Biol. Chem., Vol. 387, pp. 451–460, April 2006 • Copyright by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2006.060 2006/302 Article in press - uncorrected proof Monomeric and dimeric GDF-5 show equal type I receptor binding and oligomerization capability and have the same biological activity Christina Sieber 1,a , Frank Plo ¨ ger 2,a , Raphaela Schwappacher 1 , Rolf Bechtold 2 , Michael Hanke 2 , Shinji Kawai 3 , Yoshifumi Muraki 4 , Mieko Katsuura 5 , Michio Kimura 5 , Maya Mouler Rechtman 6 , Yoav I. Henis 6 , Jens Pohl 2, * and Petra Knaus 1, * 1 Institut fu ¨ r Chemie/Biochemie, Freie Universita ¨ t Berlin, Thielallee 63, D-14195 Berlin, Germany 2 Biopharm GmbH, Czernyring 22, D-69115 Heidelberg, Germany 3 Department of Oral Frontier Biology Osaka University, Graduate School of Dentistry, 1-8 Yamadaoka, Suita- Osaka 565-0871, Japan 4 Therapeutic Area-Oncology, Clinical Development, Scientific Affairs, Aventis Pharma Ltd., Sanofi-Aventis Group, Tokyo Opera City Tower 3-20-2, Nishi shinjyuku, Shinjyuku-ku, Tokyo 163-1488, Japan 5 DDS Institute, The Jikei University School of Medicine, 3-25-8, Nishi-Shinbashi, Minato, Tokyo 105-8461, Japan 6 Department of Neurobiochemistry, Faculty of Life Sciences, Tel Aviv University,Tel Aviv 69978, Israel * Corresponding authors e-mail: [email protected]; [email protected] Abstract Growth and differentiation factor 5 (GDF-5) is a homo- dimeric protein stabilized by a single disulfide bridge between cysteine 465 in the respective monomers, as well as by three intramolecular cysteine bridges within each subunit. A mature recombinant human GDF-5 var- iant with cysteine 465 replaced by alanine (rhGDF-5 C465A) was expressed in E. coli, purified to homogeneity, and chemically renatured. Biochemical analysis showed that this procedure eliminated the sole interchain disul- fide bond. Surprisingly, the monomeric variant of rhGDF- 5 is as potent in vitro as the dimeric form. This could be confirmed by alkaline phosphatase assays and Smad reporter gene activation. Furthermore, dimeric and monomeric rhGDF-5 show comparable binding to their specific type I receptor, BRIb. Studies on living cells showed that both the dimeric and monomeric rhGDF-5 induce homomeric BRIb and heteromeric BRIb/BRII oligomers. Our results suggest that rhGDF-5 C465A has the same biological activity as rhGDF-5 with respect to binding to, oligomerization of and signaling through the BMP receptor type Ib. These authors contributed equally to this work a Keywords: BMP; BMP receptor; GDF-5; Smad signaling; TGF-b superfamily. Introduction The transforming growth factor b (TGF-b) superfamily of cytokines consists of more than 30 mammalian family members, which are subgrouped into TGF-bs, activins, bone morphogenetic proteins (BMPs), growth and differ- entiation factors (GDFs), and several more distant rela- tives (Kingsley, 1994). Members of this superfamily have a significant impact on many different cellular functions, such as cell differentiation, proliferation, homeostasis and apoptosis (Shi and Massague, 2003). These processes strictly depend on extracellular signals, which cells read differently, depending on their molecular constitution. This allows growth factors to affect the fate of single cells during early development, as well as complex tissue in the adult organism (Piek et al., 1999; Massague, 2000). Proteins of the TGF-b superfamily are relevant for a wide range of medical applications, including promotion of cell proliferation, tissue formation and regeneration, such as wound healing. They show high amino acid homology in the mature part of the protein, which has been conserved throughout evolution. In particular, seven cysteines are highly conserved. Members of the TGF-b superfamily exist as homo- or heterodimers (Cheifetz et al., 1987) that are stabilized by a single disulfide bond. This disulfide bond is mediated by one specific, con- served in most proteins, cysteine residue on each of the respective monomers (Daopin et al., 1992; Schlunegger and Grutter, 1992). Up to now, the dimerization of two monomers was considered to be indispensable for the biological activity of the ligand. It was proposed that the ligand dimer is important for dimerization of its receptors. Structural analyzes of BMP-2 and its high-affinity recep- tor BMP receptor type Ia (BRIa), or TGF-b3 and its high- affinity receptor TbRII have shown that one dimeric ligand interacts with two receptor chains (Kirsch et al., 2000; Hart et al., 2002; Greenwald et al., 2003). This was confirmed by biosensor measurements after immobilizing the extracellular domains of receptors (De Crescenzo et al., 2003; Sebald et al., 2004). Binding of TGF-b/BMP proteins to their high-affinity receptor leads to the recruit- ment of a second (low-affinity) receptor into a hetero- oligomeric complex. Both receptor types (I and II) are needed for further signal transduction. Besides this sequential binding mode, it was shown that BMP-2 binds to preformed heteromeric receptor complexes composed of both type I and type II receptors (Gilboa et al., 2000; Nohe et al., 2002). Extensive mutagenesis analyzes of BMP-2 have revealed that two distinct epitopes deter- Unauthenticated Download Date | 4/12/16 5:21 PM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biol. Chem., Vol. 387, pp. 451–460, April 2006 • Copyright � by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2006.060
2006/302
Article in press - uncorrected proof
Monomeric and dimeric GDF-5 show equal type I receptorbinding and oligomerization capability and have the samebiological activity
Christina Sieber1,a, Frank Ploger2,a, RaphaelaSchwappacher1, Rolf Bechtold2, MichaelHanke2, Shinji Kawai3, Yoshifumi Muraki4,Mieko Katsuura5, Michio Kimura5, Maya MoulerRechtman6, Yoav I. Henis6, Jens Pohl2,* andPetra Knaus1,*1 Institut fur Chemie/Biochemie, Freie Universitat Berlin,Thielallee 63, D-14195 Berlin, Germany2 Biopharm GmbH, Czernyring 22, D-69115 Heidelberg,Germany3 Department of Oral Frontier Biology Osaka University,Graduate School of Dentistry, 1-8 Yamadaoka, Suita-Osaka 565-0871, Japan4 Therapeutic Area-Oncology, Clinical Development,Scientific Affairs, Aventis Pharma Ltd., Sanofi-AventisGroup, Tokyo Opera City Tower 3-20-2, Nishi shinjyuku,Shinjyuku-ku, Tokyo 163-1488, Japan5 DDS Institute, The Jikei University School of Medicine,3-25-8, Nishi-Shinbashi, Minato, Tokyo 105-8461,Japan6 Department of Neurobiochemistry, Faculty of LifeSciences, Tel Aviv University, Tel Aviv 69978, Israel
* Corresponding authorse-mail: [email protected]; [email protected]
Abstract
Growth and differentiation factor 5 (GDF-5) is a homo-dimeric protein stabilized by a single disulfide bridgebetween cysteine 465 in the respective monomers, aswell as by three intramolecular cysteine bridges withineach subunit. A mature recombinant human GDF-5 var-iant with cysteine 465 replaced by alanine (rhGDF-5C465A) was expressed in E. coli, purified to homogeneity,and chemically renatured. Biochemical analysis showedthat this procedure eliminated the sole interchain disul-fide bond. Surprisingly, the monomeric variant of rhGDF-5 is as potent in vitro as the dimeric form. This could beconfirmed by alkaline phosphatase assays and Smadreporter gene activation. Furthermore, dimeric andmonomeric rhGDF-5 show comparable binding to theirspecific type I receptor, BRIb. Studies on living cellsshowed that both the dimeric and monomeric rhGDF-5induce homomeric BRIb and heteromeric BRIb/BRIIoligomers. Our results suggest that rhGDF-5 C465A hasthe same biological activity as rhGDF-5 with respect tobinding to, oligomerization of and signaling through theBMP receptor type Ib.
These authors contributed equally to this worka
Keywords: BMP; BMP receptor; GDF-5; Smadsignaling; TGF-b superfamily.
Introduction
The transforming growth factor b (TGF-b) superfamily ofcytokines consists of more than 30 mammalian familymembers, which are subgrouped into TGF-bs, activins,bone morphogenetic proteins (BMPs), growth and differ-entiation factors (GDFs), and several more distant rela-tives (Kingsley, 1994). Members of this superfamily havea significant impact on many different cellular functions,such as cell differentiation, proliferation, homeostasis andapoptosis (Shi and Massague, 2003). These processesstrictly depend on extracellular signals, which cells readdifferently, depending on their molecular constitution.This allows growth factors to affect the fate of single cellsduring early development, as well as complex tissue inthe adult organism (Piek et al., 1999; Massague, 2000).
Proteins of the TGF-b superfamily are relevant for awide range of medical applications, including promotionof cell proliferation, tissue formation and regeneration,such as wound healing. They show high amino acidhomology in the mature part of the protein, which hasbeen conserved throughout evolution. In particular, sevencysteines are highly conserved. Members of the TGF-bsuperfamily exist as homo- or heterodimers (Cheifetz etal., 1987) that are stabilized by a single disulfide bond.This disulfide bond is mediated by one specific, con-served in most proteins, cysteine residue on each of therespective monomers (Daopin et al., 1992; Schluneggerand Grutter, 1992). Up to now, the dimerization of twomonomers was considered to be indispensable for thebiological activity of the ligand. It was proposed that theligand dimer is important for dimerization of its receptors.Structural analyzes of BMP-2 and its high-affinity recep-tor BMP receptor type Ia (BRIa), or TGF-b3 and its high-affinity receptor TbRII have shown that one dimericligand interacts with two receptor chains (Kirsch et al.,2000; Hart et al., 2002; Greenwald et al., 2003). This wasconfirmed by biosensor measurements after immobilizingthe extracellular domains of receptors (De Crescenzo etal., 2003; Sebald et al., 2004). Binding of TGF-b/BMPproteins to their high-affinity receptor leads to the recruit-ment of a second (low-affinity) receptor into a hetero-oligomeric complex. Both receptor types (I and II) areneeded for further signal transduction. Besides thissequential binding mode, it was shown that BMP-2 bindsto preformed heteromeric receptor complexes composedof both type I and type II receptors (Gilboa et al., 2000;Nohe et al., 2002). Extensive mutagenesis analyzes ofBMP-2 have revealed that two distinct epitopes deter-
UnauthenticatedDownload Date | 4/12/16 5:21 PM

452 C. Sieber et al.
Article in press - uncorrected proof
Figure 1 Analysis of rhGDF-5 and rhGDF-5 C465A by Westernblotting (A) and isoelectric focusing (B).(A) rhGDF-5 (lanes 1 and 2) and rhGDF-5 C465A (lanes 3 and 4)were separated by SDS gel electrophoresis with (lanes 1 and 3)and without 1% b-mercaptoethanol (lanes 2 and 4). Proteinswere transferred to a blotting membrane in the presence of10 mM DTT so that only reduced monomeric molecules weretransferred to the membrane. Marker sizes are given in kDa.(B) The rhGDF-5 (lane 1) and rhGDF-5 C465A (lane 2) proteinswere run on an immobilized pH gradient gel in the range 6.0–9.0and silver-stained.
mine the binding site for BRIa (high-affinity receptor) andBRII (low-affinity receptor) (Kirsch et al., 2000; Knaus andSebald, 2001). Since the binding site for BRIa is com-posed of residues from both monomers, it was suggest-ed that stabilization of the dimer by the intramoleculardisulfide bond is needed for this interaction. Heteromericmuteins, in which one BMP-2 monomer is defective ineither the type I or type II receptor epitope, further sug-gested that initial dimerization of BRIa by the ligand isessential for the induction of distinct signaling pathways(Knaus and Sebald, 2001; Nohe et al., 2002).
While most TGF-b/BMP proteins are active as disul-fide-linked dimers, three members of the superfamily,GDF-9, GDF-9B, and GDF-3, have a different pattern ofcysteine residues, lacking the conserved cysteine residuethat forms the disulfide link between the monomers.However, they are believed to form non-covalent dimersto achieve biological activity (McPherron and Lee, 1993;Kingsley, 1994). Mutations in activin A, as well as TGF-b1, replacing the cysteine responsible for the intramolec-ular bond with serine, were shown to affect thedimerization and led to monomeric forms with low bio-logical activity, despite high receptor affinity in the caseof mutant activin A (Amatayakul-Chantler et al., 1994;Husken-Hindi et al., 1994).
For the production of biologically active TGF-b/BMPproteins as therapeutic agents, extensive renaturationprocedures are necessary to guarantee accurately foldedligand dimers. Apart from the proper folding of eachmonomeric protein, correct cysteine bonding betweenthe two monomers is a critical step during the renatura-tion procedure. Thus, one aim of this study was to devel-op a recombinant human GDF-5 protein in a monomericform for which the biological activity essentially corre-sponded to that of the dimeric form.
The monomeric rhGDF-5 (rhGDF-5 C465A) describedhere shows that disruption of the interchain disulfidebridge does not affect the biological function of theprotein.
Results
Preparation of monomeric rhGDF-5 (rhGDF-5C465A)
Expression of rhGDF-5 and rhGDF-5 C465A was identi-cal in the E. coli strain used. Up to 90% of both proteinswas accumulated in inclusion bodies (Honda et al.,2000). Separation of renatured monomeric protein fromnon-renatured monomeric rhGDF-5 by reverse-phaseHPLC was possible by applying a very shallow gradientof 0.5%. The resulting purity of both rhGDF-5 and rh-GDF-5 C465A was greater than 90%, as determined bySDS-PAGE followed by silver staining (data not shown).Western blot analysis, performed using the monoclonalantibody P3/66 (O’Keeffe et al., 2004), revealed that bothmonomeric and dimeric rhGDF-5 are recognized by thisantibody (Figure 1A). rhGDF-5 and rhGDF-5 C465Amigrate at approximately 14 kDa under reducing condi-tions. Under non-reducing conditions, dimeric rhGDF-5migrates at approx. 25 kDa. rhGDF-5 C465A, on the oth-
er hand, migrates as a monomeric protein, but surpris-ingly slightly faster than under reducing condi-tions.
Verification by peptide mapping and isoelectricfocusing
Isoelectric focusing of the proteins rhGDF-5 and rhGDF-5 C465A (Figure 1B) displayed distinct bands. The rh-GDF-5 dimer protein gave a broad band with a pI ofapproximately 7.6, while rhGDF-5 C465A monomericprotein was focused at pI of approximately 7.5.
The substitution of cysteine by the more hydrophobicalanine at position 465 in rhGDF-5 C465A is clearlyshown by comparing the tryptic peptide maps of rhGDF-5 C465A and rhGDF-5 (Figure 2A). Tryptic peptide T11(nomenclature according to Figure 2A) comprises theamino acid at sequence position 465 and only T11 ofrhGDF-5 C465A elutes later than the T11 of rhGDF-5 (seeinsert in Figure 2B). All other tryptic peptides of rhGDF-5 C465A match the corresponding peptides of rhGDF-5with regard to retention time and mass (see Table 1), aswell as sequence (data not shown). The mass differenceof 141 Da found between T11 of rhGDF-5 and T11 ofrhGDF-5 C465A corresponds to the theoretical differenceof 137 Da between alkylated cysteine (pyridyl-ethylcys-teine) and alanine.
Solubility testing of rhGDF-5 C465A
The structural difference between rhGDF-5 and rhGDF-5C465A does not seem to have a significant impact ontheir solubility. Unlike other members of the TGF-bsuperfamily (for TGF-b3 see Jen et al., 2002), both mol-ecules are nearly insoluble at physiological pH valuesranging from pH 5.0 to 9.5 (Figure 3). The addition of150 mM NaCl does not enhance the solubility of eitherprotein (data not shown). Solubility of the ligands wasbest at pH 2. For further experiments, both rhGDF-5 andrhGDF-5 C465A were dissolved in 10 mM HCl, pH 2.
UnauthenticatedDownload Date | 4/12/16 5:21 PM

Activity of monomeric GDF-5 453
Article in press - uncorrected proof
Figure 2 Analysis of rhGDF-5 and rhGDF-5 C465A peptides.(A) Tryptic peptides of rhGDF-5 and rhGDF-5 C465A. Theoretical cleavage of rhGDF-5 and rhGDF-5 C465A by trypsin resulting intryptic peptides T1–T13. For readability, every other tryptic peptide is in gray and the single amino acid difference between rhGDF-5 and rhGDF-5 C465A at position 465 is highlighted.(B) Overlay of tryptic peptide maps of rhGDF-5 and rhGDF-5 C465A. Reversed-phase HPLC chromatograms showing the separationof tryptic digests of rhGDF-5 (lower trace) and rhGDF-5 C465A (upper trace); absorption (mAU) at 214 nm vs. time (min). Trypticfragments are labeled according to the amino acid sequence (see Figure 1) and were identified by sequencing (data not shown), aswell as by mass spectrometry (see Table 1). The tryptic peptide map of rhGDF-5 C465A is the same as the map of rhGDF-5, exceptfor peptide T11 (see insert), which contains amino acid position 465.
Table 1 Retention times and masses of the tryptic peptides found.
Tryptic peptide Retention time (min) Mass found (Da)
rhGDF-5 rhGDF-5 C465A rhGDF-5 rhGDF-5 C465A
T1* 27.30 27.27 662.4 663.0T3–4 16.02 16.02 487.2 487.3T5 18.41 18.39 375.1 374.9T7 16.38 16.37 470.0 470.8T8–9 42.45 42.38 957.8 957.7T9 44.68 44.62 829.2 829.1T10 77.35 77.30 3575.1 3568.7T11 61.61 62.28 3723.0 3581.6T12 74.06 74.04 1999.9 1993.9T13 41.37 41.31 1731.4 1730.9
*Detected as N-pyridyl-ethyl peptide.rhGDF-5 and rhGDF-5 C465A were digested with trypsin and separated by reversed-phase HPLC. The tryptic peptides were sequenced and the molecular masses weredetermined by mass spectrometry. The corresponding retention times and massesare indicated for each tryptic peptide.
Signaling induced by dimeric and monomeric GDF-5measured by alkaline phosphatase induction
It was shown that BRIb is the high-affinity type I receptorfor GDF-5 (Nishitoh et al., 1996). We used the GDF-5-responsive cell line ATDC5 to investigate the biologicalactivity of rhGDF-5 C465A in comparison to rhGDF-5. InATDC5 cells, it was shown that GDF-5 promotes the earlyphase of the chondrogenic response by inducing cellularcondensation, followed by cartilage nodule formation(Nakamura et al., 1999).
Here we demonstrate that rhGDF-5 and rhGDF-5C465A induce alkaline phosphatase activity in a dose-dependent manner (Figure 4). Similar results wereobtained using ROB-C26 cells (data not shown), which
were reported to express BRIb and to be sensitive toGDF-5 stimulation (Nishitoh et al., 1996).
Signaling induced by dimeric and monomeric GDF-5as measured in Smad-reporter gene response
As we have shown previously, rhBMP-2 activates theSmad pathway through BMP type I and type II receptors,which form heteromeric complexes prior to ligand bind-ing (Gilboa et al., 2000). To investigate whether mono-meric BMP-like growth factors such as rhGDF-5 C465Ainduce Smad signaling as efficiently as the dimeric form,we performed reporter gene assays in ATDC5 cells (Sam-mar et al., 2004). The binding of GDF-5 to its receptorsresults in the phosphorylation and activation of Smad1/
UnauthenticatedDownload Date | 4/12/16 5:21 PM
454 C. Sieber et al.
Article in press - uncorrected proof
Figure 3 Comparative solubility of dimeric rhGDF-5 and rh-GDF-5 C465A over the pH range 3.7–12.1.rhGDF-5 and rhGDF-5 C465A were dissolved in Na-acetate buf-fers ranging from pH 3.7 to 5.6 and in Na-carbonate buffersranging from pH 9.5 to 12.1. After centrifugation the supernatantwas quantified by reversed-phase HPLC. rhGDF-5 and rhGDF-5 C465A are nearly insoluble at physiological pH values.
Figure 4 Dose response of alkaline phosphatase activity inATDC5 cells.ATDC5 cells were stimulated with 1–75 nM rhGDF-5 (blacksquares) and rhGDF-5 C465A (white circles) for 72 h. Alkalinephosphatase was measured according to standard protocols.One absorbance unit at 405 nm corresponds to 1.5 nM p-nitro-phenol per minute per 104 cells wnM/(min=104 cells)x.
Figure 5 Smad-dependent reporter gene assay in ATDC5 cells.ATDC5 cells were transfected with the reporter gene constructpSBE-luc and pRL-Tk for reference. Following starvation, cellswere stimulated with 1–75 nM rhGDF-5 (black squares) or rh-GDF-5 C465A (white circles). After 24 h of stimulation, cell lysisand determination of luciferase activity were performed accord-ing to the manufacturer’s protocol.
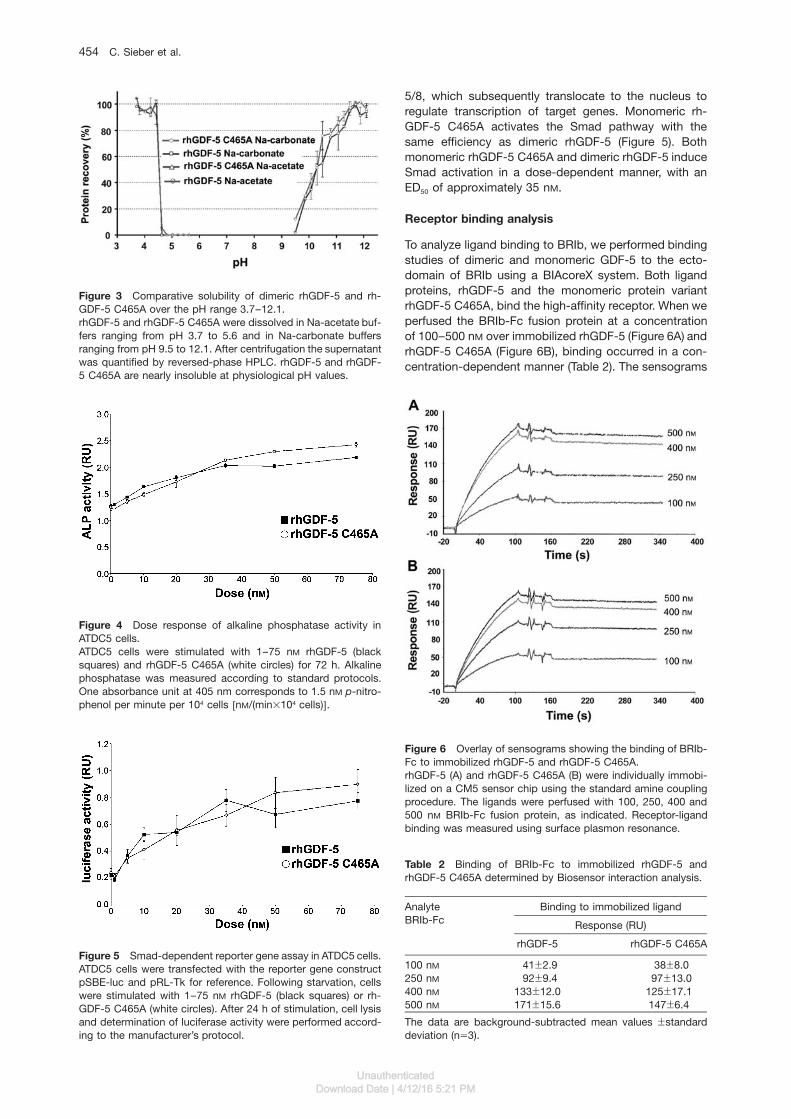
Table 2 Binding of BRIb-Fc to immobilized rhGDF-5 andrhGDF-5 C465A determined by Biosensor interaction analysis.
Analyte Binding to immobilized ligandBRIb-Fc Response (RU)
rhGDF-5 rhGDF-5 C465A
100 nM 41"2.9 38"8.0250 nM 92"9.4 97"13.0400 nM 133"12.0 125"17.1500 nM 171"15.6 147"6.4
The data are background-subtracted mean values "standarddeviation (ns3).
Figure 6 Overlay of sensograms showing the binding of BRIb-Fc to immobilized rhGDF-5 and rhGDF-5 C465A.rhGDF-5 (A) and rhGDF-5 C465A (B) were individually immobi-lized on a CM5 sensor chip using the standard amine couplingprocedure. The ligands were perfused with 100, 250, 400 and500 nM BRIb-Fc fusion protein, as indicated. Receptor-ligandbinding was measured using surface plasmon resonance.
5/8, which subsequently translocate to the nucleus toregulate transcription of target genes. Monomeric rh-GDF-5 C465A activates the Smad pathway with thesame efficiency as dimeric rhGDF-5 (Figure 5). Bothmonomeric rhGDF-5 C465A and dimeric rhGDF-5 induceSmad activation in a dose-dependent manner, with anED50 of approximately 35 nM.
Receptor binding analysis
To analyze ligand binding to BRIb, we performed bindingstudies of dimeric and monomeric GDF-5 to the ecto-domain of BRIb using a BIAcoreX system. Both ligandproteins, rhGDF-5 and the monomeric protein variantrhGDF-5 C465A, bind the high-affinity receptor. When weperfused the BRIb-Fc fusion protein at a concentrationof 100–500 nM over immobilized rhGDF-5 (Figure 6A) andrhGDF-5 C465A (Figure 6B), binding occurred in a con-centration-dependent manner (Table 2). The sensograms
UnauthenticatedDownload Date | 4/12/16 5:21 PM

Activity of monomeric GDF-5 455
Article in press - uncorrected proof
for monomeric and dimeric rhGDF-5 are comparable,with responses of the same order of magnitude. There-fore, we conclude that the C465A substitution in rhGDF-5, which results in monomerization of the protein, doesnot affect the binding to its high-affinity type I receptor.
GDF-5-induced BMP receptor oligomerizationstudied by immunofluorescence co-patching on livecells
We have recently developed a computerized quantitativeapproach to measure complex formation among mem-brane receptors situated in their native milieu (the plasmamembrane) by immunofluorescence co-patching (Lach-manovich et al., 2003). In this method (Gilboa et al., 1998,2000), two receptors with different epitope tags are co-expressed at the surface of live cells. Incubating the cellswith anti-tag IgGs in the cold (to eliminate possible endo-cytosis), one receptor is forced into micropatches by adouble layer of bivalent IgGs using a fluorescent (e.g.,coupled to Alexa488, green) secondary IgG. The co-expressed receptor, which carries a different tag, ispatched and labeled by primary IgG from another spe-cies and secondary IgGs coupled to another fluorophore(e.g., Alexa594, red). Receptors in mutual oligomers areswept into mutual micropatches, which appear yellowwhen the two CCD images are overlapped.
We have previously shown that the type I receptors ofBMP, BRIa and BRIb, reside at the cell surface as mono-meric and homodimeric oligomers prior to ligand binding(Gilboa et al., 2000; Nohe et al., 2002).
Here we compared the ability of different ligands(BMP-2, rhGDF-5 and rhGDF-5 C465A) to promote anincrease in the level of homomeric and heteromeric BMPreceptor complexes. Typical images obtained in thisseries of experiments are shown in Figure 7A–I; the aver-age data derived from studies on a large number of cellsare depicted in Figure 7J.
Figure 7 shows the actual co-patching levels betweenthe receptor pairs. As explained in detail earlier (Gilboaet al., 2000; Lachmanovich et al., 2003), in the case ofhomodimers, the percentage of homodimers is higherthan the percentage of co-patching by a factor of 3/2,because dimers containing identically tagged receptorsmay also form, but would not be swept into mutualpatches. The data in Figure 7 show that dimeric rhGDF-5 was as effective as BMP-2 in elevating the homo-olig-omerization of BRIb, but had a much weaker effect onthe homo-oligomerization of BRIa. On the other hand, thehomo-oligomerization of BRII, which exhibits low affinityto BMP-2 but higher affinity to rhGDF-5, was increasedmuch more by rhGDF-5. The monomeric derivative ofrhGDF-5, rhGDF-5 C465A, was very similar to rhGDF-5in terms of the induction of homo-oligomerization, exceptfor BRII, for which it was much less effective (Figure 7J).
Next we analyzed the hetero-oligomerization of BMPreceptors in response to the various ligands. As we haveshown previously, BMP receptors reside at the cell sur-face partially as preformed complexes composed of BRIaor BRIb with BRII (Gilboa et al., 2000; see Figure 7). WhileBRIa/BRII complex formation was significantly elevatedby rhBMP-2, only a weak increase in their hetero-oligo-merization was observed when dimeric rhGDF-5 was
applied (Figure 7). However, rhGDF-5 effectively incre-ased the level of heteromeric BRIb and BRII complexes,similar to rhBMP-2. These complexes resemble the BISColigomers (BMP-2 induced signaling complexes), whichwe have shown induces distinct Smad-independent sig-naling cascades following rhBMP-2 stimulation (Nohe etal., 2002). We therefore designated the rhGDF-5-inducedheteromeric complexes as GISCs (GDF-5-induced sig-naling complexes), as observed in Figure 7; these aremainly comprised of BRIb/BRII complexes. The mono-meric rhGDF-5 C465A has similar effects to those ofrhGDF-5 in inducing heteromeric oligomers of BRIa/BRII(no induction) and BRIb/BRII (induction), when comparedto rhGDF-5 (Figure 7).
Discussion
Bone development includes diverse processes such asthe formation of mesenchymal condensations at sites offuture skeletal elements, osteoblast and chondrocyte dif-ferentiation, and three-dimensional patterning. BMPsplay a pivotal role in these complex processes, and areinvolved in nearly all processes associated with skeletalmorphogenesis. Most BMPs are homodimeric proteinslinked by a single disulfide bridge, which transduce sig-nals via transmembrane serine/threonine kinase recep-tors to the nucleus, through both Smad and non-Smadpathways. The mode of receptor oligomerization hasbeen shown to be critical for the initiation of distinct sig-naling pathways (Nohe et al., 2002; Canalis et al., 2003).
The generation of heteromeric mutant BMP-2 (BMPmuteins) suggested that two BMP type II receptors areneeded for initiation of the Smad pathway, while twoBMP type I receptors are essential for ligand-inducedinduction of the MAPK pathway (Knaus and Sebald,2001; Nohe et al., 2002).
The structural homology of GDF-5 to BMP-2 impliesthat dimeric GDF-5 is needed for proper binding to andactivation of type I and type II receptors (Knaus andSebald, 2001; Schreuder et al., 2005). This is mainly sug-gested through studies showing that the type II receptoris recruited into a complex after ligand has bound to itshigh-affinity type I receptor (Nohe et al., 2002). A dimericligand seemed to be indispensable for signaling via thispathway. There is accumulating evidence suggesting thatnatural TGF-b heterodimers, such as TGF-b1/2, TGF-b2/3 (Ogawa et al., 1992), BMP-2/7, BMP-4/7 and the Dro-sophila homologs Decapentaplegic (Dpp) and Screw(Scw), exist that have more potent biological activity thanthe corresponding homodimers (Israel et al., 1996; Shim-mi et al., 2005).
In rhGDF-5 C465A, cysteine 465 was replaced by ala-nine to prevent formation of the disulfide bridge betweentwo rhGDF-5 monomers. In activin A and TGF-b1, forboth of which similar studies were conducted, the criticalcysteine was mutated to serine, probably owing to struc-tural considerations (Amatayakul-Chantler et al., 1994;Husken-Hindi et al., 1994). Unlike alanine, serine is apolar amino acid. To prevent changes in the charge dis-tribution, as well as the secondary and tertiary structure
UnauthenticatedDownload Date | 4/12/16 5:21 PM

456 C. Sieber et al.
Article in press - uncorrected proof
Figure 7 Co-patching of BMP type I and type II receptors upon stimulation with rhBMP-2, rhGDF-5 or rhGDF-5 C465A.COS7 cells were co-transfected transiently with pairs of receptors carrying different extracellular epitope tags, as indicated in thegraph. Live cells were labeled consecutively in the cold by a series of antibodies to mediate patching and fluorescent labeling, asdetailed in materials and methods. In the cases for which ligands were present (10 nM rhBMP-2, 10 or 100 nM rhGDF-5, or 100 nM
rhGDF-5 C465A), they were added at 48C 1.5 h prior to labeling with IgGs and included at the same concentration during subsequentincubation with primary antibody. The patching/labeling protocol results in HA-tagged receptor proteins labeled by Alexa594 (red),while myc-tagged receptors are labeled by Alexa488 (Greenwald et al., 2003). (A,D,G) Typical images of co-patching experiments oncells co-transfected with myc-BRIb and HA-BRIb. The images shown are without ligand (A), with 100 nM of rhGDF-5 (D) and with100 nM of rhGDF-5 C465A (G). The white boxes depict regions of interest (10=10 mm) that were analyzed for co-patching (B,E,H)and for randomized co-localization (C,F,I). Bars represent 20 mm (A) and 2 mm (C). (B,E,H) Zoomed images of region I in panels (A),(D) and (G), respectively. Using an algorithm written for Image Pro Plus, a green patch was defined as overlapping with a red patch(co-patched) if their centers of mass were separated by less than 0.2 mm, the resolution of the light microscope. Large patches (over5=5 pixels) were discarded by the program. (C,F,I) Zoomed control randomized images to evaluate random overlap (control foroccasional overlap owing to the high density of patches). Each random image was created from two adjacent regions of interest; thered channel of region I was merged with the unrelated green channel of region II. For a detailed explanation of this analysis, seeLachmanovich et al. (2003). (J) Averaged data (20–35 cells in each case) quantifying homo- and heteromeric complex formationbetween BMP receptors. After superposition of the green and red channels, the numbers of red (R), green (G) and overlapping (yellow,Y) patches were determined using the computerized analysis described above. The percentage co-patching (percentage of a onetagged receptor in mutual patches with the other receptor) is given by 100=wY/(YqR)x for the red-labeled receptors and by 100=wY/(YqG)x for the green-labeled receptors. Because these values were very close for each receptor pair, a single mean"SEM value isdepicted for each pair. The uncorrelated overlap (randomized control images), which was approximately 10% in all cases, wassubtracted, as it represents the percentage co-localization owing to occasional random overlap. The homomeric co-patching of BRIawas strongly enhanced (2.4-fold) by rhBMP-2 (p-10-21, Student’s t-test). The rhGDF-5 ligands were much less effective, with a small(1.4-fold) but significant (p-10-6) increase by rhGDF-5 and no significant increase by rhGDF-5 C465A (p)0.05). On the other hand,all three ligands markedly increased the BRIb homomeric co-patching (ca. 2.4-fold; p-10-14 in all cases). The effect of 10 nM rhGDF5on BRIb homo-dimerization was similar to that of 100 nM (data not shown). In the case of BRII, BMP-2 and rhGDF-5 C465A slightlybut significantly increased homomeric co-patching (ca. 1.3-fold; p-10-4), while rhGDF-5 increased this oligomerization much moreeffectively (1.9-fold; p-10-18). Heteromeric oligomerization of BRII with BRIa was strongly affected by rhBMP-2 (1.6-fold; p-10-15),while rhGDF-5 had a small but significant effect (1.4-fold; p-10-8) and rhGDF-5 C465A was ineffective (p)0.05). BRII/BRIb hetero-oligomerization was increased significantly and similarly by all three ligands (ca. 1.3-fold; p-10-5, Student’s t-test).
of rhGDF-5, we replaced cysteine 465 with the small andneutral amino acid alanine.
Since our results show that rhGDF-5 C465A is asactive as rhGDF-5, it is possible that upon receptor dime-rization, the monomeric ligand also dimerizes. As shownin Figure 1A, dimeric rhGDF-5 has a molecular weight ofapproximately 25 kDa under non-reducing conditions,whereas rhGDF-5 C465A, at least under these condi-tions, migrates faster than the reduced form of rhGDF-5,suggesting a monomeric form with a changed confor-mation without dimeric properties.
To investigate receptor oligomerization induced byrhGDF5 on live cells, receptor co-patching was per-formed on COS7 cells expressing epitope-tagged BMPreceptors as previously described (Gilboa et al., 2000;Nohe et al., 2002). The data on living cells showed onlyminor differences between rhGDF-5 and rhGDF-5 C465Afor inducing homomeric oligomers of BRIb and hetero-meric oligomers of BRIa/BRII and BRIb/BRII. In all cases,monomeric rhGDF-5 showed nearly the same activity,which hints at the possibility that it forms a dimer eitherin solution or upon dimerization of the receptors. Gel
UnauthenticatedDownload Date | 4/12/16 5:21 PM

Activity of monomeric GDF-5 457
Article in press - uncorrected proof
chromatography studies under non-dissociating condi-tions revealed distinct retention profiles for rhGDF-5C465A (retention at 11 min) and rhGDF-5 (retention at14.5 min), suggesting that rhGDF-5 C465A is monomericin solution (data not shown).
The monomeric rhGDF-5 C465A showed almost noinduction of homomeric oligomers of BRIa (Figure 7J),which is in accordance with its low affinity for this recep-tor (Nickel et al., 2005). BMP type II receptor (BRII) servesas the low-affinity receptor for BMP-2, while GDF-9, aGDF-5-related factor, was shown to bind BRII directly(Vitt et al., 2002). GDF-9 is important for ovarian follicu-logenesis, i.e., the development and release of anoocyte. GDF-9 lacks the critical cysteine and reflects anon-covalently linked dimeric ligand. Here we haveshown for the first time that dimeric rhGDF-5 causeshomo-dimerization of BRII (Figure 7; see ‘BRIIqBRII’),suggesting binding to BRII, which was not observed forrhBMP-2 (Nohe et al., 2002). Binding to and oligomeri-zation of BRIb by rhGDF-5 C465A is the same whencompared to dimeric rhGDF-5, as shown by BIAcoremeasurements of ligand binding to BRIb (Figure 6) andco-patching of receptors on live cells (Figure 7). Thisidentical activity is also reflected by functional assays inATDC5 cells. Both the Smad-dependent and -independ-ent signaling pathways (Figures 4 and 5) are induced bythe monomeric and dimeric forms of GDF-5. This sug-gests that both pathways do not require GDF-5-mediat-ed BRII dimerization, which is defective in rhGDF-5C465A-treated cells (Figure 7).
Compared to its dimeric form, activity was significantlyreduced for monomeric activin A and TGF-b1 (Amataya-kul-Chantler et al., 1994; Husken-Hindi et al., 1994). Con-trary to these results, we found that rhGDF-5 C465Aretains comparable biological activity to rhGDF-5.
It has been shown that BRII binds to the so-called‘knuckle’ epitope of each BMP-2 monomer, while BRIabinds to the ‘wrist’ epitope, composed of residues fromboth BMP-2 monomers (Kirsch et al., 2000; Knaus andSebald, 2001). Our data suggest that non-covalent bind-ing of rhGDF-5 C465A may be enough to form a dimerupon BRIb binding, along with BRIb homo-oligomeriza-tion (Figure 7, ‘BRIbqBRIb’). This event is followed byhetero-oligomerization of ligand-bound BRIb with BRII(Figure 7, ‘BRIbqBRII’). However, dimerization of rhGDF-5 C465A either does not occur when bound to BRII, oris not strong enough to induce homo-oligomerization ofBRII, as observed for the dimeric form of rhGDF-5 (Figure7, ‘BRIIqBRII’).
Both recombinant proteins, rhGDF-5 and rhGDF-5C465A, are equally soluble. The tendency of rhGDF-5C465A to precipitate upon dilution was already observedduring direct refolding (Honda et al., 2000). Structuralanalysis exploited the charge distribution of the rhGDF-5 molecule surface, which varies considerably fromBMP-2, BMP-7 and TGF-b2 (Honda et al., 2000). Thismight explain the different receptor specificity and affinityof rhGDF-5 when compared to rhBMP-2 and rhBMP-7,but also explains the different solubility profiles.
In ATDC5 cells, rhGDF-5 and rhGDF-5 C465A showedinduction of alkaline phosphatase and Smad reportergene activity in a dose-dependent manner. Taken togeth-
er, these results indicate that rhGDF-5 and rhGDF-5C465A signal via identical pathways.
More recently, we identified distinct GDF-5 mutationswithin the type I receptor binding interface that cause theskeletal malformations brachydactyly and symphalan-gism (Seemann et al., 2005). GDF-5(L441P) is an inactiveligand because of the loss of BRIb binding, while GDF-5(R438L) shows increased biological activity caused bybinding to both BRIb and BRIa receptors.
Production of recombinant TGF-b superfamily mem-bers for therapeutic purposes is usually costly and time-consuming, since they are not readily obtainable inbiologically active and sufficiently pure form in prokary-otes. A limiting factor is the separation of the active rena-tured dimeric protein from the inactive monomeric formby size exclusion.
In this study, we showed that it is possible to producea monomeric form of rhGDF-5, which has the full biolog-ical activity of the dimeric form. This finding will facilitatelarge-scale production of biologically active rhGDF5C465A at a reasonable cost.
Materials and methods
Purification of monomeric rhGDF-5 C465A fromE. coli cells
The recombinant form of mature human GDF-5 is a homodimerof 119 amino acid residues per monomer, with an approximatemolecular mass of 25 kDa (Honda et al., 2000). The first aminoacid of mature rhGDF-5 corresponds to proline at position 383of the precursor protein (Hotten et al., 1996). The mature part ofrhGDF-5 was mutated by converting cysteine to alanine at posi-tion 465 of the precursor molecule, which corresponds to cys-teine 83 of the mature protein. For this purpose, the nucleotidesat position 2032 and 2033 were converted from TG to GC. Thesubstitution of the nucleotides was performed using PCR muta-genesis and verified by sequencing.
The coding sequence of rhGDF-5 C465A was subcloned intothe plasmid pKOT245, resulting in the sequence coding for ami-no acids 383–501 of the precursor protein. Subsequently, rh-GDF-5 C465A was expressed in the E. coli strain W3110M (K12origin) as previously described (Honda et al., 2000). Purificationwas carried out on a reverse-phase column (Nucleosil 300-7C4,Macherey-Nagel, Duren, Germany) with a 0–50% buffer B (90%acetonitrile, 0.1% TFA) gradient in buffer A (0.1% TFA in water)within 50 min (flow rate 2 ml/min). The fractions containingrhGDF-5 C465A were pooled, lyophilized, and stored at -708C.rhGDF-5 C465A was solubilized in a denaturing buffer (6 M urea,500 mM NaCl, 10 mM DTT, 1 mM EDTA, 20 mM Tris, pH 8.3). Forrefolding, the solubilized probes were diluted 1:10 in a 50 mM
glycine buffer system (pH 9.8) containing CHAPS (20 mM), NaCl(500 mM) and 3 mM GSSG and gently stirred at 48C. Afterapproximately 24 h, the sample was diluted 2.8-fold with 14 mM
NaH2PO4 and subjected to isoelectric precipitation. After cen-trifugation, the pellet was dissolved in 0.1% TFA and the foldedmonomer was further purified by reverse-phase HPLC. For thispurpose, renatured rhGDF-5 C465A was loaded on a column(Aquapore Octyl, 20 mm, Applied Biosystems, Foster City, CA,USA) equilibrated with 25% buffer B in buffer A. rhGDF-5 C465Awas eluted with a 25–60% buffer B gradient within 70 min (flowrate 3 ml/min). Fractions containing purified refolded rhGDF-5C465A were pooled, lyophilized, and stored at -708C. DimericrhGDF-5 was purified accordingly and served as a positivecontrol.
UnauthenticatedDownload Date | 4/12/16 5:21 PM

458 C. Sieber et al.
Article in press - uncorrected proof
Western blotting under reducing and non-reducingconditions
Western blotting was performed as described by Hotten et al.(1996). Essentially, using 15% SDS-PAGE, the proteins wereseparated in the presence or absence of 1% b-mercaptoetha-nol. Blotting onto PVDF membranes was carried out in the pres-ence of 10 mM DTT so that only reduced monomeric GDF-5molecules were bound. These were detected with a chemilu-minescence detection kit (Applied Biosystems), using the mousemonoclonal antibody P3/66 (O’Keeffe et al., 2004).
Isoelectric focusing
Isoelectric focusing IEF) was performed using the electropho-resis system from GE Healthcare (Munich, Germany) consistingof CleanGel IEF gels and ampholines pH 6–9.
A CleanGel IEF gel was rehydrated overnight with rehydrationsolution (8 M urea, 0.1% Nonidet P-40) containing 3.1% ampho-lines pH 6.0–8.0, 3.1% ampholines pH 7.0–9.0, and 0.25%ampholines pH 3.5–10.0 under the exclusion of oxygen.
The separation was run on a Multiphor II horizontal system(GE Healthcare). After prefocusing of the gel for 20 min (voltage700 V, current 6 mA, power 4 W) the dry sample applicationpieces (GE Healthcare) were placed on the gel surface 1 cm fromthe cathode, and the samples (0.5 mg of rhGDF-5 and 0.5 mgof rhGDF-5 C465A) were applied. The samples were transferredinto the gel under the following conditions: voltage 500 V, current4 mA, power 4 W, time 60 min. Then the sample applicationpieces were removed and IEF was performed (voltage 2000 V,current 7 mA, power 7 W, time 150 min; band sharpening: volt-age 2500 V, current 7 mA, power 9 W, time 10 min).
After IEF the proteins were fixed overnight in 0.7 M TCA,0.14 M 5-sulfosalicylic acid dihydrate. For visualization, the gelwas silver-stained using the PlusOne Silver Staining Kit (GEHealthcare).
Peptide mapping
rhGDF-5 and rhGDF-5 C465A (100 mg each) were reduced withDTT under denaturing conditions and alkylated with 4-vinylpy-ridine. After desalting and lyophilization, 40 mg of each proteinwas resolved in 100 mM Tris-HCl, pH 8.5, 2 M urea, and incu-bated for 5 h with trypsin (2 mg; Merck, Darmstadt, Germany;sequencing grade) at 368C. The peptides were separated byreverse-phase HPLC using a Cosmosil column (5 mm, C18-AR-300; 4.6=250 mm; Phenomenex, Aschaffenburg, Germany)developed with a linear gradient ranging from 0 to 50% aceto-nitrile in 0.1% TFA (Pierce, Rockford, IL, USA; sequencing grade)within 90 min. Tryptic peptides were N-terminally sequenced ona protein sequencer (473 A, Applied Biosystems), and theirmasses were determined by MALDI-TOF mass spectrometry(Kompact MALDI I; Kratos Analytical, Manchester, UK).
Solubility testing
Solutions of rhGDF-5 C465A (40 mg/100 ml) in 10 mM HCl werelyophilized. To determine the solubility of rhGDF-5 C465A, twodifferent buffers with different pH values were created. Na-car-bonate buffers were adjusted to pH 9.5–12.1 by mixing 0.1 M
Na2CO3 and 0.1 M NaHCO3. For the acetate buffer, 0.1 M Na-acetate and 0.1 M acetic acid were mixed to reach pH 3.7–5.6.The buffers were added to lyophilized protein, and samples werevigorously agitated, followed by centrifugation at 13 000 g. Thesupernatant was carefully removed and quantitatively analyzedby reverse-phase HPLC and peak integration.
Alkaline phosphatase assay
The biological activity of rhGDF-5 and rhGDF-5 C465A wasdetermined using different concentrations of rhGDF-5 and rh-GDF-5 C465A on ATDC5 cells (Nakamura et al., 1999). ATDC5cells were plated at 104 cells/well in 96-well plates. After 24 h,cells were stimulated with 1–75 nM of rhGDF-5 or rhGDF-5C465A in starvation medium containing 2% FBS. After 72 h,cells were washed with PBS and extracted with alkaline phos-phate buffer 1, containing 1% Nonidet P-40, 0.1 M glycine,pH 9.6, 1 mM MgCl2 and 1 mM ZnCl2. To achieve thorough lysis,cells were incubated with shaking in buffer 1 for 1 h at roomtemperature. Enzyme activity was assayed with 10 mM p-nitro-phenylphosphate as a substrate in 0.1 M glycine, pH 9.6, 1 mM
MgCl2 and 1 mM ZnCl2. After appropriate incubation at roomtemperature (approx. 5 min), the absorbance was measured at405 nM with a Dynatech (Chantilly, VA, USA) ELISA reader.
Smad-reporter gene assay
ATDC5 cells were plated at a density of 1.3=105 cells/well in six-well plates. The following day, cells were transfected usingLipofectAMINE� (Invitrogen, Karlsruhe, Germany) with pSBE-luc (Jonk et al., 1998) and pRL-Tk for reference (Promega,Mannheim, Germany). Cells were starved in medium containing0.2% FBS for 5 h and incubated with 1–75 nM of rhGDF-5 orrhGDF-5 C465A in low-serum medium for an additional 24 h.Following starvation, cells were lysed with shaking in 400 ml of1= Passive Lysis Buffer (Promega) per well for 20–30 min. Lucif-erase activity was measured using the Dual Luciferase reporterassay system (Promega).
Receptor binding analysis
The BIAcoreX system (BIAcore, Uppsala, Sweden) was used torecord the binding of BRIb-Fc fusion protein (R&D Systems,Wiesbaden, Germany) to either immobilized rhGDF-5 or rhGDF-5 C465A.
The dimeric (rhGDF-5) or monomeric form (rhGDF-5 C465A)of GDF-5 was immobilized on biosensor CM5 chips via aminecoupling at 3000–5000 resonance units (RU). BRIb-Fc at 100,250, 400 or 500 nM in HBS buffer (10 mM HEPES, pH 7.4,500 mM NaCl, 3.4 mM EDTA, 0.005% surfactant P20) was per-fused at a flow rate of 20 ml/min at 258C. Multichannel analysisusing two flow cells was performed. One flow cell was used torecord the background sensogram (without ligand) and was sub-tracted from the sensogram obtained from the flow cell withimmobilized ligand. For each concentration, the sensogram wasrepeated in triplicate. The response in resonance units wasmeasured 315 s after injection. The immobilized ligands wereregenerated on the biosensor with 10 mM glycine, pH 2.0.
Immunofluorescence co-patching of cell-surfacereceptors
For co-patching experiments, COS7 cells were grown on glasscoverslips in Dulbecco’s modified Eagle medium (DMEM) sup-plemented with 10% FBS (Biological Industries, Beit Haemek,Israel), 100 U/ml penicillin, 100 mg/ml streptomycin, and 4 mM
glutamine (Biological Industries). Coverslips placed in 6-welldishes were transfected with 1 mg of DNA from pcDNA1 orpcDNA3.1 containing the BRIa, BRIb and BRII constructs(tagged with c-myc or HA epitope) (Gilboa et al., 2000) usingFuGene (De Crescenzo et al., 2003). Experiments were per-formed 44–48 h after transfection.
To measure oligomerization of receptors directly at the cellsurface, we used antibody-mediated immunofluorescence co-patching of epitope-tagged receptors (Henis et al., 1994; Gilboaet al., 1998, 2000). COS7 cells were co-transfected with vectors
UnauthenticatedDownload Date | 4/12/16 5:21 PM

Activity of monomeric GDF-5 459
Article in press - uncorrected proof
encoding pairs of receptors carrying different epitope tags (e.g.,myc-BRIa together with HA-BRIa). The cells were washed twicewith serum-free DMEM, and incubated for 30 min at 378C toallow digestion of serum-derived ligands. After washing twicewith cold Hank’s balanced salt solution (HBSS) supplementedwith 20 mM HEPES (pH 7.4) and 2% BSA (HBSS/HEPES/BSA),the cells were incubated with normal goat g-globulin (JacksonImmunoResearch, Soham, UK; 200 mg/ml, 45 min, 48C) to blocknon-specific binding. This was followed by successive incuba-tion in the cold (to avoid internalization and enable exclusive cellsurface labeling) in HBSS/HEPES/BSA with primary anti-tagIgGs (20 mg/ml each, 45 min): rabbit HA.11 against the HA tag(a-HA; BabCO, Berkeley, CA, USA) together with 9E10 mouseanti-myc (a-myc from Harvard Monoclonals, Boston, MA, USA)(Evan et al., 1985), followed by labeling/patching with secondaryAlexa594-goat anti-rabbit (GaR) and Alexa488-goat anti-mouse(GaM) IgG (Invitrogen; 20 mg/ml each, 30 min, 48C). The cellswere washed and fixed in methanol (5 min, -208C) and acetone(3 min, -208C), and mounted in Antifade (Invitrogen). In cases forwhich ligands were present (10 nM rhBMP-2, 100 nM rhGDF-5or 100 nM rhGDF-5 C465A), they were added at 48C 1.5 h priorto labeling with IgGs and included at the same concentrationduring subsequent incubation with the antibodies.
Fluorescence digital images were recorded using a CCD cam-era as described by Gilboa et al. (2000). The Alexa488 (green)and Alexa594 (red) images were exported in TIFF format toImage Pro Plus (Media Cybernetics, Silver Spring, MD, USA),and subjected to quantitative analysis of the extent of co-patch-ing using an algorithm we recently developed (Lachmanovich etal., 2003). Briefly, the program segments the patches in a user-defined area on the image, subtracts the background and iden-tifies the center of mass of each object in the green and redimages. The next step is nearest-neighbor analysis, calculatingthe distances from each green patch to the nearest red patch(and vice versa). Patches with a nearest neighbor within oneoptical resolution unit (up to 0.2 mm) are considered to be colo-calized (Lachmanovich et al., 2003). The percentage co-patchingof, e.g., green with red patches is given by dividing the numberof the green patches colocalized with red patches by the totalnumber of green patches. This procedure is normally applied tomore than 20 cells in each case to obtain accurate averages.
Acknowledgments
We thank Monika Jakob and Yvonne Kehl for technical assis-tance. The reporter construct pSBE was a kind gift from Peterten Dijke. This work was supported by BMBF 0312114 (to F.P.)and BMBF 0312117 (to P.K.), and the Israel Science Foundation(grant no. 185/05) (to Y.I.H.). Y.I.H. is an incumbent of the ZalmanWeinberg Chair in Cell Biology.
References
Amatayakul-Chantler, S., Qian, S.W., Gakenheimer, K., Bottinger,E.P., Roberts, A.B., and Sporn, M.B. (1994). wSer77xtransforming growth factor-b 1. Selective biological activityand receptor binding in mink lung epithelial cells. J. Biol.Chem. 269, 27687–27691.
Canalis, E., Economides, A.N., and Gazzerro, E. (2003). Bonemorphogenetic proteins, their antagonists, and the skeleton.Endocr. Rev. 24, 218–235.
Cheifetz, S., Weatherbee, J.A., Tsang, M.L., Anderson, J.K.,Mole, J.E., Lucas, R., and Massague, J. (1987). The trans-forming growth factor-b system, a complex pattern of cross-reactive ligands and receptors. Cell 48, 409–415.
Daopin, S., Piez, K.A., Ogawa, Y., and Davies, D.R. (1992). Crys-tal structure of transforming growth factor-b2: an unusualfold for the superfamily. Science 257, 369–373.
De Crescenzo, G., Pham, P.L., Durocher, Y., and O’Connor-McCourt, M.D. (2003). Transforming growth factor-b (TGF-b)binding to the extracellular domain of the type II TGF-breceptor: receptor capture on a biosensor surface using anew coiled-coil capture system demonstrates that aviditycontributes significantly to high affinity binding. J. Mol. Biol.328, 1173–1183.
Evan, G.I., Lewis, G.K., Ramsay, G., and Bishop, J.M. (1985).Isolation of monoclonal antibodies specific for human c-mycproto-oncogene product. Mol. Cell. Biol. 5, 3610–3616.
Gilboa, L., Wells, R.G., Lodish, H.F., and Henis, Y.I. (1998). Oligo-meric structure of type I and type II transforming growth fac-tor b receptors: homodimers form in the ER and persist atthe plasma membrane. J. Cell Biol. 140, 767–777.
Gilboa, L., Nohe, A., Geissendorfer, T., Sebald, W., Henis, Y.I.,and Knaus, P. (2000). Bone morphogenetic protein receptorcomplexes on the surface of live cells: a new oligomerizationmode for serine/threonine kinase receptors. Mol. Biol. Cell11, 1023–1035.
Greenwald, J., Groppe, J., Gray, P., Wiater, E., Kwiatkowski, W.,Vale, W., and Choe, S. (2003). The BMP7/ActRII extracellulardomain complex provides new insights into the cooperativenature of receptor assembly. Mol. Cell 11, 605–617.
Hart, P.J., Deep, S., Taylor, A.B., Shu, Z., Hinck, C.S., and Hinck,A.P. (2002). Crystal structure of the human TbR2 ectodo-main-TGF-b3 complex. Nat. Struct. Biol. 9, 203–208.
Henis, Y.I., Moustakas, A., Lin, H.Y., and Lodish, H.F. (1994). Thetypes II and III transforming growth factor-b receptors formhomo-oligomers. J. Cell Biol. 126, 139–154.
Honda, J., Andou, H., Mannen, T., and Sugimoto, S. (2000).Direct refolding of recombinant human growth differentiationfactor 5 for large-scale production process. J. Biosci. Bioeng.89, 582–589.
Hotten, G.C., Matsumoto, T., Kimura, M., Bechtold, R.F., Kron,R., Ohara, T., Tanaka, H., Satoh, Y., Okazaki, M., Shirai, T.,et al. (1996). Recombinant human growth/differentiation fac-tor 5 stimulates mesenchyme aggregation and chondroge-nesis responsible for the skeletal development of limbs.Growth Factors 13, 65–74.
Husken-Hindi, P., Tsuchida, K., Park, M., Corrigan, A.Z.,Vaughan, J.M., Vale, W.W., and Fischer, W.H. (1994). Mono-meric activin A retains high receptor binding affinity butexhibits low biological activity. J. Biol. Chem. 269,19380–19384.
Israel, D.I., Nove, J., Kerns, K.M., Kaufman, R.J., Rosen, V., Cox,K.A., and Wozney, J.M. (1996). Heterodimeric bone morpho-genetic proteins show enhanced activity in vitro and in vivo.Growth Factors 13, 291–300.
Jen, A., Madorin, K., Vosbeck, K., Arvinte, T., and Merkle, H.P.(2002). Transforming growth factor b-3 crystals as reservoirsfor slow release of active TGF-b3. J. Control Release 78,25–34.
Jonk, L.J., Itoh, S., Heldin, C.H., ten Dijke, P., and Kruijer, W.(1998). Identification and functional characterization of aSmad binding element (SBE) in the JunB promoter that actsas a transforming growth factor-b, activin, and bone mor-phogenetic protein-inducible enhancer. J. Biol. Chem. 273,21145–21152.
Kingsley, D.M. (1994). The TGF-b superfamily: new members,new receptors, and new genetic tests of function in differentorganisms. Genes Dev. 8, 133–146.
Kirsch, T., Sebald, W., and Dreyer, M.K. (2000). Crystal structureof the BMP-2-BRIA ectodomain complex. Nat. Struct. Biol.7, 492–496.
Knaus, P. and Sebald, W. (2001). Cooperativity of binding epi-topes and receptor chains in the BMP/TGFb superfamily.Biol. Chem. 382, 1189–1195.
UnauthenticatedDownload Date | 4/12/16 5:21 PM

460 C. Sieber et al.
Article in press - uncorrected proof
Lachmanovich, E., Shvartsman, D.E., Malka, Y., Botvin, C.,Henis, Y.I., and Weiss, A.M. (2003). Co-localization analysisof complex formation among membrane proteins by com-puterized fluorescence microscopy: application to immuno-fluorescence co-patching studies. J. Microsc. 212, 122–131.
Massague, J. (2000). How cells read TGF-b signals. Nat. Rev.Mol. Cell. Biol. 1, 169–178.
McPherron, A.C. and Lee, S.J. (1993). GDF-3 and GDF-9: twonew members of the transforming growth factor-b superfam-ily containing a novel pattern of cysteines. J. Biol. Chem.268, 3444–3449.
Nakamura, K., Shirai, T., Morishita, S., Uchida, S., Saeki-Miura,K., and Makishima, F. (1999). p38 mitogen-activated proteinkinase functionally contributes to chondrogenesis induced bygrowth/differentiation factor-5 in ATDC5 cells. Exp. Cell. Res.250, 351–363.
Nickel, J., Kotzsch, A., Sebald, W., and Mueller, T.D. (2005). Asingle residue of GDF-5 defines binding specificity to BMPreceptor IB. J. Mol. Biol. 349, 933–947.
Nishitoh, H., Ichijo, H., Kimura, M., Matsumoto, T., Makishima,F., Yamaguchi, A., Yamashita, H., Enomoto, S., and Miyazo-no, K. (1996). Identification of type I and type II serine/thre-onine kinase receptors for growth/differentiation factor-5.J. Biol. Chem. 271, 21345–21352.
Nohe, A., Hassel, S., Ehrlich, M., Neubauer, F., Sebald, W.,Henis, Y.I., and Knaus, P. (2002). The mode of bone morpho-genetic protein (BMP) receptor oligomerization determinesdifferent BMP-2 signaling pathways. J. Biol. Chem. 277,5330–5338.
Ogawa, Y., Schmidt, D.K., Dasch, J.R., Chang, R.J., and Glaser,C.B. (1992). Purification and characterization of transforminggrowth factor-b 2.3 and -b 1.2 heterodimers from bovinebone. J. Biol. Chem. 267, 2325–2328.
O’Keeffe, G.W., Hanke, M., Pohl, J., and Sullivan, A.M. (2004).Expression of growth differentiation factor-5 in the develop-ing and adult rat brain. Brain Res. Dev. Brain Res. 151,199–202.
Piek, E., Heldin, C.H., and Ten Dijke, P. (1999). Specificity, diver-sity, and regulation in TGF-b superfamily signaling. FASEB J.13, 2105–2124.
Sammar, M., Stricker, S., Schwabe, G.C., Sieber, C., Hartung,A., Hanke, M., Oishi, I., Pohl, J., Minami, Y., Sebald, W., etal. (2004). Modulation of GDF5/BRI-b signalling through inter-action with the tyrosine kinase receptor Ror2. Genes Cells 9,1227–1238.
Schlunegger, M.P. and Grutter, M.G. (1992). An unusual featurerevealed by the crystal structure at 2.2 A resolution of humantransforming growth factor-b 2. Nature 358, 430–434.
Schreuder, H., Liesum, A., Pohl, J., Kruse, M., and Koyama, M.(2005). Crystal structure of recombinant human growth anddifferentiation factor 5: evidence for interaction of the type Iand type II receptor-binding sites. Biochem. Biophys. Res.Commun. 329, 1076–1086.
Sebald, W., Nickel, J., Zhang, J.L., and Mueller, T.D. (2004).Molecular recognition in bone morphogenetic protein (BMP)/receptor interaction. Biol. Chem. 385, 697–710.
Seemann, P., Schwappacher, R., Kjaer, K.W., Krakow, D., Leh-mann, K., Dawson, K., Stricker, S., Pohl, J., Ploger, F., Staub,E., et al. (2005). Activating and deactivating mutations in thereceptor interaction site of GDF5 cause symphalangism orbrachydactyly type A2. J. Clin. Invest. 115, 2373–2381.
Shi, Y. and Massague, J. (2003). Mechanisms of TGF-b signalingfrom cell membrane to the nucleus. Cell 113, 685–700.
Shimmi, O., Umulis, D., Othmer, H., and O’Connor, M.B. (2005).Facilitated transport of a Dpp/Scw heterodimer by Sog/Tsgleads to robust patterning of the Drosophila blastodermembryo. Cell 120, 873–886.
Vitt, U.A., Mazerbourg, S., Klein, C., and Hsueh, A.J. (2002).Bone morphogenetic protein receptor type II is a receptor forgrowth differentiation factor-9. Biol. Reprod. 67, 473–480.
Received October 27, 2005; accepted January 25, 2006
UnauthenticatedDownload Date | 4/12/16 5:21 PM
Related Documents











