Mol. Nutr. Food Res. 2013, 00, 1–16 1 DOI 10.1002/mnfr.201300277 REVIEW Molecular mechanisms underlying the potential antiobesity-related diseases effect of cocoa polyphenols Faisal Ali 1 , Amin Ismail 1,2 and Sander Kersten 3 1 Department of Nutrition and Dietetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, Selangor, Malaysia 2 Halal Products Research Institute, Universiti Putra Malaysia, Selangor, Malaysia 3 Metabolism and Genomics Group, Division of Human Nutrition, Wageningen University, Wageningen, The Netherlands Obesity and related metabolic diseases (e.g., type 2 diabetes, cardiovascular diseases, and hypertension) are the most prevailing nutrition-related issues in the world. An emerging feature of obesity is their relationship with chronic inflammation that begins in white adipose tissue and eventually becomes systemic. One potential dietary strategy to reduce glucose intolerance and inflammation is consumption of polyphenol-rich cocoa-like cocoa or their by-products. In vitro as well as in vivo data indicate that cocoa polyphenols (CPs) may exhibit antioxidant and anti-inflammatory properties. Polyphenols commonly found in cocoa have been reported to regulate lipid metabolism via inducing metabolic gene expression or activating transcription factors that regulate the expression of numerous genes, many of which play an important role in energy metabolism. Currently, several molecular targets (e.g., nuclear factor Kappa B, activated protein-1, peroxisome proliferator-activated receptors, liver X receptors, and adiponectin gene) have been identified, which may explain potential beneficial obesity-associated diseases effects of CPs. Further studies have been performed regarding the protective effects of CPs against metabolic diseases by suppressing transcription factors that antagonize lipid accumulation. Thus, polyphenols-rich cocoa products may diminish obesity-mediated metabolic diseases by multiple mechanisms, thereby attenuating chronic inflammation. Keywords: Cocoa polyphenols / Inflammation / Metabolic diseases / Molecular mechanism / Obesity Received: April 16, 2013 Revised: September 2, 2013 Accepted: September 9, 2013 Correspondence: Dr. Amin Ismail, Department of Nutrition and Di- etetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia E-mail: [email protected] Fax: +60389426769 Abbreviations: ACE, angiotensin-converting enzyme:; AP-1, acti- vated protein-1; CP, cocoa polyphenol; CVD, cardiovascular dis- ease; ERK, extracellular signal-related kinase; FAS, fatty acid syn- thetase; GLUT4, glucose transporter 4; iNOS, inducible nitric oxide synthase; JNK, c-Jun-NH2 terminal kinase; LXR-, liver X receptor-; MAPK, mitogen-activated protein kinase; NF-kB, nuclear factor Kappa B; NO, nitric oxide; NOS, nitrogen oxy- gen species; NR, nuclear receptor; PGC-1, PPAR- coactivator- 1; PPAR, peroxisome proliferator-activated receptor; ROS, reac- tive oxygen species; SIRT, sirtuin1; SREBP1, sterol regulatory elevated binding protein; TNF-, tumor necrosis factor-; UCP3, uncoupling protein 1 Introduction 1.1 Cocoa polyphenols (CPs) and bioavailability Cocoa beans and their by-products derived (Theobroma cacao L.) are used for a long period of time as major ingredient of chocolate and cocoa powder [1, 2]. Cocoa has been rec- ognized as a rich source of polyphenols. Total polyphenol content of the cocoa beans is about 6–8% by dry weight and its derivative black chocolate is considered one of the ma- jor contributors of antioxidants to the American diet after fruits and vegetables [2, 3]. So far, more than 8000 pheno- lic structures are known, ranging from simple molecules as phenolic acids to highly polymerized substances, such as flavonoids. The most predominat phenolic phytochemi- cals in cocoa are flavonoids, which include mainly flavonols, flavanols, flavanones, isoflavones, and nonflavonoids, e.g., phenolic acids and the less common stilbenes as shown in Fig. 1 [3,4]. Catechins, anthocyanidins/anthocyanins, flavonol C 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mol. Nutr. Food Res. 2013, 00, 1–16 1DOI 10.1002/mnfr.201300277
REVIEW
Molecular mechanisms underlying the potential
antiobesity-related diseases effect of cocoa polyphenols
Faisal Ali1, Amin Ismail1,2 and Sander Kersten3
1 Department of Nutrition and Dietetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia,Selangor, Malaysia
2 Halal Products Research Institute, Universiti Putra Malaysia, Selangor, Malaysia3 Metabolism and Genomics Group, Division of Human Nutrition, Wageningen University, Wageningen,The Netherlands
Obesity and related metabolic diseases (e.g., type 2 diabetes, cardiovascular diseases, andhypertension) are the most prevailing nutrition-related issues in the world. An emerging featureof obesity is their relationship with chronic inflammation that begins in white adipose tissueand eventually becomes systemic. One potential dietary strategy to reduce glucose intoleranceand inflammation is consumption of polyphenol-rich cocoa-like cocoa or their by-products. Invitro as well as in vivo data indicate that cocoa polyphenols (CPs) may exhibit antioxidant andanti-inflammatory properties. Polyphenols commonly found in cocoa have been reported toregulate lipid metabolism via inducing metabolic gene expression or activating transcriptionfactors that regulate the expression of numerous genes, many of which play an important role inenergy metabolism. Currently, several molecular targets (e.g., nuclear factor Kappa B, activatedprotein-1, peroxisome proliferator-activated receptors, liver X receptors, and adiponectin gene)have been identified, which may explain potential beneficial obesity-associated diseases effectsof CPs. Further studies have been performed regarding the protective effects of CPs againstmetabolic diseases by suppressing transcription factors that antagonize lipid accumulation.Thus, polyphenols-rich cocoa products may diminish obesity-mediated metabolic diseases bymultiple mechanisms, thereby attenuating chronic inflammation.
Keywords:
Cocoa polyphenols / Inflammation / Metabolic diseases / Molecular mechanism /Obesity
Received: April 16, 2013Revised: September 2, 2013
Accepted: September 9, 2013
Correspondence: Dr. Amin Ismail, Department of Nutrition and Di-etetics, Faculty of Medicine and Health Sciences, Universiti PutraMalaysia, 43400 UPM Serdang, Selangor, MalaysiaE-mail: [email protected]: +60389426769
Abbreviations: ACE, angiotensin-converting enzyme:; AP-1, acti-vated protein-1; CP, cocoa polyphenol; CVD, cardiovascular dis-ease; ERK, extracellular signal-related kinase; FAS, fatty acid syn-thetase; GLUT4, glucose transporter 4; iNOS, inducible nitricoxide synthase; JNK, c-Jun-NH2 terminal kinase; LXR-�, liverX receptor-�; MAPK, mitogen-activated protein kinase; NF-kB,nuclear factor Kappa B; NO, nitric oxide; NOS, nitrogen oxy-gen species; NR, nuclear receptor; PGC-1�, PPAR-� coactivator-1; PPAR, peroxisome proliferator-activated receptor; ROS, reac-tive oxygen species; SIRT, sirtuin1; SREBP1, sterol regulatoryelevated binding protein; TNF-�, tumor necrosis factor-�; UCP3,uncoupling protein
1 Introduction
1.1 Cocoa polyphenols (CPs) and bioavailability
Cocoa beans and their by-products derived (Theobroma cacaoL.) are used for a long period of time as major ingredientof chocolate and cocoa powder [1, 2]. Cocoa has been rec-ognized as a rich source of polyphenols. Total polyphenolcontent of the cocoa beans is about 6–8% by dry weight andits derivative black chocolate is considered one of the ma-jor contributors of antioxidants to the American diet afterfruits and vegetables [2, 3]. So far, more than 8000 pheno-lic structures are known, ranging from simple moleculesas phenolic acids to highly polymerized substances, suchas flavonoids. The most predominat phenolic phytochemi-cals in cocoa are flavonoids, which include mainly flavonols,flavanols, flavanones, isoflavones, and nonflavonoids, e.g.,phenolic acids and the less common stilbenes as shown inFig. 1 [3,4]. Catechins, anthocyanidins/anthocyanins, flavonol
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
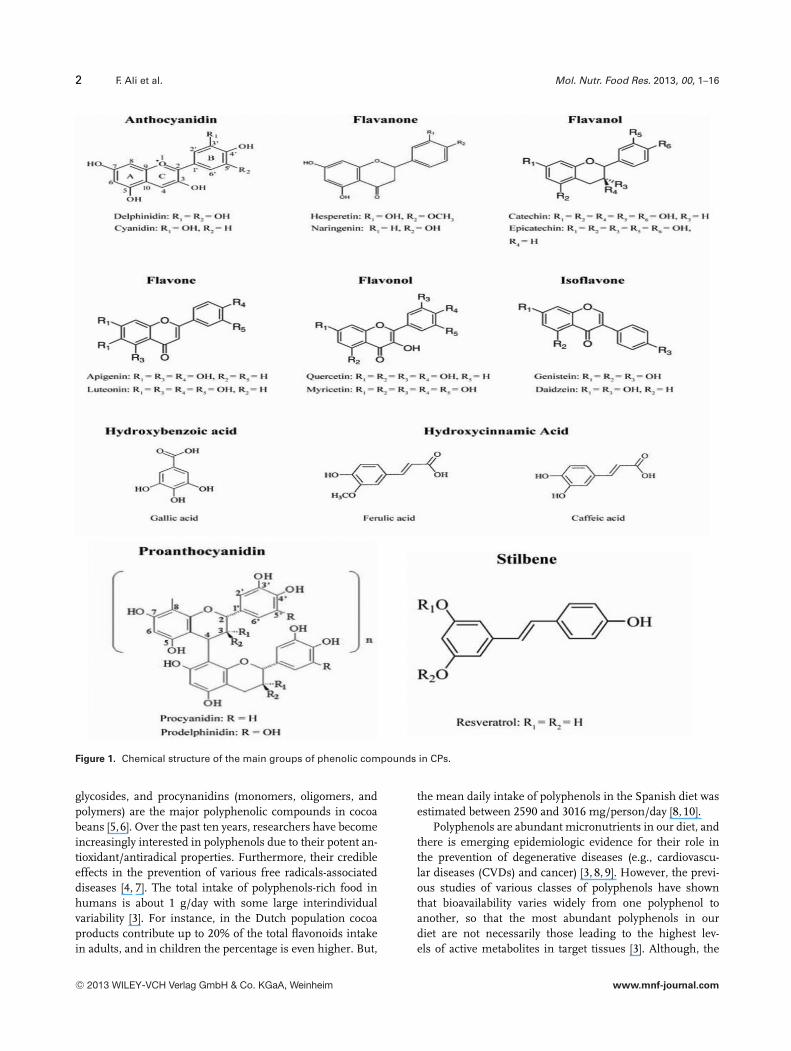
2 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
Figure 1. Chemical structure of the main groups of phenolic compounds in CPs.
glycosides, and procynanidins (monomers, oligomers, andpolymers) are the major polyphenolic compounds in cocoabeans [5,6]. Over the past ten years, researchers have becomeincreasingly interested in polyphenols due to their potent an-tioxidant/antiradical properties. Furthermore, their credibleeffects in the prevention of various free radicals-associateddiseases [4, 7]. The total intake of polyphenols-rich food inhumans is about 1 g/day with some large interindividualvariability [3]. For instance, in the Dutch population cocoaproducts contribute up to 20% of the total flavonoids intakein adults, and in children the percentage is even higher. But,
the mean daily intake of polyphenols in the Spanish diet wasestimated between 2590 and 3016 mg/person/day [8, 10].
Polyphenols are abundant micronutrients in our diet, andthere is emerging epidemiologic evidence for their role inthe prevention of degenerative diseases (e.g., cardiovascu-lar diseases (CVDs) and cancer) [3, 8, 9]. However, the previ-ous studies of various classes of polyphenols have shownthat bioavailability varies widely from one polyphenol toanother, so that the most abundant polyphenols in ourdiet are not necessarily those leading to the highest lev-els of active metabolites in target tissues [3]. Although, the
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 3
modulation effect of the food compositions on the bioavail-ability of polyphenols has not been clearly investigated. Theinteraction between polyphenolic substances and some com-ponents of food, such as binding to proteins or carbohydrates,can occur, and this interaction may affect absorption [3,9]. Inaddition, enzymes and carriers involved in polyphenol ab-sorption and metabolism may also be activated or repressedby the presence of some micronutrients or xenobiotics [9].It was reported that absorption of epicatechin from choco-late was significantly less when consumed as milk chocolate,as one of the most popular ways of cocoa powder consump-tion. The possible hypothesis is that milk proteins may bindto polyphenols from cocoa, which in turn prevents their ab-sorption in the gastrointestinal tract and thereby influencebioavailability of flavanols-rich food [8–10]. Conversely, recentresearch after this has shown that dissolved cocoa powder inmilk, did not affect the bioavailability of cocoa flavonoids inhealthy humans [11]. It must be kept in mind that one of themain aims of bioavailability studies is to determine, amongvarious classes of dietary polyphenols, which are better ab-sorbed and which lead to the production of active metabo-lites. Moreover, better knowledge of the bioavailability of thevarious polyphenols is essential to evaluate their biologicalactivity within target tissues, whatever the approach used.For these reasons, many researchers have examined the ki-netics and extent of polyphenol absorption through measur-ing plasma concentrations and urinary excretion among sub-jects after the administration of a single dose of polyphenol,supplemented as pure compound, plant extract, or wholefood/beverage [3, 10]. The polyphenol-derived metabolitesfound in blood, which result from digestive and hepatic ac-tivity, may differ from the native molecules in terms of bi-ological action. Another problem with these metabolites isthat rigorous technique to determine precise concentrationin both plasma and urine is still missing. Also many humanintervention trials with CPs seem to be underpowered. In-deed, the previous bioavailability challenges have attractedgreat interest in developing pharmacokinetic studies of CPs.Therefore, it was reported that the plasma concentrations oftotal metabolites ranged from 0 to 4 �mol/L with an intakeof 50 mg aglycone equivalents, and the relative urinary excre-tion ranged from 0.3 to 43% of the ingested dose, dependingon the polyphenol. Gallic acid and isoflavones are the well-absorbed polyphenols, followed by catechins, flavanones, andquercetin glucosides, but with different kinetics. But thesestudies found that the least well-absorbed polyphenols arethe proanthocyanidins [9, 10]. Generally, the bioavailabilityof all phenolics and their metabolites is still largely unex-plored, which demands further investigation especially withregards to its function. For example, independently of thedoses of chocolate and cocoa ingested, only 0.5% of catechinwas recovered in the free unbound form in the plasma andin the urine [10]. However, scientists later hypothesize thatsome individuals could have better absorption and tissuesdistribution than others, possibly because of particular poly-morphisms [11]. This could be the mechanism that explains
the high variability in the levels of flavonoid absorption thathave been published. Another issue to take into account whenevaluating the bioavailability of polyphenolic compounds isthe percentage of absorption in the tissues. This conditionis controlled by their hydrophilicity and binding to albumin,because they are water soluble due to the glucuronide andsulfate moieties, which makes it difficult for them to pene-trate the cell membrane to various extents; however, numer-ous studies have found these substances in such tissues asthe liver, kidneys, lungs, brain, and pancreas [8–10]. Further-more, the bioavailability of polyphenols has been reported tobe poor based on in vitro [12–15] and in vivo studies [16,17]. Aparticular focus was placed on intervention studies in the liter-ature survey to estimate the maximum concentration of CPsand their half-life time for disappearance from the circulation.Plasma concentrations of CPs were mostly in the nanomolarand low micromolar range. There are possible explanationsbehind the low concentrations of CPs in plasma (i) there isexperimental evidence that more than 90% of absorbed CPsare converted into conjugates with glucoronic acid and sul-phate [18], or (ii) polyphenols may be rapidly metabolizedby cells and, therefore, difficult to detect within the circula-tion following consumption or supplementation [21]. Mech-anisms of intestinal and hepatic conjugation (methylation,glucuronidation, sulfation) may modulate biological actions,bioavailability, and block radical scavenging capacity of CPs invivo study [3,19,20]. Therefore, careful segregation between invitro and in vivo effects is essential. Beside that, application ofnanotechnology in improving the bioavailability of uptake ofbioactive compounds to target tissue is highly recommended.Given that, any potential effect in vivo from dietary CPsshould be evaluated at the principle of low concentration ofboth conjugated and unconjugated forms. This concept mayreflect that the higher concentrations of CPs that are usuallyused in vitro experiments do not necessarily provide the sameeffect in regards to the health benefits in vivo studies. Butin vitro studies may demonstrate valuable insights into themechanisms of action [22,23]. Also, despite polyphenols hav-ing relatively poor bioavailability and low abundance, somepolyphenols possess potent biological properties in vivo stud-ies and provide potential health benefits. This may attributeto the relative high abundance and biological activity of themost effective compounds in CPs, namely, flavan-3-ols (cate-chins) [24,25]. Overall, there are several factors that may affectthe notion of bioavailability, such as the food matrix, intesti-nal absorption, excretion, intestinal and hepatic metabolism,plasma kinetics, the nature of circulating metabolites, albu-min binding, cellular uptake, accumulation in tissues, and bil-iary and urinary excretion. Therefore, it will be a big challengeto make integrating among these variables and relating themto health effects at the organ level. Thus, extensive knowl-edge of the bioavailability of polyphenols is essential if theirhealth benefits are to be understood. Nevertheless, there is re-cently experimental evidence that CPs or cocoa supplementscan act as dietary signals for direct interaction with DNA andgene expression [26]. There has, however, been relatively little
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

4 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
Obesity
GLU, FFA, TNF-α, IR
FA TG
Hypertension
Inflammation
Insulin resistance
Dyslipidemia Cardiovascular diseases
Metabolic syndromeWAT & IR
Blood
Figure 2. Simplified model illustrating the progression from obesity to CVD, overnutrition in white adipose tissue (WAT) leads to insulinresistance (IR) [(i.e., decreased free fatty acids (FFAs) and glucose (GLU) uptake into the cell) coupled with increased secretion of FFAsand proinflammatory cytokines, such as TNF-� into the blood circulation. Elevated FFAs are deposited ectopically in liver, muscle, andpancreas leading to hyperglycemia, hyperlipidemia, and systemic hyperinsulinemia. The clustering of several cardiovascular risk factorsfor abdominal obesity, dyslipidemia, glucose intolerance, and hypertension in the same individual is called the metabolic syndrome. TG =triglyceride.
study of the molecular mechanisms underlying the pro-tective effects of CPs on energy metabolism and relevantgene expression profiles. Thus, the possible mechanisms in-volved into the cellular uptake of CPs as well as its cellu-lar concentrations and distribution require to be extensivelyclarified.
1.2 Obesity-mediated metabolic diseases and
inflammation
Obesity can be defined as excess accumulation of body fatfrom an energy imbalance due to excessive energy storage andinsufficient energy expenditure [27]. Obesity- and metabolic-related diseases (e.g., type 2 diabetes, dyslipidemia, CVDs,and hypertension) are the most prevalent nutrition-related is-sues throughout the world (Fig. 2) [28]. An emerging meritof obesity and associated diseases is their linkage with in-flammation that begins in adipose tissue and ultimately be-comes systematic [29]. In recent years, there is experimentalevidence suggesting that CPs may possess the antiobese af-fect and their influence on body weight, body fat content,serum lipid and lipoprotein levels, and the expression ofenzymes regulating lipid metabolism [30–32]. On the otherhand, a group of nuclear hormone receptors (peroxisomeproliferator-activated receptors (PPARs)) have also been im-plicated. The PPARs are a group of ligand-activated tran-scription factors that govern numerous biological processes,primarily energy metabolism and inflammation [33]. Syn-thetic agonists for these receptors are used in the treatmentof type 2 diabetes mellitus and dyslipidemia [34]. More re-cently, Jonathan et al. found that flavonoid acts as an agonistof PPAR-� and a partial agonist of Liver X receptor (LXR)-�, strongly suggesting that the flavonoid may be targetingtranscriptional regulation of metabolism through nuclear re-ceptors (NRs) [35]. Thus, the present review critically sum-marizes the available studies over almost the last decade andhighlights the possible mechanisms involved in the responseto CPs consumption or supplementation. In particular, thepossible mechanisms underlying the beneficial effects of CPs
on obesity-mediated metabolic diseases and inflammation areproposed based on literature reports.
2 Potential health benefits of CPs
2.1 Lipid peroxidation and profiles in obesity-related
diseases
Cocoa procyanidins and flavanols have strong antioxidantcapacities in vitro and in vivo studies [4]. First, evidence hascome from an experiment in which isolated polyphenols fromcommercial cocoa delayed the oxidation of LDL [36]. Otherstudies reported an inhibition in the production of reactiveoxygen species (ROS) in activated leukocytes and reductionof ultraviolet-induced DNA oxidation [37, 38]. Although CPsdo not appear to be existent in the blood circulation at highenough concentrations in order to contribute to total antiox-idant activity, a number of studies have shown that CPs mayinduce the upregulation of endogenous antioxidant enzymesin vivo and thereby exert an indirect antioxidant activity.For example, flavanol-rich cocoa decreases the plasma levelof F2-isoprostanes, which act as markers of in vivo lipidperoxidation, and, therefore, increases total antioxidant activ-ity [39, 40]. In addition, several studies have been conductedto determine the role of CPs in lipid oxidation. These studieshave shown that CPs may antagonize the oxidation of LDL,which is a key event in atherosclerosis [41–44]. For instance,recent studies in hypercholesterolemic rabbits treated withdiet containing 10% cocoa powder for 24 wk suggest thatCPs decrease the formation of atherosclerotic lesions in theaorta due to the blocking of LDL oxidation [45]. However,other studies have reported that CPs have no effect onbiomarkers of lipid peroxidation in liver and heart of ratstreated with flavanol and procyanidin-rich cocoa for 2 wk [46].Moreover, numerous studies in humans indicate that CPsdecrease LDL oxidation and plasma lipid peroxdicationbiomarkers, such as F2-isoprostane and thiobarbituric acidreactive substances, following CPs consumption amongsubjects with different numbers and age range [47–49]. On
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 5
the other hand, studies in hamsters treated with 1 g of cocoapowder for 10 days show an increase in HDL along witha decrease in LDL and triglycerides levels [50]. In addition,cocoa procyanidins markedly decreased plasma cholesteroland increased steroid secretion in rats fed a high cholesteroldiet [51]. A decrease in lipid profiles, 8-isoprostane, as wellas in blood glucose levels has been reported in obese diabeticrats supplemented with cocoa extract (600 mg/kg bodyweight/day) for 4 wk [52]. Furthermore, Zucker fatty ratstreating a daily dose of quercetin (10 mg/kg body weight) for10 wk had lower blood lipid profiles and improved insulinsensitivity compared with placebo controls [53].
An increase in plasma HDL cholesterol [54, 55], whereasa decrease in plasma triglyceride [56–58], in response toCPs has been shown. To a greater extent, it was indicatedthat genistein supplementation could reduce lipid profilesof obesity phenotype and metabolic abnormalities by acti-vation of the expression of genes involved in fatty acid oxi-dation, and thus improved lipid profiles and attenuated theincreases in body weight in C57BL/6J male mice. C57BL/6J(n = 10/group) were fed a low fat diet, a high fat diet, or a highfat diet supplemented with 2 g/kg genistein for 12 wk [59,60].Changes in the expression of genes involved in fatty acidcatabolism provide interesting information about the genesassociated with the health benefits of genistein as well as themechanisms underlying the development and maintenanceof the obesity phenotype in vivo.
2.2 Obesity-associated CVDs
Inflammatory mediators released from excess white adiposetissue are the major cause of CVDs due to endothelialdysfunction and plague formation [61]. The consumption offlavanoids in cocoa and other cocoa products have been asso-ciated with reduction in the risk of CVDs [62]. For example,supplementation with quercetin 64 mg/kg body weight/dayfor 10 and 20 wk decreased inflammation and endothelialdysfunction in a mouse model of atherosclerosis [63]. Also,the consumption of flavonoid-rich beverages (one cup perday for 6 wk) decreased inflammatory cytokines and systolicblood pressure and improved blood profiles in men andpostmenopausal women [64]. In vitro studies using humanumbilical endothelial cells treated with epicatechin andflavanol metabolites mixture (0–100 �g/mL) versus controlindicate that CPs decrease arginose-2 mRNA expression andits activity levels, which may lead to a higher availability ofthe vaso-relaxing molecule nitric oxide (NO) in the vascularwall [65]. This result is in agreement with the study ofKarim et al., using cocoa procyanidins isolated rabbit aorticrings in which endothelium-dependent relaxation and anincrease in inducible nitric oxide synthase (iNOS) havebeen detected [66]. The polymeric procynidins found toactivate Ca+2-dependent iNOS activity, whereas catechinsfrom monomers to trimers exhibited no such activity [66].Moreover, cocoa procynidins (0–100 �g/mL) suppress
metalloproteinase-2 expression and activation and decreasedvascular smooth muscle cells invasion and migration. Thiseffect may reflect the anti-atherogenesis effects of cocoa [67].Surprisingly, CPs have been significantly shown to reduceangiotensin-converting enzyme (ACE) activity, which mayresult in a decrease of blood pressure. ACE is also inhibited byepicatachin. ACE increases blood pressure by causing bloodvessels constriction through converting angiotensin I toangiotensin II, which constricts the vessels [68]. As observedfor the purified ACE enzyme from rabbit aortic lung, ACEactivity in kidney membrane was inhibited by 100 �mol/L ofdimeric and hexameric epicatechin (0–500 �mol/L).
Stimulated macrophages through inflammation stimulican release large amounts of NO from L-arginine by the im-pact of iNOS. Chronic inflammation is often associated withoverproduction of NO by macrophages [69]. The transcrip-tion factor NF-�B, and activated protein-1 (AP-1), mediatesthe expression of iNOS and pro-inflammation cytokines [70].This mechanism may partly explain beneficial vascular effectsof CPs by (i) decreasing NO production by inhibiting iNOSgene expression due to NF-�B- and AP-1-dependent signaltransduction pathways [69, 70], and (ii) suppressing arginaseactivity. Flavonols-rich cocoa has been reported to lower renalarginase activity in rats supplemented with 40 g cocoa powderper kilogram diet for 4 wk and vascular arginase activity in en-dothelial cells [71]. Arginase contests with endothelial iNOSfor L-arginine as the substrate. Thus, suppression of arginaseactivity can be associated with an augmented endothelial NOproduction. Increased NO production has been reported toimprove endothelium-dependent relaxations, and thereby en-dothelial function [71]. The improvement of endothelial func-tion due the effect of dietary CPs may result in a decrease insystolic and diastolic blood pressure. However, other studiesdid not show any beneficial effect of CPs on blood pressure[72, 73]. Recent study investigated the effect of flavonol-richcocoa on vascular function in mediated diabetic patients in adouble-masked, randomized, controlled trial [74]. Significantimprovements in flow-mediated dilation, in response to theeffect of CPs on endothelial function were observed. In con-trast, other study for a 6-wk period, 48 g flavonol-containingcocoa did not improve vascular function in 34 patients (aged18–74) with coronary artery disease [75]. Moreover, currentlyFlammer and others reported that 2 h after consumption of40 g of dark chocolate containing 70% cocoa improved coro-nary vascular function and decreased platelet adhesion [76].Finally, the health benefits of natural polypheonls from theplant kingdom have been proven in reducing risk factors forCVD, such as body weight gain, oxidative stress/LDL oxida-tion, hypertension, hyperlipidemia, inflammation, and en-dothelial dysfunction [77, 78]. Further studies showed thatpolyphenols-rich cocoa extract possess various pharmacologi-cal actions, including antioxidant effects, antihyperlipidemic,antidiabetic actions, anti-atherosclerotic, and antihyperten-sive, anticancer actions [79–81]. Therefore, supplementationwith cocoa rich in polyphenols may be a useful dietary strategyfor the attenuation of CVD associated with obesity.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

6 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
2.3 Obesity-associated diabetes
Obesity and related chronic inflammation causes insulinresistance or its consequence, type 2 diabetes. For exam-ple, more than 80% of diabetic patients are overweight orobese [82]. This group of obesity-related metabolic diseasesis also known as the metabolic syndrome (Fig. 2). Insulinresistance is a prediabetic state characterized by diminishedtissue sensitivity to endogenous insulin, leading to chronicelevation in blood glucose level coupled with increased se-cretion of free fatty acids into the circulation from adiposetissue (Fig. 2) [83]. Chronic elevation in glucose and freefatty acids lead to the development of noninsulin-dependentdiabetes (type 2) and eventually pancreatic beta cell failure,resulting in insulin-dependent diabetes (type 1) [83]. Aminet al. [84] and Ruzaidi et al. [85] demonstrated that cocoa ex-tract containing 190–286 mg total polyphenol in the dosageof 1–3% to n = 10/group of normal and diabetic rats for 4wk by force feeding resulted in significantly reduced bloodglucose and lipid profiles of diabetic-induced rats comparedwith nontreated group; improved insulin sensitivity in vitrostudy using BRIN-BD11 cell lines was also reported. More-over, dietary supplementations with cocoa liquor procyani-dins prevent the development of hyperglycemia in diabeticobese mice [86]. Consumption of resveratrol (0.04% in thediet w/w for 48 wk, or 2.5–400 mg/kg body weight/day for16 wk) improved glucose, insulin levels, and lipid homeosta-sis in C57BL/6J obese mice fed high fat diet [87–89]. Overall,results suggest that polyphenol-rich cocoa products are goodcandidates for diabetic management of type 2 diabetes be-cause of their abundance of polyphenol content. However,clinical trials probing the effect of cocoa consumption on in-sulin resistance are still limited. Grassi et al. [90, 91] studiedthe effect of high-polyphenol dark chocolate administration(100 g) for 15 days in hypertensive subjects. A significantincrease in insulin sensitivity has been observed. A home-ostasis model assessment of insulin resistance is used tomeasure insulin sensitivity using fasting insulin and glu-cose concentrations [112]. This model was significantly de-creased in CPs-treated groups compared to the high fat dietgroup of KKAy mice after 7 wk, indicating for the reduc-tion of hyperglycemia and hyperinsulinemia. Subsequentstudies in vitro (0.01–10 �M) and in vivo (2.5–400 mg/kgbody weight/day) have reported that resveratrol activated sir-tuin1 (SIRT1), and insulin sensitivity was improved. Consis-tent with these data, quercetin supplementation (12.5 and25 mg/kg body weight/day) for 7 days increased SIRT1 andPPAR-� coactivator-1 (PGC-1�) expression and mitochon-drial biogenesis in male imprinting control region (ICR)mice [92, 93].
2.4 Oxidative stress in obesity-related disease
Free radicals molecules, such as ROS and nitrogen oxygenspecies (NOS), can be formed by enzyme systems, suchas iNOS, nicotinamide adenine di-nucleatide phosphate oxi-
dase, and from transition metals [94]. Oxidative stress is of-ten defined as an imbalance between the production of ROSand endogenous antioxidant defense system. Excess produc-tion of ROS is associated with metabolic disorders, includ-ing diabetes, atherosclerosis, cancer, and chronic inflamma-tion [95, 96]. Subsequent studies confirmed that obesity isassociated with increased chronic systemic oxidative stress.Oxidative stress also plays a pivotal role in the pathogenesisof obesity-mediated diseases through its effects on vascu-lar biology. Physiologically, ROS can modulate cellular func-tion, receptor signals, and immune responses on vascularcells [97, 98].
In pathophysiological status, ROS may contribute in thevascular dysfunction and inflammatory reactions throughoxidative damage caused by decreased NO bioavailability,impaired endothelium-dependent vasodilatation, endothelialcell growth, and migration [99, 100]. ROS or NOS can stim-ulate redox-sensitive kinases, such as apoptosis signal reg-ulating kinase 1, which induce NF-�B, and AP-1 transcrip-tion factors via activation mitogen-activated protein kinases(MAPKs), thereby activating inflammatory gene expression[101]. Also, recent studies have suggest that phenolic com-pounds have the capacity to increase the levels of gene andprotein expression of endogenous antioxidant enzymes, suchas �-glutamate-cysteine ligase catalytic subunit, superoxidedismutase, glutathione peroxidase, and heme oxygenase-1through activation of the transcription factor nuclear factor-erythroid 2 (as given in Fig. 4) [101, 102]. This effect mayrelatively reveal the possible mechanism by which CPs can in-directly induce the protective effects against ROS. Consistentwith antioxidant activity in vivo, pretreatment with the antho-cyanin cyanidin3-glucoside (10–40 �M) of murine adipocyte(3T3-L1 cells) exposed to H2O2 decreased ROS productioncompared with controls [103].
Quercetin and rutin (1–25 �M) blocked oxidized LDL viamodification of Jenus kinase and activation of the transcrip-tion signaling pathway. However, long-term treatment withhigh levels of quercetin (100 �M) showed signs of cytotoxicityand decreased cell viability [104]. Currently, Noe et al. havereported that the cocoa polyphenolic extract may prevent orinhibit oxidative stress associated with obesity by reducing theexpression of genes involved in the development of oxidativestress-mediated diseases, such as inflammation and CVD inhuman colon adenocarcinoma Caco-2 cells using functionalgenomics analysis [105]. In this study, Caco-2 cells (5 × 106)were incubated for 24 h with either 100 �mol/L (29.03 g/L)epicatechin or 0.2155 g/L of catechin equivalents of cocoapolyphenolic extract, both in DMSO (final concentration ofDMSO in the medium <1% v/v).
These data suggest that CPs or their metabolites have thecapacity to protect cells from oxidative damage by either neu-tralizing ROS/NOS, decreasing the activity of enzymes, suchas iNOS or nicotinamide adenine di-nucleatide phosphateoxidase, that produce free radicals or activation transcrip-tion factors, such as Nrf2, which induce the transcription ofendogenous antioxidant enzyme genes, such as superoxide
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 7
dismutase, glutathione peroxidase, and heme oxygenase-1.Hence, CPs have a potent capacity to reduce ROS and otherfree radicals, thereby preventing their activation of oxidativestress and inflammation.
3 Molecular mechanisms of theantiobesity-mediated diseases effectsby CPs
3.1 Metabolic diseases associated with obesity
The peroxisome proloferator-activated receptors (i.e., PPAR-�, PPAR-�/�, and PPAR-�) are ligand-activated transcrip-tion factors, which play an important role in the regula-tion of lipid, glucose metabolism, and even inflammatoryresponse pathway [106, 107]. PPARs belong to the family ofnuclear hormone receptors, which are activated by a varietyof compounds derived from food [108]. PPARs are inter-esting target from the perspective of pharmacological treat-ment of the conditions that contribute to the metabolic syn-drome, such as insulin resistance [109]. According to Guoand Tabrishi [110], the upregulating of PPAR-� in adiposetissue has shown to exhibit an increase in glucose trans-porter 4 (GLUT4) expression, pivotal for the enhanced glu-cose uptake and for the translocation of GLUT4 to the cellsurface, thus reduces hyperglycemia and hyperinsulinemia.Also, Chawla et al. [108] stated that increase in PPAR-� ex-pression in human skeletal muscle cell culture correlates withenhanced insulin-stimulated glucose uptake into the musclemediated by translocation of GLUT4 toward the cell mem-brane. In this context, Maeda et al. have found that PPAR-� ligands increase expression and plasma concentration ofadiponectin, a protein hormone, which exerts antiobesity andanti-inflammatory effects [111]. It has been recently demon-strated that insulin resistance can be developed by reducedadiponectin secretion and increased tumor necrosis factor-�(TNF-�) secretion in mice [112]. Interestingly, phenolic com-pounds reduced lipid deposition, hyperglycemia, and hyper-insulinemia by increasing adiponectin secretion and expres-sion in obese diabetic mice [112, 113]. Furthermore, Sujonget al. [60] have reported that genistein-derived isoflavones en-hances the expression of fatty acid oxidation genes throughthe activation of PPAR-� in HepG2 cells, when assayed byreal-time RT-PCR as well as Western blotting analysis. In-terestingly, genistein induced expression of PPAR-� at bothmRNA and protein level. Suggesting that genistein holdsgreat promise for therapeutic applications to lipid abnormal-ities, such as obesity-associated diseases in humans. Recentresearch has demonstrated that flavonoids-rich grape acti-vated PPARs, but suppressed LXRs in human and rat hepato-cytes [35]. In this study, primary hepatocytes were stimulatedwith (126–400 �M) flavonoid naringenin for 24 h in standardculture medium. The flavonoid has demonstrated to activatePPAR response element, while suppressing LXR-� responseelement. Furthermore, activated PPARs led to induction of
PPAR-�-regulated fatty acid oxidation genes. On the otherhand, reduced LXRs led to inhibition of LXR-�-regulated li-pogenesis genes and sterol regulatory elevated binding pro-tein (SREBP1). SREBP1 plays a role in cholesterol synthesis.These findings suggest a shift from lipogenesis and choles-terol synthesis into lipolysis [35]. These results are consis-tent with recent study by Sujong et al., who also use DNAmicroarray technology for analyzing the changes in hepatictranscriptional profiles, where upregulation in the expressionof genes involved in fatty acid catabolism in mice with diet-induced obesity supplemented with 2 g/kg genistein for 12wk has been observed, compared with nontreated group [59].Both studies in vivo and in vitro obviously provided a pictureof the regulatory action of CPs as an activator and a potentialligand for PPAR-� in fatty acid catabolism and potency useof polyphenols as lipid-lowering agent. Polyphenol’s myriadeffect suggests that flavonoid may be targeting transcriptionalregulation of metabolism through NRs, which play a criticalrole in the regulation of lipid metabolism. However, more re-search is needed on the regulation of PPARs by CPs to deter-mine which specific procyanidines and flavonols are involved.Recently, it has been reported that polyphenols administra-tion significantly increased mRNA and protein expressionsof ACO, CPT1 (�-oxidation enzymes), PPAR-� and PPAR-�(NRs), and UCP3 (uncoupling protein) compared to the highfat group using KKAy mice as a model of obese type 2 diabetesto induce sever obesity and diabetes. KKAy mice (n = 10 pergroup) fed either normal diet, high fat diet, or high fat dietwith additional (2.5–5%) polyphenols for 7 wk. Activation ofPPAR-� and PPAR-� expression is known to increase expres-sion of CPT1, ACO, and UCP3 to activate energy expenditure,subsequently resulting in antiobesity actions. This suggeststhat polyphenols act on skeletal muscle and liver to increaseenergy expenditure [112]. In contrast, polyphenols decreasedmRNA expression of ACC and fatty acid synthetase (FAS), therate-limiting enzymes of fatty acid synthesis in the liver, andmRNA expression of SREBP-1c. Furthermore, polyphenolssupplementation decreased the mRNA expression of FAS,PPAR-�, and lipoprotein lipase (LPL), which are related tofat intake by the liver. Reduced expressions of FAS, PPAR-�, and lipoprotein lipase in the liver suppress the onset ofobesity and fatty liver. These findings indicate that the an-tiobesity actions are due to increased expression of energyexpenditure-related genes in skeletal muscle and liver anddecreased fatty acid synthesis and fat intake in the liver asshown in Fig. 3 [29, 103]. In this study, it was also foundthat polyphenols-rich food increased mRNA expression ofadiponectin and decreased mRNA expression of TNF-� inwhite adipose tissue. In addition, this administration signif-icantly increased the mRNA expression of PPAR-�, whichfacilitates the expression of adiponectin and GLUT4 [112].
In general, overconsumption of high fat or carbohy-drate diets is considered as a major reason to increase thesize of organs or adipose tissues, body weight, and even-tually induce obesity. The obesity induced by high caloricdiets is associated with increased insulin secretion and fat
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

8 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
LXRαPPARα
L
Anti-obesity action
Insulin sensitivity Glucose uptakeAdiponectin
Fatty acid synthesis Triglyceride synthesisCholesterol synthesis
Fatty acid oxidationEnergy expenditureMetabolic genes expression
Cocoa Polyphenols
PPARγ
ipogenesis genesAdiponectin, GLUT4 genesLipolysis, UCPs genes
Figure 3. Potential molecular mecha-nisms underlying the antiobesity ef-fect by CPs. CPs may prevent vis-ceral fat deposition and attenuateobesity-mediated metabolic diseasesby transcriptional activation of PPAR-�, uncoupling proteins (UCPs) andinhibition of LXR-� target genes.CPs administration increased the ex-pression of PPAR-�, which facilitatesthe expression of adiponectin, andGLUT4 (i.e., one of the glucose trans-porters), resulting in the reductionof lipogenesis, induction of lipolysis,and adiponectin secretion, which inturn reduce lipid deposition and in-sulin resistance in tissues and circu-lation, and thereby alleviated obesity.Arrows indicate positive inputs (acti-vation), whereas perpendicular linesshow negative inputs (inhibition).
deposition as triacylglycerol, which in turn inhibits lipoly-sis or fat oxidation. In current research, Matsui et al. [32]found that visceral white adipose tissue weight tended to belower and serum triacylglycerol levels were decreased in rats(n = 10 per group) fed a cocoa containing 12.5% w/w co-coa powder diet than in those fed a cocoa-free diet for 21days. DNA microarray analysis showed that cocoa ingestionalso decreased the expression of genes involved in fatty acidsynthesis and transport systems, while it upregulated the ex-pression of genes for fatty acid �-oxidation and promote partof the thermogenesis mechanism in liver and adipose tisssue.Another study by Tomaru et al. [86] investigated that CPs havethe ability to prevent the development of diabetes in genet-ically inherited diabetics rats. These findings provide inter-esting information about the transcription factors associatedwith the beneficial effects of CPs as well as the mechanismsunderlying the development and conservation of the obesityphenotype. Taken together, CPs increase the expression ofPPARs, adiponectin, and decrease the expression of LXRs,thereby prevent obesity-mediated metabolic diseases, such asdyslipidemia, type 2 diabetes mellitus and CVDs. The pos-sible molecular mechanisms by which phenolic compoundsin cocoa antagonize obesity-mediated metabolic diseases aresummarized in Fig. 3. These mechanisms mainly depend onthe suppression or activation of transcription factors involvedin lipid metabolism and fat accumulation.
3.2 Inflammation associated with obesity
Inflammation is a protective tissue response induced by tis-sue injury or infection. The injured tissues release inflamma-tory cytokines or mediators (e.g., TNF-�, IL-1, IL-6, IL-8, andmonocyte chemoattractant protein (MCP-1)). In a complexsignal transduction pathway, these mediators can modulate
other inflammatory cytokines from activated leukocytes, lead-ing to upregulate cellular adhesion molecules in inflamed tis-sues [112]. The central regulator of the inflammatory processis NF-�B transcription factor [26,114]. CPs may act as inflam-mation modulatory agents by various mechanisms, includingdownregulation of NF-�B, and thereby reducing chronic in-flammation (Fig. 4) [115]. Further studies by using culturedcells suggest that CPs have anti-inflammatory properties byblocking the production of pro-inflammatory cytokines, suchas TNF-�, and activation of anti-inflammatory cytokines, e.g.,IL-10 [116, 117]. Several studies reported that flavonols andanthocyanins regulate inflammation associated with obesity.For example, obese patients plus type 2 diabetes have ele-vated levels of inflammatory signaling and cytokine produc-tion [118]. Supplementation of C57BL/6J mice fed a high fatdiet with anthyocyanine-rich powder for 8 wk in the diet w/whad lower expression of inflammation-related genes [119].
In this context, C57BL/6J mice fed a high fat diet treatedwith 0.8% quercetin in the diet w/w for 8 wk character-ized by lower plasma levels of IFN-�, IL-1�, compared withcontrols [120]. Moreover, quercetin (3–60 �M) suppressedTNF-�-related inflammation and insulin resistance in pri-mary human adipocytes by blocking activation of downstreamextracellular signal-related kinase (ERK), c-Jun-NH2 termi-nal kinase (JNK), NF-�B, and AP-1 signaling pathway andnegative modulators of insulin signaling pathway (i.e., phos-phorylation of serine residue on insulin receptor substrateand protein tyrosine phosphate) [121], and which subse-quently suppress insulin signaling substantial for insulin-dependent GLUT4 translocation to the plasma membrane[121]. Another study by Mao et al. indicates that the CPs(monomer–tetramer) consistently decreased IL-1� expres-sion in blood mononuclear cells, whereas the larger oligomers(monomer–tetramer) increased its expression [122]. CPshave been also exhibited inhibitory effect on 5-lipoxygenase
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 9
Figure 4. Summary of mechanisms through which CPs reduce metabolic diseases and chronic inflammation associated with obesity.CPs diffuses into the cell and may inhibit MAPKs, leading to block activation of inflammatory transcription factors, such as NF-�B andAP-1. Together, these signals repress inflammatory gene expression of many proinflammatory mediators (e.g., TNF-�, IL-6, IL-8, IL-1, andMCP-1, NO). CPS expected also to enhance Nrf2 that induce the transcription of antioxidant enzymes, such as glutathione peroxidase(GPx), superoxide dismutase (SOD), and heme oxygenase-1 (HO-1), thereby blocking ROS, NOS production, and attenuating oxidativestress. Finally, CPs may prevent lipid deposition and insulin resistance associated with obesity by activation PPARs-regulated and insulin-dependent GLUT4 genes or inhibition LXRs-regulated lipogenesis genes, thereby decreased free fatty acids (FFA), triglycerides (TG),and cholesterol synthesis and eventually prevent ectopic lipid accumulation and insulin resistance (IR). Arrows indicate positive inputs(activation), whereas perpendicular lines show negative inputs (inhibition).
activity, which plays an important role in the inflammationprocess [123]. Ki et al. found that CPs inhibit expressionof cyclooxygenase-2 and activation of NF-�B and MAPKs in12-O-tetradecanoylphorbol-13-acetate-stimulated mouse skin(40 or 200 mg/kg). This author also reported that CPs inhib-ited the catalytic activity of ERK 1/2 [124].
Furthermore, randomized clinical trials with pro-anthocyanidine-rich cocoa powder (40 g cocoa powder in 500mL skim milk/day for 4 wk) [125] and flavonol-rich bilberryjuice (330 mL/day for 4 wk) [126] caused reduced plasmamarkers of inflammation in adults with high CVD risk whoconsumed products rich in polyphenols. Consumption offlavanol-3-ols (catechins)-rich extract (0.32 mg/g diet/day for19 wk) attenuated inflammatory biomarkers in white adi-pose tissue, liver, and blood circulation in high fat diet, obesemale Zucker fatty rats [127]. Furthermore, quercetin (3–60�M) inhibited carcinogen-mediated inflammation by directlybinding to MEK (also known as MAPK), which restrains acti-vation of inflammatory MEK/MAPK signaling cascade in JB6cells [128]. MAPK regulates the activities of several transcrip-tion factors in the nucleus of the cell, such as NF-�B. Inter-estingly, kaempferol (1–3 �M) suppressed TNF-�-mediated
inflammation via blocking nuclear translocation and DNAbinding of NF-�B and AP-1 in respiratory epithelial cells [129].Furthermore, sirtuins (SIRT), are involved in protein acetyla-tion/deacetylation. SIRT has been reported to play role in theregulation of chromatin structure, gene expression, and en-ergy metabolism [130]. In vitro (0.01–10 �M) and in vivo (2.5–400 mg/kg body weight/day) analysis found that resveratrol-activated SIRT1), thereby decreasing inflammatory gene ex-pression [131–133]. Significant improvements in biochemi-cal and molecular parameters of inflammation markers inresponse to cocoa-rich polyphenols intervention or ingestionhave been observed, strongly suggesting anti-inflammationeffects of cocoa.
4 Discussion
In this section, further explanations in the crucial role ofMAPK and histone deacetylase activity in the regulatory ef-fects of CPs are discussed, with special emphasis on en-hancing energy expenditure, activation of glucose, and lipidmetabolism as well as inflammatory mediators throughMAPK pathway modulation.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

10 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
4.1 Ameliorating insulin sensitivity, glucose uptake,
and adiponectin secretion via activation MAPK
pathway
Collectively, high caloric diets increase body weight and sizesof organ or adipose tissue, which lead to some chronic dis-eases. In particular, endogenous and exogenous factors thatenhance adipocyte hypertrophy have the capacity to increasewhite adipose tissue mass and the development of obesity.It is evident that adipocytes are not just energy-storing cellsbut also secret a variety of adipocytokines, such as leptin,TNF-�, and adiponectin. Hypertrophy of visceral adiposetissue accompanies excessive secretion of some of theseadipocytokines and induces insulin resistance, which causestype 2 diabetes mellitus, dyslipidemia, hypertension, andCVDs (Fig. 2) [89].
Treatment with polyphenol-rich cocoa extract showedupregulation of those genes for beta-oxidation, energyexpenditure-related genes and downregulation of those forfatty acid synthesis and fat intake in the polyphenol-treatedgroups. In addition, the changes in messenger RNA levelsfor PPAR-�, -�/�-regulated lipolysis genes, LXR-�-regulatedlipgenesis genes, and induced PPAR-� transcriptional activityupon incubation with the cocoa polyphenolic extract, stronglysuggest that CPs prevented and ameliorated diet-induced obe-sity via the regulation of lipid metabolism-related gene ex-pression. Furthermore, CPs antagonize hyperglycemia andhyperinsulinemia, not simply through alleviated obesity, butthrough increased adiponectin secretion and suppressedTNF-� secretion in white adipose tissue, and increasedGLUT4 expression in skeletal muscle (Figs. 3 and 4) [59, 60].Many reports demonstrated that the modulation in the abovegenes and proteins expression involved in prevention obesityand its promoted diseases are mediated, principally by acti-vating the key metabolic regulator AMPK pathway. AMPKplays an important role as an intracellular energy sensor thatregulates glucose and lipid metabolism and it is consideredas one of the key targets for controlling and treating obe-sity and its related diseases [137]. In fact, it has been shownthat activation of AMPK suppresses many anabolic processes,including fatty acid synthesis and gluconeogenesis, and ac-tivates several catabolic processes, such as fatty acid oxida-tion, insulin signaling, and glucose uptake. AMPK is alsomodulated by the adipokines, e.g., leptin and adiponectin,important controllers of energy metabolism [139, 140].Published data showed that CPs significantly increase plasmaadiponectin levels (Fig. 3), an adipokine that is known to ac-tivate AMPK upon binding to adiponectin receptor R1. Con-sistent with these data, modulation of AMPK pathway by CPsin vitro and in vivo was investigated by analyzing key pro-teins of PI3K/AKT and AMPK, as well as the levels of theGLUT4, adipokines level, and the hepatic glucose produc-tion [137, 138]. It is known that insulin promotes transloca-tion of GLUT4 from intracellular compartment to the plasmamembrane. Then, GLUT4 enhances a huge amount of glu-cose into the cells to maintain glucose homeostasis. The two
main molecules that regulate translocation of GLUT4 andadipokines secretion are PI3K and AMPK [138, 139]. Severalstudies have demonstrated that activation of AMPK by CPsis involved in the promotion of insulin signaling pathway,which is potent inducer of GLUT4 translocation to the plasmamembrane in peripheral tissues, thus preventing hyper-glycemia and insulin resistance. The activation of AMPK alsopromotes the gene and protein expression of UCPs and PGC-1� in skeletal muscle, adipose tissue, and liver, which leads toprevent obesity by increasing energy expenditure and/or ther-mogenesis. UCPs comprise a family of proton transporterslocated in the mitochondrial inner membrane and enhancethermogenesis and energy expenditure. UCP-1 is mainly in-volved in thermogenesis, while UCP-2 and UCP-3 are alsoinvolved in energy metabolism and obesity. Thus, intake ofCPs likely increases thermogenesis and energy expenditure,which are involved in the reduction of white adipose tissueweight [138]. Interestingly, CPs reported to modulate the ex-pression of phosphoenol pyruvate carboxykinase, a key pro-tein involved in the gluconeogenesis, leading to a diminishedglucose production. Overall, intake of CPs can prevent highfat diet-induced hyperglycemia, hyperinsulinemia, insulin re-sistance, and obesity (Figs. 3 and 4) by activating the phospho-rylation of AMPK in skeletal muscle, liver, and adipose tissue,followed by the activation of downstream events that suppresshyperglycemia and fat deposition. Since, binding of CP to cellsurface receptors initiates a sequential pathway of phospho-rylation reactions involving phosphatidylinositol in cell mem-branes and cytosolic protein [137]. To our knowledge, activa-tion of AMPK, translocation of GLUT4, and upregulation ofUCPs expression are involved in the molecular mechanismsby which CPs prevent obesity and its related diseases.
4.2 Blocking pro-inflammatory mediators via
activation MAPK pathway
The three major subfamilies of MAPKs that are mainly ex-pressed in the immune system is ERK, JNK, and p38. All threeMAPKs are phosphorylated on the threonine and tyrosineresidues. Activation of toll-like receptor or TNF receptor mayactivate the expression of cytokines, chemokines, and otherinflammatory mediators in less than 1 h. Rapid transduc-tion of the signal from TNF receptor to the MAPK is achievedthrough the sequential activation of upstream MAPK kinases.Phosphorylation pathways are substantial for such a rapid re-sponse and both involve activating the latent transcriptionfactors, such as NF-�B and AP-1 [89, 133]. Both pathwayssynergize in inflammatory gene expression through the coor-dinate binding of the transcription factors to the NF-�B andAP-1 sites found together in the promoters of, e.g., TNF-�,IL-8, and many other genes that are up- or downregulated inresponse to toll-like receptor ligation. After proofing the anti-inflammatory effects of CPs both in vitro and in vivo, we needto gain insight into the molecular mechanism that underliesCPs anti-inflammatory effects. For this cause, we assessed
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 11
if the anti-inflammatory effect of CPs could be the result ofinhibiting the MAPK and NF-�B signaling pathways, whichare the most important pro-inflammatory pathways, that reg-ulate the production of many pro-inflammatory mediators.In this sense, the possible mechanism by which CPs preventchronic inflammation and insulin resistance-associated obe-sity might by blocking activation of MAPKs, NF-�B, and AP-1and their downstream, ERK and JNK [67,124]. This situationwould reduce the activation of inflammatory transcription fac-tors NF-�B and AP-1, which are robust inducers of inflam-matory gene expression (e.g., TNF-�, IL-6, IL-8, IL-1�, andMCP-1) and negative modulators of insulin signaling (e.g.,phosphorylation of serine residue 307 on insulin receptorsubstrate and protein tyrosine phosphatase 1B), and whichsuppress insulin signaling necessary for insulin-dependentGLUT4 translocation to the plasma membrane (Fig. 4).
4.3 Inducing metabolic or suppressing inflammatory
gene expression via activation histone
deacetylase activity
As previously mentioned, CPs may be useful in prevent-ing inflammatory-mediated insulin resistance and other re-lated metabolic diseases simultaneously by activating classIII histone deacetylase activity (sirtuins). Polyphenols fromcocoa have been reported to suppress inflammatory or in-duce metabolic gene expression-associated obesity by acti-vating SIRT1. This intercorrelation may be attributed to (i)deacetylate/inactivate NF-�B, resulting in the suppression ofinflammatory gene expression, and (ii) deacetylate/activatePGC-1�, resulting in the activation of PPARs. Activationof PPAR-� and/or PPAR-�/� transcription factors increasesoxidative phosphorylation, metabolic gene expression, mito-chondrial biogenesis, and energy expenditure. In the samecontext, activation of PPAR-� may antagonize NF-�B tran-scriptional activation of inflammatory genes, thereby attenu-ating inflammation associated with obesity. However, moreresearch is needed on the regulation of SIRT1 by CPs to de-termine which specific anthocyanidins and flavonols are in-volved [92, 93]. Collectively, the molecular mechanisms havebeen hypothesized through regulating the activity of NRs,PPARs, LXRs, NF-�B/AP-1, and Nrf2 involved in metabolicdiseases, chronic inflammation, and antioxidants, respec-tively, as shown in Fig. 4. However, in order to definitivelydetermine the precise molecular mechanisms underlyingthe protective effects of polyphenols from cocoa on lipidmetabolism and inflammation reactions, large studies withgenome-wide analysis need to be conducted in the future.
5 Cocoa consumption andcontraindications
Most often physicians tended to warn patients about the po-tential health hazards of consuming chocolate. However, the
recent discovery of biologically active phenolic compounds incocoa has changed this notice and stimulated research on itseffects in ageing, oxidative stress, blood pressure regulation,diabetes, cancer, and atherosclerosis. In addition, cocoa hasmicronutrients, such as calcium, zinc, copper, manganese,and iron, all important for good health [134, 135]. Althoughmany epidemiological studies have reported the positive ef-fects of chocolate and its ingredients in obesity-related dis-eases, precautions from its use are still recommended. Themain reason behind that is the high caloric content of com-mercial chocolates (500 kcal/100g). Furthermore, the highsugar and fat content may induce weight gain, a risk factorfor obesity. However, human study in 49 healthy women re-vealed that weight gain has not been developed after dailyconsumption of 41 g chocolate, 60 g almonds for 6 wk [136].This result reflects that weight gain may occur only withoverconsumption of high amounts of chocolate. Thus, cocoaby-products with low fat content are highly preferred. It isvery necessary to distinguish between defatted product co-coa, such as supplement-rich flavonoids, and the processedproduct chocolate, which is rich in sugar, fat, and other in-gredients into a solid product. Obviously, most of the healthbenefits of cocoa and its contents discussed in this reviewmay not be applicable to chocolate.
Furthermore, due to the limitations of data available so far,further studies should provide detailed information about theproduct of chocolate along with the exact content of flavanols.More specifically, many of the above studies suggest that thebeneficial effects of cocoa are mainly attributed to its flavanolcontent, in particular, catechins. Therefore, the food manu-facturers are encouraged to label the flavonoid content ontheir derived products. Moreover, it is important to mentionthat cocoa contains other bioactive substances, e.g., caffeine,theophyline, and theobromine, which should be discussed inmore details with another review.
6 Conclusion
The possible molecular mechanisms by which polyphenols-rich cocoa prevent metabolic diseases and chronic inflam-mation associated with obesity are summarized in Fig. 4. Inliving organisms, the mechanisms are much more complex.For this reason, arrays technology has become available to un-derstand the molecular mechanisms underlying alterationsin the pathophysiologic status of dietary obesity and createan exciting new field for nutrition and health care. However,further investigations are still needed in the nutrigenomicsanalysis to fully elucidate the genome-wide influences of CPsand patterns of global gene expression, protein expression,intracellular signaling pathways, and metabolite productionin response to particular compounds. Finally, examine the ef-fects of CPs with purified single and mixed cocoa compoundson the gene expression profiling with special emphasis onhuman intervention trials, ideally with large-scale random-ized placebo-controlled studies, in order to yield important
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

12 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
insights into their prophylactic and therapeutic uses, as wellas develop effective strategies for alleviating obesity-mediatedmetabolic diseases and chronic inflammation.
We thank our group research members for their thorough re-view and helpful discussions during the preparation of this reviewand library for assistance with collection of references.
The authors have declared no conflict of interests.
7 References
[1] Baba, S., Osakabe, N., Natsume, M., Yasuda, A. et al., Cocoapowder enhances the level of antioxidative activity in ratplasma. Br. J. Nutr. 2000, 84, 673–680.
[2] Andres-Lacueva, C., Monagas, M., Khan, N., Izquierdo-Pulido, M. et al., Flavanol and flavonol contents of cocoapowder products: influence of the manufacturing process.J. Agric. Food Chem. 2008, 56, 3111–3117.
[3] Manach, C., Scalbert, A., Morand, C., Remesy, C. et al.,Polyphenols: food sources and bioavailability. Am. J. Clin.Nutr. 2004, 79, 727–747.
[4] Dai, J., Mumper, R. J., Plant phenolics: extraction, analysisand their antioxidant and anticancer properties. Molecules2010, 15, 7313–7352.
[5] Klem, M. A., Johnson, J. C., Robbins, R. J., Hammerstone,J. F. et al., high-performance liquid chromatography sep-aration and purification of cacao (Theobroma cacao L.)procyanidins according to degree of polymerization us-ing a diol stationary phase. J. Agric. Food Chem. 2006, 54,1571–1576.
[6] Hammerstone, J. F., Lazarus, S. A., Mitchell, A. E., Rucker,R. et al., Identification of procyanidins in cocoa (Theo-broma cacao) and chocolate using high-performance liquidchromatography/mass spectrometry. J. Agric. Food Chem.1999, 47, 490–496.
[7] Halliwell, B., Plasma antioxidants: health benefits of eat-ing chocolate? Nature 2003, 426, 787, discussion 788. DOI:10.1038/426787a
[8] Tomas-Barberan, F. A., Cienfuegos-Jovellanos, E., Marin,A., Muguerza, B. et al., A new process to develop a cocoapowder with higher flavonoid monomer content and en-hanced bioavailability in healthy humans. J. Agric. FoodChem. 2007, 55, 3926–3935.
[9] Rasmussen, S. E., Frederiksen, H., Krogholm, K. S., Poulsen,L., Dietary proanthocyanidins: occurrence, dietary intake,bioavailability, and protection against cardiovascular dis-ease. Mol. Nutr. Food Res. 2005, 49, 159–174.
[10] Williamson, G., Barron, D., Shimoi, K., Terao, J., In vitrobiological properties of flavonoid conjugates found in vivo.Free Radic. Res. 2005, 39, 457–469.
[11] Roura, E., Andres-Lacueva, C., Estruch, R., Mata-Bilbao, M.L. et al., Milk does not affect the bioavailability of cocoapowder flavonoid in healthy human. Ann. Nutr. Metab.2007, 51, 493–498.
[12] Boyer, J., Brown, D., Liu, R. H., Uptake of quercetin andquercetin 3-glucoside from whole onion and apple peel
extracts by Caco-2 cell monolayers. J. Agric. Food Chem.2004, 52, 7172–7179.
[13] Yi, W., Akoh, C. C., Fischer, J., Krewer, G., Absorption ofanthocyanins from blueberry extracts by Caco-2 human in-testinal cell monolayers. J. Agric. Food Chem. 2006, 54,5651–5658.
[14] Daavalos, A., Fernaanndez-Hernando, C., Cerrato, F.,Martınez-Botas, J. et al., Red grape juice polyphenols altercholesterol homeostasis and increase LDL-receptor activityin human cells in vitro. J. Nutr. 2006, 136, 1766–1773.
[15] Faria, A., Pestana, D., Azevedo, J., Martel, F. et al., Ab-sorption of anthocyanins through intestinal epithelial cells-putative involvement of GLUT2. Mol. Nutr. Food Res. 2009,53, 1430–1437.
[16] De Pascual-Teresa, S., Moreno, D. A., Garcıa-Viguera, C.,Flavanols and anthocyanins in cardiovascular health: areview of current evidence. Int. J. Mol. Sci. 2010, 11,1679–1703.
[17] Ross, J. A., Kasum, C. M., Dietary flavonoids: bioavailability,metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22,19–34.
[18] Baba, S., Osakabe, N., Yasuda, A., Natsume, M. et al.,Bioavailability of (-)-epicatechin upon intake of chocolateand cocoa in human volunteers. Free Radic. Res. 2000, 33,635–641.
[19] Steinert, R. E., Ditscheid, B., Netzel, M., Jahreis, G., Ab-sorption of black currant anthocyanins by monolayers ofhuman intestinal epithelial Caco-2 cells mounted in usingtype chambers. J. Agric. Food Chem. 2008, 56, 4995–5001.
[20] Rimbach, G., Melchin, M., Moehring, J., Wagner, A. E.,Polyphenols from cocoa and vascular health. Int. J. Mol.Sci. 2009, 10, 4290–4309.
[21] Manach, C., Williamson, G., Morand, C., Scalbert, A. et al.,Bioavailability and bioefficacy of polyphenols in humans.Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005,81, 230S–242S.
[22] Clifford, M. N., Diet-derived phenols in plasma and tissuesand their implications for health. Planta Med. 2004, 70,1103–1114.
[23] Kroon, P. A., Clifford, M. N., Crozier, A., Day, A. J. et al.,How should we assess the effects of exposure to di-etary polyphenols in vitro? Am. J. Clin. Nutr. 2004, 80,15–21.
[24] Barger, J. L., Kayo, T., Vann, J. M., Arias, E. B. et al. A lowdose of dietary resveratrol partially mimics caloric restric-tion and retards aging parameters in mice. PLoS One 2008,3, e2264.
[25] Davison, K., Coates, A. M., Buckley, J. D., Howe, P. R., Effectof cocoa flavanols and exercise on cardiometabolic risk fac-tors in overweight and obese subjects. Int. J. Obes. 2008,32, 1289–1296.
[26] Stevenson, D. E., Hurst, R. D., Polyphenolicphytochemicals-just antioxidants or much more? Cell.Mol. Life Sci. 2007, 64, 2900–2916.
[27] Report of a World Health Organization (WHO) Consultation(WHO Technical Report Series 894) on Obesity: Preventingand Managing the Global Epidemic. WHO, Geneva 2000.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 13
[28] WHO, Obesity and overwight. Fact sheet N311 (on-line), 2006. Available at: http://www.who.int/mediacentre/factsheets/fs311/en/index.html (accessed in 20 September,2009).
[29] Bastard, J. P., Maachi, M., Lagathu, C., Kim, M. J. et al., Re-cent advances in the relationship between obesity, inflam-mation, and insulin resistance. Eur. Cytokine Netw. 2006,17, 4–12.
[30] Visioli, F., Bernaert, H., Corti, R., Ferri, C. et al., Chocolate,lifestyle, and health. Crit. Rev. Food Sci. Nutr. 2009, 49,299–312.
[31] Galleano, M., Oteiza, P. I., Fraga, C. G., Cocoa, chocolate andcardiovascular disease. J. Cardiovasc. Pharmacol. 2009, 54,483–490.
[32] Matsui, N., Ito, R., Nishimura, E., Yoshikawa, M. et al.,Ingested cocoa can prevent high-fat diet-induced obe-sity by regulating the expression of genes for fatty acidmetabolism. Nutrition 2005, 5, 594–601.
[33] Stienstra, R., Duval, C., Muller, M., Kersten, S., PPARs, Obe-sity, and Inflammation. PPAR Res. 2007, 2007, 95974.
[34] Staels, B., Auwerx, J., Regulation of apo-A-1 gene ex-pression by fibrates. J. Atherosclerosis. 1998, 137(Suppl),S19–S23.
[35] Goldwasser, J., Cohen, P. Y., Yang, E., Balaguer, P. et al.,Transcriptional regulation of human and rat hepatic lipidmetabolism by the grapefruit flavonoid Naringenin: role ofPPARs and LXR�. PLoS One 2010, 8, e12399.
[36] Waterhouse, A. L., Shirley, J. R., Donovan, J. L., Antioxi-dants in chocolate. Lancet 1996, 348, 834.
[37] Sanbongi, C., Suzuki, N., Sakane, T., Polyphenols in choco-late, which have antioxidant activity, modulate immunefunctions in humans in vitro. Cell Immunol. 1997, 177,129–136.
[38] Ottaviani, J. I., Carrasquedo, F., Keen, C. L., Lazarus,S. A. et al., Influence of flavan-3-ols and procyani-dins on UVC-mediated formation of 8-oxo-7, 8-dihydro-2’-deoxyguanosine in isolated DNA. Arch. Biochem. Biophys.2002, 406, 203–208.
[39] Wiswedel, I., Hirsch, D., Kropf, S., Gruening, M. et al.,Flavanol-rich cocoa drink lowers plasma F(2)-isoprostaneconcen-trations in humans. Free Radic. Biol. Med. 2004, 37,411–421.
[40] Rein, D., Lotito, S., Holt, R. R., Keen, C. L. et al., Epicate-chin in human plasma: in vivo determination and effect ofchocolate consumption on plasma oxidation status. J. Nutr.2000, 130, 2109–2114.
[41] Turner, R., Baron, T., Wolffram, S., Minihane, A. M. et al.,Effect of circulating forms of soy isoflavones on the oxida-tion of low density lipoprotein. Free Radic. Res. 2004, 38,209–216.
[42] Mathur, S., Devaraj, S., Grundy, S. M., Jialal, I., Cocoa prod-ucts decrease low density lipoprotein oxidative susceptibil-ity but do not affect biomarkers of inflammation in humans.J. Nutr. 2002, 132, 3663–3667.
[43] Richelle, M., Tavazzi, I., Offord, E., Comparison of the antiox-idant activity of commonly consumed polyphenolic bever-
ages (coffee, cocoa, and tea) prepared per cup serving. J.Agric. Food Chem. 2001, 49, 3438–3442.
[44] Osakabe, N., Yasuda, A., Natsume, M., Takizawa, T. et al.,Catechins and their oligomers linked by C4 → C8 bonds aremajor cacao polyphenols and protect low-density lipopro-tein from oxidation in vitro. Exp. Biol. Med. (Maywood)2002, 227, 51–56.
[45] Kurosawa, T., Itoh, F., Nozaki, A., Nakano, Y. et al., Suppres-sive effect of cocoa powder on atherosclerosis in Kurosawaand Kusanagi-hypercholesterolemic rabbits. J. Atheroscler.Thromb. 2005, 12, 20–28.
[46] Orozco, T. J., Wang, J. F., Keen, C. L., Chronic consump-tion of a flavanol- and procyanindin-rich diet is associatedwith reduced levels of 8-hydroxy-2’-deoxyguanosine in rattestes. J. Nutr. Biochem. 2003, 14, 104–110.
[47] Wan, Y., Vinson, J. A., Etherton, T. D., Proch, J. et al., Ef-fects of cocoa powder and dark chocolate on LDL oxidativesusceptibility and prostaglandin concentrations in humans.Am. J. Clin. Nutr. 2001, 74, 596–602.
[48] Kondo, K., Hirano, R., Matsumoto, A., Igarashi, O. et al.,Inhibition of LDL oxidation by cocoa. Lancet. 1996, 349,360–361.
[49] Osakabe, N., Baba, S., Yasuda, A., Iwamoto, T. et al.,Daily cocoa intake reduces the susceptibility of low-densitylipoprotein to oxidation as demonstrated in healthy humanvolunteers. Free Radic. Res.2001, 34, 93–99.
[50] Vinson, J. A., Proch, J., Bose, P., Muchler, S. et al., Choco-late is a powerful ex vivo and in vivo antioxidant, an anti-atherosclerotic agent in an animal model, and a significantcontributor to antioxidants in the European and AmericanDiets. J. Agric. Food Chem. 2006, 54, 8071–8076.
[51] Yasuda, A., Natsume, M., Sasaki, K., Baba, S. et al., Cacaoprocyanidins reduce plasma cholesterol and increase fecalsteroid excretion in rats fed a high cholesterol diet. Biofac-tors. 2008, 33, 211–223.
[52] Jalil, A. M., Ismail, A., Pei, C. P., Hamid, M. et al., Ef-fects of cocoa extract on glucometabolism, oxidative stress,and antioxidant enzymes in obese-diabetic (ob-db) rats. J.Agric. Food Chem. 2008, 56, 7877–7884.
[53] Rivera, L., Moron, R., Sanchez, M., Zarzuelo, A. et al.,Quercetin ameliorates metabolic syndrome and improvesthe inflammatory status in obese Zucker rats. Obesity. 2008,16, 2081–2087.
[54] Baba, S., Osakabe, N., Kato, Y., Natsume, M. et al., Contin-uous intake of polyphenolic compounds containing cocoapowder reduces LDL oxidative susceptibility and has ben-eficial effects on plasma HDL-cholesterol concentrations inhumans. Am. J. Clin. Nutr. 2007, 85, 709–717.
[55] Mursu, J., Voutilainen, S., Nurmi, T., Rissanen, T. H. et al.,Dark chocolate consumption increases HDL cholesterolconcentration and chocolate fatty acids may inhibit lipidperoxidation in healthy humans. Free Radic. Biol. Med.2004, 37, 1351–1359.
[56] Wang, J. F., Schramm, D. D., Holt, R. R., Ensunsa, J. L. et al.,A dose-response effect from chocolate consumption onplasma epicatechin and oxidative damage. J. Nutr. 2000,130, 2115S–2119S.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

14 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
[57] Rein, D., Paglieroni, T. G., Wun, T., Pearson, D. A. et al.,Cocoa inhibits platelet activation and function. Am. J. Clin.Nutr. 2000, 72, 30–35.
[58] Taubert, D., Berkels, R., Roesen, R., Klaus, W., Chocolate andblood pressure in elderly individuals with isolated systolichypertension. JAM. 2003, 290, 1029–1030.
[59] Kim, S., Sohn, I., Lee, Y. S., Lee, Y. S., Hepatic gene expres-sion profiles are altered by genistein supplementation inmice with diet-induced obesity. J. Nutr. 2004, 135, 33–41.
[60] Kim, S., Shin, H.-J., Kim, S. Y., Kim, J. H. et al., Genis-tein enhances expression of genes involved in fatty acidcatabolism through activation of PPAR�. Mol. Cell. En-docrinol. 2004, 220, 51–58.
[61] Lau, D. C., Dhillon, B., Yan, H., Szmitko, P. E.et al., Adipokines: molecular links between obesity andatherosclerosis. Am. J. Physiol. Heart Circ. Physiol. 2005,288, H2031–H2041.
[62] Borchers, A. A. T., Keen, C. L., Hannum, S. M., Gershwin, M.E., Cocoa and chocolate composition, bioavailability, andhealth implications. J. Med. Food. 2000, 3, 77–105.
[63] Loke, W. M., Proudfoot, J. M., Hodgson, J. M., McKin-ley, A. J. et al., Specific dietary polyphenols attenuateatherosclerosis in apolipoprotein E-knockout mice by al-leviating inflammation and endothelial dysfunction. Arte-rioscler. Thromb. Vasc. Biol. 2010, 30, 749–757.
[64] Puglisi, M. J., Vaishnav, U., Shrestha, S., Torres-Gonzalez,M. et al., Raisins and additional walking have distinct effectson plasma lipids and inflammatory cytokines. Lipids HealthDis. 2008, 16, 7–14.
[65] Schnorr, O., Brossette, T., Momma, T. Y., Kleinbongard, P.et al., Cocoa flavanols lower vascular arginase activity inhuman endothelial cells in vitro and in erythrocytes in vivo.Arch. Biochem. Biophys. 2008, 476, 211–215.
[66] Karim, M., McCormick, K., Kappagoda, C. T., Effects of co-coa extracts on endothelium dependent relaxation. J. Nutr.2000, 130, 2105S–2108S.
[67] Lee, K. W., Kang, N. J., Oak, M. H., Hwang, M. K. et al., Cocoaprocyanidins inhibit expression and activation of MMP-2 invascular smooth muscle cells by direct inhibition of MEKand MT1-MMP activities. Cardiovasc. Res. 2008, 79, 34–41.
[68] Actis-Goretta, L., Ottaviani, J. I., Keen, C. L., Fraga, C. G.,Inhibition of angiotensin converting enzyme (ACE) activ-ity by flavan-3-ols and procyanidins. FEBS Lett. 2003, 555,597–600.
[69] Guo, Q., Rimbach, G., Packer, L., Nitric oxide formationin macrophages detected by spin trapping with iron-dithiocarbamate complex: effect of purified flavonoids andplant extracts. Methods Enzymol. 2001, 335, 273–282.
[70] Saliou, C., Valacchi, G., Rimbach, G., Assessingbioflavonoids as regulators of NF-kappa B activityand inflammatory gene expression in mammalian cells.Methods Enzymol. 2001, 335, 380–387.
[71] Hermann, F., Spieker, L. E., Ruschitzka, F., Sudano, I. et al.,Dark chocolate improves endothelial and platelet function.Heart. 2006, 92, 119–120.
[72] Fisher, N. D., Hughes, M., Gerhard-Herman, M., Hollenberg,N. K., Flavanol-rich cocoa induces nitric-oxide-dependent
vasodilation in healthy humans. J. Hypertens. 2003, 21,2281–2286.
[73] Vlachopoulos, C., Aznaouridis, K., Alexopoulos, N.,Economou, E. et al., Effect of dark chocolate on arterialfunction in healthy individuals. Am. J. Hypertens. 2005, 18,785–791.
[74] Balzer, J., Rassaf, T., Heiss, C., Kleinbongard, P. et al.,Sustained benefits in vascular function through flavanol-containing cocoa in medicated diabetic patients a double-masked, randomized, controlled trial. J. Am. Coll. Cardiol.2008, 51, 2141–2149.
[75] Farouque, H. M., Leung, M., Hope, S. A., Baldi, M. et al.,Acute and chronic effects of flavanol-rich cocoa on vascu-lar function in subjects with coronary artery disease: a ran-domized double-blind placebo-controlled study. Clin. Sci.(Lond.) 2006, 111, 71–80.
[76] Flammer, A. J., Hermann, F., Sudano, I., Spieker, L.et al., Dark chocolate improves coronary vasomotionand reduces platelet reactivity. Circulation 2007, 116,2376–2382.
[77] Peng, C. H., Liu, L. K., Chuang, C. M., Chyau, C. C. et al.,Mulberry water extracts possess an anti-obesity effect andability to inhibit hepatic lipogenesis and promote lipolysis.J. Agric. Food Chem. 2011, 6, 2663–2671.
[78] Jemi, H., Bouaziz, M., Fki, I., EI Feki, A. et al., Hypolipi-demic and antioxidant activities of oleuropein—its hydrol-ysis derivatives rich extracts from Chemlali olive leaves.Chem. Biol. interact. 2008, 176, 88–98.
[79] Murase, T., Nagasawa, A., Suzuki, J., Hase, T. et al., Bene-ficial effects of tea catechins on diet-induced obesity: stim-ulation of lipid catabolism in the liver. Int. J. Obes. Relat.Metab. Disord. 2002, 26, 1459–1464.
[80] Ohta, Y., Sami, M., Kanda, T., Saito, K. et al., Gene expres-sion analysis of the anti-obesity effect by apple polyphenolsin rats fed a high fat diet or a normal diet. J. Oleo. Sci. 2006,55, 305–314.
[81] Klein, G., Kim, J., Himmeldirk, K., Cao, Y., Chen, X.,Anti-diabetes and anti-obesity activity of Lagestroemiaspeciosa. Evid. Based Complement Alternat. Med. 2007,24, 401–407.
[82] Canete, R., Gil-Campos, M., Aguilera, C. M., Gil, A., Devel-opment of insulin resistance and its relation to diet in theobese child. Eur. J. Nutr. 2007, 46, 181–187.
[83] Stumvoll, M., Goldstein, B. J., van Haeften, T. W., Type 2diabetes: principles of pathogenesis and therapy. Lancet.2005, 36, 1333–1346.
[84] Amin, I, Faizul, H. A., Azil, R., Effect of cocoa powder extracton plasma glucose levels in hyperglycemia rats. Nutr. FoodSci. 2004, 34, 116–121.
[85] Ruzaidi, A., Amin, I., Nawlyah, A. G., Hamid, M. et al., Theeffect of Malaysian cocoa extract on glucose levels andlipid profiles in diabetic rats. J. Ethnopharmacol. 2005, 98,55–60.
[86] Tomaru, M., Takano, H., Osakabe, N., Yasuda, A. et al., Di-etary supplementation with cacao liquor proanthocyani-dins prevents elevation of blood glucose levels in diabeticobese mice. Nutrition. 2007, 23, 351–355.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–16 15
[87] Baur, J. A., Pearson, K. J., Price, N. L., Jamieson, H. A.et al., Resveratrol improves health and survival of mice ona high-calorie diet. Nature. 2006, 444, 337–342.
[88] Sun, C., Zhang, F., Ge, X., Yan, T. et al., SIRT1 improvesinsulin sensitivity under insulin- resistant conditions by re-pressing PTP1B. Cell Metab. 2007, 6, 307–319.
[89] Chuang, C.-C., McIntosh, M. K., Potential mechanisms bywhich polyphenol-rich grapes prevent obesity-mediatedinflammation and metabolic diseases. Annu. Rev. Nutr.2011, 31, 155–176.
[90] Grassi, D., Desideri, G., Necozione, S., Lippi, C. et al., Bloodpressure is reduced and insulin sensitivity increased inglucose-intolerant, hypertensive subjects after 15 days ofconsuming high-polyphenol dark chocolate. J. Nutr. 2008,138, 1671–1676.
[91] Grassi, D., Lippi, C., Necozione, S., Desideri, G. et al., Short-term administration of dark chocolate is followed by a sig-nificant increase in insulin sensitivity and a decrease inblood pressure in healthy persons. Am. J. Clin. Nut. 2005,81, 611–614.
[92] Ikarashi, N., Toda, T., Okaniwa, T., Ito, K. et al., Anti-obesityand anti-diabetic effects of acacia polyphenol in obesediabetic KKAy mice fed high-fat diet. eCAM. 2010, 241,1–10.
[93] Lagouge, M., Argmann, C., Gerhart-Hines, Z., Meziane, H.et al., Resveratrol improves mitochondrial function and pro-tects against metabolic disease by activating SIRT1 andPGC-1 alpha. Cell 2006, 127, 1109–1122.
[94] Chaudhary, N., Pfluger, P. T., metabolic benefits from Sirt1and Sirt1 activators. Curr. Opin. Clin. Nutr. Metab. Care2009, 12, 431–437.
[95] Yung, L. M., Leung, F. P., Yao, X., Chen, Z. Y. et al., Reactiveoxygen species in vascular wall. Cardiovasc. Hematologic.Disorders 2006, 6, 1–19.
[96] Tang, J., Yan, H., Zhuang, S., Inflammation and oxidativestress in obesity-related glomerulopathy. Int. J. Nephrol.2012, 608397, 1–7.
[97] Bengtsson, S. H. M., Gulluyan, L. M., Dusting, G. J., Drum-mond, G. R., Novel isoforms of NADPH oxidase in vascu-lar physiology and pathophysiology. Clin. Exp. Pharmacol.Physiol. 2003, 30, 849–854.
[98] Touyz, R. M., Tabet, F., Schiffrin, E. L., Redox- dependent sig-naling by angiotensin II and vascular remodelling in hyper-tension. Clin. Exp. Pharmacol. Physiol. 2003, 30, 860–866.
[99] Schreck, C., O’Connor, P. M., NAD(P) H oxidase and re-nal epithelial ion transport. Am. J. Physiol. 2011, 300,R1023–R1029.
[100] Cai, H., Hydrogen peroxide regulation of endothelial func-tion: origins, mechanisms, and consequences. Cardiovasc.Res. 2005, 68, 26–36.
[101] Cai, H., Harrison, D. G., Endothelial dysfunction in cardio-vascular diseases: the role of oxidant stress. Circ. Res. 2000,87, 840–844.
[102] Rahman, I., Biswas, S. K., Kirkham, P. A., Regulation of in-flammation and redox signaling by dietary polyphenols.Biochem. Pharmacol. 2006, 72, 1439–1452.
[103] Ramiro-Puig, E., Castell, M., Cocoa: antioxidant and im-munomodulator. Br. J. Nutr. 2009, 101, 931–940.
[104] Guo, H., Ling, W., Wang, Q., Liu, C. et al., Cyanidin 3-glucoside protects 3T3-L1 adipocytes against H2O2- orTNF-alpha-induced insulin resistance by inhibiting c-JunNH2-terminal kinase activation. Biochem. Pharmacol. 2008,75, 1393–1401.
[105] Choi, J. S., Kang, S. W., Li, J., Kim, J. L. et al., Blockade ofoxidized LDL-triggered endothelial apoptosis by quercetinand rutin through differential signaling pathways involvingJAK2. J. Agric. Food Chem. 2009, 57, 2079–2086.
[106] Noe, V., Penuelas, S., Lamuela-Raventos, R. M., Permanyer,J., et al., Epicatechin and a cocoa polyphenolic extract mod-ulate gene expression in human Caco-2 cells. J. Nutr. 2004,134, 2509–2516.
[107] Kersten, S., Desvergne, B., Wahli, W., Roles of PPARS inhealth and disease. Nature 2000, 405, 421–424.
[108] Sugden, M. C., Caton, P. W., Holness, M. J., PPAR control: it’sSIRTainly as easy as PGC. J. Endocrinal. 2010, 204, 93–104.
[109] Chawla, I., Repa, J. J., Evans, R. M., Mangelsdrof, D. J.,Nuclear receptors and lipid physiology: opening the x-files.Science 2001, 294, 1866–1870.
[110] Patsouris, D., Muller, M., Kersten, S., Peroxisome prolifer-ator activated receptor ligands for the treatment of insulinresistance. Curr. Opin. Invest. Drugs 2004, 5, 1045–1050.
[111] Guo, L., Tabrizchi, R., Peroxisome proliferator-activatedreceptor gamma as a drug target in the pathogenesisof insulin resistance. Pharmacol. Therapeutics. 2006, 111,145–173.
[112] Maeda, N., Takahashi, M., Funahashi, T., Kihara, S. et al.,PPAR� ligands increase expression and plasma concentra-tions of adiponectin, an adipose-derived protein. Diabetes2001, 50, 2094–2098.
[113] Olholm, J., Paulsen, S. K., Cullberg, K. B., Richelsen, B.et al., Anti-inflammatory effect of resveratrol on adipokineexpression and secretion in human adipose tissue explants.Int. J. Obes. 2010, 34, 1546–1553.
[114] Huang, S. M., Wu, C. H., Yen, G. C., Effects of flavonoids onthe expression of the pro-inflammatory response in humanmonocytes induced by ligation of the receptor for AGEs.Mol. Nutr. Food Res. 2006, 50, 1129–1139.
[115] Mao, T. K., van de Water, J., Keen, C. L., Schmitz, H. H.et al., Modulation of TNF-alpha secretion in peripheralblood mononuclear cells by cocoa flavanols and procyani-dins. Dev. Immunol. 2002, 9, 135–141.
[116] Mao, T., van de Water, J., Keen, C. L., Schmitz, H. H. et al.,Cocoa procyanidins and human cytokine transcription andsecretion. J. Nutr. 2000, 130, 2093S–2099S.
[117] Hotamisligil, G. S., Inflammation and metabolic disorders.Nature 2006, 444, 860–867.
[118] DeFuria, J., Bennett, G., Strissel, K. J., Perfield, J. W. 2ndet al., Dietary blueberry attenuates whole-body insulin re-sistance in high fat-fed mice by reducing adipocyte deathand its inflammatory squeal. J. Nutr. 2009, 139, 1510–1516.
[119] Stewart, L. K., Soileau, J. L., Ribnicky, D., Wang, Z. Q.et al. Quercetin transiently increases energy expenditure
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

16 F. Ali et al. Mol. Nutr. Food Res. 2013, 00, 1–16
but persistently decreases circulating markers of inflam-mation in C57BL/6J mice fed a high-fat diet. Metabolism2008, 57, 39–46.
[120] Chuang, C. C., Bumrungpert, A., Kennedy, A., Overman,A. et al., Grape powder extract attenuates tumor necrosisfactor alpha-mediated inflammation and insulin resistancein primary cultures of human adipocytes. J. Nutr. Biochem.2011, 22, 89–94.
[121] Chuang, C. C., Martinez, K., Xie, G., Kennedy, A.et al. Quercetin is equally or more effective than resver-atrol in attenuating tumor necrosis factor-{alpha}-mediatedinflammation and insulin resistance in primary humanadipocytes. Am. J. Clin. Nutr. 2010, 92, 1511–1521.
[122] Mao, T. K., Powell, J., van de Water, J., Keen, C. L. et al., Theeffect of cocoa procyanidins on the transcription and secre-tion of interleukin 1 beta in peripheral blood mononuclearcells. Life Sci. 2000, 66, 1377–1386.
[123] Schewe, T., Sadik, C., Klotz, L. O., Yoshimoto, T. et al.,Polyphenols of cocoa: inhibition of mammalian 5-lipoxygenase. Biol. Chem. 2001, 382, 1687–1696.
[124] Lee, K. W., Kundu, J. K., Kim, S. O., Chun, K.-S., et al., Co-coa polyphenols inhibit phorbol ester-induced superoxideanion ormation in cultured HL-60 cells and expression ofcyclooxygenase-2an d activation of NF-kB and MAPKs inmouse skin in vivo. J. Nutr. 2006, 136, 1150–1155.
[125] Monagas, M., Khan, N., Andres-Lacueva, C., Casas, R. et al.,Effect of cocoa powder on the modulation of inflammatorybiomarkers in patients at high risk of cardiovascular dis-ease. Am. J. Clin. Nutr. 2009, 90, 114–150.
[126] Karlsen, A., Paur, I., Bøhn, S. K., Sakhi, A. K. et al., Bilberryjuice modulates plasma concen- tration of NF-kappa B re-lated inflammatory markers in subjects at increased risk ofCVD. Eur. J. Nutr. 2010, 49, 345–355.
[127] Terra, X., Montagut, G., Bustos, M., Llopiz, N. et al., Grape-seed procyanidins prevent low-grade inflammation bymodulating cytokine expression in rats fed a high-fat diet.J. Nutr. Biochem. 2009, 20, 210–218.
[128] Lee, K. W., Kang, N. J., Heo, Y. S., Rogozin, E. A. et al., Rafand MEK protein kinases are direct molecular targets forthe chemopreventive effect of quercetin, a major flavonolin red wine. Cancer Res. 2008, 68, 946–955.
[129] Chen, C. C., Chow, M. P., Huang, W. C., Lin, Y. C. et al.,Flavonoids inhibit tumor necrosis factor-alpha-induced
up-regulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells through activator protein-1and nuclear factor-kappa B: structure-activity relationships.Mol. Pharmacol. 2004, 66, 683–693.
[130] Yang, Z., Kahn, B. B., Shi, H., Xue, B. Z., Macrophage al-pha 1 AMP- activated protein kinase (alpha1 MAPK) antag-onizes fatty acids-induced inflammation through SIRIT. J.Biol. Chem. 2010, 285, 19051–19059.
[131] Fischer-Posovszky, P., Kukulus, V., Tews, D., Unterkircher, T.et al., Resveratrol regulates human adipocyte number andfunction in a Sirt1-dependent manner. Am. J. Clin. Nut.2010, 92, 5–15.
[132] Zhu, J., Yong, W., Wu, X., Yu, Y. et al., Anti-inflammatoryeffect of resveratrol on TNF-alpha- induced MCP-1 expres-sion in adipocytes. Biochem. Biophys. Res. Commun. 2008,369, 471–477.
[133] Lang, R., Hammer, M., Mages, J., DUSP meet immunol-ogy: dual specificity MAPK phosphatases in control of theinflammatory response. J. Immunol. 2006, 11, 7497–7504.
[134] Cordero-Herrera, I., Martın, M. A., Bravo, L., Goya, L. et al.,Cocoa flavonoids improve insulin signalling and modulateglucose production via AKT and AMPK in HepG2 cells. Mol.Nutr. Food Res. 2013, 57, 974–985.
[135] Minakawa, M., Kawano, A., Miura, Y., Yagasaki, K., Hypo-glycemic effect of resveratrol in type 2 diabetic model db/dbmice and its actions in cultured L6 myotubes and RIN-5Fpancreatic �-cells. Clin. Biochem. Nutr. 2011, 48, 237–244.
[136] Bełtowski, J., Leptin and the regulation of endothelial func-tion in physiological and pathological conditions. Clin. Exp.Pharmacol. Physiol. 2011, 4, 1440–1681.
[137] Yamashita, Y., Okabe, M., Natsume, M., Ashida, H., Preven-tion mechanisms of glucose intolerance and obesity by ca-cao liquor procyanidin extract in high-fat diet-fed C57BL/6mice. Arch. Biochem. Biophys. 2012, 527, 95–104.
[138] Corti, R., Flammer, A. J., Hollenberg, N. K., Luscher, T. F.,Cocoa and cardiovascular health. Circulation 2009, 119,1433–1441.
[139] Arts, I. C. W., Hollman, P. C. H., Polyphenols and diseaserisk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81,S317–S325.
[140] Kurlandsky, S. B., Stote, K. S., Cardioprotective effects ofchocolate and almond consumption in healthy women.Nutr. Res. 2006, 26, 509–516.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
Related Documents











