MORPHOLOGICAL CHARACTERISATION OF SELECTED SCYPHOZOAN JELLYFISH SPECIES AND GEOMETRIC MORPHOMETRIC ANALYSIS OF Chrysaora chinensis IN PENINSULAR MALAYSIA LOW LIANG BOON DISSERTATION SUBMITTED IN FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF PHILOSOPHY INSTITUTE OF GRADUATE STUDIES UNIVERSITY OF MALAYA KUALA LUMPUR 2017

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MORPHOLOGICAL CHARACTERISATION OF SELECTED
SCYPHOZOAN JELLYFISH SPECIES AND
GEOMETRIC MORPHOMETRIC ANALYSIS OF
Chrysaora chinensis IN PENINSULAR MALAYSIA
LOW LIANG BOON
DISSERTATION SUBMITTED IN FULFILLMENT OF THE
REQUIREMENTS FOR THE DEGREE OF
MASTER OF PHILOSOPHY
INSTITUTE OF GRADUATE STUDIES
UNIVERSITY OF MALAYA
KUALA LUMPUR
2017

ii
UNIVERSITI MALAYA
ORIGINAL LITERARY WORK DECLARATION
Name of Candidate: Low Liang Boon
I.C/Passport No: 761110-01-5729
Registration/Matric No: HGT 140008
Name of Degree: Master of Philosophy
Title of Project Paper/Research Report/Dissertation/Thesis (“this Work”):
MORPHOLOGICAL CHARACTERISATION OF SELECTED SCYPHOZOAN
JELLYFISH SPECIES AND GEOMETRIC MORPHOMETRIC ANALYSIS OF
Chrysaora chinensis IN PENINSULAR MALAYSIA
Field of Study: Environmental Science (Marine Sciences)
I do solemnly and sincerely declare that:
(1) I am the sole author/writer of this Work;
(2) This Work is original;
(3) Any use of any work in which copyright exists was done by way of fair dealing and
for permitted purposes and any excerpt or extract from, or reference to or
reproduction of any copyright work has been disclosed expressly and sufficiently
and the title of the Work and its authorship have been acknowledged in this Work;
(4) I do not have any actual knowledge nor do I reasonably know that the making of
this work constitutes an infringement of any copyright work;
(5) I hereby assign all and every rights in the copyright to this Work to the University
of Malaya (“UM”), who henceforth shall be owner of the copyright in this Work
and that any reproduction or use in any form or by any means whatsoever is
prohibited without the written consent of UM having been first had and obtained;
(6) I am fully aware that if in the course of making this Work I have infringed any
copyright whether intentionally or otherwise, I may be subject to legal action or any
other action as may be determined by UM.
Candidate’s Signature Date
Subscribed and solemnly declared before,
Witness’s Signature Date
Name:
Designation:
Safri
Highlight

iii
ABSTRACT
Although jellyfish blooms have significant impacts to the coastal environments, the
effective management of blooms and understanding of their proliferation are often
confounded by the lack of baseline data, which includes species identification, their
biology and ecology. In Malaysia, jellyfish blooms can negatively impact human
activities such as causing beach closures, damage to fishing nets, threats of stings and
blocking power station systems. Therefore, detail characterization and documentation of
their morphology would facilitate species identification of jellyfish in the field,
especially in Malaysian waters and this region which may harbours many undiscovered
species. In this study, the morphology of nine jellyfish species found in Peninsular
Malaysia that belong to the class Scyphozoa, namely Chrysaora chinensis, Cyanea sp.,
Versuriga anadyomene, Rhopilema hispidum, Rhopilema esculentum, Phyllorhiza
punctata, Acromitus flagellatus, Lobonemoides robustus and Lychnorhiza malayensis
were characterised in detail. Sea nettle jellyfish in Malaysia was verified as C.
chinensis. The status of Cyanea sp. found in Malaysia is uncertain as it may possibly be
a new species. This study also reported the first record of Lychnorhiza malayensis. A
total of 107 specimens of C. chinensis were obtained from four coastal areas of
Peninsular Malaysia (East-Central, East-North, West-Central, and West-North) to
compare the possible morphological variation between populations using geometric
morphometric analysis. Procrustes superimposition, Principal Component Analysis
(PCA) and Canonical Variate Analysis (CVA) were applied to the images of
gastrovascular pouches of C.chinensis to extract the shape information. There were no
significant differences in shape among all the specimens based on PCA. However, CVA
showed shape variation between populations of the four areas of Peninsular Malaysia.

iv
ABSTRAK
Walaupun ledakan obor-obor boleh memberi impak yang besar kepada persekitaran
pantai, pengurusan yang berkesan dan pemahanam tentang perkembangan mereka
sering terjejas disebabkan kekurangan data asas, termasuk pengecaman spesies, biologi
dan ekologi mereka. Di Malaysia, ledakan obor-obor boleh memberi kesan negatif
kepada aktiviti manusia seperti menyebabkan penutupan pantai, kerosakan kepada jala
ikan, ancaman sengatan dan sistem stesyen kuasa tersumbat. Oleh itu, pencirian dan
dokumentasi morfologi mereka secara terperinci akan memudahkan pengecaman
spesies obor-obor di lapangan, terutamanya di perairan Malaysia dan rantau ini yang
mungkin mengandungi pelbagai spesis yang belum ditemui. Dalam kajian ini,
morfologi sembilan spesies obor-obor di Semenanjung Malaysia yang tergolong dalam
kelas Scyphozoa iaitu Chrysaora chinensis, Cyanea sp., Versuriga anadyomene,
Rhopilema hispidum, Rhopilema esculentum, Phyllorhiza punctata, Acromitus
flagellatus, Lobonemoides robustus, dan Lychnorhiza malayensis telah dicirikan secara
terperinci. Obor-obor “sea nettle” di Malaysia telah disahkan sebagai C. Chinensis.
Status Cyanea sp. yang terdapat di Malaysia tidak dapat dipastikan dan mungkin
merupakan satu spesies yang baru. Kajian ini juga melaporkan rekod pertama bagi
Lychnorhiza malayensis. Sejumlah 107 spesimen C. chinensis diambil dari empat
kawasan persisiran pantai di Semenanjung Malaysia (East-Central, East-North,
West-Central, dan West-North) untuk membandingkan variasi morfologi spesies
dengan menggunakan analisa “geometric morphometrics”. “Procrustes
superimposition”, “principal component” (PCA) dan “canonical variate” (CVA) telah
diaplikasikan terhadap imej kantung gastrovaskular C. chinensis untuk mendapatkan
informasi bentuknya. Keputusan daripada PCA tidak menunjukkan perbezaan bentuk
yang signifikan disemua specimen. Tetapi, keputusan CVA menunjukan perbezeaan
bentuk diantara populasi dari empat lokasi di Semenanjung Malaysia.

v
ACKNOWLEDGEMENTS
I would like to express appreciation to my supervisor, Dr. Mohammed Rizman Idid and
lab mate Wan Mohd Syazwan for the supports, ideas and guidance. I am grateful to the
staff at Institute of Ocean and Earth Sciences (IOES), University of Malaya for
providing constant assistance during the study. I thank University of Malaya for
providing research grants RU006E-2014 and RG104-11SUS to my supervisor.

vi
TABLE OF CONTENTS
ORIGINAL LITERARY WORK DECLARATION ii
ABSTRACT iii
ABSTRAK iv
ACKNOWLEDGEMENTS v
TABLE OF CONTENTS vi
LIST OF FIGURES viii
LIST OF TABLES xiv
CHAPTER 1 INTRODUCTION 1
1.1 Jellyfish Studies in Malaysia 1
1.2 Taxonomy Position and Problem of the Malaysia Sea Nettles 8
1.3 Introduction to Geometric Morphometric Analysis 10
1.4 Research Aims and Questions 12
1.5 Objectives of the Study 14
1.6 Significance of the Study 15
CHAPTER 2 MATERIALS AND METHODS 16
2.1 Collection of Jellyfish Samples 16
2.2 Photography of Specimens and Collection of Morphological Data 19
2.3 Geometric Morphometric Analysis of Gastrovascular Pouch 22
2.3.1 Landmark Configurations and Photography of
Gastrovascular Pouches 23
2.3.2 Formating of Images into TPS File 26
2.3.3 Digitisation of Images 26
2.3.4 Quantification of Measurement Error 27
2.3.5 Procrustes Superimposition 28
2.3.6 Principle Component Analysis Using MorphoJ 29
2.3.7 Canonical Variate Analysis Using MorphoJ 31
2.3.8 Visualization of Shape Outline 31

vii
CHAPTER 3 RESULTS 34
3.1 Overview of Major Morphological Structures of Jellyfish 34
3.1.1 Order Semaeostomeae 34
3.1.2 Order Rhizostomeae 35
3.2 Morphological Description of Chrysaora chinensis Vanhöffen 1888 52
3.3 Morphological Description of Cyanea Sp. 56
3.4 Morphological Description of Rhopilema esculentum Kishinouye 1891 60
3.5 Morphological Description of Rhopilema hispidum Vanhöffen 1888 64
3.6 Morphological Description of Lobonemoides robustus Stiasny 1920 67
3.7 Morphological Description of Versuriga anadyomene Maas 1903 71
3.8 Morphological Description of Phyllorhiza punctata von Lendenfeld 1884 75
3.9 Morphological Description of Lychnorhiza malayensis Stiasny 1920 79
3.10 Morphological Description of Acromitus flagellates Maas 1903 82
3.11 Geometric Morphometric Analysis 85
3.11.1 Procrustes Anova 86
3.11.2 Principal Component Analysis (PCA) 86
3.11.3 Canonical Variate Analysis (CVA) 88
CHAPTER 4 DISCUSSION 93
4.1 Morphology, Diversity and Importance of Jellyfish Species of
Peninsular Malaysia 93
4.2 Lion’s mane Jellyfish (Cyanea Sp.) of Malaysia 94
4.3 Sea Nettles (C. Chinensis) of Peninsular Malaysia 96
4.4 Potential Application of GMM in Jellyfish Studies 99
CHAPTER 5 CONCLUSION 101
REFERENCES 103
APPENDICES 113

viii
LIST OF FIGURES
Figure 1.1 An adult scyphozoan jellyfish, Phyllorhiza punctata
3
Figure 1.2
Figure 1.3
Figure 1.4
Figure 1.5
Figure 1.6
Classification of Cnidarian
Life cycle of jellyfish
Bloom and stranding of Crambione mastigophora in Pulau
Ketam in April 2016
Cross section of a jellyfish
GMM used on various organisms. A: Oak leaf. B: Dog. C:
Cichlid
3
4
4
5
11
Figure 2.1 Diagram showing pompang (bag net)
18
Figure 2.2 Photography setup to float jellyfish in the tank against a
black background, with colour chart and scale. Wires were
used to suspend the specimens, and a scaled wire was also
used to aid recording of measurements.
21
Figure 2.3 Dry photography setup showing the acrylic stage, with
light source below and additional light sources at the sides.
22
Figure 2.4 An image of a subumbrella quadrant of a C. chinensis
specimen showing the 16 geometric morphometric
landmark points (indicated in red and numbered) of a
single gastrovascular pouch analysed (P1). Pouch P1 is
flanked by pouches P2 and P3. Rhopalia are indicated as R.
25
Figure 2.5 The Number of replicate for each level of the process of
specimen imaging and digitization.
28
Figure 2.6
Figure 3.1
56 landmark configurations used to create the outline of
gastrovascular pouch
Map of eight sampling sites, with number of specimen
collected for each species.
33
38
Figure 3.2
Figure 3.3
Whole medusa of an adult jellyfish, Phyllorhiza punctata
Morphology of bell of four different species of scyphozoan
jellyfish. (A): Acromitus flagellates with smooth surface
(B): Rhopilema esculentum with smooth surface (C):
Phyllorhiza punctata with warts on the surface (D):
Cyanea sp. With smooth surface
39
40

ix
Figure 3.4 Lappet of two different species of scyphozoan jellyfish.
(A): Rhopalar lappet (RL) and velar lappet (VL) of
Lychnorhiza malayensis (B): Lappet of Cyanes sp. (L).
Lappet shape broad and semi-square
40
Figure 3.5 Marginal sensory organ / rhopalium (R) of Lychnorhiza
malayensis
41
Figure 3.6 Morphology of umbrella. (A): Tentacle (T) of Chrysaora
chinensis. (B): Lychnorhiza malayensis of Rhizostomeae
without tentacles
42
Figure 3.7 Morphology of the manubrium of Chrysaora chinensis 42
Figure 3.8 Morphology of oral arm (A): Whole medusa floated in a
tank, showing four soft and curtain-like oral arms (OA).
(B): Oral arm (OA) of Rhopilema esculentum. (C): Oral
arm (OA) of Acromitus flagellatus
43
Figure 3.9 Morphology of scapulae. (A): Balde shape Scapulae of
Rhopilema hispidum. (B): Scapulae (S) attached to oral
arms of Rhopilema esculentum
44
Figure 3.10 Morphology of oral arm with terminal club. (A): Terminal
club (C) at the margin of the oral arm of Rhopilema
esculentum. (B): Terminal club (C) at the margin of the
oral arm of Phyllorhiza punctata
44
Figure 3.11 Morphology of filaments. (A): Filament (F) at the oral disk
of Phyllorhiza punctata. (B): Filament (F) at the oral arms
of Acromitus flagellatus
45
Figure 3.12 Morphology of subumbrella of Chrysaora chinensis
revealing the gastrovascular cavity (G)
46
Figure 3.13 Subumbrella morphology of subgenital fenestration (F) of
Rhopilema esculentum
47
Figure 3.14 Subumbrella morphology of papillae. (A): Three papillae
(P) at the subumbrella of Lychnorhiza malayensis. (B):
Three papillae (P) at the subumbrella of Rhopilema
hispidum
48
Figure 3.15 Network of canals of (A): Lychnorhiza malayensis injected
with red dye. (B): Phyllorhiza punctata injected with blue
dye. (C): Lobonemoides robustus injected with yellow dye
49
Figure 3.16 Subumbrella morphology of muscle. (A): Coronal muscle
(CM) of Phyllorhiza punctata. (B): Coronal muscle (CM)
and radial muscle of Cyanea sp.
50

x
Figure 3.17 Subumbrella morphology of gonad. (A): Gonad (G)
protruding from the subgenital fenestration of Cyanea sp.
(B): Gonad (G) at the base of the gastrovascular cavity of
Versuriga anadyomene
51
Figure 3.18 Gross morphology of Chrysaora chinensis. (A): Whole
medusa floated in a tank, showing four soft and curtain-
like oral arms (OA) and tentacles (T); (B): Side view of the
bell (umbrella) with reddish brown pigmentation on the
lappet (L); (C): Oral arms (OA1) without pigmentation;
(D): Oral arms (OA2) with reddish pigmentation; (E):
Subumbrella view of the central stomach (S)
54
Figure 3.19 Umbrella morphology of C.chinensis. (A): Subumbrella
view revealing 16 gastrovascular pouches, rhopalar pouch
(P1), inter-rhopalar pouch (P2), rhopalium (R), lappet (L),
tentacle (T) and oral arms (OA); (B): Subumbrellla view of
creamy white gonads (G), whereby each of the four gonads
is located inside a subgenital fenestration of the
subumbrella; (C): Subumbrella view showing the mouth
(M) bounded by four walls; (D): View of exumbrella with
numerous warts (W) densely concentrated in the centre;
(E): View of subumbrella showing lappet (L), rhopalia (R)
and three tentacles (T) following a 2-1-2 arrangement per
octant, whereby the numbers represent the ontogenetic
sequence of tentacles originating from the cleft of the
lappet
55
Figure 3.20 Gross morphology of Cyanea sp. (A): Whole medusa (B):
Numerous tentacles (T) and curtain like oral arms (OA);
(C): View of exumbrella with numerous warts (W); (D):
Subumbrella showing a group of radial muscle (RM) and a
horseshoe shaped whorl of tentacles (T) between each
group of radial muscle;. (E) Subumbrella view showing a
group of coronal muscle (CM)
58
Figure 3.21 Subumbrella morphology and structures of Cyanea sp. (A):
Subumbrella view of the medusa. (R) Rhopalium. (G)
Gonad. (T) Tentacle. (L) Lappet. (CM) Coronal muscle.
(RM) Radial muscle. (B): A whorl of tentacle (T). Radial
muscle (RM). Coronal muscle (CM), alternating between a
longer group and a shorter group. (C): Lappet shape broad
and semi-circular, with network of canal (C). (D): Gonad
(G) creamy white.
59
Figure 3.22
Gross morphology of Rhopilema esculentum (A): Whole
medusa. (B): Exumbrealla (EX) smooth. (C): Terminal
club (C). (D): 16 scapulae (S) in each medusa. (E): One
subgenital fenestration (F) for each quadrant. One papilae
(P) at the centre of the fenestration. (F): Oral arm (OA).
62

xi
Figure 3.23 Subumbrella morphology and structures of Rhopilema
esculentum (A): Gonad (G) creamy white. (B): Lappet (L).
Coronal muscle (M). Rhopalia (R). (C): Network of canals
injected with green dye
63
Figure 3.24 Gross morphology of Rhopilema hispidum (A): Whole
medusa. (B): Three papilae (P) for each quadrant where the
largest is in the centre. (F) Filaments. (C): Blade shape
scapulae (S) with numerous filaments (F). (D): Exumbrella
surface rough, with numerous warts (W). (E): Lappet (L).
(F): Network of canal (C) injected with blue dye, forming
a pyramid shape of network
66
Figure 3.25 Gross morphology of Lobonemoides robustus (A): Whole
medusa. (F) Filaments. (B): Gonad (G). (C, D): Conical
papillae on the exumbrella in water (P1), and on dry stage
(P2).
69
Figure 3.26 Morphology of Lobonemoides robustus (A): Coronal
musculatures (M) with colouration. Rhopalium (R). (B):
Spindle shape club (C). (C): Network of canal (C) with
yellow dye injected. (G) Gonad. (D): Lappet (L) elongated.
(E): Oral arm (OA) with yellow dye injected
70
Figure 3.27 Gross morphology of Versuriga anadyomene (A): Whole
medusa. Oral pillar (OP). Numerous mouthlets (MO) on
oral arms. Appendage (A). (B): Lappet (L). (C):
Portuberances (P) on exumberalla. (D): Numerous
filaments on the oral disk. Coronal muscle (M). (OA) Oral
arm.
73
Figure 3.28 Gross morphology of Versuriga anadyomene (A): Oral arm
with numerous mouthlets (M). A window on the oral arm
(W). (B): Gonad (G) brownish. (C): Network of canals,
injected with blue dye
74
Figure 3.29 Gross morphology of Phyllorhiza punctata (A): Whole
mesusa. Numerous warts (W) on the exumbrella. Terminal
club (TC). (B): Numerous mouthlets (MO) on the oral arm.
(C): Filaments (F) on oral disk. (D): Oral pillar (OP). The
white ring is a wire to suspend the jellyfish and it is not
part of the jellyfish structure
77
Figure 3.30
Gross morphology Phyllorhiza punctata (A): Coronal
musculatures (M). (R) Rhopalia (L) Lappet. (B): Warts
(W) on exumbrella. (C): Numerous appendages (A) on the
oral arms. Terminal club (C). (D): Gonad (G) brownish.
(E): Network of canals (C) injected with blue dye
78

xii
Figure 3.31 Gross morphology of Lychnorhiza malayensis (A): Whole
medusa. (OA) Oral arm. (B): Rhopalium (R). Lappet (L).
(C): Network of canals (C) injected with red dye. (D):
Gonad (G) creamy white. (P) Papillae. (E): Oral arm (OA).
(F) Papillae
81
Figure 3.32 Gross morphology of Acromitus flagelatus (A): Whole
medusa. Oral arm (OA). (B): Lappet (L) pointy.
Rhopalium (R). (C): Network of canals (C) injected with
red dye. (D): Papilae (P). Coronal muscle (M). (E): A
single terminal filament (F) on oral arm. (F): Numerous
filaments (F) on oral arms
84
Figure 3.33 Map of the sampling locations for geometric morphometric
analysis, with the number of specimens obtained. Green
circle represents East-North (EN), red circle represent
East-Centre (EC), blue represents West-Central (WC), and
purple represents West-North (WN) coastal areas of
Peninsular Malaysia
85
Figure 3.34 PCA Result – % of variation explained by components.
Two independent contrasts of the gastrovascular pouch
shape of the two main components PC1 and PC2 are
illustrated whereby light blue outline indicates the mean
shape and the dark blue outline indicates the shape change
87
Figure 3.35
Scatter plots of PC1 vs PC 2. PC1 accounted for 33.86%
and PC2 accounted for 19.41% of the total variance of the
shape change of gastrovascular pouch of specimens from
East-Central (EC), East-North (EN), West-Central (WC)
and West-North (WN) of Peninsular Malaysia.
88
Figure 3.36
Independent contrast of component using canonical variate
analysis of the gastrovascular pouch of specimens from
East-Central (EC), East-North (EN), West-Central (WC)
and West-North (WN) of Peninsular Malaysia, whereby
CV1, CV2 and CV3 accounts for 47.46%, 32.72% and
19.83% of the amount of relative between-group variation,
respectively. Light blue outline indicates the mean shape
and the dark blue outline indicates the shape change. CV1
denotes changes with blunt protuberance at the margin and
enlarging of the distal end, CV2 denotes changes with
blunt protuberance at the margin and CV3 denotes changes
with blunt protuberance at the margin and enlarging of the
distal end
90

xiii
Figure 3.37 Scatter plot of CV1 vs CV2 and illustration of mean shape
of gastrovascular pouch of specimens from four coastal
areas East-Central (EC), East-North (EN), West-Central
(WC) and West-North (WN) of Peninsular Malaysia. Plot
shows shape differences mainly between specimens of the
east and west coasts, even between those from EN and EC,
but with no distinct differences between those from WN
and WC
91
Figure 3.38 Pairwise comparisons between the pouch shape of
C.chinensis populations from East-Central (EC), East-
North (EN), West-Central (WC) and West-North (WN) of
Peninsular Malaysia
92

xiv
LIST OF TABLE
Table 2.1 The sites where samples of jellyfish were collected off the
coasts of Peninsular Malaysia, with the period, GPS and
sampling methods
16
Table 2.2 The sites where samples of C. chinensis were collected off the
coasts of Peninsular Malaysia, with the sample sizes (n) and
sampling methods. The sites are designated as coastal areas
West-North (WN), West-Central (WC), East-North (EN) and
East Central (EC) of Peninsular Malaysia
17
Table 2.3 Definition of the 16 landmark configurations
24
Table 3.1 Result of Procrustes ANOVA (SS=Sum of square, MS=Mean
Squre, df=degree of freedom)
86
Table 3.2 Mahalanobis distances among the pouch shape of C. chinensis
populations from four coastal areas designated as West-North
(WN), West-Central (WC), East-North (EN) and East Central
(EC) of Peninsular Malaysia
91
Table 4.1 Comparison of different Cyanea species 95

1
CHAPTER 1: INTRODUCTION
1.1 Jellyfish Studies in Malaysia
The term ‘jellyfish’ generally refers to free-floating gelatinous animals that belong to
the phyla Ctenophora and Cnidaria (Figure 1.1). The phylum Ctenaphora consists of
organisms such as sea gooseberry and comb jellies, whereas the phylum Cnidaria
consists of a wide range of both the sessile and floating forms of organisms that
includes jellyfish, corals and sea anemones (http://www.marinespecies.org). All
Cnidarian possess cnidae - an organelle-like capsule with eversible tubules, and it is
considered as the diagnostic feature of Cnidarian. There are three main classes under the
phylum Cnidaria: Cubozoa (46 accepted species), Hydrozoa, and Scyphozoa (187
accepted species) (Figure 1.2). All species under Scyphozoa, Hydrozoa and Cubozoa
develop the ‘medusa’ or jellyfish stage in the life cycles. Class Scyphozoa is ascribed
with four orders, namely Coronatae (crown jellyfish), Staurozoa (stalked jellyfish),
Semaeostome (sea nettle) and Rhizostomae (true jellyfish), with 65 genera and over 187
species (Mayer, 1910; Kramp, 1961; Pitt & Kingsford, 2003; Brusca & Brusca 2002;
Shao et al., 2006; Daly, 2007; Richardson et al., 2009; Bayha, 2010).
The most commonly observed jellyfish are usually those from the class
Scyphozoa, particularly from the medusa life stage. They begin their life cycle from
fertilized eggs that produces planulae. These free swimming planulae (Figure 1.3) then
settled at the substrate, becoming scyphistomas, also known as polyps. During this
sessile polyp stage, Scyphistomas asexually buds and strobilate to produce ephyra.
Strobilation is a process where each layer of the scyphistoma is separated and form a
new juvenile jellyfish, resembling a disc being liberated from a stack. This polydisc

2
strobilation only found in Scyphozoan. Although most of the scyphozoan have a polyp
stage, sometimes a direct development from planula to ephyra is also possible (Arai,
1997; Boero et al., 2008; Ceh, 2015). The adult medusa stages is dominant in the life
cycle. Therefore, this free-swimming form is most commonly seen and found stranded
on beaches (Figure 1.4).
Jellyfish are 97% water and are semi-transparent. They have two body layers,
the outer layer epidermis and the inter layer gastrodermis (Figure 1.5). Between both
layers is a thick layer of mesoglea which consists of fibres embedded in a hydrated
matrix that contains cells. These layers of tissues make up the umbrella of the jellyfish
which is usually bell shape, thus the umbrella is also known as the bell. The scyphozoan
jellyfish are tetraradially symmetrical, meaning having many structures in multiples of
four. It contains a simple gastrovascular cavity which acts as stomach. They are also
characterized by having gastric filament in the stomach. Some scyphozoan jellyfish
such as Semaestomeae contain an opening, or mouth at the subumbrella. There are four
to eight oral arms near the mouth, which functions as arms to capture and transport food
to the gastrovascular cavity. Jellyfish lack eyes, but possess many sensory receptors
capable to detect light, pressure, temperature and gravity. These sensory receptors are
concentrated in the marginal sense organ that contains the rhopalium (Nakanishi, 2015).
Not all jellyfish possess tentacles. For Semaestomeae jellyfish, tentacles can be found at
the margin of the bell or at the subumbrella whereas tentacles are absence from the
Rhizostomeae jellyfish. Jellyfish contains network of canals that usually anestomoses
with each others that formed various patterns (Hamner, 1995; Arai, 1997; Hale, 1999).

3
Figure 1.1: An adult scyphozoan jellyfish, Phyllorhiza punctata
Figure 1.2: Classification of Cnidarian

4
Figure 1.3: Life cycle of jellyfish
(http://thescyphozoan.ucmerced.edu/Biol/Ecol/LifeHistory/ScyphozoaLH.html)
Figure 1.4: Bloom and stranding of Crambione mastigophora in Pulau Ketam in April
2016 (Sin Chew Daily, 20 April 2016)

5
Figure 1.5: Cross section of a jellyfish (Hale, 1999)
The food source generated from the ocean today has been greatly reduced
compared to from decades ago, due to factors such as global warming, ocean
acidification and over fishing (Morton, 2005; Alison et al., 2009). Jellyfish has long
been a delicacy in Asia for centuries and has been increasingly important as a food
source (Hsieh et al., 2001; Pitt & Kingsford, 2003; You et al., 2007). But not all
jellyfish species are edible, and some are not economically to be produced as food due
to its size. A traditionally non edible jellyfish, Cyanea nozakii was produced into food
in China but because of the low quality and unpleasant taste, the prices of food made
from Cyanea nozakii fetched a much lower price than the more sought after Rhopilema
esculentum (Dong et al., 2010). Today, the demand of edible jellyfish is greater than
ever with more people from China, Japan, Korea, Singapore, Thailand and Malaysia
consuming it. Jellyfish are marinated with a mixture of salt and alum over a period of
approximately seven days, and the semi-dried products are sold as food ready for

6
consumption. A few commonly sought after edible jellyfish in this region are the “red
type” (Rhopilema esculentum), “white type” (Lobonemoides robustus), “river type”
(Acromitus hardenbergi), “ball type” or “sunflower type” (Crambionella annadalei),
“prigi type” (Crambione mastigophora), “sand type” (Rhopilema hispidum) (Omori &
Kitamura, 2004).
Despite its value, jellyfish on the other hand do pose treat to human and the
environment (Uye et al., 2010; Graham et al., 2015). Many jellyfish are venomous, and
some are even deadly. Beaches are forced to be closed when large quantity of venomous
jellyfish swamp the beach, causing lost in tourism revenue (Lucas, 2001; Purcell et al.,
2009; Gershwin, 2010). Jellyfish blooms have been reported extensively over the past
few decades (Brodeur, 2008; Kogovsek et al., 2010), notably the bloom of Pelagia
noctiluca throughout the Mediterranean Sea during the 1980s (Doyle et al., 2007),
bloom of Phyllorhiza punctata in the northern Gulf of Mexico in 2000 (Graham et al.,
2003), and the bloom of Nemopilema nomurai in the Sea of Japan in 2002 and 2003
(Kawahara et al., 2006). There are increasing evidence indicate that human activities
could attribute to the cause of jellyfish bloom, such as eutrophication (Purcell et al.,
2001; Parsons & Lalli, 2002; Malej et al., 2007), overfishing (Mullon et al., 2005;
Bakun & Weeks, 2006), introduction of alien species (Bolton & Graham, 2004; Graham
& Bayha, 2007; Mills, 2001), installation of artificial substrates in the ocean
(Richardson et al., 2009) and climate change (Raskoff, 2001; Attrill et al., 2007;
Gibbons & Richardson, 2008; Ruiz et al., 2012).
Beside blooms, one of the most alarming problems is the introduction of
nonindigenous jellyfish in a new region (Mills, 2001). They are believed to be
introduced to the new environment by mean of exchange of ballast water and transport
of biofouling polyps. (Richardson et al., 2009). Certain species of jellyfish previously

7
not known to other region has since appearing. These invasive species cause decline in
fisheries as they destroy the fishing net (Haddad & Nogueira, 2006).
Limited research was done on jellyfish in Malaysia. Due to the scarcity of
research effort in jellyfish, especially the morphological study of jellyfish in Malaysia
water, this region may harbor many species that yet to be discovered. Rumpet (1991)
and Daud (1998) published a field survey of scyphozoan jellyfish, whereas others
studied on their venom and toxinology (Othman & Burnett, 1990; Azila & Othman
1993; Tan et al., 1993), whereby species may have been misidentified. Jellyfish
fisheries in Malaysia are focusing on the “red type”, as it fetches the highest price due to
the high demand from China and Japan, whereas “white type”, “river type” and “sand
type” fetches relatively lower price (Omori & Nakano, 2001; Omori & Kitamura, 2004),
but these studies did not have satisfactory result on morphological descriptions and
taxonomy identification as the studies were only done on edible jellyfish. Base on
personal observations and reports, jellyfish often swamp and endanger beachgoers of
being stung, which sometimes can be lethal. They are often stuck in the cooling intake
of the power plant, causing damage to the power plant (Azila & Chong, 2010). It is also
notable that when they bloom, they were sometimes caught in the fishing net and
destroy them.
Despite their socio-economic importance in fisheries and also its treats, there
has been a lack of baseline data and documentation of jellyfish, particularly about their
diversity and ecology (Rizman-Idid et al., 2016). Jellyfish diversity studies in general
has been confounded by problems of identification. Their morphological examinations
are notoriously difficult, due to their fragile bodies, inadequate preservation and lack
of sound identification keys.

8
More recently, researchers have started using molecular genetic techniques to
facilitate the classification of jellyfish and detection of cryptic species (Dawson, 2004).
Applications of DNA sequence analysis and phylogenetics have been used to help
identify and barcode some of the Malaysian jellyfish species, including C. chinensis
(Rizman-Idid et al., 2016). Even with molecular techniques, species identification is
still based on the description of its morphology. Hence a more detailed morphological
evaluation of jellyfish in Malaysia is required.
1.2 Taxonomy Position and Problem of the Malaysia Sea Nettles
Jellyfish has a long history in evolution (Young & Hagadorn, 2010). Fossils have been
reported from as early as early Cambrian in China (Hou et al., 2005), and well-
preserved medusozoan fossils from the Middle Cambrian was reported in North
America (Cartwright et al., 2007). Ever since Linnaeus first described the popular moon
jellyfish Aurelia aurita in 1758, many studies have been done on numerous jellyfish
species throughout the years, but there are still many problems confounded with their
classification, such as the Malaysia sea nettles in the genus Chrysaora. The Chrysaora
jellyfish are classified under family Pelagidae (order Semaeostomeae) and typically
recognized by having 32-48 lappets, with eight marginal sense organs, with three or
more tentacles per octant, with 16 gastrovascular pouches, and with numerous warts on
the exumbrella (Kramp, 1961). Species of sea nettle has a worldwide distribution and
have been reported to occur in the South China Sea, North, Central and South America,
Africa, Europe and Australia (Morandini & Marques, 2010; Yap & Ong, 2012).
According to World Register of Marine Species (http://www.marinespecies.org) there
are possibly 15 -18 species, whereby 4 species have been verified; Chrysaora achlyos,
Chrysaora hysoscella, Chrysaora pacifica and Chrysaora quinquecirrha.

9
Although some regions of Southeast Asia have been reported to harbour
Chrysaora species such as - Chrysaora quinquecirrha and Chrysaora melanaster
(Kramp, 1961; Yap & Ong, 2012), there is the possibility they were misidentified base
on colour variations and warrants verification. In the past, jellyfish species were
notoriously difficult to identify as there were no reliable taxonomic keys, specimens
were badly preserved, and confounded by the existence of cryptic species complexes
that could be detected only by the application of molecular genetic techniques. For
example, Chrysaora chinensis in Malaysia has been previously identified as C.
hysoscella or C. quinquecirrha based on reports of envenomation and toxicology
studies of jellyfish stings (Azila & Othman, 1993) and those in Singapore straits as C.
melanaster (Yap & Ong, 2012) – possibly misidentified due to the colour variations.
Some features, such as nematocysts have been used to aid species identification,
whereby sea nettles from South China Sea were believed to be C. chinensis
(Morandini & Marques, 2010; Yap & Ong, 2012).
C. chinensis in Malaysia has tendencies to bloom, sometimes causing blockage
of cooling systems of coastal power plants, contamination of fishing nets and a nuisance
to fishing activities (personal observation). Beach tourism is also affected as beach
goers are often warned about its stings that are intense with painful burning sensation.
In general, jellyfish blooms have been linked to eutrophication and added nutrients to
the diet of the jellyfish (Richardson et al., 2009). Therefore, certain areas in
Malaysia were reported to have more occurances of C. chinensis. Moreover,
morphological adaptations to different localities with various ecological and
environmental conditions are well documented for jellyfish (Dawson, 2005). Thus, it
will be beneficial to study the morphological variation of C. chinensis in Malaysia.

10
1.3 Introduction to Geometric Morphometric Analysis
Shape analysis has long been an important and fundamental role in biological research
(Klingenberg, 2016). Traditionally, taxonomic classification was base mainly on
descriptive approach of morphology to formally describe species. Such shape
descriptions are sometimes ambiguous, and may not be sufficient to delineate species,
especially among closely related species that have high degree of morphological
resemblance. In the beginning of the twentieth century, researchers started employing
quantitative study of shape by meristic measurement such as length and width, and data
collected were subject to statistical analysis, using univariate, bivariate or
multivariateanalysis (Webster, 2010; Adams et al., 2013; Chen et al., 2013) to describe
the pattern of shape variation (Adams et al., 2004; Bookstein, 1998). With the
advancement of computing technology in the late 20th century multivariate
morphometrics are preferred, in which multiple measurements are analysed together
using Canonical Variates Analysis (CVA), Principal Components Analysis (PCA), and
other type of analysis (Polly et al., 2016). In the 1990’s, a new approach in studying
shape was introduced with the application of Geometric Morphometric Analysis
(GMM). GMM is the quantitative study of the biological shape, shape variation, and
covariation. Over the years, GMM has been improved and revised, with better effective
methods and softwares (Rohlf & Marcus, 1993; Adams et al., 2013). GMM have
employed outline and landmark methods.
Landmark coordinates were identified on the shape of the organism, and the
non-shape information contained in the data is removed by using the Procrustes
superimposition process. The data can then be further analyzed using statistical analysis
such as PCA and CVA.

11
One of the key advantages of GMM is that shape differences can be visually
displayed as illustrations or computer animations, making it the preferred method
compared to traditional morphometric which are usually shown as data such as length,
width and distance. (Rohlf & Marcus, 1993; Durón-Benítez & Huang, 2016). The
various methods of GMM visualization can illustrate even complex morphological
changes more effectively, making it appealing to researchers since the results are no
longer presented only as a series of statistical data but also as graphical representations
of the actual organism being studied (Rohlf & Marcus, 1993; Adams et al., 2004).
Furthermore, these visualizations provide information on morphological changes in
their immediate anatomical context (Klingenberg, 2013; Mayer et al., 2014).
GMM have been applied to a variety of organisms and structure such as oak leaf
(Viscosi & Cardini, 2011), dog (Drake & Klingenberg, 2010) and cichlid (Maderbacher
et al., 2008; Kerschbaumer & Sturmbauer, 2011) (Figure 1.6). Most organisms that
were analysed are usually rigid in structure. To date, there have yet to be any study of
shape of gelatinous organism such as jellyfish using GMM. Thus, this study is the first
of its kind to provide a more detailed morphological description of C.chinensis of
Peninsular Malaysia and to employ geometric morphometric analysis to distinguish its
populations.
Figure 1.6: GMM used on various organisms. A: Oak leaf. B: Dog. C: Cichlid

12
GMM can be performed using mathematical statistical packages such as R,
MATHEMATICA© and MATLAB©. There are also other open source integrated
software such as MorphoJ (Klingenberg, 2011), MORPHEUS (Slice, 2013) and IMP
(Sheets, 2011). R is a comprehensive GMM software because there are a number of
packages in it, and one can write script to perform analysis as wish. Thus, complex
statistical analysis can be performed in R. Since this study only require the standard
statistical analysis such as Procrustes superimposition, Canonical Variates Analysis and
Principal Components Analysis, software such as MorphoJ, MORPHEUS and IMP are
sufficient to perform the analysis needed for this study. Although these three software
have different graphical user interface, they essentially perform the same analysis with
the same numerical results. The preference of which software to use then is largely
depending on the familiarity and user-friendliness of the software. MorphoJ is choosen
to be used in this study because of the extensive training received in using the MorphoJ
from the author.
1.4 Research Aims and Questions
The aim of this research is to describe the jellyfish found in Peninsular Malaysia
water, and to compare the possible morphological variation within species or genus in
Malaysia water. Since this region may harbours many undiscovered species, detailed
documentation of morphology is required to allow for better identification and
comparison with conspecifics or congenerics in nearby waters. More recently,
applications of DNA sequence analysis and phylogenetics have been used to help
identify and barcode some of the Malaysian jellyfish species, including C. chinensis
(Rizman-Idid et al., 2016), which concurred the notion that specimens with different
colour morphs are often genetically similar and regarded as the same species. However,

13
it is important to realize that the efficiency of such DNA barcoding method relies on the
availability of reference sequences of correctly identified voucher specimens in the
Genbank database. Although the study provided 16S and ITS1 sequences, it did not
have the required cytochrome oxidase I (COI) sequences to definitively barcode C.
chinensis. Furthermore, the morphological description of the species in the study was
quite simple and preliminary, even lacking photographs, morphological illustrations and
have very simple descriptions. Thus many of previous morphological identification
needs to be reevaluated. Hence there is need to study them in more detail.
The sea nettle jellyfish of the genus Chrysaora has a worldwide distribution
and have been reported to occur in the South China Sea, North, Central and South
America, Africa, Europe and Australia (Morandini & Marques 2010; Yap & Ong
2012). Although some regions of Southeast Asia have been reported to harbour
Chrysaora species such as Chrysaora quinquecirrha and Chrysaora melanaster
(Kramp 1961; Yap & Ong 2012), there is the possibility they were misidentified base
on colour variations and warrants verification. In the past, jellyfish species were
difficult to identify as there were no reliable taxonomic keys, specimens were badly
preserved, and confounded by the existence of cryptic species complexes that could be
detected only by the application of molecular genetic techniques. Hence a more
detailed morphological evaluation of the Malaysian sea nettle (Chrysaora chinensis)
is required. Moreover, morphological adaptations to different localities with various
ecological and environmental conditions are well documented for jellyfish (Dawson,
2005). Base on personal observation, certain degrees of variations are found. For
example, gastrovascular pouches shape of C. chinensis are slightly different between
the species found in east coast and west of Peninsular Malaysia. Therefore, this
present study aims to distinguish morphologically the populations of Malaysian C.
chinensis by using geometric morphometrics, especially between populations of the

14
Straits of Malacca that are heavily impacted by anthropogenic activities from those
found in the South China Sea that has relatively better water quality. Thus, this study is
the first of its kind to provide a more detailed morphological description of
scyphozoan jellyfish in Peninsular Malaysia and to employ geometric morphometric
analysis to distinguish populations of C. chinensis.
There are two research questions in this project:
Question 1: What are the detailed morphological characteristics of different jellyfish
species in Peninsular Malaysia?
Question 2: Is there any gastrovascular pouch shape variation of Chrysaora chinensis
across different localities in Peninsular Malaysia?
1.5 Objectives of the Study
The objectives of this study are:
1. To identify, photograph and describe the detailed morphology of selected
scyphozoan jellyfish species in Peninsular Malaysia
2. To photograph the gastrovascular pouch of C. chinensis and establish its
landmark configuration for geometric morphometrics analysis
3. To discriminate between C.chinensis populations of Peninsular Malaysia by
analyzing the gastrovascular pouch shape variation using geometric
morphometrics and statistical analysis base on the configured pouch landmarks.

15
1.6 Significance of the Study
The proposed study has the following significance:
1) Some researchers, such as Kramp in his work “Synopsis of the medusae of the world
(1961)”, described various species of jellyfish in the Malayan Archipelago. But the
Malayan Archipelago encompasses a wide range of region, from Malaysia to Australia.
Thus it is unclear whether the specimen is found near Malaysia water. This research
could provide more accurate information on the jellyfish found in Malaysia water. The
findings of this study will contribute to the Malaysian checklist of marine species,
whereby baseline information of jellyfish diversity is still lacking. This study also
provide detailed description jellyfish species and relevant photographs of their
morphology that would facilitate their identification in the field.
2) This study would verify if sea nettle species in Peninsular Malaysia belong to
C.chinensis and determine if they consist of morphologically different populations.
GMM approach would also be first of its kind to be applied to jellyfish. This part of the
study would also contribute to baseline data needed for future management of sea nettle
blooms in Peninsular Malaysia.

16
CHAPTER 2: MATERIALS AND METHODS
2.1 Collection of Jellyfish Samples
Surveys and samplings in this study were carried out between April 2014 and
December 2014 from eight sites to obtain as many specimens possible that may
represent different jellyfish species and morphological variations from the east and west
coasts of Peninsular Malaysia (Table 2.1). Two sites located in the West-Central
(Sungai Janggut and Klang Power Station), three located in West-North (Pantai Kok,
Kilim and Balik Pulau), one located in East-Central (Kampung Cempaka), and two
located in East-North (Pantai Sabak, Pantai Melawi) coastal areas of Peninsular
Malaysia (Table 2.2). These sites were also chosen for the sampling of C.chinensis
samples for the GMM study. For the purpose of geometric morphometric analysis, a
minimum of 32 specimens were required, which was calculated based on 16 landmark
configurations used in the GMM.
Table 2.1: The sites where samples of jellyfish were collected off the coasts of
Peninsular Malaysia, with the period, GPS coordinates and sampling methods.
Site Period GPS Method
Sungai Janggut 9 Jul 2013* N03.16916° Bag Net
5 Dec 2013* E101.29833°
29 Apr 2014
Pantai Kok 14 Apr 2014 N06.34869° Dip Net
E99.64806°
Kilim 14 Apr 2014 N06.4766362° Dip Net E99.8042679°
Kampung. Cempaka 26 Jun 2014 N03.74365 o Dip Net
E103.32847 o

17
Table 2.1, continue
Pantai Sabak 20 Jul 2014 N06.13712o Dip Net
E102.36976o
Pantai Melawi 21 Jul 2014 N05.99612o Dip Net
E102.43714o
Klang Power 26 Jun 2014 N03.3229o Dip Net
Station E101.30108o
Balik Pulau 14 Dec 2014 N05.53972° Dip Net
E100.34888°
* Museum specimens of that was previously collected (not from this study) but
have been used for morphological examination
Table 2.2 The sites where samples of C. chinensis were collected off the coasts of
Peninsular Malaysia, with the sample sizes (n) and sampling methods. The sites are
designated as coastal areas West-North (WN), West-Central (WC), East-North (EN)
and East Central (EC) of Peninsular Malaysia.
Area Site Period GPS Method
West-North (WN) Pantai Kok 14 Apr 2014 N06.34869° Dip Net
(n = 38) (n = 5) E99.64806°
Balik Pulau 14 Dec 2014 N05.53972° Dip Net
(n = 33) E100.34888°
West-Central (WC) Sungai Janggut 9 Jul 2013* N03.16916° Bag Net
(n = 27) (n = 10 + 17*) 5 Dec 2013* E101.29833°
29 Apr 2014
East-North (EN) Pantai Sabak 20 Jul 2014 N06.13712o Dip Net
(n = 26) (n = 21) E102.36976o
Pantai Melawi 21 Jul 2014 N05.99612o Dip Net
(n = 5) E102.43714o
East-Central (EC) Kampung Cempaka 26 Jun 2014 N03.74365 o Dip Net
(n = 16) (n = 16) E103.32847 o
* Museum specimens of C. chinensis that was previously collected (not from this study)
but have been used for GMM in the present study.

18
Samples were obtained by using dip nets and bag nets. Dip nets with the mesh
size of 5mm were used to scoup out the jellyfish samples from the water surface and
kept in containers filled with seawater. Although specimens were less damaged, this
method is more labour intensive. Bag nets, also known as Pompang (Figure 2.1), were
methods used by fishermen in Sungai Janggut to collect pelagic jellyfish species.
Sixteen bag nets were deployed approximately 8m below the water surface during
spring tide and left for 6 hours before retrieving the nets. Bag nets were emptied on the
boat and jellyfish samples were sorted immediatey from the catch, and placed
temporarily into containers filled with seawater.
Figure 2.1: Diagram showing pompang (bag net)
Once at the jetty, jellyfish specimens that were relatively in good condition and
least damaged were chosen and photographed immediately to record their live
colouration by floating them in acrylic aquarium filled with seawater. Species of

19
specimens were tentatively identified in the field based on Kramp (1961). Bell diameter
and length of oral arms were measured on site using a measuring tape with the
exumbrella facing up. Specimens were cataloged and other sampling information such
as date and sampling site were recorded.
Tissue samples from oral arm and bell of each specimen were taken using
forceps and scissors that were cleaned with alcohol between each sampling to eliminate
cross contamination between samples. Tissue samples were rinsed with distilled water
and kept in vials containing 100% ethanol, cataloged and stored at 4C° for future
molecular genetic studies (not within the scope of the present study).
Each whole specimen was rinsed with distilled water to remove as much debris
as possible before being transferred into individual heavy duty plastic bag containing
5% formalin in seawater (Appendix A) and brought back to the laboratory. After seven
days of specimens being fixed in formalin, the specimens were rinsed with distilled
water to remove any remaining debris and transferred to a new container with 5%
formalin in filtered seawater to ensure better fixation.
2.2 Photography of Specimens and Collection of Morphological Data
Species identification of specimens were further verified in the laboratory based
on detailed morphological examination and taxonomic classifications of Kramp (1961)
and Morandini & Marques (2010). Prior to the examination, each specimen and its
catalog paper were removed carefully from its container, place into another container
with tap water and soaked for at least five minutes to remove the formalin as it is highly
carcinogenic. For safety measure, gloves and mask were used during handling of
specimens and examinations done in well ventilated area.

20
Morphological examinations of jellyfish specimens were done on major
structures such as bell, subumbrella, lappet, gastrovascular cavity, tentacle, muscle,
network, oral arm, scapulae, terminal club, filament and gonad. 186 detailed
morphological characters of jellyfish specimens were analysed, measured and their
respective character states recorded (Appendix B). A set of photographs were also taken
for those structures (Appendix C) to facilitate the morphological examination and
collection of morphological data. All examinations and photographs were based on a
modified data sheet guidelines of how to study the morphological characteristics of
scyphozoans, provided by courtesy of Michael Dawson and Liza Gomez Daglio from
University of California, Merced.
Morphology of specimens was photographed using a digital camera (Olympus
PEN Lite E-PL5, lense 18mm to 105mm) with additional light source, scale and colour
chart (Tiffen Q-13 Color Separation Guide) included so that measurements and the
colour code (CMYK) can be calibrated appropriately later from the digital images
(Figure 2.2). The specimens were photographed in two different methods. The first
method was by suspending the specimen using wires in a transparent glass aquarium
(50cm x 30cm) that was filled with water against a black background, so that it appears
to floating and positioned as natural as possible (Figure 2.3). The placement of the
specimen in the glass tank and the camera’s angle and distance from the specimen were
adjusted depending on the structure to be photographed, whether to obtain closeup or
overall images of the structure.
The second method is by laying the specimen on a flat transparent acrylic stage
(various sizes depending on the specimens) for dry and more detailed photography of
the morphology. A transparent acrylic glass measuring was used as the stage for
detailed photography of its morphology. A fluorescent light of 16 watt is placed 15cm

21
below the stage. Two extra light sources are placed one on each side of the stage (left
and right side) to provide extra light source. (Figure 2.3). In general, the specimen was
placed so that the orientation of rhopalia were aligned at 3, 6, 9 and 12 o’clock
accordingly, but the positioning of the specimen on the stage, the camera’s angle and
distance were adjusted depending on the various structures to be photographed.
Microsope and magnifying glass were used to observe minute structures such as
the gastric filaments. Visualisation of network of canals, gastrovascular pouches and
anastomoses in the lappets were enhanced by carefully injecting food dye (blue, green
or red) into the canals.
Figure 2.2: Photography setup to float jellyfish in the tank against a black background,
with colour chart and scale. Wires were used to suspend the specimens, and a scaled
wire was also used to aid recording of measurements.

22
Figure 2.3: Dry photography setup showing theacrylic stage, with light source below
and additional light sources at the sides.
2.3 Geometric Morphometric Analysis of Gastrovascular Pouch
Rhopalar gastrovascular pouch of C. chinensis was the chosen structure for the
geometric morphometric analysis as this internal structure often remains intact during
sampling compared to other external structures. Jellyfish are fragile organisms, and
quite often certain structures are destroyed during the sampling, such as detached
tentacles and oral arms, deformed exumbrella and others. Gastravascular pouch is
located at the underside of the bell of the jellyfish, and with the oral extending below it.
Therefore, it is relatively protected during sampling. The other reason for choosing
rhopalar gastrovascular pouch is because for Chrysaora species, they generally have two
shapes; the Pacific shape and the Atlantic shape. The Pacific septa shape is
characterized by first thinning of the tentacular pouch, then enlarging it, thus making an
“S” shape. Whereas the Atlantic septa shape is characterized by enlarging of the
tentacular pouch but gradually becomes thin, thus appearing pear shaped. Most of the
Chrysaora found in the Pacific Ocean such as C. chinensis, C. pacifica and C.
melancaster have the Pacific septa shape, whereas the Chrysaora found in the Atlantic

23
ocean such as C. quinquecirrha and C. hysoscella have the Atlantic septa shape
(Morandini & Marques, 2010). Therefore, there is reason to believe the biological
significance of the gastrovascular pouch variation base on different regions.
2.3.1 Landmark Configurations and Photography of Gastrovascular Pouches
Landmark-based geometric morphometrics methods begin with the
identification of the landmarks coordinate, either two or three-dimensional. Landmarks
are points on the Cartesian coordinates (x, y, z) that can be identified on each and every
specimen in the study to represent the shape. Although these landmark coordinates
contains information regarding the shape of the specimen, these coordinates data should
not be used directly because the effect of variation in position, orientation and size of
the specimens are still present in the coordinate. When studying shape of an organism,
the information regarding the position, orientation and size of the specimen is irrelevant.
Therefore, this non-shape information must be removed prior to the analysis of the
landmark coordinates (Klingenberg, 2013; Mitteroecker et al., 2015). Once the non-
shape variables are removed, these variables then can be used to perform statistically
analysis. The result of the analysis will be able to tell whether there is any variation in
shape comparison, in both statistical scatterplots as well as graphical representation of
the shape change (Adams et al., 2004). There are a few criteria in choosing landmarks.
Firstly, the landmarks must be of biologically significant. Secondly, landmarks must be
able to represent the morphology of the specimen, and all landmarks must be present on
all specimens, and must be reliably and repeatedly digitized for each specimen
(Klingenberg, 2013).
There are a total of 16 pouches for each specimen of C. chinensis, among them
eight rhopalar gastrovascular pouches are to the interest of this study. The reason why

24
these eight pouches were chosen over the other eight inter-rhopalar gastrovascular
pouches is because the rhopalia can be used as the guide to align the pouch more easily
along the 3 and 6 o’clock accordingly. Inter-rhopalar pouch could have been chosen to
be analised and it would have yield the same result since both pouches neighbouring
each other and shape variation on one pouch will also be shown on the neigbouring
pouch, but it will take more time to align the axis. 16 points along the edges of the
rhopalar grastovascular pouch were chosen as the landmarks for the GMM (Figure 2.4)
as those points represent the shape outline of pouch. Point 3 and 15 could have
beenomitted but they are used as the semi landmark since they are in the center of the
septa. The definition or position of the 16 landmark configurations are given in Table
2.3.
Table 2.3: Position and definition of the 16 landmark configurations.
Landmark Position
LM 1 Proximally, at the center of the rhopalar pouch nearest to the
gastrovascular cavity
LM 2 Proximally, at the upper left corner of the pouch
LM 3 Centrally, at the left septa of the pouch; half way between the top of the
pouch to the point where septa curves
LM 4 At the section where septa starting to curve outward
LM 5 Distally at the section where septa is curved at its furthest
LM 6 At the section where septa curves inward
LM 7 Distally at the lowest left point of the pouch
LM 8 Distally at the lower left of the septa next to the rhopalium
LM 9 Centrally at the rhopalium
LM 10 Distally at the lower right section of the septa next to the rhopalium
LM 11 Distally at the lowest right section of the pouch
LM 12 At the section where septa curves outward
LM 13 Distally at the section where septa is curved at its furthest
LM 14 At the section where septa starting to curve inward
LM 15 Centrally, at the right septa of the pouch; half way between the top of the
pouch to the point where septa curves
LM 16 Proximally, at the upper right corner of the rhopalar pouch
25
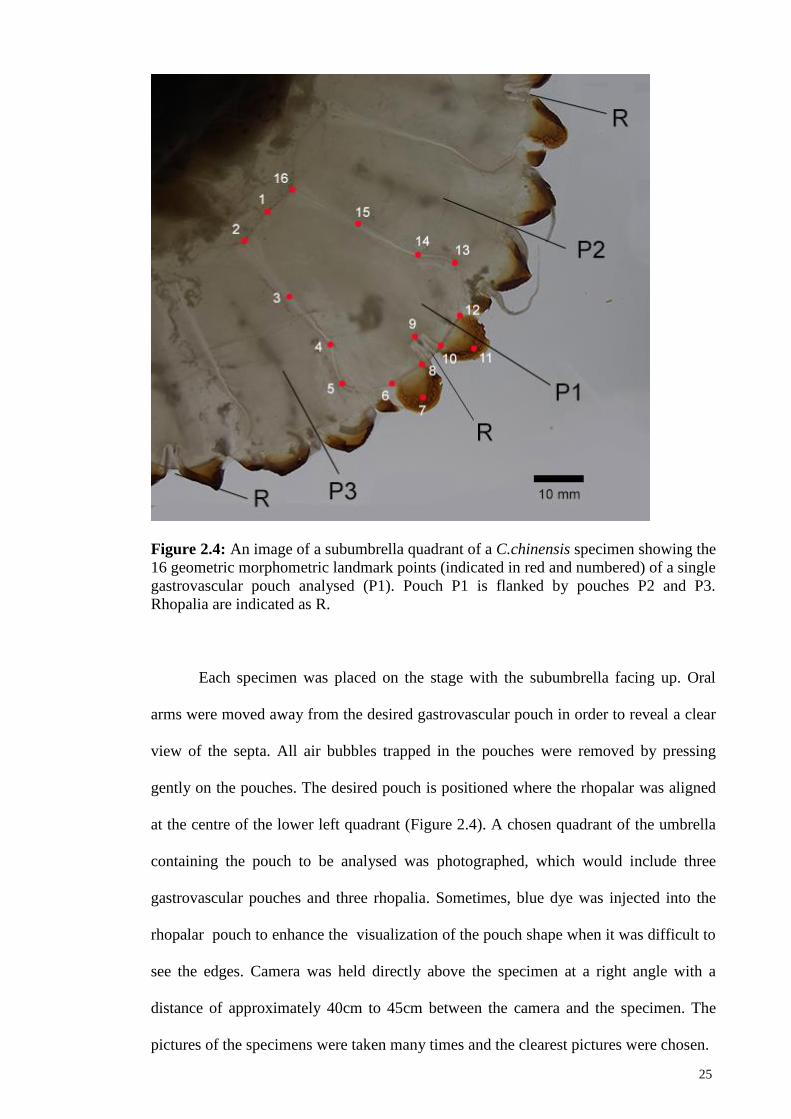
Figure 2.4: An image of a subumbrella quadrant of a C.chinensis specimen showing the
16 geometric morphometric landmark points (indicated in red and numbered) of a single
gastrovascular pouch analysed (P1). Pouch P1 is flanked by pouches P2 and P3.
Rhopalia are indicated as R.
Each specimen was placed on the stage with the subumbrella facing up. Oral
arms were moved away from the desired gastrovascular pouch in order to reveal a clear
view of the septa. All air bubbles trapped in the pouches were removed by pressing
gently on the pouches. The desired pouch is positioned where the rhopalar was aligned
at the centre of the lower left quadrant (Figure 2.4). A chosen quadrant of the umbrella
containing the pouch to be analysed was photographed, which would include three
gastrovascular pouches and three rhopalia. Sometimes, blue dye was injected into the
rhopalar pouch to enhance the visualization of the pouch shape when it was difficult to
see the edges. Camera was held directly above the specimen at a right angle with a
distance of approximately 40cm to 45cm between the camera and the specimen. The
pictures of the specimens were taken many times and the clearest pictures were chosen.

26
A total of 107 specimens were photographed (Appendix D). The minimum
requirement of specimen for a two-dimensional study is twice the number of the
landmarks. In this case, with 16 landmarks, the minimum specimen required is 32. Thus
a 107 specimen data size is well above the minimum requirement (Klingenberg, 2014).
For each specimen, two different pouches were photographed as replicates for each
specimen, making it a total of 214 images to be analysed.
2.3.2 Formating of Images Into TPS File
All 214 pouch images representing 107 specimens were converted into data
format in order to be processed by MorphoJ (Klingenberg, 2011) which is a software for
GMM. Images were saved in a folder in the computer. Images were converted to a TPS
file format using tpsUtil version 1.58 (Rohlf, 2010) (Appendix E).
2.3.3 Digitisation of Images
All pouch images were then digitised as an image information file in tpsDIG
(Rohlf, 2010). These digitised data contain information of the 16 defined landmarks that
were configured on the gastrovasular pouch. During the landmarking process of the
pouch in tpsDig software (Appendix F), the scale was adjusted so the sizes of images
were standardized. This was done by referring to the scale bar indicated in the images,
and calibrating the value of the scale into the tps file. Hence, image information of 214
pouch images representing 107 specimens, were digitized and imported into MorphoJ
for geometric morphometric analysis.

27
2.3.4 Quantification of Measurement Error
Measurement errors will always occur in any data collection procedure, and
errors can be attributed by various factors, such as imprecision of the measuring device,
the location of the landmarks, the rigidity of the specimen and the experience of the
researchers (Arnqvist & Martensson, 1998; Muñoz-Muñoz & Perpiñán, 2010;
Klingenberg, 2014). It is almost impossible to eliminate measurement errors
completely, thus it is important to reduce the measurement errors to as minimum as
possible.
A pilot study using a relatively small sample size can be performed to determine
the relative sizes of errors associated with each step. If the result shows that errors are
negligible, then replicate is not needed during the actual process.
One way of quantifying error in geometric morphometrics analysis study
involves imaging the specimen twice, and digitize (i.e. measurement) each image twice,
which build a 2 stages structure (i.e, imaging and measurement). For certain organisms
with symmetrical shape, another stage of “side” may be added to further investigate the
error associate with “side”. Procrustes ANOVA then can be performed to compute the
deviations of the individual values around the mean value at higher level (Figure 2.5).
The value at each level can then be used as relative sizes of errors associated with each
step. If the error is small, then replicate is not needed (Viscosi & Cardini, 2011).

28
Figure 2.5: The Number of replicate for each level of the process of specimen imaging
and digitization.
A preliminary quantification measurement error test was performed on 25
randomly chosen specimens using the Procrustes ANOVA analysis (Appendix G). For
each specimen, two gastrovascular pouches were photographed. Each pouch was
photographed twice, and each photograph was digitised twice. Therefore, three levels of
errors were measured: errors associated with “side”, imaging and the digitization of
landmarks, respectively. Hence, the total images used in Procrustes ANOVA for each
specimen was:
2 pouches (from each specimen) x 2 photographs x 2 digitizations = 8 images.
For 25 specimens, a tps file containing the image information of 200 images was
imported into MorphoJ to perform Procrustes ANOVA analysis.
2.3.5 Procrustes Superimposition
To remove the information of position, orientation and size of the specimens, a
procedure called the generalized Procrustes superimposition is performed.
(Klingenberg, 2013; Mitteroecker et al., 2013). The generalized Procrustes
superimposition is an iterative procedure that fits each configuration to the mean shape
in the sample as closely as possible. The end result of generalized Procrustes
superimposition is that only shape related information are extracted from the samples
landmark configurations.

29
Three stages were involved in the genaralized Procrustes superimposition. First,
variation in size was removed by scaling each configuration so that it has a centroid size
of 1.0. Secondly, variation in position was removed by shifting the landmark
configurations so that they shared the same centroid. The third procedure dealt with
rotations to find an optimal orientation for each configuration.
After the generalized Procrustes superimposition, every configuration in the
sample is then optimally aligned to the average configuration. Because the
configurations were aligned so that position, orientation and size were kept constant
according to the criterion for the least-squared fit, the remaining variation in landmark
positions will be solely the variation of shape only. The relative position of the
landmarks from the mean shape to another shape can then be used to detect the shape
variation. These relative position of the landmarks also provide a visualization of the
shape change by showing how the landmarks are reposition against each other after the
non-shape components variation of position, orientation and size are removed using the
Procrustes superimposition. (Klingenberg, 2013).
In this study, a generalized Procrustes superimposition was performed on the
gastrovascular pouch to eliminate the non-shape information (size, position and
orientation) of the image (Klingenberg, 2016). A covariance matrix was initially
generated before running the Procrustes analysis to produce a result set (Appendix H).
Once the Procrustes analysis was completed, two types of statistical analyses were
performed: the Principal Component Analysis (PCA) and Canonical Variate Analysis
(CVA).

30
2.3.6 Principle Component Analysis Using MorphoJ
PCA is a multivariate analysis whereby a numbers of factors, or components
that can be used to represent relationships among the data set with many possible
correlated variables are identified. In PCA, the basic principal is to identify a few major
components that explain most of the total variance. The first principal component (PC)
accounts for the most variability in the data set, the second PC accounts for the next
most variability, and follow by each succeeding PCs. Mathematically, this is done by
transforming the data to different axes so that those with the same pattern are aligned to
form a PC. In the context of geometric morphometrics analysis, the specimens can be
treated as points in a multivariate space where the viewing position of the data are
changed so that the most important components of the shape change are able to be
identified (Viscosi & Cardini, 2011).
PCA does not assume there any group membership in the data set. PCA will try
to maximize the variance on each of the component, and if the between-group variations
is bigger than the within-group variations, the scatterplots of PCA may show the group
variations. On the other hand, even if PCA failed to show any group varation, it doesn’t
necessary mean that there is no group variation in the data. (Strauss, 2010). Therefore,
PCA can be used as a preliminary review of the data, without making any assumptions
of group membership in the data set.
In this study, a PCA is performed for the general inspection of the gastravascular
pouch shape of C. chinensis (Appendix H).

31
2.3.7 Canonical Variate Analysis Using MorphoJ
The primary use of CVA in geometric morphometrics analysis is to determine
whether there is a significant difference in shape for pre-defined distinct groups in
multivariate data set. Therefore, in contrast to PCA, CVA makes the assumption that
there is group membership in the dataset. It tries to maximize the between-group
variation relative to the within-group variation. CVA is similar to PCA in the sense that
it realigns data to reconstructs new axes to form components, or factors.
There is a possibility that CVA will find distinction between group
memberships, even though a preliminary review by PCA may not indicate any
distinction between groups. Of course the underlying biological reasons to assign
specimens to different groups should be valid, or else the result generated by CVA may
not be significant (Sheets et al., 2001; Webster, 2010).
In this study, a CVA was performed on the gastrovascular pouches shape of C.
chinensis to distinguish populations by four coastal areas (Appendix H).
2.3.8 Visualization of Shape Outline
One of the main advantages of geometric morphometrics analysis is the ability
to visualize shape change. One of the widely used method for visualization is the
transformation grid. The deformation method is performed by placing a two-
dimensional rectangular grid over the first specimen, or the initial form, and the grid
was “stretched” to match the morphology of the second specimen, or the target form.
All parts of both specimens are still in the same grid cell after the transformation. The
change in the grid shows the differences in the shape of both specimens (Richtsmeier et
al., 2002). Later on, Bookstein (Bookstein, 1978), successfully adapt D’Arcy

32
Thompson’s transformation grid into thin plate spline, where he was able to construct
the transformation grids using an interpolation technique that fits the grids perfectly on
landmarks (Klingenberg, 2013).
Transformation grids by itself with only landmark configurations and grid can
be difficult for viewer to understand the morphological structure of the subject.
Furthermore, landmarks chosen do not sufficiently describe the morphology of the
subject of study. When all the landmarks are connected, the shape will usually not look
quite like the shape of the real organism as the connection between each landmark are
rather linear and sharp. Thus, to better visualize the shape, an outline drawing is used to
represent the organism. Sometimes, two outline drawings are imposed one on top of the
other to show the variation in shape. Although outline drawing is visually compelling in
displaying shape and variation in shape, it must be reminded that one must not rely
solely on outline drawing when analyzing shape and shape variation. Outline file is
merely a graphical representation of the shape of the specimen for the purpose of better
visualization. The disadvantage with outline file is that the points along the outline are
only assumptions, and may not represent the real shape of the specimen (Klingenberg,
2013).
In this study, an outline file was produced in tpsDIG using 56 landmark
coordination sets (Figure 2.6, Appendix I). They were able to represent the shape
outline of the gastrovascular pouch more closely to that of the specimen when compared
with outlines based on only 16 landmarks for the geometric morphometric analysis
process.

33
Figure 2.6: 56 landmark configurations used to create the outline of gastrovascular
pouch

34
CHAPTER 3: RESULTS
3.1 OVERVIEW OF MAJOR MORPHOLOGICAL STRUCTURES OF
JELLYFISH
Class Scyphozoa is ascribed with four orders, namely Stauromedusae Coronatae,
Semaeostomeae and Rhizostomeae, with 65 genera and over 187 species (Mayer, 1910;
Kramp 1961; Daly et al., 2007; Bayha et al., 2010). This study focuces on the order
Semaeostome and Rhizostomae only.
3.1.1 Order Semaeostomeae
The order Semaeostomeae composed of three families, four subfamilies, 18
genera and 56 species (Kramp, 1961). Semaestommeae jellyfish are characterized by
four oral arms around the mouth. Tentacles are found at the umbrella margin. (Arai,
1997). The two families of Semaeostomeae of the interest in this study are Cyaneidae
and Pelagiidae.
FAMILY CYANEIDAE
Family Cyaneidae is Semaeostomeae that central stomach gives rise to radiating
pouches, and it in turns gives rise to numerous blind canals extending towards the
marginal lappets; without a ring canal; with gonad completely folded; with tentacles
arising from the subumbrella distally from the margin.
Genus Cyanea – with eight rhopalia; with eight whorls of adradial tentacles, each
contains several rows of tentacles; radial and circular muscles in the subumbrella.

35
FAMILY PELAGIIDAE
Family Pelagiidae is Semaeostomeae that central stomach gives rise to radiating
pouches divided by a septa; without a ring canal; tentacles are formed at the cleft of the
bell margin; oral arms long and folded.
Genus Chrysaora – with 32 – 48 simple marginal lappets; with eight marginal sense
organs; with three or more tentacles for each octant; with 16 radial stomach pouches; in
the marginal area the eight rhopalar pouches are narrower than the eight tentacular
pouches, thus forming an “S” shape; exumbrella with numerous nematocyst warts.
3.1.2 Order Rhizostomeae
The order Rhizostomeae composed of two suborders, 10 families, 25 genera and
approximately 89 species (Kramp, 1961). Rhizostomeae jellyfish are characterized by
having bell margin cleft into lappet, with no tentacle on the bell margin, without a
central mouth, with eight oral arms extended from the subumbrella, where each oral
arms are bear numerous secondary mouths. Network of canals are found beyond the
stomach. (Kramp, 1961; Arai, 1997). This study focuces on the order Mastigiidae,
Versurigidae, Lychnorhizidea, Catostylidea, Lobonematidae and Rhizostomatidae.
FAMILY MASTIGIIDAE
Family Mastigiidae is charaterized with short, pyramidal, three-winged oral
arms; with numerous filaments on the oral disk (Kramp, 1961).

36
Genus Phyllorhiza - with broad oral arms, leaf-shaped, with window opening in the
oral arms, with numerous filaments at the oral arms; network of canals bounded by the
ring canal and do not anastomoses with the perradial rhopalar canals.
FAMILY VERSURIGIDAE
Family Versurigidae is charaterized with broad, leaf-shaped oral arms. (Kramp,
1961)
Genus Versuriga – with three-winged oral arms, with scapulaes, with terminal club,
with numerous filaments and clubs formed at the oral arms (Kramp, 1961).
FAMILY LYCHNORHIZIDAE
Family Lychnorhizidae is charaterized with centripetal, usually blinded end and
not anastomosing with the 16 canals; with broad and folded oral arms (Kramp, 1961).
Genus Lychnorhiza – with three-winged oral arms, without terminal club, with or
without filaments; with eight radial canals reaching bell margin, and another eight
reaching only the ring canal. (Kramp, 1961)
FAMILY CATOSTYLIDEA
Family Catostylidea is charaterized with intracircular network of canal
anastomosing with the ring canal but not always with the 16 radial canals; the eight
rhopalar canals reaching bell margin, and inter-rhopalar canals reaching only the ring
canal. Oral arms pyramidal (Kramp, 1961).

37
Genus Acromitus – Intracircular network of canals anastomosing with the ring canal
and the rhopalar canal, but not with the inter-rhopalar canals; oral arm is characterized
with a terminal club (Kramp, 1961).
FAMILY LOBONEMATIDAE
Family Lobonematidae is charaterized with intracircular network of canal
anastomosing with the ring canal and with some or all of the 16 – 32 radial canals, but
not with the stomach; with window openings in the membranes of the oral arms;
marginal lappets very elongated, tapering (Kramp, 1961).
Genus Lobonemoides – With large-meshed, intracircular network of canal
anastomosing with the ring canal and rhopalar canals.
FAMILY RHIZOSTOMATIDAE
Family Rhizostomatidae is charaterized with oral arms coalesced in proximal
portion only; without a mouth opening; with complicated network of canal system;
distal portion of the oral arms three winged, and usually with a terminal club (Kramp,
1961).
Genus Rhopilema – With large scapulae and long manubrium; oral arms with
numerous club and filaments, usually with a large terminal club; usually without a ring
canal; broad network of canal with numerous fine meshes; inter-rhopalar canal wide.
In this study, a total of 146 specimens (nine species from nine genera) were
obtained from the coast of Peninsular Malaysia (Figure 3.1 and Appendix J). Among

38
them, a total of 44 specimens (eight species from eight genera) were examined which
also included museum specimens (Appendix K). The nine species of scyphozoan
jellyfish described in this thesis were Chrysaora chinensis, Cyanea sp., Rhopilema
esculentum, Rhopilema hispidum, Lobonemoides robustus, Versuriga anadyomene,
Phyllorhiza punctata, Lychnorhiza malayensis and Acromitus flagellatus.
Figure 3.1: Map of eight sampling sites, with number of specimen collected for each
species.
Adult medusae of scyphozoan jellyfish (Figure 3.2) consists of a rounded bell
attached to a manubrium. Oral arms emerge from below the manubrium. In general,
jellyfish in the Semaeostomeae order possesses four oral arms, whereas those in the
Rhizostomeae order possess eight oral arms. At the edge of the bell are a series of
indentation called lappets. Tentacles are usually formed at the gap between lappets,
although not all jellyfish possess tentacle. A central stomach, or gastrovascular cavity is

39
found beneath the bell, or the subumbrella. It extends to the margin of the bell, forming
a network of canals. The complexity and the shape of the network vary between each
species.
Figure 3.2: Whole medusa of an adult jellyfish, Phyllorhiza punctata
Below is the definition of several major structure used for the description of the
morphology:
BELL - The bell, or the umbrella of adult scyphozoan medusae are usually rounded,
forming a hemispherical shape. The outer surface of the bell is the exumbrella, or also

40
known as the aboral surface; whereas the underside of the bell is the subumbrella or the
oral surface.
Figure 3.3: Morphology of bell of four different species of scyphozoan jellyfish. (A):
Acromitus flagellates with smooth surface (B): Rhopilema esculentum with smooth
surface (C): Phyllorhiza punctata with warts on the surface (D): Cyanea sp. With
smooth surface
LAPPET - The margin of the bell is usually clefted, and each of the indentation is
called the lappet. The lappet adjacent to the rhopalium is the rhopalar lappet, and the
rest of the lappets are the velar lappets. The lappets can be in many shapes, from
rounded, elongated to semi-squared.
Figure 3.4: Lappet of two different species of scyphozoan jellyfish. (A): Rhopalar
lappet (RL) and velar lappet (VL) of Lychnorhiza malayensis (B): Lappet of Cyanes sp.
(L). Lappet shape broad and semi-square
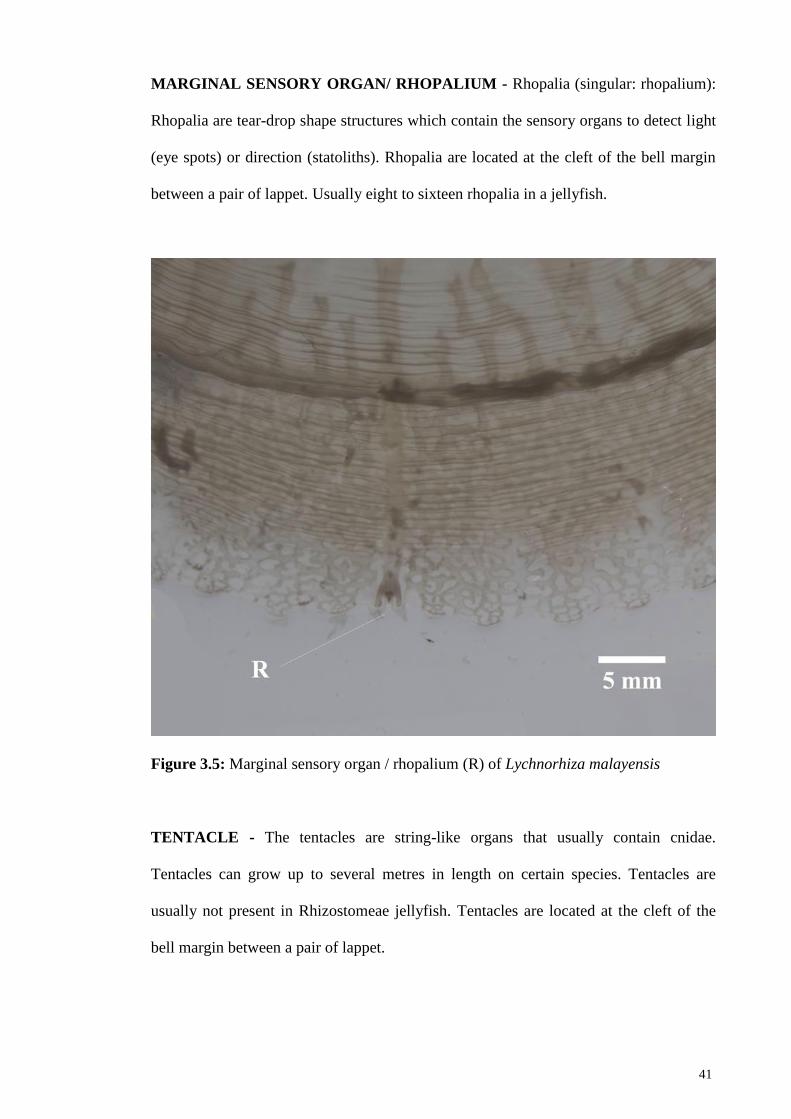
41
MARGINAL SENSORY ORGAN/ RHOPALIUM - Rhopalia (singular: rhopalium):
Rhopalia are tear-drop shape structures which contain the sensory organs to detect light
(eye spots) or direction (statoliths). Rhopalia are located at the cleft of the bell margin
between a pair of lappet. Usually eight to sixteen rhopalia in a jellyfish.
Figure 3.5: Marginal sensory organ / rhopalium (R) of Lychnorhiza malayensis
TENTACLE - The tentacles are string-like organs that usually contain cnidae.
Tentacles can grow up to several metres in length on certain species. Tentacles are
usually not present in Rhizostomeae jellyfish. Tentacles are located at the cleft of the
bell margin between a pair of lappet.
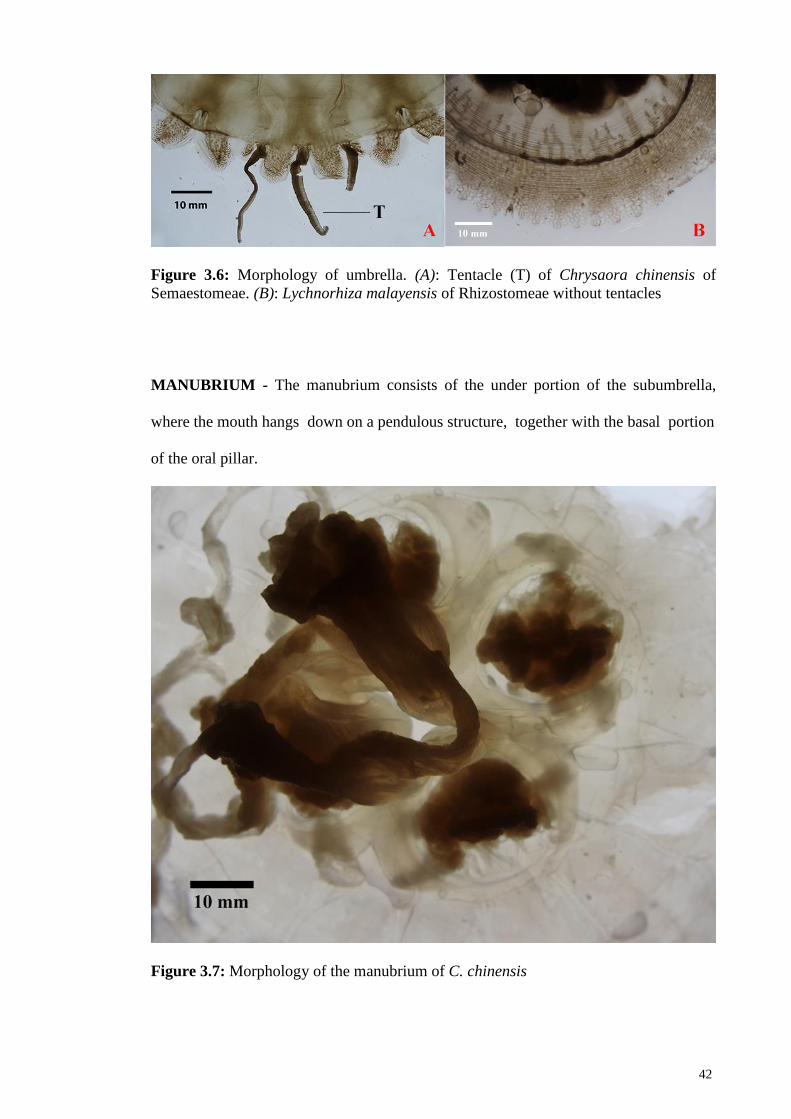
42
Figure 3.6: Morphology of umbrella. (A): Tentacle (T) of Chrysaora chinensis of
Semaestomeae. (B): Lychnorhiza malayensis of Rhizostomeae without tentacles
MANUBRIUM - The manubrium consists of the under portion of the subumbrella,
where the mouth hangs down on a pendulous structure, together with the basal portion
of the oral pillar.
Figure 3.7: Morphology of the manubrium of C. chinensis

43
ORAL ARM - Oral arm, also known as mouth arm is the organ extending from the
base of the manubrium, connected by the oral pillar. There are four oral arms for the
order Semaestomeae, and eight for the order Rhizostomeae. Oral arms are used to catch
prey for the former order, whereas the latter, food are transferred from the oral arms into
the stomach through second arms (or mouthlets) on the oral arms.
Figure 3.8: Gross morphology of oral arm. (A): Whole medusa of Chrysaora chinensis
floated in a tank, showing four soft and curtain-like oral arms (OA). (B): Oral arm (OA)
of Rhopilema esculentum. (C): Oral arm (OA) of Acromitus flagellatus

44
SCAPULAE - Scapulae are leaf-like organs attached to the manubrium. Usually two
scapulae are attached to each oral arm. Scapulae, like the oral arm, consist of numerous
mouth openings and stomach filaments.
Figure 3.9: Morphology of scapulae. (A): Blade shape scapulae of Rhopilema hispidum.
(B): Scapulae (S) attached to oral arms of Rhopilema esculentum
TERMINAL CLUB - Terminal club is the organ that originates at the margin of the
oral arm. It dangles at the tip of the oral arm.
Figure 3.10: Morphology of oral arm with terminal club. (A): Terminal club (C) at the
margin of the oral arm of Rhopilema esculentum. (B): Terminal club (C) at the margin
of the oral arm of Phyllorhiza punctata

45
FILAMENT - Filaments can be found on the oral disk and the oral arms. Filaments are
primarily used for swimming and feeding. Some species even have nematocyst on
filaments, and also used filaments to externally transport food to the mouth
Figure 3.11: Morphology of filaments. (A): Filament (F) at the oral disk of Phyllorhiza
punctata. (B): Filament (F) at the oral arms of Acromitus flagellatus
GASTROVASCULAR CAVITY - Stomach or gastrovascular cavity is the space
between the exumbrella and subumbrella that would contain products of digestions
which are distributed to the rest of the gastrovascular system. (Arai, 1997). The
distribution is done through a network of canal (see also the Network of Canal section
below). The gastravascular cavity in scyphozoan jellyfish is usually divided into four
compartments. Each compartment is separated from each other by a radial septa.

46
Figure 3.12: Morphology of subumbrella of Chrysaora chinensis revealing the
gastrovascular cavity
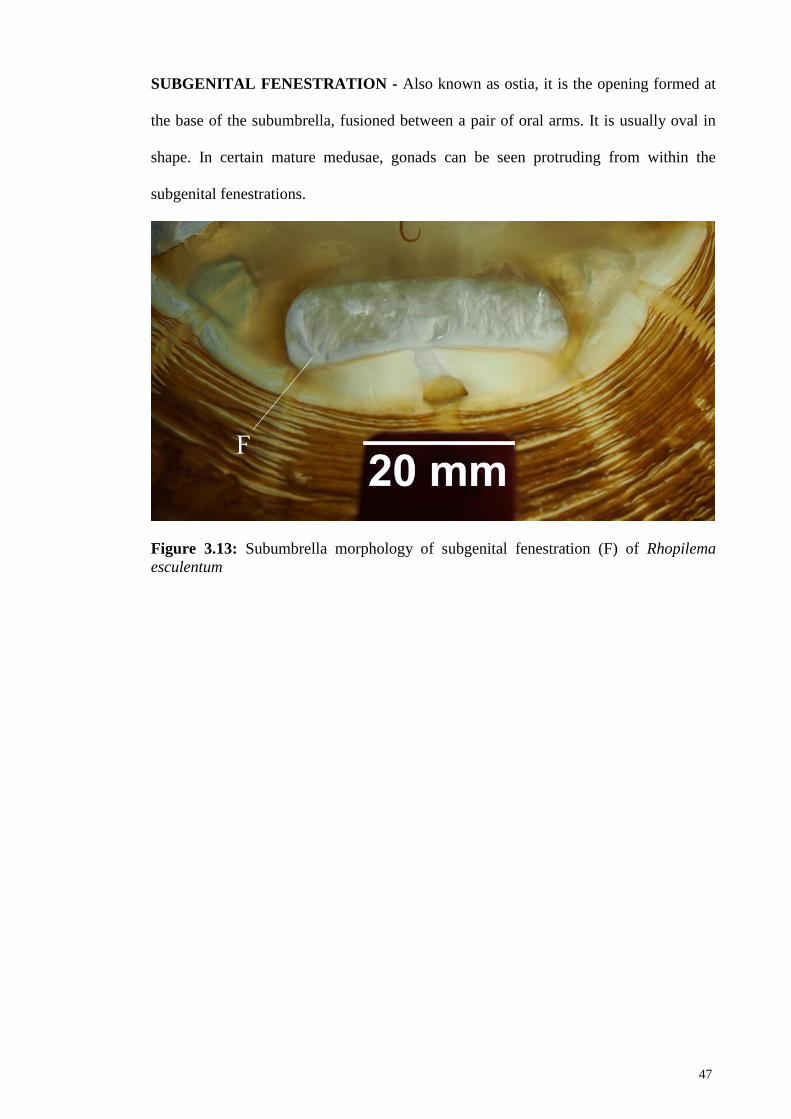
47
SUBGENITAL FENESTRATION - Also known as ostia, it is the opening formed at
the base of the subumbrella, fusioned between a pair of oral arms. It is usually oval in
shape. In certain mature medusae, gonads can be seen protruding from within the
subgenital fenestrations.
Figure 3.13: Subumbrella morphology of subgenital fenestration (F) of Rhopilema
esculentum

48
PAPILLAE - Papillae are protuberances located in the front or around the subgenital
fenestration. Some species usually have three papillaes for each subgenital fenestration.
One, usually a larger one, located at the central, and one smaller papillae one on each
side the subgenital fenestration.
Figure 3.14: Subumbrella morphology of papillae. (A): Three papillae (P) at the
subumbrella of Lychnorhiza malayensis. (B): Three papillae (P) at the subumbrella of
Rhopilema hispidum

49
NETWORK OF CANALS - Numerous tubes that radiating from the gastrovascular
cavitiy form a network of canals. The canals leading from the axis where gonads are
located are called the interradial canal. The canals leading from the axis between gonads
are called perradial canal. The canals between the interradial and perradial canals are
called the adradial canals. The complexity and the shape of the network of canals vary
between each species and can be useful for species identifcation.
Figure 3.15: Network of canals of (A): Lychnorhiza malayensis injected with red dye.
(B): Phyllorhiza punctata injected blue dye. (C): Lobonemoides robustus injected with
yellow dye
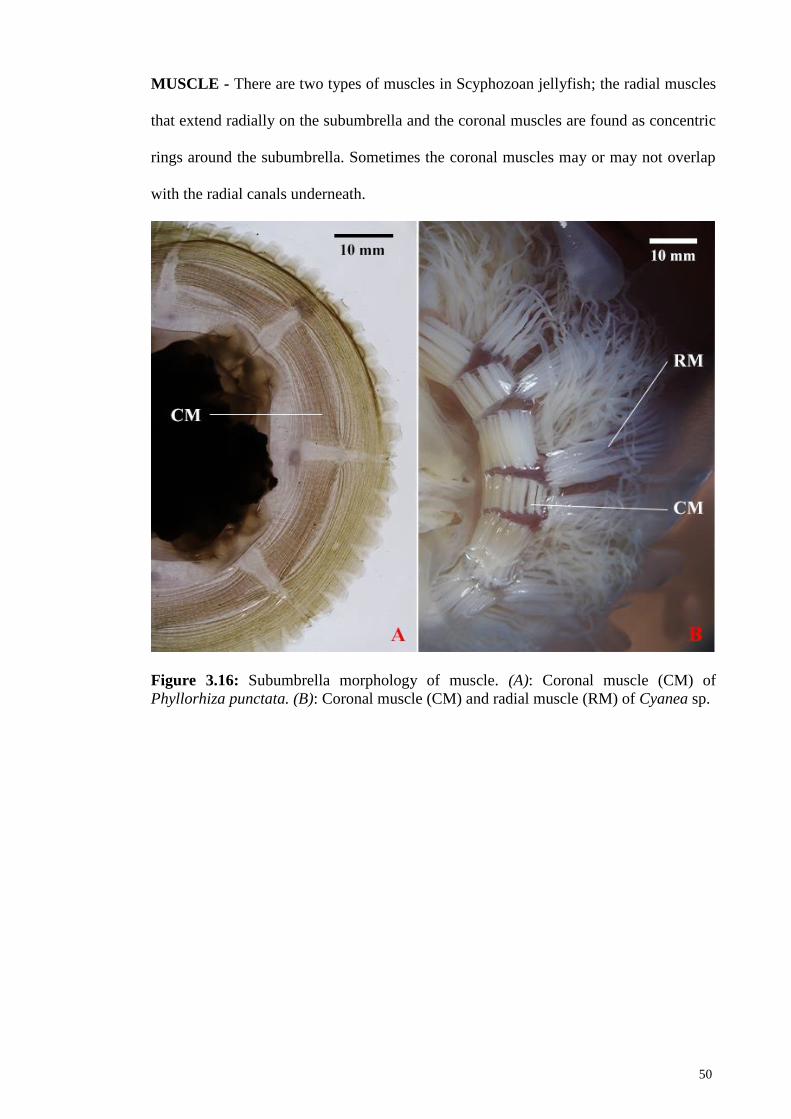
50
MUSCLE - There are two types of muscles in Scyphozoan jellyfish; the radial muscles
that extend radially on the subumbrella and the coronal muscles are found as concentric
rings around the subumbrella. Sometimes the coronal muscles may or may not overlap
with the radial canals underneath.
Figure 3.16: Subumbrella morphology of muscle. (A): Coronal muscle (CM) of
Phyllorhiza punctata. (B): Coronal muscle (CM) and radial muscle (RM) of Cyanea sp.
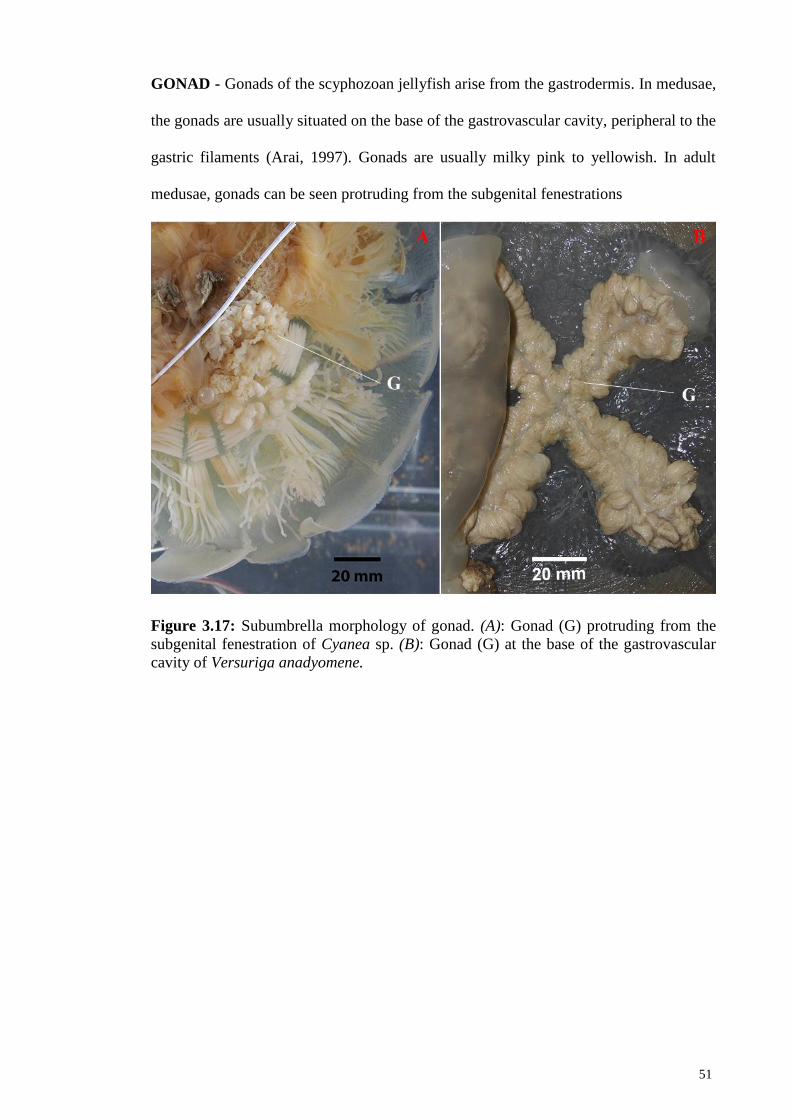
51
GONAD - Gonads of the scyphozoan jellyfish arise from the gastrodermis. In medusae,
the gonads are usually situated on the base of the gastrovascular cavity, peripheral to the
gastric filaments (Arai, 1997). Gonads are usually milky pink to yellowish. In adult
medusae, gonads can be seen protruding from the subgenital fenestrations
Figure 3.17: Subumbrella morphology of gonad. (A): Gonad (G) protruding from the
subgenital fenestration of Cyanea sp. (B): Gonad (G) at the base of the gastrovascular
cavity of Versuriga anadyomene.

52
3.2 Morphological Description of Chrysaora chinensis Vanhöffen 1888
Class SCYPHOZOA (Goette, 1887)
Order SEMAEOSTOMEAE (Agassiz, 1862)
PELAGIIDAE (Gegenbaur, 1856)
Chrysaora (Péron & Lesueur, 1809)
Chrysaora chinensis (Vanhöffen, 1888) resurrection by Morandini & Marques (2010)
(Figure 3.18 – 3.19)
Material Examined - A total of seven specimens were examined from sites Sungai
Janggut (MRI 2, MRI 65, MRI 163, MRI 171), Pantai Sabak (MRI 208), Balik Pulau
(MRI 262) and Kampung Cempaka (MRI 190).
Description of specimens- Bell: Umbrella hemispherical. For adult medusa,
exumbrella surface finely granulated; in some specimens the exumbrella is transparent
but some have light reddish spots distributed evenly on the surface. Mesoglea rigid,
thick in the centre and thinner towards the bell margin. Six lappets per octant; shape are
semi-circular, with rhopalar lappets. On certain specimens there is a small lappet
connecting to the rhopalar lappet. These lappets are loosely connected with the adjacent
lappet, and some of them appeared to be separated, therefore making eight lappets per
octant on certain specimens. Eight rhopalia present in clefts of the umbrella margin,
located at the interradial and perradial axis. Rhopalia shape is tear-drop like.
Tentacles: Three tentacles per octant (2-1-2, where the numbers represent the
ontogenetic order of the development of tentacles), originated at the cleft of the lappet.
Tentacles soft, straight and string-like. Tentacle length up to four times the diameter of

53
the bell. Personal field observations of the tentacles were much longer, more than four
times of the bell diameter, but sometimes detached when specimens were caught or
handled roughly.
Oral arms: Four oral arms, up to 200cm in length in adult medusa; oral arms formed at
the end of the oral pillar. Oral arms are transparent, soft and curtain like, with a ridge in
the centre and intricate folds at the edges. In certain specimens numerous reddish /
brownish spots are found all over the oral arm. Scapulae absent. Terminal club absent.
Filament absent.
Stomach, manubrium and radial canals: Central stomach shape slightly rounded, as
it is formed by the boundary of the 16 gastrovascular pouches, where eight are rhopalar
pouches and another eight are inter-rhopalar pouches. Radial septa present; extending
distally from stomach toward bell margin. For rhopalar pouches, septa starts widening
at approximately 1/3 from the centre, and thinning again, making an “S” shape that ends
at the base of the lappet. Subgenital fenestration shape oval, 1/8 of umbrella diameter.
Papillae absent. Network of canal absent. Radial and coronal musculatures not
prominent. Manubrium is not prominent. Oral disk at the base is about 1/3 as wide as
the bell diameter, and the base of oral arm is less than 1/3 as wide as the bell diameter.
Mouth present with four walls around the mouth. Quadralinga absent.
Gonads: Four gonads present, each located inside a subgenital fenestration of the
subumbrella; each gonad is separated with the neighbouring gonad by a septa;
associates with the subumbrella and manubrium. Colour creamy white. Gastric
filaments present.

54
Figure 3.18 Gross morphology of Chrysaora chinensis. (A): Whole medusa floated in a
tank, showing four soft and curtain-like oral arms (OA) and tentacles (T). (B): Side
view of the bell (umbrella) with reddish brown pigmentation on the lappet (L). (C): Oral
arm (OA1) without pigmentation; (D): Oral arm (OA2) with reddish pigmentation; (E):
Subumbrella view of the central stomach (S).
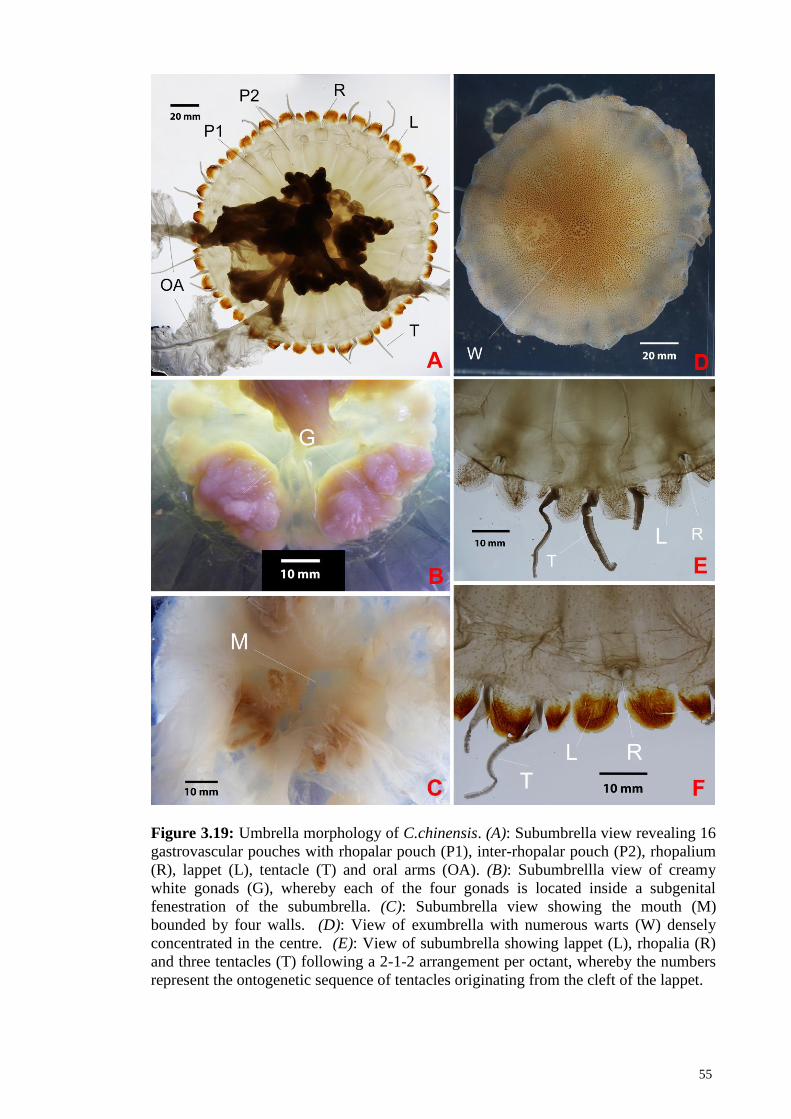
55
Figure 3.19: Umbrella morphology of C.chinensis. (A): Subumbrella view revealing 16
gastrovascular pouches with rhopalar pouch (P1), inter-rhopalar pouch (P2), rhopalium
(R), lappet (L), tentacle (T) and oral arms (OA). (B): Subumbrellla view of creamy
white gonads (G), whereby each of the four gonads is located inside a subgenital
fenestration of the subumbrella. (C): Subumbrella view showing the mouth (M)
bounded by four walls. (D): View of exumbrella with numerous warts (W) densely
concentrated in the centre. (E): View of subumbrella showing lappet (L), rhopalia (R)
and three tentacles (T) following a 2-1-2 arrangement per octant, whereby the numbers
represent the ontogenetic sequence of tentacles originating from the cleft of the lappet.

56
Figure 3.19, Continue
(F): Subumbrella view of another specimen showing distinctly pigmented lappet (L),
rhopalia (R) and tentacles (T).
3.3 Morphological Description of Cyanea Sp.
Class SCYPHOZOA (Goette, 1887)
Order SEMAEOSTOMEAE (Agassiz, 1862)
Cyaneidae (Agassiz, 1862)
Cyanea (Péron & Lesueur, 1810)
Cyanea Sp. (Figure 3.20 – 3.21)
Material Examined - A total of seven specimens were examined from sites Sungai
Janggut (MRI 55, MRI 56, MMRI 95, MRI 96, MRI 97, MRI 127 SJG 10)
Description of specimens - Bell: Flattened hemispherical. Mesoglea is rigid, thick in
the centre and thin towards the bell margin. Numerous faint warts are distributed on
exumbrella surface, denser at the centre of the bell and sparce towards the cleft of the
lappet. Sixteen equally broad sized lappets, semi square shaped along the margin of the
umbrella. All lappets overlap each other at the cleft. Rhopalar cleft is deeper than the
inter-rhopalar cleft. Eight tear drop shaped rhopalia, present in clefts of the umbrella
margin, located at the interradial and perradial axis. .
Tentacle - Eight whorls of tentacles originating from the coronal muscle at the
subumbrella. Each tentacle whorl is horseshoe shaped, consisting of two to three layers,
located between two groups of inter-rhopalar radial muscles. Tentacles are soft, straight
and string-like; up to 200 tentacles per whorl.

57
Oral arm - Four oral arms, up to four times as long as the diameter of the bell although
most were detached during the sampling process; oral arms formed at the end of the oral
pillar. Oral arms are transparent, soft and curtain like. Scapulae, terminal clubs and
filaments were absent.
Stomach and Manubrium - The digestive system of this species consists of a
gastrovascular cavity or central stomach, which is slightly round, as it is formed by the
boundary of the 16 coronal muscle groups. Sixteen pouches in the umbrella are
comprised of eight rhopalar pouches and eight tentacular pouches. Subgenital
fenestration is oval, 1/9 of the bell diameter. Papillae absent. Between six to seven
transverse “ridges” connecting both pouches below their surface. Both rhopalar pouches
and tentacular pouches also anastomoses with the lappets on both side of the cleft,
forming dense network on the lappet. Sixteen groups of coronal muscles originating
from the outer margin of the central stomach at the subumbrella, extending towards the
rim of the oral disk. Each coronal muscle group consists of six to eight muscle folds.
Bigger coronal muscle groups alternate with slightly smaller coronal muscle groups.
Muscle groups are not connected, with a ridge in between the two coronal muscle
groups. Gastrovascular pit absent. Sixteen groups of radial muscle originate at the
underside of the lappet, perpendicular to the coronal muscle. Each radial muscle group
consists of nine to eleven radiating muscle folds with different sizes and lengths, with
the longest muscle folds located in the centre of the muscle field. Manubrium thick and
smooth; oral disk at the base is about 1/3 as wide as the bell diameter.
Gonad - Gonads form below the subumbrella and attached with the subumbrella and
manubrium; each gonad is separated from the neighbouring gonads. Colour creamy
white. Gonads are big and mostly protruding out of the subgenital fenestration. Gastric
filaments present.

58
Figure 3.20: Gross morphology of Cyanea sp. (A): Whole medusa. (B): Numerous
tentacles (T) and curtain like oral arms (OA). (C): View of exumbrella with numerous
warts (W). (D): Subumbrella showing a group of radial muscle (RM) and a horseshoe
shaped whorl of tentacles (T) between each group of radial muscle. (E) Subumbrella
view showing a group of coronal muscle (CM).
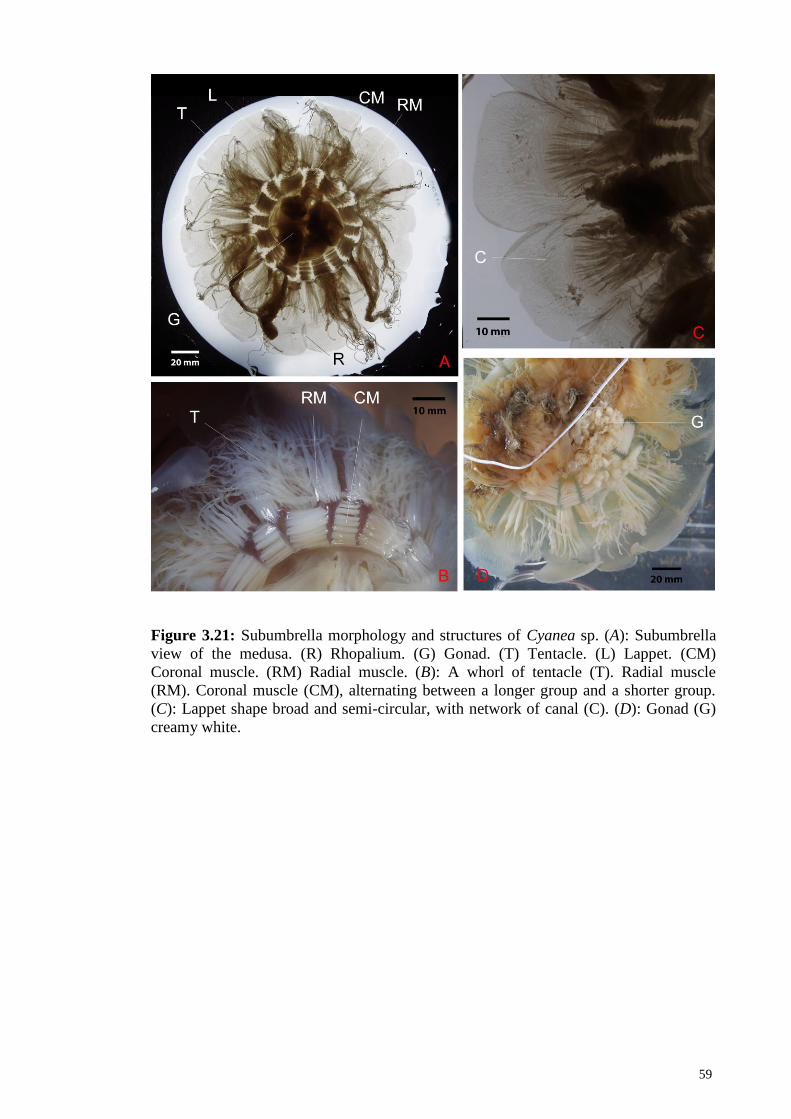
59
Figure 3.21: Subumbrella morphology and structures of Cyanea sp. (A): Subumbrella
view of the medusa. (R) Rhopalium. (G) Gonad. (T) Tentacle. (L) Lappet. (CM)
Coronal muscle. (RM) Radial muscle. (B): A whorl of tentacle (T). Radial muscle
(RM). Coronal muscle (CM), alternating between a longer group and a shorter group.
(C): Lappet shape broad and semi-circular, with network of canal (C). (D): Gonad (G)
creamy white.

60
3.4 Morphological Description of Rhopilema esculentum Kishinouye 1891
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
RHIZOSTOMATIDAE (Cuvier, 1799)
Rhopilema (Haeckel, 1880)
Rhopilema esculentum (Kishinouye, 1891) (Figure 3.22 – 3.23)
Material Examined - A total of three specimens were examined from site Sungai
Janggut (MRI 87, MRI 98, IND 3)
Description of specimens - Bell: Flattened hemispherical. Umbrella surface is smooth
and reddish. Mesoglea is rigid, thick in the centre and thin towards the bell margin.
Eight lappets per octant; shape are slightly elongated with pointy rhopalar lappets. Eight
tear-drop shaped rhopalia present in clefts of the umbrella margin, located at the
interradial and perradial axis. Tentacles absent.
Oral arm - Eight oral arms, as long as the bell diameter; oral arms formed at the end of
the oral pillar, then develop into three wings at the distal section (approximately at the
distal 1/5 of the oral arm). Two wings face away from the central axis (outer wings) and
one wing faces the central axis (inner wing). Numerous mouthlets located at the frills of
the wings. One to three spindle-shape appendages form at the frills of the wings.
Numerous filaments found at the frill of the wings. For each oral arm, two blade-shaped
but flat scapulae developed approximately 1/3 distally from the base of each oral pillar,
then develop into two wings for each scapulae. Winged portion of the scapulae is
smaller than the winged portion of the oral arms. Numerous mouthlets located at the
frill of the scapulae. Numerous short filaments located at the frills of the scapulae.
Terminal club absent.

61
Stomach and Manubrium - Gastovascular cavity or central stomach is floret shaped.
Each subgenital fenestration has one papillae located at the base of the manubrium and
near the edge of the subgenital fenestration. Papille dome shape. Papillae surface is
rough with numerous warts. Sixteen radial canals on the subumbrella are comprised of
eight rhopalar canals and eight inter-rhopalar canals; all canals fully extend to the
margin of the bell, where rhopalar canals split at the distal end and branch toward the
neighbouring lappets. Ring canal present. Rhopalar canals start anastomosing with the
neighbouring canals at about 1/3 from the centre, whereas interhopalar canals start
anastomosing with the neighbouring canals at about 1/2 from the centre. Radial muscle
absent. Sixteen coronal muscle fields cover the network mesh underneath. Coronal
muscle interrupted on the rhopalar canals but not on the inter-rhopalar canals.
Manubrium thick and smooth; oral disk at the base is about half as wide as the bell
diameter, and the base of oral arm is less than 1/6 as wide as the bell diameter.
Gonad - Gonad floret shape, formed at the subumbrella; each gonad is separated from
the neighbouring gonad; attached with the subumbrella and manubrium. Colour white.
Gastric filaments present.

62
Figure 3.22: Gross morphology of Rhopilema esculentum (A): Whole medusa. (B):
Exumbrealla (EX) smooth. (C): Terminal club (C) attached to oral arm. (D): 16
scapulae (S) in each medusa. (E): One subgenital fenestration (F) for each quadrant.
One papilae (P) at the centre of the fenestration. (F): Oral arm (OA).
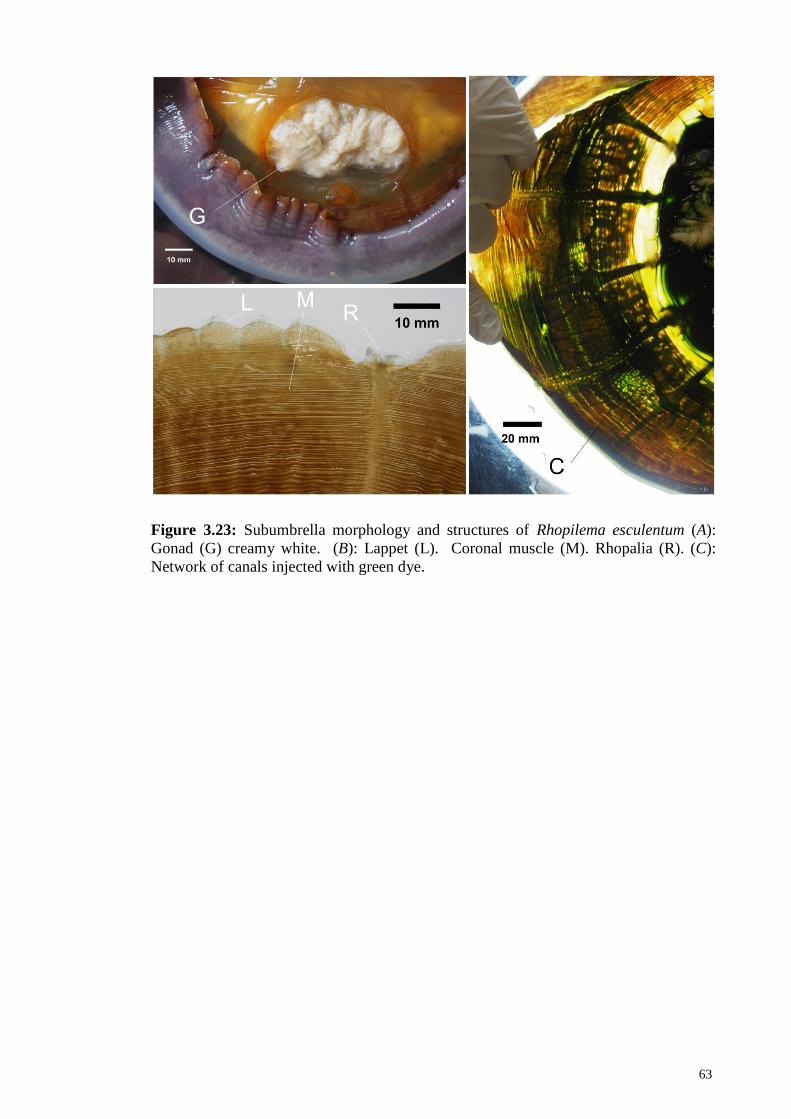
63
Figure 3.23: Subumbrella morphology and structures of Rhopilema esculentum (A):
Gonad (G) creamy white. (B): Lappet (L). Coronal muscle (M). Rhopalia (R). (C):
Network of canals injected with green dye.

64
3.5 Morphological Description of Rhopilema hispidum Vanhöffen 1888
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
RHIZOSTOMATIDAE (Cuvier, 1799)
Rhopilema (Haeckel, 1880)
Rhopilema hispidum (Vanhöffen, 1888) (Figure 3.24)
Material Examined – A total of four specimens were examined from site Sungai
Janggut (MRI 13, MRI 89, IND 9, IND 10).
Description of specimens - Bell: Umbrella hemispherical with a rough surface that
feels like sand paper. Numerous colourless warts and larger reddish brown circular
warts are found on the exumbrella and lappets. Warts are bigger and more concentrated
at the centre of the bell, decreasing in size towards the lappets. Eight slightly elongated
and pointy lappets located in each octant. The velar lappets adjacent to rhopalar lappets
are slightly bigger than the rest of the lappets. Short furrow forms between lappets,
whereas larger furrow forms between two pairs of velar lappets. Numerous warts are
concentrated on the lappets. Eight tear-drop shaped rhopalia present in clefts of the
umbrella margin, located at the interradial and perradial axis. Tentacles absent.
Oral arm - Eight oral arms, as long as the bell diameter; oral arms formed at the end of
the oral pillar, then develop to three wings at the distal section (approximately at the
distal 1/5 of the oral arm). Two wings face away from the central axis (outer wings) and
one wing faces the central axis (inner wing). Numerous mouthlets located at the frill of
the wings. Sixteen blade-shape but flat scapulae for each medusa. Four scapulae
developed approximately 1/3 distally from the base of each oral pillar, then develop to
two wings for each scapulae. Winged portion of the scapulae is smaller than the winged

65
portion of the oral arms. Numerous mouthlets located at the frill of the scapulae.
Numerous short filaments located at the frills of the scapulae. Terminal club absent
from the specimens observed, although there is a segment of left over hanging from the
end of the oral arm that may be the origin of the missing terminal club. In the field,
terminal clubs are observed. According to Omori (2001), a club shape terminal club is
found at the tip of the oral arm. Filaments are found at the scapulae.
Stomach and Manubrium - Gastrovascular cavity, or central stomach is floret shape.
Subgenital fenestration shape oval. 1/7 of the size of the bell diameter. Each subgenital
fenestration has three papillae. The largest papillae located at the upper side of the
subgenital fenestration, aligning with the rhopalar canal. One smaller on each side of the
subgenital fenestration. Papillae shape oval with rough surface. Sixteen radial canals on
the subumbrella comprised eight rhopalar canals and eight inter-rhopalar canals; all
canals fully extend to the margin of the bell, where rhopalar canals split at the distal end
and branch toward the neighbouring lappets. Inter-rhopalar doesn’t branch toward the
neighbouring lappets. Ring canal absent. A mesh of network formed between the canals,
where the proximal part of the network anastomosing more densely but decrease in size
towards the distal part. Rhopalar canal starts anastomosing with the neighbouring canals
at about 1/3 from the gastovascular cavity, whereas interhopalar canals start
anastomosing with the neighbouring canals at about ½ from the gastovascular cavity.
Three canals (two adradial, one perradial) connecting to the oral arm. Radial muscles
absent. Sixteen prominent coronal muscle fields is interrupted at the sixteen radii; each
field forms a pyramid shape. Manubrium thick and smooth; oral disk at the base is
about half as wide as the bell diameter, and the base of oral arm is less than half as wide
as the bell diameter.
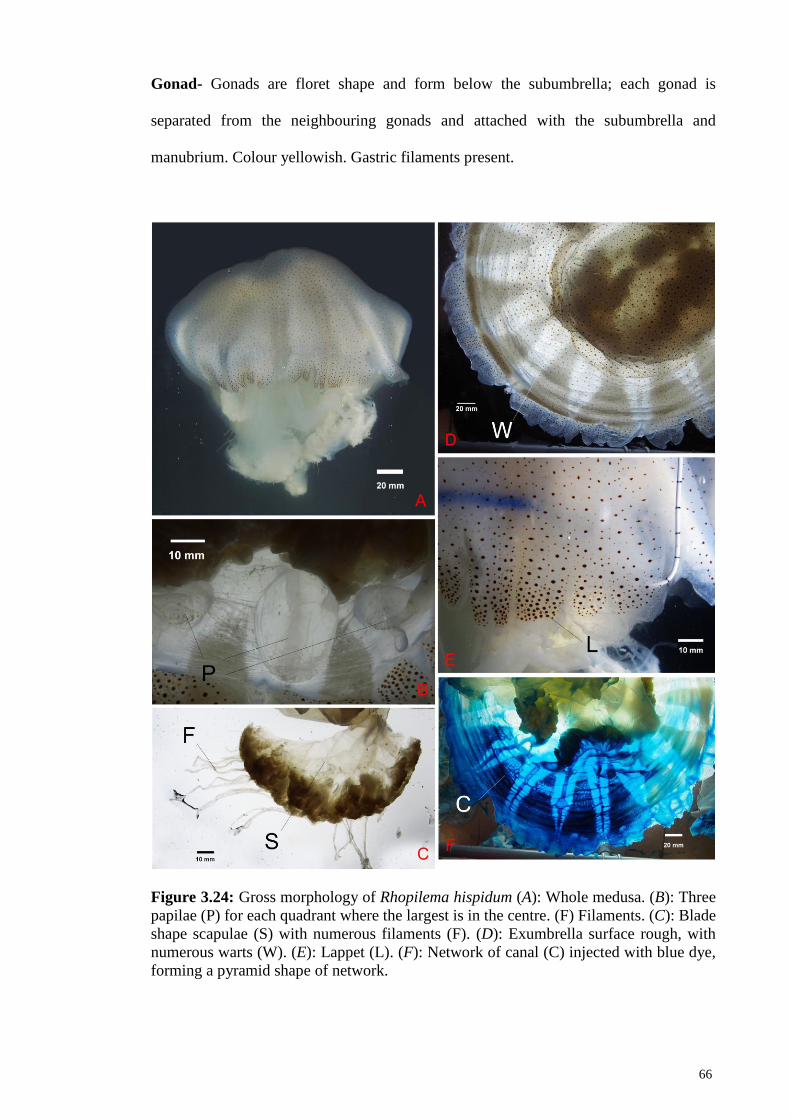
66
Gonad- Gonads are floret shape and form below the subumbrella; each gonad is
separated from the neighbouring gonads and attached with the subumbrella and
manubrium. Colour yellowish. Gastric filaments present.
Figure 3.24: Gross morphology of Rhopilema hispidum (A): Whole medusa. (B): Three
papilae (P) for each quadrant where the largest is in the centre. (F) Filaments. (C): Blade
shape scapulae (S) with numerous filaments (F). (D): Exumbrella surface rough, with
numerous warts (W). (E): Lappet (L). (F): Network of canal (C) injected with blue dye,
forming a pyramid shape of network.

67
3.6 Morphological Description of Lobonemoides robustus Stiasny 1920
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
Lobonematidae (Stiasny, 1921)
Lobonemoides (Light, 1914)
Lobonemoides robustus (Stiasny, 1920) (Figure 3.25 – 3.26)
Material Examined – A total of five specimens were examined from site Sungai
Janggut (IND 6, IND 7, IND 8, SJ 130) and Kampung Cempaka (MRI 204).
Description of specimens - Bell: Umbrella hemispherical. Numerous soft, long conical
papillae on the exumbrella, where size and density of the protuberance decreasing
towards the bell margin. Eight elongated lappets in each octant. Rhopalar lappets are
small and pointy. Short furrow between each lappets. A formation of larger furrow
between two pairs of velar lappet. 16 tear-drop shaped rhopalia present in clefts of the
umbrella margin, located at the interradial and perradial axis. Tentacles absent.
Oral arm – Eight oral arms, as long as the bell diameter; base of oral arm is less than
1/3 as wide as the bell diameter oral arms begin at the end of the oral pillar, then
develop into three wings at the distal section (approximately at the distal 1/5 of the oral
arm). Two wings facing away from the central axis (outer wings) and one wing faces
the central axis (inner wing). Numerous mouthlets located at the frills of the wings.
Numerous filaments present at the frills of the oral arms. Fenestrations, or windows are
found in the oral arm. Scapulae absent. A definite terminal club absent, but four to nine
spindle shape clubs on each oral arm that are easily detached. Numerous pigmented
spots on the clubs. A canal is found in the centre of the club, extending proximately

68
from the base of the club towards the distal end of the club. String like filaments are
found on oral arm.
Stomach and Manubrium – Almost floret shaped gastrovascular cavity, or central
stomach Subgenital fenestration is oval; 1/5 the length of the bell diameter. Three small
and non-prominent dome shaped papillae located at the lower side of the subgenital
fenestration; the papillae in the centre is smaller than the two on each side; the centre
papillae forms a ridge that extends towards the inner subgenital fenestration.
Manubrium thick and smooth; oral disk at the base is about half as wide as the bell
diameter. Thirty two radial canals on the subumbrella comprising of16 rhopalar canals
and 16 inter-rhopalar canals; all canals efface beyond the ring canal, towards fully to the
margin of the bell, where rhopalar canals split into two at the distal end and branch
toward the neighbouring lappets. Canals are found in the lappets. Within the ring canal,
three anastomoses connecting both sides of the rhopalar. Most interrhopalar canals are
without anastomoses except a few exceptions where one anastomos exist. Beyond the
ring canal, rhopalar canals anastomoses with the neighbouring canals through
approximately twelve anastomoses, whereas interophalar canal fuses with the
neighbouring canals completely. Networks are also found in the oral arm, where a main
canal connects with the oral arm from the oral disk. Radial muscles absent. Each
quadrant has one coronal muscle fieldt, comprising 80 to 96 muscle folds. The coronal
muscle folds are interrupted at the sixteen rhopalar radii. Purple spots are found in the
muscle field and the surface of the canals and lappets. Manubrium thick and smooth;
oral disk at the base is about half as wide as the bell diameter, and the base of oral arm
is less than 1/3 as wide as the bell diameter.

69
Gonad - Floret shape and bluish gonads found below the subumbrella; each gonad is
separated with the neighbouring gonad; attaches to the subumbrella and manubrium.
Gonad protruding out of the subgenital fenestration. Gastric filaments present.
Figure 3.25: Gross morphology of Lobonemoides robustus (A): Whole medusa. (F)
Filaments. (B): Gonad (G). (C, D): Conical papillae on the exumbrella in water (P1),
and on dry stage (P2).

70
Figure 3.26: Morphology of Lobonemoides robustus (A): Coronal musculatures (M)
with colouration. Rhopalium (R). (B): Spindle shape club (C). (C): Network of canal
(C) with yellow dye injected. (G) Gonad. (D): Lappet (L) elongated. (E): Oral arm (OA)
with yellow dye injected.

71
3.7 Morphological Description of Versuriga anadyomene Maas 1903
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
Mastiigidae (Stiasny, 1921)
Versuriga
Versuriga anadyomene (Maas, 1903) (Figure 3.27 – 3.28)
Material Examined – A total of two specimens were examined from sites Kilim (MRI
153) and Sungai Janggut (MRI 176).
Description – Bell - Umbrella hemispherical. Exumbrella suface grooved irregularly
where the size and quantity of the grooves decreased from the centre towards the
margin. Eight lappets in each octant; shape are slightly elongated. The lappet
neighbouring rhopalar lappets are slightly bigger than the rest of the velar lappets.
Rhopalar lappets small and pointy. Short furrow between each velar lappets. A larger
furrow formed between two pairs of velar lappet. Sometimes a small lappet formed
between two lappets. Eight rhopalia present in clefts of the umbrella margin, located at
the interradial and perradial axis. Rhopalia shape is tear-drop like. Tentacles absent.
Oral arm - Eight oral arms, as long as the bell diameter; oral arms formed at the end of
the oral pillar, then develop to three wings at the distal section (approximately at the
distal 1/5 of the oral arm). Numerous mouthlets at the frills of the wings. Numerous
appendages are found at the frills of the wings. Fenestrations, or windows are found on
the oral arms. Scapulae absent. Terminal club absent. Some club shape structures are
found at the margin of the oral arms. Filaments are found at the oral disk. Filament
shape string-like.

72
Stomach and Manubrium - Gastrovascular cavity, or centre stomach shape floret.
Subgenital fenestration shape oval; 1/5 the length of the bell diameter. Papillae absent.
Manubrium thick and smooth; oral disk at the base is about half as wide as the bell
diameter, and oral disk at the mouth is 1/3 as wide as the bell diameter. Eight radial
canals on the subumbrella. All eight rhopalar canals extending almost all the way to the
margin of the bell, where canals split at the distal end and branch toward the
neighbouring velar lappets. Ring canal present. Interradial rhopalar canals connect with
the neighbouring canals proximally, whereas perradial rhopalar canals only connected
with the neighbouring canals beyond the ring canal. Network becomes denser and
smaller towards the margin of the bell. Radial muscles absent. Eight coronal muscle
fields, each is interrupted at the eight radii. Coronal musculature starts forming
approximate 1/3 distally from the centre, covering 2/3 of the subumbrella until the
margin.
Gonad - Floret shaped and yellowish gonads found formed below the subumbrella;
each gonad is separated with the neighbouring gonads; attaches with the subumbrella
and manubrium. Gastric filaments present.
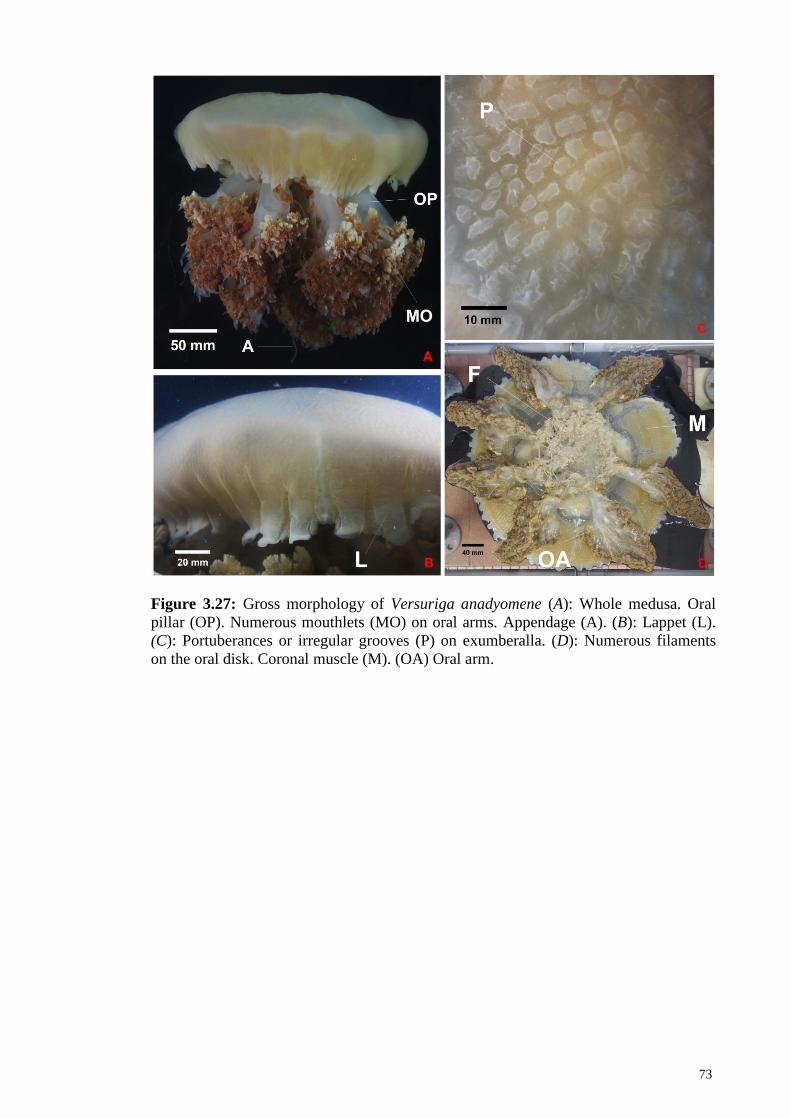
73
Figure 3.27: Gross morphology of Versuriga anadyomene (A): Whole medusa. Oral
pillar (OP). Numerous mouthlets (MO) on oral arms. Appendage (A). (B): Lappet (L).
(C): Portuberances or irregular grooves (P) on exumberalla. (D): Numerous filaments
on the oral disk. Coronal muscle (M). (OA) Oral arm.
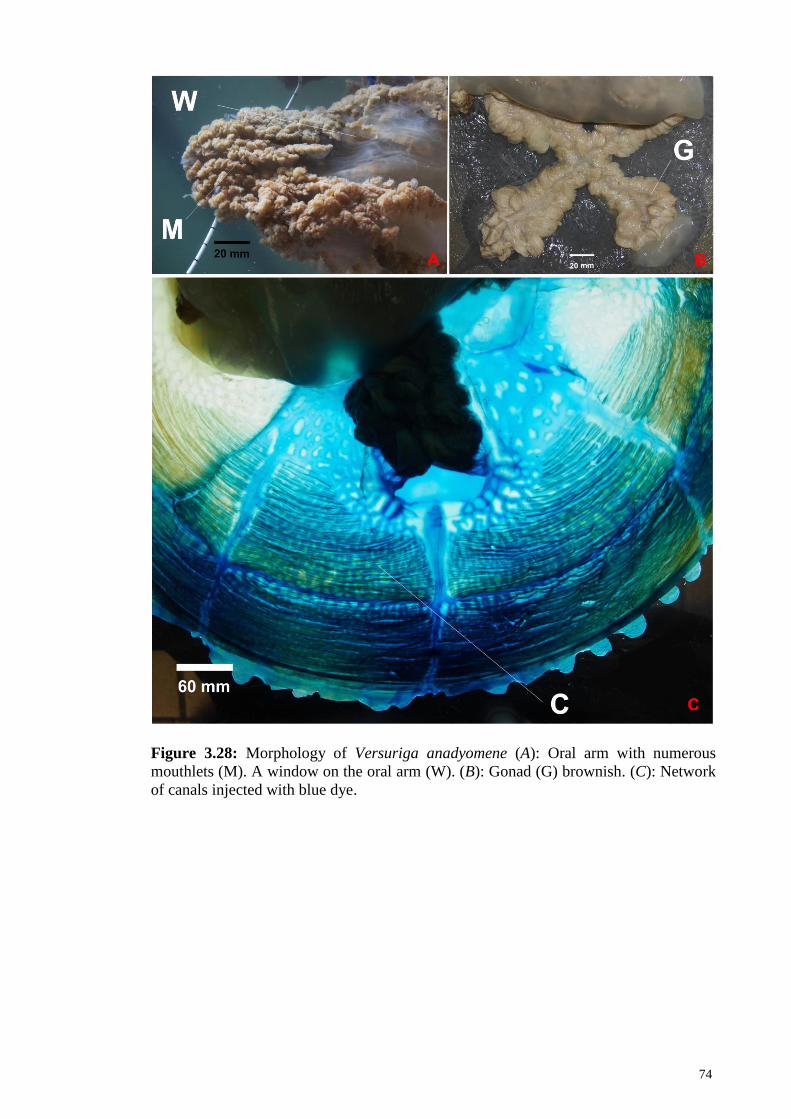
74
Figure 3.28: Morphology of Versuriga anadyomene (A): Oral arm with numerous
mouthlets (M). A window on the oral arm (W). (B): Gonad (G) brownish. (C): Network
of canals injected with blue dye.

75
3.8 Morphological Description of Phyllorhiza punctata von Lendenfeld 1884
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
Mastiigidae (Stiasny, 1921)
Phyllorhiza (Agassiz, 1862)
Phyllorhiza punctata (von Lendenfeld, 1884) (Figure 3.29 – 3.30)
Material Examined – A total of six specimens were examined from sites Sungai
Janggut (IND 2, IND 11, IND 14, MRI 14), Pantai Kok (MRI 131, MRI 139).
Description – Bell - Umbrella hemispherical. Transparent white in the centre of the
bell, gradually brownish orange towards the margin of the bell. Numerous round, white
warts are found evenly distributed on the exumbrella surface. Eight lappets in each
octant; shape are slightly elongated. Rhopalar lappets small and pointy. Short furrow
between each lappets. A larger furrow formed between two pairs of velar lappet. Eight
rhopalia present in clefts of the umbrella margin, located at the interradial and perradial
axis. Rhopalia shape is tear-drop like. Tentacle absent.
Oral arm - Eight oral arms, as long as the bell diameter; oral arms formed at the end of
the oral pillar, then develop to three wings at the distal section (approximately at the
distal 1/3 of the oral arm). Numerous mouthlets at the frills of the wings. Numerous
appendages with bulbs are found at the frills of the wings. Scapulae absent. One
terminal club on each oral arm; half as long as the bell diameter, with a bulb at the end.
Colour light brown / purple. Numerous filaments are found on oral disk.

76
Stomach and manubrium - Gastrovascular cavity, or centre stomach shape floret.
Papillae absent. Eight radial canals on the subumbrella, all eight canals extending all the
way to the margin of the bell, where rhopalar canals split at the distal end and branch
toward the neighbouring lappets. Ring canal present. Interradial rhopalar canals
connected with the neighbouring canals proximally, whereas perradial rhopalar canals
only connected with the neighbouring canals beyond the ring canal. Network becomes
denser and smaller towards the margin of the bell. Radial muscles absent. Eight coronal
muscle fields, each is interrupted at the eight radii. Up to 96 muscle folds per quadrant.
Coronal musculature starts forming approximate 1/3 distally from the centre, covering
2/3 of the subumbrella until the margin. Manubrium thick and smooth; oral disk at the
base is about half as wide as the bell diameter, and oral disk at the mouth is 1/4 as wide
as the bell diameter.
Gonad - Floret shaped and yellowish gonads found formed below the subumbrella;
each gonad is separated with the neighbouring gonads; attaches with the subumbrella.
Gastric filaments present.

77
Figure 3.29: Gross morphology of Phyllorhiza punctata (A): Whole mesusa. Numerous
warts (W) on the exumbrella. Terminal club (TC). (B): Numerous mouthlets (MO) on
the oral arm. (C): Filaments (F) on oral disk. (D): Oral pillar (OP). The white ring is a
wire to suspend the jellyfish and it is not part of the jellyfish structure.

78
Figure 3.30: Morphology of Phyllorhiza punctata (A): Coronal musculatures (M). (R)
Rhopalia (L) Lappet. (B): Warts (W) on exumbrella. (C): Numerous appendages (A) on
the oral arms. Terminal club (C). (D): Gonad (G) brownish. (E): Network of canals (C)
injected with blue dye.

79
3.9 Morphological Description of Lychnorhiza malayensis Stiasny 1920
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
Lychnorhizidae (Haeckel, 1880)
Lychnorhiza (Haeckel, 1880)
Lychnorhiza malayensis (Stiasny, 1920) (Figure 3.31)
First record of species in Malaysia
L. malayensis has a wide distribution in the Malayan Archipelago and the Indian
Ocean), where it was previously found by stiassny in 1920s & 30s in Batavia, Java, by
Menon in 1930 in Madras, India, by Nair in 1951 in Trivandrum, India, but so far have
yet to be documented or found in Malaysia. It has recently been documented in
Andaman Sea and Gulf of Thailand (http://marinegiscenter.dmcr.go.th) and in 2014
sampling by Jeyabaskaran et al (2016) from Thiruvananthapuram to Goa, India.
Material Examined - A total of six specimens were examined from sites Sungai
Janggut (IND 13, MRI 35, MRI 36, MRI 37, MRI 38, MRI 39)
Description – Bell - Umbrella hemispherical. Exumbrella surface smooth. Eight
slightly pointy lappets in each octant. Rhopalar lappets small and pointy. Eight tear-
drop shaped rhopalia present in clefts of the umbrella margin, located at the interradial
and perradial axis. Tentacles absent.
Oral arm - Eight oral arms, as long as the bell diameter; oral arms formed at the end of
the oral pillar, then develop to three wings at the distal section (approximately at the
distal 1/3 of the oral arm). Numerous mouthlets at the frills of the wings. Scapulae
absent. Terminal club absent. Filaments absent.

80
Stomach and manubrium - Stomach shape floret. Subgenital fenestration shape oval;
1/10 the length of the bell diameter. Five papillaes around each subgenital fenestration.
Three papillae located at the upper side of the margin subgenital fenestration, aligning
with the rhopalar canal. One smaller on each side of the subgenital fenestration. Papillae
surface decorated with fine dots, but smooth. Sixteen radial canals on the subumbrella.
Eight rhopalar canals extending fully to the margin of the bell. Eight inter-rhopalar
canals (perradial rhopalar canals) do not extend beyond the ring canal. Ring canal
present. Between four to five centripetal canals within the ring canal blindly ended
(except a few are not blindly ended) and do not anastomoses with the neighbouring
rhopalar nor interrhopalar canals. Canals becomes denser and smaller towards the
margin of the bell. Radial muscles absent. One coronal muscle fields not interrupted at
the eight radii. Between 80 to 96 muscle folds per quadrant. Coronal musculature starts
forming approximate 1/3 distally from the centre, covering 2/3 of the subumbrella till
the margin. Manubrium thick and smooth; oral disk at the base is about half as wide as
the bell diameter, and oral disk at the mouth is 1/4 as wide as the bell diameter.
Gonad – Gonads are floret shaped gonads and form below the subumbrella; each gonad
is separated from the neighbouring gonads; attaches with the subumbrella and
manubrium. Colour yellowish. Gastric filaments present. Gonad is completely protected
within the fenestration with outfold.

81
Figure 3.31: Gross morphology of Lychnorhiza malayensis (A) Whole medusa. (OA)
Oral arm. (B) Rhopalium (R). Lappet (L). (C) Network of canals (C) injected with red
dye. (D) Gonad (G) creamy white. (P) Papillae. (E) Oral arm (OA). (F) Three papillae
(P).

82
3.10 Morphological Description of Acromitus flagellatus Maas 1903
Class SCYPHOZOA (Goette, 1887)
Order RHIZOSTOMEAE (Agassiz, 1862)
Catostylidae (Gegenbaur, 1857)
Acromitus (Light, 1914)
Acromitus flagellatus (Maas, 1903) (Figure 3.32)
Material Examined – A total of three specimens were examined from sites Sungai
Buluh (MRI 21, MRI 23, MRI 24).
Description – Bell - Umbrella hemispherical. Exumbrella surface smooth. Numerous
red dots are distributed on the exumbrella. Eight lappets in each octant; shape are
slightly elongated. Rhopalar lappets small and pointy. Short furrow between each
lappets. A larger furrow formed between two pairs of velar lappet. Eight rhopalia
present in clefts of the umbrella margin, located at the interradial and perradial axis.
Rhopalia shape is tear-drop like. Tentacle absent.
Oral arm - Eight oral arms, 2/3 as long as the bell diameter; oral arms formed at the
end of the oral pillar, then develop to three wings at the distal section (approximately at
the distal 2/3 of the oral arm). Numerous mouthlets at the frills of the wings. Numerous
filaments at the frills of the wing. Scapulae Absent. Terminal club absent, but a single
thread-like filament is formed at the distal end of each oral arm. Numerous string-like
filaments on the oral arms.

83
Stomach and manubrium - Gastrovascular cavity, or centre stomach shape floret.
Subgenital fenestration shape oval; 1/7 the length of the bell diameter. A triangular
shape papillae is found at the centre of the subgenital fenestration. Papillae surface
smooth. 16 radial canals on the subumbrella. Eight rhopalar canals extending fully to
the margin of the bell. Eight inter-rhopalar canals do not extend beyond the ring canal.
Ring canal present. Between three to five centripetal canals within the ring canal
anastomoses with each other, and anastomoses with the neighbouring rhopalar canals,
but not with the inter-rhopalar canals. Canals becomes denser and smaller towards the
margin of the bell. Radial muscles absent. Eight coronal muscle fields covering the
network mesh underneath, each interrupted by the four interradial canals and the four
perradial canals. Coronal musculature starts forming 1/3 distally from the centre,
covering 2/3 of the subumbrella up to the margin. Muscle fields become denser beyond
the ring canal. Manubrium thick and smooth; oral disk at the base is about half as wide
as the bell diameter, and oral disk at the mouth is 1/4 as wide as the bell diameter.
Gonad – Gonads are floret shaped gonads and form below the subumbrella; each gonad
is separated from the neighbouring gonads; attaches with the subumbrella and
manubrium. Colour yellowish. Gastric filaments present.
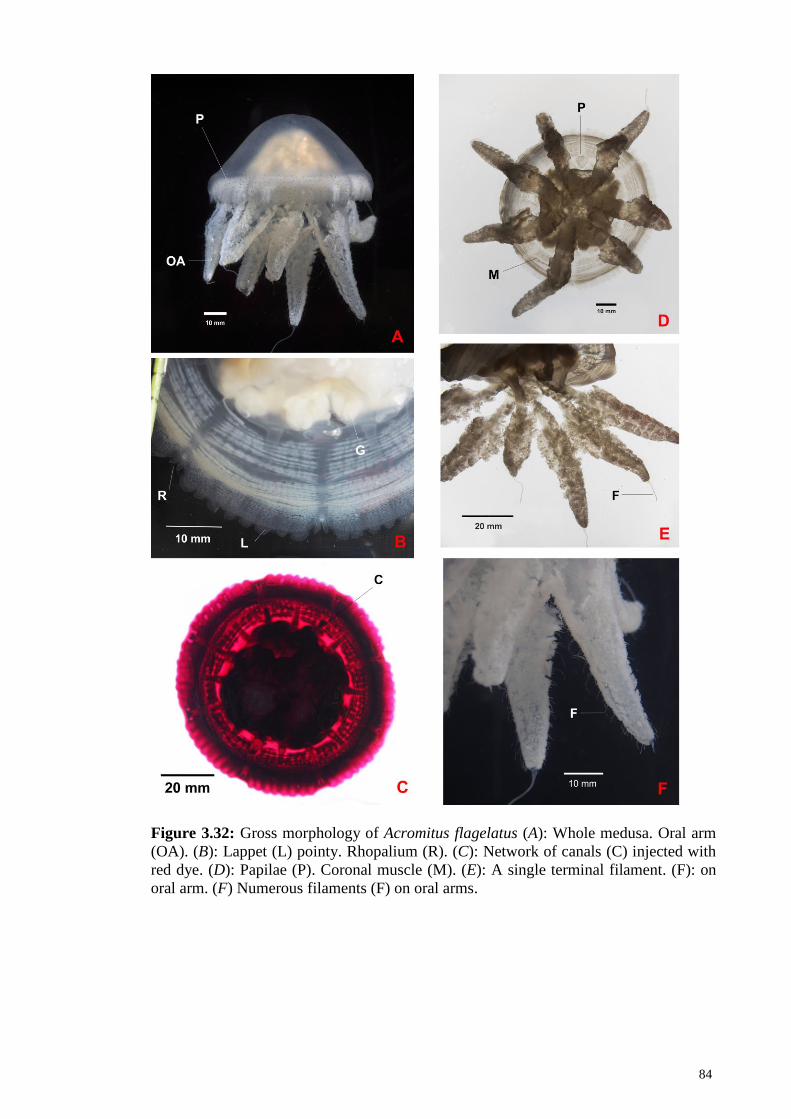
84
Figure 3.32: Gross morphology of Acromitus flagelatus (A): Whole medusa. Oral arm
(OA). (B): Lappet (L) pointy. Rhopalium (R). (C): Network of canals (C) injected with
red dye. (D): Papilae (P). Coronal muscle (M). (E): A single terminal filament. (F): on
oral arm. (F) Numerous filaments (F) on oral arms.

85
3.11 Geometric Morphometric Analysis
A total of 107 specimens of Chrysaora chinensis were used in this study. 16 are
from the East-Central of Peninsular Malaysia, 26 from East-North, 27 from West-
Central and 38 from West-North. Among them, 17 specimens were from the museum
(Appendix D, Figure 3.33).
Figure 3.33 Map of the sampling locations for geometric morphometric analysis, with
the number of specimens obtained. Green circle represents East-North (EN), red circle
represent East-Centre (EC), blue represents West-Central (WC), and purple represents
West-North (WN) coastal areas of Peninsular Malaysia.
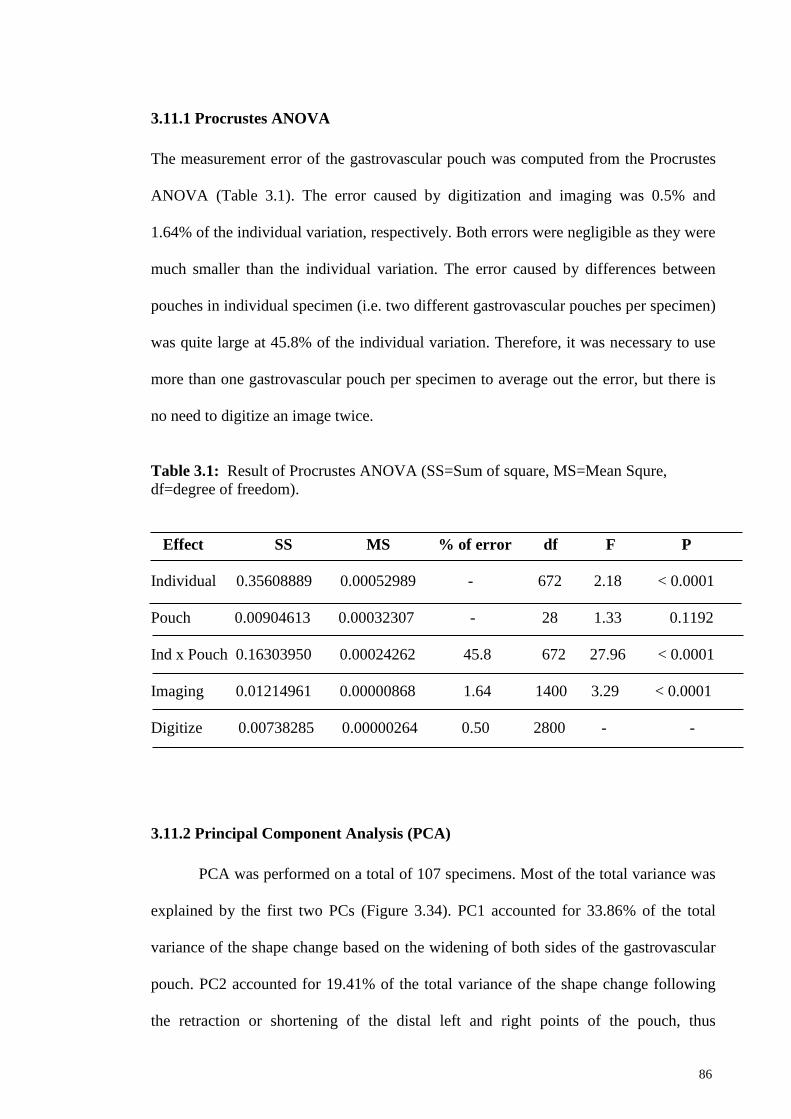
86
3.11.1 Procrustes ANOVA
The measurement error of the gastrovascular pouch was computed from the Procrustes
ANOVA (Table 3.1). The error caused by digitization and imaging was 0.5% and
1.64% of the individual variation, respectively. Both errors were negligible as they were
much smaller than the individual variation. The error caused by differences between
pouches in individual specimen (i.e. two different gastrovascular pouches per specimen)
was quite large at 45.8% of the individual variation. Therefore, it was necessary to use
more than one gastrovascular pouch per specimen to average out the error, but there is
no need to digitize an image twice.
Table 3.1: Result of Procrustes ANOVA (SS=Sum of square, MS=Mean Squre,
df=degree of freedom).
Effect SS MS % of error df F P
Individual 0.35608889 0.00052989 - 672 2.18 < 0.0001
Pouch 0.00904613 0.00032307 - 28 1.33 0.1192
Ind x Pouch 0.16303950 0.00024262 45.8 672 27.96 < 0.0001
Imaging 0.01214961 0.00000868 1.64 1400 3.29 < 0.0001
Digitize 0.00738285 0.00000264 0.50 2800 - -
3.11.2 Principal Component Analysis (PCA)
PCA was performed on a total of 107 specimens. Most of the total variance was
explained by the first two PCs (Figure 3.34). PC1 accounted for 33.86% of the total
variance of the shape change based on the widening of both sides of the gastrovascular
pouch. PC2 accounted for 19.41% of the total variance of the shape change following
the retraction or shortening of the distal left and right points of the pouch, thus
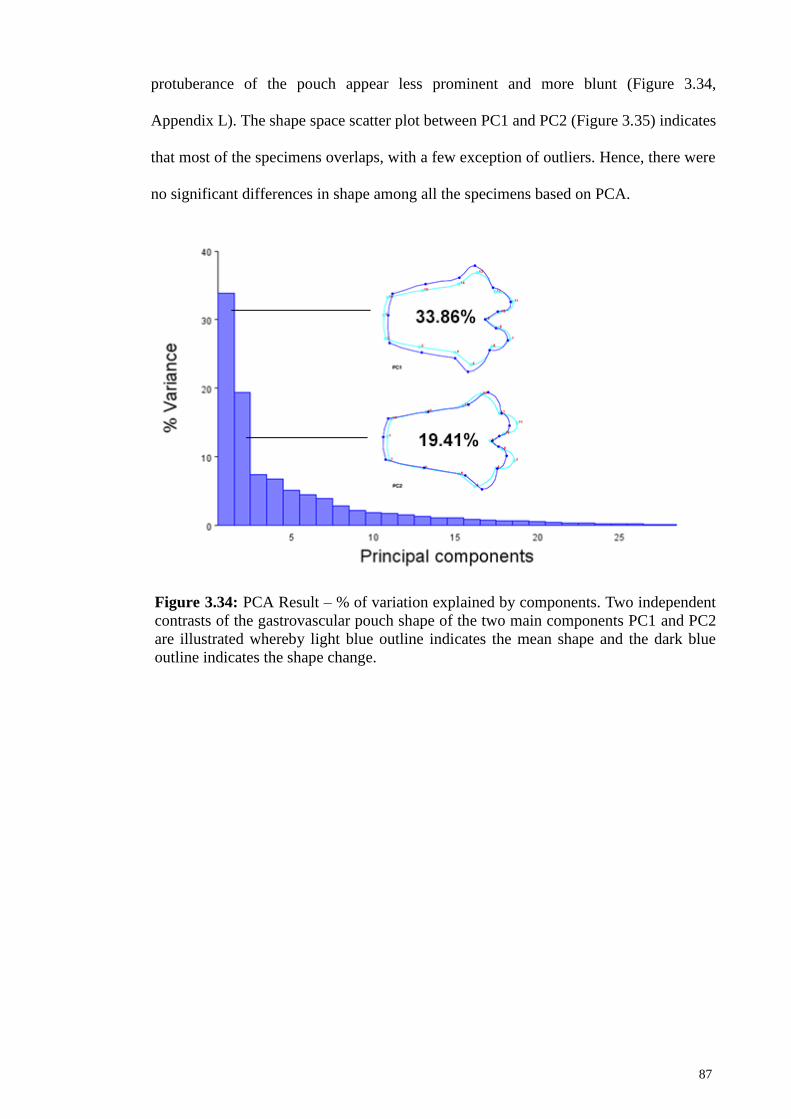
87
protuberance of the pouch appear less prominent and more blunt (Figure 3.34,
Appendix L). The shape space scatter plot between PC1 and PC2 (Figure 3.35) indicates
that most of the specimens overlaps, with a few exception of outliers. Hence, there were
no significant differences in shape among all the specimens based on PCA.
Figure 3.34: PCA Result – % of variation explained by components. Two independent
contrasts of the gastrovascular pouch shape of the two main components PC1 and PC2
are illustrated whereby light blue outline indicates the mean shape and the dark blue
outline indicates the shape change.

88
Figure 3.35: Scatter plots of PC1 vs PC 2. PC1 accounted for 33.86% and PC2
accounted for 19.41% of the total variance of the shape change of gastrovascular pouch
of specimens from East-Central (EC), East-North (EN), West-Central (WC) and West-
North (WN) of Peninsular Malaysia.
3.11.3 Canonical Variate Analysis (CVA)
CVA was performed on pouches of C.chinensis populations from four coastal
areas of the Peninsular Malaysia: East-Central (EC), East-North (EN), West-Central
(WC) and West-North (WN), with a total of 107 specimens. CV1 accounts for 47.46%,
CV2 accounts for 32.72%, and CV3 accounts for 19.83% of the amount of relative
between-group variation (Figure 3.36). Specimens from East-Central and East-North are
separated from the other two groups (West-Central and West-North). There are
considerable amount of overlapping among the group West-Central and West-North

89
(Figure 3.30). The magnitude of Mahalanobis distances is lower between West-Central
and West-North but higher between east and west coast areas, whereas East-Central and
West-North has the highest Mahalabonis distance. (Table 3.2).
The outline drawing of the visualization of the gastrovascular pouch shape of
C.chinensis populations from the four coastal areas of Peninsular Malaysia show some
variations in shape (Figure 3.37). Pairwise comparisons among the four areas show that:
EC vs WC: Protuberance at the margin of WC is more blunt
EC vs WN: Protuberance at the margin of WN is more blunt, whereas the EC’s
pouch is slightly enlarged
EC vs EN: EC’s pouch is slightly enlarged
WC vs EN: Protuberance at the margin of WC is more blunt
WC vs WN: Protuberance at the margin of WC is more blunt
WN vs EN: Protuberance at the margin of WN is more blunt

90
Fig. 3.36: Independent contrast of component using canonical variate analysis of the
gastrovascular pouch of specimens from East-Central (EC), East-North (EN), West-
Central (WC) and West-North (WN) of Peninsular Malaysia, whereby CV1, CV2 and
CV3 accounts for 47.46%, 32.72% and 19.83% of the amount of relative between-group
variation, respectively. Light blue outline indicates the mean shape and the dark blue
outline indicates the shape change. CV1 denotes changes with blunt protuberance at the
margin and enlarging of the distal end, CV2 denotes changes with blunt protuberance at
the margin and CV3 denotes changes with blunt protuberance at the margin and
enlarging of the distal end.

91
Figure 3.37: Scatter plot of CV1 vs CV2 and illustration of mean shape of
gastrovascular pouch of specimens from four coastal areas East-Central (EC), East-
North (EN), West-Central (WC) and West-North (WN) of Peninsular Malaysia. Plot
shows shape differences mainly between specimens of the east and west coasts, even
between those from EN and EC, but with no distinct differences between those from
WN and WC.
Table 3.2: Mahalanobis distances among the pouch shape of C. chinensis populations
from four coastal areas designated as West-North (WN), West-Central (WC), East-
North (EN) and East Central (EC) of Peninsular Malaysia.
EC EN WC
EN 4.3296 - -
WC 4.2151 3.4014 -
WN 4.6090 3.4034 2.6485

92
Figure 3.38: Pairwise comparisons between the pouch shape of C.chinensis populations
from East-Central (EC), East-North (EN), West-Central (WC) and West-North (WN) of
Peninsular Malaysia

93
CHAPTER 4: DISCUSSION
4.1 Morphology, Diversity and Importance of Jellyfish Species of Peninsular
Malaysia
Although the demand for jellyfish has increased, the catch had actually
decreased. Annual catch in 2000 was 179,086 metric tons (wet weight) from
Philippines, Thailand, Indonesia, Myanmar and Malaysia, and dropped drastically to
only 13,402 metric tons in 2004 (Kitamura & Omori, 2010). Personal communication
with local fishermen indicate that the supply of edible jellyfish is not enough to meet the
high demand. Despite its importance, jellyfish study in Malaysia is still relatively
scarce. Light (1914) on board the steamer Albatross to the Philippines and recorded
several jellyfish species he found during the trip (including Chrysaora, Lychnorhiza and
Acromitus species) but unfortunately he did not make his stop in Pensinsular Malaysia.
This study is aimed to try to fill in the gaps from previous studied. This studies not only
report a new record of Lychnorhiza malayensis in Malaysia, but also provided detail
description and photographs of eight scyphozoan jellyfish species previously reported
but without detail information.
Ever since Linnaeus first described the popular moon jellyfish Aurelia aurita in
1758, jellyfish taxonomy has long been subjected to disagreements and revision. Two
particular jellyfish that subject to such scrutiny are Chrysaora sp. and Cyanea capillata.
Since the description of the Genus Chrysaora in 1810 by Peron & Leseur, a number of
revisions had been done by various researchers. Eschscholtz (1829) listed six species in
the genus, Lesson (1843) listed 13, Agassiz (1862) listed nine, Haeckel (1880) listed 10,
Mayer (1910) listed 15, Kramp (1961) listed 11, and most recently Morandini &
Marques (2010) listed 15.

94
C. capillata poses another problem to the taxonomy position. It has been revised
by Linnaeus et al (1758), Mayer (1910), Kramp (1961), Dawson (2005), Sparmann
(2012) and Kolbasova et al. (2015). It was believed to be a cosmopolitan species but
recent research by Dawson (2005) has shown that it is in fact not. Thus it will be
beneficial to study jellyfish species from this region to ascertain its taxonomy position.
4.2 Lion’s Mane Jellyfish (Cyanea sp.) of Malaysia
The Cyanea species obtained in the west coast of Peninsular Malaysia were without
gastrovascular pit, and the exumbrella surface is lightly granulated with warts.
Gastrovascular pit is a prominent feature of Cyanea capillata, thus the Malaysian
specimen cannot be ascribed to be C. capillata. By injecting colour dye to the rhopalar
and tentacular pouches of Cyanea species obtained in Peninsular Malaysia show up to
five transverse “ridges” connecting both pouches, and the peripheral canals at the lappet
contains numerous networks of anastomoses, forming a dense network, which
according to Kramp (1961) is one of the characteristic of C. nozakii. But according to
Kramp (1961), the colour of C. nozakii is white, but the species obtained are brownish
and yellowish. Thus, the Cyanea species obtained in Peninsular Malaysia cannot be
ascribed to C. nozakii. C. lamarki does not seem to fit the characteristic of Cyanea
species obtained in Peninsular Malaysia because the locality of C. lamarki was reported
to be in Europe. Furthermore, its rhopalar and tentacular stomach pouches are
completely separated.
Another Cyanea species, C. barkeri from Australia (Gershwin et al., 2010) is
similar to the Cyanea species from Peninsular Malaysia in that they both lack
gastrovascular pit. But it is not clear whether C. barkeri’s rhopalar and tentacular
pouches are connected, and whether the exumbrella surface is smooth or granulated.

95
Kramp (1961) recorded C. buitendijki in Malay Archipelago but he did not describe the
exumbrella surface nor the existence of musculature intrusion. Furthermore, Malay
Archipelago encompasses a wide region ranging from Malaysia, Singapore, Indonesia
to Philippines. Thus cannot ascribe the Cyanea species from Peninsular Malaysia to
C.barkeri nor C. buitendijki.
Molecular phylogenetic evidence of Rizman-Idid et al. (2016) showed that
Malaysian Cyanea did not cluster with either C. nozakii, C. capillatta, C. rosacea nor C.
annaskala and it is very distinct from any available Cyanea sequences. Hence, with
molecular and morphological evidence it is plausible that Malaysian Cyanea is a new
species.
Table 4.1: Comparison of Cyanea sp. from Peninsular Malaysia with other Cyanea
species based on Kramp (1961).
C. capillata C. buitendijki C. lamarki C. nozakii Cyanea sp.
Musculature
intrusion
Y ? N ? N
Exumbrella
papillose
N
?
Y
N
Y
Locality
Australia,
NW Pacific,
China,
Mutsu Bay,
Japan,
Trivandrum
Coast India
Malay
Archipelago
Bay of
Biscay,
Norway,
Holland, W.
Coast of
Sweden,
Denmark,
Iceland
Japan, NW
Pacific,
Indochina,
Indian
Ocean
Malaysia
Size
up to
1000mm
up to
310mm
up to
1000mm
up to
260mm
up to
250mm
96
Table 4.1, continue
Pouch
Rhopalar and
tentacular
stomach
pouches
completely
separated
Rhopalar and
tentacular
stomach
pouches
connected by
several broad
transversed
anastomoses
Rhopalar
and
tentacular
stomach
pouches
completely
separated
Rhopalar
and
tentacular
stomach
pouches
connected by
several
broad
transversed
anastomoses
Rhopalar and
tentacular
stomach
pouches
connected by
several broad
transversed
anastomoses
Canal
Peripheral
canals more
or less
curved
Peripheral
canals without
anastomoses
Peripheral
canals more
or less
curved
Peripheral
canals with
numerous
anastomoses,
forming a
dense
network
Peripheral
canals with
numerous
anastomoses,
forming a
dense network
Colour
Reddish
brown or
yellowish
Blue
Milky white
Brownish,
yellowish
4.3 Sea Nettles (C. Chinensis) of Peninsular Malaysia
The detailed morphological description of C. chinensis in the present study represents
the first of its kind in Malaysia. C. chinensis is commonly found in most beaches in east
and west coast of Peninsular Malaysia (Rizman-Idid et al., 2016). Base on personal
observation and interviews with fishermen, they occur more abundantly at the polluted
areas such as Balik Pulau, Pantai Kok and Kilim, particularly during the dry season
(April to October). Previous study reported the occurrence of C. chinensis in South
China Sea, (Vanhöffen, 1911), but Kramp (1961) reported C. chinensis as C. helvola.
Prior to the resurrection of C. chinensis by Morandini & Marques (2010), many
Chrysaora specimens found in the South China Sea and Malay Archipelago have been
identified as either C. melanaster (Yap & Ong, 2012) or C. helvola (Kramp, 1961).

97
However, Morandini and Marques (2010) emphasized that C.chinensis is very different
from C. helvola (valid name C. fuscescens) and C. melanaster due to the difference in
tentacle and lappet number. Both the latter species are larger than in their observed C.
chinensis specimens. Recent study by Yap & Ong (2012) ascribed the specimens from
St. John’s Island in Singapore as C. chinensis. Likewise, findings from the present study
which focused on specimens obtained from the east and west coasts of Peninsular
Malaysia confirms that the sea nettles found in these waters were morphologically
identified as C. chinensis. Furthermore, DNA sequences from some of the specimens
obtained in the present study (data not shown) revealed that they are C. chinensis and
genetically similar and phylogenetically clustered to those reported by Rizman-Idid et
al., (2016). It is noteworthy that the colouration of C. chinensis varies greatly. Some
specimens are transparent whereas some are reddish with radiating stripes on the
exumbrella, with numerous pigmentations on oral arms and exumbrella, and reddish
brown lappets. Nonetheless, this colouration corroborated with the colouration
diagnosis of C. chinensis by Yap & Ong (2012) and Morandini & Marques (2010).
The shape of the gastrovascular pouch of specimens from the east coast is
slightly elongated compared to those from the west coast. Unlike Rhizostomeae
jellyfish, C. chinensis do not have well developed canal. The gastrovascular pouches of
C. chinensis are used to store and transport nutrients from the central stomach to various
parts of the jellyfish. The east coast of Peninsular Malaysia is facing the South China
Sea, whereas the west coast is facing the Straits of Malacca, one of the busiest sea
routes in the world, hosting annually about 65,000 vessels for international navigation,
and up to another additional 15,000 fishing vessel (Ibrahim & Khalid, 2007). The low
water quality of Straits of Malacca is due to pollution from shipping industries,
aquaculture, densely populated coastal areas with nutrients and effluent discharged into
the straits, causing eutrophication (Rezai et al., 2003; Praveena et al., 2011). According

98
to Bong & Lee (2008), the effects of development at nearshore increases the total
suspended solids and decreases the dissolved oxygen. Human activities that lead to
eutrophication have been associated with the bloom of the jellyfish (Arai 2001; Parsons
& Lalli, 2002; Malej et al., 2007; Richardson et al., 2009), and jellyfish appear to be
able to adapt to fluctuations of various levels of water quality and environmental
parameters, such as salinity, temperature, food source (Purcell et al., 2012). A few
geographical variation studies have shown that morphology affects the jellyfish
distribution, pulse rate and swimming speed (Dawson & Hamner, 2003).
Since the main function of the gastrovascular system is for the circulation of
nutrients (Arai, 1997), these differences in gastrovascular pouch shapes observed in the
present study may be a form of morphological adaptation of C. chinensis populations to
the different coastal areas (East-Central, East-North, West-Central, and West-North) of
Peninsular Malaysia. The landmass of Peninsular Malaysia acts as the main physical
barrier which separates the east from the west populations. The EC population also
appears distinct from the EN population. Although the distance between the sampling
areas of both populations is only approximately 350km, which may not be of significant
magnitude to explain the isolation by distance for these populations. Nonetheless, these
populations may have been kept separated due to occurrence of eddies and different
water circulations at 5°N off the east coast of Peninsular Malaysia (Daryabor et al.,
2016). According to Daryabor et al (2016), such water circulations may influence the
distribution of the nutrient balance in regulating primary productivity and the changes in
the marine ecosystem.

99
4.4 Potential Application of GMM in Jellyfish Studies
Although geometric morphometric analysis have been proven useful in analysing and
detecting even very minute shape variations in various organisms (Klingenberg, 2013,
Li et al., 2016), its application as demonstrated in the present study on gelatinous
organism such as jellyfish, may still be informative albeit a few practical challenges.
Geometric morphometric analysis seems more suited for organisms with rigid
structures, as landmarks are more easily identified and configured, thus detection of
shape variation would be attributed due to real morphological differences rather than
inconsistencies of measurements due to mishandling of specimens. Up until now, there
has been no shape variation study using geometric morphometric analysis on soft or
gelatinous organisms. Jellyfish, being gelatinous, can be quite easily distorted, either
from bad sampling methods that can cause damage to their delicate structures, or even
from prolonged preservation of specimens. In fact, formalin can contribute to the
change of shape of jellyfish, especially after a long period of time (Kapiris et al., 1997).
Therefore, using geometric morphometric analysis to detect shape variation in jellyfish
poses a higher degree of difficulties compared to rigid organisms, since even slight false
variation in the shape due to sampling or handling error can lead to the conclusion as
real variation.
The practical usage may be improved by choosing a different structure for
landmark configurations, depending on the species. For example, the present study has
successfully demonstrated that internal structure such as the gastrovascular pouch is
suitable as the result appears to be robust with low measurement errors. On the other
hand, certain structures make poor landmarks. Filaments and tentacles are easily
detached and damaged. They tend to move rather easily and it is difficult to set its

100
orientation and identify landmarks. Other jellyfish species, such as Rhopilema
esculentum of the Rhizostomeae family which is bigger in size and with more rigid
structures, may be a better target for landmark configuration and geometric
morphometric study.

101
CHAPTER 5: CONCLUSION
Morphological studies are crucial for species identification of jellyfish. Even if
molecular techniques are available, identification is firstly based on morphology.
Currently there is a revival in the taxonomy of jellyfish, with many revisions and efforts
to obtain samples world wide for comparisons. Nonetheless, there is still a lack of
jellyfish taxonomist worldwide. Hence studies like this would be helpful in providing
baseline information, including detailed morphology characterization and photographs
that could aid identification in the field for biologist and marine enthusiasts.
Below are the major findings of this study:
1) Morphology of nine jellyfish species (from eight families and eight genera)
found in Peninsular Malaysia that belong to the class Scyphozoa, namely
Chrysaora chinensis, Cyanea sp., Versuriga anadyomene, Rhopilema hispidum,
Rhopilema esculentum, Phyllorhiza punctata, Acromitus flagellatus,
Lobonemoides robustus and Lychnorhiza malayensis were characterised in
details
2) Cyanea sp. found in the Malaysia water may be new species as it could not be
morphologically ascribed to other known Cyanea species
3) A new record of Lychnorhiza malayensis in Malaysia was reported
4) The Malaysian sea nettle jellyfish was verified as C. chinensis
5) GMM analysis of the gastrovascular pouch of specimens of C. chinensis
indicates shape variation between the four coastal areas of Peninsular Malaysia,

102
especially with higher magnitudes of Mahalanobis distances between east and
west coast areas, East-Central and East-North comparisons, but lower
differences between the West-Central and West-North areas.

103
REFERENCES
Adams, D. C., Rohlf, F. J., & Slice, D. E. (2004). Geometric morphometrics: ten years
of progress following the ‘revolution’. Italian Journal of Zoolology 71: 5-16
Adams, D. C., Rohlf, F. J., & Slice, D. E. (2013). A field comes of age: geometric
morphometrics in the 21st century. Italian Journal of Mammalogy
Agassiz, L. (1862) Contributions to the natural history of the United States of America.
IV. Second monograph, In five parts, Acalephs in general, Ctenophorae,
Discophorae, Hydroidae, homologies of the Radiata. Little, Brown & Co.,
Boston, 380 pp.
Alison, E. H., Perry, A. L., Badjeck, M. C., Adger, W. N., Brown, K., Conway, D.,
Halls, A. S., Pilling, G. M., Reynolds, J. D., Andrew, N. L. & Dulvy, N. K.
(2009). Vulnerability of national economies to the impacts of climate change on
fisheries. Blackwell Publishing Ltd
Arnqvist, G. & Martensson, T. (1998). Measurement error in geometric morphometrics:
empirical strategies to assess and reduce its impact on measures of shape. Acta
Zoologica Academiae Scientiarum Hungaricae 44: 73–96.
Arai, M. N. (1997). A functional biology of Scyphozoa. London: Chapman & Hall,
(316 p)
Arai, M. N. (2001). Pelagic coelenterates and eutrophication: a review. Hydrobiologia.,
451, 69–87.
Attrill, M. J., Wright, J. & Edwards, M. (2007). Climate-related increases in jelly sh
frequency suggest a more gelatinous future for the North Sea. Limnology and
Oceanography 52:480-485.
Azila, A. & Chong, V. C. (2010). Multispecies Impingement in a Tropical Power Plant,
Straits of Malacca. Marine Environmental Research doi:
10.1016/j.marenvres.2010.02.004
Azila, N. A & Othman, I. (1993). Cases of jellyfish envenomization in Malaysia and
some cytotoxic studies on their toxin. In: Tan NH, Oo SL, Thambyrajah V, Azila
NA (eds) Advances in Venom and Toxin Research. Proceedings of the Third Asia
Pacific Congress on Animal, Plant and Microbial Toxins Malaysia. 27 June- 1
July, 1993. Kuala Lumpur. Malaysian Society on Toxinology, pp. 113-118.
Bakun, A. & Weeks, S. J. (2006). Adverse feedback sequences in exploited marine
systems: are deliberate interruptive actions warranted? Fish Fish 7:316-333.
Bayha, K. M., Dawson, M. N., Collins, A. G., Barbeitos, M. S. & Haddock, S.H.
(2010). Evolutionary relationships among scyphozoan jellyfish families based
on complete taxon sampling and phylogenetic analyses of 18S and 28S
ribosomal DNA. Integrative and comparative biology, 50 (3), 436–455.

104
Boero, F., Bouillon, J., Gravili, C., Miglietta, M. P., Parsons, T. & Piraino, S. (2008).
Gelatinous plankton: irregularities rule the world (sometimes). Marine Ecology
Progress Series 356: 299–310.
Bolton, T. F. & Graham, W. M. (2004). Morphological variation among populations of
an invasive jellyfish. Marine Ecolology Progress Series 278:125-139.
Bong, C. W. & Lee, C. W. (2008). Nearshore and Offshore Comparison of Marine
Water Quality Variables Measured During SESMA 1. Malaysian Journal of
Science 27 (3) : 25–31
Bookstein, F. L. (1997). Landmark methods for forms without landmarks:
morphometrics of group differences in outline shape. Medical Image Analysis
1:97-118.
Bookstein, F. L. (2015). The Relation Between Geometric Morphometrics and
Functional Morphology, as Explored by Procrustes Interpretation of Individual
Shape Measures Pertinent to Function. The Anatomical Record 298:314–327
Brodeur, R. D., Decker, M. B., Ciannelli, L., Purcell, J. E., Bond, N. A., Stabeno, P.
J., Acun, E. & Hunt, G. L. (2008). Rise and fall of jellyfish in the eastern
Bering Sea in relation to climate regime shifts. Progress in Oceanography 77:
103–111.
Brotz, L., Cheung, W. W. L., Kristin, K., Pakhomov, E. & Pauly, D. (2012). Increasing
jellyfish populations: trends in Large Marine Ecosystems. Hydrobiologia 690:3–
20
Brusca, R. C. & Brusca, G. J. (2002). Invertebrates. Sinauer Associates, Sunderland,
MA.
Cartwright, P, Halgedahl, S. L., Hendricks, J. R., Jarrard, R. D., Marques, A. C. et al.
(2007). Exceptionally Preserved Jellyfishes from the Middle Cambrian. PLoS
ONE 2(10): e1121
Ceh, J., Gonzalez, J., Pacheco, A. S. & Riascos, J. M. (2015). Elusive life cycle of
scyphozoan jelly sh – metagenesis revisited. Scientific. Report. 5, 12037
Chen, X., Fang, Z., Su, H., Lu, H., Liu, B. & Li, J. (2013). Review and application of
geometric morphometrics in aquatic animals Journal of Fisheries of China
Costello, J. H. & Colin, S. P. (1995). Flow and feeding by swimming scyphomedusae.
Marine Biology 124:399-406
Daly, M., Brugler, M., Cartwright, P., Collins, A. G., Dawson, M. N., Fautin, D. G.,
France, S. C., McFadden, C. S., Opresoko, D. M., Rodriguez, E., Romano, S. L.,
& Stake, J. L. (2007). The phylum Cnidaria: A review of phylogenetic patterns
and diversity 300 years after Linnaeus* Zootaxa 1668: 127–182
Daryabor, F., Ooi, S. H., Samah, A. A. & Akbari, A. (2016). Dynamics of the Water
Circulations in the Southern South China Sea and Its Seasonal Transports. PLoS
ONE 11(7): e0158415

105
Daryanabard, R. & Dawson, M. N. (2008). Jellyfish blooms: Crambionella orsini
(Scyphozoa: Rhizostomeae) in the Gulf of Oman, Iran, 2002–2003 Journal of
the Marine Biological Association of the United Kingdom 88(3), 477–483.
Daud, A. (1998). A study on Jelly sh in Sarawak waters. Fisheries Research Institute
Newsletter. Jabatan Perikanan Malaysia 4(1):4 pp.
Dawson, M. N. & Hammer, W. M. (2003). Geographic variation and behavioral
evolution in marine plankton: the case of Mastigias (Scyphozoa, Rhizostomeae)
Marine Biology 143: 1161–1174 doi 10.1007/s00227-003-1155-z
Dawson, M. N. (2004). Some implications of molecular phylogenetics for
understanding biodiversity in jelly shes, with an emphasis on Scyphozoa.
Hydrobiologia 530/531:249-260.
Dawson, M. N. (2005). Morphological variation and systematics in the Scyphozoa:
Mastigias (Rhizostomae, Mastigiidae)-a golden unstandard? Hydrobiologia
537:185-206.
Dawson, M. N. & Hamner, W. M. (2009). A character-based analysis of the evolution
of jellyfish blooms: adaptation and exaptation. Hydrobiologia 616:193–215
Dong, J., Sun, M., Wang, B., & Liu H. (2008). Comparison of life cycles and
morphology of Cyanea nozakii and other scyphozoans. Plankton Benthos
Research 3 (Suppl.) : 118–124, 2008
Dong, Z., Liu, D., & Keesing, J. K. (2010). Jellyfish blooms in China: Dominant
species, causes and consequences. Marine Pollution Bulletin 60 954–963
Doyle, T. K., Jonathan, D. R. H, Buckley, S. M., Hays, G. C. & Davenport, J. (2007).
The broad-scale distribution of five jellyfish species across a temperate coastal
environment. Hydrobiologia 579:29–39 DOI 10.1007/s10750-006-0362-2
Drake, A. G. & Klingenberg, C. P. (2010). Large-scale diversification of skull shape in
domestic dogs: Disparity and modularity. American Naturalist, 175(3), 289–
301.
Duarte, C. M., Pitt, K. A., Lucas, C. H., Purcell, J. E., Uye, S. I., Robinson, K., Brotz,
L., Decker, M. B., Sutherland, K. R., Malej, A., Madin, L., Mianzan, H., Gili, J
M., Fuentes, V., Atienza, D., Pages, F., Breitburg, D., Malek, J., Graham, W. M.
& Condon, R. H. (2012). Is global ocean sprawl a cause of jellyfish blooms?
Frontiers in Ecology and Environment doi:10.1890/110246
Durón-Benítez, Á. A. & Huang, W. C. (2016). Using Geometric Morphometrics to
Quantify Variation of Shape and Magnitude of the Pattern of Milk Production of
Dairy Cattle. Open Access Library Journal, 3: e2928.
Eschscholtz, F. (1829) System der Acalephen. Eine ausführliche Beschreibung aller
Medusenartigen Strahlthiere. Ferdi-nand Dümmler, Berlin, 190 pp.
Fenner, P. J., Lippmann, J., & Gershwin, L. A. (2009). Fatal and Nonfatal Severe
Jellyfish Stings in Thai Waters. Journal of Travel Medicine

106
Gershwin, L. A., Nardi, M. D., Winkel, K. D. & Fenner, P. J. (2010). Marine stingers:
review of an under-recognized global coastal management issue. Coastal
Management 38: 22–41.
Gershwin, L., Zeidler, W. & Davie, P.J.F. (2010). Medusae (Cnidaria) of Moreton Bay,
Queensland, Australia. In, Davie, P.J.F. & Phillips, J.A. (Eds), Proceedings of
the Thirteenth International Marine Biological Workshop, the Marine Fauna and
Flora of Moreton Bay, Queensland. Memoirs of the Queensland Museum 54(3):
47–108. Brisbane. ISSN 0079–8835.
Gibbons, M. J. & Richardson, A. J. (2008). Patterns of pelagic cnidarian abundance in
the North Atlantic. Hydrobiologia 616:51-65.
Graham, W. M. & Bayha, K. M. (2007). Biological invasions by marine jelly sh. In:
Nentwig W (ed) Biological Invasions (Ecological Studies), vol 193. Springer-
Verlag, Heidelberg, pp. 239-250
Graham, W. M., Martin, D. L., Felder, D. L., Asper, V. L., & Perry, H. M. (2003).
Ecological and economic implications of a tropical jellyfish invader in the Gulf
of Mexico. Biological Invasions Volume 5, Issue 1, pp 53-69 doi
10.1023/A:1024046707234
Graham, W. M., Gelcich, S., Robinson, K. L., Duarte, C. M., Brotz, L., Purcell, J. E.,
Madin, L. P., Mianzan, H., Sutherland, K. R., Uye, S., Pitt, K. A., Lucas, C. H.,
Bøgeberg, M., Brodeur, R. D. & Condon, R. H. (2015). Linking human well-
being and jellyfish: ecosystem services, impacts, and societal responses. Frontiers in Ecology and the Environment 12(9): 515–523, doi:10.1890/130298
Haddad, M. A. & Nogueira, M. (2006). Reappearance and seasonality of Phyllorhiza
punctata medusae in southern Brazil. Revista Brasileira De Zoologia 23: 824–
831.
Haeckel, E. (1880) Das System der Medusen. I, 2: System der Acraspeden. Gustav
Fischer, Jena, 361–672 pp.
Hale, G. (1999). The Classification and Distribution of the Class Scyphozoa
Hamner, W. M. (1995). Sensory ecology of scyphomedusae. Marine and Freshwater
Behavior and Physiology 26, 101-118
Hsieh, Y. H, Leong, P. F. M. & Rudloe, J. (2001). Jellyfish as food. Hydrobiologia
451:11-17.
Holst, S. (2012). Morphology and development of benthic and pelagic life stages of
North Sea jellyfish (Scyphozoa, Cnidaria) with special emphasis on the
identification of ephyra stages. Marine Biology 159:2707–2722
Hou, X. G, Stanley, G. D. Jr, Zhao, J, Ma, X. Y. (2005). Cambrian anemones with
preserved soft tissue from the Chengjiang biota, China. Lethaia 38: 193–203
Ibrahim, H.M. & Khalid, N. (2007). Growing Shipping Traffic in the Strait of Malacca:
Some Reflections on the Environmental Impact. Maritime Institute of Malaysia
(MIMA). Paper presented at the Global Maritime and Intermodal Logistics
Conference, Singapore, 17-18/12/2007

107
Jeyabaskaran, R., Mohan, G., Abhilash, K. S., Prema, D. & Kripa, V. (2016). Is the
scyphozoan jellyfish Lychnorhiza malayensis symbiotically associated with the
crucifix crab Charybdis feriatus? Current science, vol. 110, no. 4.
Kapiris, K., Miliou, H. & Moraitou-Apostolopoulou, M. (1997). Effects of
formaldehyde preservation on biometrical characters, biomass and biochemical
composition of Acartia dausi (Copepoda, Calanoida) HELGOLÄNDER
MEERESUNTERSUCHUNGEN Hetgoländer Meeresunters 5l, 95-106
Kawahara, M., Uye, S I., Ohtsu, K., & Iizumi, H. (2006). Unusual population explosion
of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in East
Asian waters. Marine Ecology Progress Series Vol. 307: 161–173, 2006
Kerschbaumer, M. & Sturmbauer, C. (2011). The Utility of Geometric Morphometrics
to Elucidate Pathways of Cichlid Fish Evolution. International Journal of
Evolutionary Biology Volume 2011, Article ID 290245, 8 pages
Kitamura, M. & Omori, M. (2010). Synopsis of edible jellyfishes collected from
Southeast Asia, with notes on jellyfish fisheries. Plankton and Benthos Research
5(3): 106 –118
Klingenberg, C. P. (2011). MorphoJ: an integrated software package for geometric
morphometrics. Molecular Ecology Resources 11: 353-357.
Klingenberg, C. P. (2013). Visualizations in geometric morphometrics: how to read
and how to make graphs showing shape changes Hystrix, the Italian Journal of
Mammalogy ISSN 1825-5272 Associazione Teriologica Italiana
Klingenberg, C. P. (2014). Analysis of Organismal Form (course code BIOL51111).
First semester of the 2014-15 academic year (3rd November to 12th December
2014). The University of Manchester.
Klingenberg, C. P. (2016). Size, shape, and form: concepts of allometry in geometric
morphometrics. Development Genes And Evolution 226: 113–137.
Kogovsek, T, Bogunovic, B, Malej, A (2010) Recurrence of bloom-forming
Scyphomedusae: wavelet analysis of a 200-year time series. Hydrobiologia
645:81-96
Kolbasova, G. D., Zalevsky, A. O., Gafurov, A. R., Gusev, P. O., Ezhova, M. A.,
Zheludkevich, A. A., Konovalova, O. P., Kosobokova, K. N., Kotlov, N. U.,
Lanina, N. O., Lapashina, A. S., Medvedev, D. O., Nosikova, K. S., Nuzhdina,
E. O., Bazykin, G. A., & Neretina, T. V. (2015). A new species of Cyanea
jellyfish sympatric to C. capillata in the White Sea. Polar Biology
Kramp, P. L. (1961). Synopsis of the medusae of the world. Journal of the Marine
Biological Association of the U.K, 40: 1–469.
Lesson, R. P. (1843) Histoire naturelle des Zoophytes, Acalèphes. Librairie
Encyclopédique de Roret, Paris, 596 pp.

108
Leone, A., Lecci, R. M., Durante, M. & Piraino, S. (2013). Extract from the
zooxanthellate jellyfish Cotylorhiza tuberculata modulates gap junction
intercellular communication in human cell cul- tures. Marine Drugs 11: 1728–
62.
Li, S., Ricchiardi, E., Bai, M. & Yang, X. K. (2016). A taxonomy review of Oreoderus
Burmeister, 1842 from China with a geometric morphometric evaluation
(Coleoptera, Scarabaeidae, Valgini). ZooKeys 552: 67-89
Light, S. F. (1914). Some Philippine scyphomedusae, including two new genera, five
new species, and one new variety. Philippine Journal of Science, 9: 195–231.
Lilley, M. K. S., Ferraris, M, Elineau, A., Berline, L., Cuvilliers, P., Gilletta, L., Thiéry,
A., Gorsky, G. & Lombard, F. (2014). Culture and growth of the jellyfish
Pelagia noctiluca in the laboratory. Marine Ecology Progress Series 510:265-
273
Linnaeus, C. (1758). Systema Naturae per regna tria naturae, secundum classes, ordines,
genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima,
reformata. Tomus I. Laurentii Salvii, Holmiae, 828 pp.
Lucas, C. H. (2001). Reproduction and life history strategies of common jellyfish,
Aurelia aurita, in relation to its ambient environment. Hydrobiologia 451:229-
246
Maderbacher, M., Bauer, C., Postl, H. L., Makasa, L. & Sturmbauer, C. (2008).
Assessment of traditional versus geometric morphometrics for discriminating
populations of the Tropheus moorii species complex (Teleostei: Cichlidae), a
Lake Tanganyika model for allopatric speciation. Journal of Zoological
Systematics and Evolutional Research
Malej, A., Turk, V., Lucic, D. & Benovic, A. (2007). Direct and indirect trophic
interactions of Aurelia sp. (Scyphozoa) in a stratified marine environment (Mljet
Lakes, Adriatic Sea). Marine Biology 151:827-841.
Mayer, A. G. (1910). The medusae of the world. The Scyphomedusae. Washington,
Carnegie Institution, vol. III, P. 499-735p.
Mayer, C., Metscher, B. D., Müller, G. B. & Mitteroecker, P. (2014). Studying
Developmental Variation with Geometric Morphometric Image Analysis
(GMIA). PLoS ONE 9(12): e115076
Menon, M. G. K. (1930). The Scyphomedusae of Madras and the neighbouring coast.
Bulletin of the Madras Government Museum 3: 1–28, 3 pls.
Mills, C. E. (2001). Jellyfish blooms: are populations increasing globally in response to
changing ocean conditions? Hydrobiologia 451: 55–68, 2001.
Mitteroecker, P., & Gunz, P. (2009). Advances in Geometric Morphometrics. Journal of
Evolutionary Biology 36:235–247

109
Mitteroecker, P., & Bookstein, F. (2011). Linear Discrimination, Ordination, and the
Visualization of Selection Gradients in Modern Morphometrics. Journal of
Evolutionary Biology 38:100–114
Mitteroecker, P., & Gunz, P., Windhager, S. & Schaefer, K. (2013). A brief review of
shape, form, and allometry in geometric morphometrics, with applications to
human facial morphology. Hystrix, the Italian Journal of Mammalogy. Volume
24 (1): 59–66, 2013
Mitteroecker, P., Windhager, S., Müller, G. B & Schaefer, K. (2015). The
Morphometrics of “Masculinity” in Human Faces. PLoS ONE 10(2):e0118374.
Mooney, C. J. & Kingsford, M. J. (2016). Statolith Morphometrics Can Discriminate
among Taxa of Cubozoan Jellyfishes PLoS ONE 11(5): e0155719.
doi:10.1371/journal.pone.0155719
Morandini, A. C. & Marques, A. C. (2010). Revision of the genus Chrysaora Péron &
Lesueur, 1810. Zootaxa 2464: 1–97
Morton, B. (2005). Over fishing: Hong Kong - fishing crisis finally arrives. Marine
Pollution Bulletin
Mujiono, M. (2010). Jellyfish (Crambionella sp.) Fisheries around Cilacap waters,
Centreal Java (Cnidaria: Scyphozoa). Oseanologi dan Limnologi di Indonesia
36(1): 37- 48
Mullon, C., Freon, P. & Cury, P. (2005). The dynamics of collapses in world fisheries.
Fish Fish 6:111-120.
Muñoz-Muñoz, F., & Perpiñán, D. (2010). Measurement error in morphometric studies:
comparison between manual and computerized methods. Annales Zoologici
Fennici 47: 46–56
Nair, K. K. (1951). Medusae of the Trivandrum coast. Part 1. Systematics. Bulletin of
the Central Research Institute, University of Travancore Series C Natural
Sciences 2:47– 75
Nakanishi, N., Camara, A. C., Yuan, D. C., Gold, D. A & Jacobs, D. K. (2015). Gene
Expression Data from the Moon Jelly, Aurelia, Provide Insights into the
Evolution of the Combinatorial Code Controlling Animal Sense Organ
Development PLoS One 10(7): e0132544.
Nishikawa, J., Nguyen, T. T., Tran, M. H., & Pham, T. T. (2008). Jellyfish fisheries in
northern Vietnam. Plankton and Benthos Research 3(4): 227–234
Nogueira, M. J. & Haddad, M. A. (2006). Reappareance and seasonality of Phyllorhiza
punctata von Lendefeld (Cnidaria, Scyphozae, Rhizostomaea) medusa in
southern Brazil. Revista Brasileira de Zoologia 23 (3): 824–831
Nunes, P., Loureiro, M. L., Piñol, L., Sastre, S., Voltaire, L. & Canepa, A. (2015).
Analyzing Beach Recreationists’ Preferences for the Reduction of Jellyfish
Blooms: Economic Results from a Stated-Choice Experiment in Catalonia,
Spain. PLoS ONE 10(6): e0126681

110
Omori, M., & Kitamura, M. (2004). Taxonomic review of three Japanese species of
editable jellyfish (Schyphozoa: Rhizostomeae). Plankton Biology and Ecology
51(1): 36-51
Omori, M., & Nakano, E. (2001). Jellyfish fisheries in Southeast Asia. Hydrobiologia
451: 19–26, 2001.
Othman I & Burnett J. W. 1990. Techniques applicable for the purifying of Chironex
fleckeri (box-jellyfish) venom. Toxicon 28:821-835.
Park, P. J., Aguirre, W. E., Spikes, D. A., & Miyazaki, J. M. (2013). Landmark-Based
Geometric Morphometrics: What Fish Shapes Can Tell Us about Fish Evolution.
Proceedings of the Association for Biology Laboratory Education 34, 361-371.
Parsons, T. R. & Lalli, C. M. (2002). Jellyfish population explosions: revisiting a
hypothesis of possible causes. La Mer 40:111- 121.
Pitt, K. A. & Kingsford, M. J. (2003). Temporal variation in the virgin biomass of the
edible jellyfish, Catostylus mosaicus (Scyphozoa, Rhizostomeae). Fisheries
Research 63:303-313.
Pohl, I. S. & Jarms, G. (2010). Identification key for young ephyrae: a first step for
early detection of jellyfish blooms. Hydrobiologia doi 10.1007/s10750-010-0226-
7
Polly, P. D., Lawing, A. M., Fabre, A., & Goswami, A. (2012), Phylogenetic Principal
Components Analysis and Geometric Morphometrics Hystrix. The Italian
Journal of Mammalogy Volume 24 (1): 33–41
Polly, P. D., Stayton, C. T., Dumont, E. R., Pierce, S. E., Rayfield, E. J. & Angielczyk,
K. D. (2016). Combining geometric morphometrics and finite element analysis
with evolutionary modeling: towards a synthesis, Journal of Vertebrate
Paleontology, 36:4, e1111225
Praveena, S. M., Siraj, S. S., Suleiman, A. K. & Aris, A. Z. (2011). A brush up on water
quality studies of Port Dickson, Malaysia. Research Journal of Environmental
Sciences, 5(12), 841–849.
Purcell, J. E., Hoover, R. A. & Schwarck, N. T. (2009). Interannual variation of
strobilation by the scyphozoan Aurelia labiata in relation to polyp density,
temperature, salinity, and light conditions in situ. Marine Ecology Progress Series
Vol. 375: 139–149
Purcell, J. E., Atienza, D., Fuentes, V., Olariaga, A., Tilves, U., Colahan, C. & Gili, J.
(2012). Temperature effects on asexual reproduction rates of scyphozoan species
from the northwest Mediterranean Sea. Hydrobiologiadoi 10.1007/s10750-012-
1047-7
Purcell, J. E., Uye, S I. & Lo, W. T. (2007). Anthropogenic causes of jellyfish blooms
and their direct consequences for humans: a review. Marine Ecology Progress
Series Vol. 350: 153–174, 2007

111
Raskoff, K. A. (2001). The impact of El Nino events on populations of mesopelagic
hydromedusae. Hydrobiologia 451:121- 129.
Rezai, H., Yusoff, F. M., Kawamura, A., Arshad, A. & Othman, B. H. R. (2003).
Zooplankton biomass in the Straits of Malacca. Indian Journal of Marine
Sciences, 32(3), 222-225.
Richardson, A. J., Bakun, A., Hays, G. C., & Gibbons, M. J. (2009). The jellyfish
joyride: causes, consequences and management responses to a more gelatinous
future. Trends in Ecology and Evolution Vol.24 No.6
Richtsmeier, J. T., Deleon, V. B., & Lele, S. T. (2002). The Promise of Geometric
Morphometrics. Yearbook Of Physical Anthropology 45:63–91
Rohlf, F. J., & Marcus, L. F. (1993). A Revolution in Geometric Morphometrics.
Trends in Ecology and Evolutions Vol. 8, No. 4
Rohlf, F. J. (2010). Tps Series. Department of Ecology and Evolution, State University
of New York, Stony Brook, New York. Available:
http://life.bio.sunysb.edu/morph/.
Rizman-Idid, M, Azwa, A. B. F., Chong, V.C. (2016). Preliminary Taxonomic Survey
and Molecular Documentation of Jellyfish Species (Cnidaria: Scyphozoa and
Cubozoa) in Malaysia. Zoological Studies 55: 35
Ruiz, J., Prieto, L. & Astorga, D. (2012) A model for temperature control of jellyfish
(Cotylorhiza tuberculata) outbreaks: a causal analysis in a Mediterranean coastal
lagoon. Ecological Modelling
Rumpet, R. (1991). Some aspects of the biology and fishery of jellyfish found along the
coast of Sarawak, Malaysia. Dept. of Fisheries, Ministry of Agriculture, Malaysia,
37 pp.
Shao, Z., Graf, S., Chaga, OY. & Lavrov, DV. (2006). Mitochondrial genome of the
moon jelly Aurelia aurita (Cnidaria, Scyphozoa): A linear DNA molecule
encoding a putative DNA-dependent DNA polymerase. Gene 381:92-101.
Sheets, H. D., Covino, K. M., Panasiewicz, J. M. & Morris, S. R. (2001). Comparison
of geometric morphometric outline methods in the discrimination of age-related
differences in feather shape. Frontiers in Zoology 3:15 doi:10.1186/1742-9994-
3-15
Slice, Dennis E. (2013). Morpheus et al., Java Edition. Department of Scientific
Computing, The Florida State University, Tallahassee, Florida, U.S.A. Available
from http:/morphlab.sc.fsu.edu/
Sparmann, S. F. (2012). Contributions to the molecular phylogeny, phylogeography,
and taxonomy of scyphozoan jellyfish. Dissertation, University of British
Columbia
Strauss, R. E. (2010). Morphometrics for Nonmorphometricians. Lecture Notes in Earth
Sciences 124

112
Tan, N. H., Oo, S. L, Thambyrajah, V. & Azila, N. (1993). Advances in venom and
toxin research. Proceedings of the Third Asia Pacific Congeress on Animal, Plant
and Microbial Toxins. Malaysian Society on Toxinology pp. 113-118.
Thai Marine Database. http://marinegiscenter.dmcr.go.th
Uye, S., Shimizu, M. & Watanabe, T. (2010). Tackling the giant jellyfish (Nemopilema
nomurai) plague: cause, forecast and countermeasure. Plenary presentation,
Third International Jellyfish Blooms Symposium, Mar del Plata, July 14, 2010.
Vanhöffen, E. (1888). Untersuchungen über semäostome und rhizostome Medusen.
Bibliotheca Zoologica, 1(3): 5–52.
Vanhöffen, E. (1911). Die Anthomedusen und Leptomedusen der Deutschen Tiefsee-
Expedition 1898-1899. Wissenschaftliche Ergenbnisse der Deutschen Tiefsee-
Expedition auf dem Dampfer ‘Valdivia’ 1898-1899, 19(5): 191–233, 22 pls.
Viscosi, V. & Cardini, A. (2011). Leaf Morphology, Taxonomy and Geometric
Morphometrics: A Simplified Protocol for Beginners. PLoS ONE 6(10): e25630.
Waharak, A., Ramarn, T. & Panritdam, T. (2013). Diversity and feeding of Jellyfish
(Cnidaria) at Koh Mak Noi and Koh Yao Noi, Phang-Nga Province, Southern
Thailand. 3rd International Conference on Ecological, Environmental and
Biological Sciences (ICEEBS'2013) April 29-30, 2013 Singapore
Webster, M. (2010). A Practical Introduction to Landmark-Based Geometric
Morphometric In Quantitative Methods In Paleobiology, PP. 163-188,
Paleontological society short course, october 30th, 2010. The Paleontological
society Papers volume 16, John Alroy and Gene Hunt (eds.).
WoRMS Editorial Board. (2017). World Register of Marine Species.
http://www.marinespecies.org. Accessed 10 Feb 2017.
Yap, W. L. Nicholas. & Ong, J. Y. (2012). A survey of jellyfish (Cnidaria) around St
John’s island in the Singapore Straits. Marine Science 2012: 57–74
Yonehara, Y., Konuma, J. & Klingenberg, C. P. The Use of Geometric Morphometrics
in a Study of Shape Diversity of Ground Beetles (Coleoptera: Carabidae)
You, K., Ma, C., Gao, H., Li, F., Zhang, M., Qiu, Y. & Wang, B. (2007). Research on
the jellyfish (Rhopilema esculentum Kishinouye) and associated aquaculture
techniques in China: current status. Aquaculture International 15:479–488
Young, G. A. & Hagadorn, J. W. (2010). The fossil record of cnidarian medusae.
Palaeoworld 19: 212–221.
Zelditch, M. L., Swiderski, D. L. & Sheets, H. D. A Practical Companion to Geometric
Morphometrics for Biologists: Running analyses in freely-available software

113
APPENDICES
Appendix A: Formalin preparation
1. Neutralize formalin by adding 20 table spoons of magnesium carbonate to 2.5
litres of formalin.
2. Wait for magnesium carbonate to dissolve completely, approximately 6 hours.
3. Measure the pH of the formalin. pH must be within 7 to 8. It can be slightly
more alkaline but cannot be acidic.
4. Mix neutralized formalin with the ratio of 95% seawater and 5% of neutralized
formalin. Filter the seawater prior to mixing.
5. Formalin is highly carcinogenic, thus proper precaution must be taken. Gloves
and mask must be put on when working on the specimens. Fume cupboard
should be utilized when preparing formalin.
Appendix B: List of jellyfish morphological characters analysed and their respective
character states.
Character Character Status
Symmetry of medusae radial = 0, biradial = 2, tri-radial = 3, radial
tetramerous = 4, pentamerous = 5
Pedalium absent = 0, coronate type = 1, cubozoan type =
2
Rhopaloid absent = 0, present = 1
Rhopalium absent = 0, present and simple = 1, present
and complex eyes = 2
Ocelli absent = 0, simple ocellus = 1, ocellus w/ lens
= 2, ocellus in complex eye = 3
Nerve ring absent = 0, one = 1, two = 2
Nerve net absent = 0, DNN only = 1, DNN and MNN = 2,
MNN only = 3
DNN and MNN connected in marginal
centers or ganglia
no = 0, yes = 1
Gastric filaments absent = 0, present = 1
Coronal muscle well developed = 0, marginal and tiny = 1
Velum-like structure absent = 0, velum = 1, velarium = 2
Velar canals absent = 0, present = 1
Frenulae absent = 0, present = 1
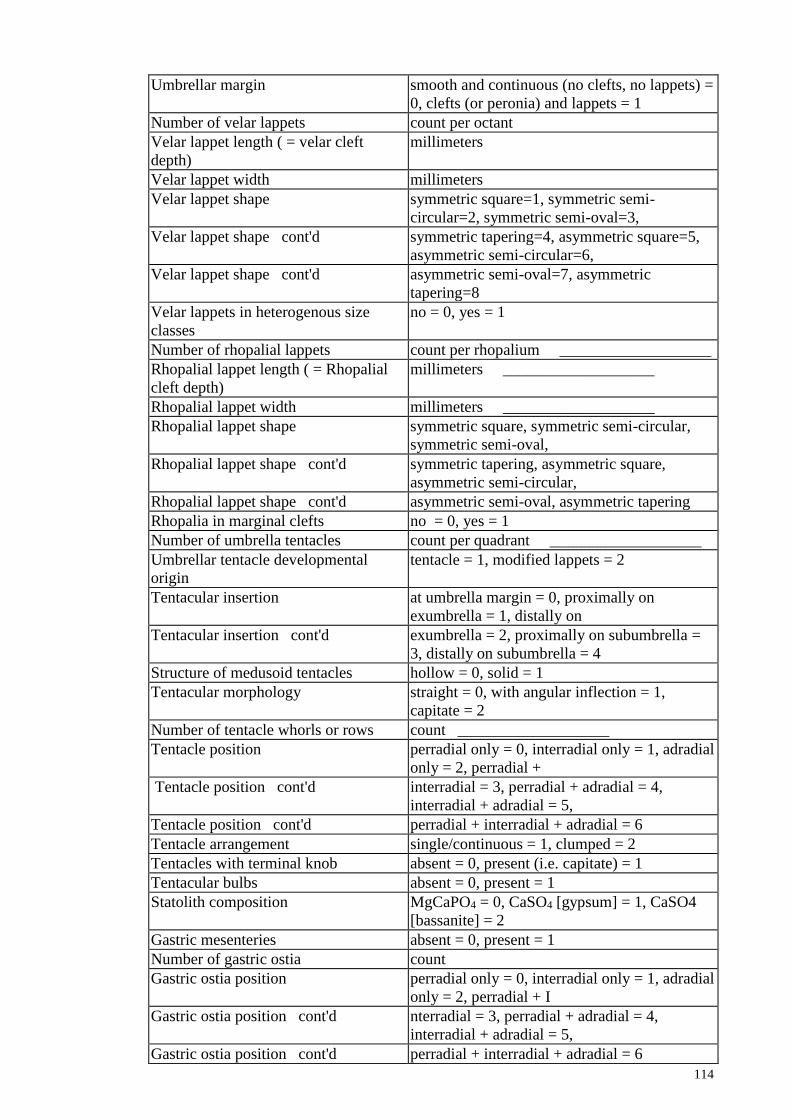
114
Umbrellar margin smooth and continuous (no clefts, no lappets) =
0, clefts (or peronia) and lappets = 1
Number of velar lappets count per octant
Velar lappet length ( = velar cleft
depth)
millimeters
Velar lappet width millimeters
Velar lappet shape symmetric square=1, symmetric semi-
circular=2, symmetric semi-oval=3,
Velar lappet shape cont'd symmetric tapering=4, asymmetric square=5,
asymmetric semi-circular=6,
Velar lappet shape cont'd asymmetric semi-oval=7, asymmetric
tapering=8
Velar lappets in heterogenous size
classes
no = 0, yes = 1
Number of rhopalial lappets count per rhopalium ___________________
Rhopalial lappet length ( = Rhopalial
cleft depth)
millimeters ___________________
Rhopalial lappet width millimeters ___________________
Rhopalial lappet shape symmetric square, symmetric semi-circular,
symmetric semi-oval,
Rhopalial lappet shape cont'd symmetric tapering, asymmetric square,
asymmetric semi-circular,
Rhopalial lappet shape cont'd asymmetric semi-oval, asymmetric tapering
Rhopalia in marginal clefts no = 0, yes = 1
Number of umbrella tentacles count per quadrant ___________________
Umbrellar tentacle developmental
origin
tentacle = 1, modified lappets = 2
Tentacular insertion at umbrella margin = 0, proximally on
exumbrella = 1, distally on
Tentacular insertion cont'd exumbrella = 2, proximally on subumbrella =
3, distally on subumbrella = 4
Structure of medusoid tentacles hollow = 0, solid = 1
Tentacular morphology straight = 0, with angular inflection = 1,
capitate = 2
Number of tentacle whorls or rows count ___________________
Tentacle position perradial only = 0, interradial only = 1, adradial
only = 2, perradial +
Tentacle position cont'd interradial = 3, perradial + adradial = 4,
interradial + adradial = 5,
Tentacle position cont'd perradial + interradial + adradial = 6
Tentacle arrangement single/continuous = 1, clumped = 2
Tentacles with terminal knob absent = 0, present (i.e. capitate) = 1
Tentacular bulbs absent = 0, present = 1
Statolith composition MgCaPO4 = 0, CaSO4 [gypsum] = 1, CaSO4
[bassanite] = 2
Gastric mesenteries absent = 0, present = 1
Number of gastric ostia count
Gastric ostia position perradial only = 0, interradial only = 1, adradial
only = 2, perradial + I
Gastric ostia position cont'd nterradial = 3, perradial + adradial = 4,
interradial + adradial = 5,
Gastric ostia position cont'd perradial + interradial + adradial = 6

115
Number of radial mesenteries count ___________________
Radial mesentery shape straight = 1, bent distally = 2, paired forming Y
proximally = 3
Radial mesentery termination percent of distance from tentacle (0%) to
rhopalium (100%) ___________
Number of radiating stomach pouches count ___________________
Radial canals absent = 0, present = 1
Number of perradial canal origins at the
gastrovascular cavity
count per quadrant ___________________
Number of interradial canal origins at
the gastrovascular cavity
count per quadrant ___________________
Number of adradial canal origins at the
gastrovascular cavity
count per quadrant ___________________
Number of perradial-perradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of interradial-interradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of adradial-adradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of perradial-interradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of perradial-adradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of interradial-adradial
anastomoses in radial canals that are
circumscribed by the ring canal
count per quadrant ___________________
Number of terminations of perradial
canals at the ring canal
count per quadrant ___________________
Number of terminations of interradial
canals at the ring canal
count per quadrant ___________________
Number of terminations of adradial
canals at the ring canal
count per quadrant ___________________
Number of perradial canals originating
distally at the circular canal
count per quadrant ___________________
Number of interradial canals
originating distally at the circular canal
count per quadrant ___________________
Number of adradial canals originating
distally at the circular canal
count per quadrant ___________________
Number of sinuses originating at the
gastrovascular cavity
count per quadrant ___________________
Number of sinuses originating at the
perradial canal
count per quadrant ___________________
Number of sinuses originating at the
interradial canals
count per quadrant ___________________
Number of sinuses originating at the
adradial canals
count per quadrant ___________________
Number of sinuses originating count per quadrant ___________________

116
proximally at the circular canal
Number of anastomoses circumscribed
by the circular canal that lead to two
sinuses
count per quadrant ___________________
Number of sinuses originating distally
at the circular canal
count per quadrant ___________________
Percentage of radius of medusa in
which there is no branching radial canal
count per quadrant ___________________
Circular canal absent = 0, weakly developed chain of enlarged
branches circumscribes
Circular canal cont'd bell = 1, a primary artery easily distinguishable
from other canals
Circular canal cont'd circumscribes bell = 2
Furrow in bell absent = 0, coronal groove = 1, laingiomedusan
type = 2
Number of gonads count
Gonads are paired no = 0, yes = 1
Gonad position axis perradial only = 0, interradial only = 1, adradial
only = 2, perradial +
Gonad position axis cont'd interradial = 3, perradial + adradial = 4,
interradial + adradial = 5,
Gonad position axis cont'd perradial + interradial + adradial = 6
Lateral distance from center to most
proximal portion of gonad
millimeters ___________________
Lateral distance from center to most
distal portion of gonad
millimeters ___________________
Gonad associated with particular
structure?
manubrium = 0; radial canals = 1; mesogleal
protrusion (gastric septa or
Gonad associated with particular
structure? cont'd
quadralinga) = 2; radial septa = 3; pouch = 4;
outfolded pockets = 5
Statocysts absent = 0, endodermal = 1, ectodermal = 2
Shape of mature medusa bell on oral-
aboral plane
semi-circular convex = 0, triangular = 1, wide
rectangular = 2,
Shape of mature medusa bell on oral-
aboral plane cont'd
tall rectangular = 3, square = 4, actinuloid = 5,
sinusoid = 6, sinusoid with
Shape of mature medusa bell on oral-
aboral plane cont'd
central globose mass
Bell thickness millimeters ___________________
Development of the umbrella fully developed = 0, aboral cone = 1
Shape of mature medusa bell on plane
perpendicular to oral-aboral axis
circular = 0, square = 1
Urticant rings absent = 0, present = 1
Mouth lips absent = 0, simple lips = 1, gelatinous or
curtain-like arms = 2,
Mouth lips cont'd oral arms with suctorial mouths = 2
Manubrium absent = 0, basal in arms = 1, basal and
extended beyond arms = 2,
Manubrium cont'd pillars and disk = 3
Manubrium depth millimeters ___________________
Manubrium width at base millimeters ___________________
Manubrium width at mouth millimeters ___________________
Length of the simple, unwinged portion millimeters ___________________

117
of the oral arm
Length of the winged portion of the
oral arm
millimeters ___________________
Oral arm width millimeters ___________________
Cross-sectional form of oral arm sheet-like = 1, two-winged = 2, three-winged =
3
Secondary structure of oral arm absent = 0, spiral =1
Number of fenestrations in oral arm count ___________________
Scapulae absent = 0, present = 1
Point of scapula attachment to oral
mass
at disk = 1, both disk and oral arm = 2, on
smooth portion of oral arm= 3
Length of attachment to oral mass millimeters ___________________
Length of scapula (smooth part) millimeters ___________________
Length of scapula (mouthed part) millimeters ___________________
Distribution of mouths on scapula top= 1; bottom = 2, entire surface = 3,
Shape of scapula straight = 1, scimitar-shaped, curved up = 2,
finger-like, curved up = 3
Scapulae occurrence per oral arm one per arm = 1; 2 per arm = 2
Scapulae branched? no= 0 ; yes = 1
Number of filaments per scapulae count ___________________
Distribution of filaments on scapulae absent = 0, scapula exterior only = 1, scapula
interior only = 2
Shape of scapular filaments rod-like = 1; tapering = 2, string-like = 3,
string-like with terminal bulb
Shape of scapular filaments (capitate) = 4, spatulate = 5
Length of scapular filaments millimeters ___________________
Width of scapular filaments millimeters ___________________
Colour of scapular filaments C-M-Y-K ___________________
Number of terminal clubs count ___________________
Cross-sectional shape of terminal clubs circular = 0, planar = 1, convex planar (ovoid)
= 2, concave planar = 3,
Cross-sectional shape of terminal clubs
cont'd
triangular = 4, convex triangular = 5, concave
triangular = 6
Longitudinal-sectional shape of
terminal clubs
rod-like = 0, tapering = 1, string-like = 3,
string-like with terminal bulb = 4,
Longitudinal-sectional shape of
terminal clubs cont'd
spatulate = 5
Length of terminal clubs millimeters ___________________
Width of terminal clubs millimeters ___________________
Length of the oral pillars millimeters ___________________
Width of the oral pillars millimeters ___________________
Depth of the oral pillars millimeters ___________________
Width of the subgenital fenestration millimeters ___________________
Perradial diameter of the oral disc millimeters ___________________
Depths of the oral disc millimeters ___________________
Distribution of intermediate filaments
on the oral arm and oral disc
absent = 0, oral arm exterior only = 1, oral arm
interior only = 2, oral disk
Distribution of intermediate filaments
on the oral arm and oral disc cont'd
only = 3, oral arm = 4, oral arm and disk = 5
Number of intermediate filaments on
the oral arm
count ___________________
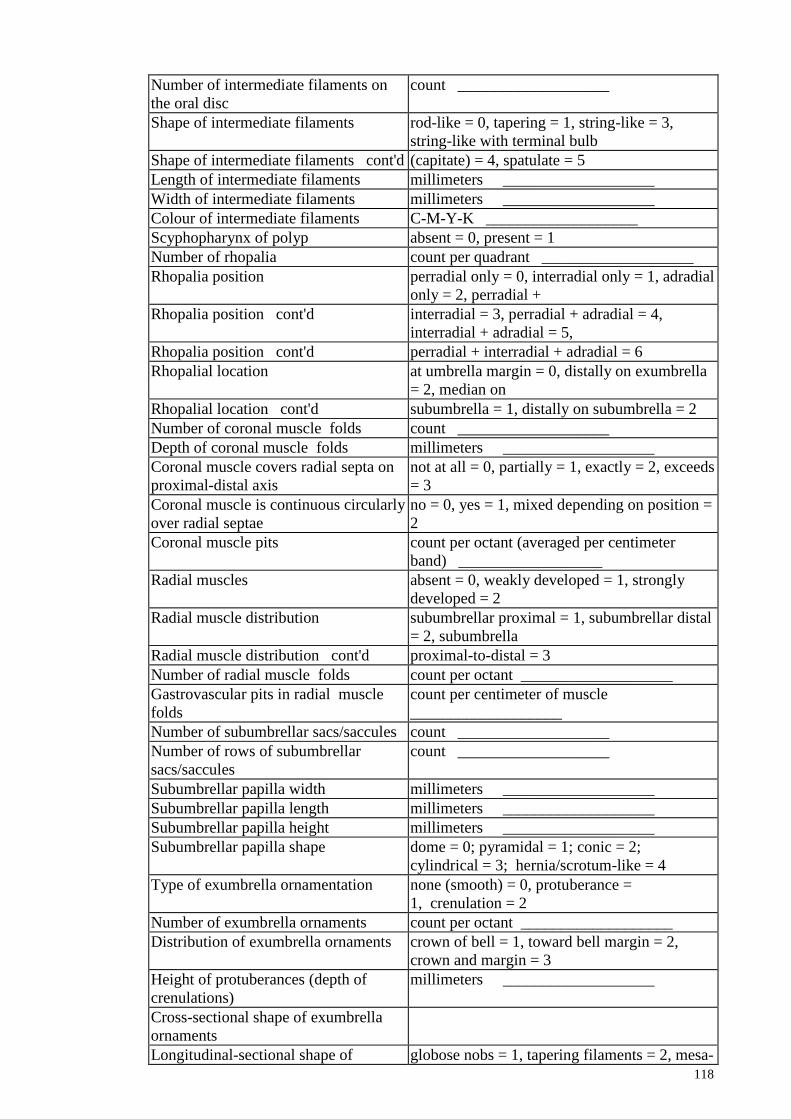
118
Number of intermediate filaments on
the oral disc
count ___________________
Shape of intermediate filaments rod-like = 0, tapering = 1, string-like = 3,
string-like with terminal bulb
Shape of intermediate filaments cont'd (capitate) = 4, spatulate = 5
Length of intermediate filaments millimeters ___________________
Width of intermediate filaments millimeters ___________________
Colour of intermediate filaments C-M-Y-K ___________________
Scyphopharynx of polyp absent = 0, present = 1
Number of rhopalia count per quadrant ___________________
Rhopalia position perradial only = 0, interradial only = 1, adradial
only = 2, perradial +
Rhopalia position cont'd interradial = 3, perradial + adradial = 4,
interradial + adradial = 5,
Rhopalia position cont'd perradial + interradial + adradial = 6
Rhopalial location at umbrella margin = 0, distally on exumbrella
= 2, median on
Rhopalial location cont'd subumbrella = 1, distally on subumbrella = 2
Number of coronal muscle folds count ___________________
Depth of coronal muscle folds millimeters ___________________
Coronal muscle covers radial septa on
proximal-distal axis
not at all = 0, partially = 1, exactly = 2, exceeds
= 3
Coronal muscle is continuous circularly
over radial septae
no = 0, yes = 1, mixed depending on position =
2
Coronal muscle pits count per octant (averaged per centimeter
band) __________________
Radial muscles absent = 0, weakly developed = 1, strongly
developed = 2
Radial muscle distribution subumbrellar proximal = 1, subumbrellar distal
= 2, subumbrella
Radial muscle distribution cont'd proximal-to-distal = 3
Number of radial muscle folds count per octant ___________________
Gastrovascular pits in radial muscle
folds
count per centimeter of muscle
___________________
Number of subumbrellar sacs/saccules count ___________________
Number of rows of subumbrellar
sacs/saccules
count ___________________
Subumbrellar papilla width millimeters ___________________
Subumbrellar papilla length millimeters ___________________
Subumbrellar papilla height millimeters ___________________
Subumbrellar papilla shape dome = 0; pyramidal = 1; conic = 2;
cylindrical = 3; hernia/scrotum-like = 4
Type of exumbrella ornamentation none (smooth) = 0, protuberance =
1, crenulation = 2
Number of exumbrella ornaments count per octant ___________________
Distribution of exumbrella ornaments crown of bell = 1, toward bell margin = 2,
crown and margin = 3
Height of protuberances (depth of
crenulations)
millimeters ___________________
Cross-sectional shape of exumbrella
ornaments
Longitudinal-sectional shape of globose nobs = 1, tapering filaments = 2, mesa-

119
exumbrella ornaments like = 3, mound = 4
Colour of exumbrella ornaments C-M-Y-K ___________________
Color of bell C-M-Y-K ___________________
Perradial canal colour C-M-Y-K ___________________
Interradial canal colour C-M-Y-K ___________________
Adradial canal colour C-M-Y-K ___________________
Number of pigmented flecks in
perradial canal
count per quadrant ___________________
Number of pigmented flecks in
interradial canal
count per quadrant ___________________
Number of pigmented flecks in
adradial canal
count per quadrant ___________________
Oral arm colour C-M-Y-K ___________________
Terminal club colour C-M-Y-K ___________________
Shape of pigment on exumbrella none = 0, dot = 1, circle = 2, uneven patch = 3,
radiating lines = 4, star = 5,
Shape of pigment on exumbrella
cont'd
wheel = 6, eyespot = 7, …
Colour of pigmentation on
the exumbrellar surface
C-M-Y-K ___________________
Number of pigmented
spots/patches /shapes on exumbrellar
surface
count per octant ___________________
Distribution of color
spots/patches/shapes on exumbrella
crown of bell = 1, toward bell margin = 2,
crown and margin = 3
Mass of the whole medusae grams
Bell diameter millimeters ___________________
Ring canal diameter millimeters ___________________
Shape of the stomach/gonadal cavity circular = 0, cruciform = 1, pouched = 3,
outfolded pockets = 4.
Perradial diameter of the stomach
cavity
millimeters ___________________
Structural form of gonad digitate = 1, ribbon = 2, floret = 3
Colour of the subgenital porticus C-M-Y-K ___________________
Thickness of the subgenital porticus millimeters ___________________
Color of gonads C-M-Y-K ___________________
Color of gastric filaments C-M-Y-K ___________________
Color of the bell margin C-M-Y-K ___________________
Rhopaliar pit depth millimeters ___________________
Quadralinga length millimeters ___________________
Quadralinga diameter millimeters ___________________
Quadralinga shape scooped = 1, tri-lobed = 3
Subumbrella radial furrows absent = 0, present = 1
Location where planulae are brooded not brooded = 0, brood filaments = 1,
proximally on oral arms = 2, on
Location where planulae are brooded manubrium = 3, on manubrium and oral arms =
4
Number of subumbrellar radial furrows count per octant ___________________

120
Appendix C: Check list of morphology photography and observation
Tank
1 Label
2 Lateral view whole animal Front
3 Lateral view whole animal Front RGB
4 Lateral view whole animal Side
5 Close-up Front/Right Quadrant -Front or side
6 Close-up Bell Margin -Front or side
7 Close-up of tentacle -Front or side
8 Close-up of Oral arms - Front or side
9 Close-up of Oral Arm Filaments
10 Close-up of terminal clubs - Front or side
11 Close-up of Mouthlets - Front
12 Close-up of Scapulae - Front or side
13 Close-up of Scapular Filaments
14 Close-up of Scapular Filaments RGB
15 Whole animal from top RGB
Lift up bell, drape oral arms to expose ends of manubrium
16 Close-up of Manubrium/Mouth
17 Close-up of gastric filaments
18 Oral side up from Top
Measure mass of the whole medusae (grams)
Measure Bell Diameter
Measure Bell thickness
Measure Oral Disk Diameter 1) Interradial 2) Perradial
Measure Depth of Oral Disc
Count number of intermediate filaments on disk
Subumbrella up, tentacles/oral arms out
19 Whole medusae - RGB with flash
20 Whole medusae - Bottom Illum/B Background
21 Whole medusae - Bottom Illum/Full Trans
Move tentacles/oa from quadrant
22 Close up of Quadrant - Bottom Illum/Full Trans
23 Close-up of Quadrant - Bottom Illum/B Background
24 Picture of center - Bottom Illum/B Background
25 Picture of center - Bottom Illum/Full Trans
26 Close-up of Velar Lappets - Bottom Illum/B Background
27 Close-up of Velar Lappets - Bottom Illum/Full Trans
Count number of Velar Lappets _______________
28 Close-up of Rhopaliar Lappets - Bottom Illum/B Background
29 Close-up of Rhopaliar Lappets - Bottom Illum/Full Trans
Count number of Rhopaliar Lappets
30 Close-up of Rhopalium - Bottom Illum/B Background
31 Close-up of Rhopalium - Bottom Illum/Full Trans
32 Close-up of Coronal Muscles - Bottom Illum/B Background

121
33 Close-up of Coronal Muscles - Bottom Illum/Full Trans
Measure number of coronal muscle folds
Measure depth of coronal muscle folds
34 Close-up of Radial Muscles - Bottom Illum/B Background
35 Close-up of Radial Muscles - Bottom Illum/Full Trans
Measure number of radial muscle folds
Measure depth of radial muscle folds
36 Close-up of canals 2 quads - Bottom Illum/Full Trans
37 Close-up of canals 2 quads - Bottom Illum/B Blackground
38 Close-up of manubrium - Bottom Illum/B Background
39 Close-up of manubrium - Bottom Illum/Full Trans
Measure Manubrium depth 1)___ 2) ___
Measure Manubrium width at base
Measure Manubrium width at mouth
Move oral arms to top in a group - reveal oral pillars
40 Close-up of Oral Pillar - Bottom Illum/B Background
41 Close-up of Oral Pillar - Bottom Illum/Full Trans
Measure depth of oral Pillar
Measure height of oral pillar
Measure the wide of oral pillar
42 Close-up of subgenital fenestration - Bottom Illum/B Background
43 Close-up of subgenital fenestration - Bottom Illum/Full Trans
Measure width of subgenital fenestration
Measure height of subgenital fenestration
44 Close-up of Subumbrellar papillae - Bottom Illum/B Background
45 Close-up of Subumbrellar papillae - Bottom Illum/Full Trans
Measure Subumbrellar papilla width
Measure Subumbrellar papilla length
Measure Subumbrellar papilla height
Observe shape of Subumbrellar papilla
Cut 1 or 2 oral pillars - reveal gonad and stomach cavity
46 Close-up of gonad - Bottom Illum/B Background
47 Close-up of gonad - Bottom Illum/Full Trans
Check gonad association with particular structure
Measure Lateral distance from center to most proximal portion of gonad
Measure Lateral distance from center to most distal portion of gonad
48 Close-up of stomach cavity - Bottom Illum/B Background
49 Close-up of stomach cavity - Bottom Illum/Full Trans
Observe shape of the stomach cavity
Measure Perradial diameter of the stomach cavity
Measure Interradial diameter of the stomach cavity
50 Close-up of gastric filaments
Observe presence of gastric filaments
Splay 1/2 of oral arms and terminal clubs out
51 Close-up of oral arms - Bottom Illum/B Background

122
52 Close-up of oral arms - Bottom Illum/Full Trans
Splay single oral arm out (winged & unwinged portion esp.) - expose
fenestrations - show the base and middle of oral arm
53
Close-up of single oral arm/terminal clubs/filaments - Bottom Illum/B
Background
54
Close-up of single oral arm/terminal clubs/filaments - Bottom Illum/Full
Trans
Measure total length of oral arms
Measure unwinged length of oral arms
Measure width of oral arms
Observe cross-sectional shape of oral arms
Observe longitudinal-sectional shape of oral arms
Count number of fenestrations in oral arms
Measure length of terminal clubs
Measure width of terminal clubs
Observe cross-sectional shape of terminal clubs
Observe longitudinal-sectional shape of terminal clubs
Measure length of filaments
Measure width of filaments
Count number of filaments
Observe cross-sectional shape of filaments
Observe longitudinal-sectional shape of filaments
Splay out scapulae as with oral arm (above)
55 Close up of scapula/filaments - Bottom Illum/Black Background
56 Close up of scapula/filaments - Bottom Illum/Full Trans
Observe cross-sectional shape of scapula
Observe longitudinal-sectional shape of scapula
Measure length of filaments
Measure width of filaments
Count number of filaments
Observe cross-sectional shape of filaments
Observe longitudinal-sectional shape of filaments
Pull up oral disk, take a look at subgenital porticus
57 Measure thickness of the subgenital porticus
Observe the colour of subgenital porticus
Flip animal so exumbrella faces up
58 Close-up of Exumbrella Quadrant - Bottom Illum/B Background
59 Close-up of Exumbrella Quadrant - Bottom Illum/Full Trans
60 Whole animal Exumbrella - Bottom Illum/B Background
61 Whole animal Exumbrella - Bottom Illum/Full Trans
Measure height of protuberances (depth of crenulations)
Observe cross-sectional shape of ornaments
Observe longitudinal-sectional shape of ornament
If dying canals possible - flip animal and dye canals

123
62 Close-up of dyed canals 2 quads
Dissect Rhopalia - place in vial
Remove tentacle(s) - place in vial
Appendix D: List of 107 specimens for geometric morphometric analysis
Specimen Site Area
MRI001WCD1A Sungai Janggut West-Central
MRI002WCD1A Sungai Janggut West-Central
MRI003WCD1A Sungai Janggut West-Central
MRI004WCD1A Sungai Janggut West-Central
MRI005WCD1A Sungai Janggut West-Central
MRI006WCD1A Sungai Janggut West -Central
MRI007WCD1A Sungai Janggut West -Central
MRI065WCR1A Sungai Janggut West-Central
MRI067WCR1A Sungai Janggut West-Central
MRI068WCR1A Sungai Janggut West-Central
MRI069WCR1A Sungai Janggut West-Central
MRI070WCR1A Sungai Janggut West-Central
MRI072WCR1A Sungai Janggut West-Central
MRI073WCR1A Sungai Janggut West-Central
MRI077WCR1A Sungai Janggut West-Central
MRI079WCR1A Sungai Janggut West-Central
MRI081WCR1A Sungai Janggut West-Central
MRI148WND1A Pantai Kok West-North
MRI149WND1A Pantai Kok West-North
MRI150WND1A Pantai Kok West-North
MRI151WND1A Pantai Kok West-North
MRI152WND1A Pantai Kok West-North
MRI162WCD1A Sungai Janggut West-Central
MRI163WCD1A Sungai Janggut West-Central
MRI164WCD1A Sungai Janggut West-Central
MRI165WCD1A Sungai Janggut West-Central
MRI166WCD1A Sungai Janggut West-Central
MRI167WCD1A Sungai Janggut West-Central
MRI168WCD1A Sungai Janggut West-Central
MRI169WCD1A Sungai Janggut West-Central
MRI170WCD1A Sungai Janggut West-Central
MRI171WCD1A Sungai Janggut West-Central
MRI178ECD1A Kg. Cempaka East-Central
MRI179ECD1A Kg. Cempaka East-Central
MRI180ECD1A Kg. Cempaka East-Central
MRI181ECD1A Kg. Cempaka East-Central
MRI183ECD1A Kg. Cempaka East-Central
MRI186ECD1A Kg. Cempaka East-Central

124
MRI188ECD1A Kg. Cempaka East-Central
MRI189ECD1A Kg. Cempaka East-Central
MRI190ECD1A Kg. Cempaka East-Central
MRI191ECD1A Kg. Cempaka East-Central
MRI192ECD1A Kg. Cempaka East-Central
MRI193ECD1A Kg. Cempaka East-Central
MRI194ECD1A Kg. Cempaka East-Central
MRI195ECD1A Kg. Cempaka East-Central
MRI196ECD1A Kg. Cempaka East-Central
MRI197ECD1A Kg. Cempaka East-Central
MRI205END1A Pantai Sabak East-North
MRI206END1A Pantai Sabak East-North
MRI207END1A Pantai Sabak East-North
MRI208END1A Pantai Sabak East-North
MRI209END1A Pantai Sabak East-North
MRI210END1A Pantai Sabak East-North
MRI211END1A Pantai Sabak East-North
MRI212END1A Pantai Sabak East-North
MRI213END1A Pantai Sabak East-North
MRI214END1A Pantai Sabak East-North
MRI215END1A Pantai Sabak East-North
MRI216END1A Pantai Sabak East-North
MRI217END1A Pantai Sabak East-North
MRI218END1A Pantai Sabak East-North
MRI219END1A Pantai Sabak East-North
MRI220END1A Pantai Sabak East-North
MRI221END1A Pantai Sabak East-North
MRI222END1A Pantai Sabak East-North
MRI223END1A Pantai Sabak East-North
MRI224END1A Pantai Sabak East-North
MRI225END1A Pantai Sabak East-North
MRI226END1A Pantai Melawi East-North
MRI227END1A Pantai Melawi East-North
MRI228END1A Pantai Melawi East-North
MRI229END1A Pantai Melawi East-North
MRI230END1A Pantai Melawi East-North
MRI237WNR1A Balik Pulau West-North
MRI238WNR1A Balik Pulau West-North
MRI239WNR1A Balik Pulau West-North
MRI240WNR1A Balik Pulau West-North
MRI241WNR1A Balik Pulau West-North
MRI242WNR1A Balik Pulau West-North
MRI243WNR1A Balik Pulau West-North
MRI244WNR1A Balik Pulau West-North
MRI245WNR1A Balik Pulau West-North
MRI246WNR1A Balik Pulau West-North
MRI247WNR1A Balik Pulau West-North
MRI248WNR1A Balik Pulau West-North

125
MRI249WNR1A Balik Pulau West-North
MRI250WNR1A Balik Pulau West-North
MRI251WNR1A Balik Pulau West-North
MRI252WNR1A Balik Pulau West-North
MRI253WNR1A Balik Pulau West-North
MRI254WNR1A Balik Pulau West-North
MRI255WNR1A Balik Pulau West-North
MRI256WNR1A Balik Pulau West-North
MRI257WNR1A Balik Pulau West-North
MRI258WNR1A Balik Pulau West-North
MRI259WNR1A Balik Pulau West-North
MRI260WNR1A Balik Pulau West-North
MRI261WNR1A Balik Pulau West-North
MRI262WNR1A Balik Pulau West-North
MRI263WNR1A Balik Pulau West-North
MRI264WNR1A Balik Pulau West-North
MRI265WNR1A Balik Pulau West-North
MRI266WNR1A Balik Pulau West-North
MRI267WNR1A Balik Pulau West-North
MRI270WNR1A Balik Pulau West-North
MRI271WNR1A Balik Pulau West-North
Appendix E: Guide for using tpsUtil
1. Place all images into a folder.
2. Open tpsUtil.
3. Click on “Select an operation” and choose “Build tps file” from the drop-down list.
4. Under Input Directory, click on the “Input” button.
5. A dialog box will pop up. Browse to the directory where the images are stored.
6. Double-click on any one image in that directory. The dialog box will close.
7. Under Output File, click on the “Output” button.
8. A dialog box will pop up. Browse to the directory where the tps file is to be stored.
9. Enter a name. Make sure the file type is “.TPS”
10. Under Actions, click on the “Setup” button.
11. Make sure all the images in the directory are relevant to the project.
12. Click on the “Create” button. A tps file should now be created.

126
Appendix F: How to use tpsDig for digitization
1. Open tpsDig
2. Open the tps file (File > Input Source > File...). A dialogue box will pop up. Browse
to the directory where the tps file was store and choose the file. All images should be
loaded and appeared on tpsDig.
To Set Scale
1. Go to Options > Set Scale.
2. In the pop-up window enter the length of your scale (e.g., if the scale in the image is
10mm, enter 10mm).
3. In the image, place the cursor on one end of the scale in the image. Left-click, then
move the arrow to the other end and left-click again.
4. The scale window should now show the scale factor.
5. Click on the “Ok” button.
To Place Landmarks
1. Select Modes > Digitize landmarks.
2. Left-click to place a landmark. All landmarks must be placed in sequence.
Landmarks can be moved or deleted if mistakes are made.
3. Once all the landmarks for the image are placed, save the landmark data. Then move
on to the next image and repeat the step. Make sure the scale is set again for every
image.
To Save Landmark Data
1. Click on File > Save data > Save > Overwrite. Make sure the file is save after each
image is digitized to avoid losing of data.
Appendix G: 25 specimens used for Procrustes ANOVA
Specimen Sites Area
MRI001 Sungai Janggut West-Central
MRI002 Sungai Janggut West-Central
MRI067 Sungai Janggut West-Central
MRI070 Sungai Janggut West-Central
MRI148 Pantai Kok West-North
MRI162 Sungai Janggut West-Central

127
MRI164 Sungai Janggut West-Central
MRI188 Kg. Cempaka East-Central
MRI205 Pantai Sabak East-North
MRI208 Pantai Sabak East-North
MRI211 Pantai Sabak East-North
MRI214 Pantai Sabak East-North
MRI218 Pantai Sabak East-North
MRI226 Pantai Melawi East-North
MRI227 Pantai Melawi East-North
MRI229 Pantai Melawi East-North
MRI239 Balik Pulau West-North
MRI244 Balik Pulau West-North
MRI246 Balik Pulau West-North
MRI249 Balik Pulau West-North
MRI251 Balik Pulau West-North
MRI254 Balik Pulau West-North
MRI259 Balik Pulau West-North
MRI260 Balik Pulau West-North
MRI271 Balik Pulau West-North
Appendix H: Guide for using MorphoJ
Create a new tps project
1. Click on (File > Create New Project). A dialogue box will pop up. Enter the name of
the project and the name of the dataset.
2. Select the tps file type.
3. Choose the tps file created from tpsDigs.
4. Click on the “Create Dataset” button. The dataset is now created.
Perform Procrustes Fit
1. Click on the (Prelimineries menu > New Procrustes Fit)
2. Select “Align by principal axes”
3. Click on the “Perform Procrustes Fit” button
Generate Covariance Matrix
1. Click on the “Generate Covariance Matrix” button
2. A dialogue box will pop up. Click on the “Execute” button

128
Create Classifier
1. To create a classifier, Click on the (Preliminaries > Extract New Classifier From ID
String)
2. A dialogue box will pop up.
3. Enter a name for the new classifier
4. Indicate the position of the first and last character of the classifier. For example, if the
name of the specimen is MRI001WC, where WC is the classifier, then the First
character is 7 and the Last character is -1. In this analysis, these two classifiers were
created:
a. Individual (1, -1)
b. Locality (7, -1)
5. Click on the “Execute” button to produce the classifier
7. Classifier may also be edited from Preliminaries > Edit Classifier
Principal Component Analysis
1. To perform Principal Component Analysis, click on the (Variation menu > Principal
Component Analysis)
2. There are three different sub tabs under the Graphic tab: PC shape changes,
Eigenvalues and PC scores
a. PC shape changes tab shows the shape changes associated with each of the
PCs (eigenvectors).
b. Eigenvalues tab contains a graphs that shows the percentages of total
variance for which the PCs are accounted for
c. PC scores tabs contains a scatter plot of the PC scores
Canonical Variate Analysis
1. To perform Canonical Variate Analysis, click on the (Comparison menu > Canonical
Variate Analysis)
2. A dialogue box will pop up. Enter a name for the analysis
3. Choose a “Dataset” and “Data type”
4. Choose the classifier in “Classification variable(s) to use for grouping:”
5. Checked on the “Permutation test for pairwise distances” check box

129
6. Click on the “Execute” button to perform the analysis
7. Under the CVA tabs, two sub tabs will be generated: CV shape changes and CV
scores
a. The CV shape changes tab shows the shape changes associated with the
canonical variates (CVs). Right click anywhere on the screen will invoke a
pop up menu which can be used to change the properties, such as the scale
factor, the type of graph and the orientation of the shape, or to export the
graph as an SVG file.
b. The CV scores tab contains the scatter plots of the CV scores. If there are
only two groups, the graph will be a histogram instead of a scatter plot.
Right click anywhere on the screen will invoke a pop up menu which can be
used to change the properties, such as too choose the CVs to be displayed, to
color the dots by group membership according to the classifiers in the
dataset, to change other options or to export the graph as an SVG file.
To save the project
1. Click on the File menu > Save project as > and enter a name for the project which
should ends with .morphoj
Appendix I: To build and import outline in tps file into MorphoJ
1. Place all of the images into the same folder.
2. Open tpsUtil (Start > All Programs > tps > tpsUtil)
3. Click on “Select an operation” and choose “Build tps file” from the drop-down list.
4. Select the input directory:
a. Click “Input”
b. Find your directory of images
c. Double-click one image in that directory
5. Name your output file:
a. Click “Output”
b. Enter a name that ends in “.tps”.
6. Build the tps file
a. Click “Setup”
b. Checked images will be used to build your tps file. Check/uncheck as needed.
c. Click “Create” to create the tps file

130
7. Open tpsDig (Start > All Programs > tps > tpsDig)
8. Open the raw tps file (File > Input Source > File...)
9. Place landmarks. Only one specimen is needed to do the landmark and save it. After
landmark, point the cursor to “Outline” to create outline and save it.
10. Remove all the decimal point and zero. Between them must separate by “Tab”
space. Below is the result of the outline file used in this study:
0 106 363
0 86 336
0 125 287
0 165 247
0 178 198
0 237 203
0 274 202
0 269 230
0 253 248
0 279 239
0 310 249
0 296 275
0 291 325
0 242 332
0 184 360
0 127 385
1 106 363
1 95 351
1 86 336
2 86 336
2 100 316
2 114 299
2 125 287
3 125 287
3 144 268
3 160 253
3 165 247
4 165 247
4 172 219
4 174 206
4 178 198
5 178 198
5 189 191
5 203 189
5 216 191
5 228 197
5 237 203
6 237 203
131
6 246 198
6 255 196
6 264 197
6 274 202
7 274 202
7 278 207
7 278 216
7 276 224
7 269 230
8 269 230
8 264 234
8 257 239
8 250 244
8 253 248
9 253 248
9 257 252
9 263 248
9 271 244
9 279 239
10 279 239
10 289 238
10 300 241
10 310 249
11 310 249
11 312 256
11 310 264
11 304 271
11 296 275
12 296 275
12 297 282
12 301 293
12 301 302
12 300 311
12 291 325
13 291 325
13 280 329
13 271 330
13 258 330
13 242 332
14 242 332
14 224 339
14 209 348
14 184 360
15 184 360
15 170 367
15 147 376
132
15 127 385
16 127 385
16 114 373
16 106 363
11. In MorphoJ, open (File > Import outline file > and choose the outline tps file)
Appendix J: List of 146 specimens collected
Catalog
No.
Species Date Site GPS Method
MRI124 Rhopilema
esculentum
02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI125 Rhopilema
esculentum
02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI126 Cyanea sp. 02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI127 Cyanea sp. 02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI128 Cyanea sp. 02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI129 Phyllorhiza
punctata
02-Mar-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI130 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI131 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI132 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI133 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI134 Acromitus
flagellatus
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI135 Lychnorhiza
malayensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI136 Lychnorhiza
malayensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI137 Lychnorhiza
malayensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI138 Lychnorhiza
malayensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI139 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI140 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI141 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI142 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net

133
MRI143 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI144 Phyllorhiza
punctata
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI146 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI147 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI148 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI149 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI150 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI151 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI152 Chrysaora
chinensis
14-Apr-14 Pantai Kok N06.34869°,
E99.64806°
Dip Net
MRI153 Versuriga
anadyomene
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI154 Chrysaora
chinensis
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI155 Acromitus
flagellatus
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI156 Rhopilema
hispidum
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI157 Rhopilema
hispidum
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI158 Rhopilema
hispidum
15-Apr-14 Kilim N06.4766362°,
E99.8042679°
Dip Net
MRI159 Cyanea sp. 29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI160 Cyanea sp. 29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI161 Cyanea sp. 29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI162 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI163 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI164 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI165 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI166 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI167 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI168 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI169 Chrysaora 29-Apr-14 Sungai Janggut N03.16916°, Bag Net

134
chinensis E101.29833°
MRI170 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI171 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI172 Chrysaora
chinensis
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI173 Phyllorhiza
punctata
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI174 Phyllorhiza
punctata
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI175 Phyllorhiza
punctata
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI176 Versuriga
anadyomene
29-Apr-14 Sungai Janggut N03.16916°,
E101.29833°
Bag Net
MRI178 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI179 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI180 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI181 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI182 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI183 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI184 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI185 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI186 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI187 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI188 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI189 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI190 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI191 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI192 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI193 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI194 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI195 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI196 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
135
MRI197 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI198 Chrysaora
chinensis
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI199 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI200 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI201 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI202 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI203 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI204 Lobonemoides
robustus
26-Jun-14 Kampung Chempaka N03.74365°,
E103.32847°
Dip Net
MRI205 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI206 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI207 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI208 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI209 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI210 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI211 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI212 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI213 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI214 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI215 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI216 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI217 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI218 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI219 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI220 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI221 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI222 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI223 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net

136
MRI224 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI225 Chrysaora
chinensis
20-Jul-14 Pantai Sabak N06.13712°,
E102.36976°
Dip Net
MRI226 Chrysaora
chinensis
21-Jul-14 Pantai Melawi N05.99612°,
E102.43714°
Dip Net
MRI227 Chrysaora
chinensis
21-Jul-14 Pantai Melawi N05.99612°,
E102.43714°
Dip Net
MRI228 Chrysaora
chinensis
21-Jul-14 Pantai Melawi N05.99612°,
E102.43714°
Dip Net
MRI229 Chrysaora
chinensis
21-Jul-14 Pantai Melawi N05.99612°,
E102.43714°
Dip Net
MRI230 Chrysaora
chinensis
21-Jul-14 Pantai Melawi N05.99612°,
E102.43714°
Dip Net
MRI231 Cyanea sp. 10-Oct-14 Klang Power Station N03.3229°,
E101.30108°
Bag Net
MRI232 Cyanea sp. 10-Oct-14 Klang Power Station N03.3229°,
E101.30108°
Bag Net
MRI233 Cyanea sp. 10-Oct-14 Klang Power Station N03.3229°,
E101.30108°
Bag Net
MRI234 Phyllorhiza
punctata
13-Oct-14 Balik Puau N05.53972°,
E100.34888°
Fish
Net
MRI235 Lychnorhiza
malayensis
13-Oct-14 Balik Puau N05.53972°,
E100.34888°
Fish
Net
MRI236 Lychnorhiza
malayensis
13-Oct-14 Balik Puau N05.53972°,
E100.34888°
Fish
Net
MRI237 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI238 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI239 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI240 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI241 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI242 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI243 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI244 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI245 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI246 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI247 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI248 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI249 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI250 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net

137
MRI251 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI252 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI253 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI254 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI255 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI256 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI257 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI258 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI259 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI260 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI261 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI262 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI263 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI264 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI265 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI267 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI268 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI269 Phyllorhiza
punctata
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI270 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
MRI271 Chrysaora
chinensis
14-Dec-14 Balik Puau N05.53972°,
E100.34888°
Dip Net
Appendix K: 44 Specimens used for morphological description. M indicates specimens
used from museum.
Catalog no. Species Museum
MRI 2 C. chinensis M
MRI 65 C. chinensis M
MRI 163 C. chinensis
MRI 163 C. chinensis

138
MRI 171 C. chinensis
MRI 208 C. chinensis
MRI 262 C. chinensis
MRI 190 C. chinensis
MRI 55 Cyanea sp. M
MRI 56 Cyanea sp. M
MRI 95 Cyanea sp. M
MRI 96 Cyanea sp. M
MRI 97 Cyanea sp. M
MRI 127 Cyanea sp.
SJG 10 Cyanea sp. M
MRI 87 R. esculentum M
MRI 98 R. esculentum M
IND 3 R. esculentum M
MRI 13 R. hispidum M
MRI 89 R. hispidum M
IND 9 R. hispidum M
IND 10 R. hispidum M
SJ 130 R. hispidum M
MRI 204 L. robustus
IND 6 L. robustus M
IND 7 L. robustus M
IND 8 L. robustus M
MRI 153 V. anadyomene
MRI 176 V. anadyomene
MRI 14 P. punctata M
MRI 131 P. punctata
MRI 139 P. punctata
IND 2 P. punctata M
IND 11 P. punctata M
IND 14 P. punctata M
MRI 35 L. malayensis M
MRI 262 L. malayensis M
MRI 37 L. malayensis M
MRI 38 L. malayensis M
MRI 39 L. malayensis M
IND 13 L. malayensis M
MRI 21 A. flagellatus M
MRI 23 A. flagellatus M
MRI 24 A. flagellatus M
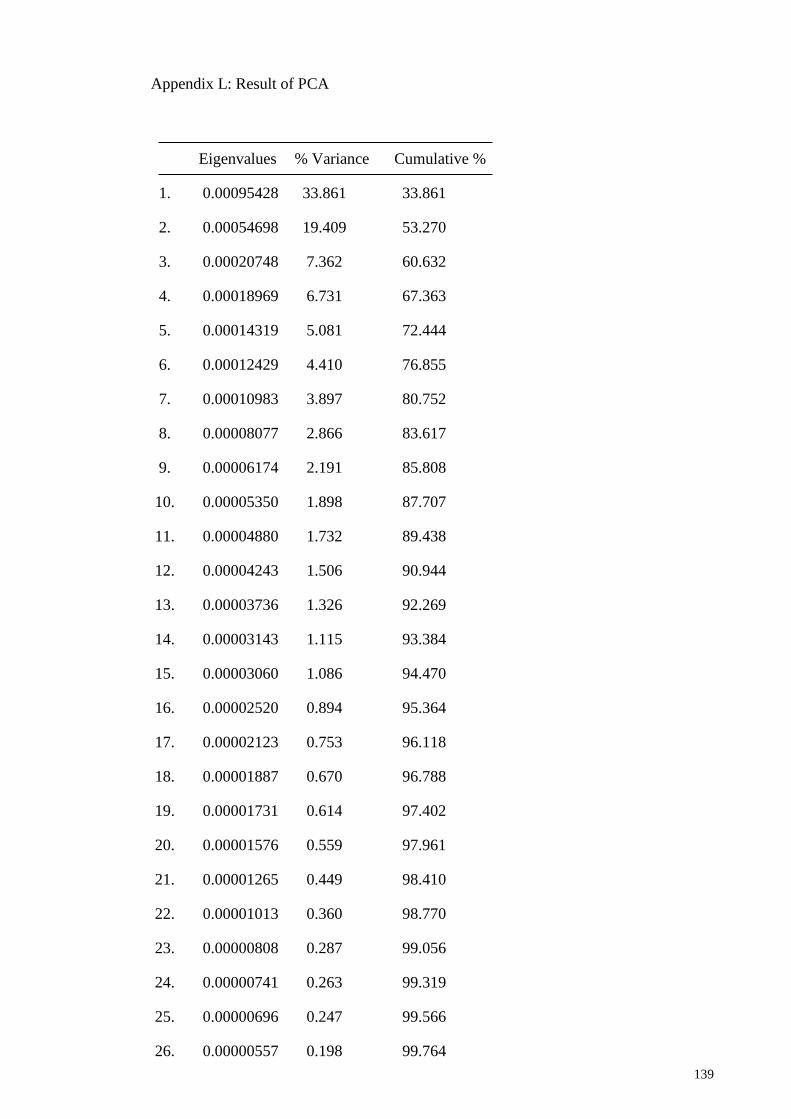
139
Appendix L: Result of PCA
Eigenvalues % Variance Cumulative %
1. 0.00095428 33.861 33.861
2. 0.00054698 19.409 53.270
3. 0.00020748 7.362 60.632
4. 0.00018969 6.731 67.363
5. 0.00014319 5.081 72.444
6. 0.00012429 4.410 76.855
7. 0.00010983 3.897 80.752
8. 0.00008077 2.866 83.617
9. 0.00006174 2.191 85.808
10. 0.00005350 1.898 87.707
11. 0.00004880 1.732 89.438
12. 0.00004243 1.506 90.944
13. 0.00003736 1.326 92.269
14. 0.00003143 1.115 93.384
15. 0.00003060 1.086 94.470
16. 0.00002520 0.894 95.364
17. 0.00002123 0.753 96.118
18. 0.00001887 0.670 96.788
19. 0.00001731 0.614 97.402
20. 0.00001576 0.559 97.961
21. 0.00001265 0.449 98.410
22. 0.00001013 0.360 98.770
23. 0.00000808 0.287 99.056
24. 0.00000741 0.263 99.319
25. 0.00000696 0.247 99.566
26. 0.00000557 0.198 99.764

140
27. 0.00000399 0.141 99.905
28. 0.00000267 0.095 100.000
Related Documents











