 × 10−2*0 C 0.7 × 10−2 ± 0.1 × 10−2 1.2 × 10−2 ± 0.1 × 10−2 1.7 × 10−2 ± 0.2 × 10−2
RA 1.8 × 10−2 ± 0.1 × 10−2‡ 1.7 × 10−2 ± 0.1 × 10−2* 2.7 × 10−2 ± 0.2 × 10−2‡1 C 1.2 × 10−2 ± 0.2 × 10−2 1.1 × 10−2 ± 0.1 × 10−2 1.8 × 10−2 ± 0.2 × 10−2
RA 2.0 × 10−2 ± 0.2 × 10−2† 1.8 × 10−2 ± 0.2 × 10−2* 2.8 × 10−2 ± 0.3 × 10−2*
Data are expressed as mean ± SEM. dpc, day post-coitum. 0, day of birth; 1, 1 day of age; C, control group; RA, RA-exposed group. Significantlyhigher results were found in the RA group at all ages in the three regions studied (* P < 0.05; †P < 0.001; ‡P < 0.0001).

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 8/9
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Mouse epidermal development after RA exposure 43
hybridization analysis of CRABPII expression in sebaceousfollicles from 13-cis retinoic acid-treated acne patients.British Journal of Dermatology 1995; 133: 241–8.
3. Hill DL, Shin T, Lin T et al. Retinoids and cancer pre-vention. Annual Review of Nutrition 1995; 12: 161–81.
4. Hendrickx AG, Hummler H. Teratogenicity of all-transretinoic acid during early embryonic development in the
cynomolgus monkey (Macaca fascicularis). Teratology1992; 45: 65–74.
5. Kochhar DM, Jiang H, Penner JD et al. Differentialteratogenic response of mouse embryos to receptorselective analogs of retinoic acid. Chemico-BiologicalInteractions 1996; 100: 1–12.
6. Rosa FW. Teratogenicity of isotretionin. Lancet 1983; 2: 513.7. Verma AK. Inhibition of both stage I and stage II mouse
skin tumor promotion by retinoic acid and the dependenceof inhibition of tumor promotion on the duration of retinoicacid treatment. Cancer Research 1987; 47: 5097–101.
8. De Luca LM, Sly L, Jones CS et al. Effects of dietaryretinoic acid on skin papilloma and carcinoma formation in
female SENCAR mice. Carcinogenesis 1993; 14: 539–42.9. Kligman AM, Fulton JE, Plewig G. Topical vitamin A acidin acne vulgaris. Archives of Dermatology 1969; 19: 469–76.
10. Peck GL, Olsen TG, Yoder FW. Prolonged remissions of cystic and conglobate acne with 13-cis-retinoic acid. NewEngland Journal of Medicine 1979; 300: 329–33.
11. García-Fernández RA, Pérez-Martínez C, Escudero-Díez A et al. Effects of retinoic acid exposure in utero
on mouse vibrissal follicle development. Anatomia,Histologia, Embryologia 2000; 29: 207–10.
12. García-Fernández RA, Pérez-Martínez C, Escudero-Díez A et al. Effects of in utero retinoic acid exposureon mouse pelage hair follicle development. VeterinaryDermatology 2002; 13: 157–63.
13. Yaar M, Stanley JR, Katz SI. Retinoic acid delays theterminal differentiation of keratinocytes in suspensionculture. Journal of Investigative Dermatology 1981; 76:363–6.
14. Stellmach V, Leask A, Fuchs E. Retinoid-mediatedtranscriptional regulation of keratin genes in humanepidermal and squamous cell carcinoma cells. Proceed-ings of the National Academy of Sciences of the UnitedStates of America 1991; 88: 4582–6.
15. Viallet JP, Dhouailly D. Retinoic acid and mouse skinmorphogenesis. I. Expression pattern of retinoic acidreceptor genes during hair vibrissa follicle, plantar, andnasal gland development. Journal of Investigative
Dermatology 1994; 103: 116–21.16. Kopan R, Fuchs E. A new look into and old problem:keratins as tools to investigate determination, morphogen-esis, and differentiation in skin. Genes and Development1989; 3: 1–15.
17. Council Directive 86/609 EEC.: Council Directive on theApproximation of Laws, Regulation and AdministrativePrevisions of the Member States Regarding the Protectionof Animals used for Experimental and other ScientificPurposes (86/609/EEC). The Council of the EuropeanCommunities. Brussels, Belgium, 1986.
18. Auld J. Antigen unmasking in routinely processedparaffin sections by pressure cooking. United KingdomNational External Quality Assessment Schemes-ICC
1994; 17: 197–209.19. Martín Andrés A, Luna del Castillo JD. Bioestadística
para las ciencias de la salud, 4th edn. Madrid, NormaSA, 1994: 383–450.
20. Eichner R, Kahn M, Capetola RJ et al. Effects of topicalretinoids on cytoskeletal proteins: implications forretinoid effects on epidermal differentiation. Journal of Investigative Dermatology 1992; 98: 154–61.
21. Craven NM, Griffiths CEM. Topical retinoids andcutaneous biology. Clinical and Experimental Derma-tology 1996; 21: 1–10.
22. Ashton RE, Connor MJ, Lowe NJ. Histologic changes inthe skin of the Rhino mouse (hrrh hrrh) induced by retinoids.Journal of Investigative Dermatology 1984; 82: 632–5.
23. Fuchs E, Byrne C. The epidermis: rising to the surface.Current Opinion in Genetics and Development 1994; 4:725–36.
24. Kopan R, Fuchs E. The use of retinoic acid to probe therelation between hyperproliferation-associated keratinsand cell proliferation in normal and malignant epidermalcell. Journal of Cell Biology 1989; 109: 295–307.
25. Fisher CJ, Esmann J, Griffiths CEM et al. Cellular,immunologic and biochemical characterization of topical retinoic acid-treated human skin. Journal of
Investigative Dermatology 1991; 96: 699–707.26. Fisher GJ, Voorhees JJ. Molecular mechanisms of retinoidactions in skin. FASEB Journal 1996; 10: 1002–13.
27. Rosenthal DS, Roop DR, Huff CA et al. Changes inphoto-aged human skin following topical applicationof all-trans retinoic acid. Journal of Investigative Derma-tology 1990; 95: 510–5.
28. Rosenthal DS, Griffiths CEM, Yuspa SH et al. Acute orchronic topical retinoic acid treatment of human skinin vivo alters the expression of epidermal transglutaminase,loricrin, involucrin, filaggrin, and keratins 6 and 13 butnot keratins 1, 10, and 14. Journal of InvestigativeDermatology 1992; 98: 343–50.
29. Moll R, Franke WW, Schiller L. The catalog of human
cytokeratins: patterns of expression in normal epithelia,tumors and cultured cells. Cell 1982; 31: 11–24.
30. Fuchs E, Green H. Regulation of terminal differentiationof cultured human keratinocytes by vitamin A. Cell1981; 25: 617–25.
31. Hieber AD, King TJ, Morioka S et al. Comparativeeffects of all-trans beta-carotene versus 9-cis beta-carotene on carcinogen-induced neoplastic transformationand connexin 43 expression in murine 10T1/2 cells andon the differentiation of human keratinocytes. Nutritionand Cancer – An International Journal 2000; 37: 234– 44.
32. Zou CP, Hong WK, Lotan R. Expression of retinoic acidreceptor beta is associated with inhibition of keratiniza-
tion in human head and neck squamous carcinoma cells.Differentiation 1999; 64: 123–32.33. Kunchala SR, Suzuki T, Murayama A. Photoisomeriza-
tion of retinoic acid and its influence on regulation of human keratinocyte growth and differentiation. IndianJournal of Bochemica and Biophysica 2000; 37: 71–6.
34. Virtanen M, Torma H, Valhquist A. Keratin 4 upregula-tion by retinoic acid in vivo: a sensitive marker for retinoidbioactivity in human epidermis. Journal of InvestigativeDermatology 2000; 114: 487–93.
35. Lenoir-Viale MC, Galup C, Darmon M et al. Epidermisreconstructed from the outer root sheath of human hairfollicle. Effect of retinoic acid. Archives of DermatologicalResearch 1993; 285: 197–204.
36. Yaar M, Stanley JR, Katz SI. Retinoic acid delays theterminal differentiation of keratinocytes in suspensionculture. Journal of Investigative Dermatology 1981; 76:363–6.

8/9/2019 Mouse Epidermal Development- Effects of Retinoic Acid Exposure in Utero (Pages 36–44)
http://slidepdf.com/reader/full/mouse-epidermal-development-effects-of-retinoic-acid-exposure-in-utero-pages 9/9
44 RA García-Fernández et al .
© 2006 The Authors. Journal compilation © 2006 European Society of Veterinary Dermatology
Résumé La morphogénèse épidermique a été étudiée in vivo en fonction de l’exposition prénatale à l’aciderétinoïque (RA). Chez les souris gestantes, une seule dose orale de RA au jour 11.5 de la gestation ne provoquaitpas d’anomalie du développement fétal, sauf au niveau de l’épaisseur cutanée. L’épaisseur cutanée a augmentée,de 16.5 jours post coitum (dpc) et par la suite et des modifications de kératines K5 et K14 à relier à la proliférationkératincoytaire ont été observées. Un effet de RA sur la prolifération cellulaire était confirmé par une augmen-tation significative du nombre de cellules épidermiques en phase S-contenant BrdU – incorporated DNA, chezles souris exposées au RA par rapport aux animaux non exposés. L’action prolongéein utero du RA sur l’activité
proliférative chez les foetus et les nouveaux nés suggère un effet à long terme du RA qui pourrait jouer un rôlesur le développement et l’évolution de maladies de la peau de l’adulte.
Resumen La morfogénesis de la epidermis se estudio in vivo en ratones tras ser expuestos durante el períodoprenatal a ácido retinoico. En ratones gestantes una sola dosis de ácido retinoico en el día 11.5 de gestación noprodujo cambios histológicos en el desarrollo fetal de la epidermis a excepción del grosor de la misma. El grosorde la epidermis aumentó desde el día 16 postcopula (dpc) en adelante, y se observaron modificaciones espacialesy temporales en la distribución de las queratinas K5 y K14, en relación con la actividad proliferativa de los que-ratinocitos. El efecto del RA en la proliferación celular en ratones expuestos a RA comparados con animales noexpuestos fue corroborado por un aumento significativo en el número de células epidérmicas en fase S (síntesis),indicado por la presencia de BrdU incorporado en el ADN. La prolongada actividad del RA in utero en laactividad proliferativa de la epidermis en fetos y neonatos sugiere un efecto prolongado del RA que puede jugarun papel relevante en el desarrollo y evolución de enfermedades en la piel de adultos.
Zusammenfassung Die epidermale Morphogenese wurde nach pränataler Exposition zu Retinolsäure (RA)invivo untersucht. Bei tragenden Mäusen konnte eine einzige orale Dosis von RA am 11,5. Tag der Trächtigkeitaußer bei der epidermalen Dicke keine histologischen Veränderungen bei der fötalen epidermalen Entwicklunginduzieren. Die epidermale Dicke nahm ab dem 16,5. Tag post coitum (dpc) zu. Es wurden zeitliche und räumlicheepidermale Modifikationen bei den Keratinen K5 und K14 beobachtet, die auf die proliferative Aktivität derKeratinozyten zurückzuführen waren. Eine Wirkung von RA auf die Zellproliferation bei RA-exponiertenMäusen im Vergleich zu nicht-exponierten Tieren wurde von einer statistisch signifikanten Zunahme der Anzahlan epidermalen S-Phase Zellen, die BrdU-inkorporierte DNA enthielten, unterstützt. Die anhaltende Wirkungvon RA in utero auf die epidermale proliferative Aktivität der Föten und Neugeborenen lässt auf eineLangzeitwirkung von RA schließen, die eine Rolle spielen könnte bei der Entwicklung und Evolution vonKrankheiten der adulten Haut.
Related Documents











