Molecular Cloning of a Human, Hemicholinium-3- Sensitive Choline Transporter Subbu Apparsundaram,* , ² ,1 Shawn M. Ferguson,* , ² , ‡ Alfred L. George, Jr.,* , ² , § and Randy D. Blakely* , ² *Department of Pharmacology, ²Center for Molecular Neuroscience, ‡Graduate Neuroscience Program, and §Division of Genetic Medicine, Vanderbilt University School of Medicine, Nashville, Tennessee 37232-3420 Received August 23, 2000 Under many physiological circumstances, Na 1 - and Cl 2 -dependent, hemicholinium-3 (HC-3)-sensitive, high-affinity choline uptake (HACU) in cholinergic neurons is thought to be rate-limiting in the biosyn- thesis of acetylcholine (ACh). Based on sequence infor- mation provided by the Human Genome Project and the recently reported rat CHT1 (rCHT1), we cloned a human CHT cDNA from spinal cord. The hCHT cDNA encodes a protein of 580 amino acids having 93% iden- tity to rCHT1 and 51% identity to the Caenorhabditis elegans homolog CHO-1, and is distantly related to members of the Na 1 -coupled glucose transporter (SGLT) gene family of Na 1 -coupled glucose (SGLT), nucleoside and iodide transporters. Northern blot analysis reveals the expression of a ;5 kb transcript in human brain regions rich in cholinergic neurons in- cluding the putamen, spinal cord, and medulla. Ex- pression of hCHT cDNA in COS-7 cells results in satu- rable, Na 1 /Cl 2 -dependent choline uptake (K m 5 1.2 mM) in membrane vesicles and [ 3 H] HC-3 binding (K d 5 4 nM) in membrane fractions, consistent with character- istics reported in mammalian cholinergic neurons. Us- ing radiation hybrid mapping techniques, we localized the hCHT gene to human chromosome 2q12. These studies elucidate the primary structure and chromo- somal localization of hCHT and provide a basis for mechanistic analysis of HACU regulation and an in- vestigation of the role of hCHT in disease states. © 2000 Academic Press Key Words: acetylcholine; choline; cholinergic; chro- mosomal mapping; gene expression; hemicholinium-3; membrane protein; transporter; gene expression. In the central and peripheral nervous system, cho- linergic neurons regulate a variety of autonomic, cog- nitive and motor functions (1). Dysfunction of cholin- ergic signaling has been implicated in Alzheimer’s disease (2), Parkinson’s disease (3), schizophrenia (4), Huntington’s disease (5) and dysautonomia (6). Whereas multiple low-affinity choline transport (CHT) pathways exist to provide choline for metabolic needs and membrane phospholipid synthesis, cholinergic neurons are endowed with a distinct high-affinity cho- line uptake (HACU) process that provides choline for the synthesis of acetylcholine (ACh) (7, 8). Selective blockade of HACU by HC-3 can reduce ACh synthesis and release in vitro and impair cholinergic function in vivo (9, 10). The activity of HACU is also highly regu- lated by neuronal activity, neurotransmitters and sec- ond messengers (11, 12), (13–16) and therefore serves as a potential mechanism for the modulation of cholin- ergic function. HACU has been defined in the rat and human brain as a Na 1 /Cl 2 -dependent process with a K m of ;1 mM for choline (7, 17). Initial attempts to identify HACU at a molecular level led to the isolation and partial purifi- cation of membrane proteins of 35 to 90 kDa from Torpedo electromotor nerve terminals, as well as from insect and rat neural tissues (18 –21). Mayser et al. (22) reported the initial cloning of a choline transporter within this gene family; however, this molecule has since been redesignated as a creatine transporter based on substrate preference and regional distribu- tion (23, 24). More recently, O’Reagan et al. (25) uti- lized complementation of choline deficiencies in yeast to identify a novel choline transporter, CTL1. However, the distribution, ion-dependence and pharmacology of this transporter differ from that expected for presyn- aptic HACU. A major breakthrough in elucidating the molecular basis of HACU was achieved by Okuda and co-workers (26) who identified a choline transporter (CHO-1) expressed in the cholinergic neurons of the nematode C. elegans. These investigators also identi- fied a rat homolog (rCHT1) and found the expression pattern and functional characteristics of rCHT1 to be consistent with the HACU in the rat brain (26). Using 1 Present address: Department of Anatomy and Neurobiology, University of Kentucky Medical Center, Lexington, KY 40536-0298. Biochemical and Biophysical Research Communications 276, 862– 867 (2000) doi:10.1006/bbrc.2000.3561, available online at http://www.idealibrary.com on 862 0006-291X/21900 $35.00 Copyright © 2000 by Academic Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MS
SA*§
R
ahntmthetem(nahcpriniitssmvA
mm
ln
U
Biochemical and Biophysical Research Communications 276, 862–867 (2000)
doi:10.1006/bbrc.2000.3561, available online at http://www.idealibrary.com on
0CA
olecular Cloning of a Human, Hemicholinium-3-ensitive Choline Transporter
ubbu Apparsundaram,*,†,1 Shawn M. Ferguson,*,†,‡lfred L. George, Jr.,*,†,§ and Randy D. Blakely*,†
Department of Pharmacology, †Center for Molecular Neuroscience, ‡Graduate Neuroscience Program, andDivision of Genetic Medicine, Vanderbilt University School of Medicine, Nashville, Tennessee 37232-3420
eceived August 23, 2000
ergic signaling has been implicated in Alzheimer’sdHWpanltbavloae
acmcTirwsbtltttamc(nfipc
Under many physiological circumstances, Na1-nd Cl2-dependent, hemicholinium-3 (HC-3)-sensitive,igh-affinity choline uptake (HACU) in cholinergiceurons is thought to be rate-limiting in the biosyn-hesis of acetylcholine (ACh). Based on sequence infor-ation provided by the Human Genome Project and
he recently reported rat CHT1 (rCHT1), we cloned auman CHT cDNA from spinal cord. The hCHT cDNAncodes a protein of 580 amino acids having 93% iden-ity to rCHT1 and 51% identity to the Caenorhabditislegans homolog CHO-1, and is distantly related toembers of the Na1-coupled glucose transporter
SGLT) gene family of Na1-coupled glucose (SGLT),ucleoside and iodide transporters. Northern blotnalysis reveals the expression of a ;5 kb transcript inuman brain regions rich in cholinergic neurons in-luding the putamen, spinal cord, and medulla. Ex-ression of hCHT cDNA in COS-7 cells results in satu-able, Na1/Cl2-dependent choline uptake (Km 5 1.2 mM)n membrane vesicles and [3H] HC-3 binding (Kd 5 4M) in membrane fractions, consistent with character-
stics reported in mammalian cholinergic neurons. Us-ng radiation hybrid mapping techniques, we localizedhe hCHT gene to human chromosome 2q12. Thesetudies elucidate the primary structure and chromo-omal localization of hCHT and provide a basis forechanistic analysis of HACU regulation and an in-
estigation of the role of hCHT in disease states. © 2000
cademic Press
Key Words: acetylcholine; choline; cholinergic; chro-osomal mapping; gene expression; hemicholinium-3;embrane protein; transporter; gene expression.
In the central and peripheral nervous system, cho-inergic neurons regulate a variety of autonomic, cog-itive and motor functions (1). Dysfunction of cholin-
1 Present address: Department of Anatomy and Neurobiology,niversity of Kentucky Medical Center, Lexington, KY 40536-0298.
862006-291X/21900 $35.00opyright © 2000 by Academic Pressll rights of reproduction in any form reserved.
isease (2), Parkinson’s disease (3), schizophrenia (4),untington’s disease (5) and dysautonomia (6).hereas multiple low-affinity choline transport (CHT)
athways exist to provide choline for metabolic needsnd membrane phospholipid synthesis, cholinergiceurons are endowed with a distinct high-affinity cho-
ine uptake (HACU) process that provides choline forhe synthesis of acetylcholine (ACh) (7, 8). Selectivelockade of HACU by HC-3 can reduce ACh synthesisnd release in vitro and impair cholinergic function inivo (9, 10). The activity of HACU is also highly regu-ated by neuronal activity, neurotransmitters and sec-nd messengers (11, 12), (13–16) and therefore servess a potential mechanism for the modulation of cholin-rgic function.HACU has been defined in the rat and human brain
s a Na1/Cl2-dependent process with a Km of ;1 mM forholine (7, 17). Initial attempts to identify HACU at aolecular level led to the isolation and partial purifi-
ation of membrane proteins of 35 to 90 kDa fromorpedo electromotor nerve terminals, as well as from
nsect and rat neural tissues (18–21). Mayser et al. (22)eported the initial cloning of a choline transporterithin this gene family; however, this molecule has
ince been redesignated as a creatine transporterased on substrate preference and regional distribu-ion (23, 24). More recently, O’Reagan et al. (25) uti-ized complementation of choline deficiencies in yeasto identify a novel choline transporter, CTL1. However,he distribution, ion-dependence and pharmacology ofhis transporter differ from that expected for presyn-ptic HACU. A major breakthrough in elucidating theolecular basis of HACU was achieved by Okuda and
o-workers (26) who identified a choline transporterCHO-1) expressed in the cholinergic neurons of theematode C. elegans. These investigators also identi-ed a rat homolog (rCHT1) and found the expressionattern and functional characteristics of rCHT1 to beonsistent with the HACU in the rat brain (26). Using

the sequence information of rCHT1 and the informa-tcitschs
M
owAfp(tnTc3aaHbstMvlSgL
hBhbe11m01wc
tFmbfc1tm
C1wh1
ml of HTE buffer, assayed for protein content (Bradford Method;BfbHC1tptarwnbtpcb
vttfFpssrPReibtretdpwnps
hBfwC1RacRHTaeiN(3tIiaa
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ion provided by Human Genome Project, we haveloned a human ortholog hCHT and identified hybrid-zing mRNAs in the human brain. Transport and an-agonist binding studies in COS-7 cells reveal a sub-trate specificity, ion-dependence and pharmacologyonsistent with HACU. Finally, through radiation-ybrid analysis, we map the hCHT gene to chromo-ome 2q12.
ATERIALS AND METHODS
Cloning of hCHT cDNA. Based on the DNA sequence informationbtained from the Human Genome Sequence Database and rCHT1,e identified genomic sequences (GenBank Accession No.C009933) predictive of a human homolog for rCHT. Based on in-
erred coding exons, we designed nondegenerate oligonucleotiderimers to isolate an hCHT cDNA. The sense primer RB855ATAAAAATGG-CTTTCCATGTGGAAGGACTG) overlaps the puta-ive start codon and extends by 16 nucleotides on the 59 end and 11ucleotides on the 39 end. The antisense primer RB852 (TCACTG-AAATTATCTTCCAGTCCCAGACCC) overlaps the putative stopodon. hCHT cDNA was amplified by PCR (5 min at 95°C followed by5 cycles of 30 s at 95°C, 30 s at 60°C, and 4 min at 68°C and finally7 min extension at 68°C) using human spinal cord cDNA (Clontech)s template with Pfu Turbo DNA polymerase and the ProStar UltraF kit (Stratagene). The PCR amplified products were insertedetween the Not1 and Xho1 sites of pcDNA3 (Invitrogen) and theequence of the inserted cDNA was determined using fluorescent dyeerminators and an ABI310 automated DNA sequencer (Center forolecular Neuroscience and Neurogenomics Core, Vanderbilt Uni-
ersity). The nucleotide sequence of hCHT cDNA and inferred trans-ation can be obtained in GenBank with the Accession No. AF276871.equence alignments of hCHT, rCHT, and CHO-1 as well as otherene family members were performed with the MegAlign module ofasergene software (DNASTAR).
Northern analysis. We examined the regional distribution ofCHT mRNA using the human brain multiple tissue Northern Brainlot II panel (Clontech). A 463 bp cDNA probe corresponding toCHT bases 1221–1684 (Kpn1/Stu1 restriction fragment) was la-eled with by random priming (Prime-It II, Stratagene) in the pres-nce of [a-32P] dCTP (Amersham). Hybridization was carried out forh at 68°C in Expresshyb (Clontech) with a probe concentration of
.5 3 106 cpm/ml. The membrane was washed at high stringency (30in at 25°C in 23 SSC 0.05% SDS followed by 1 h at 68°C in
.13 SSC 0.1% SDS) and exposed to X-ray film (Kodak X-AR) for12 h before development. To validate evenness of loading, the blotas reprobed using a human b-actin cDNA (Clontech) probe at 106
pm/ml.
Transient expression of hCHT in COS-7 cells. COS-7 cells wereransfected with a pcDNA3 vector containing hCHT cDNA using theuGene 6 transfection reagent (Roche Diagnostics) following theanufacturer’s protocols. Briefly, COS-7 cells were cultured in Dul-
ecco’s modified Eagle medium (DMEM) supplemented with 10%etal bovine serum (Hyclone), 100 mg/ml of penicillin and streptomy-in at a density of 3–4 million cells in 150 mm culture dishes or 5 304 cells per well in 24-well dishes (Falcon). After 24 h, cells wereransfected and 48 h later, the cells were harvested and processed forembrane binding and whole cell or vesicle transport assays.
Radioligand binding assays. Monolayers of hCHT transfectedOS-7 cells were washed with 30 ml of HTE buffer and harvested in0 ml of homogenization buffer (0.32 M sucrose in HTE buffer). Cellsere homogenized at 20,000 rpm for 25 s using a Polytron tissueomogenizer (Brinkman). The homogenate was centrifuged at00,000g for 45 min at 4°C. The resulting pellet was suspended in 2
863
ioRad) and 50 mg of the membrane suspension (1 mg/ml) was trans-erred to each tube in triplicate. Unless otherwise indicated, allinding assays were performed in the presence of 150 mM NaCl inTE buffer. Assays were initiated by the addition of [3H]HC-3 (127i per mmol; NEN Life Science Products, 10 nM final conc). After ah incubation at room temperature, membranes were rapidly fil-
ered through glass fiber GF/B filters (Brandel) soaked in 0.3%olyethylenimine (Sigma) in HTE buffer and accumulated radioac-ivity was quantified using liquid scintillation spectrometry (Pack-rd). To test the ion dependence of hCHT binding activity, Na1 waseplaced in the assay buffer with equimolar concentrations of Li1
hereas Cl2 was replaced with equimolar concentrations of isethio-ate. Nonspecific binding was quantified by measuring [3H] HC-3inding in the presence of 1 mM unlabeled HC-3 and subtracted fromhe total counts to yield specific binding. Data for binding and trans-ort assays (see below) were analyzed using nonlinear least squaresurve fitting and Scatchard transformations (Kaleidagraph, Abel-eck).
Transport assays. To assay CHT activity in resealed membraneesicles from hCHT-transfected mammalian cells, we transientlyransfected COS-7 cells with hCHT and prepared membrane frac-ions as described for binding assays. After obtaining the membraneractions, resealed membrane vesicles were prepared as described byisher et al., 1992 (27) with slight modifications. The membraneellets were resuspended in 2 ml of HTE buffer (;2 mg/ml) and theuspension was rotated gently at 4°C for 45 min. Membrane vesicleuspensions were centrifuged (27,000g) for 20 min at 4°C and theesulting pellet suspended in 2 ml of Na1 and Cl2 free HTE buffer.rotein content of the suspension was determined (Bradford; Bio-ad) and 50 mg of the vesicle suspension (1 mg/ml) was transferred toach tube for triplicate assays. [3H]choline transport assays werenitiated by the addition of [3H]choline and NaCl diluted in HTEuffer to reach a final concentration of 20 nM and 150 mM, respec-ively. After a 5 min incubation at room temperature, vesicles wereapidly filtered through Whatman GF/B filters soaked in 0.3% poly-thylenimine in HTE buffer and the accumulated radioactivity quan-ified as described for binding assays. In experiments designed toetermine the ion-dependence of choline transport, Na1 was re-laced with equimolar concentrations of Li1 and Cl2 was replacedith equimolar concentrations of isethionate. In all experiments,onspecific uptake was quantified by determining uptake in theresence of 1 mM HC-3 and subtracted from the total uptake to yieldpecific transport.
Chromosomal localization. The chromosomal localization of theCHT gene was determined by radiation hybrid mapping (28).riefly, oligonucleotide primers were selected to amplify a 616 bp
ragment of genomic DNA corresponding to a region that overlapsith the hCHT1 stop codon. The sense primer RB885 (59-TGTGTATGGGCTCTGGTA CC-39) is complementary to bases202–1220 of the hCHT coding sequence. The antisense primerB934 (59-GCTGCATACCATCTCTCC-39) was designed based onnalysis of the genomic sequence immediately 39 from the hCHT stopodon (GenBank Accession No. AC009963). PCR mapping withB885 and RB934 was performed using the Stanford G3 Human/amster Radiation Hybrid panel (Research Genetics) as template.he PCR conditions were: 5 min at 95°C followed by 35 cycles of 30 st 95°C; 30 sec at 60°C, and 30 s at 72°C; and finally a 7 minxtension at 72°C PCR products were denatured for 30 min at 37°Cn a solution of 0.4 N NaOH/25 mM EDTA and blotted onto Hybond
nylon membrane (Amersham). The blot was UV cross-linkedStratalinker, Stratagene) and then baked at 80°C under vacuum for0 min and then hybridized with a 463-bp hCHT Kpn1/Stu1 restric-ion fragment (bp 1221–1684), labeled by random priming (Prime-ItI, Stratagene) in the presence of [a-32P] dCTP (Amersham). Hybrid-zation was performed for 1 h at 68°C in ExpressHyb (Clontech) with
probe concentration of 106 cpm/ml. The membrane was washed tofinal stringency of 0.13 SSC 0.1% SDS for 1 h at 68°C prior to X-ray

film development. Hybridization results were submitted for scoringop
R
sSOcAdqmpsho(ctcmocA
u(bptcpttermehTCNmppTmatairy
9
identity to the C. elegans gene product CHO-1. OutsideotNi(
fcAimgopTshaptmea(tpwDbp
heemtckTtat(lcp
HC[iaf[(
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
n the Stanford Human Genome Center (SHGC) G3 radiation hybridanel (http://shgc.stanford.edu/RH/index.html).
ESULTS AND DISCUSSION
Cloning of hCHT cDNA. We used the amino acidequence of rCHT1 and searched the Human Genomeequence Database for a potential human ortholog.ur GenBank BLAST search yielded bacterial artifi-
ial chromosome (BAC) sequences (AC009963 andC023672) containing unannotated sequences thatisplay 87% nucleotide identity to rCHT coding se-uence with conserved translational initiation and ter-ination sites. Using human spinal cord cDNA as tem-
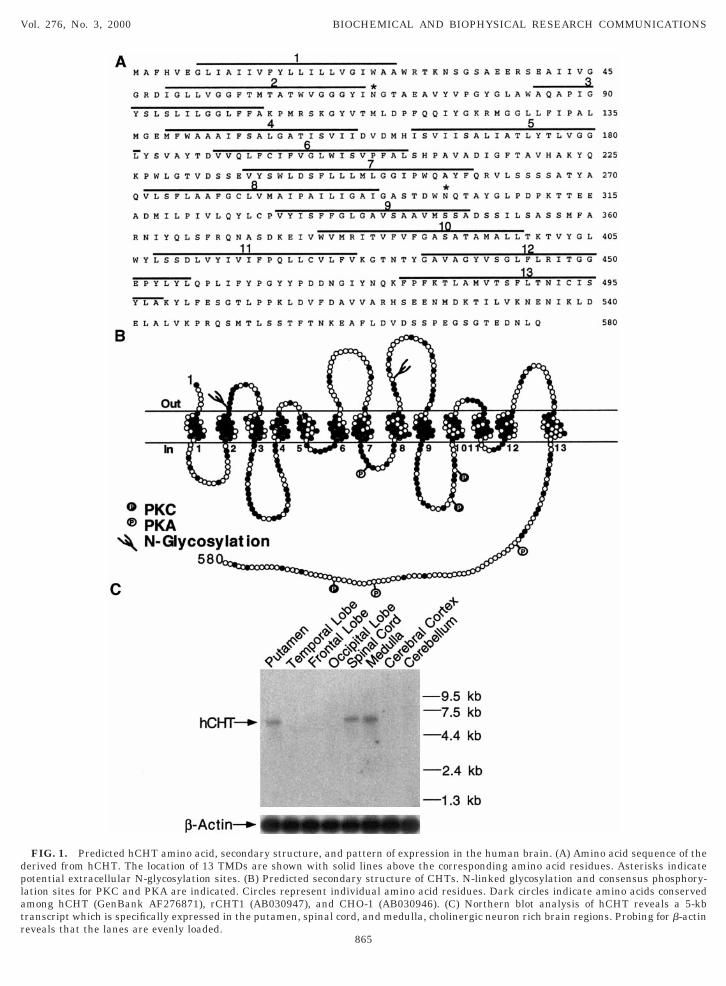
late, we amplified by PCR a 1780-bp product whoseequence matches the inferred exons of the putativeCHT gene found in the BAC sequences. The presencef an in-frame stop codon in the BAC sequenceAC009963) 9 nucleotides 59 of our predicted startodon supports the conclusion that hCHT protein ini-iates at the designated ATG. The hCHT cDNA en-odes a protein of 580 amino acids (Fig. 1A), a predictedass (Mr) of 63,203 Da and a calculated isoletric point
f 4.9. The hCHT nucleotide and amino acid sequencean be accessed at GenBank with Accession No.F276871.Sequence analysis of the predicted hCHT protein
sing Kyte-Doolittle (29), TopPred2 (30) and Signal P31) algorithms, predict the presence of 13 transmem-rane spanning domains (Figs. 1A and 1B) and theresence of a signal peptide that would orient the ini-ial 6 amino acids extracellularly. There are 3 canoni-al N-linked glycosylation residues; N69 and N301 areredicted to be localized extracellularly. It is notewor-hy that SGLT1 and NIS were also originally predictedo have 12 TMDs with NH2 and COOH terminus ori-nted towards the cytoplasm (32, 33). However, moreecent N-linked glycosylation site mutagenesis and im-unofluorescence studies reveal the presence of an
xtracellular NH2 terminus (33, 34). Recently, SGLT1as been predicted to contain 14 TMDs with the lastMD established by a hydrophobic domain at the veryOOH terminus (33). Unlike SGLT1, but more likeIS, hCHT lacks such a domain, resulting in cytoplas-ic COOH terminus (33, 34). Canonical sites for phos-
horylation by protein kinase C are present on theutative cytoplasmic domain between TMD9 andMD10 (S367 and S373) as well as in the COOH ter-inus (T558). Canonical sites for protein kinase A are
lso present between TMD7 and TMD8 (S263) and onhe COOH terminus (S522 and S550). Moreover, therere an additional 12 serine and 10 threonine residuesn predicted cytoplasmic domains of hCHT that mayepresent noncanonical sites for regulatory phosphor-lation.Sequence alignments reveal hCHT protein to exhibit
3% amino acid identity to rCHT1 and 51% amino acid
864
f putative species orthologs, hCHT is distantly relatedo the SLC5A family of solute carriers that includes thea1-coupled glucose transporter 1 (SGLT1; 25% AA
dentity) (33, 35) and the Na1/I2-iodide symporterNIS; 21% AA identity) (36).
Chromosomal localization. Altered cholinergicunction is a feature of multiple cognitive, neuromus-ular, motor and autonomic disorders (2–6, 37, 38).ssessment of the involvement of hCHT as a risk factor
n such syndromes first requires the high resolutionapping of the hCHT gene. Using human and hamster
enomic DNA, we defined PCR conditions such thatligonucleotide primers specifically amplify a 616-bproduct from human but not from hamster templates.his allowed us to assess the distribution of humanpecific amplicons on the SHGC radiation human/amster hybrid panel, with amplifications confirmeds hCHT-derived by hybridization with a hCHT cDNArobe. Our results reveal that the hCHT1 locus isightly linked to markers D2S340 and D2S176 on chro-osome 2. The proximity of hCHT to these two mark-
rs corresponds to a cytogenetic location of 2q12. Inddition, a search of BAC clone RP11-368F12AC009963) which contains the hCHT gene, revealshat this BAC also encodes a portion of the Ran bindingrotein 2 (RanBP2) gene. RanBP2 has been localizedith high resolution to 2q12 between D2S340 and2S1893 by a combination of fluorescence in situ hy-ridization (FISH) analysis, sequence analysis, andhysical mapping (39).
Northern analysis. To examine the distribution ofCHT mRNA in human brain, we performed a North-rn analysis using a 463-bp fragment comprising the 39nd of our hCHT1 cDNA. Our results show hCHTRNA to exhibit a restricted regional distribution in
he CNS, consistent with the distribution of cholinergicell bodies. Thus, we detected a single transcript of ;5b in the putamen, spinal cord, and medulla (Fig. 1C).his pattern of expression is consistent with the dis-ribution of cholinergic neurons including the large,spiny interneurons of the basal ganglia and the mo-oneurons of the midbrain, brainstem and spinal cord40–42). No hybridization was evident in temporalobe, frontal lobe, occipital lobe, cerebral cortex, anderebellum despite evidence of control hybridizationroducts with b-actin. (Fig. 1C).
Radioligand binding assays. The radiolabeledACU antagonist [3H]HC-3 has been used to identifyHT protein in human brain membranes (43). Using
3H]HC-3, we confirmed the presence of hCHT proteinn hCHT1-transfected COS-7 membranes (Figs. 2And 2B). As compared to vector-transfected cells, trans-ection of hCHT cDNA produced marked increase in3H]HC-3 binding in membrane fractions of COS-7 cellsdata not shown). The binding of [3H]HC-3 is saturable
dplatr
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Predicted hCHT amino acid, secondary structure, and paerived from hCHT. The location of 13 TMDs are shown with solidotential extracellular N-glycosylation sites. (B) Predicted secondaryation sites for PKC and PKA are indicated. Circles represent indivimong hCHT (GenBank AF276871), rCHT1 (AB030947), and CHOranscript which is specifically expressed in the putamen, spinal cord
ttern of expression in the human brain. (A) Amino acid sequence of thelines above the corresponding amino acid residues. Asterisks indicate
structure of CHTs. N-linked glycosylation and consensus phosphory-dual amino acid residues. Dark circles indicate amino acids conserved-1 (AB030946). (C) Northern blot analysis of hCHT reveals a 5-kb
, and medulla, cholinergic neuron rich brain regions. Probing for b-actineveals that the lanes are evenly loaded.
865

(iwRelTirchcT[r
c
transfected COS-7 cells demonstrated a modest in-cphcadrmrthpHtsa0e[cAfa
dcduph
A
efCN
R
1
d[stmCAt[mrsunoafS
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Fig. 2A) and Scatchard analyses of [3H]HC-3 bindingndicate single site kinetics (Hill coefficient n 5 0.99)ith an equilibrium dissociation constant (Kd) of 4 nM.eplacement of either Na1 or Cl2 with lithium or is-thionate, respectively, reduced [3H]HC-3 binding toevels seen in vector-transfected cells (data not shown).he low level of nonspecific [3H]HC-3 binding detected
n vector transfected cells is also insensitive to theemoval of either Na1 or Cl2. Unlabeled HC-3, andholine dose-dependently inhibit [3H] HC-3 binding inCHT transfected COS-7 cell membranes with Hilloefficients not significantly different from 1 (Fig. 2B).he Ki values for HC-3, choline, and ACh in inhibiting
3H]HC-3 binding were 4 nM, 21 mM and 433 mM,espectively.
Transport assays. Initial transport studies inRNA-injected Xenopus laevis oocytes and intact,
FIG. 2. [3H] HC-3 binding sites and [3H]choline uptake are evi-ent following transfection of hCHT1 in COS-7 cells. (A) Binding of
3H] HC-3 in membrane fractions of hCHT transfected COS-7 cells isaturable and has high-affinity. Binding obtained in membrane frac-ions from pcDNA3 transfected cells were subtracted to define CHT-ediated [3H] HC-3 binding. Inset: Scatchard transformation ofHT-mediated [3H] HC-3 binding. (B) Unlabeled HC-3, choline andch dose-dependently inhibit [3H] HC-3 binding in membrane frac-
ions of COS-7 cells. Inhibition assays were performed using 10 nM3H] HC-3. ACh was examined in the presence of 10 mM physostig-ine to inhibit cholinesterase activity. (C) Uptake of [3H]choline in
esealed membrane vesicles derived from transfected COS-7 cells isaturable and has high-affinity for choline. In all assays, [3H] cholineptake determined in the presence of 1 mM HC-3 is defined asonspecific uptake. Inset: Eadie-Hofstee-Scatchard transformationf CHT-mediated [3H] choline uptake. (D) Unlabeled HC-3, cholinend ACh dose-dependently inhibit [3H] HC-3 binding in membraneractions of COS-7 cells. In A–D, the results are presented as mean 6EM of triplicate experiments.
866
rease in [3H]choline uptake, with activity too low toermit extensive kinetic or pharmacologic studies. Weypothesized that intracellular retention of CHTs in aytoplasmic pool might be limiting choline transporterctivity measurements with intact cells. Therefore, weetermined CHT-mediated [3H]choline transport usingesealed membrane vesicles prepared from total cellembranes. In contrast to intact cell studies, we
eadily detected hCHT-mediated choline transport ac-ivity in resealed membrane vesicles prepared fromCHT transfected COS-7 cells but not in vesicles pre-ared from vector transfected controls and addition ofC-3 reduced [3H]choline uptake to levels seen in vec-
or transfected COS-7 membrane vesicles (data nothown). The specific uptake of [3H]choline is saturablend displays single-site kinetics (Hill coefficient n 5.99) with a Km of 1.6 mM (Fig. 2C). Replacement ofither Na1 with Li1 or Cl2 with isethionate reduced3H]choline uptake to levels seen in vector-transfectedells (data not shown). Unlabeled HC-3, choline andCh inhibited [3H]choline uptake in a dose-dependent
ashion (Fig. 2D), yielding Ki values of 5 nM, 123 nM,nd 570 nM, respectively.In summary, structural, distribution, and functional
ata provided in this report suggest that hCHT en-odes a protein responsible for human HACU. hCHT-irected antibodies (44) and gene probes should beseful in assessing molecular mechanisms that sup-ort choline transporter regulation and the role ofCHT in disease states.
CKNOWLEDGMENTS
We thank Scott Ramsey for his help in Xenopus laevis oocytexperiments. This work was supported by T32 HL07323 researchellowship to S.A., predoctoral support by the Vanderbilt Universityenter for Molecular Neuroscience to S.M.F, and Awards RO1S32387 to A.L.G. and RO1 MH58921 to R.D.B.
EFERENCES
1. Fibiger, H. C. (1991) Trends Neurosci. 14, 220–223.2. Coyle, J. T., Price, D. L., and DeLong, M. R. (1983) Science 219,
1184–1190.3. Calabresi, P., Centonze, D., Gubellini, P., Pisani, A., and Ber-
nardi, G. (2000) Trends Neurosci. 23, 120–126.4. Tandon, R. (1999) Br. J. Psychiatry Suppl. 37, 7–11.5. Lange, K. W., Javoy-Agid, F., Agid, Y., Jenner, P., and Marsden,
C. D. (1992) J. Neurol. 239, 103–104.6. Baron, R., and Engler, F. (1996) J. Neurol. 243, 18–24.7. Yamamura, H. I., and Snyder, S. H. (1972) Science 178, 626–
628.8. Blusztajn, J. K. (1998) Science 281, 794–795.9. Murrin, L. C., DeHaven, R. N., and Kuhar, M. J. (1977) J. Neu-
rochem. 29, 681–687.0. Guyenet, P., Lefresne, P., Rossier, J., Beaujouan, J. C., and
Glowinski, J. (1973) Mol. Pharmacol. 9, 630–639.

11. Atweh, S., Simon, J. R., and Kuhar, M. J. (1975) Life Sci. 17,
11
1
11
1
11
2
2
2
2
2
2
2
2
2
Lee, R., Maratukulam, A., O’Connor, K., Perkins, S., Piercy, M.,
233
333
3
33
3
3
4
4
4
4
4
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
1535–1544.2. Simon, J. R., and Kuhar, M. G. (1975) Nature 255, 162–163.3. Cancela, J. M., Bertrand, N., and Beley, A. (1995) Biochem.
Biophys. Res. Commun. 213, 944–949.4. Vogelsberg, V., Neff, N. H., and Hadjiconstantinou, M. (1997)
J. Neurochem. 68, 1062–1070.5. Breer, H., and Knipper, M. (1990) J. Neurobiol. 21, 269–275.6. Yamada, K., Saltarelli, M. D., and Coyle, J. T. (1991) Brain Res.
542, 132–134.7. Rylett, R. J., Ball, M. J., and Colhoun, E. H. (1983) Brain Res.
289, 169–175.8. Rylett, R. J. (1988) J. Neurochem. 51, 1942–1945.9. Knipper, M., Boekhoff, I., and Breer, H. (1989) Fed. Exp. Biol.
Sci. 245, 235–237.0. Knipper, M., Kahle, C., and Breer, H. (1991) Biochim. Biophys.
Acta 1065, 107–113.1. Rylett, R. J., Walters, S. A., and Davis, W. (1996) Brain Res. Mol.
Brain Res. 35, 354–358.2. Mayser, W., Schloss, P., and Betz, H. (1992) FEBS Lett. 305,
31–36.3. Guimbal, C., and Kilimann, M. W. (1993) J. Biol. Chem. 268,
8418–8421.4. Schloss, P., Mayser, W., and Betz, H. (1994) Biochem. Biophys.
Res. Commun. 198, 637–645.5. O’Regan, S., Traiffort, E., Ruat, M., Cha, N., Compaore, D., and
Meunier, F. M. (2000) Proc. Natl. Acad. Sci. USA 97, 1835–1840.6. Okuda, T., Haga, T., Kanai, Y., Endou, H., Ishihara, T., and
Katsura, I. (2000) Nat. Neurosci. 3, 120–125.7. Fisher, A. B., Dodia, C., Chander, A., and Kleinzeller, A. (1992)
Am. J. Physiol. 263, C1250–C1257.8. Stewart, E. A., McKusick, K. B., Aggarwal, A., Bajorek, E.,
Brady, S., Chu, A., Fang, N., Hadley, D., Harris, M., Hussain, S.,
867
Qin, F., Reif, T., Sanders, C., She, X., Sun, W. L., Tabar, P.,Voyticky, S., Cowles, S., Fan, J. B., Cox, D. R., et al. (1997)Genome Res. 7, 422–433.
9. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105–132.0. von Heijne, G. (1992) J. Mol. Biol. 225, 487–494.1. Nielsen, H., Engelbrecht, J., Brunak, S., and von Heijne, G.
(1997) Protein Eng. 10, 1–6.2. Dai, G., Levy, O., and Carrasco, N. (1996) Nature 379, 458–460.3. Turk, E., and Wright, E. M. (1997) J. Membr. Biol. 159, 1–20.4. Levy, O., De la Vieja, A., Ginter, C. S., Riedel, C., Dai, G., and
Carrasco, N. (1998) J. Biol. Chem. 273, 22657–22663.5. Hediger, M. A., Coady, M. J., Ikeda, T. S., and Wright, E. M.
(1987) Nature 330, 379–381.6. Dai, G., Levy, O., and Carrasco, N. (1996) Nature 379, 458–459.7. Rodriguez-Puertas, R., Pazos, A., and Pascual, J. (1994) Brain
Res. 636, 327–332.8. Alvarez, E., Ferrer, T., Perez-Conde, C., Lopez-Terradas, J. M.,
Perez-Jimenez, A., and Ramos, M. J. (1996) Neuropediatrics 27,26–31.
9. Nothwang, H. G., Rensing, C., Kubler, M., Denich, D., Brandl,B., Stubanus, M., Haaf, T., Kurnit, D., and Hildebrandt, F.(1998) Genomics 47, 383–392.
0. Kato, A. C., Touzeau, G., Bertrand, D., and Bader, C. R. (1985)J. Neurosci. 5, 2750–2761.
1. Mizukawa, K., McGeer, P. L., Tago, H., Peng, J. H., McGeer,E. G., and Kimura, H. (1986) Brain Res. 379, 39–55.
2. Mesulam, M. M., Mash, D., Hersh, L., Bothwell, M., and Geula,C. (1992) J. Comp. Neurol. 323, 252–268.
3. Pascual, J., Gonzalez, A. M., and Pazos, A. (1990) J. Neurochem.54, 792–800.
4. Apparsundaram, S., Ferguson, S., and Blakely, R. D. (2000) Soc.Neurosci. Abstr. 26, 15350.
Related Documents











