Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140 Molecular cloning of P450 aromatase from the leopard gecko and its expression in the ovary Daisuke Endo, Min Kyun Park ∗ Department of Biological Sciences, Graduate School of Sciences, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo 113-0033, Japan Received 19 November 2004; accepted 7 February 2005 Abstract In this study, we identified the cDNA of P450 aromatase in the leopard gecko, a lizard with temperature-dependent sex determination. The cDNA encodes a putative protein of 505 amino acids. The deduced amino acid sequence of leopard gecko aromatase cDNA showed 80% identity with that of turtles, 70% with humans and 77% with chickens. This is the first report of the identification of P450 aromatase cDNA in squamata species. It has been reported that this gene is expressed in different layers of cells in the ovary of mammalian species and avian species. Thus, we also investigated cells expressing the mRNA of this gene in the ovary of the leopard gecko by RT-PCR and in situ hybridization. The mRNA expression of leopard gecko P450 aromatase was localized in both the thecal and granulosa cell layers in the ovary. The expression in thecal and granulosa cell layers was examined in the largest follicle, second largest follicle and third largest follicle by RT-PCR. A higher level of mRNA expression was observed in the granulosa cell layer of the second largest follicle than in other cell layers. This result may reflect the characteristics of follicles in species with automonochronic ovulation. © 2005 Elsevier Ltd. All rights reserved. Keywords: Hybridization; Molecular cloning; RT-PCR; Ovulation; P450 aromatase; Reptiles; Squamate 1. Introduction P450 aromatase, a member of the cytochrome P450 su- perfamily, catalyzed the conversion of androgen to estrogen [1]. Estrogen plays a central role in many reproductive phe- nomena, such as sex determination, sexual differentiation [2], sexual behavior [3], and vitellogenesis [4]. The conversion of androgen to estrogen catalyzed by P450 aromatase is a rate- limiting step in estrogen synthesis. Thus, the regulation of the expression of P450 aromatase is important to consider the regulation of estrogen production. The cDNAs encoding P450 aromatase were cloned from many vertebrate species, such as humans [5], rats [6], mice [7], chickens [8], zebra finches [9], African clawed frogs [10], and zebrafish [11]. In reptiles, cDNAs were cloned from American alligators [12] and red-eared slider turtles [13] to investigate the mechanisms of temperature-dependent sex determination. However, little attention had been paid to the unique position of reptiles in ∗ Corresponding author. Tel.: +81 3 5841 4437; fax: +81 3 5841 4439. E-mail address: [email protected] (M.K. Park). vertebrates to consider the evolution of P450 aromatase. Rep- tilia are the only animal class belonging to both ectotherms and amniotes, and Aves are evolved from reptiles by one or- der. P450 aromatase is expressed in the granulosa cell layer of most mammalian species, such as humans [14], mice, rats [15], and brushtail possums [16], although in the ovary of pigs, it was expressed in not only the granulosa cell layer but also the theca cell layer [17], and in that of the tree shrew, it was expressed only in the thecal cell layer [18]. Unlike most mammalian species, P450 aromatase is expressed in the the- cal cell layer of the chicken ovary [19]. It is interesting to identify the cells that express P450 aromatase in the ovary of reptilian species, the same amniotes as in mammals and birds. The importance of reptiles is not only due to their unique position in vertebrate evolution but also their diverse repro- ductive phenomena. Particularly in squamata species, there are different types of sex determination, chromosomal sex determination and temperature-dependent sex determination [20], and different modes of reproduction, viviparity and oviparity [21], in one family. This diversity makes the molec- 0960-0760/$ – see front matter © 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jsbmb.2005.02.015

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
Molecular cloning of P450 aromatase from the leopardgecko and its expression in the ovary
Daisuke Endo, Min Kyun Park∗
Department of Biological Sciences, Graduate School of Sciences, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo 113-0033, Japan
Received 19 November 2004; accepted 7 February 2005
Abstract
In this study, we identified the cDNA of P450 aromatase in the leopard gecko, a lizard with temperature-dependent sex determination.The cDNA encodes a putative protein of 505 amino acids. The deduced amino acid sequence of leopard gecko aromatase cDNA showed80% identity with that of turtles, 70% with humans and 77% with chickens. This is the first report of the identification of P450 aromatasecDNA in squamata species. It has been reported that this gene is expressed in different layers of cells in the ovary of mammalian species andavian species. Thus, we also investigated cells expressing the mRNA of this gene in the ovary of the leopard gecko by RT-PCR and in situhybridization. The mRNA expression of leopard gecko P450 aromatase was localized in both the thecal and granulosa cell layers in the ovary.T follicle byR ell layers.T©
K
1
p[nsalttPsfiraoa
. Rep-rmse or-l layer
fr but, itte the-
varys and
quepro-therel sexationndc-
0d
he expression in thecal and granulosa cell layers was examined in the largest follicle, second largest follicle and third largestT-PCR. A higher level of mRNA expression was observed in the granulosa cell layer of the second largest follicle than in other chis result may reflect the characteristics of follicles in species with automonochronic ovulation.2005 Elsevier Ltd. All rights reserved.
eywords:Hybridization; Molecular cloning; RT-PCR; Ovulation; P450 aromatase; Reptiles; Squamate
. Introduction
P450 aromatase, a member of the cytochrome P450 su-erfamily, catalyzed the conversion of androgen to estrogen
1]. Estrogen plays a central role in many reproductive phe-omena, such as sex determination, sexual differentiation[2],exual behavior[3], and vitellogenesis[4]. The conversion ofndrogen to estrogen catalyzed by P450 aromatase is a rate-
imiting step in estrogen synthesis. Thus, the regulation ofhe expression of P450 aromatase is important to considerhe regulation of estrogen production. The cDNAs encoding450 aromatase were cloned from many vertebrate species,uch as humans[5], rats[6], mice [7], chickens[8], zebranches[9], African clawed frogs[10], and zebrafish[11]. Ineptiles, cDNAs were cloned from American alligators[12]nd red-eared slider turtles[13] to investigate the mechanismsf temperature-dependent sex determination. However, littlettention had been paid to the unique position of reptiles in
∗ Corresponding author. Tel.: +81 3 5841 4437; fax: +81 3 5841 4439.E-mail address:[email protected] (M.K. Park).
vertebrates to consider the evolution of P450 aromatasetilia are the only animal class belonging to both ectotheand amniotes, and Aves are evolved from reptiles by onder. P450 aromatase is expressed in the granulosa celof most mammalian species, such as humans[14], mice, rats[15], and brushtail possums[16], although in the ovary opigs, it was expressed in not only the granulosa cell layealso the theca cell layer[17], and in that of the tree shrewwas expressed only in the thecal cell layer[18]. Unlike mosmammalian species, P450 aromatase is expressed in thcal cell layer of the chicken ovary[19]. It is interesting toidentify the cells that express P450 aromatase in the oof reptilian species, the same amniotes as in mammalbirds.
The importance of reptiles is not only due to their uniposition in vertebrate evolution but also their diverse reductive phenomena. Particularly in squamata species,are different types of sex determination, chromosomadetermination and temperature-dependent sex determin[20], and different modes of reproduction, viviparity aoviparity[21], in one family. This diversity makes the mole
960-0760/$ – see front matter © 2005 Elsevier Ltd. All rights reserved.oi:10.1016/j.jsbmb.2005.02.015

132 D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
ular mechanisms of reproduction of squamata species impor-tant and interesting, but there have been few studies abouttheir molecular mechanisms of reproduction and no cDNAencoding P450 aromatase has been cloned from squamataspecies.
In this study, we cloned cDNA encoding P450 aromatasefrom the leopard gecko,Eublepharis macularius. This ani-mal is easily tamed and bred in many countries as a pet. Theway to breed is well established and described in the manuals[22]. Many squamata species used as experimental animalswere difficult to breed and past studies on squamata dependedon the wild caught animals. In this animal, the incubationtemperature during embryonic development determines theirgonadal sex[23]. The leopard gecko can be used not in its ori-gin. The leopard gecko has also been described as a seasonalbreeder[22], and investigations into the effect of environmen-tal factors on the expression of sex steroid-hormone receptorsand proopiomelanocortin have been reported[24,25]. Theseresults suggest that the leopard gecko is a useful experimentalmodel to study the reproduction of squamata. This is the firstreport about the cloning and characterization of P450 aro-matase from squamata species. Using the nucleotid sequenceof leopard gecko P450 aromatase, sequential, phylogenic andexpressional analyses were conducted. As previously men-tioned, there have been reports indicating the difference in thecellular expression of P450 aromatase in the ovary betweenm tifica-t wasa
2
2
theB yo.A nthso tt pro-v ater,a alsww ) andk ing.T uct,l alea def-e wereq ena asa celll themf 2)a ary.
The excised follicle was held with forceps and a cut, about1 mm long, was made with a scalpel. Both sides of the cutwere pulled with forceps and one cell layer, thecal cell layer,was obtained. Again, a cut was made on the follicle withoutthecal cell layer and its contents, yolk, was made to flow out.The cell layer without yolk, granulosa cell layer, was washedin PBS (8 g/l NaCl, 0.2 g/l KCl, 1.15 g/l Na2HPO4, and 0.2 g/lKH2PO4) several times. By histological observation of thesections of these cell layers, it was checked whether thesecell layers were truly theca and granulosa cell layers (datanot shown).
2.2. RNA extraction and cDNA synthesis
Total RNA was extracted using ISOGEN (Nippon Gene,Tokyo, Japan). The ovary cDNA used for rapid amplifica-tion of cDNA ends (RACE; see Section2.3) was synthesizedfrom 3�g of total RNA using a SMART RACE cDNA Am-plification Kit (BD Biosciences Clontech, Palo Alto, CA),according to the manufacturer’s instructions. The cDNAsused as templates for RT-PCR were synthesized from 3�gof denatured total RNA using 100 pmol oligo(dT) primer and100 units of M-MLV reverse transcriptase (Promega, Madi-son, WI) in a 20�l reaction volume with incubation at 42◦Cfor 1.5 h.
2R
atsec ge-S gion.A ina0 eachd c-t3a tedb usinge tedu ,J inatorc sys-t
tasec e de-s to ob-t dw (seeS CRcfT bove.
wasc
ammalian species and avian species. Thus, the idenion of cells that express P450 aromatase in the ovarylso conducted by in situ hybridization and RT-PCR.
. Materials and methods
.1. Animals
Animals were treated according to the guidelines ofiological Science Committee at the University of Tokdult fertile male and female leopard geckos of 15 mof age were used. They were hatched at 29◦C and raised a
he same temperature in our laboratory. Crickets wereided as the main diet three times per week and shelter, wnd a calcium supplement were supplied ad libitum. Animere maintained on a 14:10 h light/dark cycle at 29◦C. Theyere anesthetized with sodium pentobarbital (50 mg/kgilled by rapid decapitation, followed by complete bleedhe tissues, the whole brain, pituitary gland, ovary, ovid
iver, kidney, heart, skin and adrenal gland from the femdult leopard gecko, and the whole brain, testis, vasrens and kidney from the male adult leopard gecko,uickly removed and frozen immediately in liquid nitrognd stored at−70◦C until used. The left ovary was takenwhole ovary with follicles. The theca and granulosa
ayers were isolated from the right ovary according toanuscript of past researches with chicken cells[26,27], as
ollows. The largest follicle (F1), second largest follicle (Fnd third largest follicle (F3) were excised from the ov
.3. Molecular cloning of P450 aromatse cDNA byT-PCR and RACE
RT-PCR was carried out to obtain partial P450 aromDNA from ovary cDNA using degenerate primers, deE01 and dege-AS01, deduced from a conserved rell of the following PCR amplifications were performed20�l reaction mixture containing each primer at 1�M,
.25 units of TaKaRa Ex Taq (TaKaRa, Shiga, Japan),NTP at 250�M and Ex Taq buffer (TaKaRa). The rea
ion conditions for RT-PCR were as follows: 94◦C for 5 min,5 cycles of 94◦C for 40 s, 58◦C for 30 s, 72◦C for 1 min,nd 72◦C for 7 min. The amplified products were separay electrophoresis in 1.5% agarose gel and visualizedthidium bromide staining. DNA fragments were extracsing a QIA quick Gel Extraction Kit (QIAGEN K.K., Tokyoapan) and directly sequenced using a drhodamine termycle sequencing FS Ready Reaction Kit (Applied Bioems, Tokyo, Japan).
After determining the partial sequence of P450 aromaDNA, sense and antisense gene-specific primers werigned based on the sequence. RACE was carried outain the complete sequence. 3′- and 5′-RACE was performeith SE01 and nested universal primer (NUP) of the kitection2.2), and NUP and AS01, respectively. Each Pondition was as follows: 94◦C for 5 min, 35 cycles of 94◦Cor 40 s, 64◦C for 30 s, 72◦C for 2 min, and 72◦C for 7 min.he amplified products were sequenced as described a
The sequence of the entire open reading frameonfirmed with gene-specific primers, 5′-UTR-SE01 and

D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140 133
3′-UTR-AS01, using the cDNAs of ovaries from three in-dividuals.
2.4. Comparison of the amino acid sequences of variousP450 aromatase
The CLUSTAL X program (version 1.81)[28] was down-loaded fromftp://ftp-igbmc.u-strasbg.fr/pub/ClustalX/andused with default settings to align the deduced amino acidsequences of P450 aromatase of the leopard gecko and otherspecies with each other. The amino acid identity was calcu-lated between entire ORFs using GeneDoc software (version2.6.002)[29].
2.5. Molecular phylogenic analysis
The amino acid sequences of the entire ORFs of P450 aro-matase in the leopard gecko and several species from all ver-tebrate classes were aligned using CLUSTAL X with defaultsettings. The alignment of the amino acid sequences was usedto generate the phylogenic tree, using the neighbor-joiningmethod[30]. Bootstrap values were calculated with 1000replications to estimate the robustness of internal branches.The GenBank accession numbers of P450 aromatase usedin the comparisons and phylogenic analysis are as follows:h ze-b reds 8;N sh-a 968;m
2
e ofe oleb enalg layer
of the follicles, and total RNA without reverse transcriptionwere amplified using specific primers for P450 aromatase,and�-actin (Table 1). The PCR conditions were as follows:94◦C for 5 min, 25 cycles of 94◦C for 40 s, 64◦C for 30 s,72◦C for 1 min, and 72◦C for 7 min. To detect the lower ex-pression of P450 aromatase mRNA, 35 cycles of the PCRamplification were also conducted using the same primerset with cDNA from various tissues and 30 cycles of PCRwere conducted with the cDNA from the follicles. The ampli-fied products were electrophoresed on 1.5% agarose gel andstained with ethidium bromide. No band was detected usingtotal RNA without reverse transcription under PCR condi-tions (Fig. 4) with the primer set for this expression analysis.The specificity of PCR was confirmed by sequence analysis.
2.7. RNA probe synthesis for in situ hybridization
The T7 phage RNA polymerase promoter was added tothe DNA fragment amplified from the ovary cDNA with theprimer set, SE01 and 3′-UTR-AS01, using the Lig’n ScribeKit (Ambion, Austin, Tx) according to the manufacturer’smanual. Using the adaptor-ligated DNA fragment as a tem-plate for PCR, the template for the antisense probe was am-plified with the primer set, SE01 and the adaptor primer ofthe kit. For the sense probe, the template was amplified withthe adaptor primer and 3′-UTR-AS01. The sense and anti-s ed byT g la-b ny).
2
asd lde-h with7 /lN %M ed
TO thesis
N Pos
d Fromd FromS FromA FromS FromA FromS FromA FromS FromA FromS FromA FromS From5 From3 From thesis
G or T.
uman X13589; mouse D00659; Norway rat M33986;ra finch L81143; American alligator AY029233; red-ealider turtle AF178949; African clawed frog AB03127ile tilapia-a AF472620; Nile tilapia-b AF295761; zebrafiAF183906; zebrafish-b AF183908; medaka-a D82edaka-b AY319970 and Atlantic stingray AF097513.
.6. Expression analysis
RT-PCR was performed to identify the possible sourcstrogen. Twenty-five nanograms of cDNA from the whrain, pituitary, ovary, oviduct, testis, vas deferens, adrland, liver, heart, lung, skin, theca layer and garanulosa
able 1ligonucleotide primers used for RACE, RT-PCR and RNA probe syn
ame Nucleotide sequencea
ege-SE01 5′-TAYTTYGAYGCNTGGCARGC-3′ege-AS01 5′-GGYTGRTANCKCATRCTYTC-3′E01 5′-AACCTGACATCTTCTTCAAGATTTC-3′S01 5′-GTCTCTGTCACCCATTACAGCTTG-3′E02 5′-CATGCCAAAGCTGAAAGTGGTGGAG-3′S02 5′-GGCAGTGGAAAGCTTTTGTCGTTTC-3′E03 5′-CAATTTTGAGAAAAGTGTCCCCCAC-3′S03 5′-GTCGGATTTCTTTCCAAAGAGCCG-3′E04 5-CCAGATCCAAAAAGGCAAAGGTC-3′S04 5′-CCCATCCAGAGAAACCTCCAGTGCG-3′E05 5′-CTGGTGTCTCTTCTCCAAAGATGC-3′S05 5′-CCGTACGTCTCGTTGTAGTAGTTGC-3′E06 5′-CCAAATACGTTGCTGACACAGGTG-3′′UTR-SE01 5′-TCTCCCTGGTCTGCCTGGGCAACA-3′′UTR-AS01 5′-GGATGGGATGACAGTTTTTCGTTGT-3′
a Abbreviations for degenerate nucleotides: Y, C or T; R, G or A; K,
ense RNA probes for P450 aromatase were synthesiz7 RNA polymerase (TaKaRa) with the templates and Dieling mix (Roche Applied Bioscience, Penzberg, Germa
.8. In situ hybridization
For in situ hybridization, the ovary of leopard gecko wissected with a cryostat and fixed in 3% paraformayde and 0.25% glutalaldehyde for 2 h, washed5% MtOH/25% PBST (8 g/l NaCl, 0.2 g/l KCl, 1.15 ga2HPO4, 0.2 g/l KH2PO4 and 0.1% Tween 20), 50tOH/50% PBST and 25% MtOH/75% PBST, follow
ition Usage
655 to 674 For degenerate PCR1084 to 1103 For degenerate PCR686 to 710 For 3′-RACE1009 to 1022 For 5′-RACE1047 to 1071 For sequencing and RNA probe synthesis728 to 744 For sequencing1248 to 1272 For sequencing410 to 433 For sequencing1352 to 1374 For sequencing218 to 242 For sequencing1380 to 1403 For sequencing179 to 203 For sequencing1645 to 1669 For sequencing−43 to−17 For sequencing and RT-PCR1570 to 1593 For sequencing, RT-PCR and RNA probe syn
N represents all four nucleotides.

134 D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
by bleaching with 6% hydrogen peroxidase in PBST. Thesections of ovary were then digested with proteinase K(10 mg/ml) and post-fixed in 3% paraformaldehyde and0.25% glutalaldehyde for 15 min. After post-fixation, itwas washed with pre-hybridization buffer (50% formalde-hyde, 5× SSC (150 mM NaCl, 15 mM C6H5O7Na3·2H2O),10 mg/ml yeast tRNA and 1% SDS) and pre-hybridized inthis buffer for 2 h at 70◦C. Dig-labeled RNA probes wereadded and allowed to incubate at 70◦C overnight. The samplewas washed with 50% formaldehyde, 5× SSC and 1% SDSand treated by RNase A (10 mg/ml in 0.5 M NaCl, 10 mMTris–HCl, pH 7.5, and 0.1% Tween 20) for 1 h. After washingwith 50% formamyde and 2× SSC three times, it was incu-bated in blocking solution (2% blocking reagent (BoehringerMannheim, Mannheim, Germany) in TTBS (8 g/l NaCl,0.2 g/l KCl, 2.5 mM Tris–HCl, pH7.6, and 0.1% Tween 20)
for 1 h. Alkaline phosphate-labeled anti-Dig Fab fragments(Boehringer Mannheim, 1:1000) were diluted and the sam-ple was incubated at 4◦C overnight. After this incubation,the sample was washed three times in TTBS for 1 h and thenequilibrated in detection solution (100 mM NaCl, 100 mMTris–HCl, pH 9.5, 50 mM MgCl2 and 0.1% Tween 20) for30 min. NBT/BCIP substrate was used for detection. Thereaction was stopped by 3% paraformaldehyde and 0.25%glutalaldehyde after the desired signal had appeared.
3. Results
3.1. Cloning of the leopard gecko P450 aromatase cDNA
We designed degenerate primers based on the sequenceof the highly conserved region. Using these primers, 437 bp
Fa
ig. 1. Nucleotide and deduced amino acid sequences of leopard gecko Pdenylation signal.
450 aromatase cDNA. Start and stop codons are shaded. There was no authentic poly

D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140 135
of the partial sequence was acquired. Based on this partialsequence, gene-specific primers for 3′- and 5′-RACE weredesigned. The bands acquired by RACE were sequenced di-rectly using the primers SE01, SE02, SE03, SE04, SE05,SE06, AS01, AS02, AS03, AS04 and AS05. The length ofthe leopard gecko P450 aromatase cDNA identified in thisstudy was 2256 bp, except the poly(A) tail with an open read-ing frame encoding a 505 amino acid protein. There was noauthentic polyadenylation signal in the 3′-untranslated region
(Fig. 1). The sequence of the cDNA clone was deposited inGenBank (accession no. AB185924).
3.2. Structure of the P450 aromatase and comparisonwith other vertebrate P450 aromatase
As shown inFig. 2, the deduced amino acid sequenceshowed high identity with other vertebrates: 80% with red-eared slider turtles, 80% with American alligators, 77% with
Fwg
ig. 2. Alignment of the predicted amino acid sequence of leopard gecko P4as compared to alligators, red-eared slider turtles, chickens, humans, miceecko P450 aromatase are shaded. Missing residues are shown by bars.
50 aromatase with different species. The leopard gecko P450 aromatase sequence, African clawed frogs and zebrafish P450 aromatase. Identical residues to leopard

136 D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
chickens, 70% with humans, 68% with mice, 67% withAfrican clawed frogs and 49% with zebrafish. P450 aro-matase has several functional regions, such as a membrane-binding region, helix region, Ozol’s peptides, aromatic re-gion and heme-binding region. The identities in these re-gions were higher than in the entire amino acid sequences,except for the membrane-binding region. In the membrane-binding region, there were many substitutions, but mostof the substituted residues were homologous with others.Therefore, the function of this domain is thought to beconserved.
3.3. Molecular phylogenic analysis
The phylogenic tree was calculated using an amino acidalignment of the P450 aromatase of 11 selected species. Theresult is shown inFig. 3. The P450 aromatases of reptiles(leopard gecko, red-eared slider turtle and American alliga-tor) and birds (chicken and zebra finch) were clustered to-gether and that of the leopard gecko branched first off thecluster.
3.4. Expression analysis
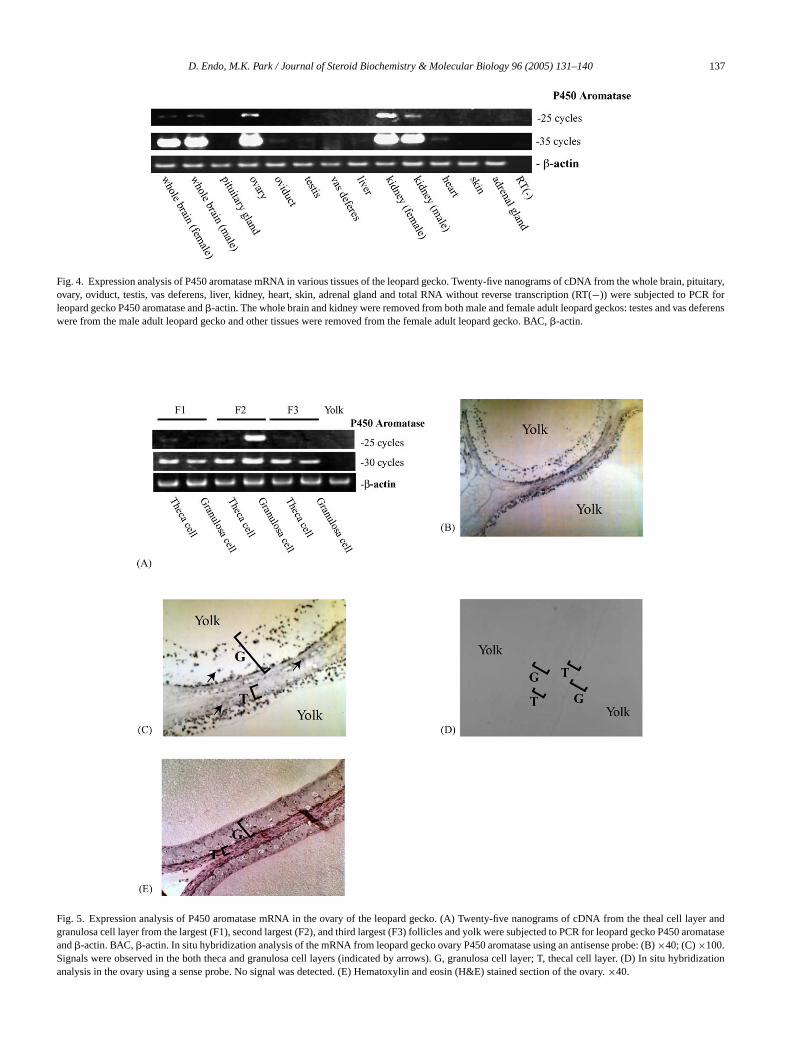
The tissue distribution of P450 aromatase was investigatedby RT-PCR using the primers 5′-UTR-SE01 and 3′-UTR-AS01. The genomic structure of this gene has not yet beencharacterized, but we found one intron (1.8 kb) between theprimers and no band was detected using the primer set withthe genomic DNA of the leopard gecko as a template (datanot shown). Therefore, the effect of genomic DNA contam-ination with cDNA could be excluded. With 25 PCR cycles,the expression in the whole brain and kidney of both male andfemale, and the ovary, was detected (Fig. 4). The expressionwas also observed in the oviduct, testis, vas deferens, liverand heart with 35 cycles of PCR (Fig. 4).
3.5. Expression in the ovary
Fig. 5E shows the hematoxylin–eosin-stained section ofthe ovary. There are follicles and each follicle had the layersof theca and granulosa cell.Fig. 5B and C shows the resultof in situ hybridization for the detection of P450 aromatase
Fb
ig. 3. Phylogenic tree of vertebrate P450 aromatase based on amino acidy the bootstrap procedure (1000 trials). For details on accession numbers i
homology. The numbers at the branches represent the confidence limits computedn the GenBank databases, see Section2.5.
D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140 137
Fig. 4. Expression analysis of P450 aromatase mRNA in various tissues of the leopard gecko. Twenty-five nanograms of cDNA from the whole brain, pituitary,ovary, oviduct, testis, vas deferens, liver, kidney, heart, skin, adrenal gland and total RNA without reverse transcription (RT(−)) were subjected to PCR forleopard gecko P450 aromatase and�-actin. The whole brain and kidney were removed from both male and female adult leopard geckos: testes and vas deferenswere from the male adult leopard gecko and other tissues were removed from the female adult leopard gecko. BAC,�-actin.
Fig. 5. Expression analysis of P450 aromatase mRNA in the ovary of the leopard gecko. (A) Twenty-five nanograms of cDNA from the theal cell layer andgranulosa cell layer from the largest (F1), second largest (F2), and third largest (F3) follicles and yolk were subjected to PCR for leopard gecko P450aromataseand�-actin. BAC,�-actin. In situ hybridization analysis of the mRNA from leopard gecko ovary P450 aromatase using an antisense probe: (B)×40; (C)×100.Signals were observed in the both theca and granulosa cell layers (indicated by arrows). G, granulosa cell layer; T, thecal cell layer. (D) In situ hybridizationanalysis in the ovary using a sense probe. No signal was detected. (E) Hematoxylin and eosin (H&E) stained section of the ovary.×40.

138 D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
mRNA expression in the F2 follicle. Strong signals were ob-served in the granulosa cell layer and weak signals in thethecal cell layer. By the sense probe, no signal was observedin both cell layers (Fig. 5D). The expressions were also de-tected by RT-PCR in the thecal and granulosa cell layers ofthree follicles examined (Fig. 5A). The F2 expression wasdetected clearly only in the granulosa cell layer by 25 cyclesof PCR, and in the thecal cell layer of F2 and other follicles,signals were detected by 30 cycles of PCR. This RT-PCRresult is consistent with that of in situ hybridization. No sig-nal was detected using the cDNA of yolk as a template. Thecontamination of yolk with the granulosa cell layer thereforehad no effect on the results.
4. Discussion
Squamata species shows diverse reproductive phenomena,such as the types of sex determination[20] and modes of re-production[21]. The molecular mechanisms underlying thisdiversity are very interesting, but little molecular informationhas been accumulated in squamata species. One reason forsuch a situation is the absence of model animal species insquamata. The leopard gecko is easily tamed, and is easy tobreed in the laboratory, unlike other squamata. Some genesrelated to reproduction, such as sex steroid-hormone recep-t -t edi lec-ur peri-m lonedt P450a manyrda ofP udiesi on.
aro-m ver-t mainst den-t ane-b ver-a 2thr P450a ce ofc icana ricanc de-p o onei es oft
se-q brate
phylum, the P450 aromatase amino acid sequences of rep-tiles and birds were clustered together and that of the leopardgecko branched off first (Fig. 3). This result is consistentwith the phylogenic trees made using other molecular mark-ers[35,36]. In these reports, the problem of whether turtles orcrocodilians are clustered with birds has not been concluded.In the phylogenic tree of this study, P450 aromatase of Amer-ican alligators was clustered with that of chickens and zebrafinches. However, there is only one species from each or-der of reptiles and two species from birds. More sequenceinformation is needed for further discussion.
With 25 cycles of RT-PCR, the mRNA expressions of leop-ard gecko P450 aromatase were detected at a high level in thewhole brain, ovary and kidney from both male and female.Weak signals were also detected in the oviduct, testis, vasdeferens, liver and heart with 35 cycles of RT-PCR (Fig. 4).Expressions in the brain and ovary have been observed in allspecies reported. In the European sea bass, it had been re-ported that P450 aromatase was expressed in the kidney andthe expression level was about 70-fold less than that of brain[37]. But, there is no report about the role of P450 aromatasein the kidney, so far as we know. Regardless of sex, the leop-ard gecko P450 aromatase mRNA was strongly expressedin the kidney. In the male leopard gecko, is the kidney oneof the major sources of circulating estrogen? Is the estrogenproduced in the kidney excreted with urine? These are veryi on int nr phe-n s alsoi eset tion,a
tedt s ex-pI gran-u -t tedt n theg layer[ rep-t l celll wase pardg am-m obidfi e iso np erentf res-s ulosac 450a toT flect
ors[31], gonadotropin-releasing hormone[32] and its recepor [33], proopiomelanocortin[25], have already been clonn this animal. Physiological experiments using this molar information have also been conducted[24,25]. Theseesults suggest that the leopard gecko will be a useful exental model. We therefore selected this species and c
he cDNA encoding leopard gecko P450 aromatase.romatase is an enzyme that plays an important role ineproductive phenomena, such as ovarian function[34], sexetermination, sex differentiation[2], sexual behavior[3],nd vitellogenesis[4]. The cloning and characterization450 aromatase in squamata species will facilitate st
nto the molecular mechanisms of squamata reproductiThe amino acid sequence of leopard gecko P450
atase showed high identity and similarity with otherebrate species. The amino acid sequences of five dohought to have an important function showed higher iity than the overall amino acid sequence (with no membrinding domain, but they showed higher similarity than oll sequences) (Fig. 2). One deletion was observed at the 1esidue of the amino acid sequence of leopard geckoromatase. This deletion was observed in the sequenhickens but not in that of other species, including Amerlligators, red-eared slider turtles, zebra finches and Aflawed frogs. Thus, this deletion is thought to occur inendently in leopard geckos and chickens. There is als
nsertion of glycine at the 328th residue in the sequenche leopard gecko, not in that of other species.
In the phylogenic tree drawn using the amino aciduences of P450 aromatase of species from all verte
nteresting questions for future research. The expressihe oviduct[38], testis[39], liver and heart[40] has beeeported in other species and its role in reproductiveomena has been suggested. In the leopard gecko, it i
nteresting to identify the regulation of the expression in thissues with specific reproductive status, such as gravitand reproductive season.
Both RT-PCR and in situ hybridization analyses indicahat the mRNA for leopard gecko P450 aromatase waressed in theca and granulosa cell layers in the ovary (Fig. 5).
t is reported that P450 aromatase was expressed in thelosa cells in mammalian species[14–16]with a few excep
ions[17,18]. In the ovary of chickens, however, it is reporhat immunoreactivity for P450 aromatase was absent iranulosa cell layer but was present in the theca cell
19]. It is interesting whether the P450 aromatase ofilian species was expressed in the granulosa or thecaayer. The results of this study showed that the mRNAxpressed in both types of cells in the ovary of the leoecko. This pattern of expression is not the same as in malian species or chickens. It is also reported that in the g
sh,Trimma okinawae, the expression of P450 aromatasbserved in both types of layers[41]. Thus, the expressioattern of leopard gecko P450 aromatase may be diff
rom that of fish. However, as mentioned above, the expion of P450 aromatase was not restricted in the granells of all mammalian species. In the porcine ovary, Promatase is expressed in both types of cells[17] and in thaf tree shrews, it is only expressed in the thecal cell layer[18].he results in chickens and leopard geckos may not re

D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140 139
the characteristics of birds and reptiles, respectively. Morestudies are needed in other species.
The existence of different expression levels between fol-licles was also suggested by RT-PCR. The expression wasdetected only in the granulosa cell layer from the secondlargest follicle with 25 cycles of PCR (Fig. 5A). The leop-ard gecko and most other geckos exhibit monoautochronicovulation[42,43]. They ovulate one egg from each ovary si-multaneously and have a clutch size of two. When femaleswere in the pre-vitellogenic phase in their estrous cycles, pre-vitellogenic follicles were arranged in a size hierarchy, andthe largest follicle began vitellogenic growth with the onsetof the vitellogenic phase. There is no study about how onefollicle is selected and becomes vitellogenic, and the differ-ence between the follicles. This study is the first to show thedifferent gene expressions between the follicles of reptiles.This is a useful first step to understand the mechanism ofmonoautochronic ovulation, and the analysis of the expres-sion of other genes related to reproduction will reveal thecharacteristics of follicles in different stages.
In conclusion, we isolated leopard gecko P450 aromataseand conducted sequential, phylogenic and expressional anal-ysis. This contributes not only to a comparative study for theevolution of P450 aromatase but also to an investigation intothe mechanisms of diverse reproduction in reptiles.
A
s.M rt-m nce,T gh-o
R
.M.her,sible
d atmor-.
sex--Horm.
en,
antify
1–
C.R.sertts ex-
pression in nonsteroidogenic cells, Proc. Natl. Acad. Sci. U.S.A. 85(23) (1988) 8948–8952.
[6] E.D. Lephart, K.G. Peterson, J.F. Noble, F.W. George, M.J. McPhaul,The structure of cDNA clones encoding the aromatase P-450 isolatedfrom a rat Leydig cell tumor line demonstrates differential processingof aromatase mRNA in rat ovary and a neoplastic cell line, Mol. Cell.Endocrinol. 70 (1) (1990) 31–40.
[7] M. Terashima, K. Toda, T. Kawamoto, I. Kuribayashi, Y. Ogawa, T.Maeda, Y. Shizuta, Isolation of a full-length cDNA encoding mousearomatase P450, Arch. Biochem. Biophys. 285 (2) (1991) 231–237.
[8] M.J. McPhaul, J.F. Noble, E.R. Simpson, C.R. Mendelson, J.D. Wil-son, The expression of a functional cDNA encoding the chicken cy-tochrome P-450arom (aromatase) that catalyzes the formation of es-trogen from androgen, J. Biol. Chem. 263 (31) (1988) 16358–16363.
[9] P. Shen, C.W. Campagnoni, K. Kampf, B.A. Schlinger, A.P. Arnold,A.T. Campagnoni, Isolation and characterization of a zebra finch aro-matase cDNA: in situ hybridization reveals high aromatase expres-sion in brain, Brain Res. Mol. Brain Res. 24 (1–4) (1994) 227–237.
[10] K. Miyashita, N. Shimizu, S. Osanai, S. Miyata, Sequence analysisand expression of the P450 aromatase and estrogen receptor genes inthe Xenopusovary, J. Steroid Biochem. Mol. Biol. 75 (2–3) (2000)101–107.
[11] S.K. Tong, E.F. Chiang, P.H. Hsiao, B. Chung, Phylogeny expressionand enzyme activity of zebrafish cyp19 (P450 aromatase) genes, J.Steroid Biochem. Mol. Biol. 79 (1–5) (2001) 299–303.
[12] W.N. Gabriel, B. Blumberg, S. Sutton, A.R. Place, V.A. Lance, Al-ligator aromatase cDNA sequence and its expression in embryosat male and female incubation temperatures, J. Exp. Zool. 290 (5)(2001) 439–448.
[13] C. Murdock, T. Wibbels, Cloning and expression of aromatase ina turtle with temperature-dependent sex determination, Gen. Comp.
[ lson,e, 17in the
[ Os-aro-989)
[ ido-
[ ofction(6)
[ oke,em-varyee)
[ enicentlop-
[ ing– 56.
[ ons,tegr.
[ ws,o,
[ cko28.
cknowledgements
We are grateful to Prof. Y. Oka, Dr. Y. Akazome, M. Kyokuwa, Mr. M. Enomoto and Mr. T. Ikemoto, Depaent of Biological Sciences, Graduate School of Sciehe University of Tokyo, for valuable discussions throuut this study.
eferences
[1] E.R. Simpson, M.S. Mahendroo, G.D. Means, M.W. Kilgore, MHinshelwood, S. Graham-Lorence, B. Amarneh, Y. Ito, C.R. FisM.D. Michael, Aromatase cytochrome P450, the enzyme responfor estrogen biosynthesis, Endocr. Rev. 15 (1994) 342–355.
[2] A. Tousignant, D. Crews, Effect of exogenous estradiol appliedifferent embryonic stages on sex determination, growth, andtality in the leopard gecko (Eublepharis macularius), J. Exp. Zool268 (1) (1994) 17–21.
[3] D.L. Flores, D. Crews, Effect of hormonal manipulation on socioual behavior in adult female leopard geckos (Eublepharis macularius), a species with temperature-dependent sex determination,Behav. 29 (4) (1995) 458–473.
[4] L.H. Herbst, L. Siconolfi-Baez, J.H. Torelli, P.A. Klein, M.J. KerbI.M. Schumacher, Induction of vitellogenesis by estradiol-17� anddevelopment of enzyme-linked immunosorbant assays to quplasma vitellogenin levels in green turtles (Chelonia mydas), Comp.Biochem. Physiol. B: Biochem. Mol. Biol. 135 (3) (2003) 55563.
[5] C.J. Corbin, S. Graham-Lorence, M. McPhaul, J.I. Mason,Mendelson, E.R. Simpson, Isolation of a full-length cDNA inencoding human aromatase system cytochrome P-450 and i
Endocrinol. 130 (2) (2003) 109–119.14] H. Sasano, M. Okamoto, J.I. Mason, E.R. Simpson, C.R. Mende
N. Sasano, S.G. Silverberg, Immunolocalization of aromatasalpha-hydroxylase and side-chain-cleavage cytochromes P-450human ovary, J. Reprod. Fertil. 85 (1) (1989) 163–169.
15] K. Ishimura, T. Yoshinaga-Hirabayashi, H. Tsuri, H. Fujita, Y.awa, Further immunocytochemical study on the localization ofmatase in the ovary of rats and mice, Histochemistry 90 (6) (1413–416.
16] L.J. Whale, D.C. Eckery, J.L. Juengel, Determination of sterogenic potential of ovarian cells of the brushtail possum (Trichosurusvulpecula), Biol. Reprod. 69 (3) (2003) 947–958.
17] E.M. Shores, M.G. Hunter, Immunohistochemical localizationsteroidogenic enzymes and comparison with hormone produduring follicle development in the pig, Reprod. Fertil. Dev. 11(1999) 337–344.
18] J. Kimura, A. Tsukise, G. Watanabe, K. Taya, W. RerkamnuaychH. Endo, M. Kurohmaru, J. Yamada, T. Nishida, Immunohistochical localization of inhibin and steroidogenic enzymes in the oof common tree shrew (Tupaia glis) and northern smooth-tailed trshrew (Dendrogale murina), Anat. Histol. Embryol. 29 (5) (2000267–271.
19] H. Nitta, Y. Osawa, J.M. Bahr, Immunolocalization of steroidogcells in small follicles of the chicken ovary: anatomical arrangemand location of steroidogenic cells change during follicular devement, Domest. Anim. Endocrinol. 8 (4) (1991) 587–594.
20] B.E. Viets, M.A. Ewert, L.G. Talent, C.E. Nelson, Sex-determinmechanisms in squamate reptiles, J. Exp. Zool. 270 (1994) 45
21] D.G. Blackburn, Reptilian viviparity: past research, future directiand appropriate models, Comp. Biochem. Physiol. A: Mol. InPhysiol. 127 (4) (2000) 391–409.
22] B.E. Viets, A. Tousignant, M.A. Ewert, C.E. Nelson, D. CreTemperature-dependent sex determination in the leopard geckEu-blepharis macularius, J. Exp. Zool. 265 (6) (1993) 679–683.
23] P. Vosjoli, B. Viets, R. Tremper, R. Klingenberg, The Leopard GeManual, Advanced Vivarium Systems, California, 1998, pp. 26–

140 D. Endo, M.K. Park / Journal of Steroid Biochemistry & Molecular Biology 96 (2005) 131–140
[24] D. Endo, M.K. Park, Quantification of three steroid hormone re-ceptors of the leopard gecko (Eublepharis macularius), a lizardwith temperature-dependent sex determination: their tissue distribu-tions and the effect of environmental change on their expressions,Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 136 (2003) 957–966.
[25] D. Endo, M.K. Park, Molecular characterization of the leopard geckoPOMC gene and expressional change in the testis by acclimationto low temperature and with a short photoperiod, Gen. Comp. En-docrinol. 138 (1) (2004) 70–77.
[26] E.S. Huang, A.V. Nalbandov, Steroidogenesis of chicken granulosaand theca cells: in vitro incubation system, Biol. Reprod. 20 (3)(1979) 442–453.
[27] A.B. Gilbert, A.J. Evans, M.M. Perry, M.H. Davidson, A methodfor separating the granulosa cells, the basal lamina and the thecaof the preovulatory ovarian follicle of the domestic fowl (Gallusdomesticus), J. Reprod. Fertil. 50 (1) (1977) 179–181.
[28] J.D. Thompson, T.J. Gibson, F. Plewniak, F. Jeanmouginm, D.G.Higgins, The CLUSTALX windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools, NucleicAcids Res. 25 (1997) 4876–4882.
[29] K.B. Nicholas, H.B. Nicholas Jr., Genedoc: a tool for editing andannotating multiple sequence alignments. Distributed by the author(1997).
[30] N. Saitou, M. Nei, The neighbor-joining method: a new methodfor reconstructing phylogenic trees, Mol. Biol. E 4 (1987) 406–425.
[31] T. Rhen, D. Crews, Distribution of androgen and estrogen receptormRNA in the brain and reproductive tissues of the leopard gecko,Eublepharis macularius, J. Comp. Neurol. 437 (4) (2001) 385–397.
[ rep-
.
[33] T. Ikemoto, M. Enomoto, M.K. Park, Identification and characteri-zation of a reptilian GnRH receptor from the leopard gecko, Mol.Cell. Endocrinol. 214 (1-2) (2004) 137–147.
[34] C.S. Rosenfeld, J.S. Wagner, R.M. Roberts, D.B. Lubahn, Intraovar-ian actions of oestrogen, Reproduction 122 (2) (2001) 215–226.
[35] S.B. Hedges, L.L. Poling, A molecular phylogeny of reptiles, Science283 (5404) (1999) 998–1001.
[36] Y. Cao, M.D. Sorenson, Y. Kumazawa, D.P. Mindell, M. Hasegawa,Phylogenic positionof turtles among amniotes: evidence from mito-chondrial and nuclear genes, Gene 259 (2000) 139–148.
[37] A. Gonzarez, F. Piferrer, Aromatase activity in the European seabass (Dicentrarchus lablaxL.) brain. Distribution and changes inrelation to age, sex, and the annual reproductive cycle, Gen. Comp.Endocrinol. 132 (2003) 223–230.
[38] F. Kobayashi, S.J. Zimniski, K.N. Smalley, Characterization ofoviductal aromatase in the northern leopard frog,Rana pipiens,Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 113 (3) (1996)653–657.
[39] S. Carreau, S. Bourguiba, S. Lambard, I. Galeraud-Denis, C. Genis-sel, J. Levallet, Reproductive system: aromatase and estrogens, Mol.Cell. Endocrinol. 193 (1–2) (2002) 137–143.
[40] E.R. Simpson, C. Clyne, G. Rubin, W.C. Boon, K. Robertson, K.Britt, C. Speed, M. Jones, Aromatase—a brief overview, Annu. Rev.Physiol. 64 (2002) 93–127.
[41] Y. Kobayashi, T. Kobayashi, M. Nakamura, T. Sunobe, C.E. Mor-rey, N. Suzuki, Y. Nagahama, Characterization of two types of cy-tochrome P450 aromatase in the serial-sex changing gobiid fish,Trimma okinawae, Zool. Sci. 21 (4) (2004) 417–425.
[42] R.E. Jones, C.H. Summers, Compensatory follicular hypertrophy dur-ing the ovarian cycle of the house gecko,Hemidactylus frenatus,Anat. Rec. 209 (1) (1984) 59–65.
[ hies:uppl.
32] T. Ikemoto, M.K. Park, Identification and characterization of thetilian GnRH-II gene in the leopard gecko,Eublepharis macularius,and its evolutionary considerations, Gene 316 (2003) 157–165
43] R.J. Etches, J.N. Petitte, Reptilian and avian follicular hierarcmodels for the study of ovarian development, J. Exp. Zool. S4 (1990) 112–122.
Related Documents











