243 Morphology and biology of Polydora rickettsi (Polychaeta: Spionidae) from Chile VASILY I. RADASHEVSKY 1,2 CÉSAR A. CÁRDENAS 2 1 Institute of Marine Biology Russian Academy of Sciences Vladivostok 690041, Russia email: [email protected] 2 Instituto de Zoología Universidad Austral de Chile Valdivia, Chile Abstract The spionid polychaete Polydora rickettsi is primarily a borer in various calcareous substrata in the eastern Pacific. Redescription of the species from the type locality in Mexico is required to better understand its distribution. In Chile, females and males become mature after growth to c. 60 seg- ments. Sex allocation in the population is close to 1:1. Gametes develop along segmental blood vessels in middle segments. Spermatogonia proliferate in temporary testes and the rest of spermatogenesis occurs in the coelomic cavity. On their release through metanephridial segmental organs, long- headed spermatozoa are packed in filiform spermatophores. Females store inactive spermatozoa in paired seminal receptacles on the dorsal side of fertile segments. Oogenesis is mostly intraovarian. Females deposit up to 4150 eggs into 65 capsules which are joined to each other in a string. Each egg capsule is attached by two stalks to the inner wall of the burrow and contains up to 65 eggs c. 95 μm in diameter. Most eggs give rise to larvae which de- velop inside egg capsules until the 3-segment stage, then hatch and continue development in sea water, feeding on plankton. Fully developed 17–18-seg- ment larvae are able to settle and metamorphose. They have one pair of phaosomes and three pairs of black eyes on the prostomium, provisional proto- nephridia in segments 1 and 2, paired dorsal melanophores from segment 3 onwards, lateral melanophores on segment 2 and from segment 10 onwards, yellow pigment in the wall of posterior gut, glandular pouches from segment 6 onwards, short branchiae on segments 7–11, gastrotrochs on seg- ments 3, 5, 7, 10, 13, 15, and 17, one pair of provi- sional modified bristles and two or three falcate spines of a “quasi-adult” kind in segment 5. Devel- opment of the “quasi-adult” spines is described for the first time in polydorid larvae. Keywords Polychaeta; Spionidae; Polydora rickettsi; morphology; ecology; reproduction; larval development; Chile INTRODUCTION The spionid polychaete Polydora rickettsi was originally described by Woodwick (1961) from Cape San Lucas, Lower California, Mexico. The species was found intertidally associated with calcareous tubes of the serpulid polychaete Spirobranchus incrassatus Mörch. Presumably, it burrowed in these tubes. No other records or any information about the reproductive biology of P. rickettsi has been provided from Mexico. Later P. rickettsi was reported by Blake (1983) from Montemar, and by Sato-Okoshi & Takatsuka (2001) (as P. cf. rickettsi) from Puerto Montt and Chiloé Island, all reports from Chile (Fig. 1). Sato-Okoshi & Takatsuka (2001) reported briefly on reproduction and larval development of the species and suggested that further studies on specimens from Chile were required to determine if they are conspecific with P. rickettsi from Mexico. Reproductive biology and larval development of polychaetes on the Pacific side of South America and in the Strait of Magellan have been studied only in one species of Sabellidae (Gambi & Patti 1999) and some species of Spionidae. Among spionids, a brief note about mature pelagic stages of two species from Peru was provided by Berkeley & Berkeley (1963); pelagic larvae of 14 species from Concepción Bay, M03061; Online publication date 8 June 2004 Received 25 September 2003; accepted 18 January 2004 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38: 243–254 0028–8330/04/3802–0243 © The Royal Society of New Zealand 2004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

243Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
Morphology and biology of Polydora rickettsi(Polychaeta: Spionidae) from Chile
VASILY I. RADASHEVSKY1,2
CÉSAR A. CÁRDENAS2
1Institute of Marine BiologyRussian Academy of SciencesVladivostok 690041, Russiaemail: [email protected]
2Instituto de ZoologíaUniversidad Austral de ChileValdivia, Chile
Abstract The spionid polychaete Polydorarickettsi is primarily a borer in various calcareoussubstrata in the eastern Pacific. Redescription of thespecies from the type locality in Mexico is requiredto better understand its distribution. In Chile, femalesand males become mature after growth to c. 60 seg-ments. Sex allocation in the population is close to1:1. Gametes develop along segmental blood vesselsin middle segments. Spermatogonia proliferate intemporary testes and the rest of spermatogenesisoccurs in the coelomic cavity. On their releasethrough metanephridial segmental organs, long-headed spermatozoa are packed in filiformspermatophores. Females store inactive spermatozoain paired seminal receptacles on the dorsal side offertile segments. Oogenesis is mostly intraovarian.Females deposit up to 4150 eggs into 65 capsuleswhich are joined to each other in a string. Each eggcapsule is attached by two stalks to the inner wall ofthe burrow and contains up to 65 eggs c. 95 µm indiameter. Most eggs give rise to larvae which de-velop inside egg capsules until the 3-segment stage,then hatch and continue development in sea water,feeding on plankton. Fully developed 17–18-seg-ment larvae are able to settle and metamorphose.They have one pair of phaosomes and three pairs ofblack eyes on the prostomium, provisional proto-nephridia in segments 1 and 2, paired dorsal
melanophores from segment 3 onwards, lateralmelanophores on segment 2 and from segment 10onwards, yellow pigment in the wall of posterior gut,glandular pouches from segment 6 onwards, shortbranchiae on segments 7–11, gastrotrochs on seg-ments 3, 5, 7, 10, 13, 15, and 17, one pair of provi-sional modified bristles and two or three falcatespines of a “quasi-adult” kind in segment 5. Devel-opment of the “quasi-adult” spines is described forthe first time in polydorid larvae.
Keywords Polychaeta; Spionidae; Polydorarickettsi; morphology; ecology; reproduction; larvaldevelopment; Chile
INTRODUCTION
The spionid polychaete Polydora rickettsi wasoriginally described by Woodwick (1961) from CapeSan Lucas, Lower California, Mexico. The specieswas found intertidally associated with calcareoustubes of the serpulid polychaete Spirobranchusincrassatus Mörch. Presumably, it burrowed in thesetubes. No other records or any information about thereproductive biology of P. rickettsi has beenprovided from Mexico. Later P. rickettsi wasreported by Blake (1983) from Montemar, and bySato-Okoshi & Takatsuka (2001) (as P. cf. rickettsi)from Puerto Montt and Chiloé Island, all reportsfrom Chile (Fig. 1). Sato-Okoshi & Takatsuka(2001) reported briefly on reproduction and larvaldevelopment of the species and suggested thatfurther studies on specimens from Chile wererequired to determine if they are conspecific with P.rickettsi from Mexico.
Reproductive biology and larval development ofpolychaetes on the Pacific side of South America andin the Strait of Magellan have been studied only inone species of Sabellidae (Gambi & Patti 1999) andsome species of Spionidae. Among spionids, a briefnote about mature pelagic stages of two species fromPeru was provided by Berkeley & Berkeley (1963);pelagic larvae of 14 species from Concepción Bay,
M03061; Online publication date 8 June 2004Received 25 September 2003; accepted 18 January 2004
New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38: 243–2540028–8330/04/3802–0243 © The Royal Society of New Zealand 2004

244 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
Chile were described by Carrasco (1976); and someinformation about four species from southern Chile,including P. rickettsi, was provided by Sato-Okoshi& Takatsuka (2001).
In the present study, P. rickettsi was found as acommon borer of various calcareous substrata insouthern Chile. The purpose of this paper is todescribe development of morphology duringontogenesis and report on biology of the species.
MATERIAL AND METHODS
Field collections were made off Chiloé Island andin Corral Bay, Tenth Region of Lakes, southernChile from May 2002 through May 2003. Variousmolluscs, barnacles, coralline algae, and spongeswere collected intertidally and shallow subtidallyusing SCUBA equipment. Shells were broken intosmall fragments with hammer and pliers andpolychaetes were removed under a stereomicroscopein the laboratory. Plankton tows were made monthlywith a 150-µm-mesh plankton net in Corral Bay.Adult worms and the larvae were relaxed in isotonicmagnesium chloride and examined alive with lightmicroscopes. Sperm, oocytes, and larvae weremeasured using an ocular micrometer to the nearest0.5, 2, and 10 µm respectively. Drawings were madeusing a camera lucida. Stages of larval developmentdescribed herein do not represent a growth seriesfrom a single specimen, but selected individuals
Fig. 1 Map showing reports of Polydora rickettsi in theeastern Pacific.
from natural conditions; hundreds of larvae from eggcapsules and more than 60 larvae on various stagesof development from the plankton were examinedwith a compound microscope. After examination, apart of adult worms was fixed in 10% formalinsolution, rinsed in fresh water, and transferred to70% ethanol. Additional material from Tongoy Bay,Fourth Region of Coquimbo, Chile, was provided byJuan I. Cañete and Carolina Olivares. Fixed materialwas deposited at the Museo Nacional de HistoriaNatural, Santiago de Chile (MNHN); NationalMuseum of Natural History, Smithsonian Institution,Washington, DC, United States (USNM); andSenckenberg Museum, Frankfurt am Main,Germany (SMF). The number of specimens in asample is given in parentheses after the museumabbreviation and catalogue number.
For scanning electron microscopy (SEM), thespecimens were critical point dried in carbondioxide, coated with gold palladium, and viewedwith a LEO 440 SEM at the National Museum ofNatural History, Smithsonian Institution, Wash-ington, DC. The digital SEM images were combinedinto plates.
RESULTS
Polydora Bosc, 1802Polydora rickettsi Woodwick, 1961
Fig. 2–6
Polydora rickettsi Woodwick, 1961: 78–81, fig. 1–7.Blake 1983: 257.?Polydora sp. II: Carrasco 1976: 34–37, fig. 12, 13L–O,21C.Polydora sp.: Basilio et al. 1995: 72–74, fig. 1A–H.Polydora cf. rickettsi: Sato-Okoshi & Takatsuka 2001:489–490, fig. 2A–D.
MaterialCHILE, Fourth Region of Coquimbo, Tongoy Bay,Coquimbo, 30º14¢S, 71º30¢W, 10 m, from shells ofscallop Argopecten purpuratus cultivated insuspended cages in the sea, coll. C. D. Basilio, 16Jun 1988, SMF 13491 (11); same locality, coll. C.Olivares, 20 Nov 2002, SMF 12307 (8). TenthRegion of Lakes, Yaldad Bay, Chiloé Island,43°06.8¢S, 73°42.8¢W, from shells of gastropodCrepipatella fecunda, intertidal, coll. C. A.Cárdenas, 7 Nov 2002, SMF 12310 (7); 30 Mar2003, USNM 1012108 (17), MNHN AN 2014 (17).Corral Bay, Morro Gonzalo, 39°50.9¢S, 73°27.6¢W,
245Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
12 m, coll. V. I. Radashevsky, 23 Nov 2002, fromshells of Chilean abalone, Concholepasconcholepas, SMF 12306 (44); from shells ofbarnacle Austromegabalanus psittacus, SMF 12308(23); from a demosponge, SMF 12309 (3). Samelocality, coll. C. A. Cárdenas, 11 Mar 2003, fromshells of C. concholepas, 10 m, USNM 1012107(12), MNHN AN 2013 (12); from shell of keyholelimpet Fissurella maxima, 12 m, SMF 13490 (5).
Adult morphologyLargest specimen 29 mm long and 0.6 mm wide onsegment 7 for 144 segments. Body light tan in life;fixed specimens usually with secretory cells visibleas small dark spots scattered on dorsal side ofposterior segments. Fine continuous black lineusually present on palps, along edges of ciliated foodgroove. Black pigment diffused on sides of
prostomium, on 2–4 anterior segments both dorsallyand ventrally, on up to 15 posteriormost segmentsand occasionally on pygidium. Pigmentation usuallyabsent in small individuals and better developed inlarge ones; occasionally, black pigment present onlyon prostomium or totally absent even in largeindividuals. No definite difference in pigmentationobserved between individuals from different hostsbut worms occurring intertidally usually morepigmented.
Prostomium usually rounded anteriorly, occasion-ally with shallow incision. Caruncle extendingposteriorly until end of segment 4, shorter in smallworms. Nuchal antenna absent (Fig. 3A,C). Two tofour black eyes usually present or eyes absent. Palpsextending posteriorly for 25–30 segments; withlongitudinal food groove lined with fine frontal cilia;latero-frontal motile compound cilia (cirri) bordering
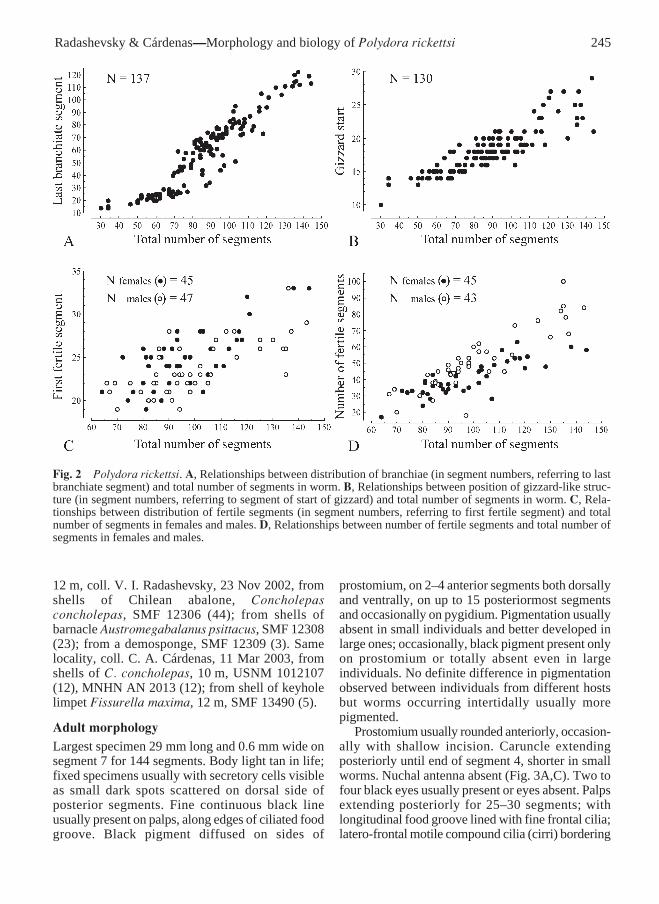
Fig. 2 Polydora rickettsi. A, Relationships between distribution of branchiae (in segment numbers, referring to lastbranchiate segment) and total number of segments in worm. B, Relationships between position of gizzard-like struc-ture (in segment numbers, referring to segment of start of gizzard) and total number of segments in worm. C, Rela-tionships between distribution of fertile segments (in segment numbers, referring to first fertile segment) and totalnumber of segments in females and males. D, Relationships between number of fertile segments and total number ofsegments in females and males.

246 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
food groove; short papillae with non-motile cirriarranged in two or three rows along either side offood groove; short compound non-motile ciliaarising directly from palp surface and scattered onlateral and abfrontal palp surfaces.
Segment 1 with small notopodial lamellae;notochaetae absent; well developed postchaetallamellae and short capillaries present in neuropodia.Prechaetal lamellae absent in all parapodia. Posteriornotopodia with only capillaries (Fig. 3F).
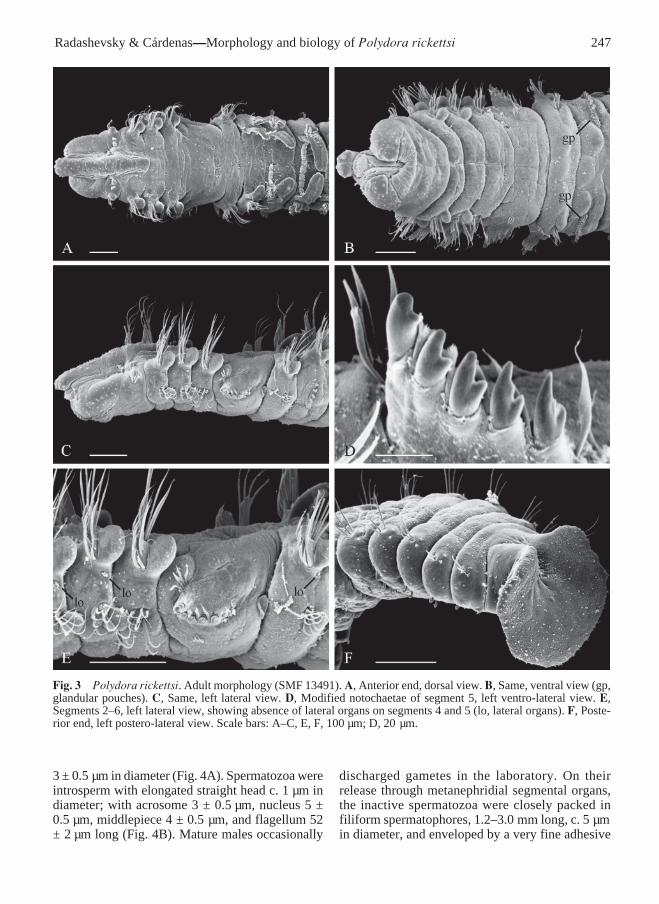
Segment 5 greatly modified, larger than segments4 or 6, without postchaetal lamellae, with 2–4 dorsalsuperior geniculate capillaries (Fig. 6L), up to 7major modified spines alternating with pennonedcompanion chaetae (Fig. 6M,N), and 4–7 ventralcapillaries (Fig. 6K). Dorsal superior and ventralcapillaries shorter and fewer than those on adjacentsegments. Major spines falcate, with lateral tooth andthin subdistal longitudinal flange located laterally onmain fang above lateral tooth (Fig. 3D); spinesarranged in a slightly curved diagonal or horizontalrow (Fig. 3E).
Hooded hooks in neuropodia from segment 7, upto 10 in a series, not accompanied by capillaries.Hooks bidentate, shaft slightly curved, with weakconstriction on shaft (Fig. 6I,J).
Branchiae from segment 7, full-sized fromsegment 8 or 9, on segment 7 half the size of thoseon segment 8, gradually diminishing in size alongposterior half of body and absent on 10–30posteriormost segments (Fig. 2A). Branchiaeflattened, with surfaces oriented parallel to bodyaxis, free from notopodial postchaetal lamellae.Nototrochs from segment 7 or 8, consisting of onerow of cilia (Fig. 3A).
Pygidium cup-shaped or disc-like with dorsal gapor incision (Fig. 3F).
Lateral organs as small ciliated pits between noto-and neuropodia on all segments but 4 and 5 (Fig. 3E).
Glandular pouches from segment 7, full-sizedfrom the beginning, diminishing in size aftersegment 10. Large, flask-shaped secretory cells ofthe pouches opening to exterior individually andappearing externally as small papillae located belowvertical rows of neurochaetae (Fig. 3B).
Gizzard-like structure present between esophagusand midgut, consisting only of weak muscular part,extending through one or two segments; secretorypart absent. Gizzard beginning in segment 10–29,occurring more posteriorly in bigger individuals(Fig. 2B).
Metanephridial segmental organs from segment7, opening to exterior laterally on anterior, sterile
segments and dorsally on fertile segments; con-segmental nephridia opening separately throughout.
HabitatPolydora rickettsi is primarily a borer of variouscalcareous substrates. It was found intertidally andshallow subtidally in shells of the barnacleAustromegabalanus psittacus (Molina, 1782),keyhole limpet Fissurella maxima (Sowerby, 1834),slipper snails Crepipatella fecunda (Gallardo, 1979)and Crepipatella dilatata (Lamarck, 1822), loco(Chilean abalone) Concholepas concholepas(Bruguière, 1789), and scallop Argopectenpurpuratus (Lamarck, 1819). Sato-Okoshi &Takatsuka (2001) reported the species also boringinto shells of the turban snail Tegula atra (Lesson,1830), keyhole limpet Fissurella nigra Lesson,1831, and commercial oysters Crassostrea gigas(Thunberg, 1793) and Ostrea chilensis (Philippi,1845). The worms resided in U-shaped burrowswithin the shells. The walls of the burrow were linedwith detritus, forming a detrital tube within theburrow. The ends of burrows were extended bysmooth silty tubes, 1–2 mm long. Tens of wormswere found in one shell, and 1–2 worms/cm werefound on the shell surface.
One group of worms was found boring into ademosponge (SMF 12309). These worms had thesame morphology as shell-boring individuals. Thesponge contained no shell fragments that mightindicate the worms initially bored into calcareoussubstratum which later became encrusted by asponge. The ability of the species to bore intosponges should be studied further.
ReproductionPolydora rickettsi is gonochoristic; of 111 examinedmature individuals, 56 were females and 55 weremales. Gametes develop along segmental bloodvessels in middle segments, from segments 19–36to 35–92 in females and from segments 19–33 to 38–122 in males. Larger worms had slightly posteriorstart of fertile segments (Fig. 2C) and a greater totalnumber of fertile segments (Fig. 2D). The middlepart of metanephridial segmental organs in fertilesegments in males was expanded, formed by largeurn-shaped cells.
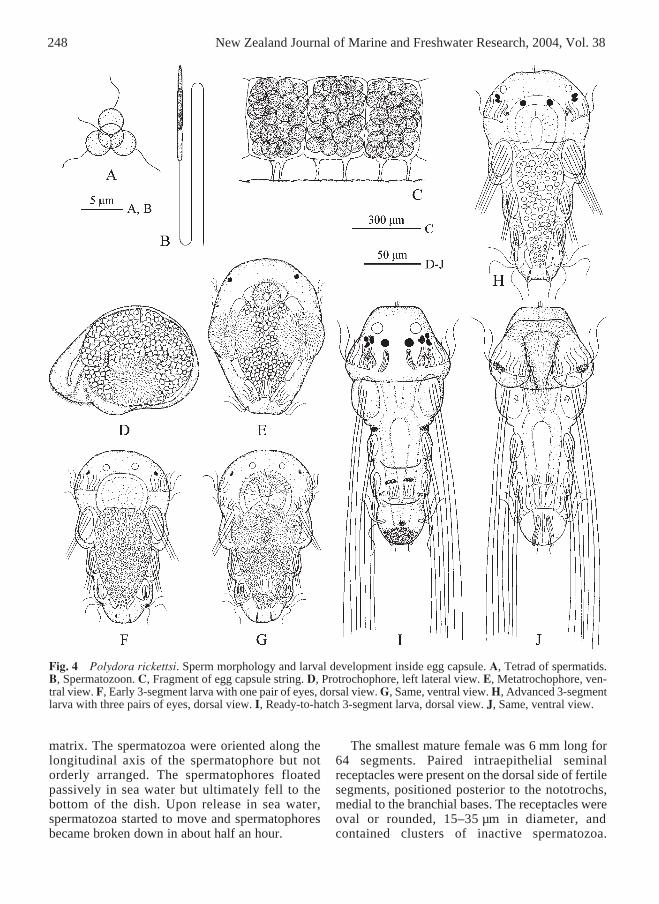
The smallest mature male was 5.5 mm long for66 segments. Temporary testis contained onlyspermatogonia. Solitary primary spermatocytes,diads of secondary spermatocytes, tetrads ofspermatids, and spermatozoa floated freely togetherin the coelomic cavity. Spermatids were spherical,
247Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
3 ± 0.5 µm in diameter (Fig. 4A). Spermatozoa wereintrosperm with elongated straight head c. 1 µm indiameter; with acrosome 3 ± 0.5 µm, nucleus 5 ±0.5 µm, middlepiece 4 ± 0.5 µm, and flagellum 52± 2 µm long (Fig. 4B). Mature males occasionally
discharged gametes in the laboratory. On theirrelease through metanephridial segmental organs,the inactive spermatozoa were closely packed infiliform spermatophores, 1.2–3.0 mm long, c. 5 µmin diameter, and enveloped by a very fine adhesive
Fig. 3 Polydora rickettsi. Adult morphology (SMF 13491). A, Anterior end, dorsal view. B, Same, ventral view (gp,glandular pouches). C, Same, left lateral view. D, Modified notochaetae of segment 5, left ventro-lateral view. E,Segments 2–6, left lateral view, showing absence of lateral organs on segments 4 and 5 (lo, lateral organs). F, Poste-rior end, left postero-lateral view. Scale bars: A–C, E, F, 100 µm; D, 20 µm.
248 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
matrix. The spermatozoa were oriented along thelongitudinal axis of the spermatophore but notorderly arranged. The spermatophores floatedpassively in sea water but ultimately fell to thebottom of the dish. Upon release in sea water,spermatozoa started to move and spermatophoresbecame broken down in about half an hour.
The smallest mature female was 6 mm long for64 segments. Paired intraepithelial seminalreceptacles were present on the dorsal side of fertilesegments, positioned posterior to the nototrochs,medial to the branchial bases. The receptacles wereoval or rounded, 15–35 µm in diameter, andcontained clusters of inactive spermatozoa.
Fig. 4 Polydora rickettsi. Sperm morphology and larval development inside egg capsule. A, Tetrad of spermatids.B, Spermatozoon. C, Fragment of egg capsule string. D, Protrochophore, left lateral view. E, Metatrochophore, ven-tral view. F, Early 3-segment larva with one pair of eyes, dorsal view. G, Same, ventral view. H, Advanced 3-segmentlarva with three pairs of eyes, dorsal view. I, Ready-to-hatch 3-segment larva, dorsal view. J, Same, ventral view.

249Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
Temporary ovaries contained previtellogenic andvitellogenic oocytes of various diameter. Largeoocytes occurred freely in the coelomic cavity.Females deposited eggs in capsules which werejoined to each other in a string. Each egg capsule wasattached by two thin stalks to the inside wall of theburrow and contained up to 65 (mean = 51.8 ± 11.1,n = 21) eggs (Fig. 4C). The total number of capsulesproduced by one female ranged from 22 to 65; thetotal number of eggs deposited per brood rangedfrom 1100 to 4150 (n = 5). The eggs were sphericaland had mean diameters of 95 ± 5 µm (n = 26).Females brooding larvae in capsules had vitellogenicoocytes up to 50 µm in diameter developing in theovaries. Capsules were found in burrows fromAugust to April.
Larval development inside egg capsuleThe majority of eggs in a brood developedsynchronously into larvae. Up to 10% of the eggs didnot undergo cleavage and were later consumed bydeveloping larvae.Protrochophores c. 125 µm long (Fig. 4D).Ventro-lateral sides of larva each with a roundedpatch of numerous short cilia, active beating ofwhich moved larva inside egg capsule. Short foregutlined with fine cilia, its posterior end joined tospherical mass of entodermal macromeres occupyingmost of inner space of larva. Lumen of midgut,posterior gut, and anus not yet developed.Metatrochophores c. 135 µm long (Fig. 4E).Prostomium with one pair of lateral eyes. Prototrochincomplete, consisting of two pairs of ciliary bandson sides of the head. One band of cilia positionedantero-dorsally and other band with shorter cilia andpositioned postero-ventrally; each band formed bytwo or three ciliated cells. Telotroch consisting offive ciliated cells on ventral and lateral sides ofpygidium. Excretory organs not observed.
As growth proceeded, the mesodermal anlagessplit up and formed internal septa that separated thehead coelom, three segmental coeloms and oneprepygidial coelom (apparently all paired). Wheninternal compartmentalisation had been completed,larval bristles started to grow in chaetal sacs, first insegment 1 and then successively in segments 2 and 3.
Early 3-segment larvae c. 140 µm long (Fig.4F,G). One or two pairs of lateral eyes. Anterior partof prostomium with one pair of phaosomesappearing as transparent spheres c. 5 µm in diameter.Nuchal organs not yet developed. Ventral ciliarypatches present on segment 1. Chaetae include short
larval bristles in notopodia, those in segment 1 thelongest. Midgut still formed by yolky macromeresand lumen not yet developed. Posterior gut notformed and anus not opened.
In further development, a median pair of eyesappeared on the head. One pair of small bossescovered by fine non-motile cilia appeared posteriorto median eyes, in front of cilia of the prototroch.Prototrochal ciliary bands on either side of the headshifted their position and became arranged in oneline (Fig. 4H). Ventral ciliary patches were reducedbut neurotroch and companion ciliated cellsappeared on segment 1.
Ready-to-hatch 3-segment larvae 200–210 µmlong (Fig. 4I,J). Three pairs of black eyes present,comprising two pairs of lateral eyes and one pair ofmedian eyes; lateral eyes on either side close to oneanother and therefore appearing as one pair of eyes.One pair of black pigment spots medial to lateraleyes. Phaosomes in anterior part of prostomium c.10 µm in diameter. Nuchal organs as roundedciliated patches on dorso-lateral sides ofprostomium, in front of prototroch. Ciliated patch ofeach nuchal organ with small boss with fine non-motile cilia. Indistinct patches of yellow pigment onlateral lips of peristomium, at level of prototroch.Lateral melanophores on segment 2. Dorsal pairedmelanophores on segment 3. Pygidium with blackpigment mid-dorsally.
Chaetae as long larval bristles in notopodia; thoseof segment 1 longest. Bristles slightly curved, withfine serrations on convex side.
Nototroch, one pair of dorsal grasping cilia andgastrotroch on segment 3. Short triangular neuro-troch on segment 1; one pair of small companionciliated cells on sides of neurotroch.
One pair of intraepithelial banana-shaped glandularcells with striated content posterior to median eyes.Similar glandular cells inside pygidium; one pair ofthose cells protruded externally, appearing as shortpapillae on either side of anus.
Lateral lips of peristomium forming voluminousvestibulum covered by numerous, actively beatingshort cilia. Longer compound, probably sensory ciliaarranged along inner sides of lips and betweenciliated cells of prototroch. Vestibulum leading intoshort esophagus separated from midgut by muscularsphincter. Intestine with wide lumen, without yolkygranules in wall. One pair of protonephridia insegment 1.
When maternal egg yolk had been depleted, the3-segment larvae started to move rapidly and
250 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
probably cut the thin wall of the capsules with theirserrated bristles. Hatched larvae were released fromthe mother’s burrow and entered the plankton.
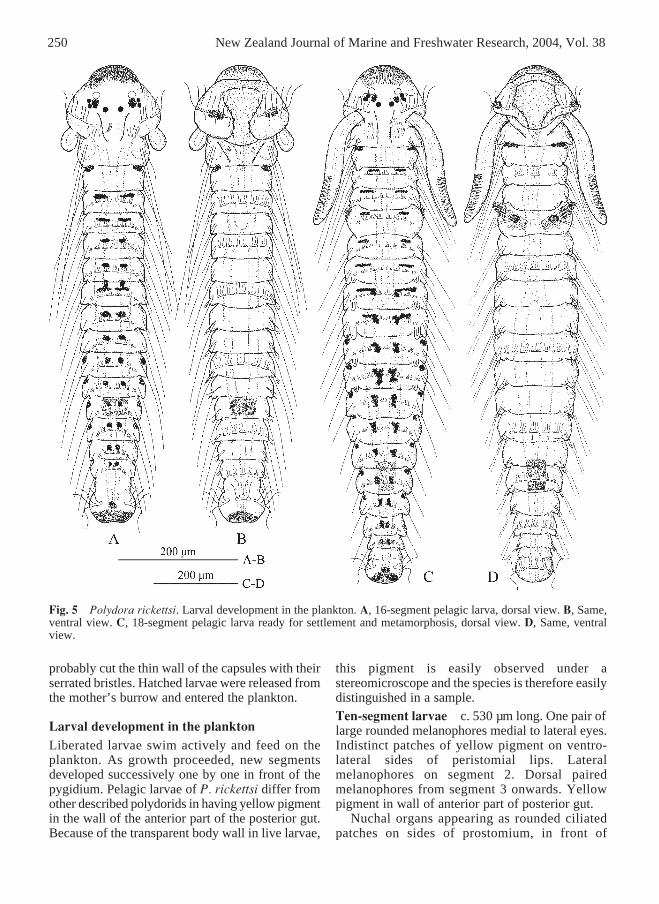
Larval development in the planktonLiberated larvae swim actively and feed on theplankton. As growth proceeded, new segmentsdeveloped successively one by one in front of thepygidium. Pelagic larvae of P. rickettsi differ fromother described polydorids in having yellow pigmentin the wall of the anterior part of the posterior gut.Because of the transparent body wall in live larvae,
this pigment is easily observed under astereomicroscope and the species is therefore easilydistinguished in a sample.Ten-segment larvae c. 530 µm long. One pair oflarge rounded melanophores medial to lateral eyes.Indistinct patches of yellow pigment on ventro-lateral sides of peristomial lips. Lateralmelanophores on segment 2. Dorsal pairedmelanophores from segment 3 onwards. Yellowpigment in wall of anterior part of posterior gut.
Nuchal organs appearing as rounded ciliatedpatches on sides of prostomium, in front of
Fig. 5 Polydora rickettsi. Larval development in the plankton. A, 16-segment pelagic larva, dorsal view. B, Same,ventral view. C, 18-segment pelagic larva ready for settlement and metamorphosis, dorsal view. D, Same, ventralview.
251Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
prototroch. Short palps on postero-lateral edges ofperistomium, behind prototroch. Chaetae still onlylarval bristles in notopodia. Neuropodia not yetdeveloped. Nototrochs and grasping cilia fromsegment 3 onwards. Gastrotrochs on segments 3, 5,7, and 10. One pair of protonephridia in segment 1.Circulatory system well developed and functional.
In further development, larval protonephridiaappeared in segment 2, adult capillaries appeared inall neuropodia and single larval serrated bristlesappeared in neuropodia from segment 3; modifiedspines appeared in notopodia of segment 5 amonglarval bristles; hooded hooks developed inneuropodia from segment 7; lateral melanophoresappeared from segment 10.Sixteen-segment larvae c. 750 µm long (Fig.5A,B). Banana-shaped glandular cells on caruncle
gradually and asynchronously became reduced, thusoften only one of them present. Prostomiumgradually elongated posteriorly as a low caruncle.Nuchal organs shifted backwards, ending posteriorto prototroch; single bosses with non-motile cilia innuchal organs reduced. Adult capillaries inneuropodia from segment 1, single larval bristles inneuropodia from segment 3, and hooded hooks inneuropodia from segment 7 onwards. One pair ofprovisional modified spines among larval bristles innotopodia of segment 5.Ready-to-settle 17–18-segment larvae c. 1100 µmlong (Fig. 5C,D). Diffuse yellow pigment on anterioredge of prostomium. Indistinct yellow spots onlateral lips of peristomium, at level of prototroch.Paired dorsal melanophores from segment 3,transversely elongated on anterior segments and
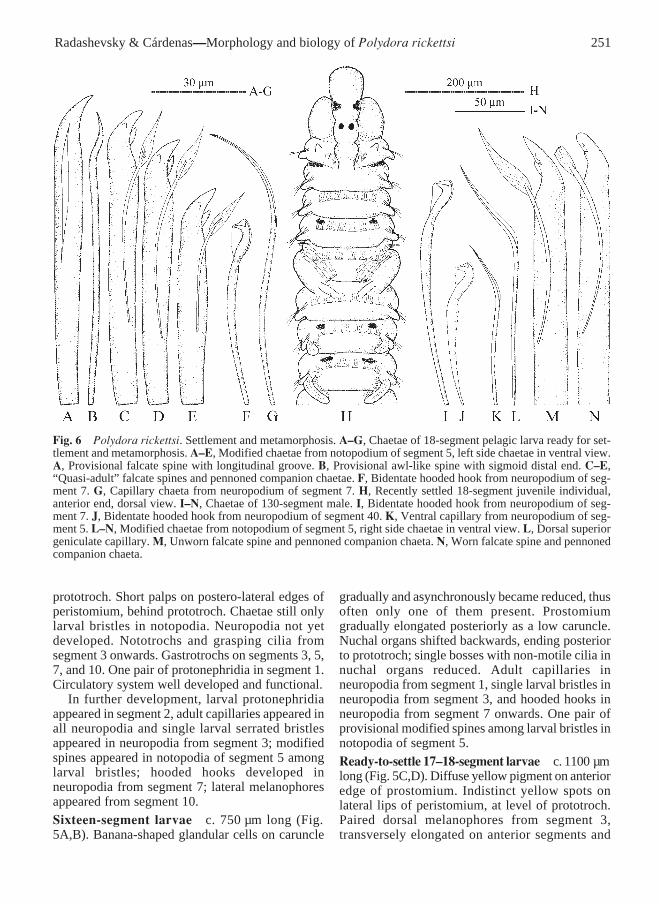
Fig. 6 Polydora rickettsi. Settlement and metamorphosis. A–G, Chaetae of 18-segment pelagic larva ready for set-tlement and metamorphosis. A–E, Modified chaetae from notopodium of segment 5, left side chaetae in ventral view.A, Provisional falcate spine with longitudinal groove. B, Provisional awl-like spine with sigmoid distal end. C–E,“Quasi-adult” falcate spines and pennoned companion chaetae. F, Bidentate hooded hook from neuropodium of seg-ment 7. G, Capillary chaeta from neuropodium of segment 7. H, Recently settled 18-segment juvenile individual,anterior end, dorsal view. I–N, Chaetae of 130-segment male. I, Bidentate hooded hook from neuropodium of seg-ment 7. J, Bidentate hooded hook from neuropodium of segment 40. K, Ventral capillary from neuropodium of seg-ment 5. L–N, Modified chaetae from notopodium of segment 5, right side chaetae in ventral view. L, Dorsal superiorgeniculate capillary. M, Unworn falcate spine and pennoned companion chaeta. N, Worn falcate spine and pennonedcompanion chaeta.

252 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
ramified from segment 7 or 8 onwards. Lateralmelanophores on segment 2 and from segment 10onwards; those on segment 2 largest, extendinglaterally on segment 1 and on ventral side betweensegments 1 and 2; those from segment 10 on frontaledges of segments. Diffuse black pigment on dorsalside of pygidium. Light yellow pigment diffused inwall of anterior part of posterior gut.
Prostomium rounded anteriorly, with three pairsof black eyes and one pair of phaosomes c. 15 µmin diameter. Caruncle low, extending posteriorlyalmost until end of segment 1. Nuchal organs as ovalor rounded ciliated patches on either side of caruncleon segment 1, without ciliated bosses. Banana-shaped cells posterior to median eyes occasionallypresent. Palps extending posteriorly to middle ofsegment 5, yet without longitudinal ciliated foodgroove. Spindle-shaped glandular cells regularlyarranged inside distal 2⁄3 of each palp.
Larval serrated bristles in all notopodia exceptthose of segment 5. Adult capillaries in all notopodiaexcept those of segment 1. Bidentate hooded hooksin neuropodia from segment 7, up to 5 in a series(Fig. 6F). Hooks accompanied by equal number ofwinged capillaries (Fig. 6G), apparently positionedin front of hooks.
Segment 5 modified, larger than segments 4 or 6,with 2 or 3 short dorsal superior capillaries, 4 or 5heavy modified spines, 2 or 3 pennoned companionchaetae, and a tuft of 4 or 5 ventral capillaries.Modified spines including one pair of provisionalbristles and 2 or 3 falcate spines of “quasi-adult”kind. Provisional bristles including a heavy falcatespine with longitudinal groove (Fig. 6A), and athinner awl-like spine with sigmoid distal end (Fig.6B). Quasi-adult modified spines with big lateraltooth on one side and small flange or tooth on otherside (Fig. 6C–E), latter structure appearing smallerin younger posterior spines; these spines alternatingwith pennoned companion chaetae.
Short branchiae on segments 7–11, those onsegment 7 almost half the length of those on segment8.
Nototrochs and grasping cilia from segment 3onwards. Short triangular neurotroch and one pairof small companion ciliated cells on sides ofneurotroch on segment 1. Ventral ciliated pit absent.Gastrotrochs on segments 3, 5, 7, 10, 13, 15, and 17.
Pygidium rounded, with dorsal incision.One pair of small glands with grey granular
content in segment 5, below set of heavy modifiedspines. Glandular pouches from segment 6 onwards,those in segment 6 consisting of one glandular cell,
in following segments of about five cells; glandslargest in segments 7–9, thereafter diminishing insize.
Larval provisional protonephridia in segments 1and 2. Adult metanephridia from segment 7.
Settlement and metamorphosisThe 17–18-segment pelagic larvae appeared readyfor settlement and metamorphosis. Smallest juvenilefound on a shell was c. 1500 µm long for 18segments (Fig. 6H). Pelagic larvae were able topostpone settlement in the absence of appropriatesubstrata; largest larva caught in the plankton wasc. 1250 µm long for 19 segments.
Settlement was accompanied by gradualmetamorphosis and loss of some larval features, e.g.,bristles, proto-, telo-, neuro-, and gastrotrochs.Nuchal organs transformed into narrow ciliatedbands on either side of caruncle. Protonephridia insegments 1 and 2 reduced and became no longerrecognisable. Nototrochs were still present fromsegment 3 (in adults they were present on branchialsegments, i.e., from segment 7 or 8 onwards) butgrasping cilia were lost; capillary chaetae in hook-bearing neuropodia were also lost. Segment 5became enlarged and lost provisional modifiedspines from notopodia and grey glands with granularcontent. Development of new segments was delayedwhile larval segments enlarged. Switch to a newmode of feeding after settlement was enabled byrapid elongation of the palps, development of theciliated food groove along their length, modificationof the prostomium, and enlargement of the ventralperistomial lip.
Newly settled individuals built small mud tubeson the shell surface and then bored into the shell.These initial mud tubes were later enlarged andappeared as a continuation of the burrow.
DISCUSSION
Sato-Okoshi & Takatsuka (2001) noticed somedifferences between Polydora specimens theycollected from Chile and the original description ofP. rickettsi from Lower California. They reportedthat Chilean worms had an incised prostomiuminstead of an entire one, and greater variability in thepigmentation. However, Woodwick’s (1961) des-cription was based on one complete individual andtwo fragments, and morphological variability ofMexican worms has not been reported since. Ourexamination of Polydora in Chile also revealed

253Radashevsky & Cárdenas—Morphology and biology of Polydora rickettsi
significant variability, including gradation of theanterior edge of the prostomium from entire toincised, and especially variations in the pigmen-tation, which exceeds variability of type specimensof P. rickettsi. However, characteristics provided byWoodwick (1961) fall within the limits of variabilityof Chilean specimens. Besides, specific details ofmorphology of modified spines on segment 5, suchas presence of a lateral tooth and fine subdistalflange, fit the diagnosis of P. rickettsi. Examinationof variability and redescription of P. rickettsi fromthe type locality will help to correct or confirm ouridentification of the Chilean worms and to betterunderstand the distribution of the species. Until thenwe consider Chilean and Mexican worms to beconspecific, as did Blake (1983).
Basilio et al. (1995) described Polydora sp., aborer of shells of the scallop Argopecten purpuratuscultivated in Tongoy Bay, Chile. They reported thatthe caruncle of these specimens extended posteriorlyto the end of segment 2. The reported material wasexamined (SMF 13491) and found to be identical toP. rickettsi specimens collected in the present study;although some individuals had short caruncle (Fig.3A), in others the caruncle extended posteriorly untilthe middle of segment 4.
Adult individuals of P. rickettsi have a gizzard-like structure positioned between the esophagus andthe midgut. Gizzards are characteristic forDipolydora and among Polydora species thestructure has only been reported for P. spongicola(Radashevsky 1988, as Polydora uschakovi). InDipolydora species, the gizzard consists of anteriormuscular and posterior secretory parts (seeRadashevsky 1993), whereas in the two Polydoraspecies it consists only of a weakly developedmuscular part. Whether the gizzards of Polydoraspecies are homologous to those in Dipolydoraremains unknown.
The 19-segment pelagic larva of P. rickettsi fromConcepción Bay, Chile was illustrated by Carrasco(1976: fig. 12, 13L–O, 21C) as Polydora sp. II.Carrasco (1976) found the larvae from December toMay and also in August, and noticed that their highestdensity in the plankton was from January to May.
Polydorid taxa are distinguished from otherspionids by the presence of modified spines innotopodia of segment 5, which usually begin todevelop in 10–15-segment larvae. The morphologyof these spines is often species specific and used toidentify larvae and adult worms. In some taxa,however, one or two first-developed spines differdrastically from those appearing later. These spines
are usually lost immediately or soon after settlementand therefore are referred to as provisional bristles.Three kinds of provisional bristles in polydoridswere described by Radashevsky & Fauchald (2000).The spines developing after these provisional bristlesusually have adult, definitive morphology. Develop-ment of spines in P. rickettsi represents a morecomplicated pattern. The first two spines areprovisional and include a falcate spine with alongitudinal lateral groove (Fig. 6A) and a thinnerpointed spine with the distal part greatly narrowedand sigmoidaly curved (Fig. 6B), as in the majorityof Polydora species (see Radashevsky & Fauchald2000: fig. 5A,B). The spines developing afterprovisional bristles resemble adult, definitive falcatespines with a large lateral tooth but differ from themin the presence of a second, smaller lateral tooth onthe side opposite to the big tooth, and in the absenceof a subdistal longitudinal flange. In P. rickettsilarvae that are ready to settle, two or threetransitional quasi-adult falcate spines are arrangedin a diagonal row (Fig. 6C–E). After settlement, twoprovisional bristles and anterior quasi-adult spinesare successively lost and newly developing spinesgradually acquire definitive appearance, with one biglateral tooth and a thin subdistal longitudinal flange(Fig. 6M,N). Gradual acquisition of definitivemorphology by successively developing falcatespines in segment 5, such as in P. rickettsi, has neverbefore been reported for polydorids. This infor-mation might be useful for phylogenetic con-siderations and should be taken into account whenidentifying polydorid larvae and early juveniles.
ACKNOWLEDGMENTS
Dr Carlos Bertrán Vives invited one of the authors (V.Radashevsky) to visit Universidad Austral de Chile inValdivia in November 2002 and provided general sup-port during this visit. Dr Juan I. Cañete and Ms CarolinaOlivares provided material from Tongoy Bay, Chile, andMs Ana Maria Ojeda provided essential help in all thisstudy. Dr Jason D. Williams provided editorial assistanceon a draft of this manuscript. Dr Mary E. Petersen andDr Geoff B. Read essentially edited and commented onthe manuscript after submission. To all these persons weexpress our sincere gratitude. Financial support was pro-vided to V. Radashevsky by the project MECESUP AUS0101 through the Instituto de Zoología, UniversidadAustral de Chile, Valdivia (2002); the National Museumof Natural History, Smithsonian Institution (VisitingScientist Fellowships, 1999); and project 03-3-A-06-125of the Far Eastern Branch of the Russian Academy ofSciences (2003).

254 New Zealand Journal of Marine and Freshwater Research, 2004, Vol. 38
REFERENCES
Basilio, C. D.; Cañete, J. I.; Rozbaczylo, N. 1995: Polydorasp. (Spionidae), un poliqueto perforador de lasvalvas del ostión Argopecten purpuratus(Bivalvia: Pectinidae) en Bahía Tongoy, Chile.Revista de Biología Marina, Valparaíso (Chile)30: 71–77.
Berkeley, E.; Berkeley, C. 1963: Neoteny in larvae oftwo species of Spionidae. Canadian Journal ofZoology 41: 149–151.
Blake, J. A. 1983: Polychaetes of the family Spionidaefrom South America, Antarctica, and adjacentseas and islands. Biology of the Antarctic SeasXIV. Antarctic Research Series 39: 205–288.
Carrasco, F. D. 1976: Larvas de la familia Spionidae(Polychaeta) en el plancton de la Bahía deConcepción, Chile. Gayana Zoológica (Chile) 38:1–63.
Gambi, M. C.; Patti, F. P. 1999: Reproductive biology ofPerkinsiana antarctica (Kinberg) (Polychaeta,Sabellidae) in the Straits of Magellan (SouthAmerica): Systematic and ecological implications.Scientia Marina 63: 253–259.
Radashevsky, V. I. 1988: Morphology, ecology, repro-duction, and larval development of Polydorauschakovi (Polychaeta, Spionidae) in the Peterthe Great Bay of the Sea of Japan. ZoologicheskiiZhurnal 67: 870–878. (In Russian with Englishsummary.)
Radashevsky, V. I. 1993: Revision of the genus Polydoraand related genera from the North West Pacific(Polychaeta: Spionidae). Publications of the SetoMarine Biological Laboratory 36: 1–60.
Radashevsky, V. I.; Fauchald, K. 2000: Chaetal arrange-ment and homology in spionids (Polychaeta:Spionidae). Bulletin of Marine Science 67: 13–23.
Sato-Okoshi, W.; Takatsuka, M. 2001: Polydora andrelated genera (Polychaeta, Spionidae) aroundPuerto Montt and Chiloé Island (Chile), with de-scription of a new species of Dipolydora. Bulletinof Marine Science 68: 485–503.
Woodwick, K. H. 1961: Polydora rickettsi, a new speciesof spionid polychaete from Lower California.Pacific Science 15: 78–81.
Related Documents











