Abstract Myeloperoxidase (MPO) belongs to a family of related proteins which also includes eosinophil, thyroid, and lactoperoxidase. The MPO gene is a 14-kb gene locat- ed on the long arm of chromosome 17. Thus far four muta- tions (R569W, Y173C, M251T and a 14-base deletion in exon 9) have been identified in patients with MPO defi- ciency. As in other genetically determined diseases, many more mutations will eventually be revealed that cause this disease. Present evidence shows that most patients are compound heterozygotes, i.e., they have inherited different mutations on their paternal and maternal MPO alleles. Un- derstanding why some patients with this genetic deficiency develop clinical symptoms while others do not requires mutation analyses of a large number of patients. This in- cludes the analysis of genotype-phenotype relationships. Genotyping has also been started in patients with EPO-de- ficiency. Key words Peroxidase deficiency · Mutation analysis · Genotype-phenotype relationship Abbreviations DGGE Denaturing gradient gel electro- phoresis · EPO Eosinophil peroxidase · LPO Lactoperoxidase MPO Myeloperoxidase · PCR Polymerase chain reaction · RFLP Restriction fragment length polymorphism · SSCP Single-strand conformation polymorphism · TPO Thyroid peroxidase Communicated by: William M. Nauseef and Petro E. Petrides P.E. Petrides Division of Oncology and Hematology, School of Medicine, Humboldt University, Charité Campus Mitte, Schumannstrasse 20/21, D-10117 Berlin, Germany E-mail: petrides@charité.de J Mol Med (1998) 76:688–698 © Springer-Verlag 1998 REVIEW Petro E. Petrides Molecular genetics of peroxidase deficiency Received: 22 January 1998 / Accepted: 13 May 1998 Introduction In humans there are several peroxidases that are important for cell physiology: myeloperoxidase (MPO) in neutro- phils, eosinophil peroxidase (EPO) in eosinophils, thyroid peroxidase (TPO) in thyroid cells, and lactoperoxidase (LPO) secreted in milk. These enzymes belong to a family of structurally and functionally related proteins. Here, I re- view present knowledge of the structure, regulation, and mutations of the MPO and EPO genes. Cloning and structure of the MPO gene MPO enzyme synthesis is restricted to late myeloblasts and promyelocytes in the bone marrow. MPO was first isolated in 1941 [1] and its deficiency described in 1966 [2], but it was only in the middle 1980s that the cDNA for this enzyme was cloned by several laboratories, when Chang et al. [3], Johnson et al. [4, 5], Weil et al. [6], Yamada et al. [7], and Morishita et al. [8] reported the characterization of human MPO cDNA. Analysis of the genomic DNA by Morishita et al. [9] revealed a single gene of approximately 14 kb, com- posed of 11 introns and 12 exons (Fig. 1). Soon thereafter the gene was localized to the long arm of chromosome 17 in segment q11–21 [6], q22–24 [10], or q21.3–23 [11, 12]. This is in close proximity to the breakpoint for the translocation PETRO E. PETRIDES received his M.D. from the University of Munich in 1975. From 1978 until 1984 he worked at the Salk Institute in La Jolla and at Stanford Uni- versity Medical School, Palo Alto, CA. From 1984 to 1998 he was on the faculty of the Grosshadern Medical School of the University of Munich. He is currently Clinical Professor of Internal Medicine in the Divi- sion of Oncology and Hematol- ogy at the Charité Medical School at the Humboldt Uni- versity of Berlin. His major re- search interests are genotype- phenotype relationships in various hematological disorders and the role of proteinases in myeloid leukemias.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abstract Myeloperoxidase (MPO) belongs to a family ofrelated proteins which also includes eosinophil, thyroid,and lactoperoxidase. The MPO gene is a 14-kb gene locat-ed on the long arm of chromosome 17. Thus far four muta-tions (R569W, Y173C, M251T and a 14-base deletion inexon 9) have been identified in patients with MPO defi-ciency. As in other genetically determined diseases, manymore mutations will eventually be revealed that cause thisdisease. Present evidence shows that most patients arecompound heterozygotes, i.e., they have inherited differentmutations on their paternal and maternal MPO alleles. Un-derstanding why some patients with this genetic deficiencydevelop clinical symptoms while others do not requiresmutation analyses of a large number of patients. This in-cludes the analysis of genotype-phenotype relationships.
Genotyping has also been started in patients with EPO-de-ficiency.
Key words Peroxidase deficiency · Mutation analysis ·Genotype-phenotype relationship
Abbreviations DGGE Denaturing gradient gel electro-phoresis · EPO Eosinophil peroxidase · LPO Lactoperoxidase MPO Myeloperoxidase · PCR Polymerase chain reaction · RFLP Restrictionfragment length polymorphism · SSCP Single-strandconformation polymorphism · TPO Thyroid peroxidase
Communicated by: William M. Nauseef and Petro E. Petrides
P.E. PetridesDivision of Oncology and Hematology, School of Medicine, Humboldt University, Charité Campus Mitte, Schumannstrasse 20/21, D-10117 Berlin, GermanyE-mail: petrides@charité.de
J Mol Med (1998) 76:688–698 © Springer-Verlag 1998
REVIEW
Petro E. Petrides
Molecular genetics of peroxidase deficiency
Received: 22 January 1998 / Accepted: 13 May 1998
Introduction
In humans there are several peroxidases that are importantfor cell physiology: myeloperoxidase (MPO) in neutro-phils, eosinophil peroxidase (EPO) in eosinophils, thyroidperoxidase (TPO) in thyroid cells, and lactoperoxidase(LPO) secreted in milk. These enzymes belong to a familyof structurally and functionally related proteins. Here, I re-view present knowledge of the structure, regulation, andmutations of the MPO and EPO genes.
Cloning and structure of the MPO gene
MPO enzyme synthesis is restricted to late myeloblasts andpromyelocytes in the bone marrow. MPO was first isolatedin 1941 [1] and its deficiency described in 1966 [2], but itwas only in the middle 1980s that the cDNA for this enzymewas cloned by several laboratories, when Chang et al. [3],Johnson et al. [4, 5], Weil et al. [6], Yamada et al. [7], andMorishita et al. [8] reported the characterization of humanMPO cDNA. Analysis of the genomic DNA by Morishita etal. [9] revealed a single gene of approximately 14 kb, com-posed of 11 introns and 12 exons (Fig. 1). Soon thereafterthe gene was localized to the long arm of chromosome 17 insegment q11–21 [6], q22–24 [10], or q21.3–23 [11, 12]. Thisis in close proximity to the breakpoint for the translocation
PETRO E. PETRIDES
received his M.D. from theUniversity of Munich in 1975.From 1978 until 1984 heworked at the Salk Institute inLa Jolla and at Stanford Uni-versity Medical School, PaloAlto, CA. From 1984 to 1998he was on the faculty of theGrosshadern Medical School ofthe University of Munich. He iscurrently Clinical Professor ofInternal Medicine in the Divi-sion of Oncology and Hematol-ogy at the Charité MedicalSchool at the Humboldt Uni-versity of Berlin. His major re-search interests are genotype-phenotype relationships invarious hematological disordersand the role of proteinases inmyeloid leukemias.

689
a
b
Fig. 1 for Continuation and legend see page 690

of acute promyelocytic leukemia, which caused a specula-tion that this gene is involved in acute promyelocytic leuke-mia [13]. This, however, has not been confirmed.
Existence of a peroxidase multigene family
Protein and gene analytical investigations have led to theconclusion that MPO, EPO, TPO, and LPO belong to a su-perfamily of related proteins [14–16]. Homology at theamino acid level between MPO and EPO is 68%, between
690
Fig. 1 a Chromosomal organization of the human MPO gene with12 exons and 11 introns. Top line, size scale (in kilobases) [9]. b,cNucleotide sequence of the human MPO gene. (Genbank entries arefor cDNA of MPO X04876 and for the gene X64647.) (From [5])
cContinuation of Fig. 1
691
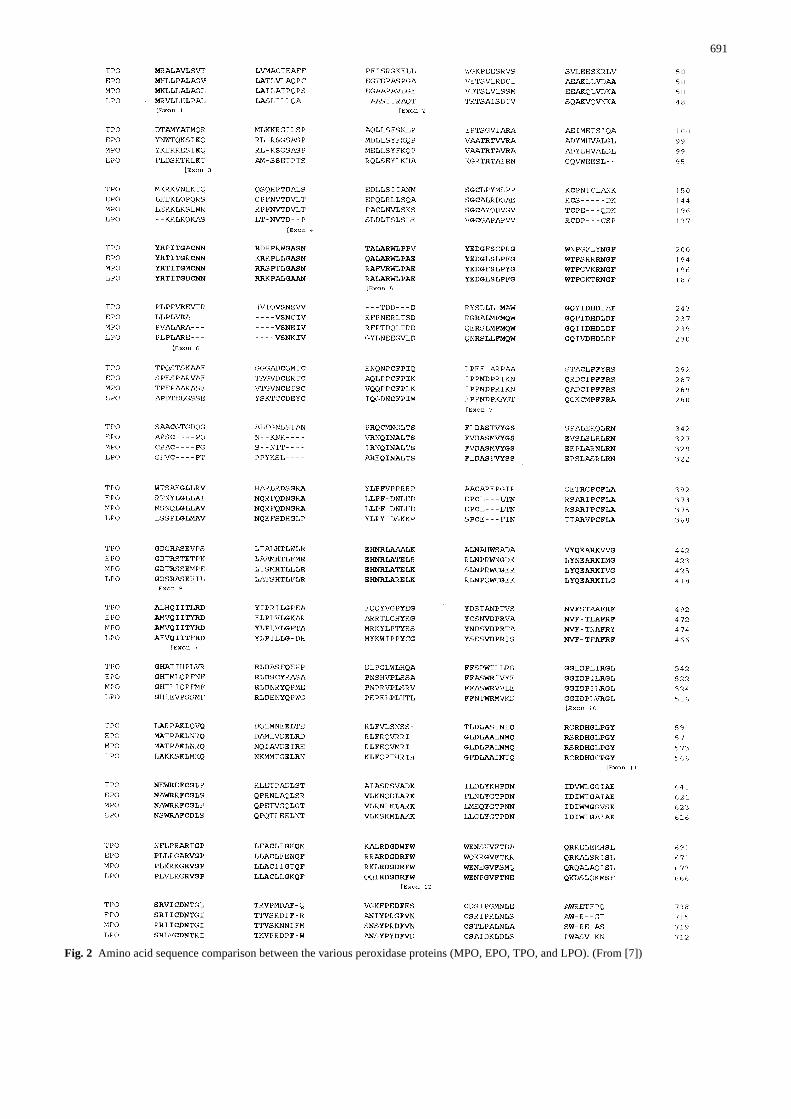
Fig. 2 Amino acid sequence comparison between the various peroxidase proteins (MPO, EPO, TPO, and LPO). (From [7])

EPO and TPO 44%, and between LPO vs. MPO/EPO 51%(Fig. 2).
Transcriptional regulation of the MPO gene
Transcription of the MPO gene is tightly regulated in a tis-sue- and differentiation-specific manner. Alternative splic-ing of hnRNA leads to various transcripts of different sizes(e.g., 3.6 and 2.9 kb) [17]. Progressive demethylation in the5’ flanking region of the MPO gene is a prerequisite fortranscription [18, 19]. Expression of the gene is regulatedby the cell-specific transcription factor AML-1; the integri-ty of the AML-1 binding site is essential for the activity ofthe MPO proximal enhancer [20, 21]. Austin et al. [22, 23]have identified the basic or minimal human MPO promotorin the proximal 5’ flanking DNA between bp –128 and +11and shown that adjacent DNA sequences enhance promotoractivity. Seven discrete nuclear binding sites (DP1–DP7)are present within the proximal 600-bp flanking region ofthe MPO gene (Table 1). Since experimental mutation ofsite DP7 stimulates and mutations of DP1–DP6 reduce pro-motor activity, it is likely that these cis elements contributein vivo to the activity of the MPO promotor [24, 25]. TheMPO promotor also contains an Alu element with bindingsites for retinoic acid and thyroid hormone receptors [26].There are two MPO alleles which differ at one known posi-tion (–463 G/A) in this region.
This single base substitution (G→A) can occur as a so-matic mutation in acute myelocytic leukemia cells [24, 25]and as an inherited polymorphism with functional signifi-cance in vitro [26]; the presence of an A rather than a Gdecreases expression by the abolition of a binding site forthe SP1 transcription factor. The wild type is thereforecalled SP, while the mutated allele has been termed N (for“no binding” of SP-1).
When HL60 cells (a leukemia cell line) are induced todifferentiate into phenotypically normal granulocytes ormonocytes by tetradecanoylphorbol acetate, dimethylsul-foxide, or retinoic acid [6, 27–30], they cease to transcribethe MPO gene. Similar results have been obtained by insitu hybridization [31]. Downregulation of the MPO genetranscription is associated with an alteration of specific nu-clease sensitive sites [32].
In normal human bone marrow cytokines such as tumornecrosis factor-α decrease MPO transcription [33]. Granu-locyte colony-stimulating factor induced differentiation ofmultipotential progenitor cells results in activation of pro-
teins of the Pu1 and C-EBP family and their recruitment tothe nucleus where they bind to a distal MPO upstream en-hancer [34]. In addition, the expression of the MPO geneis regulated via the above proximal enhancer by c-myb andAML-1 [35].
MPO gene expression in leukemias
MPO gene expression is found in acute myeloid leukemias[36–39]. Interestingly, in some patients with acute lym-phoblastic leukemia MPO expression has also been ob-served, but no MPO protein has been found either byWestern blotting or cytochemical methods [36, 40, 41].The reasons for this observation are still unclear. An asso-ciation between MPO deficiency and acute myeloid leuke-mia has been reported in one individual [42]. The SP/SPgenotype (see above) is correlated with increased MPOmRNA levels in primary myeloid leukemia cells and over-represented in AML-M3 and AML-M4 [43].
MPO genetic polymorphism and cancer risk
Since MPO activates carcinogens in tobacco smoke, in-cluding benzo[a]pyrene and aromatic amines, the hypothe-sis was tested that lower MPO transcription reduces therisk of lung cancer [44]. Restriction fragment length poly-morphism (RFLP)/polymerase chain reaction (PCR) anal-ysis of 1000 individuals revealed 8–9% persons to be ho-mozygous for the N-allele. These individuals may be at adecreased risk of lung cancer.
Genetics of peroxidase deficiency
Hereditary and acquired forms of MPO deficiency
MPO deficiency was first described in 1966 [2]. Severalinvestigators have discussed, for example, the allele fre-quency and mode of inheritance of this entity [45–48] (fordetails see the contributions by Nauseef and Kutter, thisvolume). Deficiency can be either acquired or hereditary,depending upon whether germline and/or somatic muta-tions are involved. If germline mutations are involved,both alleles must be mutated to yield the clinical pheno-type (Fig. 3).
692
Site Sequence Location (bp)
DP1 CATAA –27 to –23DP2 CACCCCACCCCCAGC –49 to –35DP3 GTGGGGAGGAGA –100 to –89DP4 CCCTCCTTCCTGCCCCTTCCCCC –160 to –138DP5 GAGCAAGATAACCGTCT –219 to –203DP6 TCCCAGCTACTCGGGAGG –285 to –268DP7 GCAGTGGATCACTTGAGGTCAGGAGTTCAAGACCAGCCTGG –466 to –425
Table 1 Cis elements(DP1–DP7) in the 5’ flankingregion of the human MPO gene(from [24])
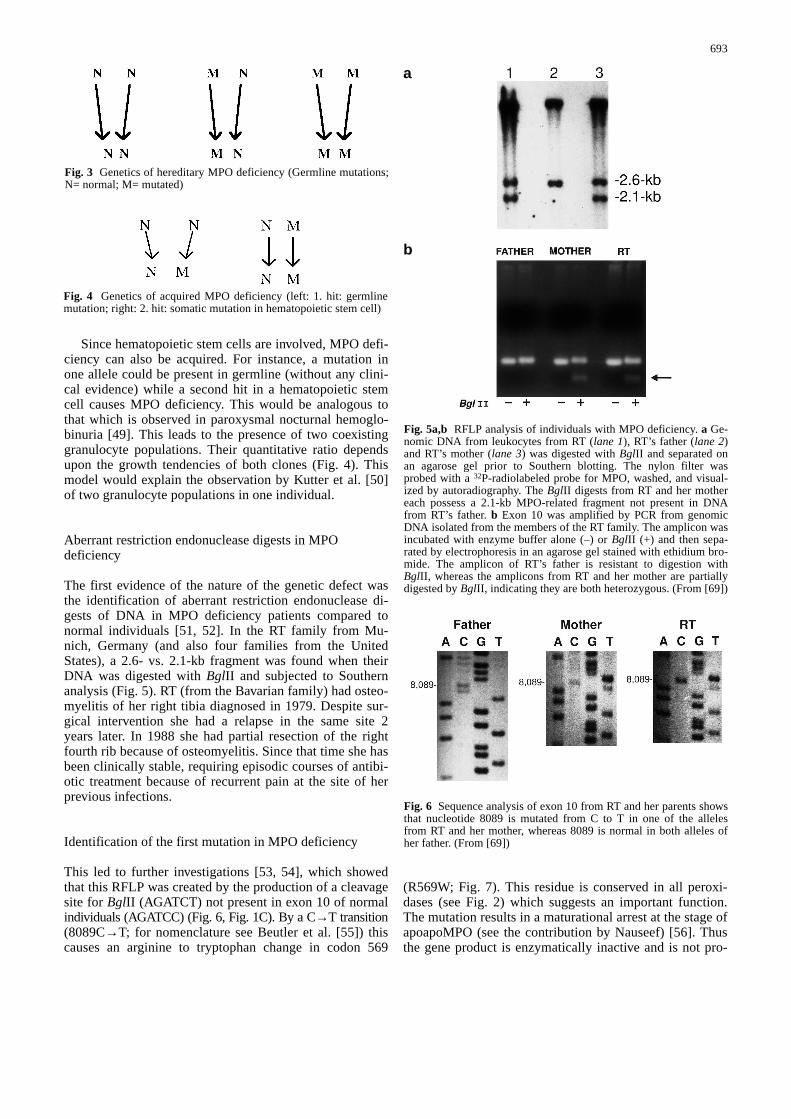
Since hematopoietic stem cells are involved, MPO defi-ciency can also be acquired. For instance, a mutation inone allele could be present in germline (without any clini-cal evidence) while a second hit in a hematopoietic stemcell causes MPO deficiency. This would be analogous tothat which is observed in paroxysmal nocturnal hemoglo-binuria [49]. This leads to the presence of two coexistinggranulocyte populations. Their quantitative ratio dependsupon the growth tendencies of both clones (Fig. 4). Thismodel would explain the observation by Kutter et al. [50]of two granulocyte populations in one individual.
Aberrant restriction endonuclease digests in MPOdeficiency
The first evidence of the nature of the genetic defect wasthe identification of aberrant restriction endonuclease di-gests of DNA in MPO deficiency patients compared tonormal individuals [51, 52]. In the RT family from Mu-nich, Germany (and also four families from the UnitedStates), a 2.6- vs. 2.1-kb fragment was found when theirDNA was digested with BglII and subjected to Southernanalysis (Fig. 5). RT (from the Bavarian family) had osteo-myelitis of her right tibia diagnosed in 1979. Despite sur-gical intervention she had a relapse in the same site 2years later. In 1988 she had partial resection of the rightfourth rib because of osteomyelitis. Since that time she hasbeen clinically stable, requiring episodic courses of antibi-otic treatment because of recurrent pain at the site of herprevious infections.
Identification of the first mutation in MPO deficiency
This led to further investigations [53, 54], which showedthat this RFLP was created by the production of a cleavagesite for BglII (AGATCT) not present in exon 10 of normalindividuals (AGATCC) (Fig. 6, Fig. 1C). By a C→T transition(8089C→T; for nomenclature see Beutler et al. [55]) thiscauses an arginine to tryptophan change in codon 569
(R569W; Fig. 7). This residue is conserved in all peroxi-dases (see Fig. 2) which suggests an important function.The mutation results in a maturational arrest at the stage ofapoapoMPO (see the contribution by Nauseef) [56]. Thusthe gene product is enzymatically inactive and is not pro-
693
a
b
Fig. 5a,b RFLP analysis of individuals with MPO deficiency. a Ge-nomic DNA from leukocytes from RT (lane 1), RT’s father (lane 2)and RT’s mother (lane 3) was digested with BglII and separated onan agarose gel prior to Southern blotting. The nylon filter wasprobed with a 32P-radiolabeled probe for MPO, washed, and visual-ized by autoradiography. The BglII digests from RT and her mothereach possess a 2.1-kb MPO-related fragment not present in DNAfrom RT’s father. b Exon 10 was amplified by PCR from genomicDNA isolated from the members of the RT family. The amplicon wasincubated with enzyme buffer alone (–) or BglII (+) and then sepa-rated by electrophoresis in an agarose gel stained with ethidium bro-mide. The amplicon of RT’s father is resistant to digestion withBglII, whereas the amplicons from RT and her mother are partiallydigested by BglII, indicating they are both heterozygous. (From [69])
Fig. 6 Sequence analysis of exon 10 from RT and her parents showsthat nucleotide 8089 is mutated from C to T in one of the allelesfrom RT and her mother, whereas 8089 is normal in both alleles ofher father. (From [69])
Fig. 3 Genetics of hereditary MPO deficiency (Germline mutations;N= normal; M= mutated)
Fig. 4 Genetics of acquired MPO deficiency (left: 1. hit: germlinemutation; right: 2. hit: somatic mutation in hematopoietic stem cell)

cessed or targeted to the mature lysosomal form of MPO.Similar R→W mutations occur in phenylketonuria(R408W) and acute intermittent porphyria (R116W,R167W, R173W, or R201W). In the latter case the porpho-bilinogen deaminase protein is involved where the loss ofthe arginine side chain leads to the interruption of stabiliz-ing salt bridges within the protein or between the enzymeprotein and its substrate. C→T transitions are mutationalhotspots; methylated cytosine residues can be deaminated,which leads through a keto-enol tautomeric change tothymine. These residues pair with adenine residues duringDNA replication, which causes the C→T transition (Fig.8). Additional mutations have recently been identified thatcause a substitution of tyrosine in position 173 by cysteine(Y173C) (F.R. DeLeo, M. Goedken, S.J. McCormick,W.M. Nauseef, submitted), a substitution of methionine bythreonine in position 251 (M251T) or a 14-base deletion inexon 9 [57]. In addition to mutations which cause mis-sense mutations, structural alterations in the regulatoryparts of the MPO gene have been postulated which causepretranslational defects [58, 59].
Methods for screening for mutations in MPO deficiency
Since patients with the R569W mutation on one allelehave a complete deficiency, either an dominant effect ormore likely a second mutation on the other allele must bepostulated to explain this complete loss of enzymatic ac-
tivity. Mutation analysis in other genetic diseases has re-vealed that more than 400 different mutations can occur ina given gene (Table 2). Some of these mutations occur at arelatively high frequency while others are observed only incertain families and are therefore referred to as private mu-tations [65].
Since DNA sequencing is time consuming and expen-sive, effective and economical DNA-scanning methods are
694
Disease Protein Gene size (kb) Number of mutations Reference
Cystic fibrosis CFTR 250 400 [60]Phenylketonuria Ph hydroxylase 90 240 [61]Gaucher disease β-Glucocerebrosidase 7.3 80 [62]Breast cancer BCRA1 8 235 [63]Acute porphyria Porphobilinogen deaminase 10 60 [64]PNH GPI synthase 17 100 [49]MPO deficiency MPO 14 4 This article
Table 2 Mutation analysis invarious genetic disorders
Fig. 7 The C to T transition incodon 569 (8089C→T) in exon10 of the MPO gene leads to asubstitution in the protein ofarginine by tryptophan, whichcannot form electrostatic bonds
Fig. 8 Deamination of cytosine or 5-methylcytosine to uracil orthymine, which leads during DNA replication to a C→T transition

necessary [66]. If the position of the mutation is alreadyknown, the exon (or intron) containing it can be amplifiedby PCR and the reaction product is then subjected toRFLP analysis if the mutation leads to an alteration of arestriction enzyme cleavage site (Fig. 5). Utilizing such anapproach, we have identified 70% of mutated alleles in 27patients with Gaucher disease when we assayed their DNAfor eight known mutations and the gene deletion [62, 67].
If there are no restriction enzyme cleavage sites in thearea of the mutation, or if the mutation is not yet known,physical methods such as denaturing gradient gel electro-phoresis (DGGE) and single-strand conformation poly-morphism (SSCP) are required. DGGE is based on theprinciple that DNA duplexes which differ by a basepairhave a different temperature or denaturant concentration atwhich they melt. When melted, they stop migration in agel. Thus duplexes differing by a basepair stop at differentpositions, which correspond to different concentrations ofdenaturant or temperature and therefore indicate the pres-ence of a mutation. The detection is more evident whenheteroduplexes are formed, and a nonmelting region or aregion difficult to melt (clamp technique) is added to oneof the amplification primers. An example of this techniqueis shown in Fig. 9.
SSCP analysis is based upon the observation that astrand of single-stranded DNA folds differently from an-other if it differs by a single base. This leads to different
mobilities for these two strands in nondenaturing gel elec-trophoresis. In addition to these techniques, mismatchcleavage methods are available.
The newly developed DNA chips [68] rely on hybrid-ization of labeled test single-strand DNA to an array ofknown oligonucleotides on a very small physical substrate.Different patterns of hybridization between wild-type andtest samples signal the presence of a mutation.
The ideal mutation scanning method, would detect100% of the mutations and would be able to screenkilobase lengths in a single step, economical, and easy tohandle; however, this has yet not been developed.
Compound heterozygosity in MPO deficiency
Analysis of various families with genetic MPO deficiencyfor peroxidase activity, immunoreactivity, and the presenceof the R569W mutation indicates that most patients arecompound heterozygotes, i.e., they have inherited differentalleles from their parents [69]. In the T family from Ger-many (Table 3) patient RT has a complete loss of MPOand immunoreactivity but is heterozygous for the R569Wmutation. She inherited this mutation from her motherRoT, who has 50% immunoreactivity and MPO activity.Since her father, GT, has approx. 75% MPO and immu-noreactivity, he must have another, still unknown, allelewhich he passed to his daughter. The combination ofR569W and this allele causes a complete loss of activity.
The presence of many mutations reported in various genet-ic disorders (see Table 1) has revealed that a large number ofdifferent allele combinations determine the genotype. Figure10 illustrates how the combination of different gene productswith differing residual enzymatic activity can yield a widespectrum of resulting total activities. This explains in part thewide clinical spectrum (phenotype) observed in various genet-ic disorders. In MPO deficiency the situation is further compli-
695
Fig. 9 DGGE analysis in a fam-ily with the autosomal dominantacute intermittent porphyria forthe identification of gene carri-ers. PCR-amplified oligonucle-otides form one duplex type(wild-type/wild-type) in normalsand three duplex types in genecarriers (wild-type/wild-type,wild-type/mutated allele andmutated/mutated allele). Sevengene carriers were identified inthe family of the patient (no.12). (From [64])
Table 3 MPO activity, immunoreactivity, and presence of theR569W mutation in a German kindred with MPO deficiency [69]
MPO activity Immunoreactivity R569W
RT (daughter) 0 0 +/–GT (father) 73 74 –/–RoT (mother) 53 47 +/–

696
cated since the enzyme is formed by proteolysis from a precur-sor and forms a tetramer which must be transported throughthe endoplasmic reticulum into the azurophilic granules.
Mutation analysis in patients with EPO deficiency
EPO is a highly basic protein present in the specific gran-ules of eosinophilic granulocytes. It is a 70-kDa dimercomposed of a 15-kDa light chain and a 55-kDa heavychain held together by a disulfide bridge. EPO deficiencywas first described by Presentey [71], and since then about100 individuals have been reported with this abnormality[72–76]. After the EPO gene had been cloned [15, 77], thefirst mutation analysis in an individual with hereditaryEPO deficiency was described. The patient was compoundheterozygous, i.e., a G→A transition caused a replacementof an arginine with a histidine (R286H) on one allele andan insertion in exon 10 caused a out of frame reading withthe production of a premature stop on the other allele.
Future directions
The cloning of MPO and EPO genes began the analysis ofthe molecular defect in individuals with MPO and EPOdeficiency. Systematic screening for mutations in a largenumber of patients will enable us to understand the molec-ular nature of these common defects and to explain theirclinical consequences. Moreover, a more detailed knowl-edge of the molecular basis of MPO gene expression andthe importance of the N/N genotype should lead to an un-derstanding of the relevance of this system to cancer de-velopment.
Acknowledgements The author’s own research reported here wassupported in part by DFG grant Pe 258/20-1 and 2.
References:
1. Agner K (1941) Verdoperoxidase. A ferment isolated from leu-kocytes. Acta Physiol Scand 2 [Suppl 8]
2. Undritz E (1966) Die Alius-Grignaschi Anomalie: der erblich-konstitutionelle Peroxydasedefekt der Neutrophilen und Monozy-ten. Blut 14:129–136
3. Chang KS, Trujillo JM, Cook RG, Stass SA (1986) Humanmyeloperoxidase gene: molecular cloning and expression in leu-kemic cells. Blood 68:1411–1414
4. Johnson KR, Nauseef WM, Care A, Wheelock MJ, Shane S, Ko-effler HP, Selsted M, Miller C, Rovera G (1987) Characterizationof cDNA clones for human myeloperoxidase: predicted aminoacid sequence and evidence for multiple mRNA species. NucleicAcids Res 15:2013–2028
5. Johnson K, Gemperlein I, Hudson S, Shane S, Rovera G (1989)Complete nucleotide sequence of human myeloperoxidase gene.Nucleic Acids Res 17:7985–7986
6. Weil SC, Rosner GL, Reid MS, et al (1987) cDNA cloning of hu-man myeloperoxidase: decrease in myeloperoxidase mRNA uponinduction of HL60 cells. Proc Natl Acad Sci USA 84:2057–2061
7. Yamada M, Hur SJ, Hashinaka K, et al (1987) Isolation and char-acterization of a cDNA coding for human myeloperoxidase. ArchBiochem Biophys 255:147–155
8. Morishita K, Kubota N, Asano S, Kaziro Y, Nagata S (1987) Mo-lecular cloning and characterization of cDNA for humanmyeloperoxidase. J Biol Chem 262:3844–3851
9. Morishita K, Tsuchiya M, Asano S, et al (1987) Chromosomalgene structure of human myeloperoxidase and regulation of itsexpression by granulocyte colony-stimulating factor. J Biol Chem262:15208–15213
10. Chang KS, Schroeder W, Siciliano MJ, et al (1987) The localiza-tion of the human myeloperoxidase gene is in close proximity tothe translocation breakpoint in acute promyelocytic leukemia.Leukemia 1:458–462
11. Inazawa J, Inoue K, Nishigaki H, Tsuda S, et al (1989) Assign-ment of the human myeloperoxidase gene (MPO) to bands q21.3-q23 of chromosome 17. Cytogen Cell Genet 50:135–136
12. Zaki SR, et al (1990) Chromosomal localization of the humanMPO gene by in situ hybridization using oligonucleotide probes.Genes Chromosomes Cancer 2:266–270
13. Van Tuinen P, et al (1987) Localization of myeloperoxidase to thelong arm of chromosome 17: relationship to the 15:17 transloca-tion of acute promyeloctic leukemia. Oncogene 1:319–322
14. Kimura S, Ikeda-Saito M (1988) Human myeloperoxidase andthyroid peroxidase, two enzymes with separate and distinct physi-ological functions, are evolutionary related members of the samegene family. Proteins 3:113–120
15. Ten RM, Pease LR, McKean DJ, Bell MP, Gleich GJ (1989) Mo-lecular cloning of the human eosinophil peroxidase. Evidence for
Fig. 10 Compound heterozygosity: combination of different allelescauses a different genotype and phenotype (e.g., resulting enzymaticactivity). (From [70])

immature myeloid cells without c-myb. Mol Cell Biol 17:5127–5135
36. Zaki SR, Austin GE, Swan D, et al (1989) Humanmyeloperoxidase gene and its expression in acute leukemia.Blood 74:2096–2102
37. Ferrari S, Tagliafico E, Ceccherrelli G, et al (1989) Expression ofthe myeloperoxidase gene in acute and chronic myeloid leuke-mias: relationship to the expression of cell cycle related genes.Leukemia 6:423–430
38. Morishita Y, Yano K (1989) Myeloperoxidase gene expression inacute leukemias. Acta Hematol 52:1459–1462
39. Ferrari S, Tagliafico E, Temperani P, et al (1990) Overexpressionof the MPO gene occurring in a case of APL without unusual ge-notypic characteristics. Leuk Res 14:735–742
40. Ferrari S, Mariano MT, Tagliafico E, et al (1988) Myeloper-oxidase gene expression in blast cells with a lymphoid pheno-type in cases of acute lymphoblastic leukemia. Blood 72:873–876
41. Crisan D, Topalovski M, O’Malley B (1996) MPO mRNA analy-sis in acute lymphoblastic leukemia. Diagn Mol Pathol 5:236–248
42. Huhn D, Belohradsky BH, Haas R (1978) FamiliärerMyeloperoxidasemangel und akute myeloische Leukämie. ActaHaematol 59:129–143
43. Reynolds W, Chang E, Douer D, Ball ED, Kanda V (1997) Anallelic association implicates myeloperoxidase in the etiology ofacute promyelocytic leukemia. Blood 90:2730–2737
44. London SJ, Lehman TA, Taylor JA (1997) Myeloperoxidase ge-netic polymorphism and lung cancer risk. Cancer Res 57:5001–5003
45. Parry MF, Root RK, Metcalf JA, et al (1981) Myeloperoxidasedeficiency. Ann Intern Med 95:293–301
46. Nauseef WM (1988) Myeloperoxidase deficiency. HematolOncol Clin North Am 2:135–158
47. Becker R, Pflüger KH (1994) Myeloperoxidase deficiency: anepidemiological study and flow cytometric detection of othergranular enzymes in myeloperoxidase deficient subjects. AnnHematol 69:199–203
48. Gerber C, et al (1996) Phagocytic activation and oxidative burstof granulocytes in persons with MPO deficiency. Eur J ClinChem Clin Biochem 34:901–908
49. Luzzatto L, Bessler M, Rotoli B (1997) Somatic mutations inparoxysmal nocturnal hemoglobinuria: a blessing disguise. Cell88:1–4
50. Kutter D, Thoma J, Al-Haidari K, Trierweiler P (1993) Coexist-ence of two distinct populations of neutrophilic granulocytes: onenormal and one partial deficiency. Acta Clin Belg 48:401–404
51. Nauseef WM (1989) Aberrant restriction endonuclease digests ofDNA from subjects with hereditary myeloperoxidase deficiency.Blood 73:290–295
52. Petrides PE, Cogley M, Bock S, Nauseef WM (1994) Molekular-patho-logie eines familiär bedingten Myeloperoxidase (MPO)-Mangels. Med Klinik 89:S1,154
53. Nauseef WM, Brigham S, Cogley M (1994) Hereditarymyeloperoxidase deficiency due to a missense mutation of argin-ine 569 to tryptophan. J Biol Chem 269:1212–1216
54. Kizaki M, Miller CW, Selsted ME, Koeffler HP (1994)Myeloperoxidase (MPO) gene mutation in hereditary MPO defi-ciency. Blood 83:1935–1940
55. Beutler E, McKusick VA, Motulsky AG, Scriver CR, HutchinsonF (1996) Mutation nomenclature: nicknames, systematic namesand unique identifiers. Hum Mutat 8:203–206
56. Nauseef WM, Cogley M, McCormick S (1996) Effect of theR569W mutation on the biosynthesis of myeloperoxidase. J BiolChem 271:9546–9549
57. Romano M, Dri P, Dalalt L, Patriarca P, Baralle FE (1997) Bio-chemical and molecular characterization of hereditarymyeloperoxidase deficiency. Blood 90:4126–4134
58. Tobler A, Selsted ME, Miller CW, et al (1989) Evidence for apretranslational defect in hereditary and acquired myeloper-oxidase deficiency. Blood 73:1980–1986
59. Selsted ME, Miller CW, Nowotny MJ, Morris WL, Koeffler HP(1993) Molecular analysis of MPO deficiency shows heteroge-
697
the existence of a peroxidase multigene family. J Exp Med169:2757–1769
16. Ueda T, et al (1997) Molecular cloning and characterization of thechromosomal gene for human lactoperoxidase. Eur J Biochem243:32–41
17. Hashikawa K, Nishio C, Hur SH, Sakiyama F, Tsunasawa S,Yamada M (1988) Multiple species of myeloperoxidase messen-ger mRNAs produced by alternative splicing and differentialpolyadenylation. Biochemistry 27:5906–5914
18. Lübbert M, Miller CW, Koeffler HP (1991) Changes of DNA me-thylation and chromatin structure in the human myeloperoxidasegene during myeloid differentiation. Blood 78:345–356
19. Hashinaka K, Yamada M (1992) Undermethylation and DNase Ihypersensitivity of myeloperoxidase gene in HL60 cells beforeand after differentiation. Arch Biochem Biophys 293:40–45
20. Suzow J, Friedman AD (1993) The murine myeloperoxidase pro-motor contains several functional elements one of which binds acell type restricted transcription factor, myeloid nuclear factor 1(MyNF1). Mol Cell Biol 13:2141–2151
21. Nuchprayoon I, Meyers S, Scott LM, et al (1994) PEBP2/CBF,the murine homologue of the human myeloid AML1 and PEBP2beta/CBF beta protooncogenes, regulates the murine myeloper-oxidase and neutrophil elastase genes in immature myeloid cells.Mol Cell Biol 14:5558–5568
22. Austin GE, Zhao WG, Zhang W, Austin ED, Findley HW, Murt-agh JJ (1995) Identification and characterization of the humanMPO promoter. Leukemia 9:848–857
23. Austin GE, et al (1993) Sequence comparison of putative regula-tory DNA of the 5’ flanking region of the myeloperoxidase genein normal and leukemic bone marrow cells. Leukemia 7:1445–1550
24. Zhao WG, Regmi A, Austin ED, et al (1996) Cis-elements in thepromoter region of the human myeloperoxidase (MPO) gene.Leukemia 10:1089–1103
25. Zhao WG, et al (1997) Identification and functional analysis ofmultiple murine MPO promotors and comparison with humanMPO promotor regions. Leukemia 11:97–105
26. Piedrafita FJ, Molander RB, Vansant G, et al (1996) An alu ele-ment in the myeloperoxidase promotor contains a composite SP1thyroid hormone retinoic acid response element. J Biol Chem271:14412–14420
27. Yamada M, Kurahashi K (1984) Regulation of myeloperoxidasegene expression during differentiation of human myeloid leuke-mia HL-60 cells. J Biol Chem 259:3021–3025
28. Tobler A, et al (1988) Regulation of gene expression ofmyeloperoxidase during myeloid differentiation. J Cell Physiol136:215–225
29. Sagoh T, Yamada M (1988) Transcriptional regulation of MPOgene expression in myeloid leukemia HL60 cells during differ-entiation into granulocytes and macrophages. Arch BiochemBiophys 262:599–604
30. Meier RW, Chen T, Friis RR, Tobler A (1991) Myeloperoxidaseis a primary response gene in HL60 cells, directly regulated dur-ing hematopoietic differentiation. Biochem Biophys ResCommun 176:1345–1350
31. Zaki SR, Austin GE, Swan DC, et al (1990) Studies onmyeloperoxidase gene expression at the cellular level by in situhybridization. Leukemia 4:813–818
32. Chang KS, Zhao S, Wang Y, et al (1991) Down regulation ofmyeloperoxidase gene associated with specific nuclease hyper-sensitive sites during TPA induced differentiation of HL60. Leu-kemia 5:205–209
33. Shrivastava CH, Rado TA, Bauerle D, Broxmeyer HE (1991)Regulation of human bone marrow lactoferrin and myeloper-oxidase gene expression by tumor necrosis factor-α. J Immunol146:1014–1019
34. Ford AM, Bennett CA, Healy LE, Towatori M, Greaves MF, En-ver T (1996) Regulation of the myeloperoxidase enhancer bind-ing proteins Pu1, C-EBP alpha, -beta and –delta during granulo-cyte lineage specification. Proc Natl Acad Sci USA 93:10838–10843
35. Britos-Bray M, Friedman AD (1997) Core binding factor cannotsynergistally activate the myeloperoxidase proximal enhancer in

68. Wallace RW (1997) DNA on a chip: serving up the genome fordiagnostics and research. Mol Med Today 3:384–389
69. Nauseef WM, Cogley S, Bock S, Petrides PE (1998) Pattern ofinheritance in hereditary MPO deficiency associated with theR569W missense mutation, J Leukoc Biol 63:264–269
70. Petrides PE (1998) Gendiagnostik und Gentherapie. In: LöfflerG, Petrides PE: Biochemie und Pathobiochemie, 6th edn.Springer, Berlin Heidelberg New York, pp 317–355
71. Presentey B (1988) A new anomaly of eosinophilic granulocytes.Am J Clin Pathol 49:887–890
72. Presentey B (1970) Partial and severe peroxidase and phospho-lipid deficiency in eosinophils. Acta Haematol 44:345–354
73. Presentey B (1984) Ultrastructure of human eosinophils geneti-cally lacking peroxidase. Acta Haematol 71:334–340
74. Lanza F, Castoldi GL, Masotti M (1988) Eosinophil peroxidasedeficiency detected by the Technicon H1 system. Blut 56:143–144
75. Zabucchi G, Soranzo MR, Menegazzi R, et al (1992) Eosinophilperoxidase deficiency: morphological and immunocytochemicalstudies of the eosinophil specific granules. Blood 80:2903–2910
76. Kutter D, Mueller-Hagedorn S, Forges T, Glaesener R (1995) Acase of eosinophil peroxidase deficiency. Ann Hematol 71:315–317
77. Romano M, et al (1994) Hereditary eosinophil peroxidase defi-ciency: immunochemical and spectroscopic studies and evidencefor a compound heterozygosity of the defect. Proc Natl Acad SciUSA 91:12496–12500
698
neous patterns of the complete deficiency state as manifested atthe genomic, mRNA and protein levels. Blood 83:1317–1322
60. Sheppard DN, Ostedgaard LS (1996) Understanding how cysticfibrosis mutations cause a loss of Cl channel function. Mol MedToday 3:290–297
61. Scriver CR, et al (1996) The phenylalanine hydroxylase locus: amarker for the history of phenylketonuria and human genetic di-versity. In: Variation of the human genome. Ciba FoundationSymposium 1996. Wiley, New York, pp 73–96
62. leCoutre P, Demina A, Beutler E, Beck M, Petrides PE (1997)Molecular analysis of Gaucher disease: distribution of eight mu-tations and the complete gene deletion in 27 patients from Ger-many. Hum Genet 99:816–821
63. Kahn P (1996) Coming to grips with genes and risk. Science274:496–498
64. Petrides PE (1997) Die akute intermittierende Porphyrie: Diag-nose und Therapie. Dtsch Ärzteblatt 94:A3407–3412
65. Petrides PE, Demina A, Müller-Höcker J, Magin E, Harzer K,Demina A, Beutler E (1998) Coincidence of Gaucher diseasedue to a private mutation and Ph’ positive CML, Am J Hematol59:87–90
66. Cotton RGH (1997) Slowly but surely towards better scanningfor mutations. Trends Genet 13:43–46
67. Petrides PE, LeCoutre P (1997) Morbus Gaucher: Klinik undStand der molekulargenetischen Diagnostik. Med Genet 9:495–500
Related Documents











