Molecular Mechanisms of Gene Regulation: The Operon (Ch7)

Molecular Mechanisms of Gene Regulation: The Operon (Ch7)
Dec 14, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Mechanisms of Gene Regulation:
The Operon (Ch7)

Operon- set of genes that are coordinately controlled by a regulatory protein AND transcribed as a single polycistronic message
Regulon- set of related genes that are transcribed as separate units but are controlled by the same regulatory protein
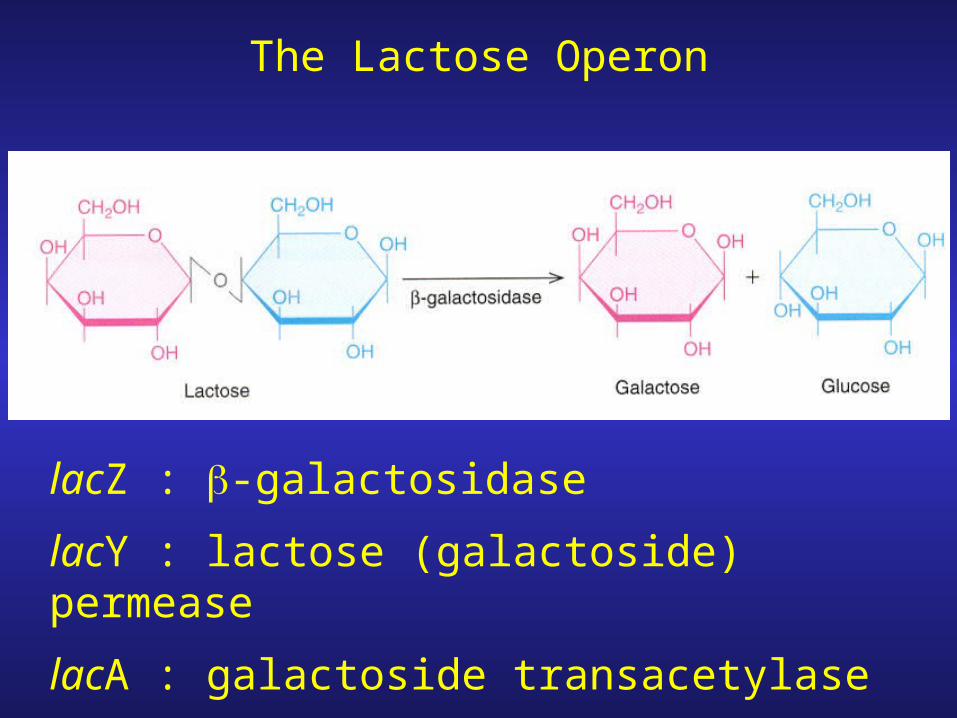
The Lactose Operon
lacZ : b-galactosidase
lacY : lactose (galactoside) permease
lacA : galactoside transacetylase
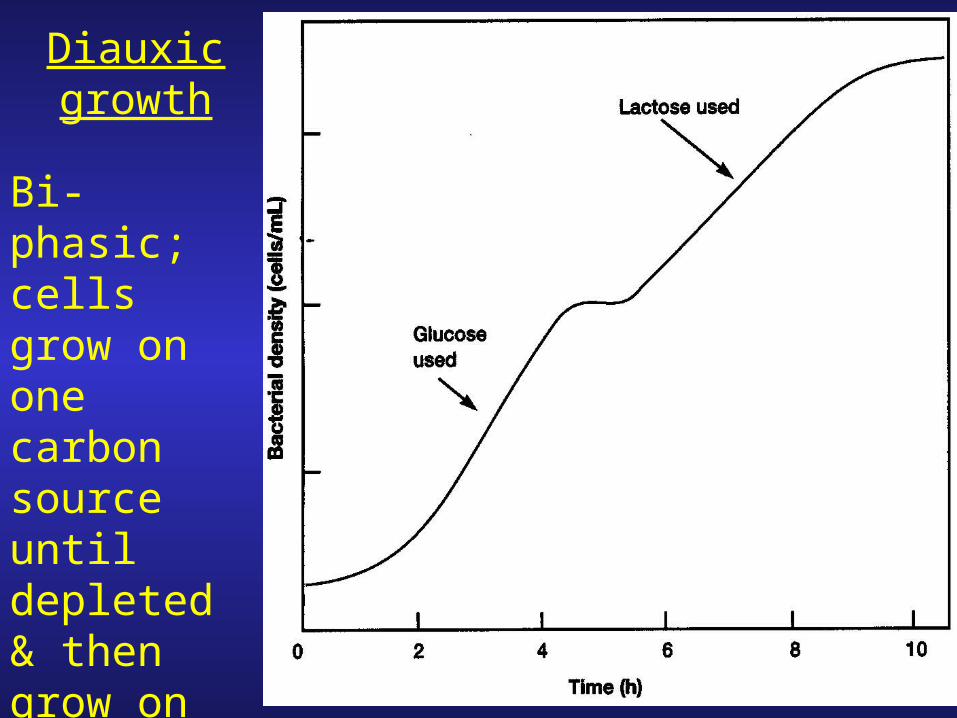
Diauxic growth
Bi-phasic; cells grow on one carbon source until depleted & then grow on the other

Francois Jacob Jaques Monod

1. Diauxic growth is dependent upon the carbon (sugar) source used.
2. In E. coli: two classes of sugar sources
(i) glucose, mannose, fructose
(ii) lactose, maltose
3. Growth on class (i) combinations, i.e. glucose + mannose no diauxic growth; same with class (ii) mixtures.
4. Diauxy is observed when cells are grown in mixtures containing (i) + (ii).
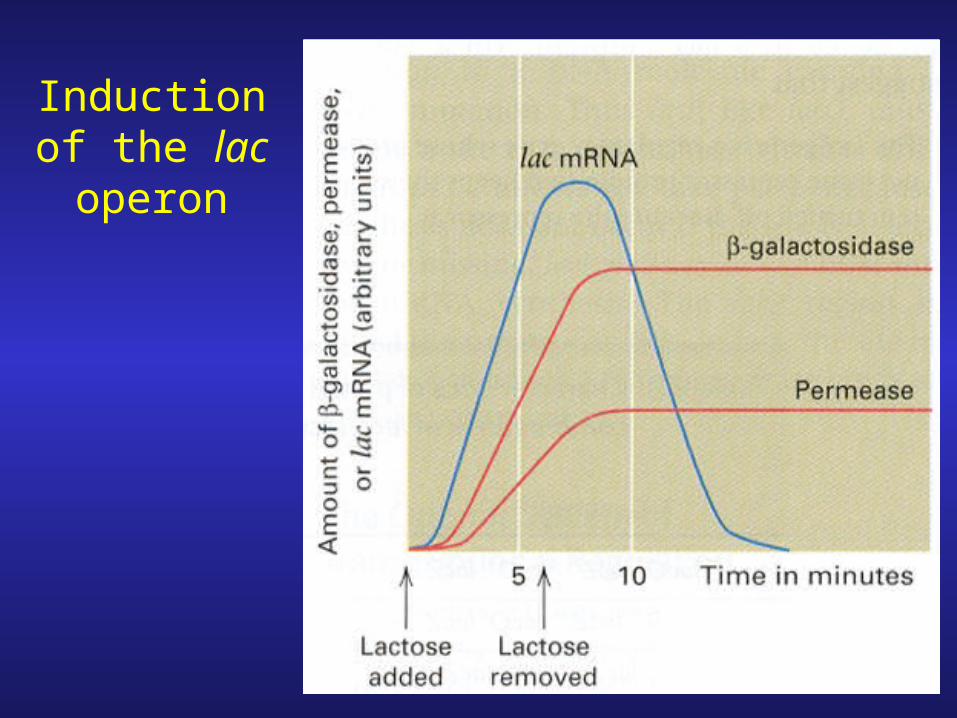
Induction of the lac operon
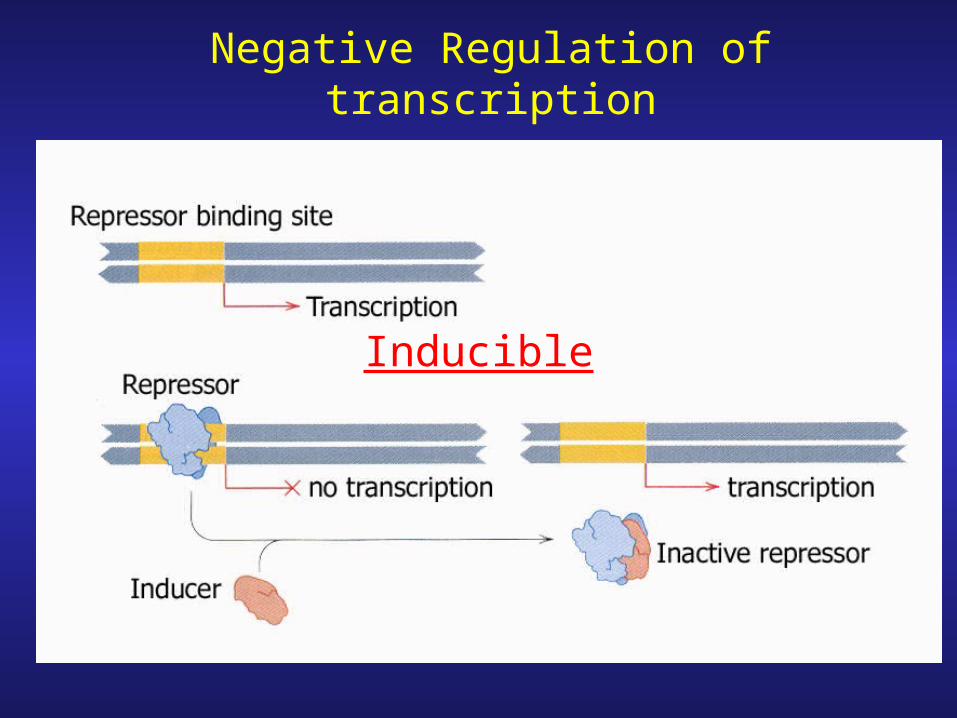
Negative Regulation of transcription
Inducible
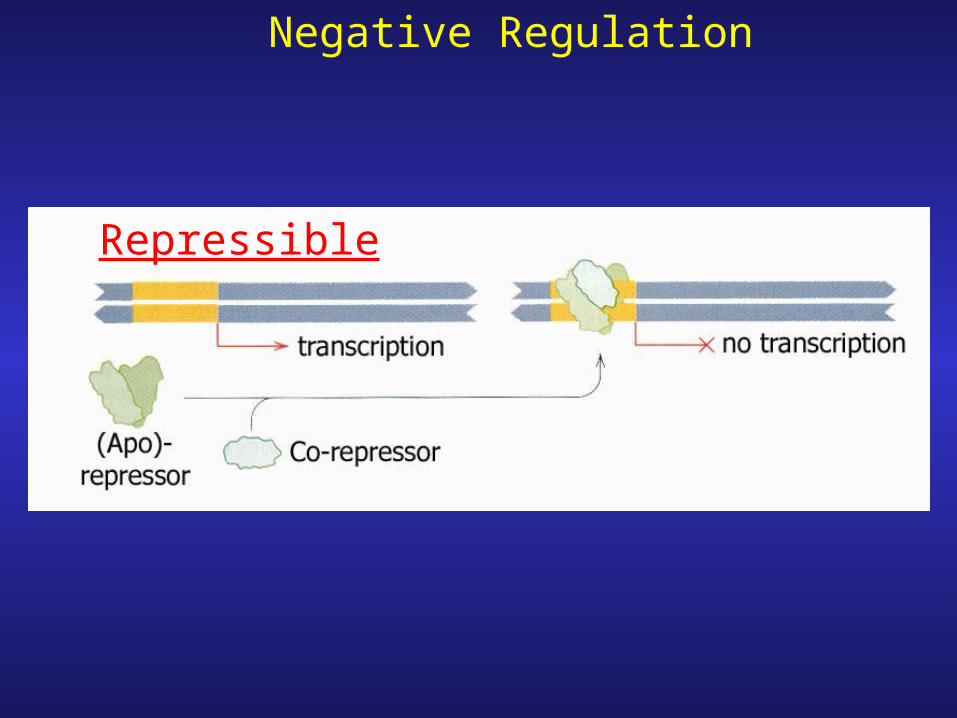
Negative Regulation
Repressible
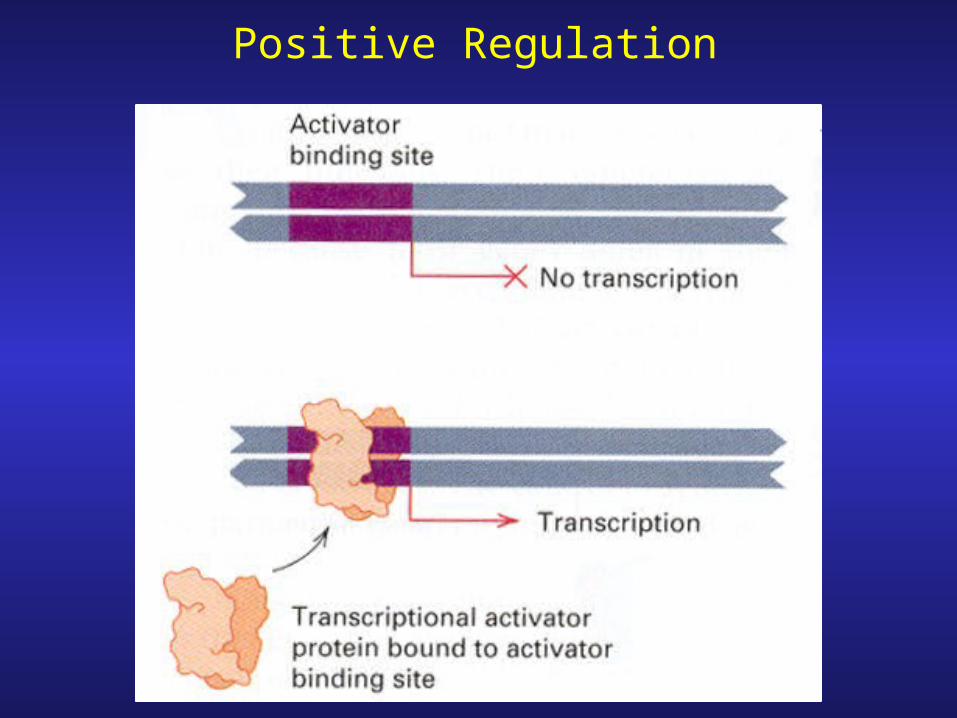
Positive Regulation
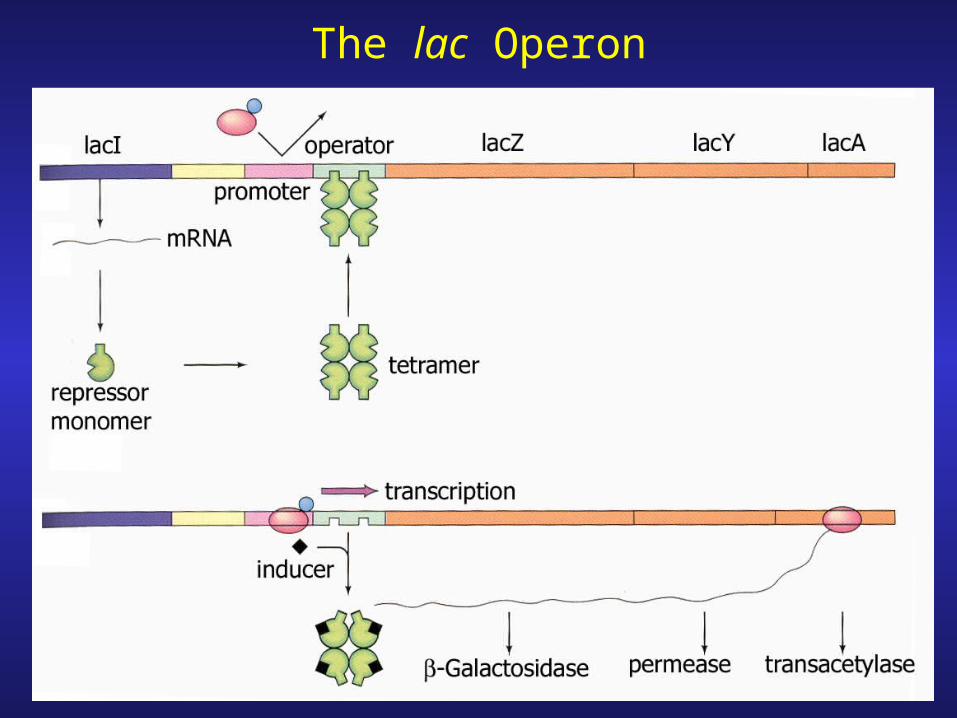
The lac Operon

The nature of the lac inducer

Complementation
1. Restoration of phenotype
2. Different types: genetic material
3. Mutation with phenotype add DNA (gene product) restores phenotype
Typical conclusion: mutation & complementing DNA encode-for or are the same gene
Alternate conclusions: compensatory affects

Complementation using two (recessive)
mutants
Interpretations very different


Mutant Repressor Gene
Y/NY/N
Lac product?
+ inducer
- inducer
Conclusion: Both lac operons are repressible
(no repressor made)
recessive
Y/NY/N

Mutant Operator (Oc)
Y/N
Y/N
Lac product
?
Conclusion: One lac operon non-repressible
cis-dominant
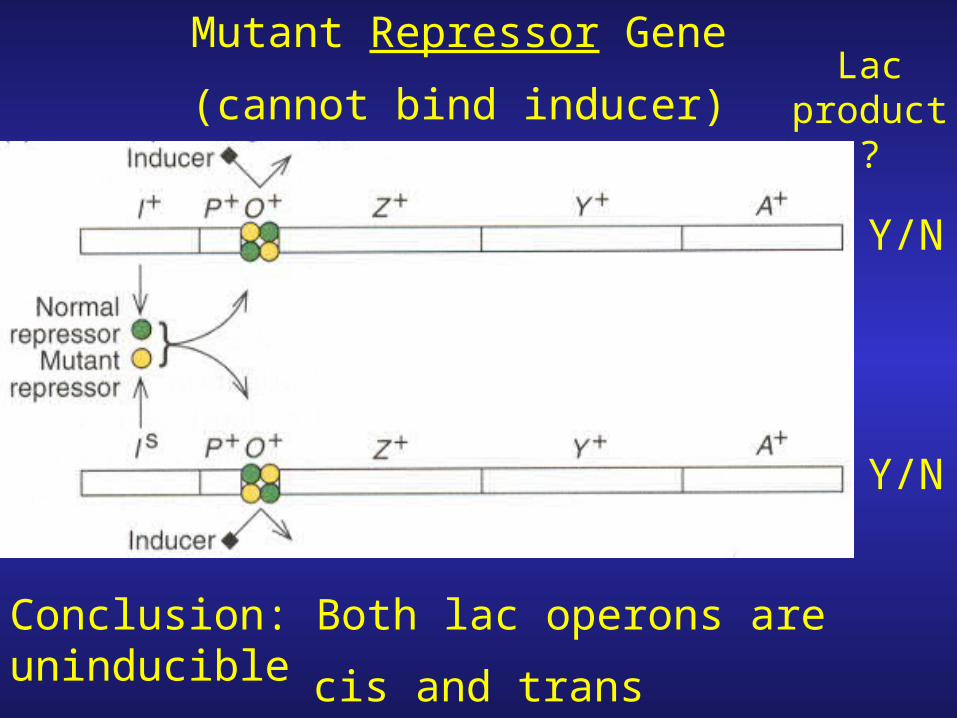
Mutant Repressor Gene
(cannot bind inducer)
Y/N
Y/N
Lac product
?
Conclusion: Both lac operons are uninduciblecis and trans dominant

Mutant Repressor Gene
Y/N
Y/N
Lac product
?
Conclusion: Both lac operons are non-repressible
(cannot bind operator sequence)
dominant-negative

Repression & Activation
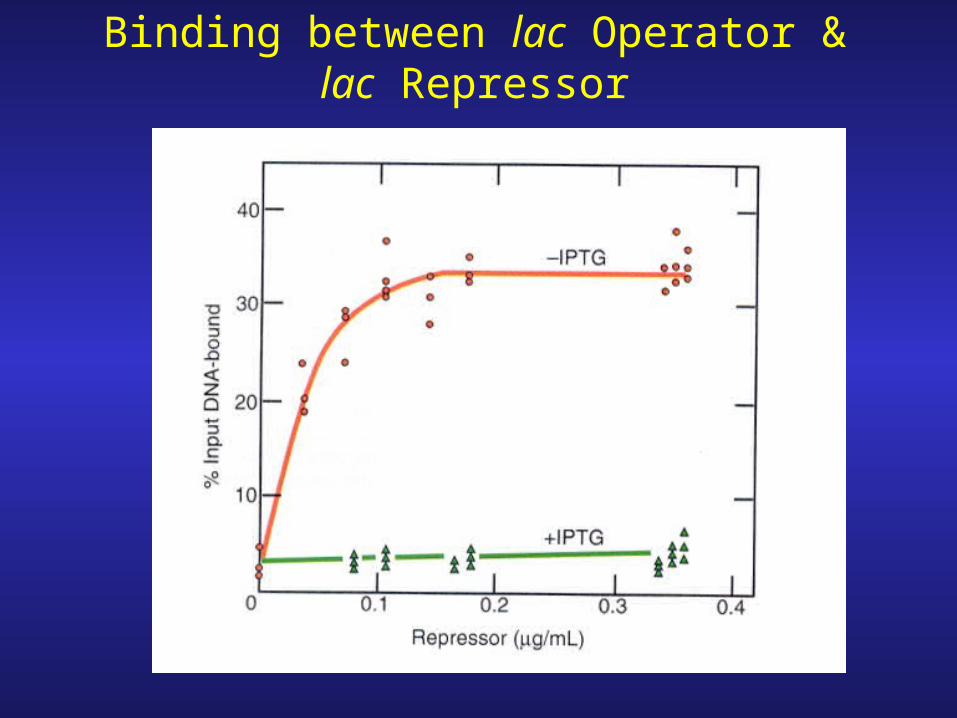
Binding between lac Operator & lac Repressor
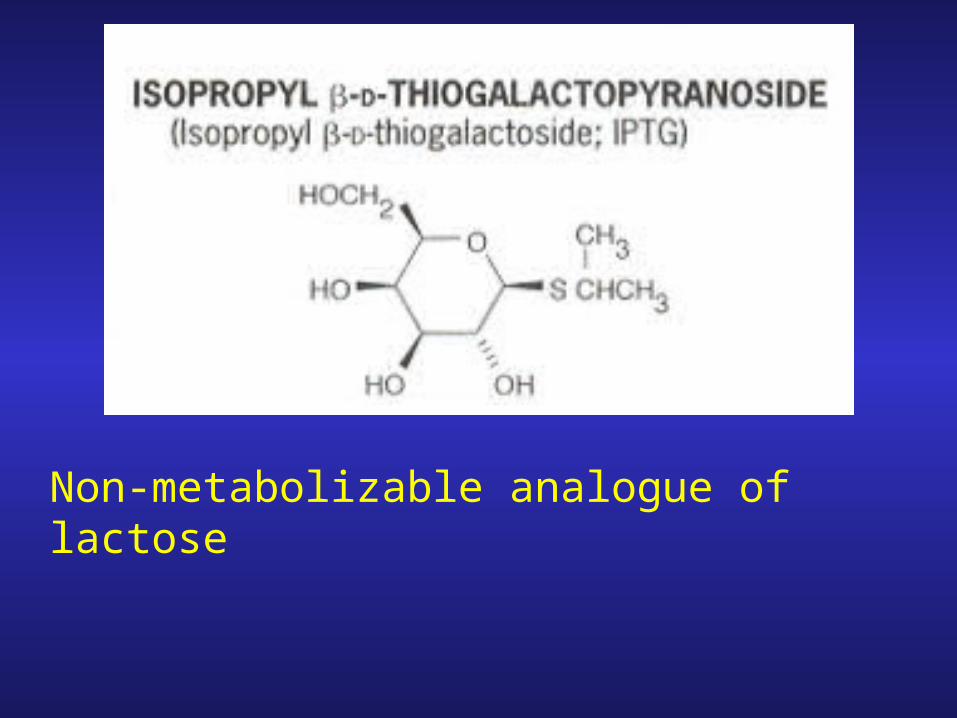
Non-metabolizable analogue of lactose
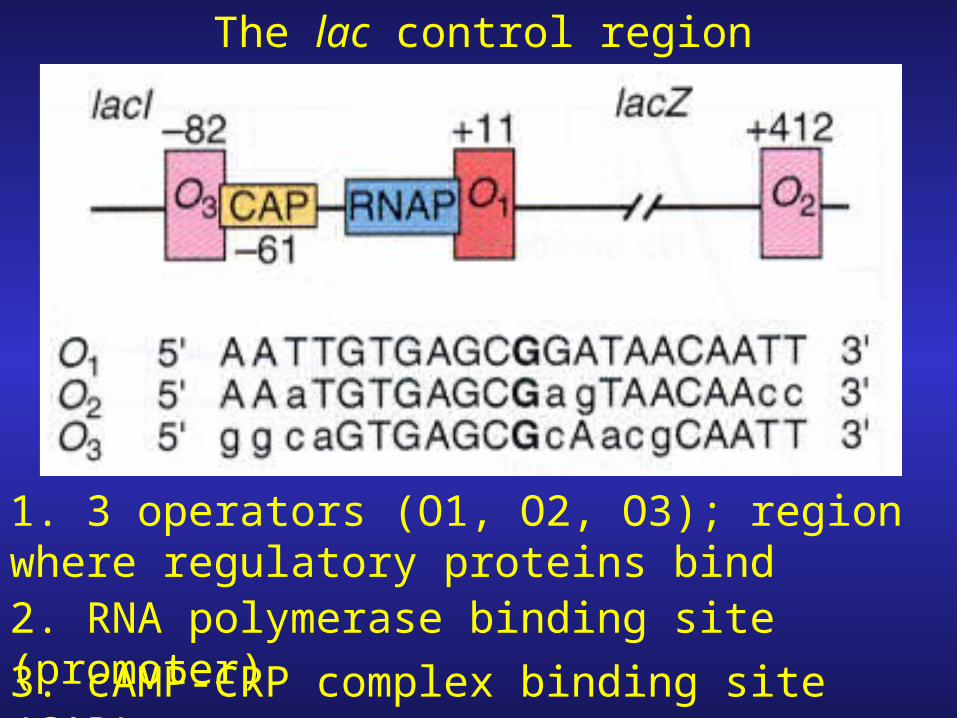
The lac control region
1. 3 operators (O1, O2, O3); region where regulatory proteins bind2. RNA polymerase binding site (promoter)
3. cAMP-CRP complex binding site (CAP)

b-Galactosidase Activity
1. Recall that the first gene in the lac operon is lacZ (b-galactosidase)
2. Enzyme activity can easily be measured using X-Gal or p-nitrophenol-galactoside (colorimetric assays that can be quantified)
3. Therefore effects on regulation can be monitored by measuring b-galactosidase activity.

Effects of Mutations in the 3 lac Operators

Positive Control of the lac Operon1. Removal of repressor is NOT enough to
activate the operon.2. The lac operon has a mechanism for reponding to glucose levels.
Why? – (i) When glucose levels are high, the cell wants to repress transcription of other operons (lactose)
(ii) When glucose levels are low & lactose present upregulate lac operon
Catabolite repression selection in favor of glucose metabolism

ATP
Adenylcyclase
Inhibited by glucose
Cyclic AMP
-cAMP responds to glucose conc.
- glucose uptake lowers the quantity of cAMP by inhibiting the enzyme adenylcyclase.

1. Addition of cAMP overcomes catabolite repression.
2. The activator is a complex between cAMP and a protein: catabolite activator protein (CAP) aka cAMP receptor protein (CRP) gene crp.
3. A mutant CRP protein with 10 lower affinity for cAMP: if cAMP-CRP complex important for activation then mutant should have reduced production of b-galactosidase
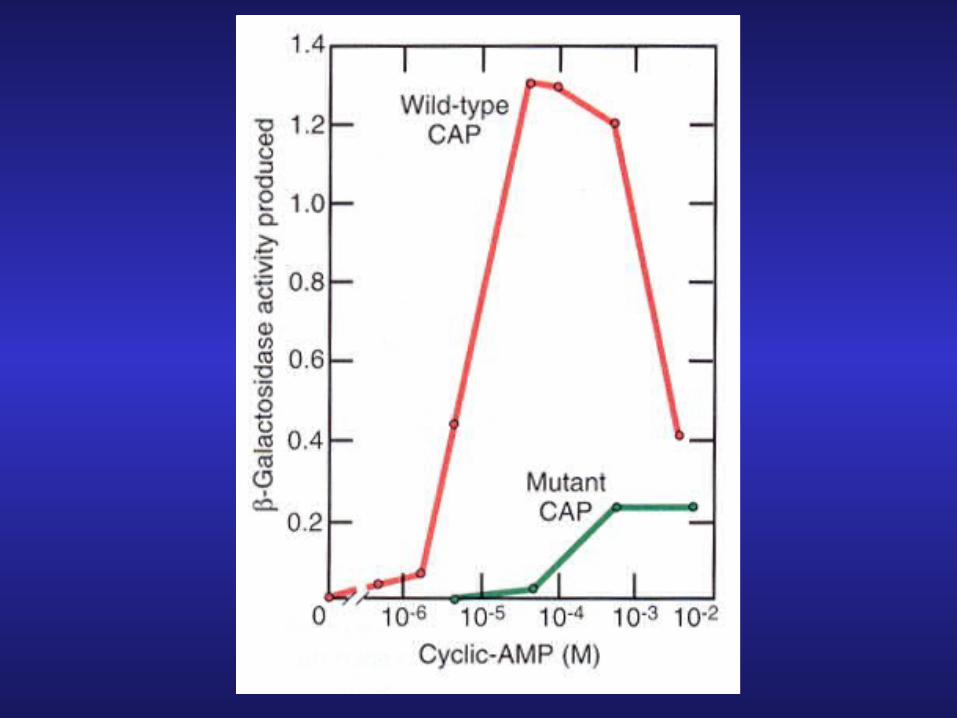

The Molecular Mechanism of c-AMP-CRP Action

1. cAMP-CRP complex stimulates transcription by binding to (activator) site adjacent to promoter.
2. cAMP-CRP recruits and helps RNA polymerase to bind to the promoter.
3. Recruitment has two steps:-formation of closed promoter complex-conversion of closed promoter complex to open promoter complex
increases rate of open promoter complex formation
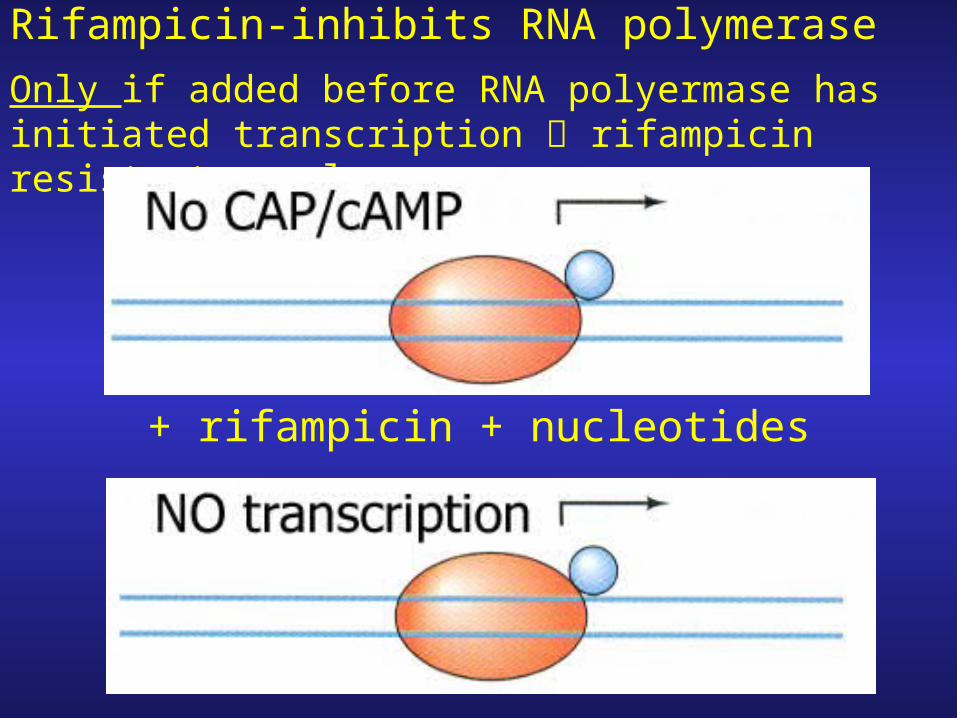
Rifampicin-inhibits RNA polymerase
Only if added before RNA polyermase has initiated transcription rifampicin resistant complex
+ rifampicin + nucleotides
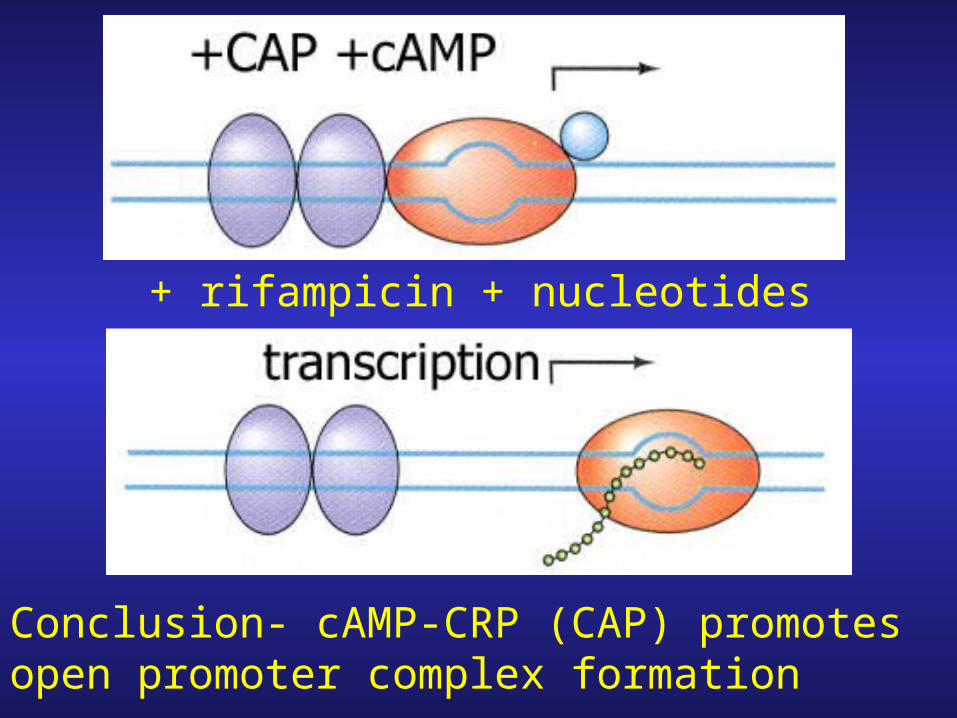
+ rifampicin + nucleotides
Conclusion- cAMP-CRP (CAP) promotes open promoter complex formation

How does cAMP-CRP binding to the activator site facilitate binding
of polymerase to the promoter?
1. cAMP-CRP complex “touches” the polymerase cooperative binding
2. cAMP-CRP causes the DNA to bend.
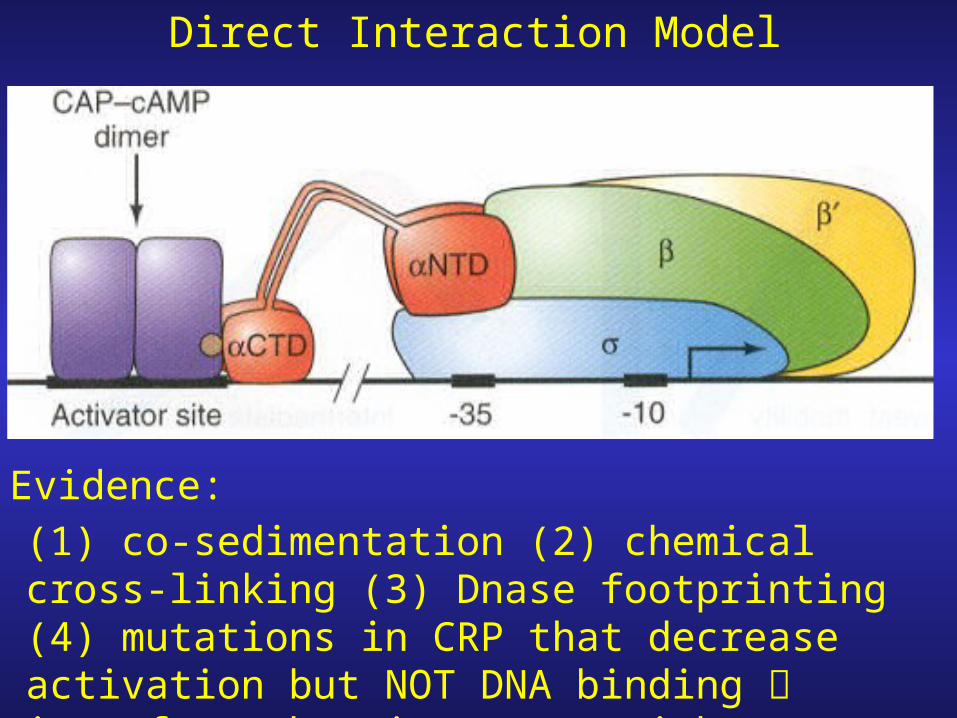
(1) co-sedimentation (2) chemical cross-linking (3) Dnase footprinting (4) mutations in CRP that decrease activation but NOT DNA binding interface that interacts with polymerase.
Direct Interaction Model
Evidence:
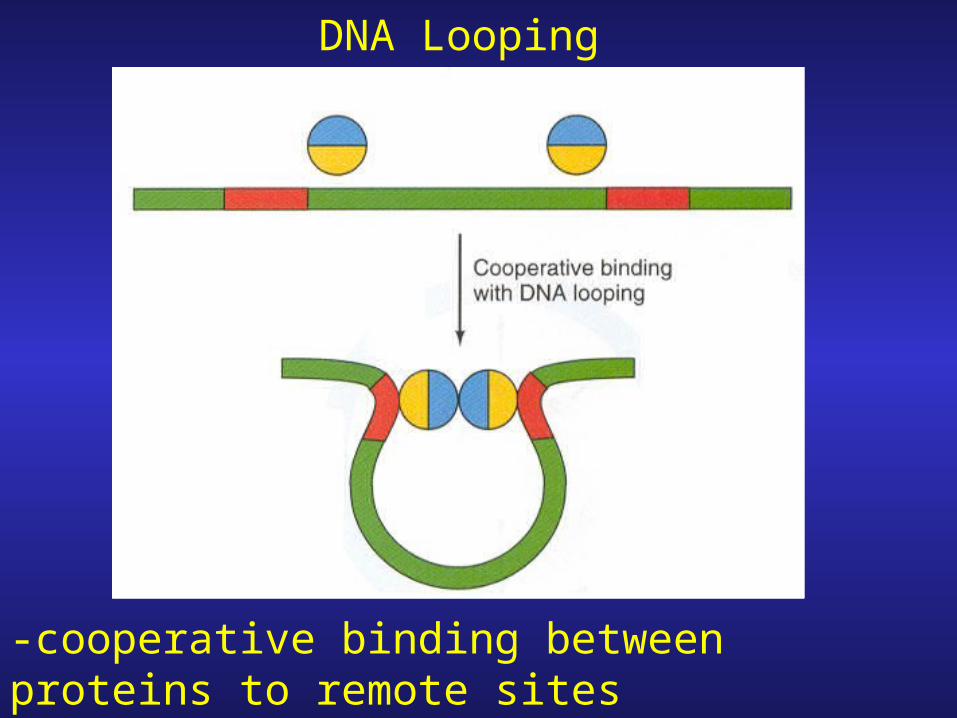
DNA Looping
-cooperative binding between proteins to remote sites
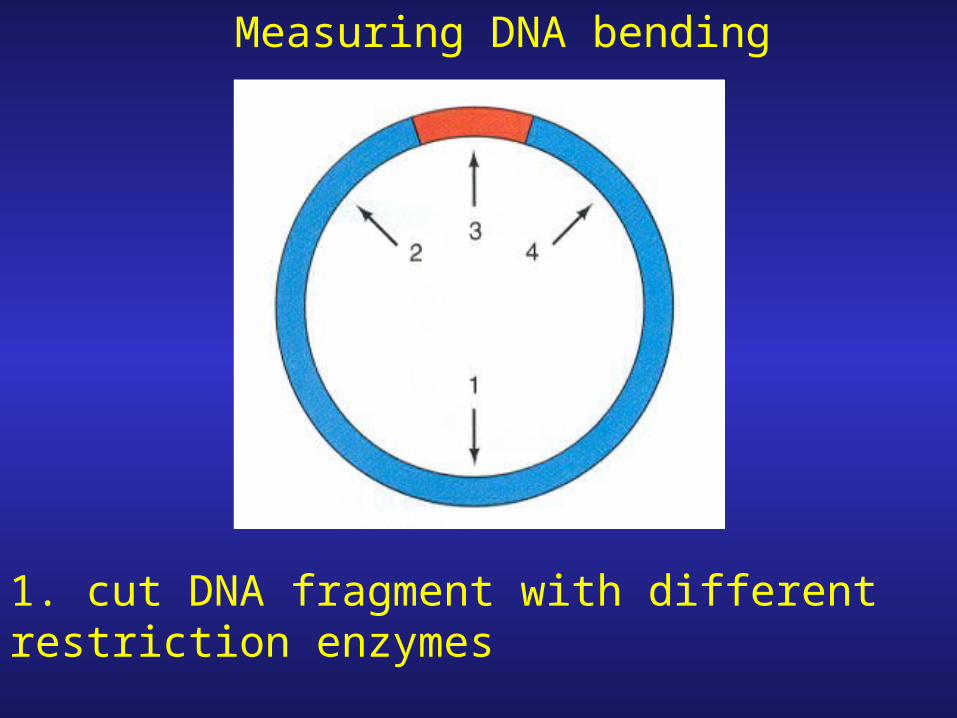
Measuring DNA bending
1. cut DNA fragment with different restriction enzymes
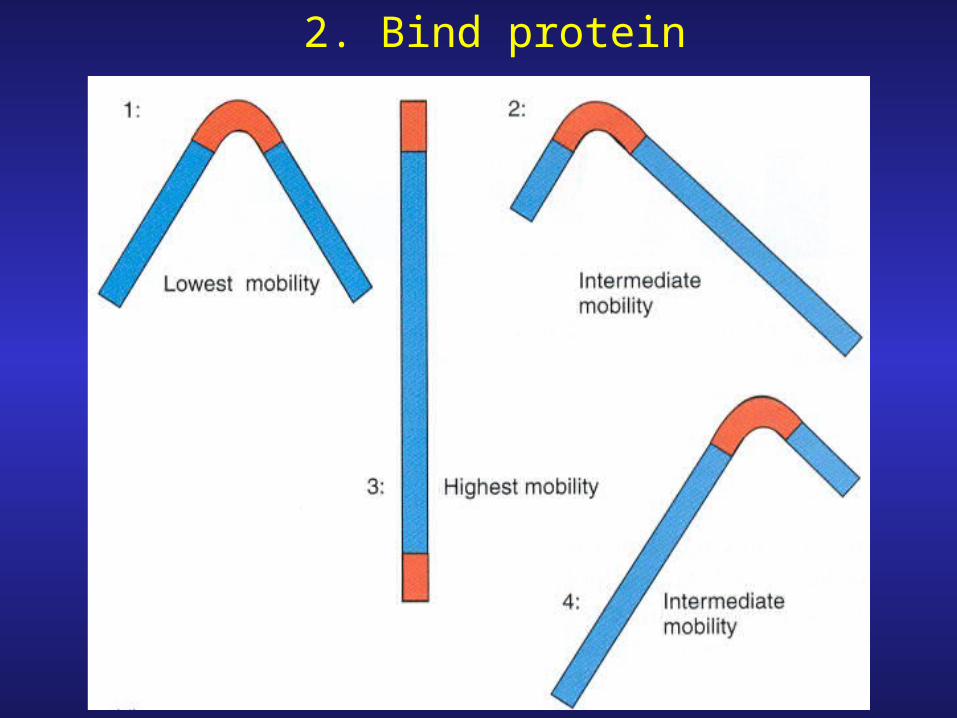
2. Bind protein
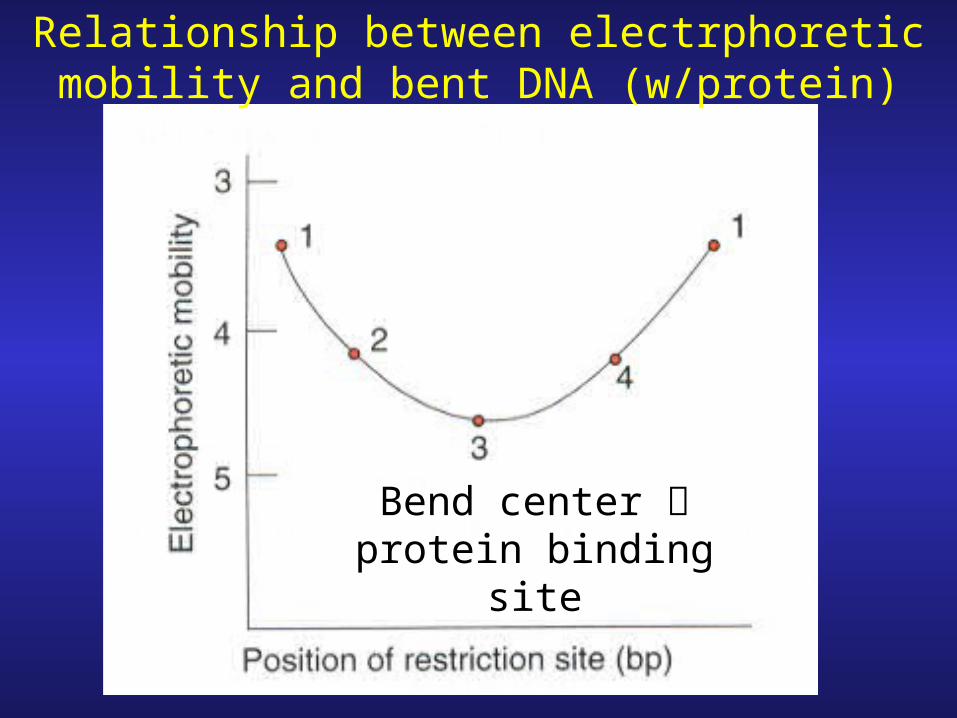
Relationship between electrphoretic mobility and bent DNA (w/protein)
Bend center protein binding site
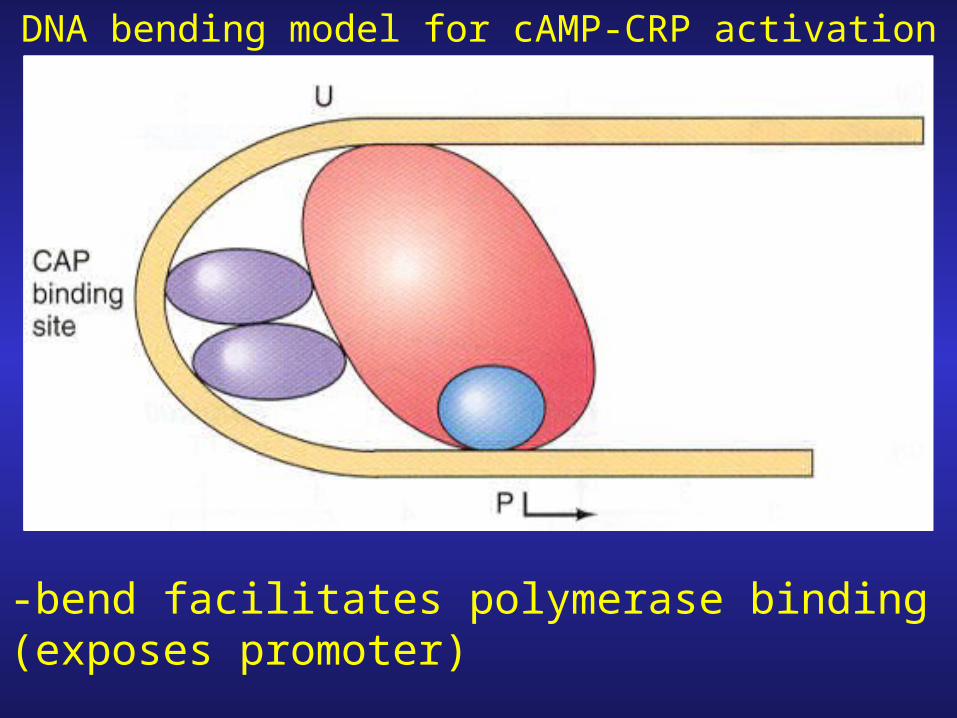
DNA bending model for cAMP-CRP activation
-bend facilitates polymerase binding (exposes promoter)

Mechanism of Repression
1. Assumption: repressor blocks polymerase access to promoter.
2. Experimental evidence, however, has shown that RNA polymerase can STILL bind to promoter in the presence of repressorRifampicin no transcription unless open promoter complex has formed
Experiment 1: DNA, polymerase, repressor
add inducer, nucleotides, & rifampicinResult :Transcription occurred repressor had not prevented formation of open complex
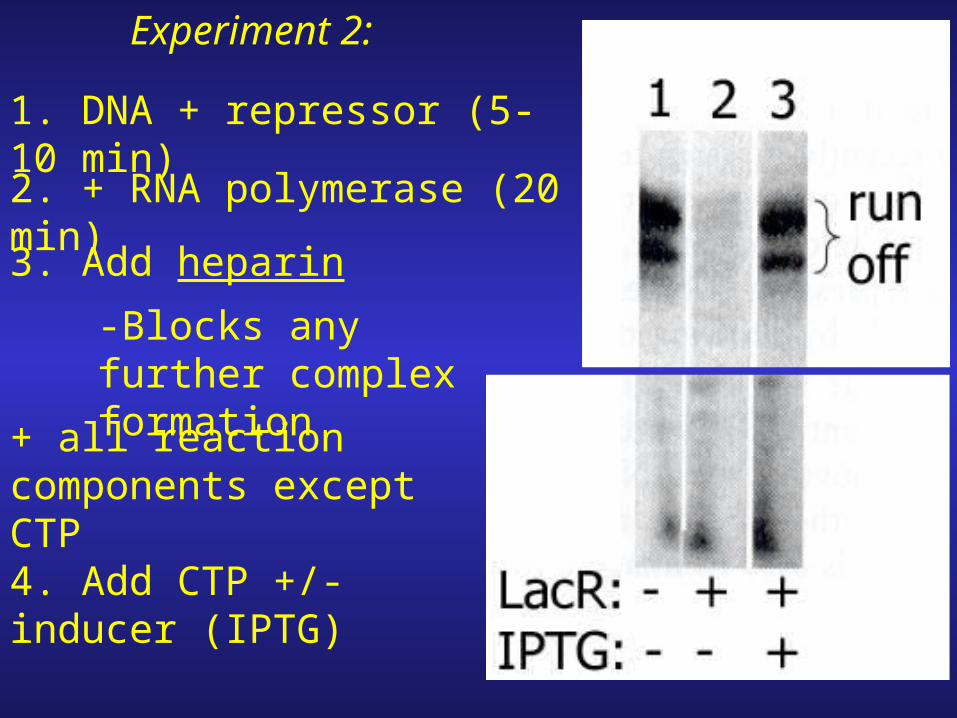
Experiment 2:
1. DNA + repressor (5-10 min)2. + RNA polymerase (20 min)3. Add heparin
-Blocks any further complex formation
+ all reaction components except CTP
4. Add CTP +/- inducer (IPTG)

-sulfated glycosoaminoglycan (chain)
-joints, vitreous humor
-viscosity increasing agent, anti-coagulant
-binds RNA polymerase inhibiting association with promoter

Further evidence showed that repressor and polymerase can bind together to lac operator.
If lac repressor does not inhibit transcription of the lac operon by
blocking access to promoter, how does it function?
Alternate theory: repressor locks RNA polymerase into a non-productive state.
Evidence: formation of abortive transcripts

HOWEVER…
More recent studies have shown that repressor/polymerase : operator interactions are in equilibrium.
Ratio of: polymerase-promoter complex and free polymerase/free promoter
And that previous experiments were simply shifting or locking this equilibrium association

Experiment:1. Add RNA polymerase + lac promoter
(used fluorescent labeled UTP analog)
2. (1) no addition (2) + heparin
(3) + repressor (4) no DNA
Analysis: (i) heparin known to prevent polymerase (re)-association
(ii) If repressor does not block access to polymerase it should not inhibit polymerase association with promoter
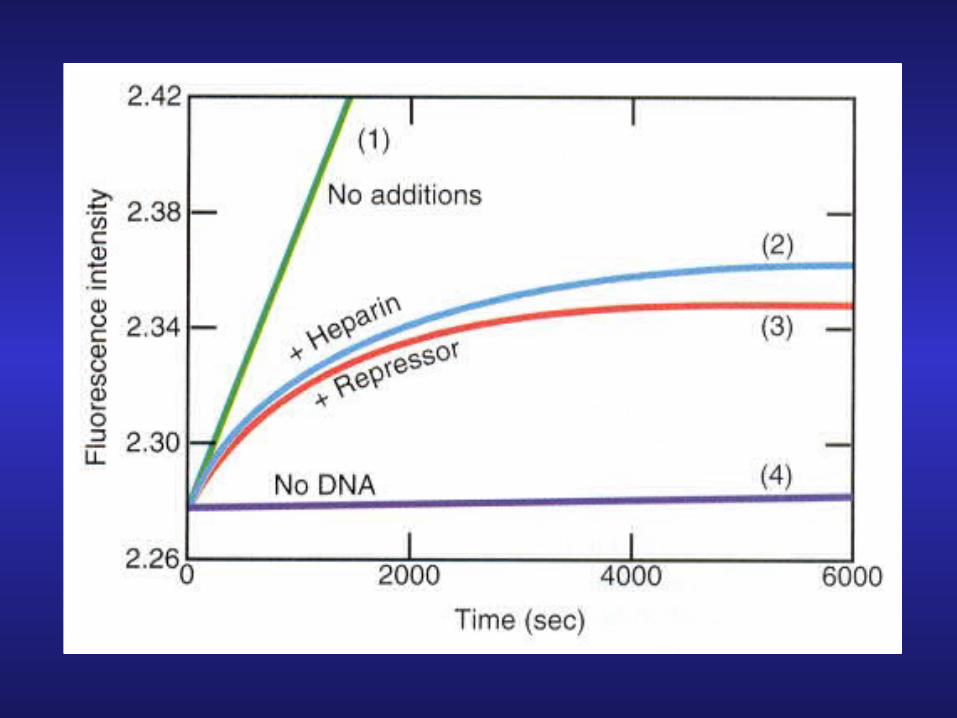

Conclusion: Original competition hypothesis may be correct!
Result: both heparin and repressor inhibits (re)-association of polymerase with promoter.
Analysis: (1) heparin binds polymerase preventing association with DNA
(2) repressor does the same by binding to the operator adjacent to the promoter and blocking access to the promoter by RNA polymerase.

Maltose Operon

1. mal regulon regulated by CRP
2. MalT also regulates the mal promoters
-requires ATP
-activated by inducer (maltotriose)
-Some mal promoters malEp & malKp use both CRP and MalT
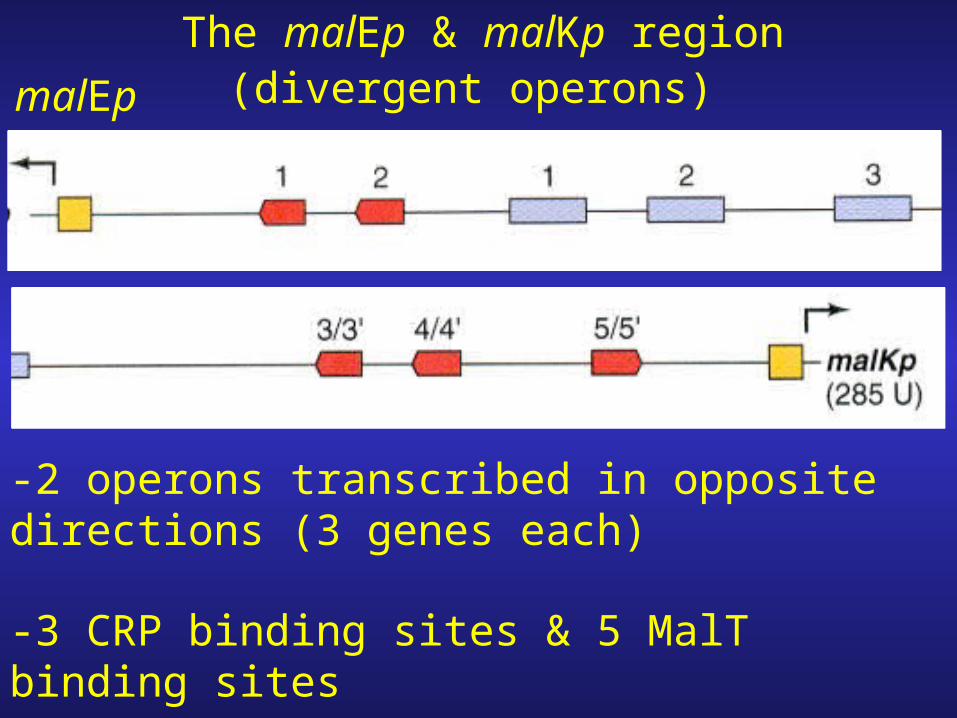
malEp
The malEp & malKp region
-2 operons transcribed in opposite directions (3 genes each)
-3 CRP binding sites & 5 MalT binding sites
(divergent operons)
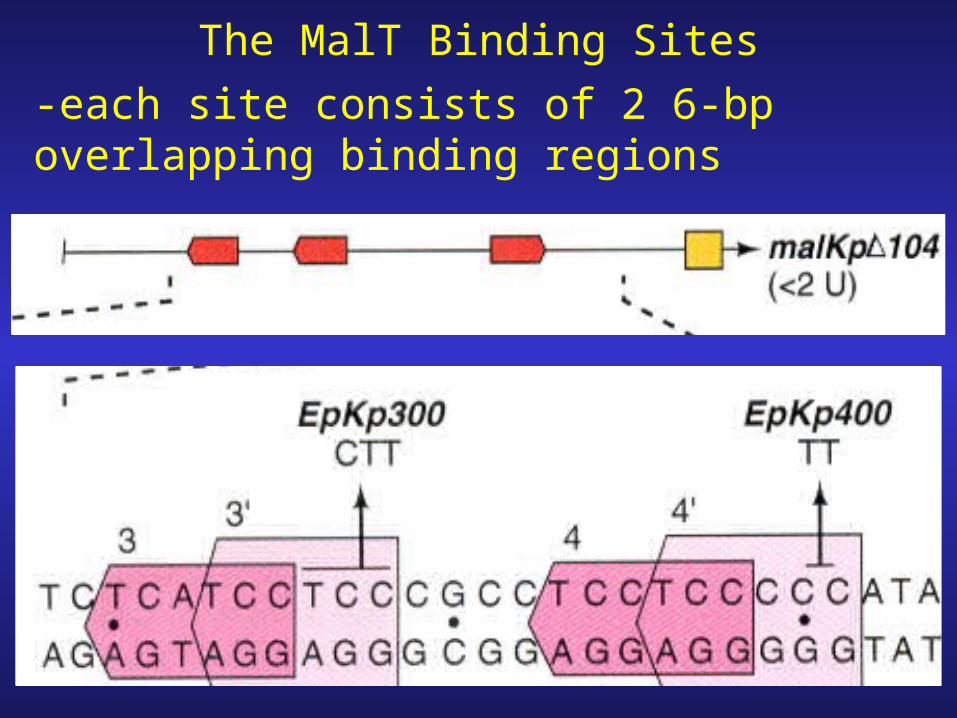
The MalT Binding Sites
-each site consists of 2 6-bp overlapping binding regions

-the third site
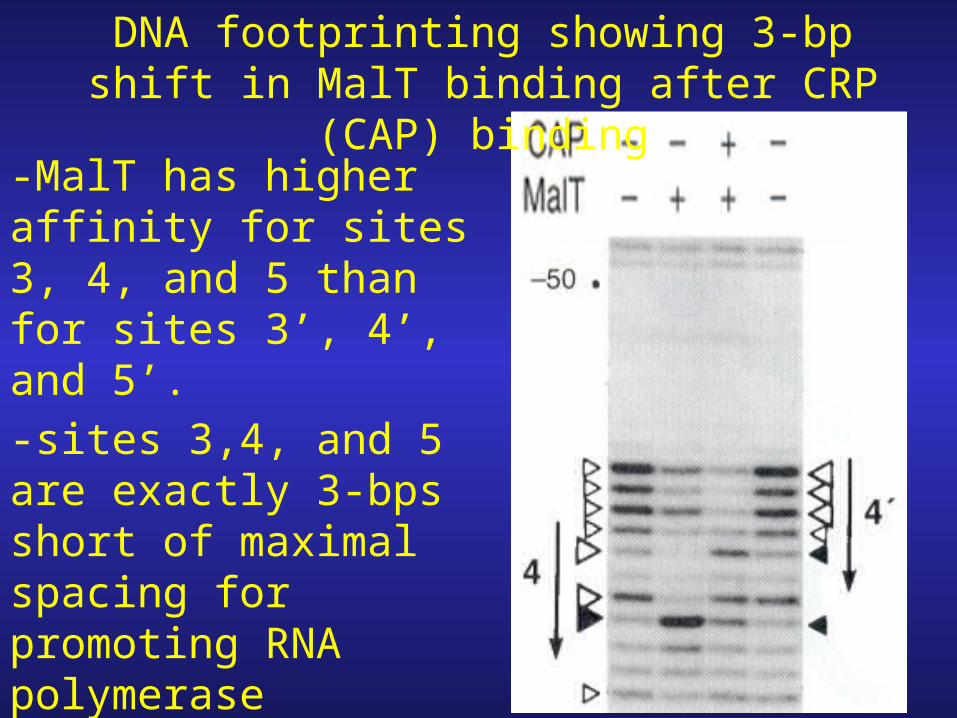
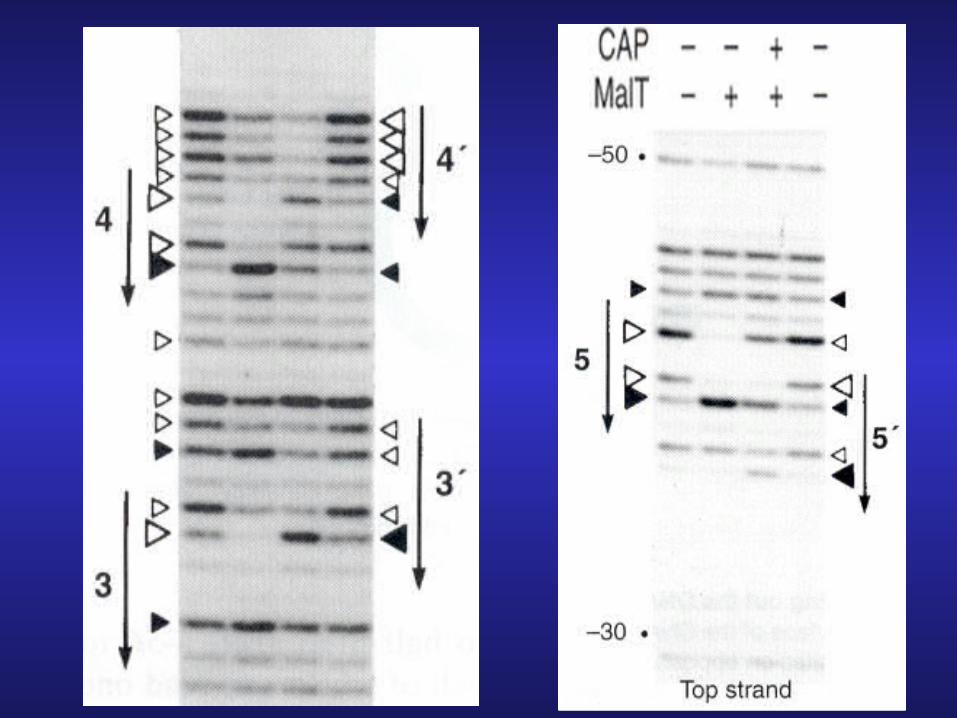
DNA footprinting showing 3-bp shift in MalT binding after CRP (CAP)
binding-MalT has higher affinity for sites 3, 4, and 5 than for sites 3’, 4’, and 5’.
-sites 3,4, and 5 are exactly 3-bps short of maximal spacing for promoting RNA polymerase binding.

Arabinose Operon
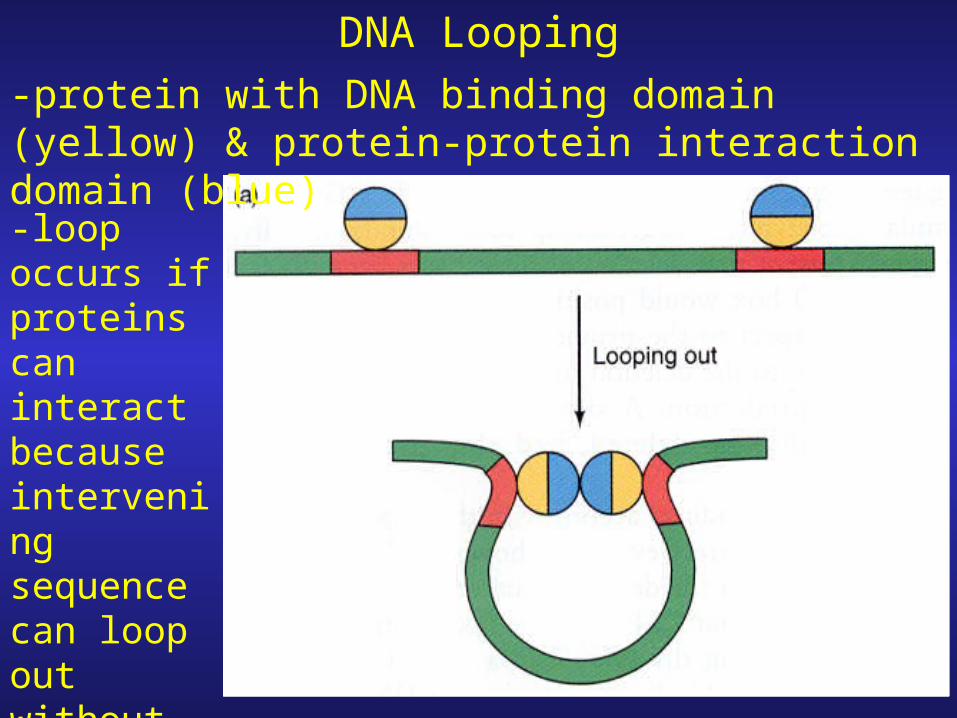
DNA Looping-protein with DNA binding domain (yellow) & protein-protein interaction domain (blue)
-loop occurs if proteins can interact because intervening sequence can loop out without twisting
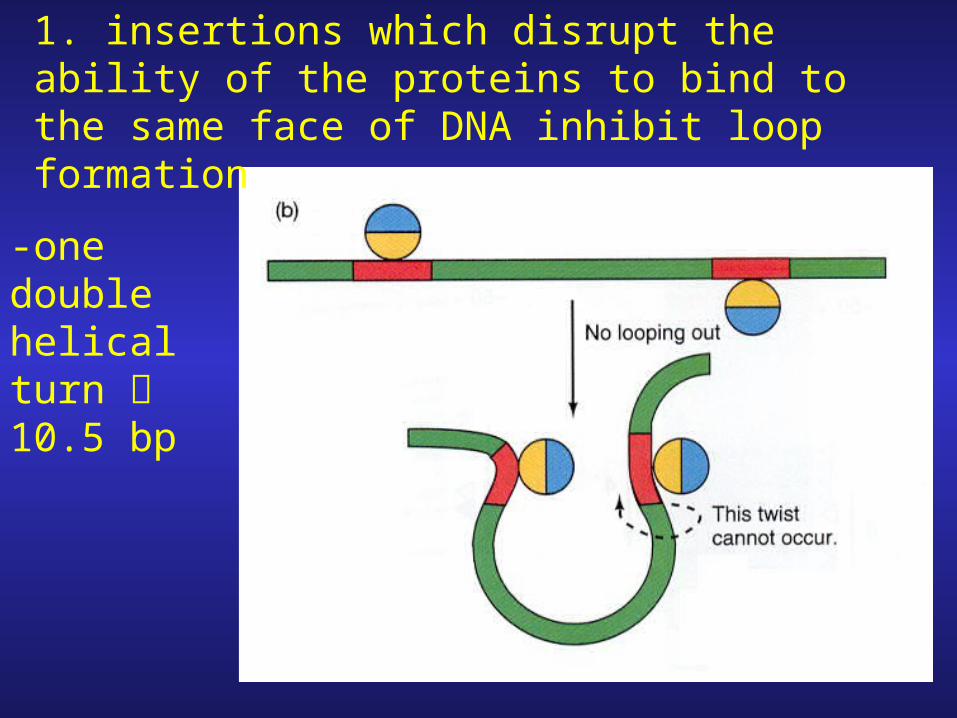
-one double helical turn 10.5 bp
1. insertions which disrupt the ability of the proteins to bind to the same face of DNA inhibit loop formation

1. Arabinose operon consists of 4 genes, 3 together transcribed in one direction (araPBAD), the fourth araC divergent (araPc)
2. AraC is the control protein, acts as repressor or activator depending upon binding conditions.
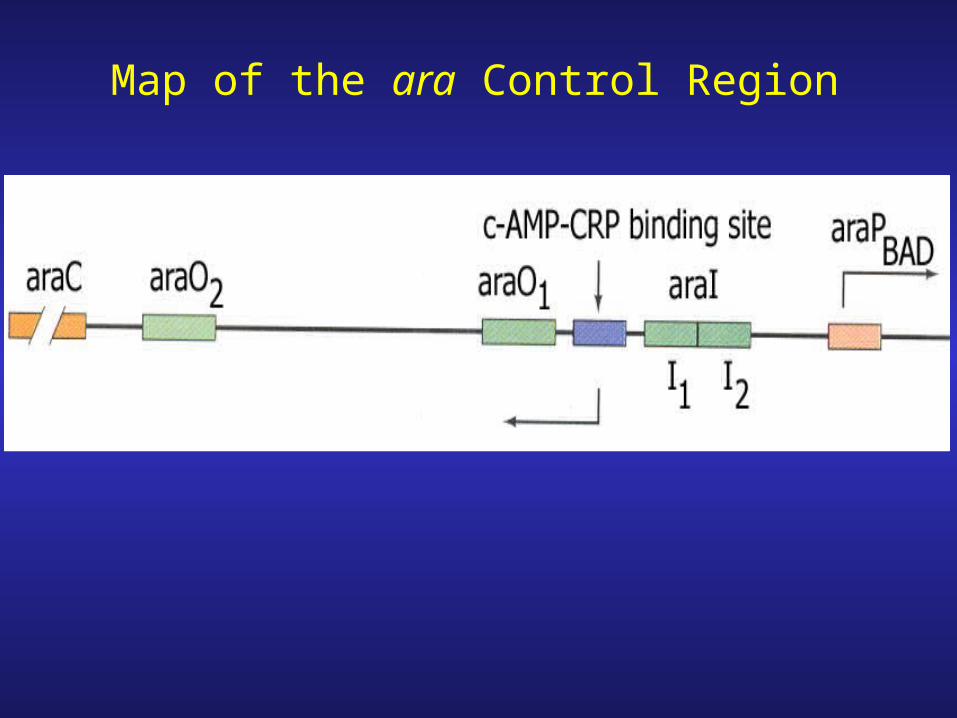
Map of the ara Control Region

Negative control- monomers of AraC bind to O2 and I1 looping out the intervening sequence (210 bp) & blocking access to the promoter by RNA polymerase
Absence of Arabinose

Positive Control1. Arabinose binds to AraC results in conformational change in AraC.
2. Arabinose-AraC complex preferentially binds to I2/I1 sequences (over O2/I1 sequence)
3. Promoter accessible to RNA polymerase
4. cAMP-CRP present (glucose absent) bind to Pc site transcription stimulated

Experimental Evidence of Looping
1. Observed by electron microscopy
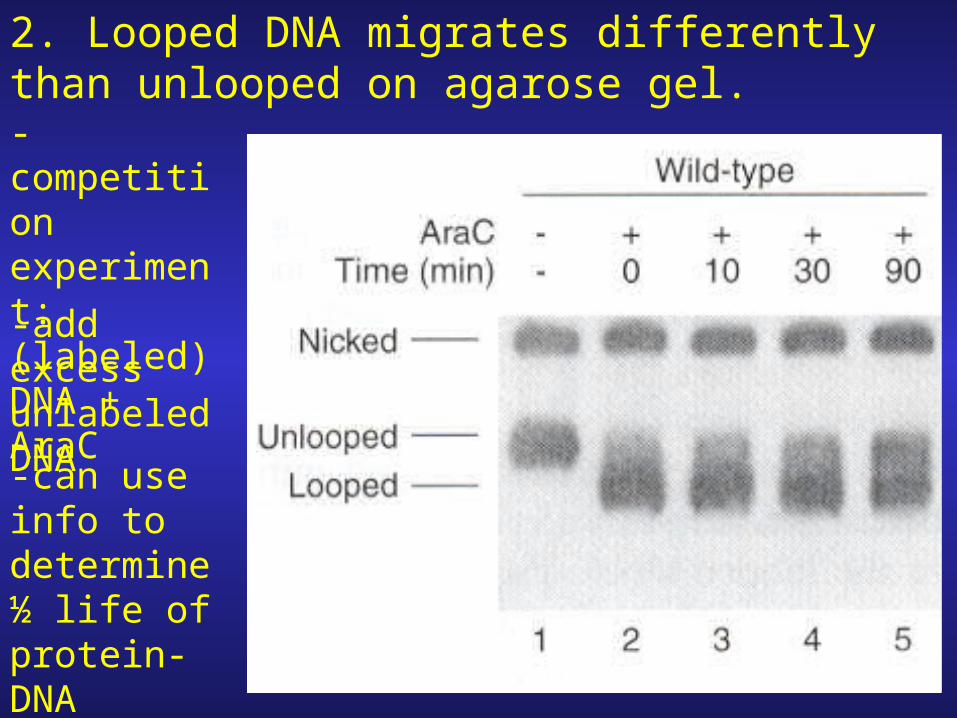
2. Looped DNA migrates differently than unlooped on agarose gel.-competition experiment: (labeled) DNA + AraC
-add excess unlabeled DNA
-can use info to determine ½ life of protein-DNA interaction
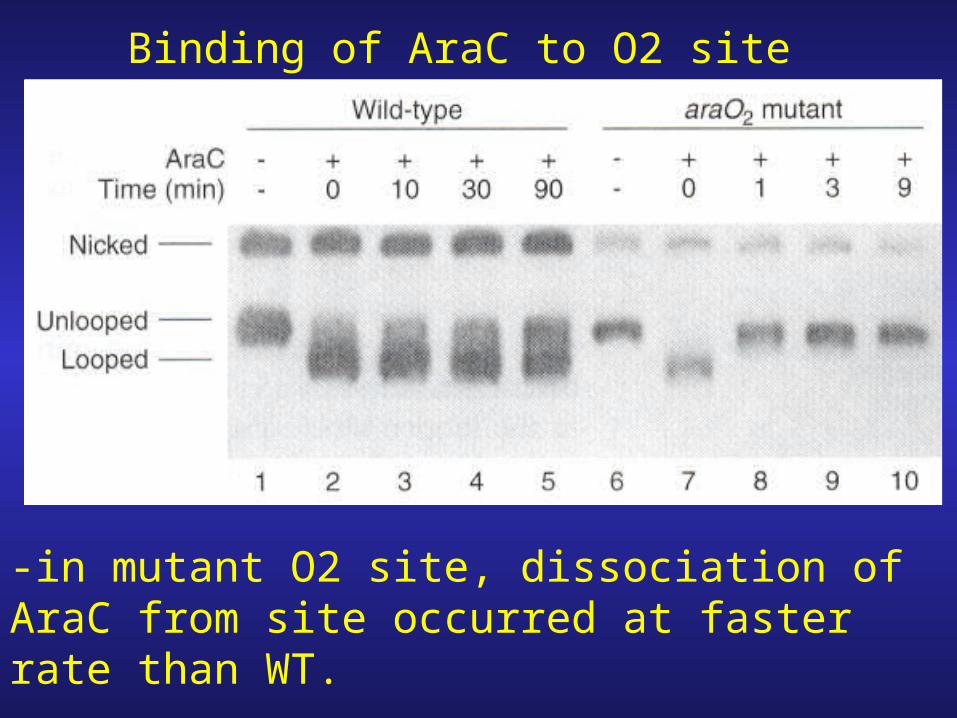
Binding of AraC to O2 site
-in mutant O2 site, dissociation of AraC from site occurred at faster rate than WT.
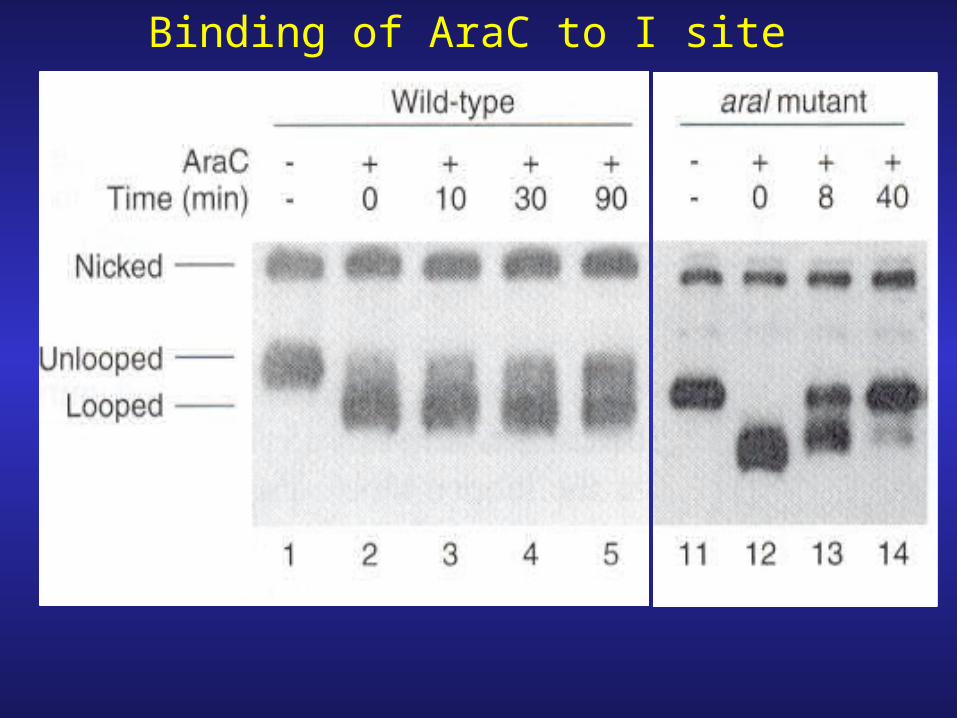
Binding of AraC to I site
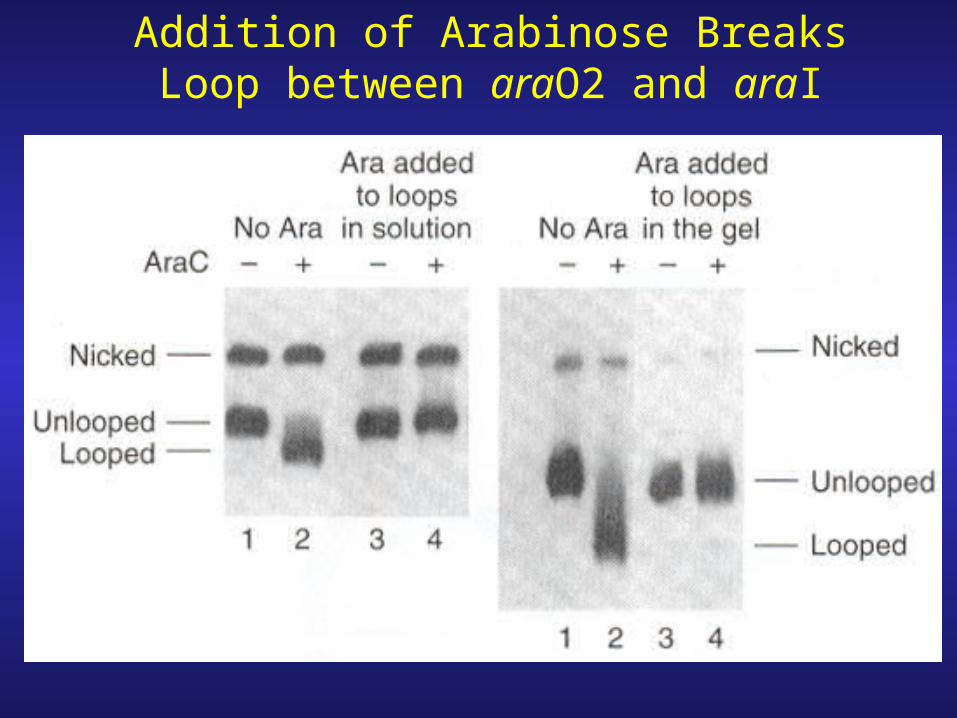
Addition of Arabinose Breaks Loop between araO2 and araI

Notes on Regulation of the Arabinose Operon
1. Looping/unlooping is reversible. Add AraC loop forms, add arabinose loop breaks, remove arabinose (dilution) loop reforms (in presence of AraC
2. AraC contacts I2 in the unlooped state but not in the looped complex.
3. A single dimer of AraC is sufficient for loop formation
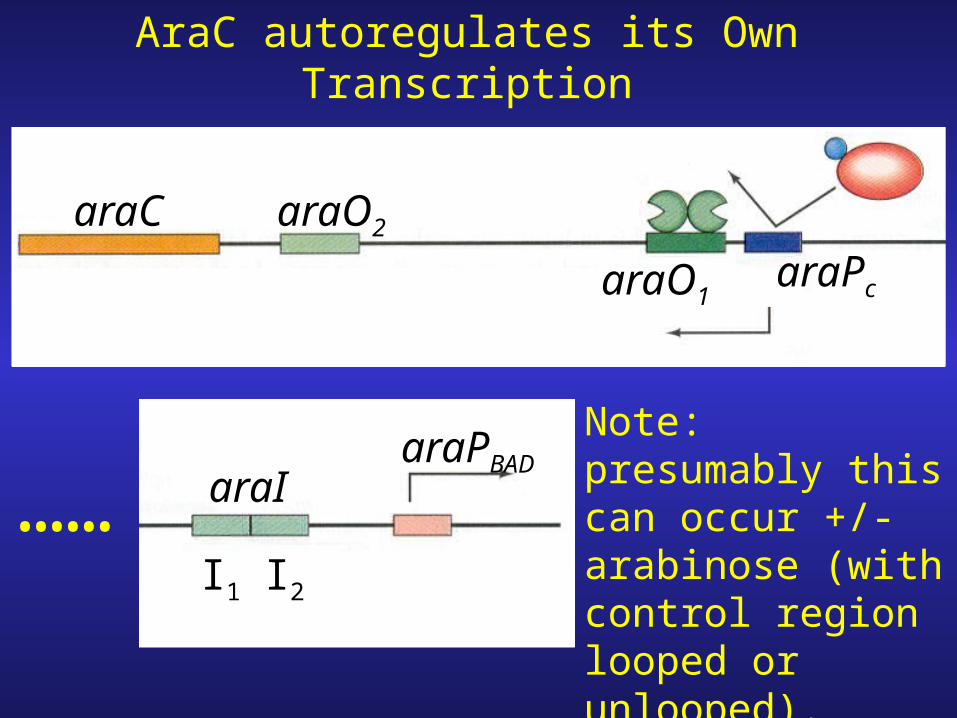
araPc
……I1 I2
araPBADaraI
araO1
araC araO2
AraC autoregulates its Own Transcription
Note: presumably this can occur +/- arabinose (with control region looped or unlooped).

Conclusions
I. Maltose Operon.
1. Mal operon controlled by CRP & MalT (transcription factor)
2. CRP stimulates transcrption by shifting MalT from one set of binding sites to another (only 3 bp away)3. Initial binding site of MalT is poorly aligned with (enhancing transcription from) the promoters
4. The “secondary” sites are better aligned with respect to the promoters and hence can facilitate transcription.
I. Arabinose Operon.
1. Ara operon controlled by AraC.
2. AraC rpresses operon by looping out the DNA between sites araO2 and araI1 (210 bp apart)
3. Arabinose derepresses the operon by causing AraC to loosen its attachment to araO2 and to bind to araI2 instead.
(beaks loop, allowing transcription)
4. cAMP-CRP further stimulates transcription by binding to a site upstream of araI.
5. AraC regulates its own transcription by binding to araO1 and preventing (leftward) transcription of the araC gene.

Tryptophan Operon
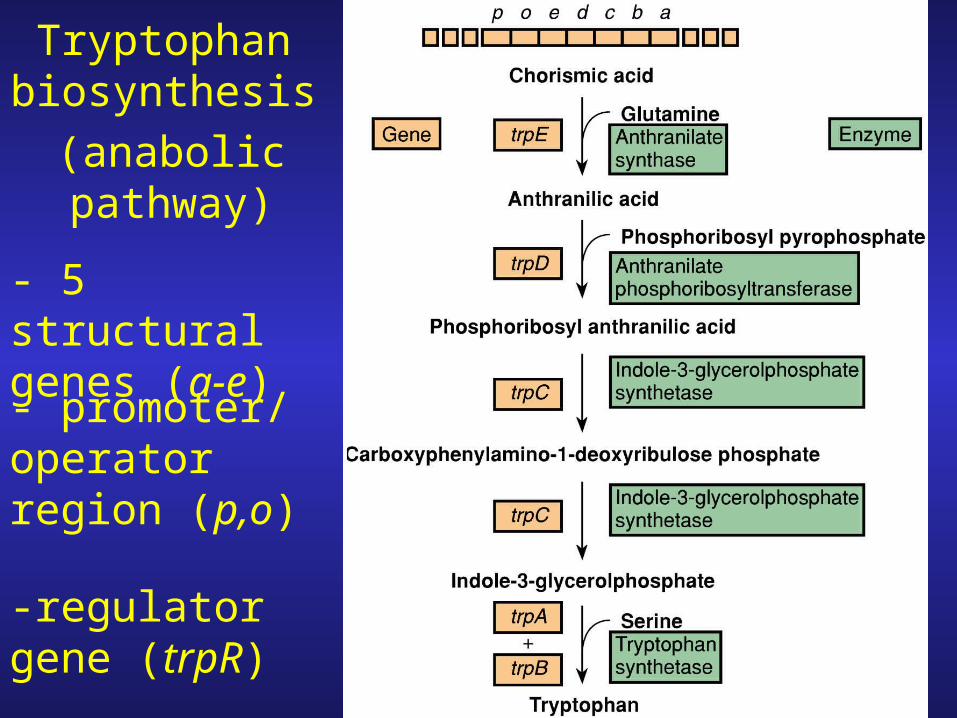
Tryptophan biosynthesis
- 5 structural genes (a-e)
- promoter/ operator region (p,o)
-regulator gene (trpR)
(anabolic pathway)
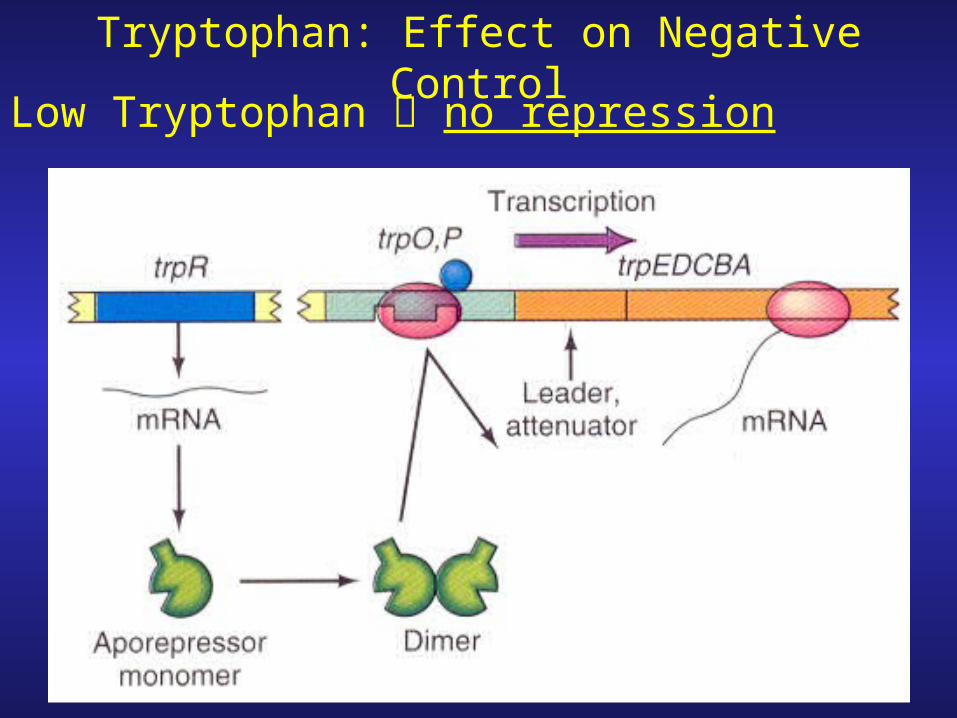
Tryptophan: Effect on Negative Control
Low Tryptophan no repression
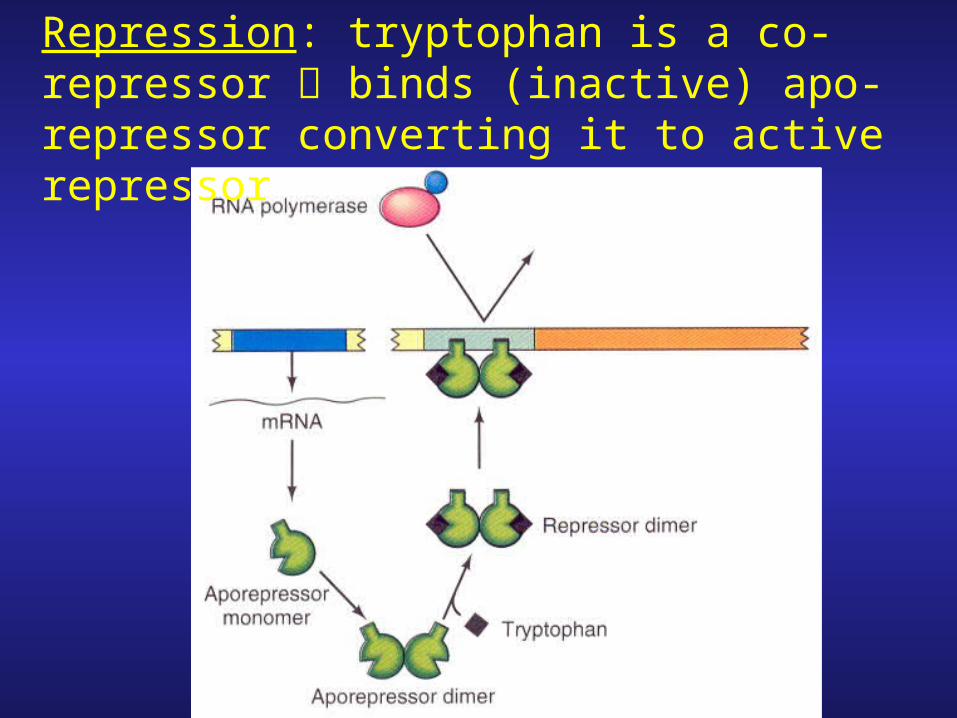
Repression: tryptophan is a co-repressor binds (inactive) apo-repressor converting it to active repressor
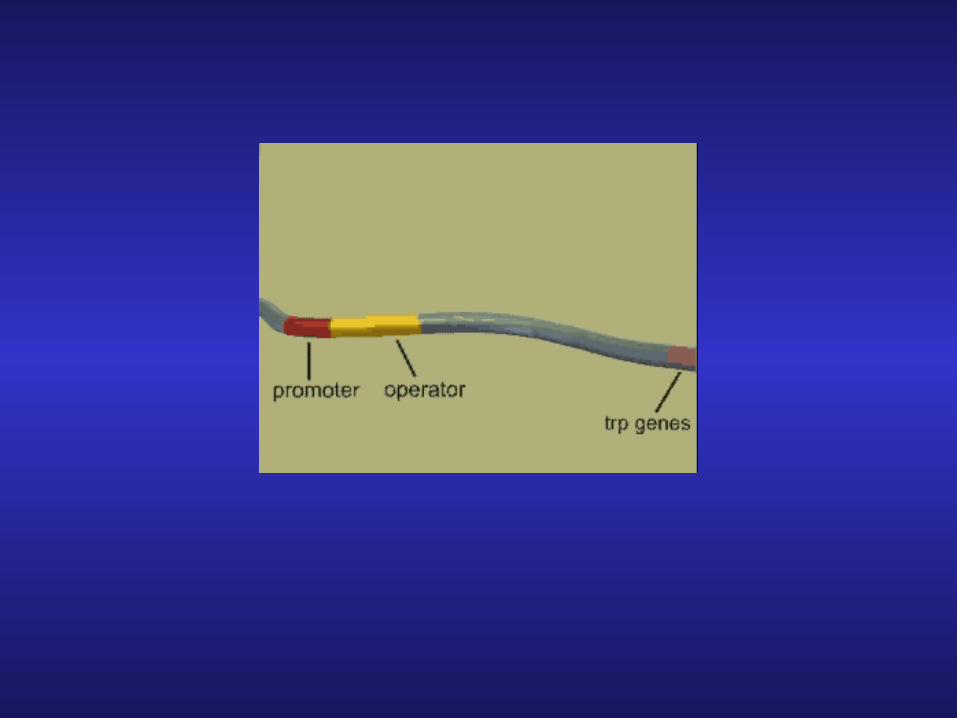
1. Operator site lies within the promoter
2. Allosteric transitionAllosteric protein-protein whose shape is changed upon binding of a particular molecule In the new conformation the protein’s ability to react to a second molecule is altered
3. Trp operon has another level of control attenuation
4. Repressor lowers transcription 70-fold (as compared to derepressed state) attentuation permits another 10-fold control total dynamic range of control = 700-fold
…
…
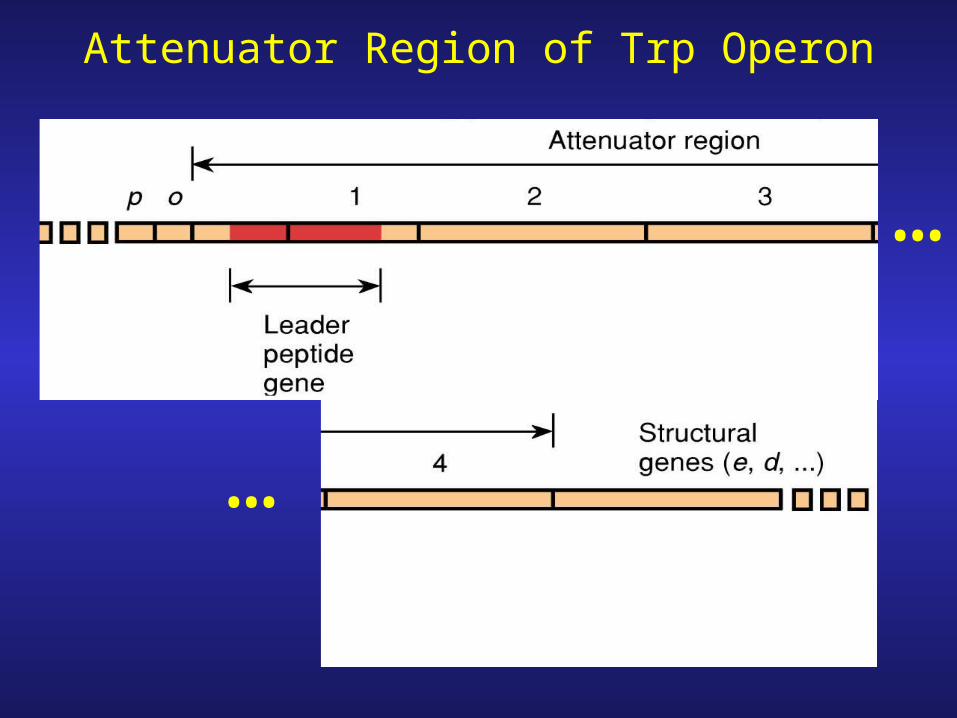
Attenuator Region of Trp Operon

Low tryptophan: transcription of trp operon genes RNA polymerase reads through attenuator.

High tryptophan: attenuation, premature termination attenuator causes premature termination of transcription
1. Attenuator region contains transcription stop signal (terminator) not STOP codon!
2. The terminator consists of an inverted repeat followed by string of eight A-T pairs.
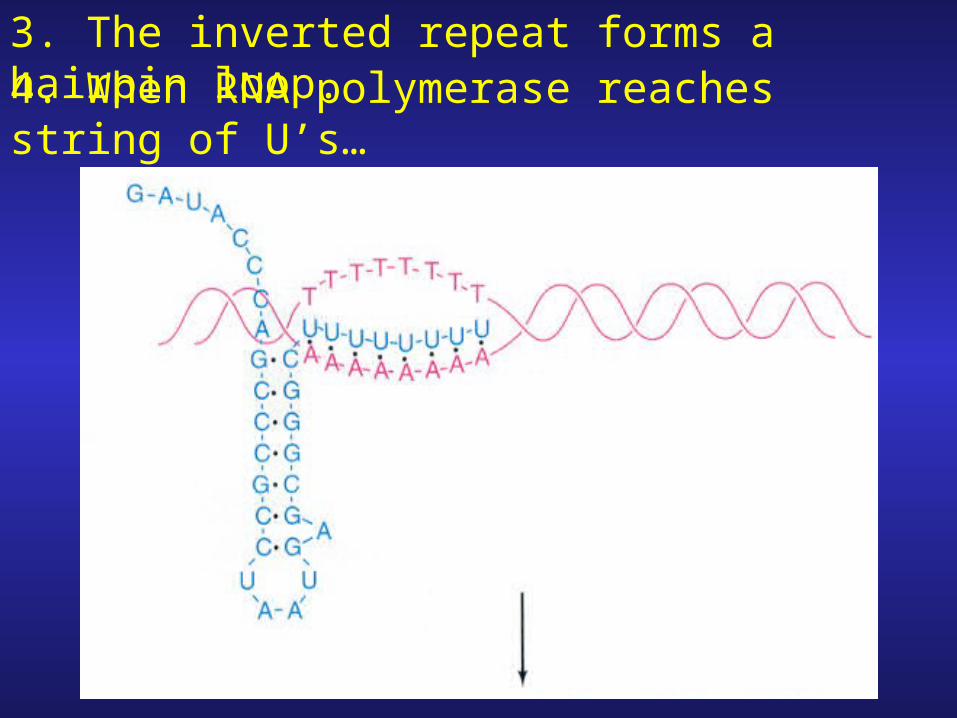
3. The inverted repeat forms a hairpin loop.4. When RNA polymerase reaches string of U’s…

…the polymerase pauses, the hairpin forms Transcript is released
Termination occurs before transcription reaches the trp (structural) genes

Attenuation gives some insight into how the operon is shut down, but how does the cell activate trp operon expression (i.e. defeat attenuation)?
preventing hairpin formation would destroy termination signal transcription would proceed

…
…
Mechanism of Attenuation
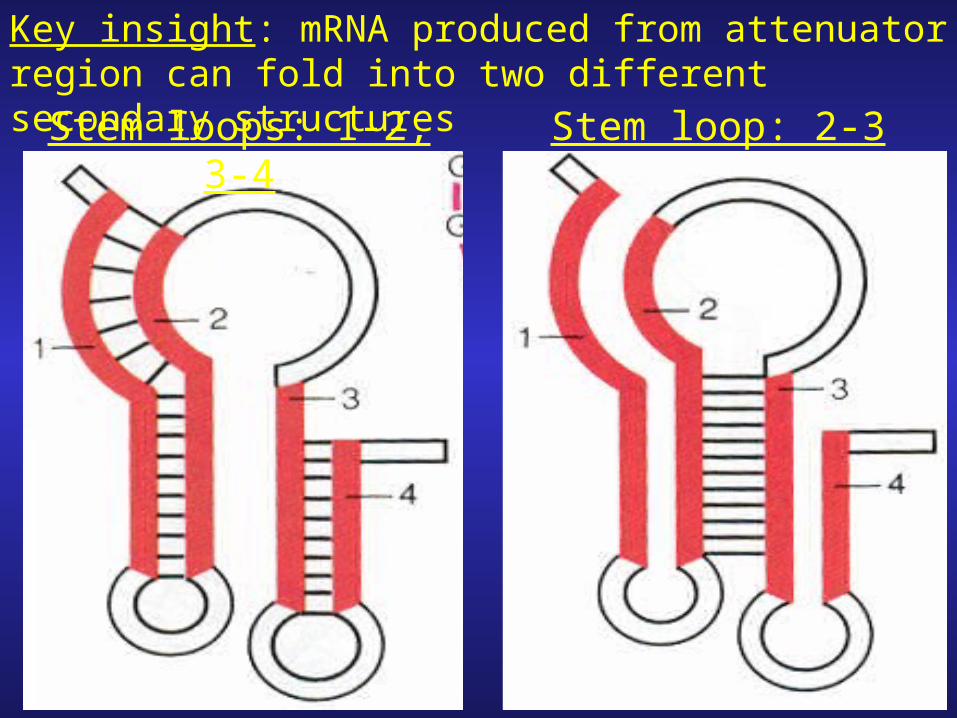
Key insight: mRNA produced from attenuator region can fold into two different secondary structuresStem loops: 1-2, 3-4 Stem loop: 2-3

1. Formation of stem loop structures; 1-2 and 3-4 is more stable and results in the formation of a termination (hairpin loop) structure/signal.
2. Formation of stem loop structure 2-3 would result in the disruption of stem loops 1-2/3-4.3. The stem loop structure formed between 2-3 does not result in termination signal transcription would proceed.
Q. becomes: How does the less stable structure (stem-loop 2-3) form?

The Importance of the Leader Region
-the 14 amino acid peptide formed from the leader sequence has 2 tryptophans.
-trp is a “rare” amino acid
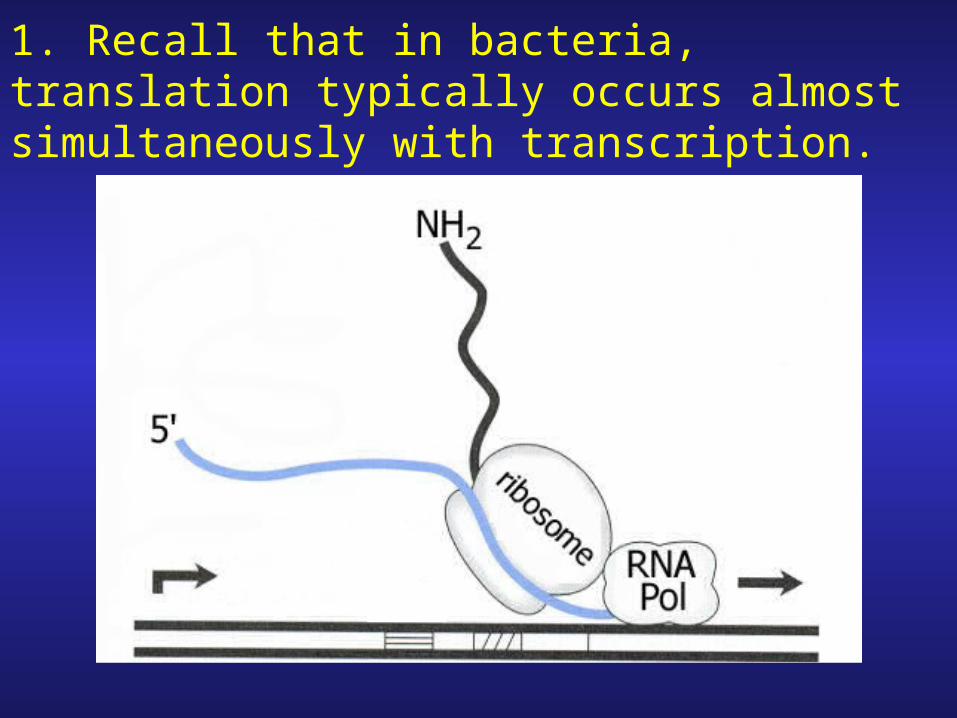
1. Recall that in bacteria, translation typically occurs almost simultaneously with transcription.

2. Thus, as soon as trp leader region is transcribed, translation begins.
3. During low tryptophan concentration, ribosome will stall at trp sites.
4. The trp site is right in the middle of region 1 of the attenuator
Meanwhile RNA polymerase continues to transcribe
Consider LOW Trp Conditions
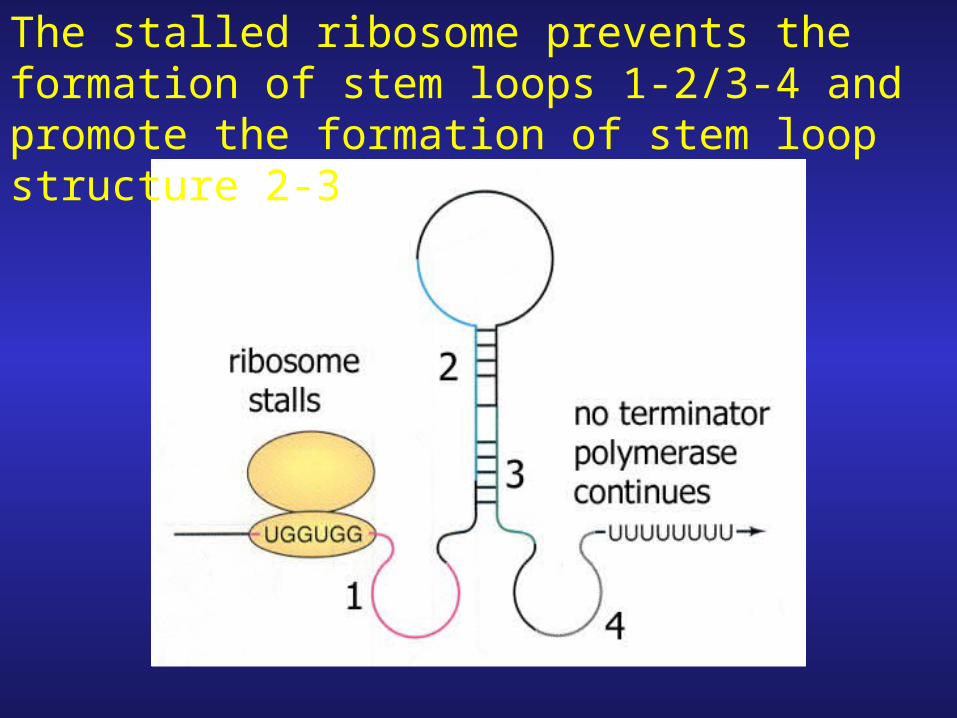
The stalled ribosome prevents the formation of stem loops 1-2/3-4 and promote the formation of stem loop structure 2-3

1. Stem loop structure 2-3 does not result in transcriptional termination whole operon mRNA made.
2. What happens to the stalled ribosome?
(i) Since the genes in the operon have their own start sites other ribosomes can come and translate those proteins
(ii) Stalled ribosome can eventually either incorporate trp-tRNA (+ 3 more a.a. before reaching stop codon) or dissociate from mRNA

At HIGH Trp Conditions
1. When high levels of Trp-tRNA are present the two tryptophan codons do not represent a barrier translation ribosome breezes through.
2. Ribosome continues through element 1 (no stalling) and reaches stop signal (UGA)
3. With no ribosome stem loops 1-2/2-3 form on the mRNA halting transcription before polymerase has chance to reach trp structural genes.
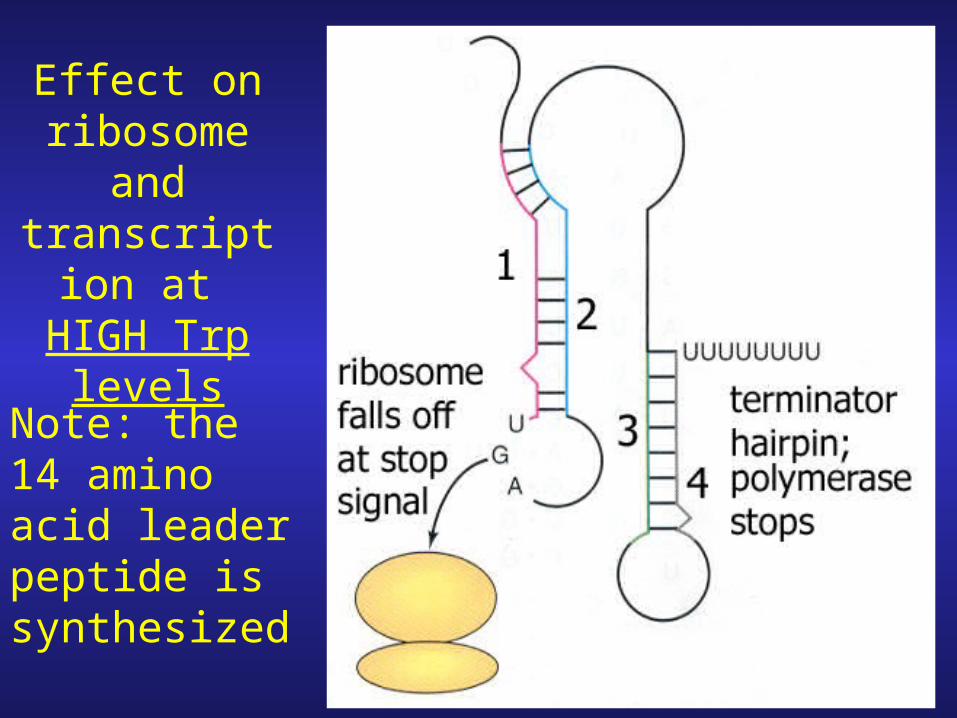
Effect on ribosome
and transcription at HIGH Trp
levels
Note: the 14 amino acid leader peptide is synthesized

-This mechanism involves: transcriptional-translational coupling.
-Relies on rate of transcription & translation to be comparable if RNA polymerase >> ribosome, it might pass through attenuator region before ribosome had a chance to stall at the tryptophan codons.

The Trp Operon of Bacillus subtilis
-mRNA secondary structure controlled by TRAP not by ribosome
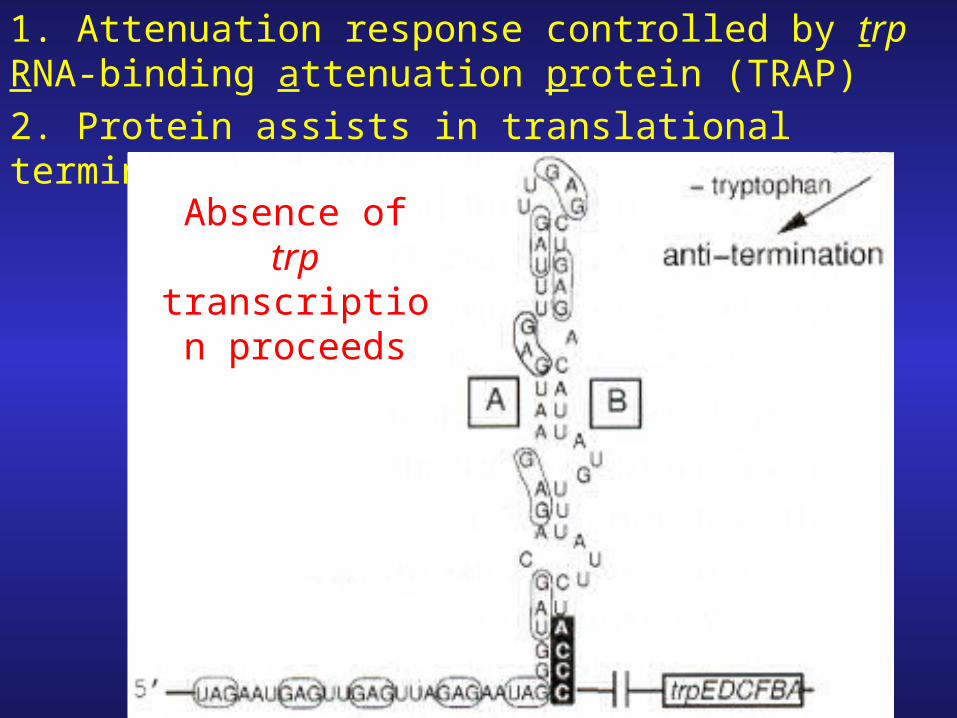
1. Attenuation response controlled by trp RNA-binding attenuation protein (TRAP)2. Protein assists in translational termination.
Absence of trp transcription
proceeds
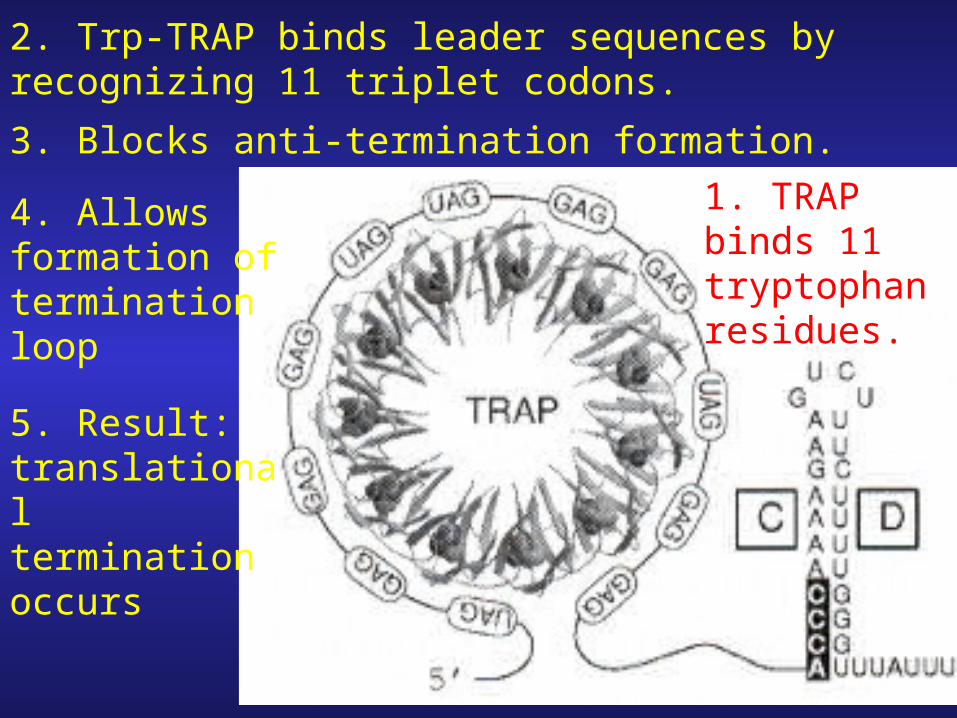
1. TRAP binds 11 tryptophan residues.
2. Trp-TRAP binds leader sequences by recognizing 11 triplet codons.
3. Blocks anti-termination formation.
4. Allows formation of termination loop
5. Result: translational termination occurs
Related Documents











