[CANCER RESEARCH 52, 267-274, January 15, 1992| Molecular Dosimetry in Rat Urine of Aflatoxin-W 7-guanine and Other Aflatoxin Metabolites by Multiple Monoclonal Antibody Affinity Chromatography and Immunoaffinity/High Performance Liquid Chromatography1 John D. Groopman,2 Julia A. Hasler, Laura J. Trudel, Anthony Pikul, Paul R. Donahue, and Gerald N. Wogan Department of Environmental Health Sciences, School of Hygiene and Public Health, The Johns Hopkins University, Baltimore, Maryland 21205 ¡J.D. G., A. P.]; Department of Biochemistry, University of Zimbabwe, Box MP 167, Harare, Zimbabwe [J. A. H.]; and Division of Toxicology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139 [L. J. T., P. R. D., C. N. W.¡ ABSTRACT The development of molecular dosimetry methods will simplify the identification of people at high risk for cancer. A combined monoclonal antibody ¡mmunoaffinitychromatography/high performance liquid chro- matography method has been devised to isolate and quantify aflatoxin- DNA adducts and other metabolites in rat urine samples. We report the production of 11 different monoclonal antibodies recognizing aflatoxin B,, aflatoxin <.),. aflatoxin <;,, aflatoxicol, and aflatoxin MI and the application of these antibodies to a multiple monoclonal antibody affinity Chromatography technique. Using the multiple monoclonal antibody af finity column with rat urines obtained from dosed animals, between 90 and 95% of total aflatoxin metabolites can be bound to the column and isolated. Analytical immunoaffinity chromatography/high performance liquid Chromatography analysis of these isolated aflatoxins reveals that more than 55% of the aflatoxins in rat urine are aflatoxin-dihydrodiol, aflatoxin-A/'-guanine, aflatoxin Qi, aflatoxin M,, aflatoxin IV and afla toxin li,, accounting for 1.5, 9.6, 1.8, 34.5, 8.0, and 1.0% of the total aflatoxins, respectively. Further, a perchloric acid digestion of the afla- toxin-yV-guanine peak was used to confirm its identity by its conversion to guanine. The measurement of aflatoxin-A/'-guanine excretion in rat urine was examined to assess its utility as a marker of DNA adduct formation in the liver, and a dose-dependent excretion in urine was found with a correlation coefficient of 0.99. A comparison of the dose-dependent residual levels of aflatoxin binding to liver DNA with the amount of aflatoxin-jV-guanine excreted in urine showed a correlation coefficient of 0.98. Besides the nucleic acid adduct excretion data, aflatoxin MI and aflatoxin I', were evaluated as molecular dosimeters in the urine. Afla toxin MI was found to be an excellent marker, whereas no linear relationship between dose and aflatoxin I', excretion in urine was found. INTRODUCTION AFBf1 is one of the most potent liver carcinogens and toxins known for experimental animals and this mycotoxin is a fre quent contaminant of the food supply as a result of mold spoilage (1). Consumption of contaminated foods by people living in some regions of Asia and Africa results in high intake levels of AFB,. Several epidemiological studies have found strong positive associations between AFB, exposure and in creased human liver cancer risk (2-4). Evidence from such studies justifies extensive research efforts undertaken by many organizations to minimize aflatoxin exposures. Received 7/23/91; accepted 10/31/91. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1This research was supported in part by USPHS Grants POI ES00597 and ROÕCA54114. J. D. G. is a recipient of Research Career Development Award KO4 CA01517. J. A. H. is a recipient of a Senior African Fulbright Fellowship from the Council for the International Exchange of Scholars. 2 To whom requests for reprints should be addressed. 3The abbreviations used are: AFB!, aflatoxin B,; AFM,, aflatoxin M,; AFG,, aflatoxin G,; AFQ,, aflatoxin Q,; AFP,, aflatoxin P,; AFB-N7-Gua, aflatoxin- /V-guanine; AFB-diol, 8,9-dihydro-8,9-dihydroxyaflatoxin; IAC/HPLC, immu noaffinity chromatography/high performance liquid Chromatography; BGG, bo vine 7-globulin; BSA, bovine serum albumin; PBS, phosphate-buffered saline, pH 7.4; DMSO, dimethyl sulfoxide; ELISA, enzyme-linked immunosorbent assay; RIA, radioimmunoassay. The toxic and carcinogenic effects of AFBi are manifested only after metabolism by members of the endogenous cyto- chrome P-450 enzyme superfamily. These enzymes produce various oxidative derivatives including: AFM,, AFQi, AFPt; a reduced aflatoxin species, aflatoxicol; and a highly reactive 8,9- epoxide metabolite, the ultimate carcinogenic compound. The reactive electrophilic epoxide can covalently interact with nu- cleophilic centers in many cellular macromolecules, including DNA and serum albumin. The two major macromolecular adducts identified are the aflatoxin-A^-guanine (AFB-N7-Gua) adduct in DNA (5) and a lysine adduct in serum albumin (6), respectively. Detoxification of the 8,9-epoxide metabolite oc curs through enzymatic reaction with glutathione mediated by glutathione 5-transferases (7). Thus, a major factor for assess ing biological hazards resulting from exposure to AFB, is the integrated balance of the genotoxic and nongenotoxic pathways of the 8,9-epoxide. Many analytical methods have been developed for the quan- titation of aflatoxin and its metabolites in foods and biological samples, including Chromatographie and immunological tech niques. We used the selectivity of monoclonal antibodies to develop an IAC/HPLC procedure to isolate and quantify afla toxin metabolites in biological samples. We previously reported that purification of aflatoxin metabolites and DNA adducts from rat urine samples was achieved using a high affinity IgM monoclonal antibody (8). A total of 65% of l4C-aflatoxins contained in the urine of a rat injected with 14C-AFB| became bound to the immunoaffinity column. HPLC of the retained metabolites showed that AFM, accounted for almost 50% of the recovered UC-AFB| and that the AFB-N7-Gua adduct was also a major metabolite in the urine. These data show the capability of IAC/HPLC to purify aflatoxin metabolites from biological samples. However, several important method prob lems remained, including optimization of the amount of a single monoclonal antibody bound to the resin, as assessed by the recovery of aflatoxin metabolites from in vivo urine samples, and the use of multiple monoclonal antibodies bound to resins as a technique to improve the total recovery of aflatoxin metab olites from urines. These two objectives have been addressed in experiments reported here. We describe the production and characterization of several monoclonal antibodies raised against various aflatoxins and metabolites and that the application of selected antibodies in IAC/HPLC results in greater than 90% recovery of aflatoxin metabolites in a rat urine sample. Our data support the utility of AFB-N7-Gua and AFM, excretion in urine as molecular dosimetry markers for assessing exposure to AFB,. MATERIALS AND METHODS Chemicals. AFB, was obtained from Aldrich Chemical Co. (Milwau kee, WI). AFB2, AFGi, and AFG2 were obtained from Sigma Chemical Co. (St. Louis, MO). The aflatoxin-DNA adduct standards were syn- 267 on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

[CANCER RESEARCH 52, 267-274, January 15, 1992|
Molecular Dosimetry in Rat Urine of Aflatoxin-W 7-guanine and Other Aflatoxin
Metabolites by Multiple Monoclonal Antibody Affinity Chromatography andImmunoaffinity/High Performance Liquid Chromatography1
John D. Groopman,2 Julia A. Hasler, Laura J. Trudel, Anthony Pikul, Paul R. Donahue, and Gerald N. Wogan
Department of Environmental Health Sciences, School of Hygiene and Public Health, The Johns Hopkins University, Baltimore, Maryland 21205 ¡J.D. G., A. P.];Department of Biochemistry, University of Zimbabwe, Box MP 167, Harare, Zimbabwe [J. A. H.]; and Division of Toxicology, Massachusetts Institute of Technology,Cambridge, Massachusetts 02139 [L. J. T., P. R. D., C. N. W.¡
ABSTRACT
The development of molecular dosimetry methods will simplify theidentification of people at high risk for cancer. A combined monoclonalantibody ¡mmunoaffinitychromatography/high performance liquid chro-matography method has been devised to isolate and quantify aflatoxin-DNA adducts and other metabolites in rat urine samples. We report theproduction of 11 different monoclonal antibodies recognizing aflatoxinB,, aflatoxin <.),. aflatoxin <;,, aflatoxicol, and aflatoxin MI and theapplication of these antibodies to a multiple monoclonal antibody affinityChromatography technique. Using the multiple monoclonal antibody affinity column with rat urines obtained from dosed animals, between 90and 95% of total aflatoxin metabolites can be bound to the column andisolated. Analytical immunoaffinity chromatography/high performanceliquid Chromatography analysis of these isolated aflatoxins reveals thatmore than 55% of the aflatoxins in rat urine are aflatoxin-dihydrodiol,aflatoxin-A/'-guanine, aflatoxin Qi, aflatoxin M,, aflatoxin IV and afla
toxin li,, accounting for 1.5, 9.6, 1.8, 34.5, 8.0, and 1.0% of the totalaflatoxins, respectively. Further, a perchloric acid digestion of the afla-toxin-yV-guanine peak was used to confirm its identity by its conversionto guanine. The measurement of aflatoxin-A/'-guanine excretion in rat
urine was examined to assess its utility as a marker of DNA adductformation in the liver, and a dose-dependent excretion in urine was foundwith a correlation coefficient of 0.99. A comparison of the dose-dependentresidual levels of aflatoxin binding to liver DNA with the amount ofaflatoxin-jV-guanine excreted in urine showed a correlation coefficientof 0.98. Besides the nucleic acid adduct excretion data, aflatoxin MI andaflatoxin I', were evaluated as molecular dosimeters in the urine. Afla
toxin MI was found to be an excellent marker, whereas no linearrelationship between dose and aflatoxin I', excretion in urine was found.
INTRODUCTION
AFBf1 is one of the most potent liver carcinogens and toxins
known for experimental animals and this mycotoxin is a frequent contaminant of the food supply as a result of moldspoilage (1). Consumption of contaminated foods by peopleliving in some regions of Asia and Africa results in high intakelevels of AFB,. Several epidemiological studies have foundstrong positive associations between AFB, exposure and increased human liver cancer risk (2-4). Evidence from suchstudies justifies extensive research efforts undertaken by manyorganizations to minimize aflatoxin exposures.
Received 7/23/91; accepted 10/31/91.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This research was supported in part by USPHS Grants POI ES00597 andROÕCA54114. J. D. G. is a recipient of Research Career Development AwardKO4 CA01517. J. A. H. is a recipient of a Senior African Fulbright Fellowshipfrom the Council for the International Exchange of Scholars.
2To whom requests for reprints should be addressed.3The abbreviations used are: AFB!, aflatoxin B,; AFM,, aflatoxin M,; AFG,,
aflatoxin G,; AFQ,, aflatoxin Q,; AFP,, aflatoxin P,; AFB-N7-Gua, aflatoxin-/V-guanine; AFB-diol, 8,9-dihydro-8,9-dihydroxyaflatoxin; IAC/HPLC, immunoaffinity chromatography/high performance liquid Chromatography; BGG, bovine 7-globulin; BSA, bovine serum albumin; PBS, phosphate-buffered saline, pH7.4; DMSO, dimethyl sulfoxide; ELISA, enzyme-linked immunosorbent assay;RIA, radioimmunoassay.
The toxic and carcinogenic effects of AFBi are manifestedonly after metabolism by members of the endogenous cyto-chrome P-450 enzyme superfamily. These enzymes producevarious oxidative derivatives including: AFM,, AFQi, AFPt; areduced aflatoxin species, aflatoxicol; and a highly reactive 8,9-
epoxide metabolite, the ultimate carcinogenic compound. Thereactive electrophilic epoxide can covalently interact with nu-cleophilic centers in many cellular macromolecules, includingDNA and serum albumin. The two major macromolecularadducts identified are the aflatoxin-A^-guanine (AFB-N7-Gua)
adduct in DNA (5) and a lysine adduct in serum albumin (6),respectively. Detoxification of the 8,9-epoxide metabolite occurs through enzymatic reaction with glutathione mediated byglutathione 5-transferases (7). Thus, a major factor for assessing biological hazards resulting from exposure to AFB, is theintegrated balance of the genotoxic and nongenotoxic pathwaysof the 8,9-epoxide.
Many analytical methods have been developed for the quan-titation of aflatoxin and its metabolites in foods and biologicalsamples, including Chromatographie and immunological techniques. We used the selectivity of monoclonal antibodies todevelop an IAC/HPLC procedure to isolate and quantify aflatoxin metabolites in biological samples. We previously reportedthat purification of aflatoxin metabolites and DNA adductsfrom rat urine samples was achieved using a high affinity IgMmonoclonal antibody (8). A total of 65% of l4C-aflatoxinscontained in the urine of a rat injected with 14C-AFB| became
bound to the immunoaffinity column. HPLC of the retainedmetabolites showed that AFM, accounted for almost 50% ofthe recovered UC-AFB| and that the AFB-N7-Gua adduct was
also a major metabolite in the urine. These data show thecapability of IAC/HPLC to purify aflatoxin metabolites frombiological samples. However, several important method problems remained, including optimization of the amount of a singlemonoclonal antibody bound to the resin, as assessed by therecovery of aflatoxin metabolites from in vivo urine samples,and the use of multiple monoclonal antibodies bound to resinsas a technique to improve the total recovery of aflatoxin metabolites from urines. These two objectives have been addressed inexperiments reported here. We describe the production andcharacterization of several monoclonal antibodies raised againstvarious aflatoxins and metabolites and that the application ofselected antibodies in IAC/HPLC results in greater than 90%recovery of aflatoxin metabolites in a rat urine sample. Ourdata support the utility of AFB-N7-Gua and AFM, excretion
in urine as molecular dosimetry markers for assessing exposureto AFB,.
MATERIALS AND METHODS
Chemicals. AFB, was obtained from Aldrich Chemical Co. (Milwaukee, WI). AFB2, AFGi, and AFG2 were obtained from Sigma ChemicalCo. (St. Louis, MO). The aflatoxin-DNA adduct standards were syn-
267
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-/V-GUANINE IN RAT URINE
thesized as described previously (9). AFM, and AFQ, were gifts fromDr. George Buchi, MIT. The aflatoxin-lysine adduci was a gift of Dr.Paul L. Skipper, MIT.
Preparation of Aflatoxin Antigens: VIH,, Aid,. AIM,, Al (,),. andAflatoxicol-modified Proteins. Aflatoxin antigens were synthesized toproduce monoclonal antibodies specific for many different aflatoxinmetabolites. All the synthesized immunogens used either BGG orthyroglobulin (both purchased from Calbiochem, La Jolla, CA) ascarrier proteins. BSA (Sigma) was used as the carrier protein to makethe screening antigens used in immunoassays. Two haptenization procedures were used, both involving the chemical activation of the unsat-urated double bond (8,9-position in AFBi) in the furo-furan rings ofthe aflatoxin molecule. One scheme used m-chloroperoxybenzoic acidand the other method produced an activated 8,9-dibromoaflatoxinderivative. All proteins for these syntheses were dissolved in PBS, pH7.4, at a concentration of 10 mg/ml.
The chemical method using m-chloroperoxybenzoic acid was modified from that previously described by Martin and Garner (10). In atypical reaction, 1 mg of AFBi (3.2 ¿miol)was dissolved in 2 ml ofméthylènechloride and added to a 5 Mexcess of m-chloroperoxybenzoicacid also dissolved in 2 ml of méthylènechloride. The protein wasadded to the reaction mixture to produce a 5 M excess of aflatoxinrelative to the lysine content of the protein. The mixture was vigorouslystirred overnight at 20°C.The reaction was terminated by centrifuging
at 2000 x g for 20 min and the aqueous epiphase containing theaflatoxin-modified protein was dialyzed against PBS, pH 7.4, at 4°C.
The level of modification of the protein was estimated by absorbanceat 362 inn. using a molar extinction coefficient of 21,800. In typicalreactions, 40-50 aflatoxin residues/molecule of BGG and 20-30 molecules of aflatoxin per molecule of BSA were found.
The second activation method of aflatoxins containing the unsatu-rated 8,9-bond in the furo-furan ring was performed with molecularbromine (Br2) as modified from the method of Gorst-Allman et al. (11).This was a rapid method to produce a direct acting reagent that inducedcovalent linkage of the aflatoxin moiety to proteins. Briefly, synthesisof the dihaloaflatoxin involved dissolving the aflatoxins in méthylènechloride that had been dried and stored over 4-Àmolecular sieves(Fisher Scientific Co., Pittsburgh, PA). The aflatoxin concentration inthe méthylènechloride was determined by UV spectroscopy using amolar extinction coefficient of 21,800 at 362 nm. Br2 (Aldrich), d =3.119 g/ml, was used for the derivatization. Ten ^1 (394.8 ^mol)bromine were dissolved in 990 n\ méthylènechloride. An aliquot ofthis dilution equal to the molar amount of aflatoxin was mixed into theaflatoxin solution. The bromine and aflatoxin were allowed to react for20 min at 20°Cunder argon. This mixture was brought to dryness
under reduced pressure and the aflatoxin derivative was dissolved inDMSO (Aldrich) and used directly for the coupling reaction. Themodified proteins were dialyzed overnight against PBS and the aflatoxin content was measured by UV spectroscopy as described above.Levels of protein modification attained by this method were similar tothose described for the m-chloroperoxybenzoic acid reactions.
Mouse Immunization Schedule. Female BALB/cJ mice (The JacksonLaboratory), 16 weeks old, were immunized with aflatoxin-modifiedbovine 7-globulin or thyroglobulin dissolved in PBS and emulsifiedwith an equal volume of complete Freund's adjuvant. For each antigen,
five mice were immunized i.p. with 100 p%of aflatoxin-modified proteinin 0.2 ml of emulsion. At 5 and 9 weeks following the initial injection,all animals received i.p. the same amount of antigen emulsified withincomplete Freund's adjuvant. About 10 days after the second booster
injection, serum samples were collected by tail bleeding and assayed byELISA for antibody activity, as described below. Mice were judged tobe producing a sufficient immune response to justify fusion when theserum titer exceeded 1:1500. Three days before fusion each mouse wasgiven a final immunization with the same aflatoxin-modified proteindissolved in 0.1 ml PBS, injected into the tail vein.
Monoclonal Antibody Production: Cell Fusion Procedures. The myeloma cell line used for fusion, SI' 2. was maintained in suspension inDulbecco's modified Eagle's medium supplemented with 20% (v/v)
fetal calf serum, glutamine, penicillin, streptomycin, and nonessential
amino acids (GIBCO). The cell fusion procedures have been describedpreviously by Groopman et al. (8).
Hybridomas secreting antibodies of interest were grown as ascitestumor cells in BALB/c mice previously given injections of 0.5 mlpristane (Aldrich). Ascites fluids collected from these animals werepooled and further purified by precipitation by addition of ice-coldammonium sulfate to a final concentration of 40% v/v. The precipitatewas centrifuged at 2000 x g for 15 min at 4°C;the pellet containing
the antibodies was redissolved in water and then dialyzed against PBSat 4"C overnight.
ELISA. ELISAs were used to determine aflatoxin-specific antibodiesin mouse sera and hybridoma media. Procedures used were modifications of previously described protocols (8). Briefly, polyvinyl chloridemicrotiter plates (Dynatech Laboratories, Alexandria, VA) containingin each well 50 n\ aflatoxin-BSA dissolved in PBS at 2 /¿g/mlwereincubated for 2 to 4 h at ambient temperature. Control plates containing50 fi\ of BSA in PBS at 2 /¿g/mlwere treated similarly. After incubationthe antigen solution was aspirated and the plates were washed threetimes with PBS-0.05% Tween 20. Solutions containing 0.2% BSA or0.2% gelatin, type IV (Sigma), in PBS was added, and the plates wereincubated for l h at ambient temperature. This procedure limits nonspecific binding of antibodies to the plates. The plates were washedwith PBS-0.05% Tween 20, and 50 ß\of mouse sera or hybridomamedia were added to each well. To titer the mouse sera, dilutions in10% horse serum were prepared over a range from 1:50 to 3:50,000 at3-fold dilutions. In the case of hybridoma media, SU-M'aliquots wereused without dilution. Plates were incubated for 90 min at 37°C,after
which they were thoroughly washed. Specific antibodies bound to theplate were detected by incubating 50 ^I/well of a 1:200 dilution of ratanti-mouse K antibody coupled with alkaline phosphatase for 2 h atambient temperature. The plates were rewashed and incubated for upto 2 h with 100 ¿il/wellof a 1-mg/ml p-nitrophenyl phosphate (Sigma)solution in 0.1 M diethanolamine buffer, pH 9.8. Quantitation of thep-nitrophenol product was performed by measuring absorbance at 405nm using a Dynatech Minireader II microtiter plate reader.
The isotypes of the monoclonal antibodies were determined in non-competitive ELISA by using the mouse immunoglobin subtype identification kit purchased from Boehringer-Mannheim (Indianapolis, IN).
Competitive RIA. Competitive RIA was used to characterize theaffinity constants of the monoclonal antibodies. An assay with a 300-ii\ total volume was used. The tracer varied with the antibody beingtested and consisted of 3H-AFB, (specific activity, 3.4 Ci/mmol) or 3H-AFQ.2, 3H-AFM2, and 3H-AFG2 (specific activity, 24 Ci/mmol) (Mo-
ravek Biochemicals, Brea, CA). The tracer was diluted in 1% normalmouse serum-0.1% BSA-PBS and 100 n\ were used in the assay. In thecompetitive RIA the monoclonal antibody added to the reaction mixture in 100 M' 10% horse serum-PBS was diluted to a concentrationthat precipitated 30-50% of the 3H-aflatoxin tracer. The test sample,consisting of nonradiolabeled aflatoxin in a series of 3-fold dilutionswere added to the tube in a 100-/jl volume. The reaction mixture wasincubated at ambient temperature for 2 h; then an equal volume of ice-cold saturated ammonium sulfate (300 /il) was added, and the samplemixed and allowed to stand at room temperature for 15 min. Thesample was centrifuged for 15 min at 2000 x g. An aliquot of thesupernatant was measured by liquid scintillation counting and thepercentage of inhibition in the reaction and affinity constants wascalculated as previously described (8).
Preparative Monoclonal Antibody Affinity Column Resin. Monoclonalantibody affinity column resins were prepared by the following procedure. The monoclonal antibody was dissolved in varying concentrationsin coupling buffer (0.1 M NaHCO,, pH 8.3, 0.5 M NaCl). This solutionwas reacted with cyanogen bromide-activated Sepharose 4B (Sigma)that had been rehydrated in 0.001 N HC1 and washed with couplingbuffer. After reaction, unbound sites of the antibody-bound gel wereblocked by incubating in 1.0 M ethanolamine, pH 8.5. All affinitycolumn procedures were performed at ambient temperature and theresin was stored with buffer adjusted to 0.02% merthiolate to preventbacterial contamination.
Rat Experimental Protocols. Male F-344 rats were purchased at 6weeks of age from Charles River Breeding Laboratories (Kingston, NY)
268
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-/V'-GUANINE IN RAT URINE
and housed singly in wire-bottomed cages in a temperature and humidity controlled environment with a 12-h light, 12-h dark photoperiod.Food and water were available ad libitum. The animals were fed anAIN-76 diet.
In one of the dosing experiments, rats received i.p. l mg 'H-AFB,/
kg body weight (155.78 ng AFB,; specific activity, 159.35 mCi/mmol)dissolved in DMSO. Rats were placed in glass metabolic cages and theurines were collected for 48 h in flasks immersed in dry ice-acetone.
In another animal experiment rat urines were collected from 12 maleF-344 rats dosed p.o. with double radiolabled 14C,'H-AFB,. The ratioof 14C to 3H ranged from 6.5 to 9.0 to facilitate tritium exchange
calculations. Three groups of four rats each were given 4.25, 10.5, and141.1 ng AFB, resulting in about a 0.03, 0.07, and 1.0 mg AFB,/kgbody weight dosage, respectively. The rats were housed in metaboliccages for 24 h postdosing and the urines were collected in flasksimmersed in a dry ice-acetone bath.
At the end of each experiment, residual aflatoxin liver DNA adducilevels in all rats were determined as described previously by Groopmanet al. (9, 12).
Monoclonal Antibody Affinity Chromatography Procedure for RatUrines. Rat urines collected from animals dosed with aflatoxins werekept frozen at -20°Cuntil use. In general, 1 ml of rat urine, containing
no more than 700 ng total aflatoxins, was used for an analysis. First, 1ml of l M ammonium formate, pH 5.2, was added to the urine samplewith thorough mixing. If the pH of the urine was greater than about5.0, 0.1 N HC1 was added in 20-^1 aliquots until the pH was about 5.0.The urine was heated at 50°Cfor 10 min to dissolve any precipitated
aflatoxin metabolites. After cooling, the volume was brought to 5 mlwith water and the sample was applied to the monoclonal antibodyaffinity column, as follows.
The monoclonal antibody column was first washed with three columnvolumes of PBS and the PBS was allowed to flow until the meniscusreached the top of the resin. The rat urine sample was layered on topof the column and allowed to flow onto the column using gravitypressure. The column was washed with three successive column volumesof PBS, and the effluents were collected. The aflatoxins were elutedfrom the column using two column volumes of 70% DMSO:30% water,after which the column was washed with water followed by PBS. Intotal, eight fractions were collected. The DMSO and two succeedingwashes containing the bound aflatoxins were pooled, and the DMSOwas reduced to a final concentration of less than 3% by dilution withwater. This sample was applied to a C,8-Sep-Pak cartridge that hadbeen successively washed with 5 ml each of 5, 80, and 100% methanoland then 5% methanohwater. The aflatoxins were eluted from the Cig-Sep-Pak cartridge using 80% methanohwater and concentrated to dry-ness using a rotary evaporator under reduced pressure. The sample wasnow ready for analytical HPLC analysis.
Analytical HPLC of Rat Urines. Material eluted from the affinitycolumn was redissolved in 100 ^1 of 0.1 N HCI with heating at 50°C
for 10 min, and the pH was adjusted to about pH 3.0 by adding anequal volume of l M ammonium formate, pH 4.5. The sample wasinjected onto the HPLC and the column effluents were monitored inseries with a Beckman Model 160 fixed wavelength detector set at 365nm and a Hewlett-Packard Model 1040A diode-array detector. TheHPLC column was a C18 5-^m 25-cm Ultrasphere analytical column(Rainin Inst. Co., Woburn, MA) and chromatography was obtained byelution at a flow rate of 1 ml/min, for 20 min at 13% ethanol followedby a 13-25% ethanol linear gradient generated over 25 min and thenisocratic elution at 25% ethanol. All mobile phases were buffered with0.01 M (ricth\[ammonium formate, pH 3.0, and the column temperature was maintained at 35°C.Authentic aflatoxin standards were used
to assign retention times of peaks and generate standard curves forindividual aflatoxin quantitation.
Conversion of AFB-yV-guanine to Guanine by Perchloric Acid Hydrolysis. Fractions from the HPLC were collected for confirmation ofthe AFB-N7-gua adduci by the conversion of the aflatoxin-DNA adduci
to guanine. A method modified from that described by Essigmann etal. (5) was used. Briefly, the HPLC fraction (about 2 ml) containingthe adduct was dried by rotary evaporation under reduced pressure, andthe adduct was redissolved in 0.1 ml 72% perchloric acid and heated at
100°Cfor 60 min. The solution was neutralized using 3 M KOH and
adjusted to pH 4.0 using 88% formic acid. After centrifugation andfiltration through a 0.45-iim nylon filter, the sample was analyzed byHPLC using reverse phase chromatography on a Ci«5-¿imUltraspherecolumn with isocratic elution at 2% ethanol at 1 ml/min. The guaninepeak was identified and quantified using the selectivity of the HP1040A Diode Array detector for the specific guanine spectrum foundunder acidic conditions.
RESULTS
In the original report of immunoaffinity chromatographyusing monoclonal antibody 2B11,65% of the radioactivity fromthe urine of an AFB,-dosed rat bound to the antibody resin (8).In these experiments, the amount of monoclonal antibodycoupled per ml Sepharose 4B resin was equivalent to 3.0 mgtotal protein from ammonium sulfate-precipitated ascites. Todetermine if greater recovery of aflatoxins from urine could beobtained by increasing the amount of antibody coupled toSepharose 4B, the following experiment was done. Monoclonalantibody 2B11 was purified from mouse ascites fluid usingammonium sulfate precipitation and then linked to cyanogenbromide-activated Sepharose 4B at levels of 5 and 10 mg totalprotein per ml resin. Rat urines obtained from animals dosedat 1 mg 'H-AFB]/kg body weight and collected over 48 h wereapplied to the affinity columns as described in "Materials andMethods." Both the 5- and 10-mg antibody/ml resin columns
modestly increased efficiency in binding radiolabeled aflatoxinmetabolites, retaining 72.6 ±2.8% (SE; four urine samples percolumn; eight urine samples total) total radioactivity from raturine. HPLC analysis (data not shown) of these samples revealed a similar pattern of metabolites similar to that seen inthe original chromatograms. Thus, despite increasing the totalamount of monoclonal antibody coupled to the resin by >3-fold, we were unable to bind more than 75% of the aflatoxinmetabolites from rat urines.
Production and Characterization of AFBi-, AFGi-, AFMi-,AFQi-, and Aflatoxicol-specific Monoclonal Antibodies. To attempt to overcome the affinity column binding problem, a seriesof new monoclonal antibodies were developed with differentspecificities. These antibodies were used for the multiple monoclonal antibody affinity column resins. Monoclonal antibodieswere produced using protein antigens haptenized with AFB,,AFQ,, AFG,, afiatoxicol, and AFM,. In total, 11 differentmonoclonal antibodies were produced as ascites and characterized. The affinity constants of monoclonal antibodies for specific aflatoxin competitors are found in Table 1. Based on thesedata, the major epitope recognized by all these aflatoxin-specificmonoclonal antibodies was shown to be the coumarin ring and"E" ring (e.g., cyclopentenone ring in AFB,) of the aflatoxin
molecule.An additional result found in Table 1 pertains to monoclonal
antibody 2B11, originally described by Groopman et al. (8),which is reported here to have an affinity constant of greaterthan 8 x IO9 liters/mol for the major aflatoxin-lysine adduct
found in serum albumin (6). All the affinity constants describedin Table 1 were calculated using tritium-based RIA data. Because of the low specific activity of tritium tracers, affinityconstants of greater than 1 x 10"' liters/mol cannot be deter
mined because the tritium tracer becomes a significant molarcontributor to the antibody competition reaction.
Finally, the isotypes of the monoclonal antibodies were determined and three antibodies (2B11, 2E4, 2D5) were found tobe IgM, six antibodies (1BB3, 2F5, 2D8,4B7,6F9, and 3BD10)
269
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-A^-GUANINE IN RAT URINE
Table 1 Affinity constants (liters/'mol x Iff) of monoclonal antibodies for
aflatoxins and metabolites
AntigenCompetitorAFB,
AFB,AFB2AFC,AFG2AFM,AFQ,AFP,AFB-N'-GuaAFB-FAPyrAFB-Lys adduci
AflatoxicolAFM,
AFM,AFB2AFG,
AFP,AFQ,
AFQ,AFB,AFM,AFG,
AFG2AFG,
AFG,AFG2AFB,AFB2AFM,2B11100
1005.03.5
1001.1
603.4
13800NR2D535
10NDND1C63.0
<1.00.20
NRNR2D825
133.03.00.751BB375
75ND°
ND3.71.0
ND3.3
10ND
ND4B71347
3.32.7ND2E42.9
4.41.41.14.40.54
NDNDNDNDND2F58.6
2.73.41.26E51.5
<1.00.10
NRNR6F914
192.12.1
ND3BD10Aflatoxicol
AflatoxicolAFB,AFG,AFM,0.35
0.400.46
0.53'ND, not determined; NR, not recognized.
IgGl, and two antibodies (6E5 and 1C6) IgG2b.Multiple Monoclonal Antibody Affinity Column. A multiple
monoclonal antibody affinity resin was produced to attempt torecover a larger percentage of aflatoxin metabolites from raturine. Our hypothesis was that the high levels of AFM, in therat urines compromised the ability to bind all aflatoxin metabolites using a single monoclonal antibody. To address thisproblem, a 7-tnl monoclonal antibody affinity column was madeconsisting of 3 ml 2F5 (anti-AFMi) bound at 5 mg protein/mlresin, 1 ml 6E5 (anti-AFQi) bound at 2.5 mg protein/ml resin,and 3 ml 2B11 (anti-AFB,) bound at 10 mg protein/ml resin.
Urine from rats dosed i.p. with 1 mg 3H-AFBi/kg body weight
were collected for 48 h and divided into four equal samples;each sample was prepared using a Sep-Pak cartridge. Thesefour samples were then separately applied to the multiantibodycolumn and 94.0 ±1.0% of the radiolabeled aflatoxins becamebound to the antibody affinity matrix. Representative HPLCdata obtained using the multicolumn technique for rat urineare shown in Fig. 1. The major metabolite quantified in thissample was AFMi (retention time, 43.5 min) (Fig. \B). Sixdifferent aflatoxins were identified by cochromatography withauthentic standards (Fig. 1C) and collectively account for 56.4%of the total aflatoxin metabolites injected. Levels of AFB-diol,AFB-N7-Gua, AFQ,, AFM,, AFP,, and AFB, were determined
using individual standard curves and 1.5, 9.6, 1.8, 34.5, 8.0,and 1.0% of the total aflatoxins in the urine, respectively, wereaccounted for by these compounds. Peak areas were used inthese calculations rather than peak heights because the combined isocratic and gradient elution scheme used for this analysis broadens the earlier eluting aflatoxins. Fig. \A depicts theUV-visible on-line spectrum of AFB-N7-Gua, identical with the
spectrum originally reported by Essigmann et al. (5).The HPLC diode-array detector used in these studies ac
quired continuous spectral absorbance information from 190to 600 nm during the Chromatographie analysis. Since mostaflatoxins have characteristic spectra in the 365 nm absorbancerange, yet unidentified aflatoxin metabolites can be detectedand measured with this instrument. Fig. 2 shows the UV-visiblespectra from 210 to 450 nm for the chromatogram found inFig. 1C for retention times between 18 and 50 min. As seen inthis diagram, all the unidentified 363 nm-absorbing peaksshown in Fig. 1C contain the characteristic absorption spectraof an aflatoxin molecule. Future experiments will need to bedone to characterize these metabolites, but their presence andthe absence of interfering compounds in the Chromatographieprofile demonstrate the ability of the immunoaffmity columnto discriminate between aflatoxins and contaminants.
The success of the multiple monoclonal antibody preparativeaffinity column containing anti-AFB,-, anti-AFM,-, and anti-AFQi-specific antibodies led to the following experiments todefine the excretion of metabolites. Rat urines were obtainedfrom 12 male F-344 rats dosed p.o. at levels ranging from 0.030to 1.00 mg AFB,/kg body weight. The AFB, used for theseexperiments was double radiolabeled with I4C and 3H, permit-
SPECTRUM OF RFB-N7-GLJR
250 300 350 400 450Nave 1ength <nm)
12B-IBB-80-SB-4B
HFB1 METHBOLITES IN RHT URINE251 ^ RFMI20- RFB-N7-GUR
Fig. 1. Analytical HPLC prillile of a rat urine sample obtained from an animaldosed at 1 mg AFB,/kg body weight and the urine collected for 48 h postdosing.The urine was preparatively prepared using three different monoclonal antibodiesbound to an immunoaffinity column. A, UV-visible spectrum of AFB-N7-Gua
adduci; B, HPLC profile at 362 nm from 0 to 70 min, showing the contributionof AFM, to total aflatoxins; C, HPLC profile at 362 nm from 0 to 70 minshowing the AFB-N7-Gua, AFQ,, AFM,, and AFP, in greater detail. The Chromatographie conditions are described in "Materials and Methods." ni-tl . milli-
absorbance units.
Fig. 2. HPLC profile between 18 and 50 min obtained from the rat urineshown in Fig. 1C. Data show the 210 to 450 nm UV-visible spectrum for each ofthe peaks.
270
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
MOLECULAR DOSIMETRY OF AFB-/V-GUANINE IN RAT URINE
ting the assessment of tritium exchange in these samples. In allcases the tritium exchange in the urines was no greater than15%. All urines were collected over the for 24 h postdosing and8.1 ±1.8% of the administered dose had been excreted in theurine during this period. Aliquots of each urine were prepara-tively purified by immunoaffinity chromatography and 89.0 ±3.0% (12 samples) of the radiolabeled aflatoxin metaboliteswere bound to the multiple antibody affinity column. Theaflatoxins were then released from the immunoaffinity columnand individual aflatoxin metabolites were separated by analytical reverse phase HPLC (data not shown). Similar to the datadescribed above, the urine from an animal dosed p.o. with 1.0mg AFB,/kg contained AFB-N7-Gua, AFQ,, AFM,, AFP,, and
AFB, accounting for 7.5, 3.0, 31.5, 2.2, and 0.3% of the totalaflatoxins injected on the HPLC, respectively.
One of the rat urine samples from the 1.0-mg/kg dosinggroup was treated with a combination of glucuronidase/sulfat-ase and chromatographed under the same conditions. Therewas no discernible change in the patterns of the aflatoxinmetabolites (data not shown). Therefore, either the conjugatescleaved by these enzymes are minor metabolites that do notcontribute to changes in the total patterns or the putativeconjugates are not bound by the antibodies used in the affinitycolumns.
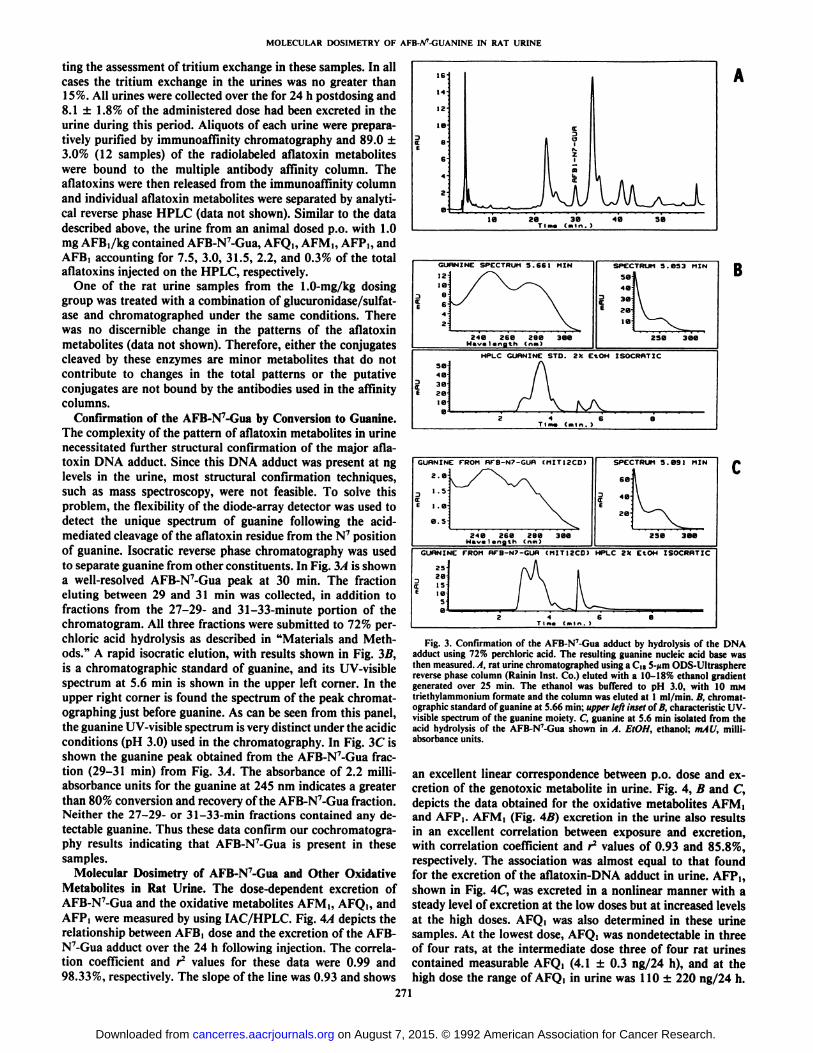
Confirmation of the AFB-N7-Gua by Conversion to Guanine.
The complexity of the pattern of aflatoxin metabolites in urinenecessitated further structural confirmation of the major aflatoxin DNA adduci. Since this DNA adduci was present at nglevels in the urine, most structural confirmation techniques,such as mass spectroscopy, were not feasible. To solve thisproblem, the flexibility of the diode-array detector was used todetect the unique spectrum of guanine following the acid-mediated cleavage of the aflatoxin residue from the N7 position
of guanine. Isocratic reverse phase chromatography was usedto separate guanine from other constituents. In Fig. 3Àis showna well-resolved AFB-N7-Gua peak at 30 min. The fraction
eluting between 29 and 31 min was collected, in addition tofractions from the 27-29- and 31-33-minute portion of thechromatogram. All three fractions were submitted to 72% perchloric acid hydrolysis as described in "Materials and Methods." A rapid isocratic elution, with results shown in Fig. 3B,
is a Chromatographie standard of guanine, and its UV-visiblespectrum at 5.6 min is shown in the upper left corner. In theupper right corner is found the spectrum of the peak chromat-ographing just before guanine. As can be seen from Ihis panel,the guanine UV-visible spectrum is very distinct under the acidicconditions (pH 3.0) used in the chromatography. In Fig. 3C isshown the guanine peak obtained from the AFB-N7-Gua fraction (29-31 min) from Fig. 3A. The absorbance of 2.2 milli-absorbance units for the guanine at 245 nm indicates a greaterthan 80% conversion and recovery of Ihe AFB-N7-Gua fraclion.Neither the 27-29- or 31-33-min fractions contained any detectable guanine. Thus these data confirm our cochromalogra-phy results indicating that AFB-N7-Gua is present in these
samples.Molecular Dosimetry of AFB-N7-Gua and Other Oxidative
Metabolites in Rat Urine. The dose-dependent excretion ofAFB-N7-Gua and the oxidative metaboliles AFM,, AFQ,, and
AFP, were measured by using IAC/HPLC. Fig. 4A depicts therelationship between AFB, dose and the excretion of the AFB-N7-Gua adduci over the 24 h following injection. The correlation coefficient and r2 values for these data were 0.99 and
98.33%, respectively. The slope of the line was 0.93 and shows
GURNINC SPECTRUM 5.661 MIN
12
240 260 280 300
SPECTRUM 5.053 MIN
58
48
Se-
zete-
250 308
HPLC GURNINE STO. 2K EtOH ISOCRHTIC
seiti3O
B
GURNINC FROM HFB-N7-GUR (MITI2CD)
2.0
3 '•=•
C£ i .a-
248 260 2BBNavelength Cnm)
250 388
GUHNINE FROM HFB-N7-GUH (MIT12CD) HPLC Z)t EtOH ISOCRRTJC
zaisles'a
Fig. 3. Confirmation of the AFB-N7-Gua adduci by hydrolysis of the DNA
adduci using 72% perchloric acid. The resulting guanine nucleic acid base wasthen measured. A, rat urine chromatographed using a d«5->imODS-Ultraspherereverse phase column (Rainin Inst. Co.) eluted with a 10-18% ethanol gradientgenerated over 25 min. The ethanol was buffered to pH 3.0, with 10 mMtriethylammonium formate and the column was eluted at l ml/min. B, Chromatographie standard of guanine at 5.66 min; upper left inset of B, characteristic UV-visible spectrum of the guanine moiety. C, guanine at 5.6 min isolated from theacid hydrolysis of the AFB-N7-Gua shown in A. EtOH, ethanol; mAU, milli-
absorbance units.
an excellenl linear correspondence belween p.o. dose and ex-crelion of Ihe genoloxic metabolite in urine. Fig. 4, B and C,depicls the dala oblained for Ihe oxidalive melaboliles AFM,and AFP,. AFM, (Fig. 4B) excretion in the urine also resultsin an excellent correlalion belween exposure and excrelion,with correlation coefficient and r2 values of 0.93 and 85.8%,
respectively. The associalion was almost equal to lhat foundfor the excretion of Ihe aflaloxin-DNA adduci in urine. AFP,,shown in Fig. 4C, was excreled in a nonlinear manner wilh asleady level of excrelion al Ihe low doses bul al increased levelsat the high doses. AFQ, was also determined in ihese urinesamples. Al Ihe lowesl dose, AFQ, was nondeleclable in ihreeof four rats, at the intermediale dose three of four rat urinesconlained measurable AFQ, (4.1 ±0.3 ng/24 h), and at Ihehigh dose the range of AFQ, in urine was 110 ±220 ng/24 h.
271
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-/V-GUANINE IN RAT URINE
1000
Do
zI
m
100
10
r=0.99
10 100
AFLATOXIN B DOSE (UG)
1000 -
ox•¿�»100 -
10
AFLATOXIN B DOSE (UG)
OI
1000
100
1010
AFLATOXIN B,
100
DOSE (UG)
Fig. 4. A, linear regression analysis of the excretion of AFB-N7-Gua in urine
over 24 h compared with the dose of AFB,. B, linear regression analysis of theexcretion of AFM, in rat urine over 24 h compared with the dose of AFB,. C,plot of the dose-dependent excretion of AFP, into rat urine.
Thus, the analytical sensitivity for AFQi quantitation precludesthe analysis of these data using linear regression.
At 24 h postdosing all of the rats were killed and the residuallevel of aflatoxin binding to liver DNA was determined. Acomparison of the level of aflatoxin adducts in liver DNA andAFB-N7-Gua excretion in urine over 24 h is found in Fig. 5.
The correlation coefficient and r values were 0.98 and 96.3%,respectively. These data support previous studies (12) that therate of removal of aflatoxin DNA adducts is constant over alarge dose range. Thus, the measurement of the AFB-N7-Gua
adduci in urine accurately reflects DNA damage at the primarysite of aflatoxin-mediated damage in the liver and this biomar-ker appears to be an excellent candidate as a molecular dosimeter for AFBi liver damage.
DISCUSSION
The development of molecular dosimetry methods to monitorhuman exposure to environmental carcinogens will require
techniques that are sensitive, specific, and amenable to largenumbers of samples. It would also be highly desirable to developa method which measures a biologically relevant metabolite,such as a DNA adduci, since these determinations may be usedfor risk assessments of genoloxic damage. Over the past 30years many analytical methods have been applied to the quan-tilation of aflatoxins and its metabolites in biological samples.These techniques include Chromatographie methods such asIhin-layer chromalography (13) and HPLC (14); immunologi-cai assays using monoclonal antibodies or anlisera such asELISA (15, 16), RIA (8), and immunohislochemislry visualization in tissues (17); and instrumentation-based methods suchas synchronous fluorescence spectroscopy (18). Each of thesemethods has its own unique characteristics for specificity andsensitivity and, depending upon the application, the user balances these inherent constraints. For example, to measure asingle aflatoxin metabolite, the Chromatographie method hasthe power to resolve mixtures of aflatoxins into individualcompounds, providing the extraction procedure does not introduce high amounts of nonspecific interfering chemicals. Theantibody-based method is more specific and selective than chro-matography, bul immunoassays are compromised by the intrin-sic cross-reactivity of an antibody for multiple metabolites. Wehave taken advantage of the inherent strengths of both antibodyselectivity and Chromatographie separations to develop a IAC/HPLC procedure to isolate and sensitively measure aflatoxinmetabolites in biological samples.
The application of preparative immunoaffinity chromalography lo Ihe analysis of environmenlal carcinogens has increased steadily and reporls of ils use in aflatoxin analysis (19),benzo(fl)pyrene metabolite delection (20), oxidative DNA damage producÃs(21), and helerocyclic amino measuremenls (22,23) have now appeared in Ihe lileralure. The major analyticaladvantage of this approach is the one-step removal of interfering materials from complex mixtures as diverse as food, serum,and urine samples. The immunoaffinity isolation procedure isnondeslruclive and subsequent chromalographic separationsfollowed by confirmatory analysis using mass spectroscopy andolher inslrumentation methods can be done. A major application of the IAC/HPLC method in our laboratories is in therapid and facile analysis of human samples containing aflatoxins from environmentally exposed people.
In the work reported here we have ulilized 3 of the 11aflaloxin-specific monoclonal anlibodies lo produce a multiplemonoclonal antibody affinity column. The use of ihese multipleantibodies improves the total recovery of aflatoxin metabolites
1000
- 100
10¿1 10 100< AFB LIVER DNA ADDUCTS AT 24 HOURS (P MOLE / MG DNA)
Fig. 5. Linear regression analysis comparing the excretion of AFB-N7-Gua inurine over a 24-h period with the residual aflatoxin-DNA adducts in the liver at24 h postdosing.
272
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-iV-GUANINE IN RAT URINE
from urine, so that 90-95% of the metabolites can be isolated.We selected several monoclonal antibodies the specificities ofwhich are different and complementary to each other for use inthe immunoaffinity preparative method. This preparative multiple antibody column technique has shown that many unknownaflatoxin metabolites exist in rat urine and only one-half of theaflatoxin derivatives chromatographed by HPLC have beenassigned structures. Thus, the development of this procedureaffords the opportunity to purify and then identify many ofthese unknown metabolites. From a molecular dosimetry perspective this knowledge should eventually simplify a morecomplete characterization of aflatoxin metabolism in high andlow risk individuals.
The strategy used to make the antigens for the monoclonalantibodies involved the derivatization of the parent aflatoxinthrough the unsaturated bond in furo-furan ring structure. Thiswas done to produce a hapten that would be similar to anepoxide-derived metabolite, such as the DNA and protein ad-ducts. As seen in the specificity data shown in Table 1, thisstrategy proved successful. However, other investigators haveused other chemistries to produce aflatoxin specific monoclonalantibodies. In particular, the keto group found in the cyclopen-tenone ring is applicable to carboxymethyloxime synthesis (24),and antibodies produced by this method show high specificityfor the furo-furan ring of the aflatoxin but did not generallycross-react with the DNA adducts (24). Thus, the epitope ofthe aflatoxin-specific monoclonal antibodies can be directed bythe synthesis of specific haptens.
The data reported in this paper provide information aboutthe molecular dosimetry of aflatoxin metabolite excretion inrat urine. Besides the dose-dependent measurements of AFB-N7-Gua in urine, other oxidative metabolites, AFM,, AFQ,,
and AFP,, were quantified. The doses used in these experimentsranged from 4.2 to 141 ¿¿g/animaland this is comparable toknown human exposures to AFB, from the diet (25). Theexcretion of the aflatoxin DNA adduci and AFMi were linearwith dose and the correlation coefficients were 0.99 and 0.93,respectively. The analyses of AFPi excretion revealed a steadylevel of excretion at the low doses but an increase at the highdose level. The lack of linearity in the excretion of this metabolite may be explained by use of the glucuronidation pathwayof this aflatoxin metabolite. It is known that the glucuronidation pathway can become saturated and at the two low doses ofAFBl5 the AFP, may be predominantly a glucuronide that isexcreted in the bile (26). Only at the high dose, when saturationof this pathway has occurred, is the AFP, excreted as anunconjugated compound in urine. AFQi was determined inthese samples, but the analytical sensitivity for AFQ, quanti-tation precluded its evaluation as a biomarker in urine.
The association between the measurement of AFB-N7-Gua
in urine and the liver DNA was assessed 24 h postdosing.Previously, we had determined that the rate of removal ofaflatoxin from DNA was dose independent with a DNA adducihalf-life of between 8 and 10 h (12). A comparison of the levelof aflatoxin adducts in liver DNA and AFB-N7-Gua excretionin urine over 24 h showed a correlation coefficient and r2 values
are 0.98 and 96.3%, respectively. These data support previousstudies (12, 27) showing that the rate of removal of aflatoxin-DNA adducts is conslant over a large dose range. As a consequence, the measurement of the AFB-N7-Gua adduci in urine
accurately reflects DNA damage at the primary site of aflatoxinmediated damage in the liver. Since these data were generaledonly following a single dose, further study remains to be done
to assess the molecular dosimetry of AFB-N7-Gua adduci fol
lowing chronic administrations, which is more reflective ofhuman exposure from the diet.
The importance of AFB-N7-Gua measurements in urine has
been stimulated by recent research of human hepatocellularcarcinomas obtained from China and Southern Africa whichhave revealed a hot spot for mutation in exon 7 at position 249of the tumor suppressor gene, p53 (28, 29). Almost one-half ofthe tumors examined had a guanine to thymine transversionmutation at this site. Since transversion mutations are causedby AFB| (30) and because these tumors were isolated fromregions of the world with high aflatoxin exposure, it has beensuggesled lhal Ihese mulations are the result of aflatoxin-DNAdamage. While the biological importance of mutant p53 isbeing suggested for a wide number of human tumors (31), it isalso important thai the molecular dosimetry methods describedfor AFB-N7-Gua may become very useful in epidemiological
investigations to examine the role of mutanl genes in livercancer. Finally, since mutation of the p53 gene can occur wellafter initialion (32), then limiting aflatoxin DNA damage maynot only lower the initiated cell population but also the progression events in human carcinogenesis.
REFERENCES
1. Busby, W. F., and Wogan, G. N. Aflatoxins. In: C. E. Searle (ed.). ChemicalCarcinogens, Ed. 2. pp. 945-1136. Washington. DC: American ChemicalSociety, 1985.
2. Van Rensburg, S. J., Cook-Mozaffari, P., Van Schalkwyk. D. J.. Van derWatt, J. J., Vincent, T. J., and Purchase, I. F. Hepatocellular carcinoma anddietary aflatoxin in Mozambique and Transkei. Br. J. Cancer, 57: 713-726,1985.
3. Peers, F., Bosch, X., Kaldor, J., Linsell, A., and Pluijmen, M. Aflatoxinexposure, hepatitis B virus infection and liver cancer in Swaziland. Int. J.Cancer, 39: 545-553, 1987.
4. Yen, F-S., Yu, M-C., Mo, C-C, Luo, S., Tong, M-J., and Henderson. B. E.Hepatitis B virus, aflatoxins, and hepatocellular carcinoma in southernGuangxi, China. Cancer Res. 49: 2506-2509, 1989.
5. Essigmann, J. M., Croy, R. G., Nadzan, A. M., Busby, W. F., Jr., Reinhold.V. N., Buchi, G., and Wogan, G. N. Structural identification of the majorDNA adduci formed by aflatoxin B, in vitro. Proc. Nati. Acad. Sci. USA, 74:1870-1874, 1976.
6. Sabbioni, G., Skipper, P.. Buchi, G., and Tannenbaum, S. R. Isolation andcharacterization of the major serum albumin adduci formed by aflatoxin B>in vivo in rats. Carcinogenesis (Lond.), 8: 819-824, 1987.
7. Moss, E. J., Judah, D. J., Przyblyski, M., and Neal, G. E. Some mass-spectral and nmr analytical studies of a glutathione conjugate of aflatoxinB,. Biochem. J., 210: 227-233, 1983.
8. Groopman, J. D., Trudel, L. J.. Donahue, P. R., Rothstein. A., and Wogan,G. N. High affinity monoclonal antibodies for aflatoxins and their applicationto solid phase immunoassay. Proc. Nati. Acad. Sci. USA. 81: 7728-7731,1984.
9. Groopman, J. D., Croy, R. G., and Wogan, G. N. In vitro reactions ofaflatoxin B,-adducted DNA. Proc. Nati. Acad. Sci. USA, 78: 5445-5449,1981.
10. Martin, C. N., and Garner, R. C. Aflatoxin B,-oxide generated by chemicalor enzymatic oxidation of aflatoxin B, causes guanine substitution in nucleicacids. Nature (Lond.), 267: 863-865. 1977.
11. Gorst-Allman, C. P., Steyn, P. S., and Wessels, P. L. Oxidation of thebisdihydrofuran moieties of aflatoxin B] and sterigmatocystin; conformationoftetrahydrofurobenzofurans. J. Chem. Soc. Perkin I, 1360-1364, 1977.
12. Groopman, J. D.. Busby. W. F., and Wogan, G. N. The nuclear distributionof aflatoxin B, and its interaction with histones in vivo. Cancer Res., 40:4343-4351, 1980.
13. Trucksess, M. W., Brumley, W. C., and Nesheim. S. Rapid quantitation andconfirmation of aflatoxins in corn and peanut butter, using a disposable silicagel column, thin layer chromatography, and gas chromatography/mass spec-trometry. J. Assoc. Offic Anal. Chem., 67:973-975. 1984.
14. Groopman, J. D., and Donahue. K. F. Aflatoxins, a human carcinogen:analyzing foods and biological samples using monoclonal antibody affinitychromatography. J. Assoc. of Offic. Anal. Chem., 71: 861-867, 1988.
15. Wild, C. P., Umbenhauer, D., Chapot, B.. and Montesano, R. Monitoringof individual human exposure to aflatoxins (AF) and /V-nitrosamines (NNO)by immunoassays. J. Cell. Biochem., 30: 171-179, 1986.
16. Hsieh. L-L., Hsu. S-W., Chen. D-S., and Santella, R. M. Immunologicaldetection of aflatoxin B.-DNA adducts formed in vivo. Cancer Res., 48:6238-6331.1988.
273
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MOLECULAR DOSIMETRY OF AFB-W-GUANINE IN RAT URINE
17. Wild, C. P., Montesano, R., Van Benthem, J., Scherer, E., and Den Engelse,L. Intercellular variation in aflatoxin B, and G.-DNA adduci levels in rattissues: a quantitative immunocytchemica! study. J. Cancer Res. Clin. Oncol.,116: 134-140, 1990.
18. Autrup, H., Bradley, K. A., Shamsuddin, A. K. M, Wakhisi, J., and Wa-sunna, A. Detection of putative adduci with fluorescence characteristicsidentical to 2,3-dihydro-2-(7'-guanyl)-3-hydroxyaflatoxin B, in human urinecollected in Murang'a district, Kenya. Carcinogenesis (Lond.), 4:1193-1195,
1983.19. Groopman, J. D., Donahue, P. R., Zhu, J., Chen, J., and Wogan, G. N.
Aflatoxin metabolism in humans: detection of metabolites and nucleic acidadducts in urine by affinity chromatography. Proc. Nati. Acad. Sci. USA, 82:6492-6497, 1985.
20. Garner, R. C., Dvorackova, I., and Turi, F. Immunoassay procedures todetect exposure to aflatoxin B> and benzo(a)pyrene in animals and man atthe DNA level. Int. Arch. Occup. Environ. Health, 60: 145-150, 1988.
21. Degan, P., Shigenaga, M. K., Park, E-M., Alperin, P. E., and Ames, B. N.Immunoaffinity isolation of urinary 8-hydroxy-2'-deoxyguanosine and 8-hydroxyguanine and quantitation of 8-hydroxy-2'-deoxyguanosine in DNAby polyclonal antibodies. Carcinogenesis (Lond.), 12: 865-871, 1991.
22. Vanderlaan, M., Alexander, J., Thomas, C., Djanegara, T., el al. Immuno-chemical detection of rodent hepatic and urinary metabolites of cooking-induced food mutagens. Carcinogenesis (Lond.), 12: 349-354, 1991.
23. Turesky, R. J., Forster, C. M., Aeschbacher, H. U., Wurzner, H. P., Skipper,P. L., Trudel, L. J., and Tannenbaum, S. R. Purification of the food-bornecarcinogens 2-amino-3-methylimidazo [4,5-/]quinoline and 2-amino-3,8-di-methylimidazo(4,5-/]quinoxaline in heated meat products by immunoaffinity
chromatography. Carcinogenesis (Lond.), 10: 151-156, 1989.24. Pestka, J. J., and Chu, F-S. Aflatoxin Bl dihydrodiol antibody: production
and specificity. Appi. Environ. Microbiol., 47: 472-477, 1984.25. Groopman, J. D., Busby, W. F., Donahue, P. R., and Wogan, G. N.
Aflatoxins as risk factors for liver cancer: an application of monoclonalantibodies to monitor human exposure. In: C. C. Harris (ed.), Biochemicaland Molecular Epidemiology of Cancer, pp. 233-256. New York: Alan R.Liss, Inc., 1986.
26. Holeski, C. J., Eaton, D. L., Monroe, D. H., and Bellamy, G. M. Effects ofphénobarbitalon the biliary excretion of aflatoxin Pi-glucuronide and aflatoxin Bi-S'-glutathione in the rat. Xenobiotica, 17: 139-153, 1987.
27. Bennett, R. A., Essigmann, J. M., and Wogan, G. N. Excretion of anaflatoxin-guanine adduct in the urine of aflatoxin B.-treated rats. CancerRes., 41: 650-654, 1981.
28. Hsu, I. C., Metcalf, R. A., Sun, T., Wesh, J. A., Wang, N. J., and Harris, C.C. Mutational hotspot in the p53 gene in human hepatocellular carcinomas.Nature (Lond.), 350:427-428, 1991.
29. Bressac, B., Kew, M., Wands, J., and Ozturk, M. Selective G to T mutationsof p53 gene in hepatocellular carcinoma from Sourthern Africa. Nature(Lond.), 350: 429-431, 1991.
30. Foster, P. L., Eisenstadt, E., and Miller. J. H. Base substitution mutationsinduced by metabolically activated aflatoxin Bi. Proc. Nati. Acad. Sci. USA,«0:2695-2698, 1983.
31. Hollstein, M., Sidransky, D., Vogelstein. B., and Harris, C. C. p53 mutationsin human cancers. Science (Washington DC), 253: 49-53, 1991.
32. Baker, S. J., Markowitz, S., Fearon, E. R., Willson, J. K., and Vogelstein,B. Suppression of human colorectal carcinoma cell growth by wild-type p53.Science (Washington DC), 249: 912-915,1990.
274
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1992;52:267-274. Cancer Res John D. Groopman, Julia A. Hasler, Laura J. Trudel, et al. Liquid ChromatographyAffinity Chromatography and Immunoaffinity/High PerformanceOther Aflatoxin Metabolites by Multiple Monoclonal Antibody
-guanine and7NMolecular Dosimetry in Rat Urine of Aflatoxin-
Updated version
http://cancerres.aacrjournals.org/content/52/2/267
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on August 7, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Related Documents











