Monoclonal antibodies generated in carbonic anhydrase IX-deficient mice recognize different domains of tumour-associated hypoxia-induced carbonic anhydrase IX $ Miriam Zat’ovic ˇova ´ a , Kvetoslava Tara ´bkova ´ a , Elis ˇka S ˇ vastova ´ a , Adriana Gibadulinova ´ a , Vojtech Mucha a , Ly ´dia Jakubı ´c ˇkova ´ a , Zuzana Biesova ´ a , Monika Rafajova ´ a , Marta Ortova Gut b , Seppo Parkkila c , Anna-Kaisa Parkkila d , Abdul Waheed e , Willam S. Sly e , Ivan Horak b , Jaromı ´r Pastorek a , Silvia Pastorekova ´ a, * a Centre of Molecular Medicine, Institute of Virology, Slovak Academy of Sciences, Du ´bravska ´ cesta 9, 845 05 Bratislava, Slovak Republic b Department of Molecular Genetics, Institute of Molecular Pharmacology and Medical Centre of Free University of Berlin, Berlin, Germany c Institute of Medical Technology, University of Tampere and Tampere University Hospital, Finland d Department of Neurology, University of Tampere and Tampere University Hospital, Finland e Edward A. Doisy Department of Biochemistry and Molecular Biology, Saint Louis University School of Medicine, St. Louis, MO 63104, USA Received 15 May 2003; received in revised form 25 July 2003; accepted 11 August 2003 Abstract Transmembrane carbonic anhydrase IX (CA IX) is frequently expressed in human tumours in response to hypoxia and may serve as a tumour marker and therapeutic target. So far, only a single monoclonal antibody (MAb) M75 with an epitope in the N-terminal proteoglycan (PG)-like region has been available for detection purposes. Attempts to produce MAbs against other parts of CA IX were unsuccessful due to the immunodominance of the PG region that significantly differs between human and mouse homologues. To overcome this problem, we used various forms of human CA IX antigen to immunize CA IX-deficient mice recently produced by targeted disruption of Car9 gene. Here, we describe new MAbs that react with human, but not mouse CA IX in different immunodetection settings, and show no cross-reactivity with CA I, II and XII. MAb IV/18 is directed to the PG region, while the other six antibodies bind to the CA domain, as determined by CA IX deletion variants. IV/18 recognizes a linear epitope, while anti-CA MAbs V/10, V/12, VII/20, VII/28, VII/32 and VII/ 38 react with conformational epitopes clustered into three antigenic sites. The new antibodies represent important tools for improving our knowledge of structure – function relationships in the CA IX molecule and a better understanding of the role 0022-1759/$ - see front matter D 2003 Elsevier B.V. All rights reserved. doi:10.1016/j.jim.2003.08.011 Abbreviations: CA, carbonic anhydrase; IHC, immunohistochemistry; MAb, monoclonal antibody; PG, proteoglycan; GST, glutathione-S transferase. $ According to the carbonic anhydrase nomenclature, human CA isoenzymes are written in capital Roman letters and numbers, while their genes are written in Italic letters and Arabic numbers. * Corresponding author. Tel.: +42-1-2-5930-2404; fax: +42-1-2-5477-4284. E-mail address: [email protected] (S. Pastorekova ´). www.elsevier.com/locate/jim Journal of Immunological Methods 282 (2003) 117 – 134

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/jim
Journal of Immunological Methods 282 (2003) 117–134
Monoclonal antibodies generated in carbonic anhydrase
IX-deficient mice recognize different domains of
tumour-associated hypoxia-induced carbonic
anhydrase IX$
Miriam Zat’ovicovaa, Kvetoslava Tarabkovaa, Eliska Svastovaa,Adriana Gibadulinovaa, Vojtech Muchaa, Lydia Jakubıckovaa,
Zuzana Biesovaa, Monika Rafajovaa, Marta Ortova Gutb, Seppo Parkkilac,Anna-Kaisa Parkkilad, Abdul Waheede, Willam S. Slye, Ivan Horakb,
Jaromır Pastoreka, Silvia Pastorekovaa,*
aCentre of Molecular Medicine, Institute of Virology, Slovak Academy of Sciences, Dubravska cesta 9, 845 05 Bratislava, Slovak RepublicbDepartment of Molecular Genetics, Institute of Molecular Pharmacology and Medical Centre of Free University of Berlin, Berlin, Germany
c Institute of Medical Technology, University of Tampere and Tampere University Hospital, FinlanddDepartment of Neurology, University of Tampere and Tampere University Hospital, Finland
eEdward A. Doisy Department of Biochemistry and Molecular Biology, Saint Louis University School of Medicine,
St. Louis, MO 63104, USA
Received 15 May 2003; received in revised form 25 July 2003; accepted 11 August 2003
Abstract
Transmembrane carbonic anhydrase IX (CA IX) is frequently expressed in human tumours in response to hypoxia and
may serve as a tumour marker and therapeutic target. So far, only a single monoclonal antibody (MAb) M75 with an epitope
in the N-terminal proteoglycan (PG)-like region has been available for detection purposes. Attempts to produce MAbs
against other parts of CA IX were unsuccessful due to the immunodominance of the PG region that significantly differs
between human and mouse homologues. To overcome this problem, we used various forms of human CA IX antigen to
immunize CA IX-deficient mice recently produced by targeted disruption of Car9 gene. Here, we describe new MAbs that
react with human, but not mouse CA IX in different immunodetection settings, and show no cross-reactivity with CA I, II
and XII. MAb IV/18 is directed to the PG region, while the other six antibodies bind to the CA domain, as determined by
CA IX deletion variants. IV/18 recognizes a linear epitope, while anti-CA MAbs V/10, V/12, VII/20, VII/28, VII/32 and VII/
38 react with conformational epitopes clustered into three antigenic sites. The new antibodies represent important tools for
improving our knowledge of structure–function relationships in the CA IX molecule and a better understanding of the role
0022-1759/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/j.jim.2003.08.011
Abbreviations: CA, carbonic anhydrase; IHC, immunohistochemistry; MAb, monoclonal antibody; PG, proteoglycan; GST, glutathione-S
transferase.$ According to the carbonic anhydrase nomenclature, human CA isoenzymes are written in capital Roman letters and numbers, while their
genes are written in Italic letters and Arabic numbers.
* Corresponding author. Tel.: +42-1-2-5930-2404; fax: +42-1-2-5477-4284.
E-mail address: [email protected] (S. Pastorekova).

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134118
of CA IX in cancer development. Moreover, the availability of the MAbs specific for distinct antigenic regions on two
separate extracellular domains offers an opportunity to elaborate a sensitive assay that could be particularly important for CA
IX detection in body fluids of cancer patients.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Monoclonal antibodies; Carbonic anhydrase IX; Cancer detection; CA IX-deficient mice; Antigenic site
1. Introduction clinical research. First, expression of CA IX is very
Carbonic anhydrase IX (CA IX, previously named
MN protein, MN/CA IX) is a highly active member
of the a carbonic anhydrase family of zinc metal-
loenzymes that catalyse the reversible conversion
between carbon dioxide and bicarbonate (Pastorek
et al., 1994; Opavsky et al., 1996; Parkkila, 2000;
Wingo et al., 2001). It is one of 14 isoforms that
exist in mammals and occupy different subcellular
positions, including cytoplasm (CA I, II, III, VII),
mitochondria (CA VA, VB), secretory vesicles (CA
VI) and the plasma membrane (CA IV, IX, XII,
XIV). Some of the isozymes are distributed over a
broad range of tissues (CA I, II, CA IV), others are
more restricted to particular organs (CA VI in sali-
vary and mammary glands), and two isoforms have
been linked to cancer tissues (CA IX, XII) (reviewed
in Parkkila, 2000; Pastorekova and Pastorek, 2003).
Enzyme activity and kinetic properties, as well as
sensitivity to sulfonamide inhibitors range from high
(CA II, CA IX, CA XII, CA IV) to low (CA III)
(Supuran and Scozzafava, 2000). Several isoforms
designated as CA-related proteins (CA-RP VIII, X,
XI) are acatalytic due to an incompletely conserved
active site (Nishimori, 2003). This extraordinary
variability among the genetically related members
of the same family of proteins creates the basis for
their involvement in diverse physiological and path-
ological processes. The catalytic activity is of fun-
damental relevance for the maintenance of acid–base
balance and the exchange of ions and water in
metabolically active tissues. Through this activity,
CAs substantially contribute to respiration, produc-
tion of body fluids (vitreous humor, gastric juice,
cerebrospinal fluid), bone resorption, renal acidifica-
tion, etc. (Parkkila, 2000).
CA IX isozyme integrates several properties that
make it an important subject of basic as well as
tightly associated with a broad variety of human
tumours, while it is generally absent from the
corresponding normal tissues (Zavada et al., 1993;
Liao et al., 1994, 1997; Turner et al., 1997; Saarnio et
al., 1998, 2001; Vermylen et al., 1999; Ivanov et al.,
2001; Bartosova et al., 2002). This is principally
related to tumour hypoxia that strongly activates
transcription of the CA9 gene via a hypoxia-inducible
transcription factor binding to a hypoxia-response
element localized just upstream of the transcription
initiation site in the CA9 promoter (Wykoff et al.,
2000). Since tumour hypoxia is an important phe-
nomenon with dramatic implications for cancer de-
velopment and therapy (Hockel and Vaupel, 2001),
CA IX offers significant potential as an intrinsic
hypoxic marker with a prognostic/predictive value
and as a promising therapeutic target (Wykoff et al.,
2000, 2001; Beasley et al., 2001; Giatromanolaki et
al., 2001; Koukourakis et al., 2001). CA IX is an
integral plasma membrane protein with a large extra-
cellular part exposed at the surface of cancer cells and
is thus accessible to many targeting tools, including
specific monoclonal antibodies. Furthermore, in con-
trast to the other CA isozymes, CA IX possesses a
unique proteoglycan (PG)-related region that forms an
N-terminal extension of the extracellular CA domain
and reduces cross-recognition with other isozymes
(Opavsky et al., 1996). Moreover, CA IX has been
functionally implicated in cell adhesion and due to its
high catalytic activity may contribute to acidification
of the extracellular microenvironment (Zavada et al.,
2000; Svastova et al., 2003; Ivanov et al., 1998). In
addition to the potential clinical exploitation of CA
IX, there is an increasing interest in resolving many
basic molecular and functional aspects of the protein
since our knowledge of its precise role in cancer cells,
the contribution of different domains/sequence motifs,
and its regulation remain insufficient.

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 119
So far, most of the basic CA IX-related studies have
been performed using a single mouse monoclonal
antibody (MAb) M75 that recognizes the N-terminal
PG region of CA IX (Pastorekova et al., 1992; Zavada
et al., 2000). This antibody proved to be highly
specific and perfectly suitable for certain purposes
including immunohistochemical analyses of cancer
tissue sections (Liao et al., 1994; Saarnio et al.,
1998; Ivanov et al., 2001 and references therein),
targeting hypoxic tumour cells in animal models
(Chrastina et al., 2003), CA IX immunodetection in
vitro and molecular characterization (Pastorek et al.,
1994; Pastorekova et al., 1997; Lieskovska et al.,
1999; Kaluz et al., 1999, 2002; Olive et al., 2001).
On the other hand, CA IX-specific monoclonal anti-
bodies with epitope specificities differing from those
of M75 have become highly desirable for those
approaches that are based on the capture-detection
principle or for studies of mutated variants of CA
IX. However, previous attempts to produce such anti-
bodies were unsuccessful apparently due to the fact
that human CA IX differs from the mouse homologue
predominantly in the N-terminal PG amino acid se-
quence (Ortova Gut et al., 2002). This sequence
appears to be strongly immunogenic possibly because
the immunized mice recognize it as non-self, while
they do not direct a humoral response to the other,
more conserved parts of the human CA IX molecule.
The availability of CA IX-deficient mice (Ortova
Gut et al., 2002) has allowed us to generate hybrid-
oma cell lines producing monoclonal antibodies spe-
cific for different extracellular regions of human CA
IX protein. Here, we provide detailed characterization
of these antibodies and an assessment of their reac-
tivity in various immunodetection procedures, and we
discuss their possible applications.
2. Materials and methods
2.1. Cell culture
Hybridoma cell lines were grown in DMEM me-
dium supplemented with 10% FCS (BioWhittaker,
Verviers, Belgium), 2 mM glutamine and 40 Ag/ml
gentamicin (Lek, Slovenia) at 37 jC in 5% CO2 in air.
The same culture conditions were applied to the
following cell lines that were used either as a source
of CA IX antigen or as negative controls: mouse NIH
3T3 fibroblasts permanently transfected with the full-
length human CA9 cDNA in pSG5C plasmid (NIH
3T3-flCA IX), with the cDNA variants in pSG5C as
described below and corresponding mock transfected
NIH 3T3-neo controls (Pastorek et al., 1994); MDCK
cells transfected with the full-length CA9 cDNA
(MDCK-flCA IX), cDNA variants in pSG5C and
corresponding MDCK-neo controls (Svastova et al.,
2003); human HT-29 colon carcinoma cells as well as
human HeLa cervical carcinoma cells naturally
expressing CA IX; and C33a cervical carcinoma cells
negative for CA IX.
For immunization purposes, the cells grown for 24–
48 h were washed twice in PBS, scraped, collected by
centrifugation and re-suspended in an appropriate
volume of PBS. For the final booster, NIH 3T3-flCA
IX cells were extracted with OCG extraction buffer
composed of 0.5M NaCl, 0.5% octyl-h-D-glucopyra-noside (Sigma, St. Louis, MO), 0.1 mMPMSF (Sigma)
for 30 min at 4 jC, scraped, centrifuged for 5 min at
13000 rpm and dialyzed against PBS for 48 h at 4 jC.
2.2. Cloning and expression of cDNA variants and
recombinant CA IX proteins
Constructs coding for CA IX-PGCA and CA IX-
DCA variants (see Fig. 1) were obtained from the full-
length CA9 cDNA in the pSG5C eukaryotic expres-
sion plasmid (Pastorek et al., 1994) by the removal of
the 3V sequence using EcoRI digestion and removal of
the central CA-coding sequence using double
EcoRV–EcoRI digestion, respectively. The construct
coding for CA IX-DPG was generated from two PCR
products by amplification of the sequences cor-
responding to amino acids 1–41 and 131–459 using
the full-length cDNA and two sets of primers: the
forward primer from the plasmid combined with a
gene-specific DPGS primer (5V-CCATCCCCA-
GAGGTTGCCCAGGGACAAAGAAGGGGATG-
3V) and the reverse primer from the plasmid combined
with DPGA that is complementary to DPGS. The
gene-specific DPGS/DPGA primers were designed so
that their arms were complementary to sequences
flanking the deletion. All the constructs were verified
by sequencing. Recombinant plasmids were co-trans-
fected to NIH 3T3 and MDCK with pSV2neo using
Gene Porter II transfection kit (Gene Therapy System,
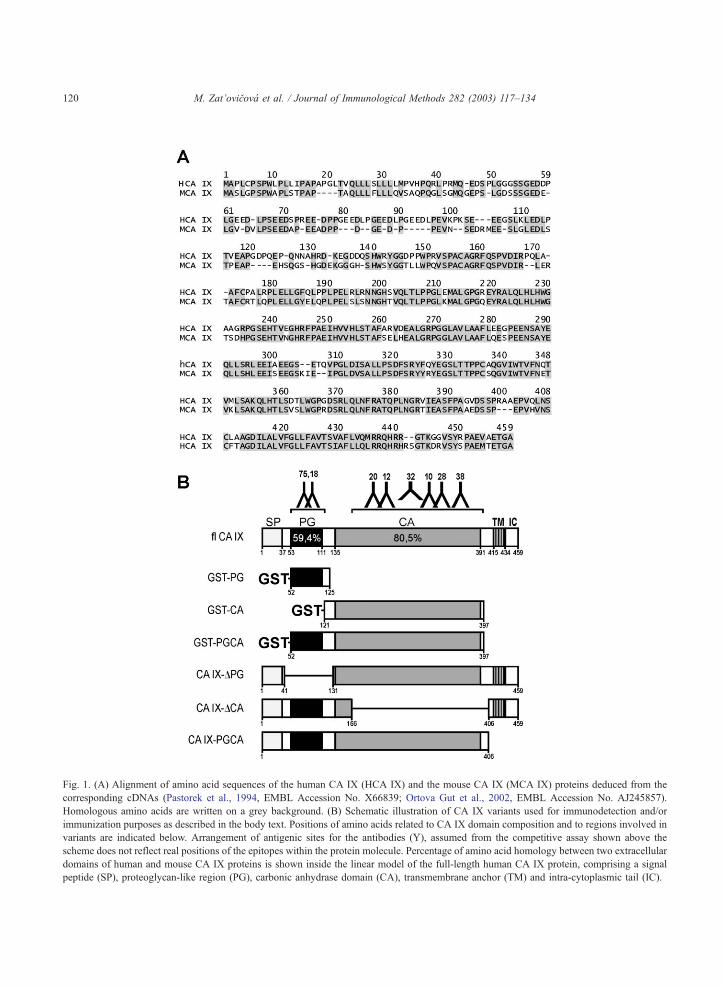
Fig. 1. (A) Alignment of amino acid sequences of the human CA IX (HCA IX) and the mouse CA IX (MCA IX) proteins deduced from the
corresponding cDNAs (Pastorek et al., 1994, EMBL Accession No. X66839; Ortova Gut et al., 2002, EMBL Accession No. AJ245857).
Homologous amino acids are written on a grey background. (B) Schematic illustration of CA IX variants used for immunodetection and/or
immunization purposes as described in the body text. Positions of amino acids related to CA IX domain composition and to regions involved in
variants are indicated below. Arrangement of antigenic sites for the antibodies (Y), assumed from the competitive assay shown above the
scheme does not reflect real positions of the epitopes within the protein molecule. Percentage of amino acid homology between two extracellular
domains of human and mouse CA IX proteins is shown inside the linear model of the full-length human CA IX protein, comprising a signal
peptide (SP), proteoglycan-like region (PG), carbonic anhydrase domain (CA), transmembrane anchor (TM) and intra-cytoplasmic tail (IC).
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134120

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 121
San Diego, CA). The transfected cells were subjected
to 2 weeks of selection in the presence of G418 (Life
Technologies, Gaithersburg, MD), cloned and tested
for expression of CA IX.
The cDNA fragments encoding PG and CA extra-
cellular domains of CA IX (see Fig. 1) were amplified
either individually or together by specific primers using
a full-length CA9 cDNA as a template. The fragment
coding for the PG domain (aa 52–125) was obtained
with PG3 (sense) 5V-TAGAATTCGGCTCTTCTGGG-GAAGAT-3V and PG4 (antisense) 5V-ATACTC-
GAGGGGTTCTTGAGGATCTCC-3V primers, and
the fragment coding for CA domain (aa 121–397)
was obtained with CA5 (sense) 5V-TAGAATTC-GATCCTCAAGAACCCCAG-3Vand CA6 (antisense)5V-AATCTCGAGACTGCTGTCCACTCCAGC-
3Vprimers. The combination of primers PG3 and CA6
was employed to produce cDNA fragment encoding
PG+CA domains (aa 52–397).
The resulting PCR products were cloned into
pGEX 4T-1 bacterial plasmid via EcoRI and XhoI
restriction sites inserted in the primer sequences.
Fusion proteins glutathione-S transferase (GST)–
PG, GST–CA, GST–PGCA were expressed in
Escherichia coli strain DH5a using a standard proce-
dure (Gibadulinova et al., 1998). The full-length
fusion GST–flCA IX protein generated earlier was
produced in parallel. The proteins were purified by
affinity chromatography using Glutathione-S Sephar-
ose (Amersham Pharmacia) and eluted with 10 mM
GSH (reduced glutathione). The proteins obtained
were utilized either as antigens for MAb testing or
as immunogen (GST–CA) for immunization.
2.3. Immunization
For immunization, CA IX-deficient mice generated
and characterized by Ortova Gut et al. (2002) were
used at the age of 8–10 weeks and immunized as
shown in Table 1. The mice were injected intraper-
itoneally (i.p.) with 5� 106 NIH 3T3-flCA IX cells in
0.5 ml PBS. Three weeks later, the mice received
either the same i.p. injection of NIH 3T3-flCA IX
cells or i.p. injection of 5� 106 HT-29 cells in 0.5 ml
PBS. After another 3 weeks, mice were boosted
alternatively as follows: (i) i.p. with 100 Ag of
GST–CA protein bound to Glutathione-S Sepharose
in 0.5 ml PBS, (ii) intravenously (i.v.) with 100 Ag of
eluted GST–CA protein in 200 Al PBS, (iii) i.p. with5� 106 NIH 3T3-flCA IX cells in 0.5 ml PBS and (iv)
i.v. with 200 Al OCG extract of NIH 3T3-CA IX cells.
Splenocytes were harvested 3 days later and fused
with the Sp2/0 myeloma cells.
2.4. Production of MAbs
The fusions of splenocytes from each of the
immunized mice with Sp2/0 cells were carried out
according to Lane et al. (1986). Hybridomas were
selected in DMEM-HAT medium containing hypo-
xanthine, aminopterin and thymidine. The superna-
tants were screened for the specific reactivity towards
CA IX by ELISA as described below. Positive hy-
bridoma cultures were cloned by limiting dilution
using Terasaki microplates. Clonal hybridoma cell
lines were expanded, subjected to freezing–refreezing
and repeatedly tested for reactivity to CA IX. Large
quantities of antibodies were purified from hybridoma
culture medium using affinity chromatography on
Protein A Sepharose CL-4B (Amersham Biosciences,
Uppsala, Sweden).
2.5. ELISA screening
Screening of positive hybridomas was performed
by a sandwich ELISA. Microplate wells were coated
overnight at 37 jC with RIPA extract (specified
below) of NIH 3T3-flCA IX cells, diluted 1/10 in
PBS, and in parallel with an extract of NIH 3T3-neo
cells as a negative control. After blocking with 10%
FCS in PBS, the coated wells were incubated with
undiluted culture media from individual hybridomas.
Peroxidase-labelled pig anti-mouse IgG (Sevapharma,
Prague, Czech Republic) was used as detector. M75
MAb was employed as the positive control.
For the differential ELISA screening, the wells
were coated with the following antigens: 10 ng/well
of GST–flCA IX, 10 ng/well of GST–PGCA, 10 ng/
well of GST–PG, RIPA extract of HeLa cells diluted
1/10 in PBS, RIPA extract of HT-29 cells diluted 1/10
in PBS, and were then assayed as above.
2.6. Determination of isotypes
MAb isotypes were determined by ELISA using
affinity purified rabbit anti-mouse IgG1, IgG2a,
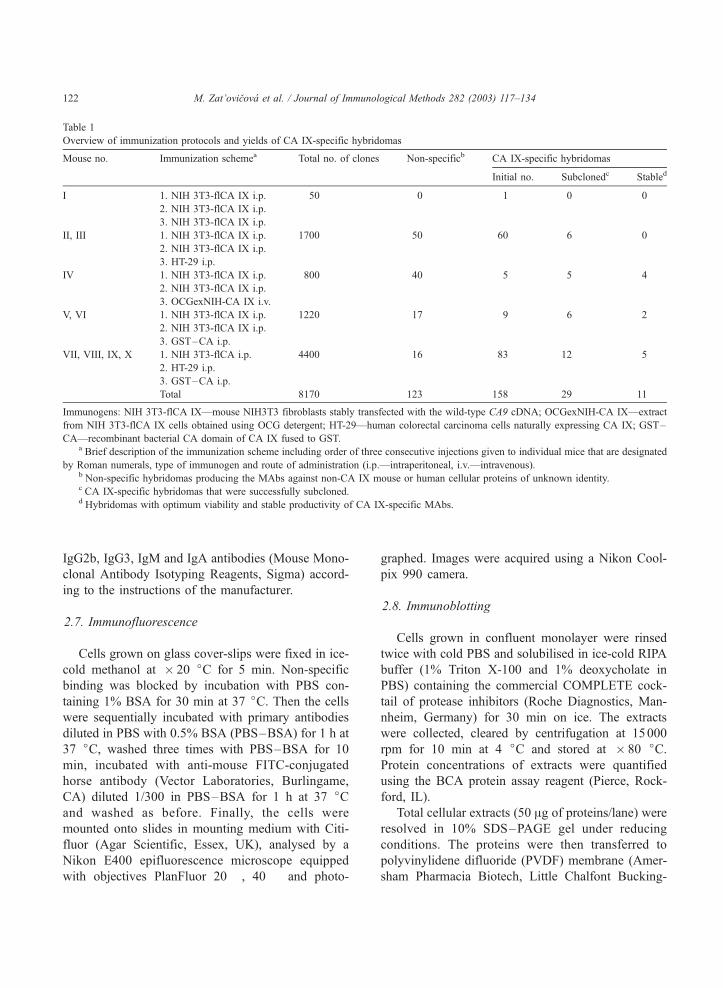
Table 1
Overview of immunization protocols and yields of CA IX-specific hybridomas
Mouse no. Immunization schemea Total no. of clones Non-specificb CA IX-specific hybridomas
Initial no. Subclonedc Stabled
I 1. NIH 3T3-flCA IX i.p. 50 0 1 0 0
2. NIH 3T3-flCA IX i.p.
3. NIH 3T3-flCA IX i.p.
II, III 1. NIH 3T3-flCA IX i.p. 1700 50 60 6 0
2. NIH 3T3-flCA IX i.p.
3. HT-29 i.p.
IV 1. NIH 3T3-flCA IX i.p. 800 40 5 5 4
2. NIH 3T3-flCA IX i.p.
3. OCGexNIH-CA IX i.v.
V, VI 1. NIH 3T3-flCA IX i.p. 1220 17 9 6 2
2. NIH 3T3-flCA IX i.p.
3. GST–CA i.p.
VII, VIII, IX, X 1. NIH 3T3-flCA i.p. 4400 16 83 12 5
2. HT-29 i.p.
3. GST–CA i.p.
Total 8170 123 158 29 11
Immunogens: NIH 3T3-flCA IX—mouse NIH3T3 fibroblasts stably transfected with the wild-type CA9 cDNA; OCGexNIH-CA IX—extract
from NIH 3T3-flCA IX cells obtained using OCG detergent; HT-29—human colorectal carcinoma cells naturally expressing CA IX; GST–
CA—recombinant bacterial CA domain of CA IX fused to GST.a Brief description of the immunization scheme including order of three consecutive injections given to individual mice that are designated
by Roman numerals, type of immunogen and route of administration (i.p.—intraperitoneal, i.v.—intravenous).b Non-specific hybridomas producing the MAbs against non-CA IX mouse or human cellular proteins of unknown identity.c CA IX-specific hybridomas that were successfully subcloned.d Hybridomas with optimum viability and stable productivity of CA IX-specific MAbs.
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134122
IgG2b, IgG3, IgM and IgA antibodies (Mouse Mono-
clonal Antibody Isotyping Reagents, Sigma) accord-
ing to the instructions of the manufacturer.
2.7. Immunofluorescence
Cells grown on glass cover-slips were fixed in ice-
cold methanol at � 20 jC for 5 min. Non-specific
binding was blocked by incubation with PBS con-
taining 1% BSA for 30 min at 37 jC. Then the cells
were sequentially incubated with primary antibodies
diluted in PBS with 0.5% BSA (PBS–BSA) for 1 h at
37 jC, washed three times with PBS–BSA for 10
min, incubated with anti-mouse FITC-conjugated
horse antibody (Vector Laboratories, Burlingame,
CA) diluted 1/300 in PBS–BSA for 1 h at 37 jCand washed as before. Finally, the cells were
mounted onto slides in mounting medium with Citi-
fluor (Agar Scientific, Essex, UK), analysed by a
Nikon E400 epifluorescence microscope equipped
with objectives PlanFluor 20� , 40� and photo-
graphed. Images were acquired using a Nikon Cool-
pix 990 camera.
2.8. Immunoblotting
Cells grown in confluent monolayer were rinsed
twice with cold PBS and solubilised in ice-cold RIPA
buffer (1% Triton X-100 and 1% deoxycholate in
PBS) containing the commercial COMPLETE cock-
tail of protease inhibitors (Roche Diagnostics, Man-
nheim, Germany) for 30 min on ice. The extracts
were collected, cleared by centrifugation at 15000
rpm for 10 min at 4 jC and stored at � 80 jC.Protein concentrations of extracts were quantified
using the BCA protein assay reagent (Pierce, Rock-
ford, IL).
Total cellular extracts (50 Ag of proteins/lane) were
resolved in 10% SDS–PAGE gel under reducing
conditions. The proteins were then transferred to
polyvinylidene difluoride (PVDF) membrane (Amer-
sham Pharmacia Biotech, Little Chalfont Bucking-

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 123
hamshire, UK). After blocking in 5% non-fat dry milk
with 0.2% Nonidet P40 in PBS, the membrane was
probed with MAbs (undiluted hybridoma medium),
washed and treated with secondary anti-mouse HRP-
conjugated swine antibody diluted 1/7500 (Seva-
pharma). The protein bands were visualized by en-
hanced chemiluminiscence using the ECL kit
(Amersham Pharmacia Biotech).
2.9. Cell biotinylation
Cells in monolayers of 70%–80% confluence
grown in a 10-cm dish were washed with ice-cold
buffer A (20 mM sodium hydrogen carbonate, 0.15 M
NaCl, pH 8.0). Immediately before use, 1 mg of NHS-
LC-Biotin (Pierce) was dissolved in 50 Al DMSO,
mixed with 4 ml buffer A, added to cells and incu-
bated for 60 min at 4 jC. After biotinylation, the cellswere washed five times with buffer A and extracted as
described above.
2.10. Immunoprecipitation
The test MAb in a volume of 1 ml culture medium
was bound to 25 Al 50% suspension of Protein-A
Sepharose, or Protein-G Sepharose for MAbs of
IgG1 isotype (Pharmacia, Uppsala, Sweden) for 2 h at
RT. Biotinylated cell extract (200 Al) was pre-clearedwith 20 Al of 50% suspension of Protein-A/G Sephar-
ose and then added to the bound MAb. Immunocom-
plexes collected on Protein-A/G Sepharose were
washed according to Williams et al. (1985), boiled 5
min in Laemmli loading buffer and separated by SDS–
PAGE gel (10%) electrophoresis. Afterwards, the pro-
teins were transferred to a PVDF membrane and
revealed with peroxidase-conjugated streptavidin (1/
1000, Pierce) followed by ECL.
2.11. Immunohistochemistry (IHC)
Dissected tumours generated from HeLa cells
xenografted to nude mice (M. Rafajova, unpublished
observations) were fixed in 4% neutral-buffered
formaldehyde, dehydrated and embedded in paraffin.
Dewaxed and rehydrated 5-Am sections were immu-
nostained using the UltraTech HRP Steptavidin–
Biotin Universal Detection System (Immunotech,
Marseilles, France) according to the instructions of
the manufacturer. Briefly, the staining procedure was
performed at room temperature as follows. First,
endogenous peroxidase was blocked by 5 min of
incubation with 3% H2O2 in distilled water. The
sections were treated with the protein blocking agent
for 5 min, incubated for 1 h with MAb in hybridoma
medium diluted 1/10 in PBS containing 1% BSA and
washed with PBS. They were then incubated for 10
min with polyvalent biotinylated secondary antibody,
washed, incubated for 10 min with streptavidin–per-
oxidase reagent, washed again, visualized for 5–15
min in diaminobenzidine solution (Sigma) and, finally,
counterstained with hematoxylin.
2.12. Evaluation of cross-reactivity of anti-CA IX
MAbs to CA I, II and XII
Microplate wells were coated overnight with the
following antigens diluted in PBS: purified CA I (200
ng/well), purified CA II (100 ng/well) and recombi-
nant CA XII (100 ng/well). Then the coated wells
were incubated with undiluted culture media from
individual CA IX-specific hybridomas and with poly-
clonal sera against CA I, CA II and CA XII (all 1/
1000), respectively, as positive controls (Parkkila et
al., 1993; Karhumaa et al., 2000). Peroxidase-labelled
pig anti-mouse IgG and pig anti-rabbit IgG diluted 1/
5000 (Sevapharma) were used as detectors.
Alternatively, the proteins immunoprecipitated
with anti-CA IX MAbs from extracts of HT-29 and
HeLa cells as described above were subjected to
SDS–PAGE and blotting. Non-precipitated extracts
were loaded in parallel to establish the expression of
CA isoforms. The blots were treated with rabbit
polyclonal antisera specific for CA II and CA XII
(1/2000), respectively, followed by HRP-conjugated
pig anti-rabbit antibody diluted 1/5000 (Sevapharma).
Protein bands were visualized by ECL.
2.13. Biotinylation of MAbs
Purified MAbs were labelled with NHS-LC-Biotin
(Pierce) according to the instructions of the manufac-
turer. Briefly, 2 mg of purified IgG dissolved in PBS,
pH 8.0, was incubated with 100 Ag NHS-LC-Biotin
for 2 h on ice. Free biotin was removed using a
microconcentrator (PALL Gelman Lab., Wien, Aus-
tria) or gel filtration.
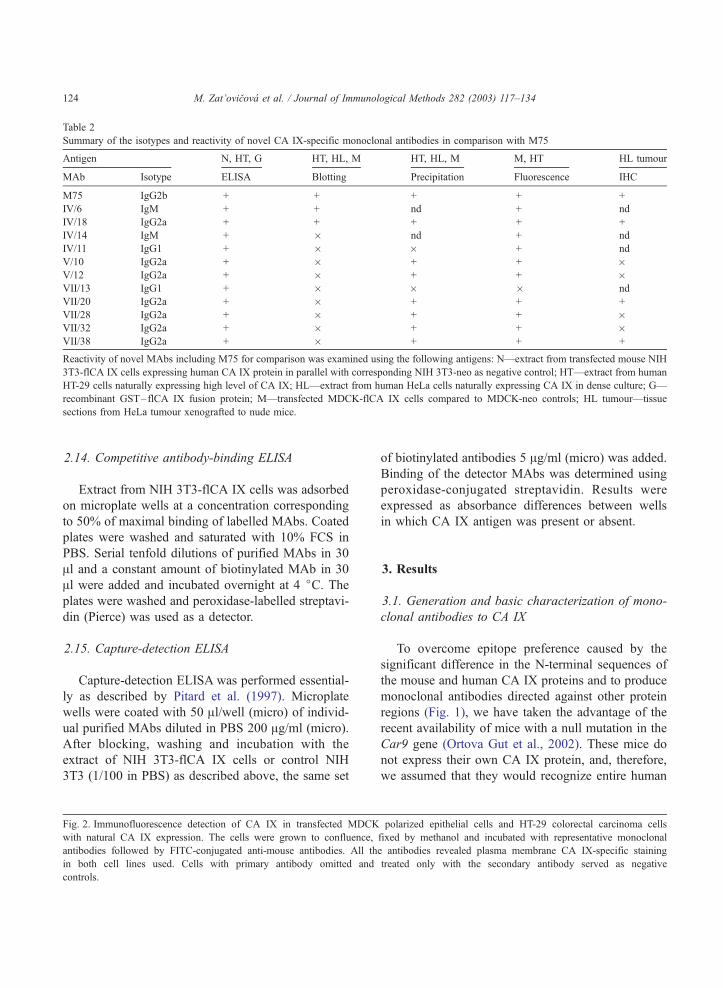
Table 2
Summary of the isotypes and reactivity of novel CA IX-specific monoclonal antibodies in comparison with M75
Antigen N, HT, G HT, HL, M HT, HL, M M, HT HL tumour
MAb Isotype ELISA Blotting Precipitation Fluorescence IHC
M75 IgG2b + + + + +
IV/6 IgM + + nd + nd
IV/18 IgG2a + + + + +
IV/14 IgM + � nd + nd
IV/11 IgG1 + � � + nd
V/10 IgG2a + � + + �V/12 IgG2a + � + + �VII/13 IgG1 + � � � nd
VII/20 IgG2a + � + + +
VII/28 IgG2a + � + + �VII/32 IgG2a + � + + �VII/38 IgG2a + � + + +
Reactivity of novel MAbs including M75 for comparison was examined using the following antigens: N—extract from transfected mouse NIH
3T3-flCA IX cells expressing human CA IX protein in parallel with corresponding NIH 3T3-neo as negative control; HT—extract from human
HT-29 cells naturally expressing high level of CA IX; HL—extract from human HeLa cells naturally expressing CA IX in dense culture; G—
recombinant GST–flCA IX fusion protein; M—transfected MDCK-flCA IX cells compared to MDCK-neo controls; HL tumour—tissue
sections from HeLa tumour xenografted to nude mice.
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134124
2.14. Competitive antibody-binding ELISA
Extract from NIH 3T3-flCA IX cells was adsorbed
on microplate wells at a concentration corresponding
to 50% of maximal binding of labelled MAbs. Coated
plates were washed and saturated with 10% FCS in
PBS. Serial tenfold dilutions of purified MAbs in 30
Al and a constant amount of biotinylated MAb in 30
Al were added and incubated overnight at 4 jC. Theplates were washed and peroxidase-labelled streptavi-
din (Pierce) was used as a detector.
2.15. Capture-detection ELISA
Capture-detection ELISA was performed essential-
ly as described by Pitard et al. (1997). Microplate
wells were coated with 50 Al/well (micro) of individ-
ual purified MAbs diluted in PBS 200 Ag/ml (micro).
After blocking, washing and incubation with the
extract of NIH 3T3-flCA IX cells or control NIH
3T3 (1/100 in PBS) as described above, the same set
Fig. 2. Immunofluorescence detection of CA IX in transfected MDCK
with natural CA IX expression. The cells were grown to confluence,
antibodies followed by FITC-conjugated anti-mouse antibodies. All th
in both cell lines used. Cells with primary antibody omitted and
controls.
of biotinylated antibodies 5 Ag/ml (micro) was added.
Binding of the detector MAbs was determined using
peroxidase-conjugated streptavidin. Results were
expressed as absorbance differences between wells
in which CA IX antigen was present or absent.
3. Results
3.1. Generation and basic characterization of mono-
clonal antibodies to CA IX
To overcome epitope preference caused by the
significant difference in the N-terminal sequences of
the mouse and human CA IX proteins and to produce
monoclonal antibodies directed against other protein
regions (Fig. 1), we have taken the advantage of the
recent availability of mice with a null mutation in the
Car9 gene (Ortova Gut et al., 2002). These mice do
not express their own CA IX protein, and, therefore,
we assumed that they would recognize entire human
polarized epithelial cells and HT-29 colorectal carcinoma cells
fixed by methanol and incubated with representative monoclonal
e antibodies revealed plasma membrane CA IX-specific staining
treated only with the secondary antibody served as negative

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 125
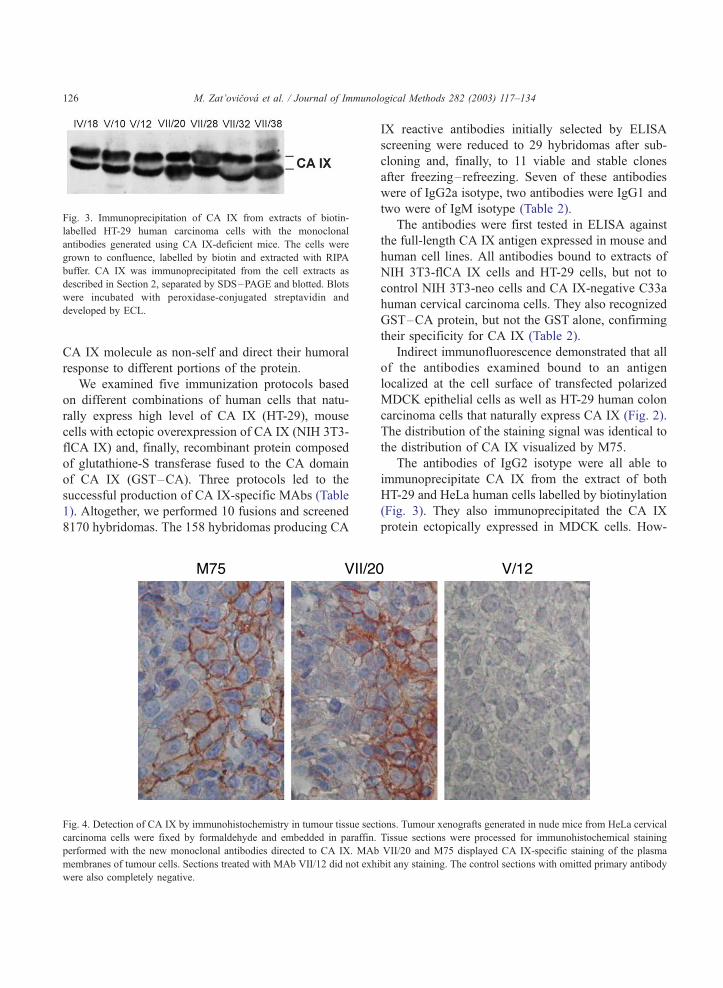
Fig. 3. Immunoprecipitation of CA IX from extracts of biotin-
labelled HT-29 human carcinoma cells with the monoclonal
antibodies generated using CA IX-deficient mice. The cells were
grown to confluence, labelled by biotin and extracted with RIPA
buffer. CA IX was immunoprecipitated from the cell extracts as
described in Section 2, separated by SDS–PAGE and blotted. Blots
were incubated with peroxidase-conjugated streptavidin and
developed by ECL.
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134126
CA IX molecule as non-self and direct their humoral
response to different portions of the protein.
We examined five immunization protocols based
on different combinations of human cells that natu-
rally express high level of CA IX (HT-29), mouse
cells with ectopic overexpression of CA IX (NIH 3T3-
flCA IX) and, finally, recombinant protein composed
of glutathione-S transferase fused to the CA domain
of CA IX (GST–CA). Three protocols led to the
successful production of CA IX-specific MAbs (Table
1). Altogether, we performed 10 fusions and screened
8170 hybridomas. The 158 hybridomas producing CA
Fig. 4. Detection of CA IX by immunohistochemistry in tumour tissue sect
carcinoma cells were fixed by formaldehyde and embedded in paraffin.
performed with the new monoclonal antibodies directed to CA IX. MAb
membranes of tumour cells. Sections treated with MAb VII/12 did not exh
were also completely negative.
IX reactive antibodies initially selected by ELISA
screening were reduced to 29 hybridomas after sub-
cloning and, finally, to 11 viable and stable clones
after freezing–refreezing. Seven of these antibodies
were of IgG2a isotype, two antibodies were IgG1 and
two were of IgM isotype (Table 2).
The antibodies were first tested in ELISA against
the full-length CA IX antigen expressed in mouse and
human cell lines. All antibodies bound to extracts of
NIH 3T3-flCA IX cells and HT-29 cells, but not to
control NIH 3T3-neo cells and CA IX-negative C33a
human cervical carcinoma cells. They also recognized
GST–CA protein, but not the GST alone, confirming
their specificity for CA IX (Table 2).
Indirect immunofluorescence demonstrated that all
of the antibodies examined bound to an antigen
localized at the cell surface of transfected polarized
MDCK epithelial cells as well as HT-29 human colon
carcinoma cells that naturally express CA IX (Fig. 2).
The distribution of the staining signal was identical to
the distribution of CA IX visualized by M75.
The antibodies of IgG2 isotype were all able to
immunoprecipitate CA IX from the extract of both
HT-29 and HeLa human cells labelled by biotinylation
(Fig. 3). They also immunoprecipitated the CA IX
protein ectopically expressed in MDCK cells. How-
ions. Tumour xenografts generated in nude mice from HeLa cervical
Tissue sections were processed for immunohistochemical staining
VII/20 and M75 displayed CA IX-specific staining of the plasma
ibit any staining. The control sections with omitted primary antibody
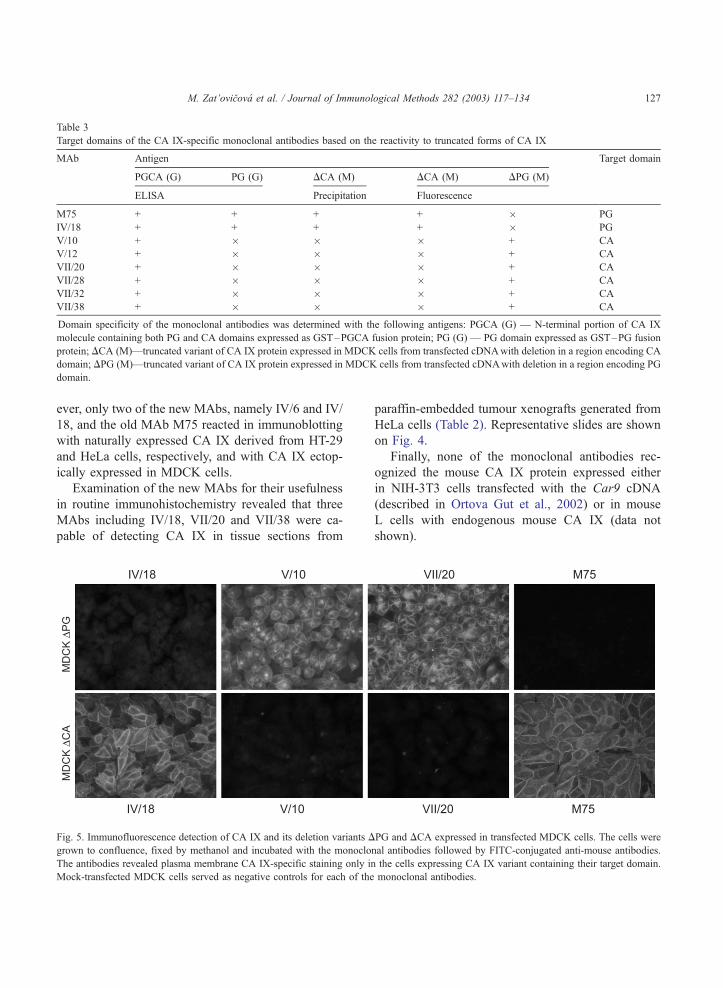
Table 3
Target domains of the CA IX-specific monoclonal antibodies based on the reactivity to truncated forms of CA IX
MAb Antigen Target domain
PGCA (G) PG (G) DCA (M) DCA (M) DPG (M)
ELISA Precipitation Fluorescence
M75 + + + + � PG
IV/18 + + + + � PG
V/10 + � � � + CA
V/12 + � � � + CA
VII/20 + � � � + CA
VII/28 + � � � + CA
VII/32 + � � � + CA
VII/38 + � � � + CA
Domain specificity of the monoclonal antibodies was determined with the following antigens: PGCA (G) — N-terminal portion of CA IX
molecule containing both PG and CA domains expressed as GST–PGCA fusion protein; PG (G) — PG domain expressed as GST–PG fusion
protein; DCA (M)—truncated variant of CA IX protein expressed in MDCK cells from transfected cDNAwith deletion in a region encoding CA
domain; DPG (M)—truncated variant of CA IX protein expressed in MDCK cells from transfected cDNAwith deletion in a region encoding PG
domain.
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 127
ever, only two of the new MAbs, namely IV/6 and IV/
18, and the old MAb M75 reacted in immunoblotting
with naturally expressed CA IX derived from HT-29
and HeLa cells, respectively, and with CA IX ectop-
ically expressed in MDCK cells.
Examination of the new MAbs for their usefulness
in routine immunohistochemistry revealed that three
MAbs including IV/18, VII/20 and VII/38 were ca-
pable of detecting CA IX in tissue sections from
Fig. 5. Immunofluorescence detection of CA IX and its deletion variants D
grown to confluence, fixed by methanol and incubated with the monoclo
The antibodies revealed plasma membrane CA IX-specific staining only in
Mock-transfected MDCK cells served as negative controls for each of the
paraffin-embedded tumour xenografts generated from
HeLa cells (Table 2). Representative slides are shown
on Fig. 4.
Finally, none of the monoclonal antibodies rec-
ognized the mouse CA IX protein expressed either
in NIH-3T3 cells transfected with the Car9 cDNA
(described in Ortova Gut et al., 2002) or in mouse
L cells with endogenous mouse CA IX (data not
shown).
PG and DCA expressed in transfected MDCK cells. The cells were
nal antibodies followed by FITC-conjugated anti-mouse antibodies.
the cells expressing CA IX variant containing their target domain.
monoclonal antibodies.
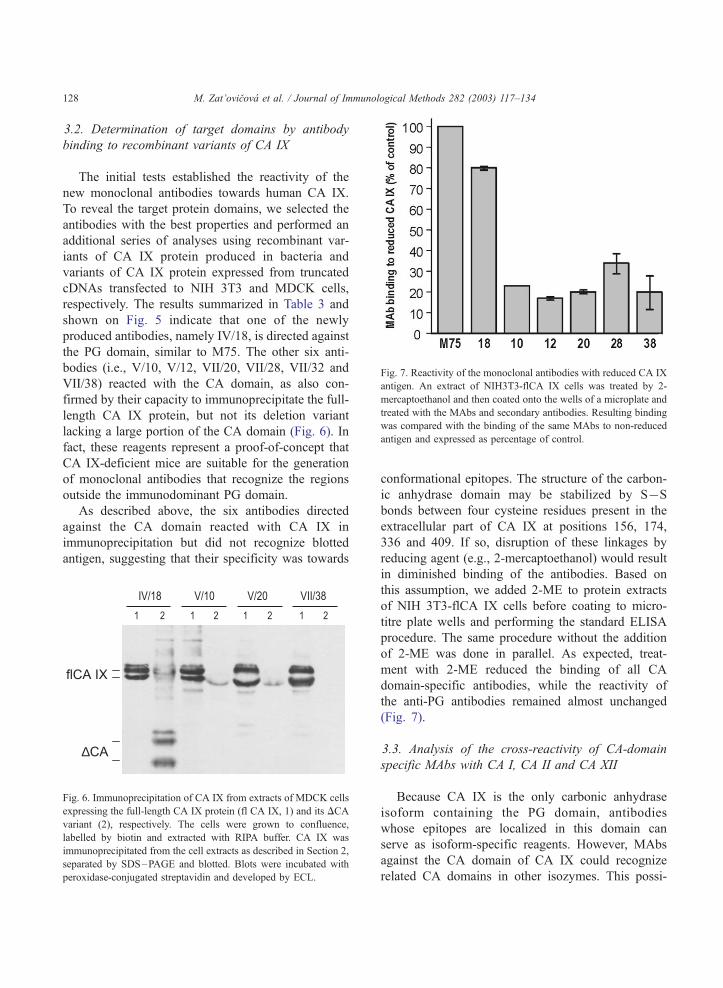
Fig. 7. Reactivity of the monoclonal antibodies with reduced CA IX
antigen. An extract of NIH3T3-flCA IX cells was treated by 2-
mercaptoethanol and then coated onto the wells of a microplate and
treated with the MAbs and secondary antibodies. Resulting binding
was compared with the binding of the same MAbs to non-reduced
antigen and expressed as percentage of control.
M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134128
3.2. Determination of target domains by antibody
binding to recombinant variants of CA IX
The initial tests established the reactivity of the
new monoclonal antibodies towards human CA IX.
To reveal the target protein domains, we selected the
antibodies with the best properties and performed an
additional series of analyses using recombinant var-
iants of CA IX protein produced in bacteria and
variants of CA IX protein expressed from truncated
cDNAs transfected to NIH 3T3 and MDCK cells,
respectively. The results summarized in Table 3 and
shown on Fig. 5 indicate that one of the newly
produced antibodies, namely IV/18, is directed against
the PG domain, similar to M75. The other six anti-
bodies (i.e., V/10, V/12, VII/20, VII/28, VII/32 and
VII/38) reacted with the CA domain, as also con-
firmed by their capacity to immunoprecipitate the full-
length CA IX protein, but not its deletion variant
lacking a large portion of the CA domain (Fig. 6). In
fact, these reagents represent a proof-of-concept that
CA IX-deficient mice are suitable for the generation
of monoclonal antibodies that recognize the regions
outside the immunodominant PG domain.
As described above, the six antibodies directed
against the CA domain reacted with CA IX in
immunoprecipitation but did not recognize blotted
antigen, suggesting that their specificity was towards
flCA IX
∆CA
IV/18 V/10 V/20 VII/38
1 1 12 2 2 1 2
Fig. 6. Immunoprecipitation of CA IX from extracts of MDCK cells
expressing the full-length CA IX protein (fl CA IX, 1) and its DCA
variant (2), respectively. The cells were grown to confluence,
labelled by biotin and extracted with RIPA buffer. CA IX was
immunoprecipitated from the cell extracts as described in Section 2,
separated by SDS–PAGE and blotted. Blots were incubated with
peroxidase-conjugated streptavidin and developed by ECL.
conformational epitopes. The structure of the carbon-
ic anhydrase domain may be stabilized by SUS
bonds between four cysteine residues present in the
extracellular part of CA IX at positions 156, 174,
336 and 409. If so, disruption of these linkages by
reducing agent (e.g., 2-mercaptoethanol) would result
in diminished binding of the antibodies. Based on
this assumption, we added 2-ME to protein extracts
of NIH 3T3-flCA IX cells before coating to micro-
titre plate wells and performing the standard ELISA
procedure. The same procedure without the addition
of 2-ME was done in parallel. As expected, treat-
ment with 2-ME reduced the binding of all CA
domain-specific antibodies, while the reactivity of
the anti-PG antibodies remained almost unchanged
(Fig. 7).
3.3. Analysis of the cross-reactivity of CA-domain
specific MAbs with CA I, CA II and CA XII
Because CA IX is the only carbonic anhydrase
isoform containing the PG domain, antibodies
whose epitopes are localized in this domain can
serve as isoform-specific reagents. However, MAbs
against the CA domain of CA IX could recognize
related CA domains in other isozymes. This possi-

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 129
bility raised the need to evaluate the cross-reactivity
of the anti-CA MAbs especially to those isoforms
that may be potentially co-expressed with CA IX.
Therefore, we have tested in ELISA the binding of
our monoclonal antibodies to purified CA I, CA II
and CA XII antigens in comparison to polyclonal
sera specific for these antigens. We found that none
of the CA IX-specific MAbs bound to any of the
heterologous CA isoforms. This finding was verified
by an experiment in which protein extracts from
HT-29 carcinoma cells that express several CA iso-
forms, including CA II, CA IX and CA XII (as was
confirmed by immunoblotting), were first immuno-
precipitated by CA IX-specific MAbs, blotted and
immunodetected with anti-CA II and anti CA XII
antisera, respectively. Neither CA II nor CA XII
were coprecipitated with CA IX by CA IX-specific
antibodies, confirming the lack of their cross-reac-
tivity at least with these isoforms (data not shown).
3.4. Mapping of antigenic sites by competitive
binding of MAbs
In order to determine the relative positions of the
antigenic sites recognized by the monoclonal anti-
bodies within their CA IX target domains, we per-
formed a competitive-binding assay. Biotin-labelled
purified antibodies were allowed to bind in the pres-
ence of increasing amounts of non-labelled competi-
tive antibodies. The extent of binding of the labelled
antibody in the presence of the non-labelled compet-
itor was expressed as percentage of binding in the
absence of the competitor. Examination of all different
pairs of the MAbs including the homologous ones and
their classification according to the extent of the
competition allowed the preparation of a basic map
indicating principal relationships between the antibod-
Fig. 8. Chequer-board maps of antigenic sites delineated on the
basis of the competitive binding ELISA (A) and capture-detection
ELISA (B). (A) Antigen-coated plates were incubated with fixed
amount of biotin-labelled antibody and increasing amounts of non-
labelled competitor antibody. Results are expressed as percentage of
absorbance measured in the absence of competitor antibody. (B)
Plates coated with capture antibody were incubated with antigen in
the cellular extract and then with biotinylated detector antibody.
Absorbance resulting from the cooperative binding of capture-
detector antibodies indicates spatial separation between the epitopes
of the paired antibodies.
ies, their clustering and relationships between the
clusters (Fig. 8A). The map shows that the M75 and
IV/18 antibodies recognize an antigenic site in a
separate region, consistent with their proven specific-

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134130
ity for the PG domain of CA IX protein. The anti-
bodies targeting the CA domain form three clusters.
Two pairs of MAbs, namely V/12 versus VII/20 and V/
10 versus VII/32, respectively, showed significant
mutual competitions that define two antigenic sites.
However, non-reciprocal competition between V/10
(label) and V/12 (competitor) indicated that the two
sites may partially extend over each other but with
differences in antibody affinity. Alternatively, binding
of the first antibody could cause an allosteric change
that prevented binding of the second one. MAb VII/38
belongs to a distinct site within the CA domain of
CA IX.
3.5. Relationships between the monoclonal antibodies
in a capture-detection assay
Clustering of the MAbs was further explored by a
sandwich ELISA in which each MAb was used
either as a plastic-coated capture or as soluble bio-
tinylated detector. This arrangement permitted spatial
orientation of the antigen bound to the capture
antibody via one antigenic site and exposure of a
distinct non-overlapping antigenic site recognized by
the detector antibody. A map of the antigenic sites
deduced from this analysis principally confirmed the
relationships among the tested antibodies with minor
differences (Fig. 8B). These could be assigned to
differences in the constitution of the assay that
generates a spatial limitation to binding of the
detector antibody to its epitope if it is localized close
to the epitope of the capture MAb. The data obtained
were evaluated on the basis of the absorbance differ-
ences resulting from the cooperation between the
capture and the detector in the presence and absence
of antigen, with the higher absorbance difference
value indicating the bigger mutual distance of the
epitopes on the folded protein molecule. On this
basis, the antigenic sites for the anti-PG antibodies
were clearly separated from the antigenic sites of the
anti-CA antibodies in accord with the competitive
assay. Moreover, the antigenic sites of VII/20 and V/
12 MAbs seemed to localize closer to the PG
domain than those of V/10, VII/28 and VII/32,
although the latter MAb appeared to interfere with
the binding of VII/20 and V/12 when used as a
capture. MAb VII/38 produced inconsistent results
and was withdrawn from this analysis.
4. Discussion
Due to a strong association with cancer tissues and
a proposed functional involvement in tumour progres-
sion, carbonic anhydrase IX has become an interesting
molecule for both basic and clinical research, as
evidenced by the growing number of CA IX-related
papers (summarized in Pastorekova and Pastorek,
2003). Most of these papers have been based on the
utilization of a CA IX-specific monoclonal antibody
M75 (Pastorekova et al., 1992). Although M75 has
facilitated the acquisition of important data, certain
aspects of CA IX could not be evaluated without
additional detection tools, including the monoclonal
antibodies directed to antigenic sites that do not
interfere with that of M75.
In this paper, we describe the generation of such
antibodies using mice deficient for CA IX as donors
of immune splenocytes for fusion and hybridoma
production. This strategy has been chosen in order
to eliminate the epitope preference possibly related to
a high regional homology between the mouse and
human proteins particularly in the sequences outside
the M75 target domain (Ortova Gut et al., 2002). Our
assumption that the mice lacking CA IX would also
recognize these regions as non-self has been con-
firmed by the production of six monoclonal antibodies
directed against the CA domain of CA IX. Interest-
ingly, the immunization protocol that led to the
successful generation of the anti-CA antibodies in-
volved the injection of a recombinant CA IX variant
containing only the CA domain as a key third-dose
component. Instead, injection with the full-length CA
IX antigen extracted from the transfected NIH 3T3
cells resulted in the production of antibodies, among
which MAb IV/18 competes with M75 and two other
antibodies (IV/6 and IV/14) also appear to bind to the
PG region, similar to M75 (not shown). This experi-
ence suggests that even in the absence of endogenous
mouse CA IX protein that would normally compro-
mise the immune recognition of highly homologous
regions of human CA IX (including the CA domain)
by the immunized mice, the M75 MAb-binding PG
domain is strongly immunodominant. The reason for
the apparent immunodominance could reside in both
the N-terminal location and the amino acid sequence
of the PG domain that is highly acidic and composed
of four perfect and two imperfect hexameric repeats

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 131
(Opavsky et al., 1996). These repetitions potentially
offer multiple binding sites and the very N-terminal
position of this region may allow for its spatial
protrusion from the presumably globular CA domain
in the extracellular portion of CA IX (Pastorek et al.,
1994).
These attributes of the PG domain may also
explain the finding that anti-PG MAbs recognize
denatured CA IX protein in immunoblotting, while
anti-CA MAbs fail to do so. One may speculate that a
relatively short PG domain with repetitive motifs
constitutes a linear antigenic region that is not sus-
ceptible to denaturation, while a presumably globular
structure of the large CA domain carrying putative
conformational epitopes can be completely disrupted.
Indeed, reduction of SUS bond between cysteine
residues in the extracellular portion of CA IX perturbs
recognition of these CA domain-related epitopes
thereby supporting this idea. It is also consistent with
the finding that both anti-PG and anti-CA are capable
of immunoprecipitating native CA IX antigen
extracted with mild detergents as well as the antigen
on the surface of transfected MDCK cells that were
fixed by methanol. Such antigen apparently possesses
intact epitopes irrespective of whether they are linear
or conformational.
As delineated by the competitive immunoassay
performed with the repertoire of new MAbs, the CA
domain contains three distinct antigenic sites, two of
which partially overlap. While it is possible to pro-
pose relative mutual positions for these sites, our data
do not permit prediction of their localization on the
CA domain. Based on the extent of the deletion
associated with the loss of binding of these MAbs,
we can at least conclude that the CA antigenic region
spans the amino acids 166–397 of CA IX. However,
it cannot be excluded that the deletion led to disloca-
tion of the antigenic site(s) partly located also on the
amino acids 135–165 that remained preserved in the
DCA deletion variant.
Furthermore, the data from capture-detection anal-
ysis indicate that the antigenic area reactive with
MAbs VII/32, V/10 and VII/28 is localized more
distantly with respect to the PG domain, when com-
pared to the antigenic area reacting with V/12 and VII/
20 MAbs. However, it is difficult to reach any
definitive conclusion regarding the mutual distance
and relationship between the antigenic sites of the CA
domain because of their conformational character.
Nevertheless, based on our data, a combination of
anti-PG MAbs M75 and IV/18 with the anti-CA
MAbs V/10 and VII/32 appears most promising for
CA IX capture-detection purposes.
The characteristics of the new CA IX-specific
monoclonal antibodies suggest their possible appli-
cations. In basic research, these antibodies represent
important reagents for the study of functional con-
tributions of the PG and CA domains, respectively.
Each of these two extracellular domains of CA IX
has been associated with different aspects of tumour
progression. The catalytically active CA domain is
thought to contribute to acidification of the tumour
microenvironment whereas the PG domain (absent
from other CA isoforms) is believed to determine
participation in adhesion-related processes. Using
the present set of MAbs, it will be possible to
differentiate between biological effects exerted by
these domains expressed separately in the form of
deletion variants or mutants of CA IX. The exclu-
sive reactivity of these MAbs with CA IX but not
with the other CAs tested here further increases
their research and clinical value.
The main potential of the new MAbs is in their
clinical application. Many clinical studies of different
tumour tissues have recently demonstrated the predic-
tive and prognostic value of CA IX, especially in
relation to tumour hypoxia (Loncaster et al., 2001;
Chia et al., 2001; Giatromanolaki et al., 2001; Kou-
kourakis et al., 2001; Hui et al., 2002). In this respect,
it is obvious that the new MAbs can be utilized for
detection and/or targeting of CA IX-expressing cancer
cells in different settings (as reviewed in von Mehren
et al., 2003). The latter option seems especially prom-
ising given the capacity of all the MAbs to recognize
CA IX antigen in its native form as confirmed in this
paper. In addition, three of the new MAbs, namely the
PG domain-specific IV/18 and the CA domain-specific
VII/20 and VII/38 antibodies can be used in immuno-
histochemical studies of cancer tissue sections. More-
over, future experiments designed to examine the
biological activity of these MAbs will show whether
they could be of any significance for antibody-medi-
ated anti-cancer therapy, analogous to the use of well-
known MAbs directed against the oncoproteins ErbB2
and EGFR (epidermal growth factor receptor). Last but
not least, the availability of non-competing antibodies

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134132
specific for distinct antigenic sites on two separate
extracellular domains offers an opportunity to elabo-
rate a sensitive assay that could be particularly impor-
tant for the detection of CA IX in the body fluids of
cancer patients. It has been shown recently that CA IX
is shed from the tumour cell surface and can be
detected in the blood and urine of cancer patients by
immunoprecipitation (Zavada et al., 2003; Chrastina et
al., 2003). Development of a fast and reliable micro-
assay based on a combination of the antibodies de-
scribed here could potentially permit non-invasive
monitoring of cancer patients. In summary, the anti-
human CA IX monoclonal antibodies generated using
CA IX-deficient mice and characterized in this paper
represent important tools for improving our knowledge
of structure–function relationships in the CA IX
molecule. They should provide a better understanding
of the role of CA IX in cancer development and
improve clinically relevant detection of CA IX in
biological materials.
Acknowledgements
This work was supported by grants from Bayer,
from the Slovak Scientific Grant Agency (VEGA-2/
3055/23 and 2025/22) and from the Science and
Technology Assistance Agency (contract APVT-51-
005802). SPc is a recipient of a grant from Sigrid
Juselius Foundation.
References
Bartosova, M., Parkkila, S., Pohlodek, K., Karttunen, T.J., Galbavy,
S., Mucha, V., Harris, A.L., Pastorek, J., Pastorekova, S., 2002.
Expression of carbonic anhydrase IX in breast is associated with
malignant tissues and related to overexpression of c-erbB2.
J. Pathol. 197, 314–321.
Beasley, N.J.P., Wykoff, C.C., Watson, P.H., Leek, R., Turley, H.,
Gatter, K., Pastorek, J., Cox, G.J., Ratcliffe, P., Harris, A.L.,
2001. Carbonic anhydrase IX, an endogenous hypoxia marker,
expression in head and neck squamous cell carcinoma and its
relationship to hypoxia, necrosis and microvessel density. Can-
cer Res. 61, 5262–5267.
Chia, S.K., Wykoff, C.C., Watson, P.H., Han, C., Leek, R.D., Pas-
torek, J., Gatter, K.C., Ratcliffe, P., Harris, A.L., 2001. Prognos-
tic significance of a novel hypoxia-regulated marker, carbonic
anhydrase IX, in invasive breast carcinoma. J. Clin. Oncol. 19,
3660–3668.
Chrastina, A., Zavada, J., Parkkila, S., Kaluz, S., Kaluzova, M.,
Rajcani, J., Pastorek, J., Pastorekova, S., 2003. Biodistribution
and pharmacokinetics of 125I-labeled monoclonal antibody M75
specific for carbonic anhydrase IX, an intrinsic marker of hypo-
xia, in nude mice xenografted with human colorectal carcinoma.
Int. J. Cancer 105, 873–881.
Giatromanolaki, A., Koukourakis, M.I., Sivridis, E., Pastorek, J.,
Wykoff, C.C., Gatter, K.C., Harris, A.L., 2001. Expression of
hypoxia-inducible carbonic anhydrase-9 relates to angiogenic
pathways and independently to poor outcome in non-small cell
lung cancer. Cancer Res. 61, 7992–7998.
Gibadulinova, A., Zelnık, V., Reiserova, L., Zavodska, E.,
Zat’ovicova, M., Ciampor, F., Pastorekova, S., Pastorek, J.,
1998. Sequence and characterization of the Z gene encoding
ring finger protein of the lymphocytic choriomeningitis virus
MX strain. Acta Virol. 42, 369–374.
Hockel, M., Vaupel, P., 2001. Tumour hypoxia: definitions and
current clinical, biologic and molecular aspects. J. Natl. Cancer
Inst. 93, 266–276.
Hui, E.P., Chan, A.T., Pezzella, F., Turley, H., To, K.F., Poon, T.C.,
Zee, B., Mo, F., Teo, P.M., Huang, D.P., Gatter, K.C., Johnson,
P.J., Harris, A.L., 2002. Coexpression of hypoxia-inducible fac-
tors 1alpha and 2alpha, carbonic anhydrase IX, and vascular en-
dothelial growth factor in nasopharyngeal carcinoma and
relationship to survival. Clin. Cancer Res. 8, 2595–2604.
Ivanov, S.V., Kuzmin, I., Wei, M.H., Pack, S., Geil, L., Johnson,
B.E., Stanbridge, E.J., Lerman, M.I., 1998. Down-regulation of
transmembrane carbonic anhydrases in renal cell carcinoma cell
lines by wild-type von Hippel–Lindau transgenes. Proc. Natl.
Acad. Sci. U. S. A. 95, 12596–12601.
Ivanov, S., Liao, S.Y., Ivanova, A., Danilkovich-Miagkova, A.,
Tarasova, N., Weirich, G., Merrill, M.J., Proescholdt, M.A.,
Oldfield, E.H., Lee, J., Zavada, J., Waheed, A., Sly, W., Lerman,
M.I., Stanbridge, E.J., 2001. Expression of hypoxia-inducible
cell-surface transmembrane carbonic anhydrases in human can-
cer. Am. J. Pathol. 158, 905–919.
Kaluz, S., Kaluzova, M., Opavsky, R., Pastorekova, S., Gibaduli-
nova, A., Dequiedt, F., Kettmann, R., Pastorek, J., 1999. Tran-
scriptional regulation of the MN/CA9 gene coding for the
tumour-associated carbonic anhydrase IX. Identification and
characterization of a proximal silencer element. J. Biol. Chem.
274, 32588–32595.
Kaluz, S., Kaluzova, M., Chrastina, A., Olive, P.L., Pastorekova, S.,
Pastorek, J., Lerman, M.I., Stanbridge, E.J., 2002. Lowered oxy-
gen tension induces expression of the hypoxia marker MN/car-
bonic anhydrase IX in the absence of hypoxia-inducible factor
1a stabilization: a role for phosphatidylinositol 3V-kinase. Can-cer Res. 62, 4469–4477.
Karhumaa, P., Parkkila, S., Tureci, O., Waheed, A., Grubb, J.H.,
Shah, G., Parkkila, A., Kaunisto, K., Tapanainen, J., Sly, W.S.,
Rajaniemi, H., 2000. Identification of carbonic anhydrase XII as
the membrane isozyme expressed in the normal human endo-
metrial epithelium. Mol. Hum. Reprod. 6, 68–74.
Koukourakis, M.I., Giatromanolaki, A., Sivridis, E., Simopoulos,
K., Pastorek, J., Wykoff, C.C., Gatter, K.C., Harris, A.L., 2001.
Hypoxia-regulated carbonic anhydrase-9 (CA9) relates to poor
vascularization and resistance of squamous cell head and neck
cancer to chemoradiotherapy. Clin. Cancer Res. 7, 3399–3403.

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134 133
Lane, R.D., Crissman, R.S., Ginn, S., 1986. High efficiency fusion
for producing monoclonal antibodies against weak immuno-
gens. Methods Enzymol. 121, 183–192.
Liao, S.Y., Brewer, C., Zavada, J., Pastorek, J., Pastorekova, S.,
Manetta, A., Berman, M.L., DiSaia, P.J., Stanbridge, E.J.,
1994. Identification of the MN antigen as a diagnostic bio-
marker of cervical intraepithelial neoplasia and cervical carcino-
ma. Am. J. Pathol. 145, 598–609.
Liao, S.Y., Aurelio, O.N., Jan, K., Zavada, J., Stanbridge, E.J.,
1997. Identification of the MN/CA9 protein as a reliable diag-
nostic biomarker of clear cell carcinoma of the kidney. Cancer
Res. 57, 2827–2831.
Lieskovska, J., Opavsky, R., Zacikova, L., Glasova, M., Pastorek,
J., Pastorekova, S., 1999. Study of in vitro conditions modulat-
ing expression of MN/CA IX protein in human cell lines derived
from cervical carcinoma. Neoplasma 46, 17–24.
Loncaster, J.A., Harris, A.L., Davidson, S.E., Logue, J.P., Hunter,
R.D., Wykoff, C.C., Pastorek, J., Ratcliffe, P., Stratford, I.J.,
West, C.M.L., 2001. Carbonic anhydrase IX expression, a po-
tential new intrinsic marker of hypoxia: correlations with tu-
mour oxygen measurements and prognosis in locally advanced
carcinoma of the cervix. Cancer Res. 61, 6394–6399.
Nishimori, I., 2003. Acatalytic CAs: Carbonic Anhydrase Related
Proteins (CARPs). In: Supuran, C.T., Scozzafava, A., Conway,
J. (Eds.), Carbonic Anhydrase, Its Inhibitors and Activators.
Taylor and Francis, London. In press.
Olive, P.L., Aquino-Parsons, C., MacPhail, S.H., Laio, S.Y., Ra-
leigh, J.A., Lerman, M.I., Stanbridge, E.J., 2001. Carbonic an-
hydrase 9 as an endogenous marker for hypoxic cells in cervical
cancer. Cancer Res. 61, 8924–8929.
Opavsky, R., Pastorekova, S., Zelnık, V., Gibadulinova, A., Stan-
bridge, E.J., Zavada, J., Kettmann, R., Pastorek, J., 1996. Hu-
man MN/CA9 gene, a novel member of the carbonic anhydrase
family: structure and exon to protein domain relationship. Ge-
nomics 33, 480–487.
Ortova Gut, M., Parkkila, S., Vernerova, Z., Rohde, E., Zavada, J.,
Hocker, M., Pastorek, J., Karttunen, T., Gibadulinova, A., Za-
vadova, Z., Knobeloch, K.P., Wiedenmann, B., Svoboda, J.,
Horak, I., Pastorekova, S., 2002. Gastric hyperplasia in mice
with targeted disruption of the carbonic anhydrase gene Car9.
Gastroenterology 123, 1889–1903.
Parkkila, S., 2000. An overview of the distribution and function of
carbonic anhydrase in mammals. In: Chegwidden, W.R., Carter,
N., Edwards, Y. (Eds.), The Carbonic Anhydrases: New Hori-
zons. Birkhauser, Basil, pp. 76–93.
Parkkila, A.K., Parkkila, S., Juvonen, T., Rajaniemi, H., 1993. Car-
bonic anhydrase isoenzymes II and I are present in the zona
glomerulosa cells of the human adrenal gland. Histochemistry
99, 37–41.
Pastorek, J., Pastorekova, S., Callebaut, I., Mornon, J.P., Zelnık,
V., Opavsky, R., Zat’ovicova, M., Liao, S., Portetelle, D., Stan-
bridge, E.J., Zavada, J., Burny, A., Kettmann, R., 1994. Clon-
ing and characterization of MN, a human tumour-associated
protein with a domain homologous to carbonic anhydrase
and a putative helix– loop–helix DNA binding segment. On-
cogene 9, 2788–2888.
Pastorekova, S., Pastorek, J., 2003. Cancer-related carbonic anhy-
drase isozymes and their inhibition. In: Supuran, C.T., Scozza-
fava, A., Conway, J. (Eds.), Carbonic Anhydrase, Its Inhibitors
and Activators. Taylor and Francis, London. In press.
Pastorekova, S., Zavadova, Z., Kost’al, M., Babusıkova, O., Zava-
da, J., 1992. A novel quasi-viral agent, MaTu, is a two-compo-
nent system. Virology 187, 620–626.
Pastorekova, S., Parkkila, S., Parkkila, A.K., Opavsky, R., Zelnık,
V., Saarnio, J., Pastorek, J., 1997. Carbonic anhydrase IX, MN/
CA IX: analysis of stomach complementary DNA sequence and
expression in human and rat alimentary tracts. Gastroenterology
112, 398–408.
Pitard, V., Taupin, J.L., Miossec, V., Blanchard, F., Cransac, M.,
Jollet, I., Vernallis, A., Hudson, K., Godard, A., Jacques, Y.,
Moreau, J.F., 1997. Production and characterization of monoclo-
nal antibodies against the leukemia inhibitory factor low affinity
receptor, gp190. J. Immunol. Methods 205, 177–190.
Saarnio, J., Parkkila, S., Parkkila, A.K., Haukipuro, K., Pastoreko-
va, S., Pastorek, J., Kairaluoma, M.I., Karttunen, T.J., 1998.
Immunohistochemical study of colorectal tumours for expres-
sion of a novel transmembrane carbonic anhydrase, MN/CA IX,
with potential value as a marker of cell proliferation. Am. J.
Pathol. 153, 279–285.
Saarnio, J., Parkkila, S., Parkkila, A.K., Pastorekova, S., Hauki-
puro, K., Pastorek, J., Juvonen, T., Karttunen, T., 2001. Trans-
membrane carbonic anhydrase, MN/CA IX, is a potential
biomarker for biliary tumours. J. Hepatol. 35, 643–649.
Supuran, C.T., Scozzafava, A., 2000. Carbonic anhydrase inhib-
itors and their therapeutic potential. Expert Opin. Ther. Pat. 10,
575–600.
Svastova, E., Zilka, N., Zat’ovicova, M., Gibadulinova, A., Ciam-
por, F., Pastorek, J., Pastorekova, S., 2003. Carbonic anhydrase
IX reduces E-cadherin-mediated adhesion of MDCK cells via
interaction with h-catenin. Exp. Cell Res. (in press).
Turner, J.R., Odze, R.D., Crum, C.P., Resnick, M.B., 1997. MN
antigen expression in normal, preneoplastic and neoplastic
esophagus: a clinicopathological study of a new cancer-associ-
ated biomarker. Human Pathol. 28, 740–744.
Vermylen, P., Roufosse, C., Burny, A., Verhest, A., Bossehaerts, T.,
Pastorekova, S., Ninane, V., Sculier, J.P., 1999. Carbonic anhy-
drase IX antigen differentiates between preneoplastic and ma-
lignant lesions in non-small cell lung carcinomas. Eur. Respir. J.
14, 806–811.
von Mehren, M., Adams, G.P., Weiner, L.M., 2003. Monoclonal
antibody therapy for cancer. Annu. Rev. Med. 54, 343–369.
Williams, D.B., Swiedler, S.J., Hart, G.W., 1985. Intracellular trans-
port of membrane glycoproteins: two closely related histocom-
patibility antigens differ in their rates of transit to the cell
surface. J. Cell Biol. 101, 725–734.
Wingo, T., Tu, C., Laipis, P.J., Silverman, D.N., 2001. The catalytic
properties of human carbonic anhydrase IX. Biochem. Biophys.
Res. Commun. 288, 666–669.
Wykoff, C., Beasley, N., Watson, P., Turner, L., Pastorek, J., Wil-
son, G., Turley, H., Maxwell, P., Pugh, C., Ratcliffe, P., Harris,
A., 2000. Hypoxia-inducible regulation of tumour-associated
carbonic anhydrases. Cancer Res. 60, 7075–7083.
Wykoff, C.C., Beasley, N., Watson, P.H., Campo, L., Chia, S.K.,
English, R., Pastorek, J., Sly, W.S., Ratcliffe, P., Harris, A.L.,

M. Zat’ovicova et al. / Journal of Immunological Methods 282 (2003) 117–134134
2001. Expression of the hypoxia-inducible and tumour-associ-
ated carbonic anhydrases in ductal carcinoma in situ of the
breast. Am. J. Pathol. 158, 1011–1019.
Zavada, J., Zavadova, Z., Pastorekova, S., Ciampor, F., Pastorek, J.,
Zelnık, V., 1993. Expression of MaTu-MN protein in human
tumour cultures and in clinical specimens. Int. J. Cancer 54,
268–274.
Zavada, J., Zavadova, Z., Pastorek, J., Biesova, Z., Jezek, K., Velek,
J., 2000. Human tumour-associated cell adhesion protein MN/
CA IX: identification of M75 epitope and of the region media-
ting cell adhesion. Br. J. Cancer 82, 1808–1813.
Zavada, J., Zavadova, Z., Zat’ovicova, M., Hyrsl, L., Kawaciuk,
I., 2003. Soluble form of carbonic anhydrase IX (CA IX) in
the serum and urine of renal carcinoma patients. Br. J. Cancer
89, 1067–1071.
Related Documents











