Molecular identification, typing and traceability of cyanobacteria from freshwater reservoirs Elisabete Vale ´ rio, 1,2 Le ´ lia Chambel, 1 Se ´ rgio Paulino, 2 Nata ´ lia Faria, 2 Paulo Pereira 2 and Roge ´ rio Tenreiro 1 Correspondence Roge ´ rio Tenreiro [email protected] 1 Universidade de Lisboa, Faculdade de Cie ˆ ncias, Centro de Biodiversidade, Geno ´ mica Integrativa e Funcional (BioFIG), Edifı ´cio ICAT, Campus da FCUL, Campo Grande, 1749-016 Lisboa, Portugal 2 Laborato ´ rio de Microbiologia e Ecotoxicologia, Instituto Nacional de Sau ´ de Dr Ricardo Jorge, Avenida Padre Cruz, 1649-016 Lisboa, Portugal Received 1 August 2008 Revised 13 October 2008 Accepted 14 October 2008 In order to assess the potential of several molecular targets for the identification, typing and traceability of cyanobacteria in freshwater reservoirs, molecular techniques were applied to 118 cyanobacterial isolates mostly sourced from Portuguese freshwater reservoirs and representative of three orders of cyanobacteria: Chroococcales (54), Oscillatoriales (15) and Nostocales (49). The isolates were previously identified by morphological methods and subsequently characterized by composite hierarchical cluster analysis of STRR and LTRR (short and long tandemly repeated repetitive sequences) PCR fingerprinting profiles. Representative isolates were selected from each cluster and their molecular identification, at the species level, was obtained or confirmed by phylogenetic positioning using 16S rRNA gene and rpoC1 phylogenies. A highly congruent association was observed between STTR- and LTRR-based clusters and taxonomic affiliation, revealing the usefulness of such PCR fingerprinting profiles for the identification of cyanobacteria. Composite analysis of hierarchical clustering of M13 and ERIC PCR fingerprints also appeared suitable for strain typing and traceability within a reservoir, indicating its potential for use in cyanobacterial monitoring, as a quality management control. Based on Simpson (D) and Shannon–Wiener (J9) indices a high diversity was observed within all species, with Planktothrix agardhii showing the lowest diversity values (D50.83; J950.88) and Aphanizomenon flos-aquae the highest ones (D5J950.99). A diagnostic key based on 16S-ARDRA, ITS amplification and ITS-ARDRA for identification purposes is also presented. INTRODUCTION Cyanobacteria are a morphologically diverse group of bacteria ranging from unicellular to colonial and filament- ous forms. Taxonomically, cyanobacteria are grouped into unicellular forms that divide by binary fission (Order Chroococcales, or Bergey’s Subsection I) or multiple fission (Order Pleurocapsales, or Bergey’s Subsection II); and filamentous forms that are non-heterocystous (Order Oscillatoriales, or Bergey’s Subsection III) or differentiate heterocysts in non-branching (Order Nostocales, or Bergey’s Subsection IV) or branching filaments (Order Stigonematales, or Bergey’s Subsection V) (Castenholz, 2001). A sixth cyanobacterial order, Gloeobacterales, was proposed by Cavalier-Smith (2002) to accommodate the genus Gloeobacter, formerly included in the Chroococcales. Traditionally, the classification of cyanobacteria has been based on morphological characters such as trichome width, cell size, division planes, shape and arrangement, pig- mentation and the presence of characters such as gas vacuoles and a sheath (Baker, 1991, 1992; Bourrelly, 1970; Koma ´rek & Anagnostidis, 1986, 1989). Beyond the considerable expertise required to identify species by such characters, subjective judgment by operators can lead to errors, resulting in incorrect assignment of isolates. Koma ´rek & Anagnostidis (1989) estimated that more than 50 % of the strains in culture collections are misidentified. Moreover, some diagnostic features, such as gas vacuoles or akinetes, can show variations with different environmental or growth conditions and even be lost during cultivation (Rudi et al., 1997; Lyra et al., 2001). Such limitations of phenotypic characters have highlighted the requirement for Abbreviations: ARDRA, amplified rDNA restriction analysis; ERIC, enterobacterial repetitive intergenic consensus; ITS, intergenic tran- scribed spacer region; LTRR, long tandemly repeated repetitive sequences; STRR, short tandemly repeated repetitive sequences; UPGMA, unweighted pair group method with arithmetic average. The GenBank/EMBL/DDBJ accession numbers for the rpoC1 and 16S rDNA sequences are EU078443 to EU078480 (rpoC1) and AY699989, DQ497635 and EU078482 to EU078548 (16S rDNA). Two supplementary tables and seven supplementary figures are available with the online version of this paper. Microbiology (2009), 155, 642–656 DOI 10.1099/mic.0.022848-0 642 022848 G 2009 SGM Printed in Great Britain

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular identification, typing and traceability ofcyanobacteria from freshwater reservoirs
Elisabete Valerio,1,2 Lelia Chambel,1 Sergio Paulino,2 Natalia Faria,2
Paulo Pereira2 and Rogerio Tenreiro1
Correspondence
Rogerio Tenreiro
1Universidade de Lisboa, Faculdade de Ciencias, Centro de Biodiversidade, Genomica Integrativa eFuncional (BioFIG), Edifıcio ICAT, Campus da FCUL, Campo Grande, 1749-016 Lisboa, Portugal
2Laboratorio de Microbiologia e Ecotoxicologia, Instituto Nacional de Saude Dr Ricardo Jorge,Avenida Padre Cruz, 1649-016 Lisboa, Portugal
Received 1 August 2008
Revised 13 October 2008
Accepted 14 October 2008
In order to assess the potential of several molecular targets for the identification, typing and
traceability of cyanobacteria in freshwater reservoirs, molecular techniques were applied to 118
cyanobacterial isolates mostly sourced from Portuguese freshwater reservoirs and representative
of three orders of cyanobacteria: Chroococcales (54), Oscillatoriales (15) and Nostocales (49).
The isolates were previously identified by morphological methods and subsequently characterized
by composite hierarchical cluster analysis of STRR and LTRR (short and long tandemly repeated
repetitive sequences) PCR fingerprinting profiles. Representative isolates were selected from
each cluster and their molecular identification, at the species level, was obtained or confirmed by
phylogenetic positioning using 16S rRNA gene and rpoC1 phylogenies. A highly congruent
association was observed between STTR- and LTRR-based clusters and taxonomic affiliation,
revealing the usefulness of such PCR fingerprinting profiles for the identification of cyanobacteria.
Composite analysis of hierarchical clustering of M13 and ERIC PCR fingerprints also appeared
suitable for strain typing and traceability within a reservoir, indicating its potential for use in
cyanobacterial monitoring, as a quality management control. Based on Simpson (D) and
Shannon–Wiener (J9) indices a high diversity was observed within all species, with Planktothrix
agardhii showing the lowest diversity values (D50.83; J950.88) and Aphanizomenon flos-aquae
the highest ones (D5J950.99). A diagnostic key based on 16S-ARDRA, ITS amplification and
ITS-ARDRA for identification purposes is also presented.
INTRODUCTION
Cyanobacteria are a morphologically diverse group ofbacteria ranging from unicellular to colonial and filament-ous forms. Taxonomically, cyanobacteria are grouped intounicellular forms that divide by binary fission (OrderChroococcales, or Bergey’s Subsection I) or multiple fission(Order Pleurocapsales, or Bergey’s Subsection II); andfilamentous forms that are non-heterocystous (OrderOscillatoriales, or Bergey’s Subsection III) or differentiateheterocysts in non-branching (Order Nostocales, or
Bergey’s Subsection IV) or branching filaments (OrderStigonematales, or Bergey’s Subsection V) (Castenholz,2001). A sixth cyanobacterial order, Gloeobacterales, wasproposed by Cavalier-Smith (2002) to accommodate thegenus Gloeobacter, formerly included in the Chroococcales.
Traditionally, the classification of cyanobacteria has beenbased on morphological characters such as trichome width,cell size, division planes, shape and arrangement, pig-mentation and the presence of characters such as gasvacuoles and a sheath (Baker, 1991, 1992; Bourrelly, 1970;Komarek & Anagnostidis, 1986, 1989). Beyond theconsiderable expertise required to identify species by suchcharacters, subjective judgment by operators can lead toerrors, resulting in incorrect assignment of isolates.Komarek & Anagnostidis (1989) estimated that more than50 % of the strains in culture collections are misidentified.Moreover, some diagnostic features, such as gas vacuoles orakinetes, can show variations with different environmentalor growth conditions and even be lost during cultivation(Rudi et al., 1997; Lyra et al., 2001). Such limitations ofphenotypic characters have highlighted the requirement for
Abbreviations: ARDRA, amplified rDNA restriction analysis; ERIC,enterobacterial repetitive intergenic consensus; ITS, intergenic tran-scribed spacer region; LTRR, long tandemly repeated repetitivesequences; STRR, short tandemly repeated repetitive sequences;UPGMA, unweighted pair group method with arithmetic average.
The GenBank/EMBL/DDBJ accession numbers for the rpoC1 and 16SrDNA sequences are EU078443 to EU078480 (rpoC1) andAY699989, DQ497635 and EU078482 to EU078548 (16S rDNA).
Two supplementary tables and seven supplementary figures areavailable with the online version of this paper.
Microbiology (2009), 155, 642–656 DOI 10.1099/mic.0.022848-0
642 022848 G 2009 SGM Printed in Great Britain

more reliable methods and promoted molecularapproaches in cyanobacterial taxonomy, including DNAbase composition (Kaneko et al., 1996, 2001), DNAhybridizations (Kondo et al., 2000), gene sequencing(Nubel et al., 1997) and PCR fingerprinting (Rasmussen& Svenning, 1998; Versalovic et al., 1991). Cyanobacterial-specific methods not requiring axenic cultures are ofutmost importance since such cultures are difficult toobtain (Choi et al., 2008).
Repetitive sequences constitute an important part of theprokaryotic genome (van Belkum et al., 1998). Despitetheir unknown function, and lack of knowledge about howthey are maintained and dispersed, the presence, wide-spread distribution and high conservation of thesesequences make them methodologically important forDNA fingerprinting and allow their use as an alternativefor the identification of species or strains and in diversitystudies. In the particular case of cyanobacteria, a family ofrepetitive sequences, the short tandemly repeated repetitivesequences (STRRs), has been described (Mazel et al., 1990).These heptanucleotide sequences have been identified inseveral cyanobacterial genera and species, so far mostly inheterocystous cyanobacteria (Rasmussen & Svenning, 1998;Zheng et al., 1999; Nilsson et al., 2000; Wilson et al., 2000;Teaumroong et al., 2002; Lyra et al., 2005; Prasanna et al.,2006), but also in some non-heterocystous ones(Rasmussen & Svenning, 1998). Furthermore, a 37 bplong tandemly repeated repetitive sequence (LTRR) hasalso been identified in some cyanobacterial species(Masepohl et al., 1996; Rasmussen & Svenning, 1998;Prasanna et al., 2006). Analysis of STRRs and LTRRs hasbeen described as a powerful tool for taxonomic studies(Mazel et al., 1990). Moreover, the specificity of thesesequences has made STRRs useful even for non-axeniccyanobacterial cultures (Nilsson et al., 2000).
A universal marker for DNA fingerprinting is theoligonucleotide csM13. It has already been tested in asmall number of cyanobacteria (Valerio et al., 2005), andhas a demonstrated ability even to discriminate strains ofthe same species. Techniques based on the enterobacterialrepetitive intergenic consensus (ERIC) have also been usedfor identification and discrimination purposes in somecyanobacteria (Rasmussen & Svenning, 1998; Lyra et al.,2001; Valerio et al., 2005; Bruno et al., 2006).
The restriction fragment length polymorphisms (RFLPs) ofparticular PCR products can provide signature profilesspecific to the genus, species, or even strain. Geneticcharacterization of cyanobacterial strains has been under-taken using RFLPs of the 16S rRNA gene (16S-ARDRA)(Lyra et al., 1997) and of the intergenic transcribed spacerregion (ITS-ARDRA) (Lu et al., 1997; West & Adams, 1997).Furthermore, amplification of the 16S–23S rRNA ITS, whichhas been shown to be variable in length (Rocap et al., 2002;Iteman et al., 2002; Neilan, 2002, Laloui et al., 2002) andnumber (West & Adams, 1997; Iteman et al., 2002) incyanobacteria, can also be used as an identification tool.
A sequential polyphasic approach was used in this study.Isolates were identified by observation of their morpho-logical features. A hierarchical analysis with STRR andLTRR PCR fingerprinting patterns was performed andrepresentatives of the clusters obtained were identified by aphylogenetic analysis using two genes, one coding for thesmall-subunit rDNA (16S rRNA gene) and the other forthe DNA-dependent RNA polymerase subunit (rpoC1).Subsequent characterization of all isolates by M13 andERIC fingerprints allowed the discrimination of strains,revealing also the traceability potential of these lastmethods for routine freshwater monitoring. Furthermore,a diagnostic key was constructed for the identification ofcyanobacterial species, based on the use of 16S-ARDRA,ITS length and ITS-ARDRA.
METHODS
Cyanobacterial strains. A total of 118 unicyanobacterial non-axenic
cultures, belonging to the orders Chroococcales (54), Oscillatoriales
(15) and Nostocales (49), mainly isolated from Portuguese freshwater
blooms and maintained in the LMECYA culture collection
(Cyanobacteria Culture Collection Estela Sousa e Silva – INSA) using
Z8 medium (Skulberg & Skulberg, 1990), were used in this study
(Supplementary Table S1, available with the online version of this
paper). Isolates were kept under a 14 : 10 h light : dark cycle
(20±4 mmol m22 s21) at 20±1 uC. Lyophilized samples of the
isolates were obtained as described by Saker et al. (2003).
Morphological identification. The morphology of cells and
filaments was studied using an Olympus BX60 light microscope with
a digital camera. The following parameters were analysed: length and
width of vegetative cells; morphology of the terminal cell; the presence
or absence of heterocysts, akinetes and gas vesicles; the distance
between heterocysts and the distance between a heterocyst and the
nearest akinete; and finally, the shape of filaments and their potential
aggregation into colonies.
DNA extraction from cyanobacteria and heterotrophic bacteria.Genomic DNA of cyanobacterial isolates was extracted according to a
previously described method (Valerio et al., 2005).
For isolation of heterotrophic (non-obligately oligotrophic) bacteria,
associated with some cyanobacterial cultures (randomly selected), an
aliquot of the supernatant of isolates LMECYA 64, AQS, LMECYA 79,
LMECYA 13 and LMECYA 173 was grown on LB agar (tryptone 1 %
w/v; yeast extract 0.05 %, w/v; NaCl 0.05 %, w/v; agar 2 %, w/v) at
22 uC for 3 days. The strains obtained were subsequently purified and
grown in LB medium (tryptone 1 %, w/v; yeast extract 0.05 %, w/v;
NaCl 0.05 %, w/v) overnight at 22 uC for DNA extraction. Cells were
recovered by centrifugation and total DNA extracted using the
guanidium thiocyanate method, according to Pitcher et al. (1989).
PCR fingerprinting and data analysis. STRR PCR fingerprinting
was performed using both STRR1F (59-CCCCARTCCCCART-39) and
STRR3F (59-CAACAGTCAACAGT-39) primers (Wilson et al., 2000).
For LTRR PCR fingerprinting both LTRR1 (59-GGATTTTTGTT-
AGTTAAAAC-39) and LTRR2 (59-CTATCAGGGATTGAAAG-39)
primers (Rasmussen & Svenning, 1998) were used.
M13 PCR fingerprinting was performed using oligonucleotide csM13
(59-GAGGGTGGCGGTTCT-39) (Huey & Hall, 1989) as single
primer. For ERIC PCR fingerprinting both ERICIR (59-ATGTAA-
Molecular diagnostics for cyanobacteria
http://mic.sgmjournals.org 643
GCTCCTGGGGATTCAC-39) and ERIC2 (59-AAGTAAGTGACT-GGGGTGAGCG-39) primers (Versalovic et al., 1991) were used.
PCRs were performed in 50 ml containing 16 PCR buffer(Invitrogen), 0.4 mM of each of the four dNTPs (Invitrogen),0.5 mM (for STRR and LTRR fingerprinting) or 1 mM (for M13 andERIC fingerprinting) of each primer, 10–15 ng genomic DNA, 3 mM(for STRR, LTRR and M13 fingerprinting) or 2 mM (for ERICfingerprinting) MgCl2, 1 mg BSA ml21 and 1 U Taq DNA polymerase(Invitrogen). Amplification was performed in a T Gradient thermo-cycler (Biometra), consisting of an initial denaturation step at 95 uCfor 10 min, followed by 35 cycles of 90 s at 95 uC, 2 min at 56 uC (forM13 fingerprinting), 40 uC (for ERIC fingerprinting) or 38 uC (forSTRR and LTRR fingerprinting) and 2 min at 72 uC, and a finalextension step of 5 min at 72 uC.
The PCR products were electrophoresed in 1.4 % (w/v) agarose gel in0.56 TBE buffer at 90 V for 3 h 30 min, using the 1 kb plus DNALadder (Gibco-BRL) as molecular size marker. After staining withethidium bromide, the image obtained under UV transilluminationwas digitized using the Kodak 1D 2.0 system.
To assess reproducibility of the fingerprinting methods according toSneath & Johnson (1972), at least 10 % of isolates were randomlyselected and analysed in duplicate in independent experiments. Thesimilarity between each pair of duplicates was obtained from theanalysis based on a dendrogram computed with Pearson’s correlationcoefficient and the unweighted pair group method with arithmeticaverage (UPGMA) as the agglomerative clustering, usingBioNumerics software v4.0 (Applied Maths) (data not shown). Thereproducibility value was determined as the average similarity for allpairs of duplicates.
The hierarchical clustering of isolates, based on their genomicfingerprinting patterns, was performed with Pearson’s correlationcoefficient and UPGMA using BioNumerics software v4.0. Theaccuracy of each dendrogram to represent the similarity matrix wasassessed by the cophenetic correlation coefficient (r), which ideallyshould be between 0.6 and 0.9 (Priest & Austin, 1993). Compositeanalyses were performed by constructing dendrograms based on thepatterns obtained with two different methods. Clusters of isolateswere correlated with taxonomic positioning, geographical origin andavailable toxicity data.
The diversity within each cyanobacterial species comprising morethan five isolates was evaluated with the indices of Simpson (Hunter& Gaston, 1988) and Shannon–Wiener (Zar, 1996). The Simpsonindex (D) is based on the number of types (fingerprinting profiles)and isolates for each type and measures the probability of two non-related strains, taken from the tested population, belonging to twodifferent genomic types. The Shannon–Wiener index (J9) alsoprovides an evenness measure, expressing the observed diversity asthe proportion of the possible maximum diversity and reflecting thehomogeneity/heterogeneity of the distribution of isolates among thegenomic types.
PCR amplification and sequencing of 16S rRNA and rpoC1
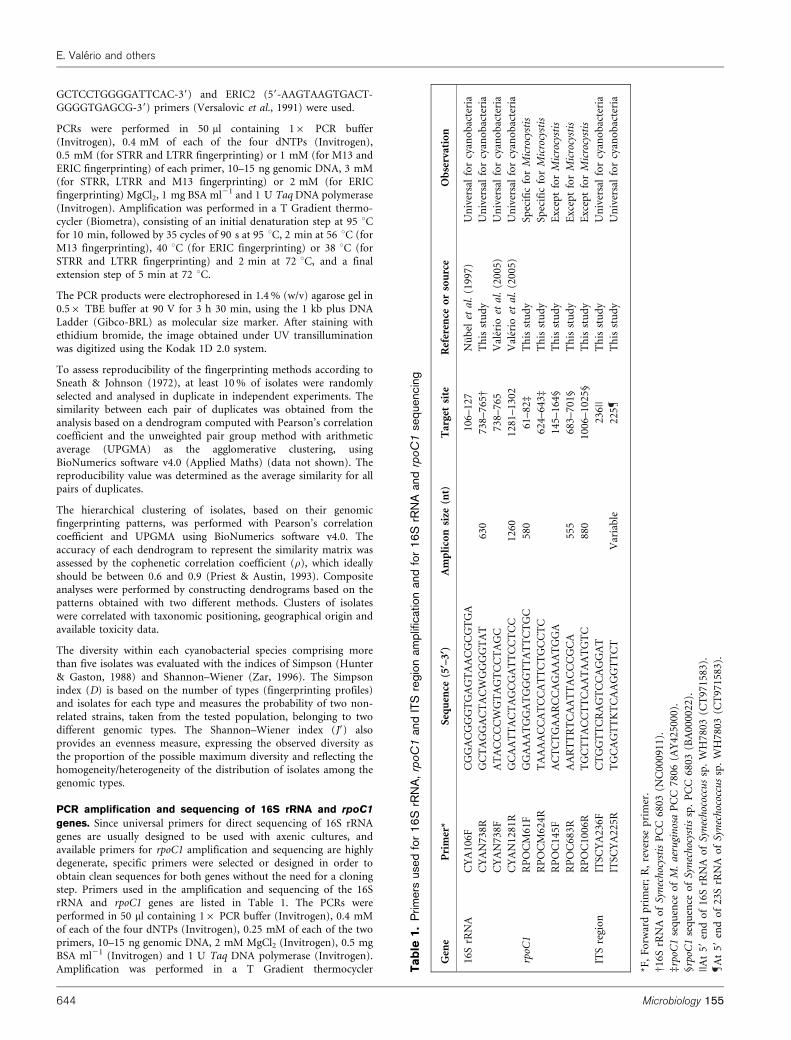
genes. Since universal primers for direct sequencing of 16S rRNAgenes are usually designed to be used with axenic cultures, andavailable primers for rpoC1 amplification and sequencing are highlydegenerate, specific primers were selected or designed in order toobtain clean sequences for both genes without the need for a cloningstep. Primers used in the amplification and sequencing of the 16SrRNA and rpoC1 genes are listed in Table 1. The PCRs wereperformed in 50 ml containing 16 PCR buffer (Invitrogen), 0.4 mMof each of the four dNTPs (Invitrogen), 0.25 mM of each of the twoprimers, 10–15 ng genomic DNA, 2 mM MgCl2 (Invitrogen), 0.5 mgBSA ml21 (Invitrogen) and 1 U Taq DNA polymerase (Invitrogen).Amplification was performed in a T Gradient thermocycler T
ab
le1.
Prim
ers
used
for
16
SrR
NA
,rp
oC
1an
dIT
Sre
gio
nam
plif
icat
ion
and
for
16
SrR
NA
and
rpo
C1
seq
uenc
ing
Gen
eP
rim
er*
Seq
uen
ce(5
§–3§
)A
mp
lico
nsi
ze(n
t)T
arge
tsi
teR
efer
ence
or
sou
rce
Ob
serv
atio
n
16
SrR
NA
CY
A1
06
FC
GG
AC
GG
GT
GA
GT
AA
CG
CG
TG
A1
06
–1
27
Nu
bel
eta
l.(1
99
7)
Un
iver
sal
for
cyan
ob
acte
ria
CY
AN
73
8R
GC
TA
GG
AC
TA
CW
GG
GG
TA
T6
30
73
8–
76
5D
Th
isst
ud
yU
niv
ersa
lfo
rcy
ano
bac
teri
a
CY
AN
73
8F
AT
AC
CC
CW
GT
AG
TC
CT
AG
C7
38
–7
65
Val
erio
eta
l.(2
00
5)
Un
iver
sal
for
cyan
ob
acte
ria
CY
AN
12
81
RG
CA
AT
TA
CT
AG
CG
AT
TC
CT
CC
12
60
12
81
–1
30
2V
aler
ioet
al.
(20
05
)U
niv
ersa
lfo
rcy
ano
bac
teri
a
rpoC
1R
PO
CM
61
FG
GA
AA
TG
GA
TG
GG
TT
AT
TC
TG
C5
80
61
–8
2d
Th
isst
ud
yS
pec
ific
for
Mic
rocy
stis
RP
OC
M6
24
RT
AA
AA
CC
AT
CC
AT
TC
TG
CC
TC
62
4–
64
3d
Th
isst
ud
yS
pec
ific
for
Mic
rocy
stis
RP
OC
14
5F
AC
TC
TG
AA
RC
CA
GA
AA
TG
GA
14
5–
16
4§
Th
isst
ud
yE
xcep
tfo
rM
icro
cyst
is
RP
OC
68
3R
AA
RT
TR
TC
AA
TT
AC
CC
GC
A5
55
68
3–
70
1§
Th
isst
ud
yE
xcep
tfo
rM
icro
cyst
is
RP
OC
10
06
RT
GC
TT
AC
CT
TC
AA
TA
AT
GT
C8
80
10
06
–1
02
5§
Th
isst
ud
yE
xcep
tfo
rM
icro
cyst
is
ITS
regi
on
ITS
CY
A2
36
FC
TG
GT
TC
RA
GT
CC
AG
GA
T2
36
||T
his
stu
dy
Un
iver
sal
for
cyan
ob
acte
ria
ITS
CY
A2
25
RT
GC
AG
TT
KT
CA
AG
GT
TC
TV
aria
ble
22
5T
his
stu
dy
Un
iver
sal
for
cyan
ob
acte
ria
*F,
Fo
rwar
dp
rim
er;
R,
reve
rse
pri
mer
.
D1
6S
rRN
Ao
fS
ynec
hoc
ysti
sP
CC
68
03
(NC
00
09
11
).
drp
oC1
seq
uen
ceo
fM
.a
eru
gin
osa
PC
C7
80
6(A
Y4
25
00
0).
§rp
oC1
seq
uen
ceo
fS
ynec
hoc
ysti
ssp
.P
CC
68
03
(BA
00
00
22
).
||A
t59
end
of
16
SrR
NA
of
Syn
ech
ococ
cus
sp.
WH
78
03
(CT
97
15
83
).
At
59
end
of
23
SrR
NA
of
Syn
ech
ococ
cus
sp.
WH
78
03
(CT
97
15
83
).
E. Valerio and others
644 Microbiology 155

(Biometra) using the following conditions: 10 min of initialdenaturation at 95 uC, followed by 35 cycles at 94 uC for 45 s,55 uC for 45 s and 72 uC for 1 min. The amplification was completedby holding for 5 min at 72 uC to allow the complete extension of thePCR product. PCR products were visualized as described above butusing 1 % (w/v) agarose gel. Amplification reaction products werepurified with a Jet Quick-PCR Purification kit (Genomed) asdescribed by the manufacturer. The purified PCR products weresequenced in both directions using an automated ABI PRISM 3130xlGenetic Analyzer (Applied Biosystems) with dye terminators, usingstandard protocols and the cyanobacterial-specific primers used foramplification, as well as primers CYAN738F and CYAN738R (Table 1)for the 16S rRNA gene sequencing.
Phylogenetic analysis and identification at species level. Atwo-step strategy was used to obtain a 16S rRNA gene phylogeny forthe cyanobacterial strains under analysis. Firstly, a separate phylogenywas constructed for each order (Chroococcales, Oscillatoriales andNostocales), including the sequences from 71 LMECYA strains andreference sequences from well-characterized and reliably identifiedstrains of validated species: sequences from completely sequencedgenomes; sequences from the strains used by Suda et al. (2002) for thetaxonomic revision of the genus Planktothrix; Limnothrix redekeisequences determined by Gkelis et al. (2005); and Microcystis spp.sequences determined by Otsuka et al. (2001).
In the next step, a subgroup of LMECYA isolates and referenceorganisms was selected from the phylogeny of each order asrepresentatives of each tree structure and phylogenetic distancesand a global 16S rRNA gene phylogeny was constructed to include thesix cyanobacterial orders. As indicated in the figure legends, thealignments used encompassed 1182 or 1203 nucleotides of the 16SrRNA gene.
Since rpoC1 sequences were determined for only 38 LMECYA isolatesand fewer reference sequences are available, a global phylogenyincluding the sequences of all these isolates and reference sequenceswas constructed using an alignment of 452 nucleotides.
All alignments were made with CLUSTAL_X version 1.83 and visuallycorrected. Phylogenetic trees were constructed based on a Bayesianapproach using MrBayes software version 3.0b4 (Huelsenbeck &Ronquist, 2001). Each analysis consisted of 2 000 000 generationsfrom a random starting tree and four Markov chains (with defaultheating values) sampled every 100 generations. The first 8000 sampledtrees were discarded, resulting in a set of 12 000 analysed treessampled after the chains became stationary. The eubacterialEscherichia coli sequence was used as an outgroup. Neighbour-joiningand maximum-parsimony trees were also constructed using the samealignments and PAUP 4.0b10 software (Swofford, 2003).
The BLAST tool of the National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov) was used to find homologous andother close sequences (97–100 % identity and E-values ¡10220) to beincluded in the phylogenies as reference sequences.
To achieve the final identification of the isolates, the following criteriawere used. When the phylogenetic positioning of an isolate retrievedidentification at the species level, this prevailed; if the phylogeneticpositioning allowed only genus determination or no molecularidentification was possible, then the identification at the species levelobtained by morphological analysis was retained.
16S rRNA gene and ITS analysis. To select the most suitablerestriction endonucleases to provide an easily identifiable 16S rRNAgene restriction profile, a theoretical study was performed withNEBcutter v2.0 software (Vincze et al., 2003), using cyanobacterial16S rRNA gene sequences available in databases, resulting in theselection of AvaII and BanII enzymes.
For 16S-ARDRA, approximately 1 mg of the amplicon of 1260 bp,
obtained with CYAN106F and CYAN1281R primers (Table 1), was
digested without further purification with AvaII (BioLabs) and BanII
(Takara) restriction endonucleases in separate reactions, according to
the manufacturer’s instructions.
The ITS region was amplified using DNA of non-axenic cultures and
two cyanobacterial-specific primers designed in this study,
ITSCYA236F and ITSCYA225R (Table 1). The PCRs were performed
in 50 ml containing 16 PCR buffer (Invitrogen), 0.4 mM of each of
the four dNTPs (Invitrogen), 0.5 mM of each primer, 10–15 ng
genomic DNA, 2.5 mM MgCl2, 0.5 mg BSA ml21 and 1 U Taq DNA
polymerase (Invitrogen). Amplification was performed in a T
Gradient thermocycler (Biometra), consisting of an initial denatura-
tion step at 95 uC for 6 min, followed by 35 cycles of 45 s at 95 uC,
45 s at 52 uC and 1 min at 72 uC, and a final extension step of 5 min
at 72 uC.
Also in this case, a theoretical study was performed with NEBcutter
v2.0 software (Vincze et al., 2003), using cyanobacterial ITS sequences
available in databases, resulting in the selection of TaqI enzyme.
For ITS-ARDRA, amplified DNA (10 ml, approximately 1 mg) was
digested without further purification with TaqI restriction endonu-
clease (Fermentas) according to the manufacturer’s instructions.
ITS amplicons and restriction products for 16S-ARDRA and ITS-
ARDRA were visualized as described above, using 1.2 % (w/v) and
2 % (w/v) agarose gels, respectively.
RESULTS
Morphological identification
Identification of isolates at the species or genus level waspreviously assessed by their morphology (SupplementaryTable S2). Out of the 118 isolates, 99 could be identified atthe species level and the remaining 19 only at the genuslevel. These results suggest that the cyanobacterial speciespresent in Portuguese freshwaters, most of which areresponsible for bloom formation, are diverse, and belong tothree of the six cyanobacterial orders: Chroococcales,Oscillatoriales and Nostocales. Of the 118 isolates understudy, 54 (45.8 %) belong to Chroococcales, 15 (12.7 %) toOscillatoriales and 49 (41.5 %) to Nostocales. The speciespredominantly found were Microcystis aeruginosa (52isolates; 44 %) and Aphanizomenon flos-aquae (13 isolates;11 %).
Assessment of PCR fingerprinting reliability
The average reproducibility of the tested fingerprintingtechniques, estimated by the similarity average value for allpairs of duplicates, was 92 %, representing a goodreproducibility (Sneath & Johnson, 1972), since thePearson’s correlation coefficient used is very sensitive tothe band intensity, leading to lower final similarity values.In all dendrograms the cophenetic value (r) is above 0.85,which corresponds to a good representation of the originaldata .
In order to assess the effect of the non-axenic nature ofthe uni-cyanobacterial cultures, the PCR fingerprinting
Molecular diagnostics for cyanobacteria
http://mic.sgmjournals.org 645

methods to be tested (STRR, LTRR, M13 and ERIC) wereapplied to a set of cyanobacterial cultures and their co-cultured heterotrophic (non-obligately oligotrophic) bac-teria. From the same culture (Fig. 1 underlined group oflanes), cyanobacteria (in bold) and cultivated hetero-trophic bacteria displayed M13 (Fig. 1c) and ERIC (Fig.1d) PCR fingerprints with very few co-migrating fragments(white boxes). However, isolates LMECYA 79 andLMECYA 173 displayed identical fingerprinting profileswith each technique, despite the fact that their corres-ponding cultivated bacteria showed M13 and ERIC profilesdifferent from each other but with some co-migratingfragments with the cyanobacteria (Fig. 1, lanes 8–11). Onthe other hand, STRR and LTRR PCR fingerprinting (Fig.1a, b) showed specificity for cyanobacteria, since noamplification was obtained for the cultivated bacteria.
STRR and LTRR PCR fingerprinting
A composite analysis of hierarchical clustering of STRRand LTRR patterns was performed with all isolates (datanot shown) and subsequently for each order as presented inFig. 2. It can be observed that the isolates groupedaccording to their genomic similarity. To confirm thespecies identification, a set of representative isolates(marked with bold italics in Fig. 2), including membersof different clusters and members of the same cluster, wasselected and their identification was obtained or confirmedby phylogenetic positioning using 16S rRNA gene andrpoC1 phylogenies.
The identification presented in Fig. 2 is in agreement withthe final identification of the isolates obtained by 16SrRNA gene sequencing (which in some cases was alsoconfirmed by the rpoC1 sequencing, as presented below). Itcan be observed that the majority of the clusterscorrespond to different species, showing the taxonomicpotential of STRR and LTRR fingerprints.
The three species (two of them represented by only oneisolate) belonging to the order Chroococcales (Fig. 2a) areintermixed and could not be separated according to theirSTRR and LTRR fingerprinting patterns. In fact, theisolates of Microcystis aeruginosa show very diverse profiles(grouping in three distinct clusters) and the isolate ofSynechocystis sp. is more similar (52.5 %) to cluster II of M.aeruginosa than the M. aeruginosa clusters are to each other(15–27 %). Synechococcus nidulans, together with oneisolate of M. aeruginosa, has the lowest similarity (10 %)with all the other isolates. Fingerprinting patterns ofisolates belonging to the order Oscillatoriales (Fig. 2b) ledto the formation of two clusters and four isolated strains at40 % similarity, while nine major clusters and four isolatedstrains were observed at 50 % similarity for isolates of theorder Nostocales (Fig. 2c). In both cases almost all theclusters correspond to different species, except forAnabaena circinalis and Aphanizomenon gracile, whichformed two distinct clusters (genotypes).
Fig. 1. Fingerprinting profiles of cyanobacteria and cultivablebacteria from each non-axenic culture. (a) STRR PCR fingerprint-ing; (b) LTRR PCR fingerprinting; (c) M13 PCR fingerprinting; (d)ERIC PCR fingerprinting. (1) LMECYA 64; (2) LMECYA 64heterotrophic bacteria #1; (3) LMECYA 64 heterotrophic bacteria#2; (4) LMECYA 64 heterotrophic bacteria #3; (5) LMECYA 64heterotrophic bacteria #4; (6) AQS; (7) AQS heterotrophicbacteria #1; (8) LMECYA 79; (9) LMECYA 79 heterotrophicbacteria #1; (10) LMECYA 173; (11) LMECYA 173 hetero-trophic bacteria #1; (12) LMECYA 13; (13) LMECYA 13heterotrophic bacteria #1. The white boxes mark the commonfragments.
E. Valerio and others
646 Microbiology 155

Fig. 2. Dendrograms obtained by composite hierarchical analysis of STRR and LTRR fingerprinting profiles using Pearson’s correlation coefficient and the UPGMA clusteringmethod for the orders used in the study: (a) Chroococcales, (b) Oscillatoriales and (c) Nostocales. The cophenetic correlations (r) of the dendrograms are shown. The scalecorresponds to global percentage similarity. gen, genotype. A ‘weight of 2’ was attributed to the STRR patterns, and a ‘weight of 1’ to the LTRR patterns.
Molecular
diagnostics
forcyanobacteria
http://mic.sg
mjournals.org
64
7

Phylogenetic analysis using the 16S rRNA gene
The phylogenetic tree presented in Fig. S1 corresponds tothe partial 16S rRNA gene phylogeny of the orderChroococcales. This phylogeny confirmed the identifica-tion of the 27 sequenced isolates only at the genus level. Infact, although the genus Microcystis forms a monophyleticcluster, all five species are highly homologous and thuscannot be distinguished at the 16S rRNA gene level.
The partial 16S rRNA gene phylogeny of the orderOscillatoriales is depicted in Fig. S2; it allowed theidentification at species level of seven sequenced isolates(out of 10), four of these (LMECYA 152, 153, 153E and153F) as Planktothrix agardhii, one (LMECYA 203) asPlanktothrix rubescens, one (LMECYA 162) as Planktothrixpseudoagardhii and one (LMECYA 145) as Limnothrixredekei. The three remaining isolates (LMECYA 79, 173and 214) were grouped in a cluster containing sequences ofPhormidium and Leptolyngbya isolates. However, takinginto account their morphological characteristics, theseisolates were identified as Phormidium sp. The distributionof Oscillatoria and Lyngbya isolates in several distinct cladesalso reveals the polyphyletic nature of these genera.
The partial 16S rRNA gene phylogeny of the orderNostocales (Fig. S3) allowed the identification of 21 outof 34 sequenced isolates. The isolate LMECYA 177 groupsin a cluster where several Anabaena species are present.This phylogenetic positioning does not allow an identifica-tion of this isolate; however, its morphological features aresimilar to those of Anabaena planctonica, which is alsopresent in the cluster. This is also the case with isolates 77A,77B, 88, 99, 125, 129 and 141, grouped in a cluster withmainly Aphanizomenon flos-aquae but also Aphanizomenongracile. However, morphological features of these isolatesare typical of Aphanizomenon flos-aquae. Isolates LMECYA9 and 33 grouped in a cluster with Anabaena flos-aquae,Aphanizomenon flos-aquae and Aphanizomenon gracilesequences, but morphological features of these isolatesare typical of Aphanizomenon gracile. The isolates LMECYA178, 182 and 185 did not group with any species or genus.In fact, their morphology is congruent with Anabaena, andLMECYA 178 is identical to Anabaena flos-aquae. Thisphylogeny also showed that Anabaena and Aphanizomenondo not form monophyletic clusters, with isolates of bothgenera intermixed along the tree.
The global 16S rRNA gene phylogeny of representativespecies of the six cyanobacterial orders, using a Bayesianapproach, is illustrated in Fig. 3(a). It shows that theNostocales form a monophyletic cluster, except for the soleScytonema sequence presented. Stigonematales are alsomonophyletic, containing the genera Chlorogloeopsis,Fischerella and Nostochopsis, except the unique Stigonemasequence here presented. The order Pleurocapsales forms amonophyletic cluster, whereas the orders Chroococcales andOscillatoriales are both divided into two distinct clades.Identical results were obtained using neighbour-joining andmaximum-parsimony trees (Supplementary Fig. S4).
Phylogenetic analysis using rpoC1
The rpoC1-based phylogeny presented in Fig. 3(b) allowedthe identification of six of the 38 sequenced isolates. Two ofthem (LMECYA 17 and 126) belong to Anabaena circinalis,one (LMECYA 134) to Cylindrospermopsis raciborskii, one(LMECYA 153E) to Planktothrix agardhii and one(LMECYA 68) to Synechocystis sp. All these identificationswere congruent with those obtained by 16S rRNA genesequencing.
Isolates LMECYA 9, 40, 64 and 148, identified bothmorphologically and by 16S rRNA gene sequencing asAphanizomenon gracile, clustered into two distinct groups,neither of them closely related to the availableAphanizomenon gracile rpoC1 sequence.
Phormidium sp. isolates (LMECYA 79 and 173) groupedinto a cluster containing another Phormidium sequence butalso Leptolyngbya and Synechococcus sequences.
Some isolates did not group with a particular species, dueto the low number of rpoC1 sequences available.
In the rpoC1 phylogeny, the Nostocales split into twoclusters, a major one containing almost all species of theclade obtained with the 16S rRNA gene and a secondgrouping the remaining species, including Scytonema sp.,with species from Stigonematales and Pleurocapsales.Oscillatoriales were separated in distinct clades, mostlyequivalent to the 16S rRNA ones. Chroococcales presenteda monophyletic cluster, except for the Prochloron and twoSynechococcus spp. sequences, which were distributed inother clusters. When this rpoC1 Bayesian tree wascompared with neighbour-joining and maximum-par-simony trees (see Supplementary Fig. S5), similar phylo-genetic relationships were observed.
Strain discrimination and traceability
Representative M13 and ERIC PCR fingerprinting profilesof the genomic types of the species comprising more thanfive isolates are presented in Supplementary Fig. S6. As thisfigure shows, the richness and diversity of the DNAbanding patterns allowed the discrimination of all distinctisolates and the detection of identical ones.
Individual dendrograms were constructed for each of theseven species (data not shown). Based on the M13 andERIC PCR fingerprints of all the Microcystis aeruginosaisolates evaluated by a composite analysis of hierarchicalclustering, a cut-off level of 75 % similarity was establishedfor definition of a genomic type. Table 2 presents thepercentage of types and values of Simpson and Shannon–Wiener diversity indices (for the species with more thanfive isolates). The lowest diversity values were obtained forPlanktothrix agardhii, where four of the nine isolates havethe same genomic type. The other four species whosediversity indices were determined revealed higher diversityvalues, with Aphanizomenon flos-aquae showing the highestone.
E. Valerio and others
648 Microbiology 155

Fig. 3. (a) Global MrBayes tree of representative species using 1182 nt of the 16S rRNA gene sequences in the alignment. (b) MrBayes tree of the rpoC1 sequences using452 nt in the alignment. Posterior probabilities values above 0.5 are indicated to provide branch support. Bars: 10 % sequence divergence. GenBank accession numbers areindicated after the species designation (names in bold correspond to sequences determined in this study).
Molecular
diagnostics
forcyanobacteria
http://mic.sg
mjournals.org
64
9
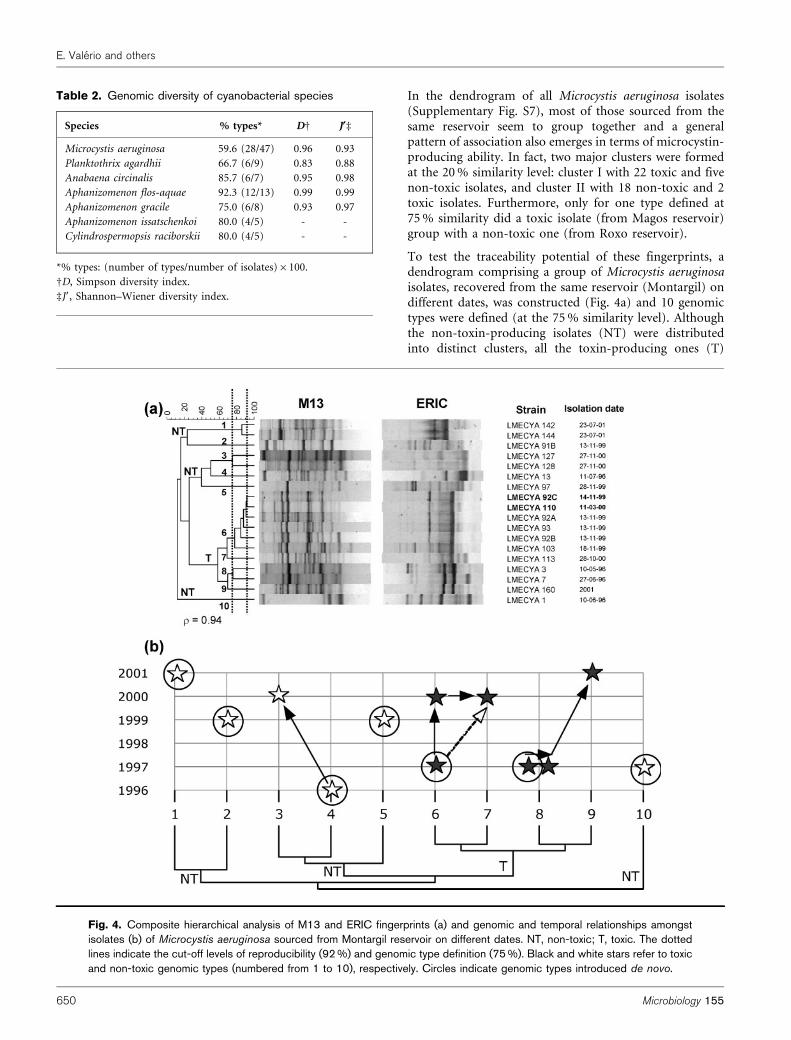
In the dendrogram of all Microcystis aeruginosa isolates(Supplementary Fig. S7), most of those sourced from thesame reservoir seem to group together and a generalpattern of association also emerges in terms of microcystin-producing ability. In fact, two major clusters were formedat the 20 % similarity level: cluster I with 22 toxic and fivenon-toxic isolates, and cluster II with 18 non-toxic and 2toxic isolates. Furthermore, only for one type defined at75 % similarity did a toxic isolate (from Magos reservoir)group with a non-toxic one (from Roxo reservoir).
To test the traceability potential of these fingerprints, adendrogram comprising a group of Microcystis aeruginosaisolates, recovered from the same reservoir (Montargil) ondifferent dates, was constructed (Fig. 4a) and 10 genomictypes were defined (at the 75 % similarity level). Althoughthe non-toxin-producing isolates (NT) were distributedinto distinct clusters, all the toxin-producing ones (T)
Table 2. Genomic diversity of cyanobacterial species
Species % types* DD J§d
Microcystis aeruginosa 59.6 (28/47) 0.96 0.93
Planktothrix agardhii 66.7 (6/9) 0.83 0.88
Anabaena circinalis 85.7 (6/7) 0.95 0.98
Aphanizomenon flos-aquae 92.3 (12/13) 0.99 0.99
Aphanizomenon gracile 75.0 (6/8) 0.93 0.97
Aphanizomenon issatschenkoi 80.0 (4/5) - -
Cylindrospermopsis raciborskii 80.0 (4/5) - -
*% types: (number of types/number of isolates)6100.
DD, Simpson diversity index.
dJ9, Shannon–Wiener diversity index.
Fig. 4. Composite hierarchical analysis of M13 and ERIC fingerprints (a) and genomic and temporal relationships amongstisolates (b) of Microcystis aeruginosa sourced from Montargil reservoir on different dates. NT, non-toxic; T, toxic. The dottedlines indicate the cut-off levels of reproducibility (92 %) and genomic type definition (75 %). Black and white stars refer to toxicand non-toxic genomic types (numbered from 1 to 10), respectively. Circles indicate genomic types introduced de novo.
E. Valerio and others
650 Microbiology 155
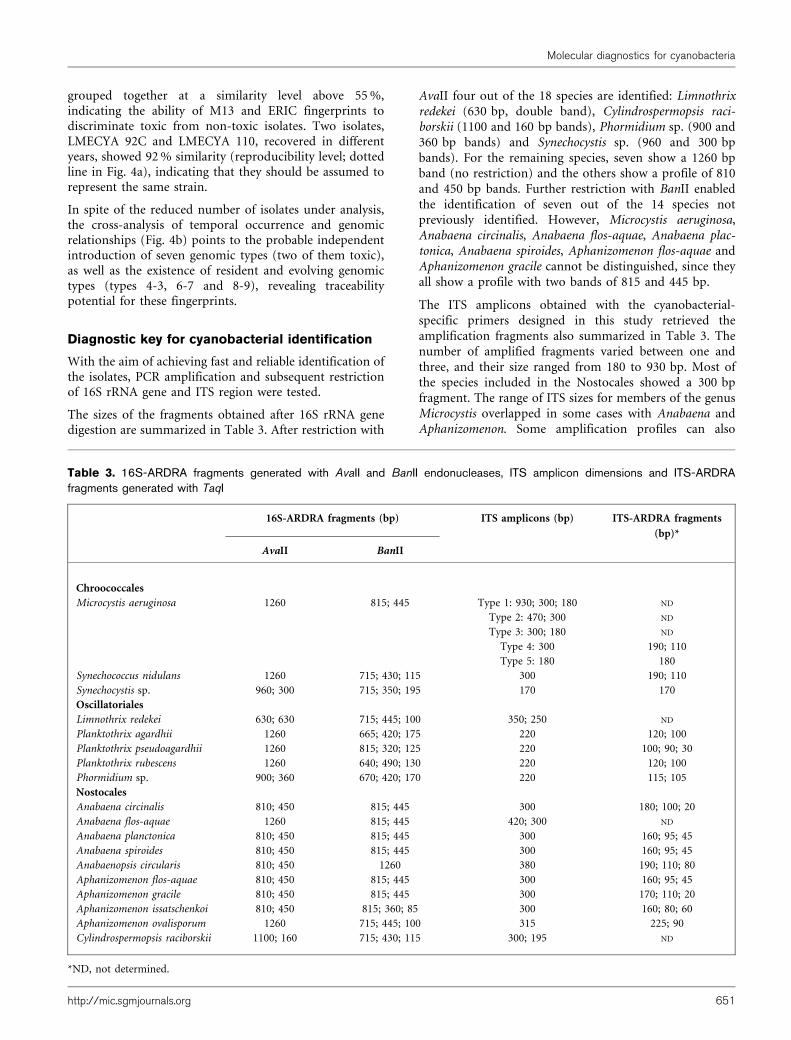
grouped together at a similarity level above 55 %,indicating the ability of M13 and ERIC fingerprints todiscriminate toxic from non-toxic isolates. Two isolates,LMECYA 92C and LMECYA 110, recovered in differentyears, showed 92 % similarity (reproducibility level; dottedline in Fig. 4a), indicating that they should be assumed torepresent the same strain.
In spite of the reduced number of isolates under analysis,the cross-analysis of temporal occurrence and genomicrelationships (Fig. 4b) points to the probable independentintroduction of seven genomic types (two of them toxic),as well as the existence of resident and evolving genomictypes (types 4-3, 6-7 and 8-9), revealing traceabilitypotential for these fingerprints.
Diagnostic key for cyanobacterial identification
With the aim of achieving fast and reliable identification ofthe isolates, PCR amplification and subsequent restrictionof 16S rRNA gene and ITS region were tested.
The sizes of the fragments obtained after 16S rRNA genedigestion are summarized in Table 3. After restriction with
AvaII four out of the 18 species are identified: Limnothrixredekei (630 bp, double band), Cylindrospermopsis raci-borskii (1100 and 160 bp bands), Phormidium sp. (900 and360 bp bands) and Synechocystis sp. (960 and 300 bpbands). For the remaining species, seven show a 1260 bpband (no restriction) and the others show a profile of 810and 450 bp bands. Further restriction with BanII enabledthe identification of seven out of the 14 species notpreviously identified. However, Microcystis aeruginosa,Anabaena circinalis, Anabaena flos-aquae, Anabaena plac-tonica, Anabaena spiroides, Aphanizomenon flos-aquae andAphanizomenon gracile cannot be distinguished, since theyall show a profile with two bands of 815 and 445 bp.
The ITS amplicons obtained with the cyanobacterial-specific primers designed in this study retrieved theamplification fragments also summarized in Table 3. Thenumber of amplified fragments varied between one andthree, and their size ranged from 180 to 930 bp. Most ofthe species included in the Nostocales showed a 300 bpfragment. The range of ITS sizes for members of the genusMicrocystis overlapped in some cases with Anabaena andAphanizomenon. Some amplification profiles can also
Table 3. 16S-ARDRA fragments generated with AvaII and BanII endonucleases, ITS amplicon dimensions and ITS-ARDRAfragments generated with TaqI
16S-ARDRA fragments (bp) ITS amplicons (bp) ITS-ARDRA fragments
(bp)*
AvaII BanII
Chroococcales
Microcystis aeruginosa 1260 815; 445 Type 1: 930; 300; 180 ND
Type 2: 470; 300 ND
Type 3: 300; 180 ND
Type 4: 300 190; 110
Type 5: 180 180
Synechococcus nidulans 1260 715; 430; 115 300 190; 110
Synechocystis sp. 960; 300 715; 350; 195 170 170
Oscillatoriales
Limnothrix redekei 630; 630 715; 445; 100 350; 250 ND
Planktothrix agardhii 1260 665; 420; 175 220 120; 100
Planktothrix pseudoagardhii 1260 815; 320; 125 220 100; 90; 30
Planktothrix rubescens 1260 640; 490; 130 220 120; 100
Phormidium sp. 900; 360 670; 420; 170 220 115; 105
Nostocales
Anabaena circinalis 810; 450 815; 445 300 180; 100; 20
Anabaena flos-aquae 1260 815; 445 420; 300 ND
Anabaena planctonica 810; 450 815; 445 300 160; 95; 45
Anabaena spiroides 810; 450 815; 445 300 160; 95; 45
Anabaenopsis circularis 810; 450 1260 380 190; 110; 80
Aphanizomenon flos-aquae 810; 450 815; 445 300 160; 95; 45
Aphanizomenon gracile 810; 450 815; 445 300 170; 110; 20
Aphanizomenon issatschenkoi 810; 450 815; 360; 85 300 160; 80; 60
Aphanizomenon ovalisporum 1260 715; 445; 100 315 225; 90
Cylindrospermopsis raciborskii 1100; 160 715; 430; 115 300; 195 ND
*ND, not determined.
Molecular diagnostics for cyanobacteria
http://mic.sgmjournals.org 651

provide an identification, as is the case for Synechocystis sp.(170 bp band), Limnothrix redekei (350 and 250 bp bands),Anabaena flos-aquae (420 and 300 bp bands), Anabaenopsiscircularis (380 bp band), Aphanizomenon ovalisporum(315 bp band) and Cylindrospermopsis raciborskii (300and 195 bp). Moreover, a 220 bp fragment was alwaysobtained for the Planktothrix spp. isolates.
The ITS-ARDRA fragments obtained after restriction of theITS with TaqI are summarized in Table 3. Some of theisolates show a profile that can be used for identification.The genotypic pattern produced using this restrictionenzyme can provide a species identification marker: forinstance Planktothrix pseudoagardhii, which showed aprofile distinct from the other two Planktothrix species,Planktothrix agardhii, difficult to distinguish from theformer by morphological characteristics, and Planktothrixrubescens, which can be identified by its cell dimensionsand brownish colour.
Based on the 16S-ARDRA, ITS amplification and ITS-ARDRA a diagnostic key was constructed (Fig. 5) allowingthe identification of 15 out of the 18 species analysed. OnlyAnabaena planctonica, Anabaena spiroides andAphanizomenon flos-aquae could not be distinguished.
DISCUSSION
In this study, several molecular techniques were used toexamine the level of genetic diversity of 118 cyanobacterialisolates for taxonomic purposes.
All the PCR fingerprinting methods showed good repro-ducibility, and the inter- and intra-specific variability,observed in all species analysed, was revealed to beappropriate for assessment of genomic relationships,isolate discrimination and traceability, in spite of thenon-axenic character of the cyanobacterial cultures. STRRfingerprints were obtained for a larger number of non-heterocystous cyanobacteria than described so far(Rasmussen & Svenning, 1998) and the LTRR PCRfingerprinting was also tested in a large number of species.Beyond some discrimination ability, the specificity of theSTRR and LTRR fingerprinting patterns revealed iden-tification potential. In fact, all species from the Nostocalesand Oscillatoriales could be discriminated by the compos-ite hierarchical analysis of the STRR and LTRR fingerprintswhen only species/genera of the same order were analysed.For most species, a good correlation was found betweenmorphological identification and genomic clustering. Inthe case of Anabaena circinalis two clusters were formed,corresponding to two genotypes whose 16S rRNA gene was96.5 % similar, whereas the isolates of the same genotypedisplayed 99.9 % similarity. A similar finding was observedfor Aphanizomenon gracile isolates, which were also dividedinto two clusters, showing 98 % 16S rRNA gene similaritybetween them and 99–100 % within the clusters. Thecomposite hierarchical analysis of the STRR and LTRRfingerprints reflects the identification results obtained by
16S rRNA gene sequencing, and thus can be used as anidentification tool, decreasing the need for 16S rRNA genesequencing, often used in association with the morpho-logical identification of a cyanobacterial strain (Nubel etal., 1997).
The genus Microcystis forms a monophyletic group distantfrom the other genera in both 16S rRNA and rpoC1phylogenies. All five Microcystis species showed a highhomology (.98.5 % with both genes), preventing theirdiscrimination. This has already been shown by Otsuka etal. (2001), who proposed the unification of all five species,with Microcystis aeruginosa having nomenclatural priority.However, despite the high sequence homology and also thehigh DNA–DNA reassociation levels found among thesespecies, Kondo et al. (2000) suggested their maintenance asdistinct taxa based on their distinct morphological features.
Both 16S rRNA and rpoC1 gene phylogenies showed thatthe genera Synechococcus and Synechocystis form distinctclades. Nevertheless, our results recommend a revision ofthe genus Synechococcus as already suggested by Robertsonet al. (2001).
Regarding the order Oscillatoriales, we also found that thegenus Planktothrix forms a monophyletic cluster closer toNostocales than to other Oscillatoriales, as alreadydescribed by Suda et al. (2002). It is also evident that thegenus Oscillatoria still needs major revision, its speciesbeing widely distributed in 16S rRNA gene trees. In fact,this genus has recently undergone revisions (Suda et al.,2002) and its polyphyly was also inferred by others (Ishidaet al., 2001; Marquardt & Palinska, 2007; Suda et al., 2002).Similarly to Joyner et al. (2008), a polyphyletic origin of thegenus Lyngbya was also inferred from the 16S rRNA genetree, justifying its taxonomic revision. This might be relatedto the fact that traditional criteria used for classification ofthe Oscillatoriales at the genus level predominantly rely onthe characteristics of external sheaths and colony forma-tion, rather than on cellular features (Marquardt &Palinska, 2007). Other 16S rRNA analyses have alreadydemonstrated the polyphyletic origin of members of theorders Chroococcales (Litvaitis, 2002; Seo & Yokota, 2003)and Oscillatoriales (Ishida et al., 2001; Litvaitis, 2002).
We also can verify from the rpoC1 phylogeny that the orderNostocales is not monophyletic and that its members areclosely related to those of the Stigonematales, givingadditional support to the statement already made byGugger & Hoffmann (2004) based on 16S rRNA genephylogeny. Anabaena and Aphanizomenon isolates weregenetically heterogeneous and intermixed in both genetrees, confirming previous results obtained with otherstrains (Lyra et al., 2001; Gugger et al., 2002; Rajaniemi etal., 2005). Furthermore, the possible presence of incorrectlyidentified sequences in databases may also distort theinferred conclusions. Although the results obtained withrpoC1 phylogeny seem to indicate possible lateral genetransfer events in Anabaena and Aphanizomenon species,the hypothesis that the morphological features are
E. Valerio and others
652 Microbiology 155
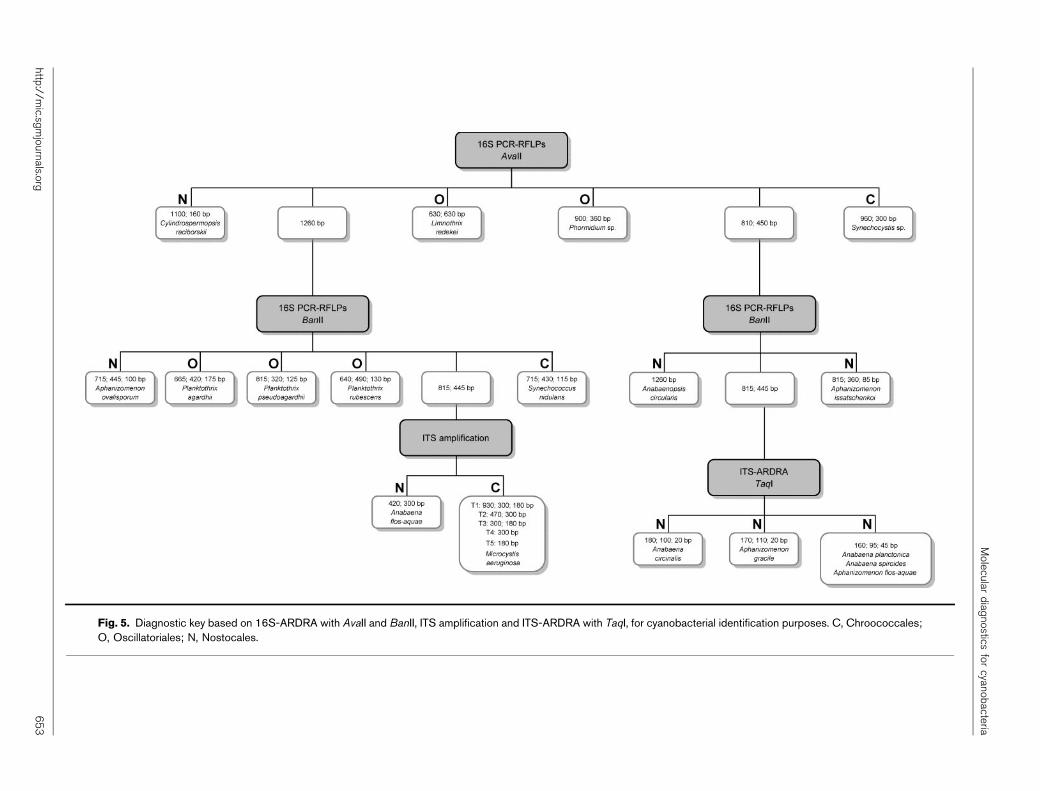
Fig. 5. Diagnostic key based on 16S-ARDRA with AvaII and BanII, ITS amplification and ITS-ARDRA with TaqI, for cyanobacterial identification purposes. C, Chroococcales;O, Oscillatoriales; N, Nostocales.
Molecular
diagnostics
forcyanobacteria
http://mic.sg
mjournals.org
65
3

insufficient to delineate the true taxonomic structure ofthese genera cannot be ruled out. Cylindrospermopsisraciborskii forms a monophyletic group, more distant fromthe other species of the Nostocales with both markers.
The identification of genomic types by hierarchicalclustering analysis of M13 and ERIC fingerprints allowedthe assessment of intra-specific diversity for the represent-ative species of this study and also revealed this approachto be a useful tool for traceability purposes within afreshwater reservoir.
Although a rather low number of isolates was analysed forthe majority of the species, except for Microcystisaeruginosa, a high diversity was observed within all species,with Planktothrix agardhii showing the lowest diversityvalues (D50.83; J950.88) and Aphanizomenon flos-aquaethe highest ones (D5J950.99). Furthermore, and exceptfor Planktothrix agardhii, a high diversity of genomic typeswas usually found amongst isolates of the same speciescollected from the same reservoir.
Cross-analysis of temporal occurrence and genomicrelationships, performed with Microcystis aeruginosa iso-lates collected over several years from Montargil reservoir,allowed inference of the dynamics of genomic types withina reservoir, illustrated the traceability potential of M13 andERIC PCR fingerprinting and, as already observed forCylindrospermopsis raciborskii isolates (Valerio et al., 2005),highlighted its usefulness for ecological studies andevaluation of populations, when high resolution is needed.
A method to identify cyanobacterial isolates was developedthrough the construction of a diagnostic key based on theamplification of the 16S rRNA gene and further digestionwith one or two restriction endonucleases. For those caseswhere no identification could still be obtained, theamplification or even further restriction of the ITS regionwith TaqI provided the identification of most species understudy. This diagnostic key provides a faster, easier andreliable method to perform cyanobacterial identificationfor 15 out of the 18 species studied, without the need forexpert experience to perform morphological identification,thus avoiding a misidentification of isolates, as frequentlyhappens (Komarek & Anagnostidis, 1989). As an exampleof the application potentiality of this method, if the keyhere presented had been applied to the 118 isolatesincluded in this study 84.7 % of them (100 out of 118)would have been immediately identified at species level.However, for the three species not discriminated by thiskey, as well as for other species where morphologicalfeatures prevail in terms of species delimitation, amorphological analysis will be more reliable or, at least,will probably be required. Therefore, and as envisaged byRita Colwell almost four decades ago, a polyphasicapproach merging traditional identification and molecularcharacterization will certainly be the most powerful optionto detect, identify and characterize cyanobacterial speciesand strains, including the harmful ones.
ACKNOWLEDGEMENTS
The authors are deeply indebted to the anonymous journal refereesfor their thorough review and helpful comments on both scientificissues and terminology. Elisabete Valerio is recipient of Fundacaopara a Ciencia e a Tecnologia grant SFRH/BD/8272/2002.
REFERENCES
Baker, P. (1991). Identification of Common Noxious CyanobacteriaPart I – Nostocales: Urban Water Research Association of Australia.Research Report no. 29.
Baker, P. (1992). Identification of Common Noxious CyanobacteriaPart II – Chroococales Oscillatoriales: Urban Water ResearchAssociation of Australia. Research Report no. 46.
Bourrelly, P. (1970). Les Algues d’Eau Douce: Les Algues Bleus etRouges, Les Eugleniens, Peridiniens, et Cryptomonadines. Paris: N.Boubee & Cie.
Bruno, L., Billi, D., Albertano, P. & Urzi, C. (2006). Geneticcharacterization of epilithic cyanobacteria and their associatedbacteria. Geomicrobiol J 23, 293–299.
Castenholz, R. W. (2001). Phylum BX. Cyanobacteria. In Bergey’sManual of Systematic Bacteriology, 2nd edn, vol. 1, pp. 473–599.Edited by D. R. Boone, R. W. Castenholz & G. M. Garrity. New York:Springer.
Cavalier-Smith, T. (2002). The neomuran origin of archaebacteria,the negibacterial root of the universal tree and bacterial mega-classification. Int J Syst Evol Microbiol 52, 7–76.
Choi, G. G., Bae, M. S., Ahn, C. Y. & Oh, H. M. (2008). Induction ofaxenic culture of Arthrospira (Spirulina) platensis based on antibioticsensitivity of contaminating bacteria. Biotechnol Lett 30, 87–92.
Gkelis, S., Rajaniemi, P., Vardaka, E., Moustaka-Gouni, M.,Lanaras, T. & Sivonen, K. (2005). Limnothrix redekei (Van Goor)Meffert (Cyanobacteria) strains from Lake Kastoria, Greece form aseparate phylogenetic group. Microb Ecol 49, 176–182.
Gugger, M. F. & Hoffmann, L. (2004). Polyphyly of true branchingcyanobacteria (Stigonematales). Int J Syst Evol Microbiol 54, 349–357.
Gugger, M., Lyra, C., Henriksen, P., Coute, A., Humbert, J. &Sivonen, K. (2002). Phylogenetic comparison of the cyanobacterialgenera Anabaena and Aphanizomenon. Int J Syst Evol Microbiol 52,1867–1880.
Huelsenbeck, J. P. & Ronquist, F. R. (2001). MrBayes: Bayesianinference of phylogenetic trees. Bioinformatics 17, 754–755.
Huey, B. & Hall, J. (1989). Hypervariable DNA fingerprinting inEscherichia coli: minisatellite probe from bacteriophage M13.J Bacteriol 171, 2528–2532.
Hunter, P. R. & Gaston, M. A. (1988). Numerical index of thediscriminatory ability of typing systems: an application of Simpson’sindex of diversity. J Clin Microbiol 26, 2465–2466.
Ishida, T., Watanabe, M. M., Sugiyama, J. & Yokota, A. (2001).Evidence for polyphyletic origin of the members of the orders ofOscillatoriales and Pleurocapsales as determined by 16S rDNAanalysis. FEMS Microbiol Lett 201, 79–82.
Iteman, I., Rippka, R., Tandeau de Marsac, N. & Herdman, M. (2002).rDNA analyses of planktonic heterocystous cyanobacteria, includingmembers of the genera Anabaenopsis and Cyanospira. Microbiology148, 481–496.
Joyner, J. J., Litaker, R. W. & Paerl, H. W. (2008). Morphological andgenetic evidence that the cyanobacterium Lyngbya wollei (Farlow exGomont) Spexiale and Dyck encompasses at least two species. ApplEnviron Microbiol 74, 3710–3717.
E. Valerio and others
654 Microbiology 155

Kaneko, T., Sato, S., Kotani, H., Tanaka, A., Asamizu, E.,Nakamura, Y., Miyajima, N., Hirosawa, M., Sugiura, M. & otherauthors (1996). Sequence analysis of the genome of the unicellular
cyanobacterium Synechocystis sp. strain PCC 6803. II. Sequence
determination of the entire genome and assignment of potential
protein-coding regions. DNA Res 3, 109–136.
Kaneko, T., Nakamura, Y., Wolk, C. P., Kuritz, T., Sasamoto, S.,Watanabe, A., Iriguchi, A., Ishikama, K., Kawashima, K. & otherauthors (2001). Complete genomic sequence of the filamentous
nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. DNA
Res 8, 205–213.
Komarek, J. & Anagnostidis, K. (1986). Modern approach to the
classification system of Cyanophytes. 2. Chroococcales. Algol Stud 43,
157–226.
Komarek, J. & Anagnostidis, K. (1989). Modern approach to the
classification system of Cyanophytes. 4. Nostocales. Algol Stud 56,
247–345.
Kondo, R., Yoshida, T., Yusi, Y. & Hiroishi, S. (2000). DNA–DNA
reassociation among a bloom-forming cyanobacterial genus,
Microcystis. Int J Syst Evol Microbiol 50, 767–770.
Laloui, W., Palinska, K. A., Rippka, R., Partensky, F., Tandeau deMarsac, N., Herdman, M. & Iteman, I. (2002). Genotyping of axenic
and non-axenic isolates of the genus Prochlorococcus and the OMF-
‘Synechococcus’ clade by size, sequence analysis or RFLP of the internal
transcribed spacer of the ribosomal operon. Microbiology 148,
453–465.
Litvaitis, M. K. (2002). A molecular test of cyanobacterial phylogeny:
inferences from constraint analyses. Hydrobiologia 468, 135–145.
Lu, W., Evans, H., McColl, S. M. & Saunders, V. A. (1997).Identification of cyanobacteria by polymorphisms of PCR-amplified
ribosomal DNA spacer region. FEMS Microbiol Lett 153, 141–149.
Lyra, C., Hantula, J., Vainio, E., Rapala, J., Rouhiainen, L. & Sivonen, K.(1997). Characterization of cyanobacteria by SDS-PAGE of whole-cell
proteins and PCR/RFLP of the 16S rRNA gene. Arch Microbiol 168,
176–184.
Lyra, C., Suomalainen, S., Gugger, M., Vezie, C., Sundman, P.,Paulin, L. & Sivonen, K. (2001). Molecular characterization of
planktic cyanobacteria of Anabaena, Aphanizomenon, Microcystis
and Planktothrix genera. Int J Syst Evol Microbiol 51, 513–526.
Lyra, C., Laamanen, M., Lehtimaki, J. M., Surakka, A. & Sivonen, K.(2005). Benthic cyanobacteria of the genus Nodularia are non-toxic,
without gas vacuoles, able to glide and genetically more diverse than
planktonic Nodularia. Int J Syst Evol Microbiol 55, 555–568.
Marquardt, J. & Palinska, K. A. (2007). Genotypic and phenotypic
diversity of cyanobacteria assigned to the genus Phormidium
(Oscillatoriales) from different habitats and geographical sites. Arch
Microbiol 187, 397–413.
Masepohl, B., Gorlitz, K. & Bohme, H. (1996). Long tandemly
repeated repetitive (LTRR) sequences in the filamentous cyanobac-
terium Anabaena sp. PCC 7120. Biochim Biophys Acta 1307, 26–30.
Mazel, D., Houmard, J., Castets, A. M. & Tandeau de Marsac, N.(1990). Highly repetitive DNA sequences in cyanobacterial genomes.
J Bacteriol 172, 2755–2761.
Neilan, B. A. (2002). The molecular evolution and DNA profiling of
toxic cyanobacteria. Curr Issues Mol Biol 4, 1–11.
Nilsson, M., Bergman, B. & Rasmussen, U. (2000). Cyanobacterial
diversity in geographically related and distant host plants of the genus
Gunnera. Arch Microbiol 173, 97–102.
Nubel, U., Garcial-Pichel, F. & Muyzer, G. (1997). PCR primers to
amplify 16S rRNA genes from Cyanobacteria. Appl Environ Microbiol
63, 3327–3332.
Otsuka, S., Suda, S., Shibata, S., Oyaizu, H., Matsumoto, S. &Watanabe, M. M. (2001). A proposal for the unification of five speciesof the cyanobacterial genus Microcystis Kutzing ex Lemmermann 1907under the rules of the bacteriological code. Int J Syst Evol Microbiol 51,873–879.
Pitcher, D., Saunders, N. & Owen, R. (1989). Rapid extraction of bacterialDNA with guanidium thyocinate. Lett Appl Microbiol 8, 151–156.
Prasanna, R., Kumar, R., Sood, A., Prasanna, B. M. & Singh, P. K.(2006). Morphological, physiochemical and molecular characteriza-tion of Anabaena strains. Microbiol Res 161, 187–202.
Priest, F. & Austin, B. (1993). Classification. In Modern BacterialTaxonomy, 2nd edn, pp. 1–13. London: Chapman & Hall.
Rajaniemi, P., Hrouzek, P., Kastovska, K., Willame, R., Rantala, A.,Hoffmann, L., Komarek, J. & Sivonen, K. (2005). Phylogenetic andmorphological evaluation of the genera Anabaena, Aphanizomenon,Trichormus and Nostoc (Nostocales, Cyanobacteria). Int J Syst EvolMicrobiol 55, 11–26.
Rasmussen, U. & Svenning, M. M. (1998). Fingerprinting ofcyanobacteria based on PCR with primers derived from short andlong tandemly repeated repetitive sequences. Appl Environ Microbiol64, 265–272.
Robertson, B. R., Tezuka, N. & Watanabe, M. M. (2001). Phylogeneticanalyses of Synechococcus strains (cyanobacteria) using sequences of16S rDNA and part of the phycocyanin operon reveal multipleevolutionary lines and reflect phycobilin content. Int J Syst EvolMicrobiol 51, 861–871.
Rocap, G., Distel, D. L., Waterbury, J. B. & Chisholm, S. W. (2002).Resolution of Prochlorococcus and Synechococcus ecotypes by using16S–23S ribosomal DNA internal transcribed spacer sequences. ApplEnviron Microbiol 68, 1180–1191.
Rudi, K., Skulberg, O. M., Larsen, F. & Jakobsen, K. S. (1997). Straincharacterization and classification of oxyphotobacteria in clonecultures on the basis of 16S rRNA sequences from the variableregions V6, V7, and V8. Appl Environ Microbiol 63, 2593–2599.
Saker, M. L., Nogueira, I. C. G., Vasconcelos, V. M., Neilan, B. A.,Eaglesham, G. H. & Pereira, P. (2003). First report and toxicologicalassessment of the cyanobacterium Cylindrospermopsis raciborskii fromPortuguese freshwaters. Ecotoxicol Environ Saf 55, 243–250.
Seo, P. S. & Yokota, A. (2003). The phylogenetic relationships ofcyanobacteria inferred from 16S rRNA, gyrB, rpoC1 and rpoD1 genesequences. J Gen Appl Microbiol 49, 191–203.
Skulberg, R. & Skulberg, O. M. (1990). Forskning med AlgekulturerNIVAs Kultursampling av Alger (Research with Algal Cultures. NIVA’sCulture Collection of Algae). Oslo, Norway: Norsk Institutt forVannforskning.
Sneath, P. H. A. & Johnson, R. (1972). The influence on numericaltaxonomic similarities of errors in microbiological tests. J GenMicrobiol 72, 377–392.
Suda, S., Watanabe, M. M., Otsuka, S., Mahakahant, A.,Yongmanitchai, W., Norpartnaraporn, N., Liu, Y. & Day, J. G.(2002). Taxonomic revision of water-bloom-forming species ofoscillatorioid cyanobacteria. Int J Syst Evol Microbiol 52, 1577–1595.
Swofford, D. L. (2003). PAUP. Phylogenetic analysis using parsimony(and other methods). Version 4.0b10. Sinauer Associates, Sunderland,Massachusetts.
Teaumroong, N., Innok, S., Chunleuchanon, S. & Boonkerd, N.(2002). Diversity of nitrogen-fixing cyanobacteria under variousecosystems of Thailand. I. Morphology, physiology and geneticdiversity. World J Microbiol Biotechnol 18, 673–682.
Valerio, E., Pereira, P., Saker, M. L., Franca, S. & Tenreiro, R. (2005).Molecular characterization of Cylindrospermopsis raciborskii strainsisolated from Portuguese freshwaters. Harmful Algae 4, 1044–1052.
Molecular diagnostics for cyanobacteria
http://mic.sgmjournals.org 655

van Belkum, A., Scherer, S., van Alphen, L. & Verbrugh, H. (1998).Short-sequence DNA repeats in prokaryotic genomes. Microbiol MolBiol Rev 62, 275–293.
Versalovic, J., Koeuth, T. & Lupski, J. R. (1991). Distribution ofrepetitive DNA sequences in eubacteria and application to finger-printing of bacterial genomes. Nucleic Acids Res 19, 6823–6831.
Vincze, T., Posfai, J. & Roberts, R. J. (2003). NEBcutter: a program tocleave DNA with restriction enzymes. Nucleic Acids Res 31, 3688–3691.
West, N. J. & Adams, D. G. (1997). Phenotypic and genotypiccomparison of symbiotic and free-living cyanobacteria from a singlefield site. Appl Environ Microbiol 63, 4479–4484.
Wilson, K. M., Schembri, M. A., Baker, P. D. & Saint, C. P. (2000).Molecular characterization of the toxic cyanobacterium Cylindrosper-
mopsis raciborskii and design of a species-specific PCR. Appl Environ
Microbiol 66, 332–338.
Zar, J. H. (1996). Biostatistical Analysis, 3rd edn. Upper Saddle River,
NJ: Prentice-Hall International.
Zheng, W. W., Nilsson, M., Bergman, B. & Rasmussen, U. (1999).Genetic diversity and classification of cyanobacteria in different Azolla
species by the use of PCR fingerprinting. Theor Appl Genet 99, 1187–
1193.
Edited by: D. J. Scanlan
E. Valerio and others
656 Microbiology 155
Related Documents











