REGULAR PAPER Molecular cloning and characterization of plasma membrane- and vacuolar-type Na + /H + antiporters of an alkaline-salt-tolerant monocot, Puccinellia tenuiflora Shio Kobayashi • Natsuki Abe • Kaoru T. Yoshida • Shenkui Liu • Tetsuo Takano Received: 31 October 2011 / Accepted: 26 December 2011 Ó The Botanical Society of Japan and Springer 2012 Abstract A better understanding of salt tolerance in plants might lead to the genetic engineering of crops that can grow in saline soils. Here we cloned and characterized plasma membrane and vacuolar Na ? /H ? antiporters of a monocotyledonous alkaline-tolerant halophyte, Puccinellia tenuiflora. The predicted amino acid sequence of the transporters were very similar to those of orthologs in monocotyledonous crops. Expression analysis showed that (1) NHA was more strongly induced by NaCl in the roots of P. tenuiflora while in rice it was rather induced in the shoots, suggesting that the role of NHA in salt excretion from the roots partly accounts for the difference in the tolerance of the two species, and that (2) NHXs were spe- cifically induced by NaHCO 3 but not by NaCl in the roots of both species, suggesting that vacuolar-type Na ? /H ? antiporters play roles in pH regulation under alkaline salt conditions. Overexpression of the antiporters resulted in increased tolerance of shoots to NaCl and roots to NaHCO 3 . Overexpression lines exhibited a lower Na ? content and a higher K ? content in shoots under NaCl treatments, leading to a higher K ? /Na ? ratio. Keywords Na ? /H ? antiporter Puccinellia tenuiflora Saline-alkali stress Salt stress Introduction Soil salinity is one of the major environmental stresses faced by crops today. More than 6% of the world’s land area ( [ 800 million hectares) is adversely affected by salt (Munns and Tester 2008). In Northeast China, about 3.2 9 10 6 ha contain elevated levels of alkaline salt. In this area, soil pH is more than 9.8, and the characteristics of the soil are classified as ‘‘alkali’’ or ‘‘saline-alkali’’ (Wang et al. 2009b). Plants in this area suffer not only from salinity stress, but also from high pH. The former is mainly caused by Na ? , and the latter is an effect of HCO 3 - and CO 3 2- . These properties make it impossible for normal crops to grow, and salt-tolerant pasture plants and weeds are dominant in the area. One of these plants is a mono- cotyledonous graminaceous halophyte, Puccinellia tenu- iflora (Griseb.) Scrib. et Merr., called alkali grass in Chinese (Peng et al. 2004). Under saline conditions, P. tenuiflora exhibits a high K ? /Na ? ratio, resulting from the high K ? /Na ? selectivity of its plasma membrane (Peng et al. 2004) and a high ability to limit Na ? influx in the roots (Wang et al. 2009a). In addition, the leaves secrete salts with wax from the stomata (Guorong et al. 2005). EST Electronic supplementary material The online version of this article (doi:10.1007/s10265-012-0475-9) contains supplementary material, which is available to authorized users. S. Kobayashi N. Abe T. Takano (&) Asian Natural Environmental Science Center (ANESC), The University of Tokyo, 1-1-1 Midori-cho, Nishitokyo, Tokyo 188-0002, Japan e-mail: [email protected] S. Kobayashi e-mail: [email protected] N. Abe e-mail: [email protected] K. T. Yoshida Department of Ecosystem Studies, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tokyo 113-8657, Japan e-mail: [email protected] S. Liu Alkali Soil Natural Environmental Science Center, Northeast Forestry University, Harbin 150040, China e-mail: [email protected] 123 J Plant Res DOI 10.1007/s10265-012-0475-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR PAPER
Molecular cloning and characterization of plasma membrane-and vacuolar-type Na+/H+ antiporters of an alkaline-salt-tolerantmonocot, Puccinellia tenuiflora
Shio Kobayashi • Natsuki Abe • Kaoru T. Yoshida •
Shenkui Liu • Tetsuo Takano
Received: 31 October 2011 / Accepted: 26 December 2011
� The Botanical Society of Japan and Springer 2012
Abstract A better understanding of salt tolerance in
plants might lead to the genetic engineering of crops that
can grow in saline soils. Here we cloned and characterized
plasma membrane and vacuolar Na?/H? antiporters of a
monocotyledonous alkaline-tolerant halophyte, Puccinellia
tenuiflora. The predicted amino acid sequence of the
transporters were very similar to those of orthologs in
monocotyledonous crops. Expression analysis showed that
(1) NHA was more strongly induced by NaCl in the roots of
P. tenuiflora while in rice it was rather induced in the
shoots, suggesting that the role of NHA in salt excretion
from the roots partly accounts for the difference in the
tolerance of the two species, and that (2) NHXs were spe-
cifically induced by NaHCO3 but not by NaCl in the roots
of both species, suggesting that vacuolar-type Na?/H?
antiporters play roles in pH regulation under alkaline salt
conditions. Overexpression of the antiporters resulted in
increased tolerance of shoots to NaCl and roots to
NaHCO3. Overexpression lines exhibited a lower Na?
content and a higher K? content in shoots under NaCl
treatments, leading to a higher K?/Na? ratio.
Keywords Na?/H? antiporter � Puccinellia tenuiflora �Saline-alkali stress � Salt stress
Introduction
Soil salinity is one of the major environmental stresses
faced by crops today. More than 6% of the world’s land
area ([800 million hectares) is adversely affected by salt
(Munns and Tester 2008). In Northeast China, about
3.2 9 106 ha contain elevated levels of alkaline salt. In this
area, soil pH is more than 9.8, and the characteristics of the
soil are classified as ‘‘alkali’’ or ‘‘saline-alkali’’ (Wang
et al. 2009b). Plants in this area suffer not only from
salinity stress, but also from high pH. The former is mainly
caused by Na?, and the latter is an effect of HCO3- and
CO32-. These properties make it impossible for normal
crops to grow, and salt-tolerant pasture plants and weeds
are dominant in the area. One of these plants is a mono-
cotyledonous graminaceous halophyte, Puccinellia tenu-
iflora (Griseb.) Scrib. et Merr., called alkali grass in
Chinese (Peng et al. 2004). Under saline conditions, P.
tenuiflora exhibits a high K?/Na? ratio, resulting from the
high K?/Na? selectivity of its plasma membrane (Peng
et al. 2004) and a high ability to limit Na? influx in the
roots (Wang et al. 2009a). In addition, the leaves secrete
salts with wax from the stomata (Guorong et al. 2005). EST
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10265-012-0475-9) contains supplementarymaterial, which is available to authorized users.
S. Kobayashi � N. Abe � T. Takano (&)
Asian Natural Environmental Science Center (ANESC),
The University of Tokyo, 1-1-1 Midori-cho,
Nishitokyo, Tokyo 188-0002, Japan
e-mail: [email protected]
S. Kobayashi
e-mail: [email protected]
N. Abe
e-mail: [email protected]
K. T. Yoshida
Department of Ecosystem Studies, The University of Tokyo,
1-1-1 Yayoi, Bunkyo-ku, Tokyo 113-8657, Japan
e-mail: [email protected]
S. Liu
Alkali Soil Natural Environmental Science Center,
Northeast Forestry University, Harbin 150040, China
e-mail: [email protected]
123
J Plant Res
DOI 10.1007/s10265-012-0475-9

and microarray analyses of NaHCO3-stressed plants
revealed two major groups of genes whose expression is
changed by the treatment, suggesting that P. tenuiflora has
at least two stress tolerance pathways (Wang et al. 2007a,
b, c). Several P. tenuiflora genes have been cloned and
characterized and have been implicated in stress tolerance
(Ardie et al. 2009, 2010, 2011; Liu et al. 2009; Wang et al.
2011). Since P. tenuiflora is a monocot, an understanding
of its tolerance mechanisms may help to engineer salt
tolerant cereal crops.
Na? adversely affects plant growth in many ways.
Overaccumulation of Na? in the cytosol may lead to a
reduced K?/Na? ratio, dissipation of the membrane
potential, inhibition of enzyme activities, and production of
reactive oxygen species (Tuteja 2007). Plants avoid Na?
accumulation in the cells mainly by three mechanisms: (1)
restriction of Na? entry into the roots, (2) sequestration of
Na? into the vacuole, and (3) pumping Na? that entered
the cytosol back to the growth medium or to apoplastic
spaces (Apse and Blumwald 2007). Cation pumps, carriers
and channels participate in these processes. Sodium/proton
exchangers, which are a group of electroneutral monova-
lent cation/proton antiporters (CPAs) belonging to the
CPA1 family, play important roles in Na? extrusion and
sequestration (Rodrıguez-Rosales et al. 2009).
NHA (Na?/H? antiporter) or SOS1 (salt overly sensitive
1) is a plasma membrane-type antiporter, and participates in
both Na? extrusion at the roots and Na? loading into the
xylem (Shi et al. 2000, 2002). A plant NHA was first cloned
and characterized in a salt-sensitive Arabidopsis mutant, salt
overly sensitive 1. The gene (AtSOS1) was shown to be
responsible for the salt-sensitive phenotype (Shi et al. 2000;
Wu et al. 1996). The Arabidopsis overexpression line of
AtSOS1 was tolerant to Na? stress (Shi et al. 2003).
Orthologous genes were cloned from crops such as rice,
tomato, and quinoa (Martınez-Atienza et al. 2007; Maughan
et al. 2009; Olıas et al. 2009) and also from halophytes such
as Populus euphratica and reed grass, and more recently
from P. tenuiflora (Takahashi et al. 2009; Wang et al. 2011;
Wu et al. 2007).
The NHX (Na?/H? exchanger) is a vacuolar-type anti-
porter. In plants, it was first reported in Arabidopsis and
designated AtNHX1 (Apse 1999). Whereas plants usually
have only one copy of a plasma-membrane type NHA,
NHXs are normally found as multiple copies: Arabidopsis
has 6 copies (AtNHX1-6) and rice has 5. AtNHX1 and its
orthologs are the best characterized. The atnhx1 mutant
was sensitive to salt stress, and the overexpression line was
tolerant to salt stress (Apse 1999; Apse et al. 2003).
In this study, we focused on the Na?/H? antiporters
NHA and NHX1 in P. tenuiflora, hypothesizing that they
account for the high salt tolerance of the plant. Wang et al.
(2011) cloned a NHA of P. tenuiflora (PtNHA1) and
revealed that it was induced by NaCl, and that Arabidopsis
plants overexpressing PtNHA1 were more tolerant to NaCl
and had higher K?/Na? ratio under NaCl treatment than
WT. We cloned PtNHA1 independently, and also cloned a
cDNA which showed high sequence similarity with rice
NHX1, and named it PutNHX. We used rice homologs for
comparison, since rice, like P. tenuiflora, is a monocot. We
used not only NaCl but also alkaline salts to characterize
the Na?/H? antiporters, since P. tenuiflora grows in a
highly alkaline condition in the wild. The roles of PtNHA1
and PutNHX in neutral or alkaline salt tolerance of plants
were examined using rice overexpression lines.
Materials and methods
Plant materials, growth conditions, and stress
treatments
Seeds of P. tenuiflora were collected in an alkaline soil area
located in North-East China. Seeds were germinated in tap
water for 2 weeks and then transferred to nutrient solution.
Seeds of Oryza sativa L. cv. Nipponbare were harvested at
Institute for Sustainable Agro-ecosystem services, Univer-
sity of Tokyo, in Tokyo, Japan. The seeds were washed with
tap water and germinated in 2.5% PPMTM (Plant Cell
Technology, USA), and subsequently grown in nutrient
solution. Nutrient solution contained 6 mg/l (NH4)2SO4,
2 mg/l K2SO4, 8.2 mg/l MgSO4, 2.3 mg/l KNO3, 7.5 mg/l
Ca(NO3)2, 3.1 mg/l KH2PO4, 10 mg/l Fe-EDTA and the pH
was adjusted to 5.7 with 1 M KOH. Growth chamber was
maintained at 28�C during the day and 22�C at night while
the daily photoperiod of 350–400 lmol m-2 s-1 was 12 h.
NaCl or NaHCO3 were added to the nutrient solution for
stress treatment.
To compare salt stress tolerance, 2-week-old plants of P.
tenuiflora and 1-week-old rice plants were transferred to
nutrient solution containing 0, 100, 300, or 1,000 mM
NaCl or 0, 100, 300, 1,000 mM NaHCO3 and grown for
another 5 days.
Cloning of PtNHA1 and PutNHX
Total RNA was extracted using ISOGEN� (NIPPON
GENE, Japan) from the shoots and roots of P. tenuiflora
plants which were sown 10 days before sampling, accord-
ing to the manufacturer’s instructions. First strand cDNA
was synthesized from the total RNA using Oligo dT primer
and PrimeScript� Reverse Transcriptase (TaKaRa, Japan).
The partial cDNA fragments were amplified by PCR, using
primers designed based on the conserved regions of NHA
and NHX (Table S1) and the first strand cDNA as a
template. Full-length cDNA was obtained with 50- and
J Plant Res
123

30-RACE techniques. 50-Full RACE CORE Set (TaKaRa,
Japan) and Cap FishingTM 30-Full-Length cDNA Premix Kit
(Seegene, Korea) were used for 50- and 30-RACE, respec-
tively. The PCR product was subcloned and sequenced.
Primers used for full-length amplification and subcloning into
Gateway� vectors (Invitrogen, USA) are shown in Table S1.
Hydropathy analyses and domain analyses were performed
using the SOSUI program (http://bp.nuap.nagoya-u.ac.
jp/sosui/) and Pfam sequence search (http://pfam.sanger.
ac.uk/), respectively.
Real-time RT-PCR
For real-time RT-PCR, total RNA was extracted by the
guanidium isothiocyanate (GTC) method from roots and
shoots of 4-week-old P. tenuiflora or 2-week-old rice
subjected to 3 days of 300 mM NaCl or 300 mM NaHCO3
treatment. DNase-treated RNA was reverse transcribed
to cDNA using High Capacity RNA-to-cDNATM Kit
(Applied Biosystems, USA) following the manufacturer’s
instructions. The cDNA was diluted 10 times and 1 ll of
the diluted cDNA was used as the template for quantitative
RT-PCR analysis. FastStart Universal SYBR Green Master
(ROX) (Roche, Switzerland) and StepOneTM Real-Time
PCR System (Applied Biosystems, USA) were used for
cDNA amplification. A tubulin gene from P. tenuiflora
(PutTubulin) was cloned and used as an internal standard
to normalize the expression data. Primers used for the
amplification of PtNHA1, PutNHX and PutTubulin are
shown in Table S2. 25S rRNA primers introduced by Jain
et al. (2006) were used for an internal standard of rice.
Primers used for the amplification of OsSOS1 and OsNHX1
is also shown in Table S2. The PCR was performed as
follows: 95�C for 10 min, followed by 40 cycles of 95�C
for 15 s and 60�C for 30 s. The experiments were carried
out in triplicate. A ten-fold serial dilution of 1/5 cDNA
mixture of all treatments were used for the standard curve.
Generation of transgenic rice plants overexpressing
PtNHA1 and PutNHX genes
PtNHA1 and PutNHX were subcloned into pActnos/Hm2
vector. The vector supports the Gateway� system (Invit-
rogen, USA), and carries a hygromycin resistance gene.
PtNHA1 and PutNHX were under the control of the actin
promoter. The resultant pActnos-PtNHA1 and pActnos-
PutNHX constructs were transformed into rice plants (cv.
Nipponbare) by Agrobacterium (strain EHA 105)-mediated
transformation of calli (Toki 1997). Transformants selected
on plates containing hygromycin were transferred to soil
and the T0 seeds were obtained by self-pollination.
Hygromycin-based selection and plant cultivation were
repeated to obtain the T1 and T2 seeds. Total RNA was
extracted from 3-week-old T2 plants using RNeasy� Plant
Mini Kit (QIAGEN, Germany) and the expression levels of
PtNHA1 or PutNHX mRNA were checked by RNA gel
blotting (Fig. S1). Probes were designed based on the
30-UTR sequences of PtNHA1 and PutNHX, and the
primers used are presented in Table S3. The T2 plants were
used for stress tolerance tests.
Characterization of transgenic rice plants
overexpressing PtNHA1 or PutNHX
T2 seeds were sterilized and grown on nutrient solution in a
growth chamber for 2 weeks before stress treatments.
To compare stress tolerance, plants were subjected to
150 mM NaCl and 60 mM NaHCO3 for 3 days, and sub-
sequently transferred to nutrient solution without NaCl/
NaHCO3, and allowed to grow for another 1 week. For the
measurement of shoot and root lengths, plants subjected to
100 mM NaCl or 50 mM NaHCO3 for 3 days were used.
To measure ion contents, 14-day-old plants were treated
with 100 mM NaCl or 50 mM NaHCO3 for 3 days. Roots
were separated from shoots and washed thoroughly by tap
water. The roots and shoots were dried in an oven at 70�C
for 3 days. Dried plants were crushed with Multi-beads
shocker� (Yasui kikai, Japan) and inubated in 1 M HCl
overnight, gently shaking. Ion concentrations were mea-
sured using an atomic absorption photometer AA-670G
(Shimadzu, Japan). The experiment was performed in
triplicate.
Results
Growth of P. tenuiflora and rice under salt stress
conditions
Treatment with 100 mM NaCl significantly inhibited
the growth of rice but slightly increased the growth of
P. tenuiflora (Fig. 1). The growth of P. tenuiflora was not
significantly inhibited by NaHCO3 at concentrations up to
300 mM. On the other hand, 100 mM NaHCO3 had det-
rimental effect on the growth of rice.
Cloning and characterization of PtNHA1 and PutNHX
We cloned NHA and NHX from P. tenuiflora. PtNHA1 has
already been cloned by Wang et al. (2011), but we inde-
pendently cloned it using our original primer pair shown in
Table S1. Primers designed to amplify the conserved
region of the NHA and NHX genes in P. tenuiflora were
based on existing plant NHA and NHX genes. They yielded
DNA fragments of 450 and 750 bp, respectively, and their
sequences confirmed that they were partial sequences of
J Plant Res
123

NHA and NHX. The full-length cDNAs were amplified by
50- and 30-RACE techniques. Since the obtained cDNA
sequence of NHA was a little different from reported
PtNHA1 sequence (the deduced amino acid sequences are
compared in Fig. S2a), we registered it at GenBank
(accession no. AB628205). However, as it has been already
shown that NHA1 of P. tenuiflora is encoded by a single
copy gene (Wang et al. 2011), ours seems to be encoded by
the same gene, PtNHA1. Therefore, we henceforth call it
PtNHA1.
The NHX was named PutNHX (accession no. AB628206).
PutNHX cDNA contained a predicted ORF of 1,617 bp that
appears to encode a protein of 538 amino acids. The protein
is predicted to have 9 transmembrane domains, and a Na?/
H? exchange domain. A putative amiloride binding domain
that is conserved among plant NHXs (Yamaguchi et al.
2003) and a potential glycosylation site, which has also been
predicted for other plant NHXs (Sato and Sakaguchi 2005),
were also conserved in PutNHX (Fig. S2b). In a phylogenic
tree (Fig. S3), PutNHX clustered in the intracellular group
class I, suggesting that it is predicted to be localized at the
vacuolar membrane. Its sequence was highly similar to those
of rice and wheat homologs.
Expression of PtNHA1 and PutNHX compared
with OsSOS1 and OsNHX1
The effect of 300 mM Na? on the expression levels of
NHA was greater in roots than in shoots in both species
(Fig. 2a, b). In rice shoots, NHA expression was increased
more by NaCl treatment than by NaHCO3 treatment, and
peaked after 24 h (Fig. 2b). In P. tenuiflora roots (Fig. 2c),
both treatments increased the PtNHA1 levels dramatically
within the first 12 h. Under NaCl treatment, the level
reached a plateau at 24 h, corresponding to an 11-fold
increase, while under NaHCO3 treatment it peaked at 12 h.
On the other hand, the increase of the expression of NHA in
the rice roots was much smaller under both treatments than
in the P. tenuiflora roots.
The NHX expression level in P. tenuiflora shoots
exposed to NaCl and NaHCO3 peaked after 1 h (Fig. 3a).
Under NaHCO3, it even decreased below the control level
after the peak. On the other hand, in rice shoots (Fig. 3b),
the expression of OsNHX1 increased dramatically with
time under both treatments. One difference was that under
NaCl treatment, the expression peaked at 24 h, whereas
Fig. 1 Growth comparison of rice (left) and P. tenuiflora (right)under salt stresses. Two-week-old P. tenuiflora and 1-week-old
rice plants were subjected to the indicated salt stresses for 5 days.
a, b NaCl-treated rice and P. tenuiflora, respectively. c, d NaHCO3
treatments. Black bars 5 cm
Fig. 2 Effects of NaCl and
NaHCO3 on expression of
NHAs. a, b The relative
expression levels in the shoots
of P. tenuiflora and rice,
respectively, and c, d the values
in the roots of P. tenuiflora and
rice. Tubulin of P. tenuifloraand 25SrRNA of rice were used
as internal controls,
respectively, and the values are
shown as the relative expression
compared to the internal
controls. The relative expression
level in untreated plants (0 h)
was defined as 1. Shown are
mean ± SD (n = 3)
J Plant Res
123

under NaHCO3 treatment, it was still increasing at 48 h.
The expression levels of NHXs in roots were drastically
higher under NaHCO3 treatment than under NaCl treatment
in both species, suggesting that this response is specific to
an alkaline salt. The increase in expression started earlier in
rice than in P. tenuiflora.
Overexpression of PtNHA1 and PutNHX in rice
PtNHA1 or PutNHX-overexpressing rice plants were gen-
erated. The mRNA expression was confirmed by RNA gel
blotting (Fig. S1). Three days of 150 mM NaCl treatment
severely damaged the WT plant, but only moderately
affected the shoots of PtNHA1-OX3 and PutNHX-OX3
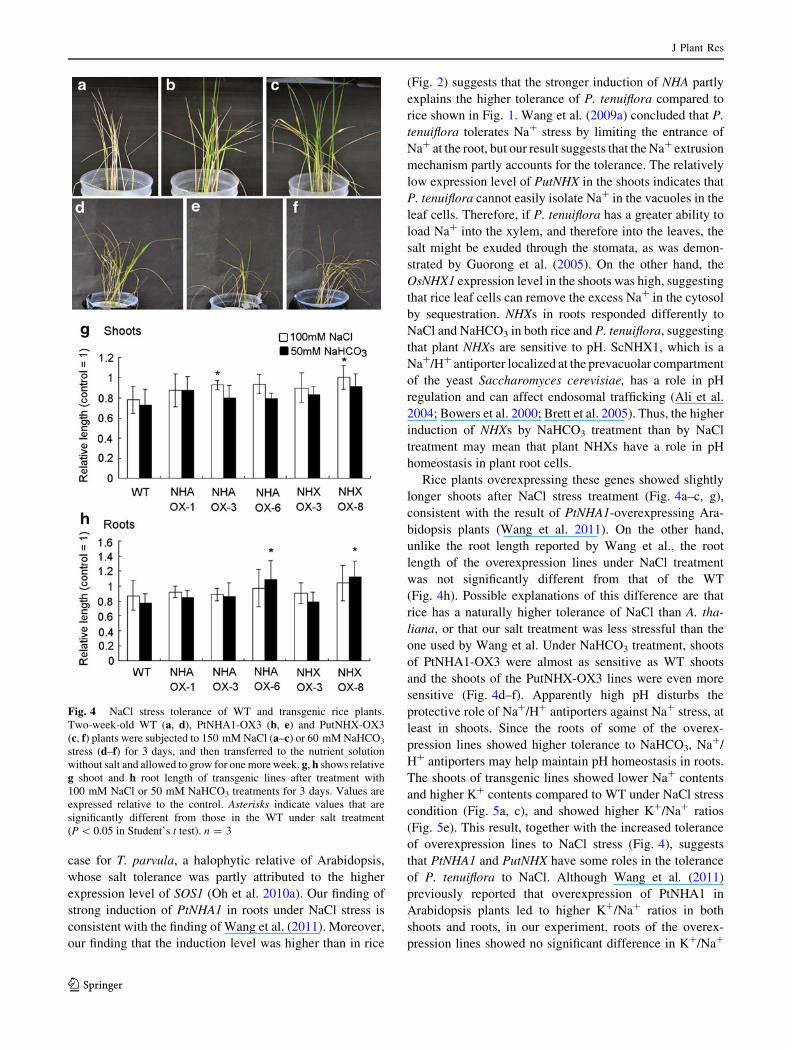
plants, both of which produced new leaves (Fig. 4a–c). On
the other hand, 3 days of 60 mM NaHCO3 treatment had a
detrimental effect on the shoots of the plants (Fig. 4d–f).
After 1 week of recovery, the WT and PtNHA1-OX3
plants produced only a few new leaves, whereas the Put-
NHX-OX3 overexpression line died. In some of the over-
expression lines, shoot length was less affected by NaCl
than it was in the WT (Fig. 4g, white bars), whereas under
NaHCO3 stress no significant difference in relative shoot
lengths was observed (Fig. 4g, black bars). On the other
hand, relative root lengths under NaCl treatment were not
significantly different between WT and the overexpression
lines (Fig. 4h, white bars), whereas the roots of NHA OX-6
and NHX OX-8 were more tolerant to NaHCO3 than the
WT (Fig. 4h, black bars).
Ion contents were measured using the overexpression
lines PtNHA1-OX1, PtNHA1-OX3, PtNHA1-OX6, Put-
NHX-OX3 and PutNHX-OX8. Shoots of the transgenic
lines had significantly lower Na? contents after 100 mM
NaCl treatment (Fig. 5a) and higher K? contents after
NaCl and 50 mM NaHCO3 treatments compared to WT
(Fig. 5c), although in NaHCO3-treated shoots and roots,
the differences were not so obvious (Fig. 5a–d). The Na?/
K? ratios in shoots were significantly lower in the over-
expression lines than in the WT under both treatments, but
in roots no significant difference was observed (Fig. 5e, f).
Discussion
We focused on the Na?/H? antiporters NHA and NHX as
candidates for the strong tolerance of P. tenuiflora to salt
stress. First we cloned NHA and NHX from P. tenuiflora.
The primary sequence of the NHA we obtained was a little
different from PtNHA1 (Wang et al. 2011), and also from
PtSOS1 (GenBank accession no. GQ452778.1; the three
sequences are shown in Fig. S2a), although it is predicted
that P. tenuiflora has only one copy of NHA (Wang et al.
2011). The minor differences may be the result of the
plants being collected in different places. The NHX-type
antiporter that we cloned seemed to be a homolog of
AtNHX1, which is the most highly expressed type of NHX
in A. thaliana. In an attempt to determine the copy number
of NHX-type antiporters in P. tenuiflora, we tried Southern
blotting using the full-length cDNA sequence of PutNHX
as a probe. However, we could detect only one band (data
not shown). In other plants, NHXs are found in multiple
copies (Fig. S3), so it is unlikely that PutNHX exists in only
one copy in P. tenuiflora. In other plants, it is known that
NHX paralogs share highly conserved sequences at their
transmembrane domains, but the homologies among their
N- and C-termini are low. This may be the reason why we
could detect only one band.
We checked the expression levels of NHA and NHX in
P. tenuiflora, based on the assumption that the expression
levels of the antiporters might be higher in P. tenuiflora
than in rice, leading to higher efficiency of P. tenuiflora to
prevent accumulation of Na? in the cytosol. This was the
Fig. 3 Effects of NaCl and
NaHCO3 on expression of
NHXs. a P. tenuiflora shoots,
b rice shoots, c P. tenuifloraroots and d rice roots. RNA
could not be extracted from the
rice roots subjected to 24 and
48 h of NaHCO3 treatment. The
values are mean ± SD (n = 3)
J Plant Res
123
case for T. parvula, a halophytic relative of Arabidopsis,
whose salt tolerance was partly attributed to the higher
expression level of SOS1 (Oh et al. 2010a). Our finding of
strong induction of PtNHA1 in roots under NaCl stress is
consistent with the finding of Wang et al. (2011). Moreover,
our finding that the induction level was higher than in rice
(Fig. 2) suggests that the stronger induction of NHA partly
explains the higher tolerance of P. tenuiflora compared to
rice shown in Fig. 1. Wang et al. (2009a) concluded that P.
tenuiflora tolerates Na? stress by limiting the entrance of
Na? at the root, but our result suggests that the Na? extrusion
mechanism partly accounts for the tolerance. The relatively
low expression level of PutNHX in the shoots indicates that
P. tenuiflora cannot easily isolate Na? in the vacuoles in the
leaf cells. Therefore, if P. tenuiflora has a greater ability to
load Na? into the xylem, and therefore into the leaves, the
salt might be exuded through the stomata, as was demon-
strated by Guorong et al. (2005). On the other hand, the
OsNHX1 expression level in the shoots was high, suggesting
that rice leaf cells can remove the excess Na? in the cytosol
by sequestration. NHXs in roots responded differently to
NaCl and NaHCO3 in both rice and P. tenuiflora, suggesting
that plant NHXs are sensitive to pH. ScNHX1, which is a
Na?/H? antiporter localized at the prevacuolar compartment
of the yeast Saccharomyces cerevisiae, has a role in pH
regulation and can affect endosomal trafficking (Ali et al.
2004; Bowers et al. 2000; Brett et al. 2005). Thus, the higher
induction of NHXs by NaHCO3 treatment than by NaCl
treatment may mean that plant NHXs have a role in pH
homeostasis in plant root cells.
Rice plants overexpressing these genes showed slightly
longer shoots after NaCl stress treatment (Fig. 4a–c, g),
consistent with the result of PtNHA1-overexpressing Ara-
bidopsis plants (Wang et al. 2011). On the other hand,
unlike the root length reported by Wang et al., the root
length of the overexpression lines under NaCl treatment
was not significantly different from that of the WT
(Fig. 4h). Possible explanations of this difference are that
rice has a naturally higher tolerance of NaCl than A. tha-
liana, or that our salt treatment was less stressful than the
one used by Wang et al. Under NaHCO3 treatment, shoots
of PtNHA1-OX3 were almost as sensitive as WT shoots
and the shoots of the PutNHX-OX3 lines were even more
sensitive (Fig. 4d–f). Apparently high pH disturbs the
protective role of Na?/H? antiporters against Na? stress, at
least in shoots. Since the roots of some of the overex-
pression lines showed higher tolerance to NaHCO3, Na?/
H? antiporters may help maintain pH homeostasis in roots.
The shoots of transgenic lines showed lower Na? contents
and higher K? contents compared to WT under NaCl stress
condition (Fig. 5a, c), and showed higher K?/Na? ratios
(Fig. 5e). This result, together with the increased tolerance
of overexpression lines to NaCl stress (Fig. 4), suggests
that PtNHA1 and PutNHX have some roles in the tolerance
of P. tenuiflora to NaCl. Although Wang et al. (2011)
previously reported that overexpression of PtNHA1 in
Arabidopsis plants led to higher K?/Na? ratios in both
shoots and roots, in our experiment, roots of the overex-
pression lines showed no significant difference in K?/Na?
Fig. 4 NaCl stress tolerance of WT and transgenic rice plants.
Two-week-old WT (a, d), PtNHA1-OX3 (b, e) and PutNHX-OX3
(c, f) plants were subjected to 150 mM NaCl (a–c) or 60 mM NaHCO3
stress (d–f) for 3 days, and then transferred to the nutrient solution
without salt and allowed to grow for one more week. g, h shows relative
g shoot and h root length of transgenic lines after treatment with
100 mM NaCl or 50 mM NaHCO3 treatments for 3 days. Values are
expressed relative to the control. Asterisks indicate values that are
significantly different from those in the WT under salt treatment
(P \ 0.05 in Student’s t test). n = 3
J Plant Res
123
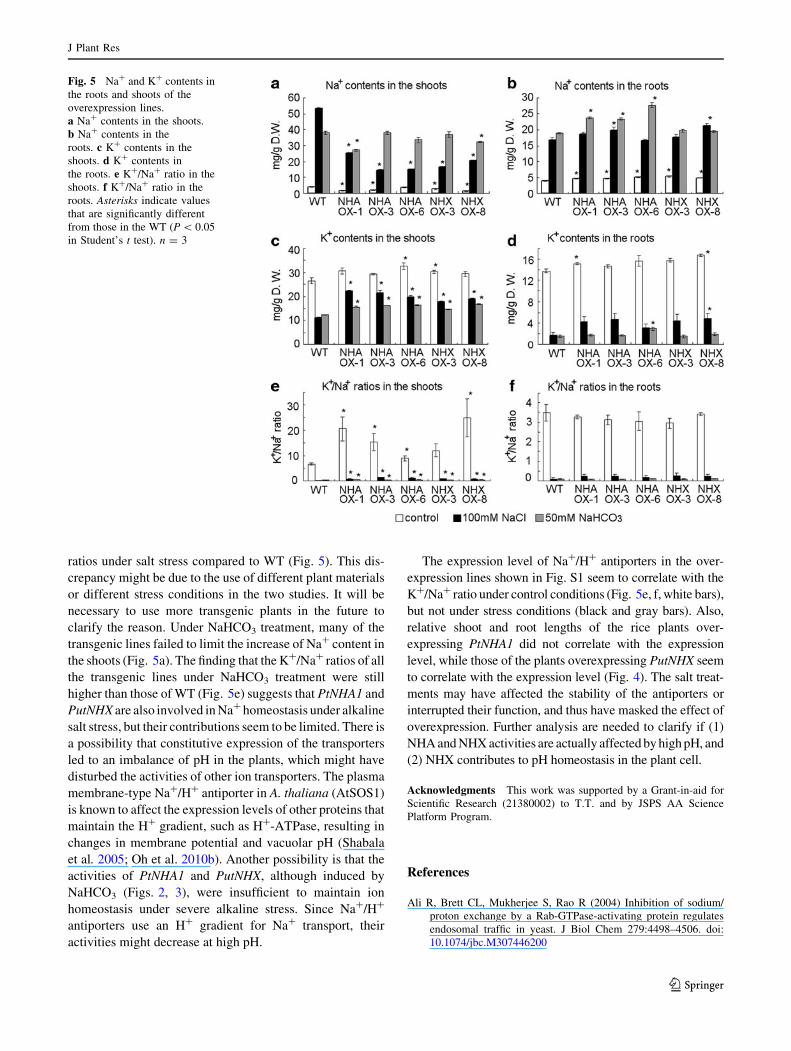
ratios under salt stress compared to WT (Fig. 5). This dis-
crepancy might be due to the use of different plant materials
or different stress conditions in the two studies. It will be
necessary to use more transgenic plants in the future to
clarify the reason. Under NaHCO3 treatment, many of the
transgenic lines failed to limit the increase of Na? content in
the shoots (Fig. 5a). The finding that the K?/Na? ratios of all
the transgenic lines under NaHCO3 treatment were still
higher than those of WT (Fig. 5e) suggests that PtNHA1 and
PutNHX are also involved in Na? homeostasis under alkaline
salt stress, but their contributions seem to be limited. There is
a possibility that constitutive expression of the transporters
led to an imbalance of pH in the plants, which might have
disturbed the activities of other ion transporters. The plasma
membrane-type Na?/H? antiporter in A. thaliana (AtSOS1)
is known to affect the expression levels of other proteins that
maintain the H? gradient, such as H?-ATPase, resulting in
changes in membrane potential and vacuolar pH (Shabala
et al. 2005; Oh et al. 2010b). Another possibility is that the
activities of PtNHA1 and PutNHX, although induced by
NaHCO3 (Figs. 2, 3), were insufficient to maintain ion
homeostasis under severe alkaline stress. Since Na?/H?
antiporters use an H? gradient for Na? transport, their
activities might decrease at high pH.
The expression level of Na?/H? antiporters in the over-
expression lines shown in Fig. S1 seem to correlate with the
K?/Na? ratio under control conditions (Fig. 5e, f, white bars),
but not under stress conditions (black and gray bars). Also,
relative shoot and root lengths of the rice plants over-
expressing PtNHA1 did not correlate with the expression
level, while those of the plants overexpressing PutNHX seem
to correlate with the expression level (Fig. 4). The salt treat-
ments may have affected the stability of the antiporters or
interrupted their function, and thus have masked the effect of
overexpression. Further analysis are needed to clarify if (1)
NHA and NHX activities are actually affected by high pH, and
(2) NHX contributes to pH homeostasis in the plant cell.
Acknowledgments This work was supported by a Grant-in-aid for
Scientific Research (21380002) to T.T. and by JSPS AA Science
Platform Program.
References
Ali R, Brett CL, Mukherjee S, Rao R (2004) Inhibition of sodium/
proton exchange by a Rab-GTPase-activating protein regulates
endosomal traffic in yeast. J Biol Chem 279:4498–4506. doi:
10.1074/jbc.M307446200
Fig. 5 Na? and K? contents in
the roots and shoots of the
overexpression lines.
a Na? contents in the shoots.
b Na? contents in the
roots. c K? contents in the
shoots. d K? contents in
the roots. e K?/Na? ratio in the
shoots. f K?/Na? ratio in the
roots. Asterisks indicate values
that are significantly different
from those in the WT (P \ 0.05
in Student’s t test). n = 3
J Plant Res
123

Apse MP (1999) Salt tolerance conferred by overexpression of a
vacuolar Na?/H? antiport in Arabidopsis. Science 285:1256–
1258. doi:10.1126/science.285.5431.1256
Apse MP, Blumwald E (2007) Na? transport in plants. FEBS Lett
581:2247–2254. doi:10.1016/j.febslet.2007.04.014
Apse MP, Sottosanto JB, Blumwald E (2003) Vacuolar cation/H?
exchange, ion homeostasis, and leaf development are altered in a
T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar
Na?/H? antiporter. Plant J 36:229–239. doi:10.1046/j.1365-
313X.2003.01871.x
Ardie SW, Xie L, Takahashi R et al (2009) Cloning of a high-affinity
K? transporter gene PutHKT2;1 from Puccinellia tenuiflora and
its functional comparison with OsHKT2;1 from rice in yeast and
Arabidopsis. J Exp Bot 60:3491–3502. doi:10.1093/jxb/erp184
Ardie SW, Liu S, Takano T (2010) Expression of the AKT1-type K?
channel gene from Puccinellia tenuiflora, PutAKT1, enhances
salt tolerance in Arabidopsis. Plant Cell Rep 29:865–874. doi:
10.1007/s00299-010-0872-2
Ardie SW, Nishiuchi S, Liu S, Takano T (2011) Ectopic expression of
the K? channel b subunits from Puccinellia tenuiflora (KPutB1)
and rice (KOB1) alters K? homeostasis of yeast and Arabidopsis.
Mol Biotechnol 48:76–86. doi:10.1007/s12033-010-9349-3
Bowers K, Levi BP, Patel FI, Stevens TH (2000) The sodium/proton
exchanger Nhx1p is required for endosomal protein trafficking in
the yeast Saccharomyces cerevisiae. Mol Biol Cell 11:4277–4294
Brett CL, Tukaye DN, Mukherjee S, Rao R (2005) The yeast
endosomal Na? (K?)/H? exchanger Nhx1 regulates cellular pH
to control vesicle trafficking. Mol Biol Cell 16:1396. doi:
10.1091/mbc.E04
Guorong S, Yongzhen P, Hongbo S et al (2005) Does Puccineliatenuiflora have the ability of salt exudation? Colloids Surf B
Biointerfaces 46:197–203. doi:10.1016/j.colsurfb.2005.11.003
Jain M, Nijhawan A, Tyagi AK, Khurana JP (2006) Validation of
housekeeping genes as internal control for studying gene
expression in rice by quantitative real-time PCR. Biochem
Biophys Res Commun 345:646–651. doi:10.1016/j.bbrc.2006.
04.140
Liu H, Zhang X, Takano T, Liu S (2009) Characterization of a
PutCAX1 gene from Puccinellia tenuiflora that confers Ca2? and
Ba2? tolerance in yeast. Biochem Biophys Res Commun
383:392–396. doi:10.1016/j.bbrc.2009.04.042
Martınez-Atienza J, Jiang X, Garciadeblas B et al (2007) Conserva-
tion of the salt overly sensitive pathway in rice. Plant Physiol
143:1001–1012. doi:10.1104/pp.106.092635
Maughan PJ, Turner TB, Coleman CE et al (2009) Characterization of
Salt Overly Sensitive 1 (SOS1) gene homoeologs in quinoa
(Chenopodium quinoa Willd.). Genome 52:647–57. doi:10.1139/
G09-041
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu
Rev Plant Biol 59:651–681. doi:10.1146/annurev.arplant.59.
032607.092911
Oh D-H, Dassanayake M, Haas JS et al (2010a) Genome structures
and halophyte-specific gene expression of the extremophile
Thellungiella parvula in comparison with Thellungiella salsugi-nea (Thellungiella halophila) and Arabidopsis. Plant Physiol
154:1040–1052. doi:10.1104/pp.110.163923
Oh D-H, Lee SY, Bressan RA, Yun DJ, Bohnert HJ (2010b)
Intracellular consequences of SOS1 deficiency during salt stress.
J Exp Bot 61:1205–1213. doi:10.1093/jxb/erp391
Olıas R, Eljakaoui Z, Li J et al (2009) The plasma membrane Na?/H?
antiporter SOS1 is essential for salt tolerance in tomato and
affects the partitioning of Na? between plant organs. Plant Cell
Environ 32:904–916. doi:10.1111/j.1365-3040.2009.01971.x
Peng Y-H, Zhu Y-F, Mao Y-Q et al (2004) Alkali grass resists salt
stress through high [K?] and an endodermis barrier to Na?.
J Exp Bot 55:939–949. doi:10.1093/jxb/erh071
Rodrıguez-Rosales MP, Galvez FJ, Huertas R et al (2009) Plant NHX
cation/proton antiporters. Plant Signal Behav 4:265–276
Sato Y, Sakaguchi M (2005) Topogenic properties of transmembrane
segments of Arabidopsis thaliana NHX1 reveal a common
topology model of the Na?/H? exchanger family. J Biochem
138:425–431. doi:10.1093/jb/mvi132
Shabala L, Cuin TA, Newman IA, Shabala S (2005) Salinity-induced ion
flux patterns from the excised roots of Arabidopsis sos mutants.
Planta 222:1041–1050. doi:10.1007/s00425-005-0074-2
Shi H, Ishitani M, Kim C, Zhu JK (2000) The Arabidopsis thalianasalt tolerance gene SOS1 encodes a putative Na?/H? antiporter.
Proc Natl Acad Sci USA 97:6896–6901. doi:10.1073/pnas.
120170197
Shi H, Quintero FJ, Pardo JM, Zhu JK (2002) The putative plasma
membrane Na?/H? antiporter SOS1 controls long-distance Na?
transport in plants. Plant Cell 14:465. doi:10.1105/tpc.010371.et
Shi H, B-ha Lee, Wu S-J, Zhu J-K (2003) Overexpression of a plasma
membrane Na?/H? antiporter gene improves salt tolerance in
Arabidopsis thaliana. Nat Biotechnol 21:81–85. doi:10.1038/
nbt766
Takahashi R, Liu S, Takano T (2009) Isolation and characterization of
plasma membrane Na?/H? antiporter genes from salt-sensitive
and salt-tolerant reed plants. J Plant Physiol 166:301–309. doi:
10.1016/j.jplph.2008.04.002
Toki S (1997) Rapid and efficient Agrobacterium-mediated transfor-
mation in rice. Plant Mol Biol Rep 15:16–21. doi:10.1016/j.cyto.
2011.05.002
Tuteja N (2007) Mechanisms of high salinity tolerance in plants.
Methods Enzymol 428:419–438. doi:10.1016/S0076-6879(07)28
024-3
Wang Y, Yang C, Liu G et al (2007a) Microarray and suppression
subtractive hybridization analyses of gene expression in Pucc-inellia tenuiflora after exposure to NaHCO3. Plant Sci 173:309–
320. doi:10.1016/j.plantsci.2007.06.011
Wang Y, Chu Y, Liu G et al (2007b) Identification of expressed sequence
tags in an alkali grass (Puccinellia tenuiflora) cDNA library. J Plant
Physiol 164:78–89. doi:10.1016/j.jplph.2005.12.006
Wang Y, Yang C, Liu G, Jiang J (2007c) Development of a cDNA
microarray to identify gene expression of Puccinellia tenuifloraunder saline-alkali stress. Plant Physiol Biochem 45:567–576.
doi:10.1016/j.plaphy.2007.05.006 PPB/Societe francaise de
physiologie vegetale
Wang C-M, Zhang J-L, Liu X-S et al (2009a) Puccinellia tenuifloramaintains a low Na? level under salinity by limiting unidirec-
tional Na? influx resulting in a high selectivity for K? over Na?.
Plant Cell Environ 32:486–496. doi:10.1111/j.1365-3040.2009.
01942.x
Wang L, Seki K, Miyazaki T, Ishihama Y (2009b) The causes of soil
alkalinization in the Songnen Plain of Northeast China. Paddy
Water Environ 7:259–270. doi:10.1007/s10333-009-0166-x
Wang X, Yang R, Wang B et al (2011) Functional characterization of
a plasma membrane Na?/H? antiporter from alkali grass
(Puccinellia tenuiflora). Mol Biol Rep 38:4813–4822. doi:
10.1007/s11033-010-0624-y
Wu SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salt
tolerance and potassium acquisition. Plant Cell 8:617–627. doi:
10.1105/tpc.8.4.617
Wu Y, Ding N, Zhao X et al (2007) Molecular characterization of
PeSOS1: the putative Na?/H? antiporter of Populus euphratica.
Plant Mol Biol 65:1–11. doi:10.1007/s11103-007-9170-y
Yamaguchi T, Apse MP, Shi H, Blumwald E (2003) Topological
analysis of a plant vacuolar Na?/H? antiporter reveals a luminal
C terminus that regulates antiporter cation selectivity. Proc Natl
Acad Sci USA 100:12510–12515. doi:10.1073/pnas.2034966100
J Plant Res
123
Related Documents











