| | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 1 KRÓTKO SZKOŁA EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA [email protected] | © for the article by the Authors © for the edition by Instytut Badań Edukacyjnych ISSN 1643-8779 NUMER 2 (46) ROK 2013 KWARTALNIK BIOLOGICZNA I ŚRODOWISKOWA Komórki macierzyste: część II Molekularny Dr Jekyll i Mr Hyde Bioróżnorodność a ochrona patentowa Osiągnięcia przyrodnicze trzecioklasistów Angielskie narzędzia dydaktyczne Czy warto studiować biologię?

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

| | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 1
KR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors © for the edition by Instytut Badań Edukacyjnych
ISSN 1643-8779
NUMER 2(46) ROK 2013
KWARTALNIK
BIOLOGICZNA I ŚRODOWISKOWA
Komórki macierzyste: część II
Molekularny Dr Jekyll i Mr Hyde
Bioróżnorodność a ochrona patentowa
Osiągnięcia przyrodnicze trzecioklasistów
Angielskie narzędzia dydaktyczne
Czy warto studiować biologię?

1
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the edition by Instytut Badań Edukacyjnych 2013
NAUKA
3 Barbara Świerczek, Damian Dudka, Karolina ArchackaKomórki macierzyste. Część II – pluripotencjalne komórki macierzyste
12 Joanna UchańskaBioróżnorodność a ochrona patentowa
23 Kamil Lisiecki Molekularny Dr Jekyll i Mr Hyde
33 Agnieszka OsówniakSiarka i jej związki jako źródła pierwiastka budulcowego dla bakterii
SZKOŁA
44 Ewa Oleńska, Alina StankiewiczMetody obliczeniowe w genetyce
51 Krzysztof KonarzewskiOsiągnięcia przyrodnicze trzecioklasistów
55 Marcin Zaród Małe eksperymenty, wielkie odkrycia – praktyka eksperymentalna w edukacji przyrodniczej
60 Urszula PoziomekAngielskie narzędzia dydaktyczne
62 Urszula PoziomekŁańcuchy i sieci pokarmowe na polu – zajęcia z biologii
69 Pracownia Przedmiotów Przyrodniczych IBENowe zadania PPP
73 Ryszard KowalskiOPINIE: Czy warto studiować biologię?
KRÓTKO
76 Biologia Campbella – recenzja książki
78 XVIII Konferencja Dydaktyków Biologii – zapowiedź
79 Nowości ze świata nauki
83 Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce
2/2013w numerze:
biologiageografia
fizykachemia
przyrodabadania
zdrowie
środowiskozadania
narzędzia w internecie
narzędzia dydaktyczne
scenariusze zajęćpomysłyjak uczyć
jak zainteresować
szykuje się ważne wydarzenie?
poinformuj nas o nim
najnowsze odkryciainformacje
recenzjewydarzenia
NAUKA – artykuły przede wszystkim o charakterze przeglądowym, adresowane do osób zainteresowanych naukami przyrodniczymi – dotyczą głównie zagadnień biologii i biochemii, ale mogą też obejmować problematykę pozostałych dyscyplin przyrodniczych. W naszym zamierzeniu mają zarówno dostarczyć rzetelną wiedzę, jak i skłonić do dyskusji, jakie treści i w jakiej formie warto proponować nauczycielom, by pomóc im w nauczaniu. Sprawia to, że dział ten ma charakter przede wszystkim pedagogiczny.
SZKOŁA – artykuły lub materiały przedstawiające rozmaite źródła informacji (np. serwisy i kursy internetowe), uwarunkowania nauczania (m.in. prawne i społeczne), a także metody pracy z uczniami, konspekty i scenariusze lekcji. W każdym numerze – najnowsze zadania Pracowni Przedmiotów Przyrodniczych IBE.
KRÓTKO – recenzje (książek, a nawet płyt z muzyką), zapowiedzi wydarzeń i relacje z nich, depesze o nowościach ze świata nauki oraz dyskusje i komentarze.
KONSPEKT
KONSPEKT
W ZAŁĄCZNIKU – MATERIAŁY
Na okładce: struktura przestrzenna włókna amyloidowego prionu HET-s określona metodą jądrowego rezonansu magnetycznego (NMR). Rysunek wykonano w programie UCSF Chimera (www.cgl.ucsf.edu/chimera) na podstawie danych zdeponowanych w Protein Data Bank (www.pdb.org) pod numerem dostępu 2RNM.

2
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the edition by Instytut Badań Edukacyjnych 2013
Od redakcjiTakao Ishikawa
Szanowni Państwo,oddajemy w Państwa ręce nu-
mer 2013/2 Edukacji Biologicznej i Środowiskowej. Znajdą w nim Państwo m.in. drugi artykuł z se-rii „komórki macierzyste”, autor-stwa Barbary Świerczek, Damiana Dudki i dr Karoliny Archackiej z Wydziału Biologii Uniwersytetu Warszawskiego. Oprócz artykułów z biologii i chemii w dziale NA-UKA publikujemy również pracę Bioróznorodnosc a ochrona paten-towa mgr Joanny Uchańskiej, która porusza zagadnienia prawne doty-czące ochrony różnorodności bio-logicznej. Każdy przyrodnik wie, że jest to temat ważny, ale rzadko rozpatruje go pod względem praw-nym. Wierzę, że artykuł ten będzie doskonałym uzupełnieniem wie-dzy o różnorodności biologicznej i jej ochronie.
Dział SZKOŁA w tym nume-rze jest reprezentowany m.in. przez artykuł dr Oleńskiej i dr Stan-kiewicz, które przybliżą wyko-rzystanie umiejętności matema-tycznych w nauczaniu genetyki. Z kolei z artykułu prof. Krzysztofa Konarzewskiego dowiemy się, co nasi trzecioklasiści mogliby osiąg-nąć, gdyby nie okrojony program i anachroniczna metodyka kształ-cenia przyrodniczego. Nie zabrakło
również propozycji metodycznych mgr. Marcina Zaroda, konspektu lekcji z biologii wykorzystującego angielskich doświadczenia w za-kresie dydaktyki oraz nowych za-dań opracowanych przez Pracow-nię Przedmiotów Przyrodniczych Instytutu Badań Edukacyjnych, które obejmują nie tylko biologię, lecz także chemię i fizykę.
Sądzę, że wielu Czytelników EBiŚ nie raz zastanawiało się, czy warto studiować biologię. Czy jest to strata czasu, czy może ra-czej obiecująca inwestycja na przyszłość? O tym pisze dr Ryszard Kowalski z Instytutu Biologii Uni-wersytetu Przyrodniczo-Humani-stycznego w Siedlcach. Publikuje-my ten tekst z nadzieją, że również Państwo podzielą się swoimi opi-niami i wrażeniami na ten temat. Oddając w Państwa ręcę ten numer EBiŚ mam nadzieję, że kwartal-nik będzie nie tylko źródłem in-formacji i nowości ze świata nauk przyrodniczych, lecz także forum osób związanych z szeroko pojętą edukacją przyrodniczą, dla których dalszy los uczniów i studentów nie jest obojętny.
Właśnie rozpoczyna się okres wakacji, dlatego życzę Państwu przede wszystkim udanego wypo-czynku. Proszę też pamiętać, że prenumerata EBiŚ pozwoli Pań-stwu zabrać kwartalnik na wakacje – w tablecie albo nawet w telefonie komórkowym!
Takao Ishikawa
Redakcja
Redaktor naczelny: Takao Ishikawa
Sekretarz redakcji: Marcin Trepczyński
Redaktorzy merytoryczni: Urszula Poziomek, Jolanta Skorupa-Korycka
Kontakt z redakcją i propozycje tekstów: [email protected] internetowa: ebis.ibe.edu.plAdres redakcji: ul. Górczewska 8, 01-180 Warszawa
Rada naukowa
przewodniczący Rady: prof. zw. dr hab. Adam Kołątaj (Instytut Genetyki i Hodowli Zwierząt PAN, Jastrzębiec),
zast. przewodniczącego: prof. dr hab. Katarzyna Potyrała (Uniwersytet Pedagogiczny w Krakowie),
a także: dr hab. Ondrej Hronec (Uniwersytet w Presowie, Słowacja), prof. dr hab. Daniel Raichvarg (Uniwersytet Burgundzki w Dijon, Francja), prof. dr hab. Valerij Rudenko (Wydział Geograficzny, Uniwersytet w Czerniowcach, Ukraina), prof. zw. dr hab. Wiesław Stawiński (emerytowany profesor Uniwersytetu Pedagogicznego w Krakowie), dr Renata Jurkowska (Uniwersytet w Stuttgarcie, Niemcy)
Poza radą czasopismo posiada również zespoły doradcze oraz stałych recenzentów – zob. na stronie: ebis.ibe.edu.pl
Wydawnictwo
Wydawca: Instytut Badań Edukacyjnych, ul. Górczewska 8, 01-180 Warszawa
Projekt okładki: Marcin Broniszewski
Redakcja techniczna: Elżbieta Gątarek
Skład i łamanie: Marcin Trepczyński
czasopismo punktowane: 4 punkty, indeksowane w bazach CEJSH i Index Copernicus
wersją referencyjną czasopisma jest wydanie elektroniczne
opublikowane na stronie: ebis.ibe.edu.pl

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 3
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Komórki macierzyste. Część II – pluripotencjalne komórki macierzysteBarbara Świerczek, Damian Dudka, Karolina Archacka
dr Karolina Archacka: Zakład Cytologii, Wydział Biologii, Uniwersytet Warszawski; [email protected]
Wstęp
W pierwszej części materiału na temat komórek macierzystych (Komórki macierzyste. Częsc I – wpro-wadzenie, EBIŚ 1/2013) przedstawiona została ich wstępna charakterystyka i klasyfikacja. Jako przykład pluripotencjalnych komórek macierzystych wymienio-no zarodkowe komórki macierzyste (komórki ES, ang. embryonic stem cells) oraz indukowane pluripotencjal-ne komórki macierzyste (komórki iPS, ang. induced pluripotent stem cells). Choć nie są to jedyne znane komórki pluripotencjalne, niniejszy artykuł dotyczyć będzie przede wszystkim komórek ES i iPS. Uzyskanie tych komórek było bez wątpienia przełomem w nauce, o czym najlepiej świadczy fakt, że badacze, którzy tego dokonali, zostali uhonorowani Nagrodą Nobla w dzie-dzinie fizjologii lub medycyny. Pluripotencjalne komór-ki macierzyste są zdolne do utworzenia wszystkich ro-dzajów komórek i tkanek budujących organizm ssaka. W związku z tym mogłyby one stanowić uniwersalne źródło materiału do przeszczepu w celu łagodzenia ob-jawów lub leczenia różnych chorób. W niniejszym ar-tykule omówimy historię badań, które doprowadziły do uzyskania pluripotencjalnych komórek macierzy-stych, a także przedstawimy możliwości ich zastosowa-nia w nauce i medycynie oraz związane z tym nadzieje i obawy.
Czy różnicowanie komórek jest procesem nieodwracalnym?
Zapłodnienie gamety żeńskiej (oocytu) przez ga-metę męską (plemnik) prowadzi do powstania pierw-szej komórki nowego organizmu – zygoty. Zygota jest komórką totipotencjalną – to z niej wywodzić się będą wszystkie tkanki i narządy rozwijającego się osob-nika, a także struktury niezbędne do prawidłowego
otrzymano: 18.04.2013; przyjęto: 20.05.2013; opublikowano: 28.06.2013
Artykuł powstał podczas realizacji projektu „Modification of myogenic potential of pluripotent stem cells – the role of sonic hedgehog and interle-ukin 4” realizowanego w ramach programu POMOST Fundacji na rzecz Nauki Polskiej współfinansowanego przez Unię Europejską ze środków Europejskiego Funduszu Rozwoju Regionalnego.
Barbara Świerczek: Zakład Cytologii, Wydział Biologii, Uniwersytet Warszawski; [email protected]
Damian Dudka: Zakład Cytologii, Wydział Biologii, Uni-wersytet Warszawski; [email protected]
Niniejszy artykuł może być pomocny przy realizacji wymagań podsta-wy programowej z biologii dla szkół ponadgimnazjalnych w zakresie rozszerzonym: treści nauczania VI.8.6 oraz VI.8.8 – Uczeń przedstawia sposoby i cele otrzymywania komórek macierzystych oraz dyskutuje problemy etyczne związane z rozwojem inżynierii genetycznej i bio-technologii, w tym przedstawia kontrowersje towarzyszące badaniom nad klonowaniem terapeutycznym człowieka i formułuje własną opinię na ten temat.
Streszczenie:
Pluripotencjalne komórki macierzyste są zdolne do różni-cowania we wszystkie rodzaje komórek i tkanek budujących organizm ssaka. Podczas rozwoju zarodkowego komórki pluripotencjalne zlokalizowane są w węźle zarodkowym blastocysty, z którego – na dalszych etapach rozwoju – wy-kształci się ciało zarodka. Z komórek węzła zarodkowego blastocysty uzyskiwane są zarodkowe komórki macierzyste (komórki ES, ang. embryonic stem cells). Innym przykła-dem pluripotencjalnych komórek macierzystych są induko-wane pluripotencjalne komórki macierzyste (komórki iPS, ang. induced pluripotent stem cells) otrzymywane w wyniku procesu określanego jako reprogramowanie komórek. Za-równo komórki ES, jak i komórki iPS stanowią cenny mo-del badawczy, a także rozważane są jako źródło materiału
do przeszczepu w medycynie regeneracyjnej. Uzyskanie obu wymienionych rodzajów pluripotencjalnych komórek ma-cierzystych jest uznawane za przełom w nauce i medycynie, i zostało uhonorowane Nagrodą Nobla w dziedzinie fizjolo-gii lub medycyny. W 2007 r. otrzymali ją Brytyjczyk Martin Evans oraz amerykańscy uczeni Mario Capecchi i Olivier Smithies, których badania doprowadziły do uzyskania komó-rek ES i otrzymania przy ich wykorzystaniu modyfikowanych genetycznie myszy. Natomiast w 2012 r. Nagrodą Nobla wy-różnieni zostali Brytyjczyk John Gurdon i Japończyk Shinya Yamanaka za prace nad reprogramowaniem komórek. Słowa kluczowe: Pluripotencjalne komórki macierzyste, komórki ES, komórki iPS, reprogramowanie, wielokierunkowe różnicowanie, medycyna regeneracyjna, terapie komórkowe

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 4
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
rozwoju: błony płodowe oraz łożysko. W trakcie roz-woju zarodkowego i płodowego, a następnie w okresie pourodzeniowym komórki stopniowo ulegają specja-lizacji, w wyniku czego zmniejsza się ich potencjał do różnicowania. Przez wiele lat uważano, że proces ten, tj. ograniczenie potencjału do różnicowania związane ze specjalizacją komórek, jest nieodwracalny. Na prze-strzeni wieków sformułowano wiele teorii dotyczących tego zagadnienia. W XIX wieku niemiecki biolog Au-gust Weismann przedstawił teorię, zgodnie z którą pro-ces różnicowania się komórek miał się wiązać z utratą części zawartego w nich materiału genetycznego. Infor-macja genetyczna, która nie była „potrzebna” komórce do pełnienia przez nią określonej funkcji w organizmie, miała być z niej „usuwana”. Komórkami, które zgodnie z teorią Weismanna zachowywały i przekazywały kom-pletną informację genetyczną, były wyłącznie komórki rozrodcze (Weismann, 1893). Warto tu przypomnieć, że w tamtym czasie nie znano jeszcze takich pojęć, jak np. „gen”. Nieznana była także budowa i rola DNA jako nośnika informacji genetycznej. Teoria Weismanna zo-stała po raz pierwszy podważona przez Yvesa Delage’a, francuskiego zoologa znanego przede wszystkim z prac poświęconych anatomii i fizjologii bezkręgowców. W opublikowanej w 1895 r. książce Delage stwierdził, że zastąpienie jądra komórkowego zygoty jądrem po-chodzącym z jakiejkolwiek innej komórki ciała zarodka nie powinno wpłynąć na prawidłowy przebieg rozwoju zarodkowego (Beetschen i Fischer, 2004). Delage oparł swoją hipotezę na założeniu, że skoro wszystkie komór-ki organizmu powstają w wyniku podziałów mitotycz-nych zygoty, to muszą one zawierać tę samą informację genetyczną. Mitoza jest bowiem procesem, w wyniku którego powstają dwie komórki potomne, identyczne pod względem genetycznym z komórką rodzicielską. Ze względu na ówczesny stan wiedzy i techniki De-lage nie był w stanie przeprowadzić eksperymentów
potwierdzających słuszność postawionej przez niego hipotezy. Dokonał tego w II połowie XX wieku John Gurdon, brytyjski biolog, który za swoje osiągnięcia został uhonorowany w 2012 r. Nagrodą Nobla w dzie-dzinie fizjologii lub medycyny. Opisane przez niego w 1962 r. doświadczenia polegały na przeniesieniu ją-dra komórkowego pobranego z komórki budującej jeli-to kijanki do oocytu pozbawionego wcześniej własnego materiału genetycznego (ryc. 1). Z tak „skonstruowane-go” oocytu Gurdon otrzymał prawidłowo rozwiniętą
kijankę (Gurdon, 1962). Oznaczało to, że na podstawie informacji genetycznej zawartej w jądrze komórkowym wyspecjalizowanej komórki może rozwinąć się w peł-ni wykształcony osobnik. Innymi słowy, różnicowanie komórek nie prowadzi do usuwania z nich „niepotrzeb-nej” informacji genetycznej. Zróżnicowana komórka nadal zawiera wszystkie informacje niezbędne do po-wstania wszystkich rodzajów komórek budujących za-rodek, a tym samym niezbędne do jego prawidłowego rozwoju. Kolejne badania wykazały, że specjalizacja ko-
Ryc. 1. Doświadczenie Gurdona
W swoim pionierskim doświadczeniu John Gurdon wykorzystał oocyty płaza pozbawione – na skutek napromienienia światłem UV – własne-go materiału genetycznego. Do przygotowanego w ten sposób oocytu Gurdon wprowadził jądro komórkowe pochodzące z komórki jelita kijanki. Na podstawie informacji genetycznej zawartej w przeszczepionym do oocytu jądrze rozwinęła się kijanka. Pod względem genetycz-nym była ona identyczna z kijanką, od której pochodziły komórki jelita.
Wszystkie zamieszczone w artykule zdjęcia i schematy zostały wykonane przez autorów tekstu.
Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 5
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
mórek związana jest z włączeniem (aktywacją) genów kluczowych dla ich różnicowania, decydujących o ich prawidłowej budowie i funkcji. Równocześnie geny ważne dla wczesnych etapów rozwoju zarodkowego zostają wyłączone (uśpione). Jednak, jak wykazały pio- nierskie doświadczenia Gurdona, a także innych ba-daczy, w odpowiednich warunkach te „wczesne” geny mogą zostać na nowo „obudzone”. Odkrycie to zapo-czątkowało intensywny rozwój badań, które z czasem pozwoliły na udoskonalenie techniki klonowania or-ganizmów, a ostatecznie doprowadziły do uzyskania w 2006 r. pierwszych komórek iPS (Ciemerych i Kubiak, 2013). Dużo wcześniej, tj. w latach 80. XX wieku bada- czom udało się otrzymać inne pluripotencjalne komór-ki macierzyste – komórki ES.
Jak uzyskano komórki ES?
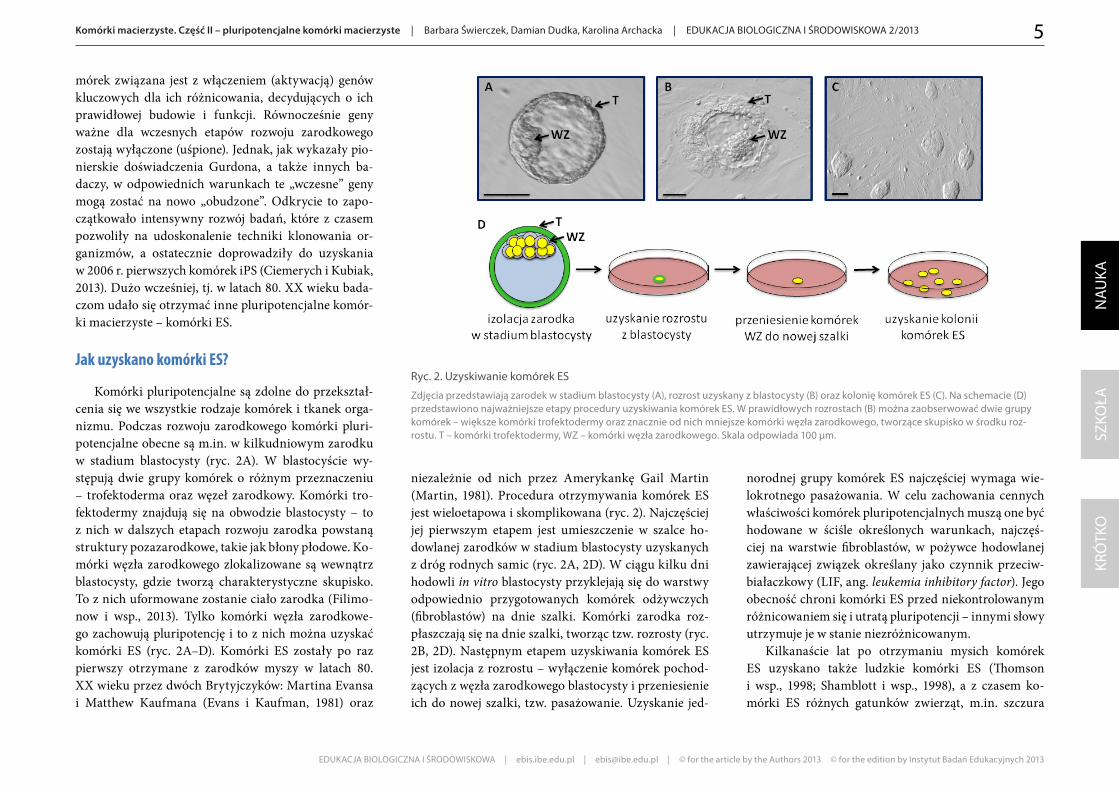
Komórki pluripotencjalne są zdolne do przekształ-cenia się we wszystkie rodzaje komórek i tkanek orga-nizmu. Podczas rozwoju zarodkowego komórki pluri-potencjalne obecne są m.in. w kilkudniowym zarodku w stadium blastocysty (ryc. 2A). W blastocyście wy-stępują dwie grupy komórek o różnym przeznaczeniu – trofektoderma oraz węzeł zarodkowy. Komórki tro-fektodermy znajdują się na obwodzie blastocysty – to z nich w dalszych etapach rozwoju zarodka powstaną struktury pozazarodkowe, takie jak błony płodowe. Ko-mórki węzła zarodkowego zlokalizowane są wewnątrz blastocysty, gdzie tworzą charakterystyczne skupisko. To z nich uformowane zostanie ciało zarodka (Filimo-now i wsp., 2013). Tylko komórki węzła zarodkowe-go zachowują pluripotencję i to z nich można uzyskać komórki ES (ryc. 2A–D). Komórki ES zostały po raz pierwszy otrzymane z zarodków myszy w latach 80. XX wieku przez dwóch Brytyjczyków: Martina Evansa i Matthew Kaufmana (Evans i Kaufman, 1981) oraz
niezależnie od nich przez Amerykankę Gail Martin (Martin, 1981). Procedura otrzymywania komórek ES jest wieloetapowa i skomplikowana (ryc. 2). Najczęściej jej pierwszym etapem jest umieszczenie w szalce ho-dowlanej zarodków w stadium blastocysty uzyskanych z dróg rodnych samic (ryc. 2A, 2D). W ciągu kilku dni hodowli in vitro blastocysty przyklejają się do warstwy odpowiednio przygotowanych komórek odżywczych (fibroblastów) na dnie szalki. Komórki zarodka roz-płaszczają się na dnie szalki, tworząc tzw. rozrosty (ryc. 2B, 2D). Następnym etapem uzyskiwania komórek ES jest izolacja z rozrostu – wyłączenie komórek pochod-zących z węzła zarodkowego blastocysty i przeniesienie ich do nowej szalki, tzw. pasażowanie. Uzyskanie jed-
Ryc. 2. Uzyskiwanie komórek ES
Zdjęcia przedstawiają zarodek w stadium blastocysty (A), rozrost uzyskany z blastocysty (B) oraz kolonię komórek ES (C). Na schemacie (D) przedstawiono najważniejsze etapy procedury uzyskiwania komórek ES. W prawidłowych rozrostach (B) można zaobserwować dwie grupy komórek – większe komórki trofektodermy oraz znacznie od nich mniejsze komórki węzła zarodkowego, tworzące skupisko w środku roz-rostu. T – komórki trofektodermy, WZ – komórki węzła zarodkowego. Skala odpowiada 100 µm.
norodnej grupy komórek ES najczęściej wymaga wie-lokrotnego pasażowania. W celu zachowania cennych właściwości komórek pluripotencjalnych muszą one być hodowane w ściśle określonych warunkach, najczęś-ciej na warstwie fibroblastów, w pożywce hodowlanej zawierającej związek określany jako czynnik przeciw-białaczkowy (LIF, ang. leukemia inhibitory factor). Jego obecność chroni komórki ES przed niekontrolowanym różnicowaniem się i utratą pluripotencji – innymi słowy utrzymuje je w stanie niezróżnicowanym.
Kilkanaście lat po otrzymaniu mysich komórek ES uzyskano także ludzkie komórki ES (Thomson i wsp., 1998; Shamblott i wsp., 1998), a z czasem ko-mórki ES różnych gatunków zwierząt, m.in. szczura

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 6
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
(Buehr i wsp., 2008; Li i wsp., 2008) oraz rezusa (Thom-son i wsp., 1995). Komórki ES różnych gatunków zwier-ząt często mają inne wymagania dotyczące warunków hodowli, a ich optymalizacja w wielu wypadkach była kluczowa dla uzyskania tych komórek. Bez względu na to, z jakiego gatunku zwierząt pochodzą komórki, ich pluripotencja musi zostać pozytywnie zweryfikowana, zanim zostaną one określone jako komórki ES.
W jaki sposób można sprawdzić pluripotencję komórek?
Pluripotencję komórek, czyli ich zdolność do różni-cowania we wszystkie rodzaje komórek i tkanek, można zweryfikować zarówno in vitro, jak i in vivo (Cieme-rych, 2008; Ciemerych i Kubiak, 2013). Komórki ES, które hodowane są w warunkach przeciwdziałających ich różnicowaniu, tworzą charakterystyczne skupiska (kolonie), zbudowane ze ściśle przylegających do siebie komórek (ryc. 2C). Jeśli natomiast komórki te będą ho-dowane w warunkach uniemożliwiających ich przykle-jenie się do szalki (czyli w zawiesinie) oraz w pożywce pozbawionej czynników utrzymujących je w stanie niezróżnicowanym (np. LIF), to rozpoczną one różni-cowanie i spontanicznie utworzą trójwymiarowe agre-gaty określane jako kule zarodkowe (EB, ang. embryoid bodies; Czerwińska i Ciemerych, 2012; ryc. 3). W kolej-nych dniach hodowli EB powiększają się, a zachodzące w nich procesy różnicowania komórek są analogiczne do tych, które mają miejsce w zarodku we wczesnych etapach jego rozwoju. Na przykład u myszy, w 8. dniu rozwoju zarodkowego zachodzi proces gastrulacji pro-wadzący do powstania trzech listków zarodkowych – ekto-, mezo- i endodermy. W kolejnych etapach roz-woju z tych trzech grup komórek będą rozwijały się określone tkanki organizmu, np. z mezodermy powsta-nie tkanka mięśniowa, a z ektodermy – tkanka nerwo-wa. Listki zarodkowe formowane są także podczas ho-dowli EB. Początkowo kule zbudowane są z ektodermy i otaczającej jej warstwy endodermy pierwotnej, nato-miast 5. dnia hodowli obecne są w nich już wszystkie trzy listki zarodkowe. Na późniejszych etapach hodowli kul można zaobserować pojawienie się komórek takich, jak np. kurczące się kardiomiocyty. Zastosowanie ściśle określonych warunków hodowli EB pozwala na uzyska-nie także wielu innych rodzajów wyspecjalizowanych
Ryc. 3. Uzyskiwanie kul zarodkowych
W celu uzyskania EB komórki ES hodowane są w tzw. wiszących kroplach. Krople pożywki zawierające komórki ES umieszczane są na wieczku szalki hodowlanej (A). Po odwróceniu wieczka komórki ES obecne w wiszących kroplach tworzą EB (B, C). Zdjęcie C przed-stawia kulę zarodkową w 5 dniu hodowli. EB umieszczone w wa-runkach umożliwiających ich przyklejenie się do szalki rozrastają się (D). Obecne w nich komórki migrują po szalce i w określonych warunkach mogą przekształcić się w komórki różnych tkanek. Skala odpowiada 100 µm.

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 7
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Myszy chimerowe można uzyskać przez połączenie ko-mórek uzyskanych z jednego zarodka z komórkami in-nego. W praktyce komórki „dawcy” są wprowadzane do blastocysty – „biorcy” za pomocą specjalnego urządze-nia nazywanego mikromanipulatorem. Do blastocysty – „biorcy” można wstrzyknąć także komórki testowane pod względem pluripotencji. Uzyskane w ten sposób za-rodki chimerowe przeszczepiane są do macicy samicy myszy, gdzie mogą kontynuować rozwój. Jeśli umiesz-czone w blastocyście komórki rzeczywiście są komór-kami pluripotencjalnymi, to wezmą one udział, razem z „oryginalnymi” komórkami budującymi blastocystę – „biorcę”, w powstawaniu wszystkich tkanek i narządów rozwijającego się osobnika. W rezultacie, tkanki te zbu-dowane będą z dwóch rodzajów komórek różniących się pod względem genetycznym. Jeśli organizm myszy będzie zbudowany wyłącznie z „oryginalnych” komó-rek blastocysty – „biorcy” lub udział wstrzykniętych do niej komórek będzie znikomy, to będzie to oznaczać bę-dzie, że testowane komórki nie wykazują pluripotencji (Ciemerych, 2008; Ciemerych i Kubiak, 2013). Niekiedy zdarza się, że badane pod względem pluripotencji ko-mórki formują EB, ale nie są w stanie utworzyć potwor-niaków czy uczestniczyć w rozwoju myszy chimerowej. Taki wynik stawia pod znakiem zapytania ich zdolność do wielokierukowego różnicowania.
Jakie jest znaczenie komórek ES dla rozwoju nauki i medycyny?
Uzyskanie komórek ES umożliwiło intensywny rozwój badań dotyczących mechanizmów regulujących pluripotencję i różnicowanie komórek. Wcześniej śle-dzenie losów komórek na wczesnych etapach rozwoju zarodkowego było ograniczone, a w przypadku zarod-ków ludzkich wręcz niemożliwe ze względu na kwestie etyczne. Podobieństwo procesów zachodzących w EB
komórek, np. neuronów (Guan i wsp., 1999). Różnico-wanie komórek w EB jest najczęściej stosowaną metodą weryfikacji ich pluripotencji in vitro. Tylko te komórki, które okażą się zdolne do utworzenia komórek wywod-zących się z trzech listków zarodkowych mogą zostać uznane za pluripotencjalne. Ostatecznym dowodem na pluripotencję analizowanych komórek są jednak wyni-ki testów in vivo, takich jak tworzenie potworniaków oraz myszy chimerowych.
Pierwsza z metod weryfikacji pluripotencji komórek in vivo polega na wstrzykiwaniu badanych komórek myszom o upośledzonym układzie odpornościowym. Takie zwierzęta nie są zdolne do usunięcia przeszcze-pionych komórek. Najczęściej testowane komórki wstrzykiwane są podskórnie, gdzie po kilku tygodniach – o ile rzeczywiście okażą się pluripotencjalne – utwo-rzą wielotkankowe guzy określane jako potworniaki lub teratomy (gr. teratos oznacza potwora, dziwo). Standar-dowa procedura obejmuje izolację potworniaka, a na-stępnie jego analizę histologiczną. Pozwala ona określić, czy w guzie obecne są komórki i tkanki wywodzące się z wszystkich trzech listków zarodkowych. Jest to dowód na to, że komórki poddane weryfikacji są zdolne do wie-lokierunkowego różnicowania, a zatem są komórkami pluripotencjalnymi (Ciemerych i Kubiak, 2013).
W przypadku mysich komórek innym sposobem weryfikacji ich pluripotencji in vivo jest sprawdzenie, czy są one w stanie uczestniczyć w rozwoju tzw. myszy chimerowych. W mitologii chimerą nazywano stwo-rzenie z głową lwa, ciałem kozy i ogonem węża. Ter-min „myszy chimerowe” stosowany jest do określania zwierząt zbudowanych z co najmniej dwóch rodza-jów komórek różniących się pod względem genetycz-nym. Pierwsze myszy chimerowe uzyskał w latach 60. XX wieku polski embriolog, profesor Andrzej Tarkow-ski, prowadzący badania dotyczące m.in. mechani-zmów kierujących rozwojem ssaków (Tarkowski, 1961).
i zarodkach dało szansę na ich lepsze poznanie i zro-zumienie (Czerwińska i Ciemerych, 2012). Nie mniej istotny jest fakt, że komórki ES można stosunkowo ła-two modyfikować genetycznie, aktywując lub wyłącza-jąc w nich wybrane geny. Daje to szansę na precyzyjne określenie funkcji danego genu w komórce. Przykłado-wo badania prowadzone z wykorzystaniem komórek ES pozbawionych funkcjonalnego genu Nanog wykazały jego kluczową rolę w utrzymaniu pluripotencji komó-rek. Wcześniej funkcja tego genu była tylko częściowo znana. Wiadomo było, że mutacja w genie Nanog pro-wadzi do poważnych zaburzeń we wczesnym etapie rozwoju zarodkowego i śmierci zarodka (Mitsui i wsp., 2003). Doświadczenia z wykorzystaniem komórek ES pozbawionych tego genu wykazały, że jego rola polega na hamowaniu aktywności genów odpowiedzialnych za różnicowanie komórek i równocześnie na stymulacji aktywności genów ważnych dla utrzymania komórek pluripotencjalnych w stanie niezróżnicowanym, takich jak Sox2 i Oct-4 (Loh i wsp., 2006). Innym przykładem mogą być badania nad rolą czynników Pax3 i Pax7 w procesie postawania komórek mięśniowych. Kontro-lowana aktywacja tych genów w komórkach ES umoż-liwiła uzyskanie z komórek pluripotencjalnych funk-cjonalnych komórek mięśniowych, co potwierdziło kluczową rolę Pax3 i Pax7 w rozwoju mięśni szkiele-towych (Darabi i wsp., 2011).
Innym, niezwykle ważnym sposobem wykorzy-stania komórek ES w badaniach naukowych jest moż-liwość uzyskiwania z ich udziałem zwierząt transge-nicznych. Organizmy transgeniczne to takie, których genom został zmodyfkowany poprzez wprowadzenie „obcego” materiału genetycznego (np. fragmentu DNA zsyntetyzowanego w laboratorium albo genu pochod-zącego z innego organizmu). Można je otrzymać w ana-logiczny sposób, jak myszy chimerowe, tzn. poprzez wprowadzenie zmodyfikowanych genetycznie komórek

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 8
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
do blastocysty-„biorcy”, a następnie przeszczepienie uzyskanego w ten sposób transgenicznego zarodka do macicy samicy (Bishop, 2001). Pionierem tej technolo-gii jest Mario Capecchi, amerykański genetyk włoskie-go pochodzenia, który w latach 80. XX wieku uzyskał myszy pozbawione funkcjonalnego genu int1 (Thomas i Capecchi, 1990). Za swoje osiagnięcia Capecchi został uhonorowany w 2007 r. Nagrodą Nobla w dziedzinie medycyny lub fizjologii. Możliwość otrzymywania my-szy pozbawionych funkcjonalnego genu daje szansę na określenie jego roli w funkcjonowaniu organizmu (Ca-pecchi, 2005). Na przykład u myszy pozbawionych genu Hoxa1 wykryto nieprawidłowości w budowie układu krążenia, m.in. w łuku aorty (Makki i Capecchi, 2012). Zwierzęta transgeniczne stanowią także cenne zwierzę-ce modele ludzkich chorób. Na przykład transgeniczne myszy, które syntetyzują nieprawidłową wersję ludzkie-go białka APP wykazują podobne zmiany w układzie nerwowym, jak te obserwowane u osób cierpiących na chorobę Alzheimera (Games i wsp., 1995). Dostępność zwierzęcych modeli ludzkich chorób daje możliwość badania ich molekularnych mechanizmów, a także tes-towania potencjalnych terapii i leków.
Otrzymanie komórek ES było ogromnym przeło-mem zarówno w nauce, jak i w medycynie, dając szan-sę na przeprowadzenie badań wcześniej niemożliwych oraz opracowanie potencjalnych terapii dotychczas nieuleczalnych chorób, takich jak cukrzyca typu I czy choroba Parkinsona. W laboratoriach na całym świecie podjęte zostały badania mające na celu stworzenie sku-tecznej terapii tych i wielu innych chorób. Należy pa-miętać, że żadna z nich nie jest jeszcze wykorzystywana do leczenia ludzi. Dotychczas przeprowadzono jedynie badania laboratoryjne, obejmujące doświadczenia in vi-tro oraz doświadczenia z wykorzystaniem zwierząt labo-ratoryjnych. Wyjątek stanowią próby kliniczne, których wyniki opisano w 2012 r. (Schwartz i wsp., 2012). W ba-
daniu uczestniczyli pacjenci cierpiący na choroby oczu: związane z wiekiem zwyrodnienie plamki żółtej oraz chorobę Stargardta, przejawiającą się uszkodzeniem centralnej części soczewki. Pacjentom tym wstrzyknię-to komórki nabłonka pigmentowego siatkówki uzyska-ne z ludzkich komórek ES. Przed transplantacją komór-ki poddano dokładnym analizom – sprawdzono m.in., czy mają one prawidłową liczbę chromosomów i czy wszystkie z nich uległy zróżnicowaniu. Należy bowiem pamiętać o tym, że wprowadzenie do organizmu nawet niewielkiej liczby niezróżnicowanych komórek pluripo-tencjalnych może doprowadzić do powstania teratom. Schwartz i współpracownicy stwierdzili, że komórki wstrzyknięte w pobliżu siatkówki zasiedliły ją i wzięły udział w tworzeniu nabłonka pigmentowego, którego rola polega na odżywianiu fotoreceptorów oka i ab-sorpcji światła. Nie zaobserwowano żadnych skutków ubocznych zastosowanej terapii, a co najistotniejsze, u pacjentów odnotowano poprawę widzenia. Chociaż długofalowe skutki tej eksperymentalnej terapii po-zostają jeszcze nieznane, dotychczas zaobserwowane efekty dają podstawę do umiarkowanego optymizmu i pozwalają myśleć o wykorzystaniu komórek macierzy-stych w leczeniu m.in. chorób siatkówki.
Należy tu przypomnieć, że możliwość wykorzysta-nia ludzkich komórek ES w medycynie regeneracyjnej budzi kontrowersje natury etycznej ze względu na ich pochodzenie. Komórki te zostały bowiem uzyskane z zarodków ludzkich otrzymanych w wyniku zapłod-nienia in vitro. W 2008 r. opracowano metodę, dzięki której komórki ES mogą być uzyskiwane w sposób, który nie zaburza dalszego rozwoju zarodka. Polega on na pobraniu z zarodka pojedynczej komórki (Chung i wsp., 2008). Pozostałe komórki są zdolne do wypeł-nienia tego „ubytku”. Mimo to „rodowód” komórek ES nadal pozostaje kontrowersyjnym zagadanieniem. Dla-tego też od wielu lat prowadzone są intensywne badania
mające na celu uzyskanie komórek pluripotencjalnych ze źródła niebudzącego kontrowesji etycznych. Badania te doprowadziły m.in. do uzyskania w 2006 r. pierws-zych komórek iPS.
Ekspresowy Nobel, czyli na czym polega fenomen komórek iPS?
Choć od momentu otrzymania pierwszych komórek iPS minęło zaledwie kilka lat, osiagnięcie to już zosta-ło uhonorowane Nagrodą Nobla w dziedzinie fizjologii i medycyny. Jej laureatem w 2012 r. został – obok wspo-mnianego wcześniej Johna Gurdona – Shinya Yamana-ka, który wraz z Kazutochi Takahashim opublikował w 2006 r. „przepis” na komórki iPS (Takahashi i Yama-naka, 2006). Dlaczego uzyskanie tych komórek zosta-ło uznane za osiągnięcie naukowe godne tej nagrody? Otóż komórki iPS, podobnie jak komórki ES, są zdolne do różnicowania we wszystkie rodzaje komórek i tka-nek, czyli wykazują pluripotencję. W przeciwieństwie do komórek ES ich pochodzenie nie budzi jednak kon-trowersji etycznych, ponieważ mogą one być uzyskane praktycznie z dowolnej komórki pochodzącej z doro-słego organizmu, np. fibroblastów pobranych ze skóry. Dodatkowo, potencjalne wykorzystanie komórek ES w medycynie regeneracyjnej związane jest z jeszcze jed-nym problemem, niewystępującym w przypadku komó-rek iPS. Liczba dostępnych rodzajów ludzkich komórek ES jest ograniczona. Oznacza to, że dla wielu pacjen-tów niemożliwe okazałoby się uzyskanie z komórek ES odpowiedniego, tj. zgodnego tkankowo, materiału do przeszczepu. Dla odmiany wyspecjalizowane komórki określonych tkanek i narządów można by potencjalnie uzyskiwać z komórek iPS otrzymanych w wyniku mo-dyfikacji komórek pochodzących od pacjenta. Innymi słowy, dawcą i biorcą materiału do przeszczepu byłaby ta sama osoba. W takiej sytuacji problem niezgodności

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 9
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
tkankowej nie występuje. Choć droga do wykorzystania komórek iPS w medycynie regeneracyjnej wydaje się jeszcze daleka, to warto pamiętać, że komórki te zostały uzyskane zaledwie kilka lat temu, a wiedza na ich temat już jest imponująca (Archacka i wsp., 2010). Jakie były początki badań nad komórkami iPS?
Reprogramowanie komórek to termin, który ozna-cza proces prowadzący do zmiany właściwości komórki w sposób, który nie zachodzi w warunkach fizjologicz-nych. Badania prowadzone przez różne zespoły badaw-cze, również Yamanakę i Takahashiego, miały na celu opracowanie metody, która pozwoliłaby na przekształ-cenie zróżnicowanych komórek w komórki o właści-wościach zbliżonych lub identycznych z komórkami ES. Japońscy badacze zdecydowali się na wprowadze-nie do zróżnicowanych komórek czynników, które są charakterystyczne dla komórek zarodka we wczesnych etapach rozwoju. W ich zamyśle taka strategia miała
doprowadzić do „zwiększenia” potencjału do różnico-wania zmodyfikowanych komórek. W pierwszym eta-pie badań naukowcy wybrali 24 geny, które umieścili w fibroblastach myszy. W wyniku tej modyfikacji fibro-blasty przekształciły się w komórki, które pod wzglę-dem morfologii i właściwości przypominały komórki ES. W kolejnych etapach badań Yamanaka i Takahashi stopniowo ograniczali liczbę wprowadzanych genów i ostatecznie udowodnili, że aktywność zaledwie czte-rech czynników wystarcza do tego, by fibroblasty prze-kształciły się w niezróżnicowane komórki pluripoten-cjalne. „Magiczny kwartet” tworzyły geny: Oct4, Sox2 i Klf4 (czynniki odgrywające kluczową rolę w zachowa-niu pluripotencji komórek) oraz c-Myc (gen promujący podziały komórkowe). Pierwsze ludzkie komórki iPS zostały uzyskane z fibroblastów rok później, w 2007 r. (Takahashi i wsp., 2007). Pionierskie prace Takahashie-go i Yamanaki otworzyły nowy rozdział w badaniach
nad komórkami macierzystymi i spowodowały lawino-wy wzrost zainteresowania „technologią iPS”. Doświad-czenia przeprowadzone przez różne grupy badawcze doprowadziły do uzyskania komórek iPS z rozmaitych rodzajów komórek, m.in. z keratynocytów (Maherali i wsp., 2008), hepatocytów (Aoi i wsp., 2008), komórek trzustki (Stadtfeld i wsp. 2008), limfocytów B (Hanna i wsp., 2008), melanocytów (Utikal i wsp., 2009) oraz komórek tkanki tłuszczowej (Sun i wsp., 2009). Zwy-kle największa wydajność reprogramowania osiągana jest wówczas, gdy geny wprowadzane są do komórek za pomocą wirusów. Jednak użycie wirusów w „technolo-gii iPS” jest równocześnie jej największym ogranicze-niem. Wirusy wykorzystywane do reprogramowania komórek wbudowują się bowiem do ich genomu, co może powodować uszkodzenie lub niekontrolowaną aktywację innych genów. Duże ryzyko wystąpienia nie-przewidzianych zmian na poziomie genomu ogranicza czy wręcz niweluje możliwość wykorzystania tak zmo-dyfikowanych komórek w medycynie regeneracyjnej. W niektórych przypadkach takie zmiany mogą bowiem prowadzić np. do niekontrolowanych podziałów komó-rek i, w konsekwencji, do powstania nowotworów po transplantacji komórek do organizmu pacjenta. Wirusy to nie jedyne ograniczenie „technologii iPS”. Naukowcy nie potrafią jeszcze uzyskiwać komórek iPS w sposób wydajny: proces ten zachodzi z wydajnością nie wyższą niż kilka procent. Równie mało skuteczne są metody uzyskiwania wybranych rodzajów wyspecjalizowanych komórek z komórek pluripotencjalych (Drews i wsp., 2012). Mimo tych ograniczeń komórki iPS już teraz zaj-mują ważne miejsce w nauce, są bowiem wykorzysty-wane m.in. jako modele in vitro różnych chorób. W tym celu od osób cierpiących na różne choroby pobierane są komórki, które następnie przekształcane są do komó-rek iPS, a te wykorzystywane są w badaniach mających na celu określenie molekularnego mechanizmu danej
Ryc. 4. Przykłady wykorzystania komórek iPS w nauce i medycynie
Pobranie komórek od pacjenta pozwala na uzyskanie – na drodze reprogramo-wania – komórek iPS. Komórki te można wykorzystać jako model in vitro choroby, którą dotknięty jest pacjent. Potencjal-nie, w przypadku, gdy pobrane komórki pochodzą od pacjenta cierpiącego na chorobę genetyczną można poddać je korekcie genetycznej, a następnie zróżni-cować w określony rodzaj komórek i prze-szczepić pacjentowi.

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 10
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
choroby czy wpływu potencjalnych leków. Przykładem mogą być badania Eberta i współpracowników, którzy uzyskali komórki iPS z fibroblastów pobranych od pa-cjenta cierpiącego na rdzeniowy zanik mięśni. Choroba ta spowodowana jest mutacją w genie SMN1 (ang. Survi-val Motor Neuron 1) i dotychczas pozostaje nieuleczal-na. Dzieci chore na rdzeniowy zanik mięśni najczęściej umierają w ciągu dwóch lat z powodu zaniku mięśni spowodowanego obumieraniem neuronów niezbęd-nych do ich właściwego funkcjonowania. W neuronach otrzymanych z komórek iPS uzyskanych od chorych na rdzeniowy zanik mięśni stwierdzono zmniejszoną ilość białka SMN1, koniecznego dla prawidłowego działania neuronów. Zbyt niski poziom tego białka w komórkach prowadził do ich degeneracji. Co ważniejsze jednak, badania przeprowadzone przez Eberta i współpra-cowników pozwoliły na wskazanie kilku czynników o potencjalnym znaczeniu terapeutycznym, np. kwasu walproinowego i tobramycyny. Ich obecność w pożyw-ce hodowlanej doprowadziła do zwiększenia poziomu białka SMN1 w neuronach (Ebert i wsp., 2009). Obecnie prowadzone są badania mające na celu określenie, w ja-kim stopniu czynniki te mogą chronić neurony przed degeneracją. Przykład ten pokazuje, że wykorzystanie „technologii iPS” może pomóc w poszukiwaniu sku-tecznego leku na tę i inne choroby degeneracyjne. Lawi-nowy postęp w badaniach nad komórkami iPS daje tak-że szansę na opracowanie skutecznych i bezpiecznych terapii różnych chorób z wykorzystaniem tych komórek (ryc. 4; Archacka i wsp., 2010).
Podsumowanie
Znaczenie komórek ES i iPS dla rozwoju nauki jest niepodważalne. Zdolność tych komórek do wielokie-runkowego różnicowania budzi również zrozumiałe nadzieje na ich wykorzystanie w medycynie regenera-
cyjnej. Jednak droga do rzeczywistego wykorzystania tych komórek w klinice jest jeszcze daleka. Alternaty-wą jest stosowanie terapii z wykorzystaniem komórek macierzystych pochodzących z organizmów dorosłych. O mocnych i słabych stronach takiego rozwiązania traktować będzie trzeci i ostatni już artykuł o komór-kach macierzystych – w numerze 3/2013 EBiŚ.
Literatura
Aoi T, Yae K, Nakagawa M, Ichisaka T, Okita K, Takahashi K, Chiba T, Yamanaka S (2008). Generation of pluripotent stem cells from adult mouse liver and stomach cells. Science 321, 699–702.
Archacka K, Grabowska I, Ciemerych MA (2010). Indukowane ko-mórki pluripotencjalne – nadzieje, obawy i perspektywy. Postępy Biologii Komórki. 37, 41-62.
Beetschen JC, Fischer JL (2004). Yves Delage (1854–1920) as a fore-runner of modern nuclear transfer experiments. Int. J. Dev. Biol. 48, 607–612.
Bishop J. (2001). Ssaki transgeniczne. Wydawnictwo Naukowe PWN, Warszawa.
Buehr M, Meek S, Blair K, Yang J, Ure J, Silva J, McLay R, Hall J, Ying QL, Smith A (2008). Capture of authentic embryo-nic stem cells from rat blastocysts. Cell 135(7), 1287-1298.
Capecchi MR (2005). Gene targeting in mice: functional analysis of the mammalian genome for the twenty-first century. Nature Re-views Genetics 6, 507-512.
Ciemerych MA (2008). Zarodkowe komórki macierzyste – w poszu-kiwaniu pluripotencji. Postępy Biologii Komórki 35, 183-205.
Ciemerych MA, Kubiak JZ (2013). Od Gurdona do Yamanaki, czyli krótka historia reprogramowania komórek. Postępy Biochemii 59 (2).
Czerwińska A, Ciemerych MA (2012). Kule zarodkowe jako me-toda różnicowania pluripotencjalnych komórek macierzystych w mioblasty. Postępy Biologii Komórki 39, 669-684.
Chung Y, Klimanskaya I, Becker S, Li T, Maserati M, Lu SJ, Zdravko-vic T, Ilic D, Genbacev O, Fisher S, Krtolica A, Lanza R (2008). Human embryonic stem cell lines generated without embryo de-struction. Cell Stem Cell 2(2), 113-117.
Darabi R, Santos FN, Filareto A, Pan W, Koene R, Rudnicki MA, Kyba M, Perlingeiro RC (2011). Assessment of the myogenic stem cell compartment following transplantation of Pax3/Pax7-indu-ced embryonicstem cell-derived progenitors. Stem Cells 29(5), 777-790.
Drews K, Jozefczuk J, Prigione A, Adjaye J (2012). Human indu-ced pluripotent stem cells -from mechanisms to clinical applica-tions. J Mol Med 90(7), 735-745.
Ebert AD, Yu J, Rose FF Jr, Mattis VB, Lorson CL, Thomson JA, Svendsen CN (2009). Induced pluripotent stem cells from a spinal muscular atrophy patient. Nature 457(7227), 277-280.
Evans MJ, Kaufman MH (1981). Establishment in culture of pluripo-tential cells from mouse embryos. Nature 292, 154-156.
Filimonow K, Krupa M, Suwińska A (2013). Pierwsze decyzje rozwo-jowe – różnicowanie komórek w przedimplantacyjnym zarodku myszy. Postępy Biochemii 59 (2).
Games D, Adams D, Alessandrini R, Barbour R, Berthelette P, Bla-ckwell C, Carr T, Clemens J, Donaldson T, Gillespie F et al. (1995). Alzheimer-type neuropathology in transgenic mice overexpress-ing V717F beta-amyloid precursor protein. Nature 373, 523-527.
Guan K, Rohwedel J, Wobus AM (1999). Embryonic stem cell diffe-rentiation models:
cardiogenesis, myogenesis, neurogenesis, epithelial and vascular smooth muscle cell differentiation in vitro. Cytotechnology 30, 211–226.
Gurdon JB (1962). The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J Embryol Exp Morphol. 10, 622-640.
Hanna J, Markoulaki S, Schorderet P, Carey BW, Beard C, Wernig M, Creyghton MP, Steine EJ, Cassady JP, Foreman R, Lengner CJ, Dausman JA, Jaenisch R (2008). Direct reprogramming of ter-minally differentiated mature B lymphocytes to pluripotency. Cel. 133(2), 250-264.
Li P, Tong C, Mehrian-Shai R, Jia L, Wu N, Yan Y, Maxson RE, Schul-ze EN, Song H, Hsieh CL, Pera MF, Ying QL (2008). Germline competent embryonic stem cells derived from rat blastocysts. Cell 135(7), 1299-1310.
Loh YH, Wu Q, Chew JL, Zhang W, Chen X, Bourque G, George J, Leong B, Liu J, Wong KY, Sung KW, Lee CW, Zhao XD, Chiu KP, Lipovich L, Kuznetsov VA, Robson P, Stanton LW, Wei CL, Ruan Y, Lim B, Ng HH (2006) The Oct4 and Nanog transcription ne-twork regulates
pluripotency in mouse embryonic stem cells. Nat Genet 38, 431-440.Maherali N, Ahfeldt T, Rigamonti A, Utikal J, Cowan C, Hochedlin-
ger K (2008). A high-efficiency system for the generation and stu-dy of human induced pluripotent stem cells. Cell Stem Cell 3(3), 340-345.
Makki N, Capecchi MR (2012) Cardiovascular defects in a mouse model of HOXA1 syndrome. Hum Mol Genet. 21(1), 26-31.
Martin GR. (1981). Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarci-noma stem cells. Proc Natl Acad Sci U S A. 78(12), 7634-7638.
Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi

Komórki macierzyste. Część II – pluripotencjalne komórki macierzyste | Barbara Świerczek, Damian Dudka, Karolina Archacka | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 11
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
K, Maruyama M, Maeda M, Yamanaka S. (2003). The homeopro-tein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 113(5), 631-642.
Shamblott MJ, Axelman J, Wang S, Bugg EM, Littlefield JW, Dono-van PJ, Blumenthal PD, Huggins GR, Gearhart JD (1998). Deri-vation of pluripotent stem cells from cultured human primordial germ cells. Proc Natl Acad Sci 95(23), 13726-13731.
Schwartz SD, Hubschman JP, Heilwell G, Franco-Cardenas V, Pan CK, Ostrick RM, Mickunas E, Gay R, Klimanskaya I, Lanza R (2012). Embryonic stem cell trials for macular degeneration: a preliminary report. Lancet 379, 713-720.
Stadtfeld M, Brennand K, Hochedlinger K (2008). Reprogramming of pancreatic β cells into induced pluripotent stem cells. Curr. Biol. 18, 890–894.
Sun N, Panetta NJ, Gupta DM, Wilson KD, Lee A, Jia F, Hu S, Cherry AM, Robbins RC, Longaker MT, Wu JC (2009). Feeder-free deri-vation of induced pluripotent stem cells from adult human adipo-se stem cells. Proc Natl Acad Sci 106(37), 15720-15725.
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131(5), 861-872.
Takahashi K, Yamanaka S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defi-ned factors. Cell 126(4), 663-676.
Tarkowski AK. (1961). Mouse chimaeras developed from fused eggs. Nature 190, 857-860.
Thomas KR, Capecchi MR. (1990) Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbra-in and cerebellar development. Nature 346, 847-850.
Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM (1998). Embryonic stem cell lines deri-ved from human blastocysts. Science 282(5391), 1145-1147.
Thomson JA, Kalishman J, Golos TG, Durning M, Harris CP, Becker RA, Hearn JP (1995). Isolation of a primate embryonic stem cell line. Proc Natl Acad Sci 92(17), 7844-7848.
Utikal J, Maherali N, Kulalert W, Hochedlinger K. (2009). Sox2 is dispensable for the reprogramming of melanocytes and melano-ma cells into induced pluripotent stem cells. J. Cell Sci. 122, 3502-3510.
Weismann A (1893). The Germ-Plasm: A Theory of Heredity. Walter Scott Ltd., Londyn.
Stem cells. Part II – pluripotent stem cells
Barbara Świerczek, Damian Dudka, Karolina Archacka
Pluripotent stem cells are able to differentiate into all types of cells and tissues buliding mammalian organism. During embryo development pluripotent cells are localized in the inner cell mass of blastocyst, which serves as a source of cells for embryo body formation. Pluripotent cells building inner cell mass can be isolated and propagated in vitro as embryonic stem cells (ES cells). Another type of pluripotent stem cells, i.e. induced pluripotent stem cells (iPS cells), is derived as a result of the process called cell reprogramming. Both ES cells and iPS cells serve as a valuable research tool. They could also be used as a potential source of cells for transplantation in regenerative medicine. Derivation of both ES and iPS cells is considered as a real breakthrough in science and medicine, and was distinguished by the Nobel Prize in physiology or medicine. In 2007 Mario Capecchi, Olivier Smithies, and Martin Evans were awarded for development of methods which enabled derivation of ES cells and genetic modi-fication of mice, while in 2012 John Gurdon and Shinya Yamanaka were awarded for research focused on cell reprogramming. Keywords: Pluripotent stem cells, ES cells, iPS cells, reprogramming, multidirectional differentiation, regenerative medicine, cell therapies

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 12
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Bioróżnorodność a ochrona patentowa Joanna Uchańska
Joanna Uchańska: doktorantka w Katedrze Prawa Własno-ści Intelektualnej Wydziału Prawa i Administracji Uniwersy-tetu Jagiellońskiego, aplikantka adwokacka w Krakowskiej Izbie Adwokackiej, członek międzynarodowego zespołu badawczego programu „Innovation Expert System”, uho-norowana nagrodą Ministra Rolnictwa i Rozwoju Wsi.
Streszczenie:
Bioróżnorodność stanowi niezgłębiony rezerwuar zaso-bów będących źródłem użytecznych w życiu człowieka rozwiązań. Stąd konieczna jest ochrona zasobów biolo-gicznych. Niniejszy artykuł prezentuje wybraną tematy-kę w przedmiocie sui generis ochrony bioróżnorodności ustanowionej w oparciu o przepisy Konwencji o różno-rodności biologicznej z 5 czerwca 1992 r. Tematyka ta została poddana analizie przez pryzmat instrumentu, jakim jest dostęp do zasobów genetycznych i równy po-dział korzyści (Access and Benefit Sharing – ABS). W tym zakresie zaprezentowano narzędzie uprzedniej zgody na dostęp do zasobów. Ponadto przeprowadzone obserwacje zostały pogłębione przez analizę relacji systemu ABS do prawa patentowego. Artykuł prezentuje dwie główne tezy w przedmiocie zrównoważonego wykorzystania bioróż-norodności oraz możliwej niezgodności między posta-nowieniami Konwencji o bioróżnorodności a prawem patentowym. Nade wszystko esej dotyka problematyki biopiractwa i wiedzy tradycyjnej.Słowa kluczowe: bioróżnorodność, dostęp do zasobów gene-tycznych i równy podział korzyści, biopiractwo, wiedza trady-cyjna, uprzednia zgoda
otrzymano: 1.04.2013; przyjęto: 22.04.2013; opublikowano: 28.06.2013
Wstęp
W 1994 r. Europejski Urząd Patentowy w Mona-chium (EPU) udzielił ochrony na metodę niszczenia grzybów na roślinach przez hydrofobowy ekstrakt z drzewa Neem (Azadirachta indica) (Nr EP 436257) na rzecz W.R. Grace & Company i Departamentu USA ds. Rolnictwa (Baharul, 2011). Rok później grupa rolników z Indii, organizacje pozarządowe oraz osoby fizyczne (m.in. Partia Zielonych przy Parlamencie Europejskim, dr Vandana Shiva, Indyjska Fundacja ds. Nauki, Tech-nologii i Ekologii, Międzynarodowa Fundacja ds. Ru-chu na rzecz rolnictwa ekologicznego) złożyły wniosek o unieważnienie tych patentów. Dowodzono w nim, że grzybobójcze właściwości ekstraktu z drzewa Neem były znane i dlatego wynalazek nie miał zdolności pa-tentowej. Wskazywano też, że drzewo Neem rośnie na terytorium południowej i południowo-wschodniej Azji, a ponadto uprawiane jest w Afryce, Ameryce Środko-wej, Karaibach, Hawajach i Arabii Saudyjskiej. Na tej podstawie w 1999 r. uznano, że rozwiązanie należało do ówczesnego stanu techniki i nie miało charakteru wynalazczego (nie posiadało tzw. poziomu wynalazcze-go). Patent został więc zrewidowany w 2000 r. (Alikhan i Mashelkar, 2004; Arihan i Gençler, 2007), zaś osta-tecznie unieważniony po orzeczeniu Izby Odwoławczej przy EPU dnia 8 marca 2005 r. (Baharul, 2011).
Kazus ten można traktować jako jedną z wielu spraw o unieważnienie patentu, w których uprawniony z pa-tentu dowodzi, że jego wynalazek jest nowy, posiada
poziom wynalazczy i charakteryzuje się przemysłową stosowalnością, zaś oponenci, wnioskujący o unieważ-nienie patentu, dowodzą, że wynalazek nie był paten-towalny, więc ochronę przyznano mu niesłusznie. Jed-nak postępowanie o unieważnienie patentu na metodę produkcji pestycydów z drzewa Neem było na tym tle szczególne. Poza argumentami, które faktycznie stały się ostatecznie podstawą wygaszenia patentu, wska-zywano bowiem, że naruszono przepisy międzynaro-dowego systemu ochrony bioróżnorodności1. System ten stanowi jeden z filarów dostępu do zasobów gene-tycznych i podziału płynących z nich korzyści (Access and Benefit Sharing – ABS), a także praw podmiotów uprawnionych do wiedzy tradycyjnej.
Czym jednak jest sama bioróżnorodność, system ABS, czy wiedza tradycyjna? Na czym, w opinii niektó-rych, polega łamanie prawa w sferze bioróżnorodności? Jak te zagadnienia wiążą się z prawem patentowym? Dlaczego system ABS w większości krajów nie wchodzi w skład przepisów będących podstawą przyznawanej ochrony patentowej? Na te oraz inne pytania staram się odpowiedzieć w niniejszym artykule, prezentując klu-czowe zagadnienia z zakresu sui generis ochrony bio-różnorodności.
Bioróżnorodność i jej ochrona sui generis
„Różnorodność biologiczna”, zgodnie art. 2 Kon-wencji o różnorodności biologicznej, sporządzonej w Rio de Janeiro dnia 5 czerwca 1992 r. (Convention on Biological Diversity – CBD), oznacza zróżnicowanie wszystkich żywych organizmów pochodzących m.in. z ekosystemów lądowych i morskich oraz innych wod-nych ekosystemów oraz zespołów ekologicznych, któ-1 Jest to system sui generis (łac. swego rodzaju), czyli system szcze-
gólny, niemający bezpośredniej podstawy w innych przepisach ani żad-nego odpowiednika i nie uzupełniany przez przepisy innych regulacji (w szczególności prawa patentowego).
Artykuł pomocny przy realizacji wymagań podstawy programowej.
Biologia – zakres rozszerzony: VIII. Różnorodność biologiczna Ziemi. 4. Uczeń przedstawia wpływ człowieka na różnorodność biologiczną, podaje przykłady tego wpływu (...).
Przyroda dla szkół ponadgimnazalnych: 15. Ochrona przyrody i śro-dowiska. 6. Uczeń określa cele zrównoważonego rozwoju i przedstawia zasady, którymi powinna kierować się gospodarka świata.

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 13
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
rych są one częścią. Dotyczy różnorodności w obrębie gatunku, pomiędzy gatunkami oraz pomiędzy ekosy-stemami (Wilson, 1992; Torrance, 2010). Zróżnicowa-nie biologicznych zasobów genetycznych jest różne na różnych obszarach Ziemi. Największe występuje w kli-macie równikowym, zwłaszcza na terytorium Brazylii, Boliwii, Peru, Kostaryki, Kongo, Indii; najmniejsze – na biegunach (Torrance, 2010; Koopman, 2003; Alikhan i Mashelkar, 2004).
Zgodnie z art. 2 CBD zasoby genetyczne to materiał genetyczny mający faktyczną lub potencjalną wartość. Z kolei materiał genetyczny to jakikolwiek materiał ro-ślinny, zwierzęcy, mikrobiologiczny lub innego pocho-dzenia zawierający funkcjonalne jednostki dziedzicz-ności. Zasoby genetyczne wchodzą zaś w skład zasobów biologicznych rozumianych jako zasoby genetyczne, or-ganizmy i ich części, populacje i jakiekolwiek inne żywe elementy ekosystemu, które faktycznie lub potencjalnie mogą być wykorzystywane lub mogą stanowić wartość dla ludzkości.
CBD wśród swych celów wymienia: ochronę bioróż-norodności, jej zrównoważone użytkowanie oraz uczci-wy i sprawiedliwy podział korzyści z wykorzystywania zasobów genetycznych, w tym przez odpowiedni dostęp do zasobów genetycznych i odpowiedni transfer właś-ciwych technologii, z uwzględnieniem wszystkich praw do tych zasobów i technologii, a także odpowiednie fi-nansowanie. Wytyczne te są realizacją Światowej Strate-gii Ochrony Przyrody, która zakłada m.in., że ochrona przyrody jest podejmowana przy udziale społeczności lokalnych. Jej celem jest budowanie odpowiednich po-staw oraz stworzenie warunków, w których ludność lokalna będzie mogła skorzystać z obszarów objętych ochroną (Jeffries, 2005). CBD jest uważana za pierwszy międzynarodowy akt, w którym zawarto zobowiązania państw do przestrzegania wymienionych postulatów. Konwencja nie jest wprawdzie kompleksową regulacją
ochrony przyrody, jednak wyznacza podstawę do ure-gulowania ochrony sui generis bioróżnorodności (Ko-opman J., 2003) i nakłada różne obowiązki na państwa, które ją podpisały. Wprowadza też takie instrumenty jak dostęp do zasobów genetycznych oraz transfer tech-nologii do krajów rozwijających się (Jeffries, 2007).
Jądrem postanowień CBD w przedmiocie ochrony bioróżnorodności jest dostęp do zasobów genetycznych. Wśród nich kluczowa jest regulacja dotycząca uprzed-niej zgody na dostęp do takich zasobów, do których – na mocy jej art. 3 w zw. z art. 15 – prawa przysługują pań-stwu, na terytorium którego zasoby te występują.
Regulacja o dostępie do zasobów genetycznych i po-dziale płynących z nich korzyści (wspomiane już Access and Benefit Sharing – ABS) związana jest z dostępem do wiedzy tradycyjnej, która towarzyszy bioróżnorodno-ści. Dr Thokozani Simelane (członek stałego Komitetu Doradczego ds. Prawa Własności intelektualnej RPA) zauważa, że CBD jest jedynym międzynarodowym ak-tem prawnym, który w tak szerokim stopniu przyznaje prawa do różnorodności biologicznej społeczności tu-bylczej i uznaje jurysdykcję krajów, z których pochodzą zasoby naturalne, w tym zasoby genetyczne (Arihan i Mine, 2007).
Eksploatacja wiedzy tradycyjnej związanej z bioróż-norodnością ma charakter pozytywny, gdy dotyczy po-szukiwania materiału biologicznego (materii ożywio-nej), modyfikowania genetycznego, opieki zdrowotnej opartej na produktach pochodzących z natury, medycy-ny naturalnej. Ma też jednak wymiar negatywny, zwią-zany z tzw. biopiractwem, o którym wspomnę w dalszej części. A zatem wiedza tradycyjna jest powiązana z bio-różnorodnością na zasadzie krzyżowania (Simelane). O ochronie bioróżnorodności i wiedzy tradycyjnej na-leży więc mówić przy zachowaniu następującego roz-graniczenia. Czym innym jest dbałość o różnorodność biologiczną jako taką, a czym innym jest zabezpieczenie
jej wraz z nawiązującą do niej wiedzą, w tym wiedzą tradycyjną, która stanowi też różnorodność kulturową (Brush, 2007). Wielu naukowców podnosi konieczność ochrony wiedzy tradycyjnej, w której zasobach można odnaleźć także recepty, instrumenty czy informacje na temat sposobów zachowania bioróżnorodności, w tym poprzez jej zrównoważone wykorzystanie czy ochronę przed zanieczyszczeniami (Zent i Zent Egleé, 2007).
Różnorodność biologiczną i kulturową traktuje się zatem obecnie jako całość (Morżoł, 2010). Podstaw tego stwierdzenia należy poszukiwać w art. 8 lit. j CBD, gdzie postanowiono, że „stosownie do swojego ustawo-dawstwa krajowego respektuje się, chroni i utrzymuje wiedzę, innowacje oraz praktyki stosowane przez tubyl-cze i lokalne społeczności, prowadzące tradycyjny tryb życia, sprzyjający ochronie i zrównoważonemu użytko-waniu różnorodności biologicznej oraz wspiera się ich szersze stosowanie za zgodą i przy udziale osób, które dysponują taką wiedzą, stosują innowacje i praktyki, oraz zachęca się do równego podziału korzyści płyną-cych z wykorzystania tej wiedzy, innowacji i praktyk”.
Czym jest jednak wiedza tradycyjna, którą chroni się wraz z bioróżnorodnością? Sama wiedza tradycyjna jest charakteryzowana jako pojęcie dynamiczne i zróż-nicowane (Mugabe, 2001), które interesuje się przeja-wami ludzkiej aktywności, takimi jak rolnictwo, nauki ścisłe, technika, ekologia, medycyna, a wreszcie wiedza powiązana z bioróżnorodnością, wyraz folkloru (muzy-ka, tańce, sztuka itd.), elementy języka (np. imion i na-zwisk), nazwy geograficzne, symbole, a nawet ruchome przedmioty własności (Alikhan i Mashelkar, 2004). Są to bowiem informacje przekazywane z pokolenia na pokolenie zwłaszcza w sposób ustny (chociaż może być spisana i nie wyklucza to jej tradycyjności). Charaktery-stycznym dla wiedzy tradycyjnej jest brak przypisania jej elementów indywidualnemu twórcy. Stanowi ona zatem anonimowy wynik kooperatywnych wysiłków

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 14
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
całej społeczności nie tylko żyjącej w konkretnej chwi-li, na konkretnym terytorium, ale ocenianej jako zbiór wielowiekowy, podporządkowany lokalnemu prawu, zwyczajom i tradycji. Dynamika rozwoju wiedzy wpły-wa na fakt, że „to co sprawia, że wiedza tradycyjna jest tradycyjna, to nie jej przeszły charakter, ale sposób jej przyswajania i użytkowania” (Mugabe, 2001).
Eksploatacja bioróżnorodności a jej ochrona
CBD przewiduje szereg działań, które wchodzą w zakres tzw. ochrony sui generis, która opiera się na trzech filarach: save – know – use. Pojęcia te oznaczają odpowiednio ochronę gatunków poprzez:
1) stworzenie obszarów chronionych,2) poznanie występujących gatunków chronionych
oraz3) używanie ich w sposób zrównoważony dla korzy-
ści społecznych i ekonomicznych (Gámez, 2007).Definicję zrównoważonego użytkowania ustanawia
art. 2 CBD i oznacza użytkowanie elementów różno-rodności biologicznej w taki sposób i z taką intensyw-nością, żeby nie prowadziło ono do jej zmniejszenia w długim czasie i tym samym pozwoliło utrzymać jej potencjał w stanie odpowiadającym potrzebom i aspi-racjom obecnych oraz przyszłych pokoleń. W jej zakres wchodzą: działania ogólne – takie jak przygotowanie krajowych strategii, planów, programów, a także szcze-gółowe – takie jak identyfikacja elementów różnorod-ności biologicznej i jej monitoring. CBD wymaga także opracowania systemu zrównoważonego użytkowania jako dostępu do jej zasobów genetycznych. W ten spo-sób proponuje się ochronę bezpośrednią bioróżnorod-ności.
Osoby dysponujące wiedzą tradycyjną związaną z bioróżnorodnością, szamani czy inni uzdrowicie-le (znachorzy), koncentrują się na widocznych gołym
okiem cechach organizmu. Wiele z nich jest dziedzicz-nych, jak np. kolor oczu, włosów, czy skłonność do otyłości (Winter i Fletcher, 2004); jak każda cecha, wi-doczna lub niewidoczna na pierwszy rzut oka, wywodzi się z konkretnego genotypu, ale w wiedzy tradycyjnej można było zwrócić uwagę tylko na te fenotypy, które dotyczyły zewnętrznych cech organizmu. Natomiast biotechnologia często skupia się na aspektach bioche-micznych, czyli tych fenotypach, których nie sposób określić na podstawie obserwacji z zewnątrz (m.in. aktywność określonych enzymów w wybranym narzą-dzie). Z tego powodu wiedza tradycyjna nie przyczynia się bezpośrednio do rozwoju biofarmacji. Niemniej jednak często jest pomocna w procesie prac badaw-czo-rozwojowych (research and development – R&D) w bioprzemyśle, ponieważ jej posiadacze znają medycz-ne zastosowanie konkretnych organizmów, co pozwala przystąpić do właściwych badań. Wiedza tradycyjna przydaje się więc w pierwszej fazie R&D – w fazie po-szukiwania. Jest też niezbędna na kolejnym etapie – owocnego wykorzystania organizmów lub ich kompo-nentów, a także na etapie ich praktycznej eksploatacji (Koopman, 2003), w tym w związku z patentowaniem.
Opracowanie strategii do identyfikacji metod gro-madzenia i stosowania materiału genetycznego oraz związanej z nim wiedzy nazwane zostało bioprospek-cją. Jest ona rozumiana jako „poszukiwanie gatunków roślin i zwierząt, z których mogą być uzyskiwane pro-dukty lecznicze lub inne związki cenne pod względem komercyjnym” (Compact Oxford English Dictionary, 2006). Kolejną zaletą wiedzy tradycyjnej jest to, że jest ona niezgłębionym i niewyczerpanym źródłem infor-macji o ochronie środowiska i jego zrównoważonym użytkowaniu. Podobnie R. Anriantsiferena (2007) pod-kreśla dwa główne elementy konstruujące pojęcie wie-dzy tradycyjnej, tj. wiedzę dotyczącą wszystkich aspek-tów życia i środowiska naturalnego.
Wiedza tradycyjna wymaga ochrony, ponieważ jednocześnie jest ona jednym z narzędzi podstawowej ochrony in situ (tzn. „w miejscu”) wyrażonej w art. 8 lit. j CBD. Ochrona in situ przewiduje szereg instru-mentów, mających na celu zachowanie gatunków występujących naturalnie w środowisku i całych eko-systemów. Oznacza więc ochronę ekosystemów i natu-ralnych siedlisk oraz utrzymanie i restytucję zdolnych do życia populacji gatunków w ich naturalnym środo-wisku, a w przypadku gatunków udomowionych lub hodowlanych – w środowisku, w którym rozwinęły swoje charakterystyczne właściwości. Pozwala też na wprowadzenie obszarów chronionych.
Pomocniczą funkcję w stosunku do ochrony in situ pełni ochrona ex situ. Należy ją rozumieć jako ochronę składników różnorodności biologicznej poza ich natu-ralnymi siedliskami (art. 2 CBD). Polega ona na bada-niu organizmów w celu przywracania naturalnych eko-systemów oraz gatunków zagrożonych wyginięciem. Wśród praktycznych działań realizujących założenia ochrony ex situ jest zakładanie ogrodów zoologicznych, a także prowadzenie banków genów. J. Chen (2007) uważa, że ochrona ex situ zachowuje jedynie niewielką część zasobów genetycznych, a kluczowa dla ochrony bioróżnorodności jest jednak ochrona in situ.
System stworzony w CBD został także uzupełnio-ny o dwa znaczące akty prawne: protokół z Kartageny (2000) oraz protokół z Nagoi (2010) – oba ratyfikowane przez Polskę. Postanowienia tego pierwszego dotyczą problematyki bezpieczeństwa biologicznego, zwłaszcza transferu pomiędzy państwami organizmów modyfi-kowanych genetycznie. Z kolei drugi dokument ustano-wił nową globalną strategię oraz instrumenty ochrony różnorodności biologicznej na lata 2011–2020 z wizją na rok 2050. Nowy Plan Strategiczny CBD określa 20 klu-czowych celów (tzw. Aichi Targets), które mają zostać osiągnięte do 2020 r. Przyjęty zaś Protokół z Nagoi ma

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 15
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
w pełniejszy i powszechniejszy sposób realizować po-stanowienia CBD o sprawiedliwym podziale korzyści wynikających z wykorzystania zasobów genetycznych.
Zasoby genetyczne w prawie patentowym
W literaturze wciąż uważa się, że ochrona zarówno bioróżnorodności, jak i wiedzy tradycyjnej w niektó-rych krajach rozwijających się jest wymierzona przede wszystkim w piractwo biologiczne oraz w narzucony, rygorystyczny system ochrony własności intelektualnej proponowany m.in. przez tzw. porozumienie TRIPS, czyli Porozumienie w sprawie handlowych aspektów praw własności intelektualnej z 15 kwietnia 1994 r. (Dz. Urz. WE L 336 z 23.12.1994 r., s. 305). Uchwale-nie CBD ma więc związek z istniejącym systemem pa-tentowania, zwłaszcza nowych związków chemicznych opartych na zasobach genetycznych wykorzystywa-nych w przemyśle farmaceutycznym. Zaproponowano zatem, by materiał roślinny odkryty przez duże firmy patentujące takie właśnie wynalazki biotechnologiczne, przynosił zyski nie tylko im, lecz także krajom, z któ-rych materiał ten pochodzi (Tritton i wsp., 2008). Na mocy CBD wprowadzono wspominany już specjalny reżim dostępu do zasobów oparty na równym podziale korzyści oraz przesłance uprzedniej zgody.
Zgodnie z CBD dysponentem zasobów genetycz-nych jest to państwo, na terytorium którego one wystę-pują. Takie stanowisko nie było jednak pierwotnie pre-zentowane. Początkowo uważano, że zasoby genetyczne są dobrem wspólnym. Rosnące znaczenie praw własno-ści intelektualnej do produktów z wykorzystaniem ro-ślin i tym samym komercyjne wykorzystanie zasobów genetycznych (Brush, 2007) konsekwentnie kierowało jednak kwestie związane z ochroną zasobów genetycz-nych w stronę koncepcji, jakie towarzyszą własności indywidualnej (Hamilton, 2006). Sygnatariuszy CBD
zobowiązano do ułatwienia innym podmiotom dostępu do swoich zasobów w sposób racjonalny z punktu wi-dzenia środowiska oraz do nie wprowadzania żadnych ograniczeń, które byłyby sprzeczne z CBD. Dostęp do zasobów genetycznych jest możliwy po uzyskaniu od-powiedniej zgody rządu oraz po zapewnieniu podziału korzyści uzyskanych z wykorzystania dóbr wytworzo-nych za pomocą lub z wykorzystaniem materiału gene-tycznego. Wymaganie zgody na dostęp do zasobów jest jednak fakultatywne, na podstawie art. 3 CBD, który przyznaje państwom suwerenne prawo do wykorzy-stania ich zasobów zgodnie z własną polityką środowi-skową. Pomimo fakultatywnego charakteru systemu regulowania dostępu do zasobów genetycznych w art. 15 ust. 7 CBD zobowiązano sygnatariuszy do podjęcia odpowiednich środków ustawodawczych, administra-cyjnych i politycznych oraz – w razie potrzeby – po-przez mechanizm finansowy mający na celu sprawied-liwy i równy dostęp do wyników badań i rozwoju oraz korzyści wynikających z komercyjnego lub innego wy-korzystania zasobów genetycznych z państwem dostar-czającym takie zasoby. Każdy sygnatariusz jest zobo-wiązany do ułatwienia dostępu i transferu technologii, które mają istotne znaczenie dla ochrony i zrównowa-żonego użytkowania różnorodności biologicznej lub wykorzystują zasoby genetyczne i nie powodują znacz-nych szkód dla środowiska. W myśl CBD sygnatariusze muszą także zapewniać ochronę patentom lub innym prawom własności intelektualnej, ponieważ wdrażanie postanowień konwencji odbywa się w zgodzie z usta-wodawstwem krajowym i prawem międzynarodowym, aby patenty oraz prawa własności intelektualnej wspie-rały cele konwencji i nie były z nimi sprzeczne.
Sprzeczność z prawem patentowym
Okazuje się, że mechanizmy dostępu do zaso-bów genetycznych wprowadzone przez wiele państw (np. Brazylii, Indiach, Kostaryce i Peru) są sprzeczne z Porozumieniem TRIPS, choć zgodne z CBD. Regu-lacje z zakresu prawa własności przemysłowej, w tym porozumienie TRIPS, chroni bowiem prawo własności uprawnionego, w tym wynalazcy. CBD skupia się zaś na ochronie zasobów genetycznych oraz wiedzy trady-cyjnej. Konflikt legislacyjny zachodzi w głównej mierze na poziomie umów międzynarodowych. Porozumienie TRIPS uniemożliwia pełną implementację CBD, gdyż oparte jest na mechanizmach własności prywatnej, podczas gdy CBD opowiada się za strukturą własności kolektywnej.
Z punktu widzenia porozumienia TRIPS przy ubie-ganiu się o ochronę patentową niewłaściwe będzie wy-maganie wykazania uzyskania uprzedniej zgody, cho-ciaż niektóre ustawodawstwa krajowe taką przesłankę wprowadziły. W Indiach uprzednią zgodę trzeba uzy-skać od krajowego organu ds. bioróżnorodności przed złożeniem aplikacji o patent. Sama CBD nie wskazuje, jaka jest sankcja dla patentu udzielonego bez uprzed-niej zgody (nieważność, bezskuteczność itp.). Mimo to niektóre państwa ustanawiają także odpowiednie normy karno-administracyjne w zakresie naruszenia postanowień chroniących zasoby genetyczne i zwią-zaną z nimi wiedzę tradycyjną. Niektóre zaś wiążą sankcje z nieważnością patentu. Takie postanowienia są sprzeczne z art. 27 porozumienia TRIPS, jeśli brak uzyskanej zgody ma stanowić przeszkodę w uzyskaniu patentu lub jego unieważnienie. Jedyne możliwe ogra-niczenia w przyznawaniu patentów, jakie dopuszczają postanowienia porozumienia TRIPS, zostały prze-widziane w ust. 2 i ust. 3 tego przepisu. Po pierwsze, zdolność patentowa może być ograniczona przez pań-

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 16
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
stwa ze względu na szczególne wyższe cele. Takie cele to np. ochrona porządku publicznego lub moralności, ochrona życia lub zdrowia ludzi, zwierząt lub roślin czy zapobieżenie poważnej szkodzie dla środowiska natu-ralnego. Do przypadku tego nie należy jednak ogra-niczenie wprowadzone tylko z tego powodu, że dany sposób wykorzystania jest zabroniony przez prawo kra-jowe. Po drugie, zdolności patentowej można odmówić diagnostycznym, terapeutycznym i chirurgicznym me-todom leczenia ludzi i zwierząt. Po trzecie zaś, państwo może nie udzielić patentu na rośliny i zwierzęta inne niż drobnoustroje oraz, zasadniczo, na biologiczne procesy służące do produkcji roślin i zwierząt inne niż procesy niebiologiczne i mikrobiologiczne. Żadne inne ograni-czenia nie są dopuszczalne na terytorium państw syg-natariuszy porozumienia TRIPS i dlatego dodatkowe postanowienia o uzyskaniu uprzedniej zgody na dostęp do zasobów genetycznych nie może być warunkiem uzyskania patentu.
Tak samo komentowano projekt nowelizacji prawa patentowego w Belgii, kiedy planowano implementować wymóg wykazania uprzedniej zgody wraz z sankcją nie-ważności patentu. Ostatecznie projekt został wycofany.
Międzynarodowe prawo patentowe nie zawiera żad-nych postanowień w sprawie ochrony zasobów gene-tycznych, ochrony wiedzy rdzennej i lokalnej społecz-ności. Wydaje się to niezrozumiałe, ponieważ Stronami CBD są 192 państwa oraz Unia Europejska, a więc re-guły w niej przyjęte można uznać bez wątpienia za po-wszechne. Mimo to system patentowy nie wprowadza żadnych postanowień w sprawie równego podziału ko-rzyści płynących z ochrony patentowej, jeśli wynalazek korzysta z zasobów genetycznych lub związanej z nimi wiedzy tradycyjnej (Anuradha i wsp., 1999). Mimo to przypadki uzyskania ochrony patentowej bez uprzed-niej zgody czy chociażby podziału korzyści płynących z ochrony patentowej spotykają się ze sprzeciwem
różnych społeczności. Oskarżenia zostały wysunięte np. przeciwko amerykańskiemu koncernowi biotech-nologicznemu Phytera. W 1996 r. koncern ten podpisał umowę z kilkoma europejskimi ogrodami botaniczny-mi. Niniejsza umowa zapewniała Phytera’i dostęp do gatunków pochodzących z ich kolekcji roślin, zaś w za-mian miała gwarantować podział korzyści płynących z wytworzonych na tej podstawie produktów. Jednak niektóre z umów pomijały klauzulę obowiązku podzia-łu korzyści z krajami, z których gatunki roślin pocho-dziły (Jeffries, 2004). Jak już wyżej wykazywano, obo-wiązek ten wynika z art. 15 CBD, który mówi o ABS.
Próby rozwiązania
Aby rozwiązać konflikt TRIPS – CBD, podjęto prace w ramach Światowej Organizacji Własności Intelektu-alnej przy ONZ (WIPO) we współpracy z Sekretaria-tem CBD, FAO i Agencji ONZ ds. Środowiska (UNEP). Postanowienie paragrafu 19 Deklaracji z Doha stanowi, że Rada TRIPS powinna zauważyć zagadnienia relacji porozumienia TRIPS do CBD oraz ochrony wiedzy tra-dycyjnej i folkloru. Ponadto w 2000 r. utworzono Mię-dzynarodowy Komitet ds. Własności Intelektualnej, Zasobów Genetycznych, Wiedzy Tradycyjnej i Folklo-ru (IGC), którego przedmiotem są: zagadnienia włas-ności intelektualnej w kontekście dostępu do zasobów genetycznych i podziału korzyści oraz ochrona wiedzy tradycyjnej, która może, ale nie musi być związana z za-sobami genetycznymi. Prace prowadzone w ramach komitetu IGC skupiają się na trzech zasadniczych kwe-stiach:• na zapewnieniu defensywnej ochrony zasobów
genetycznych poprzez eliminowanie udzielania patentów, którym brak nowości oraz poziomu wy-nalazczego;
• na wyborze właściwego modelu oraz instrumen-tów zapewniających realizację równego i spra-wiedliwego systemu ABS;
• na wprowadzeniu wymogu ujawnienia w zgłosze-niach patentowych pochodzenia materiału gene-tycznego lub źródła zasobów genetycznych (Lisow-ska, 2012).
Obecnie trwają prace mające na celu osiągnięcie po-rozumienia w zakresie harmonizacji przepisów prawa patentowego i CBD. Choć wydają się obiecujące, nie przyniosą szybkich rezultatów, ponieważ różne pań-stwa prezentują w tym zakresie rozbieżne stanowiska:• UE i Szwajcaria opowiadają się za wprowadzeniem
wymogu ujawnienia, którego naruszenie nie ma wywierać sankcji w prawie patentowym;
• kraje rozwijające się chcą, aby naruszenia wpro-wadzonego wymogu ujawnienia pociągało za sobą sankcje przy udzieleniu patentu;
• USA, Japonia, Korea i Kanada oprotestowują wy-móg ujawnienia (Lisowska, 2012) i optują za po-zostawieniem go poza systemem patentowym (Straus, 2008).
Krytyczna recenzja ABS
Dotychczasowy system ABS nie jest jednak wolny od luk i głosów krytycznych na jego temat. Po pierwsze, wprawdzie CBD opowiada się za przyznaniem prawa do dysponowania i decydowania o zasobach genetycz-nych państwu, gdzie te zasoby występują, jednak liczna grupa przedstawicieli rządów, naukowców oraz organi-zacji społecznych opowiada się za wolnym dostępem do różnorodności biologicznej, a nie jej uwłaszczeniem na czyjąkolwiek rzecz, która zagraża w ich opinii ochronie środowiska naturalnego. Problem dotyczący zwłaszcza zasobów mórz i oceanów (poza wodami terytorialnymi) czy zasobów Arktyki. Obszary te nie są bowiem objęte

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 17
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
jurysdykcją żadnego państwa. Toteż proponuje się trzy rozwiązania:
1) traktowanie zasobów jako dziedzictwo ludzkości,2) oparcie dostępu do tych zasobów na zasadzie wol-
ności przysługującej wszystkim zainteresowanym,3) „uznanie zasobów za wspólną własność ludzkości,
gdzie element «własność» będzie zobowiązywał do ustalenia zasad korzystania z niej w ramach od-rębnie ustalonego reżimu prawno międzynarodo-wego” (Szkarłat, 2012; Zimny, 2012). Takie zasoby mogą być dowolnie wykorzystywane w dziedzinie medycyny, żywienia, farmacji, przemysłu tekstyl-nego itp.
Po drugie, problem może wywoływać ustalanie, ja-kie konkretne korzyści powinny być przedmiotem po-działu oraz kto powinien być ich odbiorcą – czy kraj, z którego dany materiał pochodzi, czy społeczność lo-kalna, która ma wiedzę związaną z tym materiałem ge-netycznym.
Po trzecie, istnieją trudności z określeniem pod-miotu praw i obowiązków, gdy materiał genetyczny występuje na większej części kontynentu czy globu oraz szerokiego rozpowszechnienia wiedzy na jego temat. Komplikacje mogą być związane z ustaleniem sposobu podziału korzyści.
Po czwarte, stronami CBD pozostają państwa po-chodzące z różnych systemów prawnych, w których w różny sposób uregulowane jest prawo własności, w tym współwłasność. Ponadto nie postanowiono wy-raźnie, jak należy ustalać proporcje podziału. Często podnosi się, że posiadaczom zasobów biologicznych i wiedzy na ich temat zostają przekazane nieproporcjo-nalnie niewielkie części należnych im korzyści.
Po piąte, bioprospekcja towarzysząca systemowi ochrony bioróżnorodności przynosi w większości ko-rzyści niepieniężne, chociaż jest to ocena jedynie semi--negatywna, gdyż można oceniać pochlebnie jej wpływ
dla zbadania różnorodności genetycznej na świecie i określenia stopnia zagrożenia jej utraty.
Propozycja rozwiązania konfliktu
Proponowanym rozwiązaniem konfliktu: prawo pa-tentowe – ABS jest m.in. wprowadzenie dodatkowych przepisów do porozumienia TRIPS. Wnioskuje się za-tem uchwalenie art. 29bis w przedmiocie wymagań ujawnienia źródła pochodzenia zasobów biologicznych lub wiedzy tradycyjnej. Projekt nowej regulacji zakłada nie tylko konieczność poinformowania o źródle pocho-dzenia materiału genetycznego, uzyskaniu uprzedniej zgody. Stanowi także podstawę do wprowadzenia sank-cji zawieszenia postępowania patentowego lub nieprzy-znania prawa w sytuacji uzasadnionych wątpliwości co do spełniania przez wnioskodawcę wymagań o uprzed-niej zgodzie (Strus, 2008). Dotychczas bowiem w po-stępowaniu o unieważnienie patentu można podnieść tylko przesłanki braku zdolności patentowej: braku no-wości, nieoczywistości czy przemysłowej stosowalności.
Tak stało się np. w przypadku opatentowania w 1995 r. (Nr 540504) przez Urząd Patentowy Stanów Zjednoczonych (USPTO) metody leczenia ran za po-mocą wyciągu pochodzącego z korzenia szafranu indyj-skiego (Alikhan i Mashelkhar, 2004). Szafran indyjski był od wieków wykorzystywany w kuchni i kosmetyce azjatyckiej, w medycynie zaś jako lek m.in. na anemię, astmę, oparzenia, zapalenie spojówek, problemy stoma-tologiczne, cukrzycę, biegunkę, zwalczanie bólu (Ari-han i Mine, 2007). Patent został unieważniony 20 listo-pada 1997 r. ze względu na brak nowości, po działaniach podjętych przez Urząd Indii ds. Nauki i Przemysłu (The Council of Scientific and Industrail Research in India). Skarga na nieważność została poparta dokumentacją pochodzącą z 32 źródeł, w zakresie wiedzy tradycyj-nej, w skład której wchodziła m.in. publikacja z 1953 r.
w „The Journal of the Indian Medical Association”, inne zaś zostały spisane w lokalnych językach, takich jak sanskrit, urdu lub hindi. (Alikhan i Mashelkar, 2004). Tym samym patent został unieważniony na podstawie art. 102 i art. 103 U.S. Patent Act (Baharul, 2011).
Nie tylko jednak zmiany przepisów międzynaro-dowych mogą chronić przed łamaniem postanowień CBD. Rząd Indii podjął pracę nad stworzeniem krajo-wej Cyfrowej Biblioteki Wiedzy Tradycyjnej (TKDL). Ponadto WIPO prowadzi równoległe prace nad stwo-rzeniem i wprowadzeniem instrumentów, które mają na celu pozwolić ekspertom patentowym do wzięcia pod uwagę zasobów wiedzy tradycyjnej przy badaniu nowości i nieoczywistości. Stąd opracowuje się zaso-by światowej TKDL, która w przyszłości miałaby być zintegrowana z Cyfrową Biblioteką Własności Inte-lektualnej. Pierwsza z baz danych miałaby dostarczać zwięzłe, skategoryzowane i standaryzowane informacje na temat istniejącej wiedzy tradycyjnej. Niewątpliwie pozwoli to na wzięcie pod uwagę istniejącej wiedzy tra-dycyjnej przy prowadzeniu badań patentowych (Wyn-berg, 2003). TKDL w ramach WIPO miałaby stanowić system ogólnoświatowy. W ramach projektu, którego celem jest stworzenia TKDL dla indyjskiego systemu medycznego, pracuje interdyscyplinarny zespół składa-jący się z ekspertów ds. medycyny naturalnej, ekspertów patentowych, specjalistów ds. IT, naukowców i kadry technicznej. Biblioteka będzie dostępna w pięciu języ-kach: angielskim, niemieckim, francuskim, japońskim i hiszpańskim. TKDL zawiera już ponad 34 milionów stron i zapewnia informację na temat wiedzy tradycyj-nej, by chronić przed błędnym przyznawaniem paten-tów. (http://www.wipo.int/pressroom/en/articles/2011/article_0008.html). Dostęp do tej bazy został zagwa-rantowany dla wielu urzędów patentowych, w tym EPU i USPTO w 2009 r.

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 18
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Dodatkowe wymogi oceny zdolności patentowej, w rzeczywistości przewidziane jedynie w regulacjach niektórych państw, są jednak niewystarczające przy ochronie przed korzystaniem z zasobów genetycznych danego państwa bez uzyskania uprzedniej zgody. Otóż w wielu przypadkach prowadzonych badań R&D uzy-skane próbki są wywożone do krajów, z których nie pochodziły. Wówczas rozwiązania, które na nich ba-zują, spełnią inne przesłanki zdolności patentowej – uzyskują patent. Podobnie było np. w przypadku ryżu Basmanti. 2 września 1997 r. opatentowano ów ryż w USA (nr US 5663484). Patent obejmował 20 zastrze-żeń, wśród których znalazł się nie tylko ryż (znaczą-co podobny do indyjskiego ryżu Basmanti), lecz także znaczna liczba odmian ryżu, a także metod otrzymy-wania roślin i ziaren. Zastrzeżenia patentowe nr 15–17 stanowiły ziarna zbliżone cechami do ryżu Basmanti. Zastrzeżenia te stały się główną przeszkodą eksportu ryżu Basmantii z Indii do USA. Ryż Basmantii upra-wiany tradycyjnie w prowincji Punjab rośnie u zboczy Himalajów. Z uwagi na swą wyjątkową jakość, specjal-ny aromat i smak, a także długie ziarna, był sprzeda-wany na całym świecie. W 1993 r. przyniósł najwyższe zyski. Indie zaś, po Chinach, są (nadal) największym eksporterem ryżu na świecie (FAO). Ryż wykorzysty-wany jest w Indiach także przy modlitwach i cere-moniach religijnych, a więc ma znaczenie kulturowe. Urząd Indii ds. Badań Rolniczych (IARI) w New Delhi wystosował wniosek o unieważnienie patentu, podno-sząc, że ziarna ryżu wchodzą od 1978 r. w skład kolek-cji Wydziału ds. Badań nad Ryżem. Ziarna ryżu zaś będące przedmiotem zastrzeżeń 15–17 zostały prze-badane przez Centralny Instytut Badawczy ds. Tech-nologii Żywności. Ze względu na przeprowadzone po-nowne badanie patentowe zgłaszający (Rice Tech. Inc.) wycofał rozwiązanie w zakresie zastrzeżeń 15–17 oraz 4 (Alikhan i Mashelkar, 2004). Nazwa patentu zosta-
Legislacja w zakresie ochrony zasobów genetycz-nych jest obecnie przedmiotem dyskusji na poziomie krajowym. Jak wynika z zaprezentowanych przykła-dów, ochrona stanowi kombinację możności eksplo-atacji dóbr oraz podziału korzyści. Kraje, z których pochodzą dane zasoby naturalne są zobligowane do stworzenia regulacji w trzech sferach, a mianowicie do stworzenia przepisów na temat:
1) kolekcji genetycznych, przy zachowaniu dostępno-ści informacji oraz współpracy z lokalną ludnoś-cią,
2) transferu technologii oraz podziału korzyści, a także
3) utworzenia organów regulacyjnych odpowiedzial-nych za przestrzeganie przewidzianych regulacji. Dodatkowo, aby przepisy były skuteczne wskazuje się na konieczności ustanowienia komplementar-nych regulacji w państwach czerpiących z niniej-szych zasobów celem właściwej egzekucji praw
(Jeffries, 2004).Kompleksowa regulacja ABS nie jest jednak wy-
magana przez CBD, ale dopiero przez Protokół z Na-goi, który wejdzie w życie po 90 dniach od złożenia 50. dokumentu ratyfikacyjnego. Polska podpisała Pro-tokół z Nagoi, ale jeszcze go nie ratyfikowała. Protokół z Nagoi w szczególności wymaga, by rdzenna lub lokal-na społeczność dysponująca zasobami genetycznymi towarzyszyła w podziale korzyści. Nie postanowiono w nim jednak, aby społeczność ta miała być stroną umów czy decyzji lub innych aktów w tym zakresie. Społeczność lokalna lub rdzenna powinna być jednak zaangażowana w mechanizm przekazywania dostępu do zasobów genetycznych. Otóż w myśl art. 16 w zw. z art. 6 ust. 2 bierze ona udział w wyrażeniu zgody lub zatwierdzenia dostępu do zasobów genetycznych. Na podstawie samego porozumienia nie powiedziano tak-
ła zmieniona z „Odmiany i ziarna ryżu Basmanti” na „Odmiany Ryżu Bas867, RT117 i RT1121”. Zgłaszający zaprzeczył także, aby ziarna ryżu zostały wyizolowane ze środowiska naturalnego w Indiach czy Pakistanie, twierdząc, że są przedmiotem transformacji linii gene-tycznej ryżu. Ziarna poddane transformacji genetycz-nej miały pochodzić ze Światowej Kolekcji Materiału Genetycznego w Aberdeen (Idahoo), eksploatowanej przez Serwis Badań Rolniczych przy Depertamencie ds. Rolnictwa. Zanim trafiły do kolekcji miały być także przedmiotem wcześniejszej 10-letniej tradycyjnej i klasycznej uprawy. Mimo to argumentacja ta budzi uzasadnione wątpliwości i może świadczyć o tym, że ryż był łudząco podobny do tego uprawianego w In-diach i Pakistanie (Baharul, 2011). Na marginesie nale-ży wskazać, że Rice Tec. Inc. przed uzyskaniem paten-tu starał się wprowadzić na rynki europejskie ryż pod nazwą „Texmati”, „Jasmati”, „Kasmati”. Po uzyskaniu zaś patentu Rice Tec. postanowił wprowadzić na ry-nek ryż o nazwie „Basmanti”. Jednak 9 lipca 1998 r. Federacja USA ds. Ryżu wydała komunikat, w którym zaznaczyła, że nazwa „basmanti” odnosi się do wielu klas i gatunków aromatycznego ryżu. W konsekwen-cji, nazwa ta nie charakteryzuje produktu lub od-miany pochodzącej z danego kraju lub grupy krajów. Krytycznie w literaturze wypowiedziano się także na temat wniesienia przez Indię wniosku jedynie o re-wizję niektórych zastrzeżeń patentowych. Rząd Indii nie podniósł bowiem argumentów dotyczących na-ruszenia bioróżnorodności oraz wiedzy z nią powią-zanej, w tym praw rolników i tym samym nie wnosił o unieważnienie pozostałych zastrzeżeń patentowych
(Baharul, 2011, 42, 45-47), co jednak było poprawne, ponieważ takie argumenty nie mogły być podstawą unieważnienia patentu, jeśli nie podważały nowości, nieoczywistości lub przemysłowej stosowalności.

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 19
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
że, czy brak zgody rodzi konsekwencje dla uzyskania praw własności intelektualnej.
Unia Europejska jeszcze przed szczytem w Nagoi podkreśliła konieczność określenia zgodności protoko-łu z zasadą uprzedniej świadomej zgody społeczności rdzennych i lokalnych na skorzystanie z dostępu do ich tradycyjnej wiedzy związanej z zasobami genetycznymi Ponadto wśród pilnych potrzeb działania wyszczegól-niono te w zakresie zaniepokojenia tym, że cały czas ist-nieje możliwość nielegalnego dostępu do zasobów gene-tycznych. W projekcie rezolucji z 2 lipca 2010 r. złożony na 10. posiedzeniu Konferencji Stron Konwencji ONZ w Nagoi oceniono system uprzedniej zgody jako narzę-dzie, które pozwoli zahamować ów nielegalny dostęp.
Wydaje się, że takie poglądy są odosobnione albo pozbawione woli politycznej. Taka możliwość istnieje bowiem, jak zaznaczono wyżej, na podstawie CBD. UE i jej państwa członkowskie korzystają z tej możliwości fakultatywnie w sposób nieco odmienny oraz bardzo ograniczony. Otóż zgodnie z motywem 27 zd. 1 dyrek-tywy 44/98/WE2, „jeżeli wynalazek oparty jest na ma-teriale biologicznym pochodzenia roślinnego lub zwie-rzęcego lub jeśli wykorzystuje taki materiał, zgłoszenie patentowe powinno, tam gdzie to stosowne, zawierać informację o geograficznym pochodzeniu takiego ma-teriału, jeśli jest ono znane”. Przepisy te są jednak po-zbawione sankcji, gdyż nie mają wpływu na udzielenie prawa ochronnego. Nie określono w nich także wymo-gu przedstawienia potwierdzenia realizacji procedu-ry uzyskania zgody na dostęp do zasobów. Realizację postanowienia wyrażonego w dyrektywie 44/98/WE na poziomie krajowym wprowadziły zaś jedynie Da-nia i Belgia. W Danii przewidziano wymóg informacji o geograficznym pochodzeniu materiału, jeśli zgłasza-
2 Dyrektywa Parlamentu Europejskiego i Rady 98/44/WE z dnia 6 lipca 1998 r. w sprawie ochrony prawnej wynalazków biotechnologicznych (Dz. Urz. WE L 213 z 1998 r., s. 13).
jący posiada taką wiedzę, a jeśli nie posiada, to powi-nien to zaznaczyć. Brak informacji na ten temat nie ma jednak wpływu na postępowanie o nadanie patentu.
(van Overwalle, 2007).Sprawa „Basmanti” pokazuje, że wymogi uzyskania
uprzedniej zgody i jej wpływu na udzielenie praw włas-ności intelektualnej związane są z jurysdykcją państwa, z którego pochodzi dany materiał genetyczny. Dlate-go też nie ma przeszkód, aby uzyskać prawo wyłączne np. na terytorium UE bez uzyskania zgody na dostęp do zasobów genetycznych Brazylii, Indii, Kostaryki czy Peru. Nie ma także konieczności, np. w UE, aby po-dawać, skąd materiał genetyczny pochodzi, ponieważ przewidziana norma motywu 27 dyrektywy 98/44/WE jest normą lex imperfecta (a więc niepodanie źródła po-chodzenia materiału nie jest sankcjonowane) i taki też ma charakter w państwach członkowskich UE, które ów wymóg wprowadziły.
Reasumując, wśród państw-stron CBD istnieje kon-sensus, że powinny zostać osiągnięte następujące cele:• dostęp do zasobów genetycznych powinien być
wynikiem autoryzacji, tj. wyrażenia uprzedniej zgody przez uprawniony podmiot;
• równy podział korzyści płynących z wykorzysta-nia zasobów genetycznych i związanej z nimi wie-dzy tradycyjnej jest konieczny;
• wymagane jest przeciwdziałanie błędnemu wyda-waniu patentów.
Biopiractwo
Chociaż cele CBD zostały określone, nadal nie usta-lono ostatecznie, jak je realizować (Straus, 2008). Brak porozumienia generuje m.in. zjawisko biopiractwa. Po-jęcie to zostało po raz pierwszy użyte na początku lat 90. XX wieku. Obecnie biopiractwo definiuje się m.in. jako:
przywłaszczenie wiedzy i zasobów genetycznych pocho-dzących z upraw oraz od społeczności lokalnych przez oso-by lub instytucje poszukujące wyłącznej monopolistycznej kontroli (patentowej lub z tytułu innych praw własności intelektualnej) wśród tej wiedzy i zasobów (Drahos i Brait-hwaite, 2002; Hamilton, 2003).
Dla prawa patentowego jest nieistotne, w jaki spo-sób doszło do zdobycia danego materiału genetycznego. Dla ekspertów rozpatrujących zgłoszenie patentowe nie jest zaś możliwe dotarcie do odpowiednich informacji o znanej wiedzy tradycyjnej, co odbierałoby zgłoszeniu przymiot nowości lub nieoczywistości. Eksperci mają także problem z rozpoznaniem elementów wiedzy tra-dycyjnej w zgłoszeniu (Koopman, 2003). Może to wy-nikać z różnych systemów zapisu i stylu języka, jakimi posługują się ich autorzy. Język zgłoszenia patentowego ma często charakter techniczny, zgłoszenie jest zazwy-czaj rozbudowane i stanowi sumę różnych elementów. Jeśli miałaby pojawić się w nim wiedza tradycyjna, to byłaby ona sprecyzowana w najmniej bezpośredni spo-sób, zupełnie odbiegający od jego naturalnej, potocznej formy.
Winę za stałe istnienie biopiractwa ponosi także po-rozumienie TRIPS. Narzuca ono bowiem ścisły rygor ochrony własności intelektualnej, co z kolei prowadzi do wyeliminowania z konkurencji w dziedzinie inno-wacyjności krajów rozwijających się, z uwagi na ich techniczny i ekonomiczny status.
Wszystkie dotychczas opisane przypadki prezen-tują problematykę biopiractwa. Znanych jest wiele, wiele innych. Przykładowo przed EPU zarejestrowano siedem patentów, które wykorzystują ekstrakt z rośli-ny zwanej potocznie Maca (Lepidium meyenii). Jest to roślina występująca na terenach Andów i dlatego jest bardzo odporna na ekstremalne warunki pogodowe. Może być spożywana jako napój lub w wersji sprosz-kowanej. Od wieków stosuje się ją jako suplement diety

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 20
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
o właściwościach wzmacniających, w tym skutecznie podnoszącym płodność. Jeden z patentów został przy-znany 26 lutego 2004 r. na rzecz Pureworld Botani-cals (nr WO 2004/016216, Patent europejski przyzna-ny 8 czerwca 2005 r., nr EP 1536787 (A2); patent USA nr 2004034079) na alkaloidy imidazolu pochodzące z Lepidium Meyenii oraz sposób ich wykorzystania. W literaturze zakwestionowano jednak ważność oma-wianego patentu. Podniesiono, że wynalazek nie charak-teryzuje się ani nowością, ani nieoczywistością. Nowość była kwestionowana z uwagi na występowanie rośliny w przyrodzie przy jednoczesnym wykorzystaniu jej od wieków przez ludność Andów. Nieoczywistość kwestio-nował zaś prof. Carlos Cuirós z Uniwersytetu Kalifornii. Wskazał on, że produkt będący przedmiotem patentu niewiele różni się od tego występującego w przyrodzie. Usunięto bowiem z ekstraktu występującą naturalnie celulozę, co – zdaniem tego autora – ma jedynie znacze-nie estetyczne (Argumedo i Pimbert, 2006).
Innym przykładem jest patent na rzekomą nową odmianę rośliny Banisteriopsis caapi (B. caapi) o na-zwie „Da Vine”, przyznany obywatelowi USA Lore-nowi Milerowi 17 czerwca 1986 r. (Nr US5751)3. Kora drzewa B. caapi jest wykorzystywana od wieków przez ludy żyjące w Amazonii do wytwarzania napoju ayahu-asca. Zebrano ją w lasach Amazonii i z uwagi na od-mienny kolor kwiatów uznano, że to nowy gatunek. Opatentowana roślina zmieniała kolor wraz z wiekiem, od różowego do białego, zaś naturalnie występująca w przyrodzie – od różowego do jasnożółtego. Organ Koordynujący Rdzenne Organizacje Dorzecza Ama-zonki (The Coordinating Body of Indigenous Orga-nisations of Amazon Bine – COICA) wniósł wniosek o unieważnienie patentu, argumentując, że roślina była
3 Zob. stronę: http://patft.uspto.gov/netacgi/nph-Parser?Sect1= PTO2&Sect2=HITOFF&u=/netaht ml/search -adv.htm&r=2&p=1&f=G&l=50&d=ptxt&S1=caapi&OS=caapi&RS=caapi.
znana i wykorzystywana w medycynie przez rdzennych mieszkańców dorzecza Amazonki. USPTO unieważnił patent 2 listopada 1999 r. Jednak 17 kwietnia 2001 r. poprzednia decyzja USPTO została podważona i tym samym patent pozostaje w mocy (Alikhan i Meshelkar, 2005).
W orzecznictwie oraz doktrynie formułuje się kilka poglądów na temat potencjalnych form zapobiegania zjawisku biopiractwa. Wyróżnia się wśród nich m.in.:
1) zachowanie status quo i przyjęcie, że przepisy własności intelektualnej, mimo że nieskutecz-ne, powinny pozostać w swym dotychczasowym kształcie;
2) sformułowanie skuteczniejszych przepisów o po-dziale korzyści;
3) uregulowanie kwestii związanych z pojęciem włas-ności, w tym sformułowanie jednoznacznej tezy o zakazie patentowania życia (Koopman, 2003).
W świetle powyższych rozważań należy podnieść co następuje. W doktrynie dominuje stanowisko, że przy-jęty kształt praw własności intelektualnej nie zaspokaja potrzeb walki z biopiractwem. Wysuwa się także tezę o rozpatrzeniu różnych koncepcji własności, sposo-bów podziału dóbr, a także podjęciu decyzji, gdzie leży granica między wynalazkiem a odkryciem, a więc pro-duktem występującym naturalnie w przyrodzie (Posey, 1996).
Rozważania końcowe
Bioróżnorodność to niezgłębione zasoby będące źródłem użytecznych w życiu człowieka rozwiązań. Czerpie z niej biotechnologia, która od kilkudziesięciu lat jest jedną z najprężniej rozwijających się gałęzi na-uki i przemysłu. Zasoby genetyczne są przedmiotem ju-rysdykcji państwa na terytorium, którego się znajdują. Powszechnie zrezygnowano z koncepcji wolnego i do-
wolnego dostępu do zasobów genetycznych. Uznano, że – po pierwsze – ze względu na konieczność ochrony zasobów dostęp do nich powinien być regulowany, a po drugie – bogactwa naturalne nieożywione (węgiel, rudy metali, gaz, ropa itp.) i ożywione (zasoby genetyczne) podlegają jurysdykcji państwa, na terenie którego wy-stępują.
Potencjał bioróżnorodności powinien być wyko-rzystywany w sposób zrównoważony oraz w granicach prawa. Jednak w świetle przedstawionych rozważań można postawić dwie tezy:
1) zrównoważone wykorzystanie bioróżnorodności może stać w opozycji do innych dóbr prawnych, chronionych w świetle prawa międzynarodowego;
2) różne regulacje prawa międzynarodowego są ze sobą niespójne i dlatego przestrzeganie posta-nowień jednej z umów międzynarodowych nie pozwala na przestrzeganie postanowień innej umowy. Tak jest w przypadku niezgodności posta-nowień CBD i porozumienia TRIPS.
W opinii niektórych ochrona patentowa jest sprzecz-na z przepisami prawa i niesprawiedliwa, jeśli dotyczy ochrony leków i żywych organizmów (Gollin, 2007), sprzyja rozwojowi biopiractwa, gdyż obecnie jest nie-spójna z systemem wyznaczonym przez CBD. System patentowy wprowadzony na podstawie porozumienia TRIPS jest kolizyjny z postanowieniami ABS. Jak wska-zuje J. Koopman, na podstawie CBD i Zaleceń z Bonn4 dochodzi do błędnego wymieszania praw i obowiązków dotyczących materialnych i niematerialnych przedmio-tów intelektualnej innowacji. W konsekwencji, nie uła-twia to uzgodnienia ich przepisów z porozumieniem TRIPS (Koopman, 2003).
Regulowany dostęp do zasobów genetycznych opie-ra się na warunku uzyskania uprzedniej zgody. System
4 Zalecenia z 2002 r. nt. dostępu do zasobów genetycznych i sprawiedli-wego oraz równego podziału korzyści z ich użytkowania.

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 21
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
uprzedniej zgody został przewidziany w CBD, jed-nak ma on charakter opcjonalny i został wprowadzo-ny w niewielu krajach (np. Brazylia, Indie, Kostaryka, Peru) – tam, gdzie bioróżnorodność jest najbogatsza. Nie obowiązuje jednak na terytorium państw, w któ-rych w większości przypadków materiał biologiczny po-chodzący z eksplorowanych zasobów genetycznych jest przedmiotem wprowadzonych innowacji, prac R&D i ubiegania się o patent. Na terytorium UE na mocy mo-tywu 27 dyrektywy 44/98/WE wprowadzono jedynie warunek poinformowania, skąd dany materiał gene-tyczny pochodzi, jeśli wynalazek oparty jest na materia-le biologicznym pochodzenia roślinnego lub zwierzęce-go lub jeśli wykorzystuje taki materiał. Jest to jednak norma lex imperfecta i taką pozostaje we wszystkich państwach UE, gdzie taką regulację wprowadzono. Nie ma tym bardziej wpływu na udzielenie patentu. Dlatego też wymóg uprzedniej zgody (w krajach, w których obo-wiązuje) sam w sobie nie jest sprzeczny z prawem paten-towym. Sprzeczność taką zauważa się dopiero na etapie ubiegania się o udzielenie patentu, kiedy brak uprzed-niej zgody może zablokować uzyskanie go.
W obliczu nieskuteczności postanowień o ochro-nie zasobów genetycznych, państwa sygnatariusze CBD podpisały protokół z Nagoi, na mocy którego instrument uprzedniej zgody dostępu do zasobów ge-netycznych będzie obligatoryjny, jeśli protokół wejdzie w życie. Obecnie obowiązujące regulacje należy ocenić częściowo krytycznie. Nie chronią bowiem wystarcza-jąco przed nieuprawnionym dostępem do zasobów ge-netycznych. Natomiast w państwach, w których ograni-czane jest uzyskanie patentu na wynalazek z uwagi na brak uprzedniej zgody na dostęp, narusza się postano-wienia porozumienia TRIPS.
Powyższy konflikt można próbować rozwiązać przynajmniej na dwa sposoby: 1) nowelizując prawo pa-tentowe, które będzie spójne z CBD, lub 2) zmieniając
system CBD w harmonii z przepisami prawa patentowe-go. Nie wymaga to jednak zupełnej rezygnacji z systemu ochrony sui generis bioróżnorodności, ale takiego jej zrewidowania, które będzie zgodne zwłaszcza z porozu-mieniem TRIPS. Z pewnością konieczna byłaby rezyg-nacja z wymogu uprzedniej zgody, ale jedynie w takim zakresie, w jakim ma wpływ na udzielenie ochrony pa-tentowej. Rezygnacja z systemu ABS przynajmniej pod względem jego obligatoryjności byłaby nieunikniona. Fakultatywnie uprawniony z patentu mógłby dzielić się korzyściami, ale zależałoby to tylko od przyjętego przez niego zobowiązania w tym zakresie. Przyjęte porozu-mienie z Nagoi i propozycja art. 29 bis porozumienia TRIPS wydają się jednak zmierzać w odmiennym kie-runku – zaostrzenia przepisów o uzyskaniu uprzedniej zgody na dostęp do zasobów genetycznych oraz podzia-le korzyści płynących z prawa własności intelektualnej wykorzystujących bioróżnorodność. Kierunek ten wy-daje się zrozumiały, oczekiwany oraz potrzebny z uwagi na przedmiot ochrony, jakim jest bioróżnorodność. Po-nieważ potrzebne jest istnienie zarówno ochrony paten-towej, jak i ochrony bioróżnorodności, nie można zu-pełnie zrezygnować z żadnej z nich. Nie należy jednak zapominać, że ochrona bioróżnorodności jest kluczowa i pierwszoplanowa. Zadaniem systemu patentowego nie jest zaś tylko wynagradzanie twórcy, lecz przede wszystkim – motywowanie do tworzenia wynalazków z korzyścią dla społeczeństwa.
Literatura:
Alikhan Sh, Mashelkar R (2004). Intellectual Property and Competi-tive Strategies in the 21st Century. Hague – London – New York.
Ammann K (2007). Reconciling Traditional Knowledge with Mo-dern Agriculture: A Guide for Building Bridges. W: Krattiger A, Mohoney RT, ed. Intellectual Property Management in Health and Agricultural Innovation. Oxford – Davis.
Anriantsiferena R (2007). Traditional knowledge protection in the African
region. W: McManis ChR, ed. Biodiversity and the law: Intellectual property, biotechnology and traditional knowledge. London.
Anuradha RV, Kothari A (1999). Biodiveristy and Intellectual Pro-perty Rights: Can the Two Co-Exist? Journal of Int’ l Widlife L & Pol’y. 2:5-6.
Argumedo A, Pimbert M (2006). Protecting Indigenous Knowledge Against Biopiracy in the Andes. Peru.
Arihan O,, Gençler Özkan AM (2007). Traditional medicine and in-tellectual property. Journal of Faculty of Pharmacy of Ankara. 36.
Baharul Islam KM (2011). Defending Traditional Knowledge rights. Indians Legal Battles Against Turmeric, Basmanti Rice and Neem Patents in USA and Europe. Saarbrücken.
Brush Stephan B (2007) The demise of ‘Common Heritage’ and pro-tection for traditional agricultural knowledge. W: McManis ChR, ed. Biodiversity and the law: Intellectual property, biotechnology and traditional knowledge. London.
Compact Oxford English Dictionary (2006). Dostępny na: http://oxforddictionaries.com/definition/bioprospecting?q=bioprospecting. Dostęp 29.30.2013.
Drahos P, Braithwaite J (2002). Information feudalism. New York: The New Press
Gámez R (2007). The link between biodiversity and sustainable de-velopment: Lkessons from INBio’s bioprospecting programme in Costa Rica. W: McManis ChR, ed. Biodiversity and the law: Intellectual property, biotechnology and traditional knowledge. London.
Gollin MA (2007). Answering the call: Public Interest Intellectual Property Advisor (PIIPA). W: McManis ChR, ed. Biodiversity and the Law: Intellectual Property, Biotechnology and Traditional kno-wledge. London.
Hamilton Ch (2006). Biodiversity, Biopiracy and Benefits: What alle-gation of biopiracy tell us about intellectual property? Developing World Bioethics. 6.
Jeffries Michael J (2005). Biodiversity and conservation. London – New York.
Koopman J (2003). Biotechnology, Patent Law and Piracy: Mirroring the Interests in Resources of Life and Culture. Konferencja Ius Commune Research School workshop on Intellectual Property. Edinburgh University School of Law.
Lewis Walter H, Romani V (2007). Ethics and practice in ethnobio-logy: Analysis of the international cooperative biodiversity group project in Peru. W: McManis ChR, ed. Biodiversity and the law: Intellectual property, biotechnology and traditional knowledge, London 2007.
Lisowska E (2012). Zasoby genetyczne i ich ochrona. Kwartalnik UP RP. 1: 48.
Morżoł I. (2010). Ochrona róznorodnosci biologicznej i kulturalnej na obszarach dziedzictwa kulturowego UNESCO. Dostępny na:

Bioróżnorodność a ochrona patentowa | Joanna Uchańska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 22
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
http://www.ekoedu.uw.edu.pl/rep/pdf/07692a1d5b9e1633cbc25 740646 264c3.pdf. Dostęp 2.03.2012.
Mugabe J (2001). Intellectual Property Protection and Traditional Knowledge. An exploration in international policy disclosure. Do-stępny na: http://www.oapi.wipo.net/export/sites/www/tk/en/hr/paneldiscussion/papers/pdf/mugabe.pdf. Dostęp. 30.03.2013.
Ohmagari K, Berkes F (1997). Transmission of Indigenous Kno-wledge and Bush Skills among the Western James Bay Cree Wo-men of Subarctic Canada. Human Ecology. 25.
van Overwalle G (2007). Holder and User Perspectives in the Tradi-tional Knowledge Debate: A European View. W: McManis ChR, ed. Biodiversity and the Law: Intellectual Property, Biotechnology and Traditional Knowledge. London.
Posey D (1996). Beyond Intellectual Property: Towards Traditional Resource Rights for Indigenous Peoples and Local Communities. Ottawa: ON IDRC.
Simelane T. Traditional Knowledge linked Biodiversity. Dostępny na: http://pame.european-patentoffice.org/pubs/harare pdf/tk_lin-ked_to_biodiversity.pdf. Dostęp: 29.03.2013.
Straus J (2008). How to Break the Deadlock Preventing a Fair and Rational Use of Biodiversity. The Journal of World Intellectual Pro-perty. 11:230.
Szkarłat M (2012). Biotechnologia jako przedmiot regulacji prawno międzynarodowych. W: Twardowski T, ed. Prawne i społeczne aspekty biotechnologii. Warszawa.
Torrance A.W (2010). Patent Law, HIPPO, and the Biodiversity Cri-sis. The John Marschall Review of Intellectual Property Law. 9.
Tritton G, Davis R, Edenrorgough M, Graham J, Malynicz S, Roug-hton A (2008). Intellectual Property in Europe. London.
Wilson Edward O (1992). Diveristy of life.Winter PC, Fletcher GI (2004). Krótkie wykłady. Genetyka. Warszawa.World Conservation Strategy Living Resource Conservation for Su-
stainable Development z 1980 r., ONZWynberg R (2003). Sharing the Crumbs with the San. Barcelona:
GRAIN. Dostępny na: http://www.grain.org/bio-ipr/?id=279. Do-stęp 30.03.2013.
Zent S, Zent Egleé L (2007). On biocultural biodiversity from a Ve-nezuela Perspective: Tracing the Interrelationships among Biodi-versity, Culture Change and Legal Reform. W: McManis ChR, ed. Biodiversity and the Law: Intellectual Property, Biotechnology and Traditional Knowledge. London.
Zimny T (2012). Problemy moralne związane z inżynierią genetycz-ną. W: Twardowski T, ed. Aspekty społeczne i prawne biotechno-logii, Warszawa.
Żakowska-Henzler H (2006). Wynalazek biotechnologiczny; przed-miot patentu. Warszawa.
http://www.cbd.int. Dostęp: 29.03.2013http://patft.uspto.gov. Dostęp: 29.03.2013
Biodiversity and patent protection
Joanna Uchańska
The biodiversity is an inexhaustible reservoir of resources which ensure useful solutions of human beings. There-fore, it is necessary to protect natural resources. This pa-per presents selected issues on the sui generis protection of biodiversity established according to the Convention of Biological Diversity of the 5 June, 1992. The problem in-question was analyzed through the prism of the ABS tool – The Access to genetic resources and equitable Shar-ing of Benefits (Access and Benefit Sharing). The prior consent to resources was presented with reference to the ABS. Moreover, the analyze has been broaden by the sub-ject matter of the relationship between the ABS and the patent law. This essay presents two main thesis in the area if the sustainable use of the biodiversity, and the possi-bility of irregularity of the provisions of the Convention on Biological Diversity and the patent law. Nevertheless, the paper discusses the biopiracy and the traditional knowledge.Key words: biodiversity, access and benefit sharing, biopiracy, traditional knowledge, prior consent
http://www.europarl.europa.eu/meetdocs/2009_2014/documents/envi/re/821/821537/821537pl.pdf. Dostęp: 30.03.2013
http://faostat.fao.org/site/339/default.aspx. Dostęp: 30.03.2013http://www.cbd.int. Dostęp: 29.03.2013http://patft.uspto.gov. Dostęp: 29.03.2013http://www.europarl.europa.eu/meetdocs/2009_2014/documents/
envi/re/821/821537/821537pl.pdf. Dostęp: 30.03.2013http://faostat.fao.org/site/339/default.aspx. Dostęp: 30.03.2013

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 23
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Wstęp
Wielu spośród odbiorców tego artykułu zapewne znana jest nowela napisana w 1886 r. przez Roberta Louisa Stevensona pt. Strange Case of Dr Jekyll and Mr Hyde. Któż mógłby się spodziewać, że natura stworzyła podobne „dzieło” na poziomie molekularnym. Mowa tu o białkach prionowych – białkach, które mogą wy-stępować w dwóch jakże odmiennych postaciach: PrPC – niegroźnej dla organizmów ssaczych oraz PrPSc – po-
Kamil Lisiecki: licencjat chemii, student IV roku Między-obszarowych Indywidualnych Studiów Matematyczno--Przyrodniczych, Uniwersytet Warszawski
Molekularny Dr Jekyll i Mr HydeKamil Lisiecki
Streszczenie:
Białka prionowe stanowią jak dotąd bardzo słabo pozna-ną rodzinę białek. Jednakże ich wyjątkowe cechy sprawia-ją, iż od momentu ich odkrycia (uwieńczonego nagrodą Nobla) są intensywnie badanym obiektem. W dodatku, zaskakująca natura białek prionowych – białek o dwóch twarzach – powoduje, że są one niezwykle interesującą ciekawostką biologiczną.Słowa kluczowe: prion, choroby prionowe, PRNP, aktywność antyprionowa, choroba Creutzfeldta-Jakoba, BSE, kurkumina, dendrymery, drożdże
otrzymano: 16.04.2013; przyjęto: 9.05.2013; opublikowano: 28.06.2013
wodującej u ssaków liczne choroby (Prusiner, 1982). W tym drugim przypadku nazywane są one prionami. Pomimo tak fascynującej natury priony nie należą do często omawianych tematów, głównie za sprawą tego, że nasza wiedza na ich temat jest wciąż niewielka. W arty-kule tym postaram się przybliżyć zagadnienie prionów, przedstawić dotychczasowy stan wiedzy w tym zakresie oraz szczególnie skupić się na próbach farmakoterapii w odniesieniu do chorób prionowych.
Rys historyczny
Termin „prion” jest skrótem od angielskiego ter-minu „proteinaceous infectious particle” – „zakaźna cząsteczka białkowa”. Został on wprowadzony w 1982 r. przez amerykańskiego biochemika i neurobiologa, laureata Nagrody Nobla Stanley’a B. Prusiner’a (Prusi-ner, 1982). Warto zdać sobie sprawę, że w latach 80. XX wieku genetyka była już nauką o silnych fundamentach. Genetycy i biolodzy molekularni wierzyli, że nic nie jest już w stanie ich zaskoczyć i że ich wiedza na temat kwa-sów nukleinowych, białek oraz dziedziczenia jest prak-tycznie kompletna. Jakże wielkim zaskoczeniem było dla nich odkrycie prionu – białka, które jest w pewnym sensie wyjątkiem od dwóch obowiązujących w biologii molekularnej i genetyce zasad: 1) przekazywanie infor-macji o patogenności zachodzi przy udziale kwasów nu-kleinowych, 2) struktura drugorzędowa białek determi-nowana jest przez strukturę pierwszorzędową.
Komentarza wymaga również pochodzenie stoso-wanych skrótów (PrPC i PrPSc). PrP to skrót od angiel-skiego terminu „prion protein”, indeksy górne oznaczają natomiast: C – Cellular, czyli komórkowy, występujący naturalnie w komórce, oznaczający niepatogenną for-mę; Sc – jest to skrót od Scrapie, nazwy choroby wystę-pującej u owiec, a wywołanej przez priony, oznaczający formę patogenną tegoż białka.
Charakterystyka białka prionowego
Białko prionowe (PrP) istnieje w dwóch postaciach: fizjologicznej – PrPC, oraz patogennej – PrPSc (ryc. 1). Podstawową cechą formy patogennej jest jej słaba roz-puszczalność w środowisku wodnym oraz brak podat-ności na trawienie proteazami – enzymami hydrolizu-jącymi białka i peptydy (w przeciwieństwie do PrPC). Formy te różnią się również strukturą drugorzędową. W formie fizjologicznej dominuje struktura a-helisy, natomiast w formie patologennej struktura b-kartki (ryc. 2).
Mechanizm powstawania formy PrPSc nie jest jesz-cze do końca wyjaśniony. Naukowcy zdążyli jednak zaproponować model tłumaczący to zjawisko (ryc. 3) (Jackson i Clarke, 2000):
1. Forma fizjologiczna może w sposób spontaniczny, ale odwracalny zmienić się w monomeryczny pre-kursor PrPSc.
2. Prekursory te mogą ze sobą odwracalnie i słabo oddziaływać, dopóki nie powstanie stabilny, in-fekcyjny „zalążek” (przedstawiony na ryc. 2 jako struktura w nawiasie kwadratowym).
3. Przekroczenie ,,masy krytycznej” powoduje, że monomeryczne prekursory PrPSc mogą być przy-łączane nieodwracalnie i umożliwiany jest wzrost cząstek PrPSc.
4. Cząstki zakaźne mogą się powielać poprzez pęk-nięcia na mniejsze, stabilne cząstki.
W ten sposób powstają złogi amyloidowe (nieroz-puszczalne, włókniste agregaty białek), które uważane są za przyczynę chorób prionowych. Należy również zwrócić uwagę, że dostanie się do organizmu formy pa-togennej (zakażenie), działa jak swego rodzaju domino – powoduje indukcję konwersji formy PrPC do formy PrPSc. Nasuwa się pytanie o funkcje fizjologicznej for-my białka prionowego. Funkcje te nie są jak dotąd do
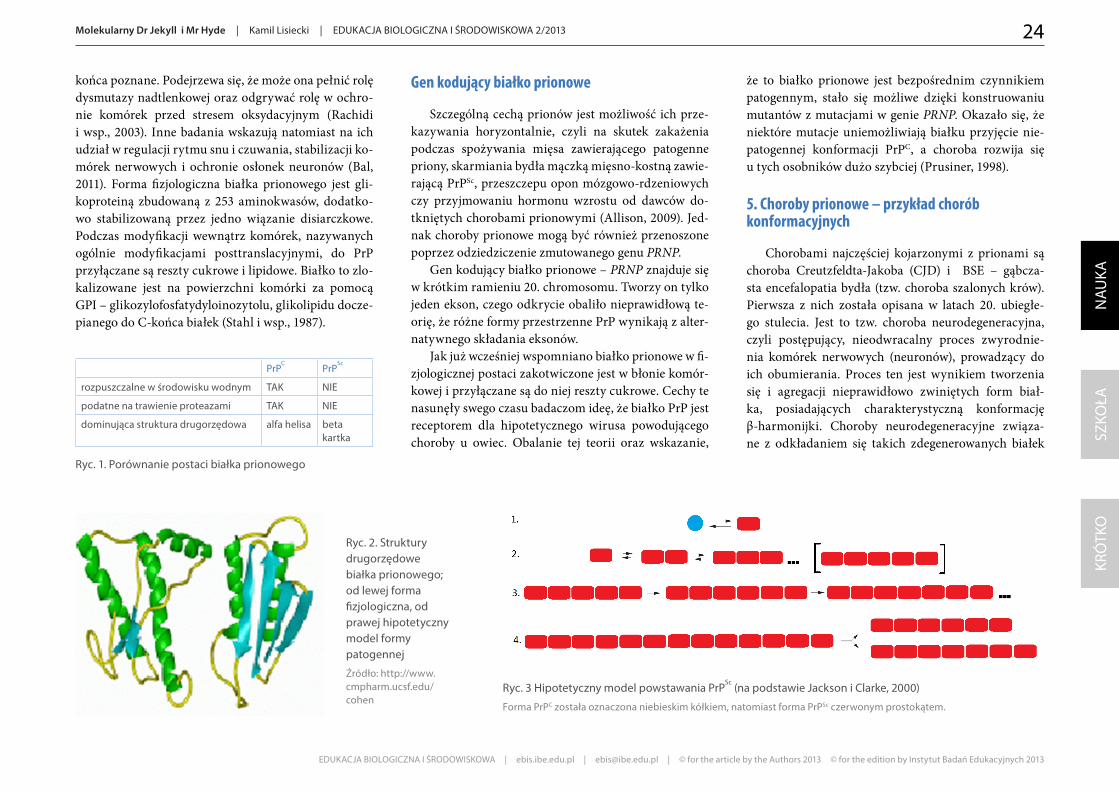
Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 24
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
końca poznane. Podejrzewa się, że może ona pełnić rolę dysmutazy nadtlenkowej oraz odgrywać rolę w ochro-nie komórek przed stresem oksydacyjnym (Rachidi i wsp., 2003). Inne badania wskazują natomiast na ich udział w regulacji rytmu snu i czuwania, stabilizacji ko-mórek nerwowych i ochronie osłonek neuronów (Bal, 2011). Forma fizjologiczna białka prionowego jest gli-koproteiną zbudowaną z 253 aminokwasów, dodatko-wo stabilizowaną przez jedno wiązanie disiarczkowe. Podczas modyfikacji wewnątrz komórek, nazywanych ogólnie modyfikacjami posttranslacyjnymi, do PrP przyłączane są reszty cukrowe i lipidowe. Białko to zlo-kalizowane jest na powierzchni komórki za pomocą GPI – glikozylofosfatydyloinozytolu, glikolipidu docze-pianego do C-końca białek (Stahl i wsp., 1987).
Gen kodujący białko prionowe
Szczególną cechą prionów jest możliwość ich prze-kazywania horyzontalnie, czyli na skutek zakażenia podczas spożywania mięsa zawierającego patogenne priony, skarmiania bydła mączką mięsno-kostną zawie-rającą PrPSc, przeszczepu opon mózgowo-rdzeniowych czy przyjmowaniu hormonu wzrostu od dawców do-tkniętych chorobami prionowymi (Allison, 2009). Jed-nak choroby prionowe mogą być również przenoszone poprzez odziedziczenie zmutowanego genu PRNP.
Gen kodujący białko prionowe – PRNP znajduje się w krótkim ramieniu 20. chromosomu. Tworzy on tylko jeden ekson, czego odkrycie obaliło nieprawidłową te-orię, że różne formy przestrzenne PrP wynikają z alter-natywnego składania eksonów.
Jak już wcześniej wspomniano białko prionowe w fi-zjologicznej postaci zakotwiczone jest w błonie komór-kowej i przyłączane są do niej reszty cukrowe. Cechy te nasunęły swego czasu badaczom ideę, że białko PrP jest receptorem dla hipotetycznego wirusa powodującego choroby u owiec. Obalanie tej teorii oraz wskazanie,
że to białko prionowe jest bezpośrednim czynnikiem patogennym, stało się możliwe dzięki konstruowaniu mutantów z mutacjami w genie PRNP. Okazało się, że niektóre mutacje uniemożliwiają białku przyjęcie nie-patogennej konformacji PrPC, a choroba rozwija się u tych osobników dużo szybciej (Prusiner, 1998).
5. Choroby prionowe – przykład chorób konformacyjnych
Chorobami najczęściej kojarzonymi z prionami są choroba Creutzfeldta-Jakoba (CJD) i BSE – gąbcza-sta encefalopatia bydła (tzw. choroba szalonych krów). Pierwsza z nich została opisana w latach 20. ubiegłe-go stulecia. Jest to tzw. choroba neurodegeneracyjna, czyli postępujący, nieodwracalny proces zwyrodnie-nia komórek nerwowych (neuronów), prowadzący do ich obumierania. Proces ten jest wynikiem tworzenia się i agregacji nieprawidłowo zwiniętych form biał-ka, posiadających charakterystyczną konformację β-harmonijki. Choroby neurodegeneracyjne związa-ne z odkładaniem się takich zdegenerowanych białek
PrPC
PrPSc
rozpuszczalne w środowisku wodnym TAK NIE
podatne na trawienie proteazami TAK NIE
dominująca struktura drugorzędowa alfa helisa beta kartka
Ryc. 1. Porównanie postaci białka prionowego
Ryc. 2. Struktury drugorzędowe białka prionowego; od lewej forma fizjologiczna, od prawej hipotetyczny model formy patogennej
Źródło: http://www.cmpharm.ucsf.edu/cohen
Ryc. 3 Hipotetyczny model powstawania PrPSc
(na podstawie Jackson i Clarke, 2000)
Forma PrPC została oznaczona niebieskim kółkiem, natomiast forma PrPSc czerwonym prostokątem.

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 25
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
noszą nazwę amyloidoz (Allison, 2009). CJD objawia się zanikiem pamięci, trzęsieniem się kończyn i utratą mimiki. W konsekwencji dochodzi do śmierci chore-go w ciągu 2 lat od pojawienia się pierwszych objawów (Prusiner, 1982). Niekorzystną cechą tej choroby jest brak odpowiedzi immunologicznej organizmu. Jest to spowodowane tym, że przyczyną choroby jest białko naturalnie występujące w organizmie ludzkim (Prusi-ner, 1998). Innymi chorobami prionowymi są: śmier-telna bezsenność rodzinna (FFI) – choroba genetyczna powodująca śmierć chorego w ciągu 36 miesięcy od wy-stąpienia objawów (Schenkein, 2006) czy choroba kuru występująca u dopuszczających się kanibalizmu człon-ków plemienia Fore, zamieszkującego Góry Wschodnie Papui Nowej Gwinei (Collinge, 2006). Podobne choroby występują również u innych organizmów, m.in. owiec, kotów lub krów. Krótką charakterystykę chorób prio-nowych przedstawia ryc. 4.
Podczas zakażenia patogenną formą białka priono-wego wyróżnia się cztery etapy infekcji (Epstein, 2005):
1. Penetracja – przedostanie się białka prionowego do komórek układu pokarmowego, a stamtąd do układu limfatycznego,
2. Translokacja – wędrówka PrPSc po układzie lim-fatycznym i krwionośnym zakończona dostaniem się do obwodowego, a następnie centralnego ukła-du nerwowego (rzadziej do śledziony),
3. Namnażanie – konwersja form fizjologicznych do form patogennych, przyłączanie kolejnych podjed-nostek białka,
4. Patogeneza – różnego rodzaju skutki uboczne wy-nikające z odkładania się złogów amyloidowych.
Choroby prionowe nie są jedynymi chorobami związanymi z błędnym zwijaniem się białek. Wszystkie takie schorzenia nazywa się chorobami konformacyj-nymi. Na ryc. 5 przedstawione jest zestawienie najważ-niejszych chorób konformacyjnych.
Choroba Gospodarz Mechanizm patogenezy
KuruCzłonkowie plemie-nia Fore
Infekcja podczas rytualnego kanibalizmu
iCJD Ludzie Infekcja poprzez zawierający priony hormon wzrostu, przy przeszczepie opony twardej
vCJD Ludzie Infekcja bydlęcymi prionami (prawdopodobnie)
FCJD Ludzie Germinalna mutacja w genie PrP
GSS Ludzie Germinalna mutacja w genie PrP
FFI Ludzie Germinalna mutacja w genie PrP (D178N, M129)
sCJD Ludzie Mutacja somatyczna lub spontaniczna konwersja PrPC w PrPSc
FSI Ludzie Mutacja somatyczna lub spontaniczna konwersja PrPC w PrPSc
Scrapie Owce Infekcja u genetycznie podatnych osobników
BSE Bydło Infekcja poprzez zawierającą priony mączkę mięsno-kostną
TME Norki Infekcja prionami od owcy lub bydła
CWD Mulaki, łosie Nieznany
FSE Koty Infekcja poprzez zawierającą priony wołowinę
Egzotyczna encefa-lopatia kopytnych
Kudu wielkie, niala, oryx
Infekcja poprzez zawierającą priony mączkę mięsno-kostną
Skróty: BSE – encefalopatia gąbczasta bydła; CJD – choroba Creutzfeldta-Jakoba; sCJD – sporadyczna CJD; fCJD – rodzinna CJD; iCJD – jatrogenna CJD; vCJD – wariant CJD; CWD – przewlekła choroba wyniszczająca; FFI – śmiertelna bezsenność rodzinna; FSE – kocia encefa-lopatia gąbczasta; FSI – śmiertelna sporadyczna bezsenność; GSS – zespół Gerstmanna-Sträusslera-Scheinkera; TME – przenośna encefalo-patia norek.
Ryc. 4. Zestawienie chorób wywoływanych przez priony
Na podstawie: Prusiner i wsp., 1998
Choroba Błędnie zwinięte białko Natura i umiejscowienie uszkodzeń
Choroba Alzheimera Peptyd A-beta Hiperufosforylowane białko TauZewnątrzkomórkowe blaszki starcze Splątki neu-rofibrylarne
Choroba Parkinsona alfa-Synukleina Cytoplazma neuronów
Choroba Huntingtona Powtórzenie poliglutaminowe w huntingtynie Jądra i cytoplazma neuronów
Cukrzyca typu II Wysepkowy polipeptyd amyloidowy (amylina) Agregaty w trzustce
Choroba Creutzfeldta-Jakoba Białko prionowe Płytki zewnątrzkomórkowe i oligomery wewnątrz i na zewnątrz neuronów
Ryc. 5. Porównanie chorób konformacyjnych
Na podstawie: Allison, 2009

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 26
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Często w literaturze można spotkać się z informa-cjami na temat aktywności antyprionowej związków wykorzystywanych w leczeniu innych chorób konfor-macyjnych. Jest to przesłanka do twierdzenia, że być może wszystkie choroby (lub większość z nich) zwią-zane z nieprawidłowym zwijaniem się białek mają wspólne podłoże. Możliwe, że istnieje dodatkowe biał-ko występujące w komórce, które pełni rolę centralnego węzła, inicjatora chorób konformacyjnych. Być może istnieje jeden, wspólny mechanizm błędnego zwijania się białek. Jednak jak do tej pory nie ma jednoznacznych informacji na ten temat.
„Stany prionowe” u drożdży
Na podstawie podanych wyżej informacji można sądzić, że priony są białkami stanowiącymi ogromne zagrożenie dla organizmów. I rzeczywiście tak jest, jeśli mamy na myśli organizmy ssacze. Okazuje się jednak, że istnieją organizmy, u których obecność tzw. stanów prinowych może odgrywać pozytywną rolę – chodzi tu o drożdże piekarnicze Saccharomyces cerevisiae. Te proste, jednokomórkowe organizmy są najlepiej pozna-nymi na poziomie genetyki i fizjologii komórki organi-zmami eukariotycznymi (Wawrzycka, 2011). Dlatego też nazywa się je organizmami modelowymi. S. cerevi-siae zawiera wiele białek, które zachowują się jak priony (Derkatch i wsp., 2001). Dodatkowo szlaki biochemicz-ne kontrolujące powstawanie agregatów prionów lub ich utrzymanie są zachowane od drożdży do człowieka (Bach i wsp., 2006).
Należy jednak wyraźnie zaznaczyć, że drożdżowe „stany prionowe” nie mają wiele wspólnego z priona-mi ssaczymi. Są to stany metaboliczne wywołane przez zmiany konformacyjne białek, niewykazujących homo-logii sekwencyjnej do PrP. Natomiast wspólną ich cechą jest możliwość przyjmowania dwóch różnych konfor-
macji bez zmiany sekwencji amino-kwasowej i odkładanie się w pew-nych warunkach w postaci złogów amyloidowych.
Najlepiej poznanym „stanem prionowym” u drożdży jest ten spo-wodowany obecnością czynnika [PSI+] (Cox i wsp., 1988). Komórki posiadające ten czynnik dużo częś-ciej pomijają kodony stop na koń-cu obszaru kodującego genu, przez co powstaje dłuższe niż powinno białko. Istotą czynnika [PSI+] jest białko Sup35. Jest ono czynnikiem terminacji translacji, składnikiem złożonego kompleksu uwalniającego rybosom od mRNA (ortolog ssacze-go eRF3) (Stansfield i wsp., 1995). Gdy dojdzie do inaktywacji Sup35, kompleks, którego jest składnikiem, przestaje działać poprawnie, powo-dując defekt w terminacji translacji. Drożdże wykazujące tę cechę ozna-cza się symbolem [PSI+], natomiast komórki bez takich zmian [psi-]. Istotę opisanego zjawiska przedsta-wia ryc. 6.
Istotnym momentem w badaniach nad czynnikiem [PSI+] było wykorzystanie mutacji w genie ADE2, któ-ry koduje jeden z enzymów niezbędnych do biosyntezy adeniny. Wykorzystywana mutacja wprowadza przed-wcześnie kodon stop – przez co powstaje krótkie, nie-funkcjonalne białko. Taki enzym nie pełni poprawnie swojej funkcji, a więc nie może dojść do dokończenia biosyntezy adeniny. Zamiast tego dochodzi do nagro-madzenia się półproduktu o czerwonej barwie (Silhan-kova, 1972). Przeprowadzono doświadczenie polegające
na wprowadzeniu czynnika [PSI+] do komórek droż-dżowych posiadających opisaną mutację w genie ADE2. Okazało się, że komórki takie w przeciwieństwie do ko-mórek [psi-] miały normalną, białą barwę (Cox i wsp., 1980). Tłumaczy się to tym, że czynnik [PSI+] umożli-wił pominięcie mutacji (przedwczesnego kodonu stop) w genie ADE2. Dzięki temu możliwe było powstanie funkcjonalnego enzymu i dokończenie biosyntezy ade-niny.
Ryc. 6. Związek Sup35 z czynnikiem [PSI+]
Na podstawie: Uptain, Lindquist, 2002

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 27
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Istnieje hipoteza o związku między czynnikiem [PSI+] a ewolucją genomu drożdży (True i wsp., 2004). Okazuje się bowiem, że z pewną częstością budowa przestrzenna białka Sup35 może się spontanicznie zmieniać i przyjmować nieaktywną formę. W zwy-kłych warunkach środowiska zmiana ta nie przynosi dla komórki korzyści. Jednak nagła zmiana warunków może spowodować, że jedyną możliwością przeżycia będzie posiadanie czynnika [PSI+]. W tych warunkach faworyzowane są oczywiście komórki [PSI+] i zaczyna-ją dominować w populacji, aczkolwiek spontanicznie powstają również komórki [psi-]. Gdy nastąpi powrót do warunków pierwotnych, lepiej przystosowanymi komórkami są te pozbawione czynnika [PSI+] i tym razem to one są faworyzowane i zaczynają dominować. Mechanizm ten umożliwia drożdżom szybkie dostoso-wanie się do zmieniających się warunków środowiska. W przeciwieństwie do zmian genetycznych przejście ze stanu [psi-] do stanu [PSI+] jest szybkie, a ponadto od-wracalne.
Strategie leczenia oraz problemy i wyzwania związane z potencjalnymi lekami antyprionowymi
Na poziomie molekularnym, terapeutyki potencjal-nie aktywne antyprionowo działają zgodnie z jedną z dwóch głównych strategii (Clare i wsp., 2006):
1) stabilizacja agregatów PrPSc (uniemożliwienie przyłączania kolejnych podjednostek) i hamowa-nie konwersji form PrPC do PrPSc,
2) niszczenie i usuwanie formy patogennej.Z poznanych do tej pory związków o aktywności
antyprionowej znakomita większość działa zgodnie z pierwszą strategią. Do związków tych należą m.in.: kurkumina, pochodne akrydyny, 2,5-diamino-1,4-ben-zochinony i naturalne polifenole.
Jednak do tej pory nie dysponujemy lekiem na cho-roby prionowe. Wszystkie związki, które dotąd zbadano pod względem aktywności antyprionowej, są aktywne jedynie in vitro. Brak aktywności in vivo podyktowany jest głównie dwoma przeszkodami:• brakiem możliwości pokonania przez cząsteczkę
bariery krew-mózg i dotarcia do neuronów (miej-sca docelowego);
• słabą biodostępnością.Pierwsza z tych przeszkód związana jest z występo-
waniem w organizmach ssaków bariery krew-mózg. Jest to fizyczna i biochemiczna bariera pomiędzy naczynia-mi krwionośnymi a tkanką nerwową. Ma ona na celu chronić układ nerwowy przed szkodliwymi czynnika-mi, a także umożliwić selektywny transport substancji z krwi do płynu mózgowo-rdzeniowego. Substancje mogą pokonać tę barierę w jeden z dwóch możliwych sposobów: na drodze dyfuzji (głównie związki drobno-cząsteczkowe) i na drodze transportu aktywnego, przy udziale białek błonowych (peptydy i białka takie jak in-sulina czy oksytocyna).
W mojej opinii istnieją dwie zasadnicze drogi, które być może ułatwią ominięcie tej przeszkody:
1) metoda biologiczna – podanie leku bezpośrednio do rdzenia kręgowego (co wiąże się z niebezpie-czeństwem jego uszkodzenia) lub w obszary, gdzie szczelność bariery krew-mózg jest osłabiona (rejo-ny tylnego płata przysadki mózgowej czy okolice splotu naczyniówkowego) (Krzymowski i Przała, 2005);
2) metoda chemiczna – przyłączenie na drodze synte-zy organicznej grupy zdolnej do pokonania bariery krew-mózg do związku o pożądanej aktywności.
Drugim poważnym problemem jest ograniczona biodostępność niektórych związków. W tym przypad-ku zasadniczą rolę odgrywa rozpuszczalność danego
związku w płynach ustrojowych, których głównym składnikiem jest woda.
Najprostszym rozwiązaniem tego problemu jest syn-teza pochodnej danego związku, która będzie wykazy-wała pożądaną aktywność, ale której rozpuszczalność w wodzie będzie dostatecznie dobra. W tym przypadku może się jednak okazać, że zwiększona biodostępność danego związku spowoduje skutki uboczne dla organi-zmu.
Testy na aktywność badanego związku
Istnieje kilka metod sprawdzenia, czy dany związek jest rzeczywiście aktywny pod względem antypriono-wym. Przedstawię dwie, w moim odczuciu, najprostsze metody.
Metoda z wykorzystaniem drożdży (Cox i wsp., 1980) – w tym przypadku wykorzystuje się opisany wy-żej fakt, że „stany prionowe” u drożdży sprzyjają pomi-janiu kodonu stop w obszarze kodującym genu, przez co powstają aktywne, niezmutowane białka. Gdy w genie ADE2, który koduje jeden z enzymów niezbędnych do syntezy adeniny wprowadzi się przedwczesny kodon stop uniemożliwi to powstanie enzymu o odpowied-niej długości. Synteza adeniny nie będzie wtedy moż-liwa i dojdzie do nagromadzenia półproduktu, który jest czerwony. W przypadku „stanów prionowych” na-stępuje supresja kodonu stop, gdyż czynnik terminacji translacji Sup35 jest nieaktywny i powstaje prawidło-wy enzym, a co za tym idzie – nia ma czerwonej bar-wy. Gdy więc badany przez nas związek jest aktywny (związkiem tym nasącza się bibułę i umieszcza się ją na szalce z hodowlą drożdżową) pojawia się czerwone za-barwienie „wyleczonych” komórek (ryc. 7).

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 28
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Metoda z wykorzystaniem proteinazy K (Caughey i wsp., 2003) – w metodzie tej wykorzystuje się fakt, że forma PrPC jest podatna na trawienie, natomiast forma PrPSc nie ulega hydrolizie pod wpływem proteinazy K (PK). Komórki zainfekowane prionami (bardzo często są to komórki ScN2a, czyli mysie komórki nerwiaka płodowego) poddaje się działaniu badanego związku w odpowiednim stężeniu, a następnie uzyskuje się lizat komórkowy, który poddawany jest działaniu proteinazy K. Jeśli związek jest aktywny, priony występujące w li-zacie zostają strawione przez PK. Jeśli jest przeciwnie – PK nie strawi prionów (związek jest nieaktywny).
Poszukiwania leków antyprionowych
W części tej opiszę kilka z licznych związków wyka-zujących in vitro aktywność antyprionową.
Prostym i szybkim sposobem weryfikacji dużej ilo-ści związków pod względem zdolności do hamowania tworzenia złogów amyloidowych jest metoda opisana w 2003 r. w Journal of Virology (Kocisko, 2003). Bada-nia te miały na celu wyłonienie klas związków, które mogą być rozpatrywane pod kątem farmakoterapii an-typrionowej. Test został wykonany na bibliotece 2000 związków, które były już stosowane jako leki w leczeniu innych chorób (niekoniecznie neurodegeneracyjnych) lub były znanymi związkami naturalnymi. Metoda po-stępowania była zmodyfikowaną wersją metody z wyko-rzystaniem proteinazy K, którą opisałem w poprzednim rozdziale. Różnica polegała na umieszczaniu zainfeko-wanych komórek na płytce zawierającej 96 wgłębień, a następnie dodawaniem do każdego z wgłębień innego związku w odpowiednim stężeniu.
Dzięki przeprowadzonym badaniom udało się wy-brać 17 najbardziej aktywnych inhibitorów (ryc. 8).
Pośród wyłonionych 17 najbardziej aktywnych in-hibitorów znalazły się związki należące do pięciu klas:
1) polifenole – związki za-wierające wiele grup feno-lowych odpowiadających za właściwości przeciwu-tleniające tych związków;
2) leki antymalaryczne – związki wykorzystywane w leczeniu malarii – tro-pikalnej choroby pasożyt-niczej;
3) leki antyhistaminowe – związki opóźniające i ła-godzące reakcje alergicz-ne poprzez blokowanie działania receptorów od-powiadających za uwal-nianie histaminy;
4) fenotiazyna i jej pochod-ne – związki wykorzysty-wane w leczeniu zaburzeń psychicznych;
5) inne.
Ryc. 7. Etapy testu na aktywność antyprionową z wykorzystaniem drożdży
Na podstawie: Bach i wsp., 2003
Ryc. 8. Schemat postępowania podczas badania biblioteki związków
Na podstawie: Kocisko, 2003

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 29
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Faktem przemawiającym za wiarygodnością uzy-skanych wyników jest to, że dwa spośród 17 wyłonio-nych, najaktywniejszych inhibitorów była znana już wcześniej ze swojej aktywności antyprionowej – były to lowastatyna i chinakryna.
Kurkumina (ryc. 9) jest głównym składnikiem kurkumy – przyprawy azjatyckiej zwanej również szafranem indyj-skim. Stosowana jest również jako żółto-pomarańczowy pigment. Strukturę związku ustalili w 1910 r. Stanisław Ko-stanecki, Janina Miłobędzka i Wiktor Lampe (Miłobędzka i wsp., 1910). Jest ona w kręgu zainteresowań naukowych już od dawna ze względu na szeroką gamę aktywności bio-logicznych: przeciwutleniających, przeciwzapalnych, prze-ciwwirusowych, antybakteryjnych i przeciwgrzybiczych, mogących przydać się w leczeniu nowotworów, cukrzycy, reumatoidalnego zapalenia stawów, Alzheimera i innych przewlekłych chorób.
Ten niezwykle intensywnie badany związek posiada kilka cech sprawiających, iż stał się on również intere-sującym obiektem badań pod kątem zastosowania jako lek antyprionowy (Demaimay i wsp., 1998):• Jest strukturalnie podobny do czerwieni Kongo
(ryc. 10) – najbardziej aktywnego, niskocząstecz-kowego inhibitora PrPSc zbadanego do roku 2003: obydwa są związkami płaskimi, zawierającymi dwa pierścienie aromatyczne bądź system pierście-ni aromatycznych połączonych łącznikami. Bada-nia wykazały, iż płaska struktura pierścieni i łącz-nika są ważne dla aktywności inhibicyjnej.
• Kurkumina jest przeciwutleniaczem – istnieją przesłanki mówiące, iż ta właściwość związków może mieć korzystny wpływ na aktywność anty-prionową (Kocisko, 2003).
• Kurkumina jest składnikiem ludzkiej diety. Wiele osób spożywa kurkuminę w dużych ilościach bez oznak jej toksyczności.
Badanie właściwości antyprionowych kurkuminy przeprowadzono zgodnie z metodą wykorzystującą proteinazę K. Okazało się, iż stężenie kurkuminy po-trzebne do 50% inhibicji wynosi 10 nM (jest to tzw. współczynnik IC50). Naukowcy wykonujący opisane badania określili działanie kurkuminy jako silne, nie-odwracalne i selektywne w komórkach ScNB (komórki nerwiaka płodowego zainfekowane prionami) (Cau-ghey, 2003).
Wykonano również badania na chomikach (kurku-mina mieszana była z pokarmem podawanym gryzo-niom) mające na celu wykazanie, czy kurkumina zdol-na jest do inhibicji konwersji (zmiany postaci) białek prionowych wewnątrz organizmu żywego (więc rów-nież czy zdolna jest do pokonania bariery krew-mózg). Niestety kurkumina nie wypadła pozytywnie w tym te-ście, w związku z czym nie jest ona zdolna do działania antyprionowego in vivo (Caughey, 2003).
Podsumowując tę część wykazano, iż kurkumina hamuje akumulację złogów amyloidowych in vitro. Jest więc jednym z najmniejszych (w sensie masy cząstecz-kowej) i najbardziej aktywnych inhibitorów formowa-nia się PrPSc. Niestety, w postaci występującej natural-nie nie jest ona zdolna do pokonania bariery krew-mózg (na co wpływ ma też jej słaba biodostępność). W moim przekonaniu badania nad kurkuminą powinny podą-żać w stronę otrzymania pochodnej, która bez utraty aktywności antyprionowej będzie wykazywała dużo większą biodostępność.
Dendrymery (gr. dendron – drzewo) są związkami, których historia liczy już około 30 lat. Po raz pierwszy zostały zsynte-tyzowane przez niemieckiego chemika Fritza Vögtla w 1978 roku (Buhleier i wsp., 1978). Pod względem chemicznym dendrymery są polimerami cechującymi się budową rozga-łęzioną, trójwymiarową, o kształcie zbliżonym do kuli.
W budowie strukturalnej dendrymerów można wy-różnić wielofunkcyjny rdzeń, od którego promieniście odchodzą „gałęzie” (ramiona) dendrymerów zwane dendronami. Na końcu dendronów znajdują się wolne grupy funkcyjne, które mogą być zmienione przez róż-nego rodzaju podstawniki modyfikujące właściwości chemiczne i fizyczne cząsteczki dendrymeru (Sękowski i wsp., 2008).
Te fraktalopodobne, regularne struktury są stosun-kowo nowymi i wysoce rozgałęzionymi polimerami, mającymi wiele interesujących właściwości. Związki te mogą być wykorzystane m.in. jako: przenośniki leków i genów, czynniki kontrastujące czy wskaźniki różnych jonów metali. Kuliste cząsteczki tych związków odzna-czają się również aktywnością farmakologiczną. Obec-nie dendrymery są intensywnie badane jako czynniki antyprionowe i zabezpieczające przed formowaniem się blaszek amyloidowych (Sękowski i wsp., 2008).
Dendrymery są jedyną, opisaną klasą związków posiadających zdolność do usuwania istniejących pa-
Ryc. 9. Struktura kurkuminy
Ryc. 10. Struktura czerwieni Kongo
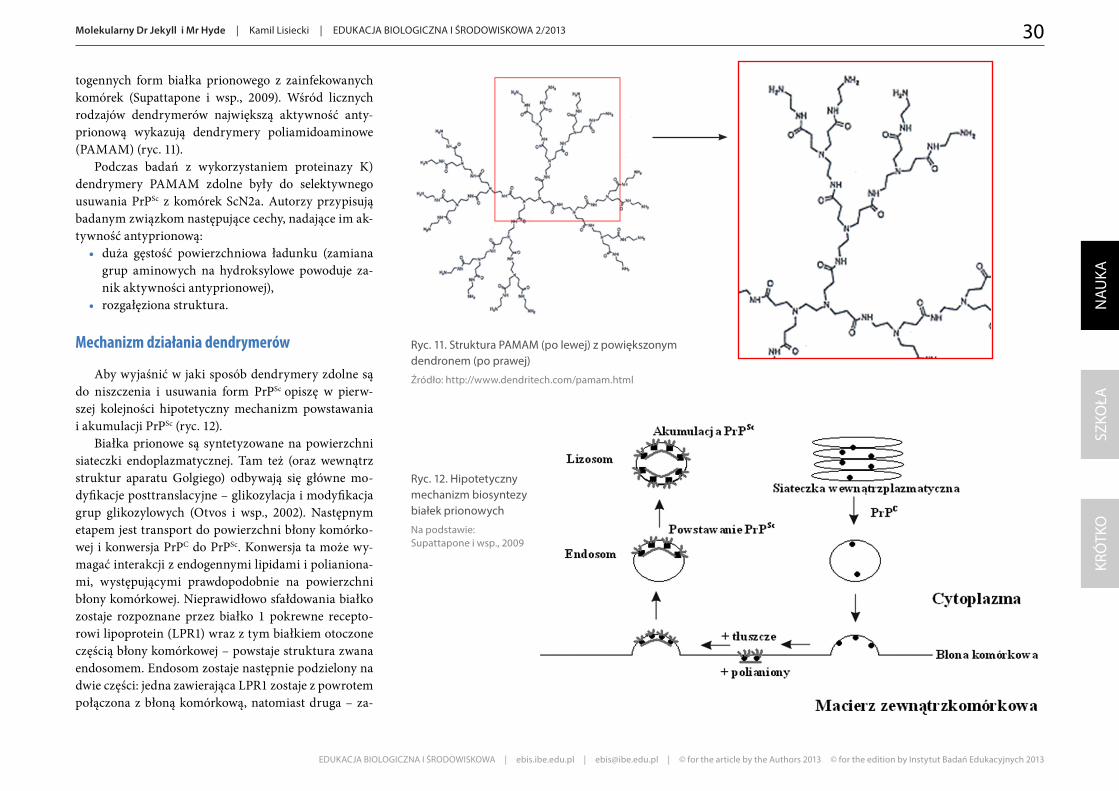
Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 30
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
togennych form białka prionowego z zainfekowanych komórek (Supattapone i wsp., 2009). Wśród licznych rodzajów dendrymerów największą aktywność anty-prionową wykazują dendrymery poliamidoaminowe (PAMAM) (ryc. 11).
Podczas badań z wykorzystaniem proteinazy K) dendrymery PAMAM zdolne były do selektywnego usuwania PrPSc z komórek ScN2a. Autorzy przypisują badanym związkom następujące cechy, nadające im ak-tywność antyprionową:• duża gęstość powierzchniowa ładunku (zamiana
grup aminowych na hydroksylowe powoduje za-nik aktywności antyprionowej),
• rozgałęziona struktura.
Mechanizm działania dendrymerów
Aby wyjaśnić w jaki sposób dendrymery zdolne są do niszczenia i usuwania form PrPSc opiszę w pierw-szej kolejności hipotetyczny mechanizm powstawania i akumulacji PrPSc (ryc. 12).
Białka prionowe są syntetyzowane na powierzchni siateczki endoplazmatycznej. Tam też (oraz wewnątrz struktur aparatu Golgiego) odbywają się główne mo-dyfikacje posttranslacyjne – glikozylacja i modyfikacja grup glikozylowych (Otvos i wsp., 2002). Następnym etapem jest transport do powierzchni błony komórko-wej i konwersja PrPC do PrPSc. Konwersja ta może wy-magać interakcji z endogennymi lipidami i polianiona-mi, występującymi prawdopodobnie na powierzchni błony komórkowej. Nieprawidłowo sfałdowania białko zostaje rozpoznane przez białko 1 pokrewne recepto-rowi lipoprotein (LPR1) wraz z tym białkiem otoczone częścią błony komórkowej – powstaje struktura zwana endosomem. Endosom zostaje następnie podzielony na dwie części: jedna zawierająca LPR1 zostaje z powrotem połączona z błoną komórkową, natomiast druga – za-
Ryc. 11. Struktura PAMAM (po lewej) z powiększonym dendronem (po prawej)
Źródło: http://www.dendritech.com/pamam.html
Ryc. 12. Hipotetyczny mechanizm biosyntezy białek prionowych
Na podstawie: Supattapone i wsp., 2009

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 31
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
wierająca błędnie zwinięte białko zostaje połączona z lizosomem – organellum komórkowym zawierają-cym kwaśne enzymy hydrolityczne rozkładające białka, kwasy nukleinowe, węglowodany i tłuszcze (Mizerski, 2004). Ponieważ jednak PrPSc nie podlegają trawieniu przez proteazy dochodzi do ich akumulacji wewnątrz lizosomu i tworzenia złogów amyloidalnych.
Proponowany mechanizm działania dendrymerów zakłada ich działanie wewnątrz lizosomów (ryc. 13). Dendrymery stają się częścią endosomu (wraz z PrPSc
i LPR1), a następnie lizosomu. Kwaśne środowisko wnę-trza lizosomu powoduje „aktywację” grup aminowych (powstają kationy) dendrymerów, które denaturują i rozbijają złogi amyloidowe. Nie wykluczone, iż rów-nież enzymy występujące wewnątrz lizosomów pełnią funkcję aktywatorów tego procesu.
Reasumując, dendrymery jako jedyne, dotąd opisa-ne związki zdolne są do usuwania z komórek patogen-
nych form białka prionowego. Ich działanie umożliwia ich akumulacja wewnątrz lizosomów (gdzie akumulują się również PrPSc) oraz kwasowe środowisko tych orga-nelli komórkowych.
Wyzwania jakie stoją przed naukowcami badający-mi dendrymery pod względem aktywności antypriono-wej to:• sposób dostarczania leków do organizmu,• zwiększenie ich biodostępności,• uniknięcie potencjalnych, neurologicznych skut-
ków ubocznych wynikających ze stosowania den-drymerów jako leku.
Podsumowanie
Podobnie jak tytułowa nowela „Strange Case of Dr Jekyll and Mr Hyde” kojarzona jest z portretem podwój-
nej osobowości, tak białka prionowe mogą stanowić molekularny przykład dwoistości natury. Dodatkowo pytanie, czy Jekyll pokonał Hyde w powieści Stevenso-na, w przypadku białek prionowych może brzmieć: czy jesteśmy w stanie „pokonać” patogenną postać białka prionowego? Jak na razie trudno jest odpowiedzieć na to pytanie ponieważ biologia i biochemia białek priono-wych, jak również aspekty fizykochemiczne związków o aktywności antyprionowej nie są w pełni wyjaśnione. Problemy te są zatem często poruszane w pracach na-ukowych z pogranicza wyżej wymienionych dziedzin nauki i jedynie interdyscyplinarne podejście do tego zagadnienia może zaowocować wymiernymi osiągnię-ciami.
Z pewnością wyjaśnienie zagadki jaką postawiła przed nami natura – białka prionowe – przyczyni się znacząco do rozwoju światowej nauki oraz myśli bio-chemicznej.
Literatura
Pruisner SB (1982). Novel proteinaceous infectious particles cause scrapie. Science. 216(4542):136-44
Jackson GS, Clarke AR. (2000). Mammalian prion proteins. Curr Opin Struct Biol. 10: 69-74
Rachidi W, Vilette D, Guiraud P, Arlotto M, Riondel J, Laude H, Leh-mann S, Favier A (2003). Expression of prion protein increases cellular copper binding and antioxidant enzyme activities but not copper delivery. J Biol Chem. 278:9064-9072
Bal J (2011). Biologia molekularna w medycynie. Warszawa: Wydaw-nictwo Naukowe PWN
Vanik DL, Surewicz WK (2002). Disease-associated F198S mutation increases the propensity of the recombinant prion protein for conformational conversion to scrapie-like form. J. Biol. Chem. 277:49065-49070
Allison LA (2009). Podstawy biologii molekularnej. Warszawa: WUW Prusiner SB (1998). Prions. Proc Natl Acad Sci USA. 95:13363-83Schenkein J (2006). Self Management of Fatal Familial Insomnia.
Part 1: What Is FFI? Medscape General Medicine. 8(3):65Collinge J, Whitfield J, McKintosh E, Beck J, Mead S, Thomas DJ,
Alpers MP (2006). Kuru in the 21st century—an acquired human
Ryc. 13. Mechanizm działania dendrymerów
Na podstawie: Supattapone i wsp., 2009

Molekularny Dr Jekyll i Mr Hyde | Kamil Lisiecki | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 32
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
prion disease with very long incubation periods. Lancet. 9528 (367):2068–2074
Epstein RJ (2005). Biologia molekularna człowieka. Lublin: Wyd. CZELEJ
Wawrzycka D (2011). Drożdże jako model w badaniach chorób neurodegeneracyjnych. Postępy Hig Med Dosw (online). 65:328-337Derkatch IL, Bradley ME, Hong JY, Liebman SW (2001). Prions af-
fect the appearance of other prions: the story of [PIN(+)]. Cell. 106(2):171-82
Bach S, Tribouillard D, Talarek N, Desban N, Gug F, Galons H, Blon-del M (2006). A yeast-based assay to isolate drugs active against mammalian prions. Methods. 39: 72–77
Cox BS, Tuite MF, McLaughin CS (1988). The psi factor of yeast: a problem in inheritance. Yeast 4:159-178
Stansfield I, Jones KM, Kushnirov VV, Dagkesamanskaya AR, Po-znyakovski AI, Paushkin SV, Nierras CR, Cox BS, Ter-Avanesyan MD, Tuite MF (1995). The products of SUP45 (cRF1) and SUP35 genes interact to mediate translation termination in Saccharomy-ces cerevisiae. EMBO J. 14(17):4365-4373
Uptain SM, Lindquist SL (2002). Prions as protein-based genetic ele-ments. Annu Rev Microbiol. 56:703-741
Silhankova L (1972). Joined suppression of rough phenotype and of red colour of ade2-1 mutants in Saccharomyces cerevisiae. Folia Microbiol. 17(6):479-489
True HL, Berlin I, Lindquist SL (2004). Epigenetic regulation of translation reveals hidden genetic variation to produce complex traits. Nature 431(7005):184-187
Clare R, Trevitt and John Collinge (2006). A systematic review of prion therapeutics in experimental models. Brain. 129:2241-2265
Krzymowski T, Przała J (2005). Fizjologia zwierząt: podręcznik dla studentów wydziałów medycyny weterynaryjnej, wydziałów biolo-gii i hodowli zwierząt akademii rolniczych i uniwersytetów. War-szawa: Państwowe Wydawnictwo Rolnicze i Leśne
Cox BS, Tuite MF, Mundy CJ (1980). Reversion from suppression to nonsuppression in SUQ5 [psi+] strains of yeast: the classificaion of mutations. Genetics. 95(3):589-609
Bach S, Talarek N, Andrieu T, Vierfond JM, Mettey Y, Galons H, Dor-mont D, Meijer L, Cullin C, Blonde M (2003). Isolation of drugs active against mammalian prions using a yeast-based screening assay. Nature Biotechnology. 21:1075 – 1081
Caughey B, Raymond LD, Raymond GJ, Maxson L, Silveira J, Baron GS (2003). Inhibition of Protease-Resistant Prion Protein Accu-mulation In Vitro by Curcumin. J Virol 77: 5499-5502
Kocisko D, Baron GS, Rubenstein R, Chen J, Kuizon S, Caughey B (2003). New Inhibitors of Scrapie-Associated Prion Ptotein For-mation in a Library of 2,000 Drugs and Natural Products. J Virol. 77:10288-10294
Miłobędzka J, Kostanecki S, Lampe V (1910). Zur Kenntnis des
Curcumins. Berichte der deutschen chemischen Gesellschaft. 43(2):2163–2170
Demaimay R, Harper J, Gordon H, Weaver D, Chesebro B, Caughey B (1998). Structural aspects of Congo red as an inhibitor of protease resistant prion protein formation. J Neurochem. 71:2534–2541.
Buhleier E, Wehner W, Vögtl F (1978). „Cascade“ and „Nonskid--Chainlike” syntheses of molecular cavity and topologies. Synthe-sis. 2:155–158
Sękowski S, Miłowska K, Gabryelak T (2008). Dendrimers in bio-medical sciences and nanotechnology. Postepy Hig Med Dosw. 62:725-733
Supattapone S, Piro JR, Rees JR (2009). Complex Polyamines: Unique Prion Disaggregating Compounds. NIH. 8:323-328
Otvos L Jr, Cudic M, (2002). Post-translational modifications in prion proteins. Curr Protein Pept Sci. 3(6):643-52.
Mizerski W (2004). Tablice biologiczne. Warszawa: Wydawnictwo Adamantan
Molecular Dr. Jekyll and Mr. Hyde
Kamil Lisiecki
Prion proteins are as yet very poorly known family of pro-teins. However, their unique characteristics make them, since their discovery (which was rewarded with the Nobel Prize) intensively investigated object. In addition, a sur-prising nature of prion proteins – proteins with two faces – makes them an extremely interesting biological curios-ity.Key words: prion, prion diseases, PRNP, antiprion activity, Creutzfeldt-Jacob disease, BSE, curcumin, dendrimers, yeast

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 33
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Agnieszka Osówniak: Uniwersytet Warszawski, Kolegium Międzywydziałowych Indywidualnych Studiów Matema-tyczno-Przyrodniczych
Siarka i jej związki jako źródła pierwiastka budulcowego dla bakteriiAgnieszka Osówniak
Streszczenie:
Siarka jest jednym z pierwiastków niezbędnych do funk-cjonowania wszystkich organizmów. W pracy opisano właściwości fizyczne i chemiczne tego pierwiastka, a tak-że związki pochodzenia biogennego, w skład których on wchodzi. Następnie scharakteryzowano związki nieorga-niczne (siarczki, siarczany, tiosiarczany) i organiczne (cy-steina, metionina), które mogą być źródłem siarki do ce-lów budulcowych i sposób ich asymilacji przez bakterie. Przedstawiono też biosyntezę L-metioniny i L-cysteiny, enzymów zawierających centra żelazo-siarkowe, biotyny, tiaminy, kwasu liponowego, koenzymu A, molibdoptery-ny i S-adenozylometioniny. Na koniec omówiono pokrót-ce udział bakterii w obiegu siarki.Słowa kluczowe: asymilacja, bakteria, biosynteza, cysteina, enzym, operon, siarczan, siarka, tiosiarczan
otrzymano: 22.04.2013; przyjęto: 18.06.2013; opublikowano: 28.06.2013
Wstęp
Siarka (ryc. 1) jest pierwiastkiem grupy 6. układu okresowego o typowych właściwościach niemetalicz-nych, zajmującym 16. miejsce wśród pierwiastków najbardziej rozpowszechnionych na Ziemi. Występuje w skorupie ziemskiej w ilości 0,035%, zarówno w for-mach nierozpuszczalnych – jako blenda cynkowa (ZnS), piryt (FeS2) czy galena ołowiana (PbS), jak i w formach rozpuszczalnych – Na2SO4, MgSO4. W przyrodzie siar-kę możemy spotkać w formie jednego z czterech stabil-nych izotopów: 32S, 33S, 34S i 36S (Canfield, 2001). Przed omówieniem właściwości chemicznych i fizycznych tego pierwiastka należy zwrócić uwagę na jego alotropię – czyli zdolność do występowania w różnych formach molekularnych o tym samym stanie skupienia. Alotro-pię wykazuje niewiele pierwiastków, np. fosfor, selen, tellur czy węgiel, a także, jak już wspomniano, siarka. Cechą wspólną tych pierwiastków, pozwalającą na po-wstawanie ich odmian alotropowych, jest zdolność do katenacji, czyli tworzenia łańcuchów lub pierścieni zbu-dowanych z bezpośrednio połączonych atomów.
Najtrwalszą i najbardziej znaną odmianą alotropo-wą siarki jest cyklo-oktasiarka (S8), występująca ma-kroskopowo jako kryształ molekularny, mikroskopowo zaś w postaci ośmiu atomów związanych w pierścień. W obrębie utworzonej cząsteczki występują wiązania kowalencyjne, natomiast kryształ utrzymywany jest dzięki słabszym wiązaniom van der Waalsa pomiędzy cząsteczkami S8. W zależności od sposobu upakowania tych cząsteczek w krysztale, wyróżniamy siarkę rombo-wą (inaczej siarkę α) i jednoskośną (siarkę β). W tem-peraturze pokojowej trwała jest siarka α, występująca w postaci jasnożółtych kryształów, która po podgrzaniu do 368,8 K ulega przemianie w siarkę β – występującą w formie żółtych igieł. Warto zauważyć, że te odmia-ny przybierają nie tylko inną postać krystaliczną, lecz
także różnią się właściwościami fizycznymi. Obie for-my stanu stałego po podgrzaniu zamieniają się w siarkę ciekłą, bardzo ruchliwą, jasnożółtą ciecz, składającą się z cząsteczek S8, natomiast w zależności od wyjściowej odmiany alotropowej, topnienie przebiega w innej tem-peraturze. Siarka rombowa topi się już w temperaturze 380 K, podczas gdy temperatura topnienia siarki jed-noskośnej jest aż o 12 K wyższa. Różnicę w tych właś-ciwościach tłumaczy się m.in. powstającą równowagą chemiczną pomiędzy liczbami cząsteczek cyklo-ok-tasiarki, a katena-oktasiarki, czyli siarki łańcuchowej. W warunkach równowagi przyjmuje się, że około 4% cyklo-oktasiarki jest rozrywane z wytworzeniem dłu-gich łańcuchów siarkowych, co stabilizuje jednocześnie jej temperaturę krzepnięcia na poziomie 386 K, a stan ten jest nazywany naturalnym punktem krzepnięcia siarki (Bielański, 2010a).
Przy dalszym ogrzewaniu dochodzi do coraz częst-szego rozrywania pierścieni siarkowych i przeważa forma łańcuchowa cząsteczek S8. Jednak, jak już wspo-mniano wcześniej, siarka ma zdolność do katenacji, co
Ryc. 1. Kryształy siarki (zdjęcie: Mills, 2007)

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 34
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
prowadzi do nadmiernego łączenia się nowopowstałych ośmioatomowych łańcuchów siarkowych ze sobą z wy-tworzeniem długich łańcuchów polimerycznych, zawie-rających aż do 105 atomów siarki (Bielański, 2010a). Siar-kowa ciecz stopniowo w trakcie tego procesu zmienia barwę z jasnożółtej aż do brunatno-czerwonej, zwięk-sza się także około 2000-krotnie jej lepkość, osiągając wartość maksymalną w temperaturze 460,15 K (Bereda, 2011). Tę odmianę siarki nazywamy siarką μ. W proce-sie dalszego ogrzewania, po przekroczeniu temperatury 470 K, poliłańcuchy siarki zaczynają się rozpadać, a jej lepkość maleje.
W stanie gazowym siarka występuje w trzech trwa-łych odmianach alotropowych. Pierwszą z nich, po-wstającą w czasie zmiany stanu skupienia, są żółte pary siarki gazowej złożonej z łańcuchowych cząsteczek S8. Druga powstaje dopiero w temperaturze 1200 K i za-wiera paramagnetyczne cząsteczki siarki dwuatomo-wej. Warto zauważyć, jak bardzo stabilne są połączenia poszczególnych atomów – do rozerwania ośmioelemen-towych łańcuchów potrzebna jest temperatura rzędu 1200 K, a nie jest to rozerwanie całkowite, raczej frag-mentacja. Dopiero w temperaturze 2500 K cząsteczki S2 ulegają rozpadowi na atomy (Bielański, 2010a).
Szereg kolejnych, nietrwałych odmian alotropo-wych siarki tworzonych jest podczas oziębiania jej roztworów. Są to np. pierścienie i łańcuchy sześcio-, siedmio-, dziesięcio-, dwunasto-, osiemnasto- czy dwu-dziestoczłonowe. Oprócz tego istnieje także wiele od-mian metastabilnych siarki.
Siarka jest pierwiastkiem mało reaktywnym w tem-peraturze pokojowej. Po podgrzaniu chętnie łączy się z tlenem tworząc ditlenek siarki SO2, a dalszej fazie utleniania także tritlenek siarki SO3, które w kontakcie z wodą tworzą kwasy tlenowe. Chętnie, a czasem wręcz gwałtownie, reaguje z metalami ziem alkalicznych, two-rzy także połączenia z metalami szlachetnymi – często
powstają nierozpuszczalne siarczki. Łatwo łączy się z fluorem w różnych stosunkach stechiometrycznych, m.in. 1:6 (SF6), a takie połączenie jest możliwe dzięki zaangażowaniu orbitali d i hybrydyzacji sp3d2 atomu siarki (Bielański, 2010b). Z wodorem siarka reaguje do-piero w podwyższonej temperaturze. Z azotem tworzy m.in. poliazyle – związki o stechiometrii (SN)x, gdzie xϵN, polimery o budowie łańcuchowej, składające się z dwóch atomów niemetali, które, dzięki delokalizacji elektronów pomiędzy atomami siarki i azotu w tempe-raturze bliskiej 4 K, przewodzą prąd elektryczny, a więc zachowują się jak metale. Co ciekawe, po schłodzeniu ich do temperatury 0,26 – 0,33 K nabierają właściwości nadprzewodzących (Bielański, 2010d).
Znaczenie i występowanie siarki i jej związków nieorganicznych
Związki powstałe z połączenia siarki z tlenem czy wodorem są substancjami gazowymi, często występują-cymi w powietrzu. Tlenki siarki: SO2 i SO3 przedostają się do atmosfery podczas spalania, np. węgla kamien-nego czy ropy naftowej, a także wskutek wybuchów wulkanów, powodując jej zanieczyszczenie. Produkty ich reakcji z wodą czy parą wodną, czyli kwasy siarko-we (IV) i (VI), powodują znaczne obniżenie pH wody, a w konsekwencji są główną przyczyną negatywnych skutków kwaśnych deszczy. Tlenki siarki wchodzą również w skład smogu. Siarkowodór jest gazem silnie trującym o bardzo charakterystycznym zapachu, prze-dostającym się do atmosfery wskutek wybuchów wul-kanów. Do jego produkcji przyczynia się także prze-mysł chemiczny (Bielański, 2010c) i mikroorganizmy (np. bakterie redukujące siarczany). W określonych stę-żeniach jest śmiertelnie niebezpieczny dla organizmów, bowiem inaktywując oksydazę cytochromową, hamuje działanie łańcucha oddechowego (Cooper i wsp., 2008).
Toksyczne stężenie wynosi ok. 1000 – 2000 ppm. Prze-prowadzone niedawno badania na myszach wykaza-ły, że w mniejszych stężeniach siarkowodór powoduje spowolnienie metabolizmu, ale nie wywołuje trwałych uszkodzeń (Bełtowski, 2004).
Tritlenku siarki, SO3, nie da się bezpośrednio otrzy-mać w procesie spalania siarki w tlenie, jak to ma miej-sce w przypadku SO2, powstaje on wskutek utlenienia tlenku siarki (IV). W kontakcie z wodą tworzy kwas siarkowy (VI) – substancję żrącą. Kwas siarkowy (IV), zwany kwasem siarkawym, powstaje wskutek reakcji tlenku siarki (IV) z wodą tylko w niewielkim stopniu. Ze względu na to, że SO2 pozostaje częściowo niezwią-zany z wodą (jest przez nią solwatowany), po reakcji nie otrzymujemy wyłącznie kwasu siarkawego i produktów jego dysocjacji, ale także SO2 i S2O5
2- (Bielański, 2010c).Oprócz wyżej wymienionych związków, siarka two-
rzy jeszcze wiele innych, np. tlenki S2O, SO, SO4 czy kwasy H2S2O7, H2S2O6, H2SO5, H4S2O8. Są to substan-cje rzadko spotykane, głównie syntetyczne, dlatego nie będą one dalej omawiane.
Siarka występuje w środowisku nie tylko w powie-trzu w formie gazów (SO2, SO3) i w środowiskach wod-nych w stanie ciekłym (H2SO3, H2SO4; jako ich jony), ale także w glebie i skałach. Można ją spotkać zarówno w postaci rodzimej (pierwiastkowej) – jest wydobywa-na także w Polsce w okolicach Tarnobrzega, jak i w po-staci minerałów, zarówno siarczków, jak i siarczanów (Pajdowski, 1976). Powszechnie znane są i wydobywane blenda cynkowa (ZnS), piryt (FeS2), chalkopiryt (Cu-FeS2), markasyt (FeS2) i galena ołowiana (PbS) oraz gips (CaSO4·2H2O), anhydryt (CaSO4), baryt (BaSO4) i cele-styn (SrSO4) (Chmielewski i wsp., 2003). Siarka rodzima jest także ważną domieszką kopalnianego węgla ka-miennego i brunatnego, gazu ziemnego i ropy naftowej (Bereda, 2011), dlatego nietrudno ją znaleźć w pobliżu kopalni. Siarka pierwiastkowa, siarczany i siarczki są

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 35
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
także obecne w osadach wód słonych, szczególnie oce-anów, a S0 i S2- są kumulowane, a następnie uwalniane w wodach gejzerów (Madsen, 2008).
Znaczenie i występowanie związków organicznych siarki
Siarka jest jednym z pierwiastków biogennych wcho-dzących w skład organizmów. Mimo, że stanowi ona je-dynie ok. 1% masy komórki, jest ona bardzo ważna dla jej prawidłowego rozwoju i funkcjonowania (Eichhorn, 2000). Atomy tego pierwiastka występują: (i) w dwóch aminokwasach – L-metioninie i L-cysteinie (a także w białkach zawierających te aminokwasy), (ii) kofak-torach enzymów, takich jak biotyna, tiamina, kwas li-ponowy, koenzym A czy molibdopteryna (iii) w S-ade-nozylometioninie, (iv) w tionukleozydach wchodzących w skład zmodyfikowanego tRNA, (v) białkach zawie-rających centra Fe-S oraz (vi) w alifatycznych sulfonia-nach, np. taurynie. Ponadto bakterie potrafią syntetyzo-wać takie związki siarki jak ergotioneina czy owotiol A, a także disiarczek glutationu.
Aminokwasy
Aminokwasy są związkami organicznymi, zawie-rającymi grupę aminową i grupę karboksylową. Wza-jemne ułożenie tych grup względem siebie określa typ aminokwasu: jeśli zarówno grupa –NH2, jak i –COOH są połączone z tym samym atomem węgla, mamy do czynienia z α-aminokwasem. Dzięki obecności obu tych grup funkcyjnych, aminokwasy klasyfikowane są jako związki amfoteryczne – czyli zdolne zarówno do reakcji z kwasami (poprzez grupę aminową), jak i z za-sadami (poprzez grupę karboksylową). W roztworze wodnym aminokwasy istnieją głównie w postaci zjoni-zowanej – jonów obojnaczych. Białka budowane są wy-
łącznie przez 20 aminokwasów należących do szeregu L (McMurry, 2005).
Znane są dwa aminokwasy białkowe zawierające siarkę – cysteina i metionina. W cysteinie występuje ona w grupie tiolowej, natomiast w metioninie w grupie sulfidowej (ryc. 2). Metionina jest pierwszym amino-kwasem, od którego zazwyczaj rozpoczyna się synteza nowego białka. Jest kodowana tylko przez jeden kodon – AUG, zwany kodonem start, od którego zaczyna się elongacja łańcucha polipeptydowego, co czyni ją bardzo ważnym elementem w syntezie białek. Ponadto może być donorem grupy metylowej w reakcjach biosyntezy tłuszczy lub innych ważnych biologicznie związków (Or-Rashid i wsp., 2001). Dwie cysteiny, poprzez utle-nienie wchodzących w skład ich struktury grup tiolo-wych, tworzą dimer zwany cystyną (ryc. 3). Taki mo-stek disulfidowy, nazywany popularnie disiarczkowym, może się tworzyć nie tylko z udziałem pary cystein, któ-re znajdują się bezpośrednio obok siebie w łańcuchu po-lipeptydowym, ale także z udziałem dwóch cząsteczek występujących w różnych łańcuchach (ryc. 4). Tak więc, cysteina, oprócz swojej roli budulcowej, ma także inne bardzo ważne zadanie – w stabilizacji trzecio- i czwar-torzędowej struktury białek (Berg i wsp., 2007). Siarkę zawiera także homocysteina – α,L-aminokwas niebiał-kowy, strukturalnie przypominający cysteinę, od któ-
rej różni się obecnością dodatkowej grupy metylenowej w łańcuchu węglowym. Związek ten może zostać wy-korzystany przez bakterie do syntezy metioniny (Or--Rashid i wsp., 2001).
Kofaktory enzymów
Wskutek dekarboksylacji cysteiny powstaje cy-steamina, która w dalszych przemianach prowadzi do powstania koenzymu A (ryc. 5a). Związek ten z kolei odgrywa bardzo ważną rolę biologiczną – jest pośred-nikiem w przenoszeniu grup acylowych. Po połączeniu koenzymu A z grupą acylową powstaje acylo-koenzym A, który umożliwia transport grup acylowych w orga-nizmie. Co jest bardzo istotne, połączenie to powstaje wskutek acylowania grupy tiolowej koenzymu A, a więc
Ryc. 2. Struktury aminokwasów siarkowych: a. cysteina, b. metionina
Ryc. 3. Reakcja tworzenia cystyny
Ryc. 4. Przykłady mostków disulfidowych
+ 2e-

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 36
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
grupy zawierającej siarkę. Acetylo-koenzym A, czy-li związek powstały w wyniku acylowania koenzymu A kwasem bądź bezwodnikiem octowym, pełni bardzo ważną rolę w metabolizmie – przenosi grupę acetylo-wą na szczawiooctan, rozpoczynając tym samym cykl kwasu cytrynowego. Bierze także udział w aktywacji kwasów tłuszczowych i ich rozkładzie. Cząsteczki acy-lo-koenzymu A są substratami w reakcjach β-oksydacji kwasów tłuszczowych (Murray i wsp., 2006a).
Innym ważnym organicznym związkiem zawierają-cym siarkę jest kwas liponowy (ryc. 5b) – pełniący, po aminacji i utlenieniu, rolę kofaktora dla dehydrogenazy pirogronianowej w reakcji dekarboksylacji oksydacyj-
nej pirogronianu. Znane są także dwie witaminy zawie-rające siarkę: biotyna i tiamina. Biotyna, znana także witaminą H lub B7, jest koenzymem niektórych enzy-mów katalizujących reakcje karboksylacji – dołączenia cząsteczki CO2. Tiamina, czyli witamina B1, pełni waż-ną rolę w przemianie węglowodanów. Jest koenzymem kilku dekarboksylaz, a difosforan tiaminy – transke-tolazy w szlaku pentozofosforanowym (Murray i wsp., 2006b).
Molidbopteryna jest niebiałkowym składnikiem miejsc aktywnych niektórych enzymów, takich jak na przykład reduktaza azotanowa czy dehydrogenaza ksantyninowa, niezbędnym do prawidłowego ich dzia-łania. W jej cząsteczce obecne są dwa atomy siarki, oba połączone są z atomem molibdenu (ryc. 6) (Hille, 2002).
Białka żelazowo-siarkowe (białka z żelazem niehemowym)
Białka żelazowo-siarkowe występują praktycznie we wszystkich organizmach, zawierają one charakte-rystyczną grupę prostetyczną: centrum żelazowo-siar-kowe. Najprostsze takie centrum znajduje się w rubre-doksynie i ma strukturę tetraedru: żelazo połączone jest wiązaniami koordynacyjnymi z czterema liganda-mi – resztami cysteiny (ryc. 7a). Najczęściej w białkach
spotykana jest jednak płaska struktura rombowa Fe2S2. W skład tej struktury wchodzą dwa atomy centralne – atomy żelaza na II stopniu utlenienia – połączone dwo-ma mostkami siarkowymi, każdy atom centralny połą-czony jest także z dwiema resztami cysteiny (ryc. 7b). Pozostałe, przedstawione w ryc. 7 oraz inne, bardziej skomplikowane, struktury powstają wskutek połącze-nia płaskich „komórek” rombowych (Kiley i wsp., 2003).
Centra żelazowo-siarkowe są częścią wielu waż-nych enzymów, głównie z grupy oksyreduktaz. Obecne w nich żelazo (II) może zostać utlenione do żelaza (III), a w reakcji odwrotnej, Fe (III) zredukowane do Fe (II). Reakcje te umożliwiają transport elektronów. Centra Fe-S, spotykane są także w miejscach aktywnych nie-których enzymów, gdzie pełnią kluczową rolę w wiąza-niu substratu poprzez polaryzację jego grup funkcyj-nych. Zaobserwowano, że centra żelazowo-siarkowe są donorami elektronu, który może zainicjować przebieg reakcji wolnorodnikowych w organizmach (Beinert,
Ryc. 5. Struktury koenzymów: a. koenzymu A, b. kwasu liponowego
Ryc. 6. Struktura molibdopteryny
Ryc. 7. Struktury najbardziej rozpowszechnionych centrów żelazowo-siarkowych występujących w białkach: a. tetraedr (rubredoksyna), b. struktura płaska rombowa typu [2Fe-2S], c. struktura przestrzenna typu [3Fe-4S], d. struktura sześcienna typu [4Fe-4S] (Kiley i wsp., 2003)

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 37
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
2000). Ponadto trzy transkrypcyjne aktywatory E. coli: SoxR, IscR oraz FNR zawierają w swojej strukturze centra żelazowo-siarkowe, które pełnią kluczową rolę w ich funkcji regulatorowej polegającej na uruchomie-niu transkrypcji wybranych genów. SoxR w warunkach normalnych jest nieaktywny, natomiast w obecno-ści np. O2
- lub NO aktywuje ekspresję białek ochrony przed stresem oksydacyjnym, naprawy DNA i innych. FNR jest globalnym regulatorem u E. coli, a IscR jest regulatorem operonu kodującego geny biorące udział w składaniu centrów Fe-S (Kiley i wsp., 2003). Centra żelazowo-siarkowe występują także w zlokalizowanych w błonach centrach reakcji fototrofów, w których ener-gia światła jest przekształcana w energię chemiczną w postaci ATP (Erickson, 1992).
Tauryna i inne alifatyczne sulfoniany
Sulfoniany to sole kwasów sulfonowych, które za-wdzięczają swój kwasowy charakter obecności dwóch wiązań S=O, a w nieznacznym stopniu reszcie węglo-wodorowej. W taurynie (ryc. 8), czyli kwasie 2-amino-etanosulfonowym, atom siarki jest częścią ugrupowania sulfonianowego, ale znane są także przypadki przyswa-jania przez bakterie estrów i kwasów sulfonowych oraz kwasów sulfaminowych (Van der Ploeg i wsp., 1996).
Zarówno tauryna, jak i wiele innych alifatycz-nych sulfonianów, takich jak koenzym M, taurocholan czy metanosulfonian, występuje naturalnie w środo-wisku. Tauryna występuje w organizmie zwierząt jako
składnik kwasu taurocholowego stanowiącego ok. 1/3 kwasów żółciowych ulegających syntezie w wątrobie. Koenzym M jest jednym z kofaktorów metanogenezy u archeonów metanogennych, metanosulfonian po-wstaje w atmosferze wyniku fotochemicznego utlenie-nia siarczku dimetylu (Eichhorn, 2000), a octanosul-fonian jest składnikiem endospor u Bacillus subtilis (Bonsen i wsp., 1969).
Istnieją także, głównie w środowiskach wodnych, organiczne związki siarki pochodzenia antropogen-nego. Są one na przykład syntetycznymi składnikami używanych przez ludzi detergentów, wybielaczy czy składnikami cementu, mowa tu głównie o alifatycz-nych i aromatycznych sulfonianach, których obecność stwierdzono w rzekach i osadach ściekowych (Kertesz, 1999). Niektóre bakterie wykształciły zdolność wyko-rzystywania tych związków jako źródła węgla, siarki i energii.
Wykorzystanie przez bakterie związków nieorganicznych siarki jako źródła pierwiastka budulcowego
Większość mikroorganizmów przyswaja siarkę do celów budulcowych w postaci siarczanów, siarczynów i tiosiarczanów w szlaku zwanym szlakiem biosyntezy cysteiny, gdzie przy pomocy licznych enzymów, trans-portowane ze środowiska zewnętrznego nieorganiczne związki siarki są redukowane i siarka jest wbudowy-wana do O-acetyloseryny z wytworzeniem L-cysteiny. Mechanizm ten został dokładnie poznany u E. coli i Sal-monella enterica sv. Typhimurium (Kredich, 1996). Cy-steina jest następnie używana nie tylko do syntezy bia-łek, ale także jest substratem do syntezy wielu ważnych związków organicznych zawierających siarkę. Ponadto niektóre mikroorganizmy posiadają także zdolność asymilacji siarczków (Dick, 1992), które mogą włączyć
się w siarczanową gałąź szlaku biosyntezy cysteiny w jej ostatnim etapie, rzadziej spotykana, ale również możli-wa, jest asymilacja siarki pierwiastkowej (Kessler, 2006).
Szlak biosyntezy cysteiny
Pierwszym krokiem w każdej ścieżce asymilacji siar-ki jest transport jej związków ze środowiska zewnętrz-nego do wnętrza komórki. Komórkom niezbędne są produkty genów cysA, cysP, cysT oraz cysW zorgani-zowanych w operonie cysPTWA-M (Eichhorn, 2000), a także produktu genu sbp. Białka kodowane przez geny cysT i cysW tworzą kanał transportowy w błonie zewnętrznej, przez który związki siarki (siarczany i tio-siarczany) znajdujące się w środowisku zewnętrznym mogą przedostawać się do wnętrza komórki. Białko CysA jest wbudowywane w błonę cytoplazmatyczną, natomiast białka Sbp oraz CysP, które wiążą odpowied-nio siarczany i tiosiarczany, występują w przestrzeni pe-ryplazmatycznej (Kredich, 1996).
Asymilacja siarczanu
Przetransportowany do cytoplazmy siarczan musi być aktywowany przed redukcją. Produktami ekspresji genów cysC, cysD i cysN (zorganizowanych w operonie cysDNC), są enzymy uczestniczące w dwóch etapach tej aktywacji: sulfurylaza ATP (cysD, cysN; E.C. 2.7.7.4) oraz kinaza APS (cysC; E.C. 2.7.1.25) (Eichhorn, 2000). Proces asymilacji został przedstawiony na ryc. 11a.
Początkowo siarczan jest aktywowany do 5’-fosfo-siarczanu adenozyny (APS) (ryc. 9) w reakcji katali-zowanej przez sulfurylazę ATP. Drugim, obok SO4
2-, substratem tej reakcji jest jedna cząsteczka wysokoener-getycznego 5’-trifosforanu adenozyny (ATP). W kolej-nym kroku kinaza APS katalizuje przeniesienie grupy fosforanowej z kolejnej cząsteczki ATP na APS z utwo-rzeniem 3’-fosfoadenozyno-5’-fosfosiarczanu (PAPS) Ryc. 8. Struktura tauryny

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 38
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
(ryc. 10), a sulfotransferaza PAPS (E.C. 1.8.99.4) prze-nosi grupę siarczanową na akceptor, którym najczęściej jest tioredoksyna. Związek ten zawiera dwie sąsiadujące ze sobą cysteiny, do jednej z nich przyłącza się grupa siarczanowa. Nowoutworzona tioredoksyna-S-SO3
- jest donorem siarczynu, a w procesie przekształcenia po-wstaje także utleniona tioredoksyna (reszty cysteiny tworzą mostek disiarczkowy), która jest regenerowana do formy zredukowanej przez reduktazę tioredoksyny (Kredich, 1996).
Sulfotransferaza PAPS jest produktem genu cysH, należącego do kolejnego operonu cysteinowego cysIJH. Pozostałe dwa geny tego operonu, cysI i cysJ oraz nie-zależny gen cysG kodują reduktazę siarczynową (E.C.
1.8.1.2), katalizującą zależną od NADPH redukcję SO3
2- do siarcz-ku (Kredich, 1996). Enzym ten ma budowę podjednostkową o stechio-metrii α8β4, gdzie α jest flawopro-teiną (cysJ), a β jest hemoprotei-ną (cysI). Flawoproteina – dzięki obecności dinukleotydu (FAD) oraz mononukleotydu (FMN) fla-winoadeninowego – spełnia rolę akceptora elektronów i przenosi je na hemoproteinę, zawierającą cen-tra Fe4S4 oraz charakterystyczną dla reduktazy azotynowej i siarczy-nowej grupę prostetyczną, okre-ślaną w literaturze anglojęzycznej jako siroheme. Gen cysG koduje enzym niezbędny do syntezy tego związku (Ostrowski i wsp., 1989). Ostatnim etapem szlaku jest prze-kształcenie siarczku w cysteinę, w którym bierze udział prekursor cysteiny, O-acetyloseryna. Powsta-je ona w reakcji przeniesienia gru-py acetylowej z acetylokoenzymu A na L-serynę katalizowanej przez transacetylazę serynową (E.C. 2.3.1.30; cysE). Synteza cysteiny odbywa się z udziałem syntazy cysteinowej A, kodowanej przez gen cysK, będącej częścią kompleksu syntaz cysteinowych (E.C. 4.2.99.8) (Kredich, 1996).
Asymilacja tiosiarczanu
Pozyskiwany ze środowiska tiosiarczan może być także źródłem siarki dla bakterii (ryc. 11b). Pierwszym etapem jego asymilacji jest, podobnie jak w przypadku asymilacji siarczanu, transport do wnętrza komórki. Następnie, kodowana przez gen cysM syntaza cysteino-
wa B, katalizuje syntezę S-sulfocysteiny z O-acetylosery-ny i tiosiarczanu. S-sulfocysteina zostaje przekształcona w L-cysteinę, a w reakcji tej uwalniany jest siarczyn, któ-ry bakterie mogą dalej przekształcić w siarczek i cyste-inę (Eichhorn, 2000). Ta gałąź szlaku biosyntezy cyste-iny jest znacznie krótsza i korzystniejsza energetycznie – przyswajanie siarczanów wymaga obecności dwóch cząsteczek ATP, zredukowanej tioredoksyny, NADPH i O-acetyloseryny, natomiast do asymilacji tiosiarczanu potrzebny jest tylko ostatni z wymienionych związków.
Ryc. 10. Struktura 3’-fosfoadenozyno-5’-fosfosiarczanu (PAPS)
Ryc. 11. Ścieżka biosyntezy cysteiny u E. coli i S. enterica sv. Typhimurium: a. biosynteza z siarczanu, b. biosynteza z tiosiarczanu. APS – 5’-fosfosiarczan adenozyny, PAPS – 3’-fosfoadenozyno-5’-fosfosiarczan
Ryc. 9. Struktura 5’-fosfosiarczanu adenozyny (APS)

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 39
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Asymilacja siarczynu i siarczku
Pozyskiwany w trakcie asymilacji tiosiarczanu jon SO3
2-, jak już wcześniej wspomniano, może także zostać wykorzystany do biosyntezy cysteiny. Jego cząsteczki występujące w komórkach są wychwytywane przez za-leżną od NADPH reduktazę siarczynową i przekształ-cane w siarczek, a następnie w cysteinę.
Podobnie jest w przypadku samego siarczku, który może włączyć się w siarczanową gałąź szlaku w jej ostat-nim etapie (ryc. 11a). Biosynteza cysteiny w obu tych przypadkach jest taka sama jak ta opisana wyżej, z po-minięciem etapów poprzedzających biosyntezę związ-ków startowych – siarczynu i siarczku. Przykładowymi mikroorganizmami zdolnymi do wykorzystania siarcz-ków są Acidithiobacillus spp. i Acidiphilium spp. (Dick, 1992), którym specjalne systemy transportu siarczków ze środowiska do wnętrza komórki umożliwiają asymi-lację S2-. Siarczki są przez Acidithiobacillus spp. (bezbar-wna bakteria siarkowa) wykorzystywane również jako źródło energii i donor elektronów.
Regulacja biosyntezy cysteiny
Szybko zmieniające się warunki środowiska, w ja-kim przyszło żyć bakteriom, zmusiły je do wykształ-cenia bardzo precyzyjnych mechanizmów regulacji genów odpowiedzialnych za adaptację do środowiska i jak najlepsze wykorzystanie dostępnych metabolitów. Biosynteza cysteiny u E. coli i S. enterica sv. Typhimu-rium zostaje prawie natychmiast zahamowana, gdy bakterie zostaną przeniesione do pożywki zawierającej ten aminokwas (Kredich, 1996). Dzieje się tak dlatego, że wolna L-cysteina jest inhibitorem transacetylazy se-rynowej odpowiedzialnej za syntezę O-acetyloseryny, która ulega przegrupowaniu do N-acetyloseryny. Za ten i inne procesy regulacyjne szlaku biosyntezy cysteiny
odpowiedzialny jest system modulacji ekspresji genów zwany regulonem cys (ryc. 12).
Jak już wcześniej wspomniano, dostępność cystei-ny hamuje proces syntezy O-acetyloseryny, a w konse-kwencji zahamowana zostaje asymilacja siarczanu i tio-siarczanu. Ponadto dostępność w komórkach siarczku i tiosiarczanu hamuje ich transport do wnętrza komór-ki. Reasumując, L-cysteina, tiosiarczan lub siarczek są negatywnymi regulatorami biosyntezy cysteiny (Kre-dich, 1996).
Asymilacja siarki elementarnej
Niektóre mikroorganizmy, oprócz możliwości po-zyskiwania siarki do celów budulcowych z jej nieorga-nicznych związków, takich jak siarczany, siarczyny, tio-siarczany czy siarczki, mają także zdolność asymilacji siarki pierwiastkowej (Kessler, 2006). Mowa tu m.in. o Acidithiobacillus thiooxidans czy A. ferrooxidans, które przy pomocy syntetyzowanych odpowiednich dioksygenaz są zdolne do utleniania siarki. Uważa się, że enzym dioksygenaza siarkowa (E.C. 1.13.11.18) ka-talizuje reakcję przekształcenia siarki elementarnej do siarczynu zgodnie z równaniem reakcji:
Jak wspomniano wcześniej, siarka w warunkach na-turalnych występuje w postaci ośmioatomowych pier-ścieni, które tworzą rombowy kryształ, dlatego postulu-je się, że przed zajściem powyższych reakcji siarka musi być aktywowana przez nukleofil, na przykład glutation, a jej pierścień musi zostać otwarty (Rohwerdert i wsp., 2003). Glutation jest syntetyzowany praktycznie w każ-dej komórce, w obecności siarki elementarnej tworzy disiarczek glutationu, a ten w reakcji następczej jest przekształcany w siarczyn:
Jak widać z powyższych równań reakcji, glutation nie jest zużywany w reakcji, odgrywa jedynie rolę ak-tywatora. Powstały w wyniku tych reakcji siarczyn jest włączany w szlak biosyntezy cysteiny (Kessler, 2006).
Ryc. 12. Interakcje pomiędzy poszczególnymi etapami szlaku biosyntezy cysteiny, regulon cysteinowy
L-cysteina jest inhibitorem współza-wodniczącym z L-seryną o miejsce aktywne transacetylazy serynowej (*), tiosiarczan i siarczek są inhibitorami aktywacji własnego transportu, a O-acetyloseryna jest prekursorem N-acetyloseryny odpowiedzialnej za ak-tywację transportu i redukcji siarczanu.

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 40
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Cysteina i metionina jako prekursory wielu ważnych związków organicznych
Bakterie potrafią wykorzystać cysteinę nie tylko do budowy białek, ale także ją przekształcić lub wy-korzystać jako donor siarki dla wielu innych ważnych dla ich funkcjonowania związków chemicznych, takich jak metionina, glutation i jego pochodne, koenzym A, S-adenozylometionina, a także L-ergotioneina i owotiol A (ryc. 13), których zadaniem jest inaktywacja takich związków jak OH· czy H2O2 powstających w czasie stre-su oksydacyjnego (Kessler, 2006).
Cysteina jest jednym z trzech aminokwasów bu-dujących ważny biologicznie i odporny na degradację enzymatyczną tripeptyd – glutation (GSH), pozosta-łe dwa to glicyna i glutaminian. Swoją unikatowość glutation zawdzięcza nietypowemu dla związków biologicznych wiązaniu izopeptydowemu, które jest tworzone przez grupę α-aminową cysteiny z grupą γ-karboksylową glutaminianu w obecności enzymu syntazy γ-glutaminocysteiny (E.C. 6.3.2.2.) – jest to pierwszy z etapów biosyntezy GSH. W kolejnym kro-ku, enzym syntetaza glutationu (E.C. 6.3.2.3.) katalizuje przyłączenie glicyny do wcześniej wspomnianego di-peptydu (Anderson, 1998). W tym przypadku cysteina nie jest jedynie donorem siarki, ale jedną ze składowych całego tworzonego związku. Glutation pełni bardzo ważną funkcję w asymilacji siarki pierwiastkowej – ak-tywuje i rozrywa jej pierścień tworząc disiarczek gluta-tionu, który następnie jest rozkładany z wydzieleniem siarczynu.
Podobnie jest w przypadku metioniny – szlak jej biosyntezy również zaczyna się od cysteiny (ryc. 14). W pierwszym etapie cysteina w reakcji z O-bursztyny-lo-homoseryną katalizowanej przez γ-syntazę cysta-tioninową (E.C. 2.5.1.48) przekształca się w cystatio-ninę, która następnie zostaje zamieniona przez enzym β-liazę cystationinową (E.C. 4.4.1.8) w homocysteinę.
Powstały w ten sposób niebiałkowy aminokwas, który jest bezpośrednim prekursorem metioniny, ulega mety-lacji w obecności syntazy metioninowej (E.C. 2.1.1.13). Ostatni z wymienionych enzymów do swej aktywności wymaga kofaktora, jakim jest witamina B12 (Or-Rashid i wsp., 2001).
Istnieją gatunki, na przykład Mycobacterium tu-berculosis (Wheeler i wsp., 2005) czy Pseudomonas pu-tida (Vermeij i wsp., 1999), które potrafią wykorzystać metioninę jako jedyne źródło siarki budulcowej. Asy-milacja metioniny polega na jej przekształceniu (przy użyciu odpowiednich enzymów) w cysteinę w procesie odwrotnym do przedstawionego na ryc. 14 (Wheeler i wsp., 2005). Prokariota wykorzystują metioninę także do syntezy S-adenozylometioniny – substratu dla wielu transferaz. Związek ten jest donorem grup metylowych
uczestniczących w procesach transmetylacji różnych związków chemicznych, na przykład kwasów nukleino-wych i białek (Chiang i wsp., 1996).
Disiarczki jako prekursory wielu ważnych związków organicznych
Duża grupa ważnych biologicznie związków che-micznych zawierających siarkę jest syntetyzowana przez mikroorganizmy z użyciem wielosiarczków, naj-częściej disiarczków (ang. persulfide). Należą do nich bez wątpienia centra żelazowo-siarkowe, tiamina, bio-tyna, kwas liponowy, a także tionukleozydy (ryc. 15).
W biosyntezie centrów Fe-S biorą udział enzymy, takie jak na przykład NifS, uczestniczący w biosyn-tezie centrów żelazowo-siarkowych nitrogenazy, czy też IscU, znaleziony u E. coli, scharakteryzowane jako
Ryc. 13. Przykłady związków syntetyzowanych z udziałem cysteiny

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 41
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
desulfurylazy cysteinowe. Enzym NifS, w obecności kofaktora – fosforanu pirydoksocysteiny, odszcze-pia od tego kofaktora ugrupowanie tiolowe i przenosi je na znajdującą się w jego centrum aktywnym resztę cysteiny z wytworzeniem disiarczku (Kessler, 2006). Disiarczki są niestabilne, ulegają spontanicznemu roz-padowi z wytworzeniem łańcuchów siarki pierwiastko-wej, która, w przypadku centrów żelazowo-siarkowych, jest substratem do ich biosyntezy (Beinert, 2000). Dzia-łanie drugiego z wymienionych enzymów jest bardzo podobne.
Rola wielosiarczków w biosyntezie tiaminy, bioty-ny, kwasu liponowego i tionukleozydów jest taka sama, jak w przypadku centrów Fe-S. Białka, takie jak biorące udział w tworzeniu tiaminy ThiS czy aktywujący siar-
kę do syntezy tionukleozydów IscS, w centrum katalitycznym tworzą wielosiarczki, które są donorami siarki w odpowiednich procesach biologicznych.
Niektóre disiarczki mogą być wykorzystywane przez bakterie jako alternatywne źródła siarki bu-dulcowej. Na przykład Acidithioba-cillus spp. i Acidiphilium spp. mogą przyswajać disiarczek glutationu, a Chromatium sp. wykorzystywać jego amid. Uważa się, że amid ten może także brać udział w transferze siarki pierwiastkowej ze środowi-ska do komórek (Kessler, 2006). Disiarczki mogą być, w razie potrzeby, wytwarzane przez niektóre bakterie, na przykład w obecności siarki elementarnej, szczepy należące do rodzaju Acidithiobacillus czy Acidiphilium, syntetyzują disiarczek glutationu (GSSH), który może być wykorzystywany przez mikroorganizmy jako alter-natywne źródło siarki do celów budulcowych.
Inne sposoby wykorzystania siarki i jej związków
Związki siarki występują w atmosferze, litosferze, hydrosferze oraz pedosferze i występują we wszystkich trzech stanach skupienia. Jak już wcześniej wspomnia-no, siarka jest jednym z budulcowych pierwiastków biogennych, niezbędnym do prawidłowego funkcjono-wania każdej komórki. Jest składnikiem dwóch białko-wych aminokwasów – cysteiny i metioniny, licznych kofaktorów, takich jak biotyna, tiamina, molibdoptery-na, kwas liponowy czy koenzym A, tionukleozydów i, jako kluczowy składnik centrów żelazowo-siarkowych, buduje centra aktywne niektórych ważnych enzymów. Mostki disiarczkowe tworzone przez dwie reszty cy-steiny stabilizują trzecio- i czwartorzędową strukturę
białek zewnątrzkomórkowych występujących przede wszystkim w warunkach utleniających. Każdy z wyżej wymienionych związków pełni bardzo ważną funkcję we wszystkich organizmach. Prokarioty nie wykorzy-stują jednak siarki i jej związków jedynie do celów bu-dulcowych, jak to ma miejsce u eukariotów. Niektóre bakterie chemolitotroficzne i archeony potrafią je utle-niać, uzyskując w ten sposób energię niezbędną do syn-tezy ATP, a inne wykorzystywać jako donor lub końco-wy akceptor elektronów.
Do pierwszej z wymienionych grup bez wątpienia należą bezbarwne bakterie siarkowe, takie jak na przy-kład Beggiatoa spp., Thiobacillus spp., Halothiobacillus spp. czy Thiovulum majus (Kelly i wsp., 2006), które można znaleźć praktycznie wszędzie, a szczególnie ob-ficie występują w wodach morskich i gorących źródłach. Pozyskują one energię (oraz elektrony niezbędne do wiązania dwutlenku węgla) w procesie utleniania siarki pierwiastkowej, a także siarczków, siarczynów, tiosiar-czanów, tetrationianów, rodanków, siarczku dimetylu czy też dwusiarczku węgla. Produktem końcowym tego utleniania jest siarczan (Dick, 1992). Siarka i jej związki są też donorami elektronów w procesie asymilacji dwu-tlenku węgla.
Ryc. 14. Szlak biosyntezy metioniny
Źródło: Grundy i wsp., 1998, zmodyf.
Ryc. 15. Przykłady związków syntetyzowanych z wykorzystaniem disiarczków jako źródła siarki; R-S-SH – disiarczek

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 42
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Druga grupa mikroorganizmów prokariotycznych wykorzystujących siarkę i jej związki (siarczki i tiosiar-czany) jako donor elektronów niezbędnych do wiązania CO2. Są to fototrofy anoksygeniczne należące do typów Proteobacteria, Chlorobi i Chloroflexi. Występują one w głębszych strefach jezior i gorących źródłach, różnią się między sobą sposobem przeprowadzania fotosynte-zy, barwnikami uczestniczącymi w tym procesie oraz miejscem w toni wodnej (głębokość), w której mogą rosnąć (Madigan i wsp., 2009).
Ostatnią grupę tworzą bakterie redukujące siar-czany, które wykorzystują siarczany jako ostateczny akceptor elektronów (proces dysymilacyjnej redukcji siarczanów). Należą do nich bakterie z rodzajów takich jak Desulfovibrio, Desulfobacter, Desulfococcus, Desul-fobulbus (typ Proteobacteria), Thermodesulfobacterium (typ Termodesulfobacteria) czy Desulfotomaculum (typ Firmicutes), i są najczęściej spotykane w beztlenowych strefach gleb i wód, ściekach oraz przewodach pokar-mowych ludzi i zwierząt. Znane są także bakterie, które potrafią wykorzystać w oddychaniu siarkę pierwiast-kową jako akceptor elektronów (np. Geobacter sul-furreducens) (Rabus i wsp., 2006). Wymienione wyżej prokarioty przeprowadzają zatem unikatowe reakcje biochemiczne w obiegu siarki, w którym uczestniczą również mikroorganizmy wykorzystujące siarkę i jej związki do celów budulcowych. Obieg siarki obejmuje również procesy fizyczne i chemiczne, a najważniejszy-mi dla niego przemianami są utlenianie, redukcja, mine-ralizacja i immobilizacja związków siarki (Dick, 1992).
Literatura
Anderson ME (1998). Glutathione: an overview of biosynthesis and modulation. Chem Biol Interact. 111-112:1-14.
Beinert H (2000). Iron-sulfur proteins: acient structures, still full of surprises. J Biol Inorg Chem.5:2-15.
Bełtowski J (2004). Hydrogen sulfide as a biologically active mediator
in the cardiovascular system. Postępy Hig Med Dosw. 58:285-291.Bereda T (2011). Siarka. W: Bilans zasobów kopalin i wód podziem-
nych w Polsce, 1. wydanie. Szuflicki M, Malon A, Tymiński M, (red.)., Argraf, 100-103.
Berg JM, Tymoczko JL, Stryer L (2007). Struktura pierwszorzędo-wa: Aminokwasy połączone wiązaniami peptydowymi tworzą łańcuchy polipeptydowe. W: Biochemia, Wydawnictwo Naukowe PWN, 51-54.
Bielański A (2010a). Odmiany alotropowe siarki. W: Podstawy chemii nieorganicznej, 6. wydanie, Wydawnictwo Naukowe PWN, 613-617.
Bielański A (2010b). Właściwości chemiczne tlenowców oraz Związ-ki tlenowców z fluorowcami. W: Podstawy chemii nieorganicznej, 6. wydanie, Wydawnictwo Naukowe PWN, 618-619, 632-634.
Bielański A (2010c). Związki tlenowców z wodorem. W: Podsta-wy chemii nieorganicznej, 6. wydanie, Wydawnictwo Naukowe PWN, 619-632.
Bielański A (2010d). Związki azotowców z siarką. W: Podstawy che-mii nieorganicznej, 6. wydanie, Wydawnictwo Naukowe PWN, 707-711.
Bonsen PPM, Spudich JA, Nelson DL, Kornberg A (1969). Biochemi-cal studies of bacterial sporulation and germination. XII. A sul-fonic acid as a major sulfur compound of Bacillus subtilis spores. J Bacteriol. 98:62-68.
Canfield DE (2001). Biogeochemistry of sulfur isotopes. GeoScience-World. 43:607-636.
Chiang PK, Gordon RK, Tal J, Zeng GC, Doctor BP, Pardhasarad-hi K, McCann PP (1996). S-adenosylmethionine and methylation. FASEB J. 10:471-480.
Chmielewski P, Jezierski A (2003). Siarka. W: Słownik chemiczny, 1. wydanie, Warszawa, 284-286.
Cooper CE, Brown GC (2008). The inhibition of mitochondrial cy-tochrome oxidase by the gases carbon monoxide, nitric oxide, hy-drogen cyanide and hydrogen sulfide: chemical mechanism and physiological significance. J Bioenerg Biomembr. 40:533-539.
Dick WA (1992). Sulfur cycle. W: Encyclopedia of microbiology, 1 wydanie. Lederberg, J., (Red.). Academic Press, Inc., 4, 123-133.
Eichhorn EE (2000). Sulfonate-sulfur assimilation in Escherichia coli. Rozprawa doktorska, Zurich, Diss. ETH Nr. 13651.
Erickson JM (1992). Photosynthesis and chloroplasts. W: Encyclo-pedia of microbiology, 1 wydanie. Lederberg, J., (Red.). Academic Press, Inc., 3, 371-401.
Grundy FJ, Henkin TM (1998). The S box regulon: a new global transcription termination control system for methionine and cy-steine biosynthesis genes in Gram-positive bacteria. Mol Micro-biol. 30(4):737-749.
Hille R (2002). Molybdenum and tungsten in biology. Trends Bio-chem Sci. 27:360-367.
Kelly DP, Wood AP (2006). The chemolithotrophic prokaryotes. W: The Prokaryotes, a handbook on the biology of bacteria, 3 wydanie. Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (red.). Springer, t. 2, 441-456.
Kertesz MA (1999). Riding the sulfur cycle – metabolism of sulfo-nates and sulfate esters in Gram-negative bacteria. FEMS Micro-biol Rev. 24, 135-175.
Kessler D (2006). Enzymatic activation of sulfur for incorporation into biomolecules in prokaryotes. FEMS Microbiol Rev. 30, 825-840.
Kiley PJ, Beinert H (2003). The role of Fe-S proteins in sensing and regulation in bacteria. Curr Opin Microbiol. 6:181-185.
Kredich NM (1996). Biosynthesis of cysteine. W: Escherichia coli and Salmonella, 2. wydanie. Neidhardt FC, Curtiss R, Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff WS, Riley M, Schaech-ter M, Umbarger HE (red.). ASM Press, 514-527.
Madigan MT, Martinko JM, Dunlap PV, Clark DP (2009). Bacteria: The Proteobacteria. W: Brock Biology of microorganisms, 399-444.
Madsen EL (2008). Selected compounds in the biosphere that partici-pate in major microbial biogeochemical processes. W: Environ-mental Microbiology from genomes to biogeochemistry, Blackwell Publishing, 296-300.
McMurry J (2005). Struktury aminokwasów. W: Chemia organicz-na 4, Wydawnictwo Naukowe PWN, 986-990.
Murray RK, Granner DK, Rodwell VW (2006a). Biosynteza kwasów tłuszczowych. W: Biochemia Harpera, PZWL, 241-256.
Murray RK, Granner DK, Rodwell VW (2006b). Mikroelementy od-żywcze: witaminy i składniki mineralne. W: Biochemia Harpera, PZWL, 588-608.
Or-Rashid M, Onodera R, Wadud S (2001). Biosynthesis of methio-nine from homocysteine, cystathionine and homoserine plus cy-steine by mixed rumen microorganisms in vitro. Appl Microbiol Biotechnol. 55:758-764.
Ostrowski J, Wu J-Y, Rueger DC, Miller BE, Siegel LM, Kredich NM (1989). Characterization of the cysJIH regions of Salmonella ty-phimurium and Escherichia coli B. DNA sequences of cysl and cysH and a model for the siroheme-Fe4S4 active center of sulfite reductase hemoprotein based on amino acid homology with spin-ach nitrite redutase. J Biol Chem. 264:15726-15737.
Pajdowski L (1976). Siarka. W: Chemia ogólna, Wydawnictwo Na-ukowe PWN, 349-355.
Rabus R, Hansen TA, Widdel F (2006). Dessimilatory sulfate- and sulfur reducting prokaryotes. W: The Prokaryotes, a handbook on the biology of bacteria, 3 wydanie. Dworkin M, Falkow S, Rosen-berg E, Schleifer KH, Stackebrandt E (red.). Springer, 2, 659-768.
Rohwerdert T, Sand W (2003). The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithio-bacillus and Acidiphilium spp. Microbiology. 149:1699-1709.

Siarka i jej związki jako źródła pierwiastka budulcowego dla bakterii | Agnieszka Osówniak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 43
NAU
KAKR
ÓTK
OSZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Van der Ploeg JR, Weiss MA, Saller E, Nashimoto H, Saito N, Kertesz MA, Leisinger T (1996). Identification of sulfate starvation-regu-lated genes in Escherichia coli: a gene cluster involved in the uti-lization of taurine as a sulfur source. J Bacteriol. 178:5438-5446.
Vermeij P, Kertesz MA (1999). Pathways of assimilative sulfur me-tabolism in Pseudomonas putida. J Bacteriol. 181:5833-5837.
Wheeler PR, Coldham NG, Keating L, Gordon SV, Wooff EE, Par-ish T, Hewinson RG (2005). Functional demonstration of reverse transsulfuration in the Mycobacterium tuberculosis complex re-veals that methionine is the preferred sulfur source for pathogenic mycobacteria. J Biol Chem. 280:8069-8078.
Sulfur and its compounds as a source of structural element for bacteria
Agnieszka Osówniak
Sulfur is one of the chemical elements, which is necessary for living organisms. In this work, not only the physical and chemical properties of the element, but also biogenic chemical compounds containing sulfur were described. Next, the role of non-organic (sulfides, sulfates, and tio-sulfates) and organic (cysteine and metionine) compo-unds containing sulfur was characterized. A biosynthesis of L-metionine, L-cysteine, enzymes containing iron-sul-fur clusters, biotin, tiamine, liponic acid, coenzyme A, molybdopterin, and S-adenosylmethionine. Finally, the role of bacteria in the sulfur circulation was described.Słowa kluczowe: assimilation, bacteria, biosynthesis, cysteine, enzyme, operon, sulfate, sulfur, tiosulfate

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 44
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Metody obliczeniowe w genetyceEwa Oleńska, Alina Stankiewicz
dr Ewa Oleńska: asystent w Zakładzie Genetyki i Ewolu-cjonizmu, Instytut Biologii Uniwersytetu w Białymstoku
Streszczenie:
Praca, kierowana do nauczycieli biologii szkół gim-nazjalnych i ponadgimnazjalnych, ukazuje zastoso-wanie podstawowych metod obliczeniowych, których znajomość jest przydatna przy rozwiązywaniu zadań genetycznych, stosowanych jako częsty sposób ewalu-acji stanu wiedzy i umiejętności uczniów. Znajomość zasad dodawania, mnożenia, permutacji, kombinacji oraz umiejętność ich stosowania jest kluczowa przy oblicza-niu prawdopodobieństwa sumy i iloczynu zdarzeń oraz zdarzeń dwumianowych, co jest niezbędne do szacowania szansy powstawania określonych gamet czy genotypów, a często stanowi trudność dla uczniów. Ponadto, praca zawiera propozycję zadania rachunkowego opartego na omawianych w pracy zasadach matematycznych wraz z kluczem odpowiedzi.Słowa kluczowe: zadanie genetyczne, zasada dodawania, za-sada mnożenia, permutacja, kombinacja, prawdopodobieństwo sumy zdarzeń, prawdopodobieństwo iloczynu zdarzeń, prawdo-podobieństwo zdarzeń o rozkładzie dwumianowym, trójkąt Pascala
otrzymano: 30.11.2012; przyjęto: 11.04.2013; opublikowano: 28.06.2013
Matura 2012 – niezadowalające wyniki z genetyki
Postęp w badaniach biologicznych nie byłby możli-wy bez zastosowania narzędzi, modeli oraz teorii z in-nych nauk przede wszystkim fizyki, chemii, matema-tyki, a także odkryć technicznych i technologicznych. Podobnie w nauczaniu biologii, do wyjaśniania pro-cesów, zjawisk czy zależności biologicznych wykorzy-stywane są narzędzia, teorie z chemii, fizyki oraz ma-tematyki. Stosowanie działań matematycznych, w tym rachunku prawdopodobieństwa, na lekcjach biologii doskonale wpisuje się w kształtowanie u uczniów umie-jętności myślenia matematycznego – jednej z najważ-niejszych umiejętności, w które powinna wyposażyć uczniów szkoła na III i IV etapie edukacyjnym (Podsta-wa programowa, 2008).
Na umiejętność myślenia matematycznego składa się szereg czynności, np. wykorzystanie narzędzi mate-matyki w życiu codziennym oraz formułowanie sądów opartych na rozumowaniu matematycznym; stawianie charakterystycznych dla matematyki pytań i wiedza, ja-kiego rodzaju odpowiedzi można oczekiwać; odróżnia-
nie rodzajów matematycznych zadań. Wykorzystanie w nauczaniu biologii działań matematycznych sprzyja pogłębieniu rozumowania przyrodniczego. Na rozu-mowanie matematyczne składają się takie czynności jak: ocena ciągu argumentów przedstawionych przez innych; wiedza, czym jest dowód matematyczny; od-krywanie podstawowych idei w danym rozumowaniu; proponowanie formalnych i nieformalnych matema-tycznych argumentów (Niss, 1997).
W nauczaniu genetyki obok podstawowych działań matematycznych szczególne znaczenie ma rachunek prawdopodobieństwa. Pozwala on przewidzieć możli-wości wystąpienia zdarzeń genetycznych i znajduje za-stosowanie w wielu rodzajach obliczeń statystycznych. Stosowanie rachunku prawdopodobieństwa w opra-cowywaniu zagadnień genetyki mendlowskiej sprzy-ja pogłębieniu rozumienia zasad dziedziczenia oraz kształtowaniu u uczniów umiejętności rozumowania i argumentacji, a szczególnie: objaśnianiu i komentowa-niu informacji; wyjaśnianiu zależności przyczynowo--skutkowych, formułowaniu wniosków, dostrzeganiu zależności między biologią a matematyką. Zagadnienia
ZagadnienieWymagania szczegółowe Gimnazjum Liceum – zakres rozszerzony
Genetyka mendlowska
Uczeń:- przedstawia dziedziczenie cech
jednogenowych posługując się podstawowymi pojęciami genetyki (fenotyp, genotyp, gen, allel, homo-zygota, heterozygota, dominacja, recesywność)
- wyjaśnia dziedziczenie grup krwi człowieka (układ ABO, czynnik Rh)
- wyjaśnia dziedziczenie płci u czło-wieka i podaje przykłady cech sprzę-żonych z płcią (hemofilia, daltonizm)
Uczeń:- przedstawia i stosuje prawa Mendla- zapisuje i analizuje krzyżówki jednogenowe i dwugenowe (z domi-
nacją zupełną i niezupełną oraz allelami wielokrotnymi, posługując się szachownicą Punnetta) oraz określa prawdopodobieństwo wy-stąpienia poszczególnych genotypów i fenotypów w pokoleniach potomnych
- przedstawia sposób dziedziczenia płci u człowieka, analizuje drze-wo rodowe, w tym dotyczące występowania chorób genetycznych człowieka
- podaje przykłady cech (nieciągłych) dziedziczących się zgodnie z prawami Mendla
Tabela 1. Wymagania szczegółowe z genetyki na poziomie gimnazjum i liceum – zakres rozszerzony (Podstawa programowa 2008), w których stosuje się działania matematyczne
dr Alina Stankiewicz: starszy wykładowca w Pracowni Dydaktyki Biologii, Instytut Biologii Uniwersytetu w Bia-łymstoku

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 45
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
z genetyki mendlowskiej, obok genetyki molekularnej, występują zarówno na poziomie gimnazjum, jak i w li-ceum (Podstawa programowa, 2008).
Wymagania szczegółowe z genetyki, których speł-nienie opiera się na wykonaniu działań matematycz-nych i zastosowaniu rachunku prawdopodobieństwa przedstawiono w tabeli 1. Wymagania te stanowią je-dyną podstawę oceniania na egzaminach zewnętrznych (gimnazjalnym i maturalnym).
Rozwiązywanie zadań z genetyki z zastosowaniem rachunku prawdopodobieństwa pozwala także na do-skonalenie umiejętności posługiwania się terminologią genetyczną (np. allel, homozygota, heterozygota, cechy recesywne, cechy dominujące, fenotyp, genotyp), rozu-mienia zasad dziedziczenia, zapisu wytworzonych ga-met, genotypów, określania fenotypów, analizowania przyczyn i przewidywania skutków zdarzeń.
Uczniowie podczas rozwiązywania zadań z ge-netyki przekonują się, jak zastosowanie matematyki wpływa na interpretację reguł dziedziczenia, jak narzę-dzia matematyczne ułatwiają wnioskowanie na temat dziedziczenia. Działania matematyczne z rachunkiem prawdopodobieństwa są niezbędne w wyjaśnianiu dzie-dziczenia cech przez pojedyncze osobniki, lecz także są konieczne w wyjaśnianiu zasad genetyki populacyjnej, zmian w puli genowej populacji. Możliwości określe-nia prawdopodobieństwa wystąpienia konkretnego fenotypu ma szczególne znaczenie w odniesieniu do ludzi, którzy posiadają niewielką liczbę potomstwa (np. w przewidywaniu płci, wystąpienia chorób sprzę-żonych z płcią i innych chorób genetycznych, takich jak np. mukowiscydoza). Do oceny prawdopodobieństwa zajścia dowolnego zdarzenia używa się ułamków zwy-kłych (np. 3/4), dziesiętnych (0,75) lub procentów (75%). Prawdopodobieństwo zdarzenia zawiera się pomiędzy wartościami 0–1 lub 0–100%. Prawdopodobieństwo za-istnienia zdarzenia pewnego jest równe 1, jeśli zdarze-
nie jest niemożliwe, prawdopodobieństwo jego zaistnie-nia jest równe 0 (Łomnicki, 2007).
Informacje o przewidywaniu zdarzeń genetycznych często pojawiają się w mediach, prasie. Mają one duże znaczenie w diagnostyce medycznej i w planowaniu ży-cia rodzinnego. Zdawałoby się, że zainteresowanie ucz-niów problemami genetyki i motywacja do uczenia się tych zagadnień jest uzasadniona i duża. Jednak, jak wy-kazały badania, nauczanie genetyki wpłynęło ujemnie na postawę uczniów względem uczenia się tych treści. Przed rozpoczęciem cyklu lekcji poświęconych genetyce 49% uczniów wykazywało pozytywny stosunek wobec nauki, a po ich zakończeniu tylko 29%. Nie stwierdzo-no dodatniej zależności między osiągnięciami uczniów z genetyki a poziomem motywacji oraz między metoda-mi nauczania genetyki a poziomem motywacji. Znaczy to, że nauczanie genetyki w szkołach objętych badania-mi nie wpłynęło pozytywnie na rozwój zainteresowań tą dziedziną wiedzy (Sternicka, 1996, 1997).
Wyniki matur z 2012 r. dotyczące genetyki są nie-zadowalające. W arkuszu podstawowym egzaminu maturalnego z biologii dwa zadania z genetyki wyma-gały zastosowania rachunku prawdopodobieństwa. Oba zadania okazały się dla uczniów trudne. Jedno z zadań wymagało zapisania krzyżówki genetycznej i określenia prawdopodobieństwa wystąpienia danej ce-chy (łatwość zadania: 0,25), drugie zadanie wymagało określenia sposobu dziedziczenia wskazanych chorób genetycznych człowieka (łatwość zadania: 0,27). Zada-nia te sprawdzały rozumienie sposobu dziedziczenia chorób genetycznych oraz umiejętność rozwiązywania zadań z zakresu dziedziczenia cech u człowieka. Nale-żało zapisać krzyżówkę genetyczną z zastosowaniem szachownicy Punnetta i określić prawdopodobieństwo wystąpienia danej cechy, zaś kolejne zadanie dotyczy-ło dziedziczenia grup krwi. Rozwiązanie krzyżówek genetycznych sprawdzających w sposób praktyczny
rozumienie zasad dziedziczenia było dla maturzystów z 2012 r. trudniejsze niż dla zdających rok wcześniej. Większość odpowiedzi osób zdających była niepełna, np. brakowało zapisu krzyżówki genetycznej, zapisa-nych gamet, lub podkreślenia genotypu chorego dzie-cka (Centralna Komisja Egzaminacyjna, 2012).
Analiza wyników na poziomie rozszerzonym po-kazuje znacznie lepsze przygotowanie maturzystów do egzaminu na tym poziomie niż zdających na poziomie podstawowym. Jednak umiejętność wyjaśniania związ-ków przyczynowo-skutkowych okazała się trudna dla maturzystów zdających egzamin na obu poziomach. Maturzyści mieli problem z określeniem zależności „przyczyna – skutek”, a najczęstszym błędem odpo-wiedzi był brak jednego z elementów tej zależności. Trudnością okazało się rozwiązanie zadania z zakre-su dziedziczenia cech u człowieka, określenie sposobu dziedziczenia opisanej cechy (łatwość zadania: 0,42), i określenia sposobu dziedziczenia opisanej cechy na podstawie analizy przedstawionych danych (łatwość zadania: 0,50).
Co jest przyczyną niezadowalających wyników maturalnych z genetyki?
Niezadowalające wyniki maturalne z genetyki mogą mieć swoje przyczyny w tym, że nie udaje się przystęp-nie powiązać zastosowania działań matematycznych z rozumieniem przez uczniów zasad dziedziczenia, czyli zastosować myślenie matematyczne w przewidy-waniu zdarzeń genetycznych. Przełożenie matematyki na zasady dziedziczenia nie jest proste i wymaga od nauczyciela stałego doskonalenia się w tym zakresie. Ważne jest również, by uczniowie przed rozpoczęciem rozwiązywania krzyżówek genetycznych, na lekcjach matematyki poznali lub przypomnieli sobie rachunek prawdopodobieństwa. Wydaje się, że korzystna by-

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 46
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
łaby także korelacja czasowa między tematami lekcji z matematyki a tematami lekcji z genetyki. Właściwe uprzystępnienie uczniom zastosowania działań mate-matycznych i rachunku prawdopodobieństwa w roz-wiązywaniu krzyżówek genetycznych z pewnością przyczyni się do podwyższenia jakości i efektywności nauczania genetyki.
Celem pracy jest przedstawienie nauczycielom za-sad matematycznych niezbędnych przy rozwiązywaniu zadań genetycznych, stosowanych jako częsty sposób ewaluacji stanu wiedzy i umiejętności uczniów szkół gimnazjalnych i ponadgimnazjalnych.
Zasady matematyczne w obliczeniach genetycznych
Obliczenia genetyczne opierają się na czterech pod-stawowych zasadach matematycznych: dodawania, mnożenia, permutacji oraz kombinacji (Elseth i Baum-gardner, 1984).
Dodawanie – zdarzenia wykluczające się
Zasada dodawania ma zastosowanie do obliczania liczby sposobów zajścia zdarzeń wykluczających się, czyli takich gdzie wynik pierwszego zdarzenia wyklu-cza wynik drugiego zdarzenia. Przykładem zdarzeń wykluczających się jest jednoczesne występowanie alle-lu A oraz a w jednej gamecie wytwarzanej przez hetero-zygotę Aa. Według zasady dodawania, jeżeli zdarzenie pierwsze zachodzi na n1 sposobów, natomiast zdarzenie drugie na n2 sposoby i oba zdarzenia wzajemnie się wy-kluczają, wówczas albo zdarzenie pierwsze, albo zda-rzenie drugie zachodzi na n1 + n2 sposoby.
Przykład 1. Na ile sposobów można uzyskać osobnika o fe-notypie dominującym, przy założeniu dominacji zupełnej (Pisum sp.) z krzyżówki Aa × Aa?
Rozwiązanie:
• zdarzenie pierwsze (n1) – uzyskanie w potomstwie AA – 1 sposób;
• zdarzenie drugie (n2) – uzyskanie w potomstwie Aa – 1 sposób;
• zdarzenie trzecie (n3) – uzyskanie w potomstwie aA – 1 sposób;
Uzyskanie fenotypu dominującego A_, gdzie _ oznacza dowolny allel tego genu, może wystąpić jako jedno z wy-kluczających się zdarzeń: AA lub Aa lub aA, gdzie każde z nich zachodzi na 1 sposób. Zatem liczbę sposobów uzyskania osobnika o fenotypie dominującym obliczamy następująco:
A_ = n1 + n2 + n3 = AA + Aa + aA = 1 + 1 + 1 = 3
Fenotyp dominujący z krzyżówki Aa × Aa można uzyskać na 3 sposoby.
Mnożenie – zdarzenia niezależne
Drugą zasadą stosowaną w obliczeniach genetycz-nych jest zasada mnożenia. Stosuje się ją przy oblicza-niu liczby zdarzeń niezależnych, czyli takich, gdy wynik jednego zdarzenia nie wpływa na wynik drugiego zda-rzenia. Przykładem zdarzeń niezależnych jest urodze-nie się pierwszego i drugiego dziecka jednakowej płci. Urodzenie pierwszego dziecka, np. dziewczynki, nie ma wpływu na płeć kolejnych dzieci. Jeżeli zdarzenie pierwsze zachodzi na n1 sposobów, natomiast zdarzenie drugie na n2 sposoby i zdarzenia te są niezależne, wów-czas zdarzenie pierwsze i drugie zachodzi na n1 × n2 sposoby.
Przykład 2. Ile rodzajów gamet może wytworzyć osobnik o genotypie DDEeFFgg, przy założeniu, że wymienione geny dziedziczą się niezależnie.
Rozwiązanie:
• zdarzenie pierwsze (n1) – allel D segreguje tylko na 1 sposób (D);
• zdarzenie drugie (n2) – allele E, e segreguje na 2 sposo-by (albo E albo e);
• zdarzenie trzecie (n3) – allel F segreguje tylko na 1 spo-sób (F);
• zdarzenie czwarte (n4) – allel g segreguje tylko na 1 sposób (g).
Każde z czterech zdarzeń zachodzi niezależnie, zatem liczba rodzajów gamet wytwarzanych potencjalnie przez tego osobnika wynosi n1 × n2 × n3 × n4 = 1 × 2 × 1 × 1 = 2.
Permutacje
Trzecią zasadą stosowaną w genetyce jest zasada permutacji. Zasada ta jest używana do obliczania liczby sposobów, na które można uzyskać żądany wynik, jeże-li wszystkie obiekty są rozróżnialne. Liczbę permutacji w próbie oblicza się na podstawie wzoru:
N! = N × (N – 1) × (N – 2) × (N – 3) × ... × 1, (1)
gdzie: N – suma obiektów w próbie.
Przykład 3. W kolbie hodowlanej znajduje się 5 samców (♂) muszki owocowej fenotypowo identycznych, lecz ge-netycznie różnych (♂1 – ♂5). Oblicz na ile sposobów można wylosować owady z kolby.
Rozwiązanie:
Pierwszy sposób rozwiązania:
5! = 5 × (5 – 1) × (5 – 2) × (5 – 3) × (5 – 4) = 5 × 4 × 3 × 2 × 1 = 120

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 47
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Drugi sposób rozwiązania:
• zdarzenie pierwsze (n1) – jako pierwszego osobnika można wylosować ♂1 albo ♂2 albo ♂3 albo ♂4 albo ♂5, czyli zgodnie z zasadą dodawania zdarzenie pierwsze zachodzi na pięć sposobów. Na przykład jako pierw-szego osobnika wylosowano ♂1;
• zdarzenie drugie (n2) – drugiego osobnika można loso-wać już tylko z czterech możliwości: ♂2 albo ♂3 albo ♂4 albo ♂5, czyli zdarzenie drugie zachodzi na cztery spo-soby. Na przykład jako drugiego osobnika wylosowano ♂2;
• zdarzenie trzecie (n3) – jako trzeciego osobnika można wylosować albo ♂3 albo ♂4 albo ♂5, czyli zdarzenie za-chodzi na trzy sposoby. Jako trzeciego osobnika wylo-sowano na przykład ♂3;
• zdarzenie czwarte (n4) – jako czwartego osobnika moż-na wylosować albo ♂4 albo ♂5, zatem zdarzenie zacho-dzi na dwa sposoby. Na przykład, jako czwartego osob-nika wylosowano ♂4;
• zdarzenie piąte (n5) – jako piątego osobnika losujemy samca, który pozostał w kolbie, czyli ♂5, zatem zdarze-nie piąte zachodzi na jeden sposób.
Ponieważ każde z pięciu zdarzeń jest zdarzeniem nie-zależnym, co należy rozumieć, że po wylosowaniu jako pierwszego osobnika ♂1 jako drugiego osobnika można wylosować dowolnego samca z pozostałych czterech (♂2 – ♂5), stosuje się zasadę mnożenia i osobniki można wylosować na n1 × n2 × n3 × n4 × n5 = 5 × 4 × 3 × 2 × 1 = 120 sposobów.
Kombinacje
Zasada kombinacji ma zastosowanie do obliczania liczby sposobów, na które można uzyskać wynik, gdy obiekty są podzielone na dwie odrębne grupy. Kombi-nacje obliczamy ze wzoru:
C = N!
–––x1!x2!
, gdzie (2)
C – liczba kombinacji, N – suma obiektów,x1 – liczba obiektów jednej grupy,x2 – liczba obiektów drugiej grupy.
Przykład 4. Na ile sposobów można uzyskać 3 samce i 3 sa-mice muszki owocowej?
Rozwiązanie:
C(3♂ i 3♀) = 6!
–––3!3!
= 120–––
6 = 20 sposobów
Zastosowanie zasad
Omówione zasady są stosowane do obliczania praw-dopodobieństw zdarzeń. Prawdopodobieństwo sukcesu (p), czyli oczekiwanego wyniku zdarzenia jest propor-cją liczby sukcesów do sumy wszystkich możliwych wy-ników (Jadwiszczak, 2010).
p = (liczba sukcesów) : (suma wszystkich wyników) (3)
Zasada mnożenia jest stosowana przy obliczaniu prawdopodobieństwa zdarzeń, które są niezależne. Prawdopodobieństwo zdarzeń niezależnych jest praw-dopodobieństwem iloczynu zdarzeń. Jeżeli zdarzenie pierwsze (E1) zachodzi na n1 sposobów, a zdarzenie dru-gie (E2) na n2 sposoby i oba zdarzenia są niezależne, to prawdopodobieństwo, że wystąpią razem, jest iloczy-nem ich indywidualnych prawdopodobieństw (Watała, 2002).
p(E1 i E 2) = p(E1) × p(E2) (4)
Przykład 5. Oblicz prawdopodobieństwo wytworzenia ga-mety AB przez osobnika o genotypie AaBB, przy założeniu, że oba geny dziedziczą się niezależnie.
Rozwiązanie:
W gamecie para alleli A, a może wystąpić na 2 sposoby, natomiast allel B na 1 sposób. Oba geny dziedziczą się niezależnie. A zatem prawdopodobieństwo, że osobnik o danym genotypie wytworzy gametę AB jest iloczynem prawdopodobieństwa wytworzenia gamety z allelem
A oraz prawdopodobieństwa wytworzenia gamety z al-lelem B. Wobec tego, że prawdopodobieństwo powsta-nia gamety z allelem A, czyli p(A) wynosi ½ (zgodnie ze wzorem (3) 1 sukces do 2 możliwych wyników – albo allel A albo allel a będzie w gamecie), natomiast prawdopodo-bieństwo powstania gamety z allelem B, czyli p(B) wynosi 1 (1 sukces do 1 możliwego wyniku), prawdopodobień-stwo, że oba zdarzenia wystąpią razem obliczamy zgod-nie ze wzorem (4) w następujący sposób:
p(AB) = p(A) × p(B) = ½ × 1 = ½
Zasada dodawania jest stosowana do obliczania prawdo-podobieństwa zdarzeń, które się wzajemnie wykluczają. Gdy zdarzenie pierwsze (E1) zachodzi na n1 sposobów, a zdarzenie drugie (E2) zachodzi na n2 sposoby i oba zda-rzenia wzajemnie się wykluczają, wówczas prawdopodo-bieństwo albo zdarzenia pierwszego albo drugiego jest sumą ich indywidualnych prawdopodobieństw, i jest na-zywane prawdopodobieństwem sumy zdarzeń.
p(E1 lub E2) = p(E1) + p(E2) (5)
Przykład 6. Galaktozemia jest chorobą autosomalną rece-sywną. Oblicz prawdopodobieństwo, że pierwsze dziecko urodzone w małżeństwie dwóch heterozygotycznych osób będzie albo zdrową dziewczynką, albo chorym chłopcem.
Rozwiązanie:
Galaktozemia jest determinowana allelem resesywnym autosomalnym g, natomiast brak choroby allelem domi-nującym G. W potomstwie dwóch heterozygot: Gg × Gg dzieci zdrowe do chorych urodzą się w proporcji 3:1.
Zdarzenie urodzenia się dziecka jako zdrowej dziewczyn-ki wyklucza, że dziecko jednocześnie będzie chorym chłopcem. A zatem prawdopodobieństwo urodzenia dziecka, które będzie albo zdrową dziewczynką, albo chorym chłopcem obliczamy jako sumę indywidualnych prawdopodobieństw obu zdarzeń.
p(G_♀ lub gg♂) = p(G_♀) + p(gg♂) = (¾×½) + (¼×½) = ⅜ + 1/8 = 4/8 = ½

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 48
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
W celu obliczenia prawdopodobieństwa zdarzeń wśród N obiektów należących do dwóch wyraźnie wyodręb-nionych grup (prawdopodobieństwo zdarzeń dwumia-nowych), stosuje się zasadę kombinacji. Prawdopodo-bieństwo zdarzenia złożonego z x1 sukcesów i x2 porażek, można obliczyć dwiema metodami. Jedna z metod opie-ra się na zastosowaniu wzoru (6), który zawiera wzór (2):
p(x1, x2) = N!
–––x1!x2!
px1qx2 , gdzie (6)
N – suma obiektów
x1 – liczba sukcesów
x2 – liczba porażek; x1 + x2 = N
p – prawdopodobieństwo sukcesu
q – prawdopodobieństwo porażki; p + q=1
Druga metoda, stosowana do obliczania prawdopodo-bieństwa wyłącznie zdarzeń dwumianowych zachodzą-cych z prawdopodobieństwem ½, opiera się na konstruk-cji trójkąta Pascala, co obrazuje rycina 1. Wykorzystanie obu metod zostanie omówione na przykładzie siódmym.
Przykład 7. Oblicz prawdopodobieństwo otrzymania dwóch samców (♂) i dwóch samic (♀) muszki owocowej przy liczebności potomstwa cztery.
Rozwiązanie:
a) metoda pierwsza
p(2♂ i 2♀) = 4!
–––2!2!
(½)2 (½)2 = 6 × ¼ × ¼ = 6/16 = 3/8
b) metoda druga oparta na trójkącie Pascala (ryc. 1)
Prawdopodobieństwo uzyskania grupy złożonej z 2 sam-ców i 2 samic oblicza się korzystając ze wzoru (3):
p(2♂ i 2♀) = 6/16 = 3/8
Zawarte w pracy przykłady stosowania podstawowych zasad matematycznych mogą ułatwić nauczycielom wy-jaśnianie trudnych zagadnień z zakresu rachunku praw-dopodobieństwa i przyczynić się do rozwoju wiedzy
genetycznej uczniów, popartej umiejętnością rozwiązy-wania zadań genetycznych. Przykład ósmy zawiera omó-wione w pracy zasady matematyczne, które nauczyciel dowolnie może wykorzystać w obliczeniach genetycz-nych.
Przykład 8. Mukowiscydoza jest chorobą autosomalną re-cesywną. Heterozygotyczni rodzice planują założenie rodzi-ny. Oblicz:
a) liczbę możliwych typów gamet wytwarzanych przez ojca dzieci;
b) na ile sposobów można uzyskać 1 dziewczynkę i 2 chłopców, przy założeniu że chłopcy nie różnią się między sobą;
c) prawdopodobieństwo urodzenia 1 dziewczynki i 2 chłopców, przy założeniu, że chłopcy nie różnią się między sobą;
d) prawdopodobieństwo, że pierwszym dzieckiem bę-dzie chłopiec, drugim dziewczynka, a trzecim chło-piec;
e) prawdopodobieństwo, że pierwsze dziecko będzie chore;
f) prawdopodobieństwo urodzenia chorego chłopca;
g) prawdopodobieństwo urodzenia pierwszego chore-go chłopca, drugiego dziecka – zdrowej dziewczynki, zaś trzeciego dziecka – zdrowego chłopca;
h) prawdopodobieństwo urodzenia trójki zdrowych dzieci;
i) prawdopodobieństwo urodzenia pierwszego dziecka jako zdrowej dziewczynki lub zdrowego chłopca.
Rozwiązanie:
Oznaczenie alleli: A – brak choroby, a – mukowiscydoza.
Krzyżówka:
P: Aa×Aa
a) Postępujemy zgodnie z zasadą dodawania:
• zdarzenie pierwsze n1 = 1 sposób (w gamecie znajduje się allel A)
• zdarzenie drugie n2 = 1 sposób (w gamecie znajduje się allel a)
Obecność w gamecie allelu A wyklucza obecność allelu a w tej samej gamecie (I prawo Mendla), zatem liczbę spo-sobów zdarzeń wykluczających się obliczamy korzystając z zasady dodawania i uzyskujemy wynik:
n1 + n2 = 1 + 1 = 2
b) Postępujemy zgodnie ze wzorem (2) lub korzystamy z trójkąta Pascala (ryc. 2):
C(1♀ i 2♂) = 3!
–––1!2!
= 3
Ryc. 1. Schemat trójkąta Pascala rozpisanego do rzędu liczącego 4 osobniki w próbie
Ryc. 2. Schemat metody obliczania liczby sposobów uzyskania zdarzenia: 1 dziewczynka i 2 chłopców przy zastosowaniu trójkąta Pascala.

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 49
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
c) Postępujemy zgodnie ze wzorem (6) lub korzystamy z trójkąta Pascala (ryc. 3):
p(1♀ i 2♂) = 3!
–––1!2!
× (1–2
)1 × (1–2
)2 = 3 × 1–2
× 1–4
= 3–8
Ryc. 3. Schemat metody obliczania prawdopodobieństwa zdarzenia: 1 dziewczynka i 2 chłopców przy zastosowaniu trójkąta Pascala.
d) Urodzenie pierwszego dziecka jako chłopca nie ma wpływu na płeć drugiego i trzeciego dziecka, zatem są to zdarzenia niezależne i korzystamy z zasady mno-żenia.
p(♂,♀,♂) = p(♂) × p(♀) × p(♂) = 1 1 1
1
– × – × – = –2 2 2 8
e) Prawdopodobieństwo urodzenia pierwszego chore-
go dziecka, czyli osobnika o genotypie aa z krzyżówki Aa×Aa obliczamy na podstawie wzoru (4), ponieważ allel a pochodzący od ojca znajduje się w zygocie nie-zależnie od allelu a z gamety pochodzącej od matki:
p(aa) = p(a) × p(a) = 1–2
× 1–2
= 1–4
f) Prawdopodobieństwo urodzenia chorego chłopca składa się z dwóch zdarzeń: urodzenia się chorego dziecka oraz urodzenia chłopca. Oba zdarzenia są nie-
zależne, zatem korzystamy z prawdopodobieństwa iloczynu zdarzeń.
p(aa♂) = 1–4
× 1–2
= 1–8
g) Urodzenie pierwszego chorego chłopca nie wpływa na urodzenie drugiego dziecka jako zdrowej dziew-czynki ani trzeciego dziecka jako zdrowego chłopca, zatem obliczenie prawdopodobieństwa tego zda-rzenia jest iloczynem indywidualnych prawdopodo-bieństw poszczególnych zdarzeń;
• prawdopodobieństwo urodzenia zdrowego dziecka
p(A_) = p(AA) + p(Aa) + p(aA) = 1 1 1 3– + – + – = –4 4 4 4
• prawdopodobieństwo urodzenia chorego dziecka
p(aa) = 1–4
• prawdopodobieństwo urodzenia chłopca p(♂) = ½
• prawdopodobieństwo urodzenia dziewczynki (♀) = ½
p(aa♂ × A_♀ × A_♂) = 1–4
× 1–2
× 3–4
× 1–2
× 3–4
× 1–2
= 9
––512
h) W zadaniu nie ma informacji co do płci dzieci. Prawdo-podobieństwo urodzenia trójki zdrowych dzieci obli-cza się z na podstawie prawdopodobieństwa zdarzeń niezależnych:
p(A_, A_, A_ ) = 3–4
× 3–4
× 3–4
= 27––64
i) Prawdopodobieństwo urodzenia pierwszego dziecka jako zdrowej dziewczynki wyklucza, że to samo dzie-cko będzie zdrowym chłopcem, zatem korzystamy ze wzoru prawdopodobieństwa sumy zdarzeń (5).
p(A_♀, A_♂) = (3–4
× 1–2
) + (3–4
× 1–2
) = 6–8
= 3–4
Podsumowanie
Rozwiązywanie zadań z genetyki przypomina roz-wiązywanie łamigłówek. Aby rozwiązywanie zadań nie sprawiało uczniom trudności i było przyjemne, po-winno się przestrzegać określonego postępowania oraz poprawnie stosować zasady matematyczne. Znajomość zasady mnożenia jest niezbędna do oceny prawdopodo-bieństwa zaistnienia zdarzeń niezależnych, natomiast zasady dodawania w celu szacowania prawdopodobień-stwa zaistnienia zdarzeń, które się wykluczają. Zasada permutacji oraz kombinacji jest przydatna przy oblicza-niu prawdopodobieństwa zdarzeń zachodzących wśród obiektów podzielonych na dwie grupy. Znajomość zasad matematycznych oraz umiejętność ich prawidłowego zastosowania może również przyczynić się do głębszego zrozumienia przez uczniów procesów i praw biologicz-nych takich jak: mejoza, losowa segregacja chromoso-mów, losowe łączenie gamet czy I i II prawo Mendla.
Literatura
Elseth GD, Baumgardner KD (1984). Genetics. Addison-Wesley Pub-lishing Company.
Jadwiszczak P (2010). Zrozumiec statystykę. Szczecin: Wyd. My Book.Łomnicki A (2007). Wprowadzenie do statystyki dla przyrodników.
Warszawa: Wyd. Naukowe PWN,.Niss M (1997). Matematyczne kompetencje. Dostępne na: www.
ap.krakow.pl/kdm/MNissOsiągnięcia maturzystów w 2012 roku. Sprawozdanie z egzaminu
maturalnego z 2012 roku. Centralna Komisja Egzaminacyjna. Do-stępne na: www.cke.edu.pl.
Podstawa programowa kształcenia ogólnego. Rozporządzenie MEN z dnia 23 grudnia 2008 r.
Sternicka A (1997). Czy genetyka może być dla uczniów ciekawsza i łatwiejsza? Biologia w Szkole. 5:286-292.
Sternicka A (1996). Postawy uczniów wobec genetyki. Biologia w Szkole. 4:198-204.
Watała C (2002) Biostatystyka – wykorzystanie metod statystycznych w pracy badawczej w naukach biomedycznych. Bielsko-Biała: Wyd. α-medica press.

Metody obliczeniowe w genetyce | Ewa Oleńska, Alina Stankiewicz | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 50
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Calculation methods in genetics
Ewa Oleńska, Alina Stankiewicz
This work, dedicated to biology teachers of high and secondary schools, presents the basic mathematic cal-culation rules applied in solving the solutions of genetic exercises which are the common evaluation measure of students’ knowledge and skills. The knowledge according to addition and multiplication rules, permutation formu-la and combination rule is applied in calculating of addi-tion, multiplication and binomial probabilities, the calcu-lating of which is crucial in determining the chances of production of specific gametes or genotypes which often is difficult for students. Moreover, present work includes proposal exercise based on presented in work mathematic ruler with the answers attached..Key words: genetic exercise, addition rule, multiplication rule, permutation formula, combination rule, addition probabil-ity, multiplication probability, binomial probability, Pascal’s triangle

Osiągnięcia przyrodnicze trzecioklasistów | Krzysztof Konarzewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 51
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Osiągnięcia przyrodnicze trzecioklasistóww perspektywie międzynarodowejKrzysztof Konarzewski
prof. Krzysztof Konarzewski: profesor pedagogiki i eks-pert oświatowy. Pracuje w Instytucie Badań Edukacyjnych w Warszawie i Wszechnicy Świętokrzyskiej w Kielcach.
W maju 2011 r. zespół badaczy powołany przez Cen-tralną Komisję Egzaminacyjną przeprowadził pomiar osiągnięć matematycznych i przyrodniczych ponad 5 tys. uczniów z losowo wybranych 257 oddziałów klasy trzeciej w 150 szkołach podstawowych. W odrębnej se-sji ci sami uczniowie wykonali ponadto test rozumie-nia pisanego tekstu. Pomiary odbyły się w ramach ba-dań TIMSS (Trends in International Mathematics and Science Study) i PIRLS (Progress in International Re-ading Literacy Study), które zorganizowało Międzyna-rodowe Towarzystwo Mierzenia Osiągnięć Szkolnych (IEA – International Association for the Evaluation of Educational Achie-vement) w 50 krajach (Martin, Mullis, Foy i Stanco, 2012). Średni wiek testowanych dzie-ci wahał się od 9,7 (we Włoszech i Norwegii) do 11,2 lat (w Jemenie), a liczba lat nauki szkol-nej – od 3 (w Polsce) do 6 lat (w Holandii i Irlan-dii Północnej). Badania dają podstawę do oceny efektywności polskiej edukacji początkowej (Konarzewski, 2012). Poniżej przedsta-wiam najważniejsze wyniki i wnioski odnoszące się do osiągnięć przyrodniczych ostatniego rocznika dzieci kształconych według podstawy programowej ministra Mirosława Handke.
Wyniki
1. Test TIMSS mierzył zasoby wiedzy przyrodniczej o życiu, Ziemi i materii nieożywionej, uznane przez ekspertów za niezbędne do kontynuowania edukacji
Streszczenie:
Artykuł przedstawia wyniki pomiaru osiągnięć przyrod-niczych uczniów klasy trzeciej szkoły podstawowej na tle wyników uczniów z 49 innych krajów, a także prawdo-podobne powody względnie niskiej pozycji polskich ucz-niów w tym pomiarze.Słowa kluczowe: TIMSS, PIRLS, międzynarodowe bada-nia osiągnięć szkolnych, osiągnięcia przyrodnicze, edukacja początkowa
otrzymano: 21.03.2013; przyjęto: 9.04.2013; opublikowano: 28.06.2013
przyrodniczej w następnych latach. Na wiedzę o życiu (biologię) złożyło się sześć zagadnień:• organizmy ludzi, zwierząt i roślin – cechy odróż-
niające je od rzeczy nieożywionych, główne układy i ich funkcje;
• rozmnażanie, dziedziczenie, cykle życiowe roślin i zwierząt;
• cechy fizyczne, zachowanie i przystosowanie orga-nizmów do życia w różnych środowiskach;
• stosunki w środowiskach naturalnych, np. zależ-ności między organizmami w lesie, sadzawce, na pustyni, proste łańcuchy pokarmowe;
• zmiany w środowisku spowodowane działalnością człowieka, np. zanieczyszczenie środowiska i spo-soby zapobiegania mu;
• zdrowie człowieka, np. symptomy zdrowia i cho-roby, przenoszenie chorób, zapobieganie choro-bom, dieta, ruch.
Wiedza o Ziemi (geografia) obejmowała sześć za-gadnień:• woda na Ziemi (umiejscowienie, rodzaje wód,
obieg) i powietrze (dowód na istnienie powietrza, skład, wykorzystanie w codziennym życiu);
• ukształtowanie powierzchni Ziemi (góry, równi-ny, pustynie, rzeki) i związki z życiem człowieka (np. rolnictwem – nawadnianie, rozwój terenów zielonych);
• warunki pogodowe w różnych porach roku;• skamieniałości zwierząt i roślin (wiek, lokalizacja,
powstawanie);• Układ Słoneczny (Słońce, planety, Księżyc);• dzień i noc, związek cienia z obrotem Ziemi.Wiedza o materii nieożywionej (fizyka) obejmowała
osiem zagadnień:• stany skupienia materii (ciała stałe, płyny i gazy)
i ich własności (kształt, objętość), w tym zmiany stanu pod wpływem ogrzewania i ochładzania;
Nasi uczniowie mogliby wypaść lepiej, gdyby nie okrojony program i anachroniczna metodyka kształcenia przyrodniczego
”

Osiągnięcia przyrodnicze trzecioklasistów | Krzysztof Konarzewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 52
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
• klasyfikowanie przedmiotów na podstawie cech fizycznych, np. ciężaru, objętości, przyciągania magnetycznego;
• tworzenie i rozdzielanie mieszanin; • typowe przemiany ciał, np. gnicie, spalanie, rdze-
wienie, gotowanie; • typowe źródła energii i ich wykorzystanie w prak-
tyce – ciepło słoneczne, elektryczność, ruch wody lub wiatru;
• światło – źródła, zachowanie się; • obwody elektryczne, właściwości magnesu; • siły poruszające ciała – grawitacja, popychanie
i ciągnięcie. 2. W międzynarodowej skali osiągnięć przyrodni-
czych o średniej 500 i odchyleniu standardowym 100 polscy uczniowie uzyskali 505 punktów z odchyleniem standardowym 78 (ryc. 1). Osiągnięcia małych Pola-ków – podobnie jak uczniów z czterech innych krajów:
Rumunii, Hiszpanii, Nowej Zelandii i Kazachstanu – nie różniły się istotnie od średniej międzynarodowej. W rankingu osiągnięć przyrodniczych dzieci z Polski zajęły 29. miejsce (ex aequo z Rumunią i Hiszpanią). Warto dodać, że w rankingu osiągnięć matematycznych wypadły wyraźnie gorzej (34. miejsce), a w rozumieniu pisanego tekstu – odrobinę lepiej (28. miejsce).
3. Polscy uczniowie należeli do najmłodszych (9,9 lat) i najkrócej kształconych (3 lata) w szkole. Czterdzie-ści trzy, a właściwie 42 kraje, bo wyniki Jemenu odsta-wały tak bardzo od pozostałych, że musiały być usunię-te z analizy, testowały czwartoklasistów. Dzieci te były w różnym wieku, zależnie od momentu rozpoczynania nauki. Jak pokazuje ryc. 2, dziesięcioletni trzecioklasi-ści z Polski uzyskali wynik typowy dla czwartoklasi-stów starszych o pół roku.
4. Cztery wartości progowe ustalone na skali mię-dzynarodowej pozwoliły podzielić wyniki na pięć prze-
działów. Do ich nazwania wykorzystuję naszą skalę ocen szkolnych od „jedynki” do „piątki”. Pod wzglę-dem odsetka piątek listę otwiera Singapur (piątki zdo-
Ryc. 1. Średnie osiągnięć przyrodniczych w 50 krajach
Źródło: Martin i wsp. (2012)
Ryc. 2. Średnie osiągnięć przyrodniczych w Polsce i 42 krajach testujących czwartoklasistów
Obliczenia własne na podstawie: Martin i wsp. (2012)
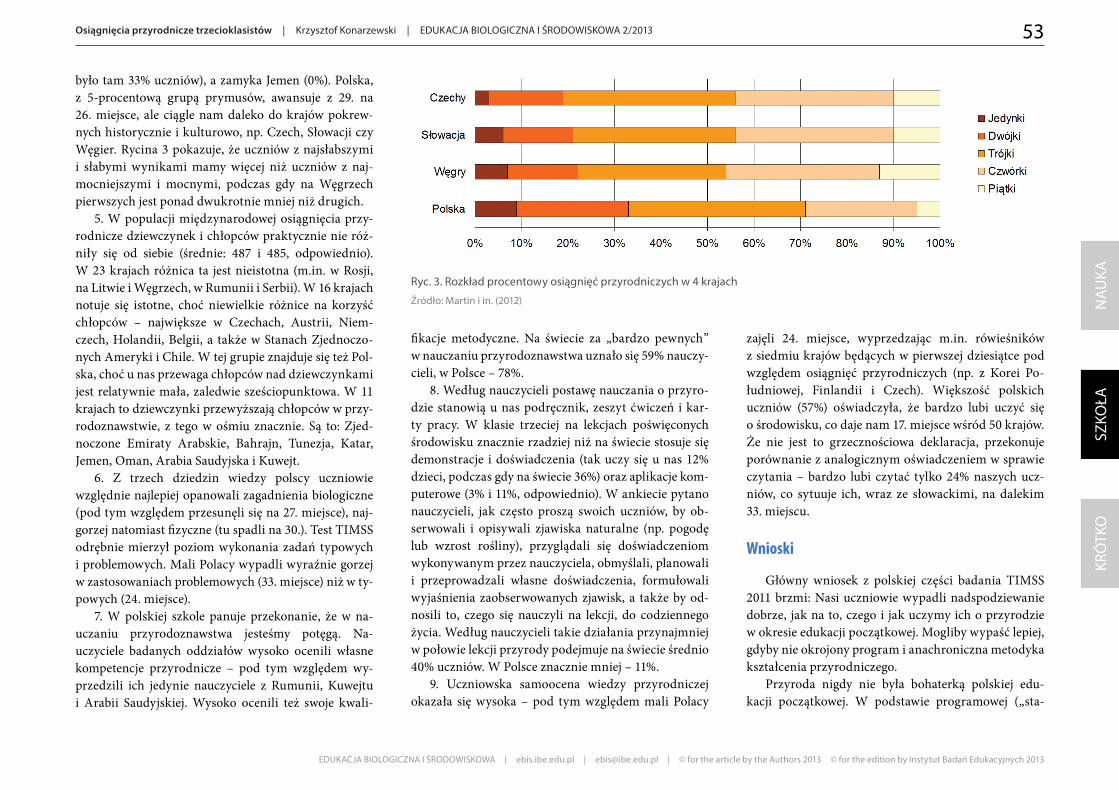
Osiągnięcia przyrodnicze trzecioklasistów | Krzysztof Konarzewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 53
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
było tam 33% uczniów), a zamyka Jemen (0%). Polska, z 5-procentową grupą prymusów, awansuje z 29. na 26. miejsce, ale ciągle nam daleko do krajów pokrew-nych historycznie i kulturowo, np. Czech, Słowacji czy Węgier. Rycina 3 pokazuje, że uczniów z najsłabszymi i słabymi wynikami mamy więcej niż uczniów z naj-mocniejszymi i mocnymi, podczas gdy na Węgrzech pierwszych jest ponad dwukrotnie mniej niż drugich.
5. W populacji międzynarodowej osiągnięcia przy-rodnicze dziewczynek i chłopców praktycznie nie róż-niły się od siebie (średnie: 487 i 485, odpowiednio). W 23 krajach różnica ta jest nieistotna (m.in. w Rosji, na Litwie i Węgrzech, w Rumunii i Serbii). W 16 krajach notuje się istotne, choć niewielkie różnice na korzyść chłopców – największe w Czechach, Austrii, Niem-czech, Holandii, Belgii, a także w Stanach Zjednoczo-nych Ameryki i Chile. W tej grupie znajduje się też Pol-ska, choć u nas przewaga chłopców nad dziewczynkami jest relatywnie mała, zaledwie sześciopunktowa. W 11 krajach to dziewczynki przewyższają chłopców w przy-rodoznawstwie, z tego w ośmiu znacznie. Są to: Zjed-noczone Emiraty Arabskie, Bahrajn, Tunezja, Katar, Jemen, Oman, Arabia Saudyjska i Kuwejt.
6. Z trzech dziedzin wiedzy polscy uczniowie względnie najlepiej opanowali zagadnienia biologiczne (pod tym względem przesunęli się na 27. miejsce), naj-gorzej natomiast fizyczne (tu spadli na 30.). Test TIMSS odrębnie mierzył poziom wykonania zadań typowych i problemowych. Mali Polacy wypadli wyraźnie gorzej w zastosowaniach problemowych (33. miejsce) niż w ty-powych (24. miejsce).
7. W polskiej szkole panuje przekonanie, że w na-uczaniu przyrodoznawstwa jesteśmy potęgą. Na-uczyciele badanych oddziałów wysoko ocenili własne kompetencje przyrodnicze – pod tym względem wy-przedzili ich jedynie nauczyciele z Rumunii, Kuwejtu i Arabii Saudyjskiej. Wysoko ocenili też swoje kwali-
fikacje metodyczne. Na świecie za „bardzo pewnych” w nauczaniu przyrodoznawstwa uznało się 59% nauczy-cieli, w Polsce – 78%.
8. Według nauczycieli postawę nauczania o przyro-dzie stanowią u nas podręcznik, zeszyt ćwiczeń i kar-ty pracy. W klasie trzeciej na lekcjach poświęconych środowisku znacznie rzadziej niż na świecie stosuje się demonstracje i doświadczenia (tak uczy się u nas 12% dzieci, podczas gdy na świecie 36%) oraz aplikacje kom-puterowe (3% i 11%, odpowiednio). W ankiecie pytano nauczycieli, jak często proszą swoich uczniów, by ob-serwowali i opisywali zjawiska naturalne (np. pogodę lub wzrost rośliny), przyglądali się doświadczeniom wykonywanym przez nauczyciela, obmyślali, planowali i przeprowadzali własne doświadczenia, formułowali wyjaśnienia zaobserwowanych zjawisk, a także by od-nosili to, czego się nauczyli na lekcji, do codziennego życia. Według nauczycieli takie działania przynajmniej w połowie lekcji przyrody podejmuje na świecie średnio 40% uczniów. W Polsce znacznie mniej – 11%.
9. Uczniowska samoocena wiedzy przyrodniczej okazała się wysoka – pod tym względem mali Polacy
zajęli 24. miejsce, wyprzedzając m.in. rówieśników z siedmiu krajów będących w pierwszej dziesiątce pod względem osiągnięć przyrodniczych (np. z Korei Po-łudniowej, Finlandii i Czech). Większość polskich uczniów (57%) oświadczyła, że bardzo lubi uczyć się o środowisku, co daje nam 17. miejsce wśród 50 krajów. Że nie jest to grzecznościowa deklaracja, przekonuje porównanie z analogicznym oświadczeniem w sprawie czytania – bardzo lubi czytać tylko 24% naszych ucz-niów, co sytuuje ich, wraz ze słowackimi, na dalekim 33. miejscu.
Wnioski
Główny wniosek z polskiej części badania TIMSS 2011 brzmi: Nasi uczniowie wypadli nadspodziewanie dobrze, jak na to, czego i jak uczymy ich o przyrodzie w okresie edukacji początkowej. Mogliby wypaść lepiej, gdyby nie okrojony program i anachroniczna metodyka kształcenia przyrodniczego.
Przyroda nigdy nie była bohaterką polskiej edu-kacji początkowej. W podstawie programowej („sta-
Ryc. 3. Rozkład procentowy osiągnięć przyrodniczych w 4 krajach
Źródło: Martin i in. (2012)

Osiągnięcia przyrodnicze trzecioklasistów | Krzysztof Konarzewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 54
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
rej”, z 2002 r., bo ta obowiązywała uczniów badanych w 2011 r.) odwołania do przyrodoznawstwa są nadzwy-czaj skąpe. Wśród pięciu celów edukacji początkowej znajdziemy „rozbudzanie potrzeby kontaktu z przyro-dą”, co z przyrodoznawstwem ma niewiele wspólnego, bo trąci romantyzmem – więcej w tym Mickiewicza niż braci Śniadeckich. W spisie zadań szkoły mamy „uczenie właściwych zachowań w stosunku do zwierząt i otaczającej przyrody” – tu też przyroda występuje jako człon relacji ze światem człowieka, a nie jako autono-miczny przedmiot obserwacji „szkiełkiem i okiem”. Na 55 jednostek treści nauczania do przyrody odwołuje się siedem. Najbliższe duchowi przyrodoznawstwa brzmi: „obserwowanie zjawisk i procesów przyrodniczych do-stępnych doświadczeniu dziecka i mówienie o nich”. Dlaczego „mówienie”, a nie „wyjaśnianie”? Zapewne dlatego, by powstrzymać inwazję teorii naukowych w wykoncypowany w romantyczno-mieszczańskim duchu świat dzieciństwa.
O okrojeniu naszych programów kształcenia przy-rodniczego najlepiej świadczy fakt, że znalazło się w nich tylko 2/5 zagadnień przyrodniczych objętych te-stem TIMSS. Najmniej wiadomości dostawały polskie dzieci o materii nieożywionej. Ponieważ mało ich też w życiu codziennym, trudno się dziwić, że w tej dzie-dzinie wypadły względnie najgorzej.
Względnie niską pozycję naszych uczniów w zada-niach problemowych najprościej tłumaczy dominują-ca, oparta na słowie i obrazku metodyka kształcenia przyrodniczego. Najnowsza metaanaliza 138 doniesień badawczych poświęconych nauczaniu opartemu na ba-daniu wykazuje, że sprzyja ono zapamiętywaniu i ro-zumieniu treści przyrodniczych. Szczególnie korzystna dla zrozumienia przyrody jest aktywność umysłowa dzieci podczas prowadzenia własnych doświadczeń i nacisk na wyciąganie wniosków z danych (Minner, Levy i Century, 2009).
Ubóstwo treści przyrodniczych i nieangażującą me-todykę po części tłumaczy niska klasa, w której byli nasi uczniowie. W pozostałych krajach badani uczniowie byli w czwartej, piątej, a nawet szóstej klasie. Wykona-nie powszechnie kontestowanej ustawy o objęciu dzieci sześcioletnich obowiązkiem szkolnym daje nadzieję na podniesienie osiągnięć przyrodniczych polskich dzie-sięciolatków – skoro bowiem dziś sporo umieją, choć były nauczane marnie i tylko przez trzy lata, to naucza-ne dobrze przez całe cztery lata powinny umieć znacz-nie więcej. Niestety, samo obniżenie wieku rozpoczyna-nia nauki nie wywoła przełomu.
Nową podstawę programową dla klas początkowych nieco poszerzono – dzięki czemu pokrywa już połowę zagadnień sprawdzanych w testach TIMSS – a także wzbogacono o zalecenie, by uczniowie obserwowali i prowadzi proste doświadczenia przyrodnicze i odwie-dzali ogród botaniczny lub gospodarstwo rolne. Nie zmierzono się jednak z problemem kwalifikacji nauczy-cieli. Od czasu wprowadzenia w latach 70. XX w. odręb-nej specjalności nauczycielskiej nazwanej nauczaniem początkowym, przyszli nauczyciele studiują filozofię i socjologię edukacji, psychologię rozwoju, komunika-cję interpersonalną, diagnostykę zaburzeń uczenia się itp., ale z przyrodoznawstwem stykają się jedynie na krótkim i uproszczonym kursie „edukacji przyrodni-czej”, z reguły niekończącym się egzaminem (na Uni-wersytecie Warszawskim trwa on 30 godzin i kończy się zaliczeniem na ocenę). W tym świetle wysoka samo-ocena naszych nauczycieli jawi się jako świadectwo nie-zrozumienia współczesnego przyrodoznawstwa i meto-dycznego tradycjonalizmu.
W klasie czwartej przyroda jest odrębnym przed-miotem nauczanym przez lepiej przygotowanych na-uczycieli. Trudno jednak mieć nadzieję, że w ciągu jednego roku uda się efektywnie zapoznać czwartokla-sistów z drugą połową zagadnień testowanych w ba-
daniu TIMSS. Na przeszkodzie stanie zapewne ostre przejście od luźnej, „zintegrowanej” edukacji począt-kowej do sztywnego nauczania przedmiotowego. Takie przejście zrywa ciągłość kształcenia, co musi utrudnić dzieciom wykorzystywanie wiedzy zdobytej w klasach początkowych. Deklarowane przez uczniów zaintereso-wanie przyrodą jak było, tak zapewne i będzie marno-wane w polskich szkołach podstawowych.
Literatura
Konarzewski K (2012). TIMSS i PIRLS 2011. Osiągnięcia szkolne polskich trzecioklasistów w perspektywie międzynarodowej. Warszawa: CKE.
Minner DD, Levy AJ i Century J (2009). Inquiry-based science in-struction – What is it and does it matter? Results from a research synthesis years 1984 to 2002. Journal of Research in Science Teaching, 47(4), 474–496.
Martin MO, Mullis IVS, Foy P i Stanco GM (2012). TIMSS 2011 in-ternational results in science. Chestnut Hill: Lynch School of Education, Boston College.
International assessment of elementary school students’ science achievement
Krzysztof Konarzewski
The paper presents elementary school third graders’ sci-ence achievement assessed against an international back-ground, and offers some reasons for relatively low posi-tion of the Polish students in the world rankings.Key words: TIMSS, PIRLS, international measurement of scho-lastic achievement, science achievement, elementary education

Małe eksperymenty, wielkie odkrycia | Marcin Zaród | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 55
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Małe eksperymenty, wielkie odkryciaPraktyka eksperymentalna w edukacji przyrodniczejMarcin Zaród
mgr inż. Marcin Zaród: pracownik Centrum Badań Mole-kularnych i Makromolekularnych PAN, stażysta w Pracowni Przedmiotów Przyrodniczych IBE, edukator w programie „Za rękę z Einsteinem”; współpracownik łódzkiej świetlicy „Krytyki Politycznej” i Fundacji Nowoczesnej Edukacji „SPUNK”
Fizyka bez eksperymentów nie ma sensu. Co jednak zrobić, a czego unikać, by eksperyment łączył zainte-resowanie z poszerzaniem wiedzy uczniów? Ile czasu powinien trwać? Jak liczne powinny być grupy? Czy do-puścić osoby z zewnątrz? Czy uczniowie powinni pisać sprawozdania? Czy do wyjaśniania zjawisk warto mie-szać Kubicę i Dodę? Jakie narzędzia wykorzystać i jakie działy nauki wybrać do eksperymentu? Bardzo wiele wniosków na ten temat można wyciągnąć obserwując realizację programów edukacyjnych, takich jak „Za rękę z Einsteinem”, oraz działalność dużych centrów edukacyjnych (np. warszawskie Centrum Nauki Koper-nik, gdańskie Hevelianum, łódzkie Experymentarium).
„Za rękę z Einsteinem” – studium przypadku
Pierwsza edycja programu „Za rękę z Einstei-nem” była prowadzona w latach 2008–2012 na terenie województw pomorskiego, kujawsko-pomorskiego i warmińsko-mazurskiego. Organizatorem programu była Politechnika Gdańska. Program był adresowany do uczniów szkół w małych miejscowościach. W jed-nym roku szkolnym z programem współpracowało ok. 120 szkół gimnazjalnych. Projekt był finansowany ze środków Europejskiego Funduszu Społecznego. Bu-dżet projektu to ok. 39 mln zł.
Pokazy eksperymentów i wykłady popularnonau-kowe były prowadzone przez studentów i pracowników Wydziału Fizyki Technicznej i Matematyki Stosowanej (w tym przez autora artykułu). Prowadzący nie mieli przygotowania pedagogicznego. Byli to raczej fizycy--naukowcy niż fizycy-pedagodzy.
W przeciwieństwie do zajęć w Centrum Nauki Ko-pernik, eksperymenty w „Einsteinie” były dopasowane do programu nauczania w danej szkole. Z tego powo-du ich wartość jako „dobrej praktyki metodycznej” jest większa niż w przypadku zajęć w CNK. Celem „Ein-
Streszczenie:
Artykuł przedstawia uwagi metodyczne związane z organizacją i przebiegiem szkolnych doświadczeń fi-zycznych na poziomie gimnazjum i szkół ponadgim-nazjalnych w ramach obecnej podstawy programowej. Pokazane są w nim propozycje łączenia fizyki z innymi przedmiotami przyrodniczymi, techniką i otocze-niem codziennym uczniów. Artykuł zawiera przykład organizacji lekcji eksperymentalnej na podstawie doświadczeń z elektrycznością na etapie gimnazjum. Przykłady i praktyki prezentowane w artykule są oparte na doświadczeniach edukacji pozaformalnej, takich jak akademickie programy popularyzatorskie i działania edukacyjne organizacji pozarządowych.Słowa kluczowe: eksperyment uczniowski, fizyka, edukacja pozaformalna, popularyzacja nauki, elektryczność
otrzymano: 15.11.2012; przyjęto: 24.04.2013; opublikowano: 28.06.2013
steina” nie była wyłącznie zabawa w fizykę, lecz także doskonalenie mniej efektownych kompetencji (np. liga zadaniowa, obliczenia uczniowskie w trakcie pokazów).
Głównym narzędziem dydaktycznym były dodat-kowe zajęcia z fizyki i matematyki prowadzone na te-renie szkół. Pojedynczy pokaz doświadczeń lub łami-główek matematycznych trwał ok. 3 godziny lekcyjne, po których następowała rotacja grup uczniowskich. Część aparatury zakupiono na potrzeby projektu, ale większość wypożyczono z politechniki lub skonstruo-wano samodzielnie. Większa część zajęć skupiała się na prostych eksperymentach z mechaniki i elektryczności. Największe zainteresowanie budziły doświadczenia z elektrostatyki (rozładowywanie butelki lejdejskiej, przesyłanie iskry przez dotyk dłońmi), mechaniki (efekt żyroskopowy, rozkład sił na rolce nici, równowaga sił) i termodynamiki (balony na ciepłe powietrze, modele silników). Eksperymenty realizowane w centrach edu-kacyjnych również koncentrują się na tych dziedzinach, ze względu na ich atrakcyjność wizualną i łatwość prze-łożenia wyników na życie codzienne widzów.
Podczas ferii i wakacji dla uczniów z wybranych klas zorganizowano kolonie naukowe w Gdańsku. Uczest-nicy nocowali w akademikach politechniki i mogli wy-konywać eksperymenty w laboratoriach studenckich (niewykorzystywanych w czasie wakacji). Po południu uczniowie zwiedzali miasto lub brali udział w progra-mie kulturalnym. Projekt przewidywał również udział uczniów w Bałtyckim Festiwalu Nauki i innych im-prezach popularyzatorskich. Laboratoria realizowa-ne w trakcie wakacji i ferii stanowiły specjalną wersję „Pierwszej pracowni fizycznej”, przystosowaną do po-trzeb uczniów. Wykorzystanie zaplecza uczelni pozwo-liło na większą liczbę eksperymentów akustycznych, optycznych i termodynamicznych z użyciem aparatury (np. przetwornik analogowo-cyfrowy, obieg skraplają-cy, ciemnia optyczna).

Małe eksperymenty, wielkie odkrycia | Marcin Zaród | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 56
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Nauczyciele ze szkół uczestniczących w projekcie brali udział w warsztatach związanych z eksperymen-tami fizycznymi oraz pełnili funkcję wychowawców na wyjazdach uczniowskich. Budżet projektu miał przewi-dziane dodatkowe środki na wynagrodzenia i szkolenia dla nauczycieli.
Eksperyment w szkole i poza szkołą – zasoby i oceny
Przenoszenie doświadczeń metodycznych między różnymi formami edukacji przyrodniczej nie zostało do-tychczas wystarczająco opisane w literaturze. Nie moż-na więc bezpośrednio przenosić wniosków płynących z edukacji pozaformalnej do postulatów metodycznych dla edukacji formalnej. Jednak wnioski płynące z czte-rech lat pracy w kilkudziesięciu szkołach mogą stanowić dobry punkt otwarcia takiej dyskusji na łamach EBiŚ.
Największą różnicą jest znacznie większa ilość cza-su, jaki realizatorzy programów pozaszkolnych mogą poświęcić na eksperymenty. Oprócz tego wspomniany wcześniej projekt dysponował zasobami leżącymi poza możliwościami wielu szkół w Polsce (6 animatorów zajęć pracujących z uczniami 2–6 godzin przez jeden dzień, nowe wyposażenie eksperymentalne). Nie spo-sób wymagać od nauczycieli eksperymentów, na które „Einstein” miał wielokrotnie większy budżet.
Ostatni element, na który warto zwrócić uwagę, to fakt, że „Einstein” funkcjonował poza codzienną ruty-ną szkolną. Nauczyciele wielokrotnie odnotowywali, że w trakcie pokazów uczniowie byli bardziej aktywni niż podczas zwykłych lekcji. Zdecydowanie chętniej zada-wali pytania i stawiali własne hipotezy. Zgodnie z rela-cjami pedagogów: rozbudzenie ciekawości poznawczej utrzymywało się przez ok. 2–4 tygodni zwykłych lekcji fizyki w okresie powakacyjnym.
Z tego powodu uważam, że rozbudzenie ciekawości poznawczej i pasji eksperymentatora jest najważniejszą funkcją szkolnego eksperymentu. Eksperyment, jako lekcja niecodzienna (niestety!) i nieobarczona negatyw-nymi skojarzeniami poznawczymi, daje szansę na wy-rwanie uczniów z monotonii.
Pamiętając o tych różnicach między dydaktyką formalną i pozaformalną, postaram się znaleźć spo-sób przeniesienia tego entuzjazmu również na zwykłe lekcje. Pamiętając o tych różnicach, spróbujmy użyć dydaktyki nieformalnej jako „poligonu metodycznego”.
Organizacja lekcji doświadczalnej
W trakcie realizacji omawianego programu testo-wane były różne modele zajęć eksperymentalnych. Na ich podstawie można sformułować następujące wnioski dla tego typu zajęć prowadzonych w szkołach: • Pokazy w formie wykładu, dla publiczności siedzą-
cej na krzesłach okazały się mniej interesujące niż zajęcia umożliwiające ruch uczniów dookoła sta-nowisk eksperymentalnych.
• Pojedynczy eksperyment nie powinien trwać dłużej niż 20 minut, po których następuje rotacja uczniów.
• W jednej grupie nie powinno być więcej niż czte-ry osoby (optymalnie trzy), aby każdy uczeń miał szansę osobiście dokonać pomiaru.
• Dla jednej klasy potrzeba więc 4–8 stanowisk po-miarowych.
• Optymalna długość lekcji eksperymentalnej to 90 minut bez przerwy, co jest trudne do osiągnięcia w gimnazjum. Proste eksperymenty da się przygo-tować i wykonać w ciągu 45 minut, jeśli nauczyciel przygotuje ławki i sprzęt na przerwie.
• Ponieważ jeden prowadzący to za mało, aby efek-tywnie koordynować lekcję laboratoryjną dla kilku grup, wskazane jest wsparcie 2–5 uczniów
pomagających nauczycielowi (tutorów, animato-rów, pomocników). Na moich zajęciach używałem określenia „tutor/tutorka”, by podkreślić nieco-dzienność lekcji eksperymentalnej i wyjątkowy status tych uczniów.
• Optymalną proporcją jest dwóch animatorów na trzy stanowiska. Mogą być nimi stażyści, studen-ci z zaprzyjaźnionego koła naukowego, uczniowie lub rodzice. Najlepszą recepcją wśród uczniów cie-szyły się te zajęcia, w których tutorami byli sami uczniowie. W praktyce można to zrealizować przy wsparciu starszych roczników (uczenie młodszych to dobra powtórka przed egzaminem) lub na za-sadzie rotacji koleżeńskiej. Przykładowo: planując w roku szkolnym pięć lekcji eksperymentalnych, każdą klasę dzielimy na pięć grup. Kilka dni przed lekcją eksperymentalną nauczyciel przygotowu-je instrukcję i omawia przebieg zajęć z wybraną grupą uczniów. W trakcie lekcji członkowie grupy nie wykonują sami eksperymentu, ale pomagają kolegom w jego przeprowadzeniu. Wcześniejsze przygotowanie spotkania z pomocnikami pozwala oszczędzić czas i ułatwia prowadzenie lekcji.
• Lekcja eksperymentalna powinna być skoncentro-wana na jednym dziale fizyki. Doświadczenia na różnych stanowiskach pomiarowych mogą się po-wtarzać (np. dwa stanowiska do pomiaru przyspie-szenia ziemskiego i dwa stanowiska do pomiaru siły odśrodkowej), ale łączenie dziedzin utrudnia dyskusję wniosków.
• Zgodnie z doświadczeniem metodycznym autora: w trakcie lekcji eksperymentalnej uczniowie mu-szą mieć prawo stawiania błędnych hipotez. Po-nieważ popełnianie błędów jest naturalną częścią eksperymentu (również w fizyce akademickiej), uczniowie nie powinni być karani za błędy, złe in-terpretacje i mylne hipotezy.

Małe eksperymenty, wielkie odkrycia | Marcin Zaród | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 57
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Wnioski te uwzględniłem w scenariuszu lekcji eks-perymentalnej „Prawo Ohma w praktyce”. Próbowałem w nim po połączyć otwartość dydaktyki pozaformalnej z wymaganiami i realiami edukacji szkolnej.
Prawo Ohma w praktyce
Cel eksperymentu: Zbadanie prostych obwodów elektrycznych. Po-znanie różnicy między połączeniem szeregowym i równoległym.Etap edukacyjny: III – gimnazjum.Podstawa programowa: Fizyka dla III etapu edukacyjnego. Wymagania eksperymentalne 9.7, 9.8. i 9.9. Wymagania teoretyczne: 4.7, 4.8 i 4.9.Czas realizacji: 15 min. – przygotowanie, 65 min. – pomiary i 20 min. – dyskusja wyników i czynności administracyjne. Sugeruję wykorzy-stanie przerwy między lekcjami (za zgodą uczniów) na cele dydaktycz-ne – eksperymenty są rzadkie i warto wykorzystać cały możliwy czas. W razie potrzeby skrócenia proponuję wykreślenie jednego z pytań w części eksperymentalnej. W siatce godzin gimnazjum trudno o taki zakres czasu, ale rezygnacja z dyskusji i części eksperymentów ograni-cza efekt dydaktyczny lekcji eksperymentalnej. Wyposażenie (na jedno stanowisko): • źródło prądu: zasilacz prądu stałego o napięciu 12 V lub układ
kilku połączonych szeregowo baterii AA (przynajmniej 5 sztuk); w tym drugim przypadku połączenie najlepiej zrobić „na stałe” (np. doklejonymi blaszkami miedzianymi), aby nie rozpadło się w trakcie lekcji;
• przewody do połączenia elementów (najlepiej z dołączonymi „kro-kodylkami”);
• 3 żarówki na napięcie 12 V (o mocy 1,5 W, najlepiej różnokoloro-we), 1 żarówka o mocy 5 W, również na napięcie 12 V. Sugerowane jest użycie żarówek mających gwinty wtykowe (m.in. typu MR16) – np. COB5SE (prod. Lumenmax);
• 2 mierniki uniwersalne;• (nieobowiązkowe): komputer z arkuszem kalkulacyjnym.Kursywą podano sugerowaną narrację ćwiczenia dla uczniów. Fizy-
ka w formie fabularyzowanej jest łatwiejsza do zrozumienia niż suche polecenia.
1. Przygotowanie (na przerwie, zanim przyjdą uczniowie):
• ustawić ławki, rozłożyć wyposażenie stano-wisk na ławkach;
• sprawdzić zasilacze i mierniki; usunąć ewentualne usterki;
• wypróbować eksperyment z grupą anima-torów, poinstruować ich o zasadach bezpie-czeństwa i celach eksperymentu.
2. Eksperymenty:
• Podział uczniów na grupy (sugerowany jest podział mieszający uczniów z różnymi wynikami w nauce, aby słabsi uczyli się od mocniejszych).
• Wprowadzenie uczniów w tematykę ćwi-czenia:
Prąd elektryczny to dla nas źródło światła i ciepła. Elektryczność zasila nasze telefony, komputery, za-bawki i dzwonki w szkole. Dzisiaj poznacie podstawy inżynierii elektrycznej. To jest źródło prądu <wskazuje-my na zasilacz lub baterię>, to są żarówki różnej mocy <wskazujemy na żarówki>, to są przewody i mierniki <demonstrujemy>. Co zrobić aby żarówka świeciła?
• Uczniowie podłączają żarówkę o mocy 1,5 W, tutorzy i nauczyciel pomagają w mia-rę potrzeby.
Świetnie, zrobiliście właśnie pierwszy krok, by zostać inżynierami elektrotechniki. Co płynie w przewodach? Co to jest prąd elektryczny i jak zmierzyć jego natęże-nie? Czy prąd płynie w otwartym obwodzie?
• Uczniowie mierzą i notują natężenie prądu, pomocnicy asystują.
Co sprawia, że prąd płynie? Dlaczego zasilacz ma dwa różne zaciski? Co oznacza plus, a co minus? Czym róż-nią się te dwa zaciski? Co wytwarza zasilacz, kiedy ob-wód jest otwarty?
• Uczniowie mierzą i notują napięcie na za-silaczu w układzie otwartym (ostrożnie!). Mierzą też napięcie na żarówce 1,5 W.
Inżynierowie używają wielu urządzeń elektrycznych, mających różne funkcje. Spróbujcie sprawdzić, czym różnią się żarówki 1,5 W i 5 W. Czy są różnice w napię-ciu i natężeniu prądu płynącego przez żarówki o mocy 1,5 W w różnych kolorach?
W jaki sposób można podłączyć do prądu dwie żarów-ki 1,5 W? Jak to zrobić, aby obie świeciły najjaśniej?
• Uczniowie eksperymentują z połączeniami żarówek.
Co się stanie, gdy podłączę trzecią żarówkę do dwóch pozostałych? Co się stanie, gdy będzie to żarówka o większej mocy?
• Pomocnicy i nauczyciel demonstrują połą-czenie równoległe. Uczniowie mierzą prąd i napięcie w połączeniu równoległym na każdej z żarówek.
Co się stanie, gdy żarówki podłączymy w inny sposób? Jak inaczej można je połączyć? Jak połączyć 3 żarówki o mocy 1,5 W, aby przez każdą z nich płynął taki sam prąd?
• Uczniowie eksperymentują z różnymi kon-figuracjami połączeń. Animatorzy i nauczy-cie asystują
Spróbujcie narysować obwody układów, które skon-struowaliście. Nanieście na schematy wyniki pomia-rów napięcia i natężenia prądu.
3. Dyskusja:
Wypróbujmy różne sposoby podłączenia żarówek? Jak wtedy mierzyć napięcia i natężenia? Jak będą różniły się wartości pomiarów? W którym połączeniu przez wszystkie żarówki płynie taki sam prąd?
• Uczniowie odpowiadają na pytania zadane przez nauczyciela w trakcie lekcji, nauczy-ciel moderuje dyskusję. Tutorzy wprowa-dzają wyniki pomiarów uczniowskich do komputera lub wypisują je na tablicy.
PRZYKŁADOWA LEKCJA

Małe eksperymenty, wielkie odkrycia | Marcin Zaród | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 58
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Fizyka codzienna
Autorzy programów pozaszkolnych, podobnie jak twórcy reformy programowej, postulują odejście od ma-tematyzacji fizyki w gimnazjum. Rezygnacja z wzorów i zadań i zwiększenie nacisku na wyjaśnienie procesów nie powinno jednak ograniczać się do lekcji ekspery-mentalnych i dyskusji teoretycznych. W „Einsteinie” dobre wyniki odnosiliśmy przy pomocy schematycz-nych rysunków i dyskusji na temat fizyki życia codzien-nego.
Z punktu widzenia ucznia Małysz, Kubica czy Doda są dużo bliższymi osobami niż Newton, Euler czy Hertz. Z tego powodu zamiast tłumaczyć fizykę na poziomie abstrakcyjnej teorii, lepiej jest odwołać się do fascynacji uczniów. Przykładowo: w jednej ze szkół uczniowie byli zainteresowani motoryzacją. Wykorzystanie przykła-du ostatniego wyścigu Roberta Kubicy do pytań o siłę odśrodkową i tarcia było dużo bardziej efektywne niż pytanie wprost o abstrakcyjne pojęcia.
Fizyka szkolna nie istnieje w próżni, więc zamiast abstrakcyjnych definicji lepiej szukać przykładów bliż-szych codzienności ucznia. Fizyka sportu, nagłośnienie koncertu, silna wichura lub głośnik komórki – nauczy-ciel powinien przed lekcją znaleźć przykład zastosowa-nia fizyki w otoczeniu uczniów, na którym oparta zosta-nie cała narracja lekcji. Zamiast teorii gazu doskonałego lepiej pokazać zamknięte pudełko z grochem i skłonić uczniów do zgadywania liczby ziarenek w środku. Po-tem porównujemy to z wielkością sali lekcyjnej i sza-cujemy liczbę atomów azotu w pomieszczeniu. Intere-sujące poznawczo jest też szacowanie wagi powietrza w klasie.
Doświadczenia edukatorów „Einsteina” pokazały, że uczniowie dużo chętniej uczestniczą w dyskusjach, eksperymentach i zajęciach, jeżeli uznają ich cel za wi-doczny w ich życiu. Wykłady dotyczące teoretycznych
konstrukcji spotkały się w programie z mniejszych za-angażowaniem niż fizyka codzienna.
Od teorii fizycznej do praktyki uczniowskiej
Każdy z działów fizyki ma bezpośrednie przełoże-nie na praktykę techniczną. Konstrukcja przykładów powinna skupiać się na przedmiotach z otoczenia ucz-niów, a nie na obiektach abstrakcyjnych. Wykorzystanie obiektów materialnych do kształtowania „wyobraźni fi-zycznej” poprawia efektywność procesów poznawczych i zachęca uczniów do uczestnictwa w lekcji. W poniż-szym rozdziale podam źródła inspiracji, z których ko-rzystałem przy przygotowywaniu warsztatów i prelekcji w programie „Za rękę z Einsteinem”.
Mechanikę (Podstawa programowa dla III etapu edukacyjnego punkty: 1. i 2.1–2.5, 9.2, 9.4) najłatwiej powiązać z wyścigami samochodowymi, konstrukcją domów i maszyn. Wyścigi samochodowe są natural-nym punktem wyjścia do dyskusji na temat tarcia, siły odśrodkowej, przyspieszenia i prędkości.
Fale i drgania (Podstawa programowa dla III etapu edukacyjnego punkt 6, 9.12 i 9.13) są bardziej proble-matyczne, zwłaszcza że eksperymenty z dyfrakcją i in-terferencją są trudniejsze i mniej efektowne. Zamiast eksperymentów z falami stojącymi (smyczek i sól) le-piej skorzystać z symulacji komputerowych dostępnych w Internecie. Zazwyczaj film lub program komputero-wy nie stanowi pełnowartościowego zamiennika żywe-go eksperymentu, ale dla tego działu fizyki warto zrobić wyjątek. Można też użyć skakanki do dyskusji o falach stojących (aut. pomysłu: Włodzimierz Natorf1).
Zajęcia z optyki (punkty 7 i 9.14) są równie trud-ne, chociaż łatwiej o konkretne przykłady zastosowań.
1 Nauczyciel fizyki w IX LO im. Klementyny Hoffmanowej w War-szawie, eskpert współpracujący z Pracownią Przedmiotów Przy-rodniczych IBE.
Dyskusja wyników eksperymentu
Eksperyment bez dyskusji wyników jest dużo mniej efektywny edukacyjnie, jednak pisemne sprawozdania nie sprawdziły się w „Einsteinie” i – zdaniem autora – nie powinny być stosowane w szkołach. Zamiast spraw-dzania prac domowych lepiej przeznaczyć czas lekcji na dyskusję wyników, zwłaszcza że kontakt z praktyką daje szansę na pobudzenie ciekawości uczniów.
Nauczyciel powinien stawiać pytania otwarte, naj-lepiej związane z ogólnym mechanizmem (np. „Po co nam izolacja w pomiarze ciepła właściwego wody?” za-miast „Jakie macie wyniki?”). Aby tutorzy nie zdomi-nowali dyskusji, można przydzielić im rolę „Telefonu do przyjaciela”, z którego grupa korzysta tylko wtedy, gdy nie ma lepszych pomysłów. Nauczyciel staje się wtedy raczej moderatorem niż źródłem wiedzy. Jeśli w klasie jest komputer, to jeden z animatorów może szukać od-powiedzi na trudniejsze pytania bez konieczności prze-rywania dyskusji całej klasy.
Dokładny wynik eksperymentu nie powinien być oceniany, podobnie jak przebieg procesu myślowego uczniów. Bycie ocenianym może ograniczać pomysło-wość i ciekawość poznawczą uczniów, co zmniejsza sku-teczność lekcji eksperymentalnej. Oczywiście wiedza zdobyta w trakcie lekcji eksperymentalnej może być przedmiotem oceny w trakcie sprawdzianu z danego działu.
Istotniejsze jest zaangażowanie w dyskusję i zrozu-mienie mechanizmu eksperymentu. Eksperyment nie powinien być pierwszą lekcją z danego działu, lepiej sprawdza się jako druga lub trzecia. Uczniowie mają wtedy podstawową wiedzę teoretyczną i ciekawość po-znania nowych rzeczy. Eksperyment na końcu cyklu sprawdza się słabiej, ze względu na słabsze zaintereso-wanie uczniów, którzy już poznali całą teorię na dany temat.

Małe eksperymenty, wielkie odkrycia | Marcin Zaród | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 59
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Można je planować zimą, kiedy łatwiej o uzyskanie ciemności w klasie szkolnej. Programy pozaszkolne kładą na optykę mniejszy nacisk niż podstawa progra-mowa. Uczniowie wykonywali doświadczenia z optyki tylko w ramach pobytu na politechnice, gdy mieli do-stęp do odpowiednich przyrządów. Eksperymenty są dłuższe i wymagają zaciemnionych pomieszczeń.
Zmniejszona liczba eksperymentów z falami i z op-tyką może zostać zrekompensowana przez dodatkowe eksperymenty z termodynamiką (2.5–2.11) i elektrycz-nością (9.7, 9.8, 9.9). Te działy fizyki są częściej wykorzy-stywane w technice, więc łatwiej o przykłady praktycz-ne i zainteresowanie uczniów.
Eksperymenty z elektrycznością stanowią stały punkt pokazu fizycznego, bo taniejące mierniki uni-wersalne i przyrządy elektroniczne zachęcają do tego, aby ten dział stał się wizytówką szkolnej fizyki. Zamiast pomiaru abstrakcyjnej mocy (9,9 W) dużo łatwiej tra-fić do uczniów z doświadczeniem porównującym koszt eksploatacji dwóch żarówek. Termodynamika świetnie splata się z chemią (reakcje spalania w naturalny sposób łączą się z cyklami termodynamicznymi). Jeżeli nauczy-ciel fizyki jedzie z klasą na wycieczkę, to jezioro, góry, las lub morze obfitują w materiał do dyskusji. Twardość kamieni, zachowanie fal morskich, mechanizm bryzy lub obieg wody w przyrodzie stanowią naturalny łącz-nik fizyki z chemią, biologią lub geografią.
Podsumowanie
Doświadczenia zebrane w trakcie festiwali nauki czy realizacji programów popularyzatorskich mogą stanowić źródło przydatnej w szkole wiedzy na temat sposobów przeprowadzania eksperymentów. Przywró-cenie w podstawie programowej do gimnazjum, fizyki eksperymentalnej to szansa na to, by szkolna fizyka była równie fascynująca, co jej akademicka odmiana.
Jeżeli nauczyciel gimnazjum ma problemy z kon-strukcją lekcji eksperymentalnej, może zwrócić się do studenckich kół naukowych na pobliskich uczelniach. Jest szansa, że ich członkowie dysponują wiedzą i za-pleczem technicznym niezbędnym do realizacji takich działań.
Small experiments, great discoveries. The practice of experiential science education
Marcin Zaród
This article presents various teaching methods that in-clude an experimental approach to the gymnasium and high-school physics curriculum. Various methods of an interdisciplinary approach between physics and other sci-ences, technology and the students’ everyday life are pre-sented. A sample laboratory lesson focusing on electrical circuits in the gymnasium curriculum is also included. Cases presented in the article are based on informal edu-cation practices, such as academic science encouragement programmes and NGO experiences.Key words: experiment for students, physics, non-formal edu-cation, popularisation of science, electricity

Angielskie narzędzia dydaktyczne | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 60
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Jednym z rezultatów wizyty studyjnej pracowników Pracowni Przedmiotów Przyrodniczych IBE, odby-tej w 2010 r., był duży zasób narzędzi dydaktycznych, uzyskanych od nauczycieli w wizytowanych szkołach i ośrodkach doskonalenia nauczycieli.
Narzędzia zbierano głównie po to, by porównać np. konstrukcję konspektu zajęć z obserwowaną przez badaczy realizacją tych zajęć podczas lekcji. Dodatko-wym, równie ważnym celem było analityczne porów-nanie tych narzędzi. Chodziło o ustalenie, czy są istot-ne różnice między narzędziami stosowanymi w Polsce i w Anglii., a jeśli tak, to na czym te różnice polegają. Miało to doprowadzić do uzyskania odpowiedzi na pytanie, czy istnieje korelacja między konstruktem narzędzia dydaktycznego a sposobem prowadzenia za-jęć przyrodniczych, głównie pod kątem kształtowania umiejętności posługiwania się metodą naukową.
Uzyskane materiały pochodzą z różnych źródeł: niektóre zostały wydane przez angielskie wydawnictwa oświatowe, inne są efektem pracy samych nauczycieli lub współpracujących z nimi pracowników technicz-nych (narzędzia autorskie).
Angielskie narzędzia dydaktyczneRefleksje z wizyty studyjnejUrszula Poziomek
mgr Urszula Poziomek: specjalista ds. badań i analiz Pracowni Przedmiotów Przyrodniczych IBE, nauczycielka dyplomowana biologii w LXXV LO im. Jana III Sobieskiego w Warszawie; współautorka podręczników szkolnych do nauki biologii na III i IV etapie edukacyjnym, współautorka raportu Eurydice Science Education in Europe, 2011.
Narzędzia pozyskiwane były w różnorodny spo-sób: po zajęciach lekcyjnych nauczyciele proszeni byli (po uprzednim wyjaśnieniu celu) o kopie materiałów, narzędzi, które były używane w trakcie lekcji. Również pracownicy techniczni proszeni byli o takie materiały, w szczególności autorskie lub pochodzące z wydawnictw instrukcje do doświadczeń, karty pracy ucznia itp. Zespół zwracał się z prośbą o te materiały również do przewodniczących zespołów przyrodniczych, którzy chętnie dzielili się swoimi zasobami.
W rezultacie uzyskano karty pracy ucznia, kartoteki nauczyciela, scenariusze lekcji, instrukcje doświadczeń przeznaczonych do realizacji w ramach różnorodnych tematów z science, biologii, chemii i fizyki dla poziomu ISCED 2 i 3 (odpowiednio dla polskich dwóch etapów edukacyjnych – III i IV). Dodatkowo uzyskano odtaj-nione testy egzaminacyjne dla III (science) i IV etapu edukacyjnego (chemia, biologia).
Najbardziej zainteresowały badaczy autorskie na-rzędzia w postaci np. instrukcji dla ucznia realizującego projekt w warunkach domowych czy karty samooceny ucznia.
Pozyskane narzędzia dydaktyczne zostały opisane pod względem zgodności z polską podstawą programo-wą właściwego przedmiotu, a także pod względem ich odmienności względem narzędzi stosowanych w pol-skich szkołach.
Wśród materiałów – narzędzi dydaktycznych do na-uczania biologii za godne uwagi ze względu na cechy nieobecne lub niewystarczająco reprezentowane w pol-skiej szkole uznano:
Dla poziomu ISCED2 czyli III etapu edukacyjnego
• Karty pracy ucznia, z pomocą których uczeń wy-konuje pewne zadania, dokonując wcześniej wy-boru poziomu, na jakim chce je zrealizować. Taki sposób oceny przez ucznia własnych możliwości można z powodzeniem stosować zarówno na lek-cjach, jak i w pracy metodą projektową. Schemat konstrukcyjny takiego materiału może służyć również przygotowywaniu narzędzi dla uczniów o specjalnych potrzebach edukacyjnych – zarów-no tych wymagających wsparcia, jak i szczególnie uzdolnionych.
• Protokoły doświadczeń lub obserwacji w formie kart pracy ucznia i instrukcji dla nauczyciela, któ-rych cechą charakterystyczną i w pewnym stopniu odmienną wobec polskich narzędzi dydaktycz-nych jest duża liczba pytań badawczych, które stopniowo prowadzą ucznia do wniosku końcowe-go opartego o analizy poszczególnych materiałów źródłowych czy danych pozyskanych z doświad-czenia lub obserwacji,
• Testy General Certificate of Secondary Education – sprawdzające poziom opanowania umiejętno-ści prostych i złożonych zadaniami wyłącznie za-mkniętymi, przy czym interesująca jest złożona konstrukcja zadań, diagnozujących poziom umie-jętności złożonych: zbudowane są one z reguły z kilku podpunktów, które z różnych stron, w róż-nych aspektach sprawdzają znajomość i rozumie-nie jednego problemu. Większość zadań (szcze-gólnie z poziomów wyższych) dotyczy metody naukowej.
Wizyta studyjną odbyto w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz insty-tucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.

Angielskie narzędzia dydaktyczne | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 61
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Dla poziomu ISCED3 czyli IV etapu edukacyjnego:
• Seria protokołów doświadczalnych w formie kart pracy ucznia i instrukcji dla nauczyciela, dotyczą-cych jednego obszaru treści podstawy programo-wej – np. enzymów i ich działania. Jest to przykład nauczania przez badanie – wielostronne i wielo-aspektowe – jednego zagadnienia, służące rzeczy-wistemu pogłębianiu wiedzy przyrodniczej z wy-korzystaniem metody laboratoryjnej. Tego typu działania są realizowane przez nauczycieli biologii w Polsce – odbywa się to jednak z reguły na za-jęciach pozalekcyjnych w ramach kół zaintereso-wań, przygotowania do udziału w olimpiadzie bio-logicznej czy też w ramach realizowania projektów grantowych z wykorzystaniem funduszy unijnych. Nie są one natomiast wpisane w codzienną realiza-cję podstawy programowej na zajęciach obowiąz-kowych głównie ze względu na nadal rozbudowane treści nauczania.
Wśród narzędzi dydaktycznych służących do na-uczania science za interesujące uznano uzyskane w The Holt School Science Department:
Dla poziomu ISCED2 czyli gimnazjum:
• Zadania testowe sprawdzające umiejętności złożone u uczniów, mające formę tzw. structured questions, w których uczniowie, na podstawie załączonego materiału źródłowego, mają za zada-nie rozwiązać problemy przedstawione w formie krótkich pytań (tzw. zadania z lukami). Ponadto powinni się wykazać umiejętnością korzystania z informacji i rysowania prostych schematów. Ce-chą wyróżniającą te zadania jest także ich odnie-sienie do sytuacji, które mogą wystąpić w życiu codziennym młodego człowieka.
Dla poziomu ISCED3:
• Karty pracy dla uczniów1, zawierające zadania składające się z dosyć obszernego tekstu źródło-wego, wzbogaconego zdjęciami i schematami, do którego dołączone są pytania kontrolne. Na margi-nesie są także wypunktowane spodziewane efekty kształcenia, wśród których dominują kształcone u uczniów umiejętności złożone. Załączono też kilka kart pracy o zróżnicowanym poziomie trud-ności. Kształcą one umiejętności złożone. Ciekawe są rozwiązania graficzne – począwszy od wyci-nanki, przy pomocy której uczniowie sami tworzą schemat przesyłu elektryczności, po analizę „przy-jemnych do percepcji” rysunków animowanych, aż po pytanie sprawdzające umiejętność uczenia się. Zwraca uwagę fakt, że wszystkie zadania na-wiązują do rzeczywistości otaczającej ucznia.
Nie chcąc ograniczać się jedynie do opisu tych na-rzędzi, zaprezentowano poniżej przykład konspektu lekcji wraz z trzema kartami pracy uczniów A, B i C, który wykorzystuje jako źródło inspiracji narzędzia an-gielskie. Pozwala on uczniom wybrać poziom zaawan-sowania, który chcą realizować w określonym temacie. Na przykładzie tego konspektu i kart pracy w wersji A, B i C można budować podobne materiały, tak by roz-wijać nie tylko umiejętności przedmiotowe ale również kompetencje społeczne w postaci trafnej samooceny swoich możliwości i podejmowaniu się zadań „na włas-ną miarę”.
Oczywiście takie podejście dydaktyczne rodzi na-tychmiast problem związany z ocenianiem – czy uczeń który wybierze poziom najniższy powinien dostać do-stateczny lub dopuszczający a ten który wybierze po-ziom najwyższy ocenę bardzo dobry? A może na każ-
1 Edexcel GCSE Science – Sample Material. Teaching and Learning Resources for the New GCSE Science 2011 specifications
dym poziomie uczeń ma szanse dostać ocenę z pełnej skali w zależności od stopnia realizacji i poprawności wykonania zadań?
Pozostawiam te pytania bez odpowiedzi i zapra-szam do dyskusji na profilu EBiŚ na facebook.com.
Źródło: Przedmioty przyrodnicze w wybranych krajach – raport z wizyty studyjnej w Anglii, http://eduentuzjasci.pl/pl/publikacje.html?start=5

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 62
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Temat: Łańcuchy i sieci pokarmowe w polu
Adresaci: uczniowie gimnazjum, III etap edukacyjny
Czas trwania zajęć: 2 x 45 minut
Cele kształcenia: • umiejętności (wymagania ogólne): IV. Rozumowanie i argumentacja. Uczeń interpretu-
je informacje i wyjaśnia zależności przyczynowo--skutkowe między faktami, formułuje wnioski.
• wiadomości (wymagania szczegółowe):IV. Ekologia. 8) wskazuje żywe i nieożywione elementy ekosyste-
mu, wykazuje, że są one powiązane różnorodnymi zależnościami,
9) opisuje zależności pokarmowe (łańcuchy i sieci po-karmowe) w ekosystemie, rozróżnia producentów, konsumentów i destruentów oraz przedstawia ich rolę w obiegu materii i przepływie energii przez ekosystem.
Metody pracy: słowna (analiza materiałów źródł.)
Formy pracy: indywidualna
Środki dydaktyczne: • karta pracy ucznia, zawierająca materiały źródło-
we
Typ zajęć: lekcja utrwalająca
Przebieg zajęć:
Faza wprowadzająca (10’)
a) Nauczyciel::• przypomina strukturę łańcucha pokarmowego na
dowolnym przykładzie, pokazuje możliwe powią-zania między dwoma lub trzema łańcuchami po-karmowymi z tego samego ekosystemu – pola;
• formułuje problem badawczy:
W jaki sposób naturalne relacje pokarmowe (kto kogo zjada?) między organizmami wpływają na produktyw-ność rolnictwa czyli ilość pokarmu, jaką można uzyskać z pola lub sadu?
• określa „Nacobezu” (na co będę zwracać uwagę): dokonanie wyboru poziomu realizacji zadań, rze-telność samooceny, umiejętność wykorzystania pozyskanych z karty pracy informacji do opraco-wania wyników analizy;
• informuje o możliwości wyboru poziomu opraco-wania – od najtrudniejszego – karta A, do najła-twiejszego – karta C;
• zachęca uczniów, by podejmowali decyzję o wybo-rze po zastanowieniu się, na ile czują się przygoto-wani do tematu lekcji.
Łańcuchy i sieci pokarmowe na polu – zajęcia z biologiiKonspekt i karta pracyUrszula Poziomek
Na podstawie Farming and food webs, Badger Publishing Ltd.
Prace U. Poziomek zostały wykonane w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowa-nego ze środków Europejskiego Funduszu Społecznego.
mgr Urszula Poziomek: specjalista ds. badań i analiz Pracowni Przedmiotów Przyrodniczych IBE, nauczycielka dyplomowana biologii w LXXV LO im. Jana III Sobieskiego w Warszawie; współautorka podręczników szkolnych do nauki biologii na III i IV etapie edukacyjnym, współautorka raportu Eurydice Science Education in Europe, 2011.
b) Uczniowie: • słuchają nauczyciela, zadają pytania, wybierają po-
ziom karty, z którą chcą pracować.
Faza realizacyjna:
a) Nauczyciel:• rozdaje uczniom wybrane przez nich karty pracy
(A, B lub C), prosi o zapoznanie się z nimi;• w trakcie pracy uczniów służy pomocą, pomaga
w razie potrzeby w wyborze właściwej odpowiedzi, zadając dodatkowe pytania, pomagające zrozu-mieć problem;
• udziela rzeczowej informacji zwrotnej. b) Uczniowie:• zapoznają się z zawartością kart pracy, zadają
w miarę potrzeby dodatkowe pytania, dotyczące realizacji zadań;
• wykorzystując informacje z karty pracy realizują wybrany poziom zadań.
Faza podsumowująca
a) Nauczyciel:• prosi wybranych lub chętnych uczniów o przedsta-
wienie rezultatów pracy w taki sposób, by zostały omówione wszystkie zadania ze wszystkich trzech kart pracy.
b) Uczniowie:• chętni lub wybrani przez nauczyciela uczniowie
przedstawiają rezultaty pracy.
Uwagi:
Uwzględniono trzy karty, różne dla trzech poziomów opracowań – A, B i C.

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 63
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom A
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 64
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
1. Narysuj schemat sieci pokarmowej, łącząc ze sobą wszystkie organizmy (żyjące na polu). Zastosuj w schemacie maksymalną liczbę możliwych do zbudowania łańcuchów pokarmowych.
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Użyj do opisu organizmu maksymalną liczbę pasujących do niego terminów.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz wpływ pestycydów na każdą z grup organizmów, uwzględnionych w sieci pokarmowej, narysowanej przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Na schemacie sieci pokarmowej, sporządzonej przez Ciebie w punkcie 1 narysuj źródło energii i oznacz strzałkami kierunki jej przepływu przez ekosystem pola.
6. Wyjaśnij szczegółowo, jak użycie przez rolnika pe-stycydów na polu może wpłynąć na funkcjonowa-nie krogulców.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
7. Sformułuj i zapisz kilka problemów, które mogą pojawić się w funkcjonowaniu ekosystemu pola po zastosowaniu na nim pestycydów oraz zaproponuj co najmniej 3 rozwiązania alternatywne, które po-mogą chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
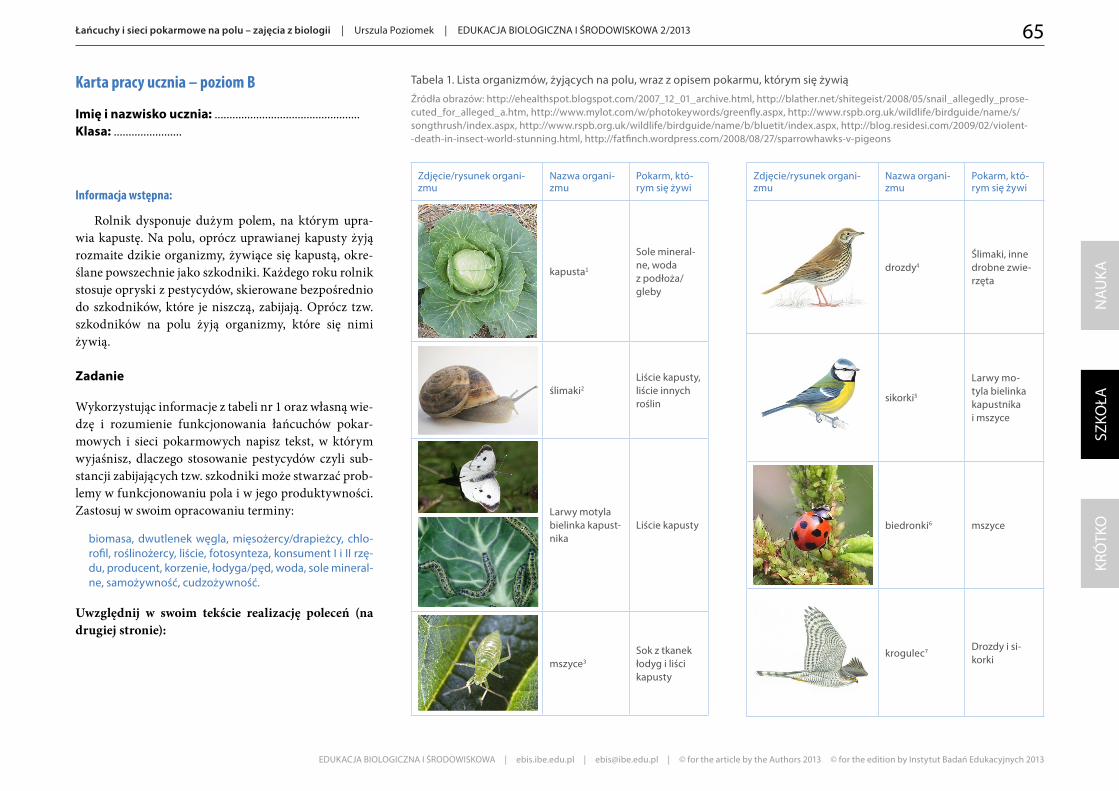
Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 65
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom B
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 66
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
1. Narysuj schemat sieci pokarmowej, łącząc ze sobą wszystkie organizmy żyjące na opisanym polu). W sieci powin-no być uwzględnionych minimum 3 łańcuchy pokarmowe (może być ich więcej).
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Powinieneś użyć więcej niż jednego terminu do opisu organizmu.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz wpływ pestycydów na każdą z grup organizmów, uwzględnionych w sieci pokarmowej, narysowanej przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Na schemacie sieci pokarmowej, sporządzonej przez Ciebie w punkcie 1 narysuj źródło energii i oznacz strzałkami kierunki jej przepływu przez ekosystem pola.
6. Sformułuj i zapisz kilka problemów, które mogą pojawić się w funkcjonowaniu ekosystemu pola po zastosowaniu na nim pestycydów oraz zaproponuj co najmniej 2 rozwiązania alternatywne, które po-mogą chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 67
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom C
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons

Łańcuchy i sieci pokarmowe na polu – zajęcia z biologii | Urszula Poziomek | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 68
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
1. Narysuj prostą sieć pokarmową, łączącą trzy różne łańcuchy pokarmowe z opisanego pola, uwzględniając w nich organizmy na nim żyjące.
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Możesz użyć kilku terminów opisując jeden organizm.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz efekty stosowania pestycydów na pozostałe organizmy tworzące sieć pokarmową, narysowaną przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Sformułuj i zapisz jeden problem, który może po-jawić się w związku ze stosowaniem pestycydów na polu i zaproponuj jedno alternatywne rozwią-zanie, które pomoże chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................

Nowe zadania PPP | Marcin Chrzanowski, Wojciech Grajkowski, Krzysztof Horodecki, Małgorzata Musialik, Maciej Wiśniewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 69
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Nowe zadania PPPMateriał przygotowują pracownicy Pracowni Przedmiotów Przyrodniczych IBE oraz eksperci zewnętrzni
Prezentowane tu zadania pochodzą z testów wy-korzystywanych w badaniu Laboratorium Myślenia. Diagnoza nauczania przedmiotów przyrodniczych w Polsce, realizowanym przez Pracownię Przedmiotów Przyrodniczych IBE. Po każdym cyklu badania (odbyły się już dwa takie cykle – w roku 2011 i 2012) część zadań jest odtajniana i są one publikowane na stronie Bazy Narzędzi Dydaktycznych IBE – bnd.ibe.edu.pl. Intencją takiego działania jest prezentacja środowisku nauczy-cielskiemu, uczniom, ich rodzicom, a wreszcie twórcom narzędzi diagnostycznych zadań, które mierzą umie-jętności ujęte w podstawie programowej przedmiotów przyrodniczych, w tym umiejętność rozumowania naukowego, posługiwania się metodą naukową w co-dziennym życiu, analizy danych, wnioskowania czy identyfikacji związków przyczynowo-skutkowych.
Więcej o badaniu Laboratorium Myślenia na stro-nie: http://eduentuzjasci.pl/pl/badania.html?id=409
Zadania powstały w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Biologia – które ciastko jest smaczniejsze?
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Komentarz
Projektowanie eksperymentów jest umiejętnością ponadprzedmiotową, a zagadnienia dotyczące formu-łowania pytania badawczego, ustalania warunków do-świadczenia czy doboru odpowiedniej próby kontrol-nej są uniwersalne i mają zastosowanie we wszystkich naukach eksperymentalnych. Opisane doświadczenie nie jest związane z konkretnymi treściami nauczania i trudno je nawet jednoznacznie przypisać do określo-nego przedmiotu. Realny, „życiowy” kontekst zadania pozwala natomiast pokazać, że zasady projektowania
eksperymentów są takie same dla badań naukowych i dla sytuacji z życia codziennego.
Pierwszym zagadnieniem poruszanym w zadaniu jest dobór właściwej próby. Ze wstępu uczeń dowiaduje się, że próbę tę stanowi „zaprzyjaźniona wielopokole-niowa rodzina”, a zasadność takiego doboru badanych ocenia, udzielając odpowiedzi w dwóch pierwszych wierszach tabeli. Pierwszy „element doświadczenia” – różny wiek badanych – jest pożądany dla rzetelności badania. Gust dzieci często bardzo wyraźnie różni się od gustu osób dorosłych, dlatego jeśli cukiernik chciał uzyskać możliwie pełną odpowiedź na pytanie, które ciastko jego potencjalni klienci uważają za smaczniej-sze, powinien przetestować osoby w różnym wieku. W punkcie pierwszym właściwa jest zatem odpowiedź „Tak” i zdecydowana większość (87,1%) absolwentów gimnazjum, którzy rozwiązywali to zadanie, tak właś-nie odpowiedziała.
W punkcie drugim należało odnieść się do infor-macji mówiącej, że wszyscy badani byli ze sobą spo-krewnieni. Jest to okoliczność obniżająca wiarygodność badania, należało zatem zaznaczyć odpowiedź „Nie”. Członkowie jednej rodziny mogą bowiem teoretycz-nie wynosić z domu, czy wręcz dziedziczyć, określone preferencje żywieniowe. Grupa bliskich krewnych jest zresztą złą reprezentacją populacji w niemal każdym badaniu. Łatwo możemy na przykład wyobrazić sobie polską rodzinę, której niemal wszyscy członkowie są wysocy, utalentowani muzycznie i mają predyspozycje genetyczne do rozwoju choroby nowotworowej. Ogra-niczając badania do tej jednej rodziny, wyciągnęlibyśmy nieprawdziwe wnioski na temat Polaków w ogóle. Za-gadnienie reprezentatywności próby było najwyraźniej obce znacznej części badanych uczniów – właściwej od-powiedzi udzieliło zaledwie 49,9% z nich. Dokładniej-sza analiza wyników pokazuje jednak, że o ile ucznio-wie słabsi (tzn. uzyskujący niższy wynik w całym teście)
Autorzy:
BIOLOGIA – komentarz i pomysl na zadanie: Wojciech Grajkowski
CHEMIA – komentarz: Małgorzata Musialik, pomysł na zadanie: Marcin Chrzanowski
FIZYKA – komentarz: Maciej Wiśniewski, Krzysztof Horodecki, pomysl na zadanie: Maciej Wiśniewski
Marcin Chrzanowski, Wojciech Grajkowski, Krzysztof Horodecki, Małgorzata Musialik, Maciej Wiśniewski

Nowe zadania PPP | Marcin Chrzanowski, Wojciech Grajkowski, Krzysztof Horodecki, Małgorzata Musialik, Maciej Wiśniewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 70
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
wyraźnie częściej zaznaczali odpowiedź błędną, o tyle w grupie uczniów najlepszych odsetek prawidłowych odpowiedzi wynosił aż 84,4%.
W podpunkcie trzecim pytano uczniów o to, czy po-dawanie każdemu badanemu napoleonki jako pierwszej jest poprawne z metodologicznego punktu widzenia. Większość (64,8%) uczniów odpowiedziała „Tak”. Być może uznali oni, że zachowanie stałej kolejności ozna-cza lepszą kontrolę warunków eksperymentu. Prawid-łową odpowiedzią jest jednak „Nie”, ponieważ zjedzenie pierwszego ciastka może bardzo istotnie wpływać na ocenę smaku drugiego – po zjedzeniu napoleonki część uczestników mogła być już najedzona lub nawet mogło ich zemdlić. W dobrze zaplanowanym doświadczeniu połowa badanych powinna otrzymać jako pierwszą na-poleonkę, a druga połowa – kremówkę, aby oba ciastka miały „równe szanse”.
Czwarty podpunkt zadania odnosi się do zagadnie-nia charakterystycznego wyłącznie dla eksperymen-tów z udziałem ludzi, czyli do nastawienia badanego. Testując wpływ różnych czynników na kiełkowanie nasion czy rdzewienie żelaza, nie musimy przejmować się nastawieniem badanych obiektów. Ludzie jednak często ulegają sugestii, co może mieć istotny wpływ na uzyskane wyniki – dlatego chociażby w badaniach medycznych nie informuje się pacjentów, czy przyjmują prawdziwy lek, czy placebo. Siła sugestii jest szczególnie istotna w sytuacji, gdy nie mierzymy pewnych obiek-tywnie istniejących zmiennych, takich jak masa, pręd-kość czy liczba kiełkujących nasion, ale prosimy bada-nych o opisanie ich subiektywnych odczuć, dotyczących na przykład smaku ciastka. W opisanym doświadcze-niu świadomość, że napoleonka kosztuje więcej niż kre-mówka mogła sprawić, że jedno ciastko podświadomie było odbierane jako bardziej „wartościowe”, a zatem smaczniejsze. Właściwa odpowiedź dla tego podpunktu brzmi zatem „Tak”. Prawidłowo odpowiedziało 72,2%
uczniów, przy czym prawdopodobieństwo udzielenia właściwej odpowiedzi silnie korelowało z wynikiem ucznia w całym teście (od 53,2% wśród uczniów z grupy najsłabszej, aż po 95,2% w grupie najlepszej).
Całe zadanie prawidłowo rozwiązało zaledwie 21,1% uczniów, co pokazuje, jak skomplikowanym zagadnie-niem jest projektowanie doświadczeń z udziałem ludzi. Warto jednak zwrócić uwagę, że wyniki takich właś-nie badań są najwyraźniej obecne w mediach. Niemal codziennie spotykamy się z informacjami o „naukowo potwierdzonej” skuteczności jakiegoś leku, kosmetyku czy diety. Dobrze, aby uczeń umiał wówczas podejść do takich informacji krytycznie i miał świadomość, jak niewłaściwy dobór próby czy sposób przeprowadzenia badania może wpływać na jego wyniki.
Chemia – naturalny plastik
Zadanie
Aby wytworzyć „naturalny plastik” domowym sposobem, można użyć mleka i octu. Należy w tym celu do litra wrzą-cego mleka dodać 40 ml octu. Tę mieszaninę dalej ogrze-wa się, mieszając aż do momentu, kiedy pojawią się w niej białożółte bryłki, a sama zrobi się przejrzysta. Wtedy należy wyłączyć palnik, a po ostygnięciu przepłukać grudki pod bieżącą wodą. Otrzymaną masę można łatwo formować, a następnie należy odstawić na kilka dni, aż stwardnieje.
Tekst na podstawie: Skamieniałość z chomika. Zrób to sam!, Mick O’Hare, Insignis Media, Kraków 2011, s. 60–62
(1) Jaki proces spowodował pojawienie się białożół-tych grudek? A. Denaturacja. B. Zobojętnienie.(2) Opisany wyżej proces wywołały dwa czynniki.
Jednym z nich była zmiana pH, a drugim zmiana: A. temperatury. B. ciśnienia.
Komentarz
Denaturacja białka jest to nieodwracalny proces po-legający na zniszczeniu przestrzennej struktury białka pod wpływem czynników fizycznych bądź chemicz-nych. Czynniki te powodują rozerwanie wiązań che-micznych i oddziaływań międzycząsteczkowych stabi-lizujących strukturę łańcuchów polipeptydowych. Na skutek denaturacji, białka tracą swoje właściwości bio-logiczne, fizyczne i chemiczne. Zmiana właściwości fi-zykochemicznych jest przyczyną agregacji i wytrącania białek z roztworów. Do najważniejszych czynników fi-

Nowe zadania PPP | Marcin Chrzanowski, Wojciech Grajkowski, Krzysztof Horodecki, Małgorzata Musialik, Maciej Wiśniewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 71
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
zycznych prowadzących do denaturacji można zaliczyć: ogrzewanie, silne mieszanie, naświetlanie promienio-waniem ultrafioletowym, rentgenowskim i jonizującym lub działanie ultradźwiękami. Denaturację mogą wy-wołać także czynniki chemiczne, m.in. kwasy, zasady, sole metali ciężkich, enzymy, alkohol i mocznik.
Zadanie to sprawdza umiejętność prostą – uczeń wnioskuje na podstawie własnych wiadomości i analizy tekstu jakie zjawisko zostało wykorzystane do wytwo-rzenia naturalnego plastiku. Aby rozwiązać prawidło-wo to zadanie uczeń powinien wiedzieć, że w mleku znajduje się białko, dokładnie przeanalizować tekst wstępny oraz rozumieć jak przebiega proces denaturacji i jakie czynniki go wywołują. Uczeń powinien się też orientować, że podczas denaturacji można zaobserwo-wać procesy agregacji i wytrącania, związane ze zmianą właściwości chemicznych białek, a więc niejako „zmia-nę stanu skupienia” białek.
Zadanie zbadano na grupie około1900 uczniów w trakcie badania „Laboratorium myślenia” prowad-zonego przez Pracownię Przedmiotów Przyrodniczych. Z grupą 15 wybranych uczniów przeprowadzono także wywiady (cognitive laby), w trakcie których poproszono o ocenę tego zadania.
Pierwsze pytanie okazało się dla uczniów bardzo trudne – 53% z nich udzieliło poprawnej odpowiedzi (prawdopodobieństwo przypadkowego udzielenia pra-widłowej odpowiedzi wynosiło 50%). Większość ucz-niów udzielała odpowiedzi w sposób przypadkowy, a tylko wśród uczniów o najwyższych umiejętnościach odsetek odpowiedzi poprawnych był wyższy. Wyniki wskazują, że badana grupa uczniów nie znała bądź nie pamiętała terminu „denaturacja”. Co więcej, uczniowie, którzy wybrali prawidłową odpowiedź, często wybierali ją drogą eliminacji, wiedząc że opisany w zadaniu pro-ces nie był reakcją zobojętniania.
Dużo lepiej wypadły wyniki dla pytania drugiego – poradziło sobie z nim 84% uczniów. Uczniowie, któ-rzy wybrali błędną odpowiedź, nie przeczytali uważnie treści zadania (mowa jest w nim wyłącznie o wysokich temperaturach) bądź nie zauważyli, że w opisanym do-świadczeniu ciśnienie nie ulega zmianie. W sumie całe zadanie prawidłowo rozwiązało 42% badanych uczniów.
Wyniki, które osiągnęli uczniowie w prezentowa-nym zadaniu, skłaniają do refleksji nad posługiwaniem się terminologią chemiczną w zadaniach. Analiza sta-tystyczna wyników wykazała bowiem, że dla uczniów biorących udział w badaniu podstawowym problemem w zadaniu wcale nie było wskazanie przyczyn zacho-dzącego procesu, lecz jego nazwanie. Płynie z tego wniosek, że uczniowie mogą mieć problemy z podsta-wową terminologią chemiczną, która jest rozbudowana, a słowa są często nieznane z życia codziennego, niezro-zumiałe i podobnie brzmiące (np. dysocjacja, dekan-tacja, destylacja, denaturacja, itd.). Nawet jeśli ucznio-wie prawidłowo rozumują i potrafią rozpoznać proces, np. denaturacji, to często zwyczajnie nie pamiętają ter-minu chemicznego, który go opisuje. Należy więc mieć zawsze świadomość tego, co się w danym zadaniu chce zmierzyć – znajomość definicji czy umiejętność wnio-skowania. Warto też zwrócić uwagę na fakt, że problemy uczniów z opanowaniem specjalistycznej terminologii mogą pojawiać się na każdym etapie kształcenia (także w szkole ponadgimnazjalnej) i dotyczyć nie tylko che-mii, ale także innych przedmiotów szkolnych.
Z wywiadów przeprowadzonych z uczniami wyni-ka, że pomimo stosunkowo długiego wstępu uczniowie czytali go wnikliwie, a większości z nich zarówno sam kontekst zadania, jak i tekst wstępny bardzo się podoba-ły. Uczniowie deklarowali, że chcieliby przeprowadzić takie doświadczenie samodzielnie w domu (starali się więc jak najdokładniej zapamiętać przepis na plastik), uważali także, że zadanie uczy czegoś ciekawego. Zada-
nie to pokazuje, że o białkach naprawdę można uczyć w sposób nieszablonowy i interesujący dla uczniów.
Zadanie jest zgodne z następującymi wymaganiami zawartymi w podstawie programowej dla III etapu edu-kacyjnego:
Cele kształcenia – wymagania ogólne:I. Pozyskiwanie, przetwarzanie i tworzenie infor-
macji.1.1. Uczeń pozyskuje i przetwarza informacje z róż-
norodnych źródeł z wykorzystaniem technologii informacyjno-komunikacyjnych.
II. Rozumowanie i zastosowanie nabytej wiedzy do rozwiązywania problemów.
2.1. Uczeń opisuje właściwości substancji i wyjaśnia przebieg prostych procesów chemicznych.
Treści nauczania – wymagania szczegółowe:9.5. Uczeń (...) opisuje właściwości kwasu octowego
(...).9.13. Uczeń bada zachowanie się białka pod wpły-
wem ogrzewania, (...) kwasów (...); opisuje róż-nice w przebiegu denaturacji i koagulacji białek; wylicza czynniki, które wywołują te procesy; wykrywa obecność białka w różnych produk-tach spożywczych.
Zadanie to można wykorzystać także przy realizacji treści nauczania obecnych na dalszym etapie kształce-nia, gdyż jest ono zgodne z następującymi podpunkta-mi z podstawy programowej dla przedmiotu chemia na IV etapie edukacyjnym (poziom podstawowy):
3.3. Uczeń wyszukuje informacje na temat skład-ników napojów dnia codziennego (mleko) w aspekcie ich działania na organizm ludzki (…).

Nowe zadania PPP | Marcin Chrzanowski, Wojciech Grajkowski, Krzysztof Horodecki, Małgorzata Musialik, Maciej Wiśniewski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 72
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
dla uczniów zupełną zagadkę, a udzielana odpowiedź była praktycznie przypadkowa.
Wykres nr 1
Wykres nr 2
Wydawałoby się, że skoro uczeń ma świadomość, że większa moc czajnika oznacza szybsze gotowanie, to ma również przynajmniej intuicyjnie poprawne wyczucie pojęcia mocy.
Przedłużając rozumowanie z pierwszego wiersza uczeń powinien bez wysiłku stwierdzić, że czajnik o mniejszej mocy będzie musiał dłużej pracować, aby zagotować litr wody. W tym kontekście uznanie za po-prawną odpowiedź, że koszt takiego dłuższego gotowa-nia może być mniejszy jest zaskakujące.
Niezależnie od mocy czajnika energia potrzebna do zagotowania litra wody jest taka sama. Nawet jeśli uwzględnić straty energii, to niewątpliwie będą one większe przy powolnym grzaniu niż przy szybkim, co tym bardziej powinno prowadzić do odrzucenia hipo-tezy o zmniejszeniu kosztów poprzez gotowanie wody w czajniku o mniejszej mocy.
Jak widać stereotyp prostego kojarzenia: mała moc – małe zużycie energii jest silniejszy niż podstawy szkol-nej wiedzy uzyskanej na lekcjach fizyki.
Zadanie diagnozuje poziom umiejętności opisanych wymaganiami ogólnymi i szczegółowymi podstawy programowej fizyki dla III etapu edukacyjnego:
Cele kształcenia:III. Wskazywanie w otaczającej rzeczywistości
przykładów zjawisk opisywanych za pomocą poznanych praw i zależności fizycznych.
Treści nauczania:4.10. Uczeń posługuje się pojęciem pracy i mocy prą-
du elektrycznego.8.2. Uczeń wyodrębnia zjawisko z kontekstu, wska-
zuje czynniki istotne i nieistotne dla wyniku do-świadczenia.
Fizyka – jaki kupić czajnik?W badaniu Laboratorium Myślenia, realizowanym
przez PPP IBE staramy się wykorzystywać możliwie szeroko zadania, dotyczące zjawisk zachodzących w bli-skim dla ucznia świecie. Jedno z takich, odtajnionych i przeznaczonych do publikacji zadań prezentujemy niżej. Dotyczy ono istotnych z punktu widzenia użyt-kownika rozmaitych urządzeń, m.in. AGD, a także fun-damentalnych w fizyce pojęć mocy i energii.
Udzielone przez badanych uczniów odpowiedzi wskazują, że edukacja szkolna w pewnych aspektach nadal jednak przegrywa ze stereotypami lub błędną in-tuicją.
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Komentarz
Pierwszy wiersz okazał się banalnie łatwy. Nawet w najsłabszej grupie uczniów poprawna odpowiedź zde-cydowanie dominowała. Widać to na wykresie nr 1. wy-kres nr 2 pokazuje natomiast, iż drugi wiersz stanowił

Opinie | Ryszard Kowalski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 73
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
OpinieCzy warto studiować biologię? Strata czasu, a może dobra inwestycja na przyszłość?Ryszard Kowalski
Jesteśmy społeczeństwem akcyjnym i pewnie dla-tego lubimy obchodzić różnorodne jubileusze. W tym tekście, łamiąc zasadę okrągłych rocznic, chciałbym przywołać pamięć o Komisji Edukacji Narodowej. Szczególny powód do przypomnienia tego pierwszego w Polsce, a może nawet w Europie, świeckiego minister-stwa oświaty, przypadnie za lat dziesięć. Może wtedy, z okazji 250-lecia od chwili jej ustanowienia, polski parlament uchwali rok KEN, a jeśli się tak nie stanie, to z pewnością przynajmniej 14 października 2023 r. bę-dzie się o tym dużo mówiło. Które z ówczesnych osiąg-nięć KEN będą wtedy nagłośnione – tego nie sposób dziś przewidzieć. Oby nie trzeba było formułować po-stulatów o przywrócenie w szkołach nauczania ojczy-stej historii i przedmiotów przyrodniczych, gdyż jak na razie trwa ich konsekwentna redukcja.
Przyznaję, że świadomie popełniam falstart i przy-wołuję czasy KEN, choć nie ma do tego specjalnej oka-zji. Czynię to, gdyż chcę przypomnieć, że wprowadziła ona do szkół ojczysty język, nauczanie historii Polski i historii naturalnej – inaczej mówiąc przyrodoznaw-stwa, i to w bardzo trudnym okresie zniewolenia naszej ojczyzny. Cenimy ją za to, że zreformowała szkolnictwo
średnie i wyższe i zajęła się kształceniem świeckiej ka-dry nauczycielskiej, gdyż takiej w tamtych czasach bra-kowało. Poprzez wprowadzenie do szkół wychowania patriotycznego przyczyniła się do kształtowania świa-domości obywatelskiej i narodowej polskiej młodzieży. To, co w tamtych czasach intelektualne elity uznawały za wartość – można dodać: ponadczasową – dziś nieste-ty traci na znaczeniu. Dlatego też należy włączyć ostrze-gawczy alarm, informując społeczeństwo o tym, że źle dzieje się w naszej oświacie.
Trudno dopatrzyć się w działaniach współczesnych władz oświatowych rozwijania szkół zgodnie z duchem, ideą i wartościami określonymi przez Komisję. Udo-wodniły to ubiegłoroczne protesty, które przetoczyły się przez nasz kraj z postulatami przywrócenia należnej rangi nauczaniu historii. W obronie ograniczanej wciąż biologii, stanowiącej już drobny margines w ogólnym wykształceniu młodzieży na poziomie średnim, na ra-zie nie protestowano, ale warto się o ten przedmiot upo-mnieć. Bios znaczy wszakże życie! Przydzielenie jednej godziny tygodniowo w pierwszej klasie szkoły ponad-gimnazjalnej, na ten podstawowy w wykształceniu ogólnym przedmiot, jest niepokojące, a jednocześnie wskazuje, jak niewiele dla współczesnych oświatowych decydentów znaczy nauka o życiu. Nie zastąpi tego przedmiotu zlepek różnych przypadkowych zagadnień opisanych w nowej podstawie programowej kształce-nia ogólnego pod nazwą „przyroda”. Wejdzie ona do liceów dopiero w następnym roku szkolnym i będzie obowiązywała wszystkich uczniów, którzy nie wybiorą biologii w zakresie rozszerzonym, ale już dziś podzieliła ona i skonfliktowała nauczycieli biologii, chemii, fizyki i geografii. Powodem jest konkurencja o godziny liczone do pensum dydaktycznego, gwarantującego przetrwa-nie w nauczycielskiej profesji. Skoro ucząc tylko biolo-gii nauczyciel nie uzbiera tyle godzin, aby wystarczyło na etat, to musi rywalizować z pozostałymi o lekcje
przyrody. Sytuacja jest niepokojąca, gdyż licea, w swo-jej nazwie i naturze ogólnokształcące, nie zapewniają ogólnego wykształcenia na poziomie średnim w zakre-sie podstawowych przedmiotów. Ewoluują w wąskie specjalności już od pierwszej klasy, co należy uznać za zbyt wczesne. Zaniedbują kształcenie w zakresie histo-rii, uważanej za podstawę kształcenia patriotycznego, ale i biologii, która jako nauka o życiu i jego różnorod-ności ma także związek z patriotyzmem i obywatelskim wychowaniem, chociażby poprzez zawarte w niej tre-ści ekologiczne i sozologiczne. Poprzez minimalizację godzinowego wymiaru wiele ważnych przedmiotów zaliczanych tradycyjnie do przyrodoznawstwa straciło na znaczeniu i znalazło się na marginesie współczesnej edukacji. Jakże trudno w tych warunkach wziąć odpo-wiedzialność za efekty nauczania i wychowania młode-go pokolenia, w tym kształtowanie prośrodowiskowych postaw. Niewielki przydział godzin przeznaczonych na przedmiot, przegęszczone klasy pomimo demograficz-nego niżu (bo tak jest taniej), ubóstwo w wyposażeniu pracowni przedmiotowych, jako rezultat niedoinwesto-wania szkół, niby państwowych, ale jednak oddanych pod opiekę samorządom gminnym i powiatowym – to wybrane czynniki degradujące polską oświatę. Efekt tego jest taki, że szkoły ponadgimnazjalne opuszczają absolwenci, którzy nie są przygotowani do świadome-go i odpowiedzialnego wyboru kierunku studiów. Po-dejmując życiową decyzję o tym, co studiować, młodzi ludzie patrzą przede wszystkim na marketingowe opa-kowanie treści w nośny w swoim wydźwięku slogan. Trudne czasy przeżywają kierunki o tradycyjnych na-zwach, np. biologia, chemia, geografia, fizyka. I nie jest to problem lokalny, lecz ogólnopolski. Niewielka grupa młodzieży chce je studiować. Większość zapisuje się na różne „biopochodne” i „biozmanipulowane” kierunki, co skrzętnie wykorzystują uczelnie, niemające już dziś obowiązku przestrzegania zatwierdzonej przez mini-
dr Ryszard Kowalski: zastępca dyrektora Instytutu Biolo-gii Uniwersytetu Przyrodniczo-Humanistyczny w Siedlcach

Opinie | Ryszard Kowalski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 74
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
sterstwo centralnej listy kierunków, kreujące w swej au-tonomii bogatą ofertę kształcenia obejmującą zarówno kierunki, jak i specjalności z przedrostkiem „bio-”.
Mając wieloletnie doświadczenie w kształceniu stu-dentów biologii, także w zakresie ich nauczycielskiego przygotowania, zastanawiam się nad współczesnym kryzysem oświaty i szkolnictwa wyższego, sztucznie rozdzielonych przez polityków na dwa ministerialne resorty. Pytanie „czy warto dziś studiować biologię?” często zadaję sam sobie. Wpisałem je także z ciekawo-ści do internetowej przeglądarki, wszechwiedzącego instrumentu, z którego szczególnie chętnie korzysta współczesna młodzież. W ułamku sekundy uzyskałem dostęp do wielu stron, które zawierały w tytule dokład-nie takie samo pytanie. Część z nich to fora interneto-we, na których młodzi ludzie dyskutują, zadają pytania, spierają się, podpowiadają, wymieniają się poglądami o warunkach nauki, programach i treściach nauczania, uczelniach, wykładowcach.
Czy warto zatem studiować biologię? Wątpiący w sens podejmowania takich studiów najczęściej po-dawali argument braku pracy dla biologów z wyższym wykształceniem. Trudny, wymagający wiele nauki kie-runek – a przecież studiowanie nie musi przypominać codziennego zajęcia dzięcioła. Wyższe wykształcenie można zdobyć niewielkim nakładem własnej pracy, studiować łatwo, korzystać z przyjemności, a ponieważ ukończenie studiów, niezależnie od kierunku i tak nie gwarantuje otrzymania pracy, to po co się przemęczać.
Czy warto zatem studiować biologię? Oddajmy głos internautom przytaczając parę zróżnicowanych opinii.
Jest to kierunek ciekawy, lecz na pewno bardzo skompliko-wany. W gimnazjum może wydawać się co prawda prosty, ale po wejściu w trudniejsze tematy, raczej taki nie jest. Jednak jeżeli nie masz z tym przedmiotem problemu, to tak, oczywiście, że warto. Można po nim znaleźć pracę dużo łatwiej niż po studiach humanistycznych.
Studiuję 3 rok biologii. Roboty jest full ciągle. W zasadzie, żeby do czegoś dojść trzeba się uczyć cały czas. Pierwsze 1,5 roku to kompletna strata czasu i energii. Same bzdury jak dla mnie – to samo, co w LO, tylko że trochę poszerzone. Ro-śliny, pierwotniaki – to była dla mnie masakra. Dopiero od tego roku jestem na specjalizacji, molekularnej i przedmio-ty są ciekawe, choć musimy teraz opanować to wszystko, co miała biotechnologia przez trzy lata w jeden rok. I znowu harówa.
Jeżeli chcesz się zająć człowiekiem, idź na medycynę. Jeśli biologią molekularną, idź na biotechnologię, a jeśli ekologia i te wszystkie dłubania w glebie, to na biologię.
W UE jest bardzo duży nacisk na środowisko. W zasadzie każda większa inwestycja musi przejść przez urząd zajmują-cy się środowiskiem. Myślę, że przez najbliższe lata ta dział-ka trochę się rozbuduje ze względu na wymagania i praca się znajdzie.
Zaczerpnięte z Internetu posty zostały w niewiel-kim stopniu zmienione, tak aby nie zdradzać nazw uczelni i poprawić interpunkcję, która w internetowych wypowiedziach nie odgrywa żadnej roli.
Jeszcze raz zatem warto powtórzyć pytanie – czy warto obecnie studiować biologię? Odpowiedzi po-winien udzielić sobie każdy młody człowiek kończący kształcenie na poziomie średnim i szukający kierunku związanego z zainteresowaniami, marzeniami, przyszłą pracą, gdyż każdy indywidualnie jest odpowiedzialny za swoje życie i swój zawodowy rozwój. Jako biolog, podsumowujący już powolutku swoje zawodowe życie, udzielam odpowiedzi twierdzącej i mam na to oczywi-ście uzasadnienie.
Stara giełdowa zasada, którą w jakimś sensie moż-na odnieść do szkolnictwa wyższego i biologii jako kierunku studiów, poucza, aby akcje kupować na doł-ku, a sprzedawać na górce, a wszystko po to, żeby jak najwięcej zarobić. Wybór kierunku studiów to także inwestycja i to najczęściej jedna w ciągu całego życia.
Zjawiska w przyrodzie, a także w społeczeństwie podle-gają ciągłym i dynamicznym zmianom. Dla przykładu, liczebność osobników należących do określonej popu-lacji przedstawiona na osi upływającego czasu układa się zazwyczaj w dość regularną sinusoidę, raz osiągając minimum, aby znów po jakimś czasie osiągnąć wartość maksymalną. Jeśli dziś biologów z wyższym wykształ-ceniem wydaje się być ponad potrzeby i nie mogą zna-leźć pracy, to nie oznacza, że jest to tendencja trwała i że absolwenci innych kierunków nie mają problemów z zatrudnieniem. Za parę lat sytuacja może się zmie-nić i biolog będzie pilnie poszukiwany. Zdaje się, że dobrze rozumie ten mechanizm jeden z internautów, pisząc w przytoczonym powyżej poście o opiniowaniu przez specjalistów przyrodników różnych projektów inwestycyjnych w Unii Europejskiej. Tę pracę powinni wykonywać biolodzy z przygotowaniem do pracy w te-renie. Trzy lata temu, na międzynarodowej konferencji w Brodnicy, zorganizowanej pod hasłem „Zielone po-nad granicami”, prezentowany był referat o wdrażaniu Natury 2000 – unijnego programu ochrony przyrod-niczego środowiska, w jednej z holenderskich gmin. Uczestnicy konferencji z Polski byli zaskoczeni tym, że w holenderskim odpowiedniku naszego samorzą-du gminnego ochroną środowiska zajmuje się 10 osób, a bywa że i więcej. U nas w przeciętnej gminie, nawet tej funkcjonującej na obszarach Natura 2000, środowi-skiem zajmuje się najczęściej jeden urzędnik, któremu jeszcze na dodatek zleca się inne zadania, np. z zakre-su budownictwa lub utrzymania dróg. Ale to się musi zmienić, gdyż w zjednoczonej Europie problemy śro-dowiskowe będą wciąż zyskiwały na znaczeniu. Biolog z wyższym wykształceniem o specjalności środowisko-wej, z umiejętnościami w zakresie waloryzacji przyrody czy zarządzania środowiskiem powinien zatem znaleźć zatrudnienie. Takich specjalistów potrzeba będzie coraz więcej w obsłudze gospodarstw rolnych, starających się

Opinie | Ryszard Kowalski | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 75
SZKO
ŁAN
AUKA
KRÓ
TKO
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
o wdrażanie programów rolno-środowiskowych, gdyż przy niektórych pakietach istnieje potrzeba wykona-nia przyrodniczej waloryzacji gospodarstwa. To ważny i rozwojowy kierunek wspólnotowego rolnictwa. Miejsc dla absolwentów uczelni z biologicznym wykształce-niem powinno przybywać zatem w gminach, w rolni-ctwie i jego otoczeniu, w różnego typu laboratoriach i tych medycznych, gdyż starzejące się społeczeństwo będzie potrzebowało intensywniejszej opieki medycz-nej, i tych środowiskowych, świadczących usługi dla służb ochrony środowiska. Należy wierzyć, że zielone światło dla biologów starających się o pracę zaświeci się także w oświacie, gdy władze przekonają się o po-pełnieniu błędu polegającego na ograniczeniu kształce-nia biologicznego i – ogólnie mówiąc – przyrodniczego w szkołach. Teraz jest właśnie dobry czas na podejmo-wanie decyzji o studiowaniu biologii.
Na pytanie, w której uczelni podjąć studia, nie chciałbym się bezpośrednio wypowiadać, aby nie być posądzonym o autoreklamę. Kierunek biologia jest prowadzony w wielu uczelniach, a w jego ramach ist-nieją rozbudowane oferty specjalnościowe, aby każdy student mógł zaspokoić potrzeby wynikające z indy-widualnych zainteresowań i życiowych planów. Moż-na także studiować w innych krajach, gdyż granice są dziś otwarte i istnieją specjalne programy stypendialne. W Uniwersytecie, w którym pracuję, studenci biologii mogą kształcić się w specjalności analitycznej, środowi-skowej, eksperymentalnej z biotechnologią, zarządza-nia środowiskiem, a także biologii w połączeniu z kul-turą fizyczną. Studiując każdą z wyżej wymienionych specjalności można bezpłatnie uzyskać dodatkowe uprawnienia do wykonywania zawodu nauczyciela, co umożliwia podjęcie pracy w szkole, lecz także w poza-szkolnych instytucjach zajmujących się edukacją przy-rodniczą, np. w domach kultury, opieki społecznej czy w nadleśnictwach.
Wybierając uczelnię, warto sprawdzić jakie ma ona tradycje kształcenia na tym kierunku. Trzeba unikać „edukacyjnych jętek”, które pojawiają się znienacka w uczelniach prywatnych i państwowych, są niestabilne kadrowo i nie gwarantują odpowiedniej jakości kształ-cenia. Sympatycznie studiuje się biologię w mniejszych ośrodkach akademickich zlokalizowanych poza duży-mi aglomeracjami miejskimi, gdyż z reguły organizuje się tam atrakcyjne z punktu widzenia przyszłej pracy zajęcia terenowe. Ponadto warto sprawdzić, czy poza zajęciami obowiązkowymi istnieje rozbudowana oferta przedmiotów do wyboru, czy w uczelni można rozwijać indywidualne zainteresowania w studenckich kołach naukowych. Warto zapytać też o możliwości wyjazdu za granicę w ramach wymiany studentów, o obowiąz-kowe praktyki, o kadrę nauczycieli akademickich, aby potem nie wyrażać swojego rozczarowania tak, jak to zrobiła w Internecie jedna z wypowiadających się osób: „Poza tym powiem, że profesorowie mają nas wszyst-kich gdzieś, chcą nam tylko dokopać przy każdej okazji i udowodnić że nic nie umiemy”. Cieszę się, że ten przy-padek nie dotyczy mojego Uniwersytetu. Młodzieży z gorzej sytuowanych rodzin radzę, aby zapoznała się wcześniej z systemem pomocy materialnej dla studen-tów. I jeszcze jedna ważna rada dla kandydatów na stu-dia: „Nie szukaj daleko tego, co masz w zasięgu ręki”.
Czy warto studiować biologię – naukę o życiu, stanowiącą podstawę wielu gospodarczych dziedzin i współczesnej ochrony przyrodniczego środowiska? Z pełnym przekonaniem, że jest to inwestycja z przy-szłością rekomenduję ten kierunek ambitnym matu-rzystom.

Recenzja książki | Weronika Wronowska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 76
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Recenzja książkiBiologia Neila A. Campbella, Jane B. Reece i wsp.Weronika Wronowska
1. Ciężka lektura?
Kiedy zobaczyłam Biologię Campbella na półce księgarni, przestraszyłam się – 5 kg papieru, 5 kg in-formacji. Nie jest to książka, którą spakujesz do bagażu podręcznego, aby mieć co czytać w podróży. To dzieło „stacjonarne” – na półkę, noszenie go w torbie gwaran-tuje skoliozę.
2. Po co książka, kiedy wszystko jest w Wikipedii?
Campbell z pewnością nie jest tak poręczny jak tab-let, ale w przeciwieństwie do Wikipedii wskazuje zagad-nienia najistotniejsze dla zrozumienia podstaw biolo-gii, określa ramy. Jeśli jakieś zagadnienie zainteresuje cię szczególnie, znajdziesz więcej na ten temat w sieci. Dzięki podręcznikowi nie ominiesz jednak tematów, które mogłyby umknąć podczas samodzielnego eksplo-rowania Internetu.
3. Książki nie nadążają za postępami nauki
Podręcznik przestaje być aktualny zanim jeszcze wydawca zdąży go wydrukować. Takie czasy, chociaż spośród dostępnych ten jest jednym z najbardziej aktu-alnych1. Obszernie omówiono w nim główne techniki
1 Ósme wydanie jest nowością na polskim rynku, tymczasem wydawnictwo Pearson już w 2011 roku wydało kolejną, dziewiątą edycję podręcznika.
stosowane we współczesnej biotechnologii, przeanali-zowano zjawisko transpozycji, przedstawiono również podstawowe fakty związane z epigenetycznymi mecha-nizmami dziedziczenia.
4. Tylko, czy to aby książka dla Ciebie?
O tym musisz przekonać się sam. Polecałabym ją przede wszystkim: • licealistom – chociaż książka znacznie wykracza
ponad podstawę programową realizowaną w pol-skich liceach, to wszystkie treści przedstawione są w przystępny sposób; każde nowe pojęcie jest na bieżąco wyjaśniane (wytłuszczenie kluczowych terminów pomaga w ich zapamiętywaniu);
• olimpijczykom – książka ta jest jedynym podręcz-nikiem polecanym przez Komitet Główny Olim-piady Biologicznej;
• nauczycielom – skoro polecam uczniom i studen-tom, trudno nie polecić nauczycielom. Większość z nich nie będzie miała żadnych wątpliwości co do przydatności tego podręcznika w nauczaniu biolo-gii. To źródło doskonałych ilustracji do lekcji, teks-tów i ciekawostek uzupełniających zajęcia;
• studentom – nie nauczysz się z tej książki do eg-zaminu z immunologii, genetyki czy botaniki, ale jeśli po jakimś czasie nie będzie ci już tak bardzo zależeć na detalach, ten podręcznik pozwoli ci od-zyskać ogólne pojęcie o temacie.
• na koniec – wszystkim tym, którzy zajmują się po-pularyzacją nauki, tak aby widzieli, co populary-zują.
BiologiaNeil A. Campbell, Jane B. Reece i wsp.Dom Wydawniczy Rebis, Poznań 2012, ss. 1416
LEKKA RECENZJA
CIĘŻKIEJ KSIĄŻKI
spis treści i szczegółowe informacje na: biologiacampbella.pl

Recenzja książki | Weronika Wronowska | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 77
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
5. Treść czy forma?
Szczęśliwie: jedno i drugie. Pod względem edytor-skim – świetna. Solidnie zszyte 1416 stron nie powin-no wypadać nawet po wielu latach używania. Zdjęcia i schematy doskonale ilustrują prezentowane treści. Nie jest to jednak kolorowy album ze zdawkowymi opisa-mi. Książka przede wszystkim zawiera obszerny, bogaty merytorycznie tekst.
6. Campbell kontra Villee
W szkolnej bibliotece obowiązkowo powinny się znaleźć oba dzieła. Biologia Campbella zawiera nieco więcej informacji szczegółowych. Dokładniej omówio-ne zostały: wybrane przemiany metaboliczne, szlaki sygnalizacji komórkowej czy regulacja ekspresji genów. Zarazem ogromnym walorem Biologii Solomon, Berg i Martin (wciąż często zwanej „Villee”) jest „odwieczna” obecność tej książki na rynku.
7. Szukanie dziury w całym
Książka jest naprawdę świetna. Jeśli jednak koniecz-nie będziemy chcieli znaleźć jakieś minusy, oto moje propozycje. • Organizacja książki – podział materiału na roz-
działy dla wielu czytelników wychowanych na polskich podręcznikach może być zaskakujący. Przyzwyczailiśmy się wrzucać mitozę i mejozę do jednego worka. Tu mitozę znajdziemy w rozdziale omawiającym cykl komórkowy, zaś mejoza opisa-na jest w całkiem innym dziale – genetyka, jako przyczyna zmienności genetycznej. Również w ge-netyce trzeba szukać rozdziału o wirusach. Jeżeli zaś chcemy przestudiować anatomię człowieka,
musimy ją wyłuskać z części o budowie i funkcjach zwierząt.
• Zgodność z wymaganiami programowymi dla li-ceum – w Biologii Campbella brakuje opisu cho-rób. Profilaktyka, drogi infekcji czy metody lecze-nia są omawiane raczej zdawkowo. Nie za wiele dowiemy się też o ukochanych przez polską eduka-cję cyklach rozwojowych pasożytów.
• Nasz system klasyfikacyjny a reszta świata – sy-stem klasyfikacji organizmów w Biologii Campbel-la jest bezpośrednio tłumaczony z języka angiel-skiego. Znajdujemy tu więc podział na jednakowe taksony, czy to dla roślin, czy zwierząt (królestwo, typ, gromada, rząd, rodzina, rodzaj, gatunek). Nie-stety nasz rodzimy system klasyfikacyjny przez wiele lat rządził się nieco odmiennymi prawidłami (np. gromada u roślin odpowiada typowi u zwie-rząt). Warto o tym wiedzieć czytając rozdział „Fi-logeneza i drzewo życia”.
• Przypisy – autorzy nie podają materiałów źródło-wych, zapewne z powodu braku miejsca. Brakuje też wskazówek dla czytelników chcących pogłębić wiedze w danym temacie oraz odniesień do litera-tury szczegółowo omawiającej wybrane zagadnie-nie.
8. Bilans końcowy
Znakomity podręcznik – esencja wiedzy biologicz-nej, dobrze napisany, ładnie wydany i jeśli w ogóle my-ślisz o kupieniu książki całościowo ujmującej biologię, „Campbell” jest doskonałym wyborem.
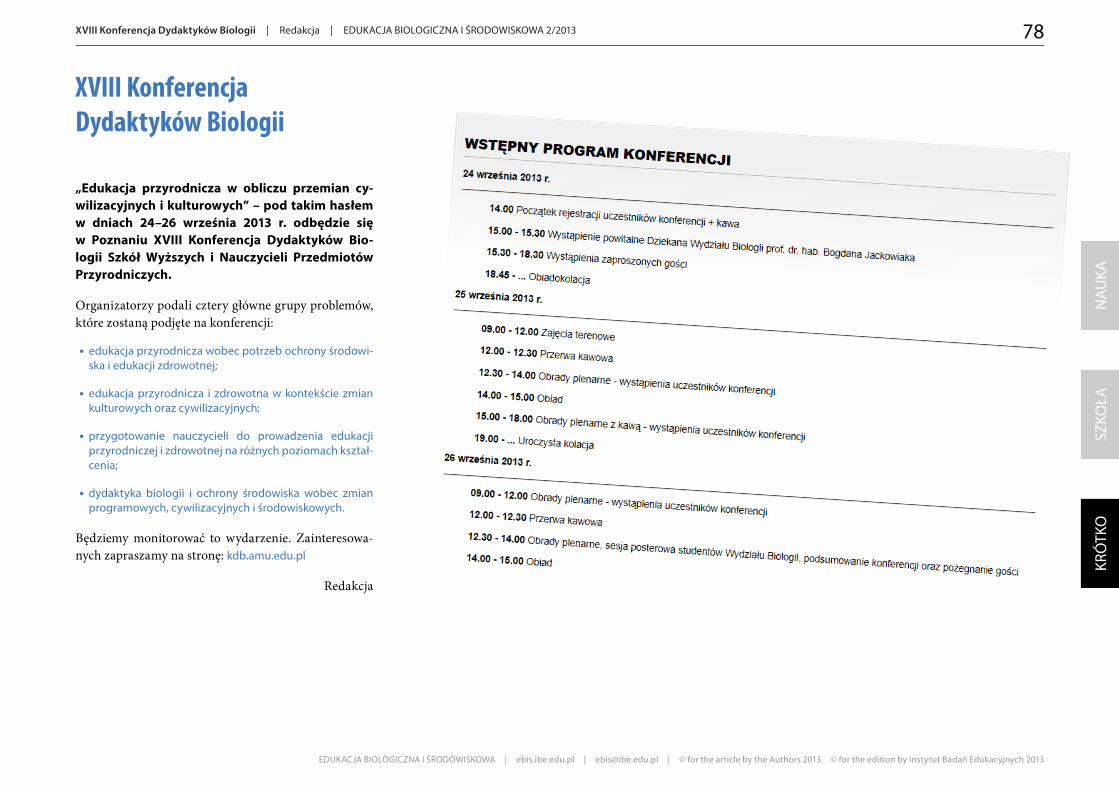
XVIII Konferencja Dydaktyków Biologii | Redakcja | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 78
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
XVIII Konferencja Dydaktyków Biologii
„Edukacja przyrodnicza w obliczu przemian cy-wilizacyjnych i kulturowych” – pod takim hasłem w dniach 24–26 września 2013 r. odbędzie się w Poznaniu XVIII Konferencja Dydaktyków Bio-logii Szkół Wyższych i Nauczycieli Przedmiotów Przyrodniczych.
Organizatorzy podali cztery główne grupy problemów, które zostaną podjęte na konferencji:
• edukacja przyrodnicza wobec potrzeb ochrony środowi-ska i edukacji zdrowotnej;
• edukacja przyrodnicza i zdrowotna w kontekście zmian kulturowych oraz cywilizacyjnych;
• przygotowanie nauczycieli do prowadzenia edukacji przyrodniczej i zdrowotnej na różnych poziomach kształ-cenia;
• dydaktyka biologii i ochrony środowiska wobec zmian programowych, cywilizacyjnych i środowiskowych.
Będziemy monitorować to wydarzenie. Zainteresowa-nych zapraszamy na stronę: kdb.amu.edu.pl
Redakcja

Nowości ze świata nauki | Redakcja i Pracownia Przedmiotów Przyrodniczych IBE | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 79
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Nowości ze świata nauki
Dep
esze
prz
ygot
ował
y:
Reda
kcja
i Pr
acow
nia
Prze
dmio
tów
Pr
zyro
dnic
zych
IBE
Bioniczne oko – nadzieja dla chorych na retinopatię barwnikową
Amerykańska Agencja do Spraw Żywności i Leków (FDA) 14 lutego 2013 r. zaaprobowała system Ar-gus® II do stosowania jako urządzenie przywraca-jące wzrok osobom niewidzącym cierpiącym na zaawansowaną postać retinopatii barwnikowej (RP) [1].
RP jest chorobą dziedziczną, polegającą na odkładania się cząsteczek barwnika wzrokowego w postaci skupisk w komórkach receptorowych. Skutkiem tego może być osłabienie widzenia wieczorem, zaburzenie tzw. widze-nia obwodowego (czyli widzenia nie tylko „na wprost”), prowadzące do widzenia lunetowego, a w konsekwencji do całkowitej utraty wzroku. Uszkodzeń wynikających z rozwoju RP w siatkówce i nerwie wzrokowym nie da się skorygować typowymi okularami. Dotąd nie pozna-no skutecznych metod zatrzymania postępu choroby ani jej leczenia.
W zdrowym oku fotoreceptory w siatkówce przekształ-cają bodźce świetlne na impulsy elektrochemiczne, któ-re przesyłane są przez nerw wzrokowy do mózgu, gdzie następnie sygnały te ulegają dekodowaniu. Jeżeli dzia-
łanie fotoreceptorów nie jest poprawne ze względu na RP, cały proces ulega przerwaniu.
Siatkówkowy system protezowy Argus® II [2] został stworzony w celu elektrycznego stymulowania pozosta-łych, działających jeszcze prawidłowo, fotoreceptorów w siatkówce oka chorego na retinopatię barwnikową
W jaki sposób działa proteza? Składa się ona z dwóch części – implantu, który wszczepia się chirurgicznie do wnętrzna oraz na powierzchnię gałki ocznej oraz przenośnego modułu zewnętrznego. W skład implantu, pokazanego na rysunku [3] wchodzą: a) układ elektro-niczny, b) elektroda szeregowa, przekazująca impulsy do fotoreceptorów oraz c) antena.
Na przenośny moduł zewnętrzny, pokazany poniżej [4], składają się: okulary zaopatrzone w miniaturową ka-merę i antenę bezprzewodową, jednostka przetwarza-jąca obraz (VPU) oraz kabel, łączący ze sobą elementy modułu.
Proces tworzenia obrazu z udziałem implantu realizo-wany jest następująco: obraz z kamery video przesyła-ny jest do mikrokomputera (VPU), który przetwarza go, a następnie wysyła za pomocą kabla z powrotem do okularów. Sygnał w postaci odpowiednich, instrukcji jest następnie przesyłany bezprzewodowo (przez nadaj-nik) i odbierany przez antenę implantu we wnętrzu oka i kierowany do elektrody. Elektroda z kolei przesyła syg-

Nowości ze świata nauki | Redakcja i Pracownia Przedmiotów Przyrodniczych IBE | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 80
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
nał w postaci serii impulsów elektrycznych do pozosta-łych, zdrowych receptorów, które przesyłają informację o obrazie do mózgu.
Pacjenci, po wszczepieniu implantu, muszą na nowo nauczyć się interpretować obrazy, jako że widzenie z sy-stemem Argus® II bardzo różni się od widzenia, którego doświadczali zanim stracili wzrok. Po pewnym czasie od zabiegu są jednak w stanie czytać nagłówki z gazet, znajdować źródło światła, lokalizować i identyfikować obiekty czy orientować się w przestrzeni. Na portalu YouTube.com można obejrzeć krótki wywiad (w języku angielskim z napisami) [5] przeprowadzony z pacjen-tem używającym systemu Argus® II. Pacjent odzyskał wzrok po 25 latach i opowiada, w jaki sposób wszcze-piony implant odmienił jego życie.
Marcin M. Chrzanowski, PPP
Literatura
[1] URL: http://www.accessdata.fda.gov/cdrh_docs/pdf11/H110002a.pdf.
[2] URL: http://2-sight.eu/en/how-is-argus-r-ii-designed-to-produ-ce-sight-en.
[3] URL: http://2-sight.eu/images/stories/2-sight/Eye.png, zmodyfi-kowany.
[4] URL: http://2-sight.eu/images/stories/2-sight/external-compo-nents_ee.png, zmodyfikowany.
[5] URL: http://www.youtube.com/watch?feature=player_embedded&v=8AWIJdayKow.
Wielofunkcyjny ditlenek tytanu ratunkiem dla środowiska?
Grafen doczekał się konku-rencji w kategorii „najbardziej obiecująca nowa substancja”. Naukowcy z Nanyang Techno-logical University (NTU) w Sin-gapurze odkryli nanomateriał o tak wszechstronnych właści-wościach, że nazwali go wielo-funkcyjnym ditlenkiem tytanu (ang. Multi-use Titanium Dioxi-de). Wynalazek może być prze-łomem w takich kwestiach jak uzdatnianie wody i przekształ-canie ścieków w odnawialne źródła energii i wodoru (Liu i wsp., 2013).
Ditlenek tytanu jest substancją chemiczną powszech-nie występującą w przyrodzie i stosowaną od dawna w przemyśle jako pigment (biel tytanowa) do produkcji żywności, kosmetyków, leków, past do zębów, papieru, tworzyw sztucznych, porcelany, farb i emalii oraz jako filtr przeciw promieniowaniu UV i stabilizator kolo-ru szkliw. W latach 70. XX wieku odkryto, że ma on wspaniałe właściwości fotokatalityczne, tzn. zdolność do przyspieszania reakcji chemicznych pod wpływem światła. Ze względu na niską cenę, trwałość i wydajność ditlenek tytanu znalazł zastosowanie jako fotokataliza-tor do oczyszczania wody, ścieków i powietrza, a nawet jako środek dezynfekcyjny zdolny do zabicia bakterii Escherichia coli oraz inaktywacji wirusa grypy. Rozwój nanotechnologii w ostatnich latach sprawił, że zaczę-to wykorzystywać ditlenek tytanu do otrzymywania
nanomateriałów, które w odróżnieniu od „klasyczne-go” materiału posiadają wiele unikalnych właściwości i jeszcze lepsze zdolności katalityczne dzięki dużemu polu powierzchni nanostruktur w stosunku do zajmo-wanej przez nie objętości.
Profesorowi Darrenowi Sunowi i jego współpracowni-kom z NTU’s School of Civil and Environmental Engi-neering udało się otrzymać z kryształów ditlenku tytanu nanomateriał, którego lista zastosowań jest naprawdę długa. Dzięki silnym właściwościom fotokatalitycznym może być wykorzystany jako fotoanoda do rozkładu wody na wodór i tlen pod wpływem promieniowania UV, jako fotokatalizator do rozkładu związków orga-nicznych w wodzie oraz innych zanieczyszczeń śro-dowiska (NOx), a także do odzysku i usuwania metali ciężkich ze ścieków. Użyty jako fotokatalizator, w pro-
Prof. Darren Sun z próbką wielofunkcyjnego ditlenku tytanu zanim zostanie on przerobiony na membranę.
Oba zdjęcia pochodzą ze strony: http://www.gizmag.com/multi-use-titanium-dioxide/26756

Nowości ze świata nauki | Redakcja i Pracownia Przedmiotów Przyrodniczych IBE | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 81
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
cesie przetwarzania ścieków w wodór i czystą wodę pod wpływem światła, jest trzykrotnie wydajniejszy od trudno dostępnej i drogiej platyny. Materiał ten nadaje się doskonale do wytwarzania wydajnych i tanich bate-rii słonecznych, membran filtracyjnych pozwalających odsalać wodę za pomocą wysokoprzepływowej wymu-szonej osmozy (ang. high-flux forward osmosis), a także membran do separacji gazów. Może posłużyć do wy-twarzania powłok samoczyszczących, antystatycznych, hydrofilowych, chroniących przed promieniowaniem UV i neutralizujących zapachy. Z kolei dzięki właści-wościom przeciwbakteryjnym może być z powodze-niem wykorzystywany do produkcji filtrów antybakte-ryjnych, a nawet elastycznych bandaży zapobiegających infekcjom i przyspieszającym leczenie ran ze względu na dobrą przepuszczalność tlenu.
Swe unikalne właściwości wielofunkcyjny ditlenek tyta-nu zawdzięcza opatentowanej strukturze nanowłókien, które w zależności od późniejszego przeznaczenia mogą być wzbogacane węglem, miedzią, cynkiem lub cyną.
Membrana
filtracyjna
wykonana
z nanocząstek
ditlenku tytanu.
Obecnie zespół prof. Suna pracuje nad opracowaniem nanomateriału z czarnego ditlenku tytanu, który ma posłużyć do produkcji baterii litowo-jonowych. Wyniki wstępnych analiz wskazują, że użycie anody zrobionej ze sferycznych nanocząstek ditlenku tytanu modyfiko-wanych węglem może podwoić pojemność baterii. Wy-dłuża to znacząco żywotność takich baterii w porówna-niu z ich standardowymi odpowiednikami
Zdaniem prof. Suna, niskie koszty i łatwość wytwa-rzania nanomateriału z ditlenku tytanu dają mu nie-samowity potencjał w rozwiązaniu najbardziej naglą-cych problemów związanych z ochroną środowiska, a zwłaszcza z zanieczyszczeniem wód, kurczących się zasobów wody pitnej i paliw kopalnych oraz wciąż wzrastającym zapotrzebowaniem na czyste źródła ener-gii, takie jak wodór.
W ciągu pięciu lat badań nad wielofunkcyjnym dit-lenkiem tytanu zespół prof. Suna opublikował ponad 70 prac naukowych w czasopismach takich jak Sepa-ration and Purification Technology, Water Research, Energy and Environmental Science oraz Journal of Ma-terials Chemistry.
Małgorzata Musialik, PPP
Literatura
http://media.ntu.edu.sg/NewsReleases/Pages/newsdetail.aspx? news=14e3b618-c71c-4f20-935c-2a566af5a298.
Liu Z, Bai H, Sun DD (2013). A general method for the fabrication of hierarchically-nanostructured membranes with multifunctional environmental applications. Separation and Purification Technol-ogy 107:324-330. http://dx.doi.org/10.1016/j.seppur.2012.12.032.
Nakata K, Fujishima A (2012). TiO2 photocatalysis: Design and applications. Journal of Photochemistry and Photobiology C: Photochemistry Reviews. 13 (3):169-189. DOI: 10.1016/j.jphoto-chemrev.2012.06.001.
Nakano R i wsp. (2012). Photocatalytic inactivation of influenza virus by titanium dioxide thin film. Photochem. Photobiol. Sci. 11:1293-1298. DOI: 10.1039/C2PP05414K.
Liu L, Liu Z, Bai H, Sun DD (2012). Concurrent filtration and solar photocatalytic disinfection/degradation using high-performance Ag/TiO2 nanofiber membrane. Water Research. 46(4):1101-1112. http://dx.doi.org/10.1016/j.watres.2011.12.009.
Liu Z, Bai H, Lee J, Sun DD (2011). A low-energy forward osmosis process to produce drinking water. Energy and Environmental Science. 4(7):2582-2585. DOI: 10.1039/C1EE01186C.
Bai H, Liu Z, Sun DD (2012). A lithium-ion anode with micro-scale mixed hierarchical carbon coated single crystal TiO2 nanorod spheres and carbon spheres. Journal of Materials Chemistry. 22(47):24552-24557. DOI: 10.1039/C2JM34142E.

Nowości ze świata nauki | Redakcja i Pracownia Przedmiotów Przyrodniczych IBE | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 82
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Wyhodują całe narządy?
Japońscy naukowcy są coraz bliżej wyhodowania w warunkach laboratoryjnych całego narządu. Dzięki wykorzystaniu specjalnej osłony żelowej udało im się już wyprodukować dobrze funkcjo-nujące włókna komórkowe.
Proste tkanki człowieka – takie jak elementy skóry, mięśnia sercowego, czy rogówka – wyhodowane w la-boratorium, coraz częściej wykorzystywane są do prze-szczepów. Taki sposób postępowania ma duże zalety, ponieważ tkanki można wyhodować z komórek opero-wanego pacjenta, dzięki czemu można zminimalizować ryzyko odrzucenia przeszczepionej tkanki. Wciąż jed-nak mamy duży problem z ukształtowaniem narządu, który jest bardziej skomplikowany pod względem bu-dowy niż warstwa tkanki. Być może przełamanie tych trudności zbliża się dużymi krokami dzięki niedawne-mu odkryciu japońskich naukowców z Uniwersytetu Tokijskiego (Onoe i wsp., 2013).
Zespół prof. Shoji Takeuchi zauważył, że wiele przed-miotów można łatwo stworzyć z włókien; można je spleść ze sobą i doprowadzić do powstania grubego włókna, ale można też utkać w płaską tkaninę. Dlacze-go nie zrobić tego samego z komórkami hodowanymi w laboratorium? To był właśnie punkt wyjścia do badań japońskiego zespołu badaczy.
Zadanie wydaje się trudne, ale nic bardziej błędnego! Uczeni z Tokio wykorzystali alginian sodu – substancję, która po zetknięciu się z chlorkiem wapnia zmienia się w żel. Zresztą jest ona szeroko wykorzystywana w me-dycynie, m.in. jako nić do zszywania ran. Japońscy na-ukowcy zaprojektowali jednak specjalną „wyciskarkę”, dzięki której potrafili stworzyć osłonkę żelową przy-pominającą spaghetti. W środku umieścili niewielką
liczbę komórek razem z macierzą zewnątrzkomórkową (siecią rozmaitych białek otaczającą komórki znajdujące się w ogramizmie) i hodowali w laboratorium, tak samo jak hoduje się inne komórki. Po pewnym czasie ich licz-ba w żelu alginianowym na tyle wzrastała, że komórki tworzyły zwarte włókno. Ostatnim krokiem było roz-puszczenie żelu, w wyniku czego zespół prof. Takeuchi otrzymał włókna komórkowe.
Metodę wypróbowano na różnych rodzajach komórek. Okazało się, że włókna komórek nerwowych przeka-zywały impuls nerwowy, komórki mięśniowe uształ-towane w ten sposób potrafiły się kurczyć, a komórki nabłonkowe naczyń krwionośnych tworzyły struktury przypominające właśnie naczynia krwionośne.
Uczeni spróbowali też zastosować tę nową metodę w leczeniu cukrzycy. Wykorzystali jako model myszy laboratoryjne z uszkodzoną trzustką, która nie wydzie-lała insuliny. Co ciekawe, gdy owinęli oni nerkę myszy włóknem zbudowanym z komórek B trzustki, które wydzielają insulinę, poziom glukozy we krwi badanych myszy wrócił całkowicie do normy. Takiego efektu nie dawało umieszczenie komórek B znajdujących się w za-wiesinie w tym samym miejscu organizmu myszy. Ba-dania te wskazują więc na to, że nie tylko rodzaj komó-rek ma znaczenie, ale ich struktura może mieć wręcz decydujący wpływ na to, czy będą one funkcjonować po przeszczepieniu tak samo, jak w narządzie, z którego zostały wyodrębnione.
Swoboda tworzenia rozmaitych struktur przestrzen-nych, jaką dają włókna komórkowe, pozwala sądzić, że znowu zbliżyliśmy się o jeden krok do przyszłości, w której będzie można „wyhodować” w laboratorium upragniony narząd. Takao Ishikawa
Prof. Shoji Takeuchi z Center for International Research on MicroMechatronics (CIRMM)
Abstrakt artykułu, w którym zespół prof. Takeuchi prezentuje swoje osiągnięcie.Dostępny na: http://www.nature.com/nmat/journal/v12/n6/full/nmat3606.html#contrib-auth
Literatura
Onoe H, Okitsu T, Itou A, Kato-Negishi M, Gojo R, Kiriya D, Sato K, Miura S, Iwanaga S, Kuribayashi-Shigetomi K, Matsunaga YT, Shi-moyama Y, Takeuchi S (2013). Metre-long cell-laden microfibres exhibit tissue morphologies and functions. Nat Mater. 12:584-590.

Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce | Adam Kowalak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 83
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce Adam Kowalak
12 maja 2013 r. minęła 91. rocznica śmierci wybitne-go niemieckiego uczonego Hugo Conwentza, światowej sławy paleobotanika, archeologa, humanisty, twórcy nowoczesnych podstaw ochrony przyrody w Europie. Jego aktywność na tak wielu płaszczyznach, była przy-kładem rzadko spotykanego zarówno na przełomie XIX i XX wieku, jak i dziś holistycznego podejścia do nauki. Wywarła ona duży wpływ na ochronę przyrody w Europie, w tym w będącej jeszcze pod zaborami Pol-sce, choć można polemizować z autorami podającymi, że działalność Conwentza stanowiła fundament ochro-ny przyrody w naszym kraju. Współpracował z kilkoma polskimi uczonymi m.in. z Ferdynandem Chłapow-skim, Marianem Raciborskim, Władysławem Szaferem. Podkreślał ścisły związek kultury z przyrodą. Postulo-wał połączenie edukacji przyrodniczej i historycznej, uznając, że jest to podstawa edukacji patriotycznej. Ści-śle współpracował ze szkołami. Uważał, że tylko świa-dome społeczeństwo może skutecznie chronić przyro-dę i kulturę. Jego stosunek do edukacji i nauki do dziś uznać można za nowatorski.
Edukacja i kariera
Hugo Wilhelm Conwentz urodził się 20 stycznia 1855 r. we wsi Święty Wojciech, położonej w pobliżu Gdańska (obecnie dzielnica Gdańska), w menonickiej rodzinie rolniczej jako jeden z siedmiorga rodzeń-stwa. Jego rodzice (Albert Wilhelm i Augusta Dyck z domu Saalfeld), byli potomkami Holendrów, którzy osiedlili się na Żuławach uciekając przed prześlado-waniami religijnymi. Wzmianki o jego rodzinie po-jawiają się po raz pierwszy w dokumentach wsi Stare Szkoty (Altschottland)1 w 1673 r.2 Zgodnie z tradycją menonicką (anabaptystów) otrzymał surowe wycho-wanie moralne. Podstawę wychowania stanowiła uczci-wość, pracowitość, wytrwałość, umiłowanie porządku i oszczędność. Tymi zasadami kierował się przez całe życie. Gdy miał kilkanaście lat, ojciec sprzedał gospo-darstwo i przeprowadził się do Gdańska na Frauengasse 28 (obecnie ul. Mariacka), gdzie zajął się handlem wę-glem i drzewem. Hugo uczęszczał m.in. do gimnazjum realnego przy kościele świętego Jana. Olbrzymi wpływ na jego rozwój wywarł profesor Carl Th. Bail, rozwi-jając jego zainteresowania naukami przyrodniczymi, a w szczególności botaniką.
Studiował we Wrocławiu i Getyndze. Mając 21 lat (w 1876 r.), rozpoczął pracę asystenta we wrocławskim ogrodzie botanicznym. W tym samym roku napisał i obronił pracę doktorską na temat Skamieniałe drze-wa północnoniemieckiego dyluwium3 (Die versteinerten Hölzer aus dem norddeutschen Diluwium). Promotorem
1 Stare Szkoty to osada na gruntach darowanych przez księcia gdańskiego Mestwina II (Mściwoja II) biskupowi kujawskiemu; zamieszkana była przez Menonitów, Żydów i Szkotów.
2 Vogel S (1957). Conventz, Hugo Wilhelm. W: Neue Deutsche Biog-raphie. 3:347. Dostępny na: http://www.deutsche-biographie.de/pnd118676806.html.
3 Dyluwium – plejstocen.
pracy był prof. Heinrich Göppert4.
Tytuł profesora otrzy-mał już w 35. roku życia. Przyczynił się do tego jego olbrzymi dorobek z pa-leobotaniki (z tej dziedzi-ny był już specjalistą świa-towej sławy) i florystyki. Jego zainteresowania na-ukowe były jednak znacz-nie szersze. Cechowało go holistyczne podejście do nauki. Postrzegał ją jako sieć wzajemnych powiązań. Prowadził szeroko zakrojone prace badawcze nie tylko z paleobotaniki i florystyki ale również geologii, fitogeografii, archeologii i etnologii.
W 1878 r. powrócił do Gdańska. W 1879 r. władze pruskie powierzyły mu kierownictwo Muzeum Prowin-cji Zachodniopruskiej (Das Westpreußische Provinzial--Museum für Naturgeschichte, Gewebe und Kunst), którego siedzibę stanowiła Zielona Brama. Oficjalne otwarcie muzeum nastąpiło w 1880 r.5 Zaraz po objęciu stanowiska dyrektora przystąpił do przeglądu i inwen-taryzacji zbiorów. W ich wyniku stwierdził, że są one zbyt mało reprezentatywne dla Prus Zachodnich. Mając do dyspozycji ograniczone środki finansowe, postano-wił pozyskiwać eksponaty głównie podczas badań tere-nowych, prowadzonych przez specjalnie przygotowa-nych wolontariuszy. Korzystając z pomocy działającego w Gdańsku Towarzystwa Przyrodniczego rozwinął sze-roko zakrojoną działalność szkoleniową i popularyza-torską. Wygłosił dziesiątki odczytów w wielu miastach,
4 Kąmpfert H. Conwentz Hugo. Botaniker. W: Ostdeutsche Bio-grephie. Personlichkeiten des historischen deutschen Ostens. Do-stępny na: http://www.ostdeutsche-biographie.de.
5 Lippky G (1980). Das Westpreusiche Provinzial-Muzeum in Danzig 1880-1945 und seine vier Direktoren. Westpreusen Jahrb. Münster, s. 30, 105–115.
Dr inż. Adam Kowalak: adiunkt w Pracowni Dydaktyki Geografii i Ochrony Środowiska, Instytut Geografii i Stu-diów Regionalnych, Akademia Pomorska w Słupsku

Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce | Adam Kowalak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 84
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
organizacjach i szkołach Prus zachodnich. Ich wyni-kiem było wyzwolenie dużej aktywności społecznej. Do muzeum zaczęły napływać liczne eksponaty od różnych instytucji, szkół, leśniczych, księży i rolników6. Szcze-gólnie aktywni okazali się nauczyciele. Conwentz regu-larnie uczestniczył w konferencjach nauczycielskich, na których zapoznawał słuchaczy z przyrodą, prehistorią i kulturą regionu. Przekonywał o konieczności moż-liwie szybkiego informowania muzeum o niszczeniu cennych obiektów przyrodniczych i stanowisk archeo-logicznych.
Z pomocą tak dużego zaplecza społecznego Hugo Conwentz wzorowo przeprowadził inwentaryzację za-bytków natury na obszarze Pomorza Gdańskiego. Wy-niki swoich badań opublikował w książce Forstbotani-sches Markbuch fur Westpreussen (Pamiętniki drzew Prus Zachodnich) wydanej w 1900 r. Książka ta zgodnie z rozporządzeniem ministra rolnictwa uznana została za wzór dla innych regionów Prus.
Był botanikiem, toteż szczególnie dużo uwagi po-święcał rzadkim roślinom takim m.in. jak cis, kotewka, brzęk czy limba. W wydanej w 1892 r. książce Die Eibe in Westpreussen, elner austerbender Waldbaum7 (Cis w Prusach Zachodnich, drzewo lesne zagrozone wymar-ciem) autor omawia zasięg i występowanie tego gatunku w Niemczech (także na terenach polskich). Szczególnie ciekawe są fragmenty opisujące miejsce tego drzewa w kulturze ludowej.
Sformułował pierwsze naukowe zasady klasyfikacji drzew i innych obiektów przyrodniczych na pomniki przyrody. Jego praca Die Gefahrung der Naturdenkma-ler und Vorschlage zu irer Erhaltung (Zagrozenia po-mników przyrody i propozycje ich utrzymania), wydana
6 Szafer W (1965). Zarys historii ochrony przyrody w Polsce. W: Ochrona przyrody i jej zasobów. Problemy i metody. Kraków, s. 71–72.
7 Conwenz H (1892). Die Eibe in Westpreussen, ein aussterbender Waldbaum, Danzig.
w 1904 r., jest uważana za osobisty manifest ochrony przyrody.
Z dziedziny paleobotaniki na uwagę zasługuje jego praca Monographie der baltischen Burnsteinbaume (1890)8. Zgromadził również zbiory bursztynów. Część tych zbiorów, uratowana od zniszczenia po drugiej woj-nie światowej9 wchodzi w skład stałej wystawy Muzeum Archeologicznego w Gdańsku „Z bursztynem przez ty-siąclecia”.
W swoich publikacjach poruszał także konieczność ochrony zwierząt (bobra, żółwia, wilka). Spis jego pub-likacji obejmował 262 pozycje. Niektóre z nich ukaza-ły się w przekładach w większości krajów Zachodniej i północnej Europy a nawet w Ameryce.
Dzięki staraniom Conwentza w październiku 1906 r. utworzony został w pierwszy w Prusach urząd konserwatora przyrody działający w ramach admi-nistracji rządowej. Urząd ten czasowo, ze względu na miejsce zamieszkania jego projektodawcy i kierownika, miał siedzibę w Gdańsku. W 1910 r. przeniesiono go do Berlina, gdzie przeprowadził się również Conwentz.
Podróże zagraniczne i ich rezultaty
Działalność Conwentza wraz z całą wszechstron-nością jego poglądów na ochronę przyrody miała ogromny wpływ na rozwój tej idei w Europie. Był on propagatorem szeroko pojętej ochrony przyrody na forum międzynarodowym. Przez odczyty wygłaszane w różnych miastach europejskich, kontakty ze światem nauki i władzami wielu państw przyczynił się w dużej
8 Conwentz H (1890). Monographie der baltischen Bernsteinböume. Gdańsk. 151 stron.
9 Zbiory Muzeum Przyrodniczego zaginęły lub uległy zniszczeniu podczas II wojny światowej w latach 1942–1944 i w czasie prze-mieszczania na Żuławy Wiślane w latach 1941–1945, gdzie mia-ły zostać ukryte. Najlepiej zachowała się stosunkowo niewielka część zbiorów wywieziona do Kaczynosa koło Starego Pola.
mierze do zaszczepienia lub rozwoju tej idei zarówno w swojej ojczyźnie, jak i w krajach sąsiednich. Wy-mowa jego argumentacji i wysoka ocena uzyskanych osiągnięć naukowych zjednały mu powszechne uznanie w świecie. I tak np. był dwukrotnie w Wiedniu w 1903 i w 1907 r. na Kongresie Gospodarki Rolnej i Leśnej. Po wizycie w 1903 r., austriackie ministerstwo oświa-ty wydało polecenie zbierania, w krajach wchodzących w skład monarchii, wiadomości o obiektach przyrodni-czych zasługujących na ochronę. Niestety w całej mo-narchii austriackiej (a więc także w polskiej Galicji) do-konana inwentaryzacja poszła na marne, ponieważ jej wyniki ugrzęzły w urzędach centralnych i nikt się nimi ze strony władz nie interesował.
Znacznie trwalsze efekty przyniosły podróże Con-wentza do Danii, Holandii i Szwecji. Jego wykłady i liczne docierające tam publikacje przyczyniły się do znacznego ożywienia ruchu ochrony przyrody w tych krajach inspirując do działania wielu naukowców, dzia-łaczy społecznych i polityków.
Jego działalność została doceniona także w winnych krajach m.in. we Francji, gdzie został zaproszony na I Kongres Ochrony Krajobrazu, który odbył się w Pary-żu w 1909 r., oraz w Rosji – został on zaproszony na 13 Konferencję Przyrodników i Lekarzy w Tbilisi (1913).
Poglądy na edukację
Zaangażowanie Conwentza w edukację stanowi-ło bardzo ważny element jego działalności naukowej. Podniesienie świadomości związku z przyrodą i kul-turą oraz obudzenie lokalnego patriotyzmu uznał on za podstawową metodę przygotowania rejonu badań. Aktywizację społeczności lokalnej uważał za klucz do zachowania dziedzictwa przyrodniczego i kulturowego. Jak już wspomniałem, gromadzenie materiałów i eks-ponatów do muzeum rozpoczął od kształcenia wolonta-

Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce | Adam Kowalak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 85
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
riuszy spośród członków towarzystw regionalnych, na-uczycieli i uczniów różnych typów szkół. Uczestniczył w dziesiątkach spotkań i szkoleń. Edukował zarówno dorosłych jak i młodzież szkolną. Od początku swojej działalności ściśle współpracował ze szkołami zarów-no w dziedzinie nauk przyrodniczych jak i humani-stycznych, utrzymując stałe kontakty z nauczycielami. Swoją działalnością przyczynił się w dużym stopniu do podniesienia kwalifikacji nauczycieli oraz wzrostu za-interesowania szkół tą problematyką. Nie były mu obce problemy prowincjonalnych placówek oświatowych. Wskazywał na konieczność przetransponowania wie-dzy z ośrodków naukowych do szkół.
Conwentz uczestniczył aktywnie zarówno w opra-cowywaniu przepisów prawnych dotyczących ochrony przyrody i zabytków jak i opracowywaniu i wprowa-dzaniu do programów szkolonych treści związanych z ta tematyką. Uważał on, że u podstaw skutecznej ochrony przyrody leży edukacja. Prezentował holi-styczne podejście do edukacji – postulował połączenie edukacji przyrodniczej (biologicznej, geograficznej) z historyczną. Wskazywał na ścisłe wzajemne powią-zania przyrody z kulturą. Swoje stanowisko prezento-wał także na wielu spotkaniach poza granicami kraju m.in. w Szwecji i Austrii10. Uznawał pomniki przyrody (zabytki przyrody) za ważny element historii kraju i re-gionu. Prowadząc badania archeologiczne starał się ich wyniki szeroko popularyzować w kontaktach z placów-kami naukowymi i oświatowymi oraz stowarzyszenia-mi regionalnymi.
W 1889 r. pruskie władze oświatowe zaleciły Con-wentzowi przygotowanie zestawów eksponatów muze-alnych dla szkół, które mogłyby być wykorzystywane dla celów edukacyjnych. Wykonanie tego postulatu już
10 Deutsches Biographies Jahrbuch. Hersusgegeben vom Verbande der Deutschen Akademien Band IV das Jadr 1922. Stuttgart, Berlin und Leipzig 1929, s. 21–25.
z założenia można było uznać za mało realne ze wzglę-du na brak możliwości pozyskania dostatecznej ilości eksponatów. W zamian opracował on zestaw ściennych tablic poglądowych i przekonał władze o celowości roz-powszechnienia ich jako pomocy dydaktycznych. Było to na owe czasy rozwiązanie nowatorskie. Tablice te doczekały się trzech wydań i używane były w szkołach różnych szczebli w tym w szkołach ludowych.
Hugo Conwentz wobec idei tworzenia parków narodowych
Conwentz, wbrew opinii prezentowanej przez wielu autorów jego biografii uznających go za „ojca parków narodowych”11, nigdy nie popierał zakładania parków narodowych. Przypisywanie mu udziału w utworzeniu w Szwecji pierwszych parków narodowych w Europie bądź też zainspirowania utworzenia parków w Danii, Norwegii czy też Białowieskiego Parku Narodowego w Polsce jest nadinterpretacją. Wprawdzie niewątpliwie jego działalność przyczyniła się do zaktywizowania ruchu ochrony przyrody na naszym kontynencie oraz rozwoju naukowych metod badawczych obiektów i terenów cennych przyrodniczo, co pośrednio przy-czyniło się do utworzenia (niejako wbrew intencjom Conwentza) wielkoobszarowych form ochrony, jakimi są parki narodowe.
Wpływ działalności Hugo Conwentza na ochronę przyrody w Polsce
W pierwszym okresie działalności Conwentza Pol-ska nie istniała jako samodzielny organizm państwowy – była pod zaborami, możemy więc mówić o ziemiach Polski a nie o państwie polskim. Jak wiadomo Con-
11 Gdańszczanie swiatu – Wydział Postaciologii. Hugo Conwentz. Dostępny na: http://rzygacz.pl/index.php?id=43,432,0,0,1,0.
wentz, zanim zyskał sławę światową jako pionier no-woczesnej ochrony przyrody, pracował w Gdańsku i na Pomorzu, a więc w zaborze pruskim. Zasadnicze zna-czenie dla ziem polskich miało uznanie przez pruskie ministerstwo rolnictwa wspomnianej już książki For-stbotanisches Merkbuch fur Westpreussen za instrukcję i wzór dla pozostałych dzielnic pruskich. Publikacja ta zainspirowała opracowanie podobnych wydawnictw na terenie ziem polskich zajętych przez Prusy. W 1904 r. poznański nauczyciel gimnazjalny i kustosz niemie-ckiego muzeum F. Pfuhl wydał Pamiętnik drzew i krze-wów Księstwa Poznańskiego (Walder und Baume der Provintz Posen), a w 1906 r. T. Schube swój Pamiętnik lasów sląskich (Waldbuch von Schlesien). W wydawni-ctwach towarzystw naukowych działających w zniem-czałych miastach zaboru pruskiego: Toruniu, Byd-goszczy, Grudziądzu i innych, w pismach codziennych i periodykach pojawiły się liczne informacje na temat osobliwości przyrodniczych, które należałoby chronić. Prowadzony przez Conwentza centralny organ rządu pruskiego dla ochrony przyrody w Berlinie (Staatliche Stelle fur Naturdenkamalpflege) objął w 1906 r. kierow-nictwo tego potężnego ruchu. Dziesiątki tysięcy odezw i z niemiecką pedanterią ułożonych kwestionariuszy (w samym Księstwie Poznańskim zebrano ich ponad 12.000) krążyło po całym kraju. Zgromadzony mate-riał pozwolił na objęcie ustawową ochroną kilka tysięcy obiektów przyrodniczych w prowincji pruskiej. Obok licznych nazwisk niemieckich w realizację idei ochro-ny przyrody w Księstwie Poznańskim włączyli się tak-że Polacy. Czołowa postacią był tu dyrektor Muzeum Wielkopolskiego Ferdynand Chłapowski, który prowa-dził ożywiona korespondencją z profesorem Racibor-skim. Obok Chłapowskiego duży wkład w dokumen-towanie walorów przyrodniczych Wielkopolski wnieśli Józef Rivoli – wybitny leśnik i Jerzy W. Szulczewski – przyrodnik.

Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce | Adam Kowalak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 86
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Conwentz wywarł widoczny wpływ na ochro-nę przyrody także w dwóch pozostałych zaborach. W 1916 r. zwiedził Puszczę Białowieską i przyczynił się do utworzenia tzw. parku natury – fragmentu puszczy położonego w widłach Hwoźnej i Narewki, gdzie odtąd nie prowadzono wyrębu drzew. W 1921 r. utworzono tam rezerwat (obecnie rezerwat ścisły w obrębie Biało-wieskiego Parku Narodowego). Jego działalność przy-czyniła się do, niestety krótkotrwałej, ochrony żubrów.
W zaborze austriackim, gdzie tradycje czynnej ochrony przyrody wyprzedzały o kilkanaście lat dzia-łalność Conwentza, inicjatywa lokalnych władz pol-skich napotykała jednak na opór ze strony centralnych władz austriackich12. Dopiero podróże tego wybitne-go uczonego do Wiednia spowodowały, czasową jak się później okazało, zmianę podejścia rządu austria-ckiego do tej problematyki. Wspomniany już reskrypt ministerstwa oświaty w Wiedniu (z 30 listopada 1903 r.), wydany po wizycie Conwentza, polecający czynni-kom rządowym zajęcie się sprawą ochrony przyrody, wymusił reakcję przedstawiciela rządowego w Gali-cji. Namiestnictwo we Lwowie zwróciło się w lutym 1904 r. do instytucji i osób prywatnych w Galicji i Lo-domerii, a także w Księstwie Krakowskim, z wezwa-niem do przekazywania mu informacje o wszelkich zabytkach przyrody, które zasługują na ochronę. Na apel odpowiedziało wiele organizacji i duża liczba osób prywatnych przesyłając opisy rozmaitych osobliwości natury. Najaktywniejszą organizacją było Towarzystwo Przyrodników im. Kopernika, które przedstawiło spis kilkudziesięciu osobliwości przyrody zasługujących na
12 Już w 1867 r. Sejm Krajowy Królestwa Galicji i Lodomerii uchwa-lił ustawę chroniącą zwierzęta alpejskie występujące w Tatrach (ustawa uzyskała moc obowiązującą w 1869 r.). Równocześ-nie Sejm Lwowski przyjął drugą analogiczna ustawę ochron-ną „o ochronie pożytecznych ptaków i innych zwierząt”, ale ta ustawa nigdy nie zyskała sankcji cesarskiej – władze centralne w Wiedniu uniemożliwiły jej wejście w życie.
ochronę i zaproponowało utworzenie kilku większych rezerwatów górskich w Karpatach (Tatry, Pieniny, Kar-paty Wschodnie) i stepowych na Podolu, w okolicy Bor-szczowa. Zbliżone żądania wysunął Wydział Filozoficz-ny Uniwersytetu Jagiellońskiego. Niestety, podobnie jak w pozostałych częściach Cesarstwa Austriackiego, wyniki podjętej z inspiracji Conwentza akcji zostały za-przepaszczone. Namiestnictwo w byłej Galicji okazało się godne swojej władzy zwierzchniej. Memoriały, plany i sporządzona inwentaryzacja obiektów i terenów cen-nych przyrodniczo złożona ad acta zaginęła.
Na szczęście znalazło się wówczas w społeczeństwie byłego zaboru austriackiego dość energii żeby zaprze-paszczoną przez rząd centralny akcję dalej prowadzić. Wiodąca rolę przejęło Towarzystwo Przyrodników im. Kopernika, które pod kierunkiem prof. Mariana Raciborskiego opracowało w latach 1906–1907 plan działalności na tym polu. W swoich działaniach Ra-ciborski ściśle współpracował z wybitnym geografem polskim Ludomirem Sawickim13. Conwentz wysoko oceniał osiągnięcia Raciborskiego – obaj uczeni prowa-dzili ożywioną korespondencję wymieniając się swoimi doświadczeniami naukowymi i organizacyjnymi. Po przedwczesnej śmierci prof. Raciborskiego współpracę z Conwentzem kontynuował jego student, współpra-cownik i następca prof. Władysław Szafer, projektodaw-ca i pierwszy przewodniczący utworzonej przez rząd II Rzeczpospolitej Tymczasowej Komisji Ochrony Przyro-dy, która w 1926 r. przekształcona została w Państwową Radę Ochrony Przyrody.
13 Szafer W (1965). Zarys historii ochrony przyrody w Polsce. W: Ochrona przyrody i jej zasobów. Problemy i metody. PAN, Zakład Ochrony Przyrody, Kraków, s. 67.
Ocena działalności
W świetle powyższych faktów można stwier-dzić, że działalność Conwentza wywarła bardzo duży wpływ na rozwój koncepcji ochrony przyrody w Pol-sce. Można jednak polemizować z Eugeniuszem No-wakiem z Bundesforschungsanstalt für Naturschutz und Landschaftsökologie, że jego działalność stanowiła fundament organizacji systemu ochrony zasobów przy-rodniczych w naszym kraju14. Należy tu zaakcentować, że to argumentacja naukowa polskich uczonych skłoni-ła Galicyjski Sejm Krajowy do uchwalenia unikalnych w skali Europy ustaw ochroniarskich w 1867 r., a więc na kilka lat przed rozpoczęciem przez Conwentza pracy naukowej. Ponadto niewyobrażalne wręcz były różnice w warunkach działalności i możliwościach realizacji koncepcji badawczych polskich uczonych (Janoty, Raci-borskiego, Pawlikowskiego i innych) działających w by-łej Galicji – kraju biednym i zacofanym, a prof. Con-wentza – kierownika państwowego urzędu w bogatym kraju, jakim były Prusy. Realizacja koncepcji ochrony przyrody w na ziemiach Polskich (poza zaborem pru-skim) wymagała zastosowania metod odbiegających znacznie od tych stosowanych przez Connwentza na Po-morzu, np. na terenie Galicji, poszukiwanie partnerów do współpracy w trzyletnich szkołach trywialnych było ze względu na ich poziom w zasadzie niemożliwe. Szafer wyprzedził również Conwentza w dążeniu do tworze-nia parków narodowych, z których ideą niemiecki uczo-ny do końca życia polemizował. Nie umniejsza to oczy-wiście zasług prof. Conwentza, słusznie uważanego za jednego z czołowych twórców konserwatorskiej ochro-ny przyrody w Europie, a więc także w Polsce.
14 Nowak E. O dziedzictwie Państwowej Placówki Zachowania Za-bytków Przyrody w Gdańsku oraz wpływie jej kierownika profe-sora Hugo Conwentza na ochronę przyrody w Polsce. W: Przyro-da Polska, Forum, Dostępny na: http://www.iop.krakow.pl.

Hugo Conwentz (1855–1922) a ochrona przyrody w Polsce | Adam Kowalak | EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013 87
KRÓ
TKO
NAU
KASZ
KOŁA
EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA | ebis.ibe.edu.pl | [email protected] | © for the article by the Authors 2013 © for the edition by Instytut Badań Edukacyjnych 2013
Zamiast zakończenia
Niespotykana aktywność zawodowa i społeczna oraz pasja dobrej pracy nie pozostawiała Conwentzo-wi zbyt wiele czasu na sprawy prywatne. Ponadto jego skrupulatność i wysokie wymagania wobec siebie i in-nych, wyniesione z menonickiego domu, sprawiały, że przez swoich współpracowników uważany był często za człowieka trudnego we współżyciu. Zarzucano mu nad-mierny upór i bezkompromisowość. Ożenił się mając 64 lata z Grete Ekelöf – bibliotekarką Szwedzkiej Biblio-teki Narodowej, która wkrótce stała się jego najbliższą współpracownicą. Zmarł bezpotomnie w Berlinie 12 maja 1922 r. W 1940 r. ukazała się powieść biograficzna opisująca życie tego wybitnego uczonego autorstwa jego wsółpracownicy Mergarete Boie15. Jego nagrobek na cmentarzu Sűdwestkirchhof Stahnsdorf został uchwa-łą Senatu Berlina w 1952 r. uznany za pomnik. 7 maja 1996 r., ustanowiony został przez ministra ochrony śro-dowiska Landu Schleswig-Holstein, medal im Hugona Conwentza (Hugo Conwentz-Medaille), wręczany za zasługi w dziedzinie ochrony przyrody16. Do jego licz-nych zasług w dziedzinie konserwatorskiej ochrony przyrody i archeologii należałoby dodać jeszcze jedną, na ogół przez biografów pomijaną – Conwentz w ciągu całej swojej działalności podkreślał rolę edukacji przy-rodniczej, historycznej i humanistycznej w zachowaniu zabytków przyrody i kultury. Podkreślał ścisły związek przyrody i kultury. Wskazywał na olbrzymie znaczenie świadomości społecznej i poziomu edukacji dla rozwo-ju nauki, a co za tym idzie dla rozwoju społeczeństwa i kraju. Jego holistyczne podejście do edukacji wykra-czało znacznie poza epokę w której żył.
15 Boie M (1940). Hugo Conwentz und seine Heimat. Stutgard.16 Kämpfert H. Ostdeutsche Biogrephie. Personlichkeiten des histo-
rischen deutschen Ostens. Conwentz Hugo. http://www: ostdeu-tsche-biographie.de s. 4.
Literatura
Boie M (1940). Hugo Conwentz und seine Heimat, Stuttgart 1940.Conwentz H (1890). Monographie der baltischen Bernsteinbäume.
Gdańsk.Conwentz H (1892). Die Eibe in Westpreussen, ein aussterbender
Waldbaum. Danzig.Deutsches Biographisches Jahrbuch. Herausgegeben vom Verbande
der deutschen Akademien. Band IV. Daz Jahr 1922. Stutgart, Ber-lin und Leipzig 1929.
Gdańszczanie swiatu – Wydział Postaciologii, Hugo Conwentz. Dostępny na: http:// rzygacz.webd.pl. Dostęp 2.09.2012.
Kampfert H. Hugo Conwentz. Botaniker. W: Ostdeutsche Biogrephie. Personlichkeiten des historischen deutschen Ostens. Dostępny na: http:// www: ostdeutsche-biographie.de (dostęp 26.09.2012)
Krzemińska E, Krzemiński W, Haenni JP, Dfour Ch (1993). W bur-sztynowej pułapce. Kraków.
Leńkowa A (1986). Ochrona przyrody w świecie. W: Człowiek prze-ciwko sobie. Warszawa: PAX.
Nowak E (2006). O dziedzictwie Państwowej Placówki Zachowania Zabytków Przyrody w Gdańsku oraz wpływie jej kierownika profesora Hugo Conwentza, na ochronę przyrody w Polcse. W: Przyroda Polska. FORUM. Dostępny na: http://www.iop.krakow.pl. Dostępny na: 2.09.2012.
Szafer W (1965). Zarys historii ochrony przyrody w Polsce. W: Ochrona przyrody i jej zasobów. Problemy i metody. Kraków: PAN, Zakład Ochrony Przyrody.
Vogel S (1957). Conwentz Hugo Wilhelm. W: Neue Deutsche Bio-graphie 3:347. Dostępny na: http:www.deutsche-biographie.de/pnd118676806.html. Dostęp: 25.08.2012.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznik | © for the by the Authors and Instytut Badań Edukacyjnych 2013
Scenariusz lekcji doświadczalnej (M. Zaród)
Prawo Ohma w praktyce
Cel eksperymentu: Zbadanie prostych obwodów elek-trycznych. Poznanie różnicy między połączeniem sze-regowym i równoległym.Etap edukacyjny: III – gimnazjum.Podstawa programowa: Fizyka dla III etapu edukacyj-nego. Wymagania eksperymentalne 9.7, 9.8. i 9.9. Wy-magania teoretyczne: 4.7, 4.8 i 4.9.Czas realizacji: 15 min. – przygotowanie, 65 min. – pomiary i 20 min. – dyskusja wyników i czynności administracyjne. Sugeruję wykorzystanie przerwy mię-dzy lekcjami (za zgodą uczniów) na cele dydaktyczne – eksperymenty są rzadkie i warto wykorzystać cały możliwy czas. W razie potrzeby skrócenia proponuję wykreślenie jednego z pytań w części eksperymental-nej. W siatce godzin gimnazjum trudno o taki zakres czasu, ale rezygnacja z dyskusji i części eksperymentów ogranicza efekt dydaktyczny lekcji eksperymentalnej. Wyposażenie (na jedno stanowisko): • źródło prądu: zasilacz prądu stałego o napięciu
12 V lub układ kilku połączonych szeregowo ba-terii AA (przynajmniej 5 sztuk); w tym drugim przypadku połączenie najlepiej zrobić „na stałe” (np. doklejonymi blaszkami miedzianymi), aby nie rozpadło się w trakcie lekcji;
• przewody do połączenia elementów (najlepiej z do-łączonymi „krokodylkami”);
• 3 żarówki na napięcie 12 V (o mocy 1,5 W, najlepiej różnokolorowe), 1 żarówka o mocy 5 W, również na napięcie 12 V. Sugerowane jest użycie żarówek mających gwinty wtykowe (m.in. typu MR16) – np. COB5SE (prod. Lumenmax);
• 2 mierniki uniwersalne;• (nieobowiązkowe): komputer z arkuszem kalkula-
cyjnym.
Kursywą podano sugerowaną narrację ćwiczenia dla uczniów. Fizyka w formie fabularyzowanej jest ła-twiejsza do zrozumienia niż suche polecenia.
1. Przygotowanie (na przerwie, zanim przyjdą uczniowie):
• ustawić ławki, rozłożyć wyposażenie stanowisk na ławkach;
• sprawdzić zasilacze i mierniki; usunąć ewentualne usterki;
• wypróbować eksperyment z grupą animatorów, poinstruować ich o zasadach bezpieczeństwa i ce-lach eksperymentu.
2. Eksperymenty:
• Podział uczniów na grupy (sugerowany jest podział mieszający uczniów z różnymi wynikami w nauce, aby słabsi uczyli się od mocniejszych).
• Wprowadzenie uczniów w tematykę ćwiczenia: Prąd elektryczny to dla nas źródło światła i ciepła. Elektrycz-ność zasila nasze telefony, komputery, zabawki i dzwonki w szkole. Dzisiaj poznacie podstawy inżynierii elektrycznej. To jest źródło prądu <wskazujemy na zasilacz lub baterię>, to są żarówki różnej mocy <wskazujemy na żarówki>, to są przewo-dy i mierniki <demonstrujemy>. Co zrobić aby żarówka świe-ciła?
• Uczniowie podłączają żarówkę o mocy 1,5 W, tuto-rzy i nauczyciel pomagają w miarę potrzeby.
Świetnie, zrobiliście właśnie pierwszy krok, by zostać inżynie-rami elektrotechniki. Co płynie w przewodach? Co to jest prąd elektryczny i jak zmierzyć jego natężenie? Czy prąd płynie w ot-wartym obwodzie?
• Uczniowie mierzą i notują natężenie prądu, po-mocnicy asystują.
Co sprawia, że prąd płynie? Dlaczego zasilacz ma dwa różne zaciski? Co oznacza plus, a co minus? Czym różnią się te dwa zaciski? Co wytwarza zasilacz, kiedy obwód jest otwarty?
• Uczniowie mierzą i notują napięcie na zasilaczu w układzie otwartym (ostrożnie!). Mierzą też na-pięcie na żarówce 1,5 W.
Inżynierowie używają wielu urządzeń elektrycznych, mających różne funkcje. Spróbujcie sprawdzić, czym różnią się żarówki 1,5 W i 5 W. Czy są różnice w napięciu i natężeniu prądu płyną-cego przez żarówki o mocy 1,5 W w różnych kolorach?
W jaki sposób można podłączyć do prądu dwie żarówki 1,5 W? Jak to zrobić, aby obie świeciły najjaśniej?
• Uczniowie eksperymentują z połączeniami żaró-wek.
Co się stanie, gdy podłączę trzecią żarówkę do dwóch pozosta-łych? Co się stanie, gdy będzie to żarówka o większej mocy?
• Pomocnicy i nauczyciel demonstrują połączenie równoległe. Uczniowie mierzą prąd i napięcie w połączeniu równoległym na każdej z żarówek.
Co się stanie, gdy żarówki podłączymy w inny sposób? Jak ina-czej można je połączyć? Jak połączyć 3 żarówki o mocy 1,5 W, aby przez każdą z nich płynął taki sam prąd?
• Uczniowie eksperymentują z różnymi konfigura-cjami połączeń. Animatorzy i nauczycie asystują
Spróbujcie narysować obwody układów, które skonstruowali-ście. Nanieście na schematy wyniki pomiarów napięcia i natę-żenia prądu.
3. Dyskusja:
Wypróbujmy różne sposoby podłączenia żarówek? Jak wtedy mierzyć napięcia i natężenia? Jak będą różniły się wartości po-miarów? W którym połączeniu przez wszystkie żarówki płynie taki sam prąd?
• Uczniowie odpowiadają na pytania zadane przez nauczyciela w trakcie lekcji, nauczyciel moderuje dyskusję. Tutorzy wprowadzają wyniki pomiarów uczniowskich do komputera lub wypisują je na tablicy.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom A
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
1. Narysuj schemat sieci pokarmowej, łącząc ze sobą wszystkie organizmy (żyjące na polu). Zastosuj w schemacie maksymalną liczbę możliwych do zbudowania łańcuchów pokarmowych.
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Użyj do opisu organizmu maksymalną liczbę pasujących do niego terminów.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz wpływ pestycydów na każdą z grup organizmów, uwzględnionych w sieci pokarmowej, narysowanej przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Na schemacie sieci pokarmowej, sporządzonej przez Ciebie w punkcie 1 narysuj źródło energii i oznacz strzałkami kierunki jej przepływu przez ekosystem pola.
6. Wyjaśnij szczegółowo, jak użycie przez rolnika pe-stycydów na polu może wpłynąć na funkcjonowa-nie krogulców.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
7. Sformułuj i zapisz kilka problemów, które mogą pojawić się w funkcjonowaniu ekosystemu pola po zastosowaniu na nim pestycydów oraz zaproponuj co najmniej 3 rozwiązania alternatywne, które po-mogą chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom B
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
1. Narysuj schemat sieci pokarmowej, łącząc ze sobą wszystkie organizmy żyjące na opisanym polu). W sieci powin-no być uwzględnionych minimum 3 łańcuchy pokarmowe (może być ich więcej).
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Powinieneś użyć więcej niż jednego terminu do opisu organizmu.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz wpływ pestycydów na każdą z grup organizmów, uwzględnionych w sieci pokarmowej, narysowanej przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Na schemacie sieci pokarmowej, sporządzonej przez Ciebie w punkcie 1 narysuj źródło energii i oznacz strzałkami kierunki jej przepływu przez ekosystem pola.
6. Sformułuj i zapisz kilka problemów, które mogą pojawić się w funkcjonowaniu ekosystemu pola po zastosowaniu na nim pestycydów oraz zaproponuj co najmniej 2 rozwiązania alternatywne, które po-mogą chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
kapusta1
Sole mineral-ne, woda z podłoża/gleby
ślimaki2Liście kapusty, liście innych roślin
Larwy motyla bielinka kapust-nika
Liście kapusty
mszyce3Sok z tkanek łodyg i liści kapusty
Zdjęcie/rysunek organi-zmu
Nazwa organi-zmu
Pokarm, któ-rym się żywi
drozdy4Ślimaki, inne drobne zwie-rzęta
sikorki5
Larwy mo-tyla bielinka kapustnika i mszyce
biedronki6 mszyce
krogulec7 Drozdy i si-korki
Karta pracy ucznia – poziom C
Imię i nazwisko ucznia: .................................................Klasa: .......................
Informacja wstępna:
Rolnik dysponuje dużym polem, na którym upra-wia kapustę. Na polu, oprócz uprawianej kapusty żyją rozmaite dzikie organizmy, żywiące się kapustą, okre-ślane powszechnie jako szkodniki. Każdego roku rolnik stosuje opryski z pestycydów, skierowane bezpośrednio do szkodników, które je niszczą, zabijają. Oprócz tzw. szkodników na polu żyją organizmy, które się nimi żywią.
Zadanie
Wykorzystując informacje z tabeli nr 1 oraz własną wie-dzę i rozumienie funkcjonowania łańcuchów pokar-mowych i sieci pokarmowych napisz tekst, w którym wyjaśnisz, dlaczego stosowanie pestycydów czyli sub-stancji zabijających tzw. szkodniki może stwarzać prob-lemy w funkcjonowaniu pola i w jego produktywności. Zastosuj w swoim opracowaniu terminy:
biomasa, dwutlenek węgla, mięsożercy/drapieżcy, chlo-rofil, roślinożercy, liście, fotosynteza, konsument I i II rzę-du, producent, korzenie, łodyga/pęd, woda, sole mineral-ne, samożywność, cudzożywność.
Uwzględnij w swoim tekście realizację poleceń (na drugiej stronie):
Tabela 1. Lista organizmów, żyjących na polu, wraz z opisem pokarmu, którym się żywią
Źródła obrazów: http://ehealthspot.blogspot.com/2007_12_01_archive.html, http://blather.net/shitegeist/2008/05/snail_allegedly_prose-cuted_for_alleged_a.htm, http://www.mylot.com/w/photokeywords/greenfly.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/s/songthrush/index.aspx, http://www.rspb.org.uk/wildlife/birdguide/name/b/bluetit/index.aspx, http://blog.residesi.com/2009/02/violent--death-in-insect-world-stunning.html, http://fatfinch.wordpress.com/2008/08/27/sparrowhawks-v-pigeons
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Karta pracy autorstwa Urszuli Poziomek powstała w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
1. Narysuj prostą sieć pokarmową, łączącą trzy różne łańcuchy pokarmowe z opisanego pola, uwzględniając w nich organizmy na nim żyjące.
2. Określ każdy z organizmów pod kątem rodzaju pokarmu, którym się żywi, używając terminów z treści zadania (zob. poprzednia strona). Możesz użyć kilku terminów opisując jeden organizm.
Organizm Termin opisujący grupę do której zalicza się organizm ze względu na pokarm, którym się żywi
kapusta
ślimaki
larwy motyla bielinka kapustnika
mszyce
drozdy
sikorki
biedronki
krogulce
3. Wyjaśnij, w jaki sposób roślinożercy – np. mszyce – wpływają na funkcjonowanie kapusty jako organizmu i dla-czego – w związku z tym – farmer decyduje się na stosowanie oprysków. Użyj terminów z treści zadania.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
4. Opisz efekty stosowania pestycydów na pozostałe organizmy tworzące sieć pokarmową, narysowaną przez Ciebie w punkcie 1.
.....................................................................................................................................................................................................................
.....................................................................................................................................................................................................................
5. Sformułuj i zapisz jeden problem, który może po-jawić się w związku ze stosowaniem pestycydów na polu i zaproponuj jedno alternatywne rozwią-zanie, które pomoże chronić uprawę kapusty przed szkodnikami.
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................
...............................................................................................

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Biologia – które ciastko jest smaczniejsze?
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie
Zadanie
Pewien cukiernik postanowił sprawdzić, które z jego ciastek jest smaczniejsze: kremówka za 6 zł czy napoleonka za 8 zł. Do badania zaprosił zaprzyjaźnio-ną wielopokoleniową rodzinę. Każda z uczestniczących w badaniu osób zjadła najpierw napoleonkę, a potem kremówkę, po czym oceniała ich smak w skali od 1 do 10. Czy wszystkie elementy tego doświadczenia zostały poprawnie zaplanowane?
Sposób przeprowadzenia do-świadczenia Czy jest to poprawne?
1. Badani byli w różnym wieku. Tak / Nie
2.Badani byli ze sobą spokrew-nieni.
Tak / Nie
3.Wszyscy otrzymywali napoleon-kę jako pierwszą.
Tak / Nie
4.Badani nie znali cen ciastek i nie musieli za nie płacić.
Tak / Nie

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zadanie
Aby wytworzyć „naturalny plastik” domowym sposobem, można użyć mleka i octu. Należy w tym celu do litra wrzącego mleka dodać 40 ml octu. Tę mieszaninę dalej ogrzewa się, mieszając aż do momentu, kiedy pojawią się w niej biało-żółte bryłki, a sama zrobi się przejrzysta. Wtedy należy wyłączyć palnik, a po ostygnięciu prze-płukać grudki pod bieżącą wodą. Otrzymaną masę można łatwo formować, a następnie nale-ży odstawić na kilka dni, aż stwardnieje.
Tekst na podstawie: Skamieniałość z chomika. Zrób to sam!, Mick O’Hare, Insignis Media, Kraków 2011, s. 60–62
(1) Jaki proces spowodował pojawienie się białożół-tych grudek? A. Denaturacja. B. Zobojętnienie.(2) Opisany wyżej proces wywołały dwa czynniki.
Jednym z nich była zmiana pH, a drugim zmiana: A. temperatury. B. ciśnienia.
Chemia – naturalny plastik
Zadanie
Aby wytworzyć „naturalny plastik” domowym sposobem, można użyć mleka i octu. Należy w tym celu do litra wrzącego mleka dodać 40 ml octu. Tę mieszaninę dalej ogrzewa się, mieszając aż do momentu, kiedy pojawią się w niej biało-żółte bryłki, a sama zrobi się przejrzysta. Wtedy należy wyłączyć palnik, a po ostygnięciu prze-płukać grudki pod bieżącą wodą. Otrzymaną masę można łatwo formować, a następnie nale-ży odstawić na kilka dni, aż stwardnieje.
Tekst na podstawie: Skamieniałość z chomika. Zrób to sam!, Mick O’Hare, Insignis Media, Kraków 2011, s. 60–62
(1) Jaki proces spowodował pojawienie się białożół-tych grudek? A. Denaturacja. B. Zobojętnienie.(2) Opisany wyżej proces wywołały dwa czynniki.
Jednym z nich była zmiana pH, a drugim zmiana: A. temperatury. B. ciśnienia.
Zadanie
Aby wytworzyć „naturalny plastik” domowym sposobem, można użyć mleka i octu. Należy w tym celu do litra wrzącego mleka dodać 40 ml octu. Tę mieszaninę dalej ogrzewa się, mieszając aż do momentu, kiedy pojawią się w niej biało-żółte bryłki, a sama zrobi się przejrzysta. Wtedy należy wyłączyć palnik, a po ostygnięciu prze-płukać grudki pod bieżącą wodą. Otrzymaną masę można łatwo formować, a następnie nale-ży odstawić na kilka dni, aż stwardnieje.
Tekst na podstawie: Skamieniałość z chomika. Zrób to sam!, Mick O’Hare, Insignis Media, Kraków 2011, s. 60–62
(1) Jaki proces spowodował pojawienie się białożół-tych grudek? A. Denaturacja. B. Zobojętnienie.(2) Opisany wyżej proces wywołały dwa czynniki.
Jednym z nich była zmiana pH, a drugim zmiana: A. temperatury. B. ciśnienia.

EDUKACJA BIOLOGICZNA I ŚRODOWISKOWA 2/2013, załącznikZadanie powstało w ramach realizowanego przez Instytut Badań Edukacyjnych projektu Badanie jakości i efektywności edukacji oraz instytucjonalizacja zaplecza badawczego, współfinansowanego ze środków Europejskiego Funduszu Społecznego.
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Fizyka – jaki kupić czajnik?
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Zadanie
Marek wybrał się z rodzicami do sklepu, by kupić czajnik elektryczny. Spodobały im się dwa czajniki o tej samej pojemności, ale różnej mocy: 1500 W i 2300 W.
W dyskusji na temat wyboru czajnika padały różne argumenty.
Rozstrzygnij, które z nich są słuszne z punktu wi-dzenia fizyki.
Stwierdzenie Czy jest słuszne?
1.„Czajnik o większej mocy szybciej zagotuje tę samą ilość wody.”
Tak / Nie
2.„W czajniku o mniejszej mocy zagoto-wanie litra wody będzie tańsze.”
Tak / Nie
Related Documents











