P.S.Z.N.: Marine Ecology, 18 (4): 361-378 (1997) 0 1997 Blackwell Wissenschafts-Verlag, Berlin ISSN 0173-9565 Accepted: June 20, 1997 Molluscan Hard-Substrate Associations in the Northern Red Sea M. ZUSCHIN & W. E. PILLER Institut filr PalSiontologie, Universitat Wien, Geozentrum, Althanstrasse 14, A-1090 Vienna. Austria. With 3 figures and 3 tables Key words: Northern Bay of Safaga, reef flats, tidal flats, scleractinian carpets, Sarcophyion carpets. Abstract. The mollusc associations of 29 hard-substrate localities in the Northern Bay of Safaga (Red Sea, Egypt) were studied. The hard substrates were classified into 7 types: reef flats, intertidal rock bottom, very shallow subtidal conglomerate, Porifes carpet, faviid carpet, platy scleractinian associ- ation, Sarcophyron carpet. Based on a cluster analysis the molluscs of these hard substrates can be classified into 5 groups: Group A is dominated by Dendropoma maxima, Chamoidea and Tridacna maxima, group B by Pedum spondyloideum, T. maxima and Coralliophila neriioidea, group C by Chamoidea and Ceriihium ruppelli, group D by Modiolus auriculaius, Chamoidea, Morula yranulara and Neriia albicilla and group E by Chamoidea and Pirar hebraea. The latter group contains only samples which are generally poor in molluscs. Group A molluscs are diagnostic for reef flats, group B for Purites and faviid carpets, group C for Sarcophyton carpets and group D for tidal/shallow subtidal rock substrates. The different hard substrates are therefore characterized by diagnostic mollusc associ- ations and some of them could also be identified in the fossil record. Problem Ecological and paleoecological studies in tropical-subtropical reefoidal environ- ments typically focus on corals and coral communities. Other than this invertebrate group, molluscs are also frequently found inhabiting a variety of ecological niches, and like corals, have considerable fossilization potential. In this context, however, only minor attention has been paid to this group and most Indo-Pacific studies focus mainly on gastropods (e.g., FRANK, 1969; HADFIELD, 1976; AUSTIN et af., 1980; TAYLOR & REID,1984; SCHUHMACHER, 1993) or cover the easily accessible intertidal areas (e.g., TAYLOR, 1971; AYAL & SAFRIEL, 1981). Studies dealing with bivalves are generally rare (for a detailed review see MORTON, 1983) or restricted to particular taxa (e.g., LEE & MORTON, 1985; ARNOUD & THOMASSIN, 1990) and to date only few studies have been published in which the total mollusc fauna of various hard substrates was treated (e.g., MASTALLER, 1978; SHEPPARD, 1984). Within the framework of an integrated project in the northern Red Sea, the study of corals represents one particular topic (KLEEMANN, 1992; RIEGL & PILLER, 1997). Several coral associations (including both hard and soft corals) were dis- U. S. Copyright Clearance Center Code Statement: 01 73-9565/97/1804-036l$14.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

P.S.Z.N.: Marine Ecology, 18 (4): 361-378 (1997) 0 1997 Blackwell Wissenschafts-Verlag, Berlin ISSN 0173-9565
Accepted: June 20, 1997
Molluscan Hard-Substrate Associations in the Northern Red Sea M. ZUSCHIN & W. E. PILLER
Institut filr PalSiontologie, Universitat Wien, Geozentrum, Althanstrasse 14, A-1090 Vienna. Austria.
With 3 figures and 3 tables
Key words: Northern Bay of Safaga, reef flats, tidal flats, scleractinian carpets, Sarcophyion carpets.
Abstract. The mollusc associations of 29 hard-substrate localities in the Northern Bay of Safaga (Red Sea, Egypt) were studied. The hard substrates were classified into 7 types: reef flats, intertidal rock bottom, very shallow subtidal conglomerate, Porifes carpet, faviid carpet, platy scleractinian associ- ation, Sarcophyron carpet. Based on a cluster analysis the molluscs of these hard substrates can be classified into 5 groups: Group A is dominated by Dendropoma maxima, Chamoidea and Tridacna maxima, group B by Pedum spondyloideum, T. maxima and Coralliophila neriioidea, group C by Chamoidea and Ceriihium ruppelli, group D by Modiolus auriculaius, Chamoidea, Morula yranulara and Neriia albicilla and group E by Chamoidea and Pirar hebraea. The latter group contains only samples which are generally poor in molluscs. Group A molluscs are diagnostic for reef flats, group B for Purites and faviid carpets, group C for Sarcophyton carpets and group D for tidal/shallow subtidal rock substrates. The different hard substrates are therefore characterized by diagnostic mollusc associ- ations and some of them could also be identified in the fossil record.
Problem
Ecological and paleoecological studies in tropical-subtropical reefoidal environ- ments typically focus on corals and coral communities. Other than this invertebrate group, molluscs are also frequently found inhabiting a variety of ecological niches, and like corals, have considerable fossilization potential. In this context, however, only minor attention has been paid to this group and most Indo-Pacific studies focus mainly on gastropods (e.g., FRANK, 1969; HADFIELD, 1976; AUSTIN et af., 1980; TAYLOR & REID, 1984; SCHUHMACHER, 1993) or cover the easily accessible intertidal areas (e.g., TAYLOR, 1971; AYAL & SAFRIEL, 1981). Studies dealing with bivalves are generally rare (for a detailed review see MORTON, 1983) or restricted to particular taxa (e .g . , LEE & MORTON, 1985; ARNOUD & THOMASSIN, 1990) and to date only few studies have been published in which the total mollusc fauna of various hard substrates was treated (e.g., MASTALLER, 1978; SHEPPARD, 1984).
Within the framework of an integrated project in the northern Red Sea, the study of corals represents one particular topic (KLEEMANN, 1992; RIEGL & PILLER, 1997). Several coral associations (including both hard and soft corals) were dis-
U. S. Copyright Clearance Center Code Statement: 01 73-9565/97/1804-036l$14.00/0

362 ZUSCHIN & PILLER
tinguished, producing a complex spatial distribution depending on several environ- mental factors such as water energy, light, nutrients and sediment suspension (RIEGL & PILLER, 1997). Parallel to investigating coral associations and mapping their spatial distribution, we also conducted a quantitative study of the molluscan associations both in coral-dominated and a range of other different hard-substrate types.
The main goals of this investigation were (1) to recognize mollusc associations, (2) to compare the associations with hard-substrate types to detect the degree of coincidence and (3) to detail parameters allowing the recognition of these hard- substrate types in the fossil record as well.
Material and Methods
1. Study area
Within the integrated project in the Northern Safaga Bay (Red Sea, Egypt; Fig. I ) (PILLER & F’ERVESLER, 1989), sediments (PILLER & MANSOUR, 1990; PILLER, 1994) as well as organism groups with considerable fossilization potential were studied. These include foraminifera (HAUNOLD el a/., 1997). corals (RIEGL e/ a/., 1996; RIEGL & PILLER, 1997) and coral-boring bivalves (KLEEMANN, 1992), molluscs (ZUSCHIN & PILLER, 1997 a, b), coralline algae (PILLER & RASER, 1996; RASSER & PILLER, in press) and echinoids (NEBELSICK, 1992a, b).
The Northern Safaga Bay represents a shallow-water area with highly structured bottom topography reaching down to more than 50 m. It is separated from the Red Sea proper by a submarine ridge (Fig. I) . This bottom topography produces a complex pattern of facies and habitats. Due to terrigenous influx the sediments reflect a mixed carbonate/siliciclastic system (PILLER & MANSOUR. 1992). The annual water temperature ranges between 21 and 2 9 ° C salinity between 40 and 4 6 L , both without any obvious depth gradient due to complete water mixing. Water energy is relatively weak, but a complex current pattern (PILLER & PERVESLER, 1989) influences facies development, particularly the distribution and development of coral reefs and coral carpets. The tidal range is < I m.
2. Sampling
Using a 0.25 m2 aluminium square frame, different inter- and subtidal hard substrates were studied at 29 sample localities (Fig. 2) with respect to shelled molluscs. At each locality the location of the first frame was selected randomly, with the following frames being positioned in an adjoining line. In mixed hard-substrate/loose-ground and/or soft-bottom habitats, frames were only taken from hard substrate. At least I m2 (up to a maximum of 5.75 m’) was investigated per locality (Table I) . The different numbers of investigated frames are due to highly variable mollusc densities (low densities require a higher number of frames and vice versa) and to shorter dive times in greater water depths. The molluscs were identified either immediately in their environment (most bivalves and sessile gastropods) or were collected (many vagile gastropods) for more detailed investigations. Some taphonomic information was collected: livedead, single valved4ouble valved, and hermit crab occupance of gastropod shells (Table 2). This information was used for characterization of the various hard substrates, but was not considered for the statistical analysis.
Taxon diversity was measured in different ways. The MARGALEF-Index was used to quantify species richness (MAGURRAN, 1988). Heterogeneity within sites was measured in using the SIMPSON Diversity Index in comparison to the SHANNON-WIENER function (MAGURRAN, 1988; KREBS, 1989).
The numerical data were averaged for each sample location and transformed to relative abundances. The average data of all molluscs (live and dead) for each sample locality were analysed by cluster analysis (WARD’S method. squared Euclidian distances) using the SPSS 6.0. I program package. Due to poor states of preservation and/or difficulties in identification, Chamoidea, Spondylidae and Ostreoidea
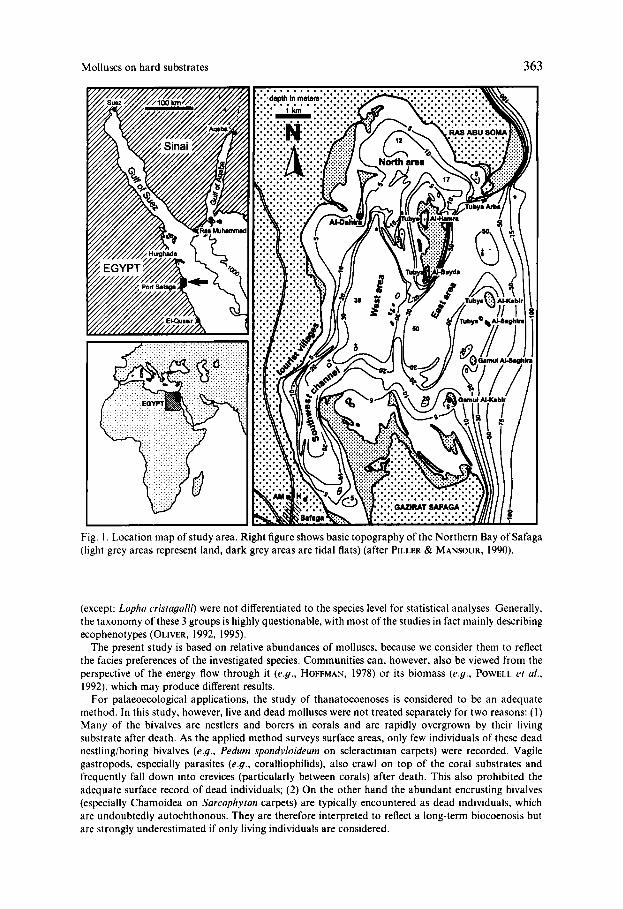
Molluscs on hard substrates 363
Fig. 1. Location map of study area. Right figure shows basic topography of the Northern Bay of Safaga (light grey areas represent land, dark grey areas are tidal flats) (after PILLER & MANSOUR, 1990).
(except: Lopho crisrogolli) were not differentiated to the species level for statistical analyses. Generally, the taxonomy of these 3 groups is highly questionable, with most of the studies in fact mainly describing ecophenotypes (OLIVER, 1992, 1995).
The present study is based on relative abundances of molluscs, because we consider them to reflect the facies preferences of the investigated species. Communities can, however, also be viewed from the perspective of the energy flow through it (e .g . , HOFFMAN, 1978) or its biomass (e.g., POWELL PI a/.. 1992), which may produce different results.
For palaeoecological applications, the study of thanatocoenoses is considered to be an adequate method. In this study, however, live and dead molluscs were not treated separately for two reasons: ( I ) Many of the bivalves are nestlers and borers in corals and are rapidly overgrown by their living substrate after death. As the applied method surveys surface areas, only few individuals of these dead nestling/boring bivalves (e.g.. Pedum spondy/oideum on scleractinian carpets) were recorded. Vagile gastropods, especially parasites (e .g . , coralliophilids), also crawl on top of the coral substrates and frequently fall down into crevices (particularly between corals) after death. This also prohibited the adequate surface record of dead individuals; (2) On the other hand the abundant encrusting bivalves (especially Chamoidea on Sorcophyron carpets) are typically encountered as dead individuals, which are undoubtedly autochthonous. They are therefore interpreted to reflect a long-term biocoenosis but are strongly underestimated if only living individuals are considered.

364 ZUSCHIN & PILLER
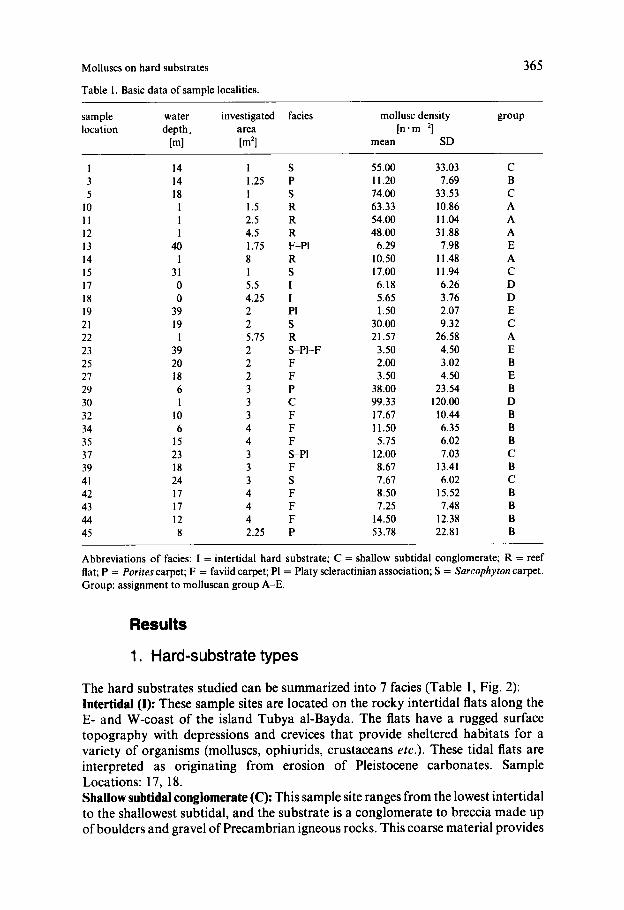
Fig. 2. Sample locations (in correspondence with Tables 1 and 2) and basic bottom topography (light grey areas represent land, dark grey areas are tidal flats).
Therefore a combination of live and dead molluscs is considered to reflect a potential fossil assemblage better than living or dead molluscs alone.
Due to data collection directly in the field, predominantly under water, micromolluscs (smallest sampled individual has a size of 4 mm) are probably underestimated. The quantitative treatment, however, was not confined to a minimum mollusc size, because cluster analyses including and excluding the rare micromolluscs produced the same results.
Most boring bivalves (e.g., gastrochaenids, lithophagins) have been excluded from the quantitative collection due to the difficulty of clearly identifying them from the surficial appearance of their bore holes. A qualitative study on boring bivalves was already presented by KLEEMANN (1992).
In contrast to a widespread, genetically defined use of the term ‘hardground’ as ‘synsedimentary lithified sea floor’ (e.g., PALMER, 1978, 1982; GOLDRING. 1995). we prefer the purely descriptive term ‘hard substrate’. Some of the studied substrates are definitely hardgrounds, for some a synsedimentary lithification cannot be demonstrated and some, for example scleractinian substrates. don’t match the definition.
Molluscs on hard substrates
Table I . Basic data of sample localities.
365
sample water investigated facies mollusc density group location depth. area [n*m-’]
[ml b 2 1 mean SD
I 3 5
10 I 1 12 13 14 I5 17 18 19 21 22 23 25 27 29 30 32 34 35 37 39 41 42 43 44 45
14 14 18
1 1 1
40 1
31 0 0
39 19
1 39 20 18 6 I
10 6
15 23 18 24 17 17 12 8
1 S 1.25 P 1 S 1.5 R 2.5 R 4.5 R 1.75 F-PI 8 R 1 S 5.5 I 4.25 I 2 PI 2 S 5.75 R 2 S-PI-F 2 F 2 F 3 P 3 C 3 F 4 F 4 F 3 s-PI 3 F 3 S 4 F 4 F 4 F 2.25 P
55.00 11.20 74.00 63.33 54.00 48.00
6.29 10.50 17.00 6.18 5.65 1.50
30.00 21.57 3.50 2.00 3.50
38.00 99.33 17.67 11.50 5.75
12.00 8.67 7.67 8.50 7.25
14.50 53.78
33.03 7.69
33.53 10.86 11.04 31.88 7.98
11.48 11.94 6.26 3.76 2.07 9.32
26.58 4.50 3.02 4.50
23.54 120.00
10.44 6.35 6.02 7.03
13.41 6.02
15.52 7.48
12.38 22.81
C B C A A A E A C D D E C A E B E B D B B B C B C B B B B
Abbreviations of facies: I = intertidal hard substrate; C = shallow subtidal conglomerate; R = reef flat; P = Porites carpet; F = faviid carpet; PI = Platy scleractinian association; S = Sarcophyron carpet. Group: assignment to molluscan group A-E.
Results
1. Hard-substrate types
The hard substrates studied can be summarized into 7 facies (Table 1 , Fig. 2): Intertidal (I): These sample sites are located on the rocky intertidal flats along the E- and W-coast of the island Tubya al-Bayda. The flats have a rugged surface topography with depressions and crevices that provide sheltered habitats for a variety of organisms (molluscs, ophiurids, crustaceans etc.). These tidal flats are interpreted as originating from erosion of Pleistocene carbonates. Sample Locations: 17, 18. Shallow subtidal conglomerate (C): This sample site ranges from the lowest intertidal to the shallowest subtidal, and the substrate is a conglomerate to breccia made up of boulders and gravel of Precambrian igneous rocks. This coarse material provides
W
m m
Tab
le 2
. Sum
mar
ized
dat
a co
rres
pond
ing
to th
e 5
grou
ps s
how
n in
Fig
. 3.
sam
ple
site
faci
es
wat
er d
epth
[m]
inve
stig
ated
are
a [m
'] in
divi
dual
s. m
-'
num
ber
of ta
xa
gast
ropo
ds
biva
lves
mol
lusc
s nu
mbe
r %
gas
trop
ods
YO b
ival
ves
YO liv
ing
YO en
crus
ting
dive
rsity
indi
ces
SIM
PSO
N
MA
RC
ALE
F In
form
atio
n
gast
ropo
ds
num
ber
% li
ving
YO
encr
ustin
g di
vers
ity in
dice
s SI
MPS
ON
M
AR
GA
LEF
Info
rmat
ion
dom
inat
ing
taxa
Gro
up A
10, 1
1, 1
2, 1
4, 2
2
R 1-2 22.2
5 29
.39
15 8 1
654 92.9
7 7.
03
95.8
7 92
.51
0.20
2.
16
0.58
608 98
.68
95.8
9
0.08
1.
09
0.24
D.
mas
ima
(95.
89 Y
O)
Gro
up B
G
roup
C
3.25
, 29
. 32,
34,
35.
39
.42.
43.4
4.45
I.
5, 1
5,21
, 37,
41
F, P
62
0
34.5
15
.15
32
13
19
523 21.9
9 78
.01
77.8
2 16
.83
0.83
4.
95
2.16
1 I5 93.9
1 0.
87
0.36
2.
53
0.93
C. ne
riro
idea
(80
YO)
s, s-
PI
14-3
1 11
23
.90
48
26
22
263 19
.39
80.6
1 25
.86
66.5
4
0.61
8.
97
I .97
51
33.3
3 1.
96
0.88
6.
36
2.12
C
. rup
pelli
(33
.33
YO
)
Gro
up D
17, 1
8. 3
0
1, K
&
2 12.7
5 27
.92
25
19* 6
356 31
.74
68.2
6 72
.75
31.7
4
0.76
4.
09
I .84
1 l3*
91
.15
3.54
0.80
3.
81
2.06
M
. gra
nula
ra (
30.9
7 YO
) N
. alh
icill
a (3
0.09
'10)
Gro
up E
13. 1
9. 2
3. 2
7 ~~
F, P
I, F-
PI,
SPI-
F
18
40
7.
75
3.61
21
I1
10
28
39.2
9 60
.71
25.0
0 25
.00
0.96
6.
00
2.85
II
27.2
7 0.
00
1 .oo
4.17
2.
40
none
N
2 3 a
L r w
3
0,
E H B sr
Tabl
e 2.
(co
ntin
ued)
5 8 - 1
Gro
up A
G
roup
B
Gro
up C
G
roup
D
10, 1
1, 1
2, 1
4,22
39
,42,
43,4
4,45
1,
5, 1
5,21
, 37,
41
17, 1
8, 3
0 3,
25,2
9, 3
2, 3
4, 3
5,
sam
ple s
ite
Gro
up E
13, 1
9,23
,27
biva
lves
nu
mbe
r liv
ing
% e
ncru
stin
g di
vers
ity in
dice
s SI
MPS
ON
MA
RG
ALE
F in
form
atio
n do
min
atin
g ta
xa
46
58.7
0 47
.83
0.75
1.
57
1.51
C
ham
oide
a (3
6.96
%)
T. m
axim
a (3
2.61
%)
408 73.2
8 21
.32
0.77
2.
99
1.83
P
. spo
ndyl
oide
um (3
7.25
%)
T. m
axim
a (2
5.74
W)
212 24
.06
82.0
8
243 64.2
0 44
.86
0.41
0.
52
4.48
0.
91
1.18
0.
82
Cha
moi
dea (
76.8
9 %
) M
. aur
icul
atus
(53.
91 %
) C
ham
oide
a (4
3.62
%)
17
23.5
3 41
.18
0.88
3.
18
2.03
C
ham
oide
a (2
9.41
%)
P. h
ebra
ea (2
3.53
%)
Abb
revi
atio
ns of
faci
es: I
= in
tert
idal
har
d su
bstr
ate;
C =
shal
low
subt
idal
cong
lom
erat
e; R
= re
ef f
lat;
P =
Por
ites c
arpe
t; F
= fa
viid
car
pet;
PI =
Pla
ty sc
lera
ctin
ian
asso
ciat
ion;
S =
Sar
coph
yion
car
pet.
Ast
eris
k in
gro
up D
= 1
indi
vidu
al o
f C
hito
n sp
. is i
nclu
ded
into
gas
trop
ods.

368 ZUSCHIN & PILLER
an uneven surface with many protected habitats. It is restricted to the northwestern- most tip of Gazirat Safaga. Sample Location: 30. Reef Flats (R): These environments are dominated by bare rock bottom with isolated scleractinian coral colonies, e.g., Stylophora pistillata, Pocillopora damicor- nis. A distinct zonation of the corals exists due to decreasing water energy from windward to leeward sides. The surface of the rock bottom is generally wavy but relatively smooth due to coverage by soft algae and coralline algal crusts. The reef flat can be exposed during low tides. The studied sites are located at the four patch reefs (Tubya al-Kabir, Tubya al-Saghira, Gamul al-Saghira, Gamul al-Kabir) which represent the boundary between Northern Safaga Bay and the open Red Sea. Sample Locations: 10, 11, 12, 14, 22. Porites carpet (P): This hard substrate is characterized by a dense coverage of scleractinians which are highly dominated by Porites. This coral coverage produces a carpet up to several metres thick with deep caverns and depressions between the stick-like Porites colonies. These caverns provide habitats for various other organisms, although at a first glance this facies is completely dominated by Porites. Due to these small habitats, a high coral diversity is developed. Sample Locations: 3, 29,45. Faviid carpet (F): Similar to the Porites carpet, scleractinians form a dense coverage of the sea bottom. The corals, however, are dominated by faviid species that also show a relatively high diversity. Besides this difference in species composition, the general appearance of the faviid carpet differs from the Porites carpet: the surface topography is very rugged here, with highly variable heights of individual colonies producing a distinct relief. Sample Locations: 25, 27, 32, 34, 35, 39,42,43,44. Platy scleractinian association (PI): This hard-substrate type is characterized by scleractinian corals with platy growth forms; one of the most typical species is Lepfoseris papyruceu. This growth form is considered to be a low-light adaptation ( RIECL & PILLER, 1997) and, consequently, this facies typically occurs below 30 m depth. It is best developed on steeper slopes, where coral coverage is highest, whereas on flat topographies the frequency of live corals is lower and rock bottom is more abundant. The studied areas are located exclusively in the ‘East area’ and mainly correspond to flat areas, where more coral-free substrate is available for mollusc settlement. Sample Locations: 13, 19, 23. Surcophyton carpet (S): Areas which are predominantly settled by soft corals, dominated by Surcophyron, are summarized in this facies. Scleractinians are also present, although their occurrence is scattered. Bare rock bottom is widespread between the soft corals, but is shaded by the ‘mushroom-like’ growth form of Surcophyton. Most of these areas occur on slopes, and the rock bottom frequently shows depressions filled with loose sediments. These sand- and gravelfilled areas represent on the one hand accumulations of sediments and on the other a different bottom type, a ‘looseground’ (GOLDRING, 1995). These sediment accumulations contain shells of epilithic organisms originally living on the neighbouring rock bottom and that were transported in by currents. The loose-ground accumulations are inhabited by epi- and even endobenthic organisms, reflecting a different habitat. The typical occurrences of this hard-substrate type are current-exposed submarine ridges, such as between Tubya al-Bayda and Gazirat Safaga, and along current- swept slopes such as on the western margin of the basin in the “Western area”. Sample Locations: I , 5, 15, 21, 3’/, 41.
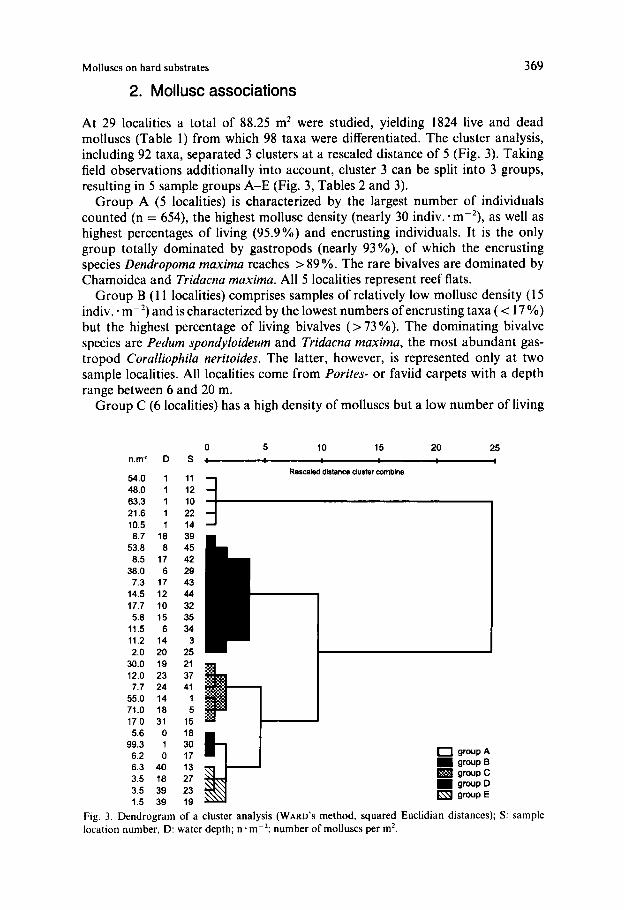
Molluscs on hard substrates 369
2. Mollusc associations
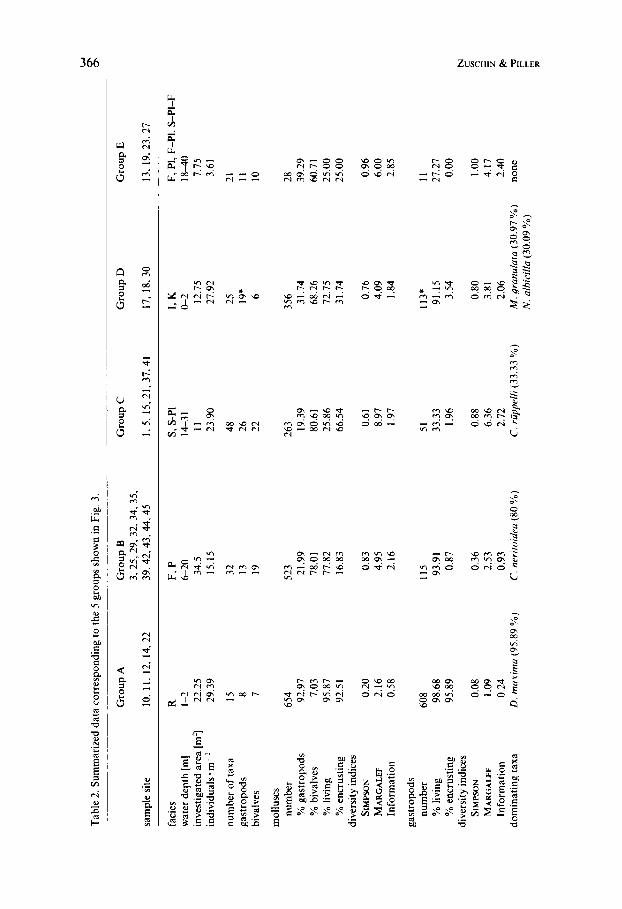
At 29 localities a total of 88.25 m2 were studied, yielding 1824 live and dead molluscs (Table 1) from which 98 taxa were differentiated. The cluster analysis, including 92 taxa, separated 3 clusters at a rescaled distance of 5 (Fig. 3). Taking field observations additionally into account, cluster 3 can be split into 3 groups, resulting in 5 sample groups A-E (Fig. 3, Tables 2 and 3).
Group A (5 localities) is characterized by the largest number of individuals counted (n = 654), the highest mollusc density (nearly 30 indiv.-m-2), as well as highest percentages of living (95.9%) and encrusting individuals. It is the only group totally dominated by gastropods (nearly 93 YO), of which the encrusting species Dendropoma maxima reaches > 89 YO. The rare bivalves are dominated by Chamoidea and Tridacna maxima. All 5 localities represent reef flats.
Group B (1 1 localities) comprises samples of relatively low mollusc density (1 5 indiv. * m-2) and is characterized by the lowest numbers of encrusting taxa ( -= I7 YO) but the highest percentage of living bivalves ( > 73 Oh). The dominating bivalve species are Pedum spondyloideum and Tridacna maxima, the most abundant gas- tropod Coralliophila neritoides. The latter, however, is represented only at two sample localities. All localities come from Porites- or faviid carpets with a depth range between 6 and 20 m.
Group C (6 localities) has a high density of molluscs but a low number of living
0 5 10 15 20 25 n.m' D S I I
54.0 1 11 48.0 1 12 63.3 1 10 21.6 1 22 10.5 1 14 8.7 18 39
53.8 0 45 8.5 17 42
38.0 6 29 7.3 17 43
14.5 12 44 17.7 10 32 5.8 15 35
11.5 6 34 11.2 14 3 2.0 20 25
30.0 19 21 za
Resealed dlslanea cluster combine w
12.0 23 37 7.7 24 41
55.0 14 1 71.0 18 5 17.0 31 15
9;:: : 3 )I I 6.2 0 17 6.3 40 13 3.5 10 27 3.5 39 23 1.5 39 19
Fig. 3. Dendrogram of a cluster analysis (WARD'S method, squared Euclidian distances); S: sample location number, D: water depth; n * rn-? number of molluscs per m2.
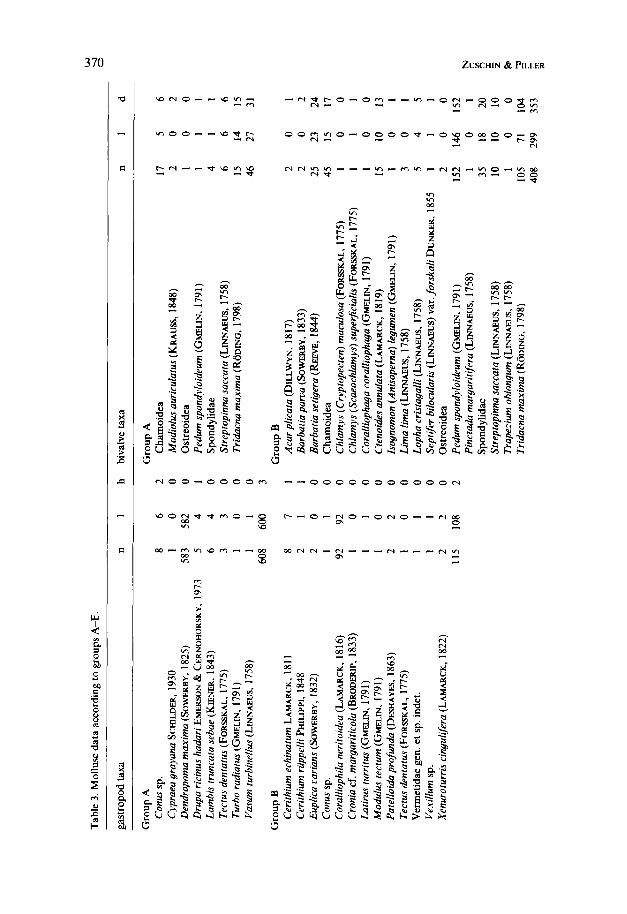
w
Tab
le 3
. Mol
lusc
dat
a ac
cord
ing
to g
roup
s A-E
.
gast
ropo
d ta
xa
n1
Gro
up A
C
onus
sp.
Cyp
raea
gra
yanu
SC
HIL
DER
, 19
30
Den
drop
oma maxima (S
OW
ERBY
, 1825
) D
rupa
ric
inus
had
ari E
~IER
SON
&
CER
NO
HO
RSK
Y,
Lum
bis
trun
cata
seb
ae (K
IEN
ER
, 1843
) Te
ctus
den
tatu
s (F
OR
SSK
AL
, 17
75)
Turb
o ra
diat
us (G
MEL
IN, 17
91)
Vm
um fu
rbin
ellu
s (LI
NN
AEU
S,
1758
)
Gro
up B
C
erith
ium
ech
inar
um L
AM
AR
CK
, 18 1 1
C
erith
ium
rup
pelli
PH
ILIP
PI,
1848
Eu
pric
a co
rian
s (SO
WER
BY, 18
32)
Con
us sp
. C
oral
lioph
ila ne
rito
idea
(LA
MA
RC
K. 18 16
) C
roni
a cf
. mar
gari
ticol
a (B
RO
DER
IP, 18
33)
Lutir
us tu
rritu
s (G
ME
LIN
, 1791
) M
odul
us te
etur
n (G
ME
LM
, 1791
) Pa
tello
ida
prof
unda
(DES
HAY
ES, 18
63)
Tert
us d
enta
tus
(FO
RSS
KA
L,
1775
) V
erm
etid
ae g
en. e
t sp.
inde
t. Ve
xillu
m sp
. Xe
nuro
turr
is c
ingu
lifer
a (L
AM
AR
CK
, 1822
)
86
1
0
583
582
1973
5
4 6
4
33
1
0
1 1
608
600
87
2
1
20
1
1
92
92
10
1
1 I
0
22
1
0
1 1
1 1
22
11
5 10
8
1 1 0 0 0 0
0 0 0 0 0 0 0 2
4 0
biva
lve
taxa
n
ld
Gro
up A
C
harn
oide
a M
odio
lus a
uric
ular
us (K
RA
USS
, 1848
) O
stre
oide
a Pe
dum
spon
dvlo
ideu
m (G
KE
LIN
, 179 1
) Sp
ondy
lidae
St
rept
opin
na s
acca
fa (L
INN
AEU
S, 17
58)
Trid
acna
max
ima
(R~D
ING
. 1798
)
Gro
up B
Ac
ar p
licat
a (D
ILLW
YN
, 1817
) Ba
rbat
ia p
ama
(SO
WER
BY. 18
33)
Barb
atia
set
iger
a (REEVE, 18
44)
Cha
rnoi
dea
Chl
amys
(Cry
ptop
ecte
n) m
acul
osa
(FO
RSS
KA
L,
1775
) C
hlam
ys (S
caeo
chla
mys
) sup
erfic
ialis
(FO
RSS
KA
L,
1775
) C
oral
lioph
aga
cora
lliop
haga
(GM
EL
IN, 1791
) C
teno
ides
unn
ufat
a (LA
MA
RC
K, 18
19)
Isog
nom
on (
Anis
oper
nu) l
egm
en (G
MEL
IN, 17
91)
Lim
a lim
a (L
INN
AFX
JS,
1758
) b
ph
a cr
ista
galli
(LIN
NA
EUS,
1758
) Se
ptife
r bi
locu
lari
s (LI
NN
AEU
S)
var.
fors
kali
DU
NK
ER
, 18
55
Ost
reoi
dea
Pedu
m sp
ondy
loid
eum
(GK
ELIN
, 1791
) Pi
ncta
da m
arga
ritif
era
(LIN
NA
EUS,
1758
) Sp
ondy
lidae
St
rept
opin
na s
acca
ta (L
INN
AEU
S, 17
58)
Trap
eziu
m ob
long
wn
(LIN
NA
EUS,
1758
) Tr
idac
na m
axim
a (R
OD
ING
, 1798
)
17
5 6
20
2
10
0
1 1
1 4
1 1
66
6
15
14
15
46
27
31
2 2 25
45 1 1 1 15 1 3 5 1 2
152 1 35
10 1
105
408
0 0 23
15 0 1 0
10 0 0 4 1 0
146 0 18
10
0 71
299
I 2 24
17 0 1 0 13 1 1 5 1 0
152 10
4 O
R
104
2 35
3
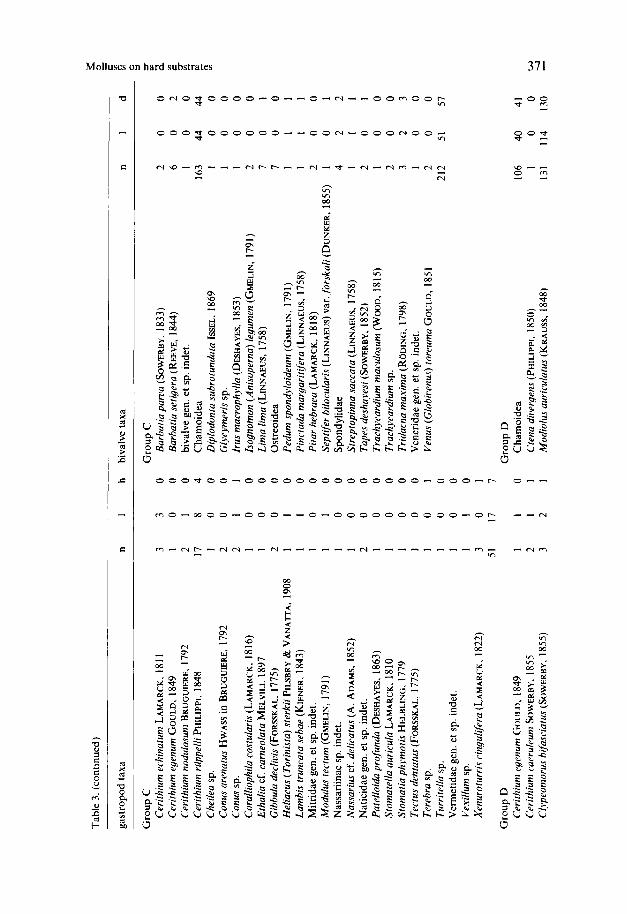
Tabl
e 3.
(con
tinue
d)
3
gast
ropo
d ta
xa
Gro
up C
C
eriih
ium
echi
natu
m L
AM
AR
CK
, 18
1 1
Cer
ithiu
m eg
enum
CO
UL
D.
1849
C
erith
iuni
nod
ulos
um B
RU
FUIE
RE,
17
92
Cer
ithiu
m ri
ippe
lli P
HIL
IPPI
. 18
48
Che
ilea
sp.
Con
us a
rena
tus
HW
ASS
in B
RU
GU
IER
E,
1792
C
onus
sp.
Cor
ullio
phila
cost
ular
is (L
AM
AR
CK
. 18
16)
Etha
lia c
f. ca
rneo
lata
ME
LV
ILL
18
97
Gib
bula
dec
licis
(FO
RSS
KA
L, 1775
) H
eliu
cus
( Tor
inis
ta) s
terk
ii PI
LSBR
Y
& V
AN
ATT
A. 19
08
Lam
his
trun
cata
seh
ae (K
IEN
ER
. 1843
) M
itrid
ae g
en. e
t sp.
inde
t. M
odul
us te
ctum
(GM
EL
IN, 17
91)
Nas
sari
inae
sp. i
ndet
. N
assa
rius
cf.
delic
atus
(A. A
DA
MS.
1852
) N
atic
idae
gen
. et s
p. in
det.
Pute
lloid
a pr
ofun
da (D
ESH
AY
ES.
18
63)
Stom
atel
la a
uric
ula
LA
MA
RC
K.
1810
St
omui
ia p
hym
otis
HE
LB
LIN
G.
1779
Te
cius
dentatus (F
OR
SSK
AL.
17
75)
Tere
bra
sp.
Turr
iiella
sp.
V
erm
etid
ae g
en. e
t sp.
inde
t. Ve
\-illu
m s
p.
Xenu
rotu
rris
cin
gulif
era
( LA
MA
RC
K.
1822
)
Gro
up D
C
erith
iuni
ege
num
CO
UL
D, 1849
C
erith
iuni
cae
rule
um S
OW
ERBY
. 1855
C
lype
onio
rus b
rfas
ciat
us (S
OW
ERBY
, 1855
)
nl
h
33
0
10
0
21
0
17
8 4
10
0
20
0
21
1
10
0
10
0
20
0
1 1
0
1 1
0
10
0
1 1
0
10
0
10
0
20
0
10
0
10
0
10
0
10
0
10
1
10
0
10
0
1 1
0
30
1
51
17
7
1 1
0
21
1 3
21
yl
biva
lve t
axa
n I
dg
0
Gro
up C
s
Barb
atia
par
t)a (
SOW
ER
BY
. 18
33)
20
0;
:
Barb
atia
set
iger
a (R
EEV
E, 18
44)
60
22
bi
valv
e gen
. et s
p. in
det.
Cha
moi
dea
163
44
44
Dip
lodo
nta
subr
oiun
dara
ISSE
L, I8
69
10
09
G
lycy
mer
is sp
. 1
00
Ir
us m
acro
ph.y
lla (
DE
SHA
YE
S,
1853
) 1
00
Is
ogno
mon
(An
isop
erna
) legu
men
(GM
ELIN
, 1791
) 2
00
Li
ma
lima
(LIN
NA
EU
S,
1758
) 7
01
O
stre
oide
a 7
00
Pe
dum
spon
dylo
ideu
m (
GM
EL
~,
1791
) 1
1 1
Pinc
tada
mar
gari
tifer
a (L
INN
AEU
S,
1758
) 1
1 1
Pita
r he
brae
a ( L
AM
AR
CK
, 18 18
)
Spon
d ylid
ae
42
2
Stre
ptop
inna
sacc
ata
(LIN
NA
EUS,
17
58)
1 I
1 Ta
pes d
esha
yesi
(SO
WE
RB
Y,
1852
) 2
01
Tr
achy
card
ium
mac
ulos
um (
WO
OD
, 181 5
) 1
00
Tr
achy
card
ium
sp.
20
0
Trid
ucna
max
ima
( RO
DIN
G, 17
98)
32
3
Ven
erid
ae g
en. e
t sp.
inde
t. 1
00
2
00
21
2 51
57
10
0%
20
0
Sept
ifer
bilo
cula
ris (
LIN
NA
EUS)
va
r. fo
rsku
li (D
UN
KE
R,
1855
) I
0 1
Venu
s (G
lobi
renu
s) io
reum
a C
OU
LD
, 1851
Gro
up D
C
ham
oide
a C
tena
div
erge
ns (P
HIL
IPPI
. 18
50)
Mod
iolu
s aur
icul
atus
(KR
AU
SS. 18
48)
106
40
41
1 o
ow
13
1 11
4 13
0 2
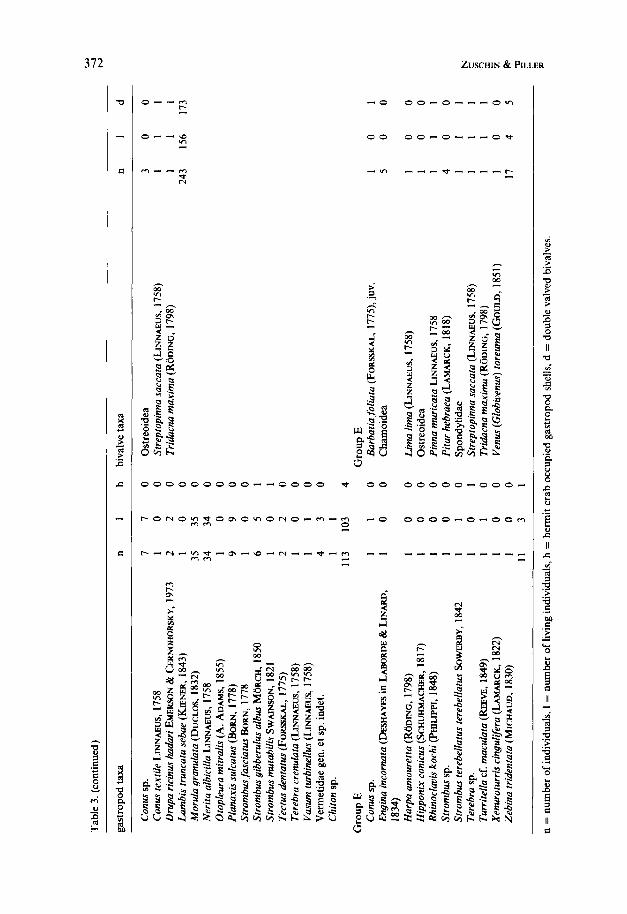
W 4
h)
Tab
le 3
. (co
ntin
ued)
gast
ropo
d ta
xa
n I
h bi
valv
e ta
xa
nl
d
Conu
s sp
. 7
70
C
onus
text
ile L
INN
AEU
S, 17
58
10
0
Dru
pa r
icin
us h
aahr
i EM
ERSO
N & C
ERN
OH
OR
SKY
, 19
73
2 2
0 La
mbi
s tr
unca
ta se
bae
(KEN
EX, 1
843)
1
00
M
orul
a gr
anul
ata
(Du
cm, 1
832)
35
35
0
Ner
ita a
lbic
illa
LINN
AEUS
, 1758
34
34
0
Oto
pleu
ra m
itrd
is (
A. A
DA
MS,
18
55)
10
0
Plan
axis
sul
caru
s (BORN, 17
78)
99
0
Siro
mbu
s farciatus B
ORN
, 177
8 1
00
St
rom
bus g
ibbe
rulu
s alb
us M
BRC
H, 1
850
65
1
Stro
mbu
s mut
abili
s SW
NN
SON
, 1821
1
01
Te
ctus
dent
atus
(FOR
SSKA
L, 17
75)
22
0
Tere
bra
cren
ulat
a (L
INN
AEU
S, 17
58)
10
0
Vmum
turb
inel
lus (
LIN
NA
EUS,
1758
) 1
10
V
erm
etid
ae g
en. e
t sp.
inde
t. C
hito
n sp
. 1
1
Con
us sp
. 1
10
En
gina
inco
rnat
a (D
EWA
YES
in L
ABO
RDE &
LIN
ARD,
1
00
18
34)
Har
pa m
oure
rta
(R~D
ING
, 1798
) 1
00
H
ippo
nix
coni
cus (SCHUHMACHER, 18
1 7)
Rhin
ocla
uis k
ochi
(PH
ILIP
PI,
1848
) 1
00
St
rom
bus s
p.
10
0
Stro
mbu
s te
rebe
llatu
s re
rebe
llatu
s SO
WER
BY, 18
42
1 1
0
Tere
bra
sp.
10
1
Turr
itella
cf.
mac
ulat
a (REEVE, 18
49)
11
0
Xenu
rotu
rris
cin
gulif
era
(LA
MA
RC
K, 18
22)
10
0
Zebi
na tr
iden
tata
(MIC
HA
UD
, 1830
) 1
00
11
3
1
43
0
113
103
4 G
roup
E
10
0 Ost
reoi
dea
Stre
ptop
inna
sar
cata
(LIN
NA
FSJS
, 1758
) Tr
idac
na m
axim
a (R
BDIN
G, 1
798)
Gro
up E
Barb
atia
folia
ta (F
OR
SSK
AL
, 17
79, j
uv.
Cha
moi
dea
Lim
a Iima (L
INN
AEU
S, 17
58)
Ost
reoi
dea
Pinn
a m
uric
ata
LIN
NA
EUS,
1758
Pi
tar
hebr
aea
(LA
MA
RC
K, 18 18
) Sp
ondy
tidae
St
rept
opin
na s
arca
ta (
LIN
NA
EUS,
1758
) Tr
idac
na m
axim
a (R
~DIN
G, 1798
) Ve
nus (
Glo
biue
nus)
tore
uma
(CO
ULD
, 1851
)
~ ~~
n =
num
ber o
f ind
ivid
uals,
1 =
num
ber o
f liv
ing
indi
vidu
als,
h =
her
mit
crab
occ
upie
d ga
stro
pod
shel
ls, d
= d
oubl
e val
ved
biva
lves
.
30
0
1 1
1 1
1 1
243
156
173
10
1
50
0
10
0
10
0
1 1
1 4
00
I
I I
N
1 1
1 2
1 1
1.
1
00
5.
r B

Molluscs on hard substrates 373
individuals (25.9 YO); this group has the highest diversity of both gastropods and bivalves. The number of living bivalves is low whereas the number of encrusting bivalves is by far the highest. The bivalves are totally dominated by Chamoidea, gastropods by Cerithium, in particular C. ruppelli. All localities represent Sar- cophyton carpets, one (LOC. 37) is transitional from a Sarcophyton carpet to a platy scleractinian association. Water depth ranges from 1 4 3 1 m.
Group D (3 localities) has a high mollusc density (nearly 28 indiv..m-2), yet with a high variance. Most general characters of this sample group are intermediate (live individuals, encrusting molluscs, gastropod/bivalve ratio); at the species level, however, an excellent discrimination is possible. Most characteristic are Modiolus auriculatus, Chamoidea, Morula granulata and Nerita albicilla. Two of the sample sites are located on intertidal flats and the third comes from the conglomerate and ranges from the intertidal to very shallow subtidal.
Group E (4 localities) comprises sample localities with lowest density (3.6 indiv. * m-2) and percentage of living molluscs (25 %). No single gastropod taxon dominates and bivalves are characterized by Chamoidea and Pitar hebraea. The samples of this group have heterogeneous origins: one stems from a faviid carpet; one from a platy scleractinian association; one from a transition between both; and one from a transition between faviid carpet-platy scleractinian-Sarcophyton carpet. The water depth also ranges widely from 18-40 m.
Discussion
1. Associations
The 5 distinguished sample groups are clearly differentiated by several parameters, and 4 of them correspond very well with a specific hard-substrate type.
Most characteristic for group A is the absolute dominance of the vermetid Dendropoma maxima (SIMPSON Index: 0.08; Information Index: 0.24) and the group’s total restriction to reef flats (Table 2). This can be easily explained because the vermetid requires agitated water conditions in order to spread its mucous net for feeding (HUGHES & LEWIS, 1974). A study of molluscs on hardgrounds in the Sudanese part of the Red Sea (TAYLOR & REID, 1984) showed similar distributions and densities of Dendropoma maxima. Such very high densities of these sessile gastropods, however, are a phenomenon largely restricted to the Red Sea, with similar abundances only being reported from the SE Pacific (TAYLOR & REID, 1984). The extremely high ratio of living Dendropoma can be explained by a rapid overgrowth by other organisms (e.g., coralline algae) after death. Generally, shells of dead vagile molluscs are washed away in this high energy environment; only larger shells (e.g., Lambis, Conus) remain.
For group B the co-dominance of Pedum spondyloideum (37.25 %) and Tridacna maxima (25.7 %) is most discriminating, whereas the very abundant occurrence of the parasitic gastropod Coralliophila (80%) is of limited importance as these gastropods were found at only two sample localities (Porites carpet). A differ- entiation between faviid and Porites carpet was not possible using relative abun- dances of molluscs, except in the case of Coralliophila violacea: the latter is restricted to Porites and Porites (Synarea) (SCHUHMACHER, 1993). Its very patchy distri-

3 74 ZUSCHIN & PILLER
bution, however, weakens its value for quantitative analyses. A similar patchy distribution was also reported from other areas of the Red Sea (TAYLOR & REID, 1984; SCHUHMACHER, 1993). The occurrence of this parasite and of coral-boring Pedum spondyloideum, however, clearly reflects the presence of specific host corals. This was pointed out for Pedum by KLEEMANN (1990). The high ratio of living Pedum is due to rapid overgrowth by the host coral after death. Nearly all vagile gastropods are alive (ca. 94%); dead individuals are interpreted as being trans- ported into depressions and crevices between coral heads and were therefore missed by the applied study method. While these vagile hard-substrate molluscs are trans- ported into adjacent loose grounds, the mollusc associations of the hard substrates are not influenced by dwellers of adjacent and interspersed loose grounds because these are topographically deeper.
Group C combines very high diversities of gastropods and bivalves (MARGALEF Index: 6.36 and 4.48), with dominance of Chamoidea (76.9% of bivalves; SIMPSON Index: 0.41) on the one hand and of Cerithium ruppelli (33.3 %) on the other hand. The high diversity may reflect the Sarcophyton carpet, which provides a variety of habitats due to a mixture of soft and hard corals, a vertical structuring owing to variable colony heights, and a high proportion of rock bottom in between. Additionally, some soft-bottom dwelling bivalves occur, e.g., venerids, Glycymeris, Diplodonta, Trachycardium (Table 3). They are all disarticulated, indicating trans- port from the neighbouring loose grounds. Although water energy is relatively low here, transport is possible due to the low relief of the rock bottom and its slope occurrence. Sarcophyton colonies are, contrary to other soft corals, considered to be long-living forms (KROLL, REINICKE & SCHUHMACHER in SCHUHMACHER et al., 1995), so that this coral carpet type does not merely represent a short-term feature. This is also reflected by the dense settlement of mostly dead (73 %) individuals of Chamoidea (Table 3). The main factor responsible for the development of a Sarcophyton carpet is considered to be the high suspension load in the water column (FABRICIUS et al., 1995; RIEGL & PILLER, 1997). The occurrence of Sar- cophyton carpets on current-exposed submarine elevations or along slopes parallel or obliquely orientated to the main currents which carry suspended material sup- ports this explanation. Such a high suspension load may also provide food for molluscs, explaining their high density. Despite a painstaking effort, no epibenthic molluscs were detected macroscopically on Sarcophyton colonies.
Group D is clearly dominated by the two bivalve taxa Chamoidea (43.6 YO) and Modiolus auriculatus (53.9 YO); the two gastropod species Morulagranulata (30.9 YO) and Ncrita albicilla (30.1 YO), however, are also very abundant and characteristic. The two gastropod species, along with the mass occurrence of Modiolus, place all the samples in extremely shallow water. Mass occurrences of Brachiodontes uari- ahilis (KRAUSS, 1848) and Cerithium caeruleum, although not documented in the studied frames, are also very typical for this facies in the Northern Bay of Safaga (ZUSCHIN & PILLER, 1997a) and additionally reported from various coastal areas of the Pacific Ocean (e.*9., ARNOUD & THOMASSIN, 1990; AYAL & SAFRIEL, 1981; LEE & MORTON, 1985; SAFRIEL ct al., 1980a, b; TAYLOR, 1971; TAYLOR & REID, 1984). More than 90% of vagile gastropods are alive, pointing to destruction and/or transport of dead shells.
Only group E is more or less negatively defined due to low mollusc numbers and no dominance of a single species (SIMPSON Index: 1.00 for gastropods, 0.88 for

Molluscs on hard substrates 315
bivalves). Its samples stem from a range of hard-substrate types, yet predominantly from transitions between faviid carpets and platy scleractinian associations. These substrate types exhibit a high surface coverage by corals and therefore provide only very limited space for molluscs. In addition, the relatively deep water reduces the light availability for zooxanthellate bivalves (Tridacna) which dominate the shallow-water faviid and Porites carpets. The lack of Coralliophila is related to the low abundance of Porites, which is considered to be the preferred substrate for this gastropod (TAYLOR & REID, 1984; SCHUHMACHER, 1993).
Initially, a differentiation of hard-substrate types based on molluscs seems prob- lematic, because of the occurrence of Chamoidea as the dominating bivalve taxon in 4 out of 5 groups (group E, however, can be excluded due to the generally very low number of molluscs). Closer examination reveals a clearer picture. Reef flats, for example, are primarily defined by Dendropoma, and gastropods generally exceed bivalves by a factor of > 10. Among bivalves, which are generally very low in abundance, Chamoidea co-occur with Tridacna maxima; this separates this assem- blage from all the others. Striking differences also exist between Sarcophyton carpets and rocky intertidal to shallow subtidal substrates. On Sarcophyton carpets, Chamoidea is the only dominating bivalve taxon, which is also reflected by a total mollusc fauna of >80% encrusting individuals. In contrast, on the rocky tidal/shallow subtidal substrates, <45 % of the bivalves are encrusting and the abundant Chamoidea are surpassed by byssally attached mytilids (Modiolus and Brach idon tes) .
Most empty shells of vagile gastropods are potentially useful for occupance by hermit crabs. In the studied material, approx. 30% were inhabited by pagurids (Table 3). This secondary use can lead to transportation of gastropod shells even to distant areas (SHIMOYAMA, 1979), producing a (pa1eo)ecological bias (WALKER, 1989).
2. Paleoecological implications
The preservational potential of most hard substrates may generally be considered as high, although extremely shallow water examples may easily be obliterated by abrasion or by erosion and dissolution due to subaerial exposure. As stated by FURSICH & PALMER (1975), encrusting organisms are often the only preserved evidence for a fossil hard substrate. In contrast to soft-bottom paleocommunities, however, sessile hard-substrate faunas are still in their life-positions (PALMER, 1982).
Based on the hard substrates studied here, the various types probably possess different degrees of recognition in the fossil record: (a) Reef flats may be preserved as hardgrounds with patches of low diversity scleractinians. Due to the composition of a very specific scleractinian association (dominated by Pocillopora, Styllophora etc.) their recognition may be relatively easy. The high numbers of the vermetid Dendropoma will be preserved as a sequence of superimposed tubes encrusted and overgrown by other sessile organisms as, for example, coralline algae. Due to the high numbers of Dendropoma maxima and its encrusting mode of life we consider the importance of these gastropods to be high for in situ carbonate production on reef flats; (b) Intertidal and very shallow submarine hardgrounds are even more

376 ZUSCHIN & PILLER
endangered by dissolution and abrasion, causing low preservation potential (Bou- COT, 1981). Additionally, the usually poor exposure of ancient rocky shores in the geologic record limits the access of paleontologists to preserved biotas (JOHNSON, 1988). Due to the lack of dead molluscs we conclude that most of the intertidal shells will predominantly be exported to the surrounding shallow marine or supratidal environments, which was confirmed by personal observations. The pronounced structures on the surface of the tidal flats may provide enough fissures, crevices and depressions to retain at least part of this community in a parautochthonous position. However, if the intertidal environment becomes preserved with its typical biota it is easily traceable by their typical bivalves (Modiolus and Bruchidontes) and gastropods (Morulu and Neritu); (c) Porites and flaviid carpets have a high fossilization potential, although the state of preservation may be poor as the skeletal aragonite will be replaced by calcitic cement. While such fossil facies are clearly recognizable by their scleractinian associations, their interpretation can be supplemented by molluscs. The boring bivalves (e.g. , Pedum) or their bore holes will be preserved inside the coral colonies, the gastropods (e.g., Coralliophilu) will perhaps occur in pockets between the corals. The latter occurrences require study- ing the surrounding soft bottoms. A differentiation between the two carpet types on the basis of mollusc associations is not possible. Due to the rapid coral growth, carbonate production of molluscs is small compared to that of scleractinians; (d) Surcophyton carpets may express themselves in the fossil record as a more or less pronounced hardground with isolated scleractinian colonies. The hardground may be overlain by shelly sediments which probably, in part, accumulated due to the baffling function of the soft corals. The poor preservation potential of soft corals will leave no distinct hint of this baffling function. Although no macroepibenthic organisms were observed to use Sarcophyton as a substrate, mollusc associations can provide additional information. The high individual numbers and densities of Chamoidea indicate a high organic suspension load and therefore sufficient nutri- ents for suspension feeders. This organic material is also considered to be important for Surcophyton (RIEGL & PILLER, 1997). The many depressions inside the Sur- cophyton carpets favour the accumulation of the upper valves of the encrusting bivalves (pers. obs.) and vagile faunal elements, particularly several Cerithium species. The high mollusc diversity reflects the co-occurring loose-ground and hard-substrate habitats. Depending on the thickness of sediment accumulations, additional elements may include epi- and infaunal loose-ground dwellers which ‘dilute’ the number of hard-substrate molluscs and render the reconstruction of the Surcophyton carpet more difficult. We consider the carbonate production by molluscs on Surcophyton carpets to be relatively high as molluscs occur in high densities and stony corals are relatively rare.
Conclusions
(a) The studied hard-substrate types show distinct mollusc associations, although a further differentiation between Porites and faviid carpet is not possible; (b) Four hard-substrate types are dominated by Chamoidea. The occurrence of typical gastropods (e.g. , Dendropoma on reef flats, Cerithium ruppelli on Sarcophyton carpets) and abundant co-occurring bivalves (e.g. , Modiolus uuriculutus on tidal

Molluscs on hard substrates 377
flats) makes their differentiation possible based on molluscs; (c) Sessile molluscs provide much better opportunities for facies reconstruction than vagile species as the latter are easily transported into adjacent environments. Therefore mainly encrusting molluscs contribute considerably to in situ carbonate production, whereas vagile species are the dominant source of molluscan carbonate export; (d) From the paleoecological perspective, 3 groups of hard substrates (1. reef flat, 2. tidal flat and shallow subtidal conglomerate, 3. coral carpet) can easily be recog- nized if they become fossilized. A reconstruction of Surcophyton carpets is doubtful; (e) Studies on fossil hard-substrate-communities should also include surrounding loose grounds and/or soft bottoms.
Acknowledgements
The authors are grateful to B. RIEGL, M. RASSER (Institute of Palaeontology, University of Vienna) and A. M. MANSOUR (South Valley University, Qena) for help with field work; R. JANSSEN, E. NEUBERT (both Senckenberg Museum, Frankfurt) and P. G. OLIVER (National Museum of Wales, Cardiff) supported the mollusc identification. Thanks are due to K. W. FLFSA (Department of Ecology and Evolutionary Biology, University of Arizona, Tucson) and P. G. OLIVER (National Museum of Wales, Cardiff) for their careful review of the manuscript. This study was supported by the ‘Hoch- schuljubiliiumsstiftung der Stadt Wien’ and by Project P10715-CEO of the ‘Fonds zur FBrderung der wissenschaftlichen Forschung’ (FWF).
References
ARNOUD, P. M. & B. A. THOMASSIN, 1990: Habits and morphological adaptations of mytilids (Mollusca: Bivalvia) from coastal and reefal environments in south-west Malagasia (Indian Ocean). In: B. MORTON (Ed.), The Bivalvia - Proceedings of a Memorial Symposium in Honour of Sir Charles MAURICE YONGE, Edinburgh, 1986. Hong Kong University Press, Hong Kong: 333-344.
AUSTIN, A. D., S. A. AUSTIN & P. F. SALE, 1980: Community structure of the fauna associated with the coral Pocillopora damicornis (L.) on the Great Barrier Reef. Aust. J. Mar. Freshwater Res., 31:
AYAL, Y. & U. N. SAFRIEL, 1981: Species composition, geographical distribution and habitat charac- teristics of rocky intertidal Cerithiidae (Gastropoda; Prosobranchia) along the Red Sea shores of Sinai. Argamon, 7: 53-72.
BOUCOT, A. J., 1981: Principles of Benthic Marine Paleoecology. Academic Press, New York; XV f 4 6 3 PP.
FABRICIUS, K. E., Y. BENAYAHU & A. GENIN, 1995: Herbivory in Asymbiotic Soft Corals. Science, 268: 9CL92.
FRANK, P. W., 1969: Growth rates and longevity of some gastropod molluscs on the coral reef at Heron Island. Oecologia, 2 232-250.
FURSICH, F. T. & T. J. PALMER, 1975: Open crustacean burrows associated with hardgrounds in the Jurassic of the Cotswolds, England. Geol. Assoc. (London), Proc., 86 171-181.
GOLDRING, R., 1995: Organisms and the substrate: response and effect: In: D. W. J. BOSENCE & P. A. ALLISON (Eds.), Marine Palaeoenvironmental Analysis from Fossils. Geol. SOC. Spec. Publ., 8 3
163- 174.
15 1-1 80. HADFIELD, M. G., 1976: Molluscs associated with living corals. Micronesica, 12: 133-148. HAUNOLD, T. G., C. BAAL & W. E. PILLER, 1997: Benthic Foraminiferan Associations in the Northern
HOFFMAN, A,, 1978: System concepts and the evolution of benthic communities. Lethaia, 11: 179-183. HUGHES, R. N. & A. H. LEWIS, 1974: On the spatial distribution, feeding and reproduction of the
JOHNSON. M. E. 1988: Why are ancient rocky shores so uncommon? J. Geol., 96: 469480. KLEEMANN, K., 1990: Coral Associations, Biocorrosion, and Space Competition in Pedum spon-
Bay of Safaga, Red Sea, Egypt. Mar. Micropaleontol., 2 9 185-210.
vermetid gastropod Dendropoma maxima. J. Zool., London, 172 53 1-547.
dvloideum (GMELIN) (Pectinacea, Bivalvia). P.S.Z.N.I.: Marine Ecology, 11: 77-94.

378 ZUSCHIN & PILLER
--, 1992: Coral Communities and Coral-Bivalve Associations in the Northern Red Sea at Safaga,
KREBS, C. J., 1989: Ecological Methodology. Harper & Row, New York; X+654 pp. LEE, S. Y. & B. MORTON, 1985: The Hong Kong Mytilidae. In: B. MORTON & D. DUDGEON (Eds.),
Proceedings of the Second International Workshop on the Malacofauna of Hong Kong and Southern China, Hong Kong, 1983. Hong Kong University Press, Hong Kong: 49-76.
MAGURRAN, A. E., 1988: Ecological Diversity and Its Measurement. Croom Helm, London - Sydney; X+179pp.
MASTALLER, M., 1978: The marine molluscan assemblages of Port Sudan, Red Sea. Zool. Meded., 53: 117-144.
MORTON, B., 1983: Coral-Associated Bivalves in the Indo-Pacific. In: W. D. RUSSEL-HUNTER (Ed.), The Mollusca. Vol. 6. Academic Press, New York: 139-224.
NEBELSICK, J. H., 1992a: The Northern Bay of Safaga (Red Sea, Egypt): An Actuopalaeontological Approach. 111. Distribution of Echinoids. Beitr. Pallontol. Gsterreich, 17: 5-79.
--, 1992b: Echinoid Distribution by Fragment Identification in the Northern Bay of Safaga. Red Sea, Egypt. Palaios, 7: 3 16-328.
OLIVER, P. G . , 1992: Bivalved Seashells of the Red Sea. Verlag Christa Hemmen, Wiesbaden; 330 pp. --, 1995: Bivalvia. In: S. P. DANCE (Ed.), Seashells of Eastern Arabia: Motivate Publishing: 194-281. PALMER, T. J . , 1978: Burrows at certain omission surfaces in the Middle Ordovician of the Upper
- -, 1982: Cambrian to Cretaceous changes in hardground communities. Lethaia, 1 5 309-323. PILLER, W. E., 1994: The Northern Bay of Safaga (Red Sea. Egypt): An Actuopaleontological Approach.
IV. Thin Section Analysis. Beitr. Palaontol., 18: 1-73. - -&A. M. MANSOUR, 1990: The Northern Bay of Safaga (Red Sea, Egypt): An Actuopalaeontological
Approach. 11. Sediment analyses and sedimentary facies. Beitr. Paliiontol. dsterreich, 1 6 1-102. -- & A. M. MANSOUR, 1992: Origin and Transport Mechanisms of Non-Carbonate Sediments in a
Carbonate-Dominated Environment (Northern Safaga Bay, Red Sea, Egypt). Abh. Geol. Bun- desanstalt, 5 0 369-379.
-- & P. PERVESLER, 1989: The Northern Bay of Safaga (Red Sea, Egypt): An Actuopalaeontological Approach. I . Topography and Bottom Facies. Beitr. Pallontol. Osterreichs, 15: 103-147.
-- & M. RASSER, 1996: Rhodolith formation induced by reef erosion in the Red Sea, Egypt. Coral Reefs, 15: 191-198.
POWELL, E. N., R. J. STANTON Jr., A. LOGAN & M. A. CRAIG, 1992: Preservation of Mollusca in Copano Bay, Texas. The long-term record. Palaeogeogr., Palaeoclimatol., Palaeoecol., 95 209-228.
RASSER, M . & W. E. PILLER, 1997: Depth distribution of calcareous encrusting associations in the northern Red Sea (Safaga, Egypt) and their geological implications: Proceedings of the 8th Inter- national Coral Reef Symposium (Panama, 1996). 1: 743-748.
RIEGL, B., W. E. PILLER & M. RASSER. 1996: Rolling stones: first report of a free living Acroporu anthocercis (BROOK) from the Red Sea. Coral Reefs, I 5 149-1 50.
-- & W. E. PILLER, 1997: Distribution of coral associations in Northern Safaga Bay (Red Sea, Egypt). Facies, 36: 141-162.
SAFRIEL, U . N.. T. FELSENBURG & A. GILBOA, 1980a: The distribution of Brachidonres variahilis (KRAUSS) along the Red Sea coasts of Sinai. Argamon. 7: 3 1 4 3 .
--, A. GILBOA & T. FELSENBURG, 1980b: Distribution of rocky intertidal mussels in the Red Sea coasts of Sinai, the Suez Canal and the Mediterranean coast of Israel, with special reference to recent colonizers. J. Biogeogr., 7: 3942.
SCHUHMACHER, H., 1993: Impact of Some Corallivorous snails on Stony Corals in the Red Sea. Proceedings of the Seventh International Coral Reef Symposium, Guam 1992.2: 840-846.
--, W. KIENE & W.-C. DULLO (coordinators), 1995: Factors Controlling Holocene Reef Growth: An Interdisciplinary Approach. Facies, 3 2 30-37.
SHEPPARD, A. L. S., 1984: The Molluscan Fauna of Chagos (Indian Ocean) and an Analysis of its Broad Distribution Patterns. Coral Reefs, 3: 43-50.
SHIMOYAMA. S.. 1979: Modification of shell distribution Datterns bv hermit crabs in a Drotected shore.
Egypt. Facies, 2 6 125-134.
Mississippi Valley. J. Paleontol., 52( I ) : 109-1 17.
Mar. Sci., I I : 527-535. TAYLOR. J. D.. 1971: Intertidal zonation of Aldebra Atoll. Philos. Trans. R. SOC. London. 254B 173-
213:
Sudanese Red Sea. J. Nat. Hist.. 1 8 175-209. -- & D. G. REID, 1984: The abundance and trophic classification of molluscs upon coral reefs in the
WALKER, S. E., 1989: Hermit Crabs as Taphonomic Agents. Palaios, 4 439452. ZUSCHIN, M. & W. E. PILLER. 1997a: Gastropod shells recycled - an example from a rocky tidal Rat in
- & W. E. PILLER, 1997b: Bivalve distribution on coral carpets in the northern Bay of Safaga (Red the northern Red Sea. Lethaia, 30 127-134.
Sea, Egypt) and its relation to environmental parameters. Facies. 37: 193-204.
Related Documents











