ENVIRONMENTAL BIOTECHNOLOGY Molecular determinants of azo reduction activity in the strain Pseudomonas putida MET94 Sónia Mendes & Luciana Pereira & Carlos Batista & Lígia O. Martins Received: 21 March 2011 /Revised: 29 April 2011 /Accepted: 2 May 2011 # Springer-Verlag 2011 Abstract Azo dyes are the major group of synthetic colourants used in industry and are serious environmental pollutants. In this study, Pseudomonas putida MET94 was selected from 48 bacterial strains on the basis of its superior ability to degrade a wide range of structurally diverse azo dyes. P . putida is a versatile microorganism with a well- recognised potential for biodegradation or bioremediation applications. P . putida MET94 removes, in 24 h and under anaerobic growing conditions, more than 80% of the majority of the structurally diverse azo dyes tested. Whole cell assays performed under anaerobic conditions revealed up to 90% decolourisation in dye wastewater bath models. The involvement of a FMN dependent NADPH: dye oxidoreductase in the decolourisation process was sug- gested by enzymatic measurements in cell crude extracts. The gene encoding a putative azoreductase was cloned from P . putida MET94 and expressed in Escherichia coli. The purified P . putida azoreductase is a 40 kDa homodimer with broad substrate specificity for azo dye reduction. The presence of dioxygen leads to the inhibition of the decolourisation activity in agreement with the results of cell cultures. The kinetic mechanism follows a ping-pong bi–bi reaction scheme and aromatic amine products were detected in stoichiometric amounts by high-performance liquid chromatography. Overall, the results indicate that P . putida MET94 is a promising candidate for bioengineer- ing studies aimed at generating more effective dye-reducing strains. Keywords Pseudomonas putida . Azoreductase . Azo dyes . Decolourisation . Whole cell catalysis Introduction Synthetic dyes are xenobiotic compounds that are being increasingly used in several industries, with special empha- sis in the textile and leather industries. Over 100,000 commercial dyes exist and more than 7×10 5 t of dyestuff is produced annually, of which 1–1.5×10 5 is released into the environment in the wastewaters (Rai et al. 2005). These dyes are hardly removed from the effluents by conventional wastewater treatment and represent anthropogenic pollu- tants causing deterioration of water quality and become a health threat because of their mutagenic or carcinogenic properties (Rai et al. 2005; van der Zee and Villaverde 2005). Biological treatment technologies are attractive alternatives to traditional physicochemical methods as they are low cost, environmentally friendly, and publicly acceptable treatment technologies (Anjaneyulu et al. 2005; Chen 2006; Husain 2006; Rodriguez-Couto 2009). In the past few years, it has been demonstrated that many microorganisms are able to transform azo dyes to non- coloured compounds or even mineralise them completely Electronic supplementary material The online version of this article (doi:10.1007/s00253-011-3366-4) contains supplementary material, which is available to authorized users. S. Mendes : L. Pereira : C. Batista : L. O. Martins (*) Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa, Av. da República, 2780-157 Oeiras, Portugal e-mail: [email protected] Present Address: C. Batista Ydreams, Madan Parque, Quinta da Torre, 2829-516 Caparica, Portugal Appl Microbiol Biotechnol DOI 10.1007/s00253-011-3366-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ENVIRONMENTAL BIOTECHNOLOGY
Molecular determinants of azo reduction activityin the strain Pseudomonas putida MET94
Sónia Mendes & Luciana Pereira & Carlos Batista &
Lígia O. Martins
Received: 21 March 2011 /Revised: 29 April 2011 /Accepted: 2 May 2011# Springer-Verlag 2011
Abstract Azo dyes are the major group of syntheticcolourants used in industry and are serious environmentalpollutants. In this study, Pseudomonas putida MET94 wasselected from 48 bacterial strains on the basis of its superiorability to degrade a wide range of structurally diverse azodyes. P. putida is a versatile microorganism with a well-recognised potential for biodegradation or bioremediationapplications. P. putida MET94 removes, in 24 h and underanaerobic growing conditions, more than 80% of themajority of the structurally diverse azo dyes tested. Wholecell assays performed under anaerobic conditions revealedup to 90% decolourisation in dye wastewater bath models.The involvement of a FMN dependent NADPH: dyeoxidoreductase in the decolourisation process was sug-gested by enzymatic measurements in cell crude extracts.The gene encoding a putative azoreductase was clonedfrom P. putida MET94 and expressed in Escherichia coli.The purified P. putida azoreductase is a 40 kDa homodimerwith broad substrate specificity for azo dye reduction. Thepresence of dioxygen leads to the inhibition of the
decolourisation activity in agreement with the results ofcell cultures. The kinetic mechanism follows a ping-pongbi–bi reaction scheme and aromatic amine products weredetected in stoichiometric amounts by high-performanceliquid chromatography. Overall, the results indicate thatP. putida MET94 is a promising candidate for bioengineer-ing studies aimed at generating more effective dye-reducingstrains.
Keywords Pseudomonas putida . Azoreductase .
Azo dyes . Decolourisation .Whole cell catalysis
Introduction
Synthetic dyes are xenobiotic compounds that are beingincreasingly used in several industries, with special empha-sis in the textile and leather industries. Over 100,000commercial dyes exist and more than 7×105 t of dyestuff isproduced annually, of which 1–1.5×105 is released into theenvironment in the wastewaters (Rai et al. 2005). Thesedyes are hardly removed from the effluents by conventionalwastewater treatment and represent anthropogenic pollu-tants causing deterioration of water quality and become ahealth threat because of their mutagenic or carcinogenicproperties (Rai et al. 2005; van der Zee and Villaverde2005). Biological treatment technologies are attractivealternatives to traditional physicochemical methods as theyare low cost, environmentally friendly, and publiclyacceptable treatment technologies (Anjaneyulu et al. 2005;Chen 2006; Husain 2006; Rodriguez-Couto 2009). In thepast few years, it has been demonstrated that manymicroorganisms are able to transform azo dyes to non-coloured compounds or even mineralise them completely
Electronic supplementary material The online version of this article(doi:10.1007/s00253-011-3366-4) contains supplementary material,which is available to authorized users.
S. Mendes : L. Pereira : C. Batista : L. O. Martins (*)Instituto de Tecnologia Química e Biológica,Universidade Nova de Lisboa,Av. da República,2780-157 Oeiras, Portugale-mail: [email protected]
Present Address:C. BatistaYdreams,Madan Parque, Quinta da Torre,2829-516 Caparica, Portugal
Appl Microbiol BiotechnolDOI 10.1007/s00253-011-3366-4

under certain environmental conditions. However, the factthat most of the dye pollutants persist in the environmentemphasises the natural inadequacy of microbial activity todeal with such xenobiotic compounds. Indeed, biologicalsystems need to exhibit not only a high catalytic versatilitytowards the degradation of a complex mixture of structur-ally different dyes but also a superior robustness against thetoxic effects of the dyes and other substances present in theeffluent (salts, detergents, surfactants, and metals), often atextreme pHs and high temperatures. Because of theserequirements, there is currently no simple solution for thebiologic treatment of dye-containing effluents (Anjaneyuluet al. 2005; Chen 2006).
Enzymatic processes are particularly sought for thetreatment of dyeing effluents because of their specificity;enzymes only attack the dye molecules, while valuabledyeing additives or fibres are kept intact and can potentiallybe re-used. Likewise, new recycling technologies will allowthe reduction of the enormous water consumption in thetextile finishing industry (e.g. up to 150 L of water to dye1 kg of cotton; Kandelbauer and Guebitz 2005). Therefore,the identification and characterisation of individualenzymes that make a discernible contribution to thedegradation of xenobiotics pave the way for the improve-ment of enzymatic systems, through protein engineeringstrategies, to maximise their biodegradation, biotransfor-mation, or bioremediation potential. Dye molecules aremainly degraded by a few redox-active enzymes, allshowing relatively wide substrate specificity: laccases,peroxidases, azoreductases, and mono- or di-oxygenases(Kandelbauer and Guebitz 2005).
The focus of our research is to investigate the enzymaticbiodegradation and bioconversion of synthetic dyes, andrecently, we have characterised the properties of a bacteriallaccase, CotA-laccase from Bacillus subtilis, in the decol-ourisation of azo and antraquinonic dyes (Pereira et al.2009a, b). In this study, the decolourisation of azo dyes bythe bacterium Pseudomonas putida MET 94 was addressed.Several bacterial species capable of decolourising dyesunder aerobic and especially under anaerobic conditionshave been reported and the ability of P. putida for dyedecolourisation was previously identified (Mansour et al.2007, 2009). However, the studies have focused mainly onthe toxicity of the dyes and their degradation products andthe physiological aspects and detailed reaction mechanismswere unknown. P. putida is known to play a key role in themaintenance of the quality of natural environments mainlybecause of its strong capability in the degradation andbiotransformation of biogenic and xenobiotic pollutants(Loh and Cao 2008). It is also considered an interestingagent for many potential biotechnological applications as ithas a robust, rapid growth, ease of lab handling, and isamenable to genetic manipulation (Timmis and Pieper
1999; de Lorenzo 2001; Nelson et al. 2002). Therefore, inthis work, we set out to fully characterise for the first timeat the physiological and molecular level, the decolourisa-tion process of P. putida MET94. A new azoreductaseP. putida azoreductase (PpAzoR) from P. putida MET94was cloned, heterologously expressed, and characterised.The enzyme broad specificity for azo dyes, under anaerobicconditions, points to a major role of PpAzoR in thebacterium decolourisation process. Moreover, whole cellsystems proved to be very effective in the decolourisationof single-dye molecules and of model dye baths composedof a mixture of commercially important dyes at a high ionicstrength. This work is expected to allow the future design,by protein engineering techniques, of optimised micro-organisms targeted for the biodegradation of wastewaterscontaining dyes.
Methods
Bacterial strains, media, and plasmids Forty-eight differentbacterial strains from different sources (Table 1S) werescreened for their ability to degrade dyes. For the presentstudy, strain P. putida MET94 (BCCM/LMG 26299=CECT7881=NCIMB 14727), an offer of M. J. Fernandes (ITQB),was used. P. putida MET94 was routinely cultivated inLuria Bertani (LB) medium at 30 °C. Escherichia coliDH5α and E. coli Tuner (DE3; Novagen) were used as hoststrains for molecular biology work. E. coli strains wereroutinely cultured at 37 °C in LB medium, which wassupplemented with ampicillin (100 μg L−1), if appropriate.The plasmid pET-21a (+) (Novagen) was used for thecloning experiments.
Chemicals All chemicals were of the highest grade avail-able commercially. The dyes tested are listed in Table 1.Three different model wastewaters, designed in the frame ofthe European Commission project SOPHIED to mimiceffluents produced during wool and cotton textile dyeingprocesses, were prepared: direct bath (for cotton) withDB71, DR80, and DY106 dyes at 1 g L−1 each and 5 g L−1
of NaCl, pH 8; acid bath (for wool) with AY49, AR266,AB210, and AB194 dyes at 0.1 g L−1 each and 2 g L−1 ofNa2SO4, pH 4; and reactive bath (for cotton) with RB222,RR195, RY145, and RB5 dyes at 1.25 g L−1 each and70 g L−1 of Na2SO4, pH 5 (Prigione et al. 2008). All dyesolutions were sterilised by tyndallisation before use.
Culture conditions Cultures of P. putida MET94 weregrown in LB medium supplemented with increasingconcentrations of dyes (0–5 mM) under aerobic andanaerobic conditions at 30 °C. Aerated cultures were grownin Erlenmeyer flasks at 150 rpm shaking. Anaerobic
Appl Microbiol Biotechnol
cultures were grown under static conditions in 100 mLrubber stopper glass bottles in media previously degassedthrough nitrogen flush. Cell growth was monitored bymeasuring the turbidity of the cultures at 600 nm (OD600).Decolourisation was assessed by measuring the absorbanceof the culture supernatants at the wavelength of maximalabsorbance for each dye tested (Table 1).
Preparation of resting whole cells Cells were harvested at thelate exponential phase of growth by centrifugation (7,000×g,10 min, 4 °C) and washed twice with a 0.9% (w/v) NaClsolution. For the enzymatic assays using resting cells, thewashed pellets were suspended in Britton Robinson (BR)buffer (0.1 M phosphoric acid, 0.1 M boric acid, and 0.1 Macetic acid titrated to the desired pH with 0.5 M NaOH),pH 8, to an OD600=6, unless otherwise stated. The cellconcentration was estimated by the standard gravimetricmethod, cell weight (dry weight; g/L)=(OD600−0.0117)/1740.
Preparation of cell-free extracts Cell suspensions preparedin BR buffer, pH 8, containing 5 mMMgCl2, 10 μg mL−1 ofDNase I, and 2 μL mL−1 of a mixture of protease inhibitors
(Complete™ mini-EDTA-free protease inhibitor mixturetablets, Roche), were disrupted in a French press. Cell debriswere removed by centrifugation (18,000×g, 2 h, 4 °C), andthe supernatants were used for the enzymatic assays.
Cloning and overexpression of ppAzoR in E. coli GenomicDNA of P. putida MET94 was obtained by centrifuging200 μL of a fresh P. putida culture for 1 min at 13,000 rpm.The pellet was washed twice and suspended in 50 μL ofMilli Q water. This sample was boiled for 5 min and theDNA was applied as a template for gene-specific primedpolymerase chain reaction (PCR) amplification. The ppAzoRgene was amplified by PCR using chromosomal DNA andthe primers PpaF (5′ GGAGAGTCATATGAAACTGTTGC3′) and PpaR (5′ CAACCAAAGGATCCCTTGATCAGG 3′).The amplified 597-bp PCR product was digested with NdeIand BamHI and inserted between the respective restrictionsites of the plasmid pET-21a (+) to yield pLP-1. Theconstruct was introduced into the host expression strain E.coli Tuner (DE3) in which the PpAzoR protein was producedunder the control of the T7lac promoter to obtain the strainLOM528. The recombinant strain was grown in LB medium
CI generic name CIconstitutionnumber
Dye content(%)
Absorptionmax (nm)
Extinctioncoefficient(M−1 cm−1)
Direct blue 1 (DB1) 24410 80 610 –
Direct red R (DRR) 22120 91 530 35,384
Direct black 38 (DB38) 30235 50 600 –
Direct blue 71 (DB71) 34140 NSa 565 –
Direct red 80 (DR80) 35780 NS 555 –
Direct yellow 106 (DY106) 1332 >50 420 –
Reactive red 4 (RR4) 18105 50 530 –
Reactive black 5 (RB5) 20505 55 600 39,850
Reactive yellow 145 (RY145) –b >50 420
Reactive yellow 81 (RY81) –b 45 360 12,991
Reactive blue 222 (RB222) –b NS 600 –
Reactive red 195 (RR195) –b >30 550 –
Acid red 299 (AR299) –b NS 440 7,280
Acid black 210 (AB210) 300285 >30 600 –
Acid yellow 49 (AY49) 18640 50 390 16,369
Acid black 194 (AB194) –b >60 570 11,927
Acid red 266 (AR266) 17101 >30 470 9,538
Acid blue 62 (AB62) 62045 100 600 –
Acid orange 7 (AO7) 15510 20–30 480 9,538
Sudan orange G (SOG) 11920 98 430 30,154
Methyl red (MR) 11920 95 430 23,360
Mordant black 3 (MB3) 14640 30 550 5,569
Mordant black 9 (MB9) 14855 60–85 550 15,641
Mordant black 17 (MB17) 15705 50 530 10,351
Table 1 Colour index (CI)generic names, CI registrationnumbers, absorption maxima,purity, and calculated molarextinction coefficients of thedyes used in this study
The dyes were purchased fromSigma-Aldrich (St. Louis, MO,USA), Merck (Darmstadt,Germany), Town End (Leeds,UK), DyStar Textilfarben(Germany), Yorkshire Europe(Belgium), and Bezema AG(Montlinglen, Switzerland). Theabsorption maxima were deter-mined in BR buffer (pH 7). Themolar extinction coefficients werecalculated by using the dye puritiesindicated. If the dye purity was notindicated by the supplier, then itwas assumed that the preparationsconsisted of a pure dyea NS, dye purity not specified bythe supplierb Confidential
Appl Microbiol Biotechnol

supplemented with ampicillin (100 μg L−1) at 37 °C.Growth was followed up to an OD600 of 0.6, and at thatpoint, 100 μM isopropyl-β-D-thiogalactopyranoside wasadded to the culture medium, and the temperature waslowered to 25 °C. Incubation was continued for a further5 h and the cells were harvested by centrifugation(8,000×g, 10 min, 4 °C).
Purification of recombinant PpAzoR The cell sediment wassuspended in 20 mM Tris–HCl buffer (pH 7.6) containingDNase I (10 μg mL−1 extract), MgCl2 (5 mM), and amixture of protease inhibitors, antipain, and leupeptin(2 μg mL−1 extract). Cells were disrupted and cell debrisremoved as described above. The cell lysate was thenloaded onto a Q-Sepharose column equilibrated with20 mM Tris–HCl buffer, pH 7.6. The active fractions werepooled, dialysed to 20 mM Tris–HCl (pH 7.6), and appliedonto a Mono-Q™ 5/50 (GE Healthcare Bio-Sciences).Active fractions were pooled and concentrated before beingapplied onto a Superdex 75 HR 10/30 column (GEHealthcare) equilibrated with 20 mM Tris–HCl buffer,pH 7.6, with 0.2 M NaCl. All purification steps werecarried out at room temperature in an AKTA-purifier (GEHealthcare).
Identification of the prosthetic group The purified PpAzoRin 20 mM Tris–HCl buffer, pH 7.6, with 0.2 M of NaCl washeated at 100 °C for 10 min in the dark. After cooling onice, the denatured protein was removed by centrifugation(13,000×g, 10 min, 4 °C). The resulting supernatant wasanalysed by high-performance liquid chromatography(HPLC). HPLC analysis was performed in an HPLC MerckHitachi with a diode-array system, using a reverse phaseC-18 column (250×4 mm length, 5 μM particle size, andpore size of 100 Å from LiChrospher 100 RP-18, Merck) asdescribed previously (Lewis and Escalante-Semerena2006). Authentic standards of flavin mononucleotide(FMN) and flavin adenine dinucleotide (FAD) were usedfor comparative purposes.
UV-visible spectrum of PpAzoR and molar extinctioncoefficient The UV-visible absorption spectra of the purifiedenzyme and the released FMN cofactor after chemicaldenaturation with 0.4% (w/v) sodium dodecyl sulfate (SDS)were recorded at room temperature by using a NicoletEvolution 300 spectrophotometer from Thermo Industries(Waltham, MA, USA). The enzyme extinction coefficient wasdetermined using the following equation: ε455=εFMN×A455/A445, where A455 is the enzyme absorbance at 455 nm, A445 isthe free FMN absorbance at 445 nm, and εFMN is the freeFMN extinction coefficient at 445 nm (12,500 M−1 cm−1).Purified enzyme concentration was then estimated from itsA455 value.
Enzyme assays Typically, the initial rates of dye decolour-isation were determined by monitoring the decrease in dyeabsorbance on a Nicolet Evolution 300 spectrophotometer(Thermo Industries) at 30 °C in BR buffer, pH 7, unlessotherwise stated. The bottles containing 0.25 mM nicotin-amide adenine dinucleotide phosphate-oxidase (NADPH)and 2 mM of the dye were sealed with rubber stoppers andmade anaerobic by argon bubbling. Reactions were initiatedby the injection of anoxic-made enzymatic preparations(cell extracts, whole cells, or purified enzyme) through theuse of a syringe. Samples were withdrawn at differenttimes, diluted, and the absorbance measured at theappropriated wavelength. The molar absorbance of thedyes in BR buffer, pH 7, is listed in Table 1. One unit (U) ofenzymatic activity was defined as the amount of enzymerequired to reduce 1 μmol of the substrate per minute. Thedependence of purified PpAzoR activity on pH (in therange of 4–10) and temperature (in the range of 20–40 °C)was measured by monitoring the decrease in the absorbanceof 0.5 mM of methyl red. All enzymatic assays wereperformed at least in triplicate.
The apparent kinetic parameters of NADPH or NADHoxidation were followed in the reactions monitored at340 nm (ε=6,200 or 6,220 M−1 cm−1 for NADH andNADPH, respectively) in the presence of 2 mM RB5. Theapparent kinetic parameters for azo dyes were determinedby following the decrease in absorbance at the maximalwavelength for each dye in the assays containing 0.25 mMNADPH and different concentrations of the dye (0.05–5mM).The apparent kinetic constantsKm and kcat were fitted directlyinto the Michaelis–Menten equation (Originlab software,Northampton, MA, USA).
The second-order kinetics were measured by monitor-ing the oxidation of NADPH in anoxic reactions containingthe BR buffer, pH 7, by varying the concentration of RB5(from 0.25 to 2 mM) or NADPH (from 0.03 to 0.5 mM), whilethe concentration of the other substrate was kept constant(NADPH or RB5). The kinetic constants Km and kcat for thereduction of RB5 or the oxidation of NADPH weredetermined from the Lineweaver–Burk double-reciprocalplots.
For the oxygen inhibition assays, a saturated solutionof dioxygen (465 nmol mL−1) was prepared by bubblingwith oxygen and different volumes of this solution wasadded to the oxygen-free reaction mixtures to give thefinal concentration of dioxygen in solution. The deaeratedreaction mixtures contained 0.1 mM RB5 and 0.25 mMNADPH in the BR buffer, pH 7, solution. The initial ratesof the reaction were monitored by following RB5reduction.
Identification of azo dye products Analysis of enzymaticdye products was carried out by HPLC (described above).
Appl Microbiol Biotechnol

The samples were eluted isocratically at a flow rate of0.2 mL min−1. The mobile phase consisted of 25 mMphosphate buffer (pH 7.5) and acetonitrile (4:6, v/v;Moutaouakkil et al. 2003). Authentic standards of N,N′-dimethyl-p-phenylenediamine, 2-aminobenzoic acid, ani-line, and 4-aminoresorcinol were used for comparativepurposes.
Other methods The sequence of the first five amino acids atthe N-terminus of the purified recombinant PpAzoR wasdetermined by stepwise Edman degradation in a pulse-liquid automated Applied Biosystem protein sequencer(model 477A) coupled with HPLC at the Instituto deTecnologia Química e Biológica microsequencing facility.The molecular mass of PpAzoR protein was determined ona gel filtration Superose 12 10/300 GL (GE Healthcare Bio-Sciences, Sweden) column equilibrated with 20mMTris–HClbuffer, pH 7.6, containing 0.2 M NaCl. Thyroglobulin(670 kDa), γ-globulin (158 kDa), ovalbumin (44 kDa),myoglobin (17 kDa), and vitamin B12 (1.35 kDa) were usedas standards (Bio-Rad Laboratories). The protein concentra-tion was determined by using the Bradford assay with bovineserum albumin as the standard.
Results
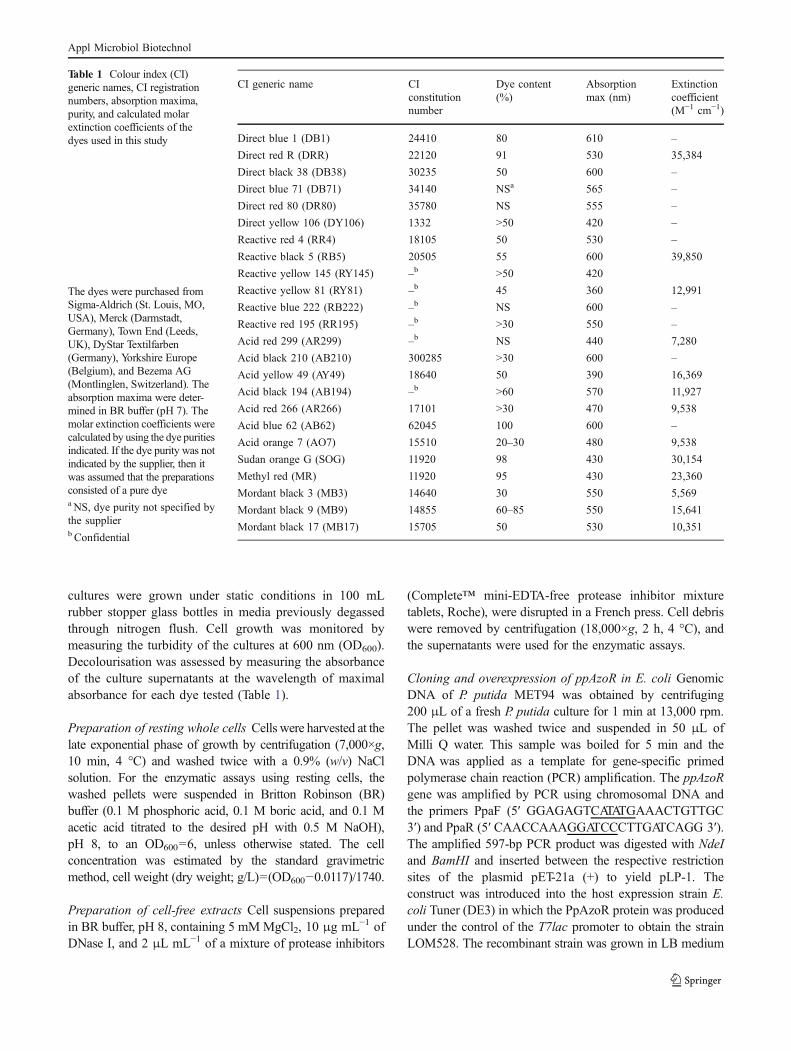
Screening for dye decolourisation The decolourisationscreening using different bacteria (Table 1S) was assessedduring growth in LB medium at 30 °C at static or stirredconditions (150 rpm) in the presence of 30 ppm of thefollowing dyes: AB62, AR299, RB19, RB5, RR4, RY81,DRR, DB1, and DB38. From this pre-screening, 10 strainswere selected for a second round of screening at higherconcentrations of the dyes (from 30 to 1,000 ppm). P. putidastrain MET94 was selected from the other bacterial strainstested for their ability to decolourise at higher extent all theazo dyes tested after 24 h of growth (Fig. 1).
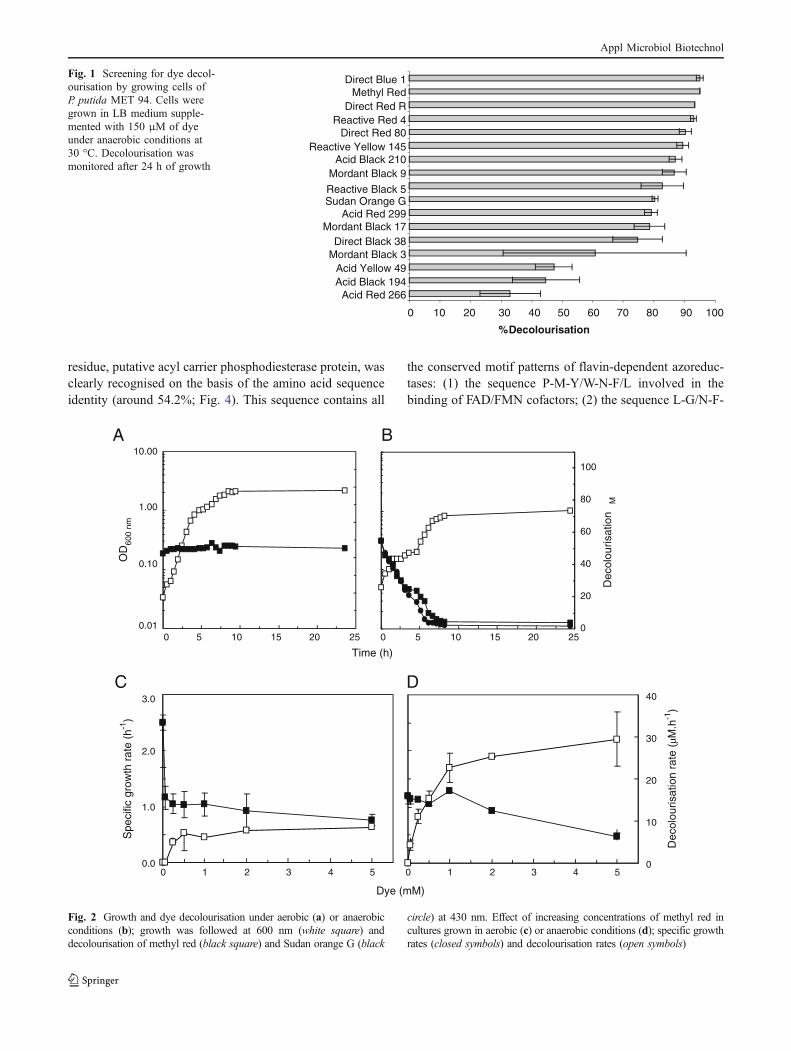
Effect of oxygen and dye concentration on growth anddecolourisation parameters To assess the physiology ofdye decolourisation by P. putida MET94, cultures weregrown under different aeration conditions and in theabsence or presence of the two structurally simple azodyes, methyl red and Sudan orange G. Decolourisation wasmostly observed in anaerobically grown cultures in agrowth-associated manner (Fig. 2a, b). Increasing theconcentration of the dyes in the culture media led to lowerspecific growth rates, especially in aerobically growncultures, most likely reflecting the toxicity that dyes exertedover cell physiology (Fig. 2c, d). The rates of decolourisa-tion shown by the growing cells increase with increasing
dye concentrations up to values where they becomeindependent of the dye concentration, in a pattern thatsuggests enzymatic dye degradation. Indeed, after culturecentrifugation, cell pellets were colourless, indicating theinexistence of dye adsorption to the bacterial cell wall.Enzymatic reactions using cell crude extracts showedincreased rates upon addition of NADH or NADPH to theassay mixture (five- and 30-fold, respectively), clearlypointing to the involvement of oxidoreductase activity inthe decolourisation process (Table 2). Moreover, slightlyhigher activities were obtained upon addition of FMN, butnot FAD, to the reaction mixture, which associate theactivity of a soluble FMN-dependent oxidoreductase in theenzymatic decolourisation system of P. putida MET94.Crude extracts from cultures grown in the presence orabsence of oxygen, with or without dye, were able todecolourise at similar rates all the azo dyes tested,suggesting that the enzymatic systems involved are consti-tutively expressed in P. putida cultures (Table 2 and data notshown). Utilising NADH and/or NADPH as an electrondonor, azoreductases can decolourise azoic dyes into thecorresponding aromatic amines by reductive cleavage of theazo bond.
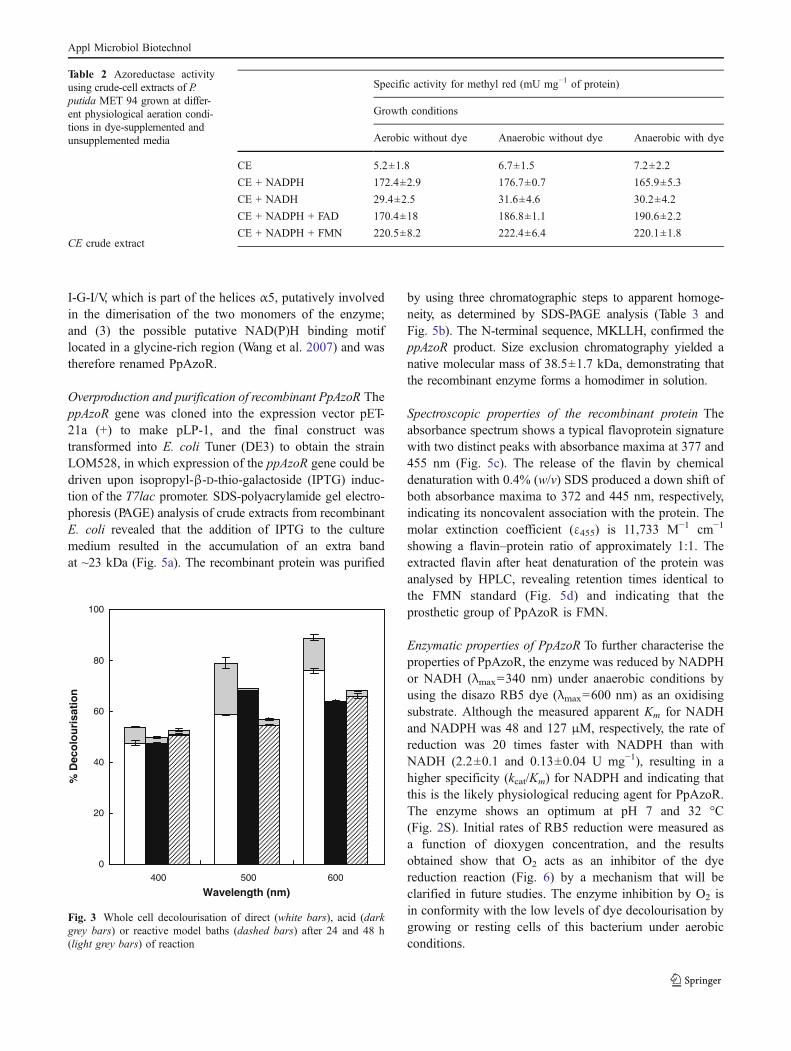
Decolourisation using resting whole cells The applicationof azoreductases in biodegradation processes requires theaddition of cofactors such as NAD(P)H in reactionmixtures, which is prohibitively expensive. Therefore, onlyif whole cells that lead to cofactor regeneration are used, dothese processes become economically feasible. Indeed, theenzymatic activity using whole cells was found to beindependent of the addition of exogenous cofactors such asNADH or NADPH (data not shown). Whole cell assayswith the model dye MR showed that reactions rates increaselinearly with the biomass concentration, following first-order kinetics, with an optimum at around pH 8 (Fig. 1S).The decolourisation of the three model dye wastewaters wasassessed using an OD600=20 (Fig. 3) that allows for maximalrates of decolourisation. These baths were designed to mimiceffluents produced during cotton or wool textile dyeingprocesses containing additives and salts in addition to dyes.The dyes selected are representative of different structuraldye types and are widely applied in the textile industry. Thelevels of decolourisation by the P. putida MET94 whole cellsystem range from 50% to 90% as measured at 400, 500, and600 nm after 72 h of reaction.
Identification of the P. putida azoreductase gene A BasicLocal Alignment Search Tool search of the P. putidaKT2440 genome was performed using template sequencesfrom bacterial azoreductases with known 3-D structures (Itoet al. 2006; Liu et al. 2007b; Wang et al. 2007). A 612-bpORF (acpD, GeneID 1042781) encoding a 203 amino acid
Appl Microbiol Biotechnol
residue, putative acyl carrier phosphodiesterase protein, wasclearly recognised on the basis of the amino acid sequenceidentity (around 54.2%; Fig. 4). This sequence contains all
the conserved motif patterns of flavin-dependent azoreduc-tases: (1) the sequence P-M-Y/W-N-F/L involved in thebinding of FAD/FMN cofactors; (2) the sequence L-G/N-F-
0 10 20 30 40 50 60 70 80 90 100
Acid Red 266Acid Black 194Acid Yellow 49
Mordant Black 3Direct Black 38
Mordant Black 17Acid Red 299
Sudan Orange GReactive Black 5
Mordant Black 9Acid Black 210
Reactive Yellow 145Direct Red 80
Reactive Red 4Direct Red R
Methyl RedDirect Blue 1
%Decolourisation
Fig. 1 Screening for dye decol-ourisation by growing cells ofP. putida MET 94. Cells weregrown in LB medium supple-mented with 150 μM of dyeunder anaerobic conditions at30 °C. Decolourisation wasmonitored after 24 h of growth
0 1 2 3 4 50
10
20
30
40
Dec
olou
risat
ion
rate
(µM
.h-1
)D
0.0
1.0
2.0
3.0
0 1 2 3 4 5
Dye (mM)
Spe
cific
gro
wth
rat
e (h
-1)
C
Time (h)
OD
600
nm
0.01
0.10
1.00
10.00
5 10 15 20 250
A
5 10 15 20 25
Dec
olou
risat
ion
µM
00
20
40
60
80
100
B
Fig. 2 Growth and dye decolourisation under aerobic (a) or anaerobicconditions (b); growth was followed at 600 nm (white square) anddecolourisation of methyl red (black square) and Sudan orange G (black
circle) at 430 nm. Effect of increasing concentrations of methyl red incultures grown in aerobic (c) or anaerobic conditions (d); specific growthrates (closed symbols) and decolourisation rates (open symbols)
Appl Microbiol Biotechnol
I-G-I/V, which is part of the helices α5, putatively involvedin the dimerisation of the two monomers of the enzyme;and (3) the possible putative NAD(P)H binding motiflocated in a glycine-rich region (Wang et al. 2007) and wastherefore renamed PpAzoR.
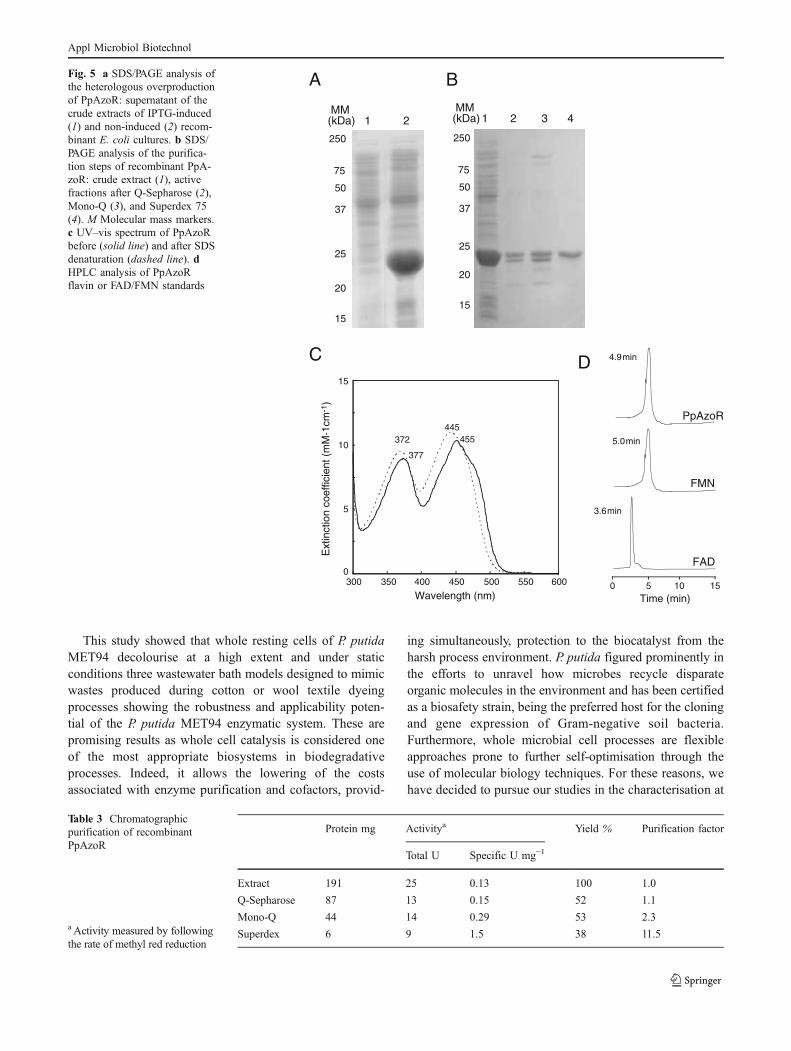
Overproduction and purification of recombinant PpAzoR TheppAzoR gene was cloned into the expression vector pET-21a (+) to make pLP-1, and the final construct wastransformed into E. coli Tuner (DE3) to obtain the strainLOM528, in which expression of the ppAzoR gene could bedriven upon isopropyl-β-D-thio-galactoside (IPTG) induc-tion of the T7lac promoter. SDS-polyacrylamide gel electro-phoresis (PAGE) analysis of crude extracts from recombinantE. coli revealed that the addition of IPTG to the culturemedium resulted in the accumulation of an extra bandat ~23 kDa (Fig. 5a). The recombinant protein was purified
by using three chromatographic steps to apparent homoge-neity, as determined by SDS-PAGE analysis (Table 3 andFig. 5b). The N-terminal sequence, MKLLH, confirmed theppAzoR product. Size exclusion chromatography yielded anative molecular mass of 38.5±1.7 kDa, demonstrating thatthe recombinant enzyme forms a homodimer in solution.
Spectroscopic properties of the recombinant protein Theabsorbance spectrum shows a typical flavoprotein signaturewith two distinct peaks with absorbance maxima at 377 and455 nm (Fig. 5c). The release of the flavin by chemicaldenaturation with 0.4% (w/v) SDS produced a down shift ofboth absorbance maxima to 372 and 445 nm, respectively,indicating its noncovalent association with the protein. Themolar extinction coefficient (ε455) is 11,733 M−1 cm−1
showing a flavin–protein ratio of approximately 1:1. Theextracted flavin after heat denaturation of the protein wasanalysed by HPLC, revealing retention times identical tothe FMN standard (Fig. 5d) and indicating that theprosthetic group of PpAzoR is FMN.
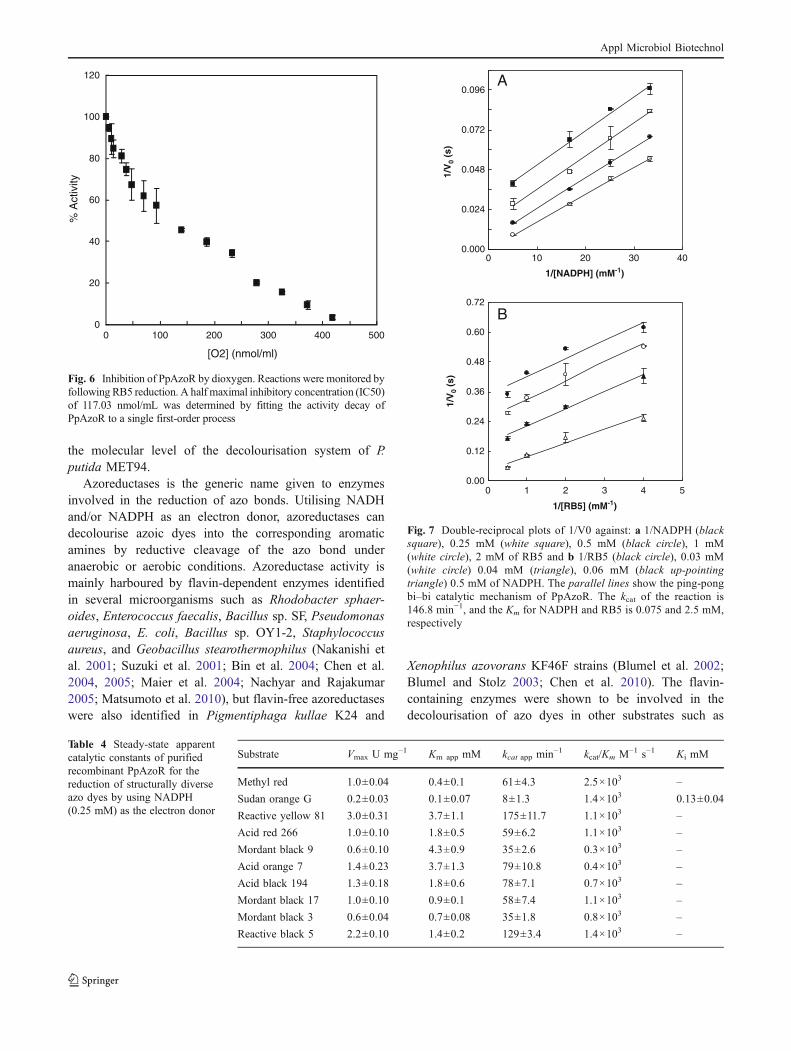
Enzymatic properties of PpAzoR To further characterise theproperties of PpAzoR, the enzyme was reduced by NADPHor NADH (λmax=340 nm) under anaerobic conditions byusing the disazo RB5 dye (λmax=600 nm) as an oxidisingsubstrate. Although the measured apparent Km for NADHand NADPH was 48 and 127 μM, respectively, the rate ofreduction was 20 times faster with NADPH than withNADH (2.2±0.1 and 0.13±0.04 U mg−1), resulting in ahigher specificity (kcat/Km) for NADPH and indicating thatthis is the likely physiological reducing agent for PpAzoR.The enzyme shows an optimum at pH 7 and 32 °C(Fig. 2S). Initial rates of RB5 reduction were measured asa function of dioxygen concentration, and the resultsobtained show that O2 acts as an inhibitor of the dyereduction reaction (Fig. 6) by a mechanism that will beclarified in future studies. The enzyme inhibition by O2 isin conformity with the low levels of dye decolourisation bygrowing or resting cells of this bacterium under aerobicconditions.
Wavelength (nm)
% D
eco
lou
risa
tio
n
0
20
40
60
80
100
400 500 600
Fig. 3 Whole cell decolourisation of direct (white bars), acid (darkgrey bars) or reactive model baths (dashed bars) after 24 and 48 h(light grey bars) of reaction
Specific activity for methyl red (mU mg−1 of protein)
Growth conditions
Aerobic without dye Anaerobic without dye Anaerobic with dye
CE 5.2±1.8 6.7±1.5 7.2±2.2
CE + NADPH 172.4±2.9 176.7±0.7 165.9±5.3
CE + NADH 29.4±2.5 31.6±4.6 30.2±4.2
CE + NADPH + FAD 170.4±18 186.8±1.1 190.6±2.2
CE + NADPH + FMN 220.5±8.2 222.4±6.4 220.1±1.8
Table 2 Azoreductase activityusing crude-cell extracts of P.putida MET 94 grown at differ-ent physiological aeration condi-tions in dye-supplemented andunsupplemented media
CE crude extract
Appl Microbiol Biotechnol

The substrate specificity of PpAzoR was investigated bymeasuring the initial rates of reduction of a set ofstructurally different azo dyes by using NADPH as theelectron donor (Table 4). Interestingly, the enzyme reducesat similar rates all the dyes tested, with reactive yellow 81and reactive black 5 representing the pre-eminent substratesand mordant black 9 and acid orange 7 representing theweaker substrates. In contrast, with the rest of the dyes, thefunction that relates the rate of reaction versus Sudanorange G concentration is a non-hyperbolic curve (data notshown), indicating enzyme inhibition at high substrateconcentration (Ki=0.13 mM), as reported previously(Zimmermann et al. 1982; Pereira et al. 2009b). Most ofthe dyes tested have –OH or –NH2 groups at the orthoposition and electron withdrawing groups, such as SO3 andCl, at the para position relatively to the azo bound,requisites previously reported for azoreductase activity(Zimmermann et al. 1982; Chen 2006; Hsueh and Chen2007). It is noteworthy that reactive yellow 81, showing thehighest rate of reduction, does not obey those conforma-tional requirements.
The catalytic mechanism of PpAzoR The reaction mecha-nism for PpAzoR was investigated by steady-state kineticanalysis. Initial velocity measurements performed in thepresence of NADPH as the electron donor and RB5 as theelectron acceptor resulted in a family of parallel lines in adouble reciprocal plot, indicating a ping-pong bi–bimechanism (Fig. 7). This shows that PpAzoR reduces azodyes in two distinct steps: first, a complete reduction of theFMN cofactor; and second, transfer of these electrons fromthe flavin to the dye substrate resulting in the formation of
the corresponding putative hydrazo derivatives. Thisreaction cycle proceed a second time and delivers thenecessary four electrons in order to obtain a completereduction of the azo dyes to amines. This mechanismwas supported by the detection of aromatic amines, theexpected reduction products of the reaction of twostructurally simple model dyes, methyl red and sudan orangeG, in stoichiometric amounts in the HPLC chromatograms ofthe reaction assays (Fig. 8).
Discussion
Azoic dyes represent more than half of the annuallyproduced dyes and are characterised by a wide structuralvariety. In this study, P. putida MET94 strain emerged as themost efficient and versatile among 48 strains screened fortheir competence in decolourising a range of structurallydiverse azo dyes. Azo dye decolourisation by P. putidaMET94 was shown to be growth associated, occurringmostly under anaerobic conditions. The enzymatic decol-ourisation systems operating in vivo seem to be constitu-tive, being expressed under aerobic or anaerobic conditionsand in the presence or absence of the dye. This is notsurprising considering that these anthropogenic syntheticcompounds are not cognate substrates in an originalphysiological context. Indeed, the observed oxygen inhibi-tion of dye decolourisation by growing cells is exerted atthe level of the enzymatic activity, as shown by kineticmeasurements performed with whole cell systems or cell-free extracts.
Fig. 4 Sequence alignment of flavin-dependent azoreductase from P.putida with azoreductases from E. coli (1V4B), S. typhimurium(1T5B), E. faecalis (2HPV), and P. aeruginosa (2V9C). The alignment
was generated by using CLUSTAL X. Similar residues are highlightedby shading and conserved residues are indicated by white lettering ona black background
Appl Microbiol Biotechnol
This study showed that whole resting cells of P. putidaMET94 decolourise at a high extent and under staticconditions three wastewater bath models designed to mimicwastes produced during cotton or wool textile dyeingprocesses showing the robustness and applicability poten-tial of the P. putida MET94 enzymatic system. These arepromising results as whole cell catalysis is considered oneof the most appropriate biosystems in biodegradativeprocesses. Indeed, it allows the lowering of the costsassociated with enzyme purification and cofactors, provid-
ing simultaneously, protection to the biocatalyst from theharsh process environment. P. putida figured prominently inthe efforts to unravel how microbes recycle disparateorganic molecules in the environment and has been certifiedas a biosafety strain, being the preferred host for the cloningand gene expression of Gram-negative soil bacteria.Furthermore, whole microbial cell processes are flexibleapproaches prone to further self-optimisation through theuse of molecular biology techniques. For these reasons, wehave decided to pursue our studies in the characterisation at
Protein mg Activitya Yield % Purification factor
Total U Specific U mg−1
Extract 191 25 0.13 100 1.0
Q-Sepharose 87 13 0.15 52 1.1
Mono-Q 44 14 0.29 53 2.3
Superdex 6 9 1.5 38 11.5
Table 3 Chromatographicpurification of recombinantPpAzoR
aActivity measured by followingthe rate of methyl red reduction
FAD
PpAzoR
3.6min
FMN
5.0min
4.9min
0 5 10 15Time (min)
D
10
15
0
5
300 350 400 450 500 550 600
C
Ext
inct
ion
coef
ficie
nt (
mM-1
cm-1
)
Wavelength (nm)
372
377
445455
MM(kDa) 1 2
A B
250
75
50
37
25
20
15
MM(kDa) 1 2 3 4
250
75
50
37
25
20
15
Fig. 5 a SDS/PAGE analysis ofthe heterologous overproductionof PpAzoR: supernatant of thecrude extracts of IPTG-induced(1) and non-induced (2) recom-binant E. coli cultures. b SDS/PAGE analysis of the purifica-tion steps of recombinant PpA-zoR: crude extract (1), activefractions after Q-Sepharose (2),Mono-Q (3), and Superdex 75(4). M Molecular mass markers.c UV–vis spectrum of PpAzoRbefore (solid line) and after SDSdenaturation (dashed line). dHPLC analysis of PpAzoRflavin or FAD/FMN standards
Appl Microbiol Biotechnol
the molecular level of the decolourisation system of P.putida MET94.
Azoreductases is the generic name given to enzymesinvolved in the reduction of azo bonds. Utilising NADHand/or NADPH as an electron donor, azoreductases candecolourise azoic dyes into the corresponding aromaticamines by reductive cleavage of the azo bond underanaerobic or aerobic conditions. Azoreductase activity ismainly harboured by flavin-dependent enzymes identifiedin several microorganisms such as Rhodobacter sphaer-oides, Enterococcus faecalis, Bacillus sp. SF, Pseudomonasaeruginosa, E. coli, Bacillus sp. OY1-2, Staphylococcusaureus, and Geobacillus stearothermophilus (Nakanishi etal. 2001; Suzuki et al. 2001; Bin et al. 2004; Chen et al.2004, 2005; Maier et al. 2004; Nachyar and Rajakumar2005; Matsumoto et al. 2010), but flavin-free azoreductaseswere also identified in Pigmentiphaga kullae K24 and
Xenophilus azovorans KF46F strains (Blumel et al. 2002;Blumel and Stolz 2003; Chen et al. 2010). The flavin-containing enzymes were shown to be involved in thedecolourisation of azo dyes in other substrates such as
0.000
0.024
0.048
0.072
0.096
40
0
0 10 20 30
1 2 3 4 50.00
0.12
0.24
0.36
0.48
0.60
0.72
A
B
1/V
0 (s
)1/
V0
(s)
1/[NADPH] (mM-1)
1/[RB5] (mM-1)
Fig. 7 Double-reciprocal plots of 1/V0 against: a 1/NADPH (blacksquare), 0.25 mM (white square), 0.5 mM (black circle), 1 mM(white circle), 2 mM of RB5 and b 1/RB5 (black circle), 0.03 mM(white circle) 0.04 mM (triangle), 0.06 mM (black up-pointingtriangle) 0.5 mM of NADPH. The parallel lines show the ping-pongbi–bi catalytic mechanism of PpAzoR. The kcat of the reaction is146.8 min−1, and the Km for NADPH and RB5 is 0.075 and 2.5 mM,respectively
0
20
40
60
80
100
120
0 100 200 300 400 500
[O2] (nmol/ml)
% A
ctiv
ity
Fig. 6 Inhibition of PpAzoR by dioxygen. Reactions were monitored byfollowing RB5 reduction. A half maximal inhibitory concentration (IC50)of 117.03 nmol/mL was determined by fitting the activity decay ofPpAzoR to a single first-order process
Substrate Vmax U mg−1 Km app mM kcat app min−1 kcat/Km M−1 s−1 Ki mM
Methyl red 1.0±0.04 0.4±0.1 61±4.3 2.5×103 –
Sudan orange G 0.2±0.03 0.1±0.07 8±1.3 1.4×103 0.13±0.04
Reactive yellow 81 3.0±0.31 3.7±1.1 175±11.7 1.1×103 –
Acid red 266 1.0±0.10 1.8±0.5 59±6.2 1.1×103 –
Mordant black 9 0.6±0.10 4.3±0.9 35±2.6 0.3×103 –
Acid orange 7 1.4±0.23 3.7±1.3 79±10.8 0.4×103 –
Acid black 194 1.3±0.18 1.8±0.6 78±7.1 0.7×103 –
Mordant black 17 1.0±0.10 0.9±0.1 58±7.4 1.1×103 –
Mordant black 3 0.6±0.04 0.7±0.08 35±1.8 0.8×103 –
Reactive black 5 2.2±0.10 1.4±0.2 129±3.4 1.4×103 –
Table 4 Steady-state apparentcatalytic constants of purifiedrecombinant PpAzoR for thereduction of structurally diverseazo dyes by using NADPH(0.25 mM) as the electron donor
Appl Microbiol Biotechnol

nitroaromatic substrates (Liu et al. 2007a) or azoic pro-drugs used in the treatment of the inflammatory boweldisease (Wang et al. 2007). Further study of the genescoding for these xenobiotic-metabolizing enzymes mayhelp to reveal factors governing the horizontal transfer ofgenetic information among bacterial species in the environ-ment. The only gene coding for an azoreductase in the P.putida genome was cloned and expressed in E. coli, and therecombinant enzyme was overproduced, purified, andcharacterised. The recombinant PpAzoR from P. putidaMET94 was shown to be a FMN-dependent homodimerwith a molecular mass around 40 kDa, as most of theflavin-dependent azoreductases that are either dimeric ortetrameric enzymes (Nakanishi et al. 2001; Chen et al.2004; Ito et al. 2006; Liu et al. 2007b). The biochemicaland kinetic properties of the recombinant enzyme indicatethat it is the main enzymatic system involved in thedecolourisation capacity of the P. putida cultures. PpAzoRis particularly unspecific with regard to the azo dyes that
can use as substrates when compared with those of othercharacterised bacterial azoreductases, even if the ratesmeasured for some dyes are relatively low (Bin et al. 2004;Chen et al. 2004, 2008; Wang et al. 2007; Liu et al. 2008). Itis noteworthy that the diazo reactive black 5, one of theoldest reactive dyes used with vast utilisation in the textiledyeing processes is reduced at a fast rate by PpAzoR. It wasshown that azo dyes are reduced by PpAzoR with bi–biping-pong (double displacement) kinetics as reported forother azoreductases (Liu et al. 2007b, 2008; Sollner et al.2007, 2009). This in accordance with the currently acceptedmechanism of azoreductases that requires two cycles ofNADPH-dependent reduction of FMN to FMNH2, reducingthe azo substrate to a hydrazine in the first cycle and thehydrazine to two amines in the second cycle (Bin et al. 2004;Ryan et al. 2010).
Microorganisms are exposed to a variety of toxic andantimicrobial compounds in the environment, which inducegeneral and specific stress response in the growing cells.Flavin-dependent azoreductases share strong similarities withregard to sequence, structure, and reaction mechanism withthe larger family of flavin-dependent quinone reductases thatincludes Lot6p from Saccharomyces cerevisiae and themammalian NQO1 (Deller et al. 2008). These enzymes areinvolved in the reduction of quinones, quinoneimines, azodyes, and nitro groups to protect the cells against the toxiceffects of free radicals and reactive oxygen species arisingfrom electron reductions. They are assumed to take part inthe organism’s enzymatic detoxification systems; e.g. theazoreductases from E. coli and B. subtilis were recentlyimplicated in the cellular response to thiol-specific stress(Towe et al. 2007; Leelakriangsak et al. 2008; Liu et al.2009) and Lot6p, the azoreductase homologue in S.cerevisiae has been implicated in the response to oxidativestress (Sollner et al. 2007, 2009). Furthermore, as additionalmembers of this family of enzymes are discovered, the list oftransformed substrates continues to grow. Evolutionarily,these enzymes may provide a selective advantage to bacteriaunder various conditions of environmental stress (Khersonskyand Tawfik 2010). The enzyme PpAzoR identified andcharacterised in the present study provides a rich opportunityto probe structure–function relationships that are determi-nants of substrate specificity and mechanisms of promiscuityamong this family of flavoproteins. Therefore, work is inprogress in our laboratory to understand the function of theenzyme in the bacterium, to fully characterise its kineticproperties, and to determine the crystallographic structure ofthis enzyme.
Acknowledgements We thank Diana Rocheta and Catarina Pimentafor their help in the preliminary studies. This work was supported by theFP6-NMP2-CT-2004-505899 (SOPHIED) and PTDC/BIO/72108/2006project grants. L. Pereira holds a post-doctoral fellowship (SFRH/BPD/20744/2004) from Fundação para a Ciência e Tecnologia.
HOOC
N=N
NH2
HOOC
0 5 10 15 20 25
1
2
Retention time (min)
NH2
30
N=N
OH
OH
3
4
0 5 10 15 20 25
Retention time (min)
OH
OH
NH2
30
A
B
N(CH3)2
N(CH3)2
H2N
Fig. 8 HPLC chromatograms of the reaction mixtures with methyl red(a) or Sudan orange G (b) after 24 h of reaction with (thin line) orwithout (thick line) PpAzoR. Products of the reaction were identified, incomparison to the standards: 2-aminobenzoic acid (1), N,N′-dimethyl-p-phenylenediamine (2), 4-aminoresorcinol (3), and aniline (4)
Appl Microbiol Biotechnol

References
Anjaneyulu Y, Chary N, Raj D (2005) Decolourization of industrialeffluents—available methods and emerging technologies—areview. Rev Environ Sci Bio/Technol 4:245–273
Bin Y, Jiti Z, Jing W, Cuihong D, Hongman H, Zhiyong S, YongmingB (2004) Expression and characteristics of the gene encodingazoreductase from Rhodobacter sphaeroides AS1.1737. FEMSMicrobiol Lett 236:129–136
Blumel S, Stolz A (2003) Cloning and characterization of the genecoding for the aerobic azoreductase from Pigmentiphaga kullaeK24. Appl Microbiol Biotechnol 62:186–190
Blumel S, Knackmuss HJ, Stolz A (2002) Molecular cloning andcharacterization of the gene coding for the aerobic azoreductasefrom Xenophilus azovorans KF46F. Appl Environ Microbiol68:3948–3955
Chen H (2006) Recent advances in azo dye degrading enzymeresearch. Curr Protein Pept Sci 7:101–111
Chen H, Wang RF, Cerniglia CE (2004) Molecular cloning, over-expression, purification, and characterization of an aerobic FMN-dependent azoreductase from Enterococcus faecalis. Protein ExprPurif 34:302–310
Chen H, Hopper SL, Cerniglia CE (2005) Biochemical and molecularcharacterization of an azoreductase from Staphylococcus aureus,a tetrameric NADPH-dependent flavoprotein. Microbiology151:1433–1441
Chen H, Xu H, Kweon O, Chen S, Cerniglia CE (2008) Functionalrole of Trp-105 of Enterococcus faecalis azoreductase (AzoA) asresolved by structural and mutational analysis. Microbiology154:2659–2667
Chen H, Feng J, Kweon O, Xu H, Cerniglia CE (2010) Identificationand molecular characterization of a novel flavin-free NADPHpreferred azoreductase encoded by azoB in Pigmentiphaga kullaeK24. BMC Biochem 11:13
de Lorenzo V (2001) Cleaning up behind us. The potential ofgenetically modified bacteria to break down toxic pollutants inthe environment. EMBO Rep 2:357–359
Deller S, Macheroux P, Sollner S (2008) Flavin-dependent quinonereductases. Cell Mol Life Sci 65:141–160
Hsueh CC, Chen BY (2007) Comparative study on reaction selectivityof azo dye decolorization by Pseudomonas luteola. J HazardMater 141:842–849
Husain Q (2006) Potential applications of the oxidoreductive enzymesin the decolorization and detoxification of textile and othersynthetic dyes from polluted water: a review. Crit Rev Biotechnol26:201–221
Ito K, Nakanishi M, Lee WC, Sasaki H, Zenno S, Saigo K, Kitade Y,Tanokura M (2006) Three-dimensional structure of AzoR fromEscherichia coli. An oxidereductase conserved in microorganisms.J Biol Chem 281:20567–20576
Kandelbauer A, Guebitz GM (2005) Bioremediation for the decolor-ization of textile dyes—a review. Env Chem 269–288
Khersonsky O, Tawfik DS (2010) Enzyme promiscuity: a mechanisticand evolutionary perspective. Annu Rev Biochem 79:471–505
Leelakriangsak M, Huyen NT, Towe S, van Duy N, Becher D, HeckerM, Antelmann H, Zuber P (2008) Regulation of quinonedetoxification by the thiol stress sensing DUF24/MarR-likerepressor, YodB in Bacillus subtilis. Mol Microbiol 67:1108–1124
Lewis JA, Escalante-Semerena JC (2006) The FAD-dependenttricarballylate dehydrogenase (TcuA) enzyme of Salmonellaenterica converts tricarballylate into cis-aconitate. J Bacteriol188:5479–5486
Liu G, Zhou J, Lv H, Xiang X, Wang J, Zhou M, Qu Y (2007a)Azoreductase from Rhodobacter sphaeroides AS1.1737 is a
flavodoxin that also functions as nitroreductase and flavin mono-nucleotide reductase. Appl Microbiol Biotechnol 76:1271–1279
Liu ZJ, Chen H, Shaw N, Hopper SL, Chen L, Chen S, Cerniglia CE,Wang BC (2007b) Crystal structure of an aerobic FMN-dependentazoreductase (AzoA) from Enterococcus faecalis. Arch BiochemBiophys 463:68–77
Liu G, Zhou J,Wang J, Yan B, Li J, Lu H, QuY, Jin R (2008) Site-directedmutagenesis of substrate binding sites of azoreductase fromRhodobacter sphaeroides. Biotechnol Lett 30:869–875
Liu G, Zhou J, Fu Q, Wang J (2009) The Escherichia coli AzoR isinvolved in resistance to thiol-specific stress caused by electrophilicquinones. J Bacteriol 191:6394–6400
Loh KC, Cao B (2008) Paradigm in biodegradation using Pseudomonasputida—a review of proteomics studies. Enzym Microb Technol42:1–12
Maier J, Kandelbauer A, Erlacher A, Cavaco-Paulo A, Gubitz GM(2004) A new alkali-thermostable azoreductase from Bacillus sp.strain SF. Appl Environ Microbiol 70:837–844
Mansour H, Corroler D, Barillier D, Ghedira K, Chekir L, Mosrati R(2007) Evaluation of genotoxicity and pro-oxidant effect of theazo dyes: acids yellow 17, violet 7, and orange 52, and of theirdegradation products by Pseudomonas putida mt-2. Food ChemToxicol 45:1670–1677
Mansour H, Mosrati R, Corroler D, Ghedira K, Barillier D, Chekir L(2009) In vitro mutagenicity of Acid Violet 7 and its degradationproducts by Pseudomonas putida mt-2: correlation with chemicalstructures. Environ Toxicol Pharmacol 27:231–236
Matsumoto K, Mukai Y, Ogata D, Shozui F, Nduko J, Taguchi S, OoiT (2010) Characterization of thermostable FMN-dependentNADH azoreductase from the moderate thermophile Geobacillusstearothermophilus. Appl Microbiol Biotechnol 86:1431–1438
Moutaouakkil A, Zeroual Y, Zohra Dzayri F, Talbi M, Lee K, BlaghenM (2003) Purification and partial characterization of azoreductasefrom Enterobacter agglomerans. Arch Biochem Biophys413:139–146
Nachyar C, Rajakumar G (2005) Purification and characterization of anoxygen insensitive azoreductase from Pseudomonas aeruginosa.Enz Microb Tecnhol 36:505–509
Nakanishi M, Yatome C, Ishida N, Kitade Y (2001) Putative ACPphosphodiesterase gene (acpD) encodes an azoreductase. J BiolChem 276:46394–46399
Nelson KE, Weinel C, Paulsen IT, Dodson RJ, Hilbert H, Martins dosSantos VA, Fouts DE, Gill SR, Pop M, Holmes M, Brinkac L,Beanan M, DeBoy RT, Daugherty S, Kolonay J, Madupu R,Nelson W, White O, Peterson J, Khouri H, Hance I, Lee P,Holtzapple E, Scanlan D, Tran K, Moazzez A, Utterback T, RizzoM, Lee K, Kosack D, Moestl D, Wedler H, Lauber J, StjepandicD, Hoheisel J, Straetz M, Heim S, Kiewitz C, Eisen JA, TimmisKN, Dusterhoft A, Tummler B, Fraser CM (2002) Completegenome sequence and comparative analysis of the metabolicallyversatile Pseudomonas putida KT2440. Environ Microbiol4:799–808
Pereira L, Coelho AV, Viegas CA, Ganachaud C, Lacazio G, Tron T,Robalo MP, Martins LO (2009a) On the mechanism ofbiotransformation of the anthraquinonic dye acid blue 62 bylaccases. Adv Synth Catal 351:1857–1865
Pereira L, Coelho AV, Viegas CA, Santos MM, Robalo MP, Martins LO(2009b) Enzymatic biotransformation of the azo dye Sudanorange G with bacterial CotA-laccase. J Biotechnol 139:68–77
Prigione V, Tigini V, Pezzella C, Anastasi A, Sannia G, Varese G(2008) Decolourisation and detoxification of textile effluents byfungal biosorption. Water Res 42:2911–2920
Rai HS, Bhattacharyya MS, Singh J, Vats P, Banerjee UC (2005)Removal of dyes from the effluent of textile and dyestuffmanufacturing industry: a review of emerging techniques with
Appl Microbiol Biotechnol

reference to biological treatment. Critical Rev Environ Sc Tech35:219–238
Rodriguez-Couto S (2009) Enzymatic biotransformation of syntheticdyes. Curr Drug Metab 10:1048–1054
Ryan A, Laurieri N, Westwood I, Wang CJ, Lowe E, Sim E (2010) Anovel mechanism for azoreduction. J Mol Biol 400:24–37
Sollner S, Nebauer R, Ehammer H, Prem A, Deller S, Palfey BA,Daum G, Macheroux P (2007) Lot6p from Saccharomycescerevisiae is a FMN-dependent reductase with a potential rolein quinone detoxification. FEBS J 274:1328–1339
Sollner S, Deller S, Macheroux P, Palfey BA (2009) Mechanism offlavin reduction and oxidation in the redox-sensing quinonereductase Lot6p from Saccharomyces cerevisiae. Biochemistry48:8636–8643
Suzuki Y, Yoda T, Ruhul A, Sugiura W (2001) Molecular cloning andcharacterization of the gene coding for azoreductase from Bacillussp. OY1-2 isolated from soil. J Biol Chem 276:9059–9065
Timmis KN, Pieper DH (1999) Bacteria designed for bioremediation.Trends Biotechnol 17:200–204
Towe S, Leelakriangsak M, Kobayashi K, Van Duy N, Hecker M,Zuber P, Antelmann H (2007) The MarR-type repressor MhqR(YkvE) regulates multiple dioxygenases/glyoxalases and anazoreductase which confer resistance to 2- methylhydroquinoneand catechol in Bacillus subtilis. Mol Microbiol 66:40–54
van der Zee FP, Villaverde S (2005) Combined anaerobic–aerobictreatment of azo dyes—a short review of bioreactor studies.Water Res 39:1425–1440
WangCJ, Hagemeier C, RahmanN, Lowe E, NobleM, CoughtrieM, SimE, Westwood I (2007) Molecular cloning, characterization andligand-bound structure of an azoreductase from Pseudomonasaeruginosa. J Mol Biol 373:1213–1228
Zimmermann T, Kulla HG, Leisinger T (1982) Properties of purifiedOrange II azoreductase, the enzyme initiating azo dye degradationby Pseudomonas KF46. Eur J Biochem 129:197–203
Appl Microbiol Biotechnol
Related Documents











